
Mecanismos de expressão genica em...
31
Mecanismo de regulação e expressão em eucariotos Alunos: Jonathan Rodrigues da Silva 8523683 Kaique Dias Galera 8523241 Luiz Otávio Pecorare 7694769 Profº Dr. Julio César Borges Bioquímica II
-
Upload
hoangkhanh -
Category
Documents
-
view
221 -
download
0
Transcript of Mecanismos de expressão genica em...

Mecanismo de regulação e expressão em eucariotos
Alunos:
Jonathan Rodrigues da Silva 8523683
Kaique Dias Galera 8523241
Luiz Otávio Pecorare 7694769
Profº Dr. Julio César Borges
Bioquímica II
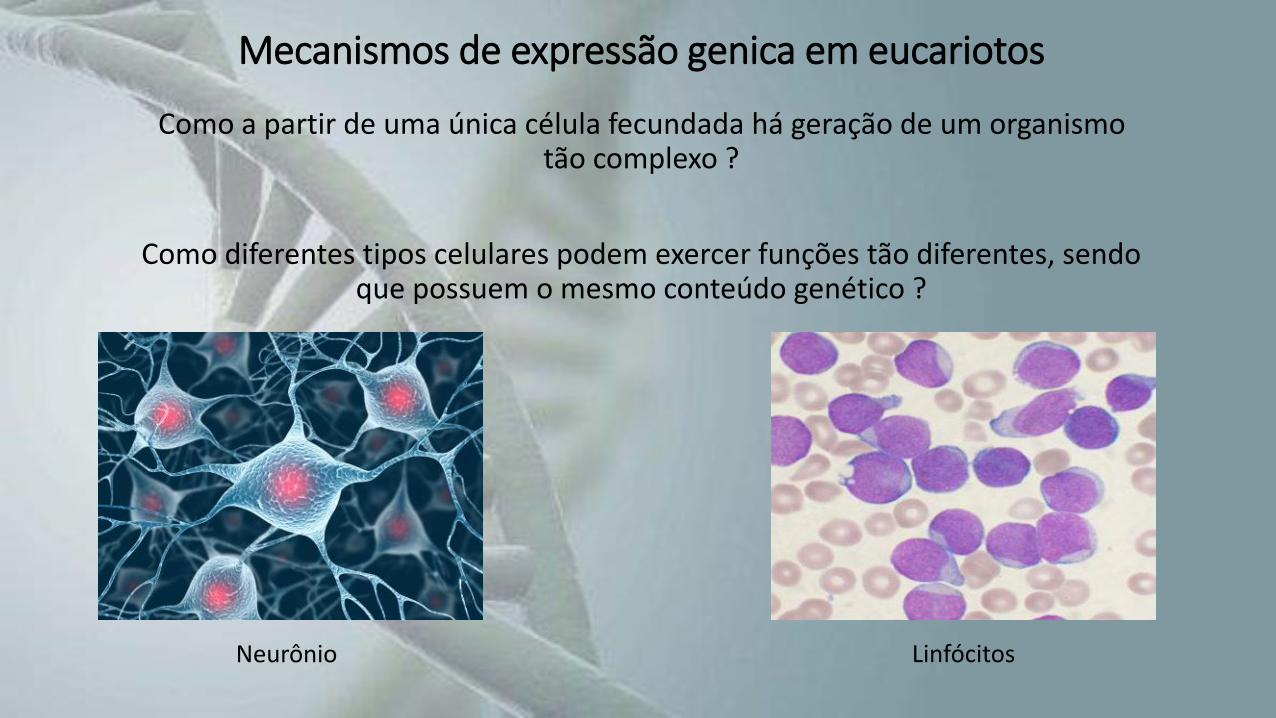
Mecanismos de expressão genica em eucariotos
Como a partir de uma única célula fecundada há geração de um organismo tão complexo ?
Como diferentes tipos celulares podem exercer funções tão diferentes, sendo que possuem o mesmo conteúdo genético ?
Neurônio Linfócitos

Mecanismos de expressão genica em eucariotos
O genoma de um chipanzé e o genoma humano diferem em cerca de 1,5%, entretanto fenotipicamente observa-se espécies completamente diferentes.

Mecanismos de expressão genica em eucariotos
Controle em dois níveis de expressão genica:
• Controle temporal
• Controle Espacial
A regulação genica se dá para todas as proteínas existentes nos indivíduos ?
• Proteínas comuns
• Proteínas específicas
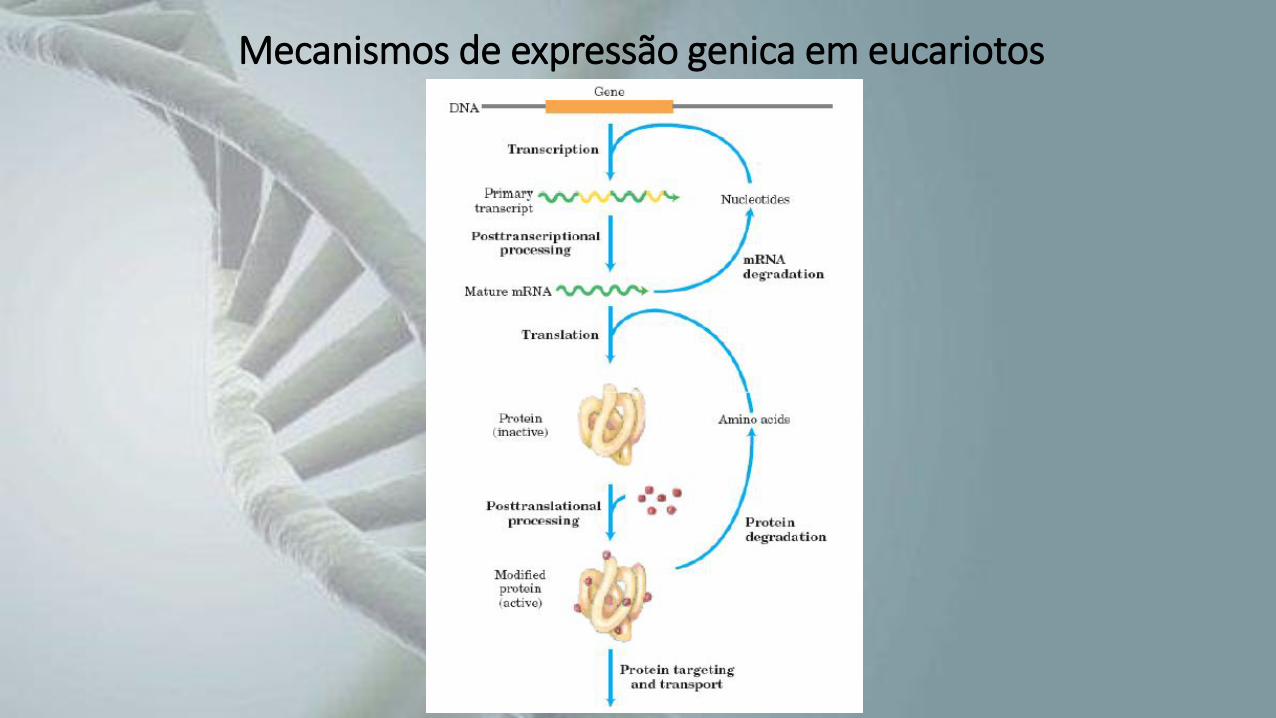
Mecanismos de expressão genica em eucariotos
