
Morfologia de serpentes aquáticas neotropicais: um estudo...
103
RODRIGO ROVERI SCARTOZZONI Morfologia de serpentes aquáticas neotropicais: um estudo comparativo São Paulo 2005
Transcript of Morfologia de serpentes aquáticas neotropicais: um estudo...

RODRIGO ROVERI SCARTOZZONI
Morfologia de serpentes aquáticas neotropicais: um estudo comparativo
São Paulo
2005

RODRIGO ROVERI SCARTOZZONI
Morfologia de serpentes aquáticas neotropicais: um estudo comparativo
Dissertação apresentada ao Instituto de Biociências da Universidade de São Paulo, para a obtenção de Título de Mestre em Ciências, na área de Ecologia.
Orientador: Prof. Dr. Marcio Roberto Costa Martins
São Paulo
2005

FICHA CATALOGRÁFICA
Scartozzoni, Rodrigo Roveri R 287m Morfologia de serpentes aquáticas neotropicais: um estudo comparativo / Rodrigo Roveri Scartozzoni. -- São Paulo: R. R. Scartozzoni, 2005. p. 102: il. Dissertação (Mestrado) – Universidade de São Paulo, Instituto de Biociências, Depto. de Ecologia, 2005.
1. Serpentes - Morfologia 2. Ambiente - Uso I. Universidade de São Paulo. Instituto de Biociências. Departamento de Ecologia. II. Título. LC : QL 666.o6

FOLHA DE APROVAÇÃO
SCARTOZZONI, Rodrigo Roveri
Morfologia de serpentes aquáticas neotropicais: um estudo comparativo
Dissertação apresentada ao Instituto de Biociências da Universidade de São Paulo, para a obtenção de Título de Mestre em Ciências, na área de Ecologia.
Aprovado em:
Banca Examinadora Prof. Dr. ___________________________________________________________ Instituição: ________________________ Assinatura: _________________________ Prof. Dr. ___________________________________________________________ Instituição: ________________________ Assinatura: _________________________ Prof. Dr. ___________________________________________________________ Instituição: ________________________ Assinatura: _________________________ Prof. Dr. ___________________________________________________________ Instituição: ________________________ Assinatura: _________________________ Prof. Dr. ___________________________________________________________ Instituição: ________________________ Assinatura: _________________________

Dedico este trabalho aos meus pais, Augusto e Cecília, pelo amor, dedicação
e apoio necessários durante todos esses anos.

AGRADECIMENTOS Ao Marcio Martins, pela orientação e por ter confiado em minha capacidade de trabalho.
À Fundação de Amparo a Pesquisa no Estado de São Paulo (FAPESP), pelo financiamento deste trabalho por meio da concessão de bolsa de estudo.
Ao Otavio Marques, pela co-orientação, amizade, encorajamento constante e exemplo profissional.
À Selma Maria de Almeida-Santos, pela amizade e por me abrir os olhos à ciência.
À Maria da Graça Salomão, pela amizade e apoio constantes.
Ao Ricardo Sawaya, pelas críticas e sugestões às versões preliminares da dissertação e pelas discussões a cerca de análises comparativas e estatísticas.
Ao Hebert Ferrarezzi, pelas críticas ao longo do trabalho e pelas várias discussões a cerca de sistemática filogenética.
Ao Wilson Fernandes e Francisco Luis Franco, pela permissão para utilização da coleção do Instituto Butantan, bem como das dependências do Laboratório de Herpetologia.
Ao Hussam Zaher, Ronaldo Fernandes e Júlio César de Moura-Leite, por permitirem o exame de exemplares preservados sobre seus cuidados.
Ao Valdir José Germano, pela amizade e apoio constante ao trabalho de coleção.
A Maria Inês Conte, bibliotecária do Instituto de Biociências da USP, pela revisão da bibliografia deste trabalho.
Ao Antonio Carlos Ribeiro da Costa, pelos desenhos desta dissertação.
A todos os colegas pesquisadores que gentilmente forneceram informações sobre atividade de serpentes na natureza: Miguel Trefaut Rodrigues, Ricardo Sawaya, Otavio Marques, Marcio Martins, Cristiano Nogueira, Donizete Pereira, Ricardo Kawashita Ribeiro, Marcos Di-Bernardo, Christine Strüssmann, Hebert Ferrarezzi, Giuseppe Puorto, Francisco Luis Franco e Renato Moraes.
A todos os colegas do Laboratório de Herpetologia, em especial a Darina, Dona Maria, Dona Vera, Mariza, Antonio Carlos Ribeiro da Costa, Einat, Alessandra, Adriana, Lílian, Myriam, Ricardo, Herbert, Lígia, Leonardo, Karina, Maryanne, Einat, Cláudio, Joãozinho, Murilo, Fátima, Fausto, Renato, Kiko, Cristina, Fernanda, Marcelo, Antonio Carlos, Zé Pedro, Eladio, Amauri, Marlene, Carlinhos, Wilson, Fátima Furtado, Silvia, Mariza...
A todos os colegas do LabVert, em especial ao Beto, Tozetti, Cínthia, Cristiano, Elaine e Carlos.
À Rita, pela paciência, cooperação e amor.
Ao meu irmão Adriano, pela amizade e apoio constante.
E finalmente, minha gratidão a todos os que, direta ou indiretamente, contribuíram para a realização deste trabalho.

“Ninguém ignora tudo, ninguém sabe tudo. Por isso, aprenderemos
sempre”.
Paulo Freire

RESUMO SCARTOZZONI, R. R. Morfologia de serpentes aquáticas neotropicais: um estudo comparativo. 2005. 102 f. Dissertação (Mestrado) – Instituto de Biociências. Universidade de São Paulo, São Paulo, 2005. O tipo de ambiente utilizado influencia fortemente a evolução da morfologia em vertebrados. Este estudo verifica a relação entre a morfologia e o uso do ambiente aquático em 37 espécies de serpentes neotropicais pertencentes às subfamílias Boinae, Elapinae e Xenodontinae. A variação da forma relativa do corpo, dentro de cada subfamília, foi verificada por análise de componentes principais (ACP). Diferenças no tamanho e forma do corpo foram testadas, dentro de cada subfamília, por análises de variância e covariância. As informações de uso do ambiente, bem como as variáveis de tamanho e forma relativa do corpo foram otimizadas em hipóteses filogenéticas das subfamílias e as relações entre elas analisadas por meio de contrastes independentes. Serpentes aquáticas das diferentes linhagens tenderam a possuir os olhos e as narinas posicionadas na região dorso-anterior da cabeça. Também apresentaram redução na largura da escama ventral e são espécies mais robustas em relação a serpentes que utilizam outros ambientes. Serpentes semi-aquáticas do gênero Hydrodynastes e algumas espécies aquáticas do gênero Liophis não apresentaram tais modificações, provavelmente devido a restrições filogenéticas. Entre os Xenodontinae, depois de eliminado o efeito da filogenia sobre as variáveis, correlações positivas e significativas foram encontradas entre o uso do ambiente e forma do corpo. Reconstruções dos estados ancestrais indicam que os ancestrais hipotéticos das serpentes analisadas eram terrícolas (Boinae e Xenodontinae) ou fossóreos (Elapinae) e não apresentavam as modificações associadas ao hábito aquático. O comprimento do corpo e o tamanho da cabeça não parecem estar relacionados ao uso do ambiente aquático, mas sim a outros fatores ecológicos (e.g., alimentação). O comprimento da cauda e o diâmetro do olho parecem estar relacionados ao uso de outros ambientes (e.g., arborícola ou fossóreo) ou a táticas defensivas, como no caso do comprimento da cauda. Nas espécies estudadas de Xenodontinae, o hábito aquático não parece ter levado a modificações na fecundidade e espaço ocupado pela ninhada na cavidade abdominal das fêmeas, diferentemente do observado para outras serpentes. Palavras-Chave: Morfologia. Serpentes. Uso do hábitat.

ABSTRACT SCARTOZZONI, R. R. Morphology of aquatic Neotropical snakes: a comparative study. 2005. 102 f. Dissertação (Mestrado) – Instituto de Biociências. Universidade de São Paulo, São Paulo, 2005.
The use of habitat is thought to strongly influence morphological evolution in vertebrates. We explored the relationships of morphology with the use of aquatic habitats in 37 species of Neotropical snakes belonging to the Boinae, Elapinae and Xenodontinae. Variation in body form within each subfamily was explored by principal components analysis (PCA). Differences in body size and body form were tested within each subfamily by variance and covariance analyses. The information on habitat use, as well as the body size and body form variables were optimized in a phylogenetic hypothesis of the subfamilies, and relationships among them were explored by independent contrasts. Aquatic snakes in different lineages tend to posses the eyes and nostrils displaced towards the anterodorsal region of the head. They also present reduction in the width of ventral scales, and are stouter than species that use other habitats. Semi-aquatic snakes of the genus Hydrodynastes and some aquatic species of the genus Liophis do not present these trends, probably because of phylogenetic constraints. Within the Xenodontinae, after removing the phylogenetic effects of the variables, significantly and positive correlations were found between habitat use and body form. Reconstruction of character states indicated that the ancestors of the each subfamily were terrestrial (Boinae and Xenodontinae) or fossorial (Elapinae) and did not present the morphological changes associated with aquatic habits. Body size and head form seem not to have any relationships with aquatic habits, but to others ecological features (e.g., feeding). Tail length and the eye diameter seem to be related to others habits (e.g., arboreal or fossorial) or to defensive tactics. In the xenodontines studied here, aquatic habits seem not to have lead to changes in fecundity and clutch position in the abdominal cavity of females, as observed in other snake lineages. Keywords: Morphology. Snakes. Habitat use.

SUMÁRIO
1 INTRODUÇÃO 10-13
2 OBJETIVOS 14
3 MATERIAIS E MÉTODOS 15
3.1 Caracterização do uso do ambiente 15-16
3.2 Dados Biométricos 16
3.3 Análise dos dados 17
3.3.1 Análises estatísticas 17-18
3.3.2 Análises comparativas 19
3.3.2.1 Otimização de caracteres 19
3.3.2.2 Contrastes independentes 19-20
3.3.2.3 Hipóteses filogenéticas 20-22
4 RESULTADOS 23
4.1 Uso do ambiente 23-24
4.2 Biometria 25
4.2.1 Tamanho do corpo 25-27
4.2.2 Forma do corpo 28-57
4.2.3 Posição e espaço ocupado pela ninhada e fecundidade 58-63
4.3 Análises comparativas 64-79
5 DISCUSSÃO 80-88
6 CONCLUSÕES 89-90
REFERÊNCIAS BIBLIOGRÁFICAS 91-98
APÊNDICE 99-102

10
1 - INTRODUÇÃO
A subordem Serpentes agrupa cerca de 2.700 espécies, distribuídas em
aproximadamente 18 famílias (GREENE, 1997; POUGH et al., 1998). É um dos grupos de maior
sucesso dentre os vertebrados. Uma evidência desse sucesso é a variedade de hábitats
ocupados pelas espécies, que apenas não ocorrem nas regiões polares devido às restrições
impostas pelo clima (GREENE, 1997). As serpentes também podem exibir grande diversidade
quanto ao tipo de ambiente utilizado (macrohábitat, cf. CADLE; GREENE, 1993). Assim, as
espécies podem ser terrícolas, arborícolas, fossóreas, criptozóicas ou aquáticas, porém muitas
serpentes podem utilizar mais que um ambiente quando ativas (CADLE; GREENE, 1993;
MARTINS, 1993; MARTINS; OLIVEIRA, 1999).
O uso do ambiente e suas implicações físicas podem influenciar fortemente a evolução
da forma do corpo em diversos grupos de vertebrados (ver KOHLSDORF; GARLAND; NAVAS,
2001; LOSOS, 1990; MACRINI; IRSCHICK; LOSOS, 2003; MILES; RICKLEFS, 1984; MOERMOND,
1979; WIKRAMANAYAKE, 1990). Síndromes morfológicas relacionadas ao uso de
determinados ambientes também evoluíram em diferentes linhagens de serpentes (ver CADLE;
GREENE, 1993; MARTINS et al., 2001). Espécies terrícolas possuem tamanho de corpo variado
e morfologia generalizada, porém modificações importantes ocorreram em serpentes que
utilizam outros ambientes (CADLE; GREENE, 1993).
Serpentes de hábitos subterrâneos (fossóreas) geralmente apresentam modificações
relacionadas ao decréscimo da fricção e que permitem a escavação de túneis em solo
compacto (GREENE, 1997). Essas espécies tendem a apresentar cabeças menores e compactas,
quase indistintas do resto do corpo, fusão e/ou redução do número de escamas da cabeça,
olhos pequenos e caudas curtas (MARX; RABB, 1972; SAVITZKY, 1983).
O ambiente arbóreo também pode impor restrições às serpentes, principalmente
devido à irregularidade e descontinuidade do substrato (LILLYWHITE; HENDERSON, 1993).

11
Serpentes arborícolas tendem a apresentar menor robustez, corpo delgado e comprimido
lateralmente, caudas compridas (muitas vezes preênsil), olhos grandes e o centro de gravidade
deslocado para a região posterior do corpo (LILLYWHITE; HENDERSON, 1993; MARTINS et al.,
2001). A coloração verde também é freqüente entre essas espécies (LILLYWHITE; HENDERSON,
1993).
Muitas serpentes são especializadas ao uso do ambiente aquático, inclusive o marinho
(GREENE, 1997). Essas espécies geralmente possuem os olhos e as narinas posicionadas no
dorso da cabeça, bem como válvulas nasais que impedem a entrada de água quando
submersas (CADLE; GREENE, 1993; GREENE, 1997). Muitos elapídeos marinhos e alguns
colubrídeos aquáticos possuem a cauda achatada lateralmente (em algumas espécies o corpo
também é achatado), em forma de remo, para facilitar a natação (GREENE, 1997). O
investimento reprodutivo em algumas serpentes aquáticas também parece ser preterido em
favor da locomoção, pois suas ninhadas tendem a ser menores, ocupando posição mais
anterior no corpo da fêmea em comparação a espécies terrícolas (SHINE, 1988).
Entretanto, mudanças morfológicas associadas ao uso do ambiente, particularmente
entre as serpentes da região neotropical, ainda são pouco conhecidas (CADLE; GREENE, 1993).
Além disso, essas mudanças são geralmente interpretadas sem que sejam consideradas as
relações filogenéticas entre espécies e linhagens abordadas (e.g., GUYER; DONNELLY, 1990;
KAUFMAN; GIBBONS, 1975; VITT; VANGILDER, 1983).
A interpretação da evolução do uso do ambiente e da variação morfológica à luz do
conhecimento filogenético é passo importante para o melhor entendimento das relações
adaptativas entre uso do ambiente e forma do corpo (BROOKS; MCLENNAN, 1991, 2002;
HARVEY; PAGEL, 1991). Desta forma, é possível a interpretação de características
morfológicas similares em espécies de diferentes linhagens, mas que ocupam o mesmo tipo de
ambiente, como possíveis evidências de adaptação (sensu CODDINGTON, 1988) ao tipo de

12
ambiente utilizado. Por outro lado, características morfológicas similares entre espécies
próximas na filogenia, mas que ocupam ambientes distintos, podem ser interpretadas como
resultado de restrições filogenéticas e/ou independência do caráter em relação às pressões
ambientais (cf. BROOKS; MCLENNAN, 1991, 2002; HARVEY; PAGEL, 1991).
A região neotropical apresenta uma fauna de serpentes extremamente diversificada,
com representantes de várias famílias. A família Boidae, basal entre as serpentes
macrostomata, inclui as subfamílias Erycinae e Boinae (KLUGE, 1991). A subfamília Boinae
possui cinco gêneros, sendo que quatro ocorrem na região neotropical (KLUGE, 1991): Boa,
Corallus, Epicrates e Eunectes. Embora não seja grupo muito diverso, seus integrantes
apresentam grande variação morfológica e ecológica, incluindo o tipo de ambiente utilizado
(ver GREENE, 1997; HENDERSON et al., 1995; MURPHY; HENDERSON, 1997).
A família Colubridae é a mais diversificada entre os Colubroidea, que inclui outras
três famílias (Elapidae, Viperidae e Atractaspididae; FERRAREZZI, 1994). Quatro subfamílias
de colubrídeos ocorrem na região neotropical: Natricinae, Colubrinae, Dipsadinae e
Xenodontinae (FERRAREZZI, 1994). O grande número de espécies, bem como a grande
diversidade morfológica e ecológica de seus integrantes, são características marcantes de
Xenodontinae (ver CADLE; GREENE, 1993; MARQUES; SAZIMA, 2004; MARTINS; OLIVEIRA,
1999).
A família Elapidae inclui cinco subfamílias. A subfamília Elapinae apresenta cinco
gêneros dos quais apenas dois (Micrurus e Micruroides) são exclusivos na região neotropical
(SLOWINSKI, 1995). O gênero Micrurus, embora seja composto por grande número de
espécies (ver CAMPBELL; LAMAR, 2004), apresenta pouca variação quanto ao uso do
ambiente; a maioria das espécies é terrícola e/ou fossórea e apenas um representante (M.
surinamensis) é aquático (ROZE, 1996).

13
A grande diversidade ecológica e morfológica das serpentes neotropicais, aliada ao
conhecimento de suas relações filogenéticas (e.g., FERRAREZZI, 1993; KLUGE, 1991; PINOU et
al., 2004; SLOWINSKI, 1995; VIDAL et al., 2000), representam grande potencial para o estudo
das relações adaptativas entre uso do ambiente e morfologia. No presente trabalho foram
estudadas 37 espécies de serpentes neotropicais, pertencentes às subfamílias Boinae, Elapinae
e Xenodontinae. Esses grupos foram escolhidos por apresentarem espécies aquáticas e/ou
semi-aquáticas e por terem suas relações filogenéticas bem estabelecidas. Buscou-se
descrever e comparar o uso do ambiente e a morfologia dessas espécies a partir de exemplares
preservados em coleções e de informações obtidas na literatura ou cedidas por outros
pesquisadores.

14
2 - OBJETIVOS
O presente estudo teve como objetivos:
1 - verificar a existência de diferenças quanto à freqüência de utilização do ambiente
aquático entre as espécies, dentro de cada grupo analisado.
2 - verificar a existência de diferenças na morfologia (tamanho e forma do corpo)
entre as espécies, dentro de cada grupo analisado.
3 - verificar se a evolução no uso do ambiente aquático foi acompanhada por
modificações na morfologia das serpentes analisadas.
4 - verificar se a evolução no uso do ambiente aquático foi acompanhada por
modificações na fecundidade e posição da ninhada nas fêmeas analisadas.
5 - desvendar possíveis agentes seletivos que levaram a modificações na morfologia,
fecundidade e posição da ninhada nas serpentes aquáticas analisadas.

15
3 - MATERIAIS E MÉTODOS
Para analisar a influência do uso do ambiente no tamanho e forma do corpo em
serpentes neotropicais, foram examinados 1.056 indivíduos pertencentes a 37 espécies das
subfamílias Boinae, Elapinae e Xenodontinae (ver Apêndice). Dados biométricos foram
obtidos de espécimes preservados das coleções do Instituto Butantan (IB), Museu de Zoologia
da Universidade de São Paulo (MZUSP), Museu de História Natural Capão da Imbuia
(MHNCI), Museu Nacional do Rio de Janeiro (MN).
3.1 Caracterização do uso do ambiente
As informações de uso do ambiente para as espécies analisadas foram cedidas por
outros pesquisadores ou obtidas de trabalhos sobre ecologia e história natural de serpentes
(e.g., BEEBE, 1946; BERNARDE; KOKUBUM; MARQUES, 2000a; BIZERRA, 1998; CUNHA;
NASCIMENTO; ÁVILA-PIRES, 1985; DIXON; SOINI, 1977; DUELLMAN, 1978; FORD; FORD,
2002; GOMES, 1918; JORGE DA SILVA, 1993; LEMA; DEIQUES, 1992; MARQUES, 1998;
MARQUES; SAZIMA, 2004; MARQUES; SOUZA, 1992; MARTINS, 1994; MARTINS; OLIVEIRA,
1999; PLATT, 1969; PORTO; CARAMASCHI, 1988; SAWAYA, 2004; SAZIMA; ABE, 1991; SAZIMA;
MARTINS, 1990; SAZIMA; STRÜSSMANN, 1990; STRÜSSMANN, 1992; VANZOLINI, 1986; VITT,
1996; ZIMMERMAN; RODRIGUES, 1990).
Espécies em que mais de 50% dos indivíduos encontrados na natureza estavam dentro
de corpos d’água foram consideradas aquáticas. Aquelas em que 20%-50% dos indivíduos
estavam dentro d’água foram classificadas como semi-aquáticas (ver procedimento
semelhante em MARTINS et al., 2001). O número de indivíduos de uma espécie encontrados
nas margens de corpos d’água foi dividido por dois e acrescentado ao número de indivíduos
encontrados dentro e fora d’água no cálculo das porcentagens acima citadas. Esse

16
procedimento foi adotado considerando a probabilidade de 50% de chances de o indivíduo
entrar na água.
3.2 Dados Biométricos
Para cada indivíduo foram obtidas as seguintes medidas: (1) comprimento rostro-
cloacal (CRC); (2) distância entre os olhos (IO); (3) distância entre o rostro e os olhos (RO);
(4) distância entre as narinas (IN); (5) distância entre o rostro e as narinas (RN); (6) diâmetro
dos olhos; (7) comprimento da cabeça (Ccab); (8) largura da cabeça (Lcab); (9) altura da
cabeça (Acab); (10) comprimento da cauda (CC); (11) massa do corpo (M); (12)
circunferência do corpo (Circ); e (13) largura da escama ventral (Lev).
Dados de fecundidade, posição e espaço ocupado pela ninhada no corpo das fêmeas
(cf. SHINE, 1988) também foram obtidos. Nas fêmeas com ovos ou embriões nos ovidutos
foram medidos: (14) comprimento anterior à ninhada (CAN, distância entre o rostro e o
primeiro embrião ou ovo); (15) comprimento da ninhada (CN, extensão da ninhada no ventre
da fêmea); (16) comprimento posterior à ninhada (CPN, distância entre o último ovo ou
embrião e a cloaca); (17) fecundidade (número de ovos ou embriões nos ovidutos).
Os comprimentos rostro-cloacal, da cauda e as medidas de posição e espaço ocupado
pela ninhada foram obtidas utilizando régua milimetrada com precisão de 0,5 mm. As
medidas do posicionamento dos olhos e narinas, o diâmetro dos olhos, o comprimento, a
largura e a altura da cabeça foram obtidas com auxílio de paquímetro digital com precisão de
0,1 mm. Para obter a massa do corpo foram usados dinamômetros portáteis Pesola® com
precisão de 0,1 ou 0,5 g. A circunferência do corpo e a largura da escama ventral foram
obtidas na região mediana do CRC, com auxílio de fita métrica ou fio flexível (cf. POUGH;
GROVES, 1983), com precisão de 0.5 mm.

