
MINERAÇÃO, POLUIÇÃO SONORA E IMPACTOS NA...
120
UNIVERSIDADE FEDERAL DE MINAS GERAIS PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA, CONSERVAÇÃO E MANEJO DA VIDA SILVESTRE MINERAÇÃO, POLUIÇÃO SONORA E IMPACTOS NA COMUNICAÇÃO ANIMAL Marina Henriques Lage Duarte Tese de Doutorado em Ecologia, Conservação e Manejo da Vida Silvestre / 2015 Orientador: Marcos Rodrigues Co-orientadores: Robert John Young e Renata Sousa-Lima Colaboradora: Nadia Pieretti Belo Horizonte 2015
Transcript of MINERAÇÃO, POLUIÇÃO SONORA E IMPACTOS NA...

UNIVERSIDADE FEDERAL DE MINAS GERAIS
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA,
CONSERVAÇÃO E MANEJO DA VIDA SILVESTRE
MINERAÇÃO, POLUIÇÃO SONORA E IMPACTOS NA
COMUNICAÇÃO ANIMAL
Marina Henriques Lage Duarte
Tese de Doutorado em Ecologia, Conservação e Manejo da Vida
Silvestre / 2015
Orientador: Marcos Rodrigues
Co-orientadores: Robert John
Young e Renata Sousa-Lima
Colaboradora: Nadia Pieretti
Belo Horizonte
2015

Marina Henriques Lage Duarte
MINERAÇÃO, POLUIÇÃO SONORA E IMPACTOS NA
COMUNICAÇÃO ANIMAL
Tese de doutorado apresentada ao
Programa de Pós-graduação em
Ecologia, Conservação e Manejo
da Vida Silvestre da Universidade
Federal de Minas Gerais, como
requisito parcial para obtenção do
título de doutor em Ecologia.
Orientador: Marcos Rodrigues
Co-orientadores: Robert John
Young e Renata Sousa-Lima
Colaboradora: Nadia Pieretti
Belo Horizonte
2015

Esta tese de doutorado foi desenvolvida no Laboratório de Ornitologia, do
Departamento de Zoologia, Universidade Federal de Minas Gerais e no Laboratório de
Bioacústica do Museu de Ciências Naturais, Pontifícia Universidade Católica de Minas
Gerais, em parceria com as seguintes instituições:
Departamento de Ciências Básicas, Universidade de Urbino, Campus Científico
"Enrico Mattei" Urbino - Itália.
Escola de Meio Ambiente e Ciências da Vida, Universidade de Salford
Manchester, Salford - Inglaterra.
Laboratório de Bioacústica (LaB), Departamento de Fisiologia, Universidade
Federal do Rio Grande do Norte.
Apoio financeiro

Dedicado ao meu pai
por guiar todos os meus passos até aqui

“A tarefa não é tanto ver aquilo que
ninguém viu, mas pensar o que
ninguém ainda pensou sobre
aquilo que todo mundo vê.”
Arthur Schopenhauer

Agradecimentos
Esta pesquisa foi concretizada graças ao apoio de diversas pessoas e instituições. O meu
eterno e carinho e gratidão dedico:
Ao meu orientador e amigo, Marcos Rodrigues. Obrigada por me aceitar como sua
aluna, por acreditar em mim e me dar credibilidade para desenvolver esta tese. Nos
momentos de tensão você me transmitiu calma e me ajudou a minimizar os contratempos.
Obrigada pelo apoio e pelas sábias palavras ditas durante estes quase quatro anos. Você foi
parte fundamental deste trabalho.
À Nadia Pieretti. Existem pessoas que surgem inesperadamente em nossas vidas e
iluminam nossos caminhos. Poucas vezes ao longo da minha trajetória profissional conheci
pessoas doces como Nadia Pieretti. Muito obrigada por ter me ensinado grande parte do que
sei hoje sobre os sons naturais. Obrigada por estar disponível a qualquer hora do dia mesmo
quando tínhamos um oceano entre nós. Nos momentos mais difíceis você foi sensível,
compreensiva, otimista, bem humorada, companheira e competente. Obrigada por me
receber com tanto carinho na Itália, por ter me apresentado sua família e seus amigos, me
fazendo sentir acolhida na ausência da minha família. Não tenho palavras para expressar o
quão importante você se tornou em minha vida. Você foi minha inspiração e meu exemplo
profissional! Quero deixar aqui eternizados, meus sentimentos sinceros de admiração,
respeito e afeto por você. Grazie per tutto.
Ao professor Robert John Young. Ainda me lembro com detalhes da primeira vez que
entrei na sua sala, em março de 2005. Eu ainda infantil e imatura, mas cheia de sonhos e
determinação. Você me acolheu com seriedade e me incentivou a desenvolver o projeto que
eu sonhava. Poucos minutos na sua sala e saí com um desenho experimental pronto, um
orientador e um amigo. Hoje, conquisto o título mais alto da carreira de pesquisador e isso só

foi possível porque eu te conheci e porque você acreditou em mim. Serei eternamente grata
pelos oito anos em que trabalhamos juntos no Brasil.
À professora Renata Sousa-Lima. Obrigada por ter participado da idealização deste
projeto, aceitando me co-orientar, sonhando junto comigo e se arriscando a fazer algo nunca
feito antes. Obrigada, porque mesmo estando longe você contribuiu de forma expressiva
nesta pesquisa.
Ao professor Almo Farina, que me recebeu como sua “ragazza brasiliana” e aceitou
dividir sua sala e sua Nadia comigo durante dois meses. Muito obrigada por ter me dado a
oportunidade de te conhecer e aprender com você.
Ao professor Bonifácio, que gentilmente cedeu uma sala no Museu de Ciências
Naturais da PUC para que pudéssemos montar o Laboratório de Bioacústica. Obrigada por
ter me dado condições logísticas e operacionais para o desenvolvimento da minha tese e
também por confiar no meu trabalho e me acolher como parte da equipe do museu.
Ao professor Carlos Augusto e seus alunos, Douglas e Alan, por todo apoio
operacional com os storages, softwares e equipamentos do projeto. Vocês foram essenciais!
Ao professor Nilo Bazzoli, por aceitar ser coordenador do projeto do qual esta tese foi
fruto.
Ao engenheiro Krisdany Cavalcante, pelo apoio e ensinamentos com os medidores de
nível sonoro.
Ao Marcelo Vasconcelos pela ajuda na identificação dos cantos das aves, sempre com
muito entusiasmo.
Aos professores do PPG ECMVS pelas disciplinas ministradas.

Aos professores e pesquisadores que gentilmente aceitaram o convite para participar
da banca e enriquecer meu trabalho.
Aos funcionários do Parque Nacional da Serra do Cipó, especialmente Ivan Campos e
Edward, pelo apoio durante as coletas de dados.
Aos funcionários do Parque Estadual do Rola Moça e da Estação Ambiental de Peti,
especialmente ao Leotacílio da Fonseca.
Aos funcionários do PPG ECMVS, Elídio, Cris, Fred pelo constante apoio.
Aos funcionários do mestrado em Zoologia, da PROPPG e Museu: Clédma, Rosa,
Elane, André e Márcio pelas gentilezas prestadas e especialmente à Ana Cristina pela
amizade.
À FAPEMIG, VALE e CNPq, pelo apoio financeiro concedido para o desenvolvimento
deste projeto e pelas bolsas de doutorado e iniciação científica.
Ao Fabrizio Frontalini, que dividiu comigo sua Nadia e muitas vezes, participou de
discussões importantes sobre o andamento da minha tese.
À Maria Ceraulo, pela doce amizade que fiz no Laboratório de Soundscape Ecology,
durante os dias na Itália.
À Regina Scarpelli por ter trazido ao mundo uma pessoinha que foi fundamental
durante minha pesquisa. Nina, me faltam palavras para expressar o quanto você é
importante para mim. Ao longo dos três anos em que trabalhamos juntas você foi fiel,
paciente, inteligente, competente, dedicada, companheira, amiga, confidente e muito mais!!!
Poucas vezes eu conheci pessoas que tivessem uma sintonia tão grande comigo. Você foi
muito mais que estagiária, porque você fez tudo com muito amor. Muito obrigada!

Às amigas Sara, Marina Nogueira e Isabella Diniz por terem sido as amigas com as
quais eu pude contar a qualquer hora. Eu amo muito vocês!
À Mari e Afiwa pelo apoio durante esta pesquisa e pela companhia no lab de
Bioacústica.
À minha amada família, papai, mamãe e Nang, vocês são o meu mundo e razão pelo
qual cheguei até aqui. Pai, obrigada por ter sempre me guiado em direção à realização dos
meus sonhos! Este título também é seu. Mãe, obrigada por cuidar de mim em todos os
momentos. Nang Pum, meu irmão, meu melhor amigo, você é meu amor maior!
Ao Rafa por ter sido a melodia doce que me muitas vezes aliviou os momentos de
estresse. Durante as adversidades você me transmitiu força. Você foi paciente, amigo,
companheiro e acima de tudo, compreensivo. Você aceitou minha ausência com toda
paciência do mundo, me encorajou e me fez acreditar que eu fosse conseguir. Obrigada pelo
amor, você foi demais!
Por fim, agradeço a Deus e à natureza e seus sons magníficos, que me deram força e
inspiração para chegar até aqui.

- SUMÁRIO -
Introdução geral e apresentação..................................................................................11
Capítulo 1. Determining temporal sampling schemes for passive acoustic studies in
different tropical ecosystems...........................................................................................16
Capítulo 2. The impact of anthropogenic noise from open cast mining on Atlantic
forest biophony................................................................................................................50
Capítulo 3. Mining noise reduces loud call by wild black-fronted titi monkeys….......81
Conclusão.....................................................................................................................105
Referências Bibliográficas..........................................................................................107

11
Introdução geral e apresentação
-Introdução geral e apresentação-
1. Atividades antrópicas e o impacto do ruído
O crescimento da população humana no mundo conduz a exploração acelerada
dos recursos naturais e a invasão das paisagens naturais, causando sérios problemas
ambientais (Cohen, 1995). Atualmente, o impacto humano sobre o meio ambiente atrai
a atenção dos cientistas para abordar os aspectos da pressão sobre os recursos naturais e
populações de animais selvagens (Cohen, 1995; Pijanowski et al., 2011).
Um dos recursos mais explorados no Brasil é o minério de ferro e a mineração é
uma das atividades econômicas mais importantes do país (IBRAM, 2011). A pressão
por extração de minério tem se tornado cada vez mais intensa e muitas minas no Brasil
estão localizadas em biomas considerados hotspots de biodiversidade, como o Cerrado e
a Mata Atlântica (Myers et al., 2000; Estrada, 2009). Entre os impactos gerados por esta
atividade estão: supressão de habitat para construção de estradas e implantação da mina,
vibrações transmitidas aos terrenos e estruturas adjacentes, e produção de ruído
proveniente das etapas de implantação da mina, extração e transporte de minérios
(Donoghue, 2004).
Métodos acústicos proporcionam uma oportunidade de monitorar o ruído
produzido por atividades antrópicas e seus efeitos nos ecossistemas com uma ampla
escala espacial e temporal, fornecendo dados relevantes para decisões sobre manejo e
uso da terra (Brown et al., 2013). Em muitas situações, o ruído antropogênico pode
mascarar sinais acústicos e impedir a capacidade dos animais em compreender,
reconhecer ou detectar sons de interesse (Warren et al., 2006; Clark et al., 2009;
Versace et al., 2008). A comunicação acústica é essencial para a sobrevivência dos
animais, pois dela dependem comportamentos sociais, como defesa de territórios,
comportamentos reprodutivos (a atração e identificação de parceiros sexuais) e também
a percepção de sinais importantes, como chamados de alarme, perigo e vocalizações
relacionadas ao cuidado parental, e detecção de presas ou predadores (Warren et al.,
2006). Além disso, a identificação de sons naturais auxilia na orientação de organismos
e seu deslocamento a locais favoráveis à sua sobrevivência e reprodução (Vermeij et al.,
2010).

12
Introdução geral e apresentação
Estudos relatam que o ruído de mineração pode afetar a reprodução das aves e
diminuir a densidade de indivíduos (Smith et al., 2005), diversidade de espécies e
tamanho populacional (Saha e Padhy, 2011). Entretanto, algumas espécies de animais
que vivem em áreas ruidosas são capazes de ajustar seus sinais acústicos para
comunicar nestes ambientes; por exemplo, aumentando a amplitude das vocalizações
(Brumm et al., 2004; Brumm et al., 2009), mudando as frequências (Slabbekorn e Peet
2003; Parks et al., 2007; Nemeth e Brumm, 2009), alterando a taxa e a duração dos
chamados (Brumm et al., 2004; Sun e Narins, 2005) ou o turno de vocalização (Fuller et
al., 2007; Sousa-Lima e Clark, 2008). Outras espécies apresentam mudanças no
comportamento, tais como evitar áreas ruidosas para forragear (Miksis-Olds et al., 2007;
Schaub et al., 2008) e desenvolver outras atividades diárias (Sousa-Lima e Clark, 2009;
Duarte et al., 2011). A evasão de áreas e os mecanismos compensatórios para reduzir os
efeitos do ruído podem alterar a complexidade acústica de uma comunidade e resultar
na diminuição de abundância e da diversidade de espécies em áreas ruidosas (Bayne et
al., 2008; Proppe et al., 2013).
Atualmente, grande parte dos esforços para reduzir a poluição acústica é
destinada a diminuir os efeitos negativos da exposição ao ruído nos seres humanos,
especialmente em comunidades urbanas expostas ao ruído proveniente de estradas e
aeroportos. No entanto, pouca atenção tem sido dedicada à regulamentação da poluição
sonora em relação aos animais (Sousa-Lima, 2007). Diante dos impactos provocados
pelo ruído proveniente das atividades mineradoras e considerando o grau de ameaça dos
biomas e das espécies inseridas em áreas com a presença desta atividade, têm-se a
necessidade de pesquisas envolvendo o impacto do ruído sobre a fauna nestes locais.
2. Monitoramento Acústico Passivo e Ecologia de Paisagem Acústica
O monitoramento acústico passivo (MAP) é uma metodologia inovadora para
ambientes terrestres, que fornece oportunidades de avaliar o grau de conservação de
ambientes e as consequências de diferentes atividades antrópicas na natureza (Blumstein
et al., 2011; Mennit e Fristrup, 2012; Brown et al., 2012, 2013). Através das técnicas de
MAP também é possível avaliar diferenças acústicas entre comunidades que ocorrem
em áreas distintas, monitorar mudanças ao longo do tempo, comprovar a ocorrência de

13
Introdução geral e apresentação
novas espécies em determinado ambiente e entender as relações entre animais e
elementos externos, por exemplo, o ruído (Pijanowski et al., 2011). Esta metodologia é
especialmente importante para ser utilizada em ambientes que são difíceis de acessar ou
monitorar usando métodos convencionais (Mellinger e Barlow 2003; Brandes, 2008).
O avanço de tecnologias e o consequente desenvolvimento de equipamentos de
MAP possibilitou o surgimento de uma nova linha de pesquisa dentro das disciplinas
ecológicas existentes: a "Ecologia de Paisagem Acústica", ou "Soundscape Ecology".
(Pijanowski et al., 2011). Esta linha de pesquisa é uma das abordagens mais recentes
para estudar o impacto das atividades antrópicas sobre o ambiente. Uma paisagem
acústica é definida como qualquer ambiente acústico natural, urbano ou rural e pode ser
composta por três elementos fundamentais: a biofonia (sons biológicos não humanos,
como vocalizações de anfíbios, aves e estridulações de insetos), a geofonia (sons físicos
da natureza como vento, trovões, cachoeiras, etc) e antropofonia (sons produzidos por
seres humanos) (Krause et al., 2011; Pijanowski et al., 2011).
A Ecologia de Paisagem Acústica ainda é uma linha de pesquisa difícil de ser
investigada devido à ampla variedade de informação disponível em cada ambiente
acústico e à dificuldade que existe na identificação de índices que possam interpretar
rapidamente a grande quantidade de informação contida nos registros de áudio. Como
toda linha de pesquisa recente, a Ecologia de Paisagem Acústica carece do
desenvolvimento de métricas e protocolos que possam otimizar o processo de análise e
interpretação de dados. Apesar do desenvolvimento de novas tecnologias e softwares
nos últimos anos, a análise de sons naturais ainda requer muito tempo para ser realizada,
o que dificulta a extração de dados ecológicos importantes em um amplo banco de
dados. Vários autores têm desenvolvido técnicas baseadas em informações bioacústicas
de uma única espécie (Klinck et al., 2008; Wolf, 2009; Bardeli et al., 2010), enquanto
índices e metodologias para o monitoramento acústico de comunidades de animais são
raramente desenvolvidos. Assim, a Ecologia de Paisagem Acústica oferece novas
perspectivas para investigações no campo da Ecologia de Paisagens, mas a
implementação de novos métodos para otimizar as pesquisas nesta área é extremamente
necessária.

14
Introdução geral e apresentação
3. Apresentação da tese
Esta tese aborda os impactos da poluição sonora na comunicação acústica da
fauna silvestre utilizando métodos de MAP e o Índice de Complexidade Acústica (ACI),
que foi recentemente desenvolvido para medir a complexidade acústica de ambientes
terrestres.
No capítulo 1, um novo método de subamostragem de dados sonoros é
apresentado, com indicações de protocolos de subamostras em diferentes ambientes em
Minas Gerais (Cerrado, Campo Rupestre e Mata Atlântica). O uso de equipamentos de
monitoramento acústico passivo permite a coleta de grande quantidade de dados, o que
faz surgir a necessidade da elaboração de métodos de subamostragem. Estes métodos
são importantes para que seja possível alcançar o compromisso entre um esforço de
amostragem rigoroso e resultados confiáveis, considerando também, questões de
armazenamento e a redução de tempo e recursos consumidos durante o processo de
análise de dados. O protocolo apresentado neste capítulo é feito com base no Índice de
Complexidade Acústica (ACI), um algoritimo criado para produzir uma medida direta
da complexidade de sons biológicos, computando a variabilidade de intensidades
registrada em arquivos sonoros, apesar da presença quase constante de ruído
antropogênico.
No capítulo 2, o ACI é novamente utilizado, porém com objetivo de analisar o
impacto da poluição sonora proveniente de atividade mineradora na biofonia de um
fragmento de Mata Atlântica, localizado próximo à mina de Brucutu, uma das maiores
minas de minério de ferro do mundo. Neste capítulo, duas áreas de mata (uma próxima
e outra distante da mina) do mesmo fragmento são comparadas em termos de ruído e
biofonia e também em riqueza, composição de espécies encontradas nas gravações e
características espectrais dos cantos. Também foi realizada a medição dos níveis
sonoros e a caracterização dos ruídos produzidos pela atividade de mineração com base
nos registros sonoros feitos na área próxima à mina.
No capítulo 3, foi analisado o impacto da poluição sonora da mina de Brucutu
nas vocalizações de guigós (Callicebus nigrifrons), um primata ameaçado de extinção,
que vive em áreas de Mata Atlântica. Neste capítulo, foram quantificadas todas as
vocalizações de guigós encontradas ao longo do dia no período de um ano, em duas

15
Introdução geral e apresentação
áreas sendo uma próxima e outra distante da mina. A duração e a taxa de ocorrência das
vocalizações foram medidas e comparadas entre as duas áreas estudadas. Além disso, o
número de caminhões de mineração que passaram ao longo do dia foi quantificado na
área próxima à mina para que fosse verificada a correlação entre a passagem dos
caminhões e ocorrência de vocalizações.
Finalmente, são apresentadas as principais conclusões do trabalho e
mencionadas as possíveis direções futuras para esta linha de pesquisa.

