Padrões sazonais e espaciais de distribuição e diversidade de ...
162
UNIVERSIDADE FEDERAL DE MINAS GERAIS INSTITUTO DE CIÊNCIAS BIOLÓGICAS PÓS-GRADUAÇÃO EM ECOLOGIA, CONSERVAÇÃO E MANEJO DA VIDA SILVESTRE Padrões sazonais e espaciais de distribuição e diversidade de herbáceas no Pantanal de Poconé-MT Luciana Rebellato Belo Horizonte, Minas Gerais 2010
Transcript of Padrões sazonais e espaciais de distribuição e diversidade de ...

UNIVERSIDADE FEDERAL DE MINAS GERAIS INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PÓS-GRADUAÇÃO EM ECOLOGIA, CONSERVAÇÃO E MANEJO DA VIDA SILVESTRE
Padrões sazonais e espaciais de distribuição e diversidade de
herbáceas no Pantanal de Poconé-MT
Luciana Rebellato
Belo Horizonte, Minas Gerais 2010

LUCIANA REBELLATO
Padrões sazonais e espaciais de distribuição e
diversidade de herbáceas no Pantanal de Poconé-MT
Orientador: Dr. José Eugênio Côrtes Figueira
Belo Horizonte, Minas Gerais 2010
Tese apresentada ao Curso de Ecologia, Conservação e Manejo da Vida Silvestre da Universidade Universidade Federal de Minas Gerais, como um dos requisitos para obtenção do título de Doutor em Ecologia, Conservação e Manejo da Vida Silvestre

Dedico este trabalho ao meu pai,
Luiz Rebellato

AGRADECIMENTOS Agradeço principalmente à minha família, minha mãe Narzira, meu pai Luiz (In
memorian) muita saudade, e irmãs Lílian e Lívia, pelo amor, carinho e apoio, sendo
todos exemplos para mim.
À Fapemat pela concessão da bolsa de estudos, fundamental para a elaboração desta
tese.
Ao Centro de Pesquisas do Pantanal (CPP) pelo apoio logístico e financeiro.
À Universidade Federal de Mato Grosso e Núcleo de Estudos do Pantanal (NEPA) pelo
espaço, laboratórios, material e herbário.
Ao curso de pós-graduação em Ecologia, Conservação e Manejo da Vida Silvestre -
ECMVS.
Ao Prof. Dr. José Eugênio por todo o apoio, dedicação, conhecimento e paciência.
Aos membros da banca examinadora, que com clareza e tranqüilidade me trouxeram
ótimas críticas e sugestões para melhoria deste trabalho.
À Profa. Cátia Nunes da Cunha por sempre me apoiar desde que cheguei ao Mato
Grosso, me oportunizando muitas conquistas.
Aos doutores Sra. Valli Joana Pott e Sr. Arnildo Pott pelas identificações de espécies e
proveitosas e divertidas tardes no herbário (UFMT).
Aos muitos amigos que fiz nesta terra quente e bela: Artema Lima, Carol Félfili, Cátia
Nunes da Cunha, Cleide Regina de Arruda, Drausio Honório de Moraes, Érica Arruda,
Fernandinho, Gilmar, Júlia Arieira, Luciana Ferraz, Mary Irantxe, Rosa Monteiro entre
tantos outros.

À minha amiga de longa data Mônica Aragona e seu companheiro pantaneiro João
Batista de Pinho por todo o carinho, apoio e peixes assados.
Aos pantaneiros do Retiro Novo: Dona Chute, Iá, Lu e Nesinho.
E também aos proprietários das Fazendas que possibilitaram tal estudo, Sr. Carlos
Maciel, Sr. Paulo das Canoas e Sra. Oreana.

A FLOR DOS AGUAÇAIS
Não posso apreciar a rosa cultivada
pela jeitosa mão do mestre jardineiro, por mais que seja bela, é pobre, é deturpada,
é sempre imitação de cunho financeiro.
A essa rosa falsa, enferma e bem tratada, prefiro a flor pagã, de porte alvissareiro,
que treme e se balouça ao som da chuvarada, que brota no negror do insípido atoleiro.
É a flor dos aguaçais... selvática, viçosa,
enfrenta a tempestade, e cresce bela, inculta, em meio ao estridor da luta tormentosa.
E quando a morte atroz perpassa-lhe a raiz, ela não chora a dor, e mesmo assim se exulta,
porque lutou, sofreu, foi livre e foi feliz
Ronaldo de Arruda Castro (poeta cuiabano)

SUMÁRIO
Padrões sazonais e espaciais de distribuição e diversidade de herbáceas no Pantanal de Poconé-MT RESUMO GERAL 1 ABSTRACT 4 INTRODUÇÃO GERAL 7 REFERÊNCIAS CITADAS 10
CAPÍTULO 1
Ordenação de tipologias vegetais e preferência de herbáceas no gradiente de inundação do Pantanal de Poconé-MT RESUMO 16 ABSTRACT 17 INTRODUÇÃO 18 MATERIAL E MÉTODOS 21 Características gerais e área de estudo 21 Desenho amostral 24 Amostragem de herbáceas 25 Identificação da tipologias vegetacionais 26 Análises estatísticas 26 RESULTADOS 27 Tipos de vegetação 27 Ordenação das comunidades de herbáceas 31 Mudanças temporais 32 Espécies dominantes e riqueza por sub-tipo de vegetação 36 Distribuição das espécies herbáceas relacionadas aos sub-tipos de vegetação 39 Distribuição das espécies herbáceas ao longo do gradiente de inundação 42 DISCUSSÃO 45 CONCLUSÕES 50 REFERÊNCIAS CITADAS 50
CAPÍTULO 2 Capítulo 2 – Efeito da sazonalidade e gradiente de inundação na riqueza, composição, cobertura e formas de vida de herbáceas do Pantanal de Poconé-MT RESUMO 57 ABSTRACT 58 INTRODUÇÃO 59 MATERIAL E MÉTODOS 61 Características gerais e área de estudo 61 Desenho amostral 64 Amostragem de herbáceas 65 Análises dos dados 66 RESULTADOS 69 Efeitos regionais da inundação sobre a florística, riqueza e diversidade de
espécies 69 Efeitos regionais da inundação sobre a cobertura de espécies 73 Efeitos regionais da inundação sobre as formas de vida 82

Efeito do gradiente de inundação na riqueza em espécies 84 Efeito do gradiente de inundação na diversidade beta 85 Efeito do gradiente de inundação na cobertura das espécies 86 Efeito do gradiente de inundação sobre as formas de vida 89 DISCUSSÃO 93 CONCLUSÕES 100 REFERÊNCIAS CITADAS 101
CAPÍTULO 3 Padrões de distribuição, riqueza e substituição de poáceas do Pantanal de Poconé-MT RESUMO 111 ABSTRACT 112 INTRODUÇÃO 113 MATERIAL E MÉTODOS 116 Características gerais e área de estudo 116 Desenho amostral 120 Amostragem de gramíneas 121 Análises dos dados 122 RESULTADOS 124 Cobertura regional de gramíneas 124 Distribuição de gramíneas ao longo do gradiente de inundação 129 Riqueza em gramíneas no gradiente de inundação 133 Diversidade Beta no gradiente 133 Formas de vida e rotas C3 e C4 134 DISCUSSÃO 138 CONCLUSÕES 143 REFERÊNCIAS CITADAS 144 ANEXOS 152

1
Padrões sazonais e espaciais de distribuição e diversidade de
herbáceas no Pantanal de Poconé-MT
RESUMO GERAL – A heterogeneidade espaço-temporal do Pantanal afeta o
estabelecimento dos organismos e, portanto, a distribuição de espécies vegetais, sejam
elas lenhosas ou herbáceas, estruturando diferenciadas comunidades vegetais. Neste
sentido, este trabalho tem como objetivo principal compreender melhor a preferência de
ocorrência de espécies herbáceas em função do gradiente hidrotopográfico, assim como
variações de composição e estrutura durante um ciclo hidrológico no Pantanal de
Poconé (MT). Nós utilizamos o sistema RAPELD, onde 30 isolinhas de 250m foram
sistematicamente distribuídas em um sistema de 6 trilhas de 5 km cada e eqüidistantes
1km, cobrindo uma área de 25 km2. A amostragem de herbáceas foi realizada pelo
método da parcela pontual ao longo das isolinhas em diferentes períodos (seca, cheia e
vazante), totalizando 209 espécies. As 30 isolinhas dispostas em variadas cotas
altimétricas diferiram quanto a profundidade e duração da inundação a que estiveram
submetidas e tipo de vegetação (florestal, campestre e savânica). O capítulo 1 teve como
objetivo verificar se diferentes sub-tipos de vegetação amostrados formam comunidades
diferenciadas no espaço e no tempo e quais espécies os caracterizam, além de verificar a
preferência das espécies herbáceas em relação ao gradiente de inundação. Foi realizada
uma ordenação das 30 isolinhas amostradas nos diferentes períodos e testou-se a
influência da profundidade da inundação em relação aos eixos, verificando também os
agrupamentos formados. Através de um Escalonamento Multidimensional Não-Métrico
(NMDS), utilizando uma matriz contendo dados de cobertura das espécies nos
diferentes períodos, constatou-se que o primeiro eixo esteve bem relacionado com a
profundidade da inundação. A maior parte das isolinhas se agrupou segundo os sub-
tipos de vegetação definidos a priori em campo. Os grupos definidos a priori (sub-tipos
de vegetação) foram significativamente distintos em relação à estrutura e composição de
espécies quando testados pela Análise de Similaridade (ANOSIM). Nos grupos
formados pelos sub-tipos de vegetação mais inundáveis, como campo de murundus,
campo limpo, campo exótico e floresta inundável, houve diferenças significativas em
relação à estrutura e composição de espécies entre os períodos, o que demonstrou o
efeito da sazonalidade hídrica nestas comunidades herbáceas. Os diferentes sub-tipos de
vegetação possuem espécies dominantes com preferências quanto ao gradiente de

2
inundação e períodos, o que provavelmente separou e definiu os grupos. Uma ordenação
direta possibilitou visualizar e agrupar as espécies quanto a preferência no gradiente.
Muitas espécies possuem ampla distribuição ao longo do gradiente, enquanto outras são
restritas aos seus extremos, ou seja, os não inundáveis e os muito inundáveis. A
determinação de espécies indicadoras quanto a sua preferência ambiental pode auxiliar
no manejo de áreas úmidas, como exemplo, na detecção de mudanças na estrutura e
composição de espécies em função de possíveis impactos em relação aos padrões do
regime hidrológico, assim como indicar rapidamente características hidrológicas de uma
determinada área a partir das espécies. No capítulo 2 o objetivo foi verificar qual o
efeito regional da inundação sazonal e do gradiente de inundação, em diferentes
períodos, sobre a riqueza, diversidade, composição, cobertura e formas de vida das
espécies herbáceas. A inundação influenciou a composição, como também a riqueza e a
diversidade (H’) em espécies, significativamente mais elevadas durante os períodos de
cheia e vazante em relação à seca. Poucas espécies foram abundantes, entre elas A.
purpusii, B. humidicola e P. lineare que mantiveram elevada cobertura regional entre os
períodos. Entretanto, inundação influencia a cobertura de muitas espécies, tendo
Eleocharis minima e Utricularia foliosa destaque em cobertura durante a cheia. As
hemicriptófitas foram dominantes e mais estáveis em relação à cobertura em todos os
períodos, sendo a participação de terófitas e geófitas vinculada aos períodos de cheia e
vazante. O gradiente crescente de inundação teve efeitos negativos sobre a riqueza em
espécies, a diversidade beta e a riqueza em hemicriptófitas em todos os períodos, e de
geófitas durante a seca. Tende a influenciar também o aumento de cobertura para
geófitas e terófitas durante a cheia. As variações nas coberturas das espécies entre os
períodos de seca, cheia e vazante, também tendem a ser maiores ao longo deste
gradiente. Desta forma, concluiu-se que as espécies possuem diferentes plasticidades ou
estratégias para que possibilitam sua manutenção neste ambiente dinâmico, tendo o
ciclo hídrico importância na determinação da riqueza e diversidade. Assim, a
manutenção do regime hidrológico nessas áreas úmidas e a variabilidade espacial na
paisagem são fundamentais para a conservação de suas biodiversidades. No capítulo 3
apenas a família Poaceae foi analisada dada a grande riqueza entre as herbáceas e
importância econômica. Nosso objetivo foi responder as seguintes questões: quais são
as espécies com maior cobertura na região, qual a influência da variação sazonal e
microtopográfica na riqueza, cobertura, distribuição, substituição de espécies, formas de
vida e rotas fotossintéticas das gramíneas? No total foram amostradas 59 espécies,

3
sendo três espécies regionalmente dominantes e pouco variáveis em cobertura entre os
três períodos, entre elas: Axonopus purpusii, Brachiaria humidicola e Paspalum lineare.
A distribuição e a cobertura das gramíneas foram influenciadas pelo gradiente de
inundação, assim como a riqueza em espécies, que diminuiu continuamente ao longo
deste gradiente. Variações na diversidade beta foram notadas no espaço e no tempo. As
gramíneas anuais tenderam a maior cobertura durante a seca nas isolinhas sujeitas a
maior profundidade de inundação. Espécies C4 estão presentes em maior número e
dominam em cobertura todo o gradiente. As espécies C3 também ocorrem ao longo de
todo gradiente com baixa cobertura e de forma mais equitativa. Entretanto, a cobertura
de C3 aumenta nos ambientes de maior profundidade durante a seca e cheia. A
cobertura média entre os períodos destas espécies C3 também revelou tendência de
aumento nos extremos, mais secos/sombreados ou mais inundáveis. Assim, foi possível
concluir que esta família se distribui por todo o gradiente de inundação, mas que este
afeta a distribuição e a riqueza em espécies de gramíneas, de acordo com suas
tolerâncias a diferentes intensidades de estresse hídrico.
Palavras-chaves: Pantanal, herbáceas, gradiente de inundação, sazonalidade

4
Seasonal and spatial distribution and diversity patterns of
herbaceous plants in the Pantanal of Poconé, Mato Grosso.
ABSTRACT – The space-time heterogeneity of the Pantanal affects the establishment
of organisms and, therefore, the distribution of plant species, be they woody or
herbaceous, allowing for differentiated plant communities. In this way, this study has
the main objective of better understanding the occurrence preference of herbaceous
species along the hydro-topographic gradient as well as the variations in composition
and structure during the water cycle of the Poconé Pantanal (Mato Grosso). We used the
RAPELD system whereby 30 isolines, 250m long each, were distributed equidistantly
throughout a system of 6 paths, 5 km long, at 1 km intervals covering an area of 25km.
The sampling of herbaceous species was carried out using the point sampling method
along the isolines during the different periods (dry, flood and drain), with a total of 209
species. The 30 isolines placed at varied altimetric positions differed in terms of the
flood depth and duration that they were submitted to as well as the vegetation type
(forest, meadow and savannah). Chapter 1 aims to verify if different sampled vegetation
sub-types form communities that are differentiated in space and time as well as
verifying which species form the communities and what the preferences are of
herbaceous plants in relation to the flood gradient. The 30 isolines were ordered
according to the different periods, and tests were made to analyse the influence of flood
depth in relation to the axis and also verifying the groups formed. By means of a
NMDS, using the data on species coverage in the different periods, it was found that the
first axis was strongly connected to flood depth. Most of the isolines were grouped
according to the vegetation sub-type defined a priori in the field. The groups defined a
priori (vegetation sub-types) were significantly distinct when tested by the ANOSIM.
For the groups formed by the more floodable vegetation sub-types, such as murundus,
open fields, exotic fields and flood forest, there were significant differences between the
periods which demonstrated the effect of the seasonal water cycle on the composition
and structure of these plant communities. The different vegetation sub-types possess
dominant species with preferences in terms of flood gradient and periods which are
probably what separated and defined the groups. Direct ordination of the species
enabled visualization and species grouping in terms of preferences on the gradient. The

5
majority of species occur in environments with intermediate flood levels followed by a
large contingency of species in non-flooding environments. Species indicators, in terms
of environmental preference, could aid habitat classification and wetland management,
such as, for example, the detection of change in the structure and composition of species
due to possible impacts on the water regime patterns as well as rapidly indicating the
hydrological characteristics of a particular area. In chapter 2, the aim was to verify the
regional effect of seasonal flooding and the flood gradient in different periods on the
flora, richness, diversity, coverage and plant life forms. Flooding influenced plant life
with the richness and diversity (H’) being most highly influenced during the flood and
drain periods. Flooding influenced the coverage of many species especially for
Eleocharis minima and Utricularia foliosa during the flood period. The
hemicryptophytes were dominant and more stable in the flood and drain periods with
the participation of geophytes and therophytes linked to the flood and drain periods. The
flood gradient has negative effects on species richness, beta diversity and
hemicryptophytes richness in all periods and geophytes during the dry period. The
variations in species coverage also tend to be more dynamic along this gradient. It was
concluded, therefore, that the species present posses plasticity or strategies to maintain
themselves in this dynamic environment and the water cycle is important for richness
and diversity reinforcing the questions of maintaining the water regime in wetlands and
its spatial variability within the landscape and conservation of its diversity. In chapter 3,
the Poaceae family alone was analysed given its great richness among herbaceous plants
and its great economic importance. The aim was to answer questions about which
species have the greatest coverage in the region, what influence seasonal changes may
have and the micro-topographics of the richness, coverage, distribution, turnover, life
forms and photosynthetic paths of grasses. In total, 59 species were sampled, three of
which are regionally dominant and showed little variation in coverage over the three
periods. These are: Axonopus purpusii, Brachiaria humidicola and Paspalum lineare.
The distribution and coverage of grasses were influenced by flood gradient as well as
species richness which diminished continuously along the flood gradient. Variations in
the beta diversity were noted in space and time. The number of annual grasses tended to
have greater coverage during the dry period along the isolines subject to greater
flooding depths. C4 species are present in greater numbers and dominate coverage along
the gradient. The C3 species also occurred along the whole gradient with less coverage
and in a more egalitarian way. However, the coverage of C3 increases in the higher

6
water depth environments during the dry and flood periods. The average coverage over
the periods of the C3 species also showed a tendency to be higher in extreme conditions
of dryness/shade and flooding. It is thus possible to conclude that this family is
distributed along the whole flood gradient, but this affects the distribution and richness
of grass species as it favours species with more adaptable characteristics that can
tolerate the different stress intensities.
Key words:Pantanal, herbaceous plants, flood gradient, seasonality

7
INTRODUÇÃO GERAL
O termo “áreas úmidas” agrupa uma ampla gama de habitats com características
comuns, sendo que a mais importante delas é a contínua, sazonal ou periódica
permanência de água ou saturação do solo (Finlayson & van der Valk 1995). De acordo
com a definição proposta na Convenção RAMSAR, áreas úmidas são "extensões de
brejos, pântanos e turfeiras, ou superfícies cobertas de água, sejam de regime natural ou
artificial, permanentes ou temporárias, estancadas ou correntes, doces, salobras ou
salgadas, incluídas as extensões de água marinha cuja profundidade na maré baixa não
exceda os seis metros". Estas áreas ocupam posições entre ambientes aquáticos e
terrestres, definidas como ATTZ (Áreas de Transição Aquático-Terrestre) e
compreendem uma variedade de habitats, entre eles, florestas, campos, brejos e
arbustais (Junk et al. 1989). Estimativas mostram que áreas úmidas podem ocupar até
12,8 milhões de quilômetros quadrados, pouco menos de 10% da superfície da Terra
(Zedler & Kersher 2005). Junk (2005) estima que até 20% da América do Sul Tropical
pode ser categorizada como áreas úmidas.
Nas últimas duas décadas, ecossistemas de áreas úmidas têm recebido forte
atenção, principalmente em relação ao entendimento de sua ecologia e em sua
conservação, devido ao reconhecimento de seus valores, funções e características
ecológicas singulares. Entre as funções chaves (serviços ambientais) que desempenham
estão: o suporte a biodiversidade (presença de água, elevada produtividade e diversidade
de hábitats), a melhoria da qualidade da água, a prevenção de inundações (armazenam e
diminuem o fluxo da água), atuação na recarga de aqüíferos e o seqüestro carbono
(Zedler & Kersher 2005; Mitsch & Gosselink 1986).
Por serem ambientes influenciados pelo regime hídrico, muitos trabalhos
realizados em áreas úmidas enfocam o papel da variabilidade espacial e temporal das
condições hidrológicas na distribuição dos organismos. Alguns estudos mostram que a
distribuição espacial de espécies de plantas é fortemente determinada pelas suas
habilidades em tolerar ou responder às condições hídricas locais, criando padrões
específicos de distribuição de plantas na paisagem (Barret et al. 2009; Whited et al.
2007; Toth 2005; Sluis & Tandarich 2004; Southal et al. 2003; Capon 2003; Townsend
2001; Kotze & O’Connor 2000; Casanova & Broke 2000; Lessen 1999; Pollock et al.
1998; Robertson et al. 1984). Em condições de inundação a deficiência de oxigênio é a

8
principal causa de estresse para as plantas (Vartapetian & Jackson 1997). A tolerância
dos organismos vegetais varia significativamente entre as espécies e nas diferentes fases
da vida. Entretanto, outros fatores, como a acumulação de liter, a textura, a compactação
e o potencial redox do solo também podem influenciar o estabelecimento de espécies e a
estruturação de comunidades vegetais (Vivian-Smith 1997). O potencial redox do solo
está associado à flutuação hídrica sazonal, tendendo a diminuir entre ambientes secos e
permanentemente inundados, onde as condições anaeróbias do solo aumentam (Dwire et
al. 2006).
Assim, os fatores que atuam estruturando comunidades vegetais e a
determinação de espécies indicadoras destes fatores se tornam importantes na
classificação de habitats (Claussen et al. 2006; Zoltai & Vitt 1995; Gopal & Sah 1995),
ajudam a fundamentar planos de recuperação e manejo destes ambientes, assim como
detectar, por exemplo, efeitos de mudanças nos ciclos de inundação (Goosle et al. 2009;
Finlayson 2005; Toth 2005; Middleton 2002).
Hábitats de áreas úmidas que apresentam sazonalidade nas condições
hidrológicas podem apresentar dinâmicas de sucessão vegetal comandadas por esta
variabilidade temporal (Finlayson 2005; Toth 2005; Capon 2003; Kotze & O’Connor
2000; Bornett & Amoros 1996; van der Valk 1981). Muitas espécies de plantas que
vivem em ambientes sazonalmente inundados dependem desta dinâmica de pulso de
inundação (Junk et al. 1989) para germinarem (van der Valk 1981). Em tais sistemas
ecológicos, o banco de sementes apresenta um papel extremamente importante na
resiliência da vegetação (Broke et al. 2003; Junk et al. 2006a). Portanto, a inundação
sazonal, considerada um distúrbio (Souza 1984), promove flutuações nas comunidades
vegetais devido às mudanças das condições ambientais que influenciam indivíduos e
populações, criando direta ou indiretamente oportunidades para o estabelecimento de
novos indivíduos (Souza 1984). As espécies de plantas herbáceas são importantes
indicadoras desta dinâmica de ambientes de áreas úmidas, pois devido a seus curtos
ciclos de vida, estratégias reprodutivas e rápidas respostas às mudanças nas condições
hídricas locais, permitem estudos para avaliar o papel da variabilidade espacial e
temporal do regime hidrológico na distribuição e dinâmica da vegetação.
A presente tese teve como principal objetivo analisar, em mesoescala, a relação
da variabilidade espacial e temporal da inundação com a variabilidade espacial e
temporal de espécies herbáceas encontradas no Pantanal ajudando assim a entender o

9
papel do pulso de inundação na promoção de riqueza e distribuição de espécies
herbáceas no Pantanal.
O Pantanal matogrossense é considerado uma das mais extensas áreas úmidas do
mundo (Zedler & Kersher 2005), de importância global (Junk et al. 2006b). As
inundações são periódicas e previsíveis, gerando adaptações e estratégias nos
organismos que se utilizam de forma eficiente os atributos da ATTZ, sendo este pulso
de inundação considerado a força motriz que mantém um equilíbrio dinâmico nas
planícies de inundação (Junk et al. 1989; 2005). A variabilidade hidrotopográfica
influencia a distribuição de espécies estruturando comunidades vegetais tolerantes a
diferentes níveis de estresse hídrico, que formam um mosaico de diferenciadas unidades
de paisagens (Nunes da Cunha et al. 2006; Nunes da Cunha & Junk. 2009). Apesar de
existirem muitos estudos enfocando as relações entre distribuição da vegetação e fatores
hidrológicos (Prance & Schaller 1982; Pinder e Rosso 1998; Zeilhofer & Schessl 1999;
Nunes da Cunha & Junk 1999 e 2001), poucos se atêm à dinâmica temporal da
vegetação já verifica (Rebellato & Nunes da Cunha 2005; Heckman 1997; Prado et al.
1994). Durante os vários meses da fase seca, a flora aquática é submetida a um forte
estresse pela falta de água, pois a pluviosidade é muito baixa. Já durante a fase aquática,
a flora terrestre sofre forte estresse pelo excesso de água (Junk & Da Silva 1996).
Entretanto, algumas espécies são comuns às duas fases (Schessl 1999, Rebellato &
Nunes da Cunha 2005). A provável causa da escassez de estudos de dinâmica temporal
da vegetação no Pantanal deve estar relacionada à dificuldade de acesso a áreas
amostrais durante a sua fase úmida. No entanto, o completo entendimento da ecologia
de plantas em sistemas de pulso de inundação necessita de abordagens que contemplem
estes diferentes períodos de seca e inundação. A omissão de observações em uma destas
fases pode também resultar em sub-estimativas da biodiversidade existente (Junk et
al.2006b).
A maior parte do Pantanal é constituída de propriedades particulares, cuja
principal atividade econômica é a pecuária extensiva de corte, desenvolvida na região
há cerca de duzentos anos. Embora esse manejo tradicional e extensivo tenha
contribuído para a conservação do Pantanal, nos últimos anos tem havido crescente
pressão para torná-lo mais competitivo. A busca por competitividade tem intensificado
os sistemas de criação e imposto algumas tecnologias não apropriadas para a região. A
avaliação do impacto destas ações sobre a biodiversidade existente necessita estar
embasado no entendimento do funcionamento do sistema. Um dos impactos está

10
relacionado ao plantio de forrageiras exóticas, principalmente do gênero Brachiaria.
Adicionalmente, registram-se efeitos da redução da capacidade produtiva das fazendas
devido a colonização de espécies lenhosas (Nunes da Cunha & Junk 2001), como
também por divisões constantes de terras por venda ou herança.
Neste sentido, o primeiro passo na conservação e promoção da integridade de
ecossistemas do Pantanal é o entendimento de como diversidade e a distribuição de
espécies herbáceas podem ser afetadas pela inundação. Tal conhecimento ajudará a
fundamentar planos de recuperação e manejo destes ambientes, assim como detectar os
efeitos de mudanças nos ciclos de inundação sobre processos.
REFERÊNCIAS CITADAS
Barret, R., Nielssen, D. L. and Croome, R. 2009. Associations between the plant
communities of floodplain wetlands, water regime and wetland type river research and
applications. Published online in Wiley InterScience (www.interscience.wiley.com).
Bornett, G. and Amoros, C.1996. Disturbance regimes and vegetation dynamics: role of
floods in riverine wetlands. Journal of Vegetation Science 7(5): 615-622
Brock, M. A., Nielsen, D. L., Shiel, R. J, Green, J. D. and Langley, J. D. 2003. Drought
and aquatic community resilience: the role of eggs and seeds in sediments of temporary
wetlands. Freshwater Biology 48:1207–1218.
Capon, S. J. 2003. Plant community responses to wetting and drying in a large arid
floodplain. River research and applications 19:509-520.
Casanova, M. T. and Brock, M. A. 2000. How do depth, duration and frequency of
flooding influence the establishment of wetland plant communities? Plant Ecology 147
(2):237 250.
Claussen, J. C., Ortega, J. M., Glaude, J. M., Relyea, R. A., Garay, G. and Guineo, O.
2006. Classification of wetlands in a Patagonian National Park, Chile. Wetlands 26(1):
217–229.