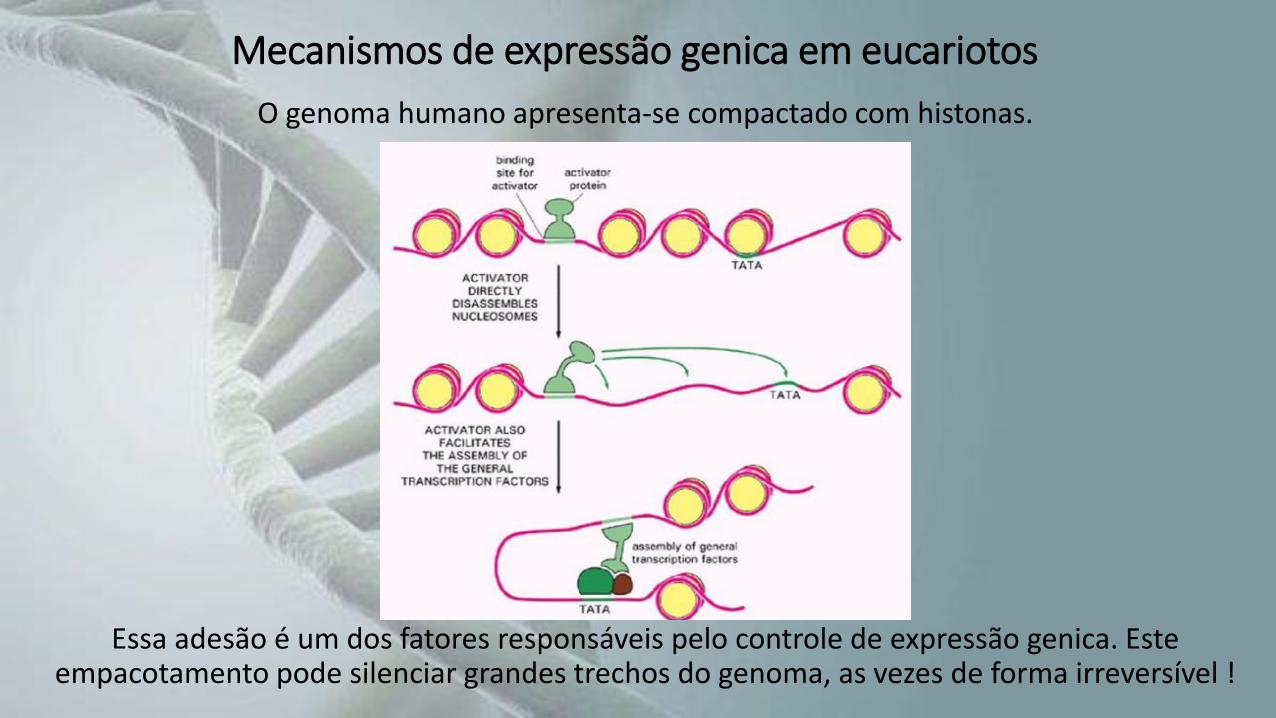
Mecanismos de expressão genica em eucariotos
O genoma humano apresenta-se compactado com histonas.
Essa adesão é um dos fatores responsáveis pelo controle de expressão genica. Este empacotamento pode silenciar grandes trechos do genoma, as vezes de forma irreversível !
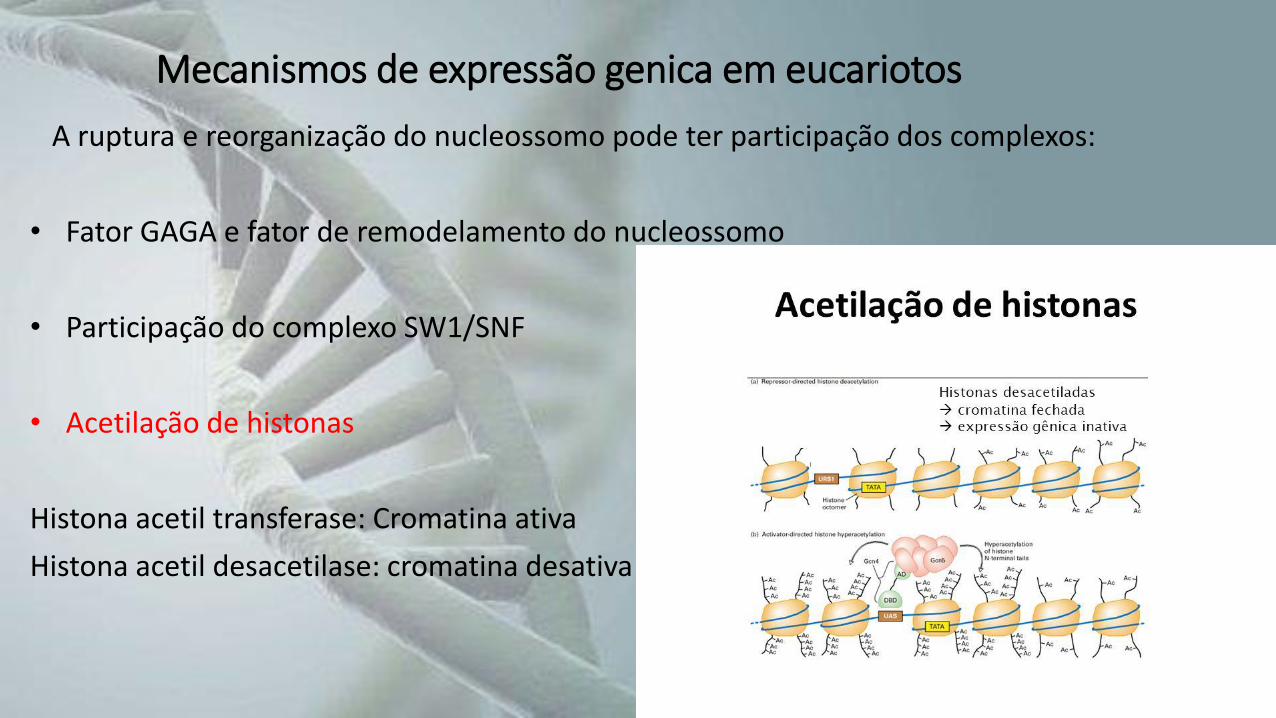
Mecanismos de expressão genica em eucariotos
A ruptura e reorganização do nucleossomo pode ter participação dos complexos:
• Fator GAGA e fator de remodelamento do nucleossomo
• Participação do complexo SW1/SNF
• Acetilação de histonas
Histona acetil transferase: Cromatina ativa
Histona acetil desacetilase: cromatina desativa

Mecanismos de expressão genica em eucariotos
Código das histonas
• Existem diferentes tipos de histonas, cada um com uma quantidade específica de Lisina.
• A quantidade e a posição das lisinas presentes nas histonas altera o fator de expressão gênica.
• Exemplo: Expressão do gene Gal 1

Mecanismos de expressão genica em eucariotos
Regiões controladoras de lócus (LCR)
• Sequencias do DNA essências para a configuração “aberta” da cromatina.
• Capazes de inibir a transcrição, de áreas relativamente grandes, com grande número de genes.
• Uma das regiões mais bem estudadas é a LCR que controla a expressão tecido específica da família da B-globina.
