
Manipulação do efeito de Warburg em células de mamífero em...
43
UNIVERSIDADE DE LISBOA FACULDADE DE CIÊNCIAS DEPARTAMENTO DE BIOLOGIA VEGETAL Manipulação do efeito de Warburg em células de mamífero em cultura: implicações para o metabolismo energético Miguel Ricardo Guerreiro MESTRADO EM BIOLOGIA MOLECULAR E GENÉTICA 2011
-
Upload
nguyenliem -
Category
Documents
-
view
212 -
download
0
Transcript of Manipulação do efeito de Warburg em células de mamífero em...

UNIVERSIDADE DE LISBOA
FACULDADE DE CIEcircNCIAS
DEPARTAMENTO DE BIOLOGIA VEGETAL
Manipulaccedilatildeo do efeito de Warburg em ceacutelulas de mamiacutefero em cultura
implicaccedilotildees para o metabolismo energeacutetico
Miguel Ricardo Guerreiro
MESTRADO EM BIOLOGIA MOLECULAR E GENEacuteTICA
2011
UNIVERSIDADE DE LISBOA
FACULDADE DE CIEcircNCIAS
DEPARTAMENTO DE BIOLOGIA VEGETAL
Manipulaccedilatildeo do efeito de Warburg em ceacutelulas de mamiacutefero em cultura
implicaccedilotildees para o metabolismo energeacutetico
Dissertaccedilatildeo de mestrado orientada por
Doutora Ana Sofia Coroadinha (Instituto de Tecnologia Quiacutemica e Bioloacutegica Instituto de
Biologia Experimental e Tecnoloacutegica Oeiras)
Doutora Maria Filomena Caeiro (Departamento de Biologia Vegetal Faculdade de Ciecircncias
da Universidade de Lisboa Lisboa)
Miguel Ricardo Guerreiro
MESTRADO EM BIOLOGIA MOLECULAR E GENEacuteTICA
2011
i
i AGRADECIMENTOS
Quero prestar os meus sinceros agradecimentos a todos aqueles que directa ou
indirectamente contribuiacuteram para o desenvolvimento deste trabalho Em particular
agrave Doutora Paula Alves pela oportunidade de realizar a minha dissertaccedilatildeo na Unidade
de Tecnologia de Ceacutelulas Animais (UTCA) e agrave Doutora Ana Sofia Coroadinha pela
oportunidade de integrar a equipa de Biotecnologia Molecular e Desenvolvimento de Linhas
Celulares e pela orientaccedilatildeo e disponibilidade
agrave Ana Filipa Rodrigues pela orientaccedilatildeo teacutecnica e cientiacutefica pela boa disposiccedilatildeo e
compreensatildeo e pelo incansaacutevel auxiacutelio no trabalho laboratorial e na elaboraccedilatildeo da presente
dissertaccedilatildeo
agrave Doutora Maria Filomena Caeiro pela orientaccedilatildeo e disposiccedilatildeo a ajudar prontamente
A todos os professores do meu percurso acadeacutemico pelas boas memoacuterias e ensinamentos que
me preparam da melhor forma possiacutevel para esta difiacutecil mas proveitosa jornada
ao Doutor Rui Gardner e agrave Telma Lopes do Laboratoacuterio de Citometria de Fluxo do
Instituto Gulbenkian de Ciecircncia pela ajuda teacutecnica necessaacuteria ao prosseguimento do
presente trabalho
a todos os colegas da UTCA por me terem recebido da melhor forma e por me terem
ajudado sempre que precisava Em particular agrave Vanessa Bandeira Heacutelio Tomaacutes Paulo
Fernandes e Doutora Rute Castro por todo o apoio e disponibilidade
a todos os meus familiares em particular aos meus pais e agrave minha irmatilde por sempre
terem acreditado em mim e pelo seu apoio nos momentos difiacuteceis
a todos os meus amigos por todo o apoio e momentos de diversatildeo e descontracccedilatildeo
A todos o meu muito obrigado
ii
ii RESUMO
A maioria das linhas celulares de mamiacutefero usadas na produccedilatildeo de biofaacutermacos
complexos apresenta um elevado consumo de glucose e acumulaccedilatildeo de quantidades
significativas de lactato ndash fenoacutetipo lactogeacutenico associado ao efeito de Warburg ndash reduzindo a
produtividade e qualidade do biofaacutermaco Este trabalho teve como objectivo a manipulaccedilatildeo
deste fenoacutetipo procurando reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do
metabolismo energeacutetico com consequente aumento da produtividade e qualidade de
partiacuteculas virais e anticorpos As linhas celulares 293 FLEX GFP (produtora de retroviacuterus)
MDCK E1 (produtora de adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram
silenciadas para o factor 1 indutiacutevel por hipoxia (HIF1) e cinases 1 e 3 da piruvato
desidrogenase (PDK13) os principais efectores deste fenoacutetipo
Nas ceacutelulas RPE A6 o silenciamento de HIF1 originou ceacutelulas com baixa capacidade
proliferativa sugerindo a reversatildeo da imortalizaccedilatildeo e entrada em senescecircncia Sem alteraccedilotildees
aparentes no crescimento celular nas ceacutelulas 293 FLEX GFP observou-se uma diminuiccedilatildeo
das taxas de consumo de glucose e de produccedilatildeo de lactato aliada a aumentos notaacuteveis na
produtividade celular (ateacute 182x em partiacuteculas totais e ateacute 216x em partiacuteculas infecciosas) e
sua qualidade o silenciamento adicional de PDK13 potenciou estes efeitos destacando-se o
aumento adicional (28x) da qualidade do produto final Nas ceacutelulas MDCK E1 natildeo se
obtiveram reduccedilotildees no consumo de glucose e produccedilatildeo de lactato e os aumentos na produccedilatildeo
viral e sua qualidade foram moderados Adicionalmente observou-se nas ceacutelulas silenciadas
aumento de poder redutor em 293 FLEX GFP e dos niacuteveis de ATP em MDCK E1 suportando
a reduccedilatildeo do fenoacutetipo lactogeacutenico e aumento de eficiecircncia do metabolismo energeacutetico
Estes resultados evidenciam a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao
efeito de Warburg como potencial ferramenta de engenharia metaboacutelica visando o aumento da
produtividade de biofaacutermacos complexos em cultura de ceacutelulas de mamiacutefero
Palavras-chave efeito de Warburg factor 1 indutiacutevel por hipoxia (HIF1) complexo
da piruvato desidrogenase (PDH) metabolismo energeacutetico produtividade celular
iii
iii ABSTRACT
The majority of mammalian cell lines used in the production of complex
biopharmaceuticals present high rates of glucose consumption accumulating significant
amounts of lactate ndash lactogenic phenotype associated to the Warburg effect ndash which results in
the reduction of cellular productivity and in final product quality The goal of this work was
the manipulation of this phenotype aiming at the reduction of lactate accumulation and the
improvement of energy metabolism efficiency to increase viral particles and antibodies
productivity and quality 293 FLEX GFP (retrovirus producer) MDCK E1 (adenovirus
producer) and RPE A6 (monoclonal antibodies producer) cell lines were silenced for hypoxia-
inducible factor 1 (HIF1) and pyruvate dehydrogenase 1 and 3 (PDK13) the major molecular
effectors of this phenotype
The silencing of HIF1 in RPE A6 cells resulted in cells with low proliferative capacity
suggesting the reversion of immortality and entry into senescence Without relevant impact in
cellular growth the 293 FLEX GFP cells showed a reduction in glucose uptake and lactate
production rates along with a significant increase of cellular productivities (up to 182x in
total and up to 216x in infectious particles) and its quality further silencing of PDK13
potentiated the effects of HIF1 silencing with special focus in the additional increase of final
product quality (28x) In MDCK E1 cells no reduction of glucose uptake and lactate
production rates was observed and viral productivity and its quality increased moderately
Additionally it was observed an increase in intracellular reducting power in 293 FLEX GFP
cells and in ATP levels in MDCK E1 cells further supporting the reduction of lactogenic
phenotype and the improvement of energy metabolism
These results highlight the manipulation of lactogenic phenotype associated to the
Warburg effect as a potential metabolic engineering tool for the improvement of complex
biopharmaceuticals production in cultured mammalian cell lines
Keywords Warburg effect hypoxia inducible factor 1 (HIF1) pyruvate
dehydrogenase complex (PDH) energetic metabolism cellular productivity
iv
iv LISTA DE ABREVIATURAS
Abreviatura Descriccedilatildeo
cDNA ADN complementar (do Inglecircs complementary DNA)
DNA Aacutecido desoxirribonucleico (do Inglecircs Deoxyribonucleic Acid)
ERO Espeacutecies reactivas de oxigeacutenio
GFP Proteiacutena fluorescente verde (do Inglecircs Green Fluorescent Protein)
HIF1 Factor 1 indutiacutevel por hipoxia (do Inglecircs Hypoxia Inducible Factor 1)
LDH Lactato desidrogenase (do Inglecircs Lactate Dehydrogenase)
mRNA ARN mensageiro (do Inglecircs messenger RNA)
ntshRNA non-target short hairpin RNA
PCR Reacccedilatildeo em cadeia da polimerase (do Inglecircs Polymerase chain reaction)
PDH Complexo da piruvato desidrogenase (do Inglecircs Pyruvate
Dehydrogenase Complex)
PDHE1α Subunidade α do componente E1 do complexo piruvato desidrogenase
(do Inglecircs E1 alpha subunit of pyruvate dehydrogenase complex)
PDK Cinase da piruvato desidrogenase (do Inglecircs Pyruvate Dehydrogenase
Kinase)
PDP Fosfatase da piruvato desidrogenase (do Inglecircs Pyruvate dehydrogenase
phosphatase)
PI Partiacuteculas infecciosas
PT Partiacuteculas totais
qRT-PCR PCR quantitativo por transcriccedilatildeo reversa (do Inglecircs Quantitative reverse
transcription polymerase chain reaction)
RFP Proteiacutena fluorescente vermelha (do Inglecircs Red Fluorescent Protein)
RNA Aacutecido ribonucleico (do Inglecircs Ribonucleic Acid)
shRNA short hairpin RNA
TCA Ciclo dos Aacutecidos Tricarboxiacutelicos (do Inglecircs Tricarboxylic Acid Cycle)
v
V IacuteNDICE
i AGRADECIMENTOS i
ii RESUMO ii
iii ABSTRACT iii
iv LISTA DE ABREVIATURAS iv
1 INTRODUCcedilAtildeO 1
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico 1
12 Imortalizaccedilatildeo celular e o efeito de Warburg 2
13 Impacto do efeito de Warburg em biotecnologia 4
14 Objectivos principais 5
2 MATERIAIS E MEacuteTODOS 6
21 Linhas celulares e meios de cultura 6
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais 6
23 Transduccedilatildeo celular selecccedilatildeo e clonagem 7
24 Estudos de crescimento e anaacutelise de metabolitos 7
25 Extracccedilatildeo de RNA total e qRT-PCR 8
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais8
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais 8
28 Determinaccedilatildeo da actividade da lactato desidrogenase9
29 Western Blot 9
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina 10
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio 10
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade 10
213 Caacutelculos 10
3 RESULTADOS 11
31 Silenciamento de HIF1 11
vi
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3 15
4 DISCUSSAtildeO E CONCLUSAtildeO 24
5 PERSPECTIVAS FUTURAS 29
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS 30
7 ANEXOS 34
1
1 INTRODUCcedilAtildeO
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico
Nos uacuteltimos anos tem-se verificado uma crescente procura de biofaacutermacos complexos
como proteiacutenas recombinantes e partiacuteculas derivadas de viacuterus quer para fins terapecircuticos quer
para fins de diagnoacutesticos in vivo O potencial dos biofaacutermacos derivados de viacuterus em
particular tem vindo a aumentar devido agraves suas diversas aplicaccedilotildees profilaacuteticas terapecircuticas e
cliacutenicas como a vacinaccedilatildeo o tratamento do cancro e a terapia geacutenica 1 Actualmente o
sistema preferencial para a manufacturaccedilatildeo destes produtos biofarmacecircuticos complexos eacute as
ceacutelulas de mamiacutefero usadas para a produccedilatildeo de mais de metade dos biofaacutermacos existentes
no mercado Tal predomiacutenio deve-se agrave sua semelhanccedila com as ceacutelulas humanas no folding de
proteiacutenas e nas modificaccedilotildees poacutes-traducionais como a glicosilaccedilatildeo aumentando a eficiecircncia
terapecircutica e diminuindo o potencial imunogeacutenico do produto final 23
As ceacutelulas de mamiacutefero em cultura agrave semelhanccedila de ceacutelulas transformadas possuem
um metabolismo energeacutetico desregulado consumindo elevadas quantidades de glucose e
acumulando quantidades significativas de lactato com reduzida entrada de carbono derivado
da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) 4 Do ponto de vista da cultura de
ceacutelulas para produccedilatildeo de biofaacutermacos este fenoacutetipo lactogeacutenico tem simultaneamente um
efeito negativo no crescimento e viabilidade celular e na produtividade e qualidade do
biofaacutermaco Este fenoacutemeno eacute ainda mais importante no caso das partiacuteculas derivadas de viacuterus
para terapia geacutenica jaacute que a sua infecciosidade eacute fortemente influenciada por variaccedilotildees no pH
do meio de cultura 5 Diversas estrateacutegias tecircm sido testadas de forma a reduzir os efeitos
nocivos da acumulaccedilatildeo de lactato no meio de cultura fornecimento de glucose de forma
controlada 67
suplementaccedilatildeo do meio de cultura com substratos alternativos agrave glucose 8
remoccedilatildeo dos metabolitos toacutexicos 9 desenvolvimento de meios de cultura com base na anaacutelise
dos fluxos metaboacutelicos permitindo adaptar os seus constituintes agraves necessidades metaboacutelicas
das ceacutelulas 1011
introduccedilatildeo da enzima piruvato carboxilase nas ceacutelulas 12-14
diminuiccedilatildeo da
expressatildeo da lactato desidrogenase atraveacutes da disrupccedilatildeo do gene Ldha por recombinaccedilatildeo
homoacuteloga 15
ou atraveacutes de RNA de interferecircncia 16
ou introduccedilatildeo de um transportador para a
frutose constituindo uma alternativa ao consumo de glucose 17
No entanto estas estrateacutegias
tecircm um sucesso limitado devido aos efeitos compensatoacuterios causados pela apertada regulaccedilatildeo
homeostaacutetica intracelular que natildeo satildeo facilmente alteraacuteveis
2
12 Imortalizaccedilatildeo celular e o efeito de Warburg
Nos anos 20 Otto Warburg observou que as ceacutelulas canceriacutegenas comparativamente
agraves ceacutelulas normais apresentavam elevadas taxas de consumo de glucose secretando a maior
parte do carbono derivado da glucose sob a forma de lactato em vez de o oxidar
completamente 1819
ndash um cenaacuterio que ficou conhecido como efeito de Warburg Estudos
recentes sobre o metabolismo energeacutetico de ceacutelulas canceriacutegenas sugerem que este resulta de
alteraccedilotildees geneacuteticas que ocorrem no processo de tumorigeacutenese principalmente durante a
imortalizaccedilatildeo e transformaccedilatildeo celulares e que levam agrave desregulaccedilatildeo do metabolismo
energeacutetico central e ao aumento da actividade glicoliacutetica 20
De facto existem diversas vias de
sinalizaccedilatildeo comuns ao controlo da proliferaccedilatildeo e do metabolismo energeacutetico que levam agrave
induccedilatildeo deste cenaacuterio como a activaccedilatildeo de oncogenes (MYC) e enzimas glicoliacuteticas ou a
supressatildeo do gene supressor de tumores P53 2021
Interessantemente muitas das vias de
imortalizaccedilatildeo activadas no processo de tumorigeacutenese satildeo tambeacutem recrutadas durante o
estabelecimento de uma linha celular e por isso tendem a exibir caracteriacutesticas metaboacutelicas
associadas ao efeito de Warburg 20
O principal efector molecular do efeito de Warburg eacute o factor 1 indutiacutevel por hipoxia
(HIF1) um factor de transcriccedilatildeo heterodimeacuterico constituiacutedo por uma subunidade β expressa
constitutivamente e uma subunidade α cuja expressatildeo estaacute dependente do oxigeacutenio Apesar
de ser expressa em condiccedilotildees de baixa disponibilidade de oxigeacutenio (hipoxia) a expressatildeo
constitutiva da subunidade α pode ocorrer em normoxia como resultado de alteraccedilotildees nos
sistemas de sinalizaccedilatildeo para degradaccedilatildeo proteica provocadas durante o processo de
imortalizaccedilatildeo celular De facto a sua sobre-expressatildeo eacute caracteriacutestica comum a muitas ceacutelulas
canceriacutegenas 2223
O factor de transcriccedilatildeo HIF1 eacute responsaacutevel pela coordenaccedilatildeo da alteraccedilatildeo
do metabolismo para glicoacutelise com formaccedilatildeo de lactato com consequente aumento do
consumo de glucose e acumulaccedilatildeo de lactato jaacute que promove a expressatildeo de vaacuterios
componentes do metabolismo energeacutetico nomeadamente transportadores de glucose (GLUT-
1) enzimas da via glicoliacutetica e de lactato desidrogenase (Figura 1) 2425
Para aleacutem do papel do HIF1 na promoccedilatildeo da glicoacutelise com formaccedilatildeo de lactato
estudos recentes revelaram uma significativa influecircncia na respiraccedilatildeo mitocondrial Nos
organismos aeroacutebios o TCA possui um papel fundamental no metabolismo central das
ceacutelulas constituindo a via de produccedilatildeo de energia mais eficiente e fornecendo o poder redutor
e intermediaacuterios necessaacuterios para as reacccedilotildees de biossiacutentese Normalmente a glucose eacute
convertida em piruvato no citoplasma o qual eacute posteriormente canalizado para o TCA ao
niacutevel mitocondrial onde eacute oxidado a dioacutexido de carbono e aacutegua A canalizaccedilatildeo do piruvato
3
para o TCA implica a sua conversatildeo em acetil-CoA numa reacccedilatildeo catalisada pelo complexo
multi-enzimaacutetico piruvato desidrogenase (PDH) Sendo esta uma das reacccedilotildees mais
importantes do metabolismo celular estaacute sujeita a um elevado niacutevel de regulaccedilatildeo Como tal o
complexo PDH eacute regulado por um ciclo reversiacutevel de fosforilaccedilatildeo caracterizado pela
fosforilaccedilatildeo (inactivaccedilatildeo) e desfosforilaccedilatildeo (activaccedilatildeo) da subunidade α do componente E1
(PDHE1α) promovida por cinases (PDK) e fosfatases (PDP) respectivamente Na PDHE1α
de mamiacuteferos existem trecircs locais de fosforilaccedilatildeo (Ser232
Ser293
e Ser300
) que podem ser
fosforilados por quatro isoenzimas (PDK1-4) sendo a PDK1 a uacutenica que consegue fosforilar
todos os trecircs locais 2627
Interessantemente a sobre-expressatildeo de PDK1 e PDK3 eacute promovida
por HIF1 levando assim agrave fosforilaccedilatildeo de PDHE1α e inibiccedilatildeo do complexo PDH (Figura 1)
Consequentemente a entrada do carbono derivado da via glicoliacutetica no TCA ocorre em menor
extensatildeo e aumenta a conversatildeo de piruvato a lactato resultando no fenoacutetipo lactogeacutenico
associado ao efeito de Warburg e na desregulaccedilatildeo do metabolismo energeacutetico 2428
Figura 1 Vias do metabolismo energeacutetico reguladas pelo factor de transcriccedilatildeo HIF1 A sobre-expressatildeo de
vaacuterias enzimas da via glicoliacutetica e da enzima lactato desidrogenase (LDH) eacute promovida pelo factor 1 indutiacutevel
por hipoxia (HIF1) levando ao aumento do consumo de glucose e agrave acumulaccedilatildeo de lactato A entrada do
carbono derivado da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) eacute realizada ao niacutevel do complexo
piruvato desidrogenase (PDH) sendo este regulado atraveacutes de um ciclo reversiacutevel de fosforilaccedilatildeo HIF1 promove
a expressatildeo das cinases da piruvato desidrogenase (PDK1-4) resultando na fosforilaccedilatildeo e consequente inibiccedilatildeo
do complexo PDH Esquema adaptado de Kondoh (2008) 20
Glucose
Glucose 6P
Fuctose 6P
Glyceraldehyde-3P
Fuctose 16 BP
Dihydroxyacetone-P
13 Biphosphoglycerate
3 - Phosphoglycerate
2 - Phosphoglycerate
Phosphoenopyruvate Pyruvate
Cytoplasm
Extracellular space
hexokinase
phosphoglucose isomerase
phosphofructokinase
GAPDH
phosphoglycerate
kinase
phosphoglucomutase
enolase
Pyruvate kinase
Pyruvate
Acetyl-CoATCA
PDH
PDH
P
LDH
Lactate
PDK
PDK2
PDK1 PDK3PDK4
aldolase
+
+
+
+
+
++
+
+
HIF1
4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