11
Dwire, K. A., Kauffman, J. B. and . Baham, J. E. 2006. Plant species distribution in
relation to water-table depth and soil redox potential in montane riparian meadows.
Wetlands 26(1):131-146.
Finlayson, C. M. 2005. Plant ecology of Australia’s tropical floodplain wetlands: A
Review. Annals of Botany 96:541–555.
Finlayson, C. M. & van der Valk, A. G. 1995. Wetland classification and inventory: A
Summary. Vegetatio 118: 185-192.
Gopal, B. & Sah, M. 1995. Inventory and classification of wetlands in India. Vegetatio
118: 39–48.
Goslee, S. C., Brooks, R. P. and Cole, C. A. 1997. Plants as indicators of wetland water
source. Plant Ecology 131: 199–206.
Heckman, C. W. 1997. Description of a dinamic ecotone in the Pantanal of Mato
Grosso, Brazil. Ecotropica 3:1-7.
Junk, W. J., Bayley, P. B. & Sparks, R. S. 1989. The flood pulse concept in river –
floodplain systems. In: Proc Int Large River Symp (LARS). D. P. Dodge (ed). Canadian
Journal of Fishers and Aquatic 106: 110-127.
Junk, W. J e Da Silva, C.J. 1999. O conceito do pulso de inundação e suas implicações
para o Pantanal de Mato Grosso. Pp.17-28. In: Anais do II Simpósio sobre Recursos
Naturais e Sócio-econômicos do Pantanal, Manejo e Conservação. Empresa Brasileira de
Pesquisa Agropecuária (Embrapa-CPAP), Corumbá. 537p.
Junk, W.J. 2005.Flood pulsing and linkages between terrestrial, aquatic and wetland
system. Verh. Internat. Verein. Limnol 29: 11-38.
Junk, W. J., Nunes da Cunha, C. N., Wantzen, K. M., Petermann, P., strüssmann, C.,
marques, M. I. & Adis, J. 2006a. Biodiversity and its conservation in the Pantanal of
Mato Grosso, Brazil. Aquatic Science 68:278–309.

12
Junk, W.J., Brown, M., Campbell, I. C., Finlayson, M, Gopal, B, Ramberg, L. and
Warner, B. G. 2006b. The comparative biodiversity of seven globally important
wetlands: a synthesis. Aquatic Science 68: 400–414.
Kotze, D. C. and O’Connor, T. G. 2000. Vegetation variation within and among
palustrine wetlands along an altitudinal gradient in KwaZulu-Natal, South Africa. Plant
Ecology 146(1):77-96
Lenssen, J., Menting, F., van der Putten, W. and Blom , K. 1999. Control of plant
species richness and zonation of functional groups along a freshwater flooding gradient.
Oikos 86(3):523-534.
Middleton, B. A. 2002. The flood pulse concept in wetland restoration. In: Flood
pulsing in wetlands: restoring the natural hydrological balance. Beth A. Middleton (ed.).
John Wiley & Sons, Inc.
Mitsch, W. J. and Gosselink, J. G. 1986. Wetlands. Van Nostrand Reinhold, New York,
USA.539p.
Nunes da Cunha, C. & Junk, W.J. 1999. Composição florística de capões e cordilheiras:
localização das espécies lenhosas quanto ao gradiente de inundação no Pantanal de
Poconé, MT-Brasil. Pp.387-405. In: Anais do II Simpósio sobre Recursos Naturais e
Sócio-econômicos do Pantanal, Manejo e Conservação. Empresa Brasileira de Pesquisa
Agropecuária (Embrapa-CPAP), Corumbá. 537p.
Nunes da Cunha, C. & Junk, W.J. 2001. Distribuition of wood plant communities along
the flood gradient in the Pantanal of Poconé, Mato Grosso, Brazil. International Journal
of Ecology and Environmental 27:63-70.
Nunes da Cunha, C. & Junk, W.J. 2004. Year-to-year changes in water level drive the
invasion of Vochysia divergens in Pantanal grasslands. Applied Vegetation Science
7:103-110.

13
Nunes da Cunha, C., Rawiel, C. P., Wantzen, K. M., Junk, W. J. & Lemes do Prado, A.
2006. Mapping and characterization of vegetation units by means of Landsat imagery
and management recommendations for the Pantanal of Mato Grosso (Brazil), north of
Poconé. Amazoniana XIX(1): 1-32.
Pinder, L. and Rosso, S. 1998. Classification and ordination of plant formations in the
Pantanal of Brazil. Plant Ecology 136: 151–165.
Pollock, M. M., Naiman, R. J. and Hanley, T. A. 1998. Plant species richness in riparian
wetlands - A test of biodiversity theory. Ecology 79:94-105.
Prado, A. L., Heckman, C. W. and Martins, F. R. 1994. The seasonal succession of
biotic communities in wetlands of the tropical wet-and-dry climatic zone: II. The
Aquatic Macrophyte Vegetation in the Pantanal of Mato Grosso, Brazil. Internationale
Revue gesamten Hydrobiologie 79(4): 569-589.
Prance, G. T. and Schaller, G. B. 1982. Preliminary study of some vegetation types of
the Pantanal, Mato Grosso, Brazil. Brittonia 34: 228-251.
Rebellato, L. e Nunes da Cunha, C. 2005. Efeito do “fluxo sazonal mínimo da
inundação” sobre a composição e estrutura de um campo inundável no Pantanal de
Poconé, MT, Brasil. Acta Botanica Brasilica 19: 789-799.
Robertson, P. A., Mackenzie, M. D. & Elliot, L. F. 1984. Gradient analysis and
classification of the woody vegetation for 4 sites in southern Illinois and adjacent
Missouri. Vegetatio 58: 87–104.
Schessl, M. 1999. Floristic composition and structure of floodplain vegetation in
northern Pantanal of Mato Grosso, Brasil. Phyton (Horn, Austria) 39(2):303-3.
Sluis, W and Tandarich, J. 2004. Siltation and hydrologic regime determine species
composition in herbaceous floodplain communities Plant Ecology 173: 115–124.

14
Sousa, P. W. 1984. The role of disturbance in natural communities. Annual Review of
Ecology and Systematics 15:353-391
Southal, E. J., Dale, M. P. and Kent, M. 2003. Spatial and temporal analysis of
vegetation mosaics for conservation: poor fen communities in a Cornish valley mire.
Journal of Biogeography 30:1427–1443.
Toth, L. A. 2005 Plant community structure and temporal variability in a channelized
subtropical floodplain. Southeastern Naturalist 4(3): 393-408.
Townsend, P. A. 2001. Relationships between vegetation patterns and hydroperiod on
the roanoke river floodplain, North Carolina. Plant Ecology 156(1): 43-58.
Vartapetian, B. B. and Jackson, M. B. 1997. Plant adaptations to anaerobic stress. _/
Annal of Botany. 79 (Suppl. A): 3-20.
van der Valk, A. G. 1981. Succession in wetlands: a Gleasonian approach. Ecology
62(3): 689-696.
Vivian-Smith, G. 1997. Microtopographic heterogeneity and floristic diversity in
experimental wetland communities. Journal of Ecology 85(1):71-82.
Zedler, J. B and Kercher, S. 2005. Wetlands resources: status, trends, ecosystem
services, and restorability. Annual Review of Environment and Resources 30:39–74
Zeilhofer, P. and Schessl, M. 1999. Relationship between vegetation and environmental
conditions in the northern Pantanal of Mato Grosso, Brazil. Journal of Biogeography
27: 59-168.
Zoltai, S. C. e Vitt, D. H.1995. Canadian wetlands: environmental gradients and
classification. Vegetatio 118(1/2):131-137. Classification and Inventory of the World's
Wetlands.

15
Whited, D. C., Lorang, M. S., Harner, M. J. Hauer, F. R., Kimball, J. S. and Stanford, J.
A. 2007. Climate, hydrologic disturbance, and succession: drivers of floodplain pattern.
Ecology, 88(4): 940-953.

16
Capítulo 1 – Ordenação de tipologias vegetais e preferência de herbáceas no gradiente de inundação do Pantanal de Poconé-MT
RESUMO – No Pantanal as espécies se agrupam de acordo com sua adaptabilidade aos
diferentes níveis da inundação sazonal. Assim, a ocorrência de diferentes tipologias
vegetais está relacionada a variabilidade hidrotopográfica. Este estudo teve como
objetivo verificar se diferentes sub-tipos de vegetação amostrados formam comunidades
diferenciadas no espaço e no tempo, quais espécies as caracterizam e qual a preferência
das herbáceas em relação ao gradiente de inundação. Este trabalho foi realizado no
Pantanal de Poconé (MT) utilizando-se o sistema RAPELD e metodologia da parcela
pontual para amostragem de herbáceas em 30 isolinhas de 250 m, situadas em
diferentes cotas altimétricas, realizadas nos períodos de seca, cheia e vazante. As 30
isolinhas passavam por vegetação florestal, campestre e savânica e diferiram quanto ao
tempo de inundação. Em campo os diferentes sub-tipos de vegetação foram definidos a
priori para cada isolinha amostrada. Uma ordenação das 30 isolinhas através de uma
NMDS foi realizada utilizando uma matriz contendo dados de cobertura das espécies
nos diferentes períodos. A ordenação agrupou as isolinhas de mesmo sub-tipo de
vegetação e diferentes períodos, estando o primeiro eixo correlacionado com a cota
altimétrica/tempo de inundação. Os grupos definidos a priori (sub-tipos de vegetação)
foram significativamente distintos quando comparados pela Análise de Similaridade
(ANOSIM). Para os grupos com sub-tipos de vegetação mais inundáveis houve
diferenças significativas entre os períodos, demonstrando o efeito da sazonalidade
hídrica sobre composição e estrutura dessas comunidades. Os diferentes sub-tipos de
vegetação possuem espécies dominantes com preferências quanto ao gradiente de
inundação e períodos, o que provavelmente separou e definiu os grupos. Uma ordenação
direta das espécies nos diferentes períodos possibilitou visualizar a preferência destas no
gradiente. Muitas espécies possuem ampla distribuição ao longo do gradiente, enquanto
outras são restritas aos seus extremos, ou seja, os não inundáveis e os muito inundáveis.
Espécies estenobiontes, ou indicadoras de condições ambientais mais restritas, podem
auxiliar na classificação de habitats e no manejo de áreas úmidas, a exemplo, na
detecção de mudanças na estrutura e composição de espécies em função de possíveis
impactos em relação aos padrões do regime hidrológico.
Palavras-chaves: Pantanal, tipos de vegetação, ordenação, gradiente de inundação e
preferência de habitas

17
Chapter 1 – Ordination of vegetation typology and herbaceous plant preferences along the flood gradient of the Pantanal of Poconé, Mato
Grosso.
ABSTRACT – In the Pantanal, species group together according to their adaptability to
the different seasonal flood levels. Thus, the occurrence of different vegetation
typologies is related to the hydro-topographic variability. The aim of this study was to
verify if different sampled vegetation sub-types form communities that are
differentiated in space and time, which species form the communities and what are the
preferences of herbaceous plants in relation to the flood gradient. The study was carried
out in the Pantanal of Poconé (Mato Grosso) using the RAPELD system and the point
sampling method along 30 isolines 250m long, during the dry, flood and drain periods.
The 30 isolines places at varied altimetric positions differed in terms of flood depth and
duration that they were submitted to as well as the vegetation type (forest, meadow and
savannah). In the field, the different vegetation sub-types were defined a priori for each
isoline sampled. The 30 isolines were ordered by means of a NMDS which was carried
out using the data on species coverage in the different periods. The ordination grouped
together the isolines of the same vegetation sub-type and different periods with the first
axis carefully and significantly set in relation to flood depth. The groups defined a
priori (vegetation sub-types) were significantly distinct when tested by the ANOSIM.
For the groups with more floodable vegetation sub-types there were significant
differences between the periods which demonstrated the effect of the seasonal water
cycle on the composition and structure of these communities. The different vegetation
sub-types possess dominant species with preferences in terms of flood gradient and
periods which are probably what separated and defined the groups. Direct ordination of
the species in the different periods enabled visualization of preferences on the gradient.
The majority of species occur in environments with intermediate flood levels followed
by a large contingency of species in non-flooding environments. Species indicators, in
terms of environmental preference, could aid habitat classification and wetland
management, such as, for example, the detection of change in the structure and
composition of species due to possible impacts on the water regime patterns.
Key words: Pantanal, vegetation type, ordination, flood gradient and habitat preference

18
INTRODUÇÃO
O Pantanal está entre as maiores e mais importantes áreas úmidas do mundo
(Junk et al. 2006), sendo considerada uma planície de inundação com padrão de
inundação monomodal e previsível influenciada pelos derrames laterais de rios e/ou
precipitação durante a estação chuvosa (Junk 2005; Hamilton et al. 2002; Junk et al.
1989). Diversas adaptações morfológicas, anatômicas, fisiológicas e comportamentais
são necessárias à biota ali presente, sendo o ciclo de cheia e seca, ou pulso de
inundação, considerado a força que dirige este sistema (Junk et al.1989).
De fato, o Pantanal é uma bacia sedimentar ativa e, apesar de seu relevo
relativamente plano e baixa declividade, apresenta uma microtopografia relacionada a
acontecimentos passados. Durante o período quaternário, oscilações climáticas
alternando períodos secos e úmidos influenciaram os padrões de descargas e deposições
diferenciais de sedimentos carreados pelo rio Paraguai e tributários, produzindo um
mosaico de feições geomórficas diferentemente afetadas pelas inundações, formando
um gradiente de inundação (Nunes da Cunha & Junk 2009; Assine & Soareas 2004). As
espécies vegetais se desenvolvem sobre as unidades geomórficas da planície de acordo
com suas tolerâncias em relação à inundação anual, desenvolvendo diferenciadas
comunidades vegetais que formam as diferentes unidades de paisagens (Nunes da
Cunha et al. 2007). Esta variabilidade ambiental, portanto, produz habitats diferenciados
que suportam, em conjunto, elevada diversidade de animais e plantas (Nunes da Cunha
& Junk 2009).
As diferentes unidades de paisagens diferem no tocante às condições edáficas e
hidrológicas e estão descritas detalhadamente em Nunes da Cunha et al. (2007) e Nunes
da Cunha & Junk (2009). Abaixo estão alguns exemplos de diferentes unidades de
paisagens com as denominações regionais que consideram características
geomorfológicas, hidrológicas e/ou aspectos vegetacionais (Nunes da Cunha & Junk
2009):
Entre as unidades de paisagens não inundáveis, constituídas sobre formas
positivas de relevo, estão as Cordilheiras, caracterizadas por elevações alongadas da
planície (cordões arenosos) originadas à partir de depósitos de sedimentos de antigos
leitos de rios, sendo cobertas por vegetação lenhosa típicas de cerrado (savana),
cerradão (savana florestada) ou floresta decídua; os Capões caracterizados por
elevações que possuem formas arredondadas ou ovais originadas à partir de processos

19
erosivos das cordilheiras, sendo cobertos por vegetação de cerrado ou florestas semi-
decíduas; os Capões de aterro ou Aterros de bugres que são elevações produzidas
artificialmente com diâmetro de dezenas de metros construídas pelas populações
indígenas para promover proteção contra as inundações. Sobre estes aterros há
vegetação arbórea com ocorrência de muitas espécies frutíferas e de produtos não
madereiros dos quais as populações faziam uso. Escavações revelam a presença de
cerâmicas e outros artefatos arqueológicos (Ponce & Nunes da Cunha 1993). Entre as
unidades de paisagens sazonalmente inundáveis, constituídas sobre as formas planas de
relevo, estão os Campos limpos com cobertura de gramíneas, cyperáceas e ervas; os
Campos sujos com cobertura de herbáceas, arbustos e algumas árvores; alguns campos
possuem elevações do terreno chamadas de “murundus”, “cocurutos” ou “monchões”
sobre os quais se desenvolvem elementos arbóreos típicos do cerrado, com cobertura
herbácea nas partes planas, sendo denominados de Campos de murundus (Ponce &
Cunha 1993). Entre outras unidades de paisagens sazonalmente inundáveis,
constituídas sobre as formas negativas de relevo, estão as Vazantes, ocorrentes em
canais rasos de drenagem e temporários cobertos por gramíneas e outras herbáceas.
Quando estes canais rasos de drenagem e temporários são cobertos por vegetação
arbórea, com elevada densidade da espécie Calophyllum brasiliensis, esta unidade de
paisagem é denominada de Landi. Entre as unidades de paisagens permanentemente
inundáveis, também constituídas sobre as formas negativas de relevo, estão as Baías ou
Lagoas, conectadas com os rios durante o período chuvoso e cobertas em suas margens
por macrófitas aquáticas flutuantes e submersas; os Corixos que são pequenos canais na
planície que podem conectar as baías umas com as outras e/ou com o canal do rio,
sendo periodicamente cobertos por macrófitas aquáticas; os Brejos são áreas cobertas
por espécies hidrófilas que permanecem com saturação hídrica do solo durante o
período de seca.
A distribuição das espécies em função do gradiente de inundação é foco de
muitos trabalhos realizados em áreas úmidas. Coenoclinas ou a zonação de
comunidades vegetais perpendicularmente ao gradiente de inundação são padrões
comuns em muitos estudos (Barret et al. 2009; Toth 2005; Sluis & Tandarich 2004;
Capon 2003; Townsend 2001; Kotze & O’Connor 2000; Casanova & Broke 2000;
Lessen 1999). Desta forma, técnicas estatísticas como a ordenação e a classificação são
ferramentas importantes nas análises diretas e indiretas de gradientes ambientais que
comandam a distribuição dos organismos vegetais.

20
No Pantanal estudos confirmam a forte correlação de diferenciados tipos de
vegetação com a duração e a capacidade de drenagem dos solos (Prance & Schaller
1982; Pinder & Rosso 1998; Zeilhofer & Schessl 1999; Nunes da Cunha & Junk 1999 e
2001). Portanto, para fins de estratégias de gestão, estudos devem elucidar as relações
espaço-temporais das comunidades vegetais nos diferentes componentes deste mosaico.
Nos ambientes sazonalmente inundados, por exemplo, a variação da posição
topográfica produz diferenças na amplitude, duração e freqüência da inundação. Em
função dessas diferenças, as comunidades herbáceas apresentam mudanças na
composição e estrutura (Rebellato & Nunes da Cunha 2005; Capon 2003; Prado et al.
1994).
A classificação e o mapeamento das unidades de vegetação da região Norte do
Pantanal de Poconé foram realizadas por Nunes da Cunha et al. (2006) na escala de
1:100.000, sendo a classe Savana (Cerrado) a de maior cobertura na região, com 41,0%,
com distribuições diferenciadas entre as sub-classes, tendo a participação da savana
arborizada com 20,6%, savana gramíneo-lenhosa sazonalmente inundável com 11,9%,
savana parque sazonalmente inundável (campo de murundu) com 7,5% e a savana
florestada (cerradão) com 1,0%. A classe sistemas de transição ocupa o segundo lugar
em cobertura na região, com 14,7%, onde a sub-classe florestas sempre verdes
sazonalmente inundáveis cobrem 12,1%. Uma tipologia preliminar da vegetação
lenhosa do Pantanal foi definida por Nunes da Cunha et al. (2007).
Numerosos estudos com a vegetação foram realizados na região do Pantanal de
Poconé. Os mesmos podem ser divididos em levantamentos de espécies (ex. Schessl
1999), fitossociologia de formações florestais, dentre elas as monodominantes de
Vochysia divergens (Nascimento & Nunes da Cunha 1989, Nunes da Cunha 1990),
caracterização de fatores ambientais determinantes dos diferentes tipos de vegetação
herbacea e arbórea (Zeilhofer & Schessl 1999), sucessão e zonação de herbáceas em
gradientes de inundação (Prado et al. 1994, Heckman 1997); distribuição de espécies
arbóreas em gradientes de inundação (Nunes da Cunha & Junk 1999, Nunes da Cunha
& Junk 2001, Fortney et al. 2004), invasibilidade de Vochysia divergens (Nunes da
Cunha & Junk 2004), a produtividade (Pozer & Nogueira 2004) e fitossociologia
(Rebellato & Nunes da Cunha 2005) das herbáceas dos campos inundáveis em
diferentes períodos hídricos. Portanto, ainda são poucos os estudos que consideram a
sazonalidade, as diferentes tipologias vegetais e preferências de espécies no gradiente
hidrotopográfico do Pantanal. Em função disso, o presente estudo teve como objetivos:

21
1) Realizar um levantamento florístico e fitossociológico em diferentes tipos de
vegetação desta região em diferentes períodos hidrológicos (seca, cheia e vazante);
2) Verificar se os diferentes sub-tipos de vegetação amostrados e definidos a priori em
campo formam grupos significativamente distintos em relação à composição e estrutura
da comunidade herbácea;
3) Verificar quais são as espécies que caracterizam estes sub-tipos de vegetação em
diferentes períodos hidrológicos;
4) Verificar se nos grupos formados há mudanças significativas na composição e
estrutura das comunidades herbáceas entre os diferentes períodos hídricos;
5) Verificar a distribuição preferencial das espécies no gradiente de inundação.
Predições a serem testadas:
1) Os tipos vegetacionais poderão ser discriminados em função das comunidades
de herbáceas (o que indicaria requerimentos ambientais/fisiológicos comuns ou
compartilhados entre várias espécies e/ou refletindo a influência da vegetação de
maior porte na definição das comunidades de herbáceas) e deverão formar um
gradiente que parte das comunidades não inundáveis para as altamente
inundáveis.
2) As variações na composição e abundância das espécies de herbáceas, em função
da variação hidrológica, serão diferenciadas entre tipos vegetacionais: as
variações serão mais pronunciadas nos tipos vegetacionais que experimentam
níveis/tempos de inundação intermediários (os extremos do gradiente
selecionariam espécies pouco ou muito tolerantes a inundação). Essas
comunidades seriam “fechadas” a invasão/manifestação de espécies transitórias
que apareceriam em cotas intermediárias.
3) A amplitude de ocorrência das espécies (indicativa de suas tolerâncias) será
restrita nos extremos e ampla em cotas intermediárias.
MATERIAL E MÉTODOS
Características gerais e área de estudo – O relevo da Bacia do Alto Paraguai possui
cotas altimétricas que oscilam entre 80 a 1200 metros, distribuídas por três regiões
fisiográficas: dos planaltos (250 a 1200 metros), das depressões (150 a 250 metros) e

22
dos pantanais (80 a 150 metros) (Alvarenga et al. 1984). Hidrologicamente é
representada, principalmente, pelos rios Paraguai, Branco, Nabilieque, Miranda,
Aquidauana, Negro, Taquari, Cuiabá, Itiquira, Bento Gomes, São Lourenço e afluentes
(Amaral Filho 1986).
Geomorfologicamente, a região dos pantanais é representada pelas unidades
planícies e pantanais mato-grossenses, onde ocorrem os maiores fenômenos das
enchentes de toda a Bacia do Alto Paraguai (Alvarenga et al. 1984). Essas planícies
deprimidas e inundáveis recebem o nome de Pantanal e se localizam na parte central da
América do Sul, entre longitudes de 16º e 22º e latitudes de 55º e 58º. A declividade é
inexpressiva: o gradiente topográfico varia de 0,3 a 0,5 cm/km no sentido leste-oeste e
0,03 a 0,15 cm/km no sentido norte-sul. Esta região tem grande expressão espacial com
configuração de um enorme anfiteatro voltado para o oeste (Franco & Pinheiro 1982).
A tendência para inundações periódicas não está ligada apenas à pluviosidade,
que é considerada baixa, mas sim à topografia plana, cotas altimétricas baixas e
predominância de solos hidromórficos que influenciam na drenagem (Amaral Filho
1986). Entretanto, a heterogeneidade topográfica da região determina áreas em que a
inundação ocorre de forma distinta no tocante a altura, época e duração das inundações
(Adámoli 1987). Outra característica é que esta vasta superfície rebaixada é recoberta
por sedimentos quaternários, oriundos de diferentes rios que formam a bacia do rio
Paraguai (Adámoli 1987), sendo sua litologia constituída por sedimentos aluviais da
formação Pantanal, que associados à dinâmica do regime de alagamento, provocam
grande variação nos solos. Ao norte predominam os solos com horizonte subsuperficial
de textura mais argilosa, na parte central, solos mais arenosos, e ao sul, solos argilosos
(Amaral Filho 1986). A heterogeneidade promovida pelos diferentes tipos de solos e
regime hídrico, que determinam diferenças fitogeográficas, levaram à divisão do
Pantanal em sub-regiões. Silva & Abdon (1998), por exemplo, delimitam o Pantanal em
138,183 km2 e o dividem em 11 sub-regiões, sendo a sub-região de Poconé (Pantanal
Norte), onde o presente estudo foi realizado, caracterizada pelo clima do tipo AW-
quente e úmido (Köppen 1948), com precipitação média de 1250 mm ao ano e
temperatura média anual de 25,8 oC. No Pantanal Norte duas estações são observadas
durante o ano, sendo uma seca no período de maio a setembro e a outra chuvosa, de
outubro a abril (Nunes da Cunha & Junk 2004). Prado et al. (1994) delimitaram quatro
fases hidrológicas do ano no Pantanal Norte: enchente (de outubro a dezembro), cheia
(de janeiro a março), vazante (de abril a junho) e seca (de julho a setembro). No

23
Pantanal Norte a inundação é do tipo monomodal ocorrendo juntamente com a estação
chuvosa (outubro a abril) (Hamilton et al. 1996). Os solos são argilosos em grande
parte, com grau de fertilidade superior à dos outros pantanais (Adámoli 1987). Possui
aportes aluviais originários quase que totalmente do rio Cuiabá com transporte e
deposição de argila, silte e areia derivados principalmente de filitos e xistos. Apresenta
grande variedade de tipos de solos, com imensas áreas de solo hidromórfico cinzento
eutrófico, vertissolos, laterita hidromórfica distrófica e planossolo distrófico (Amaral
1987). Ao norte, apresenta extensa faixa com fraca inundação, onde as cotas
altimétricas excedem 130 metros. Em direção ao sul as áreas são bastante inundadas,
pois as cotas altimétricas decrescem para 110-100 metros, (Franco e Pinheiro 1982). Na
presente área de estudo altimetria ficou entre 115 e 112 metros.
Para a parte norte do Pantanal de Poconé a savana (cerrado) é a classe
fitofisionômica que cobre a maior parte da região (41,0%), seguida pelos sistemas de
transição (ex. florestas semi-sempre verdes sazonalmente inundáveis, com 14.7%),
florestas sazonais semideciduais (6.2%), sistemas secundários (3. 3%), corpos d’água
(0.8%) e florestas sazonais decíduas (0.4%) (Nunes da Cunha et al. 2006).
A área em que foram realizadas as coletas engloba três fazendas: Nossa Senhora
da Aparecida, São Carlos e Canoas, próximas à localidade do Pirizal, município de
Nossa Senhora do Livramento (Oeste: 56º 21', Leste: 56º 18', Norte: 16º 19', Sul: 16º
22') (Fig. 1).

