
Marília Helena Fernandes Machadorepositorium.sdum.uminho.pt/bitstream/1822/35396/1/Marília...
72
outubro de 2013 Universidade do Minho Escola de Engenharia Marília Helena Fernandes Machado Produção de coalho líquido de cardo para aplicação industrial UMinho|2013 Marília Helena Fernandes Machado Produção de coalho líquido de cardo para aplicação industrial
-
Upload
nguyenhuong -
Category
Documents
-
view
215 -
download
0
Transcript of Marília Helena Fernandes Machadorepositorium.sdum.uminho.pt/bitstream/1822/35396/1/Marília...

outubro de 2013
Universidade do MinhoEscola de Engenharia
Marília Helena Fernandes Machado
Produção de coalho líquido de cardo para aplicação industrial
UM
inho
|20
13M
aríli
a H
elen
a Fe
rnan
des
Mac
hado
P
rod
uçã
o d
e c
oa
lho
líq
uid
o d
e c
ard
o p
ara
ap
lica
ção
in
du
stri
al

Dissertação de Mestrado Mestrado integrado em Engenharia Biológica Ramo Tecnologia Química e Alimentar
Trabalho efetuado sob a orientação do Doutor João Monteiro Peixoto
outubro de 2013
Universidade do MinhoEscola de Engenharia
Marília Helena Fernandes Machado
Produção de coalho líquido de cardo para aplicação industrial

DECLARAÇÃO
Nome: Marília Helena Fernandes Machado
Endereço eletrónico: [email protected]
Número do cartão de cidadão: 13728357
Título da dissertação: Produção de coalho líquido de cardo para aplicação industrial
Orientador: Doutor João Monteiro Peixoto
Ano de conclusão: 2013
Designação do Mestrado: Mestrado Integrado em Engenharia Biológica, Ramo Tecnologia
Química e Alimentar
É AUTORIZADA A REPRODUÇÃO INTEGRAL DESTA DISSERTAÇÃO, APENAS PARA EFEITOS DE
INVESTIGAÇÃO, MEDIANTE DECLARAÇÃO ESCRITA DO INTERESSADO, QUE A TAL SE
COMPROMETE.
Universidade do Minho, ___/___/____
______________________________________
(Marília Helena Fernandes Machado)

| i
Agradecimentos
Gostaria de expressar a minha gratidão a todas as pessoas que contribuíram para a
realização deste trabalho e para a conclusão de uma etapa importante e muito desejada na
minha vida.
Ao orientador, Professor João Peixoto, e ao supervisor na empresa, Rui Campos, por me
terem proporcionado sempre a liberdade de conduzir este projeto e as minhas escolhas estando,
no entanto, sempre disponíveis.
Ao Professor António Vicente pela disponibilidade e pela atenção que dedicou na
resolução de alguns problemas que foram surgindo.
À equipa de trabalho da BLC3, pelo acompanhamento e esforço ao longo destes seis
meses.
Aos meus amigos pelo apoio e pela partilha de bons momentos, em especial à minha
Pipa que me acompanha desde há muitos anos.
Agradeço às pessoas mais importantes na minha vida, os meus pais e aos meus
maninhos, a quem dedico esta dissertação. Obrigada por sempre me encorajarem e
possibilitarem este momento. Obrigada também aos meus avós e tios pelo carinho e pelos
conselhos.
A todos aqueles que direta e indiretamente marcaram a minha vida e contribuíram para
este momento, um muito Obrigada!

| iii
Sumário As proteases aspárticas (EC 3.4.23) são uma sub-subclasse de enzimas que se
encontram amplamente distribuídas na natureza. Estas enzimas desempenham um papel
importante na indústria dos laticínios, pois catalisam a coagulação do leite, podendo substituir os
coalhos de origem animal.
O objetivo do presente trabalho foi desenvolver um coagulante de origem vegetal,
extraído da planta Cynara cardunculus e avaliar a sua eficácia na coagulação do leite. As flores
desta planta, comummente designadas por flor de cardo, possuem proteases aspárticas
responsáveis pela coagulação do leite. Ao longo de várias décadas, estas têm sido usadas na
produção artesanal dos queijos de ovelha da Serra da Estrela.
Neste trabalho, para promover a extração de proteínas rompeu-se as paredes celulares
da planta por congelamento e descongelamento lento e posterior maceração do extrato de flores
de C. cardunculus. O coagulante líquido foi obtido pela precipitação com solventes orgânicos
(acetona e etanol) variando a razão de extrato bruto:solvente (1:1, 1:2, 1:4 e 1:6).
Posteriormente, foi determinado o conteúdo de proteína, o poder coagulante e a
atividade coagulante e proteolítica de cada um dos coagulantes.
O conteúdo de proteína, determinado pelo método de Lowry, variou proporcionalmente
com a razão de extrato bruto:solvente, ou seja, quanto maior foi a razão de solvente utilizado
maior foi a proteína total quantificada. O coagulante obtido pela precipitação com etanol na
razão 1:6 foi o que apresentou uma quantidade de proteínas superior, com 4,468 g/L.
O coagulante obtido pela precipitação com etanol na razão 1:4, apresentou uma razão
entre a atividade coagulante e proteolítica (R= 60,3) superior a todos os outros, assim como
também apresentou um poder coagulante superior, de 1:1624 g/mL, isto é, 1 g deste extrato
enzimático vegetal é capaz de coagular 1,6 L de leite a uma temperatura de 30 °C em 40 min.
As constantes cinéticas de Michaelis-Menten (Km e Vmáx) foram determinadas pela
linearização de Lineweaver-Burk, com posterior ajuste ao modelo não linear. O coagulante obtido
pela precipitação com etanol na razão 1:4 apresentou um valor de Km maior (38 g/L) do que o
coalho obtido pela precipitação com acetona (18 g/L), o que indica uma maior afinidade deste
último em relação ao substrato.
O conjunto de resultados mencionados acima sugere que o coagulante obtido pela
precipitação com etanol na razão 1:4 apresenta um maior potencial para ser usado como
coagulante do leite.


| v
Abstract
The aspartic proteases (EC 3.4.23) are a sub-class of enzymes that are widely
distributed in nature. These enzymes play an important role in the dairy industry as they catalyze
the coagulation of milk and therefore constitute an alternative to animal curds.
The objectives of the current work were the development of a vegetable coagulant,
extracted from the Cynara cardunculus plant and the evaluation of its efficacy in milk coagulation.
The flowers of this plant, commonly denominated as cardo flower posses the aspartic proteases
responsible for milk coagulation. Throughout the decades, these flowers have been used in the
artisanal production of Serra da Estrela sheep cheese.
In this work, the extraction of the proteins was promoted by the rupture of the plant cell
walls through freezing and slow unfreezing followed by the maceration of the C. cardunculus
flower extract. The liquid coagulant was obtained by precipitation with organic solvents (acetone
and ethanol) varying the ratio of crude extract and solvent (1:1, 1:2, 1:4 e 1:6).
Afterwards the protein content, coagulant power and proteolytic and coagulant activity of
each coagulant was determined.
The protein content determined by the Lowry method varied proportionally to the ratio
crude extract and solvent, meaning that the higher the crude extract and solvent ratio used, the
higher protein was quantified. The coagulant obtained by ethanol precipitation with a ratio of 1:6
yield higher quantities of proteins with 4,468 g/L.
The coagulant obtained by precipitation with ethanol in a 1:4 ratio presented a superior
coagulant and proteolytic activity ratio (R= 60,3) as well as a superior coagulant power of 1:1624
g/mL, i.e. 1 gram of this enzymatic vegetable extract is capable of coagulating 1,6 L of milk at a
temperature of 30 °C in 40 min.
The kinetic constants of Michaelis-Menten (Km and Vmáx) were determined by the
linearization of Lineweaver-Burk followed by an adjustment to the non-linear model. The
coagulant obtained by ethanol precipitation with a ratio of 1:4 presented a higher Km value (38
g/L) than the curds obtained by acetone precipitation (18 g/L) which indicates a higher affinity of
the latter towards the substrate.
The set of results above mentionated suggests that the coagulant obtained by etanol
precipitation with a ratio of 1:4 presents higher potential to be used as a milk coagulante.


| vii
Índice
Sumário .................................................................................................................. iii
Abstract ................................................................................................................... v
Lista de figuras ....................................................................................................... ix
Lista de tabelas ....................................................................................................... xi
Lista de abreviaturas ............................................................................................. xiii
CAPÍTULO I – Contextualização e Apresentação do Projeto ..................................... 1
1.1. Introdução................................................................................................................. 3
1.2. Objetivos do Projeto................................................................................................... 5
1.3. Motivação ................................................................................................................. 6
CAPÍTULO II – Revisão Bibliográfica ........................................................................ 7
2.1. Cynara cardunculus L. ............................................................................................... 9
2.2. Enzimas .................................................................................................................. 10
2.2.1. Classificação de enzimas ................................................................................. 11
2.2.2. Cinética Enzimática ......................................................................................... 11
2.2.3. Proteases Aspárticas........................................................................................ 13
2.2.4. Proteases Aspárticas Vegetais .......................................................................... 14
2.2.5. Cardosinas ...................................................................................................... 15
2.2.6. Purificação de proteínas por precipitação com solventes orgânicos ................... 16
2.3. Coagulação Enzimática do Leite .............................................................................. 17
2.3.1. Estrutura da micela de caseína ........................................................................ 18
2.4. Enzimas coagulantes de origem vegetal. .................................................................. 19
CAPITULO III – Materiais e Métodos ...................................................................... 21
3.1. Origem dos extratos de Cynara cardunculus ............................................................ 23
3.2. Extração e Purificação das proteases aspárticas....................................................... 23
3.3. Determinação do conteúdo em proteínas ................................................................. 24