UNIVERSIDADE DE LISBOA
FACULDADE DE CIEcircNCIAS
DEPARTAMENTO DE BIOLOGIA VEGETAL
Manipulaccedilatildeo do efeito de Warburg em ceacutelulas de mamiacutefero em cultura
implicaccedilotildees para o metabolismo energeacutetico
Dissertaccedilatildeo de mestrado orientada por
Doutora Ana Sofia Coroadinha (Instituto de Tecnologia Quiacutemica e Bioloacutegica Instituto de
Biologia Experimental e Tecnoloacutegica Oeiras)
Doutora Maria Filomena Caeiro (Departamento de Biologia Vegetal Faculdade de Ciecircncias
da Universidade de Lisboa Lisboa)
Miguel Ricardo Guerreiro
MESTRADO EM BIOLOGIA MOLECULAR E GENEacuteTICA
2011
i
i AGRADECIMENTOS
Quero prestar os meus sinceros agradecimentos a todos aqueles que directa ou
indirectamente contribuiacuteram para o desenvolvimento deste trabalho Em particular
agrave Doutora Paula Alves pela oportunidade de realizar a minha dissertaccedilatildeo na Unidade
de Tecnologia de Ceacutelulas Animais (UTCA) e agrave Doutora Ana Sofia Coroadinha pela
oportunidade de integrar a equipa de Biotecnologia Molecular e Desenvolvimento de Linhas
Celulares e pela orientaccedilatildeo e disponibilidade
agrave Ana Filipa Rodrigues pela orientaccedilatildeo teacutecnica e cientiacutefica pela boa disposiccedilatildeo e
compreensatildeo e pelo incansaacutevel auxiacutelio no trabalho laboratorial e na elaboraccedilatildeo da presente
dissertaccedilatildeo
agrave Doutora Maria Filomena Caeiro pela orientaccedilatildeo e disposiccedilatildeo a ajudar prontamente
A todos os professores do meu percurso acadeacutemico pelas boas memoacuterias e ensinamentos que
me preparam da melhor forma possiacutevel para esta difiacutecil mas proveitosa jornada
ao Doutor Rui Gardner e agrave Telma Lopes do Laboratoacuterio de Citometria de Fluxo do
Instituto Gulbenkian de Ciecircncia pela ajuda teacutecnica necessaacuteria ao prosseguimento do
presente trabalho
a todos os colegas da UTCA por me terem recebido da melhor forma e por me terem
ajudado sempre que precisava Em particular agrave Vanessa Bandeira Heacutelio Tomaacutes Paulo
Fernandes e Doutora Rute Castro por todo o apoio e disponibilidade
a todos os meus familiares em particular aos meus pais e agrave minha irmatilde por sempre
terem acreditado em mim e pelo seu apoio nos momentos difiacuteceis
a todos os meus amigos por todo o apoio e momentos de diversatildeo e descontracccedilatildeo
A todos o meu muito obrigado
ii
ii RESUMO
A maioria das linhas celulares de mamiacutefero usadas na produccedilatildeo de biofaacutermacos
complexos apresenta um elevado consumo de glucose e acumulaccedilatildeo de quantidades
significativas de lactato ndash fenoacutetipo lactogeacutenico associado ao efeito de Warburg ndash reduzindo a
produtividade e qualidade do biofaacutermaco Este trabalho teve como objectivo a manipulaccedilatildeo
deste fenoacutetipo procurando reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do
metabolismo energeacutetico com consequente aumento da produtividade e qualidade de
partiacuteculas virais e anticorpos As linhas celulares 293 FLEX GFP (produtora de retroviacuterus)
MDCK E1 (produtora de adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram
silenciadas para o factor 1 indutiacutevel por hipoxia (HIF1) e cinases 1 e 3 da piruvato
desidrogenase (PDK13) os principais efectores deste fenoacutetipo
Nas ceacutelulas RPE A6 o silenciamento de HIF1 originou ceacutelulas com baixa capacidade
proliferativa sugerindo a reversatildeo da imortalizaccedilatildeo e entrada em senescecircncia Sem alteraccedilotildees
aparentes no crescimento celular nas ceacutelulas 293 FLEX GFP observou-se uma diminuiccedilatildeo
das taxas de consumo de glucose e de produccedilatildeo de lactato aliada a aumentos notaacuteveis na
produtividade celular (ateacute 182x em partiacuteculas totais e ateacute 216x em partiacuteculas infecciosas) e
sua qualidade o silenciamento adicional de PDK13 potenciou estes efeitos destacando-se o
aumento adicional (28x) da qualidade do produto final Nas ceacutelulas MDCK E1 natildeo se
obtiveram reduccedilotildees no consumo de glucose e produccedilatildeo de lactato e os aumentos na produccedilatildeo
viral e sua qualidade foram moderados Adicionalmente observou-se nas ceacutelulas silenciadas
aumento de poder redutor em 293 FLEX GFP e dos niacuteveis de ATP em MDCK E1 suportando
a reduccedilatildeo do fenoacutetipo lactogeacutenico e aumento de eficiecircncia do metabolismo energeacutetico
Estes resultados evidenciam a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao
efeito de Warburg como potencial ferramenta de engenharia metaboacutelica visando o aumento da
produtividade de biofaacutermacos complexos em cultura de ceacutelulas de mamiacutefero
Palavras-chave efeito de Warburg factor 1 indutiacutevel por hipoxia (HIF1) complexo
da piruvato desidrogenase (PDH) metabolismo energeacutetico produtividade celular
iii
iii ABSTRACT
The majority of mammalian cell lines used in the production of complex
biopharmaceuticals present high rates of glucose consumption accumulating significant
amounts of lactate ndash lactogenic phenotype associated to the Warburg effect ndash which results in
the reduction of cellular productivity and in final product quality The goal of this work was
the manipulation of this phenotype aiming at the reduction of lactate accumulation and the
improvement of energy metabolism efficiency to increase viral particles and antibodies
productivity and quality 293 FLEX GFP (retrovirus producer) MDCK E1 (adenovirus
producer) and RPE A6 (monoclonal antibodies producer) cell lines were silenced for hypoxia-
inducible factor 1 (HIF1) and pyruvate dehydrogenase 1 and 3 (PDK13) the major molecular
effectors of this phenotype
The silencing of HIF1 in RPE A6 cells resulted in cells with low proliferative capacity
suggesting the reversion of immortality and entry into senescence Without relevant impact in
cellular growth the 293 FLEX GFP cells showed a reduction in glucose uptake and lactate
production rates along with a significant increase of cellular productivities (up to 182x in
total and up to 216x in infectious particles) and its quality further silencing of PDK13
potentiated the effects of HIF1 silencing with special focus in the additional increase of final
product quality (28x) In MDCK E1 cells no reduction of glucose uptake and lactate
production rates was observed and viral productivity and its quality increased moderately
Additionally it was observed an increase in intracellular reducting power in 293 FLEX GFP
cells and in ATP levels in MDCK E1 cells further supporting the reduction of lactogenic
phenotype and the improvement of energy metabolism
These results highlight the manipulation of lactogenic phenotype associated to the
Warburg effect as a potential metabolic engineering tool for the improvement of complex
biopharmaceuticals production in cultured mammalian cell lines
Keywords Warburg effect hypoxia inducible factor 1 (HIF1) pyruvate
dehydrogenase complex (PDH) energetic metabolism cellular productivity
iv
iv LISTA DE ABREVIATURAS
Abreviatura Descriccedilatildeo
cDNA ADN complementar (do Inglecircs complementary DNA)
DNA Aacutecido desoxirribonucleico (do Inglecircs Deoxyribonucleic Acid)
ERO Espeacutecies reactivas de oxigeacutenio
GFP Proteiacutena fluorescente verde (do Inglecircs Green Fluorescent Protein)
HIF1 Factor 1 indutiacutevel por hipoxia (do Inglecircs Hypoxia Inducible Factor 1)
LDH Lactato desidrogenase (do Inglecircs Lactate Dehydrogenase)
mRNA ARN mensageiro (do Inglecircs messenger RNA)
ntshRNA non-target short hairpin RNA
PCR Reacccedilatildeo em cadeia da polimerase (do Inglecircs Polymerase chain reaction)
PDH Complexo da piruvato desidrogenase (do Inglecircs Pyruvate
Dehydrogenase Complex)
PDHE1α Subunidade α do componente E1 do complexo piruvato desidrogenase
(do Inglecircs E1 alpha subunit of pyruvate dehydrogenase complex)
PDK Cinase da piruvato desidrogenase (do Inglecircs Pyruvate Dehydrogenase
Kinase)
PDP Fosfatase da piruvato desidrogenase (do Inglecircs Pyruvate dehydrogenase
phosphatase)
PI Partiacuteculas infecciosas
PT Partiacuteculas totais
qRT-PCR PCR quantitativo por transcriccedilatildeo reversa (do Inglecircs Quantitative reverse
transcription polymerase chain reaction)
RFP Proteiacutena fluorescente vermelha (do Inglecircs Red Fluorescent Protein)
RNA Aacutecido ribonucleico (do Inglecircs Ribonucleic Acid)
shRNA short hairpin RNA
TCA Ciclo dos Aacutecidos Tricarboxiacutelicos (do Inglecircs Tricarboxylic Acid Cycle)
v
V IacuteNDICE
i AGRADECIMENTOS i
ii RESUMO ii
iii ABSTRACT iii
iv LISTA DE ABREVIATURAS iv
1 INTRODUCcedilAtildeO 1
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico 1
12 Imortalizaccedilatildeo celular e o efeito de Warburg 2
13 Impacto do efeito de Warburg em biotecnologia 4
14 Objectivos principais 5
2 MATERIAIS E MEacuteTODOS 6
21 Linhas celulares e meios de cultura 6
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais 6
23 Transduccedilatildeo celular selecccedilatildeo e clonagem 7
24 Estudos de crescimento e anaacutelise de metabolitos 7
25 Extracccedilatildeo de RNA total e qRT-PCR 8
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais8
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais 8
28 Determinaccedilatildeo da actividade da lactato desidrogenase9
29 Western Blot 9
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina 10
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio 10
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade 10
213 Caacutelculos 10
3 RESULTADOS 11
31 Silenciamento de HIF1 11
vi
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3 15
4 DISCUSSAtildeO E CONCLUSAtildeO 24
5 PERSPECTIVAS FUTURAS 29
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS 30
7 ANEXOS 34
1
1 INTRODUCcedilAtildeO
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico
Nos uacuteltimos anos tem-se verificado uma crescente procura de biofaacutermacos complexos
como proteiacutenas recombinantes e partiacuteculas derivadas de viacuterus quer para fins terapecircuticos quer
para fins de diagnoacutesticos in vivo O potencial dos biofaacutermacos derivados de viacuterus em
particular tem vindo a aumentar devido agraves suas diversas aplicaccedilotildees profilaacuteticas terapecircuticas e
cliacutenicas como a vacinaccedilatildeo o tratamento do cancro e a terapia geacutenica 1 Actualmente o
sistema preferencial para a manufacturaccedilatildeo destes produtos biofarmacecircuticos complexos eacute as
ceacutelulas de mamiacutefero usadas para a produccedilatildeo de mais de metade dos biofaacutermacos existentes
no mercado Tal predomiacutenio deve-se agrave sua semelhanccedila com as ceacutelulas humanas no folding de
proteiacutenas e nas modificaccedilotildees poacutes-traducionais como a glicosilaccedilatildeo aumentando a eficiecircncia
terapecircutica e diminuindo o potencial imunogeacutenico do produto final 23
As ceacutelulas de mamiacutefero em cultura agrave semelhanccedila de ceacutelulas transformadas possuem
um metabolismo energeacutetico desregulado consumindo elevadas quantidades de glucose e
acumulando quantidades significativas de lactato com reduzida entrada de carbono derivado
da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) 4 Do ponto de vista da cultura de
ceacutelulas para produccedilatildeo de biofaacutermacos este fenoacutetipo lactogeacutenico tem simultaneamente um
efeito negativo no crescimento e viabilidade celular e na produtividade e qualidade do
biofaacutermaco Este fenoacutemeno eacute ainda mais importante no caso das partiacuteculas derivadas de viacuterus
para terapia geacutenica jaacute que a sua infecciosidade eacute fortemente influenciada por variaccedilotildees no pH
do meio de cultura 5 Diversas estrateacutegias tecircm sido testadas de forma a reduzir os efeitos
nocivos da acumulaccedilatildeo de lactato no meio de cultura fornecimento de glucose de forma
controlada 67
suplementaccedilatildeo do meio de cultura com substratos alternativos agrave glucose 8
remoccedilatildeo dos metabolitos toacutexicos 9 desenvolvimento de meios de cultura com base na anaacutelise
dos fluxos metaboacutelicos permitindo adaptar os seus constituintes agraves necessidades metaboacutelicas
das ceacutelulas 1011
introduccedilatildeo da enzima piruvato carboxilase nas ceacutelulas 12-14
diminuiccedilatildeo da
expressatildeo da lactato desidrogenase atraveacutes da disrupccedilatildeo do gene Ldha por recombinaccedilatildeo
homoacuteloga 15
ou atraveacutes de RNA de interferecircncia 16
ou introduccedilatildeo de um transportador para a
frutose constituindo uma alternativa ao consumo de glucose 17
No entanto estas estrateacutegias
tecircm um sucesso limitado devido aos efeitos compensatoacuterios causados pela apertada regulaccedilatildeo
homeostaacutetica intracelular que natildeo satildeo facilmente alteraacuteveis
2
12 Imortalizaccedilatildeo celular e o efeito de Warburg
Nos anos 20 Otto Warburg observou que as ceacutelulas canceriacutegenas comparativamente
agraves ceacutelulas normais apresentavam elevadas taxas de consumo de glucose secretando a maior
parte do carbono derivado da glucose sob a forma de lactato em vez de o oxidar
completamente 1819
ndash um cenaacuterio que ficou conhecido como efeito de Warburg Estudos
recentes sobre o metabolismo energeacutetico de ceacutelulas canceriacutegenas sugerem que este resulta de
alteraccedilotildees geneacuteticas que ocorrem no processo de tumorigeacutenese principalmente durante a
imortalizaccedilatildeo e transformaccedilatildeo celulares e que levam agrave desregulaccedilatildeo do metabolismo
energeacutetico central e ao aumento da actividade glicoliacutetica 20
De facto existem diversas vias de
sinalizaccedilatildeo comuns ao controlo da proliferaccedilatildeo e do metabolismo energeacutetico que levam agrave
induccedilatildeo deste cenaacuterio como a activaccedilatildeo de oncogenes (MYC) e enzimas glicoliacuteticas ou a
supressatildeo do gene supressor de tumores P53 2021
Interessantemente muitas das vias de
imortalizaccedilatildeo activadas no processo de tumorigeacutenese satildeo tambeacutem recrutadas durante o
estabelecimento de uma linha celular e por isso tendem a exibir caracteriacutesticas metaboacutelicas
associadas ao efeito de Warburg 20
O principal efector molecular do efeito de Warburg eacute o factor 1 indutiacutevel por hipoxia
(HIF1) um factor de transcriccedilatildeo heterodimeacuterico constituiacutedo por uma subunidade β expressa
constitutivamente e uma subunidade α cuja expressatildeo estaacute dependente do oxigeacutenio Apesar
de ser expressa em condiccedilotildees de baixa disponibilidade de oxigeacutenio (hipoxia) a expressatildeo
constitutiva da subunidade α pode ocorrer em normoxia como resultado de alteraccedilotildees nos
sistemas de sinalizaccedilatildeo para degradaccedilatildeo proteica provocadas durante o processo de
imortalizaccedilatildeo celular De facto a sua sobre-expressatildeo eacute caracteriacutestica comum a muitas ceacutelulas
canceriacutegenas 2223
O factor de transcriccedilatildeo HIF1 eacute responsaacutevel pela coordenaccedilatildeo da alteraccedilatildeo
do metabolismo para glicoacutelise com formaccedilatildeo de lactato com consequente aumento do
consumo de glucose e acumulaccedilatildeo de lactato jaacute que promove a expressatildeo de vaacuterios
componentes do metabolismo energeacutetico nomeadamente transportadores de glucose (GLUT-
1) enzimas da via glicoliacutetica e de lactato desidrogenase (Figura 1) 2425
Para aleacutem do papel do HIF1 na promoccedilatildeo da glicoacutelise com formaccedilatildeo de lactato
estudos recentes revelaram uma significativa influecircncia na respiraccedilatildeo mitocondrial Nos
organismos aeroacutebios o TCA possui um papel fundamental no metabolismo central das
ceacutelulas constituindo a via de produccedilatildeo de energia mais eficiente e fornecendo o poder redutor
e intermediaacuterios necessaacuterios para as reacccedilotildees de biossiacutentese Normalmente a glucose eacute
convertida em piruvato no citoplasma o qual eacute posteriormente canalizado para o TCA ao
niacutevel mitocondrial onde eacute oxidado a dioacutexido de carbono e aacutegua A canalizaccedilatildeo do piruvato
3
para o TCA implica a sua conversatildeo em acetil-CoA numa reacccedilatildeo catalisada pelo complexo
multi-enzimaacutetico piruvato desidrogenase (PDH) Sendo esta uma das reacccedilotildees mais
importantes do metabolismo celular estaacute sujeita a um elevado niacutevel de regulaccedilatildeo Como tal o
complexo PDH eacute regulado por um ciclo reversiacutevel de fosforilaccedilatildeo caracterizado pela
fosforilaccedilatildeo (inactivaccedilatildeo) e desfosforilaccedilatildeo (activaccedilatildeo) da subunidade α do componente E1
(PDHE1α) promovida por cinases (PDK) e fosfatases (PDP) respectivamente Na PDHE1α
de mamiacuteferos existem trecircs locais de fosforilaccedilatildeo (Ser232
Ser293
e Ser300
) que podem ser
fosforilados por quatro isoenzimas (PDK1-4) sendo a PDK1 a uacutenica que consegue fosforilar
todos os trecircs locais 2627
Interessantemente a sobre-expressatildeo de PDK1 e PDK3 eacute promovida
por HIF1 levando assim agrave fosforilaccedilatildeo de PDHE1α e inibiccedilatildeo do complexo PDH (Figura 1)
Consequentemente a entrada do carbono derivado da via glicoliacutetica no TCA ocorre em menor
extensatildeo e aumenta a conversatildeo de piruvato a lactato resultando no fenoacutetipo lactogeacutenico
associado ao efeito de Warburg e na desregulaccedilatildeo do metabolismo energeacutetico 2428
Figura 1 Vias do metabolismo energeacutetico reguladas pelo factor de transcriccedilatildeo HIF1 A sobre-expressatildeo de
vaacuterias enzimas da via glicoliacutetica e da enzima lactato desidrogenase (LDH) eacute promovida pelo factor 1 indutiacutevel
por hipoxia (HIF1) levando ao aumento do consumo de glucose e agrave acumulaccedilatildeo de lactato A entrada do
carbono derivado da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) eacute realizada ao niacutevel do complexo
piruvato desidrogenase (PDH) sendo este regulado atraveacutes de um ciclo reversiacutevel de fosforilaccedilatildeo HIF1 promove
a expressatildeo das cinases da piruvato desidrogenase (PDK1-4) resultando na fosforilaccedilatildeo e consequente inibiccedilatildeo
do complexo PDH Esquema adaptado de Kondoh (2008) 20
Glucose
Glucose 6P
Fuctose 6P
Glyceraldehyde-3P
Fuctose 16 BP
Dihydroxyacetone-P
13 Biphosphoglycerate
3 - Phosphoglycerate
2 - Phosphoglycerate
Phosphoenopyruvate Pyruvate
Cytoplasm
Extracellular space
hexokinase
phosphoglucose isomerase
phosphofructokinase
GAPDH
phosphoglycerate
kinase
phosphoglucomutase
enolase
Pyruvate kinase
Pyruvate
Acetyl-CoATCA
PDH
PDH
P
LDH
Lactate
PDK
PDK2
PDK1 PDK3PDK4
aldolase
+
+
+
+
+
++
+
+
HIF1
4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

i
i AGRADECIMENTOS
Quero prestar os meus sinceros agradecimentos a todos aqueles que directa ou
indirectamente contribuiacuteram para o desenvolvimento deste trabalho Em particular
agrave Doutora Paula Alves pela oportunidade de realizar a minha dissertaccedilatildeo na Unidade
de Tecnologia de Ceacutelulas Animais (UTCA) e agrave Doutora Ana Sofia Coroadinha pela
oportunidade de integrar a equipa de Biotecnologia Molecular e Desenvolvimento de Linhas
Celulares e pela orientaccedilatildeo e disponibilidade
agrave Ana Filipa Rodrigues pela orientaccedilatildeo teacutecnica e cientiacutefica pela boa disposiccedilatildeo e
compreensatildeo e pelo incansaacutevel auxiacutelio no trabalho laboratorial e na elaboraccedilatildeo da presente
dissertaccedilatildeo
agrave Doutora Maria Filomena Caeiro pela orientaccedilatildeo e disposiccedilatildeo a ajudar prontamente
A todos os professores do meu percurso acadeacutemico pelas boas memoacuterias e ensinamentos que
me preparam da melhor forma possiacutevel para esta difiacutecil mas proveitosa jornada
ao Doutor Rui Gardner e agrave Telma Lopes do Laboratoacuterio de Citometria de Fluxo do
Instituto Gulbenkian de Ciecircncia pela ajuda teacutecnica necessaacuteria ao prosseguimento do
presente trabalho
a todos os colegas da UTCA por me terem recebido da melhor forma e por me terem
ajudado sempre que precisava Em particular agrave Vanessa Bandeira Heacutelio Tomaacutes Paulo
Fernandes e Doutora Rute Castro por todo o apoio e disponibilidade
a todos os meus familiares em particular aos meus pais e agrave minha irmatilde por sempre
terem acreditado em mim e pelo seu apoio nos momentos difiacuteceis
a todos os meus amigos por todo o apoio e momentos de diversatildeo e descontracccedilatildeo
A todos o meu muito obrigado
ii
ii RESUMO
A maioria das linhas celulares de mamiacutefero usadas na produccedilatildeo de biofaacutermacos
complexos apresenta um elevado consumo de glucose e acumulaccedilatildeo de quantidades
significativas de lactato ndash fenoacutetipo lactogeacutenico associado ao efeito de Warburg ndash reduzindo a
produtividade e qualidade do biofaacutermaco Este trabalho teve como objectivo a manipulaccedilatildeo
deste fenoacutetipo procurando reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do
metabolismo energeacutetico com consequente aumento da produtividade e qualidade de
partiacuteculas virais e anticorpos As linhas celulares 293 FLEX GFP (produtora de retroviacuterus)
MDCK E1 (produtora de adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram
silenciadas para o factor 1 indutiacutevel por hipoxia (HIF1) e cinases 1 e 3 da piruvato
desidrogenase (PDK13) os principais efectores deste fenoacutetipo
Nas ceacutelulas RPE A6 o silenciamento de HIF1 originou ceacutelulas com baixa capacidade
proliferativa sugerindo a reversatildeo da imortalizaccedilatildeo e entrada em senescecircncia Sem alteraccedilotildees
aparentes no crescimento celular nas ceacutelulas 293 FLEX GFP observou-se uma diminuiccedilatildeo
das taxas de consumo de glucose e de produccedilatildeo de lactato aliada a aumentos notaacuteveis na
produtividade celular (ateacute 182x em partiacuteculas totais e ateacute 216x em partiacuteculas infecciosas) e
sua qualidade o silenciamento adicional de PDK13 potenciou estes efeitos destacando-se o
aumento adicional (28x) da qualidade do produto final Nas ceacutelulas MDCK E1 natildeo se
obtiveram reduccedilotildees no consumo de glucose e produccedilatildeo de lactato e os aumentos na produccedilatildeo
viral e sua qualidade foram moderados Adicionalmente observou-se nas ceacutelulas silenciadas
aumento de poder redutor em 293 FLEX GFP e dos niacuteveis de ATP em MDCK E1 suportando
a reduccedilatildeo do fenoacutetipo lactogeacutenico e aumento de eficiecircncia do metabolismo energeacutetico
Estes resultados evidenciam a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao
efeito de Warburg como potencial ferramenta de engenharia metaboacutelica visando o aumento da
produtividade de biofaacutermacos complexos em cultura de ceacutelulas de mamiacutefero
Palavras-chave efeito de Warburg factor 1 indutiacutevel por hipoxia (HIF1) complexo
da piruvato desidrogenase (PDH) metabolismo energeacutetico produtividade celular
iii
iii ABSTRACT
The majority of mammalian cell lines used in the production of complex
biopharmaceuticals present high rates of glucose consumption accumulating significant
amounts of lactate ndash lactogenic phenotype associated to the Warburg effect ndash which results in
the reduction of cellular productivity and in final product quality The goal of this work was
the manipulation of this phenotype aiming at the reduction of lactate accumulation and the
improvement of energy metabolism efficiency to increase viral particles and antibodies
productivity and quality 293 FLEX GFP (retrovirus producer) MDCK E1 (adenovirus
producer) and RPE A6 (monoclonal antibodies producer) cell lines were silenced for hypoxia-
inducible factor 1 (HIF1) and pyruvate dehydrogenase 1 and 3 (PDK13) the major molecular
effectors of this phenotype
The silencing of HIF1 in RPE A6 cells resulted in cells with low proliferative capacity
suggesting the reversion of immortality and entry into senescence Without relevant impact in
cellular growth the 293 FLEX GFP cells showed a reduction in glucose uptake and lactate
production rates along with a significant increase of cellular productivities (up to 182x in
total and up to 216x in infectious particles) and its quality further silencing of PDK13
potentiated the effects of HIF1 silencing with special focus in the additional increase of final
product quality (28x) In MDCK E1 cells no reduction of glucose uptake and lactate
production rates was observed and viral productivity and its quality increased moderately
Additionally it was observed an increase in intracellular reducting power in 293 FLEX GFP
cells and in ATP levels in MDCK E1 cells further supporting the reduction of lactogenic
phenotype and the improvement of energy metabolism
These results highlight the manipulation of lactogenic phenotype associated to the
Warburg effect as a potential metabolic engineering tool for the improvement of complex
biopharmaceuticals production in cultured mammalian cell lines
Keywords Warburg effect hypoxia inducible factor 1 (HIF1) pyruvate
dehydrogenase complex (PDH) energetic metabolism cellular productivity
iv
iv LISTA DE ABREVIATURAS
Abreviatura Descriccedilatildeo
cDNA ADN complementar (do Inglecircs complementary DNA)
DNA Aacutecido desoxirribonucleico (do Inglecircs Deoxyribonucleic Acid)
ERO Espeacutecies reactivas de oxigeacutenio
GFP Proteiacutena fluorescente verde (do Inglecircs Green Fluorescent Protein)
HIF1 Factor 1 indutiacutevel por hipoxia (do Inglecircs Hypoxia Inducible Factor 1)
LDH Lactato desidrogenase (do Inglecircs Lactate Dehydrogenase)
mRNA ARN mensageiro (do Inglecircs messenger RNA)
ntshRNA non-target short hairpin RNA
PCR Reacccedilatildeo em cadeia da polimerase (do Inglecircs Polymerase chain reaction)
PDH Complexo da piruvato desidrogenase (do Inglecircs Pyruvate
Dehydrogenase Complex)
PDHE1α Subunidade α do componente E1 do complexo piruvato desidrogenase
(do Inglecircs E1 alpha subunit of pyruvate dehydrogenase complex)
PDK Cinase da piruvato desidrogenase (do Inglecircs Pyruvate Dehydrogenase
Kinase)
PDP Fosfatase da piruvato desidrogenase (do Inglecircs Pyruvate dehydrogenase
phosphatase)
PI Partiacuteculas infecciosas
PT Partiacuteculas totais
qRT-PCR PCR quantitativo por transcriccedilatildeo reversa (do Inglecircs Quantitative reverse
transcription polymerase chain reaction)
RFP Proteiacutena fluorescente vermelha (do Inglecircs Red Fluorescent Protein)
RNA Aacutecido ribonucleico (do Inglecircs Ribonucleic Acid)
shRNA short hairpin RNA
TCA Ciclo dos Aacutecidos Tricarboxiacutelicos (do Inglecircs Tricarboxylic Acid Cycle)
v
V IacuteNDICE
i AGRADECIMENTOS i
ii RESUMO ii
iii ABSTRACT iii
iv LISTA DE ABREVIATURAS iv
1 INTRODUCcedilAtildeO 1
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico 1
12 Imortalizaccedilatildeo celular e o efeito de Warburg 2
13 Impacto do efeito de Warburg em biotecnologia 4
14 Objectivos principais 5
2 MATERIAIS E MEacuteTODOS 6
21 Linhas celulares e meios de cultura 6
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais 6
23 Transduccedilatildeo celular selecccedilatildeo e clonagem 7
24 Estudos de crescimento e anaacutelise de metabolitos 7
25 Extracccedilatildeo de RNA total e qRT-PCR 8
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais8
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais 8
28 Determinaccedilatildeo da actividade da lactato desidrogenase9
29 Western Blot 9
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina 10
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio 10
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade 10
213 Caacutelculos 10
3 RESULTADOS 11
31 Silenciamento de HIF1 11
vi
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3 15
4 DISCUSSAtildeO E CONCLUSAtildeO 24
5 PERSPECTIVAS FUTURAS 29
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS 30
7 ANEXOS 34
1
1 INTRODUCcedilAtildeO
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico
Nos uacuteltimos anos tem-se verificado uma crescente procura de biofaacutermacos complexos
como proteiacutenas recombinantes e partiacuteculas derivadas de viacuterus quer para fins terapecircuticos quer
para fins de diagnoacutesticos in vivo O potencial dos biofaacutermacos derivados de viacuterus em
particular tem vindo a aumentar devido agraves suas diversas aplicaccedilotildees profilaacuteticas terapecircuticas e
cliacutenicas como a vacinaccedilatildeo o tratamento do cancro e a terapia geacutenica 1 Actualmente o
sistema preferencial para a manufacturaccedilatildeo destes produtos biofarmacecircuticos complexos eacute as
ceacutelulas de mamiacutefero usadas para a produccedilatildeo de mais de metade dos biofaacutermacos existentes
no mercado Tal predomiacutenio deve-se agrave sua semelhanccedila com as ceacutelulas humanas no folding de
proteiacutenas e nas modificaccedilotildees poacutes-traducionais como a glicosilaccedilatildeo aumentando a eficiecircncia
terapecircutica e diminuindo o potencial imunogeacutenico do produto final 23
As ceacutelulas de mamiacutefero em cultura agrave semelhanccedila de ceacutelulas transformadas possuem
um metabolismo energeacutetico desregulado consumindo elevadas quantidades de glucose e
acumulando quantidades significativas de lactato com reduzida entrada de carbono derivado
da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) 4 Do ponto de vista da cultura de
ceacutelulas para produccedilatildeo de biofaacutermacos este fenoacutetipo lactogeacutenico tem simultaneamente um
efeito negativo no crescimento e viabilidade celular e na produtividade e qualidade do
biofaacutermaco Este fenoacutemeno eacute ainda mais importante no caso das partiacuteculas derivadas de viacuterus
para terapia geacutenica jaacute que a sua infecciosidade eacute fortemente influenciada por variaccedilotildees no pH
do meio de cultura 5 Diversas estrateacutegias tecircm sido testadas de forma a reduzir os efeitos
nocivos da acumulaccedilatildeo de lactato no meio de cultura fornecimento de glucose de forma
controlada 67
suplementaccedilatildeo do meio de cultura com substratos alternativos agrave glucose 8
remoccedilatildeo dos metabolitos toacutexicos 9 desenvolvimento de meios de cultura com base na anaacutelise
dos fluxos metaboacutelicos permitindo adaptar os seus constituintes agraves necessidades metaboacutelicas
das ceacutelulas 1011
introduccedilatildeo da enzima piruvato carboxilase nas ceacutelulas 12-14
diminuiccedilatildeo da
expressatildeo da lactato desidrogenase atraveacutes da disrupccedilatildeo do gene Ldha por recombinaccedilatildeo
homoacuteloga 15
ou atraveacutes de RNA de interferecircncia 16
ou introduccedilatildeo de um transportador para a
frutose constituindo uma alternativa ao consumo de glucose 17
No entanto estas estrateacutegias
tecircm um sucesso limitado devido aos efeitos compensatoacuterios causados pela apertada regulaccedilatildeo
homeostaacutetica intracelular que natildeo satildeo facilmente alteraacuteveis
2
12 Imortalizaccedilatildeo celular e o efeito de Warburg
Nos anos 20 Otto Warburg observou que as ceacutelulas canceriacutegenas comparativamente
agraves ceacutelulas normais apresentavam elevadas taxas de consumo de glucose secretando a maior
parte do carbono derivado da glucose sob a forma de lactato em vez de o oxidar
completamente 1819
ndash um cenaacuterio que ficou conhecido como efeito de Warburg Estudos
recentes sobre o metabolismo energeacutetico de ceacutelulas canceriacutegenas sugerem que este resulta de
alteraccedilotildees geneacuteticas que ocorrem no processo de tumorigeacutenese principalmente durante a
imortalizaccedilatildeo e transformaccedilatildeo celulares e que levam agrave desregulaccedilatildeo do metabolismo
energeacutetico central e ao aumento da actividade glicoliacutetica 20
De facto existem diversas vias de
sinalizaccedilatildeo comuns ao controlo da proliferaccedilatildeo e do metabolismo energeacutetico que levam agrave
induccedilatildeo deste cenaacuterio como a activaccedilatildeo de oncogenes (MYC) e enzimas glicoliacuteticas ou a
supressatildeo do gene supressor de tumores P53 2021
Interessantemente muitas das vias de
imortalizaccedilatildeo activadas no processo de tumorigeacutenese satildeo tambeacutem recrutadas durante o
estabelecimento de uma linha celular e por isso tendem a exibir caracteriacutesticas metaboacutelicas
associadas ao efeito de Warburg 20
O principal efector molecular do efeito de Warburg eacute o factor 1 indutiacutevel por hipoxia
(HIF1) um factor de transcriccedilatildeo heterodimeacuterico constituiacutedo por uma subunidade β expressa
constitutivamente e uma subunidade α cuja expressatildeo estaacute dependente do oxigeacutenio Apesar
de ser expressa em condiccedilotildees de baixa disponibilidade de oxigeacutenio (hipoxia) a expressatildeo
constitutiva da subunidade α pode ocorrer em normoxia como resultado de alteraccedilotildees nos
sistemas de sinalizaccedilatildeo para degradaccedilatildeo proteica provocadas durante o processo de
imortalizaccedilatildeo celular De facto a sua sobre-expressatildeo eacute caracteriacutestica comum a muitas ceacutelulas
canceriacutegenas 2223
O factor de transcriccedilatildeo HIF1 eacute responsaacutevel pela coordenaccedilatildeo da alteraccedilatildeo
do metabolismo para glicoacutelise com formaccedilatildeo de lactato com consequente aumento do
consumo de glucose e acumulaccedilatildeo de lactato jaacute que promove a expressatildeo de vaacuterios
componentes do metabolismo energeacutetico nomeadamente transportadores de glucose (GLUT-
1) enzimas da via glicoliacutetica e de lactato desidrogenase (Figura 1) 2425
Para aleacutem do papel do HIF1 na promoccedilatildeo da glicoacutelise com formaccedilatildeo de lactato
estudos recentes revelaram uma significativa influecircncia na respiraccedilatildeo mitocondrial Nos
organismos aeroacutebios o TCA possui um papel fundamental no metabolismo central das
ceacutelulas constituindo a via de produccedilatildeo de energia mais eficiente e fornecendo o poder redutor
e intermediaacuterios necessaacuterios para as reacccedilotildees de biossiacutentese Normalmente a glucose eacute
convertida em piruvato no citoplasma o qual eacute posteriormente canalizado para o TCA ao
niacutevel mitocondrial onde eacute oxidado a dioacutexido de carbono e aacutegua A canalizaccedilatildeo do piruvato
3
para o TCA implica a sua conversatildeo em acetil-CoA numa reacccedilatildeo catalisada pelo complexo
multi-enzimaacutetico piruvato desidrogenase (PDH) Sendo esta uma das reacccedilotildees mais
importantes do metabolismo celular estaacute sujeita a um elevado niacutevel de regulaccedilatildeo Como tal o
complexo PDH eacute regulado por um ciclo reversiacutevel de fosforilaccedilatildeo caracterizado pela
fosforilaccedilatildeo (inactivaccedilatildeo) e desfosforilaccedilatildeo (activaccedilatildeo) da subunidade α do componente E1
(PDHE1α) promovida por cinases (PDK) e fosfatases (PDP) respectivamente Na PDHE1α
de mamiacuteferos existem trecircs locais de fosforilaccedilatildeo (Ser232
Ser293
e Ser300
) que podem ser
fosforilados por quatro isoenzimas (PDK1-4) sendo a PDK1 a uacutenica que consegue fosforilar
todos os trecircs locais 2627
Interessantemente a sobre-expressatildeo de PDK1 e PDK3 eacute promovida
por HIF1 levando assim agrave fosforilaccedilatildeo de PDHE1α e inibiccedilatildeo do complexo PDH (Figura 1)
Consequentemente a entrada do carbono derivado da via glicoliacutetica no TCA ocorre em menor
extensatildeo e aumenta a conversatildeo de piruvato a lactato resultando no fenoacutetipo lactogeacutenico
associado ao efeito de Warburg e na desregulaccedilatildeo do metabolismo energeacutetico 2428
Figura 1 Vias do metabolismo energeacutetico reguladas pelo factor de transcriccedilatildeo HIF1 A sobre-expressatildeo de
vaacuterias enzimas da via glicoliacutetica e da enzima lactato desidrogenase (LDH) eacute promovida pelo factor 1 indutiacutevel
por hipoxia (HIF1) levando ao aumento do consumo de glucose e agrave acumulaccedilatildeo de lactato A entrada do
carbono derivado da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) eacute realizada ao niacutevel do complexo
piruvato desidrogenase (PDH) sendo este regulado atraveacutes de um ciclo reversiacutevel de fosforilaccedilatildeo HIF1 promove
a expressatildeo das cinases da piruvato desidrogenase (PDK1-4) resultando na fosforilaccedilatildeo e consequente inibiccedilatildeo
do complexo PDH Esquema adaptado de Kondoh (2008) 20
Glucose
Glucose 6P
Fuctose 6P
Glyceraldehyde-3P
Fuctose 16 BP
Dihydroxyacetone-P
13 Biphosphoglycerate
3 - Phosphoglycerate
2 - Phosphoglycerate
Phosphoenopyruvate Pyruvate
Cytoplasm
Extracellular space
hexokinase
phosphoglucose isomerase
phosphofructokinase
GAPDH
phosphoglycerate
kinase
phosphoglucomutase
enolase
Pyruvate kinase
Pyruvate
Acetyl-CoATCA
PDH
PDH
P
LDH
Lactate
PDK
PDK2
PDK1 PDK3PDK4
aldolase
+
+
+
+
+
++
+
+
HIF1
4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