16
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
- CAPÍTULO 1-
Determining temporal sampling schemes for passive acoustic studies in
different tropical ecosystems
Artigo aceito no periódico Tropical Conservation Science
Abstract
Among different approaches to exploring and describing the ecological complexity of
natural environments, soundscape analyses have recently provided useful proxies when
it comes to understanding and interpreting dynamic patterns and processes across a
landscape. Nevertheless, the study of soundscapes remains a new field with no
internationally accepted protocols. This work aims to provide the first guidelines for
monitoring soundscapes in three different tropical areas, specifically located in the
Atlantic Forest, Rupestrian fields and the Cerrado. Each area was investigated using
three autonomous devices recording for six entire days during a period of 15 days in
both the wet and dry seasons. The recordings were processed via a specific acoustic
index and successively subsampled in different ways to determine the degree of
information loss when reducing the number of minutes of recording used in the
analyses. We describe for the first time the temporal and spectral soundscape features of
three tropical environments and test diverse programming routines to describe the costs
and the benefits of different sampling designs, taking into consideration the pressing
issue of store and analyze extensive data sets generated by passive acoustic monitoring.
Schedule 5 (recording one minute every five) appeared to retain most of the information
contained in the continuous recordings from all the study areas. Less dense recording
schedules produced a similar level of information just in specific portions of the day.
Substantial sampling protocols such as those presented here will be useful to researchers
and wildlife managers as they will reduce time- and resource-consuming analyses,
whilst still achieving reliable results.
Keywords: environmental monitoring, animal conservation, tropical environments,
soundscape ecology, sampling protocols.

17
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
Resumo
Entre as diferentes abordagens para explorar e descrever a complexidade ecológica de
ambientes naturais, a análise de paisagem acústica tem fornecido recentemente
ferramentas úteis para o entendimento e a interpretação da dinâmica de padrões e
processos de uma paisagem. Apesar disso, o estudo de paisagens acústicas é uma nova
linha de pesquisa que ainda não possui protocolos e métricas aceitas internacionalmente.
Este estudo tem como objetivo fornecer as primeiras diretrizes para monitorar paisagens
acústicas em três diferentes áreas tropicais localizadas especificamente na Mata
Atlântica, no Campo Rupestre e no Cerrado. Cada área foi investigada usando três
equipamentos autônomos gravando por 6 dias inteiros durante um período de 15 dias
nas estações seca e chuvosa. As gravações resultantes foram processadas utilizando um
índice acústico específico e foram sucessivamente subamostradas para determinar o
grau de informação perdido quando reduzido o número de minutos de gravações usadas
nas análises. Nós descrevemos pela primeira vez, as medidas temporais e espectrais de
três ambientes tropicais e testamos rotinas de programação diversas para descrever os
custos e benefícios de diferentes desenhos de amostragem, considerando questões de
armazenamento e análise de bancos de dados extensos gerados por monitoramento
acústico passivo. A programação 5 (gravação de um minuto a cada 5 minutos) manteve
o maior número de informações contidas nos registros contínuos em todas as áreas de
estudo. Programações de gravação menos intensas produziram um nível similar de
informação apenas em porções específicas do dia. Protocolos de amostragens tais como
os apresentados aqui são úteis para pesquisadores e gestores de meio ambiente, uma vez
que eles podem reduzir tempo e recurso a ser consumido durante análise de dados e
ainda fornecer resultados confiáveis.
Palavras-chave: monitoramento ambiental, conservação animal, ambientes tropicais,
ecologia de paisagens acústicas, protocolos de amostragem.

18
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
Introduction
Nowadays, passive acoustic monitoring (PAM) is considered to be a valuable
tool for both research and management, and involves collecting acoustic data over large
spatial and temporal scales and providing detailed and long-term information on animal
distribution and variations in community dynamics. This wide-scale data collection
inevitably leads to animal populations being better understood and managed more
effectively [1]. However, to avoid time- and resource-consuming analyses, acoustic
surveys need to specifically address general guidelines that can ensure efficient
sampling on the basis of experienced protocols.
Animals produce sounds for diverse biological functions (e.g. communication,
mating, building territories, foraging) [2, 3], which can serve as proxies for estimating
species fitness and individual behavior, especially in environments that are difficult to
access or monitor using conventional methods [4, 5]. In the early 1990s, idiosyncrasies
in the study of marine mammal behavior led researchers in the field to develop
autonomous acoustic devices that enabled them to detect sounds underwater [6].
Successively, the use of acoustic recordings of the natural environment became
gradually an important technique for ecologists for monitoring all ecosystems. In
particular, passive acoustic monitoring has only recently been proposed for terrestrial
environments [7], and the study of the soundscape (soundscape ecology), which is
defined as the aggregation of sounds from physical, biological and human-made
sources, has rapidly gained attention as a potential tool to both evaluate ecosystem
health [8] and the effects of changes in land use and climate at various temporal and
spatial scales [9–11].
Advances in technology over the last decade have revolutionized the potential of
acoustic surveys. Fixed, programmable acoustic recording sensors can sample
continuously for 24 hours a day for prolonged periods of time, allowing for the non-
invasive assessment of changes in the distribution and acoustic behavior of entire
animal communities across a variety of habitats simultaneously. Moreover, all of the
recordings can be permanently stored and serve as an everlasting memory of the sounds

19
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
of the habitats [1, 5, 12].
The achievement of this temporal and spatial wide-scale application of
soundscape ecology inevitably produces an overwhelming amount of information, with
linked difficulties in data management and analysis [13]. Problems include an ever-
growing requirement for storage space and the need for time-consuming processing,
expensive power supply systems, and field personnel to periodically download data and
reinstall the equipment. Common standards and baseline data collection models could
be useful to limit unnecessary recordings and trips to the field while ensuring targeted
data are collected.
Optimizing the recording schedule by selecting specific portions of active
recording times (ON), which leaves the device off for the rest of the time (OFF),
becomes obligatory when it comes to optimizing basic resources and staff-time,
especially when constrained by limited funding. On the other hand, by reducing ON
periods, the probability of losing important information increases and may result in a
distorted description of the target community. As a consequence, identifying the
appropriate sampling period with which to conduct a study is essential for using
soundscape surveys appropriately to achieve scientific, management and conservation
objectives. To make such a decision, a good understanding of the daily and frequency
patterns of the recorded community is required.
Several acoustic surveys have been conducted in recent years to investigate
animal community dynamics and structure [14, 15], species richness and distribution
[16–18], relationships with vegetational parameters [19, 20], and human or noise impact
[21–23]. However, explicit evaluations of the survey effort required to characterize the
acoustic dynamics of different landscapes are generally lacking. Knowledge about
temporal variations in such acoustic dynamics could improve the design of future
soundscape studies and render soundscape ecology more efficient and applicable for
different categories of users (academics and other stakeholders). Our goal was to
describe the type and extent of soundscape information lost with different recording
schedules in areas located in three tropical ecoregions (Atlantic Forest, Rupestrian fields

20
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
and the Cerrado). These environments were chosen as they are priority conservation
areas, threatened environments, and contain biodiversity hotspots with high endemism
[24–26]. Additionally, studies of tropical soundscapes are limited [18].
Moreover, on the basis of the obtained results, we tried to both identify a cost-
effective scheme for surveying such areas and suggest the minimum sampling effort
required to meet the goal of characterizing the soundscape features. This was achieved
by identifying when the recording schedule loses acoustic information that is essential
for correctly describing the dynamics of the sound activity of that community and its
circadian rhythms.
Methods
Study area
The study was conducted in three threatened environments in Minas Gerais, in
the southeastern region of Brazil: Atlantic Forest, Rupestrian ferruginous fields and
Cerrado strictu sensu (Fig. 1).
Atlantic Forest – Environmental station of Peti – The Atlantic Forest is a
world biodiversity hotspot with high species richness and high levels of endemism,
which are threatened by the rapid loss of native land-cover types [25]. We collected data
in this biome at the environmental station of Peti in the municipalities of São Gonçalo
do Rio Abaixo and Santa Bárbara (19°53’57’’S and 43°22’07’’W). The reserve is
approximately 605 hectares in size and is located in the upper Rio Doce Basin (altitude
range: 630-806m). The area harbors 29 anuran species [27], 231 bird species [28] and
46 mammal species [29]. A large part of the reserve is covered by secondary arboreal
vegetation, with large trees and a continuous canopy [30].
Rupestrian fields – State Park of Rola Moça - The ecosystems found in
ferruginous outcrops known as ‘Rupestrian ferruginous fields’ or ‘Canga’ are among the
less studied and most endangered areas of Brazil due to restricted geographical

21
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
distribution and them being associated with the country’s main iron ore deposits [26].
Rupestrian fields have a relatively continuous herbaceous stratum of sclerophyllous
plants, which are small evergreen shrubs located between rocky outcrops that occur at
altitudes between 800 and 2000m. This ecosystem is highly diversified, with more than
4000 plant species along the Espinhaço Range and one of the highest levels of
endemism in Brazil [31]. We collected data in the Rupestrian fields at the State Park of
Rola Moça, which is located in the northwest of ‘Quadrilátero Ferrífero’ (20°03'60"S,
44°02'00"W) at an altitude of approximately 1450m.
The Cerrado – National Park of Serra do Cipó - The Cerrado is a biodiversity
hotspot and a highly threatened environment [25]. The Cerrado sensu strictu is
characterized by the presence of small trees with thick and twisted trunks and branches,
while grasses characterize the understory [32]. We collected data in an isolated area of
the Cerrado strictu sensu at the core of the national park of Serra do Cipó, which is
approximately 34,000 hectares in size and is situated at 19°12’19’’S and 43°30’43’’W.
This area provides habitat for 226 bird species [33] and 26 medium-large mammalian
species [34].

22
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
Figure 1. Location map of the study areas: Atlantic Forest (AF), Rupestrian fields
(RF), and Cerrado strictu sensu (CE). The photographs represent the typical
surroundings of the three habitats where the acoustic measurements were taken.
Acoustic recordings and data analyses
The climate of southeastern Brazil can be divided into two macro-climatic
seasons: a hot wet season, running from October to March, and a cooler dry season from
April to September [35]. The soundscape of the three study areas was collected by
recording for six non-consecutive days during a period of 15 days during the dry
(Cerrado: 9-23 September 2012; Rupestrian fields: 17-30 April 2013; Atlantic Forest: 4-

23
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
19 June 2013) and wet seasons (Cerrado: 15-30 March 2013; Rupestrian fields: 6-21
October 2012; Atlantic Forest: 17 October-1 November 2012). In each sampling area,
three SongMeter Digital Field Recorders (SM2) (Wildlife Acoustics, Inc.,
Massachusetts) were set to record from 00:00 to 23:59h continuously. Accordingly,
each area was recorded for 432 hours (24h * six days * three recorders) each season,
making 2592 hours in total. 06:00 and 18:00 were considered as the approximate times
of dawn and dusk, which were slightly varying among the different months. One of the
three recorders stopped recording during the wet season in the Rupestrian fields after
three days, while another recorder in the Atlantic Forest stopped working after 17 h on
the last recording day during the dry season.
The recorders were placed at a distance of approximately 300m from each other
to avoid double sampling the same sounds and intend each recorder as an independent
sampler per area. They were mounted on a tree at approximately 1.5m from the ground,
and ensured any nearby vegetation would not interfere with recordings. The SM2s
recorded at a sampling rate of 44,100Hz, set at 16 bits.
The Kaleidoscope converter utility (Wildlife Acoustics, Inc., Massachusetts) was
used to split the collected data into files of one minute in length, which were further
processed via the Wavesurfer software [36] powered by the SoundscapeMeter plug-in
[37]. One minute resolution was chosen since most of the recent literature used this time
lapse for sound assessments [18, 19, 38, 39], and to compare with previous research.
Among the variety of the available acoustic indices to directly summarize the
information in a recording (i.e. [16, 17, 22, 38, 40]), the Acoustic Complexity Index
(ACI) [11, 41] was selected for this study. The ACI was chosen since it is an algorithm
designed to measure the spectral complexity of soundscapes and was recently used to
track the dynamics of animal acoustic communities [15] and compare it with
vegetational parameters [19]. Moreover, in the recent study of Towsey et al. [38], it was
found to be one of the best indicator of the biodiversity of a bird community among a
list of 14 different acoustic indices, with weaknesses due to the sensitivity to wind
gusts. To analyze the collected acoustic data, a Fast Fourier Transform (FFT) of 512
points was applied, obtaining from every recorded minute a matrix made by 256

24
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
frequency bins of 86.13Hz and 5167 temporal intervals of 0.012s. This matrix was used
to calculate the Acoustic Complexity Index, with the following formula:
where |Ik-Ik+1| is the absolute difference between two adjacent values of amplitude along
a frequency bin, n represents the total number of temporal steps (k) contained in every
interval of time in which the calculation is made (in this study, 1s). The sum of the
results for all of the frequency bins and temporal intervals is then calculated. To avoid
bias due to background ambient noise that is inevitably present in every recording (even
if soft), we set an a priori filter on the power spectral density (SoundscapeMeter
settings: Noise filter =3000 μV2/Hz) operating on all the frequency bins, so that the ACI
did not apply to values under the selected threshold. This filter was appositely verified
for the type of recording used in order to not filter biophonies but just background noise
and to increase the signal to noise ratio.
Five different recording schedules were then chosen to be simulated: (i)
Schedule 5: recording one minute over five minutes; (ii) Schedule 10: one minute over
10 minutes; (iii) Schedule 20: one minute over 20 minutes; (iv) Schedule 30: one
minute over 30 minutes; and (v) Schedule 60: one minute over 60 minutes.
These schedules were obtained by selecting the corresponding minutes of each
simulated configuration from the continuous recording, thus simulating a recording
routine whereby the recorder was not running continuously, but intermittently, at
respectively one minute every five, 10, 20, 30 and 60 minutes. A mean of the ACI
values was then calculated for each recording hour for both the continuous recordings
and the simulated samplings in order to compare the different schedules with the
original and complete sampling. These comparisons were conducted for both the
temporal and spectral dimensions.
Rain and wind were found to be recognizable in abnormal ACI results [15, 38],
especially at lower frequencies. Consequently, when the ACI values highlighted
discrepancies with the normal acoustic behavior of the local community, the sound files

25
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
were aurally checked to verify if the anomaly was due to some atypical biophony or
anthrophony (such as insect buzzes on the microphone or transits of airplanes) or to the
influence of stormy weather. This allowed us to generate a table of the adverse weather
conditions during the recording days that was filtered from the analyses in selected
statistical tests.
Statistics
All of the statistical tests were performed using Statistica v.8.0. A non-
parametric approach was utilized, since the variables did not present a normal
distribution pattern, even after transformation of the data values. Non-parametric
correlation analyses (Spearman’s rho, p < 0.01) were conducted to investigate the
relationship between the continuous data set and the simulated recording schedules.
To quantify the relative non-conformity of the sampling schedules with the real
distribution of the ACI levels along the different hours of the day and the different
frequency bins, the percent deviation [42] was calculated using the following formula:
(1) % deviation = (actual value – expected value)/expected value x100
in which the ‘actual value’ was the ACI value calculated for a simulated configuration
(expressed as an hourly mean) and the ‘expected value’ is the ACI resulting from the
continuous recordings. Successively, the percent deviation was grouped by temporal
slots (hours of the day) and frequency bins (1kHz-wide) to determine specifically where
results from the simulated schedules differed from those from to the continuous data set.
For both the correlation and percent deviation tests, only frequencies above 500Hz were
processed since, under that threshold, the ACI could not well filter the background noise
from the environment, which, if included, could have affected the final results. At the
Rupestrian fields it was windy all year round, and so the cutoff frequency for the
temporal analyses was 1500Hz to avoid the inclusion of soft wind noise. Rain and wind
produce sounds and add complexity to soundscape analyses. As a result, to enable us to

26
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
both consider the consequences of adverse weather when searching for the best
sampling approach and focus on the acoustic behavior of the animal community, it was
decided to treat the temporal and the frequency analyses differently.The entire data set
was used for the temporal analyses in order to include realistic limitations caused by
weather conditions. On the other hand, the hours affected by rain and strong wind were
left out of the data set when considering the differences reported with respect to the
spectral distribution of sounds (the frequency footprint, sensu Farina et al. 2011a),
enabling us to reliably track the acoustic community dynamics and identify which
frequencies were most affected when the sampling was less intense.
Results
The singing community
The ACI values varied greatly from the wet to the dry season in all our study
areas, with a pronounced change between daytime and nighttime recordings. Figures 2,
3 and 4 show, respectively, the seasonal, temporal and spectral acoustic complexity
variations of the investigated environments based on the complete data set. A summary
of their main soundscape features resulting from the ACI is set out in Table 1.
Table 1. Summary of the principal soundscape features of the three environments.
Wet season
Dry season Higher
acoustic
activity Peaks of
activity (kHz)
Peaks of activity
(hours)
Peaks of activity
(kHz)
Peaks of
activity (hours)
Atlantic forest 4-6 kHz
15-16 kHz
18:00 - 01:00
07:00-08:00
1 kHz, 4 kHz
15-16 kHz 18:00 - 19:00
Wet season
Rupestrian field 3-5 kHz
9-13 kHz 19:00 - 20:00
2-4 kHz
5- 7 kHz 12:00 - 16:00
Wet season
Cerrado 5-6 kHz
10-17 kHz
18:00 - 03:00
12:00
15:00 -17:00
3 kHz - 5-6 kHz
10-14 kHz 07:00 - 17:00 Wet season

27
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
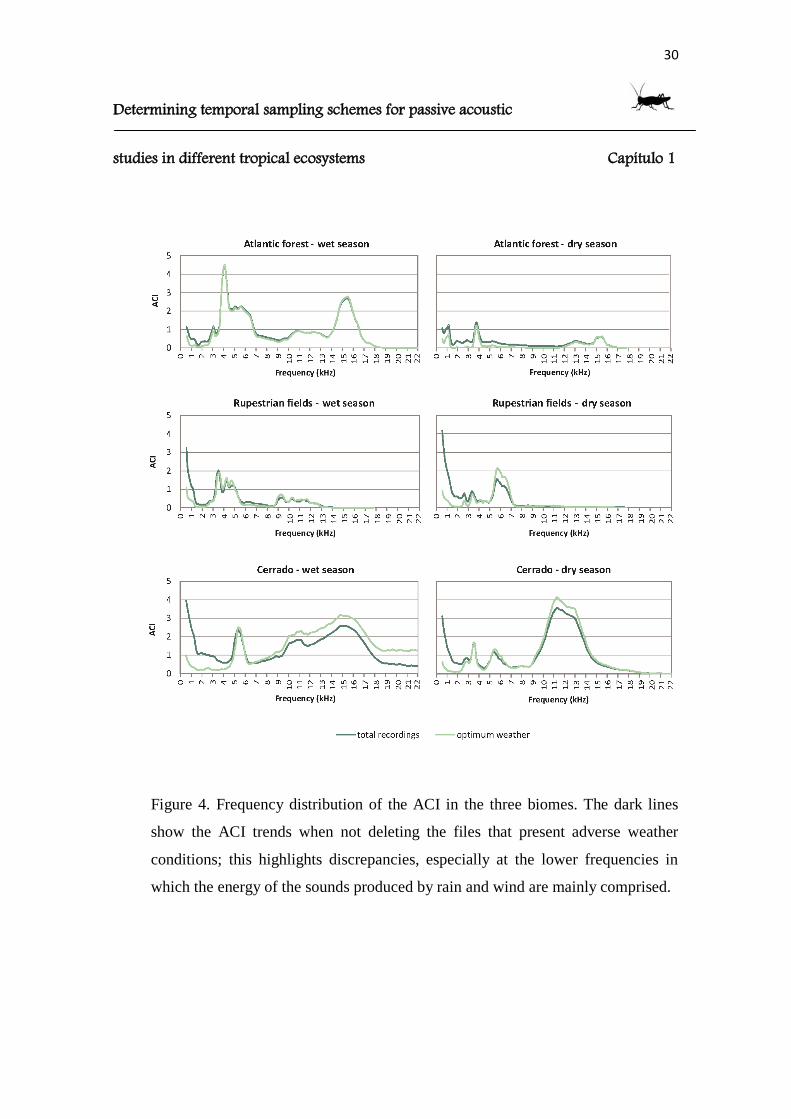
recording period, nonetheless, when filtering out the windy hours from the data set, it
was possible to register narrow peaks from 3 to 5kHz and from 9 to 13kHz in the wet
season, which switched into peaks from 2 to 4kHz and 5 to 7kHz in the dry season.
The sampled areas in Cipó (the Cerrado) had the highest ACI values, especially
in the wet season. In the wet season, most of the acoustic complexity was registered
above 10kHz, with a narrow peak from 5 to 6kHz; the ACI presented high values
preferentially during the night hours (18:00 to 03:00). In the dry season, a higher
acoustic complexity was registered during daylight hours (07:00 to 18:00) mostly
between the 10 and 14kHz frequency bands. Others peaks of ACI were found from 3 to
6kHz.

28
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
Figure 2. Box-Whisker plot of the hourly means of the ACI values in
the three environments.