24
Figura 1- Área de estudo próxima à localidade do Pirizal, município de Nossa Senhora
do Livramento, sub-região do Pantanal de Poconé-MT (Pinho 2005).
Desenho amostral – Este trabalho foi realizado utilizando-se do método RAPELD.
Este é uma modificação do método de Gentry (1982), desenvolvido para inventários
rápidos da biodiversidade em sítios de pesquisa ecológica de longa duração na
Amazônia (Magnusson et al. 2005), implementado no Pantanal de Poconé. Na área de
estudo foram instalados 30 isolinhas de 250 metros sistematicamente distribuídas em

25
um sistema de seis trilhas de 5 km cada orientadas no sentido norte-sul, ocupando uma
área de 25 Km2. Cada trilha possui uma identificação (A, B, C, D, E, F) onde foram
instaladas 5 isolinhas (Fig. 2), orientadas no sentido leste-oeste. A primeira isolinha de
cada uma das trilhas inicia-se a partir de 500 metros do início das mesmas. Assim, por
exemplo, a isolinha A1 está situada a 500 metros do início da trilha A. Cada isolinha
dista 1 Km uma da outra e foram implantadas em diferentes cotas altimétricas. A
marcação das mesmas foi feita com piquetes de ferro fixos no solo a cada 10 m, sendo a
altimetria do ponto inicial mantida ao longo dos 250 metros.
ABCDEF
500
1000
1500
2000
3000
2500
3500
4000
4500
5000
Figura 2 - Imagem de satélite mostrando a área de estudo (5 km x 5 km) com sistemas
de 6 trilhas (apenas Norte-sul) e esquema com distribuição das isolinhas de 250m
metros nas trilhas (A, B, C, D, E e F), Pantanal de Poconé-MT.
Amostragem de herbáceas – As coletas foram realizadas durante três fases
hidrológicas do Pantanal: seca (setembro/outubro de 2006), cheia (fevereiro/março de
2007) e vazante (maio de 2007). A amostragem das herbáceas foi realizada pelo método
da parcela pontual (Bullock 1996). Ao longo dos 250 metros de cada isolinha foram
distribuídas parcelas pontuais distantes 1 metro uma da outra. A parcela pontual é
considerada uma haste de metal fina posicionada verticalmente, onde qualquer
morfoespécie de herbácea que a toca é registrada. Um exemplar de cada morfo espécie
foi coletado, prensado e seco em estufa, sendo posteriormente enviado a especialistas
para identificação. Futuramente as exsicatas serão depositadas no Herbário Central da
1 km 5 km
Norte
Sul
Leste Oeste
1 km
F E D C B A
5 km

26
Universidade Federal de Mato Grosso (UFMT) e Universidade Federal de Minas Gerais
(UFMG).
Identificação da tipologias vegetacionais - Os tipos e sub-tipos de vegetação foram
classificados em função de sua fisionomia, espécies presentes, regime de inundação e
com o auxílio das descrições encontradas em Nunes da Cunha et al. (2007). A
nomenclatura dos sub-tipos também foi baseada naquela utilizada em Nunes da Cunha
et al. (2007).
Análises estatísticas – As contagens das espécies herbáceas nas parcelas pontuais
permitiram calcular suas coberturas nas 30 isolinhas nos períodos de seca, cheia e
vazante. Os dados de profundidade e duração da inundação foram coletados
quinzenalmente entre os anos de 2006 e 2007 por Fantin-Cruz (2008). As medições
foram realizadas nos marcos iniciais de cada isolinha (mourão de madeira) que
continham réguas hidrológias. O tempo de inundação variou de 0 a 200 dias (média =
90,3 dias, dp = 50,3 dias; mediana = 90 dias) e a profundidade da inundação variou
entre 0 e 63 cm (média = 25,067 cm, dp = 16,144; mediana = 24 cm). O tempo de
inundação é fortemente correlacionado com a profundidade da inundação (r2 = 0,913, p
< 0.001, n = 30).
O número de parcelas pontuais em que cada espécie ocorreu foi utilizado para o
cálculo de sua cobertura na isolinha (número de parcelas pontuais na qual a espécie
ocorreu / 250), sendo calculado para as 30 isolinhas nos três períodos.
Os cálculos da cobertura das espécies nos diferentes sub-tipos de vegetação
foram realizados da seguinte forma: (soma do número de parcelas pontuais nos quais a
espécie ocorreu / total de parcelas pontuais ocorrentes naquele sub-tipo de vegetação x
250 (= total de ocorrências / no. de isolinhas naquele sub-tipo de vegetação x 250).
Um Escalonamento Multidimensional Não-Métrico (NMDS) (Cox & Cox 2001)
foi usado para ordenar as isolinhas em um gradiente determinado pela similaridade na
cobertura das espécies herbáceas. As diferentes estações foram analisadas
conjuntamente, o que permite detectar possíveis mudanças nas similaridades das
isolinhas ao longo do tempo. NMDS tem sido uma das técnicas de ordenação mais
indicadas em análises de estruturas de comunidade, pois não requer distribuição normal
multivariada e não pressupõe relações lineares entre as variáveis (McCune & Grace
2002). Devido às grandes diferenças nos valores das coberturas, estas foram

27
estandardizadas e logaritmizadas (log (x+1)) antes do cálculo das distâncias de Bray-
Curtis, medida amplamente utilizada em análises de comunidades (McCune & Grace
2002). O teste de permutação de Monte Carlo foi realizado para verificar se os eixos da
NMDS são significativos. Para avaliar a relação entre a similaridade florística e a
distância geográfica ou a autocorrelação espacial, foi feito o teste de Mantel (1000
randomizações) utilizando-se para isso a matriz de cobertura de espécies de cada
período e a matriz de coordenadas geodésicas dos pontos iniciais de cada isolinha. Para
todas estas análises foi utilizado o pacote estatístico PCOrd 4.01 (McCune & Mefford
1999).
Para testar a influência da variável ambiental profundidade e/ou duração da
inundação na estrutura e composição de espécies utilizou-se Regressões lineares e não
lineares, tendo como variáveis dependentes os escores de cada um dos 2 primeiros eixos
da NMDS.
Posteriormente, uma análise de similaridade (ANOSIM) foi realizada para testar
i) possíveis diferenças entre os grupos formados pela NMDS, considerando-se todos os
períodos conjuntamente e ii) possíveis diferenças temporais em cada grupo (ex:
comparando todas as isolinhas de campos de murundus entre os períodos de seca, cheia
e vazante). ANOSIM é considerada o equivalente não-paramétrico da MANOVA e se
baseia no cálculo de um parâmetro estatístico (R global) de ranks de similaridade entre e
dentro dos grupos definidos a priori, com 10000 permutações. Usualmente R varia de 0
a 1 - valores próximos a 0 indicam a mesma similaridade, em média, entre e dentro dos
grupos, enquanto que valores acima de 0 indicam algum grau de discriminação entre os
grupos (Clarke 1993). Tanto para a análise entre grupos e para cada grupo entre os
diferentes períodos foi utilizada a medida de distância de Bray-Curtis. O programa
utilizado foi o PAST 1.79 (Hammer et al. 2001).
Um ordenamento direto de 40 espécies de herbáceas ao longo do gradiente de
profundidade de inundação nos três períodos foi representado graficamente. Para tanto,
utilizou-se o programa COMUNIDATA desenvolvido por Dias (2006).
RESULTADOS
Tipos de vegetação –Na área amostrada foram identificados três tipos de vegetação:
floresta sazonal, savana e campo (Tab. 1), sendo que as espécies herbáceas são bem
representadas na savana e o campo.

28
A savana apresenta nas áreas planas um tapete herbáceo com dominância de
gramíneas mais esclerófilas, entouceiradas (bunchgrasses), com altura mediana, perenes
e com rota fotossintética do tipo C4. Esta savana é sazonalmente inundável,
principalmente por precipitação local, ocorrendo na área de estudo em uma faixa de
profundidade de inundação de 14 a 44 cm, com duração de 90 a 125 dias. Nas áreas
planas são encontradas elevações (murundus, cocurutos ou monchões) formadas pela
acumulação de partículas argilosas devido à atividade de térmitas, podendo ter de 1 a 80
m2 de tamanho e cerca 0,3 a 1 metro de altura acima do nível das áreas planas (Oliveira-
Filho 1992; Schessl 1999), estando livres da inundação, podendo ser cobertos por
espécies do Cerrado (Nunes da Cunha et al. 2007; Oliveira-Filho 1992). Uma hipótese
biótica e outra abiótica explicam a formação dos murundus, e podem ser vistas na
revisão de Ponce e Nunes da Cunha (1993). Este sub-tipo de vegetação foi denominado
de savana sazonalmente inundável com térmitas (Nunes da Cunha et al. 2007),
popularmente denominada de campos de murundus.
Os campos ou campos limpos sazonalmente inundáveis são caracterizados por
inundações mais pronunciadas sob influência direta de cursos d`água associados aos
landis, ocorrendo na área de estudo em apenas uma isolinha com inundação de maior
profundidade, 63 cm, e duração de 200 dias. Esses campos apresentam um tapete
gramínoso composto por espécies menos esclerófilas e mais baixas, com proporção um
pouco maior de cobertura por espécies anuais e rota fotossintética do tipo C3.
Áreas inundáveis com diferentes tipos de vegetação sofreram alterações
antrópicas para a formação de campos com gramíneas exóticas, onde o plantio de
Brachiaria humidicola se destaca dada a sua adaptabilidade à inundação, constituindo-
se uma vegetação campestre, sendo este sub-tipo de vegetação denominado aqui de
campo exótico sazonalmente inundável. Na área de estudo os campos exóticos
ocorreram na faixa de profundidade 15 a 58 cm, com inundação durando entre 42 a 200
dias.
Além dos tipos de vegetação dominados por gramíneas (savana sazonalmente
inundável com térmitas, campo limpo sazonalmente inundável e campo exótico
sazonalmente inundável), foram classificados dois sub-tipos de floresta sazonal: as
florestas semi-sempre verdes sazonalmente inundáveis (florestas inundáveis) com
dominância de Calophyllum brasiliense (landizal), ou de Vochysia divergens
(cambarazal) ou de Licania parvifolia (pimenteira) e as florestas sazonais semi-
decíduas sobre solo seco (cerradões). O primeiro sub-tipo de floresta sazonal é formado

29
por espécies arbóreas consideradas colonizadoras de áreas abertas e inundáveis, sendo
constituídas, principalmente, por Vochysia divergens (cambarazal) e Licania parvifolia
(pimenteira) (Pott & Pott 1994; Nunes da Cunha & Junk 2004). A sinúsia herbácea se
torna menos densa e varia de acordo com o grau de invasão pelas espécies acima
citadas. Na área de estudo estas florestas ocorreram em uma faixa de profundidade de
inundação de 16 a 50 cm, com duração variando de 90 a 150 dias. O segundo sub-tipo
de floresta sazonal não é inundável, pois se encontra sobre áreas mais elevadas na
planície, uma geoforma denominada de cordilheira, que apresenta uma cobertura da
sinúsia herbácea média. Entretanto, em anos com inundações mais pronunciadas estas
podem ser inundadas. Nas inundações normais apresenta um lençol freático bastante
superficial, o que elimina espécies arbóreas sensíveis à saturação hídrica (Nunes da
Cunha et al. 2007).
Três isolinhas amostradas ocorreram entre dois sub-tipos vegetacionais, sendo
todos esses ambientes denominados de interfaces: dois ocorreram entre campo de
murundus e cerradão, e o outro entre campo limpo inundável e cerradão. A
profundidade de inundação destes ambientes variou de 7 a 26 cm, com duração entre 6 e
90 dias.
A savana sazonalmente inundável com térmitas (campo de murundus), também
denominada de savana parque, foi a mais freqüente nas trilhas do sistema RAPELD
instalado no nordeste do Pantanal de Poconé, com 40% de ocorrência (12 isolinhas),
seguida da floresta semi-sempre verde sazonalmente inundável, com 20% de ocorrência
(6 isolinhas), campo exótico sazonalmente inundável, com 16,7% (5 isolinhas),
interface, com 10% (3 isolinhas), floresta sazonal semi-decídua sobre solo seco
(cerradão), com 10% (3 isolinhas) e campo limpo sazonalmente inundável, com 3,3% (1
isolinha).
Todos estes ambientes amostrados nos diferentes períodos reuniram um total de
209 espécies, distribuídas em 45 famílias.

30
Tabela 1 – Sub-tipos de vegetação e valores de duração (dias) e profundidade da
inundação máxima (cm) das 30 isolinhas amostradas, Pantanal de Poconé-MT.
Isolinhas Código Sub-tipos de vegetação Duração
(dias) Profundidade
(cm)
A1 Flo Floresta semi-sempre verde sazonalmente inundável 111 30
A2 Flo Floresta semi-sempre verde sazonalmente inundável 104 42
A3 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
90 17
A4 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
104 21
A5 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
104 25
B1 Ce campo exótico sazonalmente inundável 42 15
B2 Cer Interface: campo exótico/floresta sazonal semi-decídua sobre solo seco (cerradão)
0 0
B3 Flo Floresta semi-sempre verde sazonalmente inundável 154 50
B4 Inter Interface: Savana sazonalmente inundável com térmitas (campo de murundus)/ floresta sazonal semi-decídua sobre solo seco (cerradão)
6 7
B5 Ce campo exótico sazonalmente inundável 56 16 C1 Ce campo exótico sazonalmente inundável 89 23
C2 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
90 16
C3 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
104 25
C4 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
110 29
C5 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
90 14
D1 Ce campo exótico sazonalmente inundável 200 58
D2 Cam Campo limpo sazonalmente inundável com invasão inicial de V. divergens
200 63
D3 Flo Floresta semi-sempre verde sazonalmente inundável 126 35
D4 Flo Floresta semi-sempre verde sazonalmente inundável 90 16
D5 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
90 26
E1 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
125 44
E2 Cer floresta sazonal semi-decídua sobre solo seco (cerradão)
0 0
E3 Inter Interface: campo limpo/ floresta sazonal semi-decídua sobre solo seco (cerradão)
90 26
E4 Inter Interface: Savana sazonalmente inundável com térmitas (campo de murundus)/ floresta sazonal semi-decídua sobre solo seco (cerradão)
23 7
E5 Flo Floresta semi-sempreverde sazonalmente inundável 125 39
F1 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
95 42
Continua...

31
Continuação
Tab. 1
F2 Cer floresta sazonal semi-decídua sobre solo seco (cerradão)
0 0
F3 Ce campo exótico sazonalmente inundável 110 18
F4 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
90 24
F5 Mur Savana sazonalmente inundável com térmitas (campo de murundus)
90 24
Com exceção do campo limpo sazonalmente inundável (n=1) e da floresta
sazonal semi-decídua sobre solo seco (não inundável) a profundidade e duração média
da inundação de cada sub-tipo de vegetação pode ser observada na figura 3.
0
10
20
30
40
50
60
70
cer inter mur ce flo cam
Pro
fundid
ade m
édia
da
inundação (
cm
)
0
50
100
150
200
250
Dura
ção m
édia
da
inundação (
dia
s)
Profundidade média Duração média
Figura 3 – Valores de profundidade e duração média para cada sub-tipo de vegetação,
Pantanal de Poconé – MT.
Ordenação das comunidades herbáceas– Os três eixos do NMDS capturaram grande
parte da variação das distâncias originais (r2=0,816) e o valor de stress (13.63) indica
que a ordenação pode ser considerada satisfatória (McCune e Grace 2002). O teste de
Monte Carlo foi significativo para os três eixos (p = 0.032). O primeiro eixo foi
correlacionado positivamente e de forma linear com a profundidade da inundação (r2 =
0.74; p = 0.00001) (Fig.4). Já o eixo 2 também foi correlacionado positivamente com a
profundidade da inundação de maneira não linear, mas quadrática (r2 = 0.70; p =
0.00001) (Fig.4). Não foram encontradas correlações significativas entre a similaridade
florística e a distância geográfica entre as 30 isolinhas amostradas para os diferentes
períodos (seca: r=0,039 e p=0,26; cheia: r=-0,012 e p=0,46; vazante: r=0,017 e p=0,36),

32
sendo assim, as amostras possuem independência estatística, ou seja, não são
consideradas pseudoréplicas. Observa-se também que a maior parte das isolinhas se
agrupam segundo o sub-tipo de vegetação (Fig.4), dadas às suas semelhanças em termos
de estrutura e composição de espécies, estando estas ordenadas positivamente ao longo
do primeiro eixo, ou seja, no gradiente de profundidade.
As diferenciadas faixas de profundidade em que os sub-tipos de vegetação
ocorrem produzem um certo espalhamento das isolinhas de cada sub-tipo de vegetação
ao longo do primeiro eixo, provavelmente, em função dos efeitos da profundidade da
inundação sobre a ocorrência e a cobertura das espécies que as caracterizam. As
florestas semi-sempre verdes sazonalmente inundáveis, por exemplo, possuem uma
distribuição mais ampla ao longo do gradiente, tendo algumas isolinhas agrupadas
juntamente com as savanas inundáveis com térmitas, ou em ambientes muito
inundáveis, onde também há a ocorrência do campo limpo sazonalmente inundável. As
isolinhas dos campos exóticos inundáveis também apresentam uma certa desagregação,
pois estes são formados sobre diferentes tipos de vegetação e cotas altimétricas,
guardando um pouco de sua identidade anterior, tendo algumas isolinhas semelhanças
maiores com as savanas inundáveis com térmitas, enquanto outras, com o campo limpo
ou florestas semi-sempre verde sazonalmente inundáveis.
Mesmo assim, a significância das diferenças entre grupos foi confirmada pela
ANOSIM (R global = 0,7486; p= 0,0001) (Tab. 2), demonstrando, portanto, que os
grupos definidos a priori em campo, e visualmente representados pelo gráfico de
dispersão na NMDS, são significativamente distintos. Na tabela 3 estão apresentados os
valores detalhados de R entre os diferentes sub-tipos de vegetação, onde observa-se a
maior dissimilaridade entre a floresta sazonal semi-decídua sobre solo seco e os demais
sub-tipos de vegetação, justamente por esta não ser inundável. O valor de R igual a 1
entre a floresta sazonal semi-decídua sobre solo seco e o campo limpo sazonalmente
inundável revela a total dissimilaridade entre estes dois sub-tipos.
Mudanças temporais - Dentro de cada grupo as mesmas isolinhas amostradas em
diferentes períodos não se mantiveram estáticas frente às mudanças hídricas sazonais,
havendo deslocamentos que as separam entre os períodos. Entretanto, uma estaticidade
está presente principalmente nas isolinhas de florestas sazonais semi-decíduas não
inundáveis. As savanas sazonalmente inundáveis com térmitas, assim como algumas
interfaces, também apresentam certa estaticidade. Aparentemente, aquelas isolinhas

33
localizadas à direita do gráfico da NMDS (Fig. 5), sofrem os maiores deslocamentos
entre os diferentes períodos. Algumas destas isolinhas sofrem mudanças notáveis em
termos de composição e estrutura de espécies, pois observa-se variações relativamente
grandes entre suas distâncias, a exemplo, estão as isolinhas A1 e B3.
A ANOSIM realizada para cada grupo entre os diferentes períodos constatou que
para alguns sub-tipos de vegetação essa variação foi significativa, como no caso dos
campos exóticos sazonalmente inundáveis, florestas semi-sempre verdes sazonalmente
inundáveis e savanas sazonalmente inundáveis com térmitas (Tab. 3). As florestas
sazonais semi-deciduais sobre solo seco (cerradões) e interfaces não demonstraram
mudanças significativas, sendo, portanto, ambientes mais estáveis ao longo do ano.
Portanto, apenas os ambientes com inundações mais pronunciadas sofrem mudanças
mais acentuadas em termos de composição e estrutura ao longo do ciclo hidrológico. As
isolinhas localizadas nas interfaces são menos inundáveis, fazendo limite com as
florestas sazonais semi-deciduais sobre solo seco (cerradões). A exceção é a interface
E3 (E2500), que tem uma maior profundidade de inundação, pois faz fronteira entre
uma floresta sazonal semi-decídual sobre solo seco (cerradão) e um campo limpo
sazonalmente inundável, ficando no gráfico da NMDS (Fig. 5) entre estes dois sub-tipos
de vegetação.

34
0 10 20 30 40 50 60 70
Profundidade (cm)
-2
-1
0
1
2
Esco
res d
o E
ixo
I
B2vB2cE2s
B2s
F2c
E2c
F2s
F2v
E2vB4vE4s
E4c
E4v
B4s
B4cC5v C5s
C5c
B1c
B1s
B1v
B5c
C2c
B5s
B5vD4s
D4v
C2v
C2s
D4c
A3s
A3c
A3v
F3s
F3vF3c
A4c
A4v
A4s
C1v
C1s
C1c
F5s
F5c
F4s
D2s
D2c
D2v
D1v
D1s
D1c
B3c
B3v
B3s
E1v
E1s
E1c
F1v
F1s
A2s
A2c
F1c
A2vE5c
E5v
E5s
D3s
D3c
D3vA1c
A1v
A1s
C4v
C4c
C4s
D5c
D5v E3s
D5s
E3v
E3c
C3v
C3c
A5c
C3s A5s
A5v
F4c
F4v
F5v
0 10 20 30 40 50 60 70
Profundidade (cm)
-2
-1
0
1
Esco
res d
o E
ixo
II
B2vB2c
E2s
B2s
F2c
E2c
F2sF2v
E2v
B4v
E4s
E4c
E4v
B4sB4c
C5vC5s
C5c
B1c
B1s
B1v
B5cC2c
B5s
B5v
D4s
D4v
C2v
C2s
D4cA3s
A3c
A3v
F3sF3v
F3c
A4c A4vA4s
C1v
C1s
C1c
F5s
F5cF4s
D2s
D2c
D2vD1v
D1s
D1c
B3c
B3v
B3s
E1v
E1s
E1c
F1vF1s
A2s
A2c
F1c
A2v
E5c
E5v
E5s
D3s
D3c
D3v
A1c
A1v
A1s
C4v
C4c C4sD5c D5v
E3s
D5s
E3vE3c
C3vC3c
A5c
C3s
A5sA5v
F4cF4v
F5v
Figura 4 – Relações entre os escores do eixo 1 e 2 com a profundidade da inundação,
Pantanal de Poconé-MT. (s = seca, c=cheia e v = vazante).