17
3.3 Análise dos dados
Todas as amostras foram constituídas de indivíduos adultos, para evitar problemas
relacionados a mudanças ontogenéticas na forma do corpo (ver KING, 1999). Esse
procedimento não foi feito para os Boinae, pois espécimes adultos são raros em coleções.
Machos de serpentes tendem a apresentar caudas maiores que fêmeas, devido principalmente
à presença do hemipênis e de seus músculos retratores (ver KING, 1989; SHINE, 1994a). Dessa
forma, a inclusão de machos pode introduzir ruídos nas análises de comprimento da cauda e
massa do corpo, já que a massa de uma serpente também inclui a massa de sua cauda (cf.
MARTINS et al., 2001). Portanto, foram utilizadas apenas fêmeas nas análises dessas variáveis.
Nas análises de massa e circunferência do corpo, foram excluídas as fêmeas com folículos
vitelogênicos, ovos ou embriões. A massa do corpo também foi tomada após drenagem do
excesso do líquido preservativo e retirada de presas do estômago, quando presentes (cf.
MARTINS et al., 2001).
3.3.1 Análises estatísticas
A variação da forma do corpo entre espécies, dentro de cada grupo analisado, foi
verificada por análise de componentes principais (ACP; MANLY, 1994) utilizando o programa
MVSP versão 3.1 (KOVACH, 1999). Nessa análise foram usados os resíduos das regressões
lineares entre as variáveis em estudo (dependentes) e as variáveis estimadoras do tamanho do
corpo (independentes), incluindo todas as espécies. Os resíduos foram usados para eliminar a
influência do tamanho do corpo sobre as variáveis estudadas (cf. LOSOS, 1990; MACRINI;
IRSCHICK; LOSOS, 2003; MARTINS et al., 2001), as quais passaram a ser consideradas variáveis
relativas da forma do corpo.
As distâncias relativas entre as narinas, entre os olhos, entre o rostro e as narinas, entre
o rostro e os olhos, e o diâmetro relativo do olho, foram obtidas a partir das regressões com o

18
Ccab. O comprimento, largura e altura relativa da cabeça foram obtidos a partir das regressões
com o comprimento do tronco (TR = CRC–Ccab). A massa relativa do corpo e a largura
relativa da escama ventral foram obtidas a partir das regressões com o comprimento total (CT
= CRC+CC) e circunferência do corpo, respectivamente. As variáveis relativas de
comprimento da cauda e circunferência do corpo foram obtidas a partir das regressões com o
CRC.
Diferenças no tamanho do corpo (CRC) foram testadas, entre espécies e entre aquelas
de diferentes hábitos, dentro de cada grupo analisado, por análise de variância. Diferenças na
forma do corpo foram testadas (a partir dos dados brutos) entre espécies e entre aquelas de
diferentes hábitos, dentro de cada grupo analisado, por análise de covariância. Nessa análise,
para as distâncias entre as narinas, entre os olhos, entre o rostro e as narinas, entre o rostro e
os olhos, e para o diâmetro do olho, a covariável foi o Ccab; o comprimento, largura e altura
da cabeça tiveram como covariável o TR; a massa do corpo teve como covariável o CT; para
a largura da escama ventral, a covariável foi o Circ; o comprimento da cauda e a
circunferência do corpo tiveram como covariável o CRC.
As razões entre as medidas de posição e espaço ocupado pela ninhada (ver acima) e o
CRC das fêmeas foram calculadas para permitir comparações com informações disponíveis na
literatura (ver SHINE, 1988). Diferenças nessas variáveis foram testadas (a partir dos dados
brutos, incluindo fecundidade) entre espécies de diferentes hábitos, dentro de cada grupo
analisado, por análise de covariância. Essas medidas tiveram como covariável o CRC.
Nas comparações múltiplas, foram realizados testes post-hoc para detectar quais
grupos diferiam entre si. Todas as análises foram baseadas em Zar (1999) e executadas no
programa Statistica (STATSOFT, 1998). Os valores de todas as variáveis foram previamente
transformados para o seu logaritmo natural (de forma a linearizar as relações e diminuir as
variâncias) e as diferenças consideradas significativas quando P < 0,05.

19
3.3.2 Análises comparativas
Nas análises comparativas descritas a seguir, o menor valor médio obtido para os
resíduos calculados de cada variável (ver procedimento para o cálculo dos resíduos acima) foi
acrescentado aos valores médios de resíduos calculados para cada espécie. Esse procedimento
foi adotado para evitar valores médios de resíduos negativos nas otimizações de caracteres e
cálculo dos contrastes filogenéticos.
3.3.2.1 Otimização de caracteres
As informações de uso do ambiente (freqüência de uso do ambiente aquático),
tamanho e forma relativa do corpo (resíduos das regressões) foram otimizadas em hipóteses
filogenéticas (cf. FARRIS, 1970; ver figuras 1, 2 e 3), por parcimônia linear, no programa
Mesquite (MADDISON; MADDISON, 2004). Na otimização, as topologias dos cladogramas
foram usadas como variáveis independentes, a partir das quais foram descritas a seqüência e
direção das transformações dos caracteres e detectados os estados ancestrais nos ramos
intermediários de cada linhagem (cf. BROOKS; MCLENNAN, 1991, 2002). A partir da
otimização de caracteres é possível distinguir similaridades decorrentes de convergências
adaptativas daquelas devidas a restrições filogenéticas (BROOKS; MCLENNAN, 1991, 2002).
3.3.2.2 Contrastes independentes
As relações entre uso do ambiente, tamanho e forma relativa do corpo, também foram
exploradas com o uso de contrastes independentes (cf. FELSENSTEIN, 1985), gerados no
módulo PDAP (MIDFORD; GARLAND; MADDISON, 2003) para o programa Mesquite. O cálculo
dos contrastes elimina o efeito da filogenia sobre as variáveis (DINIZ-FILHO, 2000). Os
contrastes para espécies irmãs e ramos intermediários na filogenia (ver FELSENSTEIN, 1985)
podem ser considerados pontos independentes em termos estatísticos e utilizados para analisar

20
a correlação evolutiva intrínseca entre caracteres quantitativos (MARTINS; GARLAND, 1991). A
relação entre os contrastes de uso do ambiente e aqueles do tamanho e forma relativa do corpo
foram analisados utilizando correlações lineares de Pearson. Para o cálculo dos contrastes,
todos os ramos da filogenia foram ajustados para o valor 1 (cf. GARLAND; HARVEY; IVES,
1992).
Os contrastes independentes foram calculados apenas para as espécies (e seus
ancestrais) de Xenodontinae devido o tamanho amostral permitir uma correlação correta (ver
DINIZ-FILHO, 2000).
3.3.2.3 Hipóteses filogenéticas
As hipóteses filogenéticas utilizadas para interpretar e discutir a evolução do uso do
ambiente, tamanho e forma relativa do corpo nas serpentes estudadas são derivadas das
análises cladísticas mais detalhadas e completas para essas espécies. As relações entre os
Boinae neotropicais (Figura 1) foram adaptadas de Kluge (1991), um dos estudos mais
abrangentes sobre a filogenia deste grupo, com dados morfológicos. As relações entre as
espécies de Elapinae (Figura 2) foram adaptadas de Jorge da Silva e Sites (2001), com dados
moleculares, e Slowinski (1995), baseado em dados morfológicos e imunológicos.
As relações entre grupos supragenéricos de Xenodontinae (Figura 3) foram adaptadas
de Ferrarezzi (1993), com dados morfológicos, as quais são congruentes com o estudo de
Vidal et al. (2000), baseado em dados moleculares. As relações entre gêneros e espécies de
Hydropsini foram adaptadas de estudo cladístico preliminar, baseado na morfologia do
hemipênis (P. M. S. NUNES, informação pessoal). O monofiletismo dos gêneros desta tribo é
corroborado por Zaher (1999), também com dados morfológicos.
O monofiletismo de Tachymenini é corroborado por Ferrarezzi (1994) e Franco
(1999), baseados em dados morfológicos, e por Pinou et al. (2004) e Vidal et al. (2000), com

21
dados moleculares. As relações entre os gêneros e espécies dessa tribo foram adaptadas de
Franco (1999). As relações entre os representantes de Pseudoboini analisados foram
adaptadas dos estudos de Pinou et al. (2004), Vidal et al. (2000) e Zaher (1994). A posição do
gênero Hydrodynastes como grupo irmão dos Pseudoboini também é corroborada por Pinou
et al. (2004), Vidal et al. (2000) e Zaher (1994 e 1999). Não foi possível a quantificação do
uso do ambiente para Hydrodynastes bicinctus e, por isso, esta espécie não foi incluída no
cladograma utilizado nas análises comparativas.
Os Xenodontini também constituem um grupo monofilético (FERRAREZZI, 1994;
MOURA-LEITE, 2001; PINOU et al., 2004; VIDAL et al., 2000). As relações entre os
representantes dessa tribo foram adaptadas do estudo de Moura-Leite (2001), baseado em
dados morfológicos.
A condição de Sordellina punctata como integrante de Hydropsini (GANDOLFI;
BARBIERI, 1996) e a posição dos Elapomorphini como grupo irmão dessas serpentes ainda não
estão bem definidas (cf. FRANCO, 1999). Além disso, Pinou et al. (2004) e Vidal et al. (2000)
não incluem essas espécies em suas análises. Por isso, S. punctata, Elapomorphus
quinquelineatus, Phalotris mertensi e Apostolepis assimilis não foram incluídas no
cladograma utilizado nas análises comparativas.
Os estudos de Cadle (1984a, b, 1985, 1988), Kelly, Barker e Villet (2003), Pinou et al.
(2004) e Vidal et al. (2000) suportam a posição dos Dipsadinae (xenodontíneos centro-
americanos, sensu CADLE, 1984b) como grupo irmão dos Xenodontinae (xenodontíneos sul-
americanos, sensu CADLE, 1984a). De acordo com esses autores, os xenodontíneos norte-
americanos (aqui representado por Heterodon platyrhinus) compõem o grupo irmão do clado
formado por Dipsadinae + Xenodontinae (sul-americanos). Com isso, Heterodon platyrhinus
(xenodontíneo norte-americano) e Leptodeira annulata (Dipsadinae) foram utilizadas como
grupo externo nas otimizações em Xenodontinae.

22
Figura 1. Hipótese filogenética usada para interpretar e discutir a evolução do uso do ambiente, tamanho e forma relativa do corpo nas espécies de Boinae analisadas (ver procedimento para sua construção no texto).
Figura 2. Hipótese filogenética usada para interpretar e discutir a evolução do uso do ambiente, tamanho e forma relativa do corpo nas espécies de Elapinae analisadas (ver procedimento para sua construção no texto).
Figura 3. Hipótese filogenética usada para interpretar e discutir a evolução do uso do ambiente, tamanho e forma relativa do corpo nas espécies de Xenodontinae analisadas (ver procedimento para sua construção no texto).

23
4 - RESULTADOS
4.1 Uso do ambiente
As espécies analisadas diferiram em relação à freqüência e tipo de ambiente utilizado
(Tabela 1; ver critérios de classificação nos Métodos). Dentre as espécies que utilizam o
ambiente aquático com maior freqüência estão um Boinae (Eunectes murinus), um Elapinae
(Micrurus surinamensis) e 12 representantes de Xenodontinae (todos os Hydropsini, Liophis
miliaris, L. cobella, L. frenata, Gomesophis brasiliensis e Ptychophis flavovirgatus). Dentre
as semi-aquáticas estão os xenodontíneos Hydrodynastes gigas e Sordellina punctata. A única
informação disponível para Hydrodynastes bicinctus é a de um indivíduo encontrado dentro
d’água. Devido à informação ser baseada em um único indivíduo, essa espécie foi considerada
a priori como sendo semi-aquática. Não foi possível a quantificação do uso do ambiente para
Elapomorphus quinquelineatus.
Tabela 1. Proporção de indivíduos encontrados no ambiente aquático (PA) e ambiente predominantemente utilizado (AP) para as serpentes estudadas. O número de espécies está indicado entre parênteses. Abreviações: A = aquático; AR = arborícola; SA = semi-aquático; F = fossóreo; T = terrícola. Abreviações em letras minúsculas denotam menor importância dentro da categoria.
Grupos/Espécies PA AP Referências para uso do ambiente Boinae Boa constrictor 0,00 (13) T Cunha, Nascimento e Ávila-Pires (1985), Martins e
Oliveira (1999), Sawaya (2004), Strüssmann (1992) Epicrates cenchria 0,00 (27) T Beebe (1946), Cunha, Nascimento e Ávila-Pires (1985),
Duellman (1978), Zimmerman e Rodrigues (1990) Corallus hortulanus 0,00 (14) AR Marques (1998), Martins e Oliveira (1999) Eunectes murinus 0,66 (9) A Dixon e Soini (1986), Martins e Oliveira (1999)
Elapinae Micrurus corallinus 0,00 (21) F Marques (1998), observação pessoal M. frontalis 0,00 (6) F Sawaya (2004), Sazima e Abe (1991) M. lemniscatus 0,07 (27) F Dixon e Soini (1986), Duellman (1978), Martins e
Oliveira (1999), Sazima e Abe (1991), Vanzolini (1986) M. surinamensis 0,81 (11) A Duellman (1978), Martins e Oliveira (1999)

24
Xenodontinae Hydropsini Helicops modestus 0,94 (17) A Sawaya (2004), Sazima e Martins (1990), Sazima e
Strüssmann (1990), D. Pereira (informação pessoal) H. carinicaudus 0,70 (5) A Marques (1998), Marques e Sazima (2004) H. infrataeniatus 0,77 (26) A M. Di-Bernardo (informação pessoal) H. angulatus 0,99 (88) A Beebe (1946), Duellman (1978), Martins e Oliveira
(1999), Vanzolini (1986) H. polylepis 0,83 (6) A Dixon e Soini (1986), C. Strüssmann (informação
pessoal), C. Nogueira (informação pessoal) Hydrops triangularis 0,95 (19) A Beebe (1946), Dixon e Soini (1986), Ford e Ford (2002),
Martins e Oliveira (1999), Vanzolini (1986) Pseudoeryx plicatilis 0,75 (8) A Dixon e Soini (1986), C. Strüssmann (informação
pessoal), R. A. K. Ribeiro (informação pessoal), M. T. Rodrigues (informação pessoal)
Elapomorphini Elapomorphus quinquelineatus - - Phalotris mertensi 0,00 (7) F Sawaya (2004) Apostolepis assimilis 0,00 (5) F H. Ferrarezzi (informação pessoal), observação pessoal
Tachymenini Thamnodynastes strigatus 0,07 (30) T, ar Bernarde, Kokubum e Marques (2000), Bizerra (1998) Tomodon dorsatus 0,00 (45) T Bizerra (1998), Marques (1998) Gomesophis brasiliensis 0,66 (6) A Gomes (1918), M. T. Rodrigues (informação pessoal) Ptychophis flavovirgatus 0,82 (11) A Lema e Deiques (1992), Porto e Caramaschi (1988)
Xenodontini Liophis frenata 0,87 (8) A G. Puorto (informação pessoal), H. Ferrarezzi
(informação pessoal) L. cobella 0,71 (7) A Duellman (1978), Ford e Ford (2002), M. T. Rodrigues
(informação pessoal) L. miliaris 0,68 (19) A Marques (1998), Marques e Sazima (2004), Marques e
Souza (1992) L. poecilogyrus 0,00 (51) T Sawaya (2004), Strüssmann (1992) L. reginae 0,00 (28) T Duellman (1978), Martins e Oliveira (1999) Xenodon neuwiedii 0,00 (42) T Marques (1998) Erythrolamprus aesculapii 0,00 (31) T Dixon e Soini (1986), Duellman (1978), Marques (1998),
Sazima e Abe (1991), Zimmerman e Rodrigues (1990) Pseudoboini Oxyrhopus guibei 0,00 (50) T Sawaya (2004), Sazima e Abe (1990) Pseudoboa nigra 0,00 (7) T Strüssmann (1992) Clelia quimi 0,00 (3) T Franco, Marques e Puorto (1997)
Outros Xenodontinae Heterodon platirhinos 0,00 (11) T Platt (1969) Sordellina punctata 0,31 (13) SA D. Pereira (informação pessoal), R. A. Moraes
(informação pessoal) Hydrodynastes gigas 0,47 (46) SA Strüssmann (1992), O. A. V. Marques (informação
pessoal) H. bicinctus 1,00 (1) SA M. Martins (informação pessoal), M. T. Rodrigues
(informação pessoal)
Dipsadinae Leptodeira annulata 0,00 (36) T, ar Vitt (1996)
Continuação da Tabela 1.

25
4.2 Biometria
4.2.1 Tamanho do corpo
O tamanho médio do corpo foi variável (Tabela 2 e Figura 4) e significativamente
diferente entre espécies e entre aquelas com diferentes hábitos (uso do ambiente) dentro de
Elapinae e Xenodontinae (Tabela 3). Dentro de cada tribo de Xenodontinae, o tamanho do
corpo também foi significativamente diferente entre espécies e entre aquelas com diferentes
hábitos, exceto para representantes terrícolas e aquáticos de Xenodontini (Tabela 3). As
espécies de Boinae não foram incluídas nessas análises devido às amostras serem
constituídas de indivíduos imaturos.
Dentro de algumas linhagens, algumas serpentes aquáticas tenderam a apresentar
menor tamanho corporal (e.g., Elapinae e Tachymenini; Figura 4). Entretanto, dentre os
Hydropsini, embora as espécies de Helicops e Hydrops triangularis apresentem tamanho
pequeno ou médio, Pseudoeryx plicatilis está entre as maiores espécies analisadas. Entre os
representantes semi-aquáticos, as espécies de Hydrodynastes foram as maiores analisadas
dentro da amostra, enquanto que Sordellina punctata apresentou tamanho de corpo próximo
ao encontrado para as menores Helicops.

26
Tabela 2. Comprimento rostro-cloacal médio (CRCm) e desvio-padrão (DP) para as espécies de serpentes estudadas. O número de espécies está indicado entre parênteses. O CRC foi tomado em milímetros. * = serpentes imaturas.
Grupos/Espécies CRCm DP Boinae Boa constrictor* 741,4 (23) 304 Epicrates cenchria* 695,2 (28) 324 Corallus hortulanus 1399 (21) 258 Eunectes murinus* 1.178,4 (25) 371 Elapinae Micrurus corallinus 702,8 (22) 106 M. frontalis 777 (32) 119 M. lemniscatus 982 (35) 245 M. surinamensis 683 (24) 202 Xenodontinae Hydropsini Helicops modestus 401,3 (27) 57 H. carinicaudus 655,8 (34) 127 H. infrataeniatus 510,8 (29) 107 H. angulatus 484 (23) 74 H. polylepis 539,7 (17) 169 Hydrops triangularis 542,3 (20) 107 Pseudoeryx plicatilis 775,2 (13) 312 Elapomorphini Elapomorphus quinquelineatus 661 (29) 106 Phalotris mertensi 943 (29) 199 Apostolepis assimilis 518,9 (30) 73 Tachymenini Thamnodynastes strigatus 501 (33) 84 Tomodon dorsatus 491,2 (28) 64 Gomesophis brasiliensis 350,2 (40) 58 Ptychophis flavovirgatus 340,8 (28) 74 Xenodontini Liophis frenata 619,3 (31) 146 L. cobella 477,4 (30) 80 L. miliaris 659 (32) 151 L. poecilogyrus 489 (30) 95 L. reginae 527,1 (39) 79 Xenodon neuwiedii 566,5 (18) 55 Erythrolamprus aesculapii 739,2 (30) 90 Pseudoboini Oxyrhopus guibei 866,9 (34) 144 Pseudoboa nigra 882,1 (35) 127 Clelia quimi 809,3 (33) 149 Outros Xenodontinae Heterodon platirhinos 445 (14) 96 Sordellina punctata 414,9 (31) 89 Hydrodynastes gigas 1.346,4 (33) 276 H. bicinctus 1.113,7 (22) 262 Dipsadinae Leptodeira annulata 540,4 (29) 113

27
Figura 4. Variação do comprimento rostro-cloacal nas espécies analisadas (exceto representantes de Boinae). Linhas dentro das caixas = médias; caixas = desvios-padrão; barras verticais = valores mínimos e máximos. Acima de cada espécie é indicado o tipo de ambiente utilizado: A = aquático; SA = semi-aquático; F = fossóreo; T = terrícola. Abreviações: Hmod = Helicops modestus, Hinf = H. infrataeniatus, Hcar = H. carinicaudus, Hang = H. angulatus, Hpol = H. polylepis, Ppli = Pseudoeryx plicatilis, Hytr = Hydrops triangularis, Spun = Sordellina punctata, Aass = Apostolepis assimilis, Equi = Elapomorphus quinquelineatus, Pmer = Phalotris mertensi, Gbra = Gomesophis brasiliensis, Pfla = Ptychophis flavovirgatus, Tdor = Tomodon dorsatus, Tstr = Thamnodynastes strigatus, Lcob = Liophis cobella, Lfre = L. frenata, Lmil = L. miliaris, Lpoe = L. poecilogyrus, Lreg = L. reginae, Eaes = Erythrolamprus aesculapii, Xnew = Xenodon neuwiedii, Hydg = Hydrodynastes gigas, Hydb = H. bicinctus, Ogui = Oxyrhopus guibei, Pnig = Pseudoboa nigra, Cqui = Clelia quimi, Hpla = Heterodon platyrhinus, Lann = Leptodeira annulata, Msur = Micrurus surinamensis, Mlem = M. lemniscatus, Mfro = M. frontalis, Mcor = M. corallinus.
Tabela 3. Resultados da análise de variância para as comparações de tamanho do corpo (CRC) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Elapinae Espécies 3 90 15,01 << 0,01 Uso do ambiente 1 13,78 << 0,01 Xenodontinae Espécies 28 514 84,9 << 0,01 Uso do ambiente 3 63,9 << 0,01 Hydropsini Espécies 15 272 49,1 << 0,01 Uso do ambiente 3 66,4 << 0,01 Tachymenini Espécies 3 70 46,8 << 0,01 Uso do ambiente 1 140,6 << 0,01 Xenodontini Espécies 6 133 24,5 << 0,01 Uso do ambiente 1 0,1 = 0,75 Pseudoboini Espécies 4 97 35,9 << 0,01 Uso do ambiente 1 115,3 << 0,01
Pfla
Gbr
aH
mod
Spu
nH
pla
Lcob
Han
gLp
oeTd
orTs
trH
inf
Aas
sLr
egH
pol
Lann
Hyt
rX
new
Lfre
Hca
rLm
ilE
qui
Msu
rM
cor
Eae
sP
pli
Mfro
Cqu
iO
gui
Pni
gP
mer
Mle
mH
ydb
Hyd
g
Espécies
0
200
400
600
800
1000
1200
1400
1600
1800
2000
Com
prim
ento
rost
ro-c
loac
al (m
m)
A
SA
SA
SA
A
A
AA
AA F
F
T
T
T
T
A
AT
FT
T
A
A
FT
TT
T
A
A
F
FP
flaG
bra
Hm
odS
pun
Hpl
aLc
obH
ang
Lpoe
Tdor
Tstr
Hin
fA
ass
Lreg
Hpo
lLa
nnH
ytr
Xne
wLf
reH
car
Lmil
Equ
iM
sur
Mco
rE
aes
Ppl
iM
froC
qui
Ogu
iP
nig
Pm
erM
lem
Hyd
bH
ydg
Espécies
0
200
400
600
800
1000
1200
1400
1600
1800
2000
Com
prim
ento
rost
ro-c
loac
al (m
m)
A
SA
SA
SA
A
A
AA
AA F
F
T
T
T
T
A
AT
FT
T
Pfla
Gbr
aH
mod
Spu
nH
pla
Lcob
Han
gLp
oeTd
orTs
trH
inf
Aas
sLr
egH
pol
Lann
Hyt
rX
new
Lfre
Hca
rLm
ilE
qui
Msu
rM
cor
Eae
sP
pli
Mfro
Cqu
iO
gui
Pni
gP
mer
Mle
mH
ydb
Hyd
g
Espécies
0
200
400
600
800
1000
1200
1400
1600
1800
2000
Com
prim
ento
rost
ro-c
loac
al (m
m)
A
SA
SA
SA
A
A
AA
AA F
F
T
T
T
T
A
AT
FT
T
A
A
FT
TT
T
A
A
F
F