ii
ii RESUMO
A maioria das linhas celulares de mamiacutefero usadas na produccedilatildeo de biofaacutermacos
complexos apresenta um elevado consumo de glucose e acumulaccedilatildeo de quantidades
significativas de lactato ndash fenoacutetipo lactogeacutenico associado ao efeito de Warburg ndash reduzindo a
produtividade e qualidade do biofaacutermaco Este trabalho teve como objectivo a manipulaccedilatildeo
deste fenoacutetipo procurando reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do
metabolismo energeacutetico com consequente aumento da produtividade e qualidade de
partiacuteculas virais e anticorpos As linhas celulares 293 FLEX GFP (produtora de retroviacuterus)
MDCK E1 (produtora de adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram
silenciadas para o factor 1 indutiacutevel por hipoxia (HIF1) e cinases 1 e 3 da piruvato
desidrogenase (PDK13) os principais efectores deste fenoacutetipo
Nas ceacutelulas RPE A6 o silenciamento de HIF1 originou ceacutelulas com baixa capacidade
proliferativa sugerindo a reversatildeo da imortalizaccedilatildeo e entrada em senescecircncia Sem alteraccedilotildees
aparentes no crescimento celular nas ceacutelulas 293 FLEX GFP observou-se uma diminuiccedilatildeo
das taxas de consumo de glucose e de produccedilatildeo de lactato aliada a aumentos notaacuteveis na
produtividade celular (ateacute 182x em partiacuteculas totais e ateacute 216x em partiacuteculas infecciosas) e
sua qualidade o silenciamento adicional de PDK13 potenciou estes efeitos destacando-se o
aumento adicional (28x) da qualidade do produto final Nas ceacutelulas MDCK E1 natildeo se
obtiveram reduccedilotildees no consumo de glucose e produccedilatildeo de lactato e os aumentos na produccedilatildeo
viral e sua qualidade foram moderados Adicionalmente observou-se nas ceacutelulas silenciadas
aumento de poder redutor em 293 FLEX GFP e dos niacuteveis de ATP em MDCK E1 suportando
a reduccedilatildeo do fenoacutetipo lactogeacutenico e aumento de eficiecircncia do metabolismo energeacutetico
Estes resultados evidenciam a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao
efeito de Warburg como potencial ferramenta de engenharia metaboacutelica visando o aumento da
produtividade de biofaacutermacos complexos em cultura de ceacutelulas de mamiacutefero
Palavras-chave efeito de Warburg factor 1 indutiacutevel por hipoxia (HIF1) complexo
da piruvato desidrogenase (PDH) metabolismo energeacutetico produtividade celular
iii
iii ABSTRACT
The majority of mammalian cell lines used in the production of complex
biopharmaceuticals present high rates of glucose consumption accumulating significant
amounts of lactate ndash lactogenic phenotype associated to the Warburg effect ndash which results in
the reduction of cellular productivity and in final product quality The goal of this work was
the manipulation of this phenotype aiming at the reduction of lactate accumulation and the
improvement of energy metabolism efficiency to increase viral particles and antibodies
productivity and quality 293 FLEX GFP (retrovirus producer) MDCK E1 (adenovirus
producer) and RPE A6 (monoclonal antibodies producer) cell lines were silenced for hypoxia-
inducible factor 1 (HIF1) and pyruvate dehydrogenase 1 and 3 (PDK13) the major molecular
effectors of this phenotype
The silencing of HIF1 in RPE A6 cells resulted in cells with low proliferative capacity
suggesting the reversion of immortality and entry into senescence Without relevant impact in
cellular growth the 293 FLEX GFP cells showed a reduction in glucose uptake and lactate
production rates along with a significant increase of cellular productivities (up to 182x in
total and up to 216x in infectious particles) and its quality further silencing of PDK13
potentiated the effects of HIF1 silencing with special focus in the additional increase of final
product quality (28x) In MDCK E1 cells no reduction of glucose uptake and lactate
production rates was observed and viral productivity and its quality increased moderately
Additionally it was observed an increase in intracellular reducting power in 293 FLEX GFP
cells and in ATP levels in MDCK E1 cells further supporting the reduction of lactogenic
phenotype and the improvement of energy metabolism
These results highlight the manipulation of lactogenic phenotype associated to the
Warburg effect as a potential metabolic engineering tool for the improvement of complex
biopharmaceuticals production in cultured mammalian cell lines
Keywords Warburg effect hypoxia inducible factor 1 (HIF1) pyruvate
dehydrogenase complex (PDH) energetic metabolism cellular productivity
iv
iv LISTA DE ABREVIATURAS
Abreviatura Descriccedilatildeo
cDNA ADN complementar (do Inglecircs complementary DNA)
DNA Aacutecido desoxirribonucleico (do Inglecircs Deoxyribonucleic Acid)
ERO Espeacutecies reactivas de oxigeacutenio
GFP Proteiacutena fluorescente verde (do Inglecircs Green Fluorescent Protein)
HIF1 Factor 1 indutiacutevel por hipoxia (do Inglecircs Hypoxia Inducible Factor 1)
LDH Lactato desidrogenase (do Inglecircs Lactate Dehydrogenase)
mRNA ARN mensageiro (do Inglecircs messenger RNA)
ntshRNA non-target short hairpin RNA
PCR Reacccedilatildeo em cadeia da polimerase (do Inglecircs Polymerase chain reaction)
PDH Complexo da piruvato desidrogenase (do Inglecircs Pyruvate
Dehydrogenase Complex)
PDHE1α Subunidade α do componente E1 do complexo piruvato desidrogenase
(do Inglecircs E1 alpha subunit of pyruvate dehydrogenase complex)
PDK Cinase da piruvato desidrogenase (do Inglecircs Pyruvate Dehydrogenase
Kinase)
PDP Fosfatase da piruvato desidrogenase (do Inglecircs Pyruvate dehydrogenase
phosphatase)
PI Partiacuteculas infecciosas
PT Partiacuteculas totais
qRT-PCR PCR quantitativo por transcriccedilatildeo reversa (do Inglecircs Quantitative reverse
transcription polymerase chain reaction)
RFP Proteiacutena fluorescente vermelha (do Inglecircs Red Fluorescent Protein)
RNA Aacutecido ribonucleico (do Inglecircs Ribonucleic Acid)
shRNA short hairpin RNA
TCA Ciclo dos Aacutecidos Tricarboxiacutelicos (do Inglecircs Tricarboxylic Acid Cycle)
v
V IacuteNDICE
i AGRADECIMENTOS i
ii RESUMO ii
iii ABSTRACT iii
iv LISTA DE ABREVIATURAS iv
1 INTRODUCcedilAtildeO 1
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico 1
12 Imortalizaccedilatildeo celular e o efeito de Warburg 2
13 Impacto do efeito de Warburg em biotecnologia 4
14 Objectivos principais 5
2 MATERIAIS E MEacuteTODOS 6
21 Linhas celulares e meios de cultura 6
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais 6
23 Transduccedilatildeo celular selecccedilatildeo e clonagem 7
24 Estudos de crescimento e anaacutelise de metabolitos 7
25 Extracccedilatildeo de RNA total e qRT-PCR 8
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais8
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais 8
28 Determinaccedilatildeo da actividade da lactato desidrogenase9
29 Western Blot 9
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina 10
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio 10
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade 10
213 Caacutelculos 10
3 RESULTADOS 11
31 Silenciamento de HIF1 11
vi
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3 15
4 DISCUSSAtildeO E CONCLUSAtildeO 24
5 PERSPECTIVAS FUTURAS 29
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS 30
7 ANEXOS 34
1
1 INTRODUCcedilAtildeO
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico
Nos uacuteltimos anos tem-se verificado uma crescente procura de biofaacutermacos complexos
como proteiacutenas recombinantes e partiacuteculas derivadas de viacuterus quer para fins terapecircuticos quer
para fins de diagnoacutesticos in vivo O potencial dos biofaacutermacos derivados de viacuterus em
particular tem vindo a aumentar devido agraves suas diversas aplicaccedilotildees profilaacuteticas terapecircuticas e
cliacutenicas como a vacinaccedilatildeo o tratamento do cancro e a terapia geacutenica 1 Actualmente o
sistema preferencial para a manufacturaccedilatildeo destes produtos biofarmacecircuticos complexos eacute as
ceacutelulas de mamiacutefero usadas para a produccedilatildeo de mais de metade dos biofaacutermacos existentes
no mercado Tal predomiacutenio deve-se agrave sua semelhanccedila com as ceacutelulas humanas no folding de
proteiacutenas e nas modificaccedilotildees poacutes-traducionais como a glicosilaccedilatildeo aumentando a eficiecircncia
terapecircutica e diminuindo o potencial imunogeacutenico do produto final 23
As ceacutelulas de mamiacutefero em cultura agrave semelhanccedila de ceacutelulas transformadas possuem
um metabolismo energeacutetico desregulado consumindo elevadas quantidades de glucose e
acumulando quantidades significativas de lactato com reduzida entrada de carbono derivado
da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) 4 Do ponto de vista da cultura de
ceacutelulas para produccedilatildeo de biofaacutermacos este fenoacutetipo lactogeacutenico tem simultaneamente um
efeito negativo no crescimento e viabilidade celular e na produtividade e qualidade do
biofaacutermaco Este fenoacutemeno eacute ainda mais importante no caso das partiacuteculas derivadas de viacuterus
para terapia geacutenica jaacute que a sua infecciosidade eacute fortemente influenciada por variaccedilotildees no pH
do meio de cultura 5 Diversas estrateacutegias tecircm sido testadas de forma a reduzir os efeitos
nocivos da acumulaccedilatildeo de lactato no meio de cultura fornecimento de glucose de forma
controlada 67
suplementaccedilatildeo do meio de cultura com substratos alternativos agrave glucose 8
remoccedilatildeo dos metabolitos toacutexicos 9 desenvolvimento de meios de cultura com base na anaacutelise
dos fluxos metaboacutelicos permitindo adaptar os seus constituintes agraves necessidades metaboacutelicas
das ceacutelulas 1011
introduccedilatildeo da enzima piruvato carboxilase nas ceacutelulas 12-14
diminuiccedilatildeo da
expressatildeo da lactato desidrogenase atraveacutes da disrupccedilatildeo do gene Ldha por recombinaccedilatildeo
homoacuteloga 15
ou atraveacutes de RNA de interferecircncia 16
ou introduccedilatildeo de um transportador para a
frutose constituindo uma alternativa ao consumo de glucose 17
No entanto estas estrateacutegias
tecircm um sucesso limitado devido aos efeitos compensatoacuterios causados pela apertada regulaccedilatildeo
homeostaacutetica intracelular que natildeo satildeo facilmente alteraacuteveis
2
12 Imortalizaccedilatildeo celular e o efeito de Warburg
Nos anos 20 Otto Warburg observou que as ceacutelulas canceriacutegenas comparativamente
agraves ceacutelulas normais apresentavam elevadas taxas de consumo de glucose secretando a maior
parte do carbono derivado da glucose sob a forma de lactato em vez de o oxidar
completamente 1819
ndash um cenaacuterio que ficou conhecido como efeito de Warburg Estudos
recentes sobre o metabolismo energeacutetico de ceacutelulas canceriacutegenas sugerem que este resulta de
alteraccedilotildees geneacuteticas que ocorrem no processo de tumorigeacutenese principalmente durante a
imortalizaccedilatildeo e transformaccedilatildeo celulares e que levam agrave desregulaccedilatildeo do metabolismo
energeacutetico central e ao aumento da actividade glicoliacutetica 20
De facto existem diversas vias de
sinalizaccedilatildeo comuns ao controlo da proliferaccedilatildeo e do metabolismo energeacutetico que levam agrave
induccedilatildeo deste cenaacuterio como a activaccedilatildeo de oncogenes (MYC) e enzimas glicoliacuteticas ou a
supressatildeo do gene supressor de tumores P53 2021
Interessantemente muitas das vias de
imortalizaccedilatildeo activadas no processo de tumorigeacutenese satildeo tambeacutem recrutadas durante o
estabelecimento de uma linha celular e por isso tendem a exibir caracteriacutesticas metaboacutelicas
associadas ao efeito de Warburg 20
O principal efector molecular do efeito de Warburg eacute o factor 1 indutiacutevel por hipoxia
(HIF1) um factor de transcriccedilatildeo heterodimeacuterico constituiacutedo por uma subunidade β expressa
constitutivamente e uma subunidade α cuja expressatildeo estaacute dependente do oxigeacutenio Apesar
de ser expressa em condiccedilotildees de baixa disponibilidade de oxigeacutenio (hipoxia) a expressatildeo
constitutiva da subunidade α pode ocorrer em normoxia como resultado de alteraccedilotildees nos
sistemas de sinalizaccedilatildeo para degradaccedilatildeo proteica provocadas durante o processo de
imortalizaccedilatildeo celular De facto a sua sobre-expressatildeo eacute caracteriacutestica comum a muitas ceacutelulas
canceriacutegenas 2223
O factor de transcriccedilatildeo HIF1 eacute responsaacutevel pela coordenaccedilatildeo da alteraccedilatildeo
do metabolismo para glicoacutelise com formaccedilatildeo de lactato com consequente aumento do
consumo de glucose e acumulaccedilatildeo de lactato jaacute que promove a expressatildeo de vaacuterios
componentes do metabolismo energeacutetico nomeadamente transportadores de glucose (GLUT-
1) enzimas da via glicoliacutetica e de lactato desidrogenase (Figura 1) 2425
Para aleacutem do papel do HIF1 na promoccedilatildeo da glicoacutelise com formaccedilatildeo de lactato
estudos recentes revelaram uma significativa influecircncia na respiraccedilatildeo mitocondrial Nos
organismos aeroacutebios o TCA possui um papel fundamental no metabolismo central das
ceacutelulas constituindo a via de produccedilatildeo de energia mais eficiente e fornecendo o poder redutor
e intermediaacuterios necessaacuterios para as reacccedilotildees de biossiacutentese Normalmente a glucose eacute
convertida em piruvato no citoplasma o qual eacute posteriormente canalizado para o TCA ao
niacutevel mitocondrial onde eacute oxidado a dioacutexido de carbono e aacutegua A canalizaccedilatildeo do piruvato
3
para o TCA implica a sua conversatildeo em acetil-CoA numa reacccedilatildeo catalisada pelo complexo
multi-enzimaacutetico piruvato desidrogenase (PDH) Sendo esta uma das reacccedilotildees mais
importantes do metabolismo celular estaacute sujeita a um elevado niacutevel de regulaccedilatildeo Como tal o
complexo PDH eacute regulado por um ciclo reversiacutevel de fosforilaccedilatildeo caracterizado pela
fosforilaccedilatildeo (inactivaccedilatildeo) e desfosforilaccedilatildeo (activaccedilatildeo) da subunidade α do componente E1
(PDHE1α) promovida por cinases (PDK) e fosfatases (PDP) respectivamente Na PDHE1α
de mamiacuteferos existem trecircs locais de fosforilaccedilatildeo (Ser232
Ser293
e Ser300
) que podem ser
fosforilados por quatro isoenzimas (PDK1-4) sendo a PDK1 a uacutenica que consegue fosforilar
todos os trecircs locais 2627
Interessantemente a sobre-expressatildeo de PDK1 e PDK3 eacute promovida
por HIF1 levando assim agrave fosforilaccedilatildeo de PDHE1α e inibiccedilatildeo do complexo PDH (Figura 1)
Consequentemente a entrada do carbono derivado da via glicoliacutetica no TCA ocorre em menor
extensatildeo e aumenta a conversatildeo de piruvato a lactato resultando no fenoacutetipo lactogeacutenico
associado ao efeito de Warburg e na desregulaccedilatildeo do metabolismo energeacutetico 2428
Figura 1 Vias do metabolismo energeacutetico reguladas pelo factor de transcriccedilatildeo HIF1 A sobre-expressatildeo de
vaacuterias enzimas da via glicoliacutetica e da enzima lactato desidrogenase (LDH) eacute promovida pelo factor 1 indutiacutevel
por hipoxia (HIF1) levando ao aumento do consumo de glucose e agrave acumulaccedilatildeo de lactato A entrada do
carbono derivado da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) eacute realizada ao niacutevel do complexo
piruvato desidrogenase (PDH) sendo este regulado atraveacutes de um ciclo reversiacutevel de fosforilaccedilatildeo HIF1 promove
a expressatildeo das cinases da piruvato desidrogenase (PDK1-4) resultando na fosforilaccedilatildeo e consequente inibiccedilatildeo
do complexo PDH Esquema adaptado de Kondoh (2008) 20
Glucose
Glucose 6P
Fuctose 6P
Glyceraldehyde-3P
Fuctose 16 BP
Dihydroxyacetone-P
13 Biphosphoglycerate
3 - Phosphoglycerate
2 - Phosphoglycerate
Phosphoenopyruvate Pyruvate
Cytoplasm
Extracellular space
hexokinase
phosphoglucose isomerase
phosphofructokinase
GAPDH
phosphoglycerate
kinase
phosphoglucomutase
enolase
Pyruvate kinase
Pyruvate
Acetyl-CoATCA
PDH
PDH
P
LDH
Lactate
PDK
PDK2
PDK1 PDK3PDK4
aldolase
+
+
+
+
+
++
+
+
HIF1
4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

iii
iii ABSTRACT
The majority of mammalian cell lines used in the production of complex
biopharmaceuticals present high rates of glucose consumption accumulating significant
amounts of lactate ndash lactogenic phenotype associated to the Warburg effect ndash which results in
the reduction of cellular productivity and in final product quality The goal of this work was
the manipulation of this phenotype aiming at the reduction of lactate accumulation and the
improvement of energy metabolism efficiency to increase viral particles and antibodies
productivity and quality 293 FLEX GFP (retrovirus producer) MDCK E1 (adenovirus
producer) and RPE A6 (monoclonal antibodies producer) cell lines were silenced for hypoxia-
inducible factor 1 (HIF1) and pyruvate dehydrogenase 1 and 3 (PDK13) the major molecular
effectors of this phenotype
The silencing of HIF1 in RPE A6 cells resulted in cells with low proliferative capacity
suggesting the reversion of immortality and entry into senescence Without relevant impact in
cellular growth the 293 FLEX GFP cells showed a reduction in glucose uptake and lactate
production rates along with a significant increase of cellular productivities (up to 182x in
total and up to 216x in infectious particles) and its quality further silencing of PDK13
potentiated the effects of HIF1 silencing with special focus in the additional increase of final
product quality (28x) In MDCK E1 cells no reduction of glucose uptake and lactate
production rates was observed and viral productivity and its quality increased moderately
Additionally it was observed an increase in intracellular reducting power in 293 FLEX GFP
cells and in ATP levels in MDCK E1 cells further supporting the reduction of lactogenic
phenotype and the improvement of energy metabolism
These results highlight the manipulation of lactogenic phenotype associated to the
Warburg effect as a potential metabolic engineering tool for the improvement of complex
biopharmaceuticals production in cultured mammalian cell lines
Keywords Warburg effect hypoxia inducible factor 1 (HIF1) pyruvate
dehydrogenase complex (PDH) energetic metabolism cellular productivity
iv
iv LISTA DE ABREVIATURAS
Abreviatura Descriccedilatildeo
cDNA ADN complementar (do Inglecircs complementary DNA)
DNA Aacutecido desoxirribonucleico (do Inglecircs Deoxyribonucleic Acid)
ERO Espeacutecies reactivas de oxigeacutenio
GFP Proteiacutena fluorescente verde (do Inglecircs Green Fluorescent Protein)
HIF1 Factor 1 indutiacutevel por hipoxia (do Inglecircs Hypoxia Inducible Factor 1)
LDH Lactato desidrogenase (do Inglecircs Lactate Dehydrogenase)
mRNA ARN mensageiro (do Inglecircs messenger RNA)
ntshRNA non-target short hairpin RNA
PCR Reacccedilatildeo em cadeia da polimerase (do Inglecircs Polymerase chain reaction)
PDH Complexo da piruvato desidrogenase (do Inglecircs Pyruvate
Dehydrogenase Complex)
PDHE1α Subunidade α do componente E1 do complexo piruvato desidrogenase
(do Inglecircs E1 alpha subunit of pyruvate dehydrogenase complex)
PDK Cinase da piruvato desidrogenase (do Inglecircs Pyruvate Dehydrogenase
Kinase)
PDP Fosfatase da piruvato desidrogenase (do Inglecircs Pyruvate dehydrogenase
phosphatase)
PI Partiacuteculas infecciosas
PT Partiacuteculas totais
qRT-PCR PCR quantitativo por transcriccedilatildeo reversa (do Inglecircs Quantitative reverse
transcription polymerase chain reaction)
RFP Proteiacutena fluorescente vermelha (do Inglecircs Red Fluorescent Protein)
RNA Aacutecido ribonucleico (do Inglecircs Ribonucleic Acid)
shRNA short hairpin RNA
TCA Ciclo dos Aacutecidos Tricarboxiacutelicos (do Inglecircs Tricarboxylic Acid Cycle)
v
V IacuteNDICE
i AGRADECIMENTOS i
ii RESUMO ii
iii ABSTRACT iii
iv LISTA DE ABREVIATURAS iv
1 INTRODUCcedilAtildeO 1
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico 1
12 Imortalizaccedilatildeo celular e o efeito de Warburg 2
13 Impacto do efeito de Warburg em biotecnologia 4
14 Objectivos principais 5
2 MATERIAIS E MEacuteTODOS 6
21 Linhas celulares e meios de cultura 6
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais 6
23 Transduccedilatildeo celular selecccedilatildeo e clonagem 7
24 Estudos de crescimento e anaacutelise de metabolitos 7
25 Extracccedilatildeo de RNA total e qRT-PCR 8
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais8
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais 8
28 Determinaccedilatildeo da actividade da lactato desidrogenase9
29 Western Blot 9
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina 10
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio 10
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade 10
213 Caacutelculos 10
3 RESULTADOS 11
31 Silenciamento de HIF1 11
vi
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3 15
4 DISCUSSAtildeO E CONCLUSAtildeO 24
5 PERSPECTIVAS FUTURAS 29
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS 30
7 ANEXOS 34
1
1 INTRODUCcedilAtildeO
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico
Nos uacuteltimos anos tem-se verificado uma crescente procura de biofaacutermacos complexos
como proteiacutenas recombinantes e partiacuteculas derivadas de viacuterus quer para fins terapecircuticos quer
para fins de diagnoacutesticos in vivo O potencial dos biofaacutermacos derivados de viacuterus em
particular tem vindo a aumentar devido agraves suas diversas aplicaccedilotildees profilaacuteticas terapecircuticas e
cliacutenicas como a vacinaccedilatildeo o tratamento do cancro e a terapia geacutenica 1 Actualmente o
sistema preferencial para a manufacturaccedilatildeo destes produtos biofarmacecircuticos complexos eacute as
ceacutelulas de mamiacutefero usadas para a produccedilatildeo de mais de metade dos biofaacutermacos existentes
no mercado Tal predomiacutenio deve-se agrave sua semelhanccedila com as ceacutelulas humanas no folding de
proteiacutenas e nas modificaccedilotildees poacutes-traducionais como a glicosilaccedilatildeo aumentando a eficiecircncia
terapecircutica e diminuindo o potencial imunogeacutenico do produto final 23
As ceacutelulas de mamiacutefero em cultura agrave semelhanccedila de ceacutelulas transformadas possuem
um metabolismo energeacutetico desregulado consumindo elevadas quantidades de glucose e
acumulando quantidades significativas de lactato com reduzida entrada de carbono derivado
da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) 4 Do ponto de vista da cultura de
ceacutelulas para produccedilatildeo de biofaacutermacos este fenoacutetipo lactogeacutenico tem simultaneamente um
efeito negativo no crescimento e viabilidade celular e na produtividade e qualidade do
biofaacutermaco Este fenoacutemeno eacute ainda mais importante no caso das partiacuteculas derivadas de viacuterus
para terapia geacutenica jaacute que a sua infecciosidade eacute fortemente influenciada por variaccedilotildees no pH
do meio de cultura 5 Diversas estrateacutegias tecircm sido testadas de forma a reduzir os efeitos
nocivos da acumulaccedilatildeo de lactato no meio de cultura fornecimento de glucose de forma
controlada 67
suplementaccedilatildeo do meio de cultura com substratos alternativos agrave glucose 8
remoccedilatildeo dos metabolitos toacutexicos 9 desenvolvimento de meios de cultura com base na anaacutelise
dos fluxos metaboacutelicos permitindo adaptar os seus constituintes agraves necessidades metaboacutelicas
das ceacutelulas 1011
introduccedilatildeo da enzima piruvato carboxilase nas ceacutelulas 12-14
diminuiccedilatildeo da
expressatildeo da lactato desidrogenase atraveacutes da disrupccedilatildeo do gene Ldha por recombinaccedilatildeo
homoacuteloga 15
ou atraveacutes de RNA de interferecircncia 16
ou introduccedilatildeo de um transportador para a
frutose constituindo uma alternativa ao consumo de glucose 17
No entanto estas estrateacutegias
tecircm um sucesso limitado devido aos efeitos compensatoacuterios causados pela apertada regulaccedilatildeo
homeostaacutetica intracelular que natildeo satildeo facilmente alteraacuteveis
2
12 Imortalizaccedilatildeo celular e o efeito de Warburg
Nos anos 20 Otto Warburg observou que as ceacutelulas canceriacutegenas comparativamente
agraves ceacutelulas normais apresentavam elevadas taxas de consumo de glucose secretando a maior
parte do carbono derivado da glucose sob a forma de lactato em vez de o oxidar
completamente 1819
ndash um cenaacuterio que ficou conhecido como efeito de Warburg Estudos
recentes sobre o metabolismo energeacutetico de ceacutelulas canceriacutegenas sugerem que este resulta de
alteraccedilotildees geneacuteticas que ocorrem no processo de tumorigeacutenese principalmente durante a
imortalizaccedilatildeo e transformaccedilatildeo celulares e que levam agrave desregulaccedilatildeo do metabolismo
energeacutetico central e ao aumento da actividade glicoliacutetica 20
De facto existem diversas vias de
sinalizaccedilatildeo comuns ao controlo da proliferaccedilatildeo e do metabolismo energeacutetico que levam agrave
induccedilatildeo deste cenaacuterio como a activaccedilatildeo de oncogenes (MYC) e enzimas glicoliacuteticas ou a
supressatildeo do gene supressor de tumores P53 2021
Interessantemente muitas das vias de
imortalizaccedilatildeo activadas no processo de tumorigeacutenese satildeo tambeacutem recrutadas durante o
estabelecimento de uma linha celular e por isso tendem a exibir caracteriacutesticas metaboacutelicas
associadas ao efeito de Warburg 20
O principal efector molecular do efeito de Warburg eacute o factor 1 indutiacutevel por hipoxia
(HIF1) um factor de transcriccedilatildeo heterodimeacuterico constituiacutedo por uma subunidade β expressa
constitutivamente e uma subunidade α cuja expressatildeo estaacute dependente do oxigeacutenio Apesar
de ser expressa em condiccedilotildees de baixa disponibilidade de oxigeacutenio (hipoxia) a expressatildeo
constitutiva da subunidade α pode ocorrer em normoxia como resultado de alteraccedilotildees nos
sistemas de sinalizaccedilatildeo para degradaccedilatildeo proteica provocadas durante o processo de
imortalizaccedilatildeo celular De facto a sua sobre-expressatildeo eacute caracteriacutestica comum a muitas ceacutelulas
canceriacutegenas 2223
O factor de transcriccedilatildeo HIF1 eacute responsaacutevel pela coordenaccedilatildeo da alteraccedilatildeo
do metabolismo para glicoacutelise com formaccedilatildeo de lactato com consequente aumento do
consumo de glucose e acumulaccedilatildeo de lactato jaacute que promove a expressatildeo de vaacuterios
componentes do metabolismo energeacutetico nomeadamente transportadores de glucose (GLUT-
1) enzimas da via glicoliacutetica e de lactato desidrogenase (Figura 1) 2425
Para aleacutem do papel do HIF1 na promoccedilatildeo da glicoacutelise com formaccedilatildeo de lactato
estudos recentes revelaram uma significativa influecircncia na respiraccedilatildeo mitocondrial Nos
organismos aeroacutebios o TCA possui um papel fundamental no metabolismo central das
ceacutelulas constituindo a via de produccedilatildeo de energia mais eficiente e fornecendo o poder redutor
e intermediaacuterios necessaacuterios para as reacccedilotildees de biossiacutentese Normalmente a glucose eacute
convertida em piruvato no citoplasma o qual eacute posteriormente canalizado para o TCA ao
niacutevel mitocondrial onde eacute oxidado a dioacutexido de carbono e aacutegua A canalizaccedilatildeo do piruvato
3
para o TCA implica a sua conversatildeo em acetil-CoA numa reacccedilatildeo catalisada pelo complexo
multi-enzimaacutetico piruvato desidrogenase (PDH) Sendo esta uma das reacccedilotildees mais
importantes do metabolismo celular estaacute sujeita a um elevado niacutevel de regulaccedilatildeo Como tal o
complexo PDH eacute regulado por um ciclo reversiacutevel de fosforilaccedilatildeo caracterizado pela
fosforilaccedilatildeo (inactivaccedilatildeo) e desfosforilaccedilatildeo (activaccedilatildeo) da subunidade α do componente E1
(PDHE1α) promovida por cinases (PDK) e fosfatases (PDP) respectivamente Na PDHE1α
de mamiacuteferos existem trecircs locais de fosforilaccedilatildeo (Ser232
Ser293
e Ser300
) que podem ser
fosforilados por quatro isoenzimas (PDK1-4) sendo a PDK1 a uacutenica que consegue fosforilar
todos os trecircs locais 2627
Interessantemente a sobre-expressatildeo de PDK1 e PDK3 eacute promovida
por HIF1 levando assim agrave fosforilaccedilatildeo de PDHE1α e inibiccedilatildeo do complexo PDH (Figura 1)
Consequentemente a entrada do carbono derivado da via glicoliacutetica no TCA ocorre em menor
extensatildeo e aumenta a conversatildeo de piruvato a lactato resultando no fenoacutetipo lactogeacutenico
associado ao efeito de Warburg e na desregulaccedilatildeo do metabolismo energeacutetico 2428
Figura 1 Vias do metabolismo energeacutetico reguladas pelo factor de transcriccedilatildeo HIF1 A sobre-expressatildeo de
vaacuterias enzimas da via glicoliacutetica e da enzima lactato desidrogenase (LDH) eacute promovida pelo factor 1 indutiacutevel
por hipoxia (HIF1) levando ao aumento do consumo de glucose e agrave acumulaccedilatildeo de lactato A entrada do
carbono derivado da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) eacute realizada ao niacutevel do complexo
piruvato desidrogenase (PDH) sendo este regulado atraveacutes de um ciclo reversiacutevel de fosforilaccedilatildeo HIF1 promove
a expressatildeo das cinases da piruvato desidrogenase (PDK1-4) resultando na fosforilaccedilatildeo e consequente inibiccedilatildeo
do complexo PDH Esquema adaptado de Kondoh (2008) 20
Glucose
Glucose 6P
Fuctose 6P
Glyceraldehyde-3P
Fuctose 16 BP
Dihydroxyacetone-P
13 Biphosphoglycerate
3 - Phosphoglycerate
2 - Phosphoglycerate
Phosphoenopyruvate Pyruvate
Cytoplasm
Extracellular space
hexokinase
phosphoglucose isomerase
phosphofructokinase
GAPDH
phosphoglycerate
kinase
phosphoglucomutase
enolase
Pyruvate kinase
Pyruvate
Acetyl-CoATCA
PDH
PDH
P
LDH
Lactate
PDK
PDK2
PDK1 PDK3PDK4
aldolase
+
+
+
+
+
++
+
+
HIF1
4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