3.4. Determinação da atividade coagulante ..................................................................... 25
3.5. Determinação da atividade proteolitica ..................................................................... 25
3.6. Determinação do poder coagulante .......................................................................... 26
3.7. Caracterização cinética ............................................................................................ 26
CAPITULO IV – Resultados e Discussão .................................................................. 27
CAPÍTULO V – Conclusões ..................................................................................... 37
CAPÍTULO VI – Recomendações ............................................................................. 41
Referências Bibliográficas ..................................................................................... 45
Anexos ................................................................................................................... 51

| ix
Lista de figuras
Figura 1- Inflorescência de C.cardunculus. .............................................................................. 9
Figura 2- Aspecto do extrato de C.cardunculus aquando da sua recepção. ............................ 23
Figura 3- Coalhos obtidos por precipitação com acetona na razão 1:1, 1:2, 1:4 e 1:6 (da
esquerda para a direita) e ressuspensão em tampão citrato de sódio a pH 6. .......................... 29
Figura 4- a) Precipitado obtido para o sistema extrato bruto:etanol na razão 1:1, após 12 h em
repouso. b) Coalhos obtidos por precipitação com etanol na razão 1:2, 1:4 e 1:6 (da esquerda
para a direita) e ressuspensão em tampão citrato de sódio a pH 6. ......................................... 30


| xi
Lista de tabelas
Tabela 1- Nomenclatura e classificação de enzimas .............................................................. 11
Tabela 2- Concentração de proteínas dos coalhos líquidos obtidos pela precipitação com
acetona e etanol variando as razões de extrato bruto:solvente. ................................................ 31
Tabela 3- Atividades coagulante e proteolítica, razão entre as atividades (R) dos coalhos obtidos
pela precipitação com acetona, variando a razão de extratobruto:acetona. ............................... 32
Tabela 4- Atividades coagulante e proteolítica, razão entre as atividades (R) dos coalhos obtidos
pela precipitação com etanol, variando a razão de extratobruto:etanol. .................................... 33
Tabela 5- Resultados do poder coagulante dos coalhos líquidos de origem vegetal obtidos pela
precipitação com acetona e etanol variando as razões de extrato bruto:solvente. ..................... 34
Tabela 6- Parâmetros cinéticos (Km e Vmáx) determinados para os coalhos obtidos pela
precipitação com acetona na razão 1:6 e pela precipitação com etanol na razão 1:4. .............. 35


| xiii
Lista de abreviaturas
a.a. Aminoácido
Abs Absorvância
AC Atividade coagulante
Ala
Alanina
AP Atividade proteolítica
Asp Ácido aspártico
BSA Albumina sérica de bovino
CaCl2 Cloreto de cálcio
CuSO4. 5H2O Sulfato de cobre penta hidratado
DOP Denominação de Origem Protegida
E
Enzima
ES Complexo enzima substrato
Glu Glutamina
GMP Glicomacropeptido
Km Constante cinética de Michaelis-Menten
Leu Leucina
Na2CO3 Carbonato de sódio
NaOH Hidróxido de sódio
P Produto
p Peso
PAs Proteases aspárticas
Phe Fenilalanina
S Substrato
t Tempo
TCA Trichloroacetic acid (ácido tricloroacético)
Tyr Tirosina
UAC Unidade de atividade coagulante
UAP
Unidade de atividade proteolitica
V Volume
Val Valina

Vmáx
Velocidade máxima de reacção

| 1
CAPÍTULO I – Contextualização e Apresentação
do Projeto
CAPÍTULO I – Contextualização e Apresentação do Projeto


| 3
1.1. Introdução
No processo de produção de queijo é necessária uma etapa de coagulação do leite, para
que ocorra a passagem do leite do estado líquido para estado sólido. Nesta etapa, ocorre a
desestabilização das caseínas do leite pela ação de enzimas proteolíticas coagulantes. Estas
enzimas são capazes de hidrolisar a cadeia de aminoácidos da κ-caseína, mais especificamente
a ligação peptídica existente entre as unidades 105 (fenilalanina) e 106 (metionina). Os dois
resíduos resultantes desta divisão são a para- κ-caseína, insolúvel, hidrofóbico, constituído pelos
aminoácidos 1-105 que permanece associada à micela de caseína e o resíduo
glicomacropeptido (GMP), solúvel, hidrofílico, constituído pelos aminoácidos 106-169. No
segundo passo, não enzimático, a estrutura micelar residual precipita na presença de cálcio,
formando o paracaseinato de cálcio. A coagulação, tem início quando cerca de 80 a 90 % da κ-
caseína for afetada, nesta fase as micelas de paracaseína começam a agregar (Almeida, 2006;
Alvarenga, 2008; Chitpinityol e Crabbe, 1997).
A renina é a enzima mais usada na produção de queijo, estando presente no coalho de
animais ruminantes. No entanto, a escassez de matéria-prima, alto preço e crescimento da
produção mundial de queijo, impulsionou a procura de substitutos de qualidade semelhante.
(Kumar et al., 2005; Nelson, 1975). Segundo Hashem (1999), um substituto adequado deve
apresentar atividade coagulante elevada e atividade proteolítica reduzida de forma a minimizar a
dissolução do coágulo.
Os substitutos de coalho presentes no mercado incluem, quimosinas produzidas por
fermentação, coagulantes fúngicos, extratos de plantas e coalhos de outros animais. Devido à
sua aplicação na indústria de laticínios, para coagulação do leite, diversos estudos tem sido
efetuados na pesquisa de proteases de origem microbiana (Cavalcanti et al., 2004; Hashem,
1999; Kumar et al., 2005), animal (Kumar et al., 2006) e de plantas (Ahmed et al., 2009;
Raposo, Domingos, 2008; Silva, Malcata, 2005).
A utilização das proteases de origem vegetal como coagulantes do leite tem despertado
interesse, uma vez que estas enzimas podem substituir o coalho de origem animal e a
quimosina recombinante, dado que a utilização destes produtos está a ser questionada pelas
entidades reguladoras da indústria dos laticínios de diversos países (Sousa, 2002).

Neste projeto irá ser desenvolvido um coagulante vegetal, extraído da espécie Cynara
cardunculus L., vulgarmente conhecida por flor de cardo, que é uma matéria-prima essencial à
indústria do queijo da serra da Estrela.
Esta planta cresce espontaneamente em Portugal Continental, na região centro e sul,
nos Arquipélagos da Madeira e das Canárias, podendo também ser encontrada nas zonas a Sul
e a Oeste da região Mediterrânica e no Norte de África (Silva, 2011; Abreu, 2009; Soares,
2008).
As flores desta planta podem ser usadas como coagulantes do leite, devido à atividade
proteolítica das suas enzimas constituintes, denominadas por cardosinas.

| 5
1.2. Objetivos do Projeto
Objetivo geral:
Desenvolver uma metodologia através do estabelecimento de determinadas condições
de processamento, de forma a obter um coagulante adequado para aplicação na produção de
queijo.
Este objetivo será concretizado através dos seguintes objetivos específicos:
- Extração das proteases aspárticas, responsáveis pela coagulação do leite, dos extratos de
Cynara cardunculus L.;
- Concentração do extrato enzimático pela precipitação com solventes orgânicos;
- Variação da razão de extrato enzimático:solventes;
- Determinação do conteúdo de proteínas, atividade coagulante e proteolítica e poder coagulante
do extrato enzimático de origem vegetal;
- Caracterização cinética do extrato enzimático.

1.3. Motivação
Este projeto foi atribuído pela Alva Solos, que se encontra integrada na BLC3 que é uma
Plataforma de Desenvolvimento da Região Interior Centro, que tem como missão desenvolver e
apoiar projetos empresariais de base tecnológica que se destaquem pela inovação e criatividade,
com o intuito de gerar emprego e valor para a região.
O desenvolvimento deste projeto possibilitou estabelecer condições ótimas de extração
das cardosinas, que conduziu por sua vez à obtenção de um produto de valor acrescentado, o
coalho líquido para queijos. Além disso, a Cynara cardunculus L. é uma espécie vegetal
característica desta zona, pelo que este projeto permitiu a valorização dos recursos da região.
De salientar ainda que o desenvolvimento de coagulantes líquidos vegetais tem elevada
importância, pois os queijos com denominação de origem protegida (DOP), como é o caso do
“Serra da Estrela”, apenas podem ser produzidos usando a Cynara cardunculus L. como agente
coagulante. Portanto, o desenvolvimento destes coagulantes permitirá a adaptação da produção
artesanal para uma produção semi-industrial, assim como poderá despertar o interesse de
produtores industriais neste tipo de coagulante.
Posto isto, considero que este projeto será uma mais valia para a região e o para o
mercado nacional, pois irá impulsionar a produção deste tipo de coalho.

| 7
CAPÍTULO II – Revisão Bibliográfica
CAPÍTULO II – Revisão Bibliográfica


| 9
2.1. Cynara cardunculus L.
A espécie Cynara cardunculus L, vulgarmente conhecida por flor de cardo (Figura 1),
classifica-se por pertencer à família Asteracea e género Cynara (Silva, 2011).
Esta planta é originária da região Mediterrânea e Norte de África, sendo já conhecida
pelos antigos Egípcios, Gregos e Romanos. Atualmente esta planta cresce espontaneamente em
Portugal Continental, na região centro e sul, nos Arquipélagos da Madeira e das Canárias,
podendo também ser encontrada nas zonas a Sul e a Oeste da região Mediterrânica e no Norte
de África (Silva, 2011; Abreu, 2009; Soares, 2008).
A C. cardunculus é uma planta herbácea perene com um caule erecto, podendo atingir
ciclos de metro e meio de altura, densamente coberto por folhas basais grandes e espinhosas.
Os caules terminam em inflorescências de capítulos globulosos de cor azul violeta, as folhas são
verdes na página superior e brancas na página inferior e muito largas. Quanto às raízes, estas
apresentam-se profundas e espessas, o que lhes permite uma boa extração de água e nutrientes
(Silva 2011, Abreu, 2009).
A flor de cardo desenvolve-se em condições muito severas, em solos pedrosos (bordos
da estrada), possuindo uma grande capacidade de adaptação climática. O ciclo desta planta tem
início com a germinação de sementes, no final do Verão aquando das primeiras chuvas, nos
meses seguintes as plantas desenvolvem-se formando uma roseta de folhas, e passando o
Inverno no estado vegetativo. A floração desta espécie é sazonal, entre os meses de Junho e
Julho (Silva, 2011).
Devido às suas características particulares esta planta é passível de ser utilizada para
diversos fins. Nomeadamente, as suas folhas podem ser consumidas cozidas ou utilizadas para
fins terapêuticos, em perturbações digestivas, redução do colesterol ou, mesmo, doenças de
Figura 1- Inflorescência de C.cardunculus.