28
4.2.2 Forma do corpo
Os resultados da análise de componentes principais (Tabelas 4 e 5, Figura 5) sobre as
12 variáveis relativas da forma do corpo (resíduos das regressões; ver Métodos) ilustram
diferenças entre as espécies de Boinae. Da variação total dos dados (12 eixos), os dois
primeiros eixos (componentes principais) explicam 81,91% da variância (Tabela 4). O eixo 1
(56,82% da variância), ilustra um gradiente relacionado principalmente ao aumento no
diâmetro do olho, no comprimento da cauda, na largura da escama ventral (sentido dos
valores positivos), no tamanho da cabeça (comprimento, largura e altura) e robustez (massa e
circunferência do corpo) (sentido dos valores negativos). O eixo 2 (25,09% da variância)
ilustra um gradiente relacionado principalmente ao aumento das distâncias entre os olhos,
entre as narinas, entre o rostro e as narinas e entre o rostro e os olhos no sentido dos valores
negativos (Tabela 5 e Figura 5).
Boa constrictor e Eunectes murinus se destacam por apresentarem maior robustez e
tamanho relativo da cabeça. Eunectes murinus ainda difere das demais por apresentar as
narinas e os olhos relativamente mais próximos entre si, no dorso da cabeça, menor diâmetro
do olho e escama ventral mais estreita. Corallus hortulanus se destaca por apresentar,
principalmente, cauda relativamente longa e menor robustez.
Tabela 4. Autovalores e porcentagens da variância explicada e acumulada pelos primeiros sete componentes principais (eixos 1 a 7) da Análise de Componentes Principais (ACP) para as 12 variáveis relativas da forma do corpo entre as espécies de Boinae.
Eixo 1 Eixo 2 Eixo 3 Eixo 4 Eixo 5 Eixo 6 Eixo 7 Autovalores 6,82 3,01 0,59 0,54 0,31 0,21 0,15 Porcentagem 56,82 25,09 4,90 4,46 2,57 1,73 1,25 Porcentagem acumulada 56,82 81,91 86,81 91,28 93,85 95,58 96,83

29
Tabela 5. Autovetores dos primeiros sete componentes principais (eixos 1 a 7) da Análise de Componentes Principais (ACP) para as 12 variáveis relativas da forma do corpo entre as espécies de Boinae. Valores em negrito indicam os autovetores mais importantes para os dois primeiros eixos (1 e 2). Abreviações: IN = distância entre as narinas, RN = entre o rostro e as narinas, IO = entre os olhos, RO = entre o rostro e os olhos, Olho = diâmetro dos olhos, Ccab = comprimento da cabeça, Lcab = largura da cabeça, Acab = altura da cabeça, CC = comprimento da cauda, M = massa do corpo, Lev = largura da escama ventral, Circ = circunferência do corpo.
Caracteres Eixo 1 Eixo 2 Eixo 3 Eixo 4 Eixo 5 Eixo 6 Eixo 7 IN 0,21 -0,45 0,18 -0,06 -0,02 -0,27 -0,17 RN -0,14 -0,44 -0,11 0,54 0,53 0,39 0,17 IO 0,11 -0,52 0,05 0,18 -0,35 -0,23 -0,13 RO 0,24 -0,41 -0,06 -0,14 -0,14 0,03 0,11 Olho 0,32 0,05 0,64 -0,08 0,18 0,21 0,39 Ccab -0,35 -0,06 -0,06 -0,31 -0,16 0,48 -0,22 Lcab -0,32 -0,18 0,21 0,08 -0,56 0,40 0,16 Acab -0,32 -0,13 0,49 -0,12 0,33 -0,07 -0,60 CC 0,29 0,25 0,42 0,30 -0,21 0,20 -0,18 M -0,36 -0,05 0,16 -0,23 0,13 -0,19 0,26 Lev 0,30 -0,21 -0,09 -0,62 0,17 0,29 0,08 Circ -0,35 -0,07 0,22 -0,06 -0,06 -0,36 0,46
Figura 5. Diagrama de ordenação (“biplot”) dos indivíduos (pontos) e espécies de Boinae analisadas em relação a 12 variáveis relativas da forma do corpo (resíduos de regressão; setas) ao longo do 1o e 2o eixos da análise de componentes principais (ACP). As abreviações das variáveis (setas) são as mesmas da Tabela 5. Abreviações das espécies: Chor = Corallus hortulanus, Emur = Eunectes murinus, Ecen = Epicrates cenchria, Bcon = Boa constrictor. Escala dos vetores = 1.0.
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
-0.6 -0.4 -0.2 0 0.2 0.4 0.6
Chor Emur Ecen Bcon
RNIO
IN
RO
Olho
Lev
Ccab
AcabLcab
CC
M
Circ
Eixo 2
Eixo 1

30
Diferenças na forma relativa do corpo entre as espécies de Elapinae também são
ilustradas pelos resultados da análise de componentes principais (Tabelas 6 e 7 e Figura 6).
Da variação total dos dados (12 eixos), os dois primeiros eixos (componentes principais)
explicam 63,79% da variância (Tabela 6). O eixo 1 (43,7% da variância), ilustra um gradiente
relacionado principalmente ao aumento na distância entre o rostro e as narinas (sentido dos
valores positivos), no tamanho da cabeça, no comprimento da cauda e na robustez (sentido
dos valores negativos). O eixo 2 (20,09% da variância) ilustra um gradiente relacionado
principalmente ao aumento nas distâncias entre os olhos, entre as narinas, entre o rostro e os
olhos, no diâmetro do olho e na largura da escama ventral no sentido dos valores positivos
(Tabela 7 e Figura 6).
As três espécies fossóreas apresentaram forma do corpo semelhante. Micrurus
surinamensis se destaca principalmente por apresentar maior robustez e maiores tamanhos
relativos de cauda e cabeça. Também é espécie que apresenta as narinas e os olhos
relativamente mais próximos entre si, no dorso da cabeça.
Tabela 6. Autovalores e porcentagens da variância explicada e acumulada pelos primeiros sete componentes principais (eixos 1 a 7) da Análise de Componentes Principais (ACP) para as 12 variáveis relativas da forma do corpo entre espécies de Elapinae.
Eixo 1 Eixo 2 Eixo 3 Eixo 4 Eixo 5 Eixo 6 Eixo 7 Autovalores 5,24 2,41 1,24 0,83 0,71 0,44 0,35 Porcentagem 43,70 20,09 10,36 6,92 5,88 3,65 2,90 Porcentagem acumulada 43,70 63,79 74,15 81,07 86,95 90,60 93,50

31
Tabela 7. Autovetores dos primeiros sete componentes principais (eixos 1 a 7) da Análise de Componentes Principais (ACP) para as 12 variáveis relativas da forma do corpo entre espécies de Elapinae. Valores em negrito indicam os autovetores mais importantes para os dois primeiros eixos (1 e 2). As abreviações das variáveis são as mesmas da Tabela 5.
Caracteres Eixo 1 Eixo 2 Eixo 3 Eixo 4 Eixo 5 Eixo 6 Eixo 7 IN 0,22 0,44 -0,13 0,07 0,43 0,05 -0,34 RN 0,28 0,22 -0,25 0,36 0,11 -0,66 0,41 IO 0,14 0,53 0,11 0,04 0,24 0,43 -0,05 RO 0,04 0,37 -0,34 0,27 -0,77 0,16 -0,14 Olho -0,09 0,39 0,14 -0,71 -0,21 -0,46 -0,18 Ccab -0,39 0,09 0,23 0,11 -0,07 0,06 0,30 Lcab -0,40 0,09 0,05 0,18 -0,03 0,05 -0,03 Acab -0,40 0,14 -0,02 0,08 0,06 0,00 0,07 CC -0,34 -0,15 -0,15 0,30 0,13 -0,32 -0,69 M -0,35 0,17 -0,33 -0,08 0,18 0,04 0,18 Lev -0,06 0,25 0,72 0,34 -0,05 -0,20 -0,00 Circ -0,36 0,17 -0,26 -0,12 0,18 -0,01 0,24
Figura 6. Diagrama de ordenação (“biplot”) dos indivíduos (pontos) e espécies de Elapinae analisadas em relação a 12 variáveis relativas da forma do corpo (resíduos de regressão; setas) ao longo do 1o e 2o eixos da análise de componentes principais (ACP). As abreviações das espécies são as mesmas da Figura 4. As abreviações das variáveis (setas) são as mesmas da Tabela 5. Escala dos vetores = 1.0.
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
-0.6 -0.4 -0.2 0 0.2 0.4 0.6
Mcor Mlem Msur Mfro
IO
INRO
RN
CC
MCircAcabCcabLcab
Lev
Olho
Eixo 2
Eixo 1

32
Os resultados da análise de componentes principais (Tabelas 8 e 9) sobre as 12
variáveis relativas ilustram diferenças na forma relativa do corpo entre espécies (Figura 7),
grupos filogenéticos (Figura 8) e grupos de espécies (quanto ao uso do ambiente) de
Xenodontinae (Figura 9). Nas figuras 8 e 9 os pontos relativos aos indivíduos não estão
representados. Da variação total dos dados (12 eixos), os dois primeiros eixos (componentes
principais) explicam 72,34% da variância (Tabela 8). O eixo 1 (45,87% da variância), ilustra
um gradiente relacionado principalmente ao aumento no tamanho da cabeça, no comprimento
da cauda e na robustez no sentido dos valores negativos. O eixo 2 (26,47% da variância)
ilustra um gradiente relacionado principalmente ao aumento das distâncias entre as narinas,
entre os olhos, entre o rostro e as narinas, entre o rostro e os olhos, no diâmetro do olho e na
largura da escama ventral no sentido dos valores positivos (Tabela 9 e Figuras 7, 8 e 9).
Pelo menos quatro grupos de espécies apresentaram características próprias de
morfologia. Em um dos grupos estão as serpentes fossóreas de Elapomorphini, com cabeça e
cauda relativamente pequena e menor massa, circunferência do corpo e diâmetro relativo do
olho (Figuras 7, 8 e 9).
Outro grupo comporta as espécies aquáticas de Hydropsini, com as narinas e os olhos
relativamente mais próximos entre si e em posição mais anterior no dorso da cabeça (Figuras
7 e 8). Em relação à maioria das outras espécies, essas serpentes também apresentaram maior
robustez e tamanho relativo da cabeça, bem como menor largura relativa da escama ventral.
Hydrops triangularis ficou em posição intermediária entre as outras espécies de Hydropsini e
os representantes de Elapomorphini (Figura 7). Gomesophis brasiliensis e Ptychophis
flavovirgatus (aquáticas) também apresentam as narinas e os olhos relativamente mais
próximos entre si, maior robustez e menor largura relativa da escama ventral, diferindo dos
representantes terrícolas de Tachymenini (Figura 7).

33
Um terceiro grupo comporta as espécies terrícolas, aquáticas e a semi-aquática
(Sordellina punctata) com morfologia generalizada, incluindo os representantes de
Pseudoboini e Xenodontini (Figuras 7, 8 e 9).
O último grupo inclui as duas espécies de Hydrodynastes, Thamnodynastes strigatus e
Heterodon platyrhinus, com cabeça maior e maior robustez relativa em comparação às
espécies terrícolas, aquáticas e a semi-aquática mencionada acima (Figuras 7, 8 e 9).
Tabela 8. Autovalores e porcentagens da variância explicada e acumulada pelos primeiros sete componentes principais (eixos 1 a 7) da Análise de Componentes Principais (ACP) para as 12 variáveis relativas de forma do corpo entre espécies de Xenodontinae.
Eixo 1 Eixo 2 Eixo 3 Eixo 4 Eixo 5 Eixo 6 Eixo 7 Autovalores 5,50 3,18 1,10 0,68 0,46 0,31 0,28 Porcentagem 45,87 26,47 9,18 5,63 3,84 2,54 2,31 Porcentagem acumulada 45,87 72,34 81,52 87,15 90,98 93,52 95,83
Tabela 9. Autovetores dos primeiros sete componentes principais (eixos 1 a 7) da Análise de Componentes Principais (ACP) para as 12 variáveis relativas de forma do corpo entre espécies de Xenodontinae. Valores em negrito indicam os autovetores mais importantes para os dois primeiros eixos (1 e 2). As abreviações das variáveis são as mesmas da Tabela 5.
Caracteres Eixo 1 Eixo 2 Eixo 3 Eixo 4 Eixo 5 Eixo 6 Eixo 7 IN 0,31 0,34 0,12 0,03 -0,13 0,09 0,07 RN 0,25 0,35 0,30 0,11 0,11 -0,12 -0,73 IO 0,26 0,31 0,31 -0,26 -0,32 0,42 0,40 RO 0,22 0,41 -0,06 -0,32 -0,11 -0,45 0,01 Olho -0,05 0,37 -0,50 -0,36 0,59 0,35 -0,03 Ccab -0,37 0,22 0,00 0,27 -0,12 0,16 -0,17 Lcab -0,37 0,20 0,08 0,10 -0,22 0,31 -0,06 Acab -0,36 0,25 0,13 0,07 -0,04 0,22 -0,11 CC -0,28 0,22 -0,46 -0,12 -0,45 -0,35 0,01 M -0,31 0,11 0,47 -0,08 0,47 -0,30 0,33 Lev 0,14 0,37 -0,19 0,72 0,16 -0,16 0,38 Circ -0,36 0,15 0,23 -0,24 -0,04 -0,26 0,05

34
Figura 7. Diagrama de ordenação (“biplot”) dos indivíduos (pontos) e espécies de Xenodontinae analisadas em relação a 12 variáveis relativas de forma do corpo (resíduos de regressão; setas) ao longo do 1o e 2o eixos da análise de componentes principais (ACP). As abreviações das espécies são as mesmas da Figura 4. As abreviações das variáveis (setas) são as mesmas da Tabela 5. Escala dos vetores = 0.5.
-0.25
-0.2
-0.15
-0.1
-0.05
0
0.05
0.1
0.15
0.2
0.25
-0.25 -0.2 -0.15 -0.1 -0.05 0 0.05 0.1 0.15 0.2 0.25
Aass Pmer Equi Hmod Hcar Hinf Hang Hpol Hytr Ppli Spun Cqui Pnig Ogui Hydb
Hydg Tstr Tdor Pfla Gbra Xnew Eaes Lreg Lpoe Lmil Lcob Lfre Lann Hpla
LevRO
RN IOIN
Olho
CCAcab
CcabLcab
CircM
Eixo 1
Eixo 2

35
Figura 8. Diagrama de ordenação (“biplot”) dos grupos filogenéticos (polígonos) de Xenodontinae em relação a 12 variáveis relativas de forma do corpo (resíduos de regressão; setas) ao longo do 1o e 2o eixos da análise de componentes principais (ACP). ▬▬ = Hydropsini; ... = Elapomorphini; --- = Tachymenini; ▬ . ▬ = Pseudoboini; ▬▬- = Xenodontini; ... = Hydrodynastes; --- = Heterodon platyrhinus; ▬▬ = Leptodeira annulata; ▬ .. ▬ = Sordellina punctata. As abreviações das variáveis (setas) são as mesmas da Tabela 5. Escala dos vetores = 0.5.
-0.25
-0.2
-0.15
-0.1
-0.05
0
0.05
0.1
0.15
0.2
0.25
-0.25 -0.2 -0.15 -0.1 -0.05 0 0.05 0.1 0.15 0.2 0.25
LevRO
RN IO IN
Olho
CCAcab
CcabLcab
CircM
Eixo 2
Eixo 1

36
Figura 9. Diagrama de ordenação (“biplot”) dos grupos ecológicos (tipo de ambiente utilizado; polígonos) de Xenodontinae em relação a 12 variáveis relativas de forma do corpo (resíduos de regressão; setas) ao longo do 1o e 2o eixos da análise de componentes principais (ACP). ▬▬ = serpentes aquáticas; .... = serpentes fossóreas; ▬▬ = serpentes terrícolas; --- = Serpentes semi-aquáticas. As abreviações das variáveis (setas) são as mesmas da Tabela 5. Escala dos vetores = 0.5.
-0.25
-0.2
-0.15
-0.1
-0.05
0
0.05
0.1
0.15
0.2
0.25
-0.25 -0.2 -0.15 -0.1 -0.05 0 0.05 0.1 0.15 0.2 0.25
LevRO
RN IO IN
Olho
CCAcab
CcabLcab
CircM
Eixo 2
Eixo 1

37
As distribuições das 12 variáveis relativas da forma do corpo entre as espécies de
Boinae estão ilustradas nas figuras 10 a 13. A análise de covariância detectou diferenças
significativas entre espécies e entre aquelas de diferentes hábitos (uso do ambiente) para todas
as variáveis (Tabelas 10 a 21). As distâncias entre as narinas, entre os olhos, entre o rostro e
as narinas, entre o rostro e os olhos e a largura da escama ventral são significativamente
menores em Eunectes murinus. Boa constrictor e Eunectes murinus não diferiram quanto ao
tamanho da cabeça (comprimento, largura e altura) ou robustez (massa e circunferência).
Corallus hortulanus apresentou menor cabeça e robustez. Eunectes murinus e Epicrates
cenchria não diferiram em relação ao diâmetro do olho e comprimento da cauda. A cauda e o
diâmetro do olho são significativamente maiores em Corallus hortulanus e menores em Boa
constrictor.
Para as espécies de Elapinae, as distribuições das 12 variáveis relativas da forma do
corpo estão ilustradas nas figuras 14 a 17. Diferenças significativas entre espécies e entre
aquelas de diferentes hábitos foram encontradas para a maioria das variáveis (Tabelas 10 a
21). Micrurus surinamensis apresentou menores distâncias entre as narinas, entre os olhos e
entre o rostro e as narinas. A distância entre o rostro e os olhos não diferiu entre as espécies
ou entre espécies de diferentes hábitos. A largura da escama ventral e o diâmetro do olho,
embora sejam distintas entre espécies, não diferiram significativamente entre a espécie
aquática e as fossóreas. Micrurus surinamensis diferiu significativamente das outras espécies
em relação à robustez, tamanho de cabeça e comprimento de cauda.
A forma do corpo também diferiu entre as espécies de Xenodontinae analisadas. Para
essas espécies, as distribuições das 12 variáveis relativas da forma do corpo estão ilustradas
nas figuras 18 a 21. Os resultados das análises de covariância, nas comparações entre espécies
e entre espécies de diferentes hábitos, revelam diferenças significativas para a maioria das
variáveis (Tabelas 10 a 21).

38
As distâncias entre as narinas, entre os olhos, entre o rostro e as narinas, entre o rostro
e os olhos e a largura da escama ventral foram, em geral, significativamente menores em
espécies aquáticas. Representantes de Hydropsini (todos aquáticos) foram comparados com
Heterodon platyrhinus, Leptodeira annulata, espécies de Pseudoboini (terrícolas) e
Elapomorphini (fossóreas) e apresentaram menores valores para essas variáveis. Espécies
aquáticas e terrícolas, dentro de Tachymenini, apenas não diferiram em relação à distância
entre o rostro e as narinas. Espécies aquáticas e terrícolas, dentro de Xenodontini, apenas não
diferiram em relação à distância entre o rostro e os olhos. As espécies de Hydrodynastes
(semi-aquáticas) somente diferiram das espécies de Pseudoboini (grupo mais próximo na
filogenia) para a distância entre os olhos.
A massa e circunferência do corpo foram, em geral, significativamente maiores nas
espécies aquáticas. A circunferência do corpo apenas não diferiu entre serpentes aquáticas e
terrícolas dentro de Xenodontini. As duas Hydrodynastes são mais robustas que os
representantes de Pseudoboini analisados.
O tamanho da cabeça (comprimento, largura e altura) foi muito variável entre as
espécies de Xenodontinae. Os representantes de Hydropsini apresentaram, em geral, cabeça
maior e diferiram significativamente em comparação a Heterodon platyrhinus, Leptodeira
annulata, espécies de Pseudoboini e Elapomorphini. Espécies terrícolas de Tachymenini
apresentaram cabeça maior em comparação às aquáticas, talvez devido a Gomesophis
brasiliensis possuir cabeça extremamente pequena. Espécies aquáticas e terrícolas, dentro de
Xenodontini, não diferiram em relação ao tamanho da cabeça, embora diferenças entre
espécie sejam evidentes. As duas Hydrodynastes tem cabeça maior que os Pseudoboini
analisados.
O diâmetro do olho também foi variável e significativamente diferente entre espécies e
entre aquelas de diferentes hábitos. Os representantes de Hydropsini apresentaram olho de

39
diâmetro intermediário aos representantes de Elapomorphini (menor diâmetro) e Pseudoboini,
Heterodon platyrhinus e Leptodeira annulata (maior diâmetro). O diâmetro do olho foi
significativamente menor nas espécies aquáticas dentro de Tachymenini e Xenodontini. As
duas Hydrodynastes também apresentaram olho menor que os Pseudoboini analisados.
O comprimento da cauda é maior nos representantes de Hydropsini e Hydrodynastes
em comparação a espécies terrícolas (comprimento de cauda intermediário) e fossóreas (cauda
curta). Essa variável não diferiu significativamente entre espécies aquáticas e terrícolas dentro
de Tachymenini e Xenodontini.