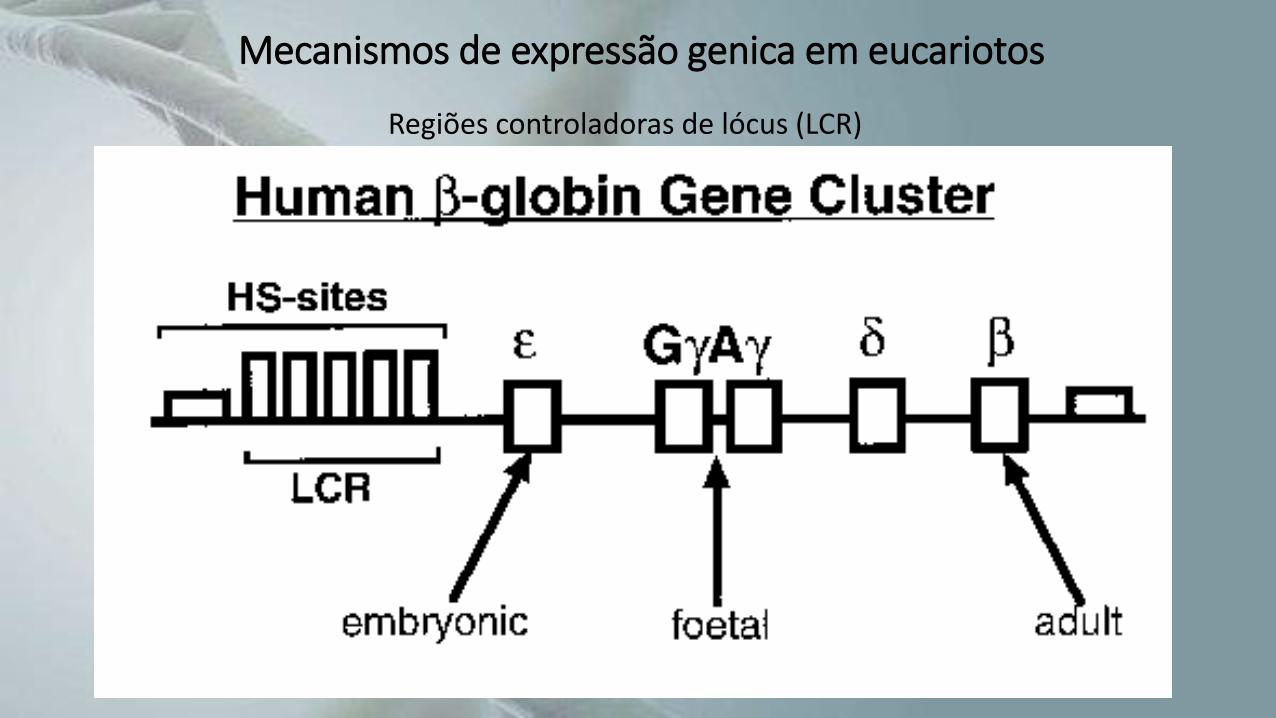
Mecanismos de expressão genica em eucariotos
Regiões controladoras de lócus (LCR)
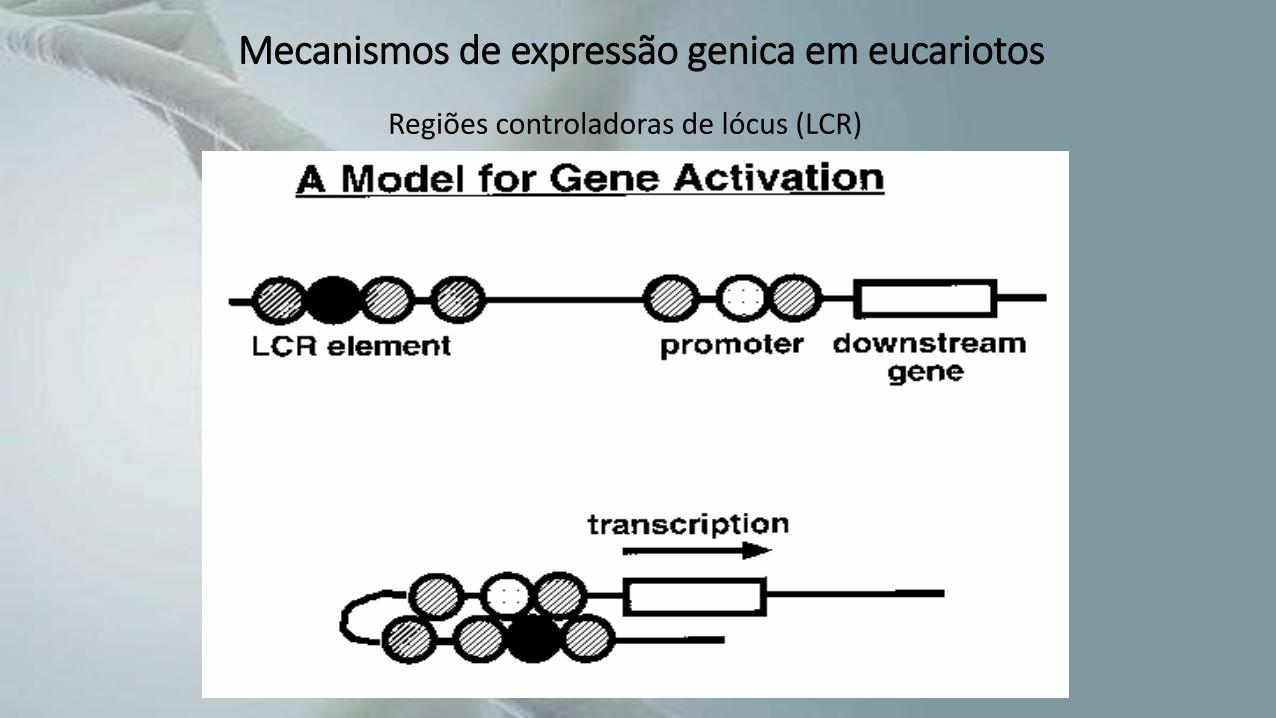
Mecanismos de expressão genica em eucariotos
Regiões controladoras de lócus (LCR)

Metilação e Inatividade Gênica
• Epigenética: qualquer mudança da expressão de umgene sem que ocorra alteração estrutural nasequência de DNA.
• A metilação é o principal fenômeno epigenéticopelo qual um gene é silenciado.
• Ela se dá pela adição covalente de um grupo metila,que é transferido de um doador (S-adenosilmetionina) à citosina presente na estruturado DNA.
• Essa reação é catalisada pela DNA-metiltransferase.
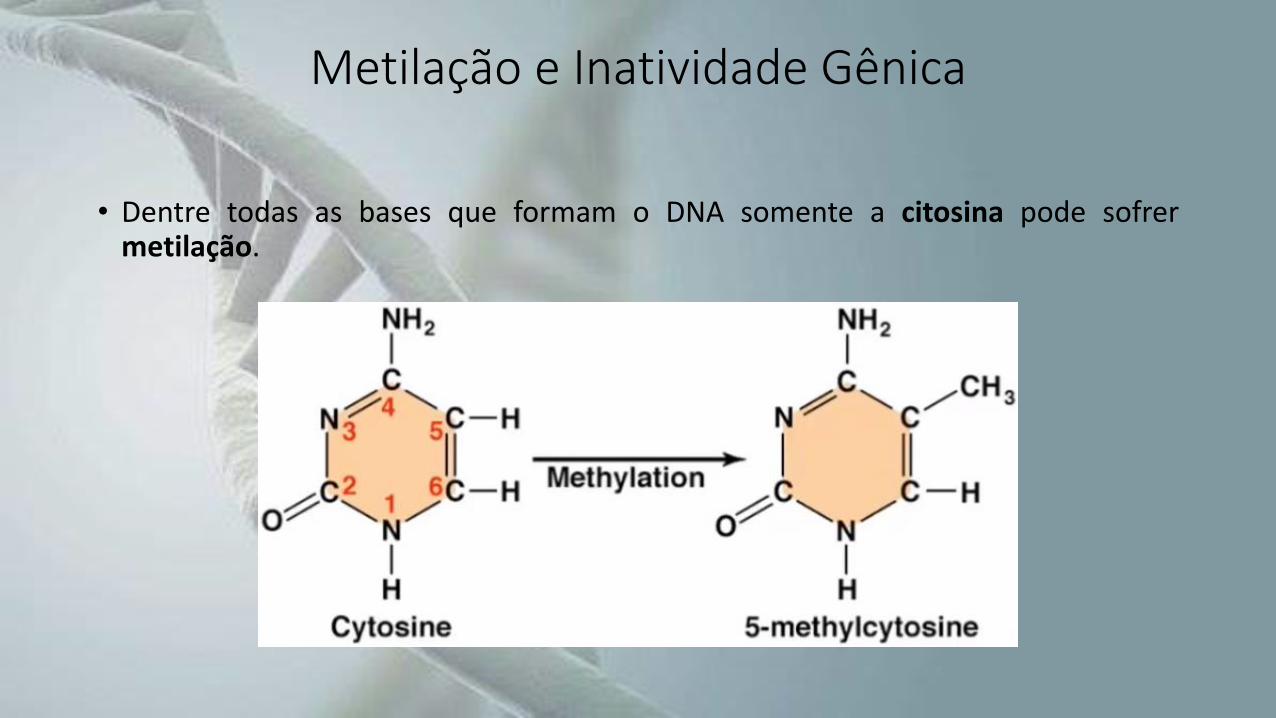
Metilação e Inatividade Gênica
• Dentre todas as bases que formam o DNA somente a citosina pode sofrermetilação.
