
Morcegos vetores de pólen e dispersores de sementes no...
31
Morcegos vetores de pólen e dispersores de sementes no Pantanal Fernando Gonçalves Dissertação apresentada ao Programa de Pós Graduação em Ecologia e Conservação, Universidade Federal de Mato Grosso do Sul, para obtenção do titulo de mestre em Ecologia. Orientador: Erich Fischer Campo Grande 2009
-
Upload
nguyenkhanh -
Category
Documents
-
view
217 -
download
0
Transcript of Morcegos vetores de pólen e dispersores de sementes no...

Morcegos vetores de pólen e dispersores de sementes no
Pantanal
Fernando Gonçalves
Dissertação apresentada ao Programa de Pós Graduação em Ecologia e Conservação, Universidade Federal de Mato Grosso do Sul, para obtenção do titulo de mestre em Ecologia.
Orientador: Erich Fischer
Campo Grande 2009

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

Aos meus pais Angela e Dorival, que sempre acreditaram no estudo como a
maior herança deixada aos filhos. Ao meu irmão Diego que caminha no mesmo passo...

Agradecimentos Ao Erich Fischer pelos ensinamentos, motivações, orientação, paciência e grande amizade.
Aos membros da minha banca de qualificação, Geraldo Damasceno, Marcelo Bordignon e
Andréa Araujo pelas criticas ao trabalho. Ao Josué Raizer e principalmente Nicolay Cunha
e pela grande ajuda nas analises estatísticas. À Ângela Maria Correa pela ajuda na
identificação das espécies de pólen e Christiane Corrêa pela identificação das sementes. A
todos os integrantes do Projeto Morcegos do Pantanal (Alan, Amanda, Camila, Carol,
Claudia, Christiane, Danielle, Gabriela, George, Gustavo Graciolli, José Milton, Luis
Felipe, Nayara, Nicolay, Priscila, Reinaldo, Roberto, Silvana) pelo aprendizado, companhia,
coleta do material e risadas. Ao Roberto, Priscila e Guile pelo auxílio na triagem e viagens
a campo. A todos os alunos, professores, funcionários e freqüentadores do laboratório de
ecologia pela ótima convivência. Agradeço à FUNDECT-MS, Earthwarch Institute e ao
CNPQ/PELD pelo financiamento e à Conservação Internacional do Brasil pelo apoio
logístico e autorização para trabalhar na RPPN Fazenda Rio Negro.

Resumo
Morcegos desempenham importante papel na estruturação das comunidades tropicais, uma
vez que são eficientes polinizadores e as sementes defecadas por estes animais possuem
altas taxas de sobrevivência. O objetivo deste trabalho é descrever quais são as espécies de
pólen e sementes endozoocóricas transportadas por morcegos, bem como quais são as
espécies de morcegos transportadores de pólen e sementes, testar a variação da composição
das espécies vegetais entre duas estações (seca e chuvosa) na região da Fazenda Rio Negro,
Pantanal da Nhecolândia. Verificar também a sobreposição entre as espécies de plantas
quanto às espécies de morcegos transportadores de pólen e semente. Em 205 amostras
fecais foram encontradas 10 espécies de sementes: Ficus crocata (78/205), Ficus insipida
(4/205), Ficus obtusifolia (22/205), Maclura tinctoria (13/205), Piper aduncum (2/205),
Piper angustifolium (2/205), Piper tuberculatum (27/205), Cecropia pachystachya (55/205),
morfoespécie 1 (1/205) e morfoespécie 2 (1/205), sendo Artibeus planirostris, Platyrrhinus
lineatus, Sturnira lilium os principais dispersores. A família Moraceae foi similar quanto
aos morcegos dispersores de sementes, já a família Piperaceae apresentou pouca
similaridade. Em 182 amostras fecais de morcegos foram identificada nove espécies de
pólen: Bauhinia ungulata (84/182), Bauhinia sp. (1/182), Hymenaea courbaril (33/182),
Hymenaea stigonocarpa (25/182), Inga vera (3/182), Caryocar brasiliense (7/182),
Pseudobombax longiflorum (8/182), Psittacanthus corynocephalus (15/182) e uma espécie
não identificada (6/182) ocorrendo pouca similaridade entre as espécies de planta com
relação aos morcegos transportadores de pólen sendo G. soricina e A.planirostris os
principais vetores de pólen. A grande quantidade de sementes e de espécies vegetais
registradas nas fezes de A. planirostris indica que esta espécie pode ser considerada espécie
chave para a manutenção da comunidade de plantas quiropterocóricas e Glossophaga
soricina é o morcego responsável pelo transporte do maior número de espécies de pólen e
pode ser considerado o principal polinizador de plantas quiropterófilas no local de estudo.
Palavras-chaves: dispersão de sementes, herbivoria, interação morcego-planta, polinizadores potenciais, sazonalidade

2
Abstract
Bats play important role in structure of tropical assemblages, whereas are efficient
pollinators and the seeds defecated by these animals have high rate of survival. The aim of
this study is describe the species of pollen and endozoochoric seeds that are transported by
bats, and which species of bats carriers of pollen and endozoochoric seeds, test the variation
of plants species composition between seasons (dry and wet) in the Fazenda Rio Negro,
Pantanal of Nhecolândia region. And also, analyze the overlapping among plants species
and bats carriers of pollen and endozoochoric seeds. In 205 fecal sample were found 10
species of seeds: Ficus crocata (78/205), Ficus insipida (4/205), Ficus obtusifolia (22/205),
Maclura tinctoria (13/205), Piper aduncum (2/205), Piper angustifolium (2/205), Piper
tuberculatum (27/205), Cecropia pachystachya (55/205), morphospecie 1 (1/205) e
morphoespécie 2 (1/205), being Artibeus planirostris, Platyrrhinus lineatus, Sturnira lilium
main dispersers. The Moraceae family was similar in relationship of bats dispersers, but the
Piperaceae family showed little similarity. In 182 fecal sample were identified nine species
of pollen: Bauhinia ungulata (84/182), Bauhinia sp. (1/182), Hymenaea courbaril (33/182),
Hymenaea stigonocarpa (25/182), Inga vera (3/182), Caryocar brasiliense (7/182),
Pseudobombax longiflorum (8/182), Psittacanthus corynocephalus (15/182) and one non-
identified (6/182) occurring little similarity among plants species in relationship of bats
carriers of pollen, being Glossophaga soricina and A.planirostris main pollen carriers. The
large amount of seeds and plant species registered in fecal sample of A. planirostris
indicates that this species can be considered key-species for the maintenance of the bat-
plant community and Glossophaga soricina is responsible for transporting the largest
number of species of pollen and can be considered the main pollinator of bat-plants in the
study site.
Key words: bat-plant interaction, herbivore, potential pollinators, seasonality, seed
dispersal,