iv
iv LISTA DE ABREVIATURAS
Abreviatura Descriccedilatildeo
cDNA ADN complementar (do Inglecircs complementary DNA)
DNA Aacutecido desoxirribonucleico (do Inglecircs Deoxyribonucleic Acid)
ERO Espeacutecies reactivas de oxigeacutenio
GFP Proteiacutena fluorescente verde (do Inglecircs Green Fluorescent Protein)
HIF1 Factor 1 indutiacutevel por hipoxia (do Inglecircs Hypoxia Inducible Factor 1)
LDH Lactato desidrogenase (do Inglecircs Lactate Dehydrogenase)
mRNA ARN mensageiro (do Inglecircs messenger RNA)
ntshRNA non-target short hairpin RNA
PCR Reacccedilatildeo em cadeia da polimerase (do Inglecircs Polymerase chain reaction)
PDH Complexo da piruvato desidrogenase (do Inglecircs Pyruvate
Dehydrogenase Complex)
PDHE1α Subunidade α do componente E1 do complexo piruvato desidrogenase
(do Inglecircs E1 alpha subunit of pyruvate dehydrogenase complex)
PDK Cinase da piruvato desidrogenase (do Inglecircs Pyruvate Dehydrogenase
Kinase)
PDP Fosfatase da piruvato desidrogenase (do Inglecircs Pyruvate dehydrogenase
phosphatase)
PI Partiacuteculas infecciosas
PT Partiacuteculas totais
qRT-PCR PCR quantitativo por transcriccedilatildeo reversa (do Inglecircs Quantitative reverse
transcription polymerase chain reaction)
RFP Proteiacutena fluorescente vermelha (do Inglecircs Red Fluorescent Protein)
RNA Aacutecido ribonucleico (do Inglecircs Ribonucleic Acid)
shRNA short hairpin RNA
TCA Ciclo dos Aacutecidos Tricarboxiacutelicos (do Inglecircs Tricarboxylic Acid Cycle)
v
V IacuteNDICE
i AGRADECIMENTOS i
ii RESUMO ii
iii ABSTRACT iii
iv LISTA DE ABREVIATURAS iv
1 INTRODUCcedilAtildeO 1
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico 1
12 Imortalizaccedilatildeo celular e o efeito de Warburg 2
13 Impacto do efeito de Warburg em biotecnologia 4
14 Objectivos principais 5
2 MATERIAIS E MEacuteTODOS 6
21 Linhas celulares e meios de cultura 6
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais 6
23 Transduccedilatildeo celular selecccedilatildeo e clonagem 7
24 Estudos de crescimento e anaacutelise de metabolitos 7
25 Extracccedilatildeo de RNA total e qRT-PCR 8
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais8
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais 8
28 Determinaccedilatildeo da actividade da lactato desidrogenase9
29 Western Blot 9
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina 10
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio 10
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade 10
213 Caacutelculos 10
3 RESULTADOS 11
31 Silenciamento de HIF1 11
vi
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3 15
4 DISCUSSAtildeO E CONCLUSAtildeO 24
5 PERSPECTIVAS FUTURAS 29
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS 30
7 ANEXOS 34
1
1 INTRODUCcedilAtildeO
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico
Nos uacuteltimos anos tem-se verificado uma crescente procura de biofaacutermacos complexos
como proteiacutenas recombinantes e partiacuteculas derivadas de viacuterus quer para fins terapecircuticos quer
para fins de diagnoacutesticos in vivo O potencial dos biofaacutermacos derivados de viacuterus em
particular tem vindo a aumentar devido agraves suas diversas aplicaccedilotildees profilaacuteticas terapecircuticas e
cliacutenicas como a vacinaccedilatildeo o tratamento do cancro e a terapia geacutenica 1 Actualmente o
sistema preferencial para a manufacturaccedilatildeo destes produtos biofarmacecircuticos complexos eacute as
ceacutelulas de mamiacutefero usadas para a produccedilatildeo de mais de metade dos biofaacutermacos existentes
no mercado Tal predomiacutenio deve-se agrave sua semelhanccedila com as ceacutelulas humanas no folding de
proteiacutenas e nas modificaccedilotildees poacutes-traducionais como a glicosilaccedilatildeo aumentando a eficiecircncia
terapecircutica e diminuindo o potencial imunogeacutenico do produto final 23
As ceacutelulas de mamiacutefero em cultura agrave semelhanccedila de ceacutelulas transformadas possuem
um metabolismo energeacutetico desregulado consumindo elevadas quantidades de glucose e
acumulando quantidades significativas de lactato com reduzida entrada de carbono derivado
da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) 4 Do ponto de vista da cultura de
ceacutelulas para produccedilatildeo de biofaacutermacos este fenoacutetipo lactogeacutenico tem simultaneamente um
efeito negativo no crescimento e viabilidade celular e na produtividade e qualidade do
biofaacutermaco Este fenoacutemeno eacute ainda mais importante no caso das partiacuteculas derivadas de viacuterus
para terapia geacutenica jaacute que a sua infecciosidade eacute fortemente influenciada por variaccedilotildees no pH
do meio de cultura 5 Diversas estrateacutegias tecircm sido testadas de forma a reduzir os efeitos
nocivos da acumulaccedilatildeo de lactato no meio de cultura fornecimento de glucose de forma
controlada 67
suplementaccedilatildeo do meio de cultura com substratos alternativos agrave glucose 8
remoccedilatildeo dos metabolitos toacutexicos 9 desenvolvimento de meios de cultura com base na anaacutelise
dos fluxos metaboacutelicos permitindo adaptar os seus constituintes agraves necessidades metaboacutelicas
das ceacutelulas 1011
introduccedilatildeo da enzima piruvato carboxilase nas ceacutelulas 12-14
diminuiccedilatildeo da
expressatildeo da lactato desidrogenase atraveacutes da disrupccedilatildeo do gene Ldha por recombinaccedilatildeo
homoacuteloga 15
ou atraveacutes de RNA de interferecircncia 16
ou introduccedilatildeo de um transportador para a
frutose constituindo uma alternativa ao consumo de glucose 17
No entanto estas estrateacutegias
tecircm um sucesso limitado devido aos efeitos compensatoacuterios causados pela apertada regulaccedilatildeo
homeostaacutetica intracelular que natildeo satildeo facilmente alteraacuteveis
2
12 Imortalizaccedilatildeo celular e o efeito de Warburg
Nos anos 20 Otto Warburg observou que as ceacutelulas canceriacutegenas comparativamente
agraves ceacutelulas normais apresentavam elevadas taxas de consumo de glucose secretando a maior
parte do carbono derivado da glucose sob a forma de lactato em vez de o oxidar
completamente 1819
ndash um cenaacuterio que ficou conhecido como efeito de Warburg Estudos
recentes sobre o metabolismo energeacutetico de ceacutelulas canceriacutegenas sugerem que este resulta de
alteraccedilotildees geneacuteticas que ocorrem no processo de tumorigeacutenese principalmente durante a
imortalizaccedilatildeo e transformaccedilatildeo celulares e que levam agrave desregulaccedilatildeo do metabolismo
energeacutetico central e ao aumento da actividade glicoliacutetica 20
De facto existem diversas vias de
sinalizaccedilatildeo comuns ao controlo da proliferaccedilatildeo e do metabolismo energeacutetico que levam agrave
induccedilatildeo deste cenaacuterio como a activaccedilatildeo de oncogenes (MYC) e enzimas glicoliacuteticas ou a
supressatildeo do gene supressor de tumores P53 2021
Interessantemente muitas das vias de
imortalizaccedilatildeo activadas no processo de tumorigeacutenese satildeo tambeacutem recrutadas durante o
estabelecimento de uma linha celular e por isso tendem a exibir caracteriacutesticas metaboacutelicas
associadas ao efeito de Warburg 20
O principal efector molecular do efeito de Warburg eacute o factor 1 indutiacutevel por hipoxia
(HIF1) um factor de transcriccedilatildeo heterodimeacuterico constituiacutedo por uma subunidade β expressa
constitutivamente e uma subunidade α cuja expressatildeo estaacute dependente do oxigeacutenio Apesar
de ser expressa em condiccedilotildees de baixa disponibilidade de oxigeacutenio (hipoxia) a expressatildeo
constitutiva da subunidade α pode ocorrer em normoxia como resultado de alteraccedilotildees nos
sistemas de sinalizaccedilatildeo para degradaccedilatildeo proteica provocadas durante o processo de
imortalizaccedilatildeo celular De facto a sua sobre-expressatildeo eacute caracteriacutestica comum a muitas ceacutelulas
canceriacutegenas 2223
O factor de transcriccedilatildeo HIF1 eacute responsaacutevel pela coordenaccedilatildeo da alteraccedilatildeo
do metabolismo para glicoacutelise com formaccedilatildeo de lactato com consequente aumento do
consumo de glucose e acumulaccedilatildeo de lactato jaacute que promove a expressatildeo de vaacuterios
componentes do metabolismo energeacutetico nomeadamente transportadores de glucose (GLUT-
1) enzimas da via glicoliacutetica e de lactato desidrogenase (Figura 1) 2425
Para aleacutem do papel do HIF1 na promoccedilatildeo da glicoacutelise com formaccedilatildeo de lactato
estudos recentes revelaram uma significativa influecircncia na respiraccedilatildeo mitocondrial Nos
organismos aeroacutebios o TCA possui um papel fundamental no metabolismo central das
ceacutelulas constituindo a via de produccedilatildeo de energia mais eficiente e fornecendo o poder redutor
e intermediaacuterios necessaacuterios para as reacccedilotildees de biossiacutentese Normalmente a glucose eacute
convertida em piruvato no citoplasma o qual eacute posteriormente canalizado para o TCA ao
niacutevel mitocondrial onde eacute oxidado a dioacutexido de carbono e aacutegua A canalizaccedilatildeo do piruvato
3
para o TCA implica a sua conversatildeo em acetil-CoA numa reacccedilatildeo catalisada pelo complexo
multi-enzimaacutetico piruvato desidrogenase (PDH) Sendo esta uma das reacccedilotildees mais
importantes do metabolismo celular estaacute sujeita a um elevado niacutevel de regulaccedilatildeo Como tal o
complexo PDH eacute regulado por um ciclo reversiacutevel de fosforilaccedilatildeo caracterizado pela
fosforilaccedilatildeo (inactivaccedilatildeo) e desfosforilaccedilatildeo (activaccedilatildeo) da subunidade α do componente E1
(PDHE1α) promovida por cinases (PDK) e fosfatases (PDP) respectivamente Na PDHE1α
de mamiacuteferos existem trecircs locais de fosforilaccedilatildeo (Ser232
Ser293
e Ser300
) que podem ser
fosforilados por quatro isoenzimas (PDK1-4) sendo a PDK1 a uacutenica que consegue fosforilar
todos os trecircs locais 2627
Interessantemente a sobre-expressatildeo de PDK1 e PDK3 eacute promovida
por HIF1 levando assim agrave fosforilaccedilatildeo de PDHE1α e inibiccedilatildeo do complexo PDH (Figura 1)
Consequentemente a entrada do carbono derivado da via glicoliacutetica no TCA ocorre em menor
extensatildeo e aumenta a conversatildeo de piruvato a lactato resultando no fenoacutetipo lactogeacutenico
associado ao efeito de Warburg e na desregulaccedilatildeo do metabolismo energeacutetico 2428
Figura 1 Vias do metabolismo energeacutetico reguladas pelo factor de transcriccedilatildeo HIF1 A sobre-expressatildeo de
vaacuterias enzimas da via glicoliacutetica e da enzima lactato desidrogenase (LDH) eacute promovida pelo factor 1 indutiacutevel
por hipoxia (HIF1) levando ao aumento do consumo de glucose e agrave acumulaccedilatildeo de lactato A entrada do
carbono derivado da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) eacute realizada ao niacutevel do complexo
piruvato desidrogenase (PDH) sendo este regulado atraveacutes de um ciclo reversiacutevel de fosforilaccedilatildeo HIF1 promove
a expressatildeo das cinases da piruvato desidrogenase (PDK1-4) resultando na fosforilaccedilatildeo e consequente inibiccedilatildeo
do complexo PDH Esquema adaptado de Kondoh (2008) 20
Glucose
Glucose 6P
Fuctose 6P
Glyceraldehyde-3P
Fuctose 16 BP
Dihydroxyacetone-P
13 Biphosphoglycerate
3 - Phosphoglycerate
2 - Phosphoglycerate
Phosphoenopyruvate Pyruvate
Cytoplasm
Extracellular space
hexokinase
phosphoglucose isomerase
phosphofructokinase
GAPDH
phosphoglycerate
kinase
phosphoglucomutase
enolase
Pyruvate kinase
Pyruvate
Acetyl-CoATCA
PDH
PDH
P
LDH
Lactate
PDK
PDK2
PDK1 PDK3PDK4
aldolase
+
+
+
+
+
++
+
+
HIF1
4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

v
V IacuteNDICE
i AGRADECIMENTOS i
ii RESUMO ii
iii ABSTRACT iii
iv LISTA DE ABREVIATURAS iv
1 INTRODUCcedilAtildeO 1
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico 1
12 Imortalizaccedilatildeo celular e o efeito de Warburg 2
13 Impacto do efeito de Warburg em biotecnologia 4
14 Objectivos principais 5
2 MATERIAIS E MEacuteTODOS 6
21 Linhas celulares e meios de cultura 6
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais 6
23 Transduccedilatildeo celular selecccedilatildeo e clonagem 7
24 Estudos de crescimento e anaacutelise de metabolitos 7
25 Extracccedilatildeo de RNA total e qRT-PCR 8
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais8
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais 8
28 Determinaccedilatildeo da actividade da lactato desidrogenase9
29 Western Blot 9
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina 10
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio 10
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade 10
213 Caacutelculos 10
3 RESULTADOS 11
31 Silenciamento de HIF1 11
vi
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3 15
4 DISCUSSAtildeO E CONCLUSAtildeO 24
5 PERSPECTIVAS FUTURAS 29
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS 30
7 ANEXOS 34
1
1 INTRODUCcedilAtildeO
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico
Nos uacuteltimos anos tem-se verificado uma crescente procura de biofaacutermacos complexos
como proteiacutenas recombinantes e partiacuteculas derivadas de viacuterus quer para fins terapecircuticos quer
para fins de diagnoacutesticos in vivo O potencial dos biofaacutermacos derivados de viacuterus em
particular tem vindo a aumentar devido agraves suas diversas aplicaccedilotildees profilaacuteticas terapecircuticas e
cliacutenicas como a vacinaccedilatildeo o tratamento do cancro e a terapia geacutenica 1 Actualmente o
sistema preferencial para a manufacturaccedilatildeo destes produtos biofarmacecircuticos complexos eacute as
ceacutelulas de mamiacutefero usadas para a produccedilatildeo de mais de metade dos biofaacutermacos existentes
no mercado Tal predomiacutenio deve-se agrave sua semelhanccedila com as ceacutelulas humanas no folding de
proteiacutenas e nas modificaccedilotildees poacutes-traducionais como a glicosilaccedilatildeo aumentando a eficiecircncia
terapecircutica e diminuindo o potencial imunogeacutenico do produto final 23
As ceacutelulas de mamiacutefero em cultura agrave semelhanccedila de ceacutelulas transformadas possuem
um metabolismo energeacutetico desregulado consumindo elevadas quantidades de glucose e
acumulando quantidades significativas de lactato com reduzida entrada de carbono derivado
da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) 4 Do ponto de vista da cultura de
ceacutelulas para produccedilatildeo de biofaacutermacos este fenoacutetipo lactogeacutenico tem simultaneamente um
efeito negativo no crescimento e viabilidade celular e na produtividade e qualidade do
biofaacutermaco Este fenoacutemeno eacute ainda mais importante no caso das partiacuteculas derivadas de viacuterus
para terapia geacutenica jaacute que a sua infecciosidade eacute fortemente influenciada por variaccedilotildees no pH
do meio de cultura 5 Diversas estrateacutegias tecircm sido testadas de forma a reduzir os efeitos
nocivos da acumulaccedilatildeo de lactato no meio de cultura fornecimento de glucose de forma
controlada 67
suplementaccedilatildeo do meio de cultura com substratos alternativos agrave glucose 8
remoccedilatildeo dos metabolitos toacutexicos 9 desenvolvimento de meios de cultura com base na anaacutelise
dos fluxos metaboacutelicos permitindo adaptar os seus constituintes agraves necessidades metaboacutelicas
das ceacutelulas 1011
introduccedilatildeo da enzima piruvato carboxilase nas ceacutelulas 12-14
diminuiccedilatildeo da
expressatildeo da lactato desidrogenase atraveacutes da disrupccedilatildeo do gene Ldha por recombinaccedilatildeo
homoacuteloga 15
ou atraveacutes de RNA de interferecircncia 16
ou introduccedilatildeo de um transportador para a
frutose constituindo uma alternativa ao consumo de glucose 17
No entanto estas estrateacutegias
tecircm um sucesso limitado devido aos efeitos compensatoacuterios causados pela apertada regulaccedilatildeo
homeostaacutetica intracelular que natildeo satildeo facilmente alteraacuteveis
2
12 Imortalizaccedilatildeo celular e o efeito de Warburg
Nos anos 20 Otto Warburg observou que as ceacutelulas canceriacutegenas comparativamente
agraves ceacutelulas normais apresentavam elevadas taxas de consumo de glucose secretando a maior
parte do carbono derivado da glucose sob a forma de lactato em vez de o oxidar
completamente 1819
ndash um cenaacuterio que ficou conhecido como efeito de Warburg Estudos
recentes sobre o metabolismo energeacutetico de ceacutelulas canceriacutegenas sugerem que este resulta de
alteraccedilotildees geneacuteticas que ocorrem no processo de tumorigeacutenese principalmente durante a
imortalizaccedilatildeo e transformaccedilatildeo celulares e que levam agrave desregulaccedilatildeo do metabolismo
energeacutetico central e ao aumento da actividade glicoliacutetica 20
De facto existem diversas vias de
sinalizaccedilatildeo comuns ao controlo da proliferaccedilatildeo e do metabolismo energeacutetico que levam agrave
induccedilatildeo deste cenaacuterio como a activaccedilatildeo de oncogenes (MYC) e enzimas glicoliacuteticas ou a
supressatildeo do gene supressor de tumores P53 2021
Interessantemente muitas das vias de
imortalizaccedilatildeo activadas no processo de tumorigeacutenese satildeo tambeacutem recrutadas durante o
estabelecimento de uma linha celular e por isso tendem a exibir caracteriacutesticas metaboacutelicas
associadas ao efeito de Warburg 20
O principal efector molecular do efeito de Warburg eacute o factor 1 indutiacutevel por hipoxia
(HIF1) um factor de transcriccedilatildeo heterodimeacuterico constituiacutedo por uma subunidade β expressa
constitutivamente e uma subunidade α cuja expressatildeo estaacute dependente do oxigeacutenio Apesar
de ser expressa em condiccedilotildees de baixa disponibilidade de oxigeacutenio (hipoxia) a expressatildeo
constitutiva da subunidade α pode ocorrer em normoxia como resultado de alteraccedilotildees nos
sistemas de sinalizaccedilatildeo para degradaccedilatildeo proteica provocadas durante o processo de
imortalizaccedilatildeo celular De facto a sua sobre-expressatildeo eacute caracteriacutestica comum a muitas ceacutelulas
canceriacutegenas 2223
O factor de transcriccedilatildeo HIF1 eacute responsaacutevel pela coordenaccedilatildeo da alteraccedilatildeo
do metabolismo para glicoacutelise com formaccedilatildeo de lactato com consequente aumento do
consumo de glucose e acumulaccedilatildeo de lactato jaacute que promove a expressatildeo de vaacuterios
componentes do metabolismo energeacutetico nomeadamente transportadores de glucose (GLUT-
1) enzimas da via glicoliacutetica e de lactato desidrogenase (Figura 1) 2425
Para aleacutem do papel do HIF1 na promoccedilatildeo da glicoacutelise com formaccedilatildeo de lactato
estudos recentes revelaram uma significativa influecircncia na respiraccedilatildeo mitocondrial Nos
organismos aeroacutebios o TCA possui um papel fundamental no metabolismo central das
ceacutelulas constituindo a via de produccedilatildeo de energia mais eficiente e fornecendo o poder redutor
e intermediaacuterios necessaacuterios para as reacccedilotildees de biossiacutentese Normalmente a glucose eacute
convertida em piruvato no citoplasma o qual eacute posteriormente canalizado para o TCA ao
niacutevel mitocondrial onde eacute oxidado a dioacutexido de carbono e aacutegua A canalizaccedilatildeo do piruvato
3
para o TCA implica a sua conversatildeo em acetil-CoA numa reacccedilatildeo catalisada pelo complexo
multi-enzimaacutetico piruvato desidrogenase (PDH) Sendo esta uma das reacccedilotildees mais
importantes do metabolismo celular estaacute sujeita a um elevado niacutevel de regulaccedilatildeo Como tal o
complexo PDH eacute regulado por um ciclo reversiacutevel de fosforilaccedilatildeo caracterizado pela
fosforilaccedilatildeo (inactivaccedilatildeo) e desfosforilaccedilatildeo (activaccedilatildeo) da subunidade α do componente E1
(PDHE1α) promovida por cinases (PDK) e fosfatases (PDP) respectivamente Na PDHE1α
de mamiacuteferos existem trecircs locais de fosforilaccedilatildeo (Ser232
Ser293
e Ser300
) que podem ser
fosforilados por quatro isoenzimas (PDK1-4) sendo a PDK1 a uacutenica que consegue fosforilar
todos os trecircs locais 2627
Interessantemente a sobre-expressatildeo de PDK1 e PDK3 eacute promovida
por HIF1 levando assim agrave fosforilaccedilatildeo de PDHE1α e inibiccedilatildeo do complexo PDH (Figura 1)
Consequentemente a entrada do carbono derivado da via glicoliacutetica no TCA ocorre em menor
extensatildeo e aumenta a conversatildeo de piruvato a lactato resultando no fenoacutetipo lactogeacutenico
associado ao efeito de Warburg e na desregulaccedilatildeo do metabolismo energeacutetico 2428
Figura 1 Vias do metabolismo energeacutetico reguladas pelo factor de transcriccedilatildeo HIF1 A sobre-expressatildeo de
vaacuterias enzimas da via glicoliacutetica e da enzima lactato desidrogenase (LDH) eacute promovida pelo factor 1 indutiacutevel
por hipoxia (HIF1) levando ao aumento do consumo de glucose e agrave acumulaccedilatildeo de lactato A entrada do
carbono derivado da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) eacute realizada ao niacutevel do complexo
piruvato desidrogenase (PDH) sendo este regulado atraveacutes de um ciclo reversiacutevel de fosforilaccedilatildeo HIF1 promove
a expressatildeo das cinases da piruvato desidrogenase (PDK1-4) resultando na fosforilaccedilatildeo e consequente inibiccedilatildeo
do complexo PDH Esquema adaptado de Kondoh (2008) 20
Glucose
Glucose 6P
Fuctose 6P
Glyceraldehyde-3P
Fuctose 16 BP
Dihydroxyacetone-P
13 Biphosphoglycerate
3 - Phosphoglycerate
2 - Phosphoglycerate
Phosphoenopyruvate Pyruvate
Cytoplasm
Extracellular space
hexokinase
phosphoglucose isomerase
phosphofructokinase
GAPDH
phosphoglycerate
kinase
phosphoglucomutase
enolase
Pyruvate kinase
Pyruvate
Acetyl-CoATCA
PDH
PDH
P
LDH
Lactate
PDK
PDK2
PDK1 PDK3PDK4
aldolase
+
+
+
+
+
++
+
+
HIF1
4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