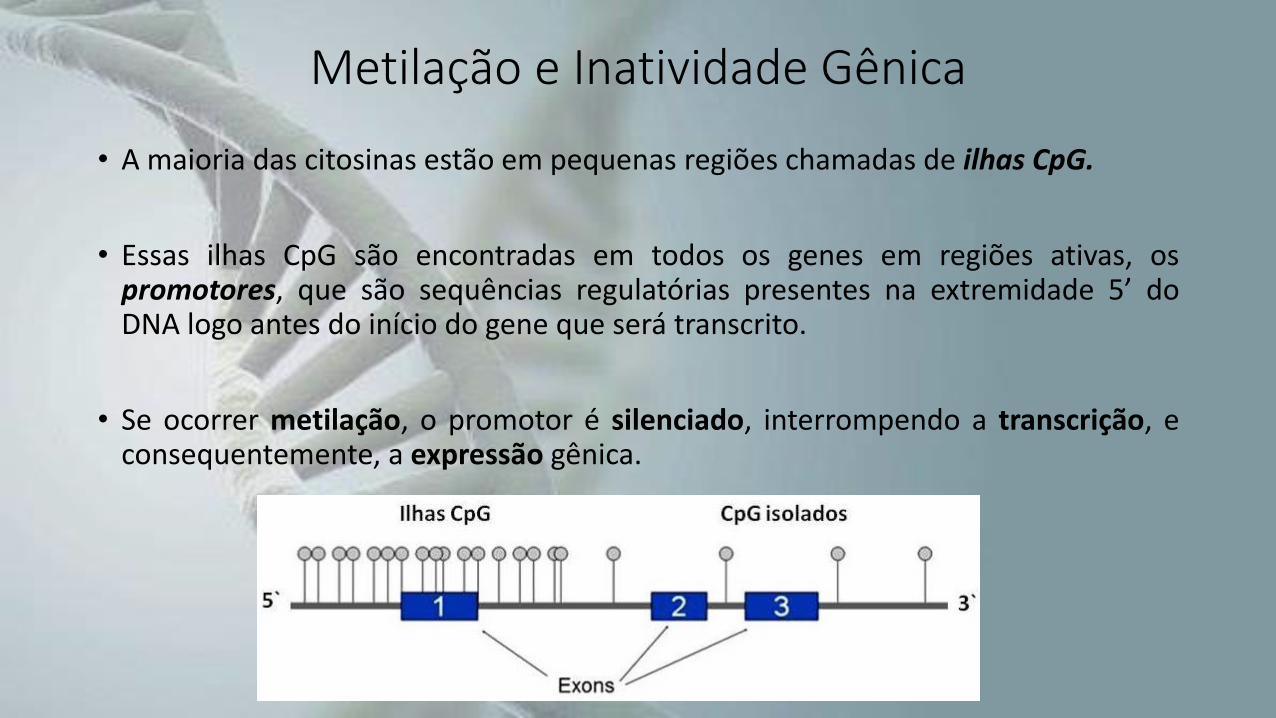
Metilação e Inatividade Gênica
• A maioria das citosinas estão em pequenas regiões chamadas de ilhas CpG.
• Essas ilhas CpG são encontradas em todos os genes em regiões ativas, ospromotores, que são sequências regulatórias presentes na extremidade 5’ doDNA logo antes do início do gene que será transcrito.
• Se ocorrer metilação, o promotor é silenciado, interrompendo a transcrição, econsequentemente, a expressão gênica.
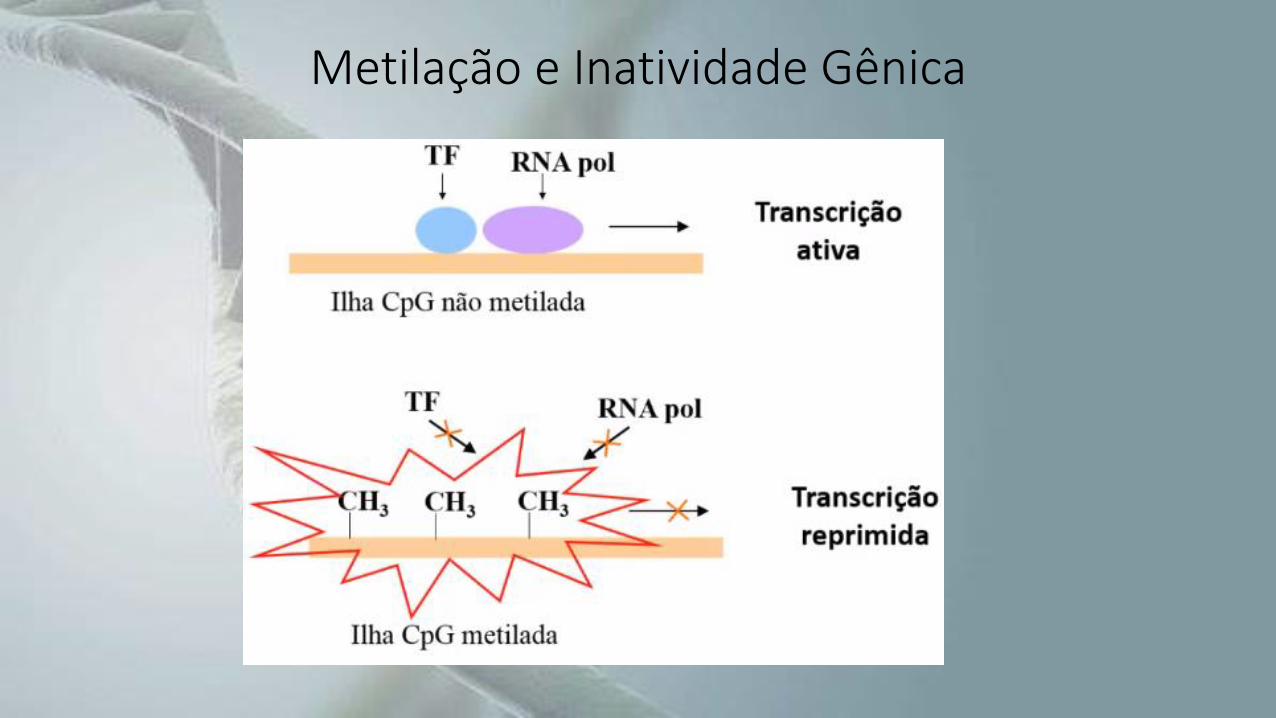
Metilação e Inatividade Gênica
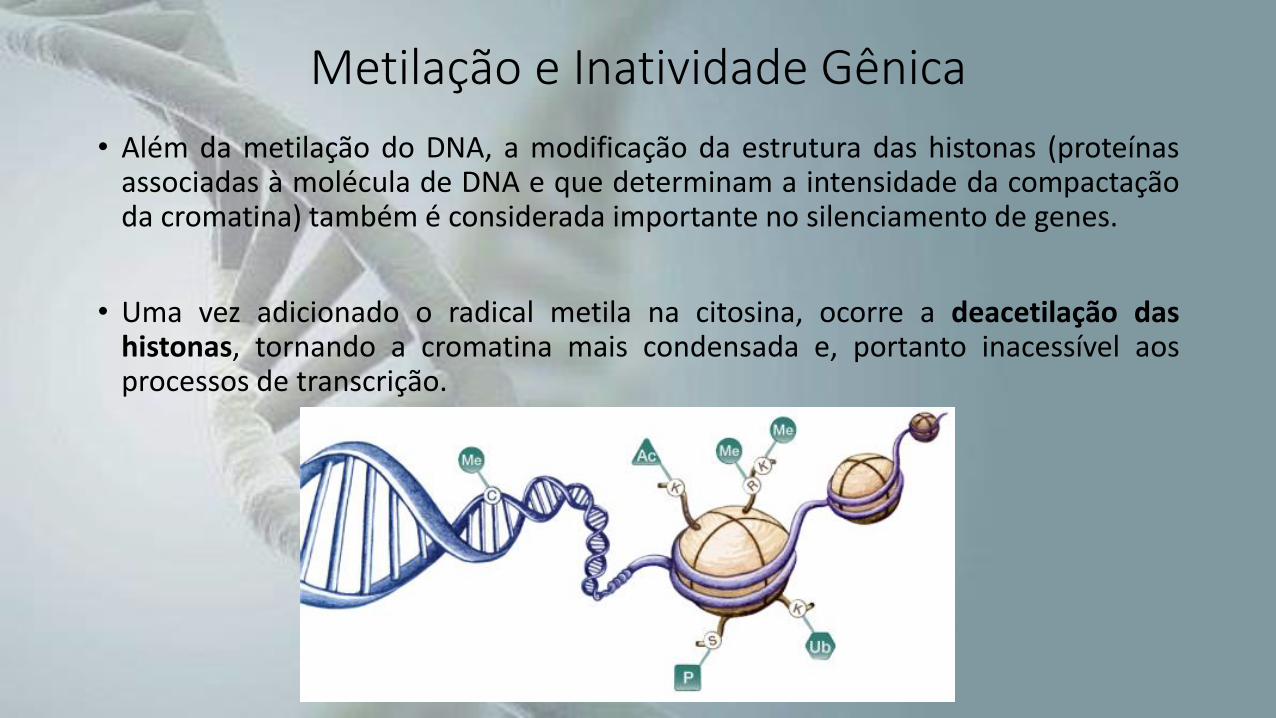
Metilação e Inatividade Gênica