fígado, assim como podem ser utilizadas no fabrico de bebidas amargas e licores. As folhas
contêm cianopicrina, taninos, ácidos orgânicos, vitamina A e compostos fenólicos.
Relativamente às flores, estas são tradicionalmente usadas como coagulante do leite
para produção de queijos, em particular aqueles que possuem Denominação de origem
protegida (DOP), devido à atividade proteolítica das enzimas constituintes, denominadas
proteases aspárticas. Estudos realizados confirmam que a maior concentração de substância
coagulante está presente na parte superior do estiolo, estando a atividade coagulante restrita ao
pistilo (Silva, 2011; Abreu, 2009, Soares, 2008; Valentão, 2002).
A utilização das proteases de origem vegetal como coagulantes do leite tem despertado
interesse, uma vez que estas enzimas podem substituir o coalho de origem bovina e a quimosina
recombinante, pois o uso destes produtos está a ser questionado pelas entidades reguladoras da
indústria dos laticínios de diversos países (Sousa, 2002).
Atualmente, esta planta pode também ser aplicada à produção de energia, eletricidade e
biodiesel (Silva, 2011).
2.2. Enzimas
As enzimas são proteínas com atividade catalítica, formadas por longas cadeias de
pequenas moléculas, designadas por aminoácidos, que estão unidos entre si por ligações
peptidícas, segundo um arranjo tridimensional. Estas ligações envolvem uma reação entre o
grupo amino (-NH2) de um a.a. e o grupo carboxilo (-COOH) de outro a.a. adjacente (Pinto,
2008).
Uma enzima pode apresentar diferentes conformações, devido à rotação dos
aminoácidos em torno das ligações covalentes simples. No entanto, apenas na conformação
nativa a enzima mantém o seu máximo potencial ativo, a sua atividade biológica (Pinto, 2008).
Dado que, existe um local na superfície das enzimas, designado por centro ativo, que
permite a ligação do substrato à enzima e o contato com o centro catalítico onde ocorre a
reação, pode-se dizer que estas são específicas para um ou vários substratos (Silva, 2011).
As enzimas são catalisadores, pois aceleraram a velocidade de uma reação reduzindo a
barreira de energia requerida para a transformação de um substrato num produto, sem alterar a
termodinâmica envolvida. Logo, quanto mais baixa for a energia de ativação a fornecer, mais
eficaz será o catalisador (Silva, 2011; Parkin, 2010).

| 11
2.2.1. Classificação de enzimas
A Comissão de Enzimas da União Internacional de Bioquímica foi a responsável pela
criação de um sistema de classificação e nomenclatura de enzimas.
Esta classificação divide as enzimas em seis classes distintas de acordo com o tipo de
reação que catalisam, como apresentado na Tabela 1.
As enzimas apresentam um código constituído por 4 números, precedido das iniciais EC.
O primeiro dígito designa a classe a que a enzima pertence, o segundo dígito indica a subclasse
o terceiro a subsubclasse e por fim o último dígito refere-se ao ordenamento da enzima dentro
da subsubclasse (Pinto, 2008; Silva, 2011).
Tabela 1- Nomenclatura e classificação de enzimas
2.2.2. Cinética Enzimática
A cinética enzimática proporciona uma melhor compreensão do mecanismo das
enzimas. A velocidade de uma reação enzimática pode definir-se como a variação do substrato
consumido ou de produto formado por unidade de tempo e é dependente da temperatura e pH
(Silva, 2011).
Dado que as enzimas são únicas no tipo de cinética que exibem, a interpretação da sua
cinética é fundamental.
Primeiro Dígito Classe de Enzima Tipo de reação Catalisada
1 Oxirredutases Reações de oxirredução
2 Transferases Transferência de grupos entre moléculas
3 Hidrolases Reações de hidrólise
4 Liases Remoção de um grupo de uma molécula
5 Isomerases Reacções de isomerização
6 Ligases Reações de síntese acopladas à hidrólise de uma
molécula de ATP

O modelo de Michaelis-Menten (Equação 1) traduz uma reação enzimática simples que
envolve a transformação de um único substrato (S), pela ação de uma enzima (E), num
complexo de associação simples (ES), resultando num único produto (P).
E + S
k−1
k1
ES kcat E + P (Equação 1)
Nos instantes iniciais da reação a quantidade de produto formada é zero, o que torna
unidirecional o segundo passo da reação. Quando a concentração de substrato é
suficientemente elevada para converter toda a enzima no complexo ES, o segundo passo da
reação torna-se limitante, de modo que a velocidade aproxima-se duma assímptota (velocidade
máxima).
A concentração de substrato necessária para obter metade da velocidade máxima é
definida como a constante de Michaelis-Menten (Km). Portanto, esta constante representa a
concentração de substrato necessária para saturar metade da quantidade de enzima e é
característica de cada enzima (Equação 2).
𝑣0 =𝑉𝑚á𝑥 ∗ [S]
𝐾𝑚 + [S] (Equação 2)
Logo, possibilita o estudo da velocidade da reação da enzima, na formação de produto
por minuto (Parkin, 2010; Costa, 2012).
Dado que a representação de v0 em função de [S] é hiperbólica, a determinação de
Vmáx é pouco exacta, pois este torna-se assimptótico. Assim, é necessário recorrer a formas
linearizadas da equação de Michaelis-Menten., sendo a representação de Lineweaver-Burk
(Equação 3) a mais usada (Evangelista, 2005).
1
𝑣0=
1
𝑉𝑚á𝑥+
𝐾𝑚
𝑉𝑚á𝑥
1
[S] (Equação 3)

| 13
2.2.3. Proteases Aspárticas
As proteases aspárticas (PAs; E.C.3.4.23) representam uma sub-subclasse de proteases,
que hidrolisam ligações peptídicas. Em 1836, Theodor Schwann, identificou a primeira PA no
estômago de animais, designada por pepsina. Com o desenvolvimento da ciência foi possível
identificar enzimas que apresentassem características comuns à classe das proteases aspárticas
em diferentes tecidos e organismos (Sousa, 2002; Pereira, 2007; Soares, 2008). Posto isto, as
PAs foram descritas num vasto grupo de organismos entre vertebrados, plantas, nemátodos,
leveduras, fungos, bactérias e vírus (Abreu, 2009;Soares, 2008).
De modo a permitir uma classificação de todas as proteases identificadas foi criada uma
base de dados de proteases, denominada por MEROPS. Nesta base de dados existem 14
famílias de proteases aspárticas que se encontram agrupadas de acordo com o grau de
homologia das suas sequências de aminoácidos, podendo estas dividir-se em sub famílias
atendendo à percentagem de similaridade da sua estrutura primária. As famílias, por sua vez
estão agrupadas em clãs, de acordo com as suas relações evolutivas e estrutura terciária (Abreu,
2009; Soares, 2008; Pereira, 2007).
Na sua maioria as enzimas desta classe exibem em comum uma atividade proteolítica
máxima a pH ácido, são inibidas pela pepstatina (hexapeptídeo produzido por streptomyces sp.),
mostram especificidade para hidrolisar ligações peptídicas entre resíduos aromáticos, e o
mecanismo catalítico é composto por dois resíduos de ácido aspártico (Asp32 e Asp215) (Abreu,
2009; Sousa, 2002; Pereira, 2007).
Estas enzimas caracterizam-se ainda pela elevada homologia que apresentam, em
termos de sequência e de estrutura. A molécula é constituída por dois domínios homólogos que
se enrolam produzindo uma estrutura terciária composta por dois domínios simétricos, que se
encontram ligados por seis sequências em folhas-β antiparalelas. Relativamente, à estrutura
secundária os dois domínios são semelhantes, compostos maioritariamente por folhas-β,
apresentando um baixo conteúdo em α-hélices. No que diz respeito à estrutura terciária, apesar
da grande homologia estrutural há diferentes estados oligoméricos (Pereira, 2007).
Apesar das características comuns presentes entre as PAs, estas exibem diferentes
propriedades catalíticas e diferentes localizações ao nível das células e tecidos, o que poderá
estar relacionado com os seus variados papéis fisiológicos.

Estas enzimas são responsáveis pela degradação de proteínas (pepsina), pelo
processamento de proteínas precursoras (renina e catepsina D) e poliproteínas (PA do vírus da
imunodeficiência humana). Para além disso, é possível salientar o seu papel em processos
fisiológicos como a regulação da pressão sanguínea (renina), da digestão (pepsina, quimosina e
gastricsina) e estão associadas a condições patológicas importantes, como a SIDA, a malária
(plasmepsinas), o cancro (catepsina D e catepsina E) e a doença de Alzheimer (proteína
precursora β-amilóide e memapsina 2). No entanto, o papel das PAs em plantas, na sua
maioria, continua ainda por caracterizar (Abreu, 2009; Sousa, 2002).
2.2.4. Proteases Aspárticas Vegetais
Um grande número de PAs vegetais foram, até à data, detetadas e/ ou purificadas,
nomeadamente a partir de Gimnospérmicas (pinheiro), Monocotiledóneas (arroz, cevada, milho e
trigo) e Dicotiledóneas (Arabidopsis, Brassica, pepino, cardo, tabaco, tomate e plantas carnívoras
Nepenthes) (Abreu, 2009; Soares, 2008).
Estas enzimas têm sido isoladas principalmente a partir de sementes em dormência, ou
de diferentes partes de sementes em germinação, mas também foram purificadas a partir de
folhas, flores e pólen (Abreu, 2009; Soares, 2008).
No entanto, comparativamente com PAs de outras origens, a informação relativa às PAs
vegetais é ainda muito reduzida, dado que apenas algumas delas se encontram completamente
caracterizadas (Abreu, 2009).
Contudo, sabe-se que a maioria das PAs vegetais possui características típicas da sua
sub-subclasse, nomeadamente a atividade proteolítica máxima a pH ácido e a inibição pela
pepstatina. Estas enzimas podem ser monoméricas, como as que estão presentes em sementes
de arroz, de trigo, de Brassica e folhas de Nicotiana ou heterodiméricas como as que estão
presentes nas flores de Cynara sp. (Soares, 2008).
Relativamente à sua localização subcelular, estudos demonstraram que a maioria das
PAs se encontram armazenadas em vacúolos de armazenamento proteico e em corpos proteicos
(fitepsina, cardosina A) ou são secretadas para o espaço extracelular (cardosina B) (Abreu,
2009; Soares, 2008).
As funções biológicas das proteases aspárticas vegetais são ainda pouco claras, ao
contrário do que sucede com as PAs de outros organismos. Contudo, as PAs vegetais têm vindo