Introdução O sucesso no estabelecimento de plantas superiores envolve uma seqüência de estágios de
regeneração, incluindo a produção e polinização de flores, desenvolvimento de frutos,
dispersão e germinação de sementes, estabelecimento de plântulas, e crescimento até a
maturidade reprodutiva (Jordano & Godoy, 2002). A ruptura em algum desses processos ou
a redução na probabilidade de transmissão entre esses estágios resulta na diminuição do
recrutamento de plântulas. A polinização e a dispersão de sementes viáveis estão entre os
fatores mais importantes para a dinâmica populacional de várias espécies de planta (Baker
1955; Barrett et al.1996, Herrera et al. 2001, Wang & Smith 2002). As plantas apresentam
diversas adaptações que favorecem o transporte de pólen entre indivíduos e a dispersão de
propágulos para longe da planta mãe (Baker et al. 1998, Stiles 2000).
Em florestas tropicais, cerca de 250 espécies de plantas dependem dos morcegos
para polinização ou dispersão de sementes (Fleming 1982, Charles-Dominique & Cockle.
2001). Flores de Bauhinia, Hymenaea, Caryocar e Pseudobombax são comumente
visitadas por morcegos do gênero Artibeus, Platyrrhinus, Sturnira, Glossophaga, Anoura,
Carollia e Phyllostomus, (Fischer 1992, Gribel 1993, Gibbs 1999, Gribel & Gibbs 2002,
Santos 2004, Oliveira 2005). Estas flores oferecem néctar (açúcar) e pólen (proteína) e são
recompensadas com a polinização. Frutos de Cecropia, Ficus, Piper, Solanum são
comumente visitados por morcegos e as sementes são dispersas em suas fezes (Lobova et al.
2003, Thies & Kalko 2004, Mello et al. 2008, Silva et al. 2008, Teixeira et al. 2009).
Outras espécies de planta, entretanto, apresentam frutos contendo uma ou poucas sementes
grandes, que são derrubadas sob poleiros de alimentação após a ingestão da polpa (Fleming
& Williams 1990, Marques & Fischer 2009). O aparelho digestório dos morcegos é
relativamente curto, ocasionando rápida eliminação das fezes (≈ 30 min), e em pleno vôo,

2
aumentando a área de dispersão de sementes endozoocóricas em comparação com sementes
grandes que não são ingeridas (Muscarella & Fleming 2007).
Os morcegos desempenham papel importante na estruturação das comunidades
tropicais, uma vez que são eficientes polinizadores (Baker 1973, Fischer 1992, Gribel &
Hay 1993) e as sementes defecadas por esses animais possuem altas taxas de sobrevivência
(Morrison 1980; Figueiredo & Perin 1995, Teixeira et al 2009). Assim, estudos sobre
transporte de pólen e dispersão de sementes são importantes para determinar com quais
espécies vegetais os animais mantêm interações e como a sazonalidade pode afetar estas
interações. A intensidade da dependência e a conectividade entre espécies de morcegos
frugívoros e espécies de plantas são aspectos muito pouco estudados; embora conhecido
que algumas espécies de plantas atuam como recurso-chave para morcegos frugívoros
(Howe 1982, 1986, Fleming 1986, Marinho-filho 1991, Giannini & Kalko 2004). O
transporte de pólen e dispersão de sementes por morcegos neotropicais tem sido bastante
estudada na América Central e América do Sul (Heithaus et al. 1975, Willig & Moulton
1989, Handley et al. 1991, Fischer 1992, 2000, Fleming 1994, Marinho-Filho & Sazima
1998, Gribel 2002), porém foram encontrados poucos estudos deste tipo para o Pantanal.
O objetivo deste trabalho é descrever quais são as espécies de pólen e sementes
endozoocóricas transportadas por morcegos, bem como quais são as espécies de morcegos
transportadores de pólen e sementes, testar a variação da composição das espécies vegetais
entre duas estações (seca e chuvosa) na região da Fazenda Rio Negro, Pantanal da
Nhecolândia. Verificar também a sobreposição entre as espécies de plantas quanto às
espécies de morcegos transportadores de pólen e semente.

3
Materiais e métodos
Local de estudo
O estudo foi realizado na Reserva Particular do Patrimônio Natural Fazenda Rio Negro
(aproximadamente 19o 34´ S, 56o 14´ W), Aquidauana, Mato Grosso do Sul, Brasil. O
clima da região varia de sub-úmido a semi-árido, com estações chuvosa e seca bem
definidas. A pluviosidade média anual é de aproximadamente 1200 mm. O período úmido
ocorre de outubro a março, com pico de chuvas em dezembro; o período seco ocorre de
abril a setembro, com menor precipitação em agosto (Allem & Valls 1987). A Fazenda Rio
Negro (7500 ha) está localizada na região do Pantanal da Nhecolândia, caracterizada pela
presença de lagos circundados por extensos cordões de florestas decíduas ou semidecíduas,
denominados cordilheiras (Adamoli, 1987). Outros ambientes florestais são as matas
ciliares ao longo do rio Negro e os “capões”, que compreendem manchas (0,2 a 4 ha) de
florestas semidecíduas com formato esférico-elíptico, circundadas por campos
sazonalmente inundáveis (Araujo & Sazima 2003).
Coleta de dados
Para amostrar as espécies de frutos e pólen consumidos foram feitas 20 expedições ao
campo entre julho de 2002 e abril de 2007. As amostragens foram em julho, agosto e
dezembro de 2002; janeiro a agosto, novembro e dezembro de 2003; janeiro a abril e agosto
a dezembro de 2004; janeiro e julho de 2005; abril e junho de 2006; e abril de 2007.
Capturas de morcegos foram feitas durante lua nova ou minguante, utilizando redes neblina
(2,6 x 12 m) abertas por 6 h a partir do anoitecer. Para reportar as espécies de sementes e
pólen e a similaridade quanto aos morcegos transportadores, foram selecionados 18 sítios

4
amostrais, sendo seis sítios em cada tipo de ambiente florestal (capões, cordilheiras e matas
ciliares) sem o conhecimento prévio da presença de espécies de morcegos ou de recursos
utilizados por morcegos. As amostragens foram realizadas mensalmente durante três noites
consecutivas, sendo uma noite em cada tipo de ambiente. Selecionei ao azar sítios
diferentes de cada ambiente entre os meses de amostragem, de modo que cada sítio foi re-
amostrado após, no mínimo, seis meses de intervalo. O número de redes abertas por noite
variou de seis a doze entre os meses de amostragem, porém foi igual entre ambientes a cada
mês.
Os morcegos capturados foram acondicionados e mantidos em sacos de pano por
pelo menos 45 min, período considerado suficiente para defecarem caso tenham se
alimentado antes da captura (Studier et al. 1983). As amostras fecais foram coletadas e
acondicionadas individualmente em microtubos herméticos contendo glicerina, para
posterior identificação de sementes e pólen. Os sacos de pano foram utilizados apenas uma
vez por expedição, e foram lavados e esterilizados entre expedições. As espécies de
morcegos foram identificadas no local e pelo menos um indivíduo de cada espécie de
morcego foi coletado para confirmação da identificação e inclusão como testemunho na
coleção de Zoologia da Universidade Federal de Mato Grosso do Sul (ZUFMS-M).
As amostras fecais foram inspecionadas em laboratório com auxílio de
estereomicroscópio para registrar a presença de sementes, e com auxilio de microscópio
para registrar a presença de pólen e preparo de lâminas frescas. As sementes e pólen
encontrados foram identificados por comparação com material coletado de plantas no local
de estudo, consulta ao material de herbário (CGMS) e auxilio de especialista.