• Além da metilação do DNA, a modificação da estrutura das histonas (proteínasassociadas à molécula de DNA e que determinam a intensidade da compactaçãoda cromatina) também é considerada importante no silenciamento de genes.
• Uma vez adicionado o radical metila na citosina, ocorre a deacetilação dashistonas, tornando a cromatina mais condensada e, portanto inacessível aosprocessos de transcrição.
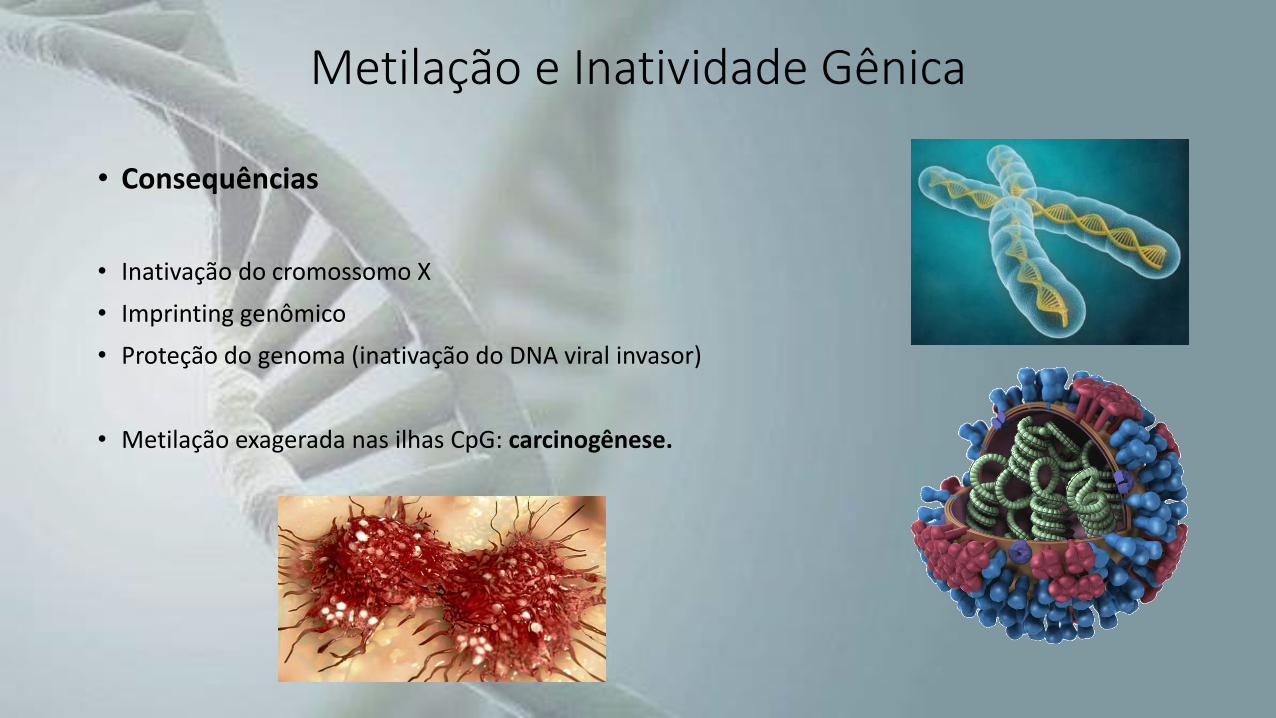
Metilação e Inatividade Gênica
• Consequências
• Inativação do cromossomo X
• Imprinting genômico
• Proteção do genoma (inativação do DNA viral invasor)
• Metilação exagerada nas ilhas CpG: carcinogênese.