35
Figura 5 – Ordenação pelo NMDS das 30 isolinhas amostradas nos períodos de seca, cheia e vazante e classificados a priori em relação ao sub-
tipo de vegetação, Pantanal de Poconé-MT. (s = seca, c=cheia e v = vazante). As setas indicam deslocamentos de isolinhas em função do período,
devido à mudanças de estrutura e composição de espécies.

36
Tabela 2 – Valores de R (global) da Análise de Similaridade (ANOSIM) e significância (p)
entre os diferentes sub-tipos de vegetação e dentre os mesmo sub-tipos entre os períodos de
seca, cheia e vazante, Pantanal de Poconé-MT. ce = campo exótico sazonalmente inundável, cer = floresta
sazonal semi-decídua sobre solo seco, flo = floresta semi-sempre verde sazonalmente inundável, inter = interfaces entre
ambientes inundáveis e não inundáveis, mur = savana sazonalmente inundável com térmitas e cam = campo limpo
sazonalmente inundável.
Entre períodos para cada sub-tipo de vegetação
Entre diferentes sub-tipos de vegetação para todos os períodos ce cer flo inter mur cam
Rank médio dentro dos grupos
819,6 43,27 23,33 59,36 34,56 250,6 *
Rank médio entre os grupos
2378 56,89 16,89 84,35 33,1 345,2 *
R 0,778 0,2596 -0,358 0,3267 0,0044 0,3002 *
p 0,0001 0,0081 0,9926 0,0007 0,5591 0,0001 *
Tabela 3 – Valores de R entre os diferentes sub-tipos de vegetação, Pantanal de Poconé-MT.
(todos os Rs foram significativos, com p< 0.0047). ce = campo exótico sazonalmente inundável, cer =
floresta sazonal semi-decídua sobre solo seco, flo = floresta semi-sempre verde sazonalmente inundável, inter = interfaces
entre ambientes inundáveis e não inundáveis, mur = savana sazonalmente inundável com térmitas e cam = campo limpo
sazonalmente inundável.
ce cer flo inter mur cam ce
cer 0.9208
flo 0.668 0.9732
inter 0.8696 0.8621 0.3924
mur 0.7838 0.9934 0.751 0.5569
cam 0.9663 1 0.5394 0.7854 0.994
Espécies dominantes e riqueza por sub-tipo de vegetação – Na tabela 4 verifica-se que em
todos os sub-tipos de vegetação as famílias Poaceae e Cyperaceae foram dominantes.
Observa-se também o incremento de espécies no sentido seca/vazante. As médias da riqueza
entre as isolinhas de um mesmo sub-tipo de vegetação para os três períodos colocam as áreas
de florestas sazonais semi-decíduas sobre solos seco e interfaces entre as mais ricas, pelo
menos nos períodos de seca e vazante.

37
Tabela 4 – Sub-tipos de vegetação, riqueza total em espécies e famílias dominantes. Para os diferentes períodos: cobertura das espécies dominantes, riqueza total, riqueza média, desvio padrão e número de cada sub-tipo de vegetação amostrado (n). ce = campo exótico sazonalmente inundável, cer = floresta sazonal semi-decídua sobre solo seco, flo = floresta semi-sempre verde sazonalmente inundável, inter = interfaces entre ambientes inundáveis e não inundáveis, mur = savana sazonalmente inundável com térmitas e cam = campo limpo sazonalmente inundável.
Sub-tipo de vegetação
Total de espécies
Famílias dominantes Períodos Espécies dominantes Riqueza
total Riqueza média
dp n
Poaceae (23), Cyperaceae (15), Euphorbiaceae (6) e Rubiaceae (5)
Seca B. humidicola (0.435); A. purpusii (0.063);
P. lineare (0.040); R. brasiliensis (0.026) 33 11.000 7.036 5
ce 79 Cheia B. humidicola (0.666); E. minima (0.168);
U. foliosa (0.138); E. acutangula (0.090) 50 19.000 5.523 5
Vazante B. humidicola (0.558); E. minima (0.119);
E. acutangula (0.075); P. lineare (0.065) 61 22.000 5.244 5
Poaceae (27), Cyperaceae (15), Leguminosae (10) e Lamiaceae (4)
Seca B. balansae (0.199); E. muticus (0.113);
B. humidicola (0.069); I. brasiliensis (0.044) 41 17.667 4.163 3
cer 93 Cheia B. balansae (0.227); E. muticus (0.139);
B. humidicola (0.064); I. brasiliensis (0.052) 53 21.667 8.386 3
Vazante B. balansae (0.212); E. muticus (0.112);
B. humidicola (0.064); I. brasiliensis (0.056) 66 26.667 7.024 3
Poaceae (25), Cyperaceae (10), Euphorbiaceae (5), Rubiaceae (5) e Alismataceae (5)
Seca
A. purpusii (0.052); P. lineare (0.018);
A. leptostachyus (0.012); C. glandulosus
(0.011)
44 12.500 2.429 6
flo 81 Cheia E. minima (0.121); U. foliosa (0.069);
C. furcata (0.052); E. acutangula (0.029) 48 18.500 6.058 6
Vazante E. minima (0.060); C. furcata (0.032); A.
purpusii (0.025); P. lineare (0.021) 53 16.500 5.468 6
Poaceae (29), Cyperaceae (11), Rubiaceae (9) e Euphorbiaceae (5)
Seca A. purpusii (0.152); R. grandiflora (0.100);
P. lineare (0.069); H. brevipes (0.044) 42 19.667 6.110 3
inter (inundáveis/não
inundáveis 90 Cheia
A. purpusii (0.155);P. lineare (0.083);
S. guyanensis (0.067); E. geniculata (0.065) 54 26.000 6.000 3
Vazante A. purpusii (0.165); P. lineare (0.061);
R. brevirostris (0.032); P. laxum (0.029) 59 28.000 5.196 3
continua......

38
continuação Tab. 4
Poaceae (31), Cyperaceae (14), Rubiaceae (8) e Euphorbiaceae (6)
Seca A. purpusii (0.270); P. lineare (0.269);
B. humidicola (0.026); P. stenodes (0.023) 49 13.833 5.006 12
mur 101 Cheia E. minima (0.278); P. lineare (0.277);
A. purpusii (0.202); U. foliosa (0.121) 70 25.083 5.230 12
Vazante P. lineare (0.266); A. purpusii (0.264);
E. minima (0.125); B. humidicola (0.056) 70 23.667 5.416 12
Poaceae(10),Cyperaceae (5), Euphorbiaceae (3) e Fabaceae (3)
Seca R. brasiliensis (0.308), S. geniculata (0.240),
M. arenosa (0.084) e D. kuntzei (0.044) 23
* * 1
cam 41 Cheia E. acutangula (0.456), D. kuntzei (0.104), S.
rhombifolia (0.092) e M. sellowiana (0.084) 26 * * 1
Vazante
C. furcata (0.156), E. acutangula (0.152), M.
sellowiana (0.096) e D. kuntzei (0.060) 26 * * 1

39
Distribuição das espécies herbáceas relacionadas aos sub-tipos de vegetação – Os diferentes
sub-tipos de vegetação inundáveis compartilham um grande número de espécies, entretanto,
possuem espécies dominantes que os caracterizam nos diferentes períodos (Tab. 4, Fig. 6). Na
figura 6 os sub-tipos de vegetação estão ordenados de forma crescente em função da
profundidade média da inundação em que ocorreram, com exceção do campo limpo
sazonalmente inundável que não possui média (n=1) mas que, entretanto, ocorreu na isolinha
de maior profundidade. As florestas semi-sempre verdes sazonalmente inundáveis apresentam
baixa cobertura de espécies herbáceas em função de efeitos do sombreamento, já que em geral
no Pantanal há um elevado contingente de espécies herbáceas do bioma Cerrado e, portanto,
adaptadas à elevada incidência solar. Já nas florestas semi-decíduas a cobertura de espécies
herbáceas é menos afetada pelo sombreamento de espécies de porte arbóreo, provavelmente
pela maior deciduidade das folhas, permitindo uma maior incidência solar. A presença de B.
humidicola (gramínea exótica) neste sub-tipo de vegetação não ocorreu de forma espontânea e
se deve ao plantio da mesma em parte da cordilheira em que foi instalada a isolinha B2. Isto
demonstra a habilidade desta espécie em ocupar tanto ambientes inundáveis como não
inundáveis.

40
Figura 6 – Cobertura das principais espécies herbáceas nos diferentes sub-tipos de vegetação nos períodos de seca, cheia e vazante, Pantanal de Poconé – MT. cer = floresta sazonal semi-decídua sobre solo seco (média 0); inter = interfaces entre ambientes inundáveis e não inundáveis (média 13,3), mur = savana sazonalmente inundável com térmitas (média 25,6); ce = campo exótico sazonalmente inundável (média 26), flo = floresta semi-sempre verde sazonalmente inundável (média 35,3), e cam = campo limpo sazonalmente inundável (63 cm).
Entre os ambientes inundáveis e não inundáveis há compartilhamento de 34 espécies.
Entretanto, muitas espécies estão distribuídas de forma restrita no gradiente de inundação,
com maior número de exclusivas dos ambientes não inundáveis (floresta sazonal semi-
decídua sobre solo seco) (Fig.7).
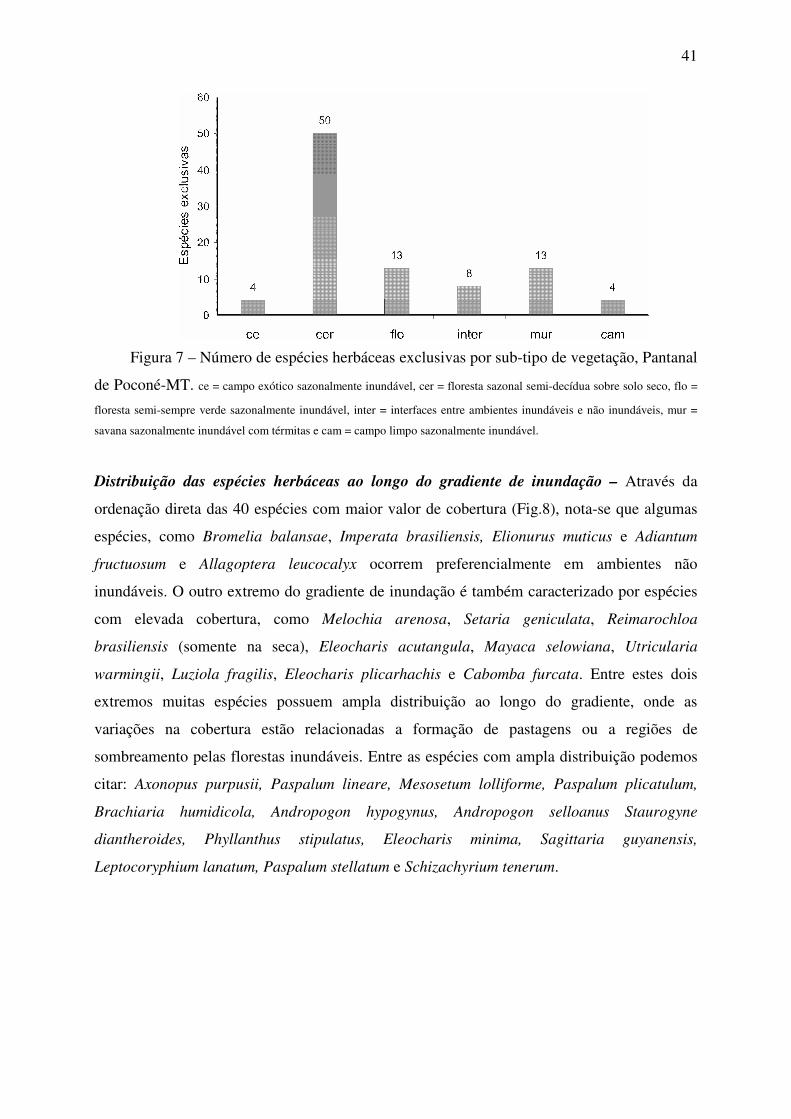
41
Figura 7 – Número de espécies herbáceas exclusivas por sub-tipo de vegetação, Pantanal
de Poconé-MT. ce = campo exótico sazonalmente inundável, cer = floresta sazonal semi-decídua sobre solo seco, flo =
floresta semi-sempre verde sazonalmente inundável, inter = interfaces entre ambientes inundáveis e não inundáveis, mur =
savana sazonalmente inundável com térmitas e cam = campo limpo sazonalmente inundável.
Distribuição das espécies herbáceas ao longo do gradiente de inundação – Através da
ordenação direta das 40 espécies com maior valor de cobertura (Fig.8), nota-se que algumas
espécies, como Bromelia balansae, Imperata brasiliensis, Elionurus muticus e Adiantum
fructuosum e Allagoptera leucocalyx ocorrem preferencialmente em ambientes não
inundáveis. O outro extremo do gradiente de inundação é também caracterizado por espécies
com elevada cobertura, como Melochia arenosa, Setaria geniculata, Reimarochloa
brasiliensis (somente na seca), Eleocharis acutangula, Mayaca selowiana, Utricularia
warmingii, Luziola fragilis, Eleocharis plicarhachis e Cabomba furcata. Entre estes dois
extremos muitas espécies possuem ampla distribuição ao longo do gradiente, onde as
variações na cobertura estão relacionadas a formação de pastagens ou a regiões de
sombreamento pelas florestas inundáveis. Entre as espécies com ampla distribuição podemos
citar: Axonopus purpusii, Paspalum lineare, Mesosetum lolliforme, Paspalum plicatulum,
Brachiaria humidicola, Andropogon hypogynus, Andropogon selloanus Staurogyne
diantheroides, Phyllanthus stipulatus, Eleocharis minima, Sagittaria guyanensis,
Leptocoryphium lanatum, Paspalum stellatum e Schizachyrium tenerum.

42
Figura 8 – Distribuição das 40 espécies de maior valor de cobertura ao longo do gradiente de profundidade da inundação, respectivamente nos períodos de seca, cheia e vazante, Pantanal de Poconé-MT.

43
Na Figura 9, entretanto, é possível observar o quão dinâmica é a variação da
cobertura das espécies em relação ao gradiente de inundação e período do ano. Espécies
como Reimarochloa brasiliensis e Setaria geniculata são preferentes de ambientes mais
profundos no período de seca, ocorrendo com baixa cobertura na cheia e vazante. Já
Melochia arenosa e Caperonia castaneifolia também ocorrem preferencialmente em
ambientes mais profundos, mas com variações na cobertura em diferentes períodos.
Cabomba furcata e Eleocharis plicarhachis, também de ambientes profundos, ocorrem
apenas durante a cheia e a vazante. Algumas espécies ocorreram preferencialmente em
ambientes entre os extremos não inundável e muito inundável, como Axonopus purpusii
e Paspalum lineare, que ocorrem nos três períodos com pouca variação na cobertura.
Eleocharis minima, também prefere ambientes entre os extremos, mas nos períodos de
cheia e vazante, enquanto Sagittaria guyanensis ocorre apenas na cheia. Um bom
exemplo de espécie com preferência de ambientes não inundáveis é Bromelia balansae
que, praticamente, não apresenta variações em cobertura. Aristida cappilaceae ocorre
preferencialmente em ambientes pouco inundáveis e nos períodos de cheia e vazante.
.

44
Figura 9 – Preferência de algumas espécies de elevada cobertura ao longo do gradiente
de inundação e dos períodos de seca, cheia e vazante, Pantanal de Poconé-MT. Números
indicam a isolinha, barras pretas indicam seca, branca a cheia e cinza a vazante. A
seqüência das isolinhas segue o gradiente sendo que a profundidade aumenta da
esquerda para a direita.

45
DISCUSSÃO
A ordenação das isolinhas situou aquelas não inundáveis em um extremo e as
muito inundáveis no outro extremo. Esse gradiente define variações na distribuição e
nas abundâncias das espécies herbáceas, que por sua vez foi influenciada pelos
diferentes sub-tipos de vegetação dominantes localmente. Apesar de muitas isolinhas de
diferentes sub-tipos de vegetação inundáveis estarem interpostas ou fazendo limites
nesse gradiente e, assim, compartilhando muitas espécies, estas se agrupam de acordo
com sua tipologia definida a priori, devido à presença de espécies características com
abundâncias semelhantes, definindo diferenciadas comunidades herbáceas. Dessa
forma, os resultados estão em acordo com a primeira predição deste trabalho, ou seja, os
sub-tipos de vegetação foram discriminados em função das comunidades herbáceas
formando um gradiente que parte de comunidades não inundáveis para as altamente
inundáveis.
Zeilhofer e Schessl (1999) encontraram poucas relações entre tipo de solos e tipo
de vegetação, como ocorre em outras planícies de inundação da Bolívia e Venezuela.
Encontraram forte relação da distribuição de diferentes tipos de vegetação,
principalmente, em relação ao regime hídrológico e textura do solo. Desta forma, uma
seqüência de ocorrência dos diferentes tipos de vegetação no gradiente crescente de
duração de inundação e drenagem do solo ocorreram da seguinte forma: os campos
limpos sazonalmente inundáveis e brejos estão relacionados a ambiente com inundações
que chegam a durar seis meses; as florestas semi-sempre verdes ocorrem em locais com
forte inundação com rápida drenagem; os campos de murundus localizam-se em
porções com inundações menos pronunciadas do que das florestas semi-sempre verdes;
e por último estão as formações arbóreas decíduas e semi-decíduas ocorrentes em cotas
altimétricas não inundáveis. Pinder e Rosso (1998) também descrevem forte influência
do gradiente hidrológico do Pantanal na formação de grupos com espécies distintas,
algumas delas características.
O gradiente limitou bem o conjunto de espécies preferenciais por ambientes não
inundáveis e altamente inundáveis, o que possivelmente ocorre devido à seleção pelo
ambiente de espécies mais adaptadas ou a não inundação ou a uma forte inundação. As
espécies dos ambiente intermediários apresentaram uma maior amplitude de
distribuição no gradiente, estando limitadas entre estes dois extremos mais seletivos, o
que está em acordo com a terceira predição deste trabalho. Assim, possivelmente os

46
mecanismos de tolerância (morfo-anatômicas e fisiológicos) de algumas espécies as
permitem ter uma amplitude de ocorrência muito maior no gradiente de inundação,
entretanto, limitadas entre dois fortes extremos de inundação.
Além de preferência em relação ao gradiente, as espécies também apresentaram
preferências entre os períodos. A distribuição e dinâmica sazonal da vegetação herbácea
sujeita ao pulso de inundação fora investigada em outros estudos realizados na região
(Prado et al. 1994, Heckman 1997; Schessl 1999; Rebellato & Nunes da Cunha 2005).
Em conjunto, esses estudos evidenciam que a diversidade dos habitats aquáticos e
terrestres em áreas úmidas é controlada e mantida pelo regime hidrológico que
influencia a distribuição de habitats e a substituição de espécies entre as diferentes fases
hídricas (Whited et al. 2007). Portanto, o pulso de inundação sazonal, estruturador de
condições físicas do ambiente, mantém elevados níveis de heterogeneidade de habitas
influenciando a distribuição da flora herbácea indicando a importância de seu papel na
manutenção da diversidade de espécies que se adapta a sua variabilidade e magnitude
(Junk 2005). A magnitude do pulso de inundação ou, o aumento do distúrbio, pôde ser
inferido em função da maior influência na substituição de espécies entre os diferentes
períodos, sendo evidenciado pelos maiores deslocamentos entre os diferentes períodos
das isolinhas situadas no extremo de maior profundidade, o que, portanto, não está em
acordo com a segunda predição deste trabalho. Ou seja, as variações de composição e
abundância das espécies herbáceas não foram mais pronunciadas nos sub-tipos
vegetacionais que experimentam níveis/tempos de inundação intermediários.
Possivelmente, se os extremos fossem uma cordilheira (não inundável) e uma lagoa
(permanentemente inundável), os ambientes sazonalmente inundáveis, situados entre
estes dois extremos, seriam aqueles que apresentariam maiores variações com relação a
composição e a abundância de espécies herbáceas, pois experimentam forte estresse
hídrico pelo excesso ou falta de água. Nos ambientes extremos as condições variariam
em termos de umidade do solo (para os ambientes não inundáveis) e em profundidade
(para os permanentemente inundáveis). Ou seja, as condições físicas seriam menos
contrastantes entre as duas fases do ano, tendo assim, menor substiuição de espécies.
Algumas formações nativas estudadas são fisionomicamente bem definidas e
bem situadas no gradiente de inundação de acordo com Zeilhofer e Schessl (1999), com
ocorrências no sentido não inundável para muito inundável na seguinte sequência:
florestas sazonais semi-decíduas sobre solo seco - savanas sazonalmente inundáveis
com térmitas - campo limpo sazonalmente inundável. Já as florestas semi-sempre verdes

47
sazonalmente inundáveis se desenvolvem sobre as savanas sazonalmente inundáveis
com térmitas ou sobre o campo limpo sazonalmente inundável. Entretanto,
apresentaram maior similaridade da composição herbácea com o campo limpo
sazonalmente inundável, o que, provavelmente, está relacionado à preferência das
espécies arbóreas que as compõem por cotas altimétricas mais baixas, que permanecem
mais tempo inundadas (Arieira & Nunes da Cunha 2006; Nunes da Cunha et al. 2007).
Dessa forma, poderia-se dizer que as espécies arbóreas invasoras possuem preferências
no gradiente de inundação semelhantes às da flora herbácea que define os campos
limpos sazonalmente inundáveis, como exemplo, as espécies Reimarochloa brasiliensis
e Setaria geniculata. As florestas semi-sempre verdes sazonalmente inundáveis são
formadas principalmente por espécies que tiveram grande expansão a partir de 1974,
ano a partir do qual o clima se tornou mais úmido com chuvas e inundações mais
pronunciadas (Nunes da Cunha & Junk 2004). Segundo Nunes da Cunha & Junk
(1999), poucas espécies arbóreas suportam elevados níveis de inundação, sendo a
maioria de ampla dispersão no gradiente de inundação. Dentre elas, Vochysia divergens
(cambará) e Licania parvifolia (pimenteira) são consideras invasoras de áreas
campestres (Pott & Pott 1994; Nunes da Cunha & Junk 2004), tendo a primeira
preferência por solos mais argilosos (Pott 1988). Esta espécie tem a capacidade de
colonizar rapidamente áreas sujeitas a inundação devido à sua elevada taxa de
crescimento sob alta luminosidade, dispersão de sementes por vento e água, elevada
taxa de recrutamento em solos descobertos e tolerância das plântulas e adultos a longos
períodos de submersão (Nunes da Cunha & Junk 2004). As densas formações
monoespecíficas desta espécie e a expansão sobre as áreas campestres têm criado
problemas aos fazendeiros locais do Pantanal de Poconé (Nunes da Cunha et al. 2007).
Pott e Pott (1999) citam Vochysia divergens e Licania parvifolia como espécies de
origem amazônica, já Calophyllum brasiliense é uma espécie com ampla dispersão,
sendo comum também à Mata Atlântica.
A savana sazonalmente inundável com térmitas (campo de murundu ou savana
parque) ocorre de forma periférica em quase todo o Pantanal de Poconé, sendo o sub-
tipo de vegetação de maior cobertura na região nordeste desta sub-região do Pantanal
(Loureiro et al. 1982). O estrato herbáceo desta fisionomia foi inventariado por Schessl
(1999), sendo identificadas 108 espécies de herbáceas. Entretanto, nenhum estudo
quantitativo foi realizado nas áreas planas deste sub-tipo de vegetação. Paspalum
lineare, uma das espécies de maior cobertura no presente estudo, não é citada por

48
Schessl (1999), sugerindo condições ambientais diferentes. A cobertura herbácea das
áreas planas deste sub-tipo de vegetação na região possui variações na composição e
estrutura em função da posição topográfica em que se encontram (Nunes da Cunha &
Junk 2006). Neste estudo é possível dizer que os campos de murundus aqui analisados
são basicamente de um único tipo, pois apresentam as mesmas espécies características
dominando, e diferentes daqueles estudados por Schessl (1999). Estes campos estão
ameaçados nesta região, principalmente, pela substituição de suas espécies pelo plantio
de Brachiaria humidicola, dada a forte dominância de gramíneas nativas extremamente
esclerófilas, de pouca aceitabilidade pelo gado da região. Outra ameaça, mas de caráter
natural e já mencionada, é a colonização destas áreas por espécies arbóreas como
Vochysia divergens e Licania parvifolia, aparentemente nas isolinhas mais inundáveis.
As florestas sazonais semi-decíduas sobre solo seco (cerradão) se desenvolvem
sobre paleo-diques muito arenosos, longos e sinuosos que alcançam de 1 a 2 metros
acima do nível da planície de inundação (Nunes da Cunha et al. 2007). A predominância
de Bromelia balansae no sub-bosque dessas formações indica condições permanentes
de não alagamento (Nunes da Cunha 1990). Neste estudo também verificou-se que
Imperata brasiliensis e Elionus muticus também ocorrem com elevada cobertura nesta
formação. Se por um lado as áreas não inundáveis apresentam espécies arbóreas mais
relacionadas ao Cerrado, sua flora herbácea também é peculiar em relação aos demais
ambientes inundáveis, com grande número de espécies exclusivas em relação aos outros
sub-tipos de vegetação inundáveis. Há um grupo característico de espécies neste sub-
tipo de vegetação, que juntamente com as espécies arbóreas, compõem uma
comunidade específica não adaptada ao pulso de inundação. Por não serem inundados,
estes ambiente provêm refúgio a muitos animais terrrestres durante o período de cheia
(Nunes da Cunha & Junk 2006).
Os campos exóticos sazonalmente inundáveis são formados pelo plantio de
espécies não nativas, principalmente, do gênero Brachiaria. A substituição da vegetação
nativa por exótica ocorre em função da crescente pressão para tornar o sistema de
produção pecuária do Pantanal mais competitivo. De modo geral, a conversão ocorre
principalmente em áreas florestadas localizadas sobre cordilheiras, livres da inundação
(Silva et al. 1999). Para todo o Pantanal do Mato Grosso, estima-se que 6% da
vegetação natural da planície pantaneira fora suprimida, principalmente pela introdução
de gramíneas exóticas, com diversos impactos sobre a fauna nativa (Harris et al. 2005).
O aumento no desmatamento de áreas não inundáveis ocupadas por florestas deciduais e