5
Análise de dados
Para avaliar a similaridade entre as espécies de plantas quanto às espécies de morcegos
visitantes de flores ou frutos foram feitas ordenações pela técnica de escalonamento
multidimensional não-métrico (NMDS). Esta técnica não assume relações lineares entre
variáveis ou distâncias métricas, apresenta mais tolerância a células vazias na matriz de
dados e pode sumarizar mais informações em menos eixos do que outras técnicas de
ordenação (Manly 1986). Para avaliar a dissimilaridade entre as espécies de plantas quanto
aos morcegos que transportam pólen ou sementes foi usado o índice de Bray-Curtis, que
não atribui similaridade devido à ausência comum de determinada espécie de morcego
visitante (Faith et al. 1987, Belbin 1992). Para avaliar a sazonalidade da ocorrência de
espécies de sementes e de pólen transportadas por morcegos foi calculada a média mensal
do número de amostras fecais contendo cada espécie de planta.
Resultados
Espécies de sementes transportadas por morcegos
Em 205 amostras fecais foram encontradas 10 espécies de sementes transportadas pelos
morcegos, sendo quatro Moraceae (Ficus crocata, Ficus insipida, Ficus obtusifolia e Maclura
tinctoria), três Piperaceae (Piper aduncum, Piper angustifolium e Piper tuberculatum), uma
Urticaceae (Cecropia pachystachya) e duas espécies não identificadas (Tabela 1). As espécies
de sementes mais freqüentes foram F. crocata (78/205), C. pachystachya (55/205), P.
tuberculatum (27/205) e F. obtusifolia (22/205). As espécies menos freqüentes foram
morfoespécie 1 e morfoespécie 2 (1/205), P. aduncum e P. angustifolium (2/205), F. insipida
(4/205) e M. tinctoria (13/205) (Tabela 1).

6
Foram registradas 10 espécies de morcegos dispersores de sementes endozoocóricas.
Artibeus planirostris e Platyrrhinus lineatus (Phyllostomidae; Stenodermatinae) foram as
espécies de morcego que dispersaram maior quantidade de espécies de sementes (n = 7 e n = 5,
respectivamente). Artibeus lituratus, Sturnira lilium (Phyllostomidae; Stenodermatinae), e
Glossophaga soricina (Phyllostomidae; Glossophaginae) dispersaram quatro espécies de
sementes; Carolia perspicillata (Phyllostomidae; Carolliinae) e Lophostoma silvicolum
(Phyllostomidae; Phyllostominae) dispersaram três espécies, e Phyllostomus discolor,
Phyllostomus hastatus (Phyllostomidae; Phyllostominae) e Noctilio albiventris (Noctilionidae)
dispersaram duas espécies de sementes (Tabela 1).
A análise de ordenação (NMDS) mostrou que espécies de Moraceae (F. crocata, F.
insipida, F. obtusifolia e M. tinctoria) são similares quanto aos morcegos dispersores de
sementes (Fig. 1), que foram principalmente A. planirostris e P. lineatus (Tabela 1). Sementes
de C. pachystachya também foram dispersas por estas duas espécies de morcegos, além de S.
lilium. Por outro lado, espécies de Piperaceae (P. aduncum, P. angustifolium e P.
tuberculatum) apresentaram pouca similaridade entre si quanto às espécies de morcegos
dispersores de sementes (Fig. 1). Piper aduncum, cujas sementes foram dispersas
principalmente por A. planirostris e G. soricina, foi mais semelhante às espécies de Moraceae;
ao passo que sementes de P. tuberculatum foram dispersas principalmente por S. lilium, e as
de P. angustifolium por C. perspicillata (Tabela 1).
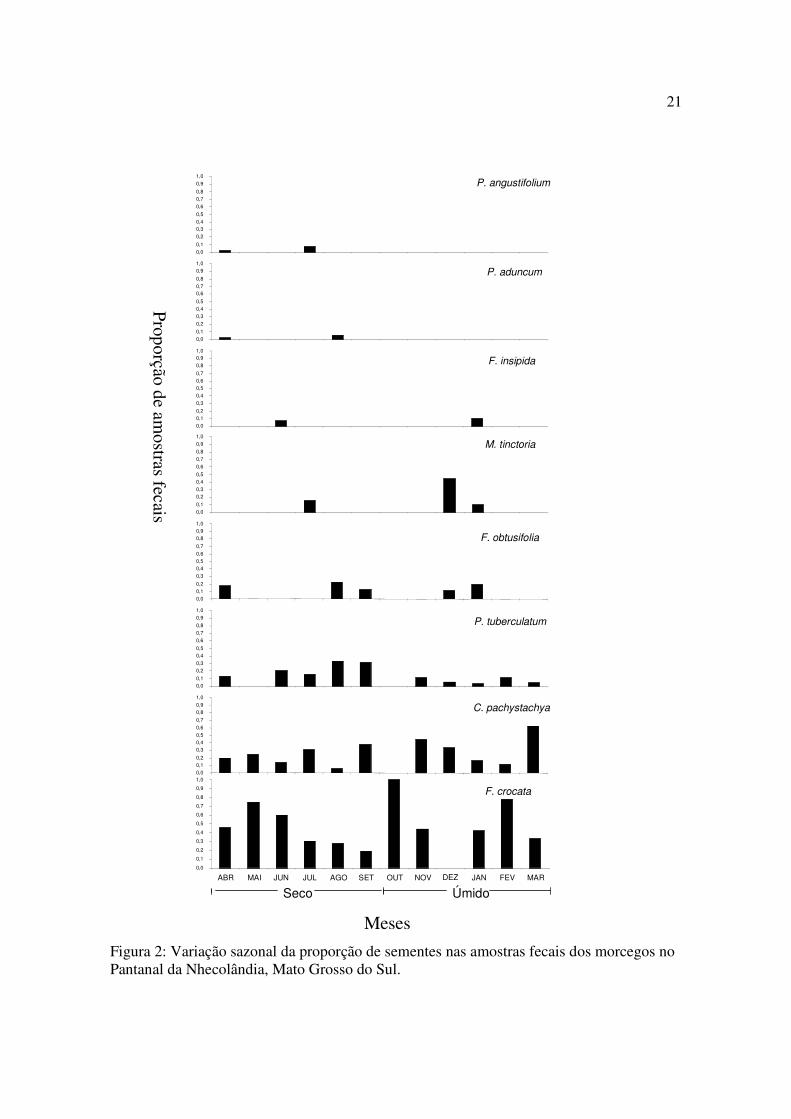
Sementes de Moraceae foram dispersas em ambas as estações. F. insipida ocorreu nas
fezes de morcegos em janeiro e junho, F. obtusifolia ocorreu em janeiro, abril, agosto,
setembro e dezembro, M. tinctoria em janeiro, julho e dezembro, e F. crocata ocorreu em
todos os meses, exceto em dezembro. C. pachystachya ocorreu em todos os meses do ano,
exceto em outubro (Fig. 2). Dentre as três espécies de Piperaceae, sementes de duas espécies