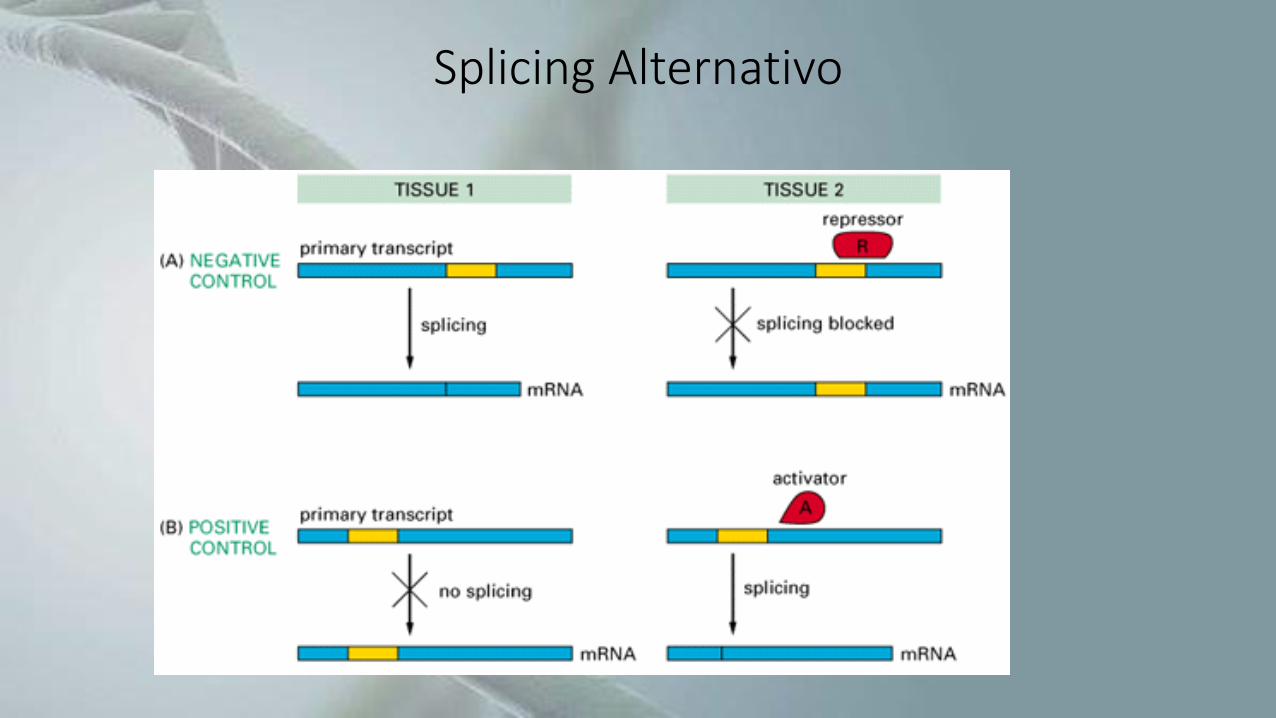
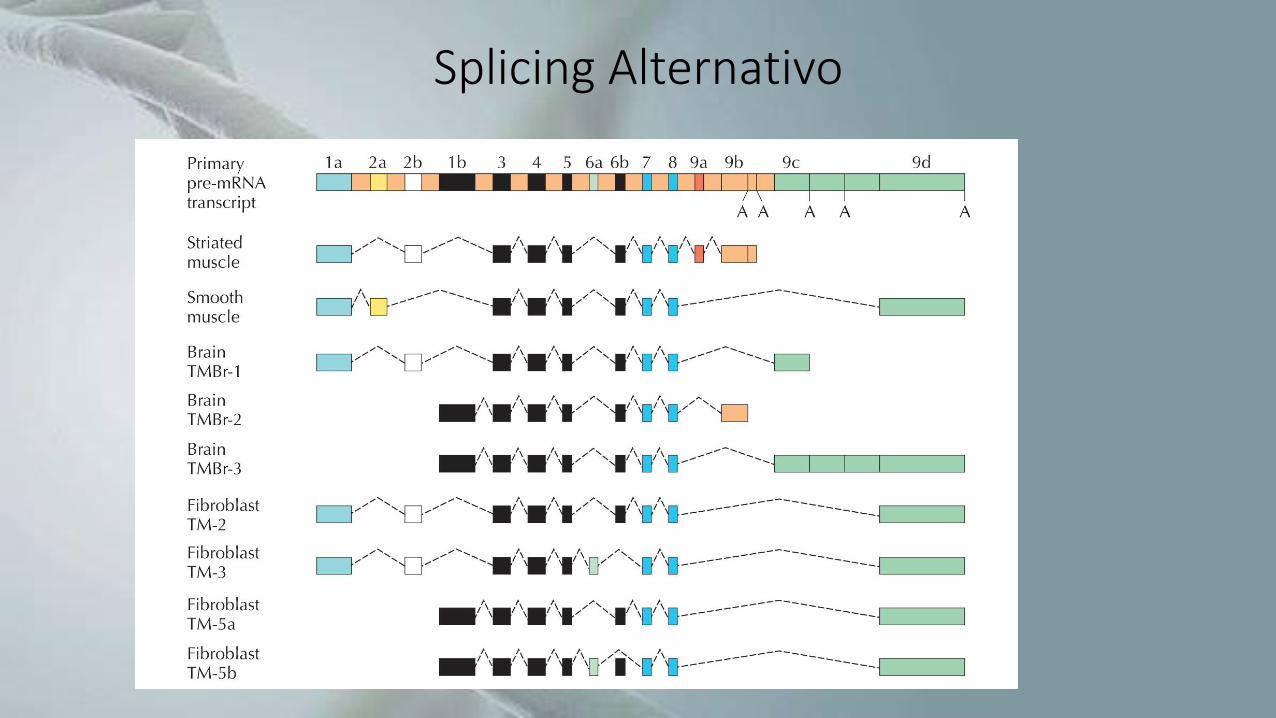
Splicing Alternativo
• Processamento de RNA no qual as sequências denominadas introns sãoremovidas, enquanto as sequências remanescentes (exons) são unidas formandoum RNA maduro, que pode ser mensageiro ou não-codificante.
• Diferentes exons de um mesmo pré-RNA podem ser utilizados na produção dediferentes RNAs maduros, e assim gerar proteínas distintas a partir de um únicogene.
• Pode levar a um grande aumento na diversidade de proteínas.
• Permite que informações específicas de um único gene se modifiquemdependendo de sinais do ambiente, gerando transcritos maduros distintos, econferindo assim uma maior plasticidade à expressão gênica
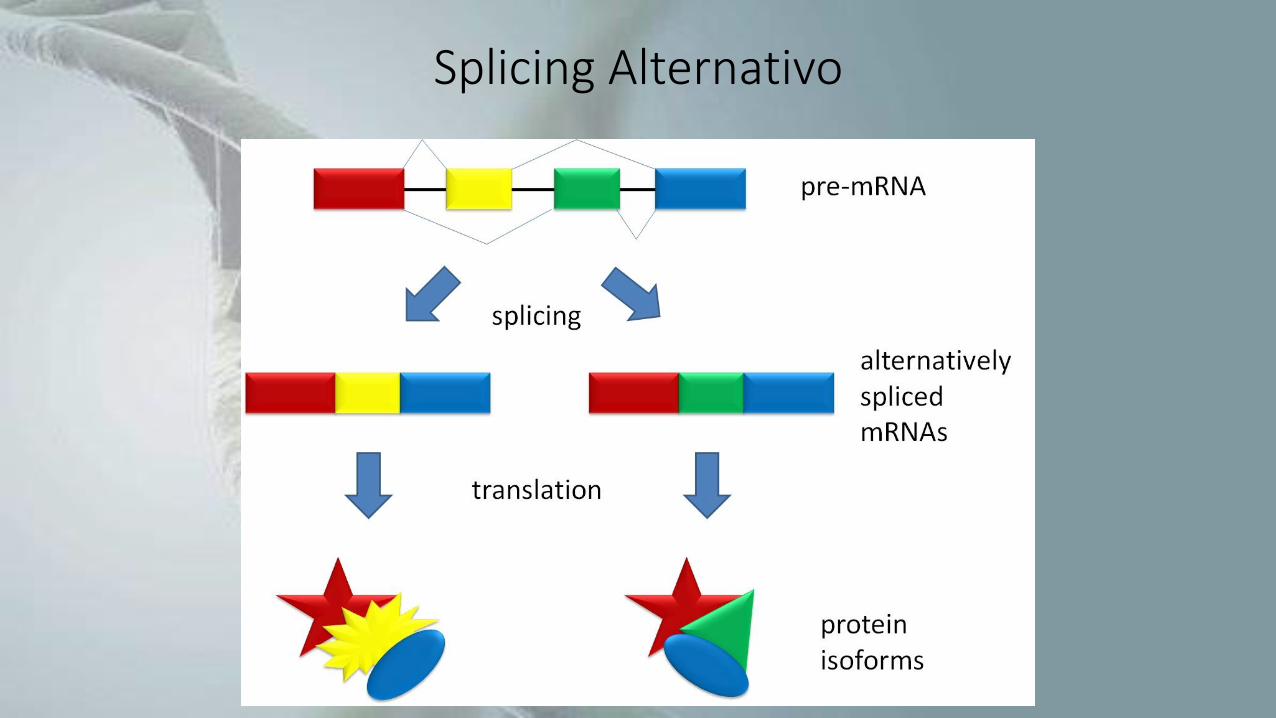
Splicing Alternativo

Splicing Alternativo
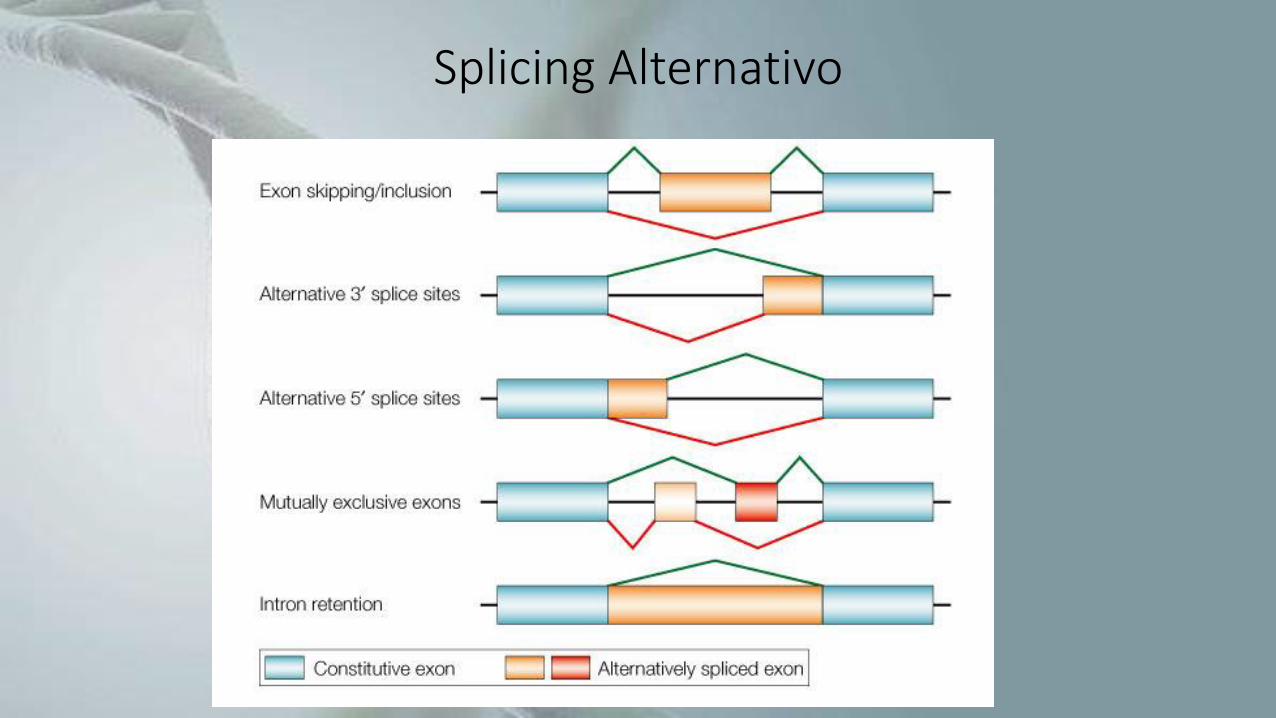
• Existem vários tipos de splicing alternativo, dentre eles os cinco principais são:
• exon skipping (uso alternativo de exon);
• alternative 3’ splice sites (sítios aceptores [3’] alternativos);
• alternative 5’ splice sites (sítios doadores [5’] alternativos);
• mutually exclusive exons (exons mutuamente excludentes);
• intron retention (retenção de intron).