vi
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3 15
4 DISCUSSAtildeO E CONCLUSAtildeO 24
5 PERSPECTIVAS FUTURAS 29
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS 30
7 ANEXOS 34
1
1 INTRODUCcedilAtildeO
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico
Nos uacuteltimos anos tem-se verificado uma crescente procura de biofaacutermacos complexos
como proteiacutenas recombinantes e partiacuteculas derivadas de viacuterus quer para fins terapecircuticos quer
para fins de diagnoacutesticos in vivo O potencial dos biofaacutermacos derivados de viacuterus em
particular tem vindo a aumentar devido agraves suas diversas aplicaccedilotildees profilaacuteticas terapecircuticas e
cliacutenicas como a vacinaccedilatildeo o tratamento do cancro e a terapia geacutenica 1 Actualmente o
sistema preferencial para a manufacturaccedilatildeo destes produtos biofarmacecircuticos complexos eacute as
ceacutelulas de mamiacutefero usadas para a produccedilatildeo de mais de metade dos biofaacutermacos existentes
no mercado Tal predomiacutenio deve-se agrave sua semelhanccedila com as ceacutelulas humanas no folding de
proteiacutenas e nas modificaccedilotildees poacutes-traducionais como a glicosilaccedilatildeo aumentando a eficiecircncia
terapecircutica e diminuindo o potencial imunogeacutenico do produto final 23
As ceacutelulas de mamiacutefero em cultura agrave semelhanccedila de ceacutelulas transformadas possuem
um metabolismo energeacutetico desregulado consumindo elevadas quantidades de glucose e
acumulando quantidades significativas de lactato com reduzida entrada de carbono derivado
da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) 4 Do ponto de vista da cultura de
ceacutelulas para produccedilatildeo de biofaacutermacos este fenoacutetipo lactogeacutenico tem simultaneamente um
efeito negativo no crescimento e viabilidade celular e na produtividade e qualidade do
biofaacutermaco Este fenoacutemeno eacute ainda mais importante no caso das partiacuteculas derivadas de viacuterus
para terapia geacutenica jaacute que a sua infecciosidade eacute fortemente influenciada por variaccedilotildees no pH
do meio de cultura 5 Diversas estrateacutegias tecircm sido testadas de forma a reduzir os efeitos
nocivos da acumulaccedilatildeo de lactato no meio de cultura fornecimento de glucose de forma
controlada 67
suplementaccedilatildeo do meio de cultura com substratos alternativos agrave glucose 8
remoccedilatildeo dos metabolitos toacutexicos 9 desenvolvimento de meios de cultura com base na anaacutelise
dos fluxos metaboacutelicos permitindo adaptar os seus constituintes agraves necessidades metaboacutelicas
das ceacutelulas 1011
introduccedilatildeo da enzima piruvato carboxilase nas ceacutelulas 12-14
diminuiccedilatildeo da
expressatildeo da lactato desidrogenase atraveacutes da disrupccedilatildeo do gene Ldha por recombinaccedilatildeo
homoacuteloga 15
ou atraveacutes de RNA de interferecircncia 16
ou introduccedilatildeo de um transportador para a
frutose constituindo uma alternativa ao consumo de glucose 17
No entanto estas estrateacutegias
tecircm um sucesso limitado devido aos efeitos compensatoacuterios causados pela apertada regulaccedilatildeo
homeostaacutetica intracelular que natildeo satildeo facilmente alteraacuteveis
2
12 Imortalizaccedilatildeo celular e o efeito de Warburg
Nos anos 20 Otto Warburg observou que as ceacutelulas canceriacutegenas comparativamente
agraves ceacutelulas normais apresentavam elevadas taxas de consumo de glucose secretando a maior
parte do carbono derivado da glucose sob a forma de lactato em vez de o oxidar
completamente 1819
ndash um cenaacuterio que ficou conhecido como efeito de Warburg Estudos
recentes sobre o metabolismo energeacutetico de ceacutelulas canceriacutegenas sugerem que este resulta de
alteraccedilotildees geneacuteticas que ocorrem no processo de tumorigeacutenese principalmente durante a
imortalizaccedilatildeo e transformaccedilatildeo celulares e que levam agrave desregulaccedilatildeo do metabolismo
energeacutetico central e ao aumento da actividade glicoliacutetica 20
De facto existem diversas vias de
sinalizaccedilatildeo comuns ao controlo da proliferaccedilatildeo e do metabolismo energeacutetico que levam agrave
induccedilatildeo deste cenaacuterio como a activaccedilatildeo de oncogenes (MYC) e enzimas glicoliacuteticas ou a
supressatildeo do gene supressor de tumores P53 2021
Interessantemente muitas das vias de
imortalizaccedilatildeo activadas no processo de tumorigeacutenese satildeo tambeacutem recrutadas durante o
estabelecimento de uma linha celular e por isso tendem a exibir caracteriacutesticas metaboacutelicas
associadas ao efeito de Warburg 20
O principal efector molecular do efeito de Warburg eacute o factor 1 indutiacutevel por hipoxia
(HIF1) um factor de transcriccedilatildeo heterodimeacuterico constituiacutedo por uma subunidade β expressa
constitutivamente e uma subunidade α cuja expressatildeo estaacute dependente do oxigeacutenio Apesar
de ser expressa em condiccedilotildees de baixa disponibilidade de oxigeacutenio (hipoxia) a expressatildeo
constitutiva da subunidade α pode ocorrer em normoxia como resultado de alteraccedilotildees nos
sistemas de sinalizaccedilatildeo para degradaccedilatildeo proteica provocadas durante o processo de
imortalizaccedilatildeo celular De facto a sua sobre-expressatildeo eacute caracteriacutestica comum a muitas ceacutelulas
canceriacutegenas 2223
O factor de transcriccedilatildeo HIF1 eacute responsaacutevel pela coordenaccedilatildeo da alteraccedilatildeo
do metabolismo para glicoacutelise com formaccedilatildeo de lactato com consequente aumento do
consumo de glucose e acumulaccedilatildeo de lactato jaacute que promove a expressatildeo de vaacuterios
componentes do metabolismo energeacutetico nomeadamente transportadores de glucose (GLUT-
1) enzimas da via glicoliacutetica e de lactato desidrogenase (Figura 1) 2425
Para aleacutem do papel do HIF1 na promoccedilatildeo da glicoacutelise com formaccedilatildeo de lactato
estudos recentes revelaram uma significativa influecircncia na respiraccedilatildeo mitocondrial Nos
organismos aeroacutebios o TCA possui um papel fundamental no metabolismo central das
ceacutelulas constituindo a via de produccedilatildeo de energia mais eficiente e fornecendo o poder redutor
e intermediaacuterios necessaacuterios para as reacccedilotildees de biossiacutentese Normalmente a glucose eacute
convertida em piruvato no citoplasma o qual eacute posteriormente canalizado para o TCA ao
niacutevel mitocondrial onde eacute oxidado a dioacutexido de carbono e aacutegua A canalizaccedilatildeo do piruvato
3
para o TCA implica a sua conversatildeo em acetil-CoA numa reacccedilatildeo catalisada pelo complexo
multi-enzimaacutetico piruvato desidrogenase (PDH) Sendo esta uma das reacccedilotildees mais
importantes do metabolismo celular estaacute sujeita a um elevado niacutevel de regulaccedilatildeo Como tal o
complexo PDH eacute regulado por um ciclo reversiacutevel de fosforilaccedilatildeo caracterizado pela
fosforilaccedilatildeo (inactivaccedilatildeo) e desfosforilaccedilatildeo (activaccedilatildeo) da subunidade α do componente E1
(PDHE1α) promovida por cinases (PDK) e fosfatases (PDP) respectivamente Na PDHE1α
de mamiacuteferos existem trecircs locais de fosforilaccedilatildeo (Ser232
Ser293
e Ser300
) que podem ser
fosforilados por quatro isoenzimas (PDK1-4) sendo a PDK1 a uacutenica que consegue fosforilar
todos os trecircs locais 2627
Interessantemente a sobre-expressatildeo de PDK1 e PDK3 eacute promovida
por HIF1 levando assim agrave fosforilaccedilatildeo de PDHE1α e inibiccedilatildeo do complexo PDH (Figura 1)
Consequentemente a entrada do carbono derivado da via glicoliacutetica no TCA ocorre em menor
extensatildeo e aumenta a conversatildeo de piruvato a lactato resultando no fenoacutetipo lactogeacutenico
associado ao efeito de Warburg e na desregulaccedilatildeo do metabolismo energeacutetico 2428
Figura 1 Vias do metabolismo energeacutetico reguladas pelo factor de transcriccedilatildeo HIF1 A sobre-expressatildeo de
vaacuterias enzimas da via glicoliacutetica e da enzima lactato desidrogenase (LDH) eacute promovida pelo factor 1 indutiacutevel
por hipoxia (HIF1) levando ao aumento do consumo de glucose e agrave acumulaccedilatildeo de lactato A entrada do
carbono derivado da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) eacute realizada ao niacutevel do complexo
piruvato desidrogenase (PDH) sendo este regulado atraveacutes de um ciclo reversiacutevel de fosforilaccedilatildeo HIF1 promove
a expressatildeo das cinases da piruvato desidrogenase (PDK1-4) resultando na fosforilaccedilatildeo e consequente inibiccedilatildeo
do complexo PDH Esquema adaptado de Kondoh (2008) 20
Glucose
Glucose 6P
Fuctose 6P
Glyceraldehyde-3P
Fuctose 16 BP
Dihydroxyacetone-P
13 Biphosphoglycerate
3 - Phosphoglycerate
2 - Phosphoglycerate
Phosphoenopyruvate Pyruvate
Cytoplasm
Extracellular space
hexokinase
phosphoglucose isomerase
phosphofructokinase
GAPDH
phosphoglycerate
kinase
phosphoglucomutase
enolase
Pyruvate kinase
Pyruvate
Acetyl-CoATCA
PDH
PDH
P
LDH
Lactate
PDK
PDK2
PDK1 PDK3PDK4
aldolase
+
+
+
+
+
++
+
+
HIF1
4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

1
1 INTRODUCcedilAtildeO
11 Cultura de ceacutelulas de mamiacutefero e o fenoacutetipo lactogeacutenico
Nos uacuteltimos anos tem-se verificado uma crescente procura de biofaacutermacos complexos
como proteiacutenas recombinantes e partiacuteculas derivadas de viacuterus quer para fins terapecircuticos quer
para fins de diagnoacutesticos in vivo O potencial dos biofaacutermacos derivados de viacuterus em
particular tem vindo a aumentar devido agraves suas diversas aplicaccedilotildees profilaacuteticas terapecircuticas e
cliacutenicas como a vacinaccedilatildeo o tratamento do cancro e a terapia geacutenica 1 Actualmente o
sistema preferencial para a manufacturaccedilatildeo destes produtos biofarmacecircuticos complexos eacute as
ceacutelulas de mamiacutefero usadas para a produccedilatildeo de mais de metade dos biofaacutermacos existentes
no mercado Tal predomiacutenio deve-se agrave sua semelhanccedila com as ceacutelulas humanas no folding de
proteiacutenas e nas modificaccedilotildees poacutes-traducionais como a glicosilaccedilatildeo aumentando a eficiecircncia
terapecircutica e diminuindo o potencial imunogeacutenico do produto final 23
As ceacutelulas de mamiacutefero em cultura agrave semelhanccedila de ceacutelulas transformadas possuem
um metabolismo energeacutetico desregulado consumindo elevadas quantidades de glucose e
acumulando quantidades significativas de lactato com reduzida entrada de carbono derivado
da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) 4 Do ponto de vista da cultura de
ceacutelulas para produccedilatildeo de biofaacutermacos este fenoacutetipo lactogeacutenico tem simultaneamente um
efeito negativo no crescimento e viabilidade celular e na produtividade e qualidade do
biofaacutermaco Este fenoacutemeno eacute ainda mais importante no caso das partiacuteculas derivadas de viacuterus
para terapia geacutenica jaacute que a sua infecciosidade eacute fortemente influenciada por variaccedilotildees no pH
do meio de cultura 5 Diversas estrateacutegias tecircm sido testadas de forma a reduzir os efeitos
nocivos da acumulaccedilatildeo de lactato no meio de cultura fornecimento de glucose de forma
controlada 67
suplementaccedilatildeo do meio de cultura com substratos alternativos agrave glucose 8
remoccedilatildeo dos metabolitos toacutexicos 9 desenvolvimento de meios de cultura com base na anaacutelise
dos fluxos metaboacutelicos permitindo adaptar os seus constituintes agraves necessidades metaboacutelicas
das ceacutelulas 1011
introduccedilatildeo da enzima piruvato carboxilase nas ceacutelulas 12-14
diminuiccedilatildeo da
expressatildeo da lactato desidrogenase atraveacutes da disrupccedilatildeo do gene Ldha por recombinaccedilatildeo
homoacuteloga 15
ou atraveacutes de RNA de interferecircncia 16
ou introduccedilatildeo de um transportador para a
frutose constituindo uma alternativa ao consumo de glucose 17
No entanto estas estrateacutegias
tecircm um sucesso limitado devido aos efeitos compensatoacuterios causados pela apertada regulaccedilatildeo
homeostaacutetica intracelular que natildeo satildeo facilmente alteraacuteveis
2
12 Imortalizaccedilatildeo celular e o efeito de Warburg
Nos anos 20 Otto Warburg observou que as ceacutelulas canceriacutegenas comparativamente
agraves ceacutelulas normais apresentavam elevadas taxas de consumo de glucose secretando a maior
parte do carbono derivado da glucose sob a forma de lactato em vez de o oxidar
completamente 1819
ndash um cenaacuterio que ficou conhecido como efeito de Warburg Estudos
recentes sobre o metabolismo energeacutetico de ceacutelulas canceriacutegenas sugerem que este resulta de
alteraccedilotildees geneacuteticas que ocorrem no processo de tumorigeacutenese principalmente durante a
imortalizaccedilatildeo e transformaccedilatildeo celulares e que levam agrave desregulaccedilatildeo do metabolismo
energeacutetico central e ao aumento da actividade glicoliacutetica 20
De facto existem diversas vias de
sinalizaccedilatildeo comuns ao controlo da proliferaccedilatildeo e do metabolismo energeacutetico que levam agrave
induccedilatildeo deste cenaacuterio como a activaccedilatildeo de oncogenes (MYC) e enzimas glicoliacuteticas ou a
supressatildeo do gene supressor de tumores P53 2021
Interessantemente muitas das vias de
imortalizaccedilatildeo activadas no processo de tumorigeacutenese satildeo tambeacutem recrutadas durante o
estabelecimento de uma linha celular e por isso tendem a exibir caracteriacutesticas metaboacutelicas
associadas ao efeito de Warburg 20
O principal efector molecular do efeito de Warburg eacute o factor 1 indutiacutevel por hipoxia
(HIF1) um factor de transcriccedilatildeo heterodimeacuterico constituiacutedo por uma subunidade β expressa
constitutivamente e uma subunidade α cuja expressatildeo estaacute dependente do oxigeacutenio Apesar
de ser expressa em condiccedilotildees de baixa disponibilidade de oxigeacutenio (hipoxia) a expressatildeo
constitutiva da subunidade α pode ocorrer em normoxia como resultado de alteraccedilotildees nos
sistemas de sinalizaccedilatildeo para degradaccedilatildeo proteica provocadas durante o processo de
imortalizaccedilatildeo celular De facto a sua sobre-expressatildeo eacute caracteriacutestica comum a muitas ceacutelulas
canceriacutegenas 2223
O factor de transcriccedilatildeo HIF1 eacute responsaacutevel pela coordenaccedilatildeo da alteraccedilatildeo
do metabolismo para glicoacutelise com formaccedilatildeo de lactato com consequente aumento do
consumo de glucose e acumulaccedilatildeo de lactato jaacute que promove a expressatildeo de vaacuterios
componentes do metabolismo energeacutetico nomeadamente transportadores de glucose (GLUT-
1) enzimas da via glicoliacutetica e de lactato desidrogenase (Figura 1) 2425
Para aleacutem do papel do HIF1 na promoccedilatildeo da glicoacutelise com formaccedilatildeo de lactato
estudos recentes revelaram uma significativa influecircncia na respiraccedilatildeo mitocondrial Nos
organismos aeroacutebios o TCA possui um papel fundamental no metabolismo central das
ceacutelulas constituindo a via de produccedilatildeo de energia mais eficiente e fornecendo o poder redutor
e intermediaacuterios necessaacuterios para as reacccedilotildees de biossiacutentese Normalmente a glucose eacute
convertida em piruvato no citoplasma o qual eacute posteriormente canalizado para o TCA ao
niacutevel mitocondrial onde eacute oxidado a dioacutexido de carbono e aacutegua A canalizaccedilatildeo do piruvato
3
para o TCA implica a sua conversatildeo em acetil-CoA numa reacccedilatildeo catalisada pelo complexo
multi-enzimaacutetico piruvato desidrogenase (PDH) Sendo esta uma das reacccedilotildees mais
importantes do metabolismo celular estaacute sujeita a um elevado niacutevel de regulaccedilatildeo Como tal o
complexo PDH eacute regulado por um ciclo reversiacutevel de fosforilaccedilatildeo caracterizado pela
fosforilaccedilatildeo (inactivaccedilatildeo) e desfosforilaccedilatildeo (activaccedilatildeo) da subunidade α do componente E1
(PDHE1α) promovida por cinases (PDK) e fosfatases (PDP) respectivamente Na PDHE1α
de mamiacuteferos existem trecircs locais de fosforilaccedilatildeo (Ser232
Ser293
e Ser300
) que podem ser
fosforilados por quatro isoenzimas (PDK1-4) sendo a PDK1 a uacutenica que consegue fosforilar
todos os trecircs locais 2627
Interessantemente a sobre-expressatildeo de PDK1 e PDK3 eacute promovida
por HIF1 levando assim agrave fosforilaccedilatildeo de PDHE1α e inibiccedilatildeo do complexo PDH (Figura 1)
Consequentemente a entrada do carbono derivado da via glicoliacutetica no TCA ocorre em menor
extensatildeo e aumenta a conversatildeo de piruvato a lactato resultando no fenoacutetipo lactogeacutenico
associado ao efeito de Warburg e na desregulaccedilatildeo do metabolismo energeacutetico 2428
Figura 1 Vias do metabolismo energeacutetico reguladas pelo factor de transcriccedilatildeo HIF1 A sobre-expressatildeo de
vaacuterias enzimas da via glicoliacutetica e da enzima lactato desidrogenase (LDH) eacute promovida pelo factor 1 indutiacutevel
por hipoxia (HIF1) levando ao aumento do consumo de glucose e agrave acumulaccedilatildeo de lactato A entrada do
carbono derivado da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) eacute realizada ao niacutevel do complexo
piruvato desidrogenase (PDH) sendo este regulado atraveacutes de um ciclo reversiacutevel de fosforilaccedilatildeo HIF1 promove
a expressatildeo das cinases da piruvato desidrogenase (PDK1-4) resultando na fosforilaccedilatildeo e consequente inibiccedilatildeo
do complexo PDH Esquema adaptado de Kondoh (2008) 20
Glucose
Glucose 6P
Fuctose 6P
Glyceraldehyde-3P
Fuctose 16 BP
Dihydroxyacetone-P
13 Biphosphoglycerate
3 - Phosphoglycerate
2 - Phosphoglycerate
Phosphoenopyruvate Pyruvate
Cytoplasm
Extracellular space
hexokinase
phosphoglucose isomerase
phosphofructokinase
GAPDH
phosphoglycerate
kinase
phosphoglucomutase
enolase
Pyruvate kinase
Pyruvate
Acetyl-CoATCA
PDH
PDH
P
LDH
Lactate
PDK
PDK2
PDK1 PDK3PDK4
aldolase
+
+
+
+
+
++
+
+
HIF1
4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

2
12 Imortalizaccedilatildeo celular e o efeito de Warburg
Nos anos 20 Otto Warburg observou que as ceacutelulas canceriacutegenas comparativamente
agraves ceacutelulas normais apresentavam elevadas taxas de consumo de glucose secretando a maior
parte do carbono derivado da glucose sob a forma de lactato em vez de o oxidar
completamente 1819
ndash um cenaacuterio que ficou conhecido como efeito de Warburg Estudos
recentes sobre o metabolismo energeacutetico de ceacutelulas canceriacutegenas sugerem que este resulta de
alteraccedilotildees geneacuteticas que ocorrem no processo de tumorigeacutenese principalmente durante a
imortalizaccedilatildeo e transformaccedilatildeo celulares e que levam agrave desregulaccedilatildeo do metabolismo
energeacutetico central e ao aumento da actividade glicoliacutetica 20
De facto existem diversas vias de
sinalizaccedilatildeo comuns ao controlo da proliferaccedilatildeo e do metabolismo energeacutetico que levam agrave
induccedilatildeo deste cenaacuterio como a activaccedilatildeo de oncogenes (MYC) e enzimas glicoliacuteticas ou a
supressatildeo do gene supressor de tumores P53 2021
Interessantemente muitas das vias de
imortalizaccedilatildeo activadas no processo de tumorigeacutenese satildeo tambeacutem recrutadas durante o
estabelecimento de uma linha celular e por isso tendem a exibir caracteriacutesticas metaboacutelicas
associadas ao efeito de Warburg 20
O principal efector molecular do efeito de Warburg eacute o factor 1 indutiacutevel por hipoxia
(HIF1) um factor de transcriccedilatildeo heterodimeacuterico constituiacutedo por uma subunidade β expressa
constitutivamente e uma subunidade α cuja expressatildeo estaacute dependente do oxigeacutenio Apesar
de ser expressa em condiccedilotildees de baixa disponibilidade de oxigeacutenio (hipoxia) a expressatildeo
constitutiva da subunidade α pode ocorrer em normoxia como resultado de alteraccedilotildees nos
sistemas de sinalizaccedilatildeo para degradaccedilatildeo proteica provocadas durante o processo de
imortalizaccedilatildeo celular De facto a sua sobre-expressatildeo eacute caracteriacutestica comum a muitas ceacutelulas
canceriacutegenas 2223
O factor de transcriccedilatildeo HIF1 eacute responsaacutevel pela coordenaccedilatildeo da alteraccedilatildeo
do metabolismo para glicoacutelise com formaccedilatildeo de lactato com consequente aumento do
consumo de glucose e acumulaccedilatildeo de lactato jaacute que promove a expressatildeo de vaacuterios
componentes do metabolismo energeacutetico nomeadamente transportadores de glucose (GLUT-
1) enzimas da via glicoliacutetica e de lactato desidrogenase (Figura 1) 2425
Para aleacutem do papel do HIF1 na promoccedilatildeo da glicoacutelise com formaccedilatildeo de lactato
estudos recentes revelaram uma significativa influecircncia na respiraccedilatildeo mitocondrial Nos
organismos aeroacutebios o TCA possui um papel fundamental no metabolismo central das
ceacutelulas constituindo a via de produccedilatildeo de energia mais eficiente e fornecendo o poder redutor
e intermediaacuterios necessaacuterios para as reacccedilotildees de biossiacutentese Normalmente a glucose eacute
convertida em piruvato no citoplasma o qual eacute posteriormente canalizado para o TCA ao
niacutevel mitocondrial onde eacute oxidado a dioacutexido de carbono e aacutegua A canalizaccedilatildeo do piruvato
3
para o TCA implica a sua conversatildeo em acetil-CoA numa reacccedilatildeo catalisada pelo complexo
multi-enzimaacutetico piruvato desidrogenase (PDH) Sendo esta uma das reacccedilotildees mais
importantes do metabolismo celular estaacute sujeita a um elevado niacutevel de regulaccedilatildeo Como tal o
complexo PDH eacute regulado por um ciclo reversiacutevel de fosforilaccedilatildeo caracterizado pela
fosforilaccedilatildeo (inactivaccedilatildeo) e desfosforilaccedilatildeo (activaccedilatildeo) da subunidade α do componente E1
(PDHE1α) promovida por cinases (PDK) e fosfatases (PDP) respectivamente Na PDHE1α
de mamiacuteferos existem trecircs locais de fosforilaccedilatildeo (Ser232
Ser293
e Ser300
) que podem ser
fosforilados por quatro isoenzimas (PDK1-4) sendo a PDK1 a uacutenica que consegue fosforilar
todos os trecircs locais 2627
Interessantemente a sobre-expressatildeo de PDK1 e PDK3 eacute promovida
por HIF1 levando assim agrave fosforilaccedilatildeo de PDHE1α e inibiccedilatildeo do complexo PDH (Figura 1)
Consequentemente a entrada do carbono derivado da via glicoliacutetica no TCA ocorre em menor
extensatildeo e aumenta a conversatildeo de piruvato a lactato resultando no fenoacutetipo lactogeacutenico
associado ao efeito de Warburg e na desregulaccedilatildeo do metabolismo energeacutetico 2428
Figura 1 Vias do metabolismo energeacutetico reguladas pelo factor de transcriccedilatildeo HIF1 A sobre-expressatildeo de
vaacuterias enzimas da via glicoliacutetica e da enzima lactato desidrogenase (LDH) eacute promovida pelo factor 1 indutiacutevel
por hipoxia (HIF1) levando ao aumento do consumo de glucose e agrave acumulaccedilatildeo de lactato A entrada do
carbono derivado da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) eacute realizada ao niacutevel do complexo
piruvato desidrogenase (PDH) sendo este regulado atraveacutes de um ciclo reversiacutevel de fosforilaccedilatildeo HIF1 promove
a expressatildeo das cinases da piruvato desidrogenase (PDK1-4) resultando na fosforilaccedilatildeo e consequente inibiccedilatildeo
do complexo PDH Esquema adaptado de Kondoh (2008) 20
Glucose
Glucose 6P
Fuctose 6P
Glyceraldehyde-3P
Fuctose 16 BP
Dihydroxyacetone-P
13 Biphosphoglycerate
3 - Phosphoglycerate
2 - Phosphoglycerate
Phosphoenopyruvate Pyruvate
Cytoplasm
Extracellular space
hexokinase
phosphoglucose isomerase
phosphofructokinase
GAPDH
phosphoglycerate
kinase
phosphoglucomutase
enolase
Pyruvate kinase
Pyruvate
Acetyl-CoATCA
PDH
PDH
P
LDH
Lactate
PDK
PDK2
PDK1 PDK3PDK4
aldolase
+
+
+
+
+
++
+
+
HIF1
4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

3
para o TCA implica a sua conversatildeo em acetil-CoA numa reacccedilatildeo catalisada pelo complexo
multi-enzimaacutetico piruvato desidrogenase (PDH) Sendo esta uma das reacccedilotildees mais
importantes do metabolismo celular estaacute sujeita a um elevado niacutevel de regulaccedilatildeo Como tal o
complexo PDH eacute regulado por um ciclo reversiacutevel de fosforilaccedilatildeo caracterizado pela
fosforilaccedilatildeo (inactivaccedilatildeo) e desfosforilaccedilatildeo (activaccedilatildeo) da subunidade α do componente E1
(PDHE1α) promovida por cinases (PDK) e fosfatases (PDP) respectivamente Na PDHE1α
de mamiacuteferos existem trecircs locais de fosforilaccedilatildeo (Ser232
Ser293
e Ser300
) que podem ser
fosforilados por quatro isoenzimas (PDK1-4) sendo a PDK1 a uacutenica que consegue fosforilar
todos os trecircs locais 2627
Interessantemente a sobre-expressatildeo de PDK1 e PDK3 eacute promovida
por HIF1 levando assim agrave fosforilaccedilatildeo de PDHE1α e inibiccedilatildeo do complexo PDH (Figura 1)
Consequentemente a entrada do carbono derivado da via glicoliacutetica no TCA ocorre em menor
extensatildeo e aumenta a conversatildeo de piruvato a lactato resultando no fenoacutetipo lactogeacutenico
associado ao efeito de Warburg e na desregulaccedilatildeo do metabolismo energeacutetico 2428
Figura 1 Vias do metabolismo energeacutetico reguladas pelo factor de transcriccedilatildeo HIF1 A sobre-expressatildeo de
vaacuterias enzimas da via glicoliacutetica e da enzima lactato desidrogenase (LDH) eacute promovida pelo factor 1 indutiacutevel
por hipoxia (HIF1) levando ao aumento do consumo de glucose e agrave acumulaccedilatildeo de lactato A entrada do
carbono derivado da glicoacutelise no Ciclo dos Aacutecidos Tricarboxiacutelicos (TCA) eacute realizada ao niacutevel do complexo
piruvato desidrogenase (PDH) sendo este regulado atraveacutes de um ciclo reversiacutevel de fosforilaccedilatildeo HIF1 promove
a expressatildeo das cinases da piruvato desidrogenase (PDK1-4) resultando na fosforilaccedilatildeo e consequente inibiccedilatildeo
do complexo PDH Esquema adaptado de Kondoh (2008) 20
Glucose
Glucose 6P
Fuctose 6P
Glyceraldehyde-3P
Fuctose 16 BP
Dihydroxyacetone-P
13 Biphosphoglycerate
3 - Phosphoglycerate
2 - Phosphoglycerate
Phosphoenopyruvate Pyruvate
Cytoplasm
Extracellular space
hexokinase
phosphoglucose isomerase
phosphofructokinase
GAPDH
phosphoglycerate
kinase
phosphoglucomutase
enolase
Pyruvate kinase
Pyruvate
Acetyl-CoATCA
PDH
PDH
P
LDH
Lactate
PDK
PDK2
PDK1 PDK3PDK4
aldolase
+
+
+
+
+
++
+
+
HIF1
4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