40
Figura 10. Variação da distância relativa entre as narinas (a), entre o rostro e as narinas (b) e entre os olhos (c) para as espécies de Boinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; Ar = arbóreo; T = terrícola. As abreviações das espécies são as mesmas da Figura 5.
Emur Ecen Bcon ChorEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Dis
tânc
ia re
lativ
a en
tre a
s na
rinas
(IN
)A
T T
Ar(a)
Emur Ecen Chor BconEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Dis
tânc
ia r
elat
iva
entre
o r
ostro
e a
s na
rinas
(RN
)
T
TA Ar
(b)
Emur Ecen Chor BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Dis
tânc
ia re
lativ
a en
tre o
s ol
hos
(IO)
T
TA
Ar(c)
Emur Ecen Bcon ChorEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Dis
tânc
ia re
lativ
a en
tre a
s na
rinas
(IN
)A
T T
Ar(a)
Emur Ecen Bcon ChorEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Dis
tânc
ia re
lativ
a en
tre a
s na
rinas
(IN
)A
T T
Ar(a)
Emur Ecen Chor BconEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Dis
tânc
ia r
elat
iva
entre
o r
ostro
e a
s na
rinas
(RN
)
T
TA Ar
(b)
Emur Ecen Chor BconEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Dis
tânc
ia r
elat
iva
entre
o r
ostro
e a
s na
rinas
(RN
)
T
TA Ar
(b)
Emur Ecen Chor BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Dis
tânc
ia re
lativ
a en
tre o
s ol
hos
(IO)
T
TA
Ar(c)
Emur Ecen Chor BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Dis
tânc
ia re
lativ
a en
tre o
s ol
hos
(IO)
T
TA
Ar(c)

41
Figura 11. Variação da distância relativa entre o rostro e os olhos (a), no diâmetro do olho (b) e no comprimento relativo da cauda (c) para as espécies de Boinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; Ar = arbóreo; T = terrícola. As abreviações das espécies são as mesmas da Figura 5.
Emur Ecen Bcon ChorEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Dis
tânc
ia re
lativ
a en
tre o
rost
ro e
os
olho
s (R
O)
T T
A
Ar(d)
Bcon Emur Ecen ChorEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Diâ
met
ro re
lativ
o do
olh
o T
T
A
Ar(e)
Bcon Emur Ecen ChorEspécies
-0,8
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
Com
prim
ento
rela
tivo
da c
auda
(CC
) T
T
A
Ar(f)
Emur Ecen Bcon ChorEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Dis
tânc
ia re
lativ
a en
tre o
rost
ro e
os
olho
s (R
O)
T T
A
Ar(d)
Emur Ecen Bcon ChorEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Dis
tânc
ia re
lativ
a en
tre o
rost
ro e
os
olho
s (R
O)
T T
A
Ar(d)
Bcon Emur Ecen ChorEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Diâ
met
ro re
lativ
o do
olh
o T
T
A
Ar(e)
Bcon Emur Ecen ChorEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Diâ
met
ro re
lativ
o do
olh
o T
T
A
Ar(e)
Bcon Emur Ecen ChorEspécies
-0,8
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
Com
prim
ento
rela
tivo
da c
auda
(CC
) T
T
A
Ar(f)
Bcon Emur Ecen ChorEspécies
-0,8
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
Com
prim
ento
rela
tivo
da c
auda
(CC
) T
T
A
Ar(f)
(a)
(b)
(c)

42
Figura 12. Variação do comprimento (a), largura (b) e altura relativa da cabeça (c) para as espécies de Boinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; Ar = arbóreo; T = terrícola. As abreviações das espécies são as mesmas da Figura 5.
Chor Ecen Emur BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Com
prim
ento
rela
tivo
da c
abeç
a (C
cab)
A
T
T
Ar
(a)
Chor Ecen Emur BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Larg
ura
rela
tiva
da c
abeç
a (L
cab)
A T
TAr
(b)
Chor Ecen Emur BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Altu
ra re
lativ
a da
cab
eça
(Aca
b)
A
TT
Ar
(c)
Chor Ecen Emur BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Com
prim
ento
rela
tivo
da c
abeç
a (C
cab)
A
T
T
Ar
(a)
Chor Ecen Emur BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Com
prim
ento
rela
tivo
da c
abeç
a (C
cab)
A
T
T
Ar
(a)
Chor Ecen Emur BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Larg
ura
rela
tiva
da c
abeç
a (L
cab)
A T
TAr
(b)
Chor Ecen Emur BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Larg
ura
rela
tiva
da c
abeç
a (L
cab)
A T
TAr
(b)
Chor Ecen Emur BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Altu
ra re
lativ
a da
cab
eça
(Aca
b)
A
TT
Ar
(c)
Chor Ecen Emur BconEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Altu
ra re
lativ
a da
cab
eça
(Aca
b)
A
TT
Ar
(c)

43
Figura 13. Variação da largura relativa da escama ventral (a), da circunferência (b) e da massa relativa do corpo (c) para as espécies de Boinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; Ar = arbóreo; T = terrícola. As abreviações das espécies são as mesmas da Figura 5.
Emur Bcon Ecen ChorEspécies
-0,7
-0,6
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Larg
ura
rela
tiva
da e
scam
a ve
ntra
l (Le
v)
A
TT
Ar(d)
Chor Ecen Emur BconEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Circ
unfe
rênc
ia re
lativ
a (C
irc)
A TT
Ar
(e)
Chor Ecen Bcon EmurEspécies
-1,4-1,2-1,0-0,8-0,6-0,4-0,20,00,20,40,60,81,01,21,4
Mas
sa re
lativ
a (M
)
A
T
Ar
T(f)
Emur Bcon Ecen ChorEspécies
-0,7
-0,6
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Larg
ura
rela
tiva
da e
scam
a ve
ntra
l (Le
v)
A
TT
Ar(d)
Emur Bcon Ecen ChorEspécies
-0,7
-0,6
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Larg
ura
rela
tiva
da e
scam
a ve
ntra
l (Le
v)
A
TT
Ar(d)
Chor Ecen Emur BconEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Circ
unfe
rênc
ia re
lativ
a (C
irc)
A TT
Ar
(e)
Chor Ecen Emur BconEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
Circ
unfe
rênc
ia re
lativ
a (C
irc)
A TT
Ar
(e)
Chor Ecen Bcon EmurEspécies
-1,4-1,2-1,0-0,8-0,6-0,4-0,20,00,20,40,60,81,01,21,4
Mas
sa re
lativ
a (M
)
A
T
Ar
T(f)
Chor Ecen Bcon EmurEspécies
-1,4-1,2-1,0-0,8-0,6-0,4-0,20,00,20,40,60,81,01,21,4
Mas
sa re
lativ
a (M
)
A
T
Ar
T(f)
(a)
(b)
(c)
Kg
Kg
Kg
Kg

44
Figura 14. Variação da distância relativa entre as narinas (a), entre o rostro e as narinas (b) e entre os olhos (c) para as espécies de Elapinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; F = fossóreo. As abreviações das espécies são as mesmas da Figura 4.
Msur Mlem Mcor MfroEspécies
-0 ,4
-0 ,3
-0 ,2
-0 ,1
0 ,0
0 ,1
0 ,2
Dis
tânc
ia re
lativ
a en
tre
as n
arin
as (I
N) F
F FA(a)
Msur Mcor Mfro MlemEspécies
-0,6
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Dis
tânc
ia r
elativ
a en
tre o
ros
tro e
as
narin
as (R
N)
F FF
A
(b)
Msur Mlem Mcor MfroEspécies
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
Dis
tânc
ia re
lativ
a e
ntre
os
olho
s (I
O)
F
F
F
A
(c)
Msur Mlem Mcor MfroEspécies
-0 ,4
-0 ,3
-0 ,2
-0 ,1
0 ,0
0 ,1
0 ,2
Dis
tânc
ia re
lativ
a en
tre
as n
arin
as (I
N) F
F FA(a)
Msur Mlem Mcor MfroEspécies
-0 ,4
-0 ,3
-0 ,2
-0 ,1
0 ,0
0 ,1
0 ,2
Dis
tânc
ia re
lativ
a en
tre
as n
arin
as (I
N) F
F FA(a)
Msur Mcor Mfro MlemEspécies
-0,6
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Dis
tânc
ia r
elativ
a en
tre o
ros
tro e
as
narin
as (R
N)
F FF
A
(b)
Msur Mcor Mfro MlemEspécies
-0,6
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Dis
tânc
ia r
elativ
a en
tre o
ros
tro e
as
narin
as (R
N)
F FF
A
(b)
Msur Mlem Mcor MfroEspécies
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
Dis
tânc
ia re
lativ
a e
ntre
os
olho
s (I
O)
F
F
F
A
(c)
Msur Mlem Mcor MfroEspécies
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
Dis
tânc
ia re
lativ
a e
ntre
os
olho
s (I
O)
F
F
F
A
(c)

45
Figura 15. Variação da distância relativa entre o rostro e os olhos (a), do diâmetro do olho (b) e do comprimento relativo da cauda (c) para as espécies de Elapinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; F = fossóreo. As abreviações das espécies são as mesmas da Figura 4.
Msur Mcor Mlem MfroEspécies
-0,16-0,14-0,12-0,10-0,08-0,06-0,04-0,020,000,020,040,060,080,10
Dis
tânc
ia re
lativ
a en
tre o
rostr
o e
os o
lhos
(RO
)
FF F
A(d)
Mlem Mcor Msur MfroEspécies
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Diâm
etro
rela
tivo
do o
lho
F
FF
A(e)
Mfro Mlem Mcor MsurEspécies
-0,8
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
1,0
Com
prim
ento
rela
tivo
da c
auda
(CC
)
F
F F
A(f)
Msur Mcor Mlem MfroEspécies
-0,16-0,14-0,12-0,10-0,08-0,06-0,04-0,020,000,020,040,060,080,10
Dis
tânc
ia re
lativ
a en
tre o
rostr
o e
os o
lhos
(RO
)
FF F
A(d)
Msur Mcor Mlem MfroEspécies
-0,16-0,14-0,12-0,10-0,08-0,06-0,04-0,020,000,020,040,060,080,10
Dis
tânc
ia re
lativ
a en
tre o
rostr
o e
os o
lhos
(RO
)
FF F
A(d)
Mlem Mcor Msur MfroEspécies
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Diâm
etro
rela
tivo
do o
lho
F
FF
A(e)
Mlem Mcor Msur MfroEspécies
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Diâm
etro
rela
tivo
do o
lho
F
FF
A(e)
Mfro Mlem Mcor MsurEspécies
-0,8
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
1,0
Com
prim
ento
rela
tivo
da c
auda
(CC
)
F
F F
A(f)
Mfro Mlem Mcor MsurEspécies
-0,8
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
1,0
Com
prim
ento
rela
tivo
da c
auda
(CC
)
F
F F
A(f)
(a)
(b)
(c)

46
Figura 16. Variação no comprimento (a), largura (b) e altura relativa da cabeça (c) para as espécies de Elapinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; F = fossóreo. As abreviações das espécies são as mesmas da Figura 4.
Mlem Mcor Mfro MsurEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Com
prim
ento
rela
tivo
da c
abeç
a (C
cab)
FF
F
A(a)
Mlem Mfro Mcor MsurEspécies
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
Larg
ura
rela
tiva
da c
abeç
a (L
cab)
FF F
A(b)
Mlem Mfro Mcor MsurEspécies
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
Altu
ra re
lativ
a da
cab
eça
(Aca
b)
FF F
A(c)
Mlem Mcor Mfro MsurEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Com
prim
ento
rela
tivo
da c
abeç
a (C
cab)
FF
F
A(a)
Mlem Mcor Mfro MsurEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
Com
prim
ento
rela
tivo
da c
abeç
a (C
cab)
FF
F
A(a)
Mlem Mfro Mcor MsurEspécies
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
Larg
ura
rela
tiva
da c
abeç
a (L
cab)
FF F
A(b)
Mlem Mfro Mcor MsurEspécies
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
Larg
ura
rela
tiva
da c
abeç
a (L
cab)
FF F
A(b)
Mlem Mfro Mcor MsurEspécies
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
Altu
ra re
lativ
a da
cab
eça
(Aca
b)
FF F
A(c)
Mlem Mfro Mcor MsurEspécies
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
Altu
ra re
lativ
a da
cab
eça
(Aca
b)
FF F
A(c)

47
Figura 17. Variação na largura relativa da escama ventral (a), circunferência (b) e massa relativa do corpo (c) para as espécies de Elapinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; F = fossóreo. As abreviações das espécies são as mesmas da Figura 4.
Msur Mlem Mfro McorEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
Larg
ura
rela
tiva
da e
scam
a ve
ntra
l (Le
v)
F
FF
A(d)
Mlem Mfro Mcor MsurEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
0,6
Circ
unfe
rênc
ia re
lativ
a do
cor
po (C
irc)
FFF
A(e)
Mlem Mfro Mcor MsurEspécies
-1,0
-0,8
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Mas
sa re
lativ
a do
cor
po (M
)
F F F
A
(f)
Msur Mlem Mfro McorEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
Larg
ura
rela
tiva
da e
scam
a ve
ntra
l (Le
v)
F
FF
A(d)
Msur Mlem Mfro McorEspécies
-0,5
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
Larg
ura
rela
tiva
da e
scam
a ve
ntra
l (Le
v)
F
FF
A(d)
Mlem Mfro Mcor MsurEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
0,6
Circ
unfe
rênc
ia re
lativ
a do
cor
po (C
irc)
FFF
A(e)
Mlem Mfro Mcor MsurEspécies
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
0,6
Circ
unfe
rênc
ia re
lativ
a do
cor
po (C
irc)
FFF
A(e)
Mlem Mfro Mcor MsurEspécies
-1,0
-0,8
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Mas
sa re
lativ
a do
cor
po (M
)
F F F
A
(f)
Mlem Mfro Mcor MsurEspécies
-1,0
-0,8
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Mas
sa re
lativ
a do
cor
po (M
)
F F F
A
(f)
(a)
(b)
(c)
Kg
Kg
Kg
Kg

48
Figura 18. Variação da distância relativa entre as narinas (a), entre o rostro e as narinas (b) e entre os olhos (c) para as espécies de Xenodontinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; SA = semi-aquático; T = terrícola, F = fossóreo. As abreviações das espécies são as mesmas da Figura 4.
Ppl
iH
mod Hin
fH
ytr
Hca
rH
ang
Hpo
lP
flaG
bra
Tstr
Tdor
Cqu
iS
pun
Lmil
Hyd
gO
gui
Aas
sLr
egLp
oeE
qui
Lcob
Hpl
aX
new
Lfre
Pm
erLa
nnP
nig
Hyd
bE
aes
Espécies
-1.2
-1.0
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
Dis
tânc
ia re
lativ
a en
tre a
s na
rinas
(IN
)
A
A A
A
A
T
T
AA
A
T T T T
T
T
TSA
SASA F F F
A
A
A
A
T T
Hm
od Ppl
iH
inf
Hpo
lH
ytr
Hca
rH
ang
Lann
Cqu
iO
gui
Gbr
aTs
trA
ass
Pfla
Tdor
Xne
wLr
egH
ydb
Hyd
gLm
ilE
qui
Lpoe
Pni
gP
mer
Lcob
Eae
sLf
reS
pun
Hpl
a
Espécies
-1.4-1.2-1.0-0.8-0.6-0.4-0.20.00.20.40.60.81.0
Dis
tânc
ia re
lativ
a en
tre o
rost
ro e
nar
inas
(RN)
A A A A
A
SA
T
T
A
A
A
T T
T
F
T
A
T
T
FF
A
A
A
T
SAT
SA T
Hm
od Hin
fH
car
Han
gH
pol
Ppl
iG
bra
Pfla
Cqu
iO
gui
Hyt
rLp
oeLm
ilLr
egP
nig
Tstr
Equ
iLf
reS
pun
Lcob
Pm
erLa
nnX
new
Tdor
Aas
sH
ydg
Hpl
aE
aes
Hyd
b
Espécies
-0.6
-0.4
-0.2
0.0
0.2
0.4
Dis
tânc
ia re
lativ
a en
tre o
s ol
hos
(IO)
T
A
A
A
T SA
SA
A
AA
T
T
F
T
T
A
A
T
FF
SAT
A A A
T T
TA
(a)
(b)
(c)

49
Figura 19. Variação da distância relativa entre o rostro e os olhos (a), no diâmetro do olho (b) e no comprimento relativo da cauda (c) para as espécies de Xenodontinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; SA = semi-aquático; T = terrícola, F = fossóreo. As abreviações das espécies são as mesmas da Figura 4.
Ppl
iH
inf
Hm
odH
car
Hpo
lP
mer
Han
gE
qui
Hyt
rA
ass
Gbr
aP
flaTs
trS
pun
Cqu
iLa
nnTd
orX
new
Hyd
gLm
ilLp
oeE
aes
Lfre
Hyd
bO
gui
Lcob
Pni
gH
pla
Lreg
Espécies
-0.4
-0.3
-0.2
-0.1
0.0
0.1
0.2
0.3
0.4
Dis
tânc
ia re
lativ
a en
tre o
rost
ro e
os
olho
s (R
O)
A A
A
TT
T
SA
A
F
A
A
AA
T
SA
SAA
T
A
A
F
F
T
T
T
TT A
T
Pm
erE
qui
Aas
sH
ytr
Han
gP
pli
Hin
fS
pun
Hpo
lH
car
Ogu
iH
ydg
Hyd
bP
nig
Hm
odC
qui
Lfre
Lcob Ts
trP
flaLm
ilLp
oeG
bra
Tdor
Eae
sH
pla
Xne
wLr
egLa
nn
Espécies
-1.0
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
Diâ
met
ro re
lativ
o do
olh
o
A
A
F
F
SA T
A A
A T T
T
F
A T
T
A A A
SA
A
T
A A
T
T T
T
SA
Pm
erA
ass
Equ
iE
aes
Gbr
aH
ytr
Spu
nP
pli
Lfre
Xne
wLp
oeLc
obLm
ilH
pla
Ogu
iTs
trH
car
Tdor
Cqu
iH
mod
Pni
gP
flaH
inf
Lann
Hyd
gLr
egH
ydb
Hpo
lH
ang
Espécies
-1.4-1.2-1.0-0.8-0.6-0.4-0.20.00.20.40.60.81.0
Com
prim
ento
rela
tivo
da c
auda
(CC
) A
F
A
T
A
T
SA SA
F
A A
A
A
T
T
AA T
T
F
AT
T T
SA
A T
A
T
(a)
(b)
(c)

50
Figura 20. Variação no comprimento (a), largura (b) e altura relativa da cabeça (c) para as espécies de Xenodontinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; SA = semi-aquático; T = terrícola, F = fossóreo. As abreviações das espécies são as mesmas da Figura 4.
Aas
sE
qui
Lfre
Pm
erS
pun
Ogu
iH
ytr
Eae
sG
bra
Cqu
iLa
nnP
nig
Lcob
Lreg
Lpoe
Tdor
Lmil
Xne
wP
flaH
pol
Hyd
bH
mod Ppl
iH
car
Tstr
Hin
fH
ydg
Han
gH
pla
Espécies
-1.0
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
Com
prim
ento
rela
tivo
da c
abeç
a (C
cab)
T
F
F
A
A
A
A
A A
T T
T T
T
T
T A
T F
AA
A
SA
T T
A
SA
A
SA
Aas
sP
mer
Equ
iO
gui
Spu
nLf
reG
bra
Eae
sC
qui
Lreg
Pni
gH
ytr
Lann
Lpoe
Lcob
Tdor
Lmil
Xne
wP
flaH
mod
Hpo
lH
car
Tstr
Hyd
bP
pli
Hyd
gH
inf
Han
gH
pla
Espécies
-1.0
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
Larg
ura
rela
tiva
da c
abeç
a (L
cab)
SA
F
A
A
T
SA A
A
T T
T
T
A
A
A
A
SA
F T F
T
T
T
T
T A
A
A
A
Aas
sO
gui
Equ
iP
mer
Lfre
Spu
nE
aes
Lann
Hyt
rP
nig
Lreg
Gbr
aC
qui
Lpoe
Lcob
Xne
wLm
ilTd
orH
car
Hm
odH
pol
Pfla Ppl
iH
ang
Hin
fH
ydb
Tstr
Hyd
gH
pla
Espécies
-1.0
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
Altu
ra re
lativ
a da
cab
eça
(Aca
b)
F
T
A
A
T
A
T
SA
F
T
A
T A
A
F
T A
SA
T SA
T
A
T
A
A
T
T A
A
(a)
(b)
(c)

51
Figura 21. Variação da largura relativa da escama ventral (a), circunferência (b) e massa relativa do corpo (c) para as espécies de Xenodontinae analisadas. Linhas dentro das caixas = médias; caixas = desvio-padrão; barras verticais = valores mínimos e máximos. Acima das espécies é indicado o tipo de ambiente utilizado: A = aquático; SA = semi-aquático; T = terrícola, F = fossóreo. As abreviações das espécies são as mesmas da Figura 4.
Ppl
iH
ytr
Aas
sH
mod
Hpo
lP
flaH
inf
Gbr
aH
car
Han
gP
mer
Hyd
bLa
nnLf
reTs
trTd
orLm
ilE
qui
Lcob
Hyd
gO
gui
Pni
gS
pun
Xne
wH
pla
Cqu
iLp
oeLr
egE
aes
Espécies
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
Larg
ura
rela
tiva
da e
scam
a ve
ntra
l (Le
v)
A
A
A
SA
T
A
A A
A
T
A F
F
T TA
T T
T
SA
A T
(a)
A
TT
SA
F
T
A
Aas
sP
mer
Lfre
Lann
Equ
iE
aes
Lreg
Spu
nO
gui
Lpoe
Pni
gC
qui
Lcob
Lmil
Hyt
rX
new
Tdor
Hca
rP
flaH
pol
Hin
fG
bra
Tstr
Hm
od Ppl
iH
ang
Hyd
bH
ydg
Hpl
a
Espécies
-1.0
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
Circ
unfe
rênc
ia re
lativ
a do
cor
po (C
irc)
A
F
F
A
A
F
T
A
A
A
T A
A
T SA
A T
SA
T
A
T
T
A
A
T
T
T
T SA
(b)
Aas
sLa
nnLr
egE
aes
Lfre
Ogu
iP
mer
Pni
gLp
oe Hyt
rE
qui
Cqu
iS
pun
Tdor
Lcob
Hpo
lX
new
Lmil
Han
gTs
trP
flaH
car
Hm
od Hin
fH
ydb
Hyd
gG
bra
Ppl
iH
pla
Espécies
-1.4-1.2-1.0-0.8-0.6-0.4-0.20.00.20.40.60.81.01.21.4
Mas
sa re
lativ
a do
cor
po (M
)
A
SA
(c) T
A
F
T A
A
A T
A
T
SA
T
A
TT
SA
T
A
A
AT
T
A
T F
A
F
Kg
Kg
Kg
Kg