• Além da possibilidade de ocorrer splicing alternativo nas sequências iniciadoras efinalizadoras da transcrição.
Splicing Alternativo
Splicing Alternativo
Splicing Alternativo
Receptores específicos
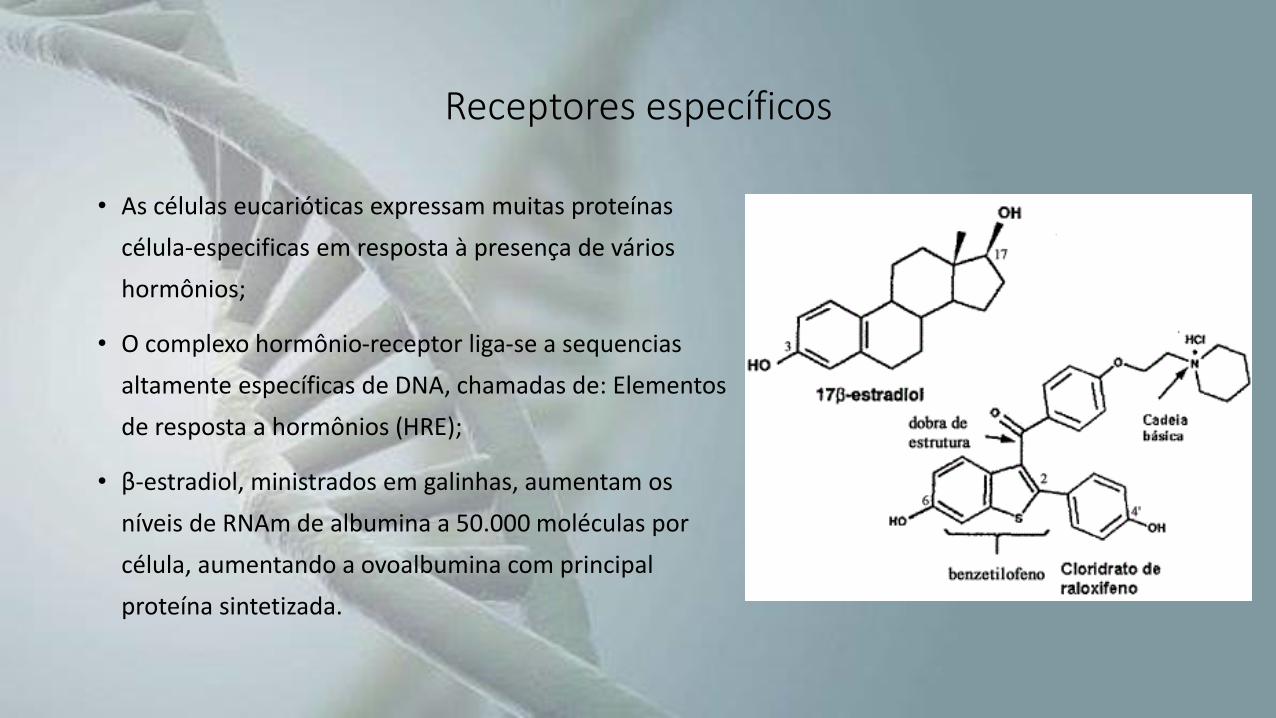
• As células eucarióticas expressam muitas proteínas
célula-especificas em resposta à presença de vários
hormônios;
• O complexo hormônio-receptor liga-se a sequencias
altamente específicas de DNA, chamadas de: Elementos
de resposta a hormônios (HRE);
• β-estradiol, ministrados em galinhas, aumentam os
níveis de RNAm de albumina a 50.000 moléculas por
célula, aumentando a ovoalbumina com principal
proteína sintetizada.

Regulação da Expressão Gênica pelo RNA
• Pode ser feita pelos pequenos RNAs (RNA formado por 20 a 25 nucleotideos),
foram descobertos recentemente mas já se sabe que possuem função
regulatória;
• Seus precursores podem ser RNAm ou fitas paralelas de um RNA viral transcrito;

• A regulação é feita a partir do impedimento da tradução do RNAm ou da indução de sua destruição;
• A interação do mRNA com o RNAm se dá, frequentemente, na região 3’UTR;
• Seus precursores possuem cerca de 70 nucleotídeos e tem forma de grampo;
• Esses RNAs são produzidos a partir da clivagem por endonucleases como Droshae Dicer.