7
foram dispersadas somente na estação seca, P. angustifolium em abril e julho, e P. aduncum
em abril e agosto. Sementes de P. tuberculatum ocorreram em todos os meses do ano, exceto
em maio e outubro (Fig 2).
Espécies de pólen transportadas por morcegos
Em 182 amostras fecais de morcegos foram identificadas a presença de nove espécies de pólen,
cinco Fabaceae (Bauhinia ungulata, Bauhinia sp., Hymenaea courbaril, Hymenaea
stigonocarpa e Inga vera), uma Caryocaraceae (Caryocar brasiliense), uma Malvaceae
(Pseudobombax longiflorum), uma Loranthaceae (Psittacanthus corynocephalus) e uma
espécie não identificada (Tabela 2). As espécies de pólen mais comuns nas fezes dos
morcegos foram B. ungulata (84/182), H. courbaril (33/182), H. stigonocarpa (25/182) e P.
corynocephalus (15/182). Cinco espécies de pólen foram pouco freqüentes, Bauhinia sp.
(1/182), I. vera (3/182), morfoespécie 1 (6/182), C. brasiliense (7/182) e P. longiflorum
(8/182) (Tabela 2).
Treze espécies de morcegos transportaram pólen. Glossophaga soricina foi a que
apresentou a maior riqueza de espécies de pólen (n = 8) nas amostras fecais; outras espécies de
morcegos que apresentaram pólen foram A. planirostris (n = 7), S. lilium (n = 6), P. discolor
(n = 5), P. lineatus e C. perspicillata (n = 4), L. silvicolum, P. hastatus (n = 3), A. lituratus, C.
auritus, L. brasiliense e N. albiventris (n = 1).
A análise de ordenação (NMDS) mostrou que há pouca similaridade entre as espécies
de planta quanto aos morcegos que transportam pólen. Inga vera e H. stigonocarpa (Fabaceae)
foram mais semelhantes, assim como B. ungulata e Morfoespécie 1. Pseudobombax
longiflorum (Malvaceae) foi mais similar a H. courbaril e Bauhinia sp. Caryocar brasiliense

8
(Caryocaraceae) e P.corynocephalus (Loranthaceae) foram as espécies que apresentaram
maior diferença quanto aos morcegos que transportam pólen (Fig. 4).
Dentre as nove espécies de pólen presentes nas amostras fecais dos morcegos, três
ocorreram somente no período seco – Bauhinia sp. (junho), P. longiflorum (abril, junho e
julho) e H. courbaril (abril, julho, agosto e setembro) e C. brasiliense ocorreu apenas no
período úmido (janeiro e fevereiro). As outras cinco espécies de pólen ocorreram em ambas as
estações, com destaque para B. ungulata que ocorreu em todos os meses do período seco (abril,
maio, junho, julho, agosto e setembro) e em dois meses do período úmido (janeiro e março)
(Fig 5).
Discussão
As espécies de Moraceae apresentaram grande semelhança em relação aos morcegos
dispersores de sementes, sendo A. planirostris e P. lineatus os principais dispersores no
local de estudo, assim como em outras localidades (Fleming 1986, Dumont 2003, Giannini
& Kalko 2004). Esta família inclui muitas espécies hemiepífitas, que possuem adaptações
para crescimento em plantas forófitas (Benzing 1990, Corrêa 2005) e dependem de
morcegos para alcançarem locais propícios para o estabelecimento inicial (Corrêa 2005,
Teixeira et al. 2009). Ficus crocata, F. insipida e F. obtusifolia possuem características
morfológicas de plantas quiropterocóricas (Kalko et al 1996) sendo preferencialmente
dispersas por morcegos.
A espécie pioneira C. pachystachya (Urticaceae), única do gênero no Pantanal (Pott
& Pott 1994), foi semelhante às espécies de Moraceae quanto aos dispersores de sementes,
sendo A. planirostris o principal dispersor, assim como em outras localidades (Lobova et al
2003, Teixeira et al. 2009). Essas plantas do gênero Cecropia desempenham importante

9
papel ecológico, pois são as primeiras a estabelecerem em áreas perturbadas e os morcegos
são os grandes responsáveis pelo movimento desses diásporos das florestas secundárias
para as primárias (Lobova et al 2003). A passagem dos frutos de Cecropia pelo trato
digestório dos morcegos remove o perianto e toda parte mucilaginosa, reduzindo assim a
aderência dos propágulos, facilitando a dispersão das sementes e aumentando a taxa de
germinação (Estrada et al. 1984, Estrada and Coates-Estrada 1986, Fleming 1988).
Espécies de Piperaceae apresentaram pouca semelhança em relação aos morcegos
dispersores, contrastando com alguns locais onde as Piperáceas são dispersas
principalmente por morcegos Carollia spp. (Fleming et al. 1972, Howell & Burch 1974,
Charles-Dominique 1986, 1991, Fleming 1988, 1991, Palmeirim et al. 1989, Marinho-Filho
1991, Muller & dos Reis 1992, Gorchov et al. 1995 Thies & Kalko 2004). Esse contraste
pode ser explicado pela baixa ocorrência de Piperaceae no Pantanal (Teixeira et al. 2009), o
que leva os animais a consumirem os frutos de acordo com a disponibilidade no local.
Sementes de P. angustifolium e P. aduncum ocorreram nas amostras fecais somente
na estação seca. A oferta de frutos maduros antes da estação chuvosa proporciona às
sementes grande chance de germinação devido à umidade favorável (van Schaik et al. 1993,
Pedroni 2002). Além disso, as plântulas poderiam desenvolver o sistema radicular durante a
estação de chuvas, antes de serem submetidas ao estresse hídrico da estação seca
subseqüente (Janzen 1967). Três espécies de Moraceae (F. insipida, F. obtusifolia e M.
tinctoria) obtiveram uma ocorrência irregular nas fezes dos morcegos, ocorrendo tanto nos
meses de período seco quanto de período úmido, porém em baixas quantidades. A
freqüência irregular de frutificação é considerada uma estratégia para reduzir a predação de
sementes através da saciação dos predadores (Janzen 1975, Tapper 1992). Períodos de
grande produção de sementes seguidos de períodos de escassa produção regulam as

10
populações de predadores (Silvertown 1980). Plantas como C. pachystachya, F. crocata e P.
tuberculatum apresentaram uma produção continua de frutos ao longo do ano e em ambas
estações. Essa estratégia garante oferta de recursos aos dispersores por longo período,
elevando as chances de dispersão (Milton et al. 1982, Pereira e Mantovani 2001).
Por ser uma formação recente, de clima semi-árido, e sob influência do cerrado e do chaco
paraguaio, o Pantanal não possui endemismos e grande riqueza de espécies zoocóricas (Pott
& Pott 1994, Ragusa-Netto 2004, Teixeira et al. 2009). Este fato pode explicar a baixa
riqueza de sementes endozoocóricas (10 espécies, três famílias) dispersas pelos morcegos
no local de estudo em comparação com florestas úmidas ou semidecíduas (Palmerin et al.
1989, Passos et al. 2003, Giannini & Kalko 2004, Lopez & Vaughan 2007, Silva et al.
2008). A grande quantidade de sementes e de espécies vegetais registradas nas fezes de A.
planirostris indica que esta espécie pode ser considerada espécie chave para a manutenção
da comunidade de plantas quiropterocóricas no local de estudo, assim como sugerido por
Teixeira et al. (2009) em região do Pantanal do Miranda.
Segundo Von Helversen (1993) morcegos com massa corpórea acima de 50 g, como
A. planirostris, A. lituratus, P. hastatus, normalmente visitam flores grandes, como aquelas
de espécies de Hymenaea, Caryocar e Pseudobombax. Os morcegos com massa corpórea
abaixo de 50 g, como G. soricina, S. lilium, C. perspicillata, P. discolor, normalmente
visitam flores menores e mais delicadas (von Helversen, 1993; Muchhala & Jarrín, 2002;
Muchhala, 2006), como as de espécies de Bauhinia, Inga, Psittacanthus. Entretanto, a
similaridade quanto aos morcegos vetores de pólen foi baixa entre as espécies de plantas,
principalmente devido a grande variação de espécies de morcegos que visitaram
ocasionalmente apenas algumas espécies de plantas.