52
Tabela 10. Resultados da análise de covariância para as comparações da distância entre as narinas (IN) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 134,2 << 0,01 Uso do ambiente 2 183,1 << 0,01 Elapinae Espécies 3 88 35,1 << 0,01 Uso do ambiente 1 85,0 << 0,01 Xenodontinae Espécies 28 514 341,1 << 0,01 Uso do ambiente 3 161,5 << 0,01 Hydropsini Espécies 15 270 361,1 << 0,01 Uso do ambiente 3 741,7 << 0,01 Tachymenini Espécies 3 68 24,7 << 0,01 Uso do ambiente 1 65,5 << 0,01 Xenodontini Espécies 6 130 59,7 << 0,01 Uso do ambiente 1 6,1 < 0,05 Pseudoboini Espécies 4 97 42,0 << 0,01 Uso do ambiente 1 0,6 = 0,42
Tabela 11. Resultados da análise de covariância para as comparações da distância entre o rostro e as narinas (RN) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 43,1 << 0,01 Uso do ambiente 2 11,9 << 0,01 Elapinae Espécies 3 88 37,0 << 0,01 Uso do ambiente 1 98,3 << 0,01 Xenodontinae Espécies 28 514 67,9 << 0,01 Uso do ambiente 3 56,9 << 0,01 Hydropsini Espécies 15 270 73,77 << 0,01 Uso do ambiente 3 198,8 << 0,01 Tachymenini Espécies 3 68 1,3 = 0,28 Uso do ambiente 1 0,7 = 0,41 Xenodontini Espécies 6 130 9,7 << 0,01 Uso do ambiente 1 6,4 < 0,05 Pseudoboini Espécies 4 97 22,8 << 0,01 Uso do ambiente 1 1,1 = 0,30

53
Tabela 12. Resultados da análise de covariância para as comparações da distância entre os olhos (IO) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 195,3 << 0,01 Uso do ambiente 2 80,0 << 0,01 Elapinae Espécies 3 88 41,1 << 0,01 Uso do ambiente 1 33,8 << 0,01 Xenodontinae Espécies 28 514 86,5 << 0,01 Uso do ambiente 3 122,8 << 0,01 Hydropsini Espécies 15 270 77,0 << 0,01 Uso do ambiente 3 118,0 << 0,01 Tachymenini Espécies 3 68 16,0 << 0,01 Uso do ambiente 1 37,0 << 0,01 Xenodontini Espécies 6 130 42,6 << 0,01 Uso do ambiente 1 7,9 < 0,05 Pseudoboini Espécies 4 97 35,8 << 0,01 Uso do ambiente 1 48,7 << 0,01
Tabela 13. Resultados da análise de covariância para as comparações da distância entre o rostro e os olhos (RO) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 337,1 << 0,01 Uso do ambiente 2 452,8 << 0,01 Elapinae Espécies 3 88 1,1 = 0,37 Uso do ambiente 1 0,3 = 0,59 Xenodontinae Espécies 28 514 55,5 << 0,01 Uso do ambiente 3 78,1 << 0,01 Hydropsini Espécies 15 270 56,4 << 0,01 Uso do ambiente 3 149,3 << 0,01 Tachymenini Espécies 3 68 4,7 < 0,01 Uso do ambiente 1 7,8 < 0,01 Xenodontini Espécies 6 130 2,4 < 0,05 Uso do ambiente 1 1,1 = 0,30 Pseudoboini Espécies 4 97 15,2 << 0,01 Uso do ambiente 1 2,8 = 0,10

54
Tabela 14. Resultados da análise de covariância para as comparações do diâmetro do olho entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 136,3 << 0,01 Uso do ambiente 2 83,9 << 0,01 Elapinae Espécies 3 88 13,4 << 0,01 Uso do ambiente 1 1,9 = 0,18 Xenodontinae Espécies 28 514 122,6 << 0,01 Uso do ambiente 3 266,3 << 0,01 Hydropsini Espécies 15 270 122,6 << 0,01 Uso do ambiente 3 161,6 << 0,01 Tachymenini Espécies 3 68 3,0 < 0,05 Uso do ambiente 1 6,0 < 0,05 Xenodontini Espécies 6 130 29,0 << 0,01 Uso do ambiente 1 58,1 << 0,01 Pseudoboini Espécies 4 97 6,0 < 0,01 Uso do ambiente 1 7,9 < 0,01
Tabela 15. Resultados da análise de covariância para as comparações do comprimento da cauda (CC) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 444,5 << 0,01 Uso do ambiente 2 118,5 << 0,01 Elapinae Espécies 3 88 270,2 << 0,01 Uso do ambiente 1 122,7 << 0,01 Xenodontinae Espécies 28 514 402,5 << 0,01 Uso do ambiente 3 219,2 << 0,01 Hydropsini Espécies 15 270 477,4 << 0,01 Uso do ambiente 3 302,6 << 0,01 Tachymenini Espécies 3 68 127,8 << 0,01 Uso do ambiente 1 1,0 = 0,31 Xenodontini Espécies 6 130 228,0 << 0,01 Uso do ambiente 1 0,16 = 0,69 Pseudoboini Espécies 4 97 59,6 << 0,01 Uso do ambiente 1 69,5 << 0,01

55
Tabela 16. Resultados da análise de covariância para as comparações do comprimento da cabeça (Ccab) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 114,6 << 0,01 Uso do ambiente 2 72,6 << 0,01 Elapinae Espécies 3 88 162,5 << 0,01 Uso do ambiente 1 172,9 << 0,01 Xenodontinae Espécies 28 514 210,4 << 0,01 Uso do ambiente 3 94,2 << 0,01 Hydropsini Espécies 15 270 229,1 << 0,01 Uso do ambiente 3 103,2 << 0,01 Tachymenini Espécies 3 68 115,6 << 0,01 Uso do ambiente 1 14,3 << 0,01 Xenodontini Espécies 6 130 102,9 << 0,01 Uso do ambiente 1 2,0 = 0,16 Pseudoboini Espécies 4 97 191,9 << 0,01 Uso do ambiente 1 306,1 << 0,01
Tabela 17. Resultados da análise de covariância para as comparações da largura da cabeça (Lcab) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 80,3 << 0,01 Uso do ambiente 2 17,6 << 0,01 Elapinae Espécies 3 88 101,0 << 0,01 Uso do ambiente 1 190,9 << 0,01 Xenodontinae Espécies 28 514 135,7 << 0,01 Uso do ambiente 3 89,7 << 0,01 Hydropsini Espécies 15 270 152,7 << 0,01 Uso do ambiente 3 115,1 << 0,01 Tachymenini Espécies 3 68 75,0 << 0,01 Uso do ambiente 1 7,6 < 0,05 Xenodontini Espécies 6 130 37,2 << 0,01 Uso do ambiente 1 0,2 = 0,67 Pseudoboini Espécies 4 97 214,5 << 0,01 Uso do ambiente 1 431,5 << 0,01
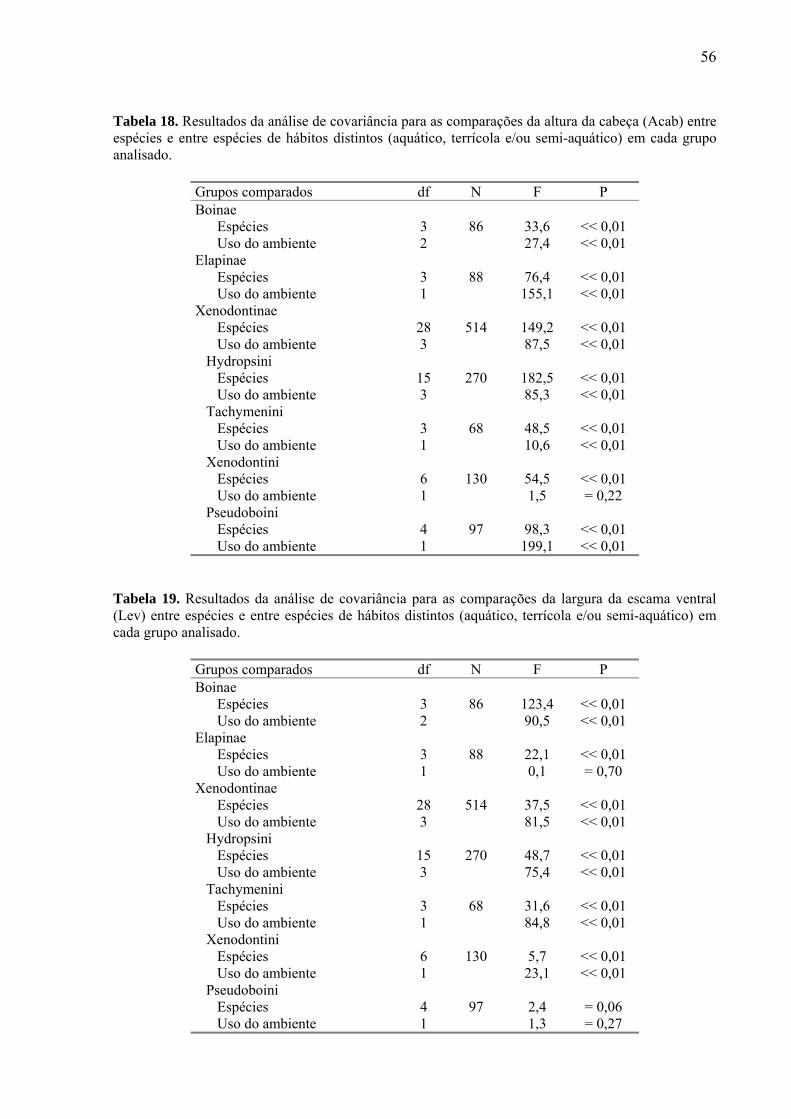
56
Tabela 18. Resultados da análise de covariância para as comparações da altura da cabeça (Acab) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 33,6 << 0,01 Uso do ambiente 2 27,4 << 0,01 Elapinae Espécies 3 88 76,4 << 0,01 Uso do ambiente 1 155,1 << 0,01 Xenodontinae Espécies 28 514 149,2 << 0,01 Uso do ambiente 3 87,5 << 0,01 Hydropsini Espécies 15 270 182,5 << 0,01 Uso do ambiente 3 85,3 << 0,01 Tachymenini Espécies 3 68 48,5 << 0,01 Uso do ambiente 1 10,6 << 0,01 Xenodontini Espécies 6 130 54,5 << 0,01 Uso do ambiente 1 1,5 = 0,22 Pseudoboini Espécies 4 97 98,3 << 0,01 Uso do ambiente 1 199,1 << 0,01
Tabela 19. Resultados da análise de covariância para as comparações da largura da escama ventral (Lev) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 123,4 << 0,01 Uso do ambiente 2 90,5 << 0,01 Elapinae Espécies 3 88 22,1 << 0,01 Uso do ambiente 1 0,1 = 0,70 Xenodontinae Espécies 28 514 37,5 << 0,01 Uso do ambiente 3 81,5 << 0,01 Hydropsini Espécies 15 270 48,7 << 0,01 Uso do ambiente 3 75,4 << 0,01 Tachymenini Espécies 3 68 31,6 << 0,01 Uso do ambiente 1 84,8 << 0,01 Xenodontini Espécies 6 130 5,7 << 0,01 Uso do ambiente 1 23,1 << 0,01 Pseudoboini Espécies 4 97 2,4 = 0,06 Uso do ambiente 1 1,3 = 0,27

57
Tabela 20. Resultados da análise de covariância para as comparações da circunferência do corpo (Circ) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 83,0 << 0,01 Uso do ambiente 2 53,0 << 0,01 Elapinae Espécies 3 88 30,1 << 0,01 Uso do ambiente 1 70,8 << 0,01 Xenodontinae Espécies 28 514 73,3 << 0,01 Uso do ambiente 3 97,3 << 0,01 Hydropsini Espécies 15 270 85,5 << 0,01 Uso do ambiente 3 117,5 << 0,01 Tachymenini Espécies 3 68 10,3 << 0,01 Uso do ambiente 1 9,4 << 0,01 Xenodontini Espécies 6 130 16,5 << 0,01 Uso do ambiente 1 0,1 = 0,77 Pseudoboini Espécies 4 97 60,4 << 0,01 Uso do ambiente 1 226,1 << 0,01
Tabela 21. Resultados da análise de covariância para as comparações massa do corpo (M) entre espécies e entre espécies de hábitos distintos (aquático, terrícola e/ou semi-aquático) em cada grupo analisado.
Grupos comparados df N F P Boinae Espécies 3 86 150,6 << 0,01 Uso do ambiente 2 107,8 << 0,01 Elapinae Espécies 3 88 23,4 << 0,01 Uso do ambiente 1 60,2 << 0,01 Xenodontinae Espécies 28 514 53,7 << 0,01 Uso do ambiente 3 58,2 << 0,01 Hydropsini Espécies 15 270 49,0 << 0,01 Uso do ambiente 3 33,5 << 0,01 Tachymenini Espécies 3 68 16,9 << 0,01 Uso do ambiente 1 7,4 < 0,05 Xenodontini Espécies 6 130 28,6 << 0,01 Uso do ambiente 1 9,2 << 0,01 Pseudoboini Espécies 4 97 59,3 << 0,01 Uso do ambiente 1 191,2 << 0,01

58
4.2.3 Posição e espaço ocupado pela ninhada e fecundidade
Dados de posição e espaço ocupado pela ninhada, bem como fecundidade, foram
obtidos para 160 indivíduos de 28 espécies de Elapinae e Xenodontinae (Tabela 22). Dentro
de Elapinae, embora comparações estatísticas não sejam possíveis devido ao pequeno número
amostral, Micrurus surinamensis apresentou fecundidade e comprimento relativo da ninhada
semelhante ao das outras espécies, porém a ninhada nessa espécie ocupa posição mais anterior
no corpo da fêmea (Figura 23).
Dentro de Xenodontinae, espécies aquáticas e terrícolas diferiram significativamente
em relação à posição ocupada pela ninhada (Figura 22 e Tabela 23), a qual está situada mais
posteriormente nas serpentes aquáticas. Essas espécies não diferiram em relação à
fecundidade ou comprimento relativo da ninhada (Figura 22 e Tabela 23). Xenodontíneos
fossóreos (não incluídos nos testes devido ao pequeno número amostral) apresentaram
ninhadas menores e que ocuparam, em média, apenas 24% do corpo das fêmeas. Serpentes
semi-aquáticas (também não incluídas nos testes) apresentaram ninhadas de tamanho e
posição intermediária a espécies aquáticas e terrícolas (Figura 22).
Em comparação aos representantes de Pseudoboini e Leptodeira annulata (todos
terrícolas), as espécies de Hydropsini (todas aquáticas) apresentaram maior fecundidade e
comprimento relativo da ninhada (Figura 26 e Tabela 23). A ninhada dos Hydropsini também
tende a se estender para a região posterior do corpo (Figura 26 e Tabela 23). Hydrodynastes
gigas (não incluída nos testes) apresentou ninhadas relativamente maiores, que se estendem
para a região anterior do corpo, em comparação aos representantes de Pseudoboini analisados
(Figura 26). Dentro de Xenodontini e Tachymenini, embora distinções sejam evidentes entre
as espécies (Figuras 24 e 25), serpentes aquáticas e terrícolas apresentaram fecundidade e
comprimento relativo da ninhada semelhante e não variaram significativamente em nenhuma
das características testadas (Tabela 23).

59
Tabela 22. Valores médios do comprimento rostro-cloacal (CRCm), fecundidade (Fec) e das razões obtidas, em relação ao CRC, para o comprimento anterior à ninhada (CAN), comprimento da ninhada (CN) e comprimento posterior à ninhada (CPN) para as espécies analisadas. N = tamanho da amostra. UA = ambiente predominantemente utilizado: A = aquático, SA = semi-aquático, T = terrícola e F = fossóreo. MR = modo reprodutivo: O = ovípara, V = vivípara. Medidas em milímetros.
Grupos/Espécies N MR Fec Espaço ocupado pela ninhada
UA CRCm
CAN CN CPN Elapinae Micrurus frontalis 1 F 912 O 8 0,74 0,22 0,04 M. lemniscatus 2 F 967,5 O 4 0,76 0,15 0,09 M. surinamensis 1 A 1040 O 4 0,60 0,26 0,13 M. corallinus 5 F 801,8 O 9 0,68 0,26 0,06 Xenodontinae Hydropsini Helicops modestus 25 A 438 V 13 0,59 0,35 0,06 H. carinicaudus 2 A 662,5 V 12 0,60 0,30 0,09 H. infrataeniatus 13 A 535,7 V 14 0,58 0,35 0,07 H. angulatus 3 A 556,3 O 13 0,52 0,37 0,11 H. polylepis 4 A 726 V 11 0,60 0,32 0,07 Pseudoeryx plicatilis 1 A 1.115 O 33 0,57 0,37 0,06 Elapomorphini Elapomorphus quinquelineatus 2 F 740 O 6 0,69 0,27 0,07 Phalotris mertensi 1 F 962 O 7 0,63 0,29 0,08 Tachymenini Thamnodynastes strigatus 13 T 525 V 16 0,46 0,44 0,10 Tomodon dorsatus 13 T 517 V 12 0,45 0,43 0,11 Gomesophis brasiliensis 4 A 357 V 6 0,51 0,35 0,15 Ptychophis flavovirgatus 6 A 370 V 7 0,45 0,46 0,09 Xenodontini Liophis cobella 1 A 440 O 4 0,61 0,35 0,04 L. miliaris 9 A 774 O 13 0,55 0,34 0,11 L. poecilogyrus 3 T 525 O 9 0,57 0,29 0,14 L. reginae 5 T 515 O 4 0,59 0,28 0,14 Erythrolamprus aesculapii 5 T 778 O 6 0,69 0,25 0,07 Xenodon neuwiedii 5 T 638 O 8 0,57 0,31 0,12 Pseudoboini Oxyrhopus guibei 10 T 944,5 O 11 0,59 0,32 0,09 Pseudoboa nigra 1 T 980 O 6 0,63 0,27 0,11 Clelia quimi 4 T 965 O 10 0,59 0,34 0,07 Outros Xenodontinae Sordellina punctata 1 SA 390 O 3 0,64 0,23 0,12 Hydrodynastes gigas 2 SA 1.260 O 13 0,50 0,39 0,10 Dipsadinae Leptodeira annulata 4 T 571 O 7 0,62 0,29 0,09

60
Semi-aquáticas
Terrestres
Aquáticas
Fossóreas
Semi-aquáticasSemi-aquáticas
TerrestresTerrestres
AquáticasAquáticas
FossóreasFossóreas
Figura 22. Variação média da posição e espaço relativo ocupado pela ninhada em serpentes de diferentes hábitos dentro de Xenodontinae.
M. corallinus
M. lemniscatus
M. frontalis
M. surinamensis
M. corallinusM. corallinus
M. lemniscatusM. lemniscatus
M. frontalisM. frontalis
M. surinamensisM. surinamensis
Figura 23. Variação média da posição e espaço relativo ocupado pela ninhada em serpentes de diferentes hábitos dentro de Elapinae.
Rostro Cloaca
Rostro Cloaca

61
L. miliaris
L. poecilogyrus
L. reginae
E. aesculapii
L. cobella
X. neuwiedii
L. miliarisL. miliaris
L. poecilogyrusL. poecilogyrus
L. reginaeL. reginae
E. aesculapiiE. aesculapii
L. cobellaL. cobella
X. neuwiediiX. neuwiedii
Figura 24. Variação média da posição e espaço relativo ocupado pela ninhada em serpentes de diferentes hábitos dentro de Xenodontini.
T. strigatus
T. dorsatus
G. brasiliensis
P. flavovirgatus
T. strigatusT. strigatus
T. dorsatusT. dorsatus
G. brasiliensisG. brasiliensis
P. flavovirgatusP. flavovirgatus
Figura 25. Variação média da posição e espaço relativo ocupado pela ninhada em serpentes de diferentes hábitos dentro de Tachymenini.
Rostro Cloaca
Rostro Cloaca

62
H. carinicaudus
H. infrataeniatus
H. polylepis
H. modestus
H. angulatus
P. plicatilis
H. carinicaudusH. carinicaudus
H. infrataeniatusH. infrataeniatus
H. polylepisH. polylepis
H. modestusH. modestus
H. angulatusH. angulatus
P. plicatilisP. plicatilis
P. nigra
C. quimi
O. guibei
H. gigas
P. nigraP. nigra
C. quimiC. quimi
O. guibeiO. guibei
H. gigasH. gigas
E. quinquelineatus
P. mertensi
L. annulata
S. punctata
E. quinquelineatusE. quinquelineatus
P. mertensiP. mertensi
L. annulataL. annulata
S. punctataS. punctata
Figura 26. Variação média da posição e espaço relativo ocupado pela ninhada em espécies de Hydropsini em comparação a representantes de Pseudoboini e outras espécies de Xenodontinae.
Rostro Cloaca

63
Tabela 23. Resultados da análise de covariância para as comparações das medidas de posição e espaço ocupado pela ninhada no corpo das fêmeas, bem como fecundidade, entre espécies de hábitos distintos (aquáticas e terrícolas) em cada grupo analisado de Xenodontinae. CAN = comprimento anterior à ninhada, CN = comprimento da ninhada, CPN = comprimento posterior à ninhada, Fec = Fecundidade.
Caracteres e grupos comparados df N F P CAN Todas as espécies Uso do ambiente 1 154 14,90 << 0,01 Hydropsini Uso do ambiente 1 71 3,33 = 0,07 Tachymenini Uso do ambiente 1 49 0,25 = 0,61 Xenodontini Uso do ambiente 1 28 2,88 = 0,10 CN Todas as espécies Uso do ambiente 1 154 0,22 = 0,64 Hydropsini Uso do ambiente 1 71 5,64 < 0,05 Tachymenini Uso do ambiente 1 49 0,07 = 0,80 Xenodontini Uso do ambiente 1 28 2,92 = 0,10 CPN Todas as espécies Uso do ambiente 1 154 32,70 << 0,01 Hydropsini Uso do ambiente 1 71 6,14 < 0,05 Tachymenini Uso do ambiente 1 49 0,62 = 0,43 Xenodontini Uso do ambiente 1 28 0,04 = 0,84 Fec Todas as espécies Uso do ambiente 1 154 3,61 = 0,06 Hydropsini Uso do ambiente 1 71 34,86 << 0,01 Tachymenini Uso do ambiente 1 49 4,21 = 0,10 Xenodontini Uso do ambiente 1 28 1,97 = 0,17

64
4.3 Análises comparativas
Depois de eliminado o efeito da filogenia sobre o tamanho e forma do corpo entre
representantes de Xenodontinae, foram encontradas correlações negativas e significativas
entre os contrastes independentes de uso do ambiente (freqüência de utilização do ambiente
aquático) e aqueles calculados para as distâncias relativas entre as narinas (r = - 0,50 P < 0,05
N = 23) e entre os olhos (r = -0,47 P < 0,05 N = 23) (Figura 27 a e b). Correlações positivas e
significativas foram encontradas entre os contrastes de uso do ambiente e aqueles obtidos para
as medidas relativas de massa (r = 0,44 P < 0,05 N = 23) e circunferência do corpo (r = 0,44 P
< 0,05 N = 23) (Figura 27 c e d). Esses resultados indicam que xenodontíneos aquáticos
possuem as narinas e os olhos mais próximos entre si, no dorso da cabeça, e são mais
robustas, assim como demonstrado nas análises estatísticas convencionais, nas quais o efeito
da filogenia não foi eliminado.
Uma correlação negativa e marginalmente significativa foi encontrada entre os
contrastes da largura relativa da escama ventral e aqueles do uso do ambiente (r = -0,36 P =
0,09 N = 23), indicando que escamas ventrais relativamente mais estreitas podem estar
associadas a hábitos aquáticos entre os xenodontíneos. Não foram encontradas correlações
significativas entre os contrastes de uso do ambiente e aqueles calculados para o comprimento
do corpo (r = -0,09 P = 0,69 N = 23), distância relativa entre o rostro e as narinas (r = -0,23 P
= 0,28 N = 23), entre o rostro e os olhos (r = -0,18 P = 0,41 N = 23), diâmetro relativo do olho
(r = -0,29 P = 0,17 N = 23), comprimento, largura e altura relativa da cabeça (r = 0,21 P =
0,33 N = 23; r = 0,22 P = 0,32 N = 23; r = 0,24 P = 0,27 N = 23; respectivamente) e
comprimento relativo da cauda (r = 0,22 P = 0,31 N = 23).