4
13 Impacto do efeito de Warburg em biotecnologia
A desregulaccedilatildeo do metabolismo energeacutetico em ceacutelulas de mamiacutefero em cultura agrave
semelhanccedila das ceacutelulas canceriacutegenas tem sido alvo de diversas hipoacuteteses explicativas A
glicoacutelise com formaccedilatildeo de lactato tira partido de um nutriente abundante a glucose pelo que
um elevado fluxo glicoliacutetico pode gerar energia mais rapidamente do que a respiraccedilatildeo
mitocondrial Como o fluxo glicoliacutetico pode superar o da oxidaccedilatildeo de piruvato pelo complexo
PDH eacute necessaacuterio um mecanismo para evitar a acumulaccedilatildeo de piruvato pelo que ocorre a
sobre-expressatildeo de lactato desidrogenase para a conversatildeo do piruvato a lactato 24
Adicionalmente ceacutelulas em contiacutenua proliferaccedilatildeo e com elevado consumo de nutrientes
teriam previsivelmente uma maior respiraccedilatildeo mitocondrial e consequente produccedilatildeo de
espeacutecies reactivas de oxigeacutenio (ERO) Como tal a diminuiccedilatildeo da respiraccedilatildeo mitocondrial
devido ao bloqueio do TCA ao niacutevel do complexo PDH pode estar associada a um aumento
da protecccedilatildeo ao stress oxidativo De facto a acumulaccedilatildeo de ERO e consequentes danos em
macromoleacuteculas a niacutevel intracelular foi envolvida no processo de senescecircncia celular 20
Como tal as alteraccedilotildees metaboacutelicas associadas ao efeito de Warburg poderatildeo natildeo soacute ter um
papel ao niacutevel de obtenccedilatildeo de energia mas tambeacutem de manutenccedilatildeo do estado proliferativo
por bypass da senescecircncia [revisto em Vander Heiden et al (2009) 29
] Esta adaptaccedilatildeo
metaboacutelica das ceacutelulas de mamiacutefero em cultura eacute no entanto desvantajosa do ponto de vista
da sua aplicaccedilatildeo biotecnoloacutegica na produccedilatildeo de biofaacutermacos complexos A acumulaccedilatildeo de
metabolitos toacutexicos como o lactato tem um efeito negativo no crescimento e viabilidade
celulares e na produtividade e qualidade do produto final Adicionalmente o bloqueio do
TCA reduz a disponibilidade de poder redutor e de intermediaacuterios necessaacuterios agraves reacccedilotildees de
biossiacutentese e indispensaacuteveis ao estabelecimento de sistemas de expressatildeo de elevada
produtividade e qualidade Como tal a manipulaccedilatildeo do efeito de Warburg em ceacutelulas de
mamiacutefero em cultura adquire grande relevacircncia no contexto biotecnoloacutegico
A linha celular RPE A6 eacute uma linha celular humana recombinante usada como sistema
de expressatildeo estaacutevel de anticorpos monoclonais humanos Para aleacutem dos referidos impactos
da acumulaccedilatildeo de lactato no crescimento e viabilidade celular a alteraccedilatildeo do pH poderaacute
induzir a agregaccedilatildeo proteica e um deficiente padratildeo de glicosilaccedilatildeo 30
diminuindo a
produtividade e qualidade do produto final e assim dificultando a sua aplicaccedilatildeo terapecircutica A
acumulaccedilatildeo de quantidades significativas de lactato tem igualmente um impacto negativo
sobre o crescimento de ceacutelulas MDCK E1 usadas na produccedilatildeo transiente de partiacuteculas
adenovirais e sobre a produtividade e qualidade dos vectores virais A linha celular 293
FLEX GFP eacute uma linha celular humana utilizada para a produccedilatildeo estaacutevel de vectores
5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

5
retrovirais recombinantes Estes vectores apresentam-se como uma ferramenta de elevada
eficiecircncia quer na transferecircncia de genes nos protocolos de terapia geacutenica quer na
apresentaccedilatildeo de epitopos em vacinaccedilatildeo 31
No entanto a sua produccedilatildeo em larga escala tem
sido dificultada pelas baixas produtividades de viacuterus infecciosos (na ordem das 106-10
7
partiacuteculas infecciosas por mL de cultura) perda acentuada da infecciosidade agrave temperatura de
cultura (t12 asymp 8h 37ordm C) e elevado grau de contaminaccedilatildeo das preparaccedilotildees virais com partiacuteculas
natildeo infecciosas (apenas 1 em cada 100 partiacuteculas eacute infecciosa) 3233
Como as aplicaccedilotildees em
terapia geacutenica humana dependendo do protocolo cliacutenico e da doenccedila requerem entre 107 e
1014
partiacuteculas infecciosas 34
com um elevado niacutevel de purificaccedilatildeo eacute imprescindiacutevel o
desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade para a
manufacturaccedilatildeo destas partiacuteculas virais Adicionalmente 30 a 40 da partiacutecula retroviral eacute
constituiacuteda por liacutepidos (viacuterus com invoacutelucro lipiacutedico) 35
Como a siacutentese lipiacutedica requer
grandes quantidades de poder redutor alteraccedilotildees a niacutevel da sua disponibilidade intracelular
teratildeo previsivelmente impacto na estabilidade do vector eou produtividade das ceacutelulas
produtoras
14 Objectivos principais
O presente trabalho teve como objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg procurando simultaneamente reduzir a acumulaccedilatildeo de lactato
e aumentar a eficiecircncia do metabolismo energeacutetico com o consequente aumento da
produtividade celular e qualidade do biofaacutermaco Para tal recorreu-se agrave tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais para o silenciamento de HIF1 PDK1 e
PDK3 os efectores do fenoacutetipo lactogeacutenico associado ao efeito de Warburg Estudos de
silenciamento de PDK1 36
e PDK3 28
utilizando a tecnologia de RNA de interferecircncia jaacute
demonstraram a viabilidade da reversatildeo do fenoacutetipo lactogeacutenico associado ao efeito de
Warburg sem que tal aparentemente afectasse o crescimento celular No entanto a
manipulaccedilatildeo efectuada neste trabalho ndash silenciamento sucessivo de HIF1 PDK1 e PDK3 ndash
nunca foi estudada
As linhas celulares 293 FLEX GFP MDCK E1 e RPE A6 foram escolhidas pela sua
importacircncia a niacutevel da produccedilatildeo de biofaacutermacos complexos tornando este projecto natildeo soacute
cientificamente atraente como tecnologicamente relevante
6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

6
2 MATERIAIS E MEacuteTODOS
21 Linhas celulares e meios de cultura
293 FLEX GFP eacute uma linha celular humana derivada de HEK293 (ATCC CRL-1573)
produtora de vectores retrovirais recombinantes baseados no viacuterus da leucemia murina
(Murine Leukemia Virus MLV) pseudotipadas com o invoacutelucro lipiacutedico do viacuterus da leucemia
do macaco gibatildeo (Gibbon Ape Leukemia Virus GaLV) e contendo um transgene codificante
para a proteiacutena fluorescente verde (Green Fluorescent Protein GFP) estabelecidas como
descrito em Coroadinha et al (2006) 37
RPE A6 eacute uma linha celular humana imortalizada
pela telomerase produtora de anticorpos monoclonais humanos por transfecccedilatildeo estaacutevel das
ceacutelulas hTERT RPE-1 (ATCC CRL-4000) MDCK E1 eacute uma linha celular canina derivada de
MDCK (ECCAC Nr 841211903) usada para produccedilatildeo de vectores adenovirais e estabelecida
como descrito em Santiago et al (em preparaccedilatildeo) 38
Estas linhas celulares foram usadas para
o estabelecimento de populaccedilotildees silenciadas respectivos clones e controlos atraveacutes de
transduccedilatildeo por infecccedilatildeo com vectores lentivirais A linha celular 293T (ATCC CRL-11268)
foi usada para produccedilatildeo de vectores lentivirais e a linha celular Te671 (ATCC CCL-136) para
titulaccedilatildeo de partiacuteculas retrovirais infecciosas Todas as linhas celulares foram mantidas em
meio de cultura DMEM (Gibco Paisley Reino Unido) suplementado com 10 (vv) de
Foetal Bovine Serum (FBS) (Gibco) a 37ordmC e atmosfera huacutemida com 5 de CO2
22 Produccedilatildeo de plasmiacutedeos e vectores lentivirais
A construccedilatildeo short hairpin RNA (shRNA) para o silenciamento de HIF1
TRCN000000309 e o controlo non-target (ntshRNA) (Anexo 1) satildeo baseados no plasmiacutedeo
pLKO1-puro 39
e contecircm um gene de resistecircncia agrave puromicina tendo sido adquiridos agrave
bdquoSigma Missionreg RNAi‟ (Sigma St Louis MO EUA) Foram produzidos vectores lentivirais
para cada um dos shRNA em sistema transiente por co-transfecccedilatildeo com pSPAX2 e pMD2G
[gentilmente cedidos por Didier Trono do Swiss Federal Institute of Technology atraveacutes do
repositoacuterio de plasmiacutedeos da Addgene (Cambridge MA EUA)] que fornecem as funccedilotildees de
empacotamento e invoacutelucro lipiacutedico respectivamente Para o silenciamento de PDK1 e PDK3
e correspondente controlo ntshRNA foram adquiridos agrave bdquoSigma Missionreg RNAi‟ vectores
lentivirais custom-made semelhantes aos anteriores mas contendo um gene que codifica a
proteiacutena fluorescente vermelha (Red Fluorescent Protein RFP) As sequecircncias usadas natildeo se
7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

7
encontram validadas tendo sido desenhadas simultaneamente para uma zona de homologia
entre os genes PDK1 e PDK3 e entre os genomas humano e canino (Anexo 1)
23 Transduccedilatildeo celular selecccedilatildeo e clonagem
Para o estabelecimento de populaccedilotildees shRNA (shHIF1 e ntshRNA) as ceacutelulas 293
FLEX GFP MDCK E1 e RPE A6 foram inoculadas em placas de 6 poccedilos a uma densidade de
1x105 ceacutelulascm
2 24 horas depois o meio de cultura foi substituiacutedo por 600 microl de
sobrenadante lentiviral preparado em meio de cultura contendo 80 μgmL de polibreno
(Sigma Steinheim Alemanha) e diluiacutedo de forma a atingir uma baixa multiplicidade de
infecccedilatildeo (01) 48 horas apoacutes a infecccedilatildeo as ceacutelulas foram colocadas sob pressatildeo selectiva do
antibioacutetico puromicina a 15 μgmL (MDCK E1 e 293 FLEX GFP) ou 30 microgmL (RPE A6) e
mantidas em cultura durante 21 dias substituindo-se o meio de cultura a cada 3 dias As
populaccedilotildees shHIF1 foram posteriormente clonadas em placas de 96 poccedilos inoculadas a uma
densidade de 05 ceacutelulaspoccedilo tendo-se isolado cerca de 30 clones para cada uma delas Para
o silenciamento de PDK1 e PDK3 os clones shHIF1 escolhidos para continuar o estudo
foram transduzidos como acima descrito usando uma multiplicidade de infecccedilatildeo superior
`anteriormente usada (lt1) As populaccedilotildees ntshRNA foram igualmente transduzidas com um
segundo vector ntshRNA (Anexo 1) A selecccedilatildeo de ceacutelulas duplamente transduzidas foi
efectuada por citometria de fluxo acoplada a sorteamento no sistema bdquoMoFlo High-Speed Cell
Sorter‟ (Dako Cytomation Glostrup Dinamarca) tirando partido da fluorescecircncia da RFP As
ceacutelulas sorteadas foram mantidas posteriormente em cultura sob pressatildeo selectiva do
antibioacutetico puromicina
24 Estudos de crescimento e anaacutelise de metabolitos
Inocularam-se tissue-flasks de 25 cm2 a uma densidade de 1x10
4 ceacutelulascm
2 (MDCK
E1) e de 4x104 ceacutelulascm
2 (293 FLEX GFP) em meio DMEM (Gibco) suplementado com
10 (vv) FBS (Gibco) Para os estudos com ceacutelulas duplamente transduzidas usou-se
adicionalmente um suplemento antioxidante a 01 (vv) (Sigma) Durante 7 dias ao longo
da curva de crescimento foram sendo retiradas amostras de sobrenadante que foram
clarificadas por filtraccedilatildeo a 045 microm e guardadas a -85ordmC ateacute posterior anaacutelise A concentraccedilatildeo
e viabilidade celulares foram determinadas pelo meacutetodo de exclusatildeo do azul de tripano
(Gibco) As concentraccedilotildees de glucose e lactato foram determinadas usando o sistema bdquoYSI
7100 Multiparameter Bioanalytical System‟ (YSI Life Sciences Yellow Springs OH EUA)
8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

8
25 Extracccedilatildeo de RNA total e qRT-PCR
Para quantificar os niacuteveis de expressatildeo geacutenica o RNA total celular foi isolado usando
o bdquoRNeasy Mini Kit‟ (Qiagen Valencia CA EUA) de acordo com as instruccedilotildees do
fabricante A transcriccedilatildeo reversa do RNA total foi efectuada segundo o protocolo de siacutentese
de cDNA do bdquoTranscriptor High Fidelity cDNA Synthesis Kit‟ (Roche Applied Science
Manheim Alemanha) usando 2 microg de RNA total e primers oligo dT O qRT-PCR foi
realizado em triplicados no termociclador bdquoLightCyclerreg 480 Real-Time PCR System‟ (Roche
Applied Science) segundo as instruccedilotildees do kit bdquoLightCyclerreg 480 SYBR Green I Master‟
(Roche Applied Science) O sinal de PCR foi definido segundo o Meacutetodo da Segunda
Derivada Maacutexima 40
e a expressatildeo geacutenica foi quantificada relativamente agrave expressatildeo do gene
housekeeping RPL22 segundo o meacutetodo 2-ΔCt
41
As sequecircncias dos primers usados neste
estudo (Anexo 2) foram desenhadas para locais de homologia entre ceacutelulas humanas e caninas
recorrendo aos softwares BLAST 42
e Primer3 43
26 Produccedilatildeo e titulaccedilatildeo de vectores retrovirais
Para a titulaccedilatildeo de partiacuteculas retrovirais infecciosas ceacutelulas Te671 foram inoculadas a
uma densidade de 5x104 ceacutelulascm
2 em placas de 24 poccedilos Apoacutes 24 horas o meio foi
removido e as ceacutelulas foram infectadas em duplicados com 150 microl de sobrenadante viral
(recolhido durante os estudos de crescimento) previamente sujeito a diluiccedilotildees em seacuterie em
meio de cultura contendo 80 microgmL de polibreno (Sigma) 48 horas apoacutes a infecccedilatildeo a
quantidade de ceacutelulas fluorescentes foi determinada por citometria de fluxo (CyFlow Space
Partec Alemanha) e o tiacutetulo viral calculado de acordo com a Equaccedilatildeo 1
(Equaccedilatildeo 1)
Para a titulaccedilatildeo de partiacuteculas retrovirais totais seguiu-se o protocolo descrito em
Carmo et al (2004) 44
com as seguintes modificaccedilotildees bdquoTranscriptor High Fidelity cDNA
Synthesis Kit‟ (Roche Applied Science) para a siacutentese de cDNA e primer reverso para a GFP
(Anexo 2) Para o qRT-PCR o mesmo sistema acima descrito foi usado utilizando o par de
primers para a GFP Diluiccedilotildees sucessivas do plasmiacutedeo pGFP foram usadas para a construccedilatildeo
da curva-padratildeo
27 Produccedilatildeo e titulaccedilatildeo de vectores adenovirais
Para a produccedilatildeo e titulaccedilatildeo de vectores adenovirais seguiram-se os protocolos
descritos em Altaras et al (2005) 45
e Kamen amp Henry (2004) 46
Resumidamente aquando
9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

9
dos estudos de crescimento ceacutelulas MDCK E1 com confluecircncia de 80 foram transduzidas
com um stock de Canine adenoviacuterus type 2 (CAV-2) contendo o transgene da GFP a uma
multiplicidade de infecccedilatildeo de 5 em meio suplementado com 1 (vv) de aminoaacutecidos natildeo
essenciais (Sigma) Apoacutes 40 horas foram recolhidas as fracccedilotildees extra (recolha do
sobrenadante) e intracelular [lise celular provocada por uma soluccedilatildeo de Triton X-100 a 01
(vv) em Tris-HCl 10 mM pH 8] dos vectores adenovirais produzidos procedendo-se agrave sua
clarificaccedilatildeo por centrifugaccedilatildeo (10 minutos 3000 g) Para a titulaccedilatildeo de partiacuteculas infecciosas
foram inoculadas ceacutelulas MDCK E1 a uma densidade de 2x104 ceacutelulascm
2 em placas de 24
poccedilos e 24 horas depois o meio de cultura foi substituiacutedo por diluiccedilotildees sucessivas dos viacuterus
recolhidos em duplicados A percentagem de ceacutelulas fluorescentes foi determinada 24 horas
apoacutes a infecccedilatildeo Para a titulaccedilatildeo de partiacuteculas adenovirais totais o DNA viral foi extraiacutedo
usando o bdquoRoche High Pure Viral DNA Kit‟ (Roche Applied Science) O qPCR foi realizado
como descrito para a titulaccedilatildeo de partiacuteculas retrovirais totais A produtividade especiacutefica de
partiacuteculas totais e infecciosas foi calculada tendo em conta o nuacutemero de partiacuteculas produzidas
e o nuacutemero de ceacutelulas no periacuteodo de produccedilatildeo
28 Determinaccedilatildeo da actividade da lactato desidrogenase
Para a determinaccedilatildeo da actividade da enzima lactato desidrogenase extractos celulares
a uma densidade de 2x106 ceacutelulasmL recolhidas ao longo das curvas de crescimento foram
preparados previamente em tampatildeo bdquoM-PER Mammalian Protein Extraction Reagent‟ (Pierce
Rockford IL EUA) de acordo com as instruccedilotildees do fabricante A actividade intracelular
enzimaacutetica foi determinada atraveacutes da monitorizaccedilatildeo espectrofotomeacutetrica a 340 nm da taxa de
oxidaccedilatildeo de NADH a NAD+ acoplada agrave reduccedilatildeo de piruvato a lactato
29 Western Blot
A expressatildeo proteica foi detectada por Western Blotting apoacutes separaccedilatildeo em gel de
poliacrilamida NuPage a 4-12 (wv) (Invitrogen Paisley Reino Unido) com running buffer
MOPS (conc 1x Invitrogen) das amostras desnaturadas a 72ordm C durante 10 minutos Cada
poccedilo foi carregado com igual quantidade de proteiacutena e as amostras correram durante 90
minutos a voltagem constante de 125 V tendo sido posteriormente transferidas durante 90
minutos para membrana de fluoreto de polivinilideno (PVDF) Estas foram reveladas com os
anticorpos listados no Anexo 3 detectadas com os respectivos anticorpos secundaacuterios
conjugados com a peroxidase de raacutebano (HRP) (diluiccedilatildeo 15000) e reveladas com bdquoAmersham
ECL Plustrade Western Blotting Detection Reagents‟ (GE Healthcare Piscataway NJ EUA)
10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

10
210 Quantificaccedilatildeo de fosfatos de adenosina e nucleoacutetidos de piridina
As razotildees ADPATP foram determinadas atraveacutes do bdquoEnzyLighttrade
ADPATP Ratio
Assay Kit‟ (BioAssay Systems Hayward CA EUA) de acordo com as instruccedilotildees do
fabricante As razotildees NAD+NADH e NADP
+NADPH foram determinadas recorrendo ao
bdquoEnzyChromtrade NADNADH Assay Kit‟ (BioAssay Systems) e bdquoEnzyChromtrade
NADP+NADPH Assay Kit‟ (BioAssay Systems) Foram seguidas as instruccedilotildees do fabricante
211 Quantificaccedilatildeo de espeacutecies reactivas de oxigeacutenio
Para determinar a carga intracelular de espeacutecies reactivas de oxigeacutenio (ERO)
quantificou-se a velocidade de formaccedilatildeo de um produto fluorescente resultante da oxidaccedilatildeo
da sonda H2DCFDA (Invitrogen) Para tal placas de 96 poccedilos foram inoculadas agrave densidade
celular acima descrita para os estudos de crescimento 48 horas depois adicionou-se uma
soluccedilatildeo de Saponina (Sigma) a uma concentraccedilatildeo final de 004 mgmL e incubou-se a 37ordmC
Apoacutes 30 minutos adicionou-se a sonda a uma concentraccedilatildeo final de 10 microM A formaccedilatildeo de
fluorescecircncia foi seguida durante 250 minutos a 485528 nm (emissatildeoexcitaccedilatildeo) A
velocidade de formaccedilatildeo de fluorescecircncia uma medida directa da quantidade de ERO foi
calculada por regressatildeo linear pelo meacutetodo de ajuste dos miacutenimos quadrados da curva de
fluorescecircncia vs tempo
212 Coloraccedilatildeo para detecccedilatildeo de actividade da β-galactosidade
Para averiguar a expressatildeo da enzima β-galactosidade inocularam-se placas de 6
poccedilos com as densidades acima descritas para os estudos de crescimento A meio da fase
exponencial da curva de crescimento as ceacutelulas foram fixadas e coradas como descrito em
Sambrook amp Russell (2001) 47
213 Caacutelculos
Para determinar as taxas especiacuteficas de crescimento celular produccedilatildeoacumulaccedilatildeo de
metabolitos e de partiacuteculas virais foi considerada a equaccedilatildeo de Boltzmann (Equaccedilatildeo 2)
(Equaccedilatildeo 2)
onde P representa os diferentes paracircmetros quer seja concentraccedilatildeo de ceacutelulas viaacuteveis
concentraccedilatildeo de metabolitos PI ou PT C representa a concentraccedilatildeo de ceacutelulas viaacuteveis t
representa o tempo de cultura e microp eacute a taxa especiacutefica celular de cada paracircmetro em estudo O
intervalo de confianccedila considerado foi de 95
11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

11
3 RESULTADOS
31 Silenciamento de HIF1
No presente trabalho foi usado o sistema bdquoSigma Missionreg RNAi‟ ndash construccedilatildeo
humana ndash para o silenciamento de genes em linhas celulares humanas e caninas A eficiecircncia
de silenciamento do sistema em ceacutelulas caninas foi previamente testada e confirmada com um
gene repoacuterter (turboGFP) (dados natildeo apresentados)
Para a primeira fase de manipulaccedilatildeo do efeito de Warburg as linhas celulares 293
FLEX GFP MDCK E1 e RPE A6 foram infectadas com vectores lentivirais transportando um
hairpin de silenciamento de HIF1 seleccionadas e clonadas por diluiccedilatildeo limitante Foram
isolados cerca de 30 clones por linha celular
O silenciamento de HIF1 induziu diferentes respostas nas linhas celulares em estudo
nos clones MDCK E1 e 293 FLEX GFP natildeo foram notadas alteraccedilotildees macroscoacutepicas
contrariamente a maioria dos clones RPE A6 apresentou alteraccedilotildees severas a niacutevel
morfoloacutegico (aumento das dimensotildees celulares e da razatildeo citoplasmanuacutecleo) (Figura 2A-C) e
do crescimento celular Alguns destes clones perderam totalmente a capacidade proliferativa
Estas alteraccedilotildees sugeriram um cenaacuterio de senescecircncia induzida pelo silenciamento de HIF1 48
Deste modo procedeu-se agrave detecccedilatildeo de actividade da enzima β-galactosidade descrita como
um biomarcador de ceacutelulas em senescecircncia 49
(Figura 2D-F) Na linha celular parental natildeo se
observou coloraccedilatildeo indicativa da actividade da β-galactosidase enquanto nos clones
silenciados para HIF1 se obteve uma elevada coloraccedilatildeo Estes resultados sugeriram que na
linha celular RPE A6 o silenciamento de HIF1 poderaacute ter induzido a entrada das ceacutelulas num
estado de senescecircncia Como tal natildeo se prosseguiu os estudos com esta linha celular
Para a continuaccedilatildeo dos estudos de caracterizaccedilatildeo do efeito do silenciamento de HIF1
foram escolhidos os clones FLEX shHIF121 FLEX shHIF125 MDCK shHIF115 e
MDCK shHIF116 com base numa anaacutelise integrada da reduccedilatildeo dos niacuteveis de lactato
desidrogenase quantificaccedilatildeo da expressatildeo de HIF1 e screening preliminar do tiacutetulo viral
(dados natildeo apresentados) Para estes clones analisaram-se as curvas de crescimento e o
metabolismo energeacutetico central (Figura 3 e Tabela 1) os niacuteveis de lactato desidrogenase
(Figura 4) e efeitos do silenciamento de HIF1 na expressatildeo geacutenica de PDHE1α LDH (A e B)
e PDK (1 e 3) (Figura 5)
12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

12
Linha celular parental Clones silenciados
Figura 2 Impacto do silenciamento de HIF1 em ceacutelulas RPE A6 Alteraccedilotildees morfoloacutegicas observadas na
linha celular parental (A) e nos clones RPE shHIF118 (B) e RPE shHIF122 (C) Detecccedilatildeo de actividade de β-
galactosidade um biomarcador de senescecircncia em RPE A6 (D) RPE shHIF13 (E) e RPE shHIF124 (F) A
barra da escala corresponde a 300 microm
Os clones silenciados para HIF1 analisados apresentaram perfis bem como taxas
especiacuteficas de crescimento idecircnticas agraves respectivas linhas celulares parentais Em termos
metaboacutelicos tambeacutem natildeo ocorreram alteraccedilotildees consideraacuteveis nas taxas de consumo de
glucose e de produccedilatildeo de lactato (Figura 3 e Tabela 1)
Figura 3 Curvas de crescimento celular Ceacutelulas 293 FLEX GFP (A) e de MDCK E1 (B) a cada ponto da
curva de crescimento estaacute associado um erro de 10 decorrente do meacutetodo de contagem celular
00
05
10
15
20
25
30
35
40
45
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
02
04
06
08
10
12
14
16
18
20
0 24 48 72 96 120 144 168
Den
sid
ad
ecelu
lar
(10
6ceacutelu
las
mL
)
Tempo de cultura (h)
MDCK E1 MDCK shHIF115 MDCK shHIF116
A B C
D E F
A B
13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

13
Tabela 1 Taxas especiacuteficas de crescimento celular e metabolismo energeacutetico central
μcrescimento
(h-1
)
γglucose
(micromol106ceacutelulash)
γlactato
(micromol106ceacutelulash)
293 FLEX GFP 0019 plusmn 0002 012 plusmn 002 019 plusmn 003 16 plusmn 04
FLEX shHIF121 0018 plusmn 0002 013 plusmn 001 022 plusmn 002 17 plusmn 03
FLEX shHIF125 0021 plusmn 0001 011 plusmn 001 017 plusmn 003 15 plusmn 04
MDCK E1 0026 plusmn 0005 030 plusmn 008 055 plusmn 011 18 plusmn 09
MDCK shHIF115 0028 plusmn 0004 036 plusmn 005 065 plusmn 004 18 plusmn 04
MDCK shHIF116 0026 plusmn 0005 039 plusmn 006 073 plusmn 005 19 plusmn 04
Os erros na μcrescimento γglucose e γlactato correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados os erros no γlactatoγglucose correspondem agrave propagaccedilatildeo de erro (μcrescimento) taxa especiacutefica de
crescimento (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de lactato
De forma a averiguar se o silenciamento de HIF1 induziu alteraccedilotildees nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 4) Nos
clones FLEX natildeo se observaram alteraccedilotildees relevantes Por outro lado nos clones MDCK
observaram-se reduccedilotildees de actividade entre os 30 e 40
Figura 4 Actividade intracelular da enzima lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas 293
FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
Em termos de expressatildeo geacutenica (Figura 5) os clones FLEX shHIF121 e FLEX
shHIF125 apresentaram uma diminuiccedilatildeo da expressatildeo de HIF1 de 628 e 500
respectivamente Para os restantes genes analisados natildeo se observaram alteraccedilotildees
significativas na sua expressatildeo Nos clones MDCK shHIF115 e MDCK shHIF116
observou-se uma diminuiccedilatildeo da expressatildeo de HIF1 (191 e 522 respectivamente) de
00
02
04
06
08
10
12
293 FLEX GFP
FLEX shHIF121
FLEX shHIF125
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
00
02
04
06
08
10
12
MDCK E1 MDCK shHIF115
MDCK shHIF116
Act
ivid
ad
e n
orm
ali
zad
a d
a
lact
ato
des
idro
gen
ase
A B
14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