29
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
Figure 3. Temporal trends of the acoustic complexity recorded in the three
environments. Each graph represents the mean pattern resulting from sampling on six
days at three recording points. The dark lines show the ACI trends when not deleting
the files with adverse weather conditions; this highlights discrepancies in the hours of
the day in which rain and wind mainly occurred. The green highlight shows the period
of the day comprised between the approximate times of dawn and dusk.
30
Determining temporal sampling schemes for passive acoustic
studies in different tropical ecosystems Capítulo 1
Figure 4. Frequency distribution of the ACI in the three biomes. The dark lines
show the ACI trends when not deleting the files that present adverse weather
conditions; this highlights discrepancies, especially at the lower frequencies in
which the energy of the sounds produced by rain and wind are mainly comprised.

31
Determining temporal sampling schemes for passive acoustic
studies in different tropical ecosystems Capítulo 1
Statistical analyses of the sampling schemes
All of the correlations between the ACI values from the scheduled and
continuous recordings were significant and positive (Fig. 5). An expected inverse
relationship between time OFF and the value of the correlation was found for both the
frequency and time analyses. The correlation coefficients were very high (r>0.90;
p<0.01) for the more intense sampling period (Schedules 5 and 10) and fell with
increasing OFF minutes, especially when considering Schedule 60. The Rupestrian field
correlations generally had the lowest values. The frequency correlations were always
found to be higher than the temporal correlations.
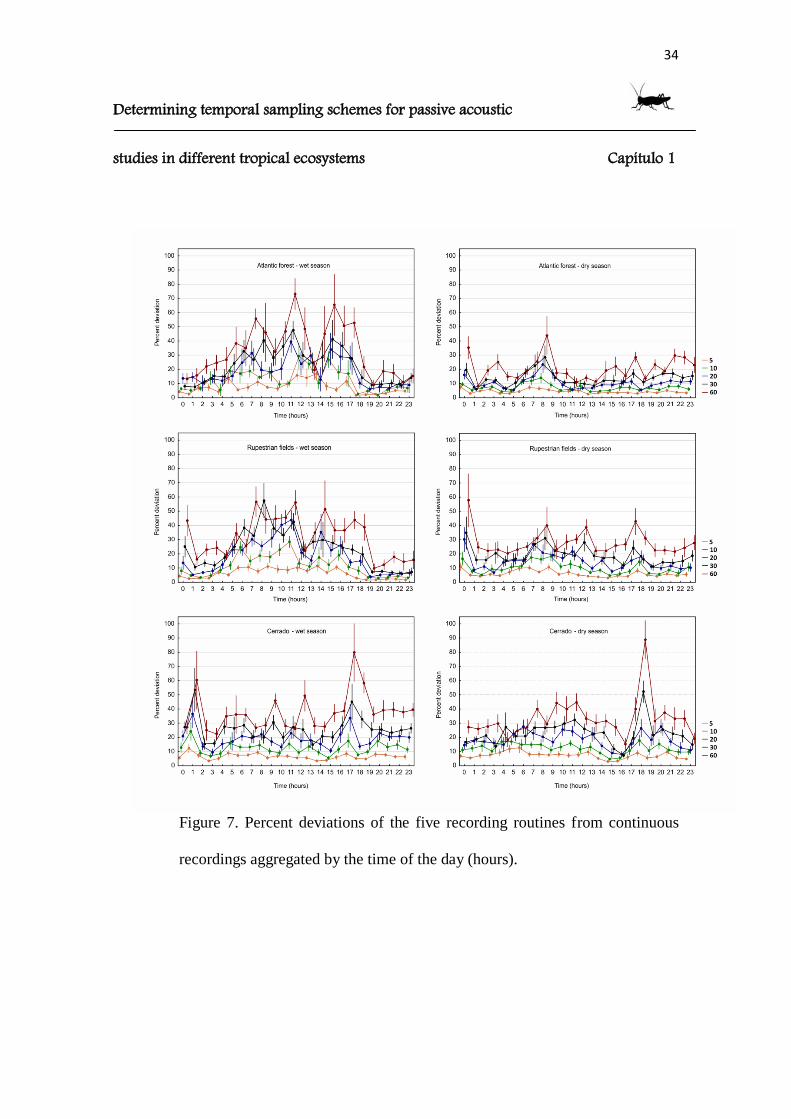
The percent deviations were low for the intense sampling schedules and tended
to increase when enlarging the OFF period (Fig. 6). As for the correlations, the temporal
analyses tended to diverge away from the continuous recordings more strongly than the
spectral analyses (Fig. 6). When categorized by hour of the day or 1kHz-frequency
bands, interesting trends on the possible major losses of information of the subsampled
recording schedules became clearly visible (Figs. 7 and 8). In particular, Schedule 5
assumed values that deviated by a maximum of 10%. Schedule 60 registered substantial
deviations of 90% and 80% at specific hours of the day (Cerrado, wet and dry seasons,
respectively), and deviations over 30% in the frequency analyses (Rupestrian fields and
Cerrado, wet season).
In the wet season, both the Atlantic Forest and the Rupestrian fields seemed to
experience a greater loss of information during daylight hours, while in the dry season
the deviations were more evenly distributed. In the Cerrado, we found peaks in the
deviations at 17-18:00 (both seasons) and 01:00 (wet season). The highest frequency
bands registered null deviations in the Atlantic Forest (dry season) and Rupestrian fields
(wet season). In the Cerrado during the dry season, low variations were found in the
frequencies around 11-13kHz.

32
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
Figure 5. Spearman Rank correlations of the ACI results according to the temporal
and spectral comparisons of the different schedules (p< 0.01). The ACI results were
grouped by hour, comparing the mean value registered each hour by the different
recording schedules, or by the frequency bin (1kHz), comparing the mean value
registered for every frequency band by the different recording schedules.

33
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
Figure 6. Percent deviations of each subsampling category with respect to the
continuous recording. The unfiltered data set and the recordings with the
weather perturbations removed (i.e. optimum weather) are shown.
34
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
Figure 7. Percent deviations of the five recording routines from continuous
recordings aggregated by the time of the day (hours).

35
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
Figure 8. Percent deviations of the five recording routines from continuous
recordings aggregated by frequency (1kHz).

36
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
Discussion
Soundscape studies can be particularly useful for exploring fragile and
endangered ecosystems that need special attention for their conservation [18]. Our
results from simulating recording schedules provided information useful for suggesting
the first guidelines for soundscape studies in three tropical areas considered as
threatened environments. These suggestions are based upon our overall description of
the main dynamics recorded in the three study areas, evaluation of the different
sampling schedules as representative of the real acoustic dynamics, and the percentage
of information lost when reducing the recording time.
Soundscape characterization
Temporal and spectral characteristics of the soundscape for each study area were
unique and largely specific to the climate season. Generally we found a comparatively
higher ACI in the wet season which, in Brazil, coincides with the breeding season for
most species [33, 43, 44], when anurans, birds, and insects produce sounds to achieve
mating success. Acoustic complexity differences were clearly noticeable from the
diverse trend across the temporal domain (Fig. 3), and by delineating habitat and
season-specific frequency footprints (Fig. 4) (sensu [11]) depending on the singing
behavior of the emitters acting in each season and environment. Evidence of habitat
type acoustic signatures was also found in temperate environments in four forest and
two grassland habitat types in Northern Greece [45].
The lower acoustic complexity of the Rupestrian fields is probably related to
their high altitude, which directly influences vegetation structure (fewer trees, open
areas and strong winds). This leads to lower species richness and, consequently, lower
acoustic diversity (Figure 2).

37
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
General considerations on the different sampling schedules
All study areas were characterized by falling Spearman Rank correlation
coefficients with increasing OFF minutes, showing that there was a gradual loss of
correspondence with the real soundscape. This testifies that, in these environments, it is
difficult to provide a perfect picture of the acoustic dynamics of the community if the
sampling becomes too sparse. It is therefore likely that there will be a loss of important
data that could be essential for conservation issues.
This decreasing trend is shown by both temporal and spectral correlations, even
if the correlation coefficients are always very high in the latter. We hypothesize that this
is probably because the frequency bins have a lower degree of freedom than the
temporal analyses, since the spectral emissions were strictly linked to the acoustical
organs of indigenous species. Accordingly, animals cannot vary the spectral properties
of their emissions, which over the entire day are likely to be registered by less intense
sampling, but they can vary the moment and the length of a singing period. In other
words, the presented temporal analyses depend on what was singing across all of the
spectrum at a certain temporal interval, while the frequency analyses depend on what
was singing in the 24 hours of one day in a fixed frequency band. The frequency
footprint is thus less variable than the temporal trend across time. The Rupestrian fields
were the most critical environment, since the lowering of the sampled files
corresponded to very low correlations with the continuous recordings. The main reason
probably lies in the lower acoustic activity presented by the area (Fig. 2), which has a
higher risk of not being recorded and, thus, needs a greater sampling effort to be
captured and measured.
More detailed evidence about the loss of acoustic information is given by the
percent deviation analyses. In general, lower percent temporal deviations were found
where the sound emissions were more constant and prolonged in time, such as during
the night in the wet season in the Atlantic Forest and Rupestrian fields. At these times,
insects are the main protagonists of the acoustic performances, and tended to produce a

38
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
longer sonic performance than other taxa (mammals, birds). This makes them more
easily detectable in all of the sampling schedules, thus minimizing the percent
deviations. In contrast, during the day, birds also sung abundantly, but were less
constant in their acoustic emissions and more variable over time than the insects,
meaning that they may or may not be detected by less intense sampling. This
consideration suggests the need for more cautionary sampling during specific hours of
the day and a less intense effort at other times, which are typically characterized by the
greater constancy of sounds produced by the community.
The percent spectral deviation was found to be at a minimum where the
frequency bins were unoccupied (or rarely occupied) by some species, such as in the
Atlantic Forest (dry season) and Rupestrian fields (wet season). The narrow peaks
visible on the lower frequencies all referred to insects, most likely crickets, while
cicadas presented a broader frequency band. In the Cerrado (dry season), the reduction
in variation from 10 to 15kHz is related to the continuous and abundant sound
emissions of cicadas.
Which sampling routine is better?
The choice of the type of sampling will always depend on the principal focus of
the investigation, and so these results may help researchers to opt for the best sampling
protocol according their principal goals. Our findings show that there are preferential
recording schedules for each of the three investigated ecosystems. When the mean
soundscape of the community across the six recording days shows a high and
continuous presence of sounds, it may be preferable to use less dense recording
schedules, since the acoustic information is going to be captured anyway and will be
representative of the community. On the other hand, when the acoustic emissions are
occasional or intermittent and impossible to predict, the sampling should be more
intense to ensure a reliable representation of the soundscape.

39
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
Schedule 5 seemed to most reliably depict the soundscapes captured by
continuous recordings in all of our study areas. This schedule, which is the most
conservative, results already in an 80% storage space and battery power reduction with
respect to the continuous sampling. Schedule 10 seems often to represent a good
compromise, which will correspond to a 90% reduction in respect to the continuous
sampling, and to a 50% reduction of energy and storage space occupied by Schedule 5.
Following these considerations, and with the intention of creating an effective
reproduction of the soundscapes, it could be possible to design robust sampling for the
Atlantic Forest from 06:00 to 17:00, such as Schedule 5 (to avoid important gaps in the
6-8kHz frequency band) for the wet season, while an even less dense sampling routine
could be used from 18:00 to 03:00 without registering a major loss of information
(Schedule 30). In the dry season, it could be enough to record one minute in every 10,
or even every 20, although this risks losing some sounds at 7kHz at dawn.
For the Rupestrian fields in the wet season, a similar solution to that for the
Atlantic Forest wet season should be applied, with the exception being the early
morning hours when it is necessary to record one minute in every 30. In the dry season,
Schedule 10 should be adopted, which would be a good compromise in both the
temporal and spectral domains.
In the Cerrado, schedules 5 or 10 will provide reliable insights into the acoustic
diversity of the community, both for the dry and wet seasons. Schedules 60 and 30
should be avoided, especially when recording at 18:00 (dusk).
Additional insights
Soundscape information can sometimes be misleading and interpreted
incorrectly. Where the weather intervenes significantly in the soundscape of the
environment, as in the Rupestrian fields or the Cerrado (wet season), sounds produced
by the rain and wind mask and interrupt the soundscape of the community, meaning that
weather condition is an additional variable to take into account, with all of its
unpredictability. Moreover, Towsey et al. [38] found that ACI was responsive to wind

40
Determining temporal sampling schemes for passive acoustic
studies in different tropical ecosystems Capítulo 1
gusts, and that it was not inherent to the biological community in adverse
meteorological conditions. Certainly, before conducting brief acoustic surveys, which
rely on only recording for a few days, it is advisable to select the days that may be less
demanding from this point of view. Nevertheless, in the case of long-term investigations
(as in the case of fixed stations detection of acoustic dynamics), a precautionary
schedule should be adopted. In the Atlantic Forest in both the dry and wet seasons, we
found a lower degree of bias due to sounds from adverse weather conditions. Moderate
or strong wind was not noticed in this location, and the rain was easily detectable
because of its natural broadband and dominance signal across the spectrogram. In
contrast, the Rupestrian fields were always very windy, consequently having an
influence on the distribution of sounds across time.
Moreover, we need to underline that it is not possible to extend the results of this
study for all locations in all weather conditions. Soundscape dynamics vary enormously
from an ecosystem to another, and they even tend to differ between two recording
points with the same macroscopic vegetation features on the base of the therein
established animal community. Thus, the analyses here proposed can be representative
of just the three localities taken under investigation and cannot necessarily be extended
to all ecosystems.
Despite of this limitation in the present methodology and the relatively small
number of studied days per season, we believe that our results can provide useful
insights in how to approach the problem of choosing the correct sampling of the sounds
of an ecosystem. Moreover, we trust that the three recording points randomly chosen in
each area were so spaced to be independent and to be good representatives of the
variability of those selected environments.
Clearly more work could be done with other acoustic indices besides the ACI.
Adding further elaborations including a number of other indices could certainly improve
the herein presented results and add more information, so that researchers and field
technicians can have a better understanding of the impact of a particular sampling

41
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
strategy. This could lead to a better support to establish an international accepted
sampling methodology.
As a final consideration, in the present study we tested different sampling
schedules keeping the ON duration fixed to one minute, and varying the OFF period
from four to 59 minutes. It would also be interesting to test whether the one minute
resolution is the best setting for soundscape investigations or if a shorter/longer
recording interval could be more cost-effective.
Implications for conservation
Sounds are valuable scientific specimens that provide an indirect source of
information with which to survey animal dynamics and diversity in particular regions of
interest [3, 7]. The assessment of acoustic temporal and spectral changes offers a new
way to interpret the dynamics of animal communities and, consequently, understand or
address spatio-temporal variations in community structure across space and time [8, 11,
22]. Given the urgency of the issue of climate change and the loss of habitats,
understanding normal levels of variation in acoustic complexity could be fundamental
for conservation efforts, enabling managers to decide whether changes in acoustic
dynamics warrant further investigation.
Herein, we have produced a starting point for what could be a series of research-
guidelines to improve the efficiency of acoustic surveys using analytical methods, by
suggesting the sampling effort needed for planning biologically robust investigations of
animal communities in three tropical environments.
This could be especially useful for wildlife managers who have their choices
linked to economic and staff constrictions. If non-optimal sampling schedules were to
be adopted, our results will help to identify the most critical points, both temporal and
spectral, when the risk of the loss of information is highest.
Future research may focus on the sampling efforts required in temperate areas or
in different tropical ecosystems. Additional insights could be provided by the use of
other indices besides the ACI, or by testing variations in length of the ON period (here

42
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
kept constant to one minute). Suggesting the ideal number of days needed to represent
the acoustic community reliably in different seasons throughout the year would also be
another important step when it comes to designing the best protocol for soundscape
investigation.
These kinds of study are particularly important at this early stage of soundscape
ecology research, since this discipline is demonstrating its suitability to both interpret
the state of health of environments and monitor the anthropogenic challenges that
natural environments face today.
Acknowledgements
We would like to thank the editor, Dr. Alejandro Estrada, and one anonymous
referee for useful comments and constructive suggestions on this manuscript. We
warmly thank all of the staff at the national park of Serra do Cipó, the environmental
station of Peti and the state park of Rola Moça who assisted with our study. We are also
grateful to Marina Scarpelli, Mariane Kaizer and Renan Duarte for their help during the
data acquisition. This study was funded by FAPEMIG and VALE S.A. We would also
like to thank CNPq for their continuing support. R.J.Y. and M.R. were financially
supported by CNPq and FAPEMIG (PPM). The authors declare that there are no
conflicts of interest, financial or otherwise.

43
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
References
[1] Van Parijs, S., Clark, C., Sousa-Lima, R., Parks, S., Rankin, S., Risch, D. and Van
Opzeeland, I. 2009. Management and research applications of real-time and archival
passive acoustic sensors over varying temporal and spatial scales. Marine Ecology
Progress Series 395:21–36.
[2] Luther, D. A. 2008. The evolution of communication in a complex acoustic
environment. ProQuest.
[3] Laiolo, P. 2010. The emerging significance of bioacoustics in animal species
conservation. Biological Conservation 143:1635–1645.
[4] Mellinger, D. K. and Barlow, J. 2003. Future Directions for Acoustic Marine
Mammal Surveys: Stock Assessment and Habitat Use. NOAA OAR Special Report.
Retrieved from:
http://www.beamreach.org/research/whales/Mellinger_AcousticAssessmentWorkshopR
eport.pdf
[5] Brandes, T. S. 2008. Automated sound recording and analysis techniques for bird
surveys and conservation. Bird Conservation International 18(S1).
[6] Sousa-Lima, R. S., Norris, T. F., Oswald, J. N. and Fernandes, D. P. 2013. A review
and inventory of fixed autonomous recorders for passive acoustic monitoring of marine
mammals. Aquatic Mammals 39:23–53.
[7] Blumstein, D. T., Mennill, D. J., Clemins, P., Girod, L., Yao, K., Patricelli, G.,
Deppe, J.L., Krakauer, A.H., Clark, C., Cortopassi, K.A., Hanser, S.F., Mccowan, B.,
Ali, A.M. and Kirschel, A.N.G. 2011. Acoustic monitoring in terrestrial environments

44
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
using microphone arrays: Applications, technological considerations and prospectus.
Journal of Applied Ecology 48:758–767.
[8] Pijanowski, B. C., Farina, A., Gage, S. H., Dumyahn, S. L. and Krause, B. L. 2011.
What is soundscape ecology? An introduction and overview of an emerging new
science. Landscape Ecology 26:1213–1232.
[9] Farina, A. 2014. Soundscape Ecology - Principles, Patterns, Methods and
Applications. Springer.
[10] Farina, A., Lattanzi, E., Malavasi, R., Pieretti, N. and Piccioli, L. 2011. Avian
soundscapes and cognitive landscapes: Theory, application and ecological perspectives.
Landscape Ecology 26:1257–1267.
[11] Farina, A., Pieretti, N. and Piccioli, L. 2011. The soundscape methodology for
long-term bird monitoring: A Mediterranean Europe case-study. Ecological Informatics
6:354–363.
[12] Hobson, K. A., Rempel, R. S., Greenwood, H., Turnbull, B. and Van Wilgenburg,
S. L. 2002. Acoustic surveys of birds using electronic recordings: New potential from
an omnidirectional microphone system. Wildlife Society Bulletin 30:709–720.
[13] Aide, T. M., Corrada-Bravo, C., Campos-Cerqueira, M., Milan, C., Vega, G. and
Alvarez, R. 2013. Real-time bioacoustics monitoring and automated species
identification. PeerJ 1:e103.
[14] Luther, D. 2009. The influence of the acoustic community on songs of birds in a
neotropical rain forest. Behavioral Ecology 20:864–871.