65
Figura 27. Correlações entre os contrastes independentes de uso do ambiente (PA) e aqueles calculados para as distâncias relativas entre as narinas (a), entre os olhos (b), massa (c) e circunferência relativa do corpo (d) para as 24 espécies de Xenodontinae analisadas.
Os resultados das otimizações do uso do ambiente e forma relativa do corpo nas
espécies de Boinae analisadas são apresentados nas figuras 28, 29 e 30. Seus ancestrais
hipotéticos eram terrícolas, com tamanho da cauda e largura relativa da escama ventral de
valores intermediários (Figura 28). A cauda aumentou em Corallus hortulanus e a largura da
escama ventral sofreu diminuição em Eunectes murinus. Seus ancestrais também possuíam
valores intermediários para as distâncias relativas entre as narinas, entre os olhos e entre o
rostro e os olhos, com diminuição em Eunectes murinus (Figura 29). A distância relativa entre
o rostro e as narinas manteve-se constante entre as espécies, porém aumentou em Boa
constrictor (Figura 29). O diâmetro do olho aumentou em Corallus hortulanus e sofreu
(a)
(b)
(c)
(d)

66
diminuição em Boa constrictor (Figura 30). O comprimento da cabeça, circunferência e
massa relativa do corpo provavelmente sofreram redução independente em Corallus
hortulanus e Epicrates cenchria (Figura 30).
As otimizações para o uso do ambiente, tamanho e forma do corpo nas espécies de
Elapinae são apresentadas nas figuras 31, 32 e 33. Seus ancestrais eram fossóreos e de
tamanho de corpo moderado. O CRC aumentou em Micrurus lemniscatus e diminuiu em M.
surinamensis (Figura 31). A largura da escama ventral diminuiu e o comprimento da cauda
aumentou em Micrurus surinamensis (Figura 31). Seus ancestrais possuíam valores
intermediários para as distâncias relativas entre as narinas, entre os olhos e entre o rostro e as
narinas, com diminuição em M. surinamensis (Figura 32). A distância entre o rostro e os
olhos manteve-se constante em M. corallinus e M. surinamensis (e seus ancestrais) e
aumentou em M. lemniscatus e M. frontalis (Figura 32 d). O diâmetro do olho aumentou em
M. frontalis e diminuiu em M. lemniscatus. O tamanho da cabeça, circunferência e massa
relativa do corpo sofreram aumento em M. surinamensis (Figura 33).
Os resultados das otimizações do uso do ambiente, tamanho e forma do corpo para as
espécies de Xenodontinae são apresentados nas figuras 34, 35 e 36. Apesar da falta de
resolução para muitos estados ancestrais em muitos dos caracteres, os resultados indicam que
os ancestrais hipotéticos de cada linhagem eram terrícolas e que o hábito aquático surgiu pelo
menos quatro vezes (Figura 34). Os ancestrais de cada linhagem eram de tamanho moderado a
grande, com aumento ou diminuição em cada grupo, independente do tipo de ambiente
utilizado (Figura 34).
Os ancestrais hipotéticos de cada linhagem de Xenodontinae apresentavam caudas de
tamanho intermediário, com diminuição em Gomesophis brasiliensis e Erythrolamprus
aesculapii e aumento em Helicops angulatus, H. polylepis, Hydrodynastes gigas e Liophis
reginae (Figura 35a). Apesar das indefinições dos estados ancestrais para a largura relativa da

67
escama ventral, as otimizações indicam que essa variável tendeu a sofrer redução em
Hydropsini e em representantes aquáticos de Tachymenini e Xenodontini (Figura 35b).
As distâncias relativas entre as narinas e entre os olhos tenderam a sofrer redução pelo
menos entre os Hydropsini e representantes aquáticos de Tachymenini (Figuras 36a e 37a). As
distâncias relativas entre o rostro e as narinas e entre o rostro e os olhos parecem ter
permanecido constantes nas diferentes linhagens, porém sofreram redução nos Hydropsini.
(Figuras 36b e 37b).
Ancestrais hipotéticos de cada linhagem de Xenodontinae apresentavam o olho de
diâmetro moderado a grande, com redução nos Hydropsini e pequena diminuição em seus
valores relativos em outras linhagens com representantes aquáticos (Tachymenini e
Xenodontini) (Figura 38a). Os ancestrais dos grupos analisados apresentavam cabeça de
tamanho moderado, com aumento em alguns Hydropsini, Hydrodynastes gigas e
Thamnodynastes strigatus e redução em Hydrops triangularis, Gomesophis brasiliensis,
Erythrolamprus aesculapii, Liophis frenata e espécies de Pseudoboini (Figura 38b).
Os ancestrais das linhagens analisadas eram medianamente robustos (Figura 39). A
circunferência e a massa relativa do corpo tenderam aumentar em alguns Hydropsini,
Hydrodynastes gigas, representantes aquáticos de Tachymenini e Thamnodynastes strigatus e
sofrer redução em Hydrops triangularis e em algumas espécies de Xenodontini (Figura 39).
68
Figura 28. Otimização do uso do ambiente (proporção de indivíduos encontrados no ambiente aquático) (a), largura relativa da escama ventral (b) e comprimento relativo da cauda (c) na hipótese filogenética das 04 espécies de Boinae analisadas. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(a)
(c) (b)

69
Figura 29. Otimização das distâncias relativas entre as narinas (a), entre o rostro e as narinas (b), entre os olhos (c) e entre o rostro e os olhos (d) na hipótese filogenética das 04 espécies de Boinae analisadas. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(a) (c)
(b) (d)

70
Figura 30. Otimização do diâmetro relativo do olho (a), comprimento relativo da cabeça (b), massa (c) e circunferência relativa do corpo (d) na hipótese filogenética das 04 espécies de Boinae analisadas. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(a)
(d) (b)
(c)
Kg
Kg

71
Figura 31. Otimização do tipo de ambiente utilizado (freqüência de utilização do ambiente aquático) (a), logaritmo do comprimento rostro-cloacal (b), largura relativa da escama ventral (c) e comprimento relativo da cauda (d) na hipótese filogenética das 04 espécies de Elapinae analisadas. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(a)
(b)
(c)
(d)

72
Figura 32. Otimização das distâncias relativas entre as narinas (a), entre o rostro e as narinas (b), entre os olhos (c) e entre o rostro e os olhos (d) na hipótese filogenética das 04 espécies de Elapinae analisadas. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(a) (c)
(b) (d)

73
Figura 33. Otimização do diâmetro relativo do olho (a), comprimento relativo da cabeça (b), massa (c) e circunferência relativa do corpo (d) na hipótese filogenética das 04 espécies de Elapinae analisadas. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(a)
(d) (b)
(c)
Kg
Kg

74
Figura 34. Otimização do tipo de ambiente utilizado (freqüência de utilização do ambiente aquático) (a) e logaritmo do comprimento rostro-cloacal (b) na hipótese filogenética das espécies de Xenodontinae analisadas, com Heterodon platyrhinus e Leptodeira annulata como grupo externo. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(a)
(b)

75
Figura 35. Otimização do comprimento relativo da cauda (a) e da largura relativa da escama ventral (b) na hipótese filogenética das espécies de Xenodontinae analisadas, com Heterodon platyrhinus e Leptodeira annulata como grupo externo. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(a)
(b)

76
Figura 36. Otimização das distâncias relativas entre as narinas (a) e entre o rostro e as narinas (b) na hipótese filogenética das espécies de Xenodontinae analisadas, com Heterodon platyrhinus e Leptodeira annulata como grupo externo. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(b)
(a)

77
Figura 37. Otimização das distâncias relativas entre os olhos (a) e entre o rostro e os olhos (b) na hipótese filogenética das espécies de Xenodontinae analisadas, com Heterodon platyrhinus e Leptodeira annulata como grupo externo. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(b)
(a)

78
Figura 38. Otimização do diâmetro relativo dos olhos (a) e comprimento relativo da cabeça (b) na hipótese filogenética das espécies de Xenodontinae analisadas, com Heterodon platyrhinus e Leptodeira annulata como grupo externo. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(a)
(b)
79
Figura 39. Otimização da massa (a) e circunferência relativa do corpo (b) na hipótese filogenética das espécies de Xenodontinae analisadas, com Heterodon platyrhinus e Leptodeira annulata como grupo externo. Ramos mais escuros indicam maiores valores relativos (resíduos das regressões) para essas variáveis; ramos rachurados indicam estados duvidosos. Os valores nos ramos terminais e intermediários são as médias dos resíduos obtidos para essas variáveis.
(a)
Kg
Kg
(b)

80
5 - DISCUSSÃO
Apesar da falta de informações detalhadas para algumas serpentes, a freqüência e tipo de
ambiente utilizado foram variáveis entre as espécies analisadas. O hábito aquático surgiu apenas
uma vez entre os Boinae, nas serpentes do gênero Eunectes, a despeito da grande diversidade de
hábitos encontrada nos outros gêneros da subfamília (MURPHY; HENDERSON, 1997). O tipo de
ambiente utilizado parece ser conservativo dentro de algumas linhagens. Por exemplo, hábitos
fossóreos são predominantes entre as mais de 50 espécies existentes de Micrurus (CAMPBELL;
LAMAR, 2004; ROZE, 1996). Embora algumas espécies do gênero possam estar eventualmente
associadas a ambientes aquáticos (e.g., M. lemniscatus; BEEBE, 1946), Micrurus surinamensis é a
única serpente integralmente aquática dentro da subfamília (MARTINS; OLIVEIRA, 1999; ROZE,
1996).
Dentro de Xenodontinae, o hábito aquático também é conservativo entre as espécies de
Hydropsini. O uso do ambiente aquático surgiu pelo menos outras três vezes na subfamília: entre
os Tachymenini (Ptychophis flavovirgatus e Gomesophis brasiliensis) e provavelmente duas
vezes entre os Xenodontini, nas espécies do gênero Liophis (Liophis frenata – L. miliaris e em L.
cobella). Entretanto, informações detalhadas de uso do ambiente para outras espécies de Liophis
(gênero composto por aproximadamente 40 espécies; DIXON, 1980) ainda são escassas ou
inexistentes. Informações dessa natureza serão importantes para determinar o número de vezes
em que o hábito aquático surgiu dentro do grupo.
Hábitos semi-aquáticos são características derivadas nas serpentes Hydrodynastes gigas e
Sordellina punctata. Embora existam informações limitadas para caracterizar a utilização do
ambiente por Hydrodynastes bicinctus, as informações disponíveis para H. gigas permitem
algumas inferências. Essa última espécie inclui peixes na dieta e utiliza tanto o ambiente aquático

81
quanto o terrícola (STRÜSSMANN; SAZIMA, 1993), o que também pode ser esperado para H.
bicinctus, que se alimenta de peixes e crustáceos decápodes (CUNHA; NASCIMENTO, 1978).
Os resultados indicam que o tamanho do corpo não parece estar relacionado ao uso do
ambiente aquático. Apesar de algumas serpentes aquáticas apresentarem menor tamanho corporal
que espécies relacionadas (e.g., Micrurus surinamensis, Gomesophis brasiliensis, Ptychophis
flavovirgatus e algumas espécies de Helicops), outras (e.g., Pseudoeryx plicatilis) apresentaram
tamanho próximo daquelas que usam outros ambientes. Entre as serpentes semi-aquáticas, as
duas espécies de Hydrodynastes analisadas foram as maiores da amostra. Entre os Boinae,
embora não tenha sido possível medir o tamanho do corpo em indivíduos adultos, Eunectes
murinus também atinge maior porte que as outras espécies da subfamília (MURPHY; HENDERSON,
1997).
Entre as espécies fossóreas de Elapomorphini, Apostolepis assimilis apresentou menor
tamanho corporal que Elapomorphus quinquelineatus e Phalotris mertensi. A tendência na
redução do tamanho do corpo em serpentes fossóreas foi sugerida por Cadle e Greene (1993).
Apostolepis assimilis é provavelmente a forma mais especializada ao hábito subterrâneo entre os
Elapomorphini (FERRAREZZI, 1993) e apresenta tal característica.
Entretanto, o tamanho do corpo em serpentes pode estar relacionado à ação de outros
agentes seletivos (e.g., um corpo maior pode comportar ninhadas e/ou capturar e ingerir presas
relativamente maiores ou mais robustas; MARTINS; MARQUES; SAZIMA, 2002; SHINE, 1991,
1994b).
Entre as espécies de Boinae analisadas, Boa constrictor pode se alimentar de aves e
pequenos mamíferos e apresenta menor tamanho corporal que Eunectes murinus, espécie que
inclui grandes répteis e mamíferos em sua dieta (e.g., jacarés e capivaras; MURPHY; HENDERSON,
1997; L. PIZZATO, informação pessoal). Entre os Tachymenini, Gomesophis brasiliensis se

82
alimenta de minhocas (OLIVEIRA; BORGES; MARQUES, 2003) e é menor que Thamnodynastes
strigatus, que preda vertebrados (BERNARDE et al., 2000; RUFFATO; DI-BERNARDO; MASCHIO,
2003). Porém, Ptychophis flavovirgatus, espécie-irmã de G. brasiliensis (FRANCO, 1999),
também apresenta pequeno tamanho corporal e aparentemente se alimenta de peixes ou anfíbios
(no tubo digestivo de alguns indivíduos foram encontrados vestígios de pequenos artrópodes e
fibras vegetais - provavelmente itens secundários decorrentes da ingestão desses vertebrados;
observação pessoal).
Os resultados indicam que determinadas características da forma do corpo nas espécies
estudadas estão relacionadas à evolução do uso do ambiente aquático. Tais características foram
encontradas em serpentes aquáticas pertencentes a quatro linhagens independentes: (1) Eunectes
murinus, (2) Micrurus surinamensis, (3) Gomesophis brasiliensis e Ptychophis flavovirgatus e
(4) representantes de Hydropsini. Essas características incluem: (a) as narinas e os olhos
relativamente mais próximos entre si, na região dorso-anterior da cabeça, (b) maior robustez
(massa e circunferência relativa do corpo) e (c) menor largura relativa da escama ventral. As
serpentes aquáticas do gênero Liophis, embora sejam mais robustas e apresentem menor largura
da escama ventral, não diferiram quanto ao posicionamento dos olhos e narinas em comparação
às espécies terrícolas relacionadas, indicando que essas características são conservativas dentro
do grupo. Entre as espécies semi-aquáticas, Sordellina punctata ocupou posição intermediária
entre espécies aquáticas e aquelas que utilizam outros ambientes quanto ao posicionamento dos
olhos e narinas, bem como para a largura da escama ventral, mas não é uma espécie robusta.
Hydrodynastes gigas e H. bicinctus são parecidas quanto à posição dos olhos e narinas, bem
como para a largura da escama ventral, em relação às espécies terrícolas, porém, são serpentes
mais robustas.

83
As narinas e os olhos mais próximos entre si e em posição mais anterior no dorso da
cabeça são características existentes em diversas linhagens de serpentes aquáticas, como em
Acrochordidae, outros Colubridae (e.g., Natricinae e Homalopsinae) e em alguns Elapidae
marinhos (CADLE; GREENE, 1993; GREENE, 1997; MARX; RABB, 1972). Serpentes aquáticas,
quando no interior de corpos d’água, podem emergir apenas pequena porção da cabeça para
poderem respirar, talvez ficando pouco expostas a predadores aéreos (GREENE, 1997). Assim, o
deslocamento das narinas e dos olhos para a região dorso-anterior da cabeça seria vantajoso nas
espécies que utilizam este ambiente (GREENE, 1997).
A redução na largura da escama ventral também é característica comum em outras
linhagens de serpentes aquáticas (GREENE, 1997). Em serpentes terrícolas, as forças propulsoras
durante a locomoção por ondulação lateral são transmitidas contra irregularidades do solo e as
escamas ventrais largas, geralmente em forma de grandes placas, possuem a função de diminuir a
fricção durante o deslocamento (GANS, 1974). No ambiente aquático, escamas ventrais menores
devem melhorar a performance para o deslocamento, talvez facilitando a locomoção por
ondulação lateral.
A maior robustez em serpentes aquáticas já havia sido sugerida por outros autores (cf.
GREENE, 1997; PARKER; GRANDISON, 1986) e relatada para algumas espécies neotropicais (cf.
MARQUES, 1998; MARTINS, 1994). As características físicas do ambiente aquático devem permitir
que os animais que vivem nesse meio apresentem maior robustez, uma vez que a demanda
energética para a natação é menor que aquela para a locomoção no ambiente terrestre, devido ao
menor esforço requerido para o suporte do corpo e pelo decréscimo da fricção (LILLYWHITE,
1987).
Contudo, a variação na robustez em serpentes também pode ser explicada pela ação de
outros agentes seletivos. Espécies especializadas em se alimentar de mamíferos (presas robustas)

84
tendem a possuir maior massa e circunferência relativa do corpo que espécies com dieta
generalista (MARTINS; MARQUES; SAZIMA, 2002; SHINE, 1991). Porém, independente do tipo de
alimento ingerido, as serpentes aquáticas analisadas apresentaram maior robustez que espécies
que usam outros ambientes, reforçando a hipótese de que a maior robustez nessas serpentes seja
devida às características físicas do ambiente aquático.
O tamanho da cabeça em muitas serpentes parece estar relacionado à ingestão de presas
robustas, sendo geralmente maior em espécies que se alimentam de mamíferos (ver GREENE,
1983; MARTINS; MARQUES; SAZIMA, 2002; POUGH; GROVES, 1983; SHINE, 1991). A ingestão de
peixes também parece ter conferido modificações semelhantes em algumas serpentes aquáticas.
Savitsky (1983) faz comentários sobre modificações no crânio de serpentes piscívoras
relacionadas à retenção e ingestão de suas presas. Os peixes possuem estrutura óssea rígida e a
ingestão de espécies com corpos fusiformes (largos ou altos) pode ter levado a modificações no
tamanho da cabeça em serpentes aquáticas. Recentemente, Vincent, Herrel e Irschick (2004)
estudaram o dimorfismo sexual em Agkistrodon piscivorus. Machos dessa espécie incluem peixes
na dieta com maior freqüência e possuem cabeças relativamente maiores que fêmeas. Quanto ao
tamanho relativo dos peixes ingeridos, machos também ingerem peixes mais robustos que
fêmeas.
Evidências de que a ingestão de diferentes tipos de peixes pode ter levado a modificações
no tamanho da cabeça em serpentes foram levantadas nesse estudo. Por exemplo, Micrurus
surinamensis pode se alimentar de várias espécies de peixes (ROZE, 1996) e possui cabeça
relativamente maior que as outras espécies analisadas do gênero e que se alimentam basicamente
de outros vertebrados alongados, como anfisbenídeos (e.g., M. frontalis e M. corallinus;
MARQUES, 1996; ROZE, 1996). Entre os Xenodontinae, o tamanho da cabeça também é maior em
espécies que incluem peixes robustos na dieta. Entre os Hydropsini, as espécies do gênero

85
Helicops analisadas e Pseudoeryx plicatilis se alimentam de ampla variedade de peixes (AGUIAR;
DI-BERNARDO, 2004; CUNHA; NASCIMENTO, 1978) e possuem cabeça maior que Hydrops
triangularis que se alimenta de peixes alongados (muçuns; ALBUQUERQUE; CAMARGO, 2004).
Liophis miliaris também pode se alimentar de vários tipos de peixes (O. A. V. MARQUES,
informação pessoal) e possui cabeça maior que as outras espécies do gênero e que se alimentam
de anfíbios (e.g., Liophis poecilogyrus; VITT, 1983), outras serpentes (e.g., Erythrolamprus
aesculapii; MARQUES; PUORTO, 1994) ou peixes alongados (e.g., Liophis frenata; observação
pessoal).
O comprimento da cauda nas serpentes analisadas foi bastante variado e não parece estar
relacionado ao uso do ambiente aquático. Embora um estudo com serpentes amazônicas tenha
indicado a tendência de espécies aquáticas apresentarem caudas maiores (cf. MARTINS, 1994), as
serpentes aquáticas aqui analisadas apresentaram, independente da linhagem a que pertencem,
caudas relativamente curtas (e.g., Gomesophis brasiliensis, Hydrops triangularis e Pseudoeryx
plicatilis) ou longas (Helicops angulatus, H. polylepis) em relação às espécies que utilizam outros
ambientes. Da mesma forma, serpentes semi-aquáticas apresentaram caudas curtas (Sordellina
punctata) ou longas (Hydrodynastes).
As serpentes fossóreas analisadas apresentaram caudas extremamente curtas, assim como
outras serpentes que utilizam este ambiente (CADLE; GREENE, 1993; MARX; RABB, 1972).
Gomesophis brasiliensis possui o hábito de se enterrar em meio a solo lamacento (AMARAL,
1932), o que poderia explicar a cauda curta nessa espécie.
Caudas longas em serpentes geralmente estão associadas às espécies que utilizam o
substrato arbóreo (LILLYWHITE; HENDERSON, 1993; MARTINS et al., 2001). Corallus hortulanus é
espécie arborícola e tem cauda maior que os outros Boinae analisados. Contudo, o tamanho da
cauda em serpentes pode refletir ação de outros agentes seletivos. Caudas extremamente longas