14
LDHA (385 e 176 respectivamente) e de PDK1 (640 e 544 respectivamente) A
expressatildeo geacutenica de LDHA observada para as ceacutelulas 293 FLEX GFP e MDCK E1 estaacute de
acordo com a quantificaccedilatildeo de actividade intracelular da enzima (Figura 4)
Figura 5 Expressatildeo geacutenica apoacutes silenciamento de HIF1 Ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a
quantificaccedilatildeo da expressatildeo foi feita por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada
para as respectivas linhas celulares parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados (HIF1) factor 1 indutiacutevel por
hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato desidrogenase (LDHA e LDHB)
lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato desidrogenase
Um dos principais objectivos do presente estudo foi avaliar o impacto da manipulaccedilatildeo
do efeito de Warburg na produtividade celular Neste sentido quantificou-se a produccedilatildeo de
partiacuteculas virais totais e infecciosas (Figura 6) O silenciamento de HIF1 ainda que moderado
(maacuteximo de 628 de acordo com a Figura 5) resultou em alteraccedilotildees notaacuteveis na
produtividade das ceacutelulas 293 FLEX GFP com especial destaque para o clone FLEX
shHIF121 em que a taxa de produccedilatildeo de partiacuteculas totais aumentou 68x e a taxa de
partiacuteculas infecciosas aumentou 212x comparativamente agrave linha celular parental Tambeacutem
nos clones MDCK silenciados para HIF1 se observou um aumento da produtividade viral
ainda que em menor extensatildeo particularmente o clone MDCK shHIF115 em que a
produtividade especiacutefica de partiacuteculas totais aumentou 15x e a de partiacuteculas infecciosas
aumentou 20x A anaacutelise das razotildees PIPT (Tabela 2) demonstrou ainda que para ambas as
linhas celulares o silenciamento de HIF1 resultou numa produccedilatildeo viral de qualidade superior
isto eacute com um maior nuacutemero de partiacuteculas infecciosas por nuacutemero de partiacuteculas totais
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a
no
rma
liza
da
293 FLEX GFP FLEX shHIF121 FLEX shHIF125A B
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100
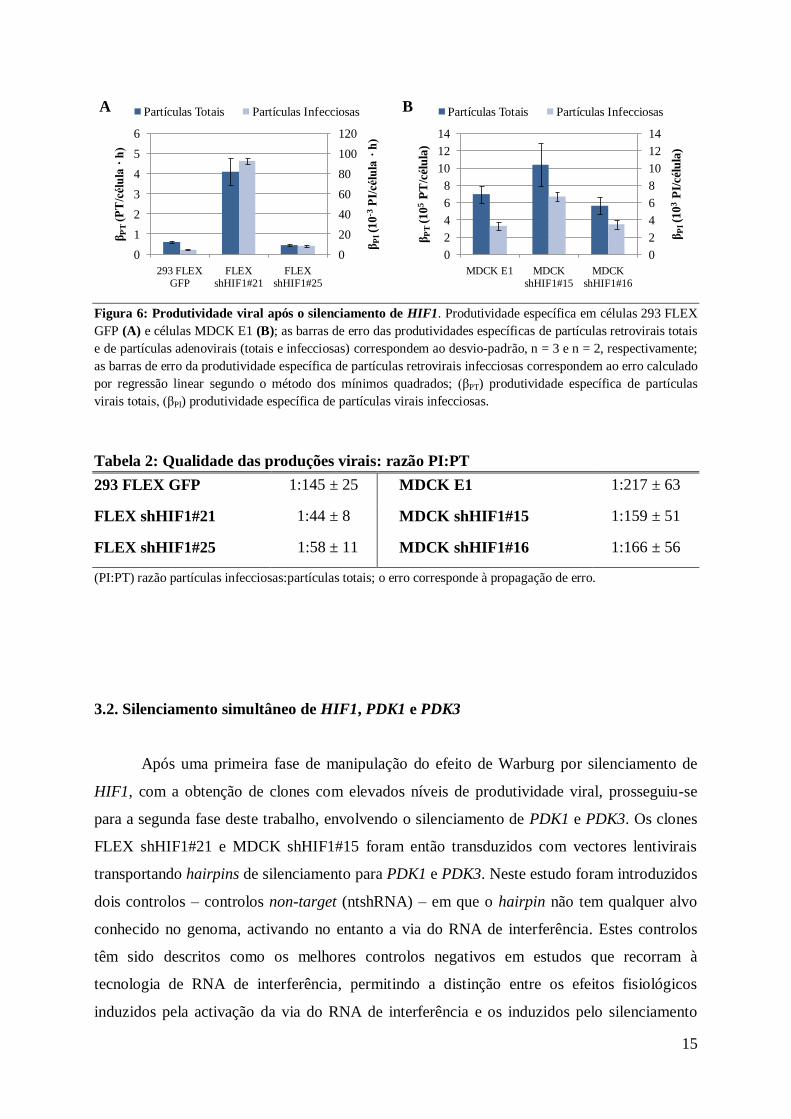
15
Figura 6 Produtividade viral apoacutes o silenciamento de HIF1 Produtividade especiacutefica em ceacutelulas 293 FLEX
GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais
e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n = 3 e n = 2 respectivamente
as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas correspondem ao erro calculado
por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT) produtividade especiacutefica de partiacuteculas
virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
Tabela 2 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1145 plusmn 25 MDCK E1 1217 plusmn 63
FLEX shHIF121 144 plusmn 8 MDCK shHIF115 1159 plusmn 51
FLEX shHIF125 158 plusmn 11 MDCK shHIF116 1166 plusmn 56
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
32 Silenciamento simultacircneo de HIF1 PDK1 e PDK3
Apoacutes uma primeira fase de manipulaccedilatildeo do efeito de Warburg por silenciamento de
HIF1 com a obtenccedilatildeo de clones com elevados niacuteveis de produtividade viral prosseguiu-se
para a segunda fase deste trabalho envolvendo o silenciamento de PDK1 e PDK3 Os clones
FLEX shHIF121 e MDCK shHIF115 foram entatildeo transduzidos com vectores lentivirais
transportando hairpins de silenciamento para PDK1 e PDK3 Neste estudo foram introduzidos
dois controlos ndash controlos non-target (ntshRNA) ndash em que o hairpin natildeo tem qualquer alvo
conhecido no genoma activando no entanto a via do RNA de interferecircncia Estes controlos
tecircm sido descritos como os melhores controlos negativos em estudos que recorram agrave
tecnologia de RNA de interferecircncia permitindo a distinccedilatildeo entre os efeitos fisioloacutegicos
induzidos pela activaccedilatildeo da via do RNA de interferecircncia e os induzidos pelo silenciamento
0
20
40
60
80
100
120
0
1
2
3
4
5
6
293 FLEX
GFP
FLEX
shHIF121
FLEX
shHIF125
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14
MDCK E1 MDCK
shHIF115
MDCK
shHIF116
βP
I(1
03
PI
ceacutelu
la)
βP
T(1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

16
geacutenico 5051
Ceacutelulas 293 FLEX GFP e MDCK E1 foram transduzidas com um hairpin non-
target (populaccedilotildees 1xntshRNA) e apoacutes selecccedilatildeo estas populaccedilotildees foram novamente
transduzidas com outro hairpin non-target (populaccedilotildees 2xntshRNA) As populaccedilotildees
duplamente transduzidas foram seleccionadas por citometria de fluxo acoplada a sorteamento
celular Parte destas populaccedilotildees foram clonadas directamente por sorteamento e outra parte
foi mantida como populaccedilatildeo Por questotildees de limitaccedilatildeo de tempo o trabalho descrito nesta
dissertaccedilatildeo engloba apenas os resultados relativos agraves populaccedilotildees
Avaliou-se o impacto da activaccedilatildeo da via do RNA de interferecircncia (populaccedilotildees
ntshRNA) do silenciamento de HIF1 e do silenciamento duplo HIF1+PDK13 no
crescimento celular natildeo tendo sido observadas diferenccedilas significativas (Tabela 3)
Metabolicamente as ceacutelulas FLEX shHIF121 e as duplamente silenciadas apresentaram
reduccedilotildees evidentes das taxas de consumo de glucose e de produccedilatildeo de lactato sem no
entanto existir alteraccedilatildeo nas razotildees lactatoglucose (Figura 7 e Tabela 4) Nas populaccedilotildees
FLEX ntshRNA em particular a populaccedilatildeo duplamente transduzida foram observadas
alteraccedilotildees na morfologia celular e visiacutevel aumento na concentraccedilatildeo de lactato no meio de
cultura observaccedilotildees confirmadas pela obtenccedilatildeo de maiores taxas de produccedilatildeo de lactato As
ceacutelulas MDCK silenciadas pelo contraacuterio apresentaram um ligeiro aumento nas taxas de
consumo de glucose e de produccedilatildeo de lactato ainda que sem alteraccedilatildeo na razatildeo
lactatoglucose
Tabela 3 Taxas de crescimento celular
μcrescimento (h-1
) μcrescimento (h-1
)
293 FLEX GFP 0021 plusmn 0002 MDCK E1 0033 plusmn 0003
FLEX 1xntshRNA 0021 plusmn 0002 MDCK 1xntshRNA 0029 plusmn 0002
FLEX 2xntshRNA 0027 plusmn 00004 MDCK 2xntshRNA 0037 plusmn 0003
FLEX shHIF121 0022 plusmn 0001 MDCK shHIF115 0031 plusmn 0003
FLEX shHIF121
+shPDK13 0024 plusmn 0001
MDCK shHIF115
+shPDK13 0037 plusmn 0005
Os erros correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
(μcrescimento) taxa de crescimento celular
17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

17
Figura 7 Impacto nas taxas de consumo de glucose e de produccedilatildeo de lactato Ceacutelulas 293 FLEX GFP (A) e
MDCK E1 (B) as barras de erro correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos
miacutenimos quadrados (γmetaboacutelica) taxa de consumo de glucose ou taxa de produccedilatildeo de lactato conforme o caso
Tabela 4 Metabolismo energeacutetico central razotildees γlactatoγglucose
293 FLEX GFP 16 plusmn 03 MDCK E1 15 plusmn 03
FLEX 1xntshRNA 17 plusmn 03 MDCK 1xntshRNA 15 plusmn 05
FLEX 2xntshRNA 17 plusmn 03 MDCK 2xntshRNA 16 plusmn 02
FLEX shHIF121 16 plusmn 01 MDCK shHIF115 15 plusmn 03
FLEX shHIF121+shPDK13 15 plusmn 01 MDCK shHIF115+shPDK13 15 plusmn 02
Os erros correspondem agrave propagaccedilatildeo de erro (γglucose) taxa de consumo de glucose (γlactato) taxa de produccedilatildeo de
lactato
De forma a averiguar o impacto do silenciamento de PDK1 e PDK3 nos niacuteveis de
lactato desidrogenase quantificou-se a actividade intracelular da enzima (Figura 8)
Verificou-se uma reduccedilatildeo de actividade total da enzima lactato desidrogenase nas ceacutelulas 293
FLEX GFP silenciadas No entanto nas ceacutelulas MDCK E1 natildeo foram observadas diferenccedilas
na actividade enzimaacutetica
Figura 8 Impacto na actividade intracelular da lactato desidrogenase Actividade enzimaacutetica nas ceacutelulas
293 FLEX GFP (A) e MDCK E1 (B) normalizada para as respectivas linhas celulares parentais barras de erro
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados
00
01
02
03
04
05
06
07
08
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
14
16
18
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
γm
eta
boacute
lica
(microm
ol
10
6ceacute
lula
s ∙
h)
Consumo de glucose Produccedilatildeo de lactato
00
02
04
06
08
10
12
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp64h Tempo de cultura asymp118h
00
02
04
06
08
10
12
14
16
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+siPDIK13
Act
ivid
ad
e n
orm
aliza
da d
a
lact
ato
des
idro
gen
ase
Tempo de cultura asymp68h Tempo de cultura asymp120h
A B
A B
18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

18
Nesta segunda fase quantificaram-se igualmente os niacuteveis de expressatildeo geacutenica de
HIF1 PDHE1α LDH (A e B) e PDK (1 e 3) (Figura 9A-B) Contrariamente aos resultados
anteriormente obtidos (Figura 5) o clone FLEX shHIF121 revelou alteraccedilotildees para aleacutem das
observadas no HIF1 nomeadamente a diminuiccedilatildeo da expressatildeo dos genes da LDHA (reduccedilatildeo
de 308) e PDK1 (reduccedilatildeo de 289) relativamente agrave linha celular parental em linha com o
que acontecera anteriormente nos clones MDCK silenciados para HIF1 Na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa da expressatildeo de PDK1
(reduccedilatildeo de 630) e de PDK3 (reduccedilatildeo de 427) relativamente agrave linha celular parental No
clone MDCK shHIF115 os resultados anteriormente obtidos foram validados na populaccedilatildeo
duplamente silenciada observou-se uma diminuiccedilatildeo significativa de PDK1 (reduccedilatildeo de
841) e de PDK3 (reduccedilatildeo de 218) relativamente agrave linha celular parental A expressatildeo
geacutenica nos controlos ntshRNA foi idecircntica agrave das respectivas linhas celulares parentais
excepccedilatildeo feita agrave quantificaccedilatildeo de HIF1 na populaccedilatildeo MDCK 1xntshRNA que poderaacute dever-
se a um erro de execuccedilatildeo da teacutecnica jaacute que tal resultado natildeo eacute reprodutiacutevel na populaccedilatildeo
MDCK 2xntshRNA
Para aleacutem das alteraccedilotildees em termos de expressatildeo geacutenica analisaram-se os efeitos na
quantidade intracelular das respectivas proteiacutenas (Figura 9C) Natildeo foi possiacutevel detectar
diferenccedilas nos niacuteveis proteicos de HIF1 possivelmente porque a teacutecnica natildeo tem
sensibilidade suficiente para detectar pequenas variaccedilotildees de expressatildeo num factor de
transcriccedilatildeo que per se eacute transcrito em pequenas quantidades Devido agrave elevada quantidade de
proteiacutena carregada que provoca a saturaccedilatildeo do sinal e impossibilita a detecccedilatildeo de pequenas
variaccedilotildees tambeacutem natildeo se conseguiram observar alteraccedilotildees na quantidade proteica da
subunidade α do componente E1 do complexo piruvato desidrogenase e na enzima lactato
desidrogenase No entanto reduccedilotildees evidentes de actividade intracelular da lactato
desidrogenase jaacute tinham sido observadas (Figuras 4 e 8) De salientar ainda as reduccedilotildees
evidentes na quantidade proteica de PDK1 e PDK3 nas ceacutelulas silenciadas particularmente as
duplamente silenciadas
19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

19
Figura 9 Expressatildeo geacutenica e proteica apoacutes silenciamento de HIF1 e PDK1 e PDK3 Quantificaccedilatildeo da
expressatildeo geacutenica por qRT-PCR em ceacutelulas 293 FLEX GFP (A) e MDCK E1 (B) a quantificaccedilatildeo da expressatildeo
foi realizada relativamente ao gene housekeeping RPL22 e normalizada para as respectivas linhas celulares
parentais as barras de erro correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para
amostras natildeo emparelhadas entre 3 replicados (C) Western Blot para detecccedilatildeo da expressatildeo proteica cada poccedilo
foi carregado com a mesma quantidade de proteiacutena e os anticorpos utilizados encontram-se listados no Anexo 3
(HIF1) factor 1 indutiacutevel por hipoxia (PDHE1α) subunidade α do componente E1 do complexo piruvato
desidrogenase (LDHA e LDHB) lactato desidrogenase A e B (PDK1 e PDK3) cinase 1 e 3 da piruvato
desidrogenase (loading control) β-actina
00
05
10
15
20
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
00
05
10
15
20
25
30
HIF1 PDHE1α LDHA LDHB PDK1 PDK3
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
MDCK E1 MDCK 1xntshRNA MDCK 2xntshRNA
MDCK shHIF115 MDCK shHIF115+shPDK13
HIF1
PDK1
PDK3
PDHE1α fosforilado
PDHE1α
LDH
Loading control
A
B
C
20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

20
Para avaliar o impacto desta manipulaccedilatildeo adicional do efeito de Warburg na
produtividade celular quantificou-se a produccedilatildeo de partiacuteculas totais e partiacuteculas infecciosas
(Figura 10) Nas ceacutelulas 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA apresentaram
uma produccedilatildeo viral muito reduzida comparativamente agrave linha celular parental No clone
FLEX shHIF121 agrave semelhanccedila dos resultados anteriormente obtidos (Figura 6) observou-se
um notaacutevel aumento da produtividade A populaccedilatildeo duplamente silenciada apesar dos
silenciamentos moderados de PDK1 e PDK3 (Figura 9) apresentou aumentos tambeacutem eles
notaacuteveis na produccedilatildeo viral 59x na taxa de produccedilatildeo de partiacuteculas totais e 185x na taxa de
partiacuteculas infecciosas relativamente agrave linha celular parental Nas ceacutelulas MDCK E1 e ao
contraacuterio do observado para as 293 FLEX GFP as duas populaccedilotildees controlo ntshRNA
apresentaram aumentos na produccedilatildeo viral No clone MDCK shHIF115 natildeo se observou um
aumento da produtividade de partiacuteculas totais como anteriormente (Figura 6) no entanto a
produtividade especiacutefica de partiacuteculas infecciosas foi 35x superior relativamente agrave linha
celular parental A populaccedilatildeo duplamente silenciada apesar de um ligeiro decreacutescimo na
produccedilatildeo de partiacuteculas totais apresentou um aumento da produccedilatildeo de partiacuteculas infecciosas
de 22x relativamente agrave linha celular parental
A anaacutelise das razotildees PIPT (Tabela 5) confirmou que o silenciamento de HIF1
resultou num aumento de qualidade da produccedilatildeo viral para ambas as linhas celulares No
entanto apenas para as ceacutelulas 293 FLEX GFP o segundo silenciamento (shPDK13) resultou
num melhoramento adicional notaacutevel relativamente ao obtido com o silenciamento de HIF1
(aumento de 28x na razatildeo PIPT)
Figura 10 Produtividade viral apoacutes o silenciamento de HIF1 e PDK1 e PDK3 Produtividade especiacutefica em
ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro das produtividades especiacuteficas de partiacuteculas retrovirais totais e de partiacuteculas adenovirais (totais e infecciosas) correspondem ao desvio-padratildeo n =
3 e n = 2 respectivamente as barras de erro da produtividade especiacutefica de partiacuteculas retrovirais infecciosas
correspondem ao erro calculado por regressatildeo linear segundo o meacutetodo dos miacutenimos quadrados (βPT)
produtividade especiacutefica de partiacuteculas virais totais (βPI) produtividade especiacutefica de partiacuteculas virais infecciosas
0
10
20
30
40
50
60
70
80
90
100
0
1
2
3
4
5
6
7
8
9
10
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF21
+shPDK13
βP
I(1
0-3
PI
ceacutelu
la ∙
h)
βP
T(P
Tc
eacutelu
la ∙
h)
Partiacuteculas Totais Partiacuteculas Infecciosas
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
βP
I(10
3P
Iceacute
lula
)
βP
T (1
05
PT
ceacutel
ula
)
Partiacuteculas Totais Partiacuteculas InfecciosasA B
21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

21
Tabela 5 Qualidade das produccedilotildees virais razatildeo PIPT
293 FLEX GFP 1 123 plusmn 15 MDCK E1 1 286 plusmn 22
FLEX 1xntshRNA 1 1248 plusmn 434 MDCK 1xntshRNA 1 126 plusmn 30
FLEX 2xntshRNA 1 282 plusmn 209 MDCK 2xntshRNA 1 118 plusmn 10
FLEX shHIF121 1 108 plusmn 33 MDCK shHIF115 1 88 plusmn 6
FLEX
shHIF121+shPDK13 1 39 plusmn 9
MDCK
shHIF115+shPDK13 1 107 plusmn 10
(PIPT) razatildeo partiacuteculas infecciosaspartiacuteculas totais o erro corresponde agrave propagaccedilatildeo de erro
Observou-se que as populaccedilotildees controlo ntshRNA apresentavam uma intensidade de
fluorescecircncia reduzida enquanto as ceacutelulas silenciadas apresentavam uma intensidade de
fluorescecircncia muito acentuada comparativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia conferida pela expressatildeo do transgene viral avaliou-se o impacto das
manipulaccedilotildees efectuadas na expressatildeo deste e das outras duas construccedilotildees virais (gag-pol e
env GaLV) (Figura 11) As populaccedilotildees controlo ntshRNA apresentaram uma reduccedilatildeo muito
acentuada na expressatildeo dos genes gag-pol e do transgene sem que a expressatildeo de env GaLV
fosse afectada de forma relevante Contrariamente o clone FLEX shHIF121 apresentou um
aumento notaacutevel da expressatildeo de gag-pol (174x) e de transgene (139x) e em menor escala
do env GaLV (22x) comparativamente agrave linha celular parental Estes aumentos foram
igualmente observados na populaccedilatildeo duplamente silenciada e vatildeo de encontro aos
anteriormente obtidos para as produccedilotildees virais (Figura 10) Semelhante anaacutelise para o
componente E1 da linha celular MDCK E1 natildeo foi realizada por limitaccedilatildeo de tempo
Figura 11 Expressatildeo de genes virais em ceacutelulas 293 FLEX GFP A quantificaccedilatildeo da expressatildeo foi realizada
por qRT-PCR relativamente ao gene housekeeping RPL22 e normalizada para a linha celular parental as barras
de erro correspondem ao desvio-padratildeo n = 3 (gag-pol e transgene) n = 2 (env GaLV) p lt 005 dado por um
teste t bilateral para amostras natildeo emparelhadas entre 3 ou 2 replicados
0
2
4
6
8
10
12
14
16
18
gag-pol transgene env GaLV
Ex
pre
ssatilde
o r
ela
tiv
a n
orm
ali
zad
a
293 FLEX GFP FLEX 1xntshRNA FLEX 2xntshRNA
FLEX shHIF121 FLEX shHIF121+shPDK13
22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

22
O fenoacutetipo de glicoacutelise com formaccedilatildeo de lactato foi sugerido por Kondoh et al (2005)
52 como um mecanismo de protecccedilatildeo contra o stress oxidativo devido agrave diminuiccedilatildeo de fluxo
atraveacutes da cadeia respiratoacuteria Para avaliar um possiacutevel cenaacuterio de stress oxidativo nas ceacutelulas
silenciadas quantificou-se as espeacutecies reactivas de oxigeacutenio intracelularmente (Figura 12)
Interessantemente quer o clone FLEX shHIF121 como a populaccedilatildeo duplamente silenciada
apresentaram valores ligeiramente inferiores agrave linha celular parental e controlos ntshRNA
Nas ceacutelulas MDCK natildeo foram observadas alteraccedilotildees
Figura 12 Detecccedilatildeo de espeacutecies reactivas de oxigeacutenio (ERO) Taxa de detecccedilatildeo de ERO em ceacutelulas 293
FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-padratildeo n = 12 p lt 005
dado por um teste t bilateral para amostras natildeo emparelhadas entre 12 replicados
De forma a determinar se os silenciamentos efectuados induziram um desbloqueio do
TCA e consequente melhoria do estado energeacutetico ndash razatildeo ADPATP ndash e do poder redutor
disponiacutevel ndash razotildees NAD+NADH e NADP
+NADPH ndash quantificaram-se os fosfatos de
adenosina e os nucleoacutetidos de piridina intracelularmente Nas ceacutelulas 293 FLEX GFP natildeo
foram observadas diferenccedilas nas razotildees ADPATP entre a linha celular parental controlos
ntshRNA e ceacutelulas silenciadas (Figura 13) Nas ceacutelulas MDCK E1 observou-se uma reduccedilatildeo
da razatildeo ADPATP de 604 no clone silenciado para HIF1 e de 661 na populaccedilatildeo
duplamente silenciada relativamente agrave linha celular parental indicativo de uma maior
disponibilidade intracelular de ATP
0000
0001
0002
0003
0004
0005
0006
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(m
in-1
)
0000
0001
0002
0003
0004
0005
0006
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
Taxa
de
det
ecccedilatilde
o d
e E
RO
(min
-1)A B
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100
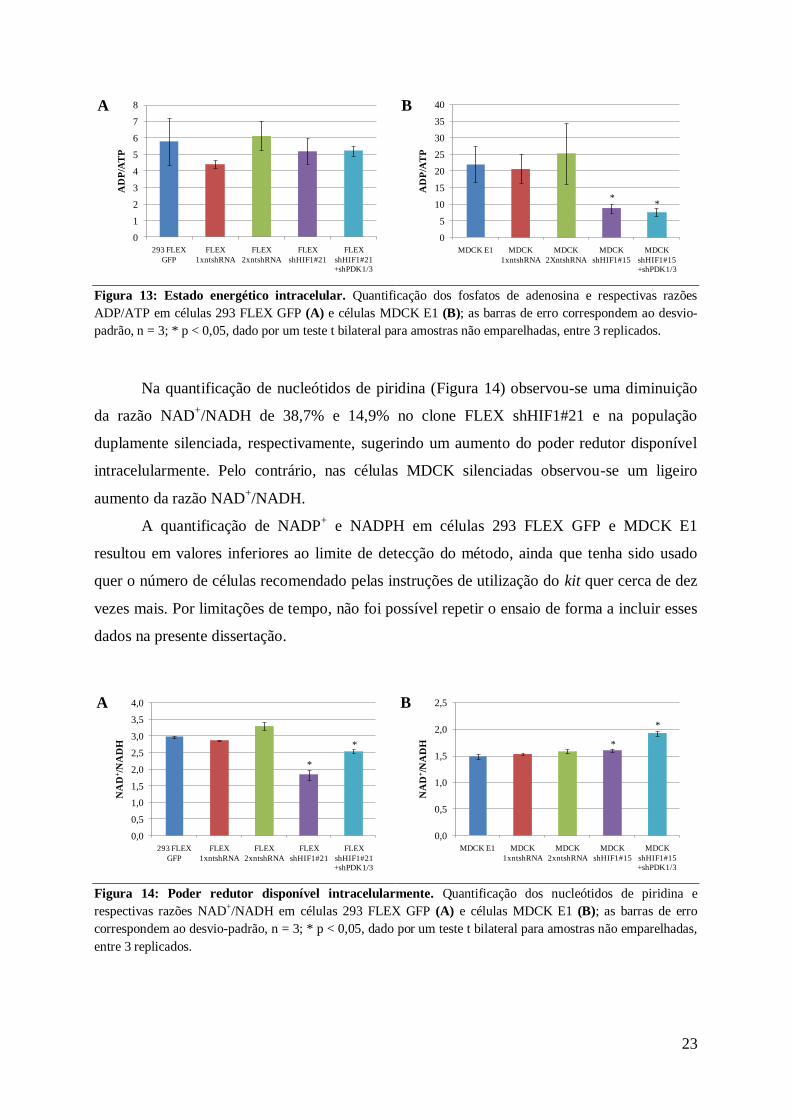
23
Figura 13 Estado energeacutetico intracelular Quantificaccedilatildeo dos fosfatos de adenosina e respectivas razotildees
ADPATP em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro correspondem ao desvio-
padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas entre 3 replicados
Na quantificaccedilatildeo de nucleoacutetidos de piridina (Figura 14) observou-se uma diminuiccedilatildeo
da razatildeo NAD+NADH de 387 e 149 no clone FLEX shHIF121 e na populaccedilatildeo
duplamente silenciada respectivamente sugerindo um aumento do poder redutor disponiacutevel
intracelularmente Pelo contraacuterio nas ceacutelulas MDCK silenciadas observou-se um ligeiro
aumento da razatildeo NAD+NADH
A quantificaccedilatildeo de NADP+ e NADPH em ceacutelulas 293 FLEX GFP e MDCK E1
resultou em valores inferiores ao limite de detecccedilatildeo do meacutetodo ainda que tenha sido usado
quer o nuacutemero de ceacutelulas recomendado pelas instruccedilotildees de utilizaccedilatildeo do kit quer cerca de dez
vezes mais Por limitaccedilotildees de tempo natildeo foi possiacutevel repetir o ensaio de forma a incluir esses
dados na presente dissertaccedilatildeo
Figura 14 Poder redutor disponiacutevel intracelularmente Quantificaccedilatildeo dos nucleoacutetidos de piridina e
respectivas razotildees NAD+NADH em ceacutelulas 293 FLEX GFP (A) e ceacutelulas MDCK E1 (B) as barras de erro
correspondem ao desvio-padratildeo n = 3 p lt 005 dado por um teste t bilateral para amostras natildeo emparelhadas
entre 3 replicados
0
1
2
3
4
5
6
7
8
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
AD
PA
TP
0
5
10
15
20
25
30
35
40
MDCK E1 MDCK
1xntshRNA
MDCK
2XntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
AD
PA
TP
00
05
10
15
20
25
30
35
40
293 FLEX
GFP
FLEX
1xntshRNA
FLEX
2xntshRNA
FLEX
shHIF121
FLEX
shHIF121
+shPDK13
NA
D+N
AD
H
00
05
10
15
20
25
MDCK E1 MDCK
1xntshRNA
MDCK
2xntshRNA
MDCK
shHIF115
MDCK
shHIF115
+shPDK13
NA
D+N
AD
H
A B
A B
24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

24
4 DISCUSSAtildeO E CONCLUSAtildeO
A crescente procura de biofaacutermacos complexos produzidos em ceacutelulas animais requer
o desenvolvimento de processos robustos aliados a ceacutelulas de elevada produtividade Poreacutem a
maioria das linhas celulares de mamiacutefero possuem um metabolismo energeacutetico desregulado
consumindo elevadas quantidades de glucose e acumulando quantidades significativas de
lactato 4 com a consequente reduccedilatildeo na produtividade tempo uacutetil de cultura e qualidade do
produto final Este trabalho teve por objectivo a manipulaccedilatildeo do fenoacutetipo lactogeacutenico
associado ao efeito de Warburg em ceacutelulas de mamiacutefero em cultura procurando
simultaneamente reduzir a acumulaccedilatildeo de lactato e aumentar a eficiecircncia do metabolismo
energeacutetico com o consequente aumento da produtividade celular A tecnologia de RNA de
interferecircncia via transduccedilatildeo com vectores lentivirais foi escolhida como uma ferramenta
raacutepida e versaacutetil para o silenciamento de HIF1 PDK1 e PDK3 os principais efectores do
fenoacutetipo lactogeacutenico associado ao efeito de Warburg
As linhas celulares 293 FLEX GFP (produtora de retroviacuterus) MDCK E1 (produtora de
adenoviacuterus) e RPE A6 (produtora de anticorpos monoclonais) foram numa primeira fase
transduzidas com vectores lentivirais transportando um hairpin de silenciamento de HIF1
Este processo foi realizado a uma baixa multiplicidade de infecccedilatildeo (01) para minimizar a
activaccedilatildeo da via de interferatildeo uma resposta celular de defesa contra infecccedilatildeo de viacuterus de
RNA que leva a uma inibiccedilatildeo natildeo especiacutefica da traduccedilatildeo e agrave possiacutevel apoptose celular 53
Apoacutes o silenciamento de HIF1 as alteraccedilotildees morfoloacutegicas de crescimento celular e a
detecccedilatildeo de actividade da β-galactosidase na maioria dos clones RPE A6 (Figura 2) sugeriram
a induccedilatildeo de um cenaacuterio de senescecircncia 4849
Esta linha celular foi imortalizada por
transfecccedilatildeo com a subunidade cataliacutetica da telomerase humana (hTERT) o HIF1 foi descrito
como tendo um papel criacutetico na actividade desta enzima 54-57
Assim os resultados obtidos
parecem sugerir que o seu silenciamento levou agrave inactivaccedilatildeo da telomerase reversatildeo da
imortalizaccedilatildeo e consequente entrada em senescecircncia replicativa Nas restantes linhas celulares
estas alteraccedilotildees natildeo foram observadas as ceacutelulas MDCK natildeo possuem actividade
telomeraacutesica detectaacutevel 58
as ceacutelulas derivadas de 293 apesar de possuiacuterem actividade
telomeraacutesica 59
natildeo aparentaram sofrer alteraccedilotildees decorrentes do silenciamento de HIF1
Sem que houvesse alteraccedilotildees de relevacircncia no crescimento celular nas ceacutelulas 293
FLEX GFP silenciadas foram observadas diminuiccedilotildees no consumo de glucose e na produccedilatildeo
de lactato especialmente apoacutes o silenciamento adicional de PDK1 e PDK3 acopladas agrave
25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