45
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
[15] Farina, A., Pieretti, N. and Morganti, N. 2013. Acoustic patterns of an invasive
species: The Red-billed Leiothrix (Leiothrix lutea Scopoli 1786) in a Mediterranean
shrubland. Bioacoustics 22:175–194.
[16] Sueur, J., Pavoine, S., Hamerlynck, O. and Duvail, S. 2008. Rapid acoustic survey
for biodiversity appraisal. PLoS One 3:e4065.
[17] Depraetere, M., Pavoine, S., Jiguet, F., Gasc, A., Duvail, S. and Sueur, J. 2012.
Monitoring animal diversity using acoustic indices: Implementation in a temperate
woodland. Ecological Indicators 13:46–54.
[18] Rodriguez, A., Gasc, A., Pavoine, S., Grandcolas, P., Gaucher, P. and Sueur, J.
2013. Temporal and spatial variability of animal sound within a neotropical forest.
Ecological Informatics 21 doi:10.1016/j.ecoinf.2013.12.006
[19] Farina, A. and Pieretti, N. 2014. Sonic environment and vegetation structure: A
methodological approach for a soundscape analysis of a Mediterranean maqui.
Ecological Informatics 21:120-132.
[20] Pekin, B. K., Jung, J., Villanueva-Rivera, L. J., Pijanowski, B. C. and Ahumada, J.
A. 2012. Modeling acoustic diversity using soundscape recordings and LIDAR-derived
metrics of vertical forest structure in a neotropical rainforest. Landscape Ecology
27:1513–1522.
[21] Barber, J. R., Fristrup, K. M., Brown, C. L., Hardy, A. R., Angeloni, L. M. and
Crooks, K. R. 2009. Conserving the wild life therein - Protecting park fauna from
anthropogenic noise. Park Science 26(3).

46
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
[22] Joo, W., Gage, S. H. and Kasten, E. P. 2011. Analysis and interpretation of
variability in soundscapes along an urban–rural gradient. Landscape and Urban
Planning 103:259–276.
[23] Pieretti, N. and Farina, A. 2013. Application of a recently introduced index for
acoustic complexity to an avian soundscape with traffic noise. Journal of the Acoustical
Society of America 134: 891–900.
[24] Giulietti, A., Pirani, J. and Harley, R. 1997. Espinhaço range region, eastern Brazil.
Centres of plant diversity: a guide and strategy for their conservation 3:397–404.
[25] Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. B. and Kent,
J. 2000. Biodiversity hotspots for conservation priorities. Nature 403:853–858.
[26] Jacobi, C. M. and Carmo, F. 2008. The contribution of ironstone outcrops to plant
diversity in the Iron Quadrangle, a threatened Brazilian landscape. AMBIO: A Journal
of the Human Environment 37:324–326.
[27] Bertoluci, J., Canelas, M. A. S., Eisemberg, C. C., Palmuti, C. F. de S. and
Montingelli, G. G. 2009. Herpetofauna of Estação Ambiental de Peti, an Atlantic
Rainforest fragment of Minas Gerais State, southeastern Brazil. Biota Neotropica
9:147–155.
[28] Faria, C. M., Rodrigues, M., do Amaral, F. Q., Módena, É. and Fernandes, A. M.
2006. Aves de um fragmento de Mata Atlântica no alto Rio Doce, Minas Gerais:
colonização e extinção. Revista Brasileira de Zoologia 23:1217–1230.
[29] Paglia, A. P., Lopes, M. O. G., Perini, F. A. and Cunha, H. M. 2005. Mammals of
the Estação de Preservação e Desenvolvimento Ambiental de Peti (EPDA-Peti), São
Gonçalo do Rio Abaixo, Minas Gerais, Brazil. Lundiana 6:89–96.

47
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
[30] Nunes, Y. R. F. and Pedralli, G. 1995. Desenvolvimento de metodologia para
adensamento e recomposição da vegetação na EPDA-Peti, MG. BIOS, Cadernos do
Departamento de Ciências Biológicas da PUC-MG, Belo Horizonte 2:53–61.
[31] Jacobi, C. M., do Carmo, F. F., Vincent, R. C. and Stehmann, J. R. 2007. Plant
communities on ironstone outcrops: a diverse and endangered Brazilian ecosystem.
Biodiversity and Conservation 16:2185–2200.
[32] Oliveira, P. S. and Marquis, R. J. 2002. The cerrados of Brazil: ecology and
natural history of a neotropical savanna. Columbia University Press, New York.
[33] Rodrigues, M., Carrara, L. A., Faria, L. P. and Gomes, H. B. 2005. The birds of“
Parque Nacional da Serra do Cipó”: the Rio Cipó valley, Minas Gerais, Brazil. Revista
Brasileira de Zoologia 22:326–338.
[34] Oliveira, V. B., Câmara, E. M. and Oliveira, L. C. 2009. Composiçao e
caracterizaçao da mastofauna de médio e grande porte do Parque Nacional da Serra do
Cipó, Minas Gerais, Brasil. Mastozoología neotropical 16:355–364.
[35] Minuzzi, R. B., Sediyama, G. C., Barbosa, E. and Melo Júnior, J. 2007.
Climatologia do comportamento do período chuvoso da região sudeste do Brasil.
Revista Brasileira de Meteorologia 22:338–344.
[36] Sjölander, K. and Beskow, J. 2000. Wavesurfer-an open source speech tool. In
INTERSPEECH 464–467.
[37] Farina, A., Lattanzi, E., Piccioli, L. and Pieretti, N. 2012. The soundscapemeter
user manual. www.disbef.uniurb.it.

48
Determining temporal sampling schemes for passive acoustic studies in different tropical ecosystems Capítulo 1
[38] Towsey, M., Wimmer, J., Williamson, I. and Roe, P. 2014. The use of acoustic
indices to determine avian species richness in audio-recordings of the environment.
Ecological Informatics 21:110-119.
[39] Wimmer, J., Towsey, M., Roe, P. and Williamson, I. 2013. Sampling
environmental acoustic recordings to determine bird species richness. Ecological
Applications 23:1419–1428.
[40] Sueur, J., Farina, A., Gasc, A., Pieretti, N. and Pavoine, S. 2014. Acoustic indices
for biodiversity assessment and landscape investigation. Acta Acustica united with
Acustica 100:772–781.
[41] Pieretti, N., Farina, A. and Morri, D. 2011. A new methodology to infer the singing
activity of an avian community: The Acoustic Complexity Index (ACI). Ecological
Indicators 11:868–873.
[42] Bennett, J. O. and Briggs, W. L. 2008. Using and understanding mathematics: A
quantitative reasoning approach. Pearson Addison Wesley.
[43] Aichinger, M. 1987. Annual activity patterns of anurans in a seasonal neotropical
environment. Oecologia 71:583–592.
[44] Haddad, C., Sazima, I. and Morellato, L. 1992. Anfíbios anuros da Serra do Japi.
História natural da Serra do Japi: Ecologia e preservação de uma área florestal no
sudeste do Brasil (LPC Morellato, ed.). Editora da Unicamp/FAPESP, Campinas188–
211.
[45] Bormpoudakis, D., Sueur, J., Pantis, J.D. 2013. Spatial heterogeneity of ambient
sound at the habitat type level: ecological implications and applications. Landscape
Ecology 28:495–506.

49
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
- CAPÍTULO 2-
The impact of anthropogenic noise from open cast mining on Atlantic
forest biophony
Artigo submetido ao periódico Biological Conservation
Abstract
Anthropogenic noise (anthropophony) is known to cause negative impacts on animal
communication and wellbeing. Mining is an important economic activity in Brazil,
which is often conducted close to forested areas and produces a diffuse noise. In this
study we investigated the impact of such noise on biophony (biological sounds) by
characterizing and comparing the soundscapes of two different sites (close versus far
from an open cast mine) in the same Atlantic forest fragment matched for habitat type in
Southeast Brazil. Six Song Meters (SM2) were installed in each site and programmed to
record continuously during seven continuous days every two months from October 2012
to August 2013. Anthropophony and biophony values were derived from power spectra
and the Acoustic Complexity Index (ACI). As predicted, anthropophony was
significantly higher closer to the mine site. Biophony was significantly higher in the wet
season at both sites. Anthropophony was significantly higher in the wet season close to
the mine. The soundscape of the site close to the mine presented higher biophony during
the day and higher anthropophony levels at night whereas the site far from the mine
showed higher biophony during the night. Potential species richness was higher at the
site far from the mine. The animal community composition and the spectral
characteristics of the calls were different between the two sites. Thus, here we have
shown that mining noise can affect biophony dynamics by modifying the temporal
distribution and daily patterns of animal sounds. These results provide important
information to be taken into consideration during the regulation of the use of natural
areas for mining.
Keywords: Acoustic Complexity Index, Atlantic forest, biophony, mining activity.

50
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
1. Introduction
Opencast mining can produce high sound pressure levels by exploratory and
production drilling, blasting, cutting, handling of materials, ventilation, crushing,
conveying and ore processing (Donoghue, 2004). This massive noise pollution can
negatively impact wildlife. Mining has been shown to impact breeding birds by
reducing their density (Smith et al., 2005), their species diversity, and their population
sizes (Saha and Padhy, 2011). Ant species richness also decreases due to mining activity
(Queiroz, 2013). Despite the evidence that noise pollution negatively affects wildlife
reproduction and longevity (Warren et al., 2006; Slabbekoorn and Ripmeester, 2008;
Barber et al., 2009; Francis et al 2011; Kight and Swaddle 2011), sound pollution from
mining activity is still poorly regulated around the world (Hessel and Sluis-Cremer,
1987; Frank et al., 2003).
Many animal species depend on acoustic signals for intraspecific communication
(Catchpole and Slater, 2008). Several studies have shown that noise may reduce habitat
quality for many species (Bayne et al., 2008) by masking sound signals and decreasing
the efficiency of animal communication (Langemann, et al 1998; Lohr et al., 2003;
Brumm, 2004; Bee and Swanson, 2007). Noise can also decrease reproductive success
(Halfwerk et al., 2011), alter mating systems (Swaddle and Page, 2007; Habib et al.,
2007) and parental care in bird species (Schroeder et al., 2012). Nonetheless, some
animal species are capable of adjusting their acoustic signals to communicate in noisy
environments, for example, increasing their amplitude (Brumm et al., 2004; Brumm et
al., 2009), shifting frequencies (Slabbekorn and Peet 2003; Parks and Clark 2007;
Nemeth and Brumm, 2009), calling rates (Sun and Narins, 2005), call duration (Brumm
et al., 2004) or time of calling (Fuller et al., 2007; Sousa-Lima and Clark, 2008). Other
species present changes in behavior by avoidance of noisy areas for foraging (Miksis-
Olds et al., 2007; Schaub et al., 2008), and other daily activities (Sousa-Lima and Clark,
2009; Duarte et al., 2011). Area avoidance and acoustic compensatory mechanisms to
reduce or offset the effects of noise may alter the acoustic complexity of a community
in a given location and result in a decrease in species’ abundance (Bayne et al., 2008)

51
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
and/or diversity (Proppe et al., 2013) in noise polluted sites.
Anthropogenic noise has become omnipresent in natural soundscapes (Barber et
al., 2011) and despite evidence showing negative impacts on animals, there is still a lack
of official regulation of noise produced by industrial and exploratory activities in
terrestrial natural areas. The Atlantic forest in Brazil is one of the richest and most
endangered biomes of the world (Myers et al., 2000) where much mining activity takes
place. Despite this, there are no laws regulating sound pollution levels allowed in this
biome. In many countries of the world, noise monitoring from industrial activities is
required only in respect to its impacts on human health. Thus, the already known
impacts of noise on wildlife should drive efforts to develop environmental legislation to
protect wildlife (Brown et al., 2013).
Passive acoustic monitoring (PAM) methods provide opportunities to evaluate
the consequences of different land use decisions (Blumstein et al., 2011; Mennit and
Fristrup 2012; Brown et al., 2012 and 2013), especially in environments such as mines,
that are difficult to access or monitor using conventional methods (Mellinger and
Barlow 2003; Scott Brandes, 2008). PAM devices can record data during several days
continuously and hence, a large amount of information can be collected from the
acoustic environment. As a result, special software and indices to rapidly and efficiently
process audio files are required (Sueur et al., 2014). In this context, Pieretti et al. (2011)
introduced the Acoustic Complexity Index (ACI), which allows an indirect and rapid
measure of the complexity of the soundscape. The ACI has been proven to be a useful
tool in tracking the dynamics of the sounds produced by animal communities (Farina et
al., 2013) by describing the spectral complexity of the biophony of soundscapes,
through the intrinsic variability of biotic sounds. This index has already been used in
noisy environments (Pieretti et al., 2011; Pieretti and Farina, 2013) and Towsey et al.
(2014) indicate ACI as one of the best indicators of bird biodiversity among 14 different
acoustic indices.
There are no studies investigating how anthropogenic noise affects soundscapes
and biophony in mining areas. The aim of this study was to investigate noise effects on

52
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Atlantic forest soundscape dynamics by comparing biophony and anthropophony in a
site close to an active opencast mine and at a habitat matched site far from any mine or
anthropogenic activities.
2. Methodology
2.1 Study area
Data were collected at the Environmental station of Peti in the municipalities of
São Gonçalo do Rio Abaixo and Santa Bárbara, Minas Gerais state, Brazil (centered at
19°53’57’’S and 43°22’07’’W). The climate of southeastern Brazil can be divided into
two macro-climatic seasons: a hot wet season, from October to March, and a cooler dry
season from April to September (Minuzzi et al., 2007).
The reserve is an Atlantic forest fragment of approximately 605 ha located in the
upper Rio Doce Basin (altitude range: 630-806m). It is estimated that the area harbors
approximately 29 species of anurans (Bertoluci et al., 2009), 231 species of birds (Faria
et al., 2006) and 46 species of mammals (Paglia et al., 2005). A large part of the reserve
is covered by secondary arboreal vegetation of continuous canopy and large trees
(Nunes and Pedralli, 1995).
Peti is surrounded by small farms and is contiguous with the Brucutu Mine,
which occupies an area of 8km2 and produces noise through road traffic, sirens and
explosions during the day and night (Roberto, 2010). Brucutu’s iron ore extraction
started in 1992 and currently is one of the largest mines of the world (Roberto, 2010).
2.2 Acoustic recordings and data analysis
Sensor arrays comprised by six Song Meter Digital Field Recorders (SM2)
(Wildlife Acoustics, Inc., Massachusetts) distributed in two triangles were installed in
two sites and programmed to record continuously during seven days every two months

53
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
from October 2012 to August 2013 (six recording sessions). Both sites were matched by
habitat and located in the same Atlantic forest fragment. The 6-SM2 array close to the
active opencast mine was installed at a distance of 500m from the mine and 25m from
the closest mining road. The 6-SM2 array far from the mine was installed at a distance
of approximately 2 500m from the mine and 25m from a rarely used road in order to
control for a potential border effect due to the physical structure of the road (Fig 1).
Figure 1. Position of the passive acoustic monitoring devices close (1) an far (2) to the
mine site at Peti Environmental station, Southeast Brazil.
To avoid overlap of sounds recorded, each SM2 within each sensor triangle was
placed 80m from each other. This distance between recorders was established during a
pilot study conducted in the area. The distance between two SM2 triangles was at least
100m in order to have two independent recording samples in each site (close and far
from the mine). The distance between arrays (far and close sites) was approximately 2
300m (Fig 1). The triangular array geometry was chosen to have one SM2 at the forest
border and two located 80m towards the interior of the forest.

54
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Each SM2 was fixed on a tree at 1.5m above the ground and placed in order to
have the two lateral microphones clear of any surface that could be an obstacle to
incoming sound waves. They were configured to record in wave format at a sampling
rate of 44.1 kHz, at 16 bits. One SM2 was stolen during the 5th session (at the site close
to the mine), and the 2nd session was not considered for one SM2 installed in the site
far from the mine because the geophony produced by a flooded river masked all
incoming sounds.
The collected data were sub sampled by analyzing the first two minutes of
recordings every hour. The resulting 23 520 minutes (392 hours) were further processed
using the Wavesurfer software (Sjölander and Beskow 2000) powered by the
SoundscapeMeter plug-in (Farina et al., 2012). A Fast Fourier Transform (FFT) of 512
points was applied to obtain from every two-minute file a matrix made by 256
frequency bins of 86.13Hz and 10 335 time intervals of 0.012s. The resulting database
of power spectra (i.e., the sound energy values along a frequency axis in each temporal
interval) was used to analyze and describe two sonic components of the soundscape in
each site: anthropophony and biophony.
All files were separated into two frequency bands: 1) 0–1.5 kHz (predominantly
occupied by noise or anthropophony) and 2) 1.5–22.05 kHz (mainly occupied by
biophony). The lower frequency band was used to characterize noise by considering the
raw values expressed in the power spectrum and the second band was further processed
to extract values of the Acoustic Complexity Index (ACI) (Farina et al., 2011; Pieretti et
al., 2011). The threshold of 1.5 kHz was chosen since most of the energy of the
anthropogenic noise is mainly concentrated under 2kHz (Warren at al., 2006), and
lowered 500Hz to prevent the exclusion of some important biophonies from the ACI
calculations that were just above 1.5 kHz (Pieretti and Farina, 2013). Nonetheless, in the
site closest to the mine, noise produced by truck transits often covered frequencies up to
7-8 kHz, sometimes reaching upwards of 21 kHz. To avoid bias from these specific
events in the ACI estimations of the biological sound expression, a specific routine was
elaborated in JustBasic v.1.01 to recognize and eliminate from the recordings every
recorded truck passing. This allowed us to focus only on the features of biophony

55
The impact of anthropogenic noise from open cast mining
on Atlantic forest biophony Capítulo 2
above the threshold of 1.5 kHz.
Sound recording files dominated by rain or wind, which can also influence ACI
estimations, were eliminated from the analysis.
Noise levels at the sites close and far from the mine were compared by
conducting two 20 minutes measurements of the background sound pressure levels at
each SM2 recording point using a A-weighted B&K2270 sound level meter. We
excluded from the recordings all the animal sounds close to the microphone using the
BZ5503 software. The standard sound pollution measurements of minimum (Lmin),
maximum (Lmax) and equivalent sound levels (Leq) were then extracted from the
recordings (Rossing 2007). The number of passings of the mining trucks per day was
determined by listening to the recordings made at the site close to mine during 24 hours
in two days in each recording session using Raven Pro 1.5. Recurrent sounds produced
by the mine were classified and characterized. Different types of anthropogenic sounds
were selected from two days of recordings (48 hours) from one SM2 at the site close to
the mine in each recording session. The two most frequent types of noise were truck
passing noise and reverse warning sound of trucks. Twenty noise events were randomly
selected per day, totalizing 240 truck traffic events and 240 reversing sound events. For
the less frequent noises, such as explosions, horns, and sirens, all events heard in two
days were selected. These noise events were described using Raven Pro 1.5 by
measuring their Min, Max, Peak frequency and Duration.
Along with noise, variables such as species richness, species composition and
abundance could have influenced the acoustic dynamics of the two sites. To account for
such differences between the sites close and far from the mine, species richness was
obteined for each site by the aural identification of animals’ sounds using Raven Pro 1.5
software. A single day of recording per session in four points (two in the close and two
in the far site) were randomly selected for species identification surveys. Sounds
emitted by amphibians, birds, mammals, and insects were identified by specialists in the
two minute files from 0500 to 0700 hours, 1000 to 1200 hours, and from 1800 to 2200
hours, totalizing 528 analyzed minutes.

56
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
The above time slots were chosen to include: the dawn and dusk choruses of
birds and the midday and night activity of insects. It was not possible to determine
species abundance aurally due to the huge number of calls recorded. Insects and bats
sounds were classified as soundmorphs (different sound emissions or codas). This
procedure was essential to identify potential species since biodiversity in the Atlantic
forest is so high that it is impossible to identify every species by aural census – and
there is a chance that some species recorded are not yet taxonomically classified.
We extracted from the recordings the bandwidth, minimum and maximum
frequencies of bird vocalizations and insect stridulations (most representative groups),
that were identified only at one of the two sites (close or far from the mine) to compare
the acoustic niche occupation of the singing community between the two sites.
2.3 Statistical analysis
Data analyses were separated into wet and dry seasons, and into day (5 am to 5
pm) and night (6 pm to 4 am). We conducted two analyses considering the time of the
day: (1) comparison of ACI day x ACI night in each site separately; and (2) comparison
between sites of ACI day and ACI night.
Preliminary analyses showed that sample points on the border (closer to the
roads), at both sites, were noisier than the other points, therefore data analyses also
included the groups: border and forest points.
All the statistical tests were performed using Statistica v.8.0. A non-parametric
approach was used, since the variables were not normally distributed, even after
attempted transformation of the data values. Mann Whitney U tests were conducted to
test for differences in ACI and noise values between: sites (close and far from the
mine); seasons (wet and dry); and time period (day and night).