86
em serpentes não arborícolas, como observado para Helicops angulatus, H. polylepis,
Hydrodynastes spp., e Liophis reginae, podem estar relacionadas a outros fatores ecológicos
(e.g., estratégias de forrageio). Hydrodynastes gigas possui cauda longa e a utiliza para
esquadrinha o substrato à procura de anfíbios, dos quais pode se alimentar (ver STRÜSSMANN;
SAZIMA, 1990). O tamanho da cauda em serpentes também pode estar relacionado à defesa
(GREENE, 1988). Como constatado por Guyer e Donnelly (1990), espécies terrícolas com caudas
extremamente grandes tendem a apresentar maior freqüência na ocorrência de quebra de cauda
que espécies com caudas menores (ver também MARQUES; SAZIMA 2004; MARTINS; OLIVEIRA,
1999, para outros exemplos).
Modificações no diâmetro do olho devem estar associadas ao uso do ambiente
subterrâneo. As serpentes fossóreas apresentaram o diâmetro do olho reduzido, característica
comum a diversas outras espécies que utilizam esse ambiente (CADLE; GREENE, 1993; MARX;
RABB, 1972; SAVITSKY, 1983). Porém, todos os representantes de Hydropsini também
apresentaram redução no diâmetro dos olhos, ao passo que a maioria dos Xenodontini e
Tachymenini apresentou olhos relativamente grandes, indicando que esta característica pode ser
conservativa dentro dos grupos analisados. O olho muito reduzido em Hydrops triangularis
poderia ser explicado pelo tipo de ambiente utilizado para forrageio. Hydrops triangulares se
alimenta de peixes alongados (muçuns) que geralmente vivem enterrados (ALBUQUERQUE;
CAMARGO, 2004; BEEBE, 1946). Em regiões de igapó na Amazônia Central, essa espécie tem sido
freqüentemente observada deslocando-se entre a serapilheira acumulada no fundo de lagoas, bem
como sob e entre as ranhuras de casca de árvores na floresta inundada (M. MARTINS, informação
pessoal).
O uso do ambiente aquático não parece ter implicado em modificações importantes na
fecundidade, espaço e posição ocupada pela ninhada no corpo das fêmeas. Nas serpentes

87
aquáticas e terrícolas de Xenodontinae analisadas, a ninhada tende a ocupar o mesmo espaço e
posição relativa no corpo das fêmeas. Porém, a ninhada está em posição mais posterior nos
Hydropsini. Essa característica é contrária àquela constatada para outros grupos de serpentes do
sudeste da Ásia e Austrália (ver SHINE, 1988).
Em pelo menos quatro linhagens monofiléticas de serpentes onde ocorre o hábito aquático
(elapídeos marinhos das subfamílias Hrydrophiinae e Laticaudinae, colubrídeos Homalopsinae e
em Acrochordidae), a ninhada ocupa menor espaço (menor fecundidade e comprimento relativo
da ninhada) e está em posição mais anterior no corpo das fêmeas (maior espaço posterior à
ninhada) em relação a espécies terrícolas relacionadas (SHINE, 1988). Dentre as espécies
analisadas no presente estudo, somente Micrurus surinamensis e Gomesophis brasiliensis
parecem obedecer a esse padrão.
Carregar os filhotes ou ovos pode ser desfavorável se a região do corpo usada para isso
for importante na locomoção (SHINE, 1988). A amplitude e a forma das ondas do corpo de uma
serpente aquática, durante a natação, aumentam da região anterior para a posterior, sendo esta
última porção responsável em gerar a propulsão (JAYNE, 1985). Isto não ocorre em serpentes
terrícolas, onde as ondulações laterais do corpo da serpente são mais regulares e a propulsão
transmitida contra irregularidades do solo (GANS, 1974). Portanto, ter a ninhada menor e
posicionada na região anterior do corpo deve facilitar a natação e tal característica foi
interpretada como uma adaptação à vida aquática nas serpentes analisadas por Shine (1988).
Algumas serpentes analisadas no presente estudo (e.g., Helicops spp., Liophis miliaris,
Ptychophis flavovirgatus e Hydrodynastes gigas) utilizam ambientes aquáticos pouco profundos
(e.g., brejos ou regiões alagadas e pequenas lagoas; LEMA; DEIQUES, 1992; MARQUES, 1998;
MARTINS; OLIVEIRA, 1999; STRÜSSMANN; SAZIMA, 1992) e a região posterior do corpo, nestas
serpentes, talvez não seja a mais importante em gerar propulsão durante a locomoção. Por isso, a

88
ocupação da região posterior do corpo com a ninhada poderia não ter implicações importantes à
locomoção dessas serpentes. Entretanto, estudos que analisem a locomoção dessas espécies em
ambientes aquáticos são necessários para confirmar ou não essa hipótese.
Nas linhagens analisadas por Shine (1988), espécies aquáticas e terrícolas não diferem em
relação ao volume disponível na cavidade abdominal (SHINE, 1992). Portanto, o espaço
disponível no abdômen das fêmeas não seria restrição para o investimento reprodutivo nessas
linhagens (SHINE, 1992). Se as serpentes analisadas no presente estudo não estão sujeitas às
forças seletivas relacionadas à acomodação da ninhada na região posterior do corpo, por outro
lado, elas são relativamente mais robustas (maior massa e circunferência relativa do corpo) que
espécies que utilizam outros ambientes, o que também poderia permitir a acomodação de
ninhadas maiores.

89
6 - CONCLUSÕES
1. A freqüência no uso do ambiente aquático variou entre as espécies analisadas, dentro
de cada linhagem.
2. Entre as serpentes estudadas, hábitos aquáticos parecem ter evoluído pelo menos
quatro vezes entre os Xenodontinae (em Hydropsini, entre os Tachymenini e duas
vezes dentro do gênero Liophis) e uma vez em Elapinae (Micrurus surinamensis) e
Boinae (Eunectes murinus).
3. Os resultados indicam que a evolução do hábito aquático foi acompanhada de
mudanças na forma do corpo entre as espécies, dentro de cada linhagem. Essas
mudanças incluem os olhos e as narinas deslocadas para a região dorso-anterior da
cabeça, maior robustez (massa e circunferência do corpo) e menor largura da escama
ventral.
4. O tamanho do corpo não parece estar relacionado ao uso do ambiente aquático, mas
sim a outros fatores, como dieta e/ou reprodução.
5. Outras variáveis da forma do corpo não parecem estar relacionadas aos hábitos
aquáticos nas serpentes analisadas, incluindo o diâmetro do olho, o tamanho da cabeça
e o comprimento da cauda.
6. O menor diâmetro dos olhos está relacionado à utilização do ambiente subterrâneo nas
espécies aqui estudadas (representantes de Elapomorphini e Elapinae), assim como
constatado em outras serpentes fossóreas.
7. O tamanho da cabeça (comprimento, largura e altura) parece estar relacionado ao tipo
de alimento ingerido pelas serpentes analisadas. Cabeças pequenas estão relacionadas
à ingestão de invertebrados ou vertebrados de corpo alongado. Cabeças grandes estão

90
relacionadas à ingestão de presas robustas (mamíferos) ou com corpos fusiformes
(e.g., peixes).
8. Nas espécies analisadas, o menor comprimento da cauda está relacionado ao hábito
fossóreo, assim como constatado em outras serpentes.
9. Caudas longas, como observado em algumas serpentes aquáticas (Helicops angulatus,
H. polylepis, Hydrodynastes e Liophis reginae) podem estar relacionadas a táticas de
forrageio ou defensivas.
10. Nas espécies estudadas, os hábitos aquáticos parecem não ter levado a modificações
na fecundidade e no espaço ocupado pela ninhada, diferentemente do observado para
outras serpentes.

91
REFERÊNCIAS BIBLIOGRÁFICAS
AGUIAR, L. F. S.; DI-BERNARDO, M. Diet and feeding behavior of Helicops infrataeniatus (Serpentes: Colubridae: Xenodontinae) in southern Brazil. Studies of Neotropical Fauna and Environment, Lisse, v. 39, p. 7-14, 2004. ALBUQUERQUE, N. R.; CAMARGO, M. Hábitos alimentares e comentários sobre a predação e reprodução das espécies do gênero Hydrops Wagler, 1830 (Serpentes: Colubridae). Comunicações do Museu de Ciências e Tecnologia da PUC-RS: Série Zoológica, Porto Alegre, v. 17, p. 21-32, 2004. AMARAL, A. Contribuição à biologia dos ophidios do Brasil III: hábitos curiosos da espécie Tachymenis brasiliensis Gomes (Colubridae, Boiginae). Memórias do Instituto Butantan, São Paulo, v. 7, p. 3-4, 1932. BEEBE, W. Field notes on the snakes of Kartabo, British Guiana, and Caripito, Venezuela. Zoologica, New York, v. 31, p. 11-52, 1946. BERNARDE, P. S.; KOKUBUM, M. N. C.; MARQUES, O. A. V. Utilização de hábitat e atividade em Thamnodynastes strigatus (Günther, 1858) no sul do Brasil (Serpentes, Colubridae). Boletim do Museu Nacional, Rio de Janeiro, v. 428, p 1-8, 2000. BERNARDE, P. S.; MOURA-LEITE, J. C.; MACHADO, R. A.; KOKUBUM, N. M. C. Diet of the colubrid snake Thamnodynastes strigatus (Günter, 1858) from Parana State, Brazil: with field notes on anuran predation. Revista Brasileira de Biologia, São Carlos, v. 60, p. 695-699, 2000. BIZERRA, A. História Natural de Tomodon dorsatus (Serpentes: Colubridae). 1998. 102 f. Dissertação (Mestrado em Ciências) - Instituto de Biociências, Universidade de São Paulo, São Paulo, 1998. BROOKS, D. R.; MCLENNAN, D. A. Phylogeny, ecology and behavior: a research programme in comparative biology. Chicago: University of Chicago Press, 1991. 434 p. BROOKS, D. R.; MCLENNAN, D. A. The nature of diversity: an evolutionary voyage of discovery. Chicago: University of Chicago Press, 2002. 668 p. CADLE, J. E. Molecular systematics of Neotropical xenodontine snakes: I. South American Xenodontines. Herpetologica, Bainbridge, v. 40, p. 8-20, 1984a. CADLE, J. E. Molecular systematics of Neotropical xenodontine snakes: II. Central American Xenodontines. Herpetologica, Bainbridge, v. 40, p. 21-30, 1984b. CADLE, J. E. The Neotropical colubrid snake fauna: lineage components and biogeography. Systematic Zoology, Levittown, v. 31, p. 1-20, 1985. CADLE, J. E. Phylogenetic relationships among advanced snakes: a molecular perspective. University of California Publications in Zoology, Berkeley, v. 119, p. 1-77, 1988.

92
CADLE, J. E.; GREENE, H. W. Phylogenetic patterns, biogeography, and the ecological structure of Neotropical snake assemblages. In: RICKLEFS, R. E.; SCHLUTER, D. (eds.). Species diversity in ecological communities: historical and geographical perspectives. Chicago: University of Chicago Press, 1993. p. 281-293. CAMPBELL, J. A.; LAMAR, W. W. The venomous reptiles of the western hemisphere. Ithaca: Cornell University Press, 2004. v.1. 475 p. CODDINGTON, J. A. Cladistic tests of adaptational hypothesis. Cladistics, New York, v. 4, p. 3-22, 1988. CUNHA, O. R.; NASCIMENTO, F. P. Ofídios da Amazônia X: as cobras da região Leste do Pará. Publicações Avulsas do Museu Paraense Emílio Goeldi, Pará, v. 31, p. 1-218, 1978. CUNHA, O. R.; NASCIMENTO, F. P.; ÁVILA-PIRES, T. C. S. Os répteis da área de Carajás, Pará, Brasil (Testudines e Squamatas). Publicações Avulsas do Museu Paraense Emílio Goeldi, Pará, v. 40, p. 10-92, 1985. DINIZ-FILHO, J. A. F. Métodos filogenéticos comparativos. Ribeirão Preto: Holos, 2000. 162 p. DIXON, J. R. The Neotropical colubrid snake genus Liophis: the generic concept. Milwaukee Public Museum, Contribution in Biology and Geology, Milwaukee, v. 31, p. 1-40, 1980. DIXON, J. R.; SOINI, P. The reptiles of the upper Amazon basin, Iquitos region, Peru: II. Crocodilians, turtles and snakes. Milwaukee Public Museum, Contribution in Biology and Geology, Milwaukee, v. 12, p 1-91, 1977. DUELLMAN, W. E. The biology of an equatorial herpetofauna in Amazonian Equador. University of Kansas, Museum of Natural History, Miscellaneous Publications, Kansas, v. 65, p. 1-352, 1978. FARRIS, J. S. Methods for computing Wagner trees. Systematic Zoology, Levittown, v. 19, p. 83-92, 1970. FELSENSTEIN, J. Phylogenies and the comparative method. American Naturalist, Chicago, v. 125, p. 1-15, 1985. FERRAREZZI, H. Sistemática filogenética de Elapomorphus, Phalotris e Apostolepis (Serpentes, Colubridae, Xenodontinae). 1993. Dissertação (Mestrado em Ciências) - Instituto de Biociências, Universidade de São Paulo, São Paulo, 1993. FERRAREZZI, H. Uma sinopse dos gêneros e classificação das serpentes (Squamata): II. Família Colubridae. In: NASCIMENTO, L. B.; BERNARDES, A. T.; COTTA, G. A. (eds.). Herpetologia no Brasil 1. Belo Horizonte: PUC-MG, 1994. p. 81-91. FORD, N. B.; FORD, D. F. Notes on the ecology of the South American water snake Helicops angulatus (Squamata: Colubridae) in Nariva Swamp, Trinidad. Caribbean Journal of Science, Mayagüez, v. 38, p. 129-132, 2002.

93
FRANCO, F. L. Relações filogenéticas entre os gêneros da tribo Tachymenini Bailey, 1967 (Serpentes: Colubridae). 1999. 252 f. Tese (Doutorado em Ciências) - Instituto de Biociências, Universidade de São Paulo, São Paulo, 1999. GANDOLFI, M. B.; BARBIERI, R. F. Relações filogenéticas entre as cobras d’água sul americanas (Serpentes: Colubridae: Xenodontinae). In: CONGRESSO LATINO AMERICANO DE HERPETOLOGIA, 4., 1996, Santiago. Anais… Santiago, Sociedade Brasileira de Herpetologia, 1996. p. 139. GANS, C. Biomechanics: an approach to vertebrate biology. Philadelphia: J. B. Lippincott, 1974. 261 p. GARLAND, T. JR.; HARVEY, P. H.; IVES, A. R. Procedures for the analysis of comparative data using phylogenetically independent contrasts. Systematic Biology, McLean, v. 41, p. 18-32, 1992. GOMES, J. F. Contribuição para o conhecimento dos ofídios do Brasil – II. Descrição de duas espécies novas. Memórias do Instituto Butantan, São Paulo, v. I, p. 79, 1918. GREENE, H. W. Dietary correlates of the origin and radiation of snakes. American Zoologist, Seattle, v. 23, p. 431-441, 1983. GREENE, H. W. Antipredator mechanisms in reptiles. In: GANS, C.; HUEY, R. B. (eds.). Biology of the Reptilia. Defense and life history. New York: Alan R. Liss. 1988, v. 16. p. 1-152. GREENE, H. W. Snakes: The evolution of mystery in nature. Berkeley: University of California Press, 1997. 351 p. GUYER, C.; DONNELLY, M. A. Length-mass relationship among an assemblage of tropical snakes in Costa Rica. Journal of Tropical Ecology, Winchelsea, v. 6, p. 65-76, 1990. HARVEY, P. H.; PAGEL, M. The comparative method in evolutionary biology. Oxford: Oxford University Press, 1991. 239 p. HENDERSON, R. W.; MICUCCI, T. W. P.; PUORTO, G.; BOURGEOIS, R. W. Ecological Correlates and patterns in the distribution of Neotropical boines (Serpentes: Boidae): a preliminary assessment. Herpetological Natural History, Corona, v. 3, p. 15-27, 1995. JAYNE, B. C. Swimming in constricting (Elaphe g. guttata) and nonconstricting (Nerodia fasciata pictiventris) colubrids snakes. Copeia, p. 195-208, 1985. JORGE DA SILVA, N. JR. The snakes from Samuel hydroelectric power plant and vicinity, Rondônia, Brazil. Herpetological Natural History, Corona, v. 1, p. 37-86, 1993. JORGE DA SILVA, N. JR.; SITES, J. W. JR. Phylogeny of South American triad coral snakes (Elapidae: Micrurus) based on molecular characters. Herpetologica, Bainbridge, v. 57, p. 1-22, 2001.

94
KAUFMAN, G. A.; GIBBONS, J. W. Weight - length relationships in thirteen species of snakes in the southeastern United States. Herpetologica, Bainbridge, v. 31, p. 31-37, 1975. KELLY, C. M.; BARKER, N. P.; VILLET, M. H. Phylogenetics of advanced snakes (Caenophidia) based on four mitochondrial genes. Systematic Biology, McLean, v. 52, p. 439-459, 2003. KING, R. B. Sexual dimorphism in snake tail length: sexual selection, natural selection, or morphological constraint? Biological Journal of the Linnean Society, London, v. 38, p. 133-154, 1989. KING, R. B. Sexual dimorphism in neonate and adult snakes. Journal of Zoology, London, v. 247, p. 19-28, 1999. KLUGE, A. G. Boine snake phylogeny and research cycles. Miscellaneous Publications, Museum of Zoology, University of Michigan, Michigan, v. 178, p. 1-58, 1991. KOHLSDORF, T.; GARLAND, T. JR.; NAVAS, C. A. Limb and tail lengths in relation to substrate usage in Tropidurus lizards. Journal of Morphology, v. 248, p. 151-164, 2001. KOVACH, W. L. MVSP: a mult-variate statistical package for Windows. Version 3.1. Penthraeth: Kovach Computing Services, 1999. 02 disquetes. LEMA, T.; DEIQUES, C. H. Contribuição ao conhecimento da “cobra espada d’água”, Ptychophis flavovirgatus Gomes, (Serpentes, Colubridae, Xenodontinae, Tachymenini). Comunicações do Museu de Ciências e Tecnologia da PUC-RS: Série Zoológica, Porto Alegre, v. 5, p. 55-83, 1992. LILLYWHITE, H. B. Temperature, energetics, and physiological ecology. In: SEIGEL, R. A.; COLLINS, J. T.; NOVAK, S. S. (eds.). Snakes: ecology and evolutionary biology. New York: MacMillan, 1987. p. 422-477. LILLYWHITE, H. B.; HENDERSON, R. W. Behavioral and functional ecology of arboreal snakes. In: SEIGEL, R. A.; COLLINS, J. T. (eds.). Snakes: ecology and behavior. New York: MacGraw-Hill, 1993. p. 1-48. LOSOS, J. B. Ecomorphology, performance capability, and scaling of West Indian Anolis lizards: an evolutionary analysis. Ecological Monographs, Washington, v. 60, p. 369-388, 1990. MACRINI, T. E.; IRSCHICK, D. J.; LOSOS, J. B. Ecomorphological differences in toepad characteristics between mainland and island Anolis. Journal of Herpetology, New Haven, v. 37, p. 52-58, 2003. MADDISON, W. P.; MADDISON, D. R. Mesquite: a modular system for evolutionary analysis. Version 1.05. 2004. Disponível em http://mesquiteproject.org. MANLY, B. F. J. Multivariate Statistical Methods. A Primer. London: Chapman & Hall, 1994. p. 214.

95
MARQUES, O. A. V. Composição faunística, história natural e ecologia de serpentes da Mata Atlântica, na região da Estação Ecológica de Juréia-Itatins, São Paulo. 1998. 135 f. Tese (Doutorado em Ciências) - Instituto de Biociências, Universidade de São Paulo, São Paulo, 1998. MARQUES, O. A. V.; SAZIMA, I. História natural dos répteis da Estação Ecológica Juréia-Itatins. In: MARQUES, O. A. V.; DULEBA, W. (eds.). Estação Ecológica Juréia-Itatins: ambiente físico, flora e fauna. Ribeirão Preto: Holos, 2004. p. 254-274. MARQUES, O. A. V.; PUORTO, G. Dieta e comportamento alimentar de Erytholamprus aesculapii, uma serpente ofiófaga. Revista Brasileira de Biologia, São Carlos, v. 54, p. 253-259, 1994. MARQUES, O. A. V.; SOUZA, V. C. Nota sobre a atividade alimentar de Liophis miliaris no ambiente marinho (Serpentes, Colubridae). Revista Brasileira de Biologia, São Carlos, v. 53, p. 645-648, 1992. MARTINS, E. P.; GARLAND, T. JR. Phylogenetic analyses of the correlated evolution of continuous characters: a simulation study. Evolution, Lawrence, v. 45, p. 534-557, 1991. MARTINS, M. Why do snakes sleep on the vegetation in central Amazonia? Herpetological Review, Saint Louis, v. 24, p. 83-84, 1993. MARTINS, M. História natural de uma taxocenose de serpentes de mata na região de Manaus, Amazônia Central, Brasil. 1994. 98 f. Tese (Doutorado em Ciências) - Instituto de Biologia, Universidade Estadual de Campinas, Campinas, 1994. MARTINS, M.; OLIVEIRA, M. E. Natural history of snakes in forests of the Manaus region, Central Amazonia, Brazil. Herpetological Natural History, Corona, v. 6, p. 78-150, 1999. MARTINS, M.; ARAÚJO, M. S.; SAWAYA, R. J.; NUNES, R. Diversity and evolution of macrohabitat use, body size and morphology in a monophyletic group of Neotropical pitvipers (Bothrops). Journal of Zoology, London, v. 254, p. 529-538, 2001. MARTINS, M.; MARQUES, O. A. V.; SAZIMA, I. Ecological and phylogenetic correlates of feeding habits in Neotropical pitvipers of the genus Bothrops. In: SCHUETT, G. W.; HÖGGREN, M.; DOUGLAS, M. E.; GREENE, H. W. (eds.). Biology of the vipers. Eagle Mountain, Eagle Mountain Publishing, 2002. p. 307-328. MARX, H.; RABB, G. B. Phyletic analysis of fifty characters of advanced snakes. Fieldiana: Zoology, Chicago, v. 63, p. 1-320, 1972. MIDFORD, P. E.; GARLAND, T. JR.; MADDISON, W. P. PDAP: PDTREE Package for Mesquite. Version 1.00. 2003. Disponível em http://mesquiteproject.org. MILES, D. B.; RICKLEFS, R. E. The correlation between ecology and morphology in deciduous forest passerine birds. Ecology, Washington, v. 65, p. 1629-1640, 1984. MOERMOND, T. C. Habitat constraints on the behavior, morphology, and community structure of Anolis lizards. Ecology, Washington, v. 60, p. 152-164, 1979.