| 15
a ser associadas a funções de processamento e degradação de proteínas, senescência,
respostas ao stress, morte celular programada e reprodução nas células vegetais (Abreu, 2009;
Soares, 2008).
2.2.5. Cardosinas
As cardosinas são proteases aspárticas presentes nas flores de Cynara cardunculus L.,
sendo que até à data, foram isoladas e caracterizadas duas formas de cardosinas, A e B,
responsáveis pela atividade proteolítica (Abreu, 2009; Sousa, 2002; Andrade, 2006).
Estas cardosinas são as que mais contribuem para o conteúdo em proteínas solúveis
totais presentes nos estigmas maduros da flor de cardo, pois juntas correspondem a cerca de
60 % deste conteúdo. Sendo assim, estas são expressas em quantidades pouco usuais, em
relação às restantes PAs de plantas (Abreu, 2009; Andrade, 2006).
Todas as cardosinas identificadas e caracterizadas até ao momento, mostram ser
heterodiméricas e glicosiladas. Estas, como na maioria das outras proteases aspárticas, clivam
preferencialmente péptidos em bandas entre resíduos hidrofóbicos, a cardosina A hidrolisa o
péptido na banda Leu15-Tyr16, Leu17-Val18 e Phe25-Tyr26, a clivagem pela cardosina B foi
encontrada em Glu13-Ala14, Ala14- Leu15, Leu15-Tyr16, Leu17- Val18, Phe24-Phe25 e Phe25
(Pereira, 2007).
Além disso, apresentam diferenças ao nível da sequência aminoacídica, ao nível da
localização histológica e ao nível da especificidade enzimática (Sousa, 2002).
Ao nível da localização histológica e citológica, estudos demonstraram que a expressão
da cardosina A restringe-se ao pistilo da flor, acumulando-se, predominantemente, nos vacúolos
de armazenamento proteico das papilas estigmáticas em menor abundância nos vacúolos das
células epidérmicas do estilete (Abreu, 2009; Sousa, 2002; Soares, 2008; Pereira, 2007;
Andrade, 2006).
Ao contrário da cardosina A que é maioritariamente direcionada para o vacúolo, a
cardosina B é secretada para a matriz extracelular e paredes celulares do tecido de transmissão
do estilete durante a maturação da flor (Abreu, 2009).
A cardosina B, apesar de menos abundante na flor do cardo, apresenta uma maior
atividade proteolítica e menor especificidade em relação à cardosina A (Abreu, 2009; Andrade,
2006).

A comparação das sequências de aminoácidos das cadeias de ambas as cardosinas
indica que são produtos de genes diferentes, embora apresentem um elevado grau de homologia
e como tal pensa-se que tenham surgido por um fenómeno de duplicação genética (Abreu,
2009; Pereira, 2007; Andrade, 2006).
Estudos demonstram que as cardosinas apresentam atividade em pH que varia entre 2
e 7, sendo que a atividade máxima verifica-se para um pH de aproximadamente 5, utilizando a
caseína como substrato. Já em condições de pH alcalino as cardosinas apresentam elevada
instabilidade (Soares, 2008).
Relativamente à temperatura sabe-se que as cardosinas mantêm actividade a 60 °C, no
entanto a estabilidade a temperaturas elevadas é relativamente baixa.
2.2.6. Purificação de proteínas por precipitação com solventes
orgânicos
A precipitação de proteínas de uma solução aquosa concentrada (extrato) é uma técnica
de purificação.
A acetona e o etanol são solventes orgânicos miscíveis em água que possuem a
capacidade de precipitar proteínas. Estes solventes apresentam um valor de constante dielétrica
bastante inferior ao da água, logo ao serem acrescentados à solução aquosa possibilitam que as
interações proteína-proteína prevaleçam sobre o poder de solvatação da água, ocorrendo assim
a precipitação de proteínas.
Esta técnica geralmente é muito dependente da temperatura a que é realizada, pois os
solventes quando são usados a temperaturas negativas são muito eficazes na separação de
proteínas de misturas aquosas. Por outro lado, quando são usados a temperaturas elevadas
podem provocar a desnaturação das proteínas, pois quebram-se as pontes de hidrogénio e
estabelecem-se interacções apolares, importantes na manutenção da conformação proteica
(Leite, 2011; Voet D., Voet J., 2005).

| 17
2.3. Coagulação Enzimática do Leite
A coagulação enzimática da proteína do leite pode ser dividida em duas fases e é
efectuada por meio de enzimas de origem animal, vegetal e microbiana.
Numa primeira fase, designada por primária ou enzimática, após a adição do agente
coagulante, ocorre a hidrólise da ligação peptídica existente entre a fenilalanina (105) e
metionina (106) da κ-caseína (Phe105-Met106) e a consequente libertação do
glicomacropeptido (GMP) (Almeida, 2006; Alvarenga, 2008).
Sendo assim, esta ação promove a divisão da κ-caseína em dois resíduos, o resíduo
glicomacropeptido (GMP), solúvel, hidrofilico, constituído pelos aminoácidos 106-169, o qual fica
no soro e o resíduo para-κ-caseína, insolúvel, hidrofóbico, constituído pelos aminoácidos 1-105,
o qual permanece ligado as αS1- e β-caseínas e, por ser altamente hidrofóbico e alcalino,
conduz a destabilização das micelas (Alvarenga, 2008).
Quando ocorre a libertação do GMP verifica-se uma redução importante da carga das
micelas de caseína e uma diminuição das forças de repulsão eletrostáticas que, no estado inicial
são responsáveis pele manutenção do estado coloidal, destabilizando as micelas de caseína.
Nesta fase, os fatores relacionados com a atividade enzimática têm maior importância, como a
concentração e o tipo de enzima, a temperatura e o pH (Almeida, 2006).
A segunda fase da coagulação, designada de fase secundária ou de agregação micelar,
tem início quando cerca de 80 a 90 % da κ-caseína for afetada. Nesta fase ocorre a agregação
das micelas de caseína destabilizadas e a formação de uma rede, inicialmente débil, que vai
aprisionando os glóbulos de gordura, a água e os materiais solúveis em água (Almeida, 2006;
Alvarenga, 2008).
Em seguida, as partículas começam a agregar-se em cadeias cada vez maiores
verificando-se uma reorganização das micelas agregadas e formando-se por fim uma rede
proteica, designada de coalhada. Esta fase caracteriza-se por ser muito sensível à concentração
de Ca2+ presente, à temperatura e ao pH (Almeida, 2006; Alvarenga, 2008).
A fase final da coagulação assenta num fenómeno conhecido por sinérese, no qual
ocorre uma expulsão espontânea do soro na sequência do aumento da rigidez do gel (Almeida,
2006; Alvarenga, 2008).

2.3.1. Estrutura da micela de caseína
Para compreender melhor a dinâmica da coagulação é essencial compreender o
comportamento das moléculas de proteína bem como as alterações que se vão verificando ao
longo do processo.
As moléculas de proteína formam complexos que são conhecidos como micelas de
caseína. A sua estrutura é coloidal, a forma pode ser considerada esférica e são compostas por
várias moléculas agrupadas em subunidades (αS1-, αS2-, β-, e κ-caseínas) (Alvarenga, 2008).
A κ-caseína encontra-se na superfície da micela e desempenha um papel fundamental
na estabilidade do coloide, dado que estabiliza as micelas num estado coloidal, devido ao seu
comportamento simultaneamente hidrofóbico e hidrofilico, conferindo-lhe propriedades de
superfície (Alvarenga, 2008).
As αS1-, αS2- e β-caseínas localizam-se no centro hidrofóbico, ligando-se entre si por
ligações fosfato de cálcio, rodeadas por uma camada rica em κ-caseínas, uma vez que estas têm
propriedades de superfície (Alvarenga, 2008).
A estrutura da micela de caseína é sensível a fatores externos, como por exemplo a
presença de enzimas, o pH e a temperatura (Alvarenga, 2008).
Estudos demonstram que à medida que o pH do leite vai descendo, entre 6,7 e 5,9,
observa-se uma dissociação de 40 a 50 % dos minerais da micela e uma solubilização lenta das
caseínas. Para valores de pH inferiores a 5,3, próximos do ponto isoelétricos da caseína (pH
4,6), observa-se a precipitação da caseína (Alvarenga, 2008).

| 19
2.4. Enzimas coagulantes de origem vegetal.
Coagulantes de origem vegetal incluem enzimas de Cynara cardunculus, sete espécies
de papilionoideae (Eriosema shirense, E. ellipticum, E. pauciflorum, E. gossweilleri, E.
psoraleoides, Adenolichos anchietae e Droogmansia megalantha), Solanum dobium fresen e
Centaurea calcitrapa, Cynara scolymus, Calotropis procera, Helianthus annuus, Lactuca sativa,
figueira (Ficus carica), papaia (Carica papaya), abacaxi (Ananas sativa) e sementes de mamona
(Ricinus communis) com capacidade de coagular o leite (El- Sayed et al, 2013).
Muitas espécies de plantas podem dar origem a coagulação do leite, no entanto as
enzimas extraídas têm uma atividade proteolítica relativamente intensa em relação à atividade
coagulante.
As proteases de plantas podem ter um efeito proteolítico desfavorável na produção de
queijo, pois podem gerar sabores indesejáveis. Duas exceções conhecidas são as proteases
extraídas das flores do cardo (Cynara cardunculus e Cynara humilis) e das bagas (Withania
coagulans) que podem ser adequadas para o fabrico de queijo cottage ou queijo macio (Yusef,
2004).
As enzimas utilizadas na produção de queijo devem apresentar um elevado poder
coagulante e baixa atividade proteolítica, isto é, as enzimas devem romper rapidamente a ligação
Phe105-Met106 da κ-caseína, mas devem possuir baixa atividade proteolítica em relação às
restantes caseínas (Brutti et al 2012).
A atividade proteolítica das enzimas exerce grande influência sobre as características
sensoriais do queijo, como sabor e textura, pois para além da hidrólise da ligação Phe105-
Met106 na κ-caseína, outras ligações peptídicas são quebradas a taxas que variam de acordo
com a enzima utilizada. Uma elevada atividade proteolíca pode ser responsável pelo
desenvolvimento de sabores amargos durante a maturação, além de modificações indesejáveis
na textura do queijo (Vasconcelos M. et al 2004).


| 21
CAPITULO III- Materiais e Métodos
3.
CAPITULO III- Materiais e Métodos


| 23
3.1. Origem dos extratos de Cynara cardunculus
Os extratos secos da planta utilizada neste trabalho, nomeadamente a espécie
C.cardunculus, foram cedidos pela empresa Alva Solos. Após a sua receção, os extratos de
C.cardunculus (Figura 2) foram mantidos em local seco e arejado.
3.2. Extração e Purificação das proteases aspárticas
4. Para efetuar a extração, pesou-se 5 g de extrato de C.cardunculus aos quais foram
adicionados 50 mL de tampão citrato de sódio 0,1 mol/L a pH=5,9. Para promover a extração
de proteínas rompeu-se as paredes celulares da planta por congelamento seguido de
descongelamento à temperatura ambiente. Posteriormente o extrato bruto foi homogeneizado e
centrifugado a 3800 g durante 30 min para eliminar o material insolúvel. O sobrenadante foi
recuperado e filtrado.
5. Os sobrenadantes foram concentrados por precipitação com solventes, para remover os
hidratos de carbono solúveis e pigmentos. A 30 mL de extrato enzimático foi adicionado acetona
a -15 °C com agitação suave. Para a operação foram utilizadas as seguintes razões de extrato
enzimático:acetona, 1:1, 1:2, 1:4 e 1:6.
6. Após completa solubilização, as amostras foram mantidas no congelador over night, e de
seguida foram centrifugadas a 3800 g por 10 min.
Figura 2- Aspecto do extrato de C.cardunculus aquando da sua recepção.