57
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
3 Results
3.1 Mining noise characterization
Sites close and far from the mine differed significantly in terms of background
noise. The site close to the mine showed levels 3 to 36 dB(A) higher in comparison to
the site far from the mine (T=2.94, DF=5, p<0.05). Mean Leq, Lmax and Lmin of each
type of soundscape are shown in Table 1. The noise measured using power spectral
density confirmed the results of the noise level measurements, showing that noise was
significantly higher at the site close to the mine both on the border and on the interior of
the forest (Border: U=118, Z= 20.70, p<0.01, Nclose=288, Nfar=288; Forest: U=3556,
Z=28.4, p<0.01, Nclose=552, Nfar=575).
Table 1. Mean noise levels measurements in sites close and
far from an opencast mine at Peti environmental station,
Minas Gerais, Brazil.
Site Leq dB(A) L max dB(A) L min dB(A)
Close (border) 51.7 78.0 30.9
Close (forest) 41.2 64.1 30.1
Far (border) 35.5 41.3 27.4
Far (forest) 33.2 48.9 23.3
The five most frequent mining noise sources identified in the site close to the
mine were: trucks passing, reversing alarm of trucks, work sirens, horns, and explosions
(Fig. 2). The most frequent noise was truck transiting. A mean of 700±43.8 (mean±SD)
trucks passed daily (29.91±1.82 trucks/hour) in the wet season and 244.6±57
(10.91±2.37 trucks/hour) in the dry season. The descriptive statistics of the acoustic
parameters of each noise event type are presented in Table 2.
58
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Table 2. Acoustic variables of most frequent noise sources from mining activity at Peti
environmental station, Minas Gerais, Brazil.
Noise source Duration (s)
Average ±SD
Peak Freq.(Hz)
Average ±SD
Max Freq.(Hz)
Average ±SD
Min Freq.(Hz)
Average ±SD
Truck
N=240
20.2±8.9 553.9±38.6 15291.0±43.8 0.0±0.0
Reversing
N=240
10.56±2.54 1314.6±91.4 1373.9±90.6 1255.1±91.4
Siren
N=75
17.9±4.6 1229.4±136.1 1393.6±101.4 872.3±144.9
Horn
N=19
6.7±14.5 1219.0±98.5 4747.0±55.4 781.0±75.9
Explosion
N=19
5.9±3.2 197.2±117.8 4353.0±206.0 90.4±30.7

59
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Figure 2. Spectrograms of the noise produced by: A- transit of trucks, B- explosion, C-
work sirens, D-horns, E-reversing alarm of trucks on a mining road at Peti
environmental station, Southeast Brazil. In the background of the spectrograms there is
also biophony.

60
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Considering that a mean of 700 trucks passed per day in wet season with a mean
duration of 20.2 seconds, this means 16.2% of the time of the day was occupied by
truck transiting noise. The mean maximum frequency of this type of noise event was
15.2 kHz, meaning that the noise occupied 68% of the full spectrogram bandwidth
(22.05 kHz).
3.2 Soundscape dynamics
3.2.1 Wet versus dry season
The ACI was significantly higher in the wet season than in the dry season at both
sites (Close border: U=4824, Z= 7.84, p<0.01; Nwet=144; Ndry=144; close forest: U=
20759, Z= 8.04, p<0.01; Nwet=264; Ndry=264; Far border: U= 5915, Z= 6.3, p<0.01;
Nwet=144, Ndry= 144; Far forest: U= 23689, Z= 7.65, p<0.01; Nwet= 288, Ndry=264).
Noise showed a similar trend except for the interior of the forest at the site close the
mine (Close border: U=6531, Z= 5.42, p<0.01; Nwet=144, Ndry 144; Close forest:
U=37359, Z=-0.35, p= 0.72; Nwet=288, Ndry=264).
3.2.2 Day versus night
In the wet season, at the site far from the mine, the ACI values were significantly
higher during the night (Border: U=1220, Z=5.42, p<0.01; Nday=78 Nnight=66; Forest:
U=3386, Z=8.51, p<0.01; Nday= 143, Nnight=121). Contrastingly, there was no difference
between the ACI values of day and night at the site close to the mine (Border: U=2481,
Z=0.37, p=0.70; Nday=78, Nnight=66; Forest: U=8498, Z=-0.24, p=0.80; Nday=143,
Nnight=121). Noise values were significantly higher during the night at the site close to
the mine (Border: U=1884, Z=2.76, p<0.01; Nday= 78, Nnight=66; Forest: U=7304,
Z=4.24, p<0.01; Nday=156, Nnight=132).

61
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
In the dry season, the ACI values were significantly higher during the night at
both sites except on border points at the site close to the mine (Far border U=1987,
Z=2.35, p<0.01; Nday=78, Nnight=66; far forest: U=6840, Z=4.9, p<0.01; Nday=156,
Nnight=132; close border U= 2279, Z= 1.18, p=0.23; Nday=78, Nnight=66; close forest:
U=5595, Z=4.94, p<0.01; Nday=143, Nnight=121). At the site close to the mine, noise
values were also significantly higher during the night, but only on the forest points
(Forest: U=6764, Z=3.05, p<0.01; Nday=143, Nnight=121; Border: U=2276, Z=1.19,
p=0.23; Nday=78, Nnight=66).
3.2.3 Close versus Far site
In the wet season during the night the ACI was significantly higher at the site far
from the mine (Fig.3, A,B) (Border: U=1510, Z=-3.04, p<0.01; Nclose=66, Nfar=66;
Forest: U=5145 Z=-3.99, p<0.01; Nclose=121, Nfar=121) and during the day at the site
close to the mine (Border: U=2032, Z=3.57, p<0.01; Nclose=78, Nfar=78; Forest: U=
6191, Z=5.76, p<0.01, Nclose=143, Nfar=143).
In the dry season, there was no significant difference in ACI values at night
between the two sites (Fig 4, A,B) (Border: U=2070, Z=-0.49, p=0.62; Nclose=66
Nfar=66; Forest: U= 7156, Z=-1.42, p=0.15; Nclose=121, Nfar=132), while during the day
ACI values were significantly higher at the close site except at the border points
(Border: U= 2633, Z= -1.44, p=0.14; Nclose=78, N far=78; Forest: U= 8989, Z= -2.89,
p<0.01; Nclose=143, Nfar=156).

62
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Figure 3. Temporal distribution of the ACI values at border (A) and forest (B) points at sites
close and far from an opencast mine during the wet season at Peti Environmental Station,
Brazil.
Figure 4. Temporal distribution of the ACI values at border (A) and forest (B) points at sites
close and far from an opencast mine during the dry season at Peti Environmental Station,
Brazil.
3.3 Characterization of fauna
A total of 91 bird species (16 were classified as "not identified" due to the short
duration of the song or long distance from the microphone, which prevented
identification), 84 different soundmorphs of insects, 9 of bats, 3 species of frogs and 2
species of primates were identified. In both sites, the insect community was particularly
acoustically active during the night although cicadas were highly active during the day.

63
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
The bird community showed greater acoustic activity during the day, mainly
during the dawn chorus, with few species singing during the night. Primates were
especially vocal early in the morning. Insect species were estimated to be higher in the
wet season and especially during the night. Bat calls were detected only at night. The
richness of species was higher: (1) during the wet season on both sites; (2) during the
day in both seasons and sites; and (3) on the site far from the mine, especially in wet
season. Species richness results from the aural survey are shown in Table 3. Insects
species detected only at the site far from the mine presented stridulations with
significantly larger bandwidth (bandwidthclose=1777Hz±378, bandwidthfar=3233Hz±517;
U=3391.5, Z=-3.88, p<0.01, Nclose=80, Nfar=125), higher maximum (max.
freq.close=6117Hz±360, max.freq.far=8560Hz±615; U=3893, Z=-2.67, p<0.01, Nclose=80,
Nfar=125) and minimum frequencies (min. freq.close=4340Hz±213 min.freq.far=5326Hz ±
262; U=3877, Z=-2.71 p<0.01, Nclose=80, Nfar=125) than species which occurred only at
the site close to the mine. The opposite occurred with bird species. Species recorded
close to the mine presented significantly larger bandwidth (bandwidthclose=2189±164,
bandwidthfar=2088±212; U=11723, Z=2.16, p<0.05, Nclose=160, Nfar=170), higher
maximum (max. freq.close=5832Hz ±243, max.freq.far=3425Hz ± 249, U=6039, Z=8.72,
p<0.01, Nclose=160, Nfar=170) and minimum frequencies than species from the site far
from the mine (min. freq.close= 3643Hz ±172, min.freq.far =3425H ± 249, U=11110,
Z=2.87, p<0.01, Nclose=160, Nfar=170).
64
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Table 3. Potential number of species at close and far sites from an opencast mine at the Peti Environmental Station, Brazil by season (wet
versus dry) and time of day (day versus night).
CLOSE FAR
WET DRY WET DRY
Taxonomic
Group Day Night Total Day Night Total Day Night Total Day Night Total
Species
in
common
Species
only in
close
Species
only in
far
Insects 25 36 49 15 23 33 26 42 54 12 27 36 43 16 25
Amphibians - 1 1 - - - - 2 2 - 1 1 1 - 1
Birds 40 - 40 34 - 34 43 - 43 34 1 35 25 32 34
Bats - 4 4 - 7 7 - 5 5 - 2 2 6 2 1
Primates 2 - 2 2 - 2 2 - 2 2 - 2 2 - -
Total species 67 41 96 51 30 76 71 49 106 48 31 76 77 50 62

65
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
4 Discussion
Large scale anthropogenic activities can have a considerable impact on the
accomplishment of daily ecological functions of a community (Francis et al 2011);
especially on acoustic communication processes (Rabin et al., 2003, Slabbekoorn and
Ripmeester, 2008). Noise is one of the most common threats to landscapes around the
world due to its established negative impacts on fauna (Brown et al., 2013, Pieretti and
Farina, 2013). Although mining is an important economic activity in many parts of the
world, its subtle effects on animal ecosystems are still poorly understood. The approach
taken in this study of investigating acoustic dynamics has recently been considered as a
proxy for biodiversity measurement (Krause, 1987; Sueur et al., 2008) and can also
provide additional information related to species’ adaptation and the well being of
animal communities (Farina et al., 2011).
Our results showed that there were significant differences between the sites close
and far from the mine in terms of anthropogenic noise levels. Considering that the
mining activity has been ongoing in the study area for decades, changes in the behavior
of the animal community could be interpreted as long term responses to mining impacts.
Noise sources in our study area were diverse, continuous and occupied a wide
frequency bandwidth potentially masking many animal sounds and affecting their
behavior and distribution. For example, some birds with low frequency vocalizations
were only recorded far from the mine such as Patagioenas plumbea, Leptotila sp.,
Leptotila verreauxi, and Ramphastos toco (see supplementary material - table 1).
During the wet season almost 70% of the frequency bandwidth (up to 22kHz) was
completely occupied by the truck transit noise for 16% of the day time, interfering with
the acoustic space used by the animal community.
As expected, the acoustic complexity registered during the wet season was much
higher than in the dry season. The wet season in Brazil coincides with breeding season
of insects, amphibians and birds; thus, animals are more acoustically active (Aichinger,
1987; Haddad et al., 1992; Rodrigues et al., 2005). This result was confirmed by the

66
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
species count, since a higher number of species from all of the animal groups was
detected during the wet season.
Our results showed a change in soundscape dynamics resulting from proximity
to the mine. In the wet season, the site close to the mine lost the diel pattern of having
higher acoustic activity at night found at the site far from the mine (Fig. 3), presenting a
tendency of higher acoustic complexity along the day and lower along the night. This
phenomenon can be explained by two possible reasons: (1) noise levels are lower during
daytime and the acoustic community can be more acoustically active during the day
when there is more available acoustic space and less competition with anthropogenic noise; (2) acoustic activity might be higher than expected during the day simply because
there are more singing species, which affects the ACI values. The first explanation is
most likely because at the site far from the mine, species richness was higher during the
day and ACI was higher during the night. Greater acoustic activity during the night was
expected at both sites because the majority of biophony was due to the insects. This
animal group is mostly active during the night and produces long songs with high
amplitude that results in high ACI values (Farina et al. 2011).
Comparison of the same period of the day between sites (close day versus far day and
close night versus far night) yielded similar results. Higher acoustic activity at the site
close to the mine during the day could also be explained by anthropogenic noise. During
the day the number of species recorded was higher at the site far from the mine and we
expected a higher ACI value as well. However, we observed the opposite. We suggest
that this result might be related to compensatory mechanisms of individuals trying to
propagate their signals with greater emphasis (higher amplitude or repetition of the
strophes or syllables) to override the masking effect of anthropogenic noise. The
number of individuals singing might have an effect on ACI values, but unfortunately,
species abundance was impossible to assess. Other studies have found similar results in
different environments. Pieretti and Farina (2013) found that both ACI values resulting
from birds and noise were significantly higher with greater proximity to a road,
indicating a more active singing/calling community where noise is more intense.

67
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
The animal community in an urban forest in Brazil also presented higher activity
in noisy sites (Santos, 2013). Birds and mammals can present a behavior known as the
Lombard Effect in which animals increase the amplitude of their calls in the presence of
high levels of competing environmental noise (Cynx et al., 1998; Brumm et al., 2004,
Brumm and Slater, 2006). Additionally, many species are capable of increasing the rate
and duration of their vocalizations to ensure the efficiency of their communication
(Brumm et al., 2004, Sun and Narins, 2005). Greater amplitudes, higher emission rates
and longer call duration could all lead to an increase in ACI values and explain our
results.
The higher ACI values at night in the site far from the mine in comparison with the
values from the same period in the site close to the mine may be a direct effect of a
higher number of species vocalizing, especially insects. Noise can affect species
diversity and population density of birds in areas close to mining activity (Saha and
Padhy 2011). Our results show that the number of species was lower at the site close to
the mine and species composition was different between the two sites. Many studies
concerning the effects of road noise on animals show that there is a strong negative
relationship between traffic intensity and species richness with changes in composition
and density of individuals (Forman et al., 2002; Rheindt, 2003). Bayne et al.
(2008) showed that near noiseless energy facilities passerine density was 1.5 times
higher than areas near noise-producing energy sites. Other factors can contribute to a
low species richness, abundance and diversity in noisy environments; for example,
quality of habitat, vegetation characteristics, air and chemical pollution, soil vibration
and others (Summers et al., 2011). Therefore, due to the importance of acoustic
communication, which animals use to find food (Elowson et al., 1991; Slabbekoorn and
Ripmeester, 2008) and reproductive partners (Patricelli et al., 2002); to escape from
predators (Greig-Smith, 1980; Chan et al., 2010); to defend resources (Zuberbuehler et
al., 1997) just to name the main functions, it is expected that noise will affect species
richness, abundance and community composition. The acoustic measurements of the
insect stridulations and bird vocalizations showed other possible effects of
anthropogenic noise on the animal communication.

68
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
The greater bandwidth of the insect stridulations in the site far from the mine
could be interpreted as the natural pattern that evolved in the absence of anthropogenic
noise. In contrast, the noise present in the site close to the mine could be selecting for
species with narrowband stridulations, since a considerable part of the local acoustic
space is occupied by anthropogenic noise. Species that produce sounds which occupy
less acoustic space can better cope with the competition from noise given that the
probability of overlap would be reduced, especially if the spectral occupancy pattern of
these animals’ sounds overlaps with less intense bands of noise. Bird vocalization
analyses showed different results. Species with sounds with larger bandwidths were
recorded in the site close to the mine. Additionally, bird calls at the site close to the
mine presented higher maximum and minimum frequencies in comparison with species
from the site far from the mine. Hence, it could be speculated that the pervasive noise in
the site close to the mine could be selecting species which vocalize at higher frequencies
and are less masked by the noise. This can be confirmed by the absence of species,
which vocalize at very low frequencies in that site. Rheindt (2003) has shown that there
is a significant relationship between dominant frequency of bird vocalizations and
decline in abundance towards the motorway, indicating that having a higher pitched
song with frequencies above those of traffic noise makes birds less susceptive to
anthropogenic noise. Hence, our result supports the hypothesis that noise can affect the
animal community by changing its singing dynamics.
5. Conclusion
Many studies have shown the negative impact of noise pollution on animal
acoustic communication and other studies have shown negative impacts on species
diversity, richness and abundance. Nevertheless, studies about the impact of
anthropogenic noise on the biophony in the terrestrial soundscapes in tropical
environments are still inexistent. Here we have shown that sound pollution from
opencast mining activities has a significant impact on the biophonical soundscape of a
neighboring tropical forest. Differences found in soundscape complexity were probably

69
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
related to lower species richness at the site close to the mine, changes in animal
community composition, spectral characteristics of calls between the two sites and
possible animal adaptive responses to noise. Given that opencast mining is a major
global economic activity, which frequently occurs in natural areas, these results show
the need for its noise impact to be taken into consideration during the elaboration of
conservation and management strategies of natural areas close to mining activity.
Beside this, we provide data to enhance the importance of the elaboration of laws and
regulations to monitor and control noise close to natural areas.
6. Acknowledgements
We would like to thank all of the staff at the environmental station of Peti who
assisted with our study, especially Leotacílio da Fonseca. We are also grateful to Marina
Scarpelli, Mariane Kaizer and Renan Duarte for their help during data acquisition and
the engineer Krisdany Cavalcante for help with the noise level measurements.
7. Role of the funding source
We would like to thank CNPq for their continuing support. RJY and MR were
financially supported by CNPq and FAPEMIG (PPM). MHLD was supported by a
FAPEMIG postgraduate scholarship during this research. This study was funded
competitively by FAPEMIG from a financial donation made by VALE, but VALE did
not in any way restrict our research or contribute to its design, execution or publication.

70
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
8. References
Aichinger, M., 1987. Annual activity patterns of anurans in a seasonal
neotropical environment. Oecologia. 71, 583-592.
Barber, J.R., Crooks, K.R., Fristrup, K.M., 2009. The costs of chronic noise
exposure for terrestrial organisms. Trends in Ecology and Evolution. 25, 180-189.
Barber, J.R., Burdett, C.L., Reed, S.E., Warner, K.A., Formichella, C., Crooks,
K.R., 2011. Anthropogenic noise exposure in protected natural areas: estimating the
scale of ecological consequences. Landscape Ecology. 26, 1281-1295.
Bayne, E.M., Habib, L., Boutin, S., 2008. Impacts of chronic anthropogenic
noise from energy-sector activity on abundance of songbirds in the boreal forest.
Conservation Biology. 22,1186-1193.
Bee, M.A., Swanson, E.M., 2007. Auditory masking of anuran advertisement
calls by road traffic noise. Animal Behaviour. 74, 1765-1776.
Bertoluci, J., Canelas, M.A.S., Eisemberg, C.C., Palmuti, C.F. de S.,
Montingelli, G.G.,2009. Herpetofauna of Estação Ambiental de Peti, an Atlantic
Rainforest fragment of Minas Gerais State, southeastern Brazil. Biota Neotropica. 9,
147-155.
Blumstein, D.T., Mennill, D.J., Clemins, P., Girod, L., Yao, K., Patricelli, G.,
Deppe, J.L., Krakauer, A.H., Clark, C., Cortopassi, K.A., Hanser, S.F., McCowan, B.,
Ali, A.M., Kirschel, A.N.G., 2011. Acoustic monitoring in terrestrial environments
using microphone arrays: applications, technological considerations and prospectus.
Journal of Applied Ecology. 48, 758-767.

71
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Brown, C.L., Hardy, A.R., Barber, J.R, Fristrup, K.M, Crooks, K.R, et al., 2012.
The effect of human activities and their associated noise on ungulate behavior. Plos
One, 7.
Brown, C.L., Reed, S.E., Dietz, M.S, Fristrup, K.M., 2013. Detection and
classification of motor vehicle noise in a forested landscape. Environmental
Management. 52,1262-1270.
Brumm, H., 2004. The impact of environmental noise on song amplitude in a
territorial bird. Journal of Animal Ecology. 73, 434.
Brumm, H., Slater, P.J.B., 2006. Animals can vary signal amplitude with
receiver distance: evidence from zebra finch song. Animal behaviour. 72, 699-705.
Brumm, H., Schmidt, R, Schrader L., 2009. Noise-dependent vocal plasticity in
domestic fowl. Animal Behaviour. 78, 741-746.
Catchpole, C.K., Slater, P.J.B., 2008. Bird Song: Biological themes and
variations. Cambridge University Press, Cambridge.
Chan, A.A.Y-H, Giraldo-Perez, P., Smith, S., Blumstein, D.T., 2010.
Anthropogenic noise affects risk assessment and attention: the distracted prey
hypothesis. Biology Letters. 6, 458-61.
Cynx, J., Lewis, R., Tavel, B., Tse, H.,1998. Amplitude regulation of
vocalizations in noise by a songbird, Taeniopygia guttata. Animal Behaviour. 56, 107-
113.
Donoghue, A.M, 2004. Occupational health hazards in mining: an overview.
Occupational Medicine. 54, 283-289.