96
MOURA-LEITE, J. C. Sistemática e filogenia da tribo Xenodontini Bonaparte, 1845 (Serpentes, Colubridae). 2001. 99 f. Tese (Doutorado) - Universidade Federal do Paraná, Curitiba, 2001. MURPHY, J. C.; HENDERSON, R. W. Tales of giants snakes: a historical natural history of anacondas and pythons. Malabar: Krieger Publishing, 1997. 215 p. OLIVEIRA, J. L.; BORGES, M.; MARQUES, O. A. V. Gomesophis brasiliensis (NCN). Reproduction and diet. Herpetological Review, Saint Louis, v. 34, p. 251-252, 2003. PARKER, H. W.; GRANDISON, A. G. C. Snakes - A natural history. Itacha: Cornell University Press, 1986. PINOU, T.; VICARIO, S.; MARSCHNER, M.; CACCONE, A. Relict snakes of North America and their relationships within Caenophidia, using likelihood-based Bayesian methods on mitochondrial sequences. Molecular Phylogenetics and Evolution, Oxford, v. 32, p. 563-574, 2004. PLATT, D. R. Natural History of the hognose snakes, Heterodon platyrhinus and H. nasicus. University of Kansas, Museum of Natural History, Miscellaneous Publications, Kansas, v. 18, p. 253-420, 1969. PORTO, M.; CARAMASCHI, U. Notes on the taxonomic status, biology, and distribution of Ptychophis flavovirgatus Gomes, 1915 (Ophidia, Colubridae). Anais da Academia Brasileira de Ciências, Rio de Janeiro, v. 60, p. 471-475, 1988. POUGH, F. H.; GROVES, J. D. Specialization of the body form and food habits of snakes. American Zoologist, Seattle, v. 23, p. 443-454, 1983. POUGH, F. H.; ANDREWS, R. M.; CADLE, J. E.; CRUMP, M. L.; SAVITZKY, A. H.; WELLS, K. D. Herpetology. New Jersey: Prentice-Hall International, 1998. ROZE, J. A. Coral snakes of the Americas: biology, identification, and venoms. Malabar: Krieger Publishing, 1996. 328 p. RUFFATO, R.; DI-BERNARDO, M.; MASCHIO, G. F. Dieta de Thamnodynastes strigatus (Serpentes, Colubridae) no sul do Brasil. Phyllomedusa, Belo Horizonte, v. 2, p. 27-34, 2003. SAVITZKY, A. H. Coadapted character complexes among snakes: fossoriality, piscivory, and durophagy. American Zoologist, Seattle, v. 23, p. 397-409, 1983. SAWAYA, R. J. História natural e ecologia das serpentes de Cerrado da região de Itirapina, SP. 2004. 159 f. Tese (Doutorado) - Instituto de Biologia, Universidade Estadual de Campinas, Campinas, 2004. SAZIMA, I.; ABE, A. S. Habits of five Brazilian snakes with coral-snake pattern, including a summary of defensive tactics. Studies of Neotropical Fauna and Environment, Lisse, v. 26, p. 159-164, 1991.

97
SAZIMA, I.; MARTINS, M. Presas grandes e serpentes jovens: quando os olhos são maiores que a boca. Memórias do Instituto Butantan, São Paulo, v. 52, p. 73-79, 1990. SAZIMA, I.; STRÜSSMANN, C. Necrofagia em serpentes brasileiras: exemplos e previsões. Revista Brasileira de Biologia, São Carlos, v. 50, p. 463-468, 1990. SHINE, R. Constraints on reproductive investment: a comparison between aquatic and terrestrial snakes. Evolution, Lawrence, v. 42, p. 17-27, 1988. SHINE, R. Why do larger snakes eats larger pry items? Functional Ecology, v. 5, p. 493-502, 1990. SHINE, R. Relative clutch mass and body shape in lizards and snakes: is reproductive investment constrained or optimized? Evolution, Lawrence, v. 46, p. 828-833, 1992. SHINE, R. Sexual size dimorphism in snakes revisited. Copeia, Miami, p. 326-346, 1994a. SHINE, R. Allometric patterns in the ecology of Australian snakes. Copeia, Miami, p. 851-867, 1994b. SLOWINSKI, J. B. A phylogenetic analysis of the New Word coral snakes (Elapidae: Leptomicrurus, Micruroides, and Micrurus) based on allozymic and morphological characters. Journal of Herpetology, New Haven, v. 29, p. 325-338, 1995. STATSOFT. Statistica for Windows. Version 5.1. Tulsa: Statsoft Incorporation, 1998. 01 CD STRÜSSMANN, C. Serpentes do pantanal de Poconé, Mato Grosso: composição faunística, história natural e ecologia comparada. 1992. 125 f. Dissertação (Mestrado) - Instituto de Biologia, Universidade Estadual de Campinas, Campinas, 1992. STRÜSSMANN, C.; SAZIMA, I. Esquadrinhar com a cauda: uma tática de caça da serpente Hydrodynastes gigas no Pantanal, Mato Grosso. Memórias do Instituto Butantan, São Paulo, v. 52, p. 57-61, 1990. STRÜSSMANN, C.; SAZIMA, I. The snake assemblage of the Pantanal at Poconé, western Brazil: faunal composition and ecological summary. Studies of Neotropical Fauna and Environment, Lisse, v. 28, p. 157-168, 1993. VANZOLINI, P. E. Levantamento herpetológico da área do Estado de Rondônia sob a influência da rodovia BR364. Brasília: CNPq, 1986. (Programa Polonoroeste, subprograma ecologia animal, relatório de pesquisa nº 1). VIDAL, N.; KINDL, S. G.; WONG, A.; HEDGES, S. B. Phylogenetic relationships of Xenodontine snakes inferred from 12S and 16S ribosomal RNA sequences. Molecular Phylogenetics and Evolution, Oxford, v. 14, p. 389-402, 2000.

98
VINCENT, S. E.; HERREL, A.; IRSCHICK, D. J. Sexual dimorphism in head shape and diet in the cottonmouth snake (Agkistrodon piscivorus). Journal of Zoology, London, v. 264, p. 53-59, 2004. VITT, L. J. Ecology of an anuran-eating guild of terrestrial tropical snake. Herpetologica, Bainbridge, v. 39, p. 52-66, 1983. VITT, L. J. Ecological observations on the tropical colubrid snake Leptodeira annulata. Herpetological Natural History, Corona, v. 4, p. 69-76, 1996. VITT, L. J.; VANGILDER, L. D. Ecology of snake community in the northeastern Brazil. Amphibia-Reptilia, Laiden, v. 4, p. 273-296, 1983. WIKRAMANAYAKE, E. D. Ecomorphology and biogeography of a tropical stream fish assemblage: evolution of assemblage structure. Ecology, Washington, v. 71, p. 1756-1764, 1990. ZAHER, H. Phylogénie des Pseudoboini et evólution des Xenodontinae sudaméricains (Serpentes, Colubridae). 1994. Dissertation - Muséum National d’Histoire Naturelle de Paris, 1994. ZAHER, H. Hemipenial Morphology of the South American Xenodontinae snakes, with a proposal for a monophyletic Xenodontinae and a reappraisal of Colubroid hemipenis. Bulletin Of the American Museum of Natural History, New York, v. 240, p. 1-168, 1999. ZAR, J. H. Biostatistical Analysis. 4º ed. New Jersey: Prentice-Hall International, 1999. 718 p. ZEMMERMANN, B. L.; RODRIGUES, M. T. Frogs, snakes, and lizards of the INPA/WWF reserves near Manaus, Brazil. In: GENTRY, A. H. (ed.). Four Neotropical Rainforests. New York: Yale University Press, 1990. p. 426-454.

99
APÊNDICE Apêndice: Localização das espécies e indivíduos analisados. Subfamília Xenodontinae Apostolepis assimilis. Minas Gerais: IB23985, IB44222, IB49374, IB49666, IB64671 e IB66376. São Paulo: IB6532, IB8506, IB9463, IB10022, IB23523, IB23889, IB24548, IB24873, IB27309, IB30283, IB32563, IB34182, IB41401, IB42513, IB42970, IB43026, IB44793, IB45207, IB45633, IB46016, IB46527, IB49354, IB51160, IB54208 e IB58341. Elapomorphus quinquelineatus. Espírito Santo: IB1018, IB9273, IB10057, IB30662, IB40225, IB50287 e IB62251. Minas Gerais: IB114, IB913, IB8833, IB9519, IB9572, IB9591, IB9656, IB10483, IB1290, IB1454, IB8255, IB25817, IB25976, IB27963, IB32130, IB37380, IB37534 e IB37539. Rio de Janeiro: IB6537, IB8370, IB9008, IB12334 e IB31447. Phalotris mertensi. Goiás: IB27933 e IB67616. Mato Grosso do Sul: IB16651. Minas Gerais: IB6842, IB7879 e IB23406. São Paulo: IB16650, IB16711, IB18885, IB19890, IB21690, IB24476, IB28031, IB30107, IB30634, IB30722, IB33922, IB34349, IB40539, IB41270, IB42514, IB42652, IB42965, IB43137, IB47501, IB50225, IB50282, IB57467, IB58449 e IB62515. Pseudoeryx plicatilis. Acre: IB28062 e MZUSP7369. Amapá: IB26276. Mato Grosso do Sul: IB4783 e IB41329. Pará: IB5090, IB14871, IB17640 e MZUSP4812. Roraima: MZUSP9769. Suriname: IB20463. Sem Procedência: IB68261e IB68262. Hydrops triangularis. Amazonas: IB40811, MZUSP7679, MZUSP7834, MZUSP8432 e MZUSP5478. Maranhão: IB55162. Mato Grosso do Sul: IB29171. Pará: IB14677, IB22174, MZUSP4220 e MZUSP9054. Rondônia: MZUSP8780. Tocantins: IB65080, IB65594, IB65641, IB65680, IB65810, IB65821 e IB65822. Colômbia: MZUSP5994. Guiana Francesa: IB13760. Helicops modestus. São Paulo: IB430, IB745, IB748, IB753, IB4522, IB5502, IB5700, IB6988, IB7547, IB7724, IB16528, IB16534, IB16536, IB16550, IB16551, IB16778, IB16779, IB16797, IB16815, IB16818, IB16821, IB16828, IB20847, IB20849, IB56819, IB58133, IB59527, IB60221 e IB63669. Helicops infrataeniatus. Mato Grosso do Sul: IB15972, IB30262, IB31729, IB37131, IB37660, IB37856, IB48983, IB59428, IB59429, IB59453, IB59458, IB59461, IB59658, IB59677, IB59712, IB59738, IB59758, IB59820, IB59853, IB59855, IB63023, IB63024 e IB63032. São Paulo: IB4722, IB25650, IB29037, IB29046, IB29525, IB30248 e IB31800. Helicops carinicaudus. Paraná: IB12325. Santa Catarina: IB3062, IB26221 e IB31827. São Paulo: 05 indivíduos não tombados, IB1383, IB4437, IB7094, IB8473, IB17180, IB17368, IB27038, IB27821, IB28035, IB29514, IB31135, IB31738, IB31882, IB32229, IB32242, IB32484, IB32989, IB34095, IB34309, IB46631, IB46632, IB49630, IB56405, IB57751 e IB59517. Helicops angulatus. Acre: IB55247. Amapá: IB45709. Amazonas: 02 indivíduos não tombados. Bahia: IB1760. Ceará: IB20158. Goiás: IB34387, IB42605, IB45962 e IB50288. Maranhão: IB20673. Mato Grosso: IB12831 e IB12851. Pará: IB18502. Paraíba: IB51817. Rondônia: IB46912 e IB47031. Roraima: IB63870. Tocantins: IB46165, IB66445 e IB66878. Helicops polylepis. Amazonas: 11 indivíduos não tombados. Tocantins: IB66244. Sordellina punctata. Mato Grosso do Sul: IB10464. Paraná: IB20481. São Paulo: IB6791, IB22313, IB22936, IB24313, IB29487, IB32749, IB33201, IB34286, IB40285, IB40160, IB41071, IB41186,

100
IB41375, IB41377, IB41408, IB42518, IB44176, IB46054, IB46023, IB46611, IB48744, IB54552, IB55084, IB55332, IB56066, IB56068, IB62203 e MZUSP2003. Gomesophis brasiliensis. Minas Gerais: IB9746 e IB43209. Paraná: IB4323, IB5065, IB6943, IB18815 e IB45993. Rio Grande do Sul: IB6484, IB8498 e IB18343. Santa Catarina: IB22945 e IB32115. São Paulo: IB3083, IB5200, IB9462, IB9648, IB10982, IB16909, IB16910, IB16953, IB16954, IB16971, IB17103, IB17104, IB17105, IB17106, IB17107, IB17108, IB17109, IB17110, IB17111, IB17737, IB18402, IB19694, IB22105, IB23282, IB26638, IB40494, IB54906 e MZUSP5933. Ptychophis flavovirgatus. Minas Gerais: IB10548, IB13204, MNRJ3276 e MNRJ3277. Paraná: IB5635, IB7821, IB12551, IB25958, IB25970, IB29881, IB32531, MHNCI3364, MHNCI3490, MHNCI7844, MHNCI7803 e MCP10979. Rio Grande do Sul: IB7924, IB8326, IB8451, MCP1526, MCP2606, MCP2607, MCP2798 e MCP2809. Santa Catarina: IB6967, IB11343, IB11343, IB53568 e IB53995. Tomodon dorsatus. Minas Gerais: IB60626. Rio Grande do Sul: IB32151. São Paulo: IB25725, IB29549, IB29659, IB29618, IB30575, IB30576, IB30611, IB31213, IB31285, IB31391, IB31666, IB44007, IB58624, IB58849, IB58850, IB59417, IB59423, IB59984, IB60003, IB60080, IB60083, IB60085, IB60561, IB60606, IB61514 e IB61647. Thamnodynastes strigatus. Minas Gerais: IB9433, IB29389, IB30017, IB30287, IB56879, IB57547, IB62023 e IB62025. São Paulo: IB11651, IB11930, IB23663, IB23811, IB28075, IB28111, IB28235, IB29460, IB28640, IB28642, IB28643, IB28644, IB28646, IB29541, IB29615, IB29692, IB30238, IB60309, IB61007, IB61140, IB61958, IB68147, IB68148, IB68297, IB68303 e IB68501. Liophis frenata. Bahia: IB2537. Mato Grosso do Sul: IB19138, IB19907, IB36376, IB48940, IB59441 e MZUSP10132. São Paulo: IB5768, IB8522, IB8536, IB10236, IB16709, IB17313, IB17620, IB24614, IB27848, IB28295, IB32665, IB34153, IB34406, IB35992, IB41616, IB41751, IB41848, IB41914, IB41919, IB41921, IB42748, IB46606, IB54052, IB55704 e IB59991. Liophis cobella. Alagoas: MZUSP2927, MZUSP2930, MZUSP2934 e MZUSP2935. Amapá: IB13796, IB14960, IB24833, IB24853, IB24854, IB24857 e IB24859. Amazonas: IB22176. Bahia: IB1358 e IB3213. Maranhão: IB21747, IB21748 e IB56907. Pará: IB14791, IB17703, IB17704, IB25483, IB25484, IB25485, IB25487 e IB46536. Pernambuco: IB50478. Tocantins: IB63345, IB65801 e IB65802. Liophis miliaris. São Paulo: IB9242, IB11437, IB11586, IB11587, IB11593, IB11596, IB12080, IB12130, IB12203, IB12206, IB12207, IB12327, IB15589, IB15642, IB15678, IB15832, IB15833, IB15991, IB17613, IB19486, IB22182, IB26570, IB26755, IB27670, IB27706, IB29435, IB30317, IB31918, IB31928, IB52592 e IB68052. Liophis poecilogyrus. São Paulo: IB4684, IB4968, IB5013, IB6415, IB6416, IB6441, IB6464, IB6594, IB6598, IB6850, IB6908, IB6917, IB6990, IB7015, IB7017, IB7090, IB7135, IB7369, IB7412, IB7437, IB7443, IB7925, IB13159, IB13167, IB21920, IB27662, IB27880, IB32700, IB61867, IB62170 e IB62851. Liophis reginae. Ceará: IB20276. São Paulo: IB5716, IB6958, IB7390, IB8253, IB9849, IB10233, IB12583, IB15299, IB16620, IB17032, IB17290, IB18371, IB18975, IB19121, IB22238, IB22500, IB22587, IB22588, IB22947, IB27383, IB27702, IB27703, IB27704, IB27777, IB28559, IB28637, IB29631, IB30872, IB31394, IB31639, IB31760, IB32350, IB32569, IB33115, IB41958, IB44018, IB46008, IB46283 e IB56264. Erythrolamprus aesculapii. São Paulo: IB2611, IB4435, IB4562, IB4568, IB4668, IB5138, IB5167, IB5644, IB5656, IB5690, IB5935, IB5948, IB6074, IB6557, IB11032, IB19919, IB21373, IB27690,

101
IB29187, IB29459, IB29479, IB29632, IB33773, IB37271, IB41812, IB51588, IB58661, IB61234, IB64794, IB68591 e IB68608. Xenodon neuwiedii. São Paulo: IB18154, IB18941, IB24334, IB26551, IB27207, IB28038, IB28297, IB56486, IB56490, IB56491, IB57850, IB57895, IB57899, IB57906, IB57974, IB58633, IB60938, IB60253. Hydrodynastes bicinctus. Amazonas: 01 indivíduo não tombado, IB31794, IB32002, IB32012 e MZUSP5358. Goiás: IB32560, IB33323 e MZUSP3838. Mato Grosso: IB27652, IB41160, IB41232, IB42063 e MZUSP4752. Pará: IB46235 e IB46883. Roraima: MZUSP9125. São Paulo: IB160, IB18159, IB28240, IB29293, IB42136 e IB42167. Hydrodynastes gigas. Amazonas: IB40895. Mato Grosso do Sul: IB17550, IB18264, IB22214, IB22600, IB59437, IB59475, IB59826, IB63203, IB63265 e IB63859. Mato Grosso: IB46997. Pará: IB19843 e IB41123. São Paulo: IB226, IB20720, IB36715, IB37132, IB38008, IB38179, IB38682, IB38683, IB38684, IB38770, IB38790, IB50676, IB41623, IB42194, IB42541, IB52248, IB52250, IB52253, IB52254 e IB54012. Pseudoboa nigra. Alagoas: IB48629. Espírito Santo: IB7758, IB8808, IB9092, IB25147, IB26072, IB29983, IB37408, IB48631, IB50042 e IB60653. Goiás: IB4849, IB5668, IB7772 e IB12043. Mato Grosso do Sul: IB15853, IB16078, IB24468, IB24893, IB31131, IB33457, IB59427, IB63297 e IB63874. Minas Gerais: IB5870, IB28402, IB54335 e IB67000. Rio de Janeiro: IB10135, IB27158 e IB30333. Tocantins: IB65403, IB66083, IB66170, IB66265 e IB66266. Oxyrhopus guibei. São Paulo: IB387, IB4366, IB4404, IB4405, IB4406, IB4844, IB5519, IB14347, IB52530, IB52534, IB53632, IB57586, IB59621, IB60019, IB60158, IB60560, IB60994, IB61214, IB64141, IB64147, IB64622, IB64663, IB64664, IB64709, IB64818, IB64820, IB64823, IB64873, IB66726, IB67410, IB67422, IB67425, IB67626, IB67865 e IB68813. Clelia quimi. São Paulo: IB711, IB1813, IB880, IB8752, IB9074, IB11327, IB14331, IB22445, IB22485, IB22490, IB24323, IB25605, IB25606, IB26906, IB27128, IB27442, IB30029, IB33216, IB34293, IB37255, IB40747, IB41066, IB42616, IB42617, IB44141, IB52644, IB54883, IB54903, IB54963, IB59363, IB61152, IB61154 e IB61155. Leptodeira annulata. Mato Grosso do Sul: IB36876, IB36877, IB63322, IB63326, IB63327, IB63329, IB63330, IB63331, IB63332, IB63733, IB63736, IB63737. São Paulo: IB20511, IB31224, IB34431, IB36878, IB36884, IB37623, IB41533, IB41544, IB41615, IB41738, IB45735, IB46548, IB55561, IB56338, IB60109, IB63274, IB69866. Heterodon platyrhinus. Estados Unidos. Alabama: IB4104, IB4105. Carolina do Sul: IB4113, IB4114, IB4116. Iowa: IB4094. Texas: IB4080, IB4081, IB4082, IB4083, IB4084. Sem procedência: IB2295, IB2299 IB2359. Subfamília Boinae Boa constrictor. São Paulo: IB10902, IB15221, IB18632, IB31166, IB31402, IB38754, IB52218, IB53119, IB55788, IB57479, IB60073, IB60075, IB60076, IB6135, IB63744, IB64913, IB7148, IB7544, IB7546, IB7903. Minas Gerais: IB46150, IB46151 e IB46152. Epicrates cenchria. Amapá: IB14035, IB19126, IB24828 e IB25425. Amazonas: IB24484, IB44169 e IB51494. Bahia: IB50983. Espírito Santo: IB33018, IB51234 e IB51765. Mato Grosso: IB41463, IB49979, IB51358, IB51359, IB51360, IB51362, IB51367, IB51370, IB51371 e IB55245. Maranhão:

102
IB41275. Minas Gerais: IB33315. Pará: IB14687, IB14691, IB41037, IB46662. Pernambuco: IB46547. Eunectes murinus. Mato Grosso do Sul: IB7489, IB18697, IB18760, IB18841, IB19699, IB40774 e IB59464. Minas Gerais: IB16005. São Paulo: IB8700, IB19062, IB19063, IB19064, IB21945, IB27736, IB33959, IB51421, IB52424, IB58674 e IB63882. Corallus hortulanus. Rio de Janeiro: IB28941, IB30610, IB42607, IB43859, IB44402, IB44231. São Paulo: IB19168, IB28919, IB33652, IB45634, IB53397, IB55134. Tocantins: IB65746, IB65748, IB65749, IB65750, IB66278, IB66279, IB66285, IB66291, IB66292. Subfamília Elapinae Micrurus surinamensis. Amapá: IB13769, IB13880 e IB14862. Amazonas: IB52225 e IB52327. Mato Grosso: IB41205 e IB68050. Pará: IB2193, IB14657, IB14858, IB14864, IB25430, IB33438, IB44101 e IB46764. Rondônia: MZUSP8716. Roraima: MZUSP10703 e MZUSP10704 Tocantins: IB65619, IB66246 e IB66889. Colômbia: MZUSP6107 e MZUSP6124. Micrurus lemniscatus. Bahia: IB67647. Minas Gerais: IB40178, IB40182, IB42642 e IB46333. Paraná: IB46796. São Paulo: IB1220, IB1308, IB3051, IB3054, IB4975, IB5578, IB5808, IB6132, IB7519, IB7793, IB7864, IB8125, IB9324, IB9929, IB9990, IB16611, IB33999, IB37440, IB40146, IB42457, IB43601, IB44642, IB46339, IB46388, IB46765 e IB53965. Micrurus frontalis. Minas Gerais: IB32977, IB33237, IB33334 e IB55990. São Paulo: IB410, IB4976, IB32309, IB33436, IB33517, IB37364, IB40132, IB42930, IB43196, IB43609, IB43986, IB 46572, IB47652, IB48810, IB49451, IB49669, IB50134, IB50326, IB50675, IB50882, IB51081, IB51132, IB51592, IB52161, IB54820, IB58271 e IB 68218. Micrurus corallinus. São Paulo: IB42230, IB44088, IB46475, IB46557, IB46569, IB46769, IB53039, IB53040, IB53041, IB53042, IB53043, IB53045, IB53046, IB53048, IB53051, IB53061, IB53363, IB53364, IB54459, IB54813, IB54874, IB55981.


