7. O sobrenadante foi removido e o precipitado ressuspendido em 30 mL de tampão citrato
de sódio a pH=5,9.
8. Posteriormente, foi adicionado benzoato de sódio a 10 g/L.
9. O procedimento foi repetido utilizando o etanol como solvente, para as razões de extrato
enzimático:etanol de 1:1, 1:2, 1:4 e 1:6.
10.
3.3. Determinação do conteúdo em proteínas
O conteúdo de proteínas foi determinado pelo método de Lowry, cujo princípio é o
desenvolvimento de um complexo de cor azul, devido à reação das ligações peptídicas de
aminoácidos das proteínas com o reagente de Folin Ciocalteau.
Inicialmente, foram medidos 0,6 mL do extrato enzimático aos quais foram adicionados
3 mL da solução C, composta por A (Na2CO3 20 g/L dissolvido em NaOH 0,1 mol/L) e B (CuSO4.
5H2O a 10 g/L e tartarato de Na e K a 20 g/L), numa proporção de 50:1. Após 10 min foram
adicionados 0,3 mL do reagente Folin- Ciocalteau 1 mol/L e deixou-se reagir durante 30 min, à
temperatura ambiente. Por fim, a absorvância foi lida a 750 nm e determinada a concentração
de proteína, usando a reta padrão previamente estabelecida. Os ensaios foram realizados em
triplicado.
Curva de calibração para a determinação de proteínas
Para a preparação da curva padrão, adicionou-se a tubos de ensaio uma solução de BSA
a 2 mg/mL em quantidades de 10, 20, 30, 40, 50 e 60 µL e completou-se com água destilada
até perfazer 0,6 mL.
A cada tubo adicionou-se 3 mL da solução C e deixou-se 10 min à temperatura
ambiente. De seguida, adicionou-se 0,3 mL do reagente Folin- Ciocalteau 1 mol/L. Após 30 min
mediu-se a absorvância a 750 nm. Os ensaios foram realizados em triplicado.

| 25
3.4. Determinação da atividade coagulante
A atividade coagulante (AC) foi determinada de acordo com Dini (2010), com
modificações. Uma solução de 5 mL de leite, cedido por uma queijaria local, com CaCl2 0,01
mol/L foi pré incubada a 35 °C por 10 min. Adicionou-se 0,5 mL da solução enzimática e
iniciou-se a contagem do tempo. A formação do coágulo foi observada enquanto o tubo de
ensaio era agitado manualmente. O tempo em que as primeiras partículas se formaram foi
medido.
Uma unidade de atividade coagulante (UAC) foi definida como a quantidade de enzima
necessária presente em 1 mL de extrato que coagulou 10 mL de substrato em 40 min e foi
calculada pela Equação 4. Nesta, t é o tempo necessário para formação do coágulo, Vleite é o
volume de leite e Venzima é o volume de enzima.
UAC
mL=
2400
t∗
Vleite
Venzima (Equação 4)
3.5. Determinação da atividade proteolitica
A atividade proteolitica (AP) foi determinada de acordo com Merheb et al. (2007), com
modificações. A mistura reacional era composta por 0,4 mL de caseína a 5 g/L em tampão
citrato de sódio 0,1 mol/L pH=5,9 e 0,2 mL de extrato enzimático. A mistura de reacão foi
incubada a 35 °C e ao final de 30 min a reacão foi interrompida pela adição de 1 mL de TCA
(ácido tricloroacético) a 10 %. As amostras foram centrifugadas a 2800 g durante 10 min. Um
controle foi preparado, onde o TCA foi adicionado antes do extrato enzimático. Fez-se a leitura da
absorvância a 280 nm.
Uma unidade de atividade proteolitica (UAP) e definida arbitrariamente como a
quantidade de enzima necessária para causar um aumento de 0,1 na absorvância a 280 nm,
nas condições de ensaio.
A atividade proteolítica foi calculada pela equação 5. Nesta, Venzima é o volume de enzima e
t e o tempo da reacão.
UAP
mL=
∆Abs ∗ 10 ∗ fator de diluição
Venzima ∗ t (Equação 5)

3.6. Determinação do poder coagulante
O poder coagulante foi determinado de acordo com Yusef (2004). Num recipiente com
um orifício de 1 mm, na parte inferior, foram colocados 100 mL de leite e 0,02 g de CaCl2, a
uma temperatura de 30 °C. Adicionou-se 5 mL de extrato enzimático sob agitação e deixou-se
escorrer o leite pelo orifício até que o gotejamento parasse. O tempo desde que se adicionou e
coagulante até que a última gota caiu foi registado. O leite utilizado nos ensaios foi fornecido por
uma queijaria local.
O poder coagulante foi determinado pela equação 6. Nesta, Vleite é o volume de leite,
pcoagulante é o peso do coagulante e t é o tempo.
Poder coagulante =Vleite ∗ 2400
pcoagulante ∗ t (Equação 6)
3.7. Caracterização cinética
Para a determinação das constantes cinéticas, preparou-se várias soluções de caseína a
diferentes concentrações, 10, 20, 30, 50 e 100 g/L. Reagiu-se 0,4 mL de cada uma das
soluções de substrato com 0,2 mL de extrato enzimático à temperatura de 30 °C. A reação foi
interrompida pela adição de 1 mL de TCA a 10 %. As amostras foram centrifugadas a 2800 g
durante 10 min. Recuperou-se os sobrenadantes e fez-se a leitura da absorvância a 280 nm.
Para a curva de calibração, preparou-se uma serie de soluções de GMP em
concentrações de 1, 2, 3, 4, 5, 6, 7, 8, 9 e 10 %.

| 27
CAPITULO IV- Resultados e Discussão
CAPITULO IV- Resultados e Discussão


| 29
4. Extração de proteínas da C. cardunculus
Para promover a extração de proteínas rompeu-se as paredes celulares da planta por
congelamento e descongelamento lento e posterior homogeneização do extrato de flores.
4.1. Precipitação de proteínas com acetona e etanol
Os coalhos líquidos obtidos pela precipitação, com acetona e etanol, dos extratos de
flores de C. cardunculus são apresentados nas figuras 3 e 4, respectivamente. Foram testadas
várias razões de extrato enzimático bruto:acetona/etanol, (1:1, 1:2, 1:4 e 1:6), de modo a
apurar qual seria a técnica mais eficiente para concentração e purificação das enzimas
presentes, investigando assim a ação de solventes orgânicos (acetona e etanol) que são
compostos comummente usados para esta finalidade.
Após a adição do solvente o sistema foi mantido em repouso por 12 h a -15 °C. O
precipitado foi recuperado, sendo que este variou com a razão de extrato bruto:acetona, ou seja,
para as razões 1:1 e 1:2 verificou-se que a quantidade de precipitado recuperado foi mais
reduzida do que para as razões 1:4 e 1:6. Este facto foi evidenciado pela cor dos coalhos
apresentados na figura 3, pois quanto maior a quantidade de precipitado recuperado mais
escuro foi o coalho obtido.
Figura 3- Coalhos obtidos por precipitação com acetona na razão 1:1, 1:2, 1:4 e 1:6 (da esquerda
para a direita) e ressuspensão em tampão citrato de sódio a pH 5,9.

Portanto, a avaliação visual dos coalhos demonstrou que à medida que o valor da
concentração de acetona aumentava, o sobrenadante obtido ficava mais claro e a altura
(quantidade) do precipitado aumentava.
No caso da precipitação com etanol, após a adição do solvente o sistema foi mantido em
repouso over night a -15 °C. O precipitado foi recuperado, sendo que, também variou com a
razão de extrato bruto:etanol. No entanto, para a razão 1:1 a quantidade de precipitado obtido,
após 12 h em repouso, foi insignificante, como se pode verificar na figura 4 a), acabando
mesmo por se dissolver aquando da remoção do sobrenadante. Assim, a não obtenção de
precipitado para a razão 1:1, poderá ser explicada pela desnaturação das proteínas no decorrer
da aplicação desta técnica, pois geralmente é efectuada a temperaturas abaixo dos 0 °C, dado
que a temperaturas mais altas os solventes tendem a desnaturar proteínas. Como tal, a solução
extrato bruto:etanol deve ter sido sujeita a temperaturas acima de 0 °C, conduzindo a uma
precipitação residual de proteína para a razão 1:1.
Para os sistemas restantes foi possível recuperar o precipitado formado, sendo que foi
para a razão 1:6 que se verificou uma quantidade de precipitado mais elevada. Este facto pode
ser comprovado pela cor dos coalhos apresentados na figura 4 b), pois quanto maior foi a
quantidade de precipitado recuperado mais escuro foi o coalho obtido.
a) b)
Figura 4- a) Precipitado obtido para o sistema extrato bruto:etanol na razão 1:1, após 12 h em repouso.
b) Coalhos obtidos por precipitação com etanol na razão 1:2, 1:4 e 1:6 (da esquerda para a direita) e
ressuspensão em tampão citrato de sódio a pH 5,9.