72
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Duarte, M.H.L., Vecci, M.A., Hirsch, A., Young, R.J. , 2011. Noisy human
neighbours affect where urban monkeys live. Biology Letters. 7, 840-842.
Elowson, A.M., Tannenbaum, P.L., Snowdon, C.T., 1991. Food associated calls
correlate with food preferences on cotton-top tamarins. Animal Behaviour. 42, 931-937.
Faria, C.M., Rodrigues, M., do Amaral, F.Q., Módena, É., Fernandes, A.M.,
2006. Aves de um fragmento de Mata Atlântica no alto Rio Doce, Minas Gerais:
colonização e extinção. Revista Brasileira de Zoologia. 23, 1217-1230.
Farina, A., Pieretti, N., Piccioli, L., 2011. The soundscape methodology for
long-term bird monitoring: A Mediterranean Europe case-study. Ecological Informatics.
6, 354-363.
Farina A, Lattanzi E, Piccioli L, Pieretti N., 2012. The soundscapemeter user
manual.
Farina, A., Pieretti, N., Morganti, N., 2013. Acoustic patterns of an invasive
species: The Red-billed Leiothrix (Leiothrix lutea Scopoli 1786) in a Mediterranean
shrubland. Bioacoustics. 22, 175-194.
Forman, R. T. T., Reineking, B., Hersperger, A. M., 2002. Road traffic and
nearby grassland bird patterns in a suburbanizing landscape. Environmental
Management. 29, 782-800.
Francis. C.D., Ortega, C.P., Cruz, A., 2011. Noise pollution filters bird
communities based on vocal frequency. Plos One. 6.
Frank, T, Bise, C.J., Michael, K., 2003. A hearing conservation program for coal
miners. Occup Health Safety.72, 106-110.
Fuller, R.A, Warren, P.H, Gaston, K.J., 2007. Daytime noise predicts nocturnal
singing in urban robins. Biology Letters. 3, 368-70.

73
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Greig-Smith, P., 1980. Parental investment in nest defense by Stonechats
(Saxicola torquata). Animal Behavior. 28, 604-619.
Habib, L., 2007. Chronic industrial noise affects pairing success and age
structure of ovenbirds Seiurus aurocapilla. Journal of Applied Ecology. 44,176-184.
Haddad, C., Sazima, I., Morellato, L., 1992. Anfíbios anuros da Serra do Japi.
História natural da Serra do Japi: Ecologia e preservação de uma área florestal no
sudeste do Brasil. First ed. Campinas, São paulo.
Halfwerk, W., Holleman, L.J.M., Lessells, C.K.M, Slabbekoorn, H., 2011.
Negative impact of traffic noise on avian reproductive success: Traffic noise and avian
reproductive success. Journal of Applied Ecology. 48, 210-219.
Hessel, P.A, Sluis-Cremer, G.K., 1987. Hearing loss in white South African
goldminers. South African Medical Journal. 71, 364-367.
Kight, C.R, Swaddle, J.P., 2011. How and why environmental noise impacts
animals: an integrative, mechanistic review: Environmental noise and animals. Ecology
Letters.14, 1052-1061.
Krause, B., 1987. Bioacoustics, habitat ambience in ecological balance. Whole
Earth Review. 57,1-6
Langemann, U., Gauger, B. Klump, G.M.,1998. Auditory sensitivity in the great
tit: perception of signals in the presence and absence of noise. Animal Behaviour. 56,
763-769.
Lohr, B., Wright, T.F., Dooling, R.J., 2003. Detection and discrimination of
natural calls in masking noise by birds: estimating the active space of a signal. Animal
Behaviour. 65, 763-77.
Mellinger, D., Barlow, J., 2003. Future directions for acoustic marine mammal

74
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
surveys: stock assessment and habitat use. Noaa oar Special Report, Noaa/pmel
Contribution, 2557, 37.
Mennitt, D.J., Fristrup, K.M., 2012. Obtaining calibrated sound pressure levels
from consumer digital audio recorders. Applied Acoustics. 73, 1138-1145.
Miksis-Olds, J.L., Donaghay, P.L., Miller, J.H., Tyack, P.L., Nystuen, J.A.,
2007. Noise level correlates with manatee use of foraging habitats. The Journal of the
Acoustical Society of America. 121, 3011.
Minuzzi, R.B., Sediyama, G.C., Barbosa, E., Melo Júnior, J., 2007. Climatologia
do comportamento do período chuvoso da região sudeste do Brasil. Revista Brasileira
de Meteorologia. 22, 338-344.
Myers, N. Mittermeier, R. A., Mittermeier,C. G.; Fonseca, G. A. B., Kent, J.,
2000. Biodiversity hotspots for conservation priorities. Nature. 403, 853-858.
Nemeth, E., Brumm, H., 2009. Blackbirds sing higher-pitched songs in cities:
adaptation to habitat acoustics or side-effect of urbanization? Animal Behaviour. 78,
637-41.
Nunes, Y.R.F., Pedralli, G.,1995. Desenvolvimento de metodologia para
adensamento e recomposição da vegetação na EPDA-Peti, MG. BIOS-Cadernos do
Departamento de Ciências Biológicas da PUC-MG. 2, 53–61.
Paglia, A.P., Lopes, M.O.G., Perini, F.A., Cunha, H.M., 2005. Mammals of the
Estação de Preservação e Desenvolvimento Ambiental de Peti (EPDA-Peti), São
Gonçalo do Rio Abaixo, Minas Gerais, Brazil. Lundiana, 6, 89-96.
Parks, E.S., Clark, C.W., Tyack, P.L., 2007. Short-and long-term changes in
right whale calling behavior: The potential effects of noise on acoustic communication.
Journal of the Acoustical Society of America.122, 3725-3731.

75
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Patricelli, G. L., Uy, J. A. C., Walsh, G., Borgia, G., 2002. Male displays
adjusted to female's response. Nature. 415, 279-280.
Pieretti, N., Farina, A., Morri, D., 2011. A new methodology to infer the singing
activity of an avian community: The Acoustic Complexity Index (ACI). Ecological
Indicators. 11, 868-873.
Pieretti N, Farina A., 2013. Application of a recently introduced index for
acoustic complexity to an avian soundscape with traffic noise. The Journal of the
Acoustical Society of America.134, 891.
Proppe, D.S, Sturdy, C.B, St. Clair, C.C., 2013. Anthropogenic noise decreases
urban songbird diversity and may contribute to homogenization. Global Change
Biology. 19, 1075-1084.
Queiroz, A.C.M., 2013. Formigas como indicadoras de impacto e reabilitação
em áreas de mineração. Master dissertation. Universidade Federal de Lavras.
Rabin,L.A., McCowan,B., Hooper,S.L., Owings, D.H., 2003. Anthropogenic
noise and its effect on animal communication: An Interface Between Comparative
Psychology and Conservation Biology. International Journal of Comparative
Psychology. 16, 172-192.
Rheindt, F.E., 2003. The impact of roads on birds: Does song frequency play a
role in determiing susceptibility to noise pollution? Journal of Ornithology. 144, 295-
306.
Roberto, J.B., 2010. Influência dos diversos tipos litológicos nas operações de
concentração da instalação de beneficiamento de Brucutu. Master dissertation.
Universidade Federal de Minas Gerais.
Rodrigues, M., Carrara, L.A., Faria, L.P., Gomes, H.B. , 2005. The birds of

76
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Parque Nacional da Serra do Cipó: the Rio Cipó valley, Minas Gerais, Brazil.
Revista Brasileira de Zoologia. 22, 326-338.
Rossing, T.D., 2007. Springer Handbook of Acoustics. Springer, New York,
NY.
Saha, D. C, Padhy, P.K., 2011. Effect of air and noise pollution on species
diversity and population density of forest birds at Lalpahari, West Bengal, India.
Science of the Total Environment. 409, 5328-5336.
Santos, S. G., 2013. The sound of the suburbs: Investigações sobre a biofonia e a
antropofonia em uma mata urbana. Master Dissertation. Pontificia Universidade
Católica de Minas Gerais.
Schaub, A., Ostwald,J. Siemers, B.M., 2008. Foraging bats avoid noise. Journal
of Experimental Biology. 211, 3174-3180.
Schroeder, J, Nakagawa, S, Cleasby, I.R, Burke, T., 2012. Passerine birds
breeding under chronic noise experience reduced fitness. Plos One. 7.
Scott Brandes T., 2008. Automated sound recording and analysis techniques for
bird surveys and conservation. Bird Conservation International.18.
Sjölander, K; Beskow, J., 2000. Wavesurfer-an open source speech tool.
Interspeech. 464-467.
Slabbekoorn, H. Peet, M., 2003. Birds sing at a higher pitch in urban noise.
Nature. 424, 267.
Slabbekoorn, H. Ripmeester, E.A., 2008. Birdsong and anthropogenic noise:
implications and applications for conservation. Molecular Ecology. 17, 72-83.

77
The impact of anthropogenic noise from open cast mining
on Atlantic forest biophony Capítulo 2
Smith, A.C., Virgl, J.A., Panayi, D., Armstrong, A.R., 2005. Effects of a
diamond mine on Tundra breeding birds. Arctic. 58, 295-304.
Sousa-Lima, R., Clark, C., 2008. Modeling the effect of boat traffic on the
fluctuation of humpback whale singing activity in the Abrolhos National Park, Brazil.
Canadian Acoustics. 36, 174-181.
Sousa-Lima, R.S, C. W. Clark., 2009. Whale sound recording technology as a
tool for assessing the effects of boat noise in a Brazilian marine park. Park Science. 26,
59.
Sueur, J., Pavoine, S., Hamerlynck, O., Duvail, S., 2008. Rapid acoustic survey
for biodiversity appraisal. Plos One. 3.
Sueur, J., Farina, A., Gasc, A., Pieretti, N., Pavoine, S., 2014. Acoustic Indices
for Biodiversity Assessment and Landscape Investigation. Acta Acustica united with
Acustica. 100, 772-781.
Sun, J. W. C., Narins, P. M., 2005. Anthropogenic sounds differentially affect
amphibian call rate. Biological Conservation. 121, 419-427.
Summers, P.D., Cunnington, G.M, Fahrig, L., 2011. Are the negative effects of
roads on breeding birds caused by traffic noise? Journal of Applied Ecology. 48, 1527-
1534.
Swaddle, J.P, Page, L.C., 2007. High levels of environmental noise erode pair
preferences in zebra finches: implications for noise pollution. Animal Behaviour.74,
363-368.
Towsey, M., Zhang, L., Cottman-Fields,M., Wimmer,J., Zhang, J., Roe, P.
Visualization of long-duration acoustic recordings of the environment. Procedia
Computer Science. 29, 703-712.

78
The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Warren, P.S. Katti, M., Ermann, M., Brazel., 2006. Urban bioacoustics: it’s not
just noise. Animal Behaviour.71, 491-502.
Zuberbuehler, K., Noe, R., Seyfarth, R.M., 1997. Diana monkey long-distance
calls: Messages for conspecifics and predators. Animal Behaviour. 53, 589-604.

The impact of anthropogenic noise from open cast mining on Atlantic forest biophony Capítulo 2
Supplementary material:
Table S1. Potential species richness in sites close and far from an opencast mine
at Peti Environment Station, Brazil by season and time of day.
Taxonomic group CLOSE FAR
Insects WET DRY WET DRY
Day Night Day Night Day Night Day Night
Insect 1 x x x
Insect 2 x x
Insect 3 x x x x x x
Insect 4 x x x
Insect 5 x x x x x
Insect 6 x x x
Insect 7 x x x x x x x x
Insect 8 x x x x x x x x
Insect 9 x x x x x x
Insect 10 x x
Insect 11 x x x

80
Insect 12 x x
Insect 13 x x
Insect 14 x
Insect 15 x x
Insect 16 x x
Insect 17 x
Insect 18 x x x x
Insect 19 x x x x
Insect 20 x
Insect 21 x x x x
Insect 22 x x
Insect 23 x x x
Insect 24 x x x
Insect 25 x x x x x x
Insect 26 x x x
Insect 27 x x x x x
Insect 28 x x x
Insect 29 x x x

81
Insect 30 x x
Insect 31 x
Insect 32 x
Insect 33 x x
Insect 34 x x x
Insect 35 x x x
Insect 36 x x x
Insect 37 x x x
Insect 38 x x
Insect 39 x
Insect 40 x
Insect 41 x
Insect 42 x
Insect 43 x x
Insect 44 x x x x
Insect 45 x
Insect 46 x x x x x x
Insect 47 x

82
Insect 48 x
Insect 49 x x
Insect 50 x x
Insect 51 x x
Insect 52 x
Insect 53 x x x
Insect 54 x
Insect 55 x
Insect 56 x x x x x x x
Insect 57 x x x x
Insect 58 x x
Insect 59 x x x
Insect 60 x x
Insect 61 x x x x
Insect 62 x
Insect 63 x x x
Insect 64 x x
Insect 65 x x

83
Insect 66 x x x
Insect 67 x
Insect 68 x
Insect 69 x
Insect 70 x x
Insect 71 x
Insect 72 x
Insect 73 x
Insect 74 x
Insect 75 x x
Insect 76 x
Insect 77 x x x
Insect 78 x x
Insect 79 x
Insect 80 x
Insect 81 x
Insect 82 x
Insect 83 x

84
Insect 84 x
Amphibians x x
Hypsiboas faber x
Hypsiboas lundii x x x
Birds
Arremon sp. x
Amazilia lactea x
Automolus
leucophthalmus
x
Basileuterus
culicivorus
x
Camptostoma
obsoletum
x
Capsiempis
flaveola
x
Casiornis rufus x
Chiroxiphia
caudata
x x x
Conopophaga
lineata
x x
Crypturellus
obsoletus
x x
Drymophila
ochropyga
x
Dysithamnus
mentalis
x

85
Dysithamnus sp. x
Elaenia sp. x x
Euphonia
chlorotica
x
Formicivora
serrana
x
Hemithraupis
ruficapilla
x x x
Herpsilochmus
atricapillus
x x x x
Hylophilus
amaurocephalus
x x x x
Lanio melanops x x x
Lanio pileatus x
Legatus
leucophaius
x
Leptotila sp. x
Leptotila verreauxi x x
Manacus manacus x x x
Micrastur
semitorquatus
x
Milvago
chimachima
x
Myiarchus ferox x
Myiarchus sp. x
Myiodynastes x

86
maculatus
Myiopagis
viridicata
x x
Myiornis
auricularis
x
Myiotlhypis
flaveola
x x x x
Nyctiphrynus
ocellatus
x
Patagioenas
plumbea
x x
Phaeomyias
murina
x x
Phyllomyias
fasciatus
x x x
Picumnus cirratus x x
Platyrinchus
mystaceus
x x
Psittacara
leucophtalma
x
Pyriglena
leucoptera
x x
Ramphastos toco x
Saltator similis x x x
Schiffornis
virescens
x
Sporophila
nigricollis
x x
Synallaxis
cinerascens
x

87
Syndactyla
rufosuperciliata
x
Tachyphonus
coronatus
x
Tangara cayana x x x
Tangara
cyanoventris
x
Tangara sp. x x
Thamnophilidae x
Thamnophilus
caerulescens
x x
Thlypopsis sordida x
Thraupidae 1 x x x
Thraupidae 2 x x x x
Thraupidae 3 x x x
Thraupidae 4 x x x x
Thraupidae 5 x x x
Thraupidae 6 x
Thraupidae 7 x
Thraupidae 8 x
Thraupidae 9 x x
Thraupidae 10 x

88
Thraupis sp. x
Tolmomyias
sulphurescens
x x x
Trochilidae x x
Trogon surrucura x x x x
Turdus albicollis x x x
Turdus leucomelas x
Turdus rufiventris x
Turdus sp. x
Tyrannidae x x x
Vireo chivi x x
Zonotrichia
capensis
x
NI1 x x
NI2 x x
NI3 x
NI4 x
NI5 x
NI6 x
NI7

89
NI8 x
NI9 x
NI10 x
NI11 x
NI12 x
NI13 x
NI14 x
NI15 x
NI16 x
Bats
Bat 1 x
Bat 2 x x x
Bat 3 x x
Bat 4 x x x
Bat 5 x x
Bat 6 x x
Bat 7 x x
Bat 8 x x

90
Bat 9 x
Primates
Callithrix
penicillata
x x x x
Callicebus
nigriffrons
x x x x

91
Mining noise reduces loud call by wild black-fronted titi monkeys Capítulo 3
- CAPÍTULO 3-
Mining noise reduces loud calls by wild black-fronted titi monkeys
Artigo submetido ao periódico International Journal of Primatology
Abstract
Human activity has resulted in increased anthropogenic noise on soundscapes. Noise
pollution can constrain acoustic communication and prevent animals to effectively
communicate. Our aim was to investigate how the black-fronted titi monkey (Callicebus
nigrifrons) is affected by noise produced by mining activity in a fragment of Atlantic
forest in Brazil. We installed two passive acoustic monitoring devices to record
24h/day, 7 days every two months, for a year, one unit close to an opencast mine and
the other 2.5km away. Both sites presented similar habitat structures and were inhabited
by multiple groups of C. nigrifrons. Sound pressure levels measurements were
undertaken six times for 20 minutes on different days at both sites. The number of
Callicebus loud calls was quantified at both sites by analyzing the recorded files. The
site close to the mine presented higher noise levels than the one further away. More
black-fronted titi loud calls were detected at the far site and many vocalisations
(20.32%) from the site close to the mine were masked by noise. Duration of loud calls
was longer at the site far from the mine and the diel pattern of vocalisations was
different between the two sites. Our results indicate that mining noise can constrain
Callicebus long distance vocal activity, probably because their loud calls occupy a
similar frequency band of the noise. Given that vocalisations are important regulators of
social behavior in primates, consideration should be given to the impact of mining noise
on their behavior in impact evaluations and mitigation recommendations.
Keywords: Animal communication, anthropogenic activity, primates, social behavior,
sound masking

92
Mining noise reduces loud call by wild black-fronted titi monkeys Capítulo 3
Introduction
Beyond the effect of deforestation caused by mining, one less obvious impact on
wildlife is the noise produced by such activity. Mining noise, especially if produced at
the same frequencies that animals use in their vocalisations can mask important calls
and, consequently, greatly reduce the efficiency of animal communication (Foote et al.
2004; Bee and Swanson 2007).
Acoustic communication is essential in the lives of many species as they use
such signals to transmit biologically relevant information; for example, to find
reproductive partners (Brumm et al. 2009), to escape from predators (Greig-Smith 1980;
Chan et al. 2010) and defend resources (Zuberbuehler et al. 1997). However,
anthropogenic noise has become a common impact on animal communication systems
(Slabbekoorn and Ripmeester 2008; Barber et al. 2009; Laiolo 2010). Noise can
interfere with the propagation and detection of signals by masking animal sounds and
thus, preventing effective species communication (Foote et al. 2004; Bee and Swanson
2007).
Noise pollution can also affect the behaviour of many species. Studies have
shown that animals avoid foraging in noisy areas (Schaub et al. 2008), increase their
vigilance behaviour in presence of noise (Delaney et al. 1999; Karp and Root 2009),
select quiet areas to perform their daily activities (Sousa-Lima and Clark 2009, Duarte
et al. 2011) and can be distracted by noise, thereby increasing the risk of predation
(Chan et al. 2010). Noise can also cause physiological stress (Campo et al. 2005, Kight
and Swaddle 2011) and impact on ecological aspects of animals lives such as population
distribution (Reijen et al. 1998; Bejder et al. 2006), species abundance (Bayne et al.
2008) and diversity (Proppe et al. 2013).
Animal livelihoods can be severely impaired by anthropogenic noise,
nonetheless many studies have documented a range of adaptive responses to minimize
the immediate impact of noise of communication systems including: changes in
frequency (Slabbekoorn and Peet 2003; Parks et al. 2007; Nemeth and Brumm 2009),