11
Pólen de Bauhinia sp., P. longiflorum e H. courbaril ocorreram somente na estação
seca nas amostras fecais dos morcegos. Por ser um período de baixa disponibilidade de
recursos alimentares aos morcegos (Teixeira et al. 2009), plantas que florescem nessa
estação conseqüentemente atraem mais espécies de animais visitantes florais (Janzen 1997).
Pólen de C. brasiliense ocorreu somente na estação úmida, plantas que florescem neste
período possuem características que as favorecem, pois nesta estação há um aumento nas
populações e atividade dos animais vetores de pólen (Wikander 1984), e os elevados
índices pluviométrico e de temperaturas nesta estação, acarretam no aumento na
decomposição da serapilheira e nos teores de nutrientes disponíveis para as plantas
(Morellato 1992). Pólen de B. ungulata, P. corynocephalus, ocorreram em ambas as
estações nas amostras fecais dos morcegos com B. ungulata e P. corynocephalus sendo
mais abundantes na estação seca e G. soricina o maior responsável pelo transporte de pólen,
de ambas, 30% e 80 % respectivamente e I. vera e H. stigonocarpa sendo mais abundantes
na estação chuvosa e A. planirostris o maior responsável pelo transporte de pólen de ambas
as espécies, 50 %. Esse pico de floração em determinadas estações pode ser uma estratégia
das plantas para atrair determinadas espécies de morcegos vetores de pólen.
Nos neotrópicos, são conhecidas aproximadamente 40 famílias de plantas que
possuem flores visitadas por morcegos (Fleming & Muchhala 2007), porém no Pantanal
ocorreram apenas quatro famílias. Este resultado poderia ser devido ao fato deste estudo
considerar apenas a presença de pólen nas amostras fecais. Entretanto, por ser uma
formação recente e apresentar condições estressantes, o Pantanal possui baixa riqueza de
plantas (Pott & Pott 1994). Glossophaga soricina é o morcego responsável pelo transporte
do maior número de espécies de pólen (n = 8) e pode ser considerado o principal
polinizador de flores quiropterófilas no local de estudo.

12
Literatura Citada Adamoli J (1987) Fisiografia do Pantanal. In: Allem AC, Valls JFM (eds.) Recursos
Forrageiros Nativos do Pantanal Matograssense. DDT-EMBRAPA, Brasília, pp. 23–25 Allem AC, Valls JFM (1987) Recursos forrageiros nativos do Pantanal Mato-Grossense.
Cenagem/ Embrapa-Cpap Brasília Araujo AC, Sazima M (2003) The assemblage of flowers visited by hummingbirds in the
capões of Southern Pantanal, Mato Grosso do Sul, Brazil. Flora 198: 427-435 Baker HG (1955) Self compatibility and establishment after long-distance dispersal.
Evolution 9: 347–349 Baker HG (1973) Evolutionary relationships between plants and animals in American and
African forest. In: Meggers BJ, Ayensu ES, Duckwortii WD (eds.) Tropical forest eeosystems in Africa and South Ameriean: A Comparative review. Washington, Smithsoniam Institution Press, pp 145-159
Baker HG, Baker I, Hodges SA (1998) HodgesSugar Composition of Nectars and Fruits
Consumed by Birds and Bats in the Tropics and Subtropics. Biotropica 30(4): 559-586 Belbin L (1992) User’s guide PATN. Commonwealth Scientific Industrial Research
Oragnization (CSI-RO). Australian Capital Territory, Canberra Benzing DH (1990) Vascular epiphytes - General biology and realted biota. Cambridge
University Press, New York Charles-Dominique P (1986) Inter-relations between frugivorous vertebrates and pioneer
plants: Cecropia, birds and bats in French Guiana. In: Estrada A, Fleming TH (eds) Frugivores and seed dispersal. Dr. W. Junk, Dordrecht, pp. 119–135
Charles-Dominique P (1991) Feeding strategy and activity budget of the frugivorous bat
Carollia perspicillata (Chiroptera: Phyllostomidae) in French Guiana. Journal of Tropical Ecology 7: 243–256
Charles-Dominique P, Cockle A (2001) Frugivory and seed dispersal by bats. In: Bongers F,
Charles-Dominique P, Forget PM, Thery M (eds.). Dynamics and plant-animal interactions in a Neotropical rainforest. Kluwer Academic Publishers. Nouragues pp. 206-215
Corrêa CE (2005) Comunidade de sementes em caules de attalea phalerata mart. ex.
spreng. (ARECACEAE) no Pantanal Sul. Dissertação de mestrado, Universidade Federal de Mato Grosso do Sul, Campo Grande
Dumont ER (2003) Bats and fruits: An Ecomorphological Approach. In: Kunz TH, Fenton
MB (eds.) Bat ecology. The University of Chicago Press, Chicago, pp 398-429

13
Estrada A, Coates-Estrada R, Vásquez-Yanes C. (1984) Observations on fruiting and dispersers of Cecropia obtusifolia at Los Tuxtlas, Mexico. Biotropica 16: 315–318
Estrada A, Coates-Estrada R (1986) Frugivory by howling monkeys (Allouata palliata) at
Los Tuxtlas, Mexico: dispersal and fate of seeds. In: Estrada A, Fleming TH (eds.) Frugivores and seed dispersal. Dr. W. Junk, Dordrecht, Netherlands, pp. 93–104
Faith DP, Minghin PR, Belbin L (1987) Composition dissimilarity as a robust measure of
ecological distance: a theoretical model and computer simulations. Vegetatio 69: 57-58 Figueiredo RA, Perin E (1995) Germination ecology of Fícus luschnathiana druplets after
bird and bat ingestion. Acta Oecologica, 16:71-75 Fischer E (1992) Foraging of nectarivorous bats on Bauhinia ungulata. Biotropica 24: 579-
582 Fischer E (2000) Polinização por morcegos Glossophaginae versus Phyllostominae em
floresta de terra firme na Amazônia Central. Tese de doutorado, Universidade Estadual de Campinas, Campinas
Fleming TH, Hooper ET, Wilson DE (1972) Three Central American bat communities,
structure, reproductive cycles and movement patterns. Ecology 53: 555–569 Fleming TH (1982) Foraging strategies of plant-visiting bats. In: Kunz TH (ed) Ecology of
bats. Plenum Press, New York. pp 287–325 Fleming TH, Heithaus ER (1986) Seasonal foranging behavior of the frugivorous bat
Carollia perspicillata. Journal of Mammalogy 67 (4): 660-671 Fleming TH (1988) The short-tailed fruit bat. University of Chicago Press, Chicago Fleming TH, Williams CF (1990) Phenology, seed dispersal, and recruitment in Cecropia
peltata (Moraceae) in Costa Rican tropical dry forest. Journal of Tropical Ecology 6:163–178
Fleming TH, Maurice S,. Buchmann SL,. Tuttle MD (1994) Reproductive biology and
relative male and female fitness in a trioecious cactus, Pachycereus pringlei (Cactaceae). American Journal of Botany 81: 858–867
Fleming TH, Muchhala N (2008) Nectar-feeding bird and bat niches in two worlds:
pantropical comparisons of vertebrate pollination systems. Journal of Biogeography 35: 764–780
Giannini NP, Kalko EKV (2004) Trophic structure in a large assemblage of Phyllostomid
bats in panama. Oikos 105: 209-220