| 31
4.2. Conteúdo em proteína dos coalhos de origem vegetal
O conteúdo em proteínas, determinado pelo método de Lowry, dos coalhos de origem
vegetal obtidos pela precipitação com acetona e etanol, para as várias razões de extrato
enzimático bruto:solvente, encontra-se apresentado na tabela 2.
Tabela 2- Concentração de proteínas dos coalhos líquidos obtidos pela precipitação com acetona e
etanol variando as razões de extrato bruto:solvente
O conteúdo de proteína de uma preparação enzimática está diretamente relacionado
com a probabilidade de encontrar uma maior quantidade de enzimas coagulantes do leite, e por
sua vez, encontrar uma maior quantidade de enzimas implica o aumento do poder coagulante,
que conduzirá a uma diminuição do tempo de coagulação do leite.
A precipitação de proteínas de extrato bruto com acetona apresenta uma concentração
de proteínas em solução inferior à precipitação de proteínas com etanol.
Os resultados apresentados na tabela 2 revelam ainda que a concentração de proteína
total varia proporcionalmente com a razão de extrato bruto:solvente, pois quanto maior foi a
razão de solvente utilizado maior foi a proteína total quantificada. Este facto verifica-se pois a
acetona e o etanol possuem uma constante dielétrica menor que a da água, aumentando assim
a atração entre as moléculas proteicas, conduzindo à precipitação das mesmas.
Claramente o coalho obtido pela precipitação com etanol na razão 1:6 apresenta uma
quantidade de proteínas maior, com 4,468 g/L, do que o coalho obtido pela precipitação com
acetona, com 1,418 g/L.
Razão de extrato bruto:solvente
Proteína total ACETONA (g/L)
Proteína total ETANOL
(g/L) 1:1 0,198 -
1:2 0,301 0,805
1:4 0,901 2,086
1:6 1,418 4,468

4.3. Atividade Coagulante e Proteolítica
Na tabela 3 e 4 encontram-se apresentadas as atividades coagulante e proteolítica dos
coalhos obtidos pela precipitação com acetona e etanol, respectivamente. Os resultados
determinados para a razão entre a atividade coagulante e proteolítica (R) também são
apresentados na tabela 3 e 4.
Tabela 3- Atividades coagulante e proteolítica, razão entre as atividades (R) dos coalhos obtidos pela
precipitação com acetona, variando a razão de extratobruto:acetona
A atividade coagulante foi determinada pela medição do tempo de formação dos
primeiros coágulos, o que não foi possível no caso do coalho obtido pela precipitação com
acetona na razão 1:1, dado que a adição deste coalho ao leite não originou a formação de
partículas em suspensão. Relativamente à atividade proteolítica este coalho apresentou um valor
residual (0,079 UAP/mL), que pode estar associado à clivagem de alguma ligação peptídica,
exceptuando a ligação Phe105-Met106, ou à presença de alguma partícula em suspensão.
Um coalho adequado deve possuir intensa atividade coagulante e baixa atividade
proteolítica para minimizar a dissolução do coágulo, como tal, pela análise dos restantes
resultados é possível constatar que o coalho obtido pela precipitação com acetona na razão 1:6
apresentar maior atividade coagulante (86,44 UAC/mL) e a atividade proteolítica menor (0,841
UAP/mL) foi conseguida para o coalho obtido pela precipitação com acetona na razão 1:2.
A Tabela 3 também mostra os resultados obtidos para as determinações de R para as
várias razões de extrato bruto:acetona. Nota-se que em todas as razões o valor de R foi maior
que 1, ou seja, a atividade coagulante foi maior que a proteolítica.
Razão de extrato bruto:acetona
Atividade Coagulante (UAC/mL)
Atividade Proteolítica (UAP/mL)
R
1:1 - 0,079 -
1:2 32,54 0,841 38,68
1:4 57,74 1,181 48,91
1:6 86,44 1,757 49,20

| 33
Dado que, a razão entre atividade coagulante e proteolitica (R) é usada como um índice
para justificar a adequabilidade de um extrato enzimático, quanto maior for o seu valor, melhor.
Portanto, a análise da tabela 3 revela que o melhor coalho foi o obtido pela precipitação com
acetona na razão 1:6, pois este possui o maior valor de R (49,2).
Tabela 4- Atividades coagulante e proteolítica, razão entre as atividades (R) dos coalhos obtidos pela
precipitação com etanol, variando a razão de extratobruto:etanol
Relativamente aos resultados dos coalhos obtidos pela precipitação com etanol,
apresentados na tabela 4, é possível constatar que o coalho que apresenta maior atividade
coagulante foi o obtido pela precipitação com etanol na razão 1:4 (122,66 UAC/mL) e o que
possui menor atividade proteolítica foi o coalho obtido pela precipitação com etanol na razão 1:2
(0,963 UAP/mL).
No caso dos coalhos obtidos pela precipitação com etanol, também se verifica que para
as várias razões de extrato bruto:etanol o valor de R foi sempre maior que 1, ou seja, a atividade
coagulante foi superior à proteolítica.
A análise da tabela 4 revela ainda que o melhor coalho foi o obtido pela precipitação
com etanol na razão 1:4, pois este possui o maior valor de R (60,32). Portanto, foram
necessários quatro volumes de etanol para um volume de extrato bruto para obter uma boa
precipitação.
Os resultados referentes aos coalhos obtidos pela precipitação com acetona foram
insatisfatórios quando comparados com os dados do etanol, pois utilizando o etanol como
solvente atingem-se valores mais elevados de R.
Razão de extrato bruto:etanol
Atividade Coagulante (UAC/mL)
Atividade Proteolítica (UAP/mL)
R
1:2 38,20 0,963 39,65
1:4 122,66 2,033 60,32
1:6 67,80 1,342 50,51

4.4. Poder Coagulante dos Coalhos de Origem Vegetal
O poder coagulante é um dos fatores mais importantes na produção queijo, em termos
de qualidade e desempenho, dado que expressa a capacidade de coagulação do leite. Os
resultados do poder coagulante de cada um dos coalhos de origem vegetal obtidos pela
precipitação com acetona e etanol, para as várias razões de extrato enzimático bruto:solvente,
encontram-se apresentados na tabela 5.
Tabela 5- Resultados do poder coagulante dos coalhos líquidos de origem vegetal obtidos pela
precipitação com acetona e etanol variando as razões de extrato bruto:solvente
A comparação dos resultados relativos ao poder coagulante dos coalhos obtidos pela
precipitação com acetona (1:595, 1:875 e 1:1416 g/mL) com os resultados relativos ao
conteúdo de proteína das mesmas amostras (0,301, 0,901 e 1,418 g/L) revelaram que quanto
maior o conteúdo de proteínas maior o poder coagulante do coalho. De salientar ainda, que para
o coalho obtido pela precipitação com acetona na razão 1:1, que possui um conteúdo em
proteína de 0,198 g/L (Tabela 2), não foi possível obter o valor do poder coagulante, pois após a
adição deste coalho ao leite não ocorreu a formação de coágulos. Sendo assim, pode-se afirmar
que as proteínas precipitadas pela acetona na razão 1:1 não apresentavam capacidade de
coagulação, ou seja, este coalho não possuía proteases aspárticas ou então possuía-as numa
quantidade residual, incapaz de provocar a coagulação do leite.
No caso dos resultados relativos ao poder coagulante dos coalhos obtidos pela
precipitação com etanol (1:752, 1:1624 e 1:1134 g/mL), não se verifica a mesma tendência
apresentada para os coalhos obtidos pela precipitação com acetona, pois o coalho que
apresenta maior conteúdo em proteína (4,469 g/L) não corresponde ao coalho que possui maior
Razão de extrato bruto:solvente
Poder coagulante ACETONA (g/mL)
Poder coagulante ETANOL (g/mL)
1:1 - -
1:2 1:594,9 1:751,6
1:4 1:875 1:1623,8
1:6 1:1417,6 1:1134,2

| 35
poder coagulante. Logo, um maior conteúdo em proteínas nem sempre implica uma maior
quantidade de enzimas coagulantes, responsáveis pelo aumento do poder coagulante.
A análise da tabela 5 revela ainda que os solventes usados na razão 1:1, não são
capazes de precipitar as proteases aspárticas responsáveis pela coagulação do leite, pois a
adição destes coalhos não induziu qualquer alteração no leite.
Portanto, os resultados, apresentados na tabela 5, para o poder coagulante dos coalhos
enzimáticos indicam que o maior poder coagulante, 1:1624 g/mL, foi o do coalho obtido pela
precipitação com etanol na razão 1:4, isto é, 1 g deste extrato enzimático vegetal é capaz de
coagular 1,6 L de leite a uma temperatura de 30 °C em 40 min.
4.5. Cinética enzimática
A determinação dos parâmetros cinéticos, apresentados na tabela 6, foi conseguida pela
linearização de Lineweaver-Burk, com posterior ajuste a um modelo não linear. Esta
determinação foi efectuada apenas para os dois coalhos que se revelaram mais eficientes, no
caso da precipitação com acetona, o mais eficaz foi o coalho obtido na razão 1:6 e no caso do
etanol foi o obtido na razão 1:4.
Tabela 6- Parâmetros cinéticos (Km e Vmáx) determinados para os coalhos obtidos pela precipitação
com acetona na razão 1:6 e pela precipitação com etanol na razão 1:4
Como pode ser observado na tabela 6, o valor de Km para o coalho obtido pela
precipitação com etanol na razão 1:4 (38 g/L) é maior do que o atingido para o coalho obtido
pela precipitação com acetona (18 g/L), o que indica uma maior afinidade deste último em
relação ao substrato. Isso denota que as proteases do coalho obtido pela precipitação com
acetona na razão 1:6 são menos específicas, pois apresentam mais opções de sítios de hidrólise
na caseína.
Km (gCaseína/L) Vmáx (gGMP/L.min)
Coalho acetona 1:6 18 ± 11 0,239 ± 0,048
Coalho etanol 1:4 38 ± 14 0,327 ± 0,053