93
Mining noise reduces loud call by wild black-fronted titi monkeys Capítulo 3
amplitude (Brumm 2004; Brumm et al. 2009; Hage et al. 2013), calling rate (Sun and
Narins 2005) number of notes (Slabbekoorn and Boer-Visser 2006), timing (Fuller et al.
2007) and duration of the calls (Brumm et al. 2004). The direct impact of noise on
animal behaviour and ecology and incidental costs of maintaining an efficient
communication system through compensatory mechanisms can impose fitness costs on
affected individuals (senders and receivers) and consequently on their survival and
reproduction (Chan et al. 2010; Schroeder et al. 2012).
The effects of mining noise on animals have been poorly documented, especially
in the Neotropical region. Smith et al. (2005) showed that diamond mines affect tundra
birds by lowering breeding bird densities. In India, stone mining and crushing affected
bird species diversity and population density in the areas adjacent to crushers (Saha and
Padhy 2011). Thus, studies involving mining noise impact in terrestrial mammals and
their communication systems are still lacking.
Species of titi monkeys (genus Callicebus) exchange loud calls (duets) to either
defend territories or food resources in their home-ranges; thus, these vocalisations are
important regulators of their social behaviour (Robinson 1979, 1981; Kinsey and
Robinson 1983; Prince and Piedade 2001; Caselli et al. 2014). Many of the forests in
South America, where titi monkeys live suffer from large scale mining (Estrada 2009).
In the state of Minas Gerais, Brazil, mining is an important economic activity and is
commonly conducted close to Atlantic forest region, one of the world’s richest
biodiversity hotspots (Myers et al. 2000). The Atlantic forest is one of the most
impacted habitats of the world retaining only 7% of its primary vegetation (Myers et al.
2000) and is home to the black-fronted titi monkey (Callicebus nigrifrons), an endemic
primate classified as Near Threatened on the IUCN’s Red List (Veiga et al. 2008).
Primates of the genus Callicebus live in monogamous family groups, consisting
of a reproductive pair and up to four generations of offspring (Kinzey and Becker 1983;
Mendoza and Mason 1986; Valeggia et al. 1999). Titi monkeys are morphologically
cryptic primates, which hinders surveying them using traditional methods such as linear

94
Mining noise reduces loud call by wild black-fronted titi monkeys Capítulo 3
transects (Aldrich et al. 2008). Mated pairs of Callicebus species regularly emit loud
and coordinated calls (duets), which permit researchers to use an alternative and
potentially more accurate method to monitor populations based in call surveys (Melo
and Mendes 2000; Aldrich et al. 2008). Duetting is commonly used by many bird and
primate species for both within and between group communication (Hall 2004; Oliveira
and Ades 2004). Studies of Callicebus species show that their duets have a role in group
location and avoidance of intergroup aggressive encounters (C. lucifer, previously C.
torquatus, Kinzey and Robinson 1983; C. personatus, Kinzey and Becker 1983; Price
and Piedade 2001), in territory establishment and probably mate defense (C. ornatus,
previously C. moloch, Mason 1968; Robinson 1979, 1981). Black-fronted titi monkeys
(C. nigrifrons) loud calls are used during intergroup communication to regulate access
to important food resources, such as fruits. There is also some evidence that loud calls
are used for mate defence (Caselli et al. 2014). Typically, titi monkeys vocalise mostly
at dawn, but also during the day when another group is sighted or heard (Kinzey et al.
1977, Kinzey and Robinson 1983; Melo and Mendes 2000).
Due to spectral characteristics of titi monkey loud calls such as high amplitude
and low frequency, these calls can be heard over long distances (Melo and Mendes
2000; Caselli et al. 2014). Unfortunately, the same acoustic characteristics that were
adaptive for long distance communication are now bringing these sounds into
competition with mining noise.
In this study, we investigated how the noise produced by one of the largest
opencast mines of the world affects acoustic communication of C. nigrifrons in an
Atlantic forest fragment in Southeast Brazil. Here we tested the following hypotheses:
(1) noise levels are different in the sites close and far from the mine; (2) temporal
acoustic parameters, duration and diel pattern of titi monkey loud vocalisations would
change between the areas due to noise exposure.

95
Mining noise reduces loud call by wild black-fronted titi monkeys Capítulo 3
Material and Methods
Study area
This study was conducted at Peti environmental station, which is located in an
Atlantic forest fragment of approximately 605 hectares. The reserve is located in the
upper Rio Doce Basin (altitude range: 630-806m) in the municipalities of São Gonçalo
do Rio Abaixo and Santa Bárbara, Minas Gerais state, Brazil (19°53’57’’S and
43°22’07’’W), one of the most fragmented Atlantic forest regions of Brazil (Machado
and Fonseca 2000). Peti environmental station harbors approximately 46 species of
mammals (Paglia et al. 2005), 231 species of birds (Faria et al. 2006) and 29 species of
anurans (Bertoluci et al. 2009).
A large part of the reserve is covered by secondary arboreal vegetation, and is
surrounded by a matrix mainly composed by Eucalyptus, small farms and areas of
exposed soil due to the activities of the Brucutu mine. Mining activity occupies an area
of approximately 8km2 and produces noise through road traffic, sirens and explosions
along the day and night (Roberto 2010). Brucutu’s iron ore extraction started in 1992.
To increase the capacity of iron production, expansion projects started in 2004 placing
Brucutu among the largest opencast mines in the world (Roberto 2010).
Data collection
To record black fronted titi monkey loud calls, one song meter (SM2, Wildlife
Acoustics) was installed in the forest fragment close to the mine site at a distance of
100m from the closest mining road (Fig.1). Another song meter was installed far from
the mine at a distance of 2,500m and 100m away from a low traffic (‘quiet’) road (to
control for a potential border effect at both sites in the same Atlantic forest fragment).
Both sites were habitat matched; they presented similar floristic compositions and

96
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
habitat structures and were inhabited by multiple groups of C. nigrifrons. Groups of titi
monkeys were sighted many times at both sites during fieldwork. At both sites the
passive acoustic monitoring devices were programmed to record 24h/day during seven
days every two months from October 2012 to August 2013, in a total of six sessions and
2,016 hours of recordings. Each SM2 was fixed on a tree 1.5m above the ground,
leaving the two lateral microphones free from any surface that could be an obstacle to
incoming sound waves. They were configured to record in wave format at a sampling
rate of 44,100Hz, at 16 bits, and with a 36% microphone gain. This configuration had
been found in pilot studies to be optimal for recording the soundscape of the Atlantic
forest (Pieretti et al. in press). The sound pressure levels at both sites were characterized
by using B&K2270 (Denmark) sound level meter configured on the A curve to conduct
6 measurements of 20 minutes at each site on different days and time.
This research adhered to the Brazilian legal requirements and to the American
Society of Primatologists (ASP) Principles for the Ethical Treatment of Non Human
Primates.
Fig.1 Sites close to and far from the Brucutu mine at Peti Environmental station,
southeast Brazil. Red lines represent the limit of each site by considering the geographic
barriers.

97
Mining noise reduces loud call by wild black-fronted titi monkeys Capítulo 3
Data analyses
To test for a difference in noise levels between the sites close and far from the
mine we extracted data from the sound level pressure measurements and analyzed them
using BZ 5503 software (Bruel and Kjaer). To avoid bias in the measured levels we
excluded all recordings, which included loud animal sounds (i.e., animals close to the
microphone).
The rate of occurrence and the duration of black-fronted titi monkey loud calls
were measured in both sites during seven days by session from 0500 to 1700 hours
totalizing 1,092 analyzed hours. This procedure was done using the band limited energy
detector in Raven Pro 1.5 but resulted in a large number of false positives and misses.
Therefore all sound files used for analyses had to be visually and aurally checked in
Raven. We also manually detected all the loud calls, which were partially masked by
anthropogenic noise at the site close to the mine.
To verify a possible association between the noise produced by mining truck traffic at
the site close to the mine and the occurrence of the loud calls, we quantified all trucks
passing from 0500 to 1700 hours at the road in front of the sampling site. This
procedure was done by audio and visual identification of the trucks’ noise pattern in
spectrograms. An FFT size of 1024 points was used for all analyses in Raven. We used
a nonparametric statistical approach with our data analyses since data did not meet the
requirements for parametric statistics even after data transformations. All the statistical
analyses were performed in Statistica version.8.0.
Results
Sound pressure (noise) levels were significantly higher at the site close to the
mine (Mann-Whitney U-test: U=1, Z=2.72, N=6, p<0.01), as expected (Table 1).

98
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
Table 1. Equivalent sound pressure levels (Leq) at sites close to and far from an
opencast mine site near Peti environmental station, southeast Brazil.
Close
Leq dB(A)
Far
Leq dB(A)
42.6 33.8
38.7 30.3
42.0 30.1
60.9 37.2
42.9 38.8
41.2 33.3
Black fronted titi monkeys emitted more loud calls than expected at the site far
from the mine (Chi-squared test: X2= 339.96, df=1, P<0.001 Nclose=187, Nfar=752). A
considerable part (20.32%) of the vocalisations found in the site close to the mine was
partially masked by noise from mining activity. Duration of loud calls were also
significantly longer at the site far from the mine (Mann-Whitney U test: U= 29142.5,
Z= 12.40, P<0.01; Medianclose=1.77, Medianfar=16.33).
The temporal distribution pattern of the vocalisations was also different between
the two sites (Fig. 2). At the site far from the mine, titi monkeys were more vocally
active early in the morning (from 0600 to 1000 hours, with peak vocal activity around
0700 hours), while at the site close to the mine they presented a constant but very low
activity from 0700 to 1000 hours with peak vocal activity occurring around 1300 hours.
The time period of highest truck passing activity coincided with the time period
of the lower number of loud vocalisations at the site close to the mine and the peak of
loud calls also occurred when there was a decrease in trucks passing (Fig 3). Despite
this, a Spearman rank test showed no significant correlation between the number of
trucks and number of vocalisations (rs = -0.21, t= -0.71, P>0.05).

99
Mining noise reduces loud call by wild black-fronted titi monkeys Capítulo 3
Fig 2. Daily distribution of the mean (±SD) number of loud calls emitted by black
fronted titi monkeys at sites close to and far from an opencast mine site near Peti
environmental station, southeast, Brazil.

100
Mining noise reduces loud call by wild black-fronted titi monkeys Capítulo 3
Fig 3. Daily distribution of mean mining truck activity (number passing a fixed point)
and mean frequency of loud calls of black-fronted titi monkey close to an opencast
mine site near Peti Environmental station, southeast Brazil.

101
Mining noise reduces loud call by wild black-fronted titi monkeys Capítulo 3
Discussion
The higher rate of loud calls found at the far site could be explained by several
non-exclusive hypotheses, such as: (1) more titi monkey groups are present at the far
site; (2) more encounters between titi groups at the far site; (3) titi monkeys from the
close site reducing their emission of calls due to masking caused by mining noise; (4)
call emissions masked by noise decreasing detection of vocal activity at the site close to
the mine. All of these hypotheses are possible; however, habitat matching means that
there should be very similar numbers of groups at both sites. The area monitored by the
passive acoustic monitoring devices was the same at both sites. Home range sizes of C.
nigrifrons ranged from 8.35 to 9.23 ha in an Atlantic forest field site approximately
25km far from our study site (Santos 2012). The positions of geographic barriers such
as roads and a river that surround close and far sites suggest that groups, which inhabit
the close site are isolated from the groups that inhabit the far site. Thus, while there will
be some differences between sites, these are unlikely to be the major factors affecting
differences in the rate of loud call emissions. Another noteworthy factor is the longer
duration of calls at the far site. This fact supports the third hypothesis and not the first
two: as it indisputably demonstrates the impact of mining noise on titi monkey’s loud
vocalisations.
A decrease of animal call rate in presence of noise has already been established
in other studies and can be interpreted as a response to avoid interference from
anthropogenic noise (Miksis-Olds and Tyack 2009; Sun and Narins 2005; Parks et al.
2007; Sousa-Lima and Clark 2008). This pattern may indicate that animals wait until it
is quiet to vocalize, exhibiting only minimal vocalisation effort during periods of
masking noise (Miksis-Olds and Tyack 2009; Sousa-Lima and Clark 2008). In this
study, at the close site many loud calls (20%) were partially masked by noise, thereby
potentially disturbing the exchange of acoustic information and preventing titi monkeys
from communicating effectively (Lohr et al. 2003; Foote et al. 2004; Bee and Swanson
2007). One particularly important factor driving vocalisation effort is the range over

102
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
which the signaller and receiver must effectively communicate (Miksis-Olds and Tyack
2009). In this context, when noise masks the vocalisations there is a decrease in the
acoustic space over which the information can reach.
The longer duration of titi monkey loud calls at the far site is further evidence of
the noise impact from mining. Researches have already documented that some species
adjust their vocal behaviour to compensate for anthropogenic noise by increasing or
decreasing the duration of the calls. Studies with Saguinus oedipus showed a decreasing
in the average call duration to avoid overlapping the noise (Egnor et al. 2007).
However, common marmosets Callithrix jacchus increase the duration of their calls in
presence of noise, but they use higher vocal frequencies (Brumm et al. 2004). Our
results, suggest that there is more available acoustic space at the far site, especially in
the lower frequencies, which are naturally occupied by titi monkeys. At the close site,
noise from the mine could be excluding titi monkeys from an acoustic niche. Thus, they
probably are emitting calls with shorter duration to save energy since acoustic
communication is an energetically expensive behaviour and vocalisation effort is
increased by increasing call duration (Miksis-Olds and Tyack 2009).
The difference in the diel pattern of loud calls between the two sites can be also
a consequence of the mining noise disturbance on titi monkeys’ vocal behaviour. C.
nigrifrons are vocally active mainly during the first hours of the day (Melo and Mendes
2000) and this natural pattern was observed only at the far site. At the site close to the
mine, animals presented very low vocal activity in the first hours of the day and peak of
activity at 1300 hours, which coincides with the lower traffic of trucks and mining
activities due to lunch time of mine employees. Although there was no significant
negative correlation between the number of trucks passing and the number of loud calls
at the site close. Titi monkeys vocalised less in the first hours of the day when there
were a higher number of trucks and presented a peak of vocal activity around 1300
when the number of trucks passing was lower. Many mammals affected by
anthropogenic noise have limited developmental capacity to change the acoustic

103
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
parameters of their calls to avoid the masking by noise such as some birds can do
(Weiss et al. 2014). On the other hand, mammals may avoid noise with other
behavioural modifications, such as vocalizing during periods of low noise (Rabin et al.
2003) or moving to quieter areas (Duarte et al. 2011).
Loud vocalisations are key factors involved in the regulation of titi monkey
social behaviour. One consequence of the masking of such calls can be increased
territory invasion by neighbouring group and consequently increased rates of inter-
group agonistic encounters. Such changes could impact on the survival and reproductive
success of the affected individuals. Future studies should investigate the other
behavioural effects of sound pollution on primates, for example, do they try and
compensate by using more ‘exaggerated’ visual signals, and/or do they may alter their
acoustics parameters, such as amplitude or the frequency of their calls when it is noisy?
Overall our results suggest that although several factors can influence the rate
and duration of loud calls – the principal cause in difference between sites close and far
from an opencast mine was the noise being emitted by mining activities.
Conclusion
In the last decade, many studies have documented the impact of anthropogenic
noise on animal communication systems and wellbeing. Most of the studies have
focussed on bird communication and there are no studies about noise impact from
mining activity on primates communication. Here, we have shown for the first time how
a noise disturbance affects black fronted-titi monkey communication. Our results
provide important information to be considered during the elaboration of conservation
strategies in natural areas affected by mining activity. Furthermore, we suggest that
noise monitoring plans for wildlife should be part of the process of licensing large scale
anthropogenic activities such as mining.

104
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
Acknowledgements
We thank all of the staff at the environmental station of Peti, especially
Leotacílio da Fonseca. We are also grateful to Marina Scarpelli and Renan Duarte for
their help during data acquisition and the engineer Krisdany Cavalcante for help with
the noise level measurements. MHLD and MCK were supported by a Fundação de
Amparo à Pesquisa de Minas Gerais (FAPEMIG) postgraduate and technical
scholarship respectively. MR, RJY and RSL received financial support from FAPEMIG
and Conselho Nacional de Pesquisa (CNPq).

105
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3 References
Aldrich, B. C., Molleson, L., & Nekaris, K. A. I. (2008). Vocalizations as a
conservation tool : an auditory survey of the Andean titi monkey Callicebus oenanthe
Thomas, 1924 (Mammalia: Primates: Pitheciidae) at Tarangue, Northern Peru.
Contributions to Zoology, 77, 1–6.
Barber, J. R., Crooks, K .R., & Fristrup, K. M. (2009). The costs of chronic
noise exposure for terrestrial organisms. Trends in Ecology and Evolution, 25, 180-189.
Bayne, E. M., Habib, L., & Boutin, S. (2008). Impacts of chronic anthropogenic
noise from energy-sector activity on abundance of songbirds in the boreal forest.
Conservation Biology, 22, 1186-1193.
Bee, M. A., & Swanson, E. M. (2007). Auditory masking of anuran
advertisement calls by road traffic noise. Animal Behaviour, 74, 1765-1776.
Bejder, L., Samuels, A., Whitehead, H., & Gales, N. (2006). Interpreting short-
term behavioural responses to disturbance within a longitudinal perspective. Animal
Behaviour, 72, 1149-1158.
Bertoluci, J, Canelas, M. A. S., Eisemberg, C. C., Palmuti, C. F. de S., &
Montingelli, G. G.(2009). Herpetofauna of Estação Ambiental de Peti, an Atlantic
Rainforest fragment of Minas Gerais State, southeastern Brazil. Biota Neotropica, 9,
147-155.
Brumm, H. (2004). The impact of environmental noise on song amplitude in a
territorial bird. Journal of Animal Ecology, 73, 434-440.

106
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
Brumm, H., Voss,K., Köllmer,I., & Todt, D. (2004). Acoustic communication in
noise: regulation of call characteristics in a New World monkey. The Journal of
Experimental Biology, 207, 443-448.
Brumm, H., Schmidt, R, & Schrader L., (2009). Noise-dependent vocal
plasticity in domestic fowl. Animal Behaviour, 78, 741-746.
Campo, J.L., Gil, M. G. & Dávila, S. G. (2005). Effects of specific noise and
music stimuli on stress and fear levels of laying hens of several breeds. Applied. Animal
Behavaviour. Science, 91: 75-84.
Caselli, C. B., Mennill, D. J., Bicca-Marques, J. C., & Setz, E. Z. F. (2014).
Vocal behavior of black-fronted titi monkeys (Callicebus nigrifrons): acoustic
properties and behavioral contexts of loud calls. American Journal of Primatology, 76,
788–800.
Chan, A. A. Y-H, Giraldo-Perez, P., Smith, S., & Blumstein, D. T., (2010).
Anthropogenic noise affects risk assessment and attention: the distracted prey
hypothesis. Biology Letters, 6, 458-61.
Delaney, D. K., Gurbb, T. G., Seibr, P., Pater, L. L. & Reiser, M.H. (1999).
Effects of helicopter noise on Mexican spotted owls. Journal of Wildlife Management,
63, 60–76.
Duarte, M. H. L., Vecci, M. A., Hirsch, A., & Young, R. J. (2011). Noisy human
neighbours affect where urban monkeys live. Biology Letters, 7, 840-842.
Egnor, S. E. R., Wickelgren, J. G. & Hauser, M. D. (2007). Tracking silence:
adjusting vocal production to avoid acoustic interference. Journal of Comparative
Physiology, 193, 477-483.