49
semi-deciduais ocorreu a partir de 1974, quando uma seqüência de anos mais úmidos,
com inundações mais pronunciadas, tornou grandes extensões de campos nativos
inundáveis menos disponíveis ao longo do ano (Silva et al. 1999). Neste estudo,
entretanto, constatou-se que os campos nativos sazonalmente inundáveis também têm
sido convertidos em pastagens exóticas, principalmente em áreas com domínio de
gramíneas mais esclerófilas e pouco palatáveis. Brachiaria humidicola, a espécie
exótica disseminada nestes ambientes inundáveis, é extremamente adaptada à variação
hídrica sazonal (Dias-Filho & Carvalho 2000), e vêm se destacando em termos de área
plantada no Pantanal como um todo (Santos et al. 2005).
Os campos limpos são ambientes onde se alternam períodos de pronunciada
inundação e seca durante o ano (Schessl 1999), onde dominam Reimarochloa
brasiliensis, Setaria geniculata, Panicum laxum, Cyperus haspam e Eleochris minima
(Rebellato e Nunes da Cunha 2005; Schessl 1999). Entre 40 de 50% das espécies
detectadas nesses campos são terófitas de lâminas curtas (Schessl 1999; Rebellato e
Nunes da Cunha 2005). As gramíneas desses campos são excelentes para a pecuária, por
serem menos esclerófilas, mais nutritivas e mais palatáveis. Neste trabalho este tipo de
vegetação ocorreu em apenas uma isolinha, onde foram encontradas centenas de
plântulas de Vochysia divergens, o que indica estágio inicial de colonização.
Assim, o pulso de inundação afeta as comunidades inundáveis e sua
variabilidade espacial determina a ocorrência de diferentes tipos de vegetação em
função da diferenciada distribuição de preferência de espécies. O conhecimento de
espécies indicadoras quanto a sua preferência ambiental pode auxiliar no manejo de
áreas úmidas, como na detecção de mudanças na estrutura e composição de espécies em
função de possíveis impactos em relação aos padrões do regime hidrológico, assim
como indicar rapidamente características hidrológicas de uma determinada área (Goslee
et al. 1997).
Um bom exemplo sobre os efeitos de mudanças hidrológicas na vegetação é
revelado por Zeilhofer e Schessl (1999) e Nunes da Cunha (com. pessoal). Estes autores
comentam que mudanças ambientais que possam aumentar o nível / duração da
inundação (aumento do distúrbio) afetam o contingente de espécies das savanas
inundáveis com térmitas que, então, é substituído por um contingente de espécies
comuns de campos limpos inundáveis. Assim, as diferentes comunidades inundáveis no
Pantanal flutuam, mas estão em equilíbrio com seu ambiente dinâmico. Entretanto, a
quebra de determinados limites ambientais dá início a uma sucessão de espécies

50
descaracterizando os ambientes ou tipos de vegetação anteriormente existentes. Desta
forma, os diferentes ambientes possuem diversos graus de resiliência, podendo absorver
certos níveis de distúrbios sem ser deslocados de seus estados de equilíbrio ou domínios
de atração (Holling 1973). Isso reforça Pinder & Rosso (1998) que sugerem que o
manejo de atividades econômicas e conservação em áreas úmidas poderiam ser
planejados de forma mais eficiente a partir de classificações de habitats que utilizam
dados fitossociológicos quantitativos.
CONCLUSÕES
• Os sub-tipos de vegetação formam comunidades diferenciadas em termos de
composição e estrutura de espécies herbáceas;
• Nos ambientes mais inundáveis a composição e estrutura destas comunidades de
herbáceas são afetadas pelo pulso de inundação, e isoladamente de forma mais
pronunciada naquelas isolinhas com maior profundidade de inundação;
• A riqueza em espécies nos diferentes sub-tipos de vegetação se eleva em função
da sazonalidade hídrica, sendo maior nos períodos de cheia e vazante;
• As diferentes espécies herbáceas mostram preferências em relação ao gradiente
de profundidade, assim como em relação aos aos diferentes períodos;
REFERÊNCIAS CITADAS
Adámoli, J. 1987. Fisiografia do Pantanal. Pp. 15-22. In: Recursos forrageiros nativos
do Pantanal Mato-Grossense. Empresa Brasileira de Pesquisa Agropecuária/Cernagem,
Brasília. Documento 8. 339p.
Amaral Filho, Z. P. 1986. Solos do Pantanal Mato-Grossense. Pp 91-104. In: Simpósio
sobre recursos naturais e sócio-econômico do pantanal. Empresa Brasileira de Pesquisa
Agropecuária (Embrapa-CPAP), Brasília. 265 p.
Alvarenga, S. M. M., Brasil, A. E., Pinheiro, R. e Kux, H. J. H. 1984. Pp 90-183.
Estudo geomorfológico aplicado a Bacia do Alto Paraguai e Pantanais Mato-grossenses.
Boletim Técnico do Projeto RADAMBRASIL (Série Geomorfologia,1). 187p.

51
Arieira, J. & C. Nunes da Cunha. 2006. Fitossociologia de uma floresta inundável
monodominante de Vochysia divergens Pohl. (Vochysiaceae), no Pantanal Norte, MT,
Brasil. Acta Botânica Brasileira 20 (3): 569-580.
Assine, M. L. and Soares, P. C. 2004. Quaternary of the Pantanal, west-central Brazil.
Quaternary International 114(1): 23-34.
Barret, R., Nielssen, D. L. and Croome, R. 2009. Associations between the plant
communities of floodplain wetlands, water regime and wetland type river research and
applications. Published online in Wiley InterScience (www.interscience.wiley.com).
Bullock, J. 1996. Plants. In: Ecological census techniques. W.J. Sutherland (ed). Pp.
111-138. Cambridge University Press, Cambridge. 336p.
Capon, S. J. 2003. Plant community responses to wetting and drying in a large arid
floodplain. River research and applications 19:509-520.
Casanova, M. T. and Brock, M. A. 2000. How do depth, duration and frequency of
flooding influence the establishment of wetland plant communities? Plant Ecology 147
(2):237 250.
Clarke, K.R. 1993. Non-parametric multivariate analysis of changes in community
structure. Australian Journal of Ecology 18:117-143.
Cox, T.F. & Cox, M.A.A. 2001. Multidimensional Scaling. Chapman & Hall, Boca
Raton.
Dias, L. R. 2006. Comunidata. Versão 1.6. Manaus.
Dias-Filho, M. B. & Carvalho, C. J. R. 2000. Physiological and morphological
responses of Brachiaria spp. to flooding. Pesquisa Agropecuária Brasileira 35:.1959-
1966.

52
Fantim–Cruz, I. 2008. Dinâmica da Inundação em Meso-Escala na Planície do Rio
Cuiabá, Pantanal, Brasil. Dissertação de Mestrado apresentada ao Programa de Pós-
graduação em Ecologia e Conservação da Biodiversidade - Universidade Federal de
Mato Grosso - Instituto de ciências biológicas, Cuiabá.
Fortney, R. F., Benedict, M., Gottgens, J. F., Walters, T. L., Leady, B. S. and Rentch, J.
2004. Aquatic plants community composition and distribution along an inundation
gradient at two ecologically-distinct sites in the Pantanal region of Brasil. Wetlands
Ecology and Management 12:575-585.
Franco, M. S. M. e Pinheiro, R. 1982. Geomorfologia. Pp.161-224. In: Brasil.
Ministério das Minas e Energia. Departamento Nacional da Produção Mineral. Projeto
Radam-brasil. Folha SE.21 Corumbá. Rio de Janeiro.
Goslee, S. C., Brooks, R. P. and Cole, C. A. 1997. Plants as indicators of wetland water
source. Plant Ecology 131: 199–206.
Hammer, Ø.; Harper, D. A. T. & Ryan, P. D. 2001. Past Palaeontological Statistics, ver.
1.79. Disponível em: <http://folk.uio.no/ohammer/past>.
Hamilton, S. K., Sippel, S. J., Melack, J. M. 1996. Inundation patterns in the Pantanal
wetland of South America determined form passive microwave remote sensing.
Archives of Hydrobiology 137:1-23.
Hamilton, S. K., Sippel, S. J. and Melack, J. M. 2002. Comparison of inundation
patterns among major South American floodplains. Journal of Geophysical Research
107.
Harris, M. B.; Arcangelo, C.; Pinto, E.C.T.; Camargo, G.; Ramos Neto, M.B.; Silva,
S.M. 2005. Estimativas de perda da área natural da Bacia do Alto Paraguai e Pantanal
Brasileiro. Relatório Técnico não publicado. Consevação Internacional, Campo Grande,
MS.
Heckman, C. W. 1997. Description of a dinamic ecotone in the Pantanal of Mato
Grosso, Brazil. Ecotropica 3:1-7.

53
Holling, C. S. 1973. Resilience and stability of ecological systems. Annual Review of
Ecology and Systematics 4: 1–23.
Junk, W.J. 2005. Flood pulsing and linkages between terrestrial, aquatic and wetland
system. Verh. Internat. Verein. Limnol 29: 11-38.
Junk, W. J., Bayley, P. B. & Sparks, R. S. 1989. The flood pulse concept in river –
floodplain systems. In: Proc Int Large River Symp (LARS). D. P. Dodge (ed). Canadian
Journal of Fishers and Aquatic 106: 110-127.
Junk, W.J., Brown, M., Campbell, I. C., Finlayson, M, Gopal, B., Ramberg, L. and
Warner, B. G. 2006. The comparative biodiversity of seven globally important
wetlands: a synthesis. Aquatic Science 68: 400–414.
Köppen, G. W. 1948. Climatologia. Fundo de Cultura Econômica. México. Pp. 242-
248.
Kotze, D. C. and O’Connor, T. G. 2000. Vegetation variation within and among
palustrine wetlands along an altitudinal gradient in KwaZulu-Natal, South Africa. Plant
Ecology 146(1):77-96.
Lenssen, J., Menting, F., van der Putten, W. and Blom , K. 1999. Control of plant
species richness and zonation of functional groups along a freshwater flooding gradient.
Oikos 86(3):523-534.
Loureiro, R.L., Souza Lima, J.P., Fronzar, P.C. Vegetação. Pp. 329-372. In: Brasil:
Ministério das Minas e Energia. Departamento Nacional da Produção Mineral. Projeto
Radam-brasil. Folha SE.21 Corumbá. Rio de Janeiro. (Levantamento de Recursos
Naturais, 27).
Magnusson, W. E., Lima, A. P., Luizão, R., Luizão,F., Costa, F. R. C., Castilho, C. V
and Kinupp, V. F. 2005. Rapeld: A modification of the Gentry method for biodiversity
surveys in long-term ecological research sites. Biota Neotropica 5(2): 1-6.

54
McCune, B. & Mefford, M.J. 1999. PC-ORD. Multivariate analysis of ecological data,
version 4.0. Oregon Glaneden Beach, MjM Software Desing.
McCune, B. and J. B. Grace. 2002. Analysis of Ecological Communities . MjM
Software, Gleneden Beach , Oregon . 304 pages.
Nascimento, M.T. & Nunes da Cunha, C. 1989. Estrutura e composição florística de um
cambarazal no Pantanal de Poconé - MT. Acta Botanica Brasilica 3(1): 3-11.
Nunes da Cunha, C. 1990. Estudo florístico e fitofisionômico das principais formações
do Pantanal de Poconé-Mato Grosso. Dissertação de mestrado apresentada ao Instituto
de Biologia da Universidade Estadual de Campinas (Unicamp). 133p
Nunes da Cunha, C. 1998. Comunidades arbustivo-arbóreas de Capões e Diques
marginais no Pantanal de Poconé-MT: Caracterização e análise de gradiente. Tese
apresentada ao Programa de Pós-graduação em Ecologia e Recursos Naturais,
Universidade de São Carlos (UFSCAR).
Nunes da Cunha, C. & Junk, W.J. 1999. Composição florística de capões e cordilheiras:
localização das espécies lenhosas quanto ao gradiente de inundação no Pantanal de
Poconé, MT-Brasil. Pp.387-405. In: Anais do II Simpósio sobre Recursos Naturais e
Sócio-econômicos do Pantanal, Manejo e Conservação. Empresa Brasileira de Pesquisa
Agropecuária (Embrapa-CPAP), Corumbá. 537p.
Nunes da Cunha, C. & Junk, W.J. 2001. Distribuition of wood plant communities along
the flood gradient in the Pantanal of Poconé, Mato Grosso, Brazil. International Journal
of Ecology and Environmental 27:63-70.
Nunes da Cunha, C. & Junk, W.J. 2004. Year-to-year changes in water level drive the
invasion of Vochysia divergens in Pantanal grasslands. Applied Vegetation Science
7:103-110.

55
Nunes da Cunha, C., Rawiel, C. P., Wantzen, K. M., Junk, W. J. & Lemes do Prado, A.
2006. Mapping and characterization of vegetation units by means of Landsat imagery
and management recommendations for the Pantanal of Mato Grosso (Brazil), north of
Poconé. Amazoniana XIX(1): 1-32.
Nunes da Cunha C., Junk W.J., Leitão Filho, H. F. 2007. Floristic and physiognomic
types of arboreal vegetation of the Pantanal of Pocone, Mato Grosso. Amazoniana
XIX(3/4): 159-184.
Nunes da Cunha, C. & Junk, W.J. 2009. Landscape units of the Pantanal: structure,
function, and human use. Pp. 127-141. In: The Pantanal: Ecology, biodiversity and
sustainable management of a large neotropical seasonal wetland. Junk, W.J., Da Silva,
C.J., Nunes da Cunha, C., Wantzen, K.M. (eds). Pensoft Publishers, Sofi a–Moscow.
Oliveira-Filho, A. T. 1992. Floodplain ‘murundus’ of Central Brazil: evidence for the
termite-origin hypothesis. Journal of Tropical Ecology 8 (1): 1-19.
Pinder, L. and Rosso, S. 1998. Classification and ordination of plant formations in the
Pantanal of Brazil. Plant Ecology 136: 151–165.
Pinho, J. B. 2005. Comunidades de aves, biologia reprodutiva e dieta alimentar de aves
de sub-bosque em quatro áreas fitofisionomias diferentes no Pantanal de Mato Grosso.
Tese de doutorado apresentada ao Programa de Pós-graduação em Ecologia,
Conservação e Manejo da Vida Silvestre. Universidade Federal de Minas Gerais
(UFMG), Instituto de biociência, Belo Horizonte.
Ponce, V. M. & Cunha, C. N. 1993. Vegetated earthmounds in tropical savannas of
Central Brazil: a synthesis. Journal of Biogeography 20: 219-225.
Pott, A. 1988. Pastagens no Pantanal. Empresa Brasileira de Pesquisa Agropecuária
(Embrapa-CPAP), Corumbá. Documentos 7. 58p.

56
Pott, A. 2000. Dinâmica da vegetação do Pantanal. In: Cavalcanti, T.C., Walter, B.M.T.
(org.) Tópicos atuais em Botânica. Brasília: Embrapa Recursos Genéticos e
Biotecnologia/Sociedade Botânica do Brasil. p. 172-182.
Pott, A. & Pott, V.J. 1994. Plantas do Pantanal. Empresa Brasileira de Pesquisa
Agropecuária (Embrapa-CPAP), Corumbá. 320 p.
Pott, A. & Pott, V. J. 1999. Flora do Pantanal - Listagem Atual de Fanerógamas. Pp.
297-325. In: Simpósio sobre Recursos Naturais e Sócio-Econômicos do Pantanal:
Manejo e Conservação. Empresa Brasileira de Pesquisa Agropecuária (Embrapa-CPAP),
Corumbá. 537p.
Pozer, C.G. & Nogueira, F. 2004. Pastagens nativas inundáveis da região norte do
Pantanal de Mato Grosso: variações de biomassa e produtividade primária. Brazilian
Journal of Biology 64(4): 859-866.
Prado, A. L., Heckman, C. W., E Martins, F. R. 1994. The seasonal succession of biotic
communities in wetlands of the tropical wet-and-dry climatic zone: II. The Aquatic
Macrophyte Vegetation in the Pantanal of Mato Grosso, Brazil. Internationale Revue
gesamten Hydrobiologie 79(4): 569-589.
Prance, G. T. and Schaller, G. B. 1982. Preliminary study of some vegetation types of
the Pantanal, Mato Grosso, Brazil. Brittonia 34: 228-251.
Rebellato, L. e Nunes da Cunha, C. 2005. Efeito do “fluxo sazonal mínimo da
inundação” sobre a composição e estrutura de um campo inundável no Pantanal de
Poconé, MT, Brasil. Acta Botanica Brasilica 19: 789-799.
Santos, S. A., Crispim, S. M. A., Comastri-Filho, J. A., Pott, A. e Cardoso, E. L. 2005.
Substituição de pastagens nativas de baixo valor nutritivo por forrageiras de melhor
qualidade no Pantanal. Empresa Brasileira de Pesquisa Agropecuária (Embrapa-CPAP),
Corumbá. Circular técnica, 62, 5p.

57
Schessl, M. 1999. Floristic Composition and Structure of Floodplain Vegetation in
Northern Pantanal of Mato Grosso, Brasil. Phyton (Horn, Austria) 39(2):303-3.
Silva, J. S. V. e Abdon, M. M. 1998. Delimitação do Pantanal brasileiro e suas sub-
regiões. Pesquisa agropecuária brasileira, Brasília-DF 33: 1703-1711.
Silva, M. P., Mauro, R., Mourão, G. and Coutinho, M. 1999. Conversion of forests and
woodlands to cultivated pastures in the wetland of Brazil. Ecotropicos 12(2):101-108.
Sluis, W and Tandarich, J. 2004. Siltation and hydrologic regime determine species
composition in herbaceous floodplain communities Plant Ecology 173: 115–124.
Southal, E. J., Dale, M. P. and Kent, M. 2003. Spatial and temporal analysis of
vegetation mosaics for conservation: poor fen communities in a Cornish valley mire.
Journal of Biogeography 30:1427–1443.
Townsend, P. A. 2001. Relationships between vegetation patterns and hydroperiod on
the roanoke river floodplain, North Carolina. Plant Ecology 156(1): 43-58.
Whited, D. C., Lorang, M. S., Harner, M. J. Hauer, F. R., Kimball, J. S. and Stanford, J.
A. 2007. Climate, hydrologic disturbance, and succession: drivers of floodplain pattern.
Ecology 88(4): 940-953
Zeilhofer, P. & Schessl, M. 1999. Relationship between vegetation and environmental
conditions in the northern Pantanal of Mato Grosso, Brazil. Journal of Biogeography
27: 59-168.

58
Capítulo 2 – Efeito da sazonalidade e gradiente de inundação na riqueza, composição, cobertura e formas de vida de herbáceas do
Pantanal de Poconé-MT RESUMO - A variabilidade espaço-temporal influencia a distribuição, a riqueza e a
substituição de espécies em áreas úmidas. Sendo o Pantanal uma planície sujeita a
inundação anual previsível e monomodal com elevada variabilidade hidrotopográfica,
objetivamos neste trabalho verificar qual efeito regional da inundação sazonal e do
gradiente de inundação, em diferentes períodos, sobre a riqueza, diversidade,
composição, cobertura e formas de vida das espécies herbáceas.Este trabalho foi
realizado no Pantanal de Poconé (MT) utilizando-se o sistema RAPELD e metodologia
da parcela pontual para amostragem de herbáceas em 30 isolinhas de 250 m, realizadas
nos períodos de seca, cheia e vazante. As contagens das espécies nas parcelas pontuais
permitiram estimar a riqueza, a diversidade e a cobertura das espécies. A inundação
influenciou a composição, como também, a riqueza e diversidade (H’),
significativamente mais elevadas durante os períodos de cheia e vazante. Poucas
espécies foram abundantes, entre elas A. purpusii, B. humidicola e P. lineare que
mantiveram elevada cobertura regional entre os períodos. Entretanto, a inundação
influencia a cobertura de muitas espécies, tendo Eleocharis minima e Utricularia
foliosa destaque durante a cheia. As hemicriptófitas são dominantes e mais estáveis em
todos os períodos, e a participação de terófitas e geófitas esteve vinculada aos períodos
de cheia e vazante. O gradiente de inundação tem efeitos negativos sobre a riqueza em
espécies e diversidade beta, onde a substituição de espécies é elevado na transição entre
os ambientes não inundáveis e pouco inundáveis, tendendo a diminuir durante os
períodos de cheia e vazante em função da queda na riqueza e aumento da similaridade.
O gradiente de inundação também influencia as formas de vida, com quedas
significativas na riqueza em hemicriptófitas em todos os períodos, e em geófitas durante
a seca. Tende a influenciar também o aumento de cobertura para geófitas e terófitas
durante a cheia. As variações nas coberturas das espécies entre os períodos de seca,
cheia e vazante, também tendem a ser maiores ao longo deste gradiente.
Desta forma, concluiu-se que as espécies possuem diferentes plasticidades ou estratégias para que possibilitam sua manutenção neste ambiente dinâmico, tendo o ciclo hídrico importância na determinação da riqueza e diversidade. Assim, a manutenção do regime hidrológico nessas áreas úmidas e a variabilidade espacial na paisagem são fundamentais para a conservação de suas biodiversidades. Palavras-chaves: Pantanal, herbáceas, inundação sazonal, gradiente de inundação, riqueza,
formas de vida

59
Chapter 2 – The effect of seasonality and flood gradient in the
structure and composition of herbaceous plants in the Pantanal of
Poconé, Mato Grosso.
ABSTRACT – The space-time variables influence the distribution, richness and
turnover of species in wetlands. Since the Pantanal is a plain which is subject to
predictable and mono-modal annual flooding with a high degree of hydro-topographic
variability, the objectives set for this study were to verify the effect of seasonal flooding
on a regional scale and the flood gradient in different periods on the flora, richness,
diversity, coverage and life forms. The study was carried out in the Pantanal of Poconé
(Mato Grosso) using the RAPELD system and the point sampling method for
herbaceous species along 30 isolines 250m long, during the dry, flood and drain
periods. The species count at the sample points allowed for estimates to be made of
species richness, diversity and coverage. Flooding influenced the richness and diversity
(H’) which were significantly higher during the flood and drain periods. Few species
were abundant, among which were A. purpusii, B. humidicola and P. lineare and these
maintained a high level of regional coverage over all the periods. However, flooding
influenced coverage for many species especially for Eleocharis minima and Utricularia
foliosa during the flood period. The hemicryptophytes are dominant and more stable in
the flood and drain periods. The flood gradient has negative effects on species richness
and beta diversity where species turnover is high in the transition areas between non-
flooded and slightly flooded environments with a tendency to diminish during the flood
and drain periods due to the fall in richness and an increase in similarity. The flood
gradient also influences life forms with significant drops in the richness of
hemicryptophytes in all periods and of geophytes during the dry period. It also tends to
influence the increase of coverage by geophytes and therophytes during the flood
period. It was concluded, therefore, that the species present posses plasticity or
strategies to maintain themselves in this dynamic environment and the water cycle is
important for richness and diversity reinforcing the questions of maintaining the water
regime in wetlands and its spatial variability within the landscape and conservation of
its diversity.
Key words: Pantanal, herbaceous plants, seasonal flooding, flood gradient, richness, life forms

60
INTRODUÇÃO
Em áreas úmidas, o regime hidrológico, com variações na duração, amplitude e
freqüência de inundações, é determinante na distribuição e riqueza em espécies de
plantas (Robertson et al. 1984; Pollock et al. 1998; Lenssen et al. 1999; Casanova &
Broke 2000; Townsend 2001; Capon 2003; Southal et al. 2003). Em razão de variações
das condições hidrológicas, as espécies se agrupam de acordo com suas habilidades em
tolerar ou responder à dinâmica de cheias e secas (Casanova & Broke 2000). Em
relação à diversidade, alguns trabalhos relatam queda gradativa com o aumento do
estresse causado pela duração e/ou profundidade das inundações (Robertson et al. 1984;
Lenssen et al. 1999; Casanova & Broke 2000; Capon 2003), ou mesmo, aumento da
diversidade em níveis intermediários deste distúrbio (Pollock et al. 1998). Segundo a
hipótese do distúrbio intermediário (Connell 1978) em locais mais estáveis, as espécies
competitivamente mais aptas eliminam aquelas com inferior capacidade de competição,
reduzindo a riqueza. Em níveis intermediários de distúrbio um balanço é alcançado
entre as espécies adaptadas a maior estabilidade e aquelas a maiores níveis de distúrbio,
onde, portanto, a riqueza deverá ser máxima. Em ambientes com elevado nível de
distúrbio, apenas poucas espécies podem resistir e persistir, e a riqueza novamente
decai.
Adaptações de plantas ao regime hidrológico geram não apenas variações
espaciais na riqueza, composição e cobertura de espécies, mas mudanças temporais e
previsíveis nestes atributos de espécies perenes e anuais (van der Valk 1981). A forma
de vida pode ser um atributo que ajuda a caracterizar as respostas das comunidades
vegetais em função de distúrbios ambientais (McIntyre et al. 1995).
O Pantanal, por ser uma planície sedimentar inundável, possui deposições
aluviais diferenciadas de sedimentos sobre o terreno, produzindo feições
geomorfológicas que formam gradientes hidrotopográficos. Sobre este gradiente, várias
comunidades vegetais tolerantes a diferentes níveis de estresse causados pelas cheias e
secas anuais, tais como campos inundáveis, florestas monodominantes, cerradão e
florestas aluviais formam um mosaico de unidades de paisagens (Nunes da Cunha et al.
2006). Nestas diferentes fisionomias vegetais, as proporções de espécies lenhosas e
herbáceas refletem a ocorrência e intensidade da inundação. Isto é evidenciado, por
exemplo, na transição entre cerradão, que ocupa áreas mais elevadas do gradiente
hidrotopográfico, cerrado sensu stricto situado em posições intermediárias, e às