14
Gibbs PE, Oliveira PE, Bianchi MB (1999) Post-zygotic control of selfing in Hymenaea
stigonocarpa (Leguminosae-Caesalpinioideae), a bat-pollinated tree of the Brazilian cerrados. International Journal of Plant Science 160: 72–78
Gorchov DL, Cornejo F, Ascorra CF (1995) Dietary overlap between frugivorous birds and
bats in the Peruvian Amazon. Oikos 74: 235–250 Gribel R, Hay JD (1993) Pollination ecology of Caryocar brasiliense (Cariocaraceae) in
Central Brazil cerrado vegetation. Journal of Tropical Ecology 9: 199-211 Gribel R, Gibbs PE (2002) High outbreeding as a consequence of selfed ovule mortality
and single vector bat pollination in the Amazonian tree Pseudobombax munguba
(Bombacaceae). International Journal of Plant Science 163: 1035-1043 Gribel R, Gibbs PE, Queiroz AL (2002) Flowering phenology and pollination biology of
Ceiba pentandra (Bombacaceae) in central Amazonia. Journal of Tropical Ecology 15: 247–263
Handley CO, Wilson DE, Gardner AL (1991) Demography and natural history of the
common fruit bat, Artibeus jamaicensis, on the Barro Colorado Island, Panama. Smithsonian Institution, Washington
Herrera CM, Sanchez-Lafuente AM, Medrano M,. Guitia JN, Cerda X, Rey P (2001)
Geographical variation in autonomous selfpollination levels unrelated to pollinator service in Helleborus foetidus (Ranunculaceae). American Journal of Botany 88: 1025–1032
Howell DJ, Burch D (1974) Food habits of some Costa Rican bats. Revista de Biologia
Tropical 21: 281–294 Howe HF, Smallwood J (1982) Ecology of seed dispersal. Annual Review of Ecology and
Systematic 13: 201-228 Janzen D (1967) Synchronization of sexual reproduction of trees within the dry season in
Central America. Evolution 21: 620-637 Janzen D (1975) Behaviour of Hymenaea courbaril when its predispersal seed predator is
absent. Science 189:145-147 Jordano P, Godoy JA (2002) Frugivore-generated seed shadows: a landscape view of
demographic and genetics effects. In: Levey DJ, Silva WR, Galetti M (eds) Seed Dispersal and frugivory: ecology, evolution and conservation. CAB Interanational Publishing, Wallingford. Brasil, pp 305-321
Kalko EKV, Herre EA, Handley CO (1996) Relation of fig fruit characteristics to fruit-
eating bats in the New and Old World tropics Journal of Biogeography. 23: 565-576

15
Lobova TA, Moris SA, Blanchard F, Peckham H, Charles-Dominique P (2003) Cecropia as a food resource for bats in French Guiana and the significance of fruit structure in seed dispersal and longevity. American Journal of Botany 90: 388-403
Lopez JE, Vaughan C (2007) Food niche overlap among neotropical frugivorous bats in
Costa Rica. Revista de Biologia Tropical 55: 301-313 Manly BJ (1986) Multivariate statistical methods primer. Caphman and Hall, New York. Marinho-Filho JS (1991) The coexistence of two frugivorous bat species and the phenology
of their food plants in Brazil. Journal of Tropical Ecology 7: 59–67 Marinho-Filho JS, Sazima I (1998) Brazilian bats and conservation. In: Kunz TH, Racey P
(eds.) Bat biology and conservation. Smithsonian Institutional Press Marques MCM, Fischer E (2009) Effect of bats on seed distribution and germination of
Calophyllum brasiliense (Clusiaceae). Ecotropica 15:1-6 Mello MAR, Kalko EKV, Silva WR (2008). Diet and abundance of the bat Sturnira Lilium
(CHIROPTERA) in Brazilian Montane Atlantic Forest. Journal of Mammalogy 89(2): 485-492
Milton K, Windsor DM, Morrison DW, Estribi MA (1982) Fruiting phenologies of two
tropical Ficus species. Ecology 63: 752-762 Morellato LPC (1992) Sazonalidade e dinâmica de ecossistemas florestais na Serra do Japi.
In: Morellato LPC (ed) História natural da Serra do Japi: ecologia e preservação de uma área florestal no Sudeste do Brasil. Universidade Estadual de Campinas/Fundação de Amparo à Pesquisa do Estado de São Paulo, Campinas, pp. 98-110
Morrison DW (1980) Foraging and day-roosting dynamics of canopy fruit bats in Panama.
Journal of Mammalogy 61: 20–29 Muchhala N, Jarrín VP (2002) Flower visitation by bats in cloud forests of western Ecuador.
Biotropica 34: 387–395 Muchhala N (2006) The pollination biology of Burmeistera (Campanulaceae):
specialization and syndromes. American Journal of Botany 93 1081–1089 Muller MF, dos Reis NR (1992) Partição de recursos alimentares entre quatro especies de
morcegos frugívoros (Chiroptera, Phyllostomidae). Revista Brasleira Zoologia 9: 345–355
Muscarella R, Fleming TH (2007) The Role of Frugivorous Bats in Tropical Forest
Succession. Biological Reviews 82: 573–590