| 37
CAPÍTULO V – Conclusões
CAPÍTULO V – Conclusões


| 39
Neste projeto foi feita a avaliação do potencial de obtenção de um coagulante líquido a
partir de extratos de C. cardunculus. Várias estratégias foram definidas, nomeadamente a
precipitação das proteases asparticas, responsáveis pela coagulação do leite, com recurso a
solventes orgânicos (acetona e etanol). Avaliou-se também o efeito da variação da razão de
extrato bruto:solvente na efetividade dos coagulantes obtidos.
Numa fase inicial procedeu-se à extração proteínas, uma vez que rompeu-se as paredes
celulares da planta por congelamento e descongelamento lento e posterior homogeneização do
extrato de flores. Aos extratos brutos obtidos foi adicionado solvente nas razões 1:1, 1:2, 1:4 e
1:6. A avaliação visual das amostras precipitadas com acetona revelou que à medida que a
razão de acetona aumentava, o sobrenadante obtido ficava mais claro e a altura (quantidade) do
precipitado aumentava. Relativamente às amostras precipitadas com etanol, foi possível verificar
a mesma tendência, exceto para a razão 1:1, pois neste caso não foi possível recuperar o
precipitado obtido, dado que foi residual, acabando por ser descartado. A quantidade de
precipitado recuperado foi mais elevada para o etanol na razão 1:6.
Os resultados relativos ao conteúdo de proteína dos coalhos revelam que, as amostras
obtidas pela precipitação com acetona apresentam uma concentração de proteínas em solução
inferior às obtidas pela precipitação com etanol. A maior concentração de proteínas foi
conseguida para o coalho obtido pela precipitação com etanol na razão 1:6, com 4,468 g/L.
A atividade coagulante máxima foi conseguida para o coalho obtido pela precipitação
com etanol na razão 1:4 (122,66 UAC/mL) e a atividade proteolítica mínima (0,841 UAP/mL)
foi conseguida para o coalho obtido pela precipitação com acetona na razão 1:2. No entanto, a
razão entre atividade coagulante e proteolitica (R) foi superior para o coalho obtido pela
precipitação com etanol na razão 1:4.
O coalho obtido pela precipitação com etanol na razão 1:4 atingiu um poder coagulante
superior a todos os outros, pois um grama deste extrato enzimático vegetal foi capaz de coagular
1,6 L de leite a uma temperatura de 30 °C em 40 min. Este coalho apresentou um valor de Km
maior do que o atingido para o coalho obtido pela precipitação com acetona (18 g/L), o que
indica uma maior afinidade deste último em relação ao substrato, ou seja as proteases do coalho
obtido pela precipitação com acetona na razão 1:6 são menos específicas, pois apresentam
mais opções de sítios de hidrólise na caseína.
Assim, os resultados do coagulante obtido pela precipitação com etanol na razão 1:4,
sugerem que este realmente tenha potencial tecnológico para ser aplicado na produção de

queijo, pois exibiu elevada atividade coagulante e baixa atividade proteolítica. O valor de R, usado
como um índice para justificar a adequabilidade de um extrato enzimático, e o poder coagulante
também foram elevados.

| 41
CAPÍTULO VI – Recomendações
CAPÍTULO VI – Recomendações


| 43
No decorrer do desenvolvimento deste projeto revelaram-se algumas necessidades de
eventuais linhas de investigação para projetos futuros.
Como tal, seria pertinente avaliar a influência de outras variáveis, tais como o pH e a
temperatura, no processo de extração das proteases aspárticas, de forma a otimizar o processo
de extração.
No que diz respeito à purificação das proteases aspárticas, seria interessante testar
outras técnicas, tais como a ultrafiltração e/ou técnicas cromatográficas, de forma a obter um
extrato enzimático mais puro.
Também seria importante avaliar os efeitos dos coagulantes obtidos sobre os
parâmetros de textura (dureza, elasticidade e coesividade) e sabor do queijo assim como avaliar
os efeitos de um coagulante vegetal comercial sobre esses mesmos parâmetros, de forma a
efectuar a sua comparação.

No decorrer do desenvolvimento deste projeto revelaram-se algumas necessidades de
eventuais linhas de investigação para projetos futuros.
Como tal, seria pertinente avaliar a influência de outras variáveis, tais como o pH e a
temperatura, no processo de extração das proteases aspárticas, de forma a otimizar o processo
de extração.
No que diz respeito à purificação das proteases aspárticas, seria interessante testar
outras técnicas, tais como a ultrafiltração e/ou técnicas cromatográficas, de forma a obter um
extrato enzimático mais puro.
Também seria importante avaliar os efeitos dos coagulantes obtidos sobre os
parâmetros de textura (dureza, elasticidade e coesividade) e sabor do queijo assim como avaliar
os e coagulante vegetal comercial sobre esses mesmos parâmetros, de forma a efectuar a sua
comparação.

| 45
Referências Bibliográficas
Referências bibliográficas


| 47
Abreu S. (2009). Obtenção e caracterização de linhas celulares de BY-2 a expressar
cardosinas. Tese de Mestrado, Universidade do Minho, Portugal.
Ahmed I. (2009) Characterisation of partially purified milk-clotting enzyme from Solanum
dubium Fresen seeds. Food Chemistry, v. 116, p. 395-400.
Almeida M. (2006) Caracterização da lactação e do leite de ovelhas da raça Assaf.
Universidade Técnica de Lisboa, Portugal.
Alvarenga N. (2008) Introdução da tecnologia de congelação na produção de queijo de
ovelha. Universidade Técnica de Lisboa, Portugal.
Andrade A. (2006) A associação cardosina-quitosano como potencial biomaterial na
prevenção das fibroses peritoneais.Estudo toxicológlco em ratinhos. Universidade de Aveiro,
Portugal.
Brutti C. et al. (2012) Onopordum acanthium L. (Asteraceae) flowers as coagulating agent for
cheesemaking. Food Science and Technology, v. 45, p. 172-179.
Cavalcanti M. et al. (2004) Partial purification of new milk-clotting enzyme produced by
Nocardiopsis sp. Bioresource Technology, v. 93, p. 29-35.
Chitpinityol S., Crabbe M. J. (1997) Chymosin and aspartic proteinases. Food Chemistry,
v. 61, p. 395-418.
Costa P. (2012) Modelo cinético estocástico para a transcrição considerando colisões entre
moléculas de RNA Polimerase. Tese de Mestrado, Universidade Estadual Paulista, Brasil.
Dini C. (2010) PRODUÇÃO, PURIFICAÇÃO E CARACTERIZAÇÃO DA PROTEASE DE
THERMOMUCOR INDICAE-SEUDATICAE N31 E AVALIAÇÃO DE SUA APLICAÇÃO NA
FABRICAÇÃO DE QUEIJO MATURADO. Tese de Doutoramento, Universidade Estadual Paulista,
Brasil.
El- Sayed S. et al. (2013) Purification and Characterization of a Novel Milk-Clotting Enzyme
from Brassica napus Seeds. Australian Journal of Basic and Applied Sciences, v. 7, p. 482-493.
Evangelista A. et al. (2005) Produção e estudo do potencial de hidrólise de uma nova fonte
de enzimas amilolíticas a partir do malte de milho (Zea mays). Revista Brasileira de Produtos
Agroindustriais, v 7, p.1-14.
Hashem A. (1999) Optimization of milk-clotting productivity by Penicillium oxalicum.
Bioresource Technology, v. 70, p. 203-207.

Kumar S. et al. (2005) Extracellular acid protease from Rhizopus oryzae: purification and
characterization. Process Biochemistry, v. 40, p. 1701-1705.
Kumar A. et al. (2006) Purification and characterization of milk clotting enzyme from goat
(Capra hircus). Comparative Biochemistry and Physiology, Part B, v. 145, p. 108-113.
Leite M. (2011) Desenvolvimento e Optimização de uma Metodologia Analítica para a
Determinação de α- e β-Amanitina em urina humana por LC-MS/MS. Tese de Mestrado,
Universidade de Coimbra, Portugal.
Merheb C. et al. (2007) Partial characterization of protease from thermophilic fungus
Thermoascus aurantiacus and its hydrolytic activity on bovine casein. Food Chemistry, v.104, p.
127-131.
Nelson J. H. (1975) Impact o f new milk clotting enzymes on cheese technology. Journal of
Dairy Science, v. 58, p. 1739-1750.
Parkin K. (2010) Enzimas in Damodaran S., Parkin K., Fennema O., p.265-281. Quimica de
Alimentos de Fennema. Brasil.
Pereira A. (2007) Cardosina A como modelo para o estudo da estabilidade conformacional de
proteinases aspárticas. Universidade de Aveiro, Portugal Pinto A. (2008) Efeito da alta pressão na actividade da enzima peroxidase. Tese de Mestrado,
Universidade de Aveiro, Portugal.
Raposo S., Domingos A. (2008) Purification and characterization milk-clotting aspartic
proteinases from Centaurea calcitrapa cell suspension cultures. Process Biochemistry, v. 43, p.
139-144.
Silva R. (2011) Produção de biodiesel por catálise enzimática a partir de óleo de cardo.
Universidade Técnica de Lisboa, Portugal.
Silva R. (2011) Produção de biodiesel por catálise enzimática a partir de óleo de cardo.
Dissertação de Mestrado, Universidade Técnica de Lisboa, Portugal.
Silva S., Malcata F. (2005) Studies pertaining to coagulant and proteolytic activities of plant
proteases from Cynara cardunculus. Food Chemistry, v. 89, p. 19-26.
Soares R. (2008) Estudo da acção anti-tumoral do extracto de ciprosina, isolado de Cynara
cardunculus. Tese de Mestrado, Universidade do Algarve, Portugal.
Sousa A. (2002) Proteinases asparticas em flores de Cynara Humius: Estudos de expressão
durante o desenvolvimento floral. Faculdade de Ciências da Universidade do Porto, Portugal.

| 49
Valentão P. (2002) Limonete, Hiperecão-do-Gêres, Cardo-do-Coalho, Fel-da-Terra -
Metodologias de controlo de qualidade com base na fracção fenólica - Estudos de acção
antioxidante e hepatoprotectora. Universidade do Porto, Portugal.
Vasconcelos M. et al. (2004) EFEITO DO pH DE COAGULAÇÃO DO LEITE E DO TIPO DE
COALHO SOBRE O RENDIMENTO DE MASSA NA PRODUÇÃO DE QUEIJO. Agrociência, v.10, p.
499-502.
Voet D., Voet J. (2005) Techniques de purification des proteins et des acids nucleiques in
Voet D., Voet J., p. 132. Biochimie. França.
Yusef J. (2004) Extracción de Proteasas de Ulex europaeus L. y su potencial utilización como
sustituto de cuajo. Universidad Austral de Chile.