107
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
Estrada, A. (2009). Primate Conservation in South America: The Human
and Ecological Dimensions of the Problem. In P. A. Garber, A. Estrada, J. C.
Bicca-Marques, E. W. Heymann, K. B. Strier (Ed.), South American
primates.(pp. 463-505). New York: Springer.
Faria, C. M., Rodrigues, M., do Amaral, F. Q., Módena, É., & Fernandes,
A. M., (2006). Aves de um fragmento de Mata Atlântica no alto Rio Doce,
Minas Gerais: colonização e extinção. Revista Brasileira de Zoologia, 23, 1217-
1230.
Foote, A. D., Osborne, R. W., & Hoelzel, A. R. (2004). Whale-call
response to masking boat noise. Nature, 428, 910.
Fuller, R. A, Warren, P. H, & Gaston, K. J. (2007). Daytime noise
predicts nocturnal singing in urban robins. Biology Letters, 3, 368-70.
Greig-Smith, P. (1980). Parental investment in nest defense by
Stonechats (Suxicolu torquata). Animal Behavior, 28, 604-619.
Hage, S. R., Jiang, T., Berquist, S. W., Feng, J. & Metzner, W. (2013).
Ambient noise induces independent shifts in call frequency and amplitude within
the Lombard effect in echolocating bats. Proceedings of the National Academic
os Sciences of the United States of America. 110, 4063-4068.
Hall, M. L. (2004). A review of hypotheses for the functions of avian
duetting. Behavioural Ecology and Sociobiology, 55, 415- 430.

108
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
Karp, D. S, & Root, T. L. (2009). Sound the stressor: how Hoatzins
(Opisthocomus hoazin) react to ecotourist conversation. Biodiversity Conservation, 18,
3733–3742.
Kight, C. R, & Swaddle, J. P. (2011). How and why environmental noise
impacts animals: an integrative, mechanistic review: Environmental noise and animals.
Ecology Letters.14, 1052-1061.
Kinzey, W. G, & Becker, M. (1983). Activity pattern of the masked titi monkey,
Callicebus personatus. Primates, 24, 337–343.
Kinzey, W. G, & Robinson, J. G. (1983). Intergroup loud calls, range size, and
spacing in Callicebus torquatus. American Journal of Physical Anthropology, 60, 539–
544.
Kinzey, W. G, Rosenberger, A. L, Heisler, P. S, Prowse, D. L, & Trilling, J. S.
(1977). A preliminary field investigation of the yellow handed titi monkey, Callicebus
torquatus torquatus, in northern Peru. Primates, 18, 159–181.
Laiolo, P. (2010). The merging significance of bioacoustics in animal species
conservation. Biological Conservation, 143, 1635-1645.
Lohr, B., Wright, T. F. & Dooling, R. J. (2003). Detection and discrimination of
natural calls in masking noise by birds: estimating the active space signal. Animal
Behaviour, 65, 763-777.
Machado, R. B. & G. A. B, Fonseca. (2000). The avifauna of rio Donde Valley,
southeastern Brazil, a highly fragmented area. Biotropica, 32, 914-924.

109
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
Mason, W. A. (1968). Use of space by Callicebus groups. In P.C. Jay (Ed),
Primates: studies in adaptation and variability. (pp. 200-216). New York: Holt,
Rinehart and Winston.
Melo, F. R. & Mendes, S. L. (2000). Emissão de gritos longos por grupos de
Callicebus nigrifrons e suas reações a playbacks. In C. Alonso & A. Langgut (Eds), A
Primatologia no Brasil - 7 (pp 215-222). João Pessoa, Paraíba: Sociedade Brasileira de
Primatologia and Editoria Universitária.
Mendoza, S. P, & Mason, W. A. (1986). Parental division-of-labor and differ-
entiation of attachments in a monogamous primate (Callicebus moloch). Animal
Behaviour, 34, 1336–1347.
Miksis-Olds, J. L. & Tyack, P. L., (2009). Manatee (Thichechus manatus)
vocalization usage in relation to environmental noise levels. The Journal of the
Acoustical Society of America, 125, 1806-1815.
Myers, N. Mittermeier, R. A., Mittermeier,C. G.; Fonseca, G. A. B., & Kent, J.
(2000). Biodiversity hotspots for conservation priorities. Nature, 403, 853-858.
Nemeth, E., & Brumm, H. (2009). Blackbirds sing higher-pitched songs in
cities: adaptation to habitat acoustics or side-effect of urbanization? Animal Behaviour,
78, 637-41.
Oliveira D. A, & Ades C. (2004). Long‐distance calls in Neotropical primates.
Anais da Academia Brasileira de Ciências, 76, 393-398.
Paglia, A. P., Lopes, M. O. G., Perini, F. A., & Cunha, H. M. (2005). Mammals

110
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
of the Estação de Preservação e Desenvolvimento Ambiental de Peti (EPDA-Peti), São
Gonçalo do Rio Abaixo, Minas Gerais, Brazil. Lundiana, 6, 89-96.
Parks, E. S., Clark, C. W., & Tyack, P. L. (2007). Short-and long-term changes
in right whale calling behavior: The potential effects of noise on acoustic
communication. Journal of the Acoustical Society of America, 122, 3725-3731.
Pieretti, N., Duarte, M. H. L., Sousa-Lima, R. S., Young, Robert J., Rodrigues,
M., Farina, A. (In press). Determining sampling schemes for passive acoustic studies in
different tropical environments. Tropical Conservation Science.
Price, E. C., & Piedade, H. M. (2001). Ranging behavior and intraspecific
relationships of masked titi monkeys (Callicebus personatus personatus). American
Journal of Primatology, 53, 87–92.
Proppe, D. S, Sturdy, C. B, St & Clair, C. C. (2013). Anthropogenic noise
decreases urban songbird diversity and may contribute to homogenization. Global
Change Biology, 19, 1075-1084.
Rabin, L. A., McCowan, B., Hooper, S. L. & Owings, D. H. (2003).
Anthropogenic noise and its effect on animal communication: an interface between
comparative psychology and conservation biology. International Journal of Psychology,
16, 172–192.
Reijen, R., Foppen, R., Braak, C. T., Thissen, J. (1998). The effects of car traffic
and breeding bird populations in woodland. III. Reduction of density in relation to the
proximity of main roads. Journal of Applied Ecology, 32, 187–202.

111
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
Roberto, J. B. (2010). Influência dos diversos tipos litológicos nas operações de
concentração da instalação de beneficiamento de Brucutu. Master dissertation.
Universidade Federal de Minas Gerais.
Robinson, J. G. (1979). Vocal regulation of use of space by groups of titi
monkeys Callicebus moloch. Behavioral Ecology and Sociobiology, 5, 1–15.
Robinson, J. G. (1981). Vocal regulation of inter‐ and intra‐group spacing during
boundary encounters in the titi monkey, Callicebus moloch. Primates, 22, 161–172.
Saha, D. C, & Padhy, P. K. (2011). Effect of air and noise pollution on species
diversity and population density of forest birds at Lalpahari, West Bengal, India.
Science of the Total Environment, 409, 5328-5336.
Santos, R. V. (2012). Área de vida e uso do espaço por Callicebus nigrifrons
Spix, 1823 (Primates: Pitheciidae). Master dissertation. Pontifícia Universidade Católica
de Minas Gerais.
Schaub, A., Ostwald, J. & Siemers, B. M. (2008). Foraging bats avoid noise.
Journal of Experimental Biology, 211, 3174-3180.
Schroeder, J., Nakagawa, S., Cleasby, I. R., Burke, T. (2012). Passerine birds
breeding under chronic noise experience reduced fitness. Plos One. 7.
Slabbekoorn, H. & Peet, M. (2003). Birds sing at a higher pitch in urban noise.
Nature, 424, 267.

112
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
Slabbekoorn, H., & den Boer-Visser, A. (2006). Cities change the songs of birds.
Current Biology ,16, 2326–31.
Slabbekoorn, H. & Ripmeester, E. A. (2008). Birdsong and anthropogenic noise:
implications and applications for conservation. Molecular Ecology, 17, 72-83.
Smith, A. C., Virgl, J. A., Panayi, D., & Armstrong, A. R., (2005). Effects of a
diamond mine on Tundra breeding birds. Arctic, 58, 295-304.
Sousa-Lima, R., & Clark, C. (2008). Modeling the effect of boat traffic on the
fluctuation of humpback whale singing activity in the Abrolhos National Park, Brazil.
Canadian Acoustics, 36, 174-181.
Sousa-Lima, R. S, C. W. & Clark. (2009). Whale sound recording technology as
a tool for assessing the effects of boat noise in a Brazilian marine park. Park Science,
26, 59.
Sun, J. W. C., & Narins, P. M. (2005). Anthropogenic sounds differentially
affect amphibian call rate. Biological Conservation, 121, 419-427.
Veiga, L. M., Kierulff, C. M., de Oliveira, M. M. & Mendes, S. L.
(2008). Callicebus nigrifrons. The IUCN Red List of Threatened Species. Version
2014.3. <www.iucnredlist.org>. Downloaded on 17 December 2014.
Valeggia, C. R., Mendoza, S. P., Fernandez-Duque, E., Mason, W. A., & Lasley,
B. (1999). Reproductive biology of female titi monkeys (Callicebus moloch) in
captivity. American Journal of Primatology, 47, 183–195.

113
Mining noise reduces loud call by wild black-fronted
titi monkeys Capítulo 3
Weiss, D. J., Hotchkin, C. F., Parks, S. E. (2014). Modification of spectral
features by nonhuman primates. Behavioral and brain Sciences, 37, 574-576.
Zuberbuehler, K., Noe, R., & Seyfarth, R. M. (1997). Diana monkey long-
distance calls: Messages for conspecifics and predators. Animal Behaviour, 53, 589-
604.

114
Conclusão
A atividade de mineração gera diversos tipos de ruído que atuam como fator
estressor e cria uma nova pressão seletiva sobre as espécies que usam sinais acústicos
para se comunicar. Um sistema efetivo de monitoramento acústico de populações
animais se faz necessário em áreas com a presença desta atividade, devido às mudanças
que o ruído pode causar na dinâmica de comunicação das espécies.
Nesta tese de doutorado, a abordagem da Ecologia de Paisagem Acústica foi
utilizada para monitorar ambientes naturais e ruidosos. Em particular, as pesquisas
mostradas aqui, apresentam uma nova forma de coleta, análise e interpretação de dados
acústicos para monitorar as alterações antrópicas no ambiente, avaliar diferenças entre
comunidades e monitorar a dinâmica acústica no espaço e no tempo.
O estudo de paisagens acústicas ainda é um tópico difícil de investigar,
especialmente em áreas tropicais, devido à quantidade de informações contidas em cada
ambiente e seu estudo requer o desenvolvimento de técnicas de subamostragem, a fim
de otimizar o processo de análise e interpretação de dados. Neste contexto, a
metodologia apresentada no capítulo 1 fornece critérios para subamostragem de dados
em alguns biomas tropicais e sugere rotinas de programação baseadas nas características
de cada ambiente estudado. Em conclusão, ambientes que mostraram alta e contínua
presença de sons podem ser amostrados de forma menos intensa, enquanto aqueles que
apresentam emissões acústicas ocasionais ou imprevisíveis devem ser amostrados de
forma mais intensa para garantir uma representação confiável da paisagem acústica.
O capítulo 2 apresentou resultados significativos sobre o impacto da poluição
sonora proveniente de atividade mineradora na dinâmica de comunicação acústica dos
animais. A maior complexidade acústica durante o dia na área próxima à mina pode
estar relacionada com os níveis de ruído desta área. Além disso, a menor riqueza de
espécies na área próxima à mina, as diferenças na composição de espécies e nas
características espectrais dos cantos registrados nas duas áreas são outros indicadores do
impacto da poluição sonora. Assim, é possível concluir que o ruído pode contribuir para
alterar aspectos ecológicos de comunidades e a dinâmica da comunicação acústica dos
animais.
No capítulo 3 foi encontrada uma menor taxa de ocorrência das vocalizações de
guigós na área próxima à mina, maior duração dos chamados na área distante e uma

115
Conclusão diferença na distribuição diária das vocalizações ao longo do dia entre as duas áreas
estudadas. Estes resultados também são indicadores de que o ruído altera o
comportamento natural da espécie e afeta diretamente na sua comunicação acústica.
Futuras pesquisas deveriam focar na elaboração de metodologias e índices para
análise de paisagens acústicas em ambientes tropicais, uma vez que todas as ferramentas
existentes atualmente foram elaboradas em ambientes temperados, onde o ambiente
acústico é consideravelmente menos complexo. O estudo da paisagem acústica mostrou
ser uma ferramenta eficaz para monitoramento acústico em áreas afetadas por ruído
antrópico, mas pesquisas envolvendo cada espécie individualmente também podem
fornecer dados extremamente relevantes para estratégias de conservação e manejo. Por
exemplo, uma informação importante seria a identificação das espécies mais sensíveis
ao ruído e daquelas que conseguem se adaptar e desenvolver estratégias de comunicação
para sobreviver em áreas ruidosas. Outro dado que pode direcionar ações mais efetivas
de manejo é a identificação dos níveis de ruído acima dos quais cada espécie começa a
ter sua comunicação afetada.
Esta tese de doutorado apresenta uma contribuição inédita sobre o impacto da
poluição sonora proveniente da atividade mineradora na comunicação acústica da fauna
silvestre, um impacto ainda não estudado no Brasil. Os resultados desta tese fornecem
informações importantes para fomentar políticas públicas durante a elaboração de
estratégias de conservação e manejo de áreas naturais. Desta forma, o monitoramento do
ruído em áreas naturais afetadas por mineração deveria ser incluído como uma das
exigências dos órgãos ambientais durante o processo de licenciamento desta atividade.

116
Referências bibliográficas
Bardeli, R., Wolff, D., Kurth, F., Koch, M., Tauchert, K.H., Frommolt, K.H.
2010. Detecting bird sounds in a complex acoustic environment and application to
bioacoustic monitoring. Pattern Recognition Letters. 3, 1524–1534.
Bayne, E.M., Habib, L., Boutin, S. 2008. Impacts of chronic anthropogenic
noise from energy-sector activity on abundance of songbirds in the boreal forest.
Conservation Biology. 22, 1186-1193.
Blumstein, D.T., Mennill, D.J., Clemins, P., Girod, L., Yao, K., Patricelli, G.,
Deppe, J.L., Krakauer, A.H., Clark, C., Cortopassi, K.A., Hanser, S.F., McCowan, B.,
Ali, A.M., Kirschel, A.N.G. 2011. Acoustic monitoring in terrestrial environments
using microphone arrays: applications, technological considerations and prospectus.
Journal of Applied Ecology. 48, 758-767.
Brandes, T. S. 2008. Automated sound recording and analysis techniques for
bird surveys and conservation. Bird Conservation International. 18.
Brown, C.L., Hardy, A.R., Barber, J.R, Fristrup, K.M, Crooks, K.R. 2012. The
effect of human activities and their associated noise on ungulate behavior. Plos One. 7.
Brown, C.L., Reed, S.E., Dietz, M.S, Fristrup, K.M. 2013. Detection and
classification of motor vehicle noise in a forested landscape. Environmental
Management. 52, 1262-1270.
Brumm, H., Voss,K., Köllmer,I., Todt, D. 2004. Acoustic communication in
noise: regulation of call characteristics in a New World monkey. The Journal of
Experimental Biology. 207, 443-448.
Brumm, H., Schmidt, R, Schrader L. 2009. Noise-dependent vocal plasticity in
domestic fowl. Animal Behaviour. 78, 741-746.

117
Referências bibliográficas
Clark, C.W., Ellison, W.T., Southall, B.L., Hatch, L., Parijs, S.M.V., Frankel,
A., Ponirakis, D. 2009. Acoustic masking in marine ecosystems: intuitions, analysis,
and implication. Marine ecology progress series. 395, 201-222.
Cohen, J. How many people can the earth support? W.W. Norton & Co., New
York. 1995.
Donoghue, A.M. 2004. Occupational health hazards in mining: an overview.
Occupational Medicine. 54, 283-289.
Duarte, M.H.L., Vecci, M.A., Hirsch, A., Young, R.J. 2011. Noisy human
neighbours affect where urban monkeys live. Biology Letters. 7, 840-842.
Estrada, A. 2009. Primate Conservation in South America: The Human and
Ecological Dimensions of the Problem. In P. A. Garber, A. Estrada, J. C. Bicca-
Marques, E. W. Heymann, K. B. Strier (Ed.), South American primates (pp. 463-505).
Fuller, R.A, Warren, P.H, Gaston, K.J., 2007. Daytime noise predicts nocturnal
singing in urban robins. Biology Letters. 3, 368-70.
IBRAM. 2011. Informações e análises da economia mineral brasileira. 6ª edição.
Klinck, H., Kindermann, L., Boebel, O. 2008. Detection of leopard seal
(Hydrurga leptonyx) vocalizations using the Envelope–Spectrogram Technique (TEST)
in combination with a Hidden Markov model. Canadian Acoustics – Acoustique
Canadienne. 36, 118–124.
Krause, B., Gage, S.H., Joo, W. 2011. Measuring and interpreting the
soundscape at four places in Sequoia National Park. Landscape Ecology. 26, 1247–
1256.

118
Referências bibliográficas
Mellinger, D., Barlow, J. 2003. Future directions for acoustic marine mammal
surveys: stock assessment and habitat use. Noaa oar Special Report, Noaa/pmel
Contribution, 2557, 37.
Mennitt, D.J., Fristrup, K.M. 2012. Obtaining calibrated sound pressure levels
from consumer digital audio recorders. Applied Acoustics. 73, 1138-1145.
Miksis-Olds, J.L., Donaghay, P.L., Miller, J.H., Tyack, P.L., Nystuen, J.A. 2007.
Noise level correlates with manatee use of foraging habitats. The Journal of the
Acoustical Society of America. 121, 3011.
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. B. and
Kent, J. 2000. Biodiversity hotspots for conservation priorities. Nature. 403, 853-858.
Nemeth, E., Brumm, H. 2009. Blackbirds sing higher-pitched songs in cities:
adaptation to habitat acoustics or side-effect of urbanization? Animal Behaviour. 78,
637-41.
Parks, E.S., Clark, C.W., Tyack, P.L. 2007. Short-and long-term changes in right
whale calling behavior: The potential effects of noise on acoustic communication.
Journal of the Acoustical Society of America.122, 3725-3731.
Pijanowski, B. C., Farina, A., Gage, S. H., Dumyahn, S. L. and Krause, B. L.
2011. What is soundscape ecology? An introduction and overview of an emerging new
science. Landscape Ecology. 26, 1213-1232.
Proppe, D.S, Sturdy, C.B, St. Clair, C.C. 2013. Anthropogenic noise decreases
urban songbird diversity and may contribute to homogenization. Global Change
Biology. 19, 1075-1084.

119
Referências bibliográficas
Saha, D. C, Padhy, P.K. 2011. Effect of air and noise pollution on species
diversity and population density of forest birds at Lalpahari, West Bengal, India.
Science of the Total Environment. 409, 5328-5336.
Schaub, A., Ostwald,J. Siemers, B.M., 2008. Foraging bats avoid noise. Journal
of Experimental Biology. 211, 3174-3180.
Slabbekoorn, H. Peet, M. 2003. Birds sing at a higher pitch in urban noise.
Nature. 424, 267.
Smith, A.C., Virgl, J.A., Panayi, D., Armstrong, A.R. 2005. Effects of a
diamond mine on Tundrabreeding birds. Arctic. 58, 295-304.
Sousa-Lima, R.S. Acoustic Ecology of Humpback Whales (Megaptera
novaeangliae) in the Abrolhos National Marine Park, Brazil. Ph.D. Dissertation, Field
of Zoology, Cornell University. 205 pp. 2007.
Sousa-Lima, R., Clark, C. 2008. Modeling the effect of boat traffic on the
fluctuation of humpback whale singing activity in the Abrolhos National Park, Brazil.
Canadian Acoustics. 36, 174-181.
Sousa-Lima, R.S, C. W. Clark. 2009. Whale sound recording technology as a
tool for assessing the effects of boat noise in a Brazilian marine park. Park Science. 26,
59.
Sun, J. W. C., Narins, P. M. 2005. Anthropogenic sounds differentially affect
amphibian call rate. Biological Conservation. 121, 419-427.
Vermeij A.J. M. Marhaver, K.L., Huijbers,C.M., Nagelkerken, I., Simpson, S.D.
2010. Coral larvae move toward reef sounds. Plos One 5.

120
Referências bibliográficas
Versace, E., Endress, A., Hauser, M. 2008. Pattern recognition mediates flexible
timing of vocalizations in nonhuman primates: experiments with cottontop tamarins.
Animal Behaviour. 76, 1885-1892.
Warren, P.S., Katti, M., Ermann, M., Brazel. 2006. Urban bioacoustics: it’s not
just noise. Animal Behaviour.71, 491-502.
Wolf, K. 2009. Bird song recognition through spectrogram processing and
labelling. Available on line at: http://www.tc.umn.edu/~wolfx265/.


