16
Oliveira R (2006) Secreção de néctar e atividade de morcegos em Hymenaea stigonocarpa (Leguminosae-Caesalpinioideae) no Pantanal da Nhecolândia e remanescente urbano de cerrado, Mato Grosso do Sul . Dissertação de mestrado, Universidade Federal de Mato Grosso do Sul, Campo Grande
Palmeirim J, Gorchov DL, Stoleson S (1989) Trophic structure of a neotropical frugivore
community: is there competition between birds and bats? Oecologia 79: 403–411 Passos FC, Silva WR, Pedro WA, Bonin MR (2003) Frugivoria em Morcegos (Chiroptera,
Phyllostomidae) no Parque Estadual Intervales, sudeste do Brasil. Revista Brasileira de Zoologia 20: 511-517
Pedroni F, Sanches M, Santos FAM (2002) Fenologia da copaíba (Copaifera langsdorffii
Desf. Leguminosae, Caesalpinioideae) em uma floresta semidecídua no sudeste do Brasil. Revista Brasileira de Botânica 25: 183-194
Pereira ST, Mantovani W (2001) Maturação e dispersão de Miconia cinnamomifolia (DC)
Naud. na Reserva Biológica de Poço das Antas, município de Silva Jardim, RJ, Brasil. Acta Botânica Brasília 15:335-348
Pott A, Pott VJ (1994) Plantas do Pantanal. Empresa Brasileira de Pesquisa Ragusa-Netto J (2004) Flowers, fruits, and the abundance of the Yellow-chevroned
Parakeet (Brotogeris chiriri) at a gallery forest in the south Pantanal. Brazilian Journal of Biology 64: 867-877
Santos CF (2004) Produção de néctar e visitas de morcegos a caryocar brasiliense no
Pantanal da Nhecolândia, Dissertação de mestrado, Universidade Federal de Mato Grosso do Sul, Campo Grande
Silva AG, Gaona O, Medellín RA (2008) Diet and trophic structure in a community of
fruit-eating bats in lacandon forest Mèxico. Journal of Mammalogy 89(1):43–49 Silvertown JW (1980) The evolutionary ecology of mast seeding in trees. Biological
Journal of the Linnean Society 14: 235-250 Stiles EW (2000) Animals as Seed Dispersers. In: Fenner M (ed) Seeds: The Ecology of
Regeneration in Plant Communities. UK, CAB International., pp 111-124 Studier EH, Boyd BC, Felman AT, Dapson R, Wilson DE (1983) Renal function in the
Neotropical bat, Artibeus jamaicensis. Comparative Biochemistry and Physiology 74:199–209.
Tapper PG (1992) Irregular fruiting in Fraxinus excelsior. Journal of Vegetation Science 3:
41-46

17
Teixeira RC, Corrêa CE, Fischer E (2009) Frugivory by Artibeus jamaicensis
(Phyllostomidae) bats in the Pantanal, Brazil. Studies on Neotropical Fauna and Environment 44: 7-15
Thies W, Kalko EKV (2004) Phenology of neotropical pepper plants (Piperaceae) and their
association with their main dispersers, two short-tailed fruit bats, Carollia perspicillata and C. castanea (Phyllostomidae). Oikos 104: 362-376
Van Schaik CP, Terborgh JW, Wright SJ (1993) The phenology of tropical forest:
adaptative significance and consequences of consumers. Annual Review of Ecology and Systematics 24: 353-377
von Helversen O (1993) Adaptations of flowers to the pollination by glossophagine bats. In:
Barthlott W, Naumann CM, Schmidt-Loske K, Schuchmann K (eds) Plant–animal interactions in tropical environments. Museum Alexander Koenig, Bonn, Germany. pp. 41–59
Wang BC, Smith TB (2002) Closing the seed dispersal loop. Trends in Ecology &
Evolution 17: 379-385 Wikander T (1984) Mecanismos de dispersión de diásporas de una selva decidua en
Venezuela. Biotropica 16: 276-283 Willig MR, Moulton MP (1989) The role of stochastic and deterministic processes in
structuring Neotropical bat communities. Journal of Mammalogy 70: 323-329
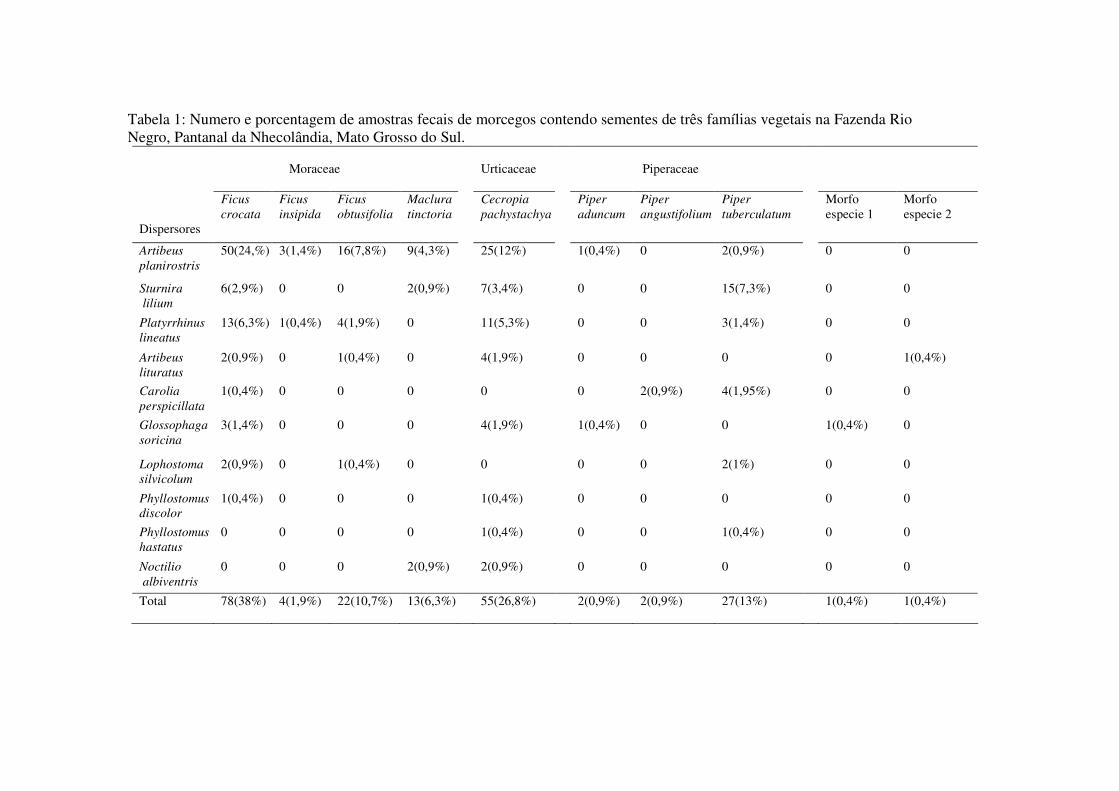
Tabela 1: Numero e porcentagem de amostras fecais de morcegos contendo sementes de três famílias vegetais na Fazenda Rio Negro, Pantanal da Nhecolândia, Mato Grosso do Sul.
Moraceae
Urticaceae
Piperaceae
Dispersores
Ficus
crocata
Ficus
insipida
Ficus
obtusifolia
Maclura
tinctoria
Cecropia
pachystachya
Piper
aduncum
Piper
angustifolium
Piper
tuberculatum
Morfo especie 1
Morfo especie 2
Artibeus
planirostris
50(24,%) 3(1,4%) 16(7,8%) 9(4,3%) 25(12%) 1(0,4%) 0 2(0,9%) 0 0
Sturnira
lilium
6(2,9%) 0 0 2(0,9%) 7(3,4%) 0 0 15(7,3%) 0 0
Platyrrhinus
lineatus
13(6,3%) 1(0,4%) 4(1,9%) 0 11(5,3%) 0 0 3(1,4%) 0 0
Artibeus
lituratus
2(0,9%) 0
1(0,4%) 0 4(1,9%) 0 0 0 0 1(0,4%)
Carolia
perspicillata
1(0,4%) 0 0 0 0 0 2(0,9%) 4(1,95%) 0 0
Glossophaga
soricina
3(1,4%) 0 0 0 4(1,9%) 1(0,4%) 0 0 1(0,4%) 0
Lophostoma
silvicolum
2(0,9%) 0 1(0,4%) 0 0 0 0 2(1%) 0 0
Phyllostomus
discolor
1(0,4%) 0 0 0 1(0,4%) 0 0 0 0 0
Phyllostomus
hastatus
0 0 0 0 1(0,4%) 0 0 1(0,4%) 0 0
Noctilio
albiventris
0 0 0 2(0,9%) 2(0,9%) 0 0 0 0 0
Total 78(38%) 4(1,9%) 22(10,7%) 13(6,3%) 55(26,8%)
2(0,9%) 2(0,9%) 27(13%) 1(0,4%) 1(0,4%)