| 51
Anexos
Anexos


| 53
y = 1,650x + 0,068R² = 0,994
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0
0 0,1 0,2 0,3 0,4 0,5 0,6
Ab
s (n
m)
CBSA (g/L)
Figura i- Curva de calibração obtida pelo método de Lowry para a concentração de proteínas.
Tabela i- Absorvâncias dos coagulantes obtidos pela precipitação com acetona, para determinação
da concentração de proteínas.
Razão de extrato
bruto:acetona Absorvância (nm) Média
1:1 0,394 0,374 0,417 0,395
1:2 0,553 0,595 0,548 0,565
1:4 0,825 0,733 0,879 0,812
1:6 0,627 0,683 0,651 0,654
Tabela ii- Absorvâncias dos coagulantes obtidos pela precipitação com etanol, para
determinação da concentração de proteínas.
Razão de extrato
bruto:etanol Absorvância (nm) Média
1:1 - - - -
1:2 0,713 0,736 0,751 0,733
1:4 0,518 0,499 0,480 0,499
1:6 0,540 0,534 0,515 0,530
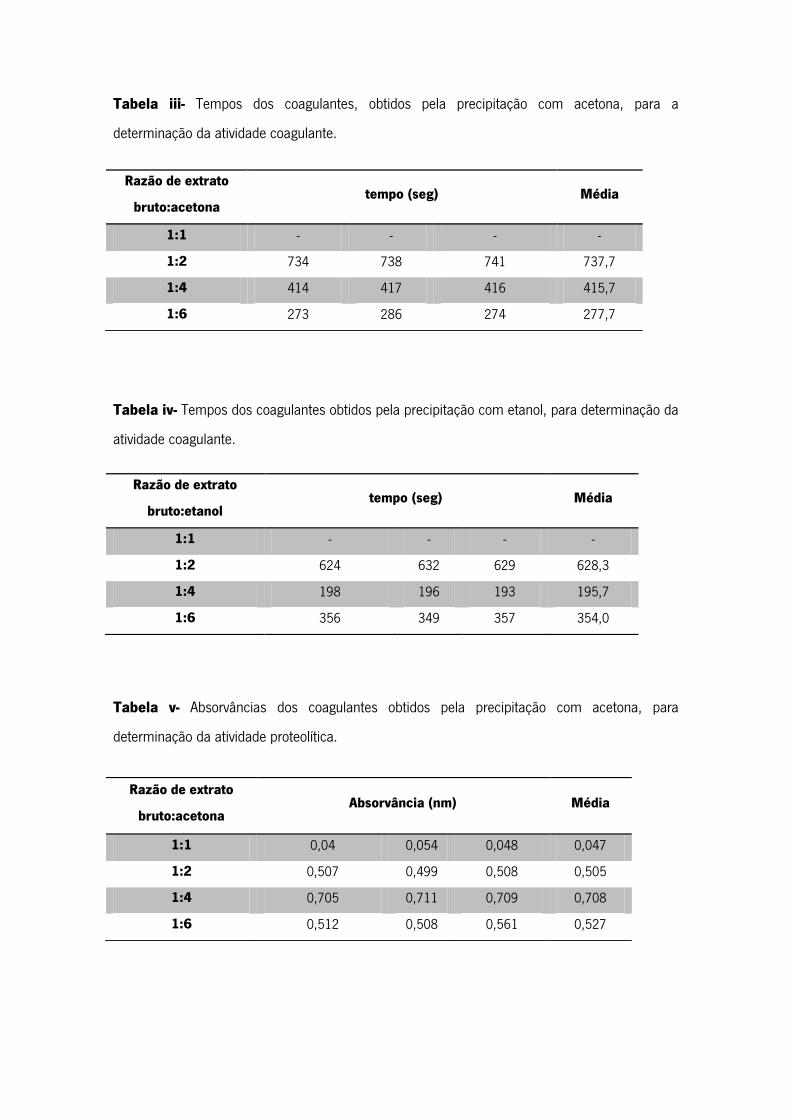
Tabela iii- Tempos dos coagulantes, obtidos pela precipitação com acetona, para a
determinação da atividade coagulante.
Tabela iv- Tempos dos coagulantes obtidos pela precipitação com etanol, para determinação da
atividade coagulante.
Tabela v- Absorvâncias dos coagulantes obtidos pela precipitação com acetona, para
determinação da atividade proteolítica.
Razão de extrato
bruto:acetona tempo (seg) Média
1:1 - - - -
1:2 734 738 741 737,7
1:4 414 417 416 415,7
1:6 273 286 274 277,7
Razão de extrato
bruto:etanol tempo (seg) Média
1:1 - - - -
1:2 624 632 629 628,3
1:4 198 196 193 195,7
1:6 356 349 357 354,0
Razão de extrato
bruto:acetona Absorvância (nm) Média
1:1 0,04 0,054 0,048 0,047
1:2 0,507 0,499 0,508 0,505
1:4 0,705 0,711 0,709 0,708
1:6 0,512 0,508 0,561 0,527
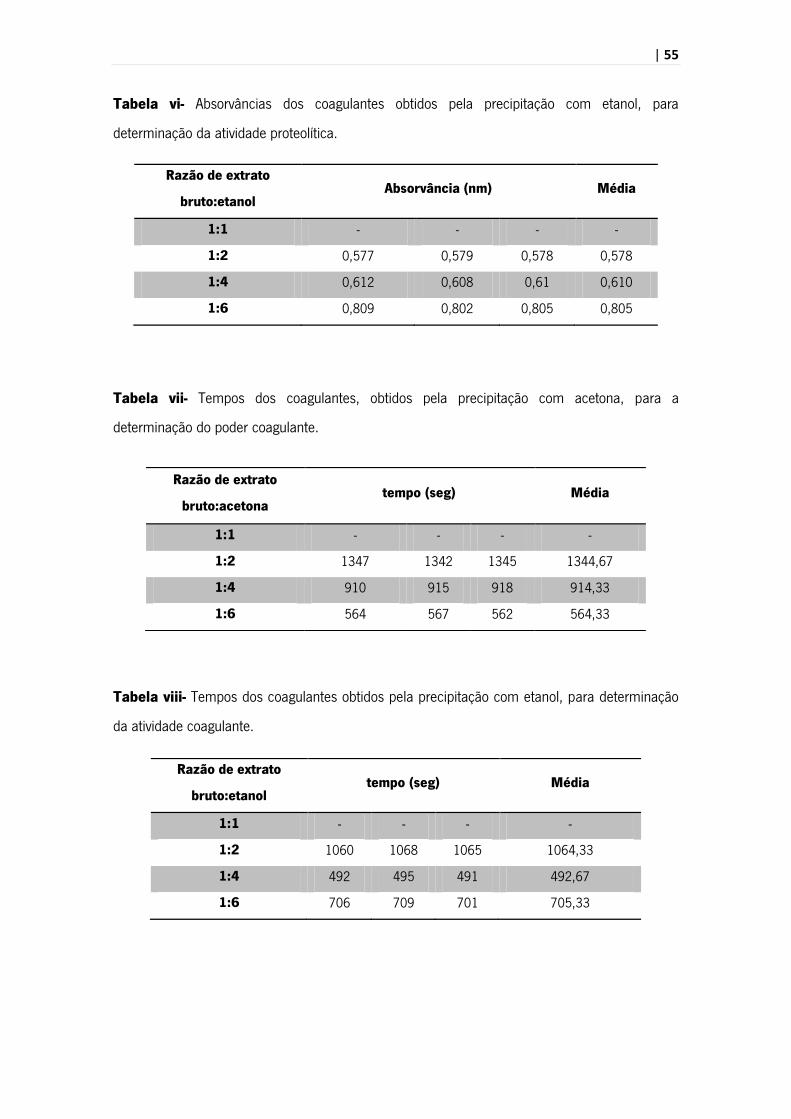
| 55
Tabela vi- Absorvâncias dos coagulantes obtidos pela precipitação com etanol, para
determinação da atividade proteolítica.
Razão de extrato
bruto:etanol Absorvância (nm) Média
1:1 - - - -
1:2 0,577 0,579 0,578 0,578
1:4 0,612 0,608 0,61 0,610
1:6 0,809 0,802 0,805 0,805
Tabela vii- Tempos dos coagulantes, obtidos pela precipitação com acetona, para a
determinação do poder coagulante.
Tabela viii- Tempos dos coagulantes obtidos pela precipitação com etanol, para determinação
da atividade coagulante.
Razão de extrato
bruto:acetona tempo (seg) Média
1:1 - - - -
1:2 1347 1342 1345 1344,67
1:4 910 915 918 914,33
1:6 564 567 562 564,33
Razão de extrato
bruto:etanol tempo (seg) Média
1:1 - - - -
1:2 1060 1068 1065 1064,33
1:4 492 495 491 492,67
1:6 706 709 701 705,33

y = 0,908x + 0,004R² = 0,999
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
0 0,2 0,4 0,6 0,8 1 1,2
Ab
s (n
m)
C GMP (g/L)
Tabela ix- Valores de absorvância do coagulante obtido pela precipitação com acetona na razão 1:6, para caracterização cinética
Tabela x- Valores de absorvância do coagulante obtido pela precipitação com etanol na razão 1:4, para caracterização cinética
t C (min) (g/L)
0 3 6 9 12 15
10 0,057 0,06 0,298 0,23 0,483 0,487 0,719 0,738 0,845 0,761 0,525 0,56
20 0,062 0,061 0,383 0,41 0,7 0,731 1,018 1,069 0,625 0,549 0,791 0,728
30 0,058 0,061 0,57 0,541 0,263 0,254 0,352 0,365 0,458 0,465 0,507 0,532
50 0,065 0,06 0,16 0,168 0,288 0,246 0,335 0,341 0,463 0,479 0,529 0,498
100 0,063 0,052 0,144 0,116 0,207 0,229 0,316 0,33 0,452 0,414 0,524 0,528
t (min)
C (g/L) 0 3 6 9 12 15
10 0,046 0,054 0,141 0,14 0,261 0,271 0,371 0,435 0,551 0,56 0,699 0,729
20 0,048 0,057 0,278 0,259 0,529 0,525 0,839 0,858 0,593 0,586 0,754 0,729
30 0,055 0,068 0,334 0,317 0,685 0,706 0,618 0,598 0,812 0,7911 0,522 0,552
50 0,06 0,056 0,379 0,323 0,461 0,481 0,716 0,78 0,401 0,464 0,624 0,522
100 0,051 0,059 0,335 0,382 0,444 0,475 0,775 0,758 0,46 0,501 0,609 0,6
Figura ii- Curva de calibração de GMP, para determinação dos parâmetros cinéticos













![MARÍLIA OLIVEIRA RIBEIRORibeiro, Marília Oliveira sangramento em usuárias do sistema intra-uterino liberador de levonorgestrel. / Marília Oliveira Ribeiro. Campinas, SP : [s.n.],](https://static.fdocumentos.com/doc/165x107/60e9b2ecfb907352a7417919/marlia-oliveira-ribeiro-ribeiro-marlia-oliveira-sangramento-em-usurias-do.jpg)