61
formações campestres, encontradas áreas nas partes mais baixas e inundadas do
gradiente (Silva et at. 2000). Segundo Silva et al. (2000), o Cerrado é a formação
vegetacional mais representativa do Pantanal (36%), sendo o cerradão representado por
22% e o cerrado sensu stricto por 14%.
Portanto, em termos espaciais, há diferenças na composição de espécies e nas
proporções entre lenhosas e herbáceas das diferentes fitofisionomias, dispostas no
gradiente hidrotopográfico (Nunes da Cunha & Junk 1999; Zeilhofer & Schessl 1999;
Silva et at. 2000; Nunes da Cunha & Junk 2001;). Em termos temporais, a flora
herbácea responde de forma dinâmica à variação cíclica de seca e cheia ao longo do
ano. Durante os vários meses da fase seca, a flora herbácea aquática é submetida a um
forte estresse pela falta de água. Já durante a fase aquática, a flora herbácea terrestre
sofre forte estresse pelo excesso de água (Junk & Da Silva 1999). Entretanto, algumas
espécies são comuns às duas fases (Schessl 1999; Rebellato & Nunes da Cunha 2005).
No Pantanal de Poconé, estudos verificaram zonação de espécies herbáceas ao longo do
gradiente de inundação (Prado et al. 1994; Heckman 1997; Fortney et al. 2004) bem
como, mudanças sazonais nas comunidades vegetais sujeitas ao pulso de inundação
(Prado et al. 1994, Heckman 1997; Schessl 1999; Rebellato & Nunes da Cunha 2005).
O pulso de inundação é considerado a força que dirige o ecossistema pantaneiro (Junk et
al. 1989), sendo monomodal e previsível (Junk 2005), promovendo mudanças temporais
na estrutura e na composição das comunidades vegetais estabelecidas no gradiente
hidrotopográfico.
O Pantanal está entre os ecossistemas mais ricos em espécies de plantas
herbáceas, quando comparado com outras seis áreas úmidas de importância global,
equiparando-se a área úmida do Delta do Okavango na Botswana (Junk et al. 2006a). A
maior parte das espécies tem ampla dispersão, com muitas gramíneas, cyperáceas e
outras herbáceas neotropicais, seguidas por um maior contingente da flora do Cerrado
(Pott & Pott 1999; Pott & Pott 2009). Já comparado com outros biomas brasileiros, o
Pantanal é considerado pobre em espécies vegetais (Pott & Pott 2009). Vários fatores
explicam a diversidade florística atual do Pantanal. A fronteira com o Chaco, o Cerrado
e a Amazônia explicam à origem da flora (Prance & Shaller 1982; Loureiro et al. 1982;
Nunes da Cunha 1998; Prado et al. 1994; Schessl 1999). A menor riqueza em espécies
comparada a flora de origem e a falta de endemismos podem ser atribuídas à falta de
isolamento geográfico e origem geologicamente recente deste ecossistema. Somado a
isto, o clima progressivamente mais úmido na transição entre o Pleistoceno e o

62
Holoceno, associado às pronunciadas oscilações climáticas deste período, promoveram
extinções maciças e atuaram como um filtro para o estabelecimento de espécies (veja
Browm 1986; Junk et al. 2006a; Junk et al. 2006b).
Atualmente a flora herbácea do Pantanal é estimada em 1147 espécies, entre
ervas, graminóides (poáceas e cyperáceas), trepadeiras e parasitas. Destas, 900 espécies
são consideradas terrestres e 247 macrófitas aquáticas (Pott & Pott 1999; Junk et al.
2006b). O grande número de espécies herbáceas na região se deve à predominância de
áreas inundáveis, ocupadas por formações aquáticas, campestres e savânicas (Pott &
Pott 1999). Resistência de sementes e esporos à inundação e dessecação, ciclos
reprodutivos curtos, altas taxas reprodutivas e elevada produtividade são alguns dos
atributos das herbáceas que se desenvolvem em áreas sujeitas ao pulso de inundação
(Junk e Piedade 1993).
Para o aumento no conhecimento sobre a flora herbácea e efeitos do pulso de
inundação e gradiente hidrotopográfico sobre a riqueza, composição, cobertura e formas
de vida do Pantanal de Poconé, este trabalho teve como objetivos:
1) verificar qual o efeito regional da inundação sazonal sobre a riqueza,
diversidade, composição de espécies, cobertura de espécies e formas de vida;
2) verificar qual o efeito do gradiente de inundação em diferentes períodos sobre
a riqueza, diversidade, composição de espécies, cobertura de espécies e formas de vida;
MATERIAL E MÉTODOS
Características gerais e área de estudo – O relevo da Bacia do Alto Paraguai possui
cotas altimétricas que oscilam entre 80 a 1200 metros, distribuídas por três regiões
fisiográficas: dos planaltos (250 a 1200 metros), das depressões (150 a 250 metros) e
dos pantanais (80 a 150 metros) (Alvarenga et al. 1984). Hidrologicamente é
representada, principalmente, pelos rios Paraguai, Branco, Nabilieque, Miranda,
Aquidauana, Negro, Taquari, Cuiabá, Itiquira, Bento Gomes, São Lourenço e afluentes
(Amaral Filho 1986).
Geomorfologicamente, a região dos pantanais é representada pelas unidades
planícies e pantanais mato-grossenses, onde ocorrem os maiores fenômenos das

63
enchentes de toda a Bacia do Alto Paraguai (Alvarenga et al. 1984). Essas planícies
deprimidas e inundáveis recebem o nome de Pantanal e se localizam na parte central da
América do Sul, entre longitudes de 16º e 22º e latitudes de 55º e 58º. A declividade é
inexpressiva: o gradiente topográfico varia de 0,3 a 0,5 cm/km no sentido leste-oeste e
0,03 a 0,15 cm/km no sentido norte-sul. Esta região tem grande expressão espacial com
configuração de um enorme anfiteatro voltado para o oeste (Franco & Pinheiro 1982).
A tendência para inundações periódicas não está ligada apenas à pluviosidade,
que é considerada baixa, mas sim à topografia plana, cotas altimétricas baixas e
predominância de solos hidromórficos que influenciam na drenagem (Amaral Filho
1986). Entretanto, a heterogeneidade topográfica da região determina áreas em que a
inundação ocorre de forma distinta no tocante a altura, época e duração das inundações
(Adámoli 1987). Outra característica é que esta vasta superfície rebaixada é recoberta
por sedimentos quaternários, oriundos de diferentes rios que formam a bacia do rio
Paraguai (Adámoli 1987), sendo sua litologia constituída por sedimentos aluviais da
formação Pantanal, que associados à dinâmica do regime de alagamento, provocam
grande variação nos solos. Ao norte predominam os solos com horizonte subsuperficial
de textura mais argilosa, na parte central, solos mais arenosos, e ao sul, solos argilosos
(Amaral Filho 1986). A heterogeneidade promovida pelos diferentes tipos de solos e
regime hídrico, que determinam diferenças fitogeográficas, levaram à divisão do
Pantanal em sub-regiões. Silva & Abdon (1998), por exemplo, delimitam o Pantanal em
138,183 km2 e o dividem em 11 sub-regiões, sendo a sub-região de Poconé (Pantanal
Norte), onde o presente estudo foi realizado, caracterizada pelo clima do tipo AW-
quente e úmido (Köppen 1948), com precipitação média de 1250 mm ao ano e
temperatura média anual de 25,8 oC. No Pantanal Norte duas estações são observadas
durante o ano, sendo uma seca no período de maio a setembro e a outra chuvosa, de
outubro a abril (Nunes da Cunha & Junk 2004). Prado et al. (1994) delimitaram quatro
fases hidrológicas do ano no Pantanal Norte: enchente (de outubro a dezembro), cheia
(de janeiro a março), vazante (de abril a junho) e seca (de julho a setembro). No
Pantanal Norte a inundação é do tipo monomodal ocorrendo juntamente com a estação
chuvosa (outubro a abril) (Hamilton et al. 1996). Os solos são argilosos em grande
parte, com grau de fertilidade superior à dos outros pantanais (Adámoli 1987). Possui
aportes aluviais originários quase que totalmente do rio Cuiabá com transporte e
deposição de argila, silte e areia derivados principalmente de filitos e xistos. Apresenta
grande variedade de tipos de solos, com imensas áreas de solo hidromórfico cinzento

64
eutrófico, vertissolos, laterita hidromórfica distrófica e planossolo distrófico (Amaral
1987). Ao norte, apresenta extensa faixa com fraca inundação, onde as cotas
altimétricas excedem 130 metros. Em direção ao sul as áreas são bastante inundadas,
pois as cotas altimétricas decrescem para 110-100 metros, (Franco e Pinheiro 1982). Na
presente área de estudo altimetria ficou entre 115 e 112 metros.
Para a parte norte do Pantanal de Poconé a savana (cerrado) é a classe
fitofisionômica que cobre a maior parte da região (41,0%), seguida pelos sistemas de
transição (ex. florestas semi-sempre verdes sazonalmente inundáveis, com 14.7%),
florestas sazonais semideciduais (6.2%), sistemas secundários (3. 3%), corpos d’água
(0.8%) e florestas sazonais decíduas (0.4%) (Nunes da Cunha et al. 2006).
A área em que foram realizadas as coletas engloba três fazendas: Nossa Senhora
da Aparecida, São Carlos e Canoas, próximas à localidade do Pirizal, município de
Nossa Senhora do Livramento (Oeste: 56º 21', Leste: 56º 18', Norte: 16º 19', Sul: 16º
22') (Fig. 1).

65
Figura 1- Área de estudo, fazendas Nossa Senhora Aparecida, São Carlos e Canoas,
próximas à localidade do Pirizal, município de Nossa Senhora do Livramento, sub-
região do Pantanal de Poconé-MT (Pinho 2005).
Desenho amostral – Este trabalho foi realizado utilizando-se do método RAPELD.
Este é uma modificação do método de Gentry (1982), desenvolvido para inventários
rápidos da biodiversidade em sítios de pesquisa ecológica de longa duração na
Amazônia (Magnusson et al. 2005), implementado no Pantanal de Poconé. Na área de
estudo foram instalados 30 isolinhas de 250 metros sistematicamente distribuídas em
um sistema de seis trilhas de 5 km cada orientadas no sentido norte-sul, ocupando uma
área de 25 Km2. Cada trilha possui uma identificação (A, B, C, D, E, F) onde foram

66
instaladas 5 isolinhas (Fig. 2), orientadas no sentido leste-oeste. A primeira isolinha de
cada uma das trilhas inicia-se a partir de 500 metros do início das mesmas. Assim, por
exemplo, a isolinha A1 está situada a 500 metros do início da trilha A. Cada isolinha
dista 1 Km uma da outra e foram implantadas em diferentes cotas altimétricas. A
marcação das mesmas foi feita com piquetes de ferro fixos no solo a cada 10 m, sendo a
altimetria do ponto inicial mantida ao longo dos 250 metros.
ABCDEF
500
1000
1500
2000
3000
2500
3500
4000
4500
5000
Figura 2- Imagem de satélite mostrando a área de estudo (5 km x 5 km) com sistemas de
6 trilhas (apenas Norte-sul) e esquema com distribuição das isolinhas para cada trilha,
Pantanal de Poconé-MT.
Amostragem de herbáceas – As coletas foram realizadas durante três fases
hidrológicas do Pantanal: seca (setembro/outubro de 2006), cheia (fevereiro/março de
2007) e vazante (maio de 2007). A amostragem das herbáceas foi realizada pelo método
da parcela pontual (Bullock 1996). Ao longo dos 250 metros de cada isolinha foram
distribuídas parcelas pontuais distantes 1 metro uma da outra. A parcela pontual é
considerada uma haste de metal fina posicionada verticalmente, onde qualquer
morfoespécie de herbácea que a toca é registrada. Um exemplar de cada morfo espécie
foi coletado, prensado e seco em estufa, sendo posteriormente enviado a especialistas
para identificação. Futuramente as exsicatas serão depositadas no Herbário Central da
Universidade Federal de Mato Grosso (UFMT) e Universidade Federal de Minas Gerais
(UFMG). As formas de vida das espécies identificadas foram determinadas segundo
1 km 5 km
Norte
Sul
Leste Oeste
1 km
F E D C B A
5 km

67
Raunkier (1934), seguindo as classificações feitas nas listagens florísticas de Schessl
(1999), Pott e Adámoli (1999) e Pott (comunicação pessoal).
Análises dos dados
As contagens das espécies nas parcelas pontuais permitiram estimar a riqueza, a
diversidade e a cobertura das espécies nas 30 isolinhas amostradas. Os dados de
profundidade e duração da inundação foram coletados quinzenalmente entre os anos de
2006 e 2007 por Fantin-Cruz (2008). As medições foram realizadas nos marcos iniciais
de cada isolinha (mourão de madeira) que continham réguas hidrológias. O tempo de
inundação variou de 0 a 200 dias (média = 90,3 dias, dp = 50,3 dias; mediana = 90 dias)
e a profundidade da inundação variou entre 0 e 63 cm (média = 25,067 cm, dp = 16,144;
mediana = 24 cm). O tempo de inundação é fortemente correlacionado com a
profundidade da inundação (r2 = 0,913, p < 0.001, n = 30).
Análises para os efeitos regionais da inundação sazonal:
Para avaliar o efeito sazonal da inundação na cobertura das espécies de forma
regional, calculou-se o valor da cobertura regional das espécies da seguinte forma:
soma do número de parcelas pontuais nos quais a espécie ocorreu / total de parcelas
pontuais (= total de ocorrências / 30 x 250).
Curvas de distribuição de abundâncias das espécies, ranqueadas da mais
abundante para a menos abundante - uma representação visual da diversidade - foram
comparadas entre os três períodos. Para isso, foram utilizados os valores de cobertura
regional de cada espécie em cada período. A partir dos valores de cobertura regional de
cada espécie calculou-se o índice de diversidade de Shannon-Wienner (H’) para os três
períodos, aqui denominada diversidade regional. A magnitude da diversidade está
relacionada com a riqueza em espécies e a abundância relativa de cada espécie na
amostra (Magurran 1988).
O número de parcelas pontuais em que cada espécie ocorreu em cada isolinha foi
utilizado para o cálculo de sua cobertura na isolinha (número de parcelas pontuais na
qual a espécie ocorreu na isolinha / 250). Estes foram utilizados para o cálculo da
diversidade alfa, ou diversidade local das 30 isolinhas, utilizando o índice de Shannon-
Wienner (H’), e foram realizados para os três períodos amostrados. Os índices foram
calculados através do Pacote estatístico MVSP 3.13 (MultiVariate Statistical Package)
(Kovach 2007). Para testarmos a hipótese de que a sazonalidade anual da inundação

68
influencia a riqueza e a diversidade em espécies, as possíveis diferenças na riqueza em
espécies e diversidade alfa entre os períodos seca, cheia e vazante foram testadas por
meio de uma análise de variância considerando as 30 isolinhas amostradas (ANOVA
unifatorial).
Uma classificação através do método de Análise de Agrupamento Hierárquico
(cluster) foi utilizado para se verificar a similaridade florística entre os diferentes
períodos. Foi utilizado o índice de similaridade de Sorensen para as matrizes de
presença/ausência, segundo técnicas de Agrupamento pela Associação Média Não-
Ponderada (UPGMA – Unweighted Pair Groups Method with Arithmatic Mean) (Kent
& Coker 1994). Esta análise de classificação foi realizada através do Pacote estatístico
MVSP 3.13 (MultiVariate Statistical Package) (Kovach 2007).
A cobertura regional pelas formas de vida caméfita, fanerófita, geófita,
hemicriptófita e terófita (Raunkiaer 1934) seguiu o mesmo cálculo da cobertura
regional: soma do número de parcelas pontuais nas quais a forma de vida ocorreu / 30 x
250.
Análises para os efeitos ao longo do gradiente de inundação em diferentes períodos:
Para avaliarmos o efeito da profundidade da inundação na riqueza, diversidade,
cobertura e formas de vida, as isolinhas foram agrupadas em classes de profundidade de
9 cm, totalizando seis classes. Para tanto, foram utilizados os valores máximos de
profundidade a que as isolinhas estiveram submetidas durante o ano de estudo. Portanto,
esta profundidade faz referência a que cota altimétrica estas se situam.
Para testarmos o efeito do gradiente de inundação e períodos sobre a riqueza em
herbáceas, as relações entre riqueza, profundidade e períodos amostrados (seca, cheia e
vazante) foram investigadas através de uma Análise de Covariância (ANCOVA),
utilizando o modelo: Y(riqueza em herbáceas) = constante + classes de
profundidade (covariável) + período (fator). O teste de Kolmogov-Smirnov foi
realizado para verificar se a distribuição dos resíduos era normal, utilizando-se para
tanto os valores de probabilidades de Lilliefors. Para esta análise utilizou-se o pacote
estatístico SYSTAT 10.2.
A fim de se evitar o efeito do número de isolinhas por classe de profundidade (o
qual variou) sobre a riqueza em espécies para cada uma dessas classes foi realizada uma
rarefação que comparou a riqueza em espécies estimada para estas 6 classes em função
de um mesmo número de indivíduos registrados. Consideramos o número de toques de

69
herbáceas por classe como sendo o número de indivíduos por classe. A riqueza estimada
para cada classe em 1000 toques foi utilizada para verificarmos posteriormente as
relações entre riqueza e profundidade, utilizando-se para isso a correlação de Spearman.
Aqui não foram realizadas comparações entre períodos. Para a análise de rarefação
utilizou-se o pacote estatístico EstimateS 7.5.2 (Colwell 2006).
Para medir as variações na composição das espécies ao longo do gradiente de
inundação foi calculada a diversidade beta (β) através do índice βT (beta turnover). βT =
[g(H) + l(H)] / 2αm, onde g(H) é o número de espécies novas adicionadas e l(H) o
número de espécies perdidas ao longo do gradiente, sendo αm a média da riqueza das 30
isolinhas. O βT é indicado para amostras que podem ser ordenadas ao longo de um
gradiente dominante, sendo estimado entre pares de amostras adjacentes ou não (Wilson
e Shmida 1984). A diversidade beta foi analisada entre pares de isolinhas adjacentes
ordenados no gradiente de profundidade da inundação. A variação de βT entre pares de
isolinhas adjacentes foi realçada por meio de uma função Lowess.
A cobertura das espécies nas diferentes classes de profundidade para os períodos
de seca, cheia e vazante foram calculadas através da soma do número de parcelas
pontuais nas quais a espécie ocorreu em cada classe de profundidade / total de isolinhas
daquela classe de profundidade x 250. Posteriormente, com estes valores de cobertura
foi realizada uma análise de agrupamento entre as diferentes classes de profundidade
para avaliarmos o efeito do gradiente de inundação na estrutura e composição de
espécies. Para tanto, utilizou o índice de similaridade de Sorensen (Bray-Curtis) e
método de ligação entre grupos beta flexível. Para esta análise foi utilizado o pacote
estatístico PCOrd versão 4.01 (McCune & Mefford 1999).
Para avaliarmos o efeito da profundidade da inundação na cobertura das formas
de vida, o cálculo foi feito da seguinte forma: soma do número de parcelas pontuais nas
quais a forma de vida ocorreu em cada classe de profundidade / total de isolinhas
daquela classe de profundidade x 250, sendo realizados para os três períodos.
As relações entre as formas de vida dominantes e profundidade foram
investigadas através de uma Análise de Correlação (Spearman) para os três períodos
amostrados (seca, cheia e vazante), onde diferentes parâmetros, como riqueza por forma
de vida, cobertura por forma de vida e porcentagem de espécies por forma de vida em
cada classe de profundidade, foram correlacionados com as classes de profundidade.
Para estas análises utilizou-se o pacote estatístico SYSTAT 10.2.

70
RESULTADOS
Efeitos regionais da inundação sazonal sobre a composição, riqueza e diversidade em
espécies – Para os três períodos amostrados, o que totaliza 22.500 parcelas pontuais em
uma área de 25Km2, foram inventariadas 209 espécies, distribuídas em 45 famílias. A
estimativa com Jackknife de primeira ordem foi de 268,3 espécies, enquanto o de
segunda ordem foi de 302,8 espécies. Sessenta espécies ocorreram em apenas uma única
isolinha e 25 espécies em apenas duas isolinhas. Entre os períodos houve variação da
riqueza, entretanto, mantendo as mesmas famílias dominantes, sendo estas Poaceae,
Cyperaceae, Leguminosae e Rubiaceae. O período da seca foi o de menor riqueza, com
113 espécies e 30 famílias, com 25 espécies exclusivas. A riqueza da cheia foi
intermediária, com 139 espécies e 35 famílias, com 18 espécies exclusivas. A vazante
foi o período de maior riqueza com 160 espécies, 41 famílias, com 35 espécies
exclusivas (Tab.1). Esta variação da riqueza esteve relacionada a um incremento de
espécies nas 30 isolinhas nos períodos de cheia e vazante (Tab. 2). Todas as espécies e
famílias estão listadas na tabela 3.
Tabela 1 – Riqueza, diversidade regional (H’), famílias mais ricas e número de espécies
exclusivas por período e geral entre todos os períodos, Pantanal de Poconé-MT.
Seca Cheia Vazante Todos os períodos
Riqueza 113 139 160 209
Jack1 =268,3 Jack2 = 302,8
H’ 2,82 3,13 3,22
Total famílias 30 35 41 45
Famílias mais ricas
Poaceae(41), Cyperaceae (14), Leguminosae (9): Fabaceae (6) e Caesalpiniaceae (3),Rubiaceae (7), Euphorbiaceae (6) e Lamiaceae (5)
Poaceae (41), Cyperaceae (23), Leguminosae (8): Fabaceae (5), Caesalpiniaceae (3) e Mimosaceae (1), Rubiaceae (6), Euphorbiaceae (6), Lamiaceae (5)
Poaceae (48), Cyperaceae (20), Leguminosae (10): Fabaceae (6), Caesalpiniaceae (2) e Mimosaceae (2), Rubiaceae (8), Euphorbiaceae (7), Lamiaceae (6), Commelinaceae (5) e Alismataceae (5)
Poaceae (59), Cyperaceae (30), Leguminosae (14): Fabaceae (9), Caesalpiniaceae (3) e Mimosaceae (2) , Rubiaceae (10), Euphorbiaceae (8), Lamiaceae (8), Scrophulariaceae (6), Commelinaceae (5) e Alismataceae (5)
Espécies exclusivas
25 18 35

71
A similaridade florística entre os períodos foi maior entre cheia e a vazante,
76,9%, seguida de seca e cheia, 61,9%, e seca e vazante, 60,1% (Fig. 3). Ao todo 72
espécies são comuns aos três períodos. A cheia e a vazante compartilham um grande
número de espécies (115 espécies).
Figura 3 – Dendrograma de similaridade florística de herbáceas entre os períodos de
seca, cheia e vazante, Pantanal de Poconé-MT.

72
Tabela 2 – Sub-tipos de vegetação, valores de duração e profundidade da inundação, classes de profundidade da inundação, riqueza em espécies, diversidade de Shannon-wienner (H’) e equitabilidade (J) para as 30 isolinhas amostrados nos diferentes períodos (seca, cheia e vazante), Pantanal de Poconé-MT.
SECA CHEIA VAZANTE
Isolinhas Sub-tipos de vegetação Duração Profundidade Classes H' J Riqueza H' J Riqueza H' J Riqueza
A0500 Floresta semi-sempre verde sazonalmente inundável
111 30 4 2.087 0.84 12 2.207 0.749 19 2.446 0.903 15
A1500 Floresta semi-sempre verde sazonalmente inundável
104 42 5 2.061 0.804 13 2.037 0.692 19 1.919 0.692 16
A2500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 17 2 1.502 0.604 12 2.267 0.688 27 1.721 0.557 22
A3500 Savana sazonalmente inundável com térmitas (campo de murundus)
104 21 3 1.084 0.471 10 2.005 0.659 21 1.731 0.56 22
A4500 Savana sazonalmente inundável com térmitas (campo de murundus)
104 25 3 1.235 0.497 12 2.376 0.692 31 2.196 0.639 31
B0500 Campo exótico sazonalmente inundável 42 15 2 1.045 0.502 8 1.63 0.527 22 1.855 0.557 28
B1500 Floresta sazonal semi-decídua sobre solo seco (cerradão)
0 0 1 1.833 0.623 19 2.388 0.733 26 2.242 0.688 26
B2500 Floresta semi-sempre verde sazonalmente inundável
154 50 6 1.967 0.895 9 2.233 0.758 19 2.179 0.754 18
B3500
Interface: Savana sazonalmente inundável com térmitas (campo de murundus)/ floresta sazonal semi-decídua sobre solo seco (cerradão)
6 7 1 1.715 0.669 13 2.335 0.674 32 2.644 0.75 34
B4500 Campo exótico sazonalmente inundável 56 16 2 0.677 0.42 5 1.299 0.506 13 1.779 0.642 16
C0500 Campo exótico sazonalmente inundável 89 23 3 0.541 0.26 8 1.62 0.572 17 1.347 0.458 19
C1500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 16 2 1.063 0.414 13 1.761 0.562 23 1.743 0.582 20
C2500 Savana sazonalmente inundável com térmitas (campo de murundus)
104 25 3 1.251 0.777 5 1.977 0.66 20 2.161 0.68 24
C3500 Savana sazonalmente inundável com térmitas (campo de murundus)
110 29 4 1.918 0.677 17 2.596 0.742 33 2.607 0.759 31
continua....