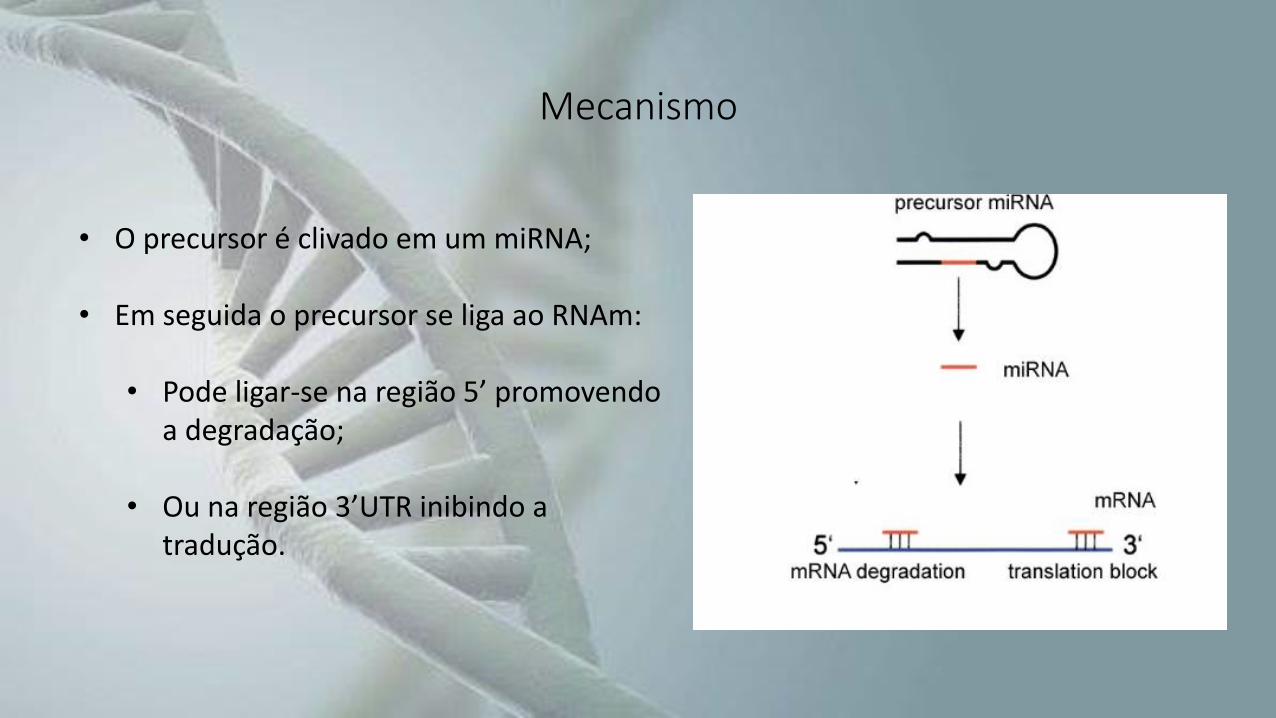
Mecanismo
• O precursor é clivado em um miRNA;
• Em seguida o precursor se liga ao RNAm:
• Pode ligar-se na região 5’ promovendo a degradação;
• Ou na região 3’UTR inibindo a tradução.

Mecanismos pós-traducionais
• Mesmo após sua tradução, a proteína pode estar inativa, necessitando de alguma
modificação estrutural para ativá-la;
• Como exemplo, o tripsinogêneo, a forma inativa da tripsina;
• É necessária a retirada de uma parte da sua cadeia polipeptídica para tornar a
pepsina ativa.
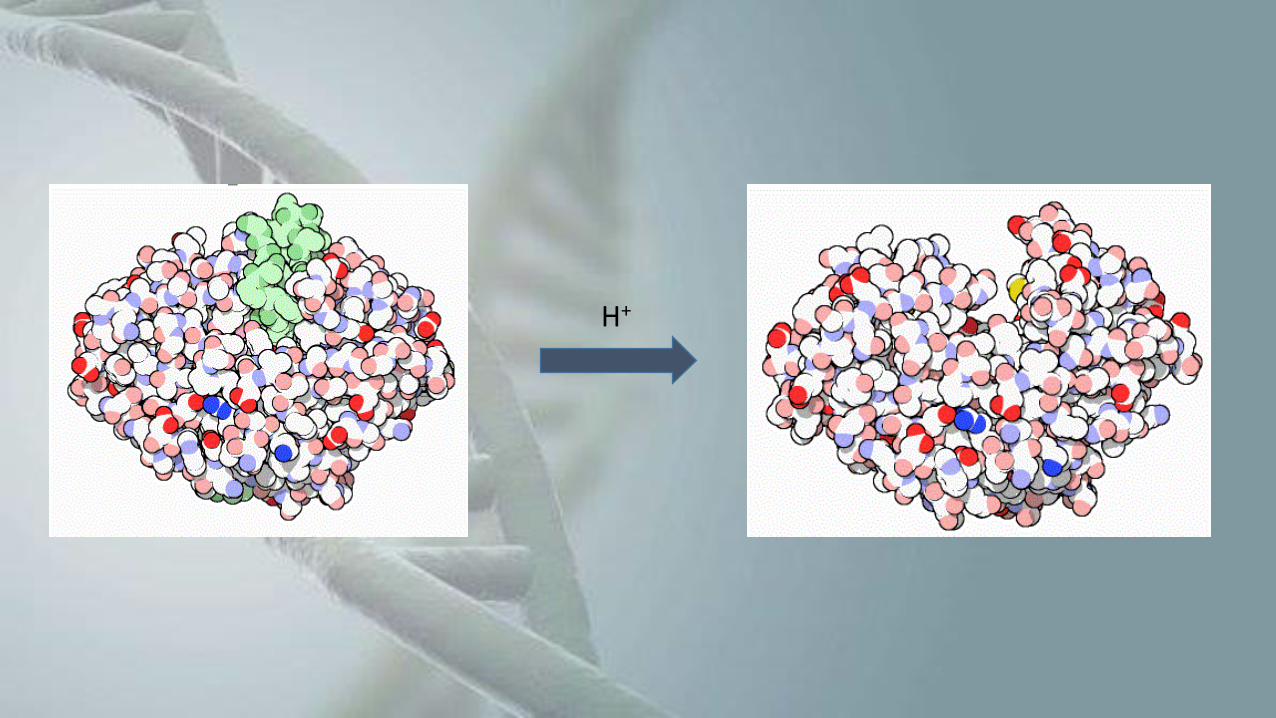
H+

Referências• Disciplina de Genética Humana - Monitora: Scheila Seady. HIPERMETILAÇÃO E
CÂNCER Epigenética. (n.d.), 5–6.
• http://www.rcsb.org/pdb/101/motm.do?momID=12 , acessado em 23/11/2015.
• Tomotani, B. M. (2010). Aspectos evolutivos do splicing alternativo. Revista Da Biologia, 4, 44–49. http://doi.org/10.7594/revbio.04.09
• Hammond S, Bernstein E, Beach D, Hannon G (2000). "An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells". Nature 404 (6775): 293–6.
• Voet, D.; Voet, J.; Pratt, C. W. Fundamentos de bioquímica : a vida em nível molecular. 4. ed. Porto Alegre: Artmed, 2014.
• LEHNINGER, A. L. & NELSON, D. L. & COX, M. M. - Princípios de Bioqumica. Princípios de Bioquímica de Lehninger. Editora: Sarvier, 4ª edição, 2007.

Muito Obrigado!


