25
diminuiccedilatildeo de actividade da enzima lactato desidrogenase (Figuras 4 7 e 8 e Tabelas 1 e 4)
Estes resultados estatildeo de acordo com a quantificaccedilatildeo da expressatildeo geacutenica onde foram
observadas reduccedilotildees na expressatildeo dos genes HIF1 LDHA PDK1 e PDK3 e com os
resultados de Western Blot onde se observaram reduccedilotildees evidentes de PDK1 e PDK3 (Figura
9) Nas ceacutelulas MDCK E1 silenciadas apesar de se terem observado diminuiccedilotildees na
expressatildeo dos genes atraacutes referidos bem como na quantidade de proteiacutena por eles codificada
tal natildeo teve repercussatildeo no fenoacutetipo jaacute que natildeo se observaram reduccedilotildees nas taxas de consumo
e produccedilatildeo de glucose e lactato respectivamente (Tabela 1 e Figura 7) As diferenccedilas nos
resultados obtidos entre a primeira fase de caracterizaccedilatildeo (silenciamento de HIF1) e a
segunda (silenciamento de HIF1 PDK1 e PDK3) como ao niacutevel da expressatildeo geacutenica ou de
actividade da lactato desidrogenase poderatildeo ser explicadas natildeo soacute como decorrentes das
diferentes condiccedilotildees de cultura como de erros inerentes agraves proacuteprias teacutecnicas Adicionalmente
podem tambeacutem dever-se ao padratildeo oscilatoacuterio do silenciamento via RNA de interferecircncia
durante o crescimento celular 60
Os resultados acima descritos suportam a reduccedilatildeo do fenoacutetipo lactogeacutenico associado
ao efeito de Warburg com base no silenciamento sucessivo de HIF1 PDK1 e PDK3 O
impacto desta reduccedilatildeo na produtividade celular foi adicionalmente avaliado O silenciamento
de HIF1 levou natildeo soacute ao aumento da produtividade viral como tambeacutem da sua qualidade
avaliada pela razatildeo PIPT Este efeito foi particularmente notaacutevel no clone FLEX shHIF121
ndash com aumento ateacute 212x na produtividade de partiacuteculas infecciosas ndash e em menor escala no
clone MDCK shHIF115 ndash com um aumento ateacute 35x na produtividade de partiacuteculas
infecciosas (Figuras 6 e 10 e Tabelas 2 e 5) O silenciamento adicional de PDK1 e PDK3
nestes clones ainda que natildeo se traduzindo num aumento de tiacutetulo viral infeccioso
relativamente aos correspondentes clones silenciados para HIF1 levou agrave obtenccedilatildeo de
produccedilotildees virais de qualidade superior De salientar que os resultados observados para a
populaccedilatildeo duplamente silenciada reflectem valores meacutedios resultantes da heterogeneidade
fenotiacutepica pelo que eacute de esperar que alguns dos clones desta populaccedilatildeo apresentem um
fenoacutetipo superior Estes resultados demonstram que a manipulaccedilatildeo do efeito de Warburg em
particular nas ceacutelulas 293 FLEX GFP resultou natildeo soacute na obtenccedilatildeo de ceacutelulas mais produtivas
mas tambeacutem mais eficientes na medida em que para o mesmo nuacutemero de partiacuteculas totais
produzidas corresponde uma maior fracccedilatildeo de partiacuteculas infecciosas Uma maior eficiecircncia de
produccedilatildeo nestas ceacutelulas foi de resto anteriormente descrita como decorrente de condiccedilotildees
nutricionais oacuteptimas capazes de alterar o seu estado metaboacutelico 33
Neste sentido avaliou-se o
potencial de desbloqueio do TCA induzido pelo silenciamento de HIF1 PDK1 e PDK3 e
26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

26
consequente melhoria no estado energeacutetico e do poder redutor disponiacutevel intracelularmente
Os resultados obtidos (Figuras 13 e 14) sugerem um aumento dos niacuteveis de ATP nas ceacutelulas
MDCK E1 e aumento de poder redutor nas ceacutelulas 293 FLEX GFP Como a produccedilatildeo de ATP
estaacute acoplada ao fluxo de prototildees doados pelo NADH ao niacutevel da cadeia respiratoacuteria
mitocondrial entatildeo eacute expectaacutevel que o aumento dos niacuteveis de ATP nas ceacutelulas MDCK E1
resulte de um maior poder redutor disponiacutevel intracelularmente de igual forma eacute expectaacutevel
que o aumento do poder redutor nas ceacutelulas 293 FLEX GFP resulte em niacuteveis superiores de
produccedilatildeo de ATP O facto de natildeo terem sido observadas alteraccedilotildees relevantes nas razotildees
ADPATP nas ceacutelulas 293 FLEX GFP e NAD+NADH nas ceacutelulas MDCK E1 pode dever-se a
um maior consumo ndash associado a uma maior disponibilidade ndash mantendo-se as razotildees
inalteradas De resto o aumento consideraacutevel de partiacuteculas totais principalmente nas ceacutelulas
293 FLEX GFP (Figuras 6 e 10) estaraacute necessariamente associado a um maior consumo de
ATP e disponibilidade de poder redutor suportando esta hipoacutetese
O aumento do fluxo metaboacutelico atraveacutes do TCA e da cadeia respiratoacuteria sugerido
pelos resultados anteriores colocam a hipoacutetese de uma maior sobrecarga oxidativa 52
Os
resultados obtidos (Figura 12) mostraram que esse efeito natildeo se fez sentir De facto um
aumento dos niacuteveis intracelulares de espeacutecies reactivas de oxigeacutenio (ERO) pode facilmente
culminar na morte celular pelo que eacute razoaacutevel aceitar que as ceacutelulas sobreviventes agraves
manipulaccedilotildees induzidas representam apenas eventos onde o desequiliacutebrio na carga oxidativa
natildeo ocorreu em extensatildeo apreciaacutevel Interessantemente no clone FLEX shHIF121 e na
respectiva populaccedilatildeo duplamente silenciada observou-se uma ligeira diminuiccedilatildeo das ERO
quantificadas relativamente agrave linha celular parental e aos controlos ntshRNA Estes
resultados colocam a hipoacutetese de que para as ceacutelulas sobreviventes um silenciamento
moderado de HIF1 PDK1 e PDK3 acoplado a um pequeno aumento da sobrecarga oxidativa
possam induzir um cenaacuterio de preacute-condicionamento descrito para outros sistemas celulares 61
em que um pequeno ldquoinsultordquo oxidativo activa sistemas de protecccedilatildeo resultando em ceacutelulas
mais resistentes ao stress oxidativo
Os resultados obtidos decorrentes do silenciamento moderado de HIF1 PDK1 e
PDK3 mostram que a manipulaccedilatildeo do efeito de Warburg resultou numa reduccedilatildeo do efeito
lactogeacutenico e sugerem um aumento da eficiecircncia do metabolismo energeacutetico por desbloqueio
do TCA ao niacutevel do complexo piruvato desidrogenase Niacuteveis de silenciamento superiores aos
obtidos poderiam resultar num fenoacutetipo de qualidade superior No entanto um elevado
silenciamento destes genes poderaacute estar associado a uma desvantagem proliferativa HIF1
controla ateacute 5 do genoma 62
pelo que um elevado silenciamento deste factor de transcriccedilatildeo
27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

27
poderaacute levar agrave desregulaccedilatildeo de outras vias onde estaacute envolvido e consequentemente da
homeostasia celular um silenciamento total de PDK1 e PDK3 poderaacute desregular o sistema no
sentido oposto ou seja a super-activaccedilatildeo do complexo piruvato desidrogenase Tal facto
poderaacute natildeo soacute explicar o enriquecimento em ceacutelulas com maior vantagem proliferativa (menor
silenciamento) decorrente do alargado periacuteodo temporal desde a transduccedilatildeo com vectores
lentivirais selecccedilatildeo da populaccedilatildeo e subsequente clonagem como tambeacutem a obtenccedilatildeo de
apenas dois clones duplamente silenciados dos cerca de 200 clones isolados e mantidos em
paralelo agrave realizaccedilatildeo do presente estudo
Durante este trabalho notaram-se alteraccedilotildees na intensidade de fluorescecircncia de ceacutelulas
293 FLEX GFP transduzidas relativamente agrave linha celular parental (Anexo 4) Sendo a
fluorescecircncia resultado da expressatildeo de uma de trecircs construccedilotildees virais introduzidas nesta
linha celular ndash transgene ndash avaliou-se o impacto das manipulaccedilotildees efectuadas nas respectivas
expressotildees geacutenicas (Figura 11) Os resultados obtidos sugeriram um efeito toacutexico em ceacutelulas
293 FLEX GFP do hairpin usado nos controlos ntshRNA o que corrobora a reduccedilatildeo draacutestica
observada na produccedilatildeo viral (Figura 10) No entanto nas populaccedilotildees controlo MDCK E1 natildeo
se observou o mesmo efeito pelo que estudos posteriores seratildeo necessaacuterios para tentar
compreender este efeito antagoacutenico Pelo contraacuterio os elevados aumentos da expressatildeo de
gag-pol do transgene e em menor escala de env GaLV observados no clone FLEX
shHIF121 sugerem um cenaacuterio de alteraccedilatildeo transcricional no qual a expressatildeo das trecircs
construccedilotildees retrovirais usadas para estabelecer esta linha celular deixou de estar sob o efeito
inibitoacuterio do factor de transcriccedilatildeo HIF1 De facto e apesar de ser uma condiccedilatildeo necessaacuteria
mas natildeo suficiente existem elementos de resposta agrave hipoxia via HIF1 63
nas trecircs construccedilotildees
suportando a sugestatildeo de que a sua expressatildeo se encontra sob influecircncia de HIF1 Estes
resultados podem explicar em parte os elevados aumentos da produccedilatildeo viral nas ceacutelulas
silenciadas No entanto o aumento de 28x na qualidade da produccedilatildeo viral na populaccedilatildeo
duplamente silenciada (Tabela 5) natildeo pode ser explicado como resultado do efeito
transcricional jaacute que os niacuteveis de expressatildeo dos genes virais satildeo idecircnticos aos do clone
silenciado para HIF1 Assim este aumento deveraacute estar relacionado com a reduccedilatildeo adicional
na acumulaccedilatildeo de lactato e com um aumento de eficiecircncia na assemblagem de partiacuteculas
virais como resultado da reduccedilatildeo do fenoacutetipo lactogeacutenico e do aumento de eficiecircncia do
metabolismo energeacutetico Um estudo detalhado da estabilidade das partiacuteculas virais permitiraacute
avaliar tambeacutem o impacto da manipulaccedilatildeo do efeito de Warburg na qualidade intriacutenseca das
mesmas
28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

28
Globalmente os resultados obtidos neste trabalho em particular para a linha celular
293 FLEX GFP demonstram que o silenciamento sucessivo de HIF1 PDK1 e PDK3 permite
a manipulaccedilatildeo do fenoacutetipo lactogeacutenico associado ao efeito de Warburg com evidentes
reduccedilotildees no consumo de glucose e de acumulaccedilatildeo de lactato Apesar de natildeo ter sido possiacutevel
observar alteraccedilotildees na razatildeo complexo PDH fosforiladoPDH natildeo fosforilado (Figura 9C) os
notaacuteveis aumentos de produccedilatildeo viral e da sua qualidade sugerem que a manipulaccedilatildeo efectuada
desbloqueou o TCA com o consequente aumento de eficiecircncia do metabolismo energeacutetico e
da produtividade celular A obtenccedilatildeo destas ceacutelulas de fenoacutetipo superior agrave linha celular
parental evidencia a manipulaccedilatildeo do efeito de Warburg como uma potencial ferramenta de
engenharia metaboacutelica com vista ao aumento da produtividade de biofaacutermacos complexos em
ceacutelulas de mamiacutefero em cultura De facto durante a execuccedilatildeo do trabalho descrito nesta
dissertaccedilatildeo Zhou et al (2011) 64
obtiveram aumentos consideraacuteveis na produccedilatildeo de
anticorpos monoclonais em ceacutelulas Chinese Hamster Ovary (CHO) pela reduccedilatildeo da expressatildeo
de Ldh e Pdk1-3
29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

29
5 PERSPECTIVAS FUTURAS
Apesar dos resultados obtidos durante o presente estudo suportarem a hipoacutetese de que
a manipulaccedilatildeo de HIF1 PDK1 e PDK3 poderaacute conduzir a um metabolismo mais eficiente
resultados ainda mais expressivos poderatildeo advir da caracterizaccedilatildeo dos clones duplamente
silenciados jaacute que a anaacutelise aqui efectuada incidiu sobre a populaccedilatildeo duplamente silenciada
Os clones de fenoacutetipo superior seratildeo bons candidatos para a realizaccedilatildeo de um estudo de
Anaacutelise de Fluxos Metaboacutelicos acoplado a substratos marcados o que permitiraacute confirmar em
definitivo se as manipulaccedilotildees geneacuteticas efectuadas induziram um aumento do fluxo
metaboacutelico atraveacutes do TCA como consequecircncia do desbloqueio ao niacutevel do complexo
piruvato desidrogenase Adicionalmente estes clones seratildeo caracterizados pela teacutecnica de
microarrays para avaliar o impacto global na expressatildeo geacutenica
A integraccedilatildeo dos dados do transcriptoma e metaboloma constituiraacute uma poderosa
ferramenta natildeo soacute do ponto de vista tecnoloacutegico podendo ser extensiacutevel agrave manipulaccedilatildeo de
outras linhas celulares de mamiacutefero utilizadas na produccedilatildeo de biofaacutermacos complexos como
tambeacutem de um ponto de vista fundamental na medida em que forneceraacute uma perspectiva
global e integrada do efeito de Warburg e da sua manipulaccedilatildeo na fisiologia e metabolismo
celulares
30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

30
6 REFEREcircNCIAS BIBLIOGRAacuteFICAS
1 Mountain A Gene therapy the first decade Trends in Biotechnology 18 119-128
(2000)
2 Browne SM amp Al-Rubeai M Selection methods for high-producing mammalian cell
lines Trends in Biotechnology 25 425-32 (2007)
3 Kraumlmer O Klausing S amp Noll T Methods in mammalian cell line engineering from
random mutagenesis to sequence-specific approaches Applied Microbiology and
Biotechnology 88 425-36 (2010)
4 Neermann J amp Wagner R Comparative analysis of glucose and glutamine metabolism
in transformed mammalian cell lines insect and primary liver cells Journal of Cellular
Physiology 166 152-69 (1996)
5 Higashikawa F amp Chang L Kinetic analyses of stability of simple and complex
retroviral vectors Virology 280 124-31 (2001)
6 Zhou W Rehm J Europa A amp Hu W-S Alteration of mammalian cell metabolism
by dynamic nutrient feeding Cytotechnology 24 99-108 (1997)
7 Sanfeliu A Identification of key patterns in the metabolism of hybridoma cells in
culture Enzyme and Microbial Technology 21 421-428 (1997)
8 Coroadinha AS et al Effect of medium sugar source on the production of retroviral
vectors for gene therapy Biotechnology and Bioengineering 94 24-36 (2006)
9 Nadeau I et al Improvement of recombinant protein production with the human
adenovirus293S expression system using fed-batch strategies Biotechnology and
Bioengineering 51 613-623 (1996)
10 Poumlrtner R Schilling A Luumldemann I amp Maumlrkl H High density fed-batch cultures for
hybridoma cells performed with the aid of a kinetic model Bioprocess Engineering 15
117-124 (1996)
11 Xie L amp Wang DIC Material balance studies on animal cell metabolism using a
stoichiometrically based reaction network Biotechnology and Bioengineering 52 579-
590 (1996)
12 Elias CB et al Improving glucose and glutamine metabolism of human HEK 293 and
Trichoplusia ni insect cells engineered to express a cytosolic pyruvate carboxylase
enzyme Biotechnology Progress 19 90-7 (2003)
13 Irani N Wirth M van den Heuvel J amp Wagner R Improvement of the primary
metabolism of cell cultures by introducing a new cytoplasmic pyruvate carboxylase
reaction Biotechnology and Bioengineering 66 238-246 (1999)
14 MB Wagner R Etcheverrigaray M amp Kratje R Impact of temperature
reduction and expression of yeast pyruvate carboxylase on hGM-CSF-producing CHO
cells Journal of Biotechnology 109 179-191 (2004)
15 Chen K Liu Q Xie L Sharp P a amp Wang DI Engineering of a mammalian cell
line for reduction of lactate formation and high monoclonal antibody production
Biotechnology and Bioengineering 72 55-61 (2001)
31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

31
16 Kim SH amp Lee GM Down-regulation of lactate dehydrogenase-A by siRNAs for
reduced lactic acid formation of Chinese hamster ovary cells producing thrombopoietin
Applied Microbiology and Biotechnology 74 152-9 (2007)
17 Wlaschin KF amp Hu W-S Engineering cell metabolism for high-density cell culture via
manipulation of sugar transport Journal of Biotechnology 131 168-76 (2007)
18 Warburg O On the Origin of Cancer Cells Science 123 309-314 (1956)
19 Warburg O Uumlber den Stoffwechsel der Carcinomzelle Klinische Wochenschrift 4 534-
536 (1925)
20 Kondoh H Cellular life span and the Warburg effect Experimental Cell Research 314
1923-8 (2008)
21 Fridman AL amp Tainsky MA Critical pathways in cellular senescence and
immortalization revealed by gene expression profiling Oncogene 27 5975-87 (2008)
22 Zhong H et al Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases Cancer Research 59 5830-5 (1999)
23 Talks KL et al The Expression and Distribution of the Hypoxia-Inducible Factors HIF-
1α and HIF-2α in Normal Human Tissues Cancers and Tumor-Associated Macrophages
The American Journal of Pathology 157 411-421 (2000)
24 DeBerardinis RJ Lum JJ Hatzivassiliou G amp Thompson CB The biology of
cancer metabolic reprogramming fuels cell growth and proliferation Cell Metabolism 7
11-20 (2008)
25 Weidemann A amp Johnson RS Biology of HIF-1alpha Cell Death and Differentiation
15 621-7 (2008)
26 Patel MS amp Korotchkina LG The biochemistry of the pyruvate dehydrogenase
complex Biochemistry and Molecular Biology Education 31 5-15 (2003)
27 Rardin MJ Wiley SE Naviaux RK Murphy AN amp Dixon JE Monitoring
phosphorylation of the pyruvate dehydrogenase complex Analytical Biochemistry 389
157-64 (2009)
28 Lu C-W Lin S-C Chen K-F Lai Y-Y amp Tsai S-J Induction of pyruvate
dehydrogenase kinase-3 by hypoxia-inducible factor-1 promotes metabolic switch and
drug resistance The Journal of Biological Chemistry 283 28106-14 (2008)
29 Vander Heiden MG Cantley LC amp Thompson CB Understanding the Warburg
effect the metabolic requirements of cell proliferation Science 324 1029-33 (2009)
30 Zhu J Mammalian cell protein expression for biopharmaceutical production
Biotechnology Advances (2011)doi101016jbiotechadv201108022
31 Rodrigues T Carrondo MJT Alves PM amp Cruz PE Purification of retroviral
vectors for clinical application biological implications and technological challenges
Journal of Biotechnology 127 520-41 (2007)
32 Coroadinha AS et al Retrovirus producer cell line metabolism implications on viral
productivity Applied Microbiology and Biotechnology 72 1125-35 (2006)
33 Rodrigues AF Carmo M Alves PM amp Coroadinha AS Retroviral vector
production under serum deprivation The role of lipids Biotechnology and
Bioengineering 104 1171-81 (2009)
34 Coroadinha AS et al Effect of osmotic pressure on the production of retroviral vectors
Enhancement in vector stability Biotechnology and Bioengineering 94 322-9 (2006)
32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

32
35 Amaral AI Coroadinha AS Merten O-W amp Alves PM Improving retroviral
vectors production role of carbon sources in lipid biosynthesis Journal of Biotechnology
138 57-66 (2008)
36 McFate T et al Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells The Journal of Biological Chemistry 283 22700-8
(2008)
37 Coroadinha AS et al The use of recombinase mediated cassette exchange in retroviral
vector producer cell lines predictability and efficiency by transgene exchange Journal of
Biotechnology 124 457-68 (2006)
38 Santiago VM et al Development of a MDCK-CAV-2 E1 transcomplementing cell line
the impact of E1 gene expression in viral amplification
39 Moffat J et al A lentiviral RNAi library for human and mouse genes applied to an
arrayed viral high-content screen Cell 124 1283-98 (2006)
40 Rasmussen R Quantification on the LightCycler Rapid Cycle Real-Time PCR-Methods
and Applications 21-34 (2001)
41 Livak KJ amp Schmittgen TD Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method Methods 25 402-8 (2001)
42 Altschul SF et al Gapped BLAST and PSI-BLAST a new generation of protein
database search programs Nucleic Acids Research 25 3389-402 (1997)
43 Rozen S amp Skaletsky H Primer3 on the WWW for general users and for biologist
programmers Bioinformatics Methods and Protocols Methods in Molecular Biology
132 365-86 (2000)
44 Carmo M et al Quantitation of MLV-based retroviral vectors using real-time RT-PCR
Journal of Virological Methods 119 115-9 (2004)
45 Altaras NE et al Production and formulation of adenovirus vectors Advances in
Biochemical EngineeringBiotechnology 99 193-260 (2005)
46 Kamen A amp Henry O Development and optimization of an adenovirus production
process The Journal of Gene Medicine 6 Suppl 1 S184-92 (2004)
47 Sambrook J amp Russell D Molecular Cloning A Laboratory Manual (Cold Spring
Harbor Laboratory Press 2001)
48 Beacuterubeacute NG Smith JR amp Pereira-Smith OM The genetics of cellular senescence
American Journal of Human Genetics 62 1015-9 (1998)
49 Dimri GP et al A biomarker that identifies senescent human cells in culture and in
aging skin in vivo Proceedings of the National Academy of Sciences of the United States
of America 92 9363-7 (1995)
50 Scacheri PC et al Short interfering RNAs can induce unexpected and divergent
changes in the levels of untargeted proteins in mammalian cells Proceedings of the
National Academy of Sciences of the United States of America 101 1892-7 (2004)
51 Editorial Whither RNAi Nat Cell Biol 5 489-490 (2003)
52 Kondoh H et al Glycolytic enzymes can modulate cellular life span Cancer Research
65 177-85 (2005)
53 Lehner B Fraser AG amp Sanderson CM Technique review how to use RNA
interference Briefings in Functional Genomics amp Proteomics 3 68-83 (2004)
33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

33
54 Nishi H et al Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT)
Molecular and Cellular Biology 24 6076-83 (2004)
55 Coussens M et al RNAi screen for telomerase reverse transcriptase transcriptional
regulators identifies HIF1alpha as critical for telomerase function in murine embryonic
stem cells Proceedings of the National Academy of Sciences of the United States of
America 107 13842-7 (2010)
56 Bodnar a G Extension of Life-Span by Introduction of Telomerase into Normal Human
Cells Science 279 349-352 (1998)
57 Matsunaga H et al Beta-galactosidase histochemistry and telomere loss in senescent
retinal pigment epithelial cells Investigative Ophthalmology amp Visual Science 40 197-
202 (1999)
58 Nasir L Devlin P Mckevitt T Rutteman G amp Argyle DJ Telomere lengths and
telomerase activity in dog tissues a potential model system to study human telomere and
telomerase biology Neoplasia 3 351-9 (2001)
59 Kim N et al Specific association of human telomerase activity with immortal cells and
cancer Science 266 2011-2015 (1994)
60 Rodrigues AF et al Down-regulation of CD81 tetraspanin in human cells producing
retroviral-based particles Tailoring vector composition Biotechnology and
Bioengineering 1-11 (2011)doi101002bit23231
61 Vieira HL a Queiroga CSF amp Alves PM Pre-conditioning induced by carbon
monoxide provides neuronal protection against apoptosis Journal of Neurochemistry
107 375-84 (2008)
62 Manalo DJ et al Transcriptional regulation of vascular endothelial cell responses to
hypoxia by HIF-1 Blood 105 659-69 (2005)
63 Semenza GL Hypoxia-inducible factor 1 and the molecular physiology of oxygen
homeostasis The Journal of Laboratory and Clinical Medicine 131 207-14 (1998)
64 Zhou M et al Decreasing lactate level and increasing antibody production in Chinese
Hamster Ovary cells (CHO) by reducing the expression of lactate dehydrogenase and
pyruvate dehydrogenase kinases Journal of Biotechnology 153 27-34 (2011)
34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

34
7 ANEXOS
Anexo 1 Mapas dos vectores lsquoSigma Mission
reg RNAi shRNArsquo Mapa do vector shHIF1 e primeiro ntshRNA
(A) e shPDK13 e segundo ntshRNA (B) Os hairpins satildeo clonados a montante do promotor humano U6 para o
silenciamento de PDK13 foram usados 3 vectores lentivirais cada um transportando um hairpin diferente
(CTGTCACCAGCCAGAATGTTC TAGGCGTCTGTGTGATTTGTA E GTGATTGAATACAAGGAGAGC)
e que hibridam totalmente com o mRNA de PDK1 e com os primeiros 19 pares de base de PDK3
Anexo 2 Lista de primers usados no presente estudo
Gene Sequecircncia directa Sequecircncia reversa
HIF1 GAAACCTACTGCAGGGTGAA GTAATGAGCCACCAGTGTCC
PDHE1α TTCGTGGTTTCTGTCACTTG CCTGTAAGCTCTGCGAGAAT
LDHA TTGAAGGGAGAGATGATGGA CGTTACGCTGGACCAAATTA
LDHB GAAAAACTTGGCATTCATCC GAGAAACACCTGCCACATTC
PDK1 GGTGTGATTGAATACAAGGAG CTTCCTTTGCCTTTTCCAC
PDK3 TGAAAATAAGAGCCCTGAGGA ATCACTCCTTGTGCCATTGT
RPL22 CTGCCAATTTTGAGCAGTTT CTTTGCTGTTAGCAACTACGC
Gfp CAGAAGAACGGCATCAAGGT CTGGGTGCTCAGGTAGTGG
env GaLV GGACCAAAATAGCGAATGGA GGTGAACTGTACGCCTGGAT
gag-pol GTCCACTATCGCCAGTTGCT CTGGGTCCTCAGGGTCATAA
Todas as sequecircncias estatildeo indicadas no sentido 5‟ndash3‟ e foram desenhadas para locais de homologia entre ceacutelulas
humanas e caninas recorrendo aos softwares BLAST 42 e Primer3 43
A B
35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100

35
Anexo 3 Lista de anticorpos usados em Western Blot
Anticorpo ClonalidadeModelo Fabricante Diluiccedilatildeo
α-HIF1 Monoclonal Ratinho Abcam 1500
α-PDHE1α Policlonal Coelho Abcam 11000
α-PDHE1α (fosf S293) Policlonal Coelho Abcam 11000
α-LDH Monoclonal Ratinho Abcam 11000
α-PDK1 Policlonal Coelho Novus Biologicals 12000
α-PDK3 Monoclonal Ratinho Abcam 11000
α-βactina Monoclonal Ratinho Abcam 12000
Anexo 4 Intensidade de fluorescecircncia em ceacutelulas 293 FLEX GFP detectada por citometria de fluxo
Ceacutelulas 293 foram usadas como controlo negativo de fluorescecircncia
0
100
200
300
400
500
600
0 64 128 192
Ev
ento
s
Intensidade da fluorescecircncia (log10)
293 293 FLEX GFP FLEX 1xntshRNA
FLEX 2xntshRNA FLEX shHIF121 FLEX shHIF121+shPDK13
01 1 10 100


