73
continuação
Tab. 2
C4500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 14 2 1.614 0.701 10 2.372 0.747 24 2.127 0.638 28
D0500 Campo exótico sazonalmente inundável 200 58 6 0.912 0.38 11 1.794 0.647 16 1.683 0.562 20
D1500 Interface: campo limpo/Floresta semi-sempre verde sazonalmente inundável
200 63 6 2.154 0.687 23 2.27 0.697 26 2.437 0.777 23
D2500 Floresta semi-sempre verde sazonalmente inundável
126 35 4 2.031 0.847 11 2.113 0.824 13 2.479 0.875 17
D3500 Floresta semi-sempre verde sazonalmente inundável
90 16 2 2.168 0.782 16 2.633 0.782 29 2.579 0.801 25
D4500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 26 3 0.96 0.401 11 1.901 0.702 15 1.457 0.552 14
E0500 Savana sazonalmente inundável com térmitas (campo de murundus)
125 44 5 1.807 0.625 18 2.381 0.74 25 2.067 0.702 19
E1500 Floresta sazonal semi-decídua sobre solo seco (cerradão)
0 0 2 1.528 0.596 13 1.286 0.518 12 1.645 0.549 20
E2500 Interface: campo limpo/ floresta sazonal semi-decídua sobre solo seco (cerradão)
90 26 3 2.256 0.701 25 2.337 0.78 20 2.761 0.858 25
E3500
Interface: Savana sazonalmente inundável com térmitas (campo de murundus)/ floresta sazonal semi-decídua sobre solo seco (cerradão)
23 7 1 2.245 0.737 21 2.451 0.752 26 2.23 0.693 25
E4500 Floresta semi-sempreverde sazonalmente inundável
125 39 5 1.707 0.647 14 2.028 0.816 12 1.803 0.867 8
F0500 Savana sazonalmente inundável com térmitas (campo de murundus)
95 42 5 1.676 0.58 18 2.089 0.641 26 2.247 0.661 30
F1500 Floresta sazonal semi-decídua sobre solo seco (cerradão)
0 0 1 1.706 0.56 21 2.023 0.614 27 2.618 0.742 34
F2500 Campo exótico sazonalmente inundável 110 18 2 2.186 0.697 23 2.516 0.763 27 2.774 0.842 27
F3500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 24 3 2.073 0.652 24 2.143 0.674 24 1.771 0.601 19
F4500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 24 3 1.466 0.529 16 2.288 0.66 32 1.912 0.602 24
Média 90.267 25.067 1.616 0.619 14.367 2.112 0.686 22.533 2.105 0.685 22.667
Desvio padrão 50.308 16.144 0.493 0.154 5.5241 0.343 0.085 6.1797 0.398 0.117 6.1886

74
Uma visualização gráfica da diversidade entre os períodos demonstra que poucas
espécies foram abundantes, sendo bastante elevado o número de espécies raras ou
relativamente raras (Fig.4). Em termos de diversidade regional houve variação entre os
períodos de seca, cheia e vazante, sendo os valores do índice de Shannon-wienner (H’),
respectivamente 2,82, 3,13 e 3,22 (Tab.1). As médias da riqueza e da diversidade alfa (H’) das
30 isolinhas (Tab.2) comparadas pela ANOVA entre os períodos mostrou diferenças
significativas entre os períodos de seca e cheia como também entre seca e vazante (p<0,05).
Figura 4 – Distribuição de abundâncias das espécies herbáceas ocorrentes nas 30 isolinhas e
períodos amostrados, Pantanal de Poconé – MT.
Efeitos regionais da inundação sobre a cobertura de espécies –. Na tabela 3 estão listadas as
espécies presentes nos diferentes períodos com respectivos valores de cobertura regional.
Entre as 5 espécies com elevada cobertura regional durante o período de seca, podemos citar
Axonopus purpusii (0.1441), Paspalum lineare (0.1246), Brachiaria humidicola (0.0904),
Bromelia balansae (0.0206) e Reimarochloa brasiliensis (0.0205). Algumas destas espécies
persistem durante a cheia, mas ocorre o estabelecimento de espécies aquáticas que alcançam
elevada cobertura, e dentre as 5 espécies de maior valor de cobertura regional para este
período podemos citar: Eleocharis minima (0.1674), Brachiaria humidicola (0.1428),
Paspalum lineare (0.1297), Axonopus purpusii (0.1062) e Utricularia foliosa (0.0852). Na da
vazante, as espécies com maior cobertura regional foram Axonopus purpusii (0.1358),
0 50 100 150 200
Rank das Espécies
0.0001
0.0010
0.0100
0.1000
Co
be
rtu
ra r
eg
ion
al
VazanteCheiaSeca

75
Paspalum lineare (0.1276), Brachiaria humidicola (0.1218), Eleocharis minima (0.0824),
Eleocharis acutangula (0.0824) e Bromelia balansae (0.0232). Portanto, Axonopus purpusii,
Paspalum lineare e Brachiaria humidicola possuem elevada cobertura regional nos três
períodos. A variação do valor de cobertura regional destas espécies pode ser melhor
observada na Fig. 5. Entre as espécies com maior cobertura durante os três períodos,
Axonopus purpusii e Brachiaria humidicola, apresentaram variações mais acentuadas,
enquanto que a cobertura de Paspalum lineare praticamente não variou.
Algumas espécies são altamente variáveis, ocorrendo com baixa cobertura (ou não
registradas) durante a seca e a vazante aumentando durante a cheia, como Eleocharis minima,
Cabomba furfucata, Eleocharis acutangula, Sagittaria guyanensis e Utricularia foliosa.
Espécies como Cabomba furfucata, Sagittaria guyanensis e Utricularia foliosa não ocorreram
na seca. Outras como Croton glandulosus, Reimarochloa brasiliensis, Setaria geniculata e
Richardia grandiflora tiveram maior cobertura durante a seca e diminuíram ou desapareceram
durante a cheia e a vazante.
Tabela 3 – Espécies ocorrentes nas 30 isolinhas amostradas nos períodos de seca, cheia e
vazante, valores de cobertura regional e formas de vida, Pantanal de Poconé-MT. C = caméfita,
F = fanerófita, G = geófita; H = hemicriptífita, T = terófita, H/C = hemicriptófita/ caméfita, T/C =
terófita/caméfita e N = não identificada.
Família Nome científico Seca Cheia Vazante Forma de
vida Acanthaceae Ruellia erythropus (Nees) Lindau 0 0.00053 0.00093 H
Acanthaceae Ruellia gemminiflora H.B.K 0.0004 0.00013 0.00027 H
Acanthaceae Staurogyne diantheroides Lindau 0.002 0.008 0.00493 H
Alismataceae Echinodorus lanceolatus Rataj 0.00013 0.00253 0.00347 G
Alismataceae Echinodorus macrophyllus (Kunth) Micheli
0 0 0.00013 H
Alismataceae Echinodorus tenellus (Mart. ex Schult. & Schult. f.) Buchenau
0 0 0.00133 T
Alismataceae Sagittaria guyanensis H.B.K 0 0.04213 0.0004 G
Alismataceae Sagittaria rhombifolia Cham. 0 0.00667 0.002 H
Araceae Dracontium margaretae Bogner 0 0.00013 0.00013 G
Arecaceae Allagoptera leucocalyx (Drude) Kuntze
0.0012 0.00107 0.0012 G
Asteraceae Bidens gardneri Baker 0 0.00027 0.00027 C
Asteraceae Eupatorium squalidum DC. 0.00013 0.00053 0.00067 F
Asteraceae Praxelis clematidea (Griseb.) R.M. King & H. Rob.
0.00493 0 0.0008 H
continua.....

76
continuação Tab.3
Asteraceae Vernonia rubricaulis Bonpl. 0 0.0004 0.00067 H
Boraginaceae Heliotropium filiforme Lehm. 0 0 0.00013 T
Boraginaceae Heliotropium paradoxum Gürke 0.0028 0 0 T
Bromeliaceae Bromelia balansae Mez 0.02067 0.02453 0.0232 G
Cabombaceae Cabomba furcata Schult. & Schult. f.
0 0.022 0.0196 T
Caesalpiniaceae Chamaecrista desvauxii (Collad.) Killip var. glauca (Hassl.) H.S. Irwin & Barneby
0.00013 0.00027 0 H
Caesalpiniaceae Chamaecrista serpens (L.) Greene 0.00053 0.0004 0.0004 H
Caesalpiniaceae Senna splendida (Vogel) H.S. Irwin & Barneby
0.00013 0.00013 0.00013 H
Commelinaceae Commelina benghalensis L.* 0 0 0.00013 G
Commelinaceae Commelina cf. erecta L. 0.00067 0.00013 0.0004 G
Commelinaceae Commelina sp. 0 0 0.00093 N
Commelinaceae Murdannia nudiflora (L.) Brenan 0 0.00187 0.00027 G
Commelinaceae Murdannia sp. 0 0 0.00133 T
Convolvulaceae Ipomoea asarifolia (Desr.) Roem. & Schult.
0.00053 0.0016 0.00093 H
Convolvulaceae Ipomoea carnea Jacq. 0 0.00013 0 H
Convolvulaceae Ipomoea cf. hirsuta R. Br. 0 0.00013 0.00013 N
Cyperaceae Bulbostylis sp. 0.00013 0 0 N
Cyperaceae Cyperaceae1 0.00027 0 0 N
Cyperaceae Cyperaceae2 0 0.00013 0 N
Cyperaceae Cyperus aggregatus (Willd.) Endl.
0.00027 0 0 G
Cyperaceae Cyperus brevifolius (Rottb.) Endl. ex Hassk.
0.00053 0.00053 0.00053 G
Cyperaceae Cyperus cf. cornellii-ostenii Kük. 0.00027 0.00027 0.0004 H
Cyperaceae Cyperus cf. iria L. 0.00027 0 0.0004 T
Cyperaceae Cyperus esculentus L. 0 0.00013 0 G
Cyperaceae Cyperus haspan ssp. juncoides (Lam.) Kük.
0.0012 0.00507 0.00253 G
Cyperaceae Cyperus incomptus Kunth. 0 0 0.00013 H
Cyperaceae Cyperus luzulae (L.) Rottb. ex Retz.
0.00013 0.0008 0.00027 G
Cyperaceae Cyperus schomburgkianus (Schr.) Kuk.
0 0.00027 0 H
Cyperaceae Cyperus sesquiflorus (Torr.) Mattf. & Kük.
0.00093 0.00173 0.00067 H
Cyperaceae Eleocharis acutangula (Roxb.) Schult.
0.00173 0.04613 0.0248 G
Cyperaceae Eleocharis geniculata (L.) Roem. & Schult.
0.00107 0.0088 0.00227 H
Cyperaceae Eleocharis minima Kunth 0.00453 0.16747 0.0824 T
Cyperaceae Eleocharis nudipes (Kunth) Palla 0 0.00027 0.00027 H
continua......
77
continuação Tab. 3
Cyperaceae Eleocharis plicarhachis (Griseb.) Svenson
0 0.01733 0.01307 H
Cyperaceae Fimbristylis dichotoma (L.) Vahl 0 0 0.00053 T
Cyperaceae Fimbristylis miliacea (L.) Vahl 0 0.00067 0 H
Cyperaceae Fimbristylis sp. 0 0.00013 0 N
Cyperaceae Hemicarpha micrantha (Vahl) Pax
0 0.00013 0 T
Cyperaceae Lipocarpha humboldtiana Nees 0 0 0.00027 T
Cyperaceae Rhynchospora brevirostris Griseb.
0 0.0012 0.0124 G
Cyperaceae Rhynchospora hirta (Nees) Boeck.
0 0.0028 0.0076 G
Cyperaceae Rhynchospora nervosa (Vahl) Boeckeler
0.00013 0.00027 0.00027 G
Cyperaceae Scleria leptostachya Kunth 0 0.00013 0 H/T
Cyperaceae Scleria melaleuca Rchb. ex Schltdl. & Cham.
0.0012 0.00147 0.00213 H
Cyperaceae Scleria reticularis Michx. 0 0.00187 0.0016 G
Cyperaceae Scleria sp. 0 0.00013 0.00013 N
Eriocaulaceae Philodice hoffmannseggii Mart. 0 0.00533 0.00147 T
Eriocaulaceae Syngonanthus gracilis (Bong.) Ruhland
0 0.00067 0.00107 T
Euphorbiaceae Caperonia castaneifolia (L.) A. St.-Hil.
0.00067 0.0024 0.0012 H
Euphorbiaceae Croton glandulosus (L.) Muell. Arg.
0.0108 0 0.00133 T
Euphorbiaceae Euphorbia hyssopifolia L. 0.0004 0.00013 0 T
Euphorbiaceae Euphorbia thymifolia L. 0.0012 0 0.00013 H
Euphorbiaceae Phyllanthus hyssopifolioides Kunth
0 0.00027 0.0004 T
Euphorbiaceae Phyllanthus stipulatus (Raf.) G.L. Webster
0.00067 0.00573 0.0144 T
Euphorbiaceae Sebastiania hispida (Mart.) Pax ex Engl.
0.00013 0.00013 0.00053 T
Euphorbiaceae Sebastiania salicifolia (Mart.) Pax 0 0.00133 0.00133 H
Fabaceae Aeschynomene fluminensis Vell. 0.00053 0 0.00027 C
Fabaceae Aeschynomene sensitiva Sw. 0 0 0.00013 C
Fabaceae Alysicarpus vaginalis (L.) DC.* 0 0 0.00013 H
Fabaceae Arachis kuhlmannii Krapov. & W.C. Gregory
0.00027 0.0004 0 H
Fabaceae Calopogonium mucunoides Desv. 0 0.00013 0.00067 H
Fabaceae Desmodium barbatum (L.) Benth. 0.00027 0 0 H
Fabaceae Discolobium psoraliaefolium Benth.
0.0004 0.00227 0.00133 H
Fabaceae Eriosema platycarpon Micheli 0.00013 0.00013 0 H
Fabaceae Zornia latifolia Sm. 0.00027 0 0.00013 H
continua.....

78
continuação Tab. 3
Gentianaceae Coutoubea ramosa Aubl. 0.00013 0 0 T
Hydrophyllaceae Hydrolea spinosa L. 0.00013 0 0.00013 C
Iridaceae Cipura paludosa Aubl. 0 0.00013 0.00027 G
Iridaceae Trimezia sp. 0 0.00013 0 G
Lamiaceae Hyptis brevipes Poit. 0.00547 0.0008 0.0004 H
Lamiaceae Hyptis cf. peduncularis Benth. 0.00027 0.00053 0.00187 N
Lamiaceae Hyptis crenata Pohl ex Benth. 0.00173 0.00067 0.0012 H
Lamiaceae Hyptis lorentziana O. Hoffm. 0.00133 0.0012 0.00093 T
Lamiaceae Hyptis microphylla Pohl ex Benth.
0 0.00027 0 C
Lamiaceae Hyptis mutabilis (Rich.) Briq. 0 0 0.00013 T
Lamiaceae Hyptis suaveolens (L.) Poit.* 0 0 0.00013 C
Lamiaceae Peltodon tomentosus Pohl 0.00027 0 0 H
Lentibulariaceae Utricularia breviscapa Wright ex Griseb.
0 0 0.00027 T
Lentibulariaceae Utricularia foliosa L. 0 0.0852 0.00107 G
Lentibulariaceae Utricularia hydrocarpa Vahl 0 0 0.00053 G
Lentibulariaceae Utricularia warmingii Kamienski 0 0.00267 0.00093 T
Limnocharitaceae/Butomaceae
Hydrocleys nymphoides (Willd.) Buchenau
0 0.00013 0 N
Limnocharitaceae/Butomaceae
Hydrocleys parviflora Seub. 0 0.00027 0.00067 T
Lythraceae Cuphea carthagenensis (Jacq.) J.F. Macbr.
0 0.00093 0.00053 H
Lythraceae Cuphea cf. retrorsicarpilla Koehne
0.00053 0 0.0004 C
Lythraceae Rotala mexicana Schltdl. & Cham.
0 0 0.0004 T
Malpighiaceae Galphimia brasiliensis (L.) A. Juss.
0.00013 0 0 H
Malvaceae Pavonia angustifolia Benth. 0 0.00013 0 C
Malvaceae Sida santaremensis Monteiro 0.00067 0.00093 0.00013 C
Mayacaceae Mayaca sellowiana Kunth 0 0.0048 0.00627 T
Mimosaceae Mimosa debilis Humb. & Bonpl. ex Willd.
0 0 0.00013 F
Mimosaceae Mimosa polycarpa Kunth 0 0.00027 0.00027 H
Nymphaeaceae Nymphaea gardneriana/jasmesoniana
0 0.01227 0.00053 G
Nymphaeaceae Nymphaea oxypetala Planch. 0 0.00227 0 G
Ochnaceae Sauvagesia erecta L. 0.00013 0 0.00053 H
Onagraceae Ludwigia longifolia (DC.) H. Hara
0 0.00027 0.0052 H
Onagraceae Ludwigia octovalvis (Jacq.) P.H. Raven
0.00147 0.00733 0.01573 T
Orquidaceae Habenaria cf. orchiocalcar Hoehne
0 0.00013 0 G
continua......

79
continuação Tab. 3
Orquidaceae Habenaria repens Nutt. 0 0.00027 0 H
Oxalidaceae Oxalis physocalyx Zucc. ex Progel
0.00013 0.0004 0.00093 H
Passifloraceae Passiflora tricuspis Mast. 0.00013 0 0 H
Poaceae Andropogon bicornis L. 0.00187 0.00093 0.0012 H
Poaceae Andropogon hypogynus Hack. 0.0044 0.0044 0.00493 H
Poaceae Andropogon selloanus (Hack.) Hack.
0.0044 0.0072 0.00587 H
Poaceae Aristida cappilacea Lam. 0 0.00027 0.00213 T
Poaceae Aristida cf. riparia Trin. 0 0 0.00027 H
Poaceae Aristida sp. 0.0004 0 0 H
Poaceae Axonopus cf. brasiliensis (Spreng.) Kuhlm.
0 0 0.00013 H
Poaceae Axonopus chrysoblepharis (Lag.) Chase
0.00027 0.00027 0.0008 H
Poaceae Axonopus compressus (Sw.) P. Beauv.
0.0004 0.0004 0.0004 H
Poaceae Axonopus leptostachyus (Flüggé) Hitchc.
0.0036 0.00147 0.00173 H
Poaceae Axonopus purpusii (Mez) Chase 0.14413 0.10627 0.13587 H
Poaceae Brachiaria humidicola (Rendle) Schweick.
0.0904 0.1428 0.12187 H
Poaceae Coelorachis aurita (Steud.) A. Camus
0 0.00013 0.00013 H
Poaceae Digitaria ciliaris (Retz.) Koeler 0 0 0.00013 T
Poaceae Digitaria sp.3 0.0004 0.00013 0.00013 N
Poaceae Digitaria sp.4 0 0 0.00013 N
Poaceae Echinolaena gracilis Swallen 0.00027 0.00187 0.00187 H
Poaceae Elionurus muticus (Spreng.) Kuntze
0.0124 0.0152 0.01253 H
Poaceae Eragrostis articulata (Schrank) Nees
0.00067 0 0 T
Poaceae Eragrostis rufescens Schrad. ex Schult.
0.00067 0.00027 0.00067 T
Poaceae Ichnanthus procurrens (Nees ex Trin.) Swallen
0.00707 0.00467 0.00347 H
Poaceae Imperata brasiliensis Trin. 0.0044 0.00547 0.00573 H
Poaceae Imperata tenuis Hack. 0.00013 0.0004 0.00027 H
Poaceae Lasiacis sorghoidea (Desv. ex Ham.) Hitchc. & Chase
0 0 0.00013 H
Poaceae Leersia hexandra Sw. 0.00013 0 0 H
Poaceae Leptochloa virgata (L.) P. Beauv. 0.0004 0 0 H
Poaceae Leptocoryphium lanatum (Kunth) Nees
0.0036 0.00453 0.00347 H
Poaceae Luziola fragilis Swallen 0 0.00333 0.00067 H
Poaceae Mesosetum ansatum (Trin.) Kuhlm.
0.00453 0.0108 0.00253 H
continua.....
80
continuação Tab. 3
Poaceae Mesosetum chaseae Luces 0.00107 0.0004 0 H
Poaceae Mesosetum loliiforme (Hochst. ex Steud.) Chase
0.00813 0.01053 0.00987 H
Poaceae Otachyrium versicolor (Döll) Henrard
0 0.00013 0.0016 H
Poaceae Panicum dichotomiflorum Michx. 0.00013 0 0 T
Poaceae Panicum discrepans Döll 0.00148 0 0 T
Poaceae Panicum laxum Sw. 0.00507 0.01173 0.0092 H
Poaceae Panicum pilosum Sw. 0.00107 0.0008 0.00067 H
Poaceae Panicum stenodes Griseb. 0.01 0.01947 0.01493 H
Poaceae Panicum trichanthum Nees 0 0.0004 0.00013 T
Poaceae Paratheria prostrata Griseb. 0.00013 0 0 T
Poaceae Paspalum carinatum Humb. & Bonpl. ex Flüggé
0.00307 0.00067 0.00107 H
Poaceae Paspalum delicatum Swallen 0 0 0.00013 T
Poaceae Paspalum gardnerianum Nees 0.00013 0 0 T
Poaceae Paspalum lineare Trin. 0.12467 0.12973 0.1276 H
Poaceae Paspalum multicaule Poir. 0 0.00027 0.00093 T
Poaceae Paspalum plicatulum Michx. 0.0056 0.0048 0.01053 H
Poaceae Paspalum sp.1 0.00147 0.00147 0.0012 H
Poaceae Paspalum sp.2 0 0.00053 0.0016 H
Poaceae Paspalum stellatum Humb. & Bonpl. ex Flüggé
0 0.00067 0.00333 H
Poaceae Paspalum subciliatum Chase 0.0012 0 0 H
Poaceae Poaceae21 0 0 0.00067 N
Poaceae Poaceae31 0 0 0.00013 N
Poaceae Poaceae9 0.00013 0 0 H
Poaceae Reimarochloa brasiliensis (Spreng.) Hitchc.
0.02053 0.00013 0.00027 T
Poaceae Sacciolepis myuros (Lam.) Chase 0 0.0004 0.00133 T
Poaceae Schizachyrium microstachyum (Desv. ex Ham.) Roseng.
0.0008 0.00053 0.00027 H
Poaceae Schizachyrium tenerum Nees 0.0056 0.00733 0.00987 H
Poaceae Setaria geniculata P. Beauv. 0.01053 0.00013 0.00253 T
Poaceae Setaria vulpiseta (Lam.) Roem. & Schult.
0 0.00013 0.00027 H
Poaceae Sorghastrum setosum (Griseb.) Hitchc.
0.006 0.00627 0.0072 H
Primulaceae Centunculus minimus L. 0 0 0.00053 T
Pteridaceae Adiantum fructuosum Poepp. ex Spreng.
0.00213 0.00227 0.00427 H
Rubiaceae Borreria quadrifaria E.L. Cabral 0.00013 0.00493 0.00093 H
Rubiaceae Diodella radula (Willd. & Hoffmanns. ex Roem. & Schult.) Delprete
0.00013 0 0 H
continua......