Tabela 2: Numero e porcentagem de amostras fecais de morcegos contendo pólen de cinco famílias vegetais na Fazenda Rio Negro, Pantanal da Nhecolândia, Mato Grosso do Sul.
Fabaceae
Caryocaraceae
Malvaceae
Loranthaceae
Transportadores
Bauhinia
ungulata Bauhinia
Sp. Hymenaea
courbaril Hymenaea
stigonocarpa
Inga vera Caryocar
brasiliense Pseudobombax
longiflorum
Psittacanthus
corynocephalus Morfo
espécie 1
Artibeus planirostris
25(13,7%) 0
4(2,1%) 12(6,5%) 2(1%) 5(2,6%) 1(0,5%) 0 1(0,5%)
Sturnira lilium
10(5,5%) 0 3(1,6%) 2(1%) 0 1(0,5%) 1(0,5%) 0 2(1%)
Platyrrhinus lineatus
9(4,9 %) 0 5(2,6%) 2(1%) 0 0 0 2(1%) 0
Artibeus lituratus
2(1%) 0 0 0 0 0 0 0 0
Carolia perspicillata
3(1,6%) 0 1(0,5%) 0 0 0 1(0,5%) 2(1%) 0
Glossophaga soricina
26(14,2%) 1(0,5%) 16(8,7%) 6(3,2 %) 1(0,5%) 0 5(2,6%) 8(4,3%) 3(1,5%)
Lophostoma silvicolum
2(1%) 0 1(0,5%) 1(0,5%) 0 0 0 0 0
Phyllostomus hastatus
3(1,6%) 0 1(0,5%) 0 0 0 0 2(1%) 0
Phyllostomus discolor
1(0,5%) 0 2(1%) 1(0,5%) 0 1(0,5%) 0 1(0,5%) 0
Crothopterus auritus
1(0,5%) 0 0 0 0 0 0 0 0
Lophostoma brasiliense
0 0 0 1(0,5%) 0 0 0 0 0
Noctilio albiventris
2(1%) 0 0 0 0 0 0 0 0
Total 84(46,1%) 1(0,5%) 33(18,1%) 25(13,7%) 3(1,6%) 7(3,8 %) 8(4,3%) 15(8,2%) 6(3,2%)

Figura 1: A: similaridade entre plantas quanto aos morcegos dispersores (NMDS uma dimensão, stress 18% e variância recuperada de 53%). B: proporções das espécies de sementes registradas nas amostras fecais dos morcegos no Pantanal da Nhecolândia, Mato Grosso do Sul. P.adu = Piper aduncum, C. pac = Cecropia pachystachya, F. obt = Ficus obtusifolia, F. cro = Ficus crocata, M. tin = Maclura tinctoria, F. ins = Ficus insipida, P.tub = Piper
tuberculatum, P ang = Piper angustifolium.
Morfoespécie 1
0 0,2 0,4 0,6 0,8
1
Morfoespécie 2
0 0,2 0,4 0,6 0,8
1
Piper aduncum
0 0,2 0,4 0,6 0,8
1
Ficus obtusifolia
0 0,2 0,4 0,6 0,8
1
Cecropia pachystachya
0 0,2 0,4 0,6 0,8
1
Ficus insipida
0 0,2 0,4 0,6 0,8
1
Ficus crocata
0 0,2 0,4 0,6 0,8
1
Piper angustifolium
0 0,2 0,4 0,6 0,8
1
Alit Gsor Pdis Nalb Apla Plin Lsil Phas Slil Cper
Piper tuberculatum
0 0,2 0,4 0,6 0,8
1
-1,5 -1,0 -0,5 0,0 0,5 1,0 1,5
NMDS
M2 M1 Padu
Cpac
Fcroc Fins
Ptub Pang
Maclura tinctoria
0 0,2 0,4 0,6 0,8
1
Espécies de plantas
B
Proporção de am
ostras fecais
Mtin
Fobt
A
21
Figura 2: Variação sazonal da proporção de sementes nas amostras fecais dos morcegos no Pantanal da Nhecolândia, Mato Grosso do Sul.
Meses
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 F. insipida
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 C. pachystachya
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 F. obtusifolia
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 M. tinctoria
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 P. aduncum
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 P. angustifolium
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 P. tuberculatum
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0
ABR MAI JUN JUL AGO SET OUT NOV DEZ JAN FEV MAR
F. crocata
Seco Úmido
Proporção de amostras fecais

Figura 3: A: similaridade entre plantas quanto aos morcegos vetores de pólen (NMDS uma dimensão, stress 11% e variância recuperada de 88%). B: proporções de espécies de pólen encontrados nas amostras fecais dos morcegos no Pantanal da Nhecolândia, Mato Grosso do Sul C. bra = Caryocar brasiliense, I. ver = Inga vera, H. stil = Hymenaea stigonocarpa, B. ung = Bauhinia ungulata, H cou = Hymenaea courbaril, P. lon = Pseudobombax longiflorum, Bau sp. = Bauhinia sp., P. cor = Psittacanthus corynocephalus.
Psittacanthus corynocephalus
0 0,2 0,4 0,6 0,8
1
Lbra Apla Phas Pdis Nalb Caur Alit Lsil Plin Slil Gsor Cper
Hymenaea courbaril
0 0,2 0,4 0,6 0,8
1
Inga vera
0 0,2 0,4 0,6 0,8
1
Caryocar brasiliense
0 0,2 0,4 0,6 0,8
1
Hymenaea stigonocarpa
0 0,2 0,4 0,6 0,8
1
Morfoespécie 1
0 0,2 0,4 0,6 0,8
1
Bauhinia ungulata
0 0,2 0,4 0,6 0,8
1
Bauhinia sp.
0 0,2 0,4 0,6 0,8
1
-0,8 -0,6 -0,4 -0,2 0 0,2 0,4 0,6
NMDS
Cbra Iver
Hist Bung M1 Hcou Plon Bau sp. Pcor
Pseudobombax longiflorum
0 0,2 0,4 0,6 0,8
1
Espécies de morcegos
Proporção de amostras fecais
A
B

Figura 4: Variação sazonal da proporção de pólen nas amostras fecais dos morcegos no Pantanal da Nhecolândia, Mato Grosso do Sul.
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 H. stigonocarpa
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 I. vera
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 P. corynocephalus
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 H. courbaril
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 P. longiflorum
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 Bauhinia sp.
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 B. ungulata
C. brasiliense
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0
ABR MAI JUN JUL AGO SET OUT NOV DEZ JAN FEV MAR
Úmido
Meses
Proporção de amostras fecais
Seco
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo


