81
continuação Tab. 3
Rubiaceae Diodia kuntzei K. Schum. 0.00373 0.0072 0.00547 H
Rubiaceae Limnosipanea cf. ternifolia (Pittier)
0 0 0.0048 T
Rubiaceae Limnosipanea schomburgkii Hook. f.
0 0 0.00173 T
Rubiaceae Mitracarpus parvulus K. Schum. 0.00027 0.00053 0.00013 N
Rubiaceae Richardia grandiflora Cham. & Schltdl.) Steud.
0.01093 0.00013 0.00067 H
Rubiaceae Sipanea biflora (L. f.) Cham. & Schltdl.
0.0028 0.0012 0.00147 H
Rubiaceae Spermacocedes glabrum (Michx.) 0 0.0004 0 T
Rubiaceae Staelia virgata (Link ex Roem. & Schult.) K. Schum.
0.0004 0 0.00027 T
Schizaeaceae Lygodium sp. 0.00053 0.00093 0.00067 G
Scrophulariaceae Agallinis glandulosa (Barroso) V.C. Souza
0.00027 0 0 T
Scrophulariaceae Bacopa monnierioides (Cham.) B.L. Rob.
0 0.00093 0.00013 H
Scrophulariaceae Bacopa myriophylloides Wettst. 0 0 0.00187 T
Scrophulariaceae Bacopa reflexa (Benth.) Edwall 0 0.00507 0 T
Scrophulariaceae Bacopa salzmanii (Benth.) Wettst. ex Edwall
0 0.00227 0.00053 T
Scrophulariaceae Buchnera palustris (Aubl.) Spreng.
0 0 0.00013 H
Solanaceae Schwenckia angustifolia Benth. 0 0.0004 0.00013 T
Solanaceae Solanum viarum Dunal 0 0.00013 0 H
Sterculiaceae Byttneria genistella Triana & Planch.
0.00027 0.00173 0.00093 H
Sterculiaceae Melochia arenosa Benth. 0.0044 0.0012 0.00213 C
Sterculiaceae Melochia graminifolia A. St.-Hil. 0.00027 0 0 H
Sterculiaceae Melochia villosa (Mill.) Fawc. & Rendle
0.00093 0.00133 0.00173 H
Turneraceae Piriqueta cistoides (L.) Griseb. 0 0.00027 0.0004 T/C
Turneraceae Turnera melochioides A. St.-Hil. & Cambess.
0.00013 0.00027 0 H
Verbenaceae Lippia alba (Mill.) N.E. Br. 0 0 0.00013 C
Vitaceae Cissus erosa Rich. 0.00013 0.00093 0.00093 F
Xyridaceae Xyris savannensis Miq. 0 0 0.00013 H
Zingiberaceae Costus arabicus L. 0 0 0.00013 G
Indeterminada (Calathea) 0 0 0.00027 N
Indeterminada1* 0.00013 0 0 N
Indeterminada17 coletada 0 0 0.00027 N
Indeterminada5 0.00013 0 0 N
Indeterminada8* 0.00013 0 0 N
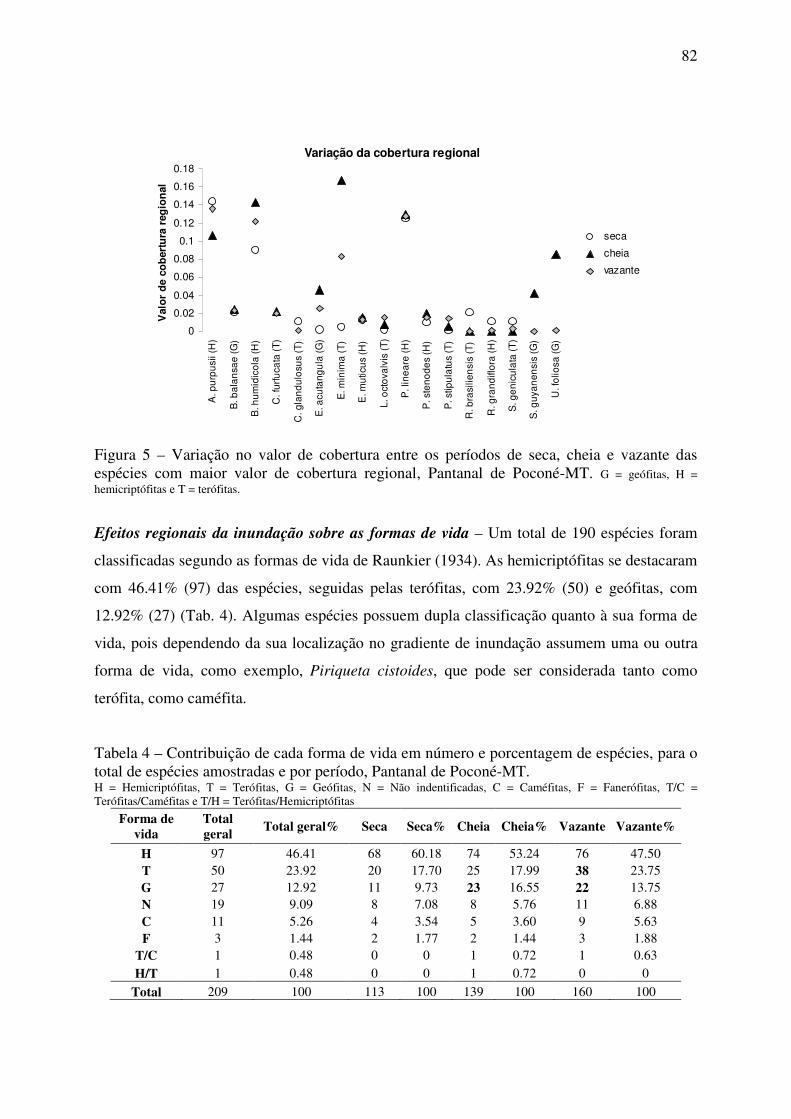
82
Variação da cobertura regional
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
A. p
urp
usii (
H)
B. b
ala
nsa
e (
G)
B. h
um
idic
ola
(H
)
C. fu
rfu
ca
ta (
T)
C. g
lan
du
losu
s (
T)
E. a
cu
tan
gu
la (
G)
E. m
inim
a (
T)
E. m
uticu
s (
H)
L. o
cto
va
lvis
(T
)
P. lin
ea
re (
H)
P. ste
no
de
s (
H)
P. stip
ula
tus (
T)
R. b
rasilie
nsis
(T
)
R. g
ran
diflo
ra (
H)
S. g
en
icu
lata
(T
)
S. g
uya
ne
nsis
(G
)
U. fo
lio
sa
(G
)
Valo
r d
e c
ob
ert
ura
reg
ion
al
seca
cheia
vazante
Figura 5 – Variação no valor de cobertura entre os períodos de seca, cheia e vazante das espécies com maior valor de cobertura regional, Pantanal de Poconé-MT. G = geófitas, H = hemicriptófitas e T = terófitas.
Efeitos regionais da inundação sobre as formas de vida – Um total de 190 espécies foram
classificadas segundo as formas de vida de Raunkier (1934). As hemicriptófitas se destacaram
com 46.41% (97) das espécies, seguidas pelas terófitas, com 23.92% (50) e geófitas, com
12.92% (27) (Tab. 4). Algumas espécies possuem dupla classificação quanto à sua forma de
vida, pois dependendo da sua localização no gradiente de inundação assumem uma ou outra
forma de vida, como exemplo, Piriqueta cistoides, que pode ser considerada tanto como
terófita, como caméfita.
Tabela 4 – Contribuição de cada forma de vida em número e porcentagem de espécies, para o total de espécies amostradas e por período, Pantanal de Poconé-MT. H = Hemicriptófitas, T = Terófitas, G = Geófitas, N = Não indentificadas, C = Caméfitas, F = Fanerófitas, T/C = Terófitas/Caméfitas e T/H = Terófitas/Hemicriptófitas
Forma de vida
Total geral
Total geral% Seca Seca% Cheia Cheia% Vazante Vazante%
H 97 46.41 68 60.18 74 53.24 76 47.50 T 50 23.92 20 17.70 25 17.99 38 23.75 G 27 12.92 11 9.73 23 16.55 22 13.75 N 19 9.09 8 7.08 8 5.76 11 6.88 C 11 5.26 4 3.54 5 3.60 9 5.63 F 3 1.44 2 1.77 2 1.44 3 1.88
T/C 1 0.48 0 0 1 0.72 1 0.63
H/T 1 0.48 0 0 1 0.72 0 0
Total 209 100 113 100 139 100 160 100

83
O aumento da riqueza entre os períodos de cheia e vazante em relação à seca está
ligado ao estabelecimento ou restabelecimento de espécies hemicriptófitas, geófitas e terófitas
relacionadas ao pulso de inundação (Tab.4), algumas com aumentos notáveis na cobertura na
transição seca/cheia. Especialmente as geófitas foram extremamente vinculadas ao período de
cheia e vazante, participando com maior riqueza e elevada cobertura regional, principalmente,
durante a cheia (Fig. 6 e 7). As terófitas tiveram forte incremento de espécies durante o
período de vazante, entretanto, com menor cobertura regional do que na cheia. Na figura 7
nota-se que muitas hemicriptófitas, comuns aos períodos nas transições seca/cheia e
cheia/vazante, possuem menor variação na cobertura, sendo estas basicamente gramíneas
(destaque em vermelho) com maior estabilidade na cobertura frente a mudança hídrica
sazonal. As três espécies em destaque e já citadas anteriormente são Axonopus purpusii,
Paspalum lineare e Brachiaria humidicola. Além destas, podemos citar: Panicum stenodes,
Elionurus muticus, Mesosetum lolliforme, Panicum laxum, Schizachyrium tenerum, Imperata
brasiliensis e Andropogon selloanus.
Figura 6 – Valor de cobertura regional das formas de vida nos períodos de seca, cheia e
vazante, Pantanal de Poconé-MT.

84
1 10 100
Cobertura regional da seca
1
10
100
Co
be
rtu
ra r
eg
i on
al d
a c
he
i a
GG
G
G
G
G
G
G
G
G
G
GGGG
G
G
G
G
G
G
G
G
G
HHH HHH HH HHHH HHHH
HH
HH
H
HH
H
H
H
H
H
H
HH
HH
HH
H
H
H
H
H
H
H
HH
H
H
H
H
H
H
H
H
H
HH
HH
H
HH
H
H
HHH
H
H
H
HH
H
HH
H
H
HHH
H
HH
HHH
H
H
HHH
TT TT T TT TTT TTTT
T
T
T
TT
T
T
T
T
T
TT
T
TT
T
TTTTT
T
1 10 100
Cobertura regional da cheia
1
10
100
Cobert
ura
regio
nal da v
azante
G
GG
G
G
GG
G
G
G
G
G
G
GGG
GGGG
G
G G
GG
G
G
H
H
H
H
H
H HH
HH
H
H
H
HH
HHH
HH H
H H
H
H
H
HHHH
H
HHH
H
H
H
H
H HH
HH
H HH HHHH
H
HHHH
HHH HH HH
H HHH
H HHHHHH
H
HH
H
HH
HHH
H
H
HH
T
T
T
T
T
TTT
T
TT
T
T
TT
T
T
T
T
T
TT
T
T
TT
TTT
TT TT
T
T
T
TT
T
T
TT
Figura 7 – Cobertura regional das espécies, representadas por símbolos de suas respectivas
forma de vida, nas transições entre seca / cheia e cheia / vazante, Pantanal de Poconé-MT. H
= hemicriptófitas, T = terófitas e G = geófitas. Em vermelho destaque às gramíneas e em azul as
demais herbáceas.

85
No resumo da tabela 5, relacionada à figura 7, verifica-se que para todas as formas de
vida há espécies comuns e exclusivas entre os diferentes períodos. Na transição seca/cheia,
portanto, o período de seca é desfavorável a um certo grupo de espécies de hemicriptófitas,
terófitas e geófitas, enquanto a cheia a outro grupo destas formas de vida, entretanto, há
espécies comuns. Na transição cheia/vazante há um maior compartilhamento de espécies,
entretanto, com um elevado número de terófitas exclusivas da vazante, pois, possivelmente, o
banco de sementes é ativado após a cheia.
Tabela 5 – Tabela resumo com totalizações do número de espécies entre as transições
seca/cheia e cheia vazante, Pantanal de Poconé – MT. Hemi = hemicriptófitas, Tero = terófitas e Geo
= geófitas.
Hemi Tero Geo
Total na transição seca/cheia 89 36 24
Exclusiva da seca 15 11 1
Exclusiva da cheia 22 16 13
Comuns 52 9 10
Total na transição cheia/vazante 86 42 26
Exclusiva da cheia 10 4 4
Exclusiva da vazante 12 17 3
Comuns 64 21 19
Efeito do gradiente de inundação na riqueza em espécies- A riqueza em herbáceas diminui
significativamente com o aumento da profundidade das isolinhas (p < 0,001; r2 = 0.84) sendo
esta variação dependente dos períodos (p =0.001) (Fig. 8). A correlação de Spearman entre a
riqueza estimada por rarefação para 1000 indivíduos (toques) e a profundidade foi
significativa para o teste unicaudal (rs = 0.82.9).

86
0 1 2 3 4 5 6 7
Classes de profundidade
20
30
40
50
60
70
80
90
100
Riq
ue
za
em
he
rbáce
as
VazanteCheiaSeca
Períodos
Figura 8 – Relação entre a riqueza acumulada de herbáceas nas classes de profundidade das
30 isolinhas agrupadas a cada 9 cm de profundidade para os três períodos, Pantanal de
Poconé-MT. Classe 1 (0 a 9 cm), Classe 2 (9 a 18cm), Classe 3 (18 a 27 cm), Classe 4 (27 a 36 cm), Classe 5
(36 a 45 cm) e Classe 6 (>45 cm).
Efeito do gradiente de inundação na diversidade beta– O gradiente de inundação tem efeitos
sobre a variação na composição das espécies, sendo os valores de βT mais elevados,
principalmente, entre as isolinhas não inundáveis e pouco inundáveis, com queda gradativa ao
longo do gradiente de inundação nos períodos de cheia e vazante, sendo as correlações de
Spearman negativas e significativas para estes períodos em relação a este gradiente, com rs=-
0,57 e rs=-0,58 (p = 0,05) (Fig. 9). De outra forma, observa-se também um efeito temporal
(entre os períodos) em relação ao comportamento dos pares de isolinhas comparados, onde o
decréscimo de βT se acentua no sentido seca / cheia com posterior tendência ao retorno
durante a vazante. Portanto, a substiuição de espécies progressivamente diminui muito
durante o período de cheia ao longo do gradiente, devido às condições ambientais mais
semelhantes entre os pares de isolinhas comparadas. Durante o período de seca é possível que
o contraste das condições ambientais no gradiente, ou seja, da umidade, seja mais marcante,
restringindo espécies a determinados sítios neste gradiente.

87
0 4 8 12 16
Pares adjacentes
0.0
0.5
1.0
1.5
2.0
2.5
Bt
VazanteCheiaSeca
Períodos
Figura 9 – Diversidade Beta (βT) comparando pares de isolinhas adjacentes ordenadas pelo
gradiente de duração da inundação, Pantanal de Poconé-MT.
Efeito do gradiente de inundação na cobertura das espécies: Para as todas as 6 classes de
profundidade foram eleitas as cinco espécies com os maiores valores de cobertura para cada
período. Em praticamente todas estas classes há flutuações nas coberturas das espécies entre
os períodos, entretanto, de forma menos pronunciada para a maioria das espécies da classe 1,
que reúne as isolinhas não inundáveis e pouco inundáveis. A partir da classe 2 as variações na
cobertura entre os três períodos se intensificam para as espécies. Por exemplo, espécies como
Eleocharis minima apresentou forte variação na cobertura entre os períodos nas classes 3, 4 e
5, com elevada cobertura durante a cheia. Axonopus purpusii tem forte queda na cheia e na
vazante na classe 5, ocorrendo com baixa cobertura na classe 6, onde provavelmente não
suporta a forte variação hídrica. Este padrão também ocorre com Paspalum lineare. Já
Utricularia foliosa, que não ocorre na seca, tem cobertura elevada durante a cheia a partir da
classe 3 e se mantém apenas na última classe durante a vazante (Fig. 10). Na última classe a
instabilidade na cobertura das espécies se torna mais intensa e espécies estritamente aquáticas,
dependentes de lâmina de água, se manifestam em maior número com elevada cobertura
como Cabomba furcata, Utricularia foliosa e Mayaca sellowiana. No dendrograma da figura
11 observa-se a formação de basicamente três grupos, com dois extremos. Um grupo extremo
formado pela classe 1 e outro formado pela classe 6, sendo o restante das classes um grupo
(rs= -0,054)
(rs= -0,57)
(rs= -0,58)

88
intermediário. A elevada dissimilaridade, em relação à composição e cobertura, entre a última
classe e as demais classes de profundidade, assim como, a elevada variabilidade entre os
períodos de seca e cheia e seca e vazante também pode ser observada.

89
Figura 10 – Variação no valor de cobertura nas classes de profundidade nos períodos de
seca, cheia e vazante, Pantanal de Poconé-MT. Classe 1 (0 a 9 cm), Classe 2 (9 a 18cm), Classe
3 (18 a 27 cm), Classe 4 (27 a 36 cm), Classe 5 (36 a 45 cm) e Classe 6 (>45 cm).

90
Análise de Agrupamento
Distance (Objective Function)
Information Remaining (%)
3.6E-02
100
7.8E-01
75
1.5E+00
50
2.3E+00
25
3E+00
0
CL1c
CL1s
CL1v
CL2c
CL2v
CL2s
CL3s
CL3v
CL5s
CL4s
CL3c
CL5c
CL4c
CL4v
CL5v
CL6c
CL6v
CL6s
Figura 11 – Análise de agrupamento representada por um dendrograma comparando as
6 classes de profundidade em diferentes períodos. Classe 1 (0 a 9 cm), Classe 2 (9 a 18cm),
Classe 3 (18 a 27 cm), Classe 4 (27 a 36 cm), Classe 5 (36 a 45 cm) e Classe 6 (>45 cm).
Efeito do gradiente de inundação sobre as formas de vida: Diferentes parâmetros
quanto à forma de vida, como riqueza, cobertura e porcentagem de espécies por classe
de profundidade apresentaram variações ao longo das classes de profundidade nos três
períodos. Entretanto, apenas as correlações com riqueza em hemicriptófitas foram
significativas, diminuindo significativamente com a profundidade (seca rs = -0.886,
cheia rs = -0.942, e para a vazante rs = -0.886), assim como para geófitas durante o
período de seca (seca rs = -0.883) (Fig. 12 A, B e C). Com relação à cobertura e
porcentagem de espécies por forma de vida em cada classe de profundidade, não houve
variações significativas com a profundidade e período (p > 0.10), contudo, algumas
tendências foram importantes. As geófitas tiveram tendência de aumento em cobertura
ao longo do gradiente de profundidade durante o período de cheia (rs = 0,7714), como
também as terófitas para os períodos de cheia (rs = 0,714) e vazante (rs = 0,828) (Fig.
13 A, B e C).
Entre as principais espécies responsáveis pelo aumento na cobertura de geófitas
na cheia, que em conjunto tendem a um aumento no gradiente de inundação, estão
Eleocharis acutangula, Sagittaria guyanensis e Utricularia foliosa. Estas espécies
apresentam formas biológicas emergente, flutuante fixa e submersa livre,
respectivamente. Entre as terófitas estão Cabomba furcata e Eleocharis minima, com as

91
respectivas formas biológicas, submersa fixa e anfíbia e/ou flutuante fixa. Na vazante as
terófitas ainda possuem uma elevada cobertura, com tendência de aumento no gradiente
de inundação, onde espécies como Cabomba furcata e Eleocharis minima contribuem
para isto.

92
A
B
C Figura 12 – Número de espécies por forma de vida nas classes de profundidades nos
períodos de seca, cheia e vazante, Pantanal de Poconé-MT. Classe 1 (0 a 9 cm), Classe 2 (9 a
18cm), Classe 3 (18 a 27 cm), Classe 4 (27 a 36 cm), Classe 5 (36 a 45 cm) e Classe 6 (>45 cm).
0 1 2 3 4 5 6 7
Classes
0
10
20
30
40
50
Núm
ero
de e
spécie
s
GeoHemiTero
Vazante
0 1 2 3 4 5 6 7
Classe
0
10
20
30
40
50
Núm
ero
de
espé
cie
s
GeoHemiTero
Cheia
0 1 2 3 4 5 6 7
Classes
0
10
20
30
40
50
Núm
ero
de
espé
cie
s
GeoHemiTero
Seca

93
A
B
C Figura 13 – Valor de cobertura por forma de vida nas classes de profundidades nos
períodos de seca, cheia e vazante, Pantanal de Poconé-MT. Classe 1 (0 a 9 cm), Classe 2 (9 a
18cm), Classe 3 (18 a 27 cm), Classe 4 (27 a 36 cm), Classe 5 (36 a 45 cm) e Classe 6 (>45 cm).
0 1 2 3 4 5 6 7
Classes de profundidade
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Va
lor
de
co
be
r tura
GeoHemiTero
Vazante
0 1 2 3 4 5 6 7
Classes de profundidade
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Valo
r d
e c
obe
rtura
GeoHemiTero
Cheia
0 1 2 3 4 5 6 7
Classes de profundidade
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Valo
r d
e c
ob
ert
ura
GeoHemiTero
Seca

94
DISCUSSÃO
As espécies identificadas neste levantamento correspondem a 18,2% do total de
espécies herbáceas conhecidas para o Pantanal como um todo. Os estimadores Jackknife
1 e 2 indicam que se podería chegar a 23,39% e 26,40% das espécies, respectivamente
Na listagem da flora do Pantanal (Pott & Pott 1999), que inclui todos os tipos de hábitos
vegetais, Leguminosae é a famílias mais numerosa (240 espécies), seguida de Poaceae
(212 espécies), Cyperaceae (92 espécies), Asteraceae (87 espécies), Euphorbiaceae (78
espécies) e Rubiaceae (62 espécies). Considerando apenas as espécies de hábito
herbáceo e terrestres, as famílias mais ricas em espécies são Poaceae, Cyperaceae,
Leguminosae e Asteraceae (Pott & Pott 1999; Junk et al. 2006b). Um levantamento de
Schessl (1999) realizado em diferentes fitofisionomias ao longo da rodovia
transpantaneira no Pantanal de Poconé coloca as famílias Poaceae (104 espécies),
Leguminosae (57 espécies), Cyperaceae (56 espécies), além das famílias Rubiaceae (29
espécies), Asteraceae (25 espécies) e Euphorbiaceae (21 espécies) como as mais ricas.
Segundo este autor, as principais famílias que compõem diferentes áreas inundáveis da
América do Sul são Poaceae, Leguminosae e Cyperaceae. Portanto, a riqueza por
família da flora em geral do Pantanal e estudo de Schessl (1999) na região Poconé é
semelhante a este estudo, não seguindo a mesma ordem devido a não inclusão de
espécies de hábito arbóreo, o que aumentaria a contribuição de espécies de
Leguminosae. Nenhuma espécie foi acrescentada à listagem da flora do Pantanal. As
famílias Poaceae e Cyperaceae estão entre as mais ricas em macrófitas aquáticas (Pott &
Pott 1997), tendo um bom incremento de espécies nos períodos de cheia e vazante.
Algumas famílias são características de ambientes inundáveis e só ocorreram durante a
cheia e/ou vazante (somente nos sítios com lâmina de água), como Cabombaceae,
Lentibulariaceae, Limnocharitaceae, Mayacaceae e Nymphaeaceae, aumentado a
riqueza e diversidade em espécies destes períodos. A família Poaceae é a melhor
representada em termos de cobertura regional.
Algumas espécies são consideradas subespontâneas, ou seja, foram introduzidas
acidental ou propositadamente na região, com posterior propagação sem qualquer
intervenção do homem. Pott & Pott (1999) e Schneider (2007) citam como espécies
subespontâneas Alysicarpus vaginalis, Commelina benghalensis, Digitaria ciliaris,
Hyptis suaveolens, Paspalum multicaule, Solanum viarum, Murdannia nudiflora,
Cyperus esculentus, Cyperus iria, Fimbristylis miliacea, Euphorbia hyssopifolia,

95
Leptochloa virgata e Centunculus minimus. Este conjunto de espécies, entretanto, é raro
e ocorre com baixa cobertura. Já Brachiaria humidicola, intencionalmente introduzida,
tem elevada cobertura na região, pois é cultivada para formação de extensas pastagens
nesta região do Pantanal de Poconé por ser adaptada a ambientes inundáveis. Nesta área
de estudo também foi registrada Habranthus pantanalensis (Amaryllidaceae), uma das
espécies citadas por Pott & Pott (2000) como endêmicas do Pantanal. Por ter sido
coletada de forma aleatória, não foi incluída na listagem.
Neste trabalho foi possível verificar que na escala regional o pulso de inundação
influencia a riqueza, a diversidade, a composição, a cobertura e as formas de vida das
herbáceas no Pantanal, promovendo aumento na riqueza em espécies durante os
períodos de cheia e vazante. A maior similaridade e riqueza em espécies entre cheia e
vazante, assim como, variações na cobertura de espécies entre períodos também foi
verificada por Prado et al. (1994) em trabalho realizado em um gradiente de inundação
nos diferentes períodos hidrológicos do Pantanal de Poconé. Segundo Brook et al.
(2003) a sazonalidade hídrica é considerada importante na manutenção da diversidade
em comunidades de ambientes temporariamente inundáveis.
Diferentes adaptações e estratégias das espécies frente às mudanças hidrológicas
determinam um grupo de espécies capaz de se manter apenas em determinados períodos
e outro que se mantém durante todo o ano, mesmo nos ambientes sujeitos ao pulso de
inundação. Sendo 78% das espécies de ervas e gramíneas do Pantanal consideradas
terrestres, a ocorrência de microsítios e, principalmente, a plasticidade morfológicas e
fisiológica (Junk et al. 2006b) são fatores determinantes na manutenção destas espécies
ao longo de todo o ano. Uma das conseqüências da inundação é a diminuição da difusão
de oxigênio no solo, sujeitanto as raízes das plantas (órgão aeróbio) a condições de
hipoxia (pouco oxigênio) ou anoxia (ausência de oxigênio), que de forma indireta
também influencia os ramos em função da perda de funções das raízes das quais
dependem (Vartapetian & Jackson 1996). Assim, muitas espécies toleram a inundação
em função de adaptações morfológicas e anatômicas que representam estratégias para se
evitar o déficit de oxigênio, como por exemplo: a formação de aerênquima em raízes,
caule e/ou folhas e de raízes adventícias (Armstrog et al. 1994 apud Dias-Filho 2005).
Além disso, a habilidade das plantas em tolerar as condições de hipoxia ou anoxia
também está relacionada a ajustes das funções fisiológicas com modificações na taxa
fotossintética, condutância estomática, taxa de transpiração, taxa de renovação foliar,
entre outras (Chapin III et al. 1993). Há, portanto, há uma diversidade de respostas e

96
uma multiplicidade de estratégias envolvidas que asseguram a manutenção das plantas
em ambientes inundáveis (Crawford 1996). As espécies tolerantes a inundação devem
ter habilidades para resistir à inundação, assim como para retornar às condições de não
inundação, já que pode haver desvantagens de determinadas adaptações morfológicas à
inundação quando a lâmina de água decai (Crawford 1996). Por exemplo, o
engrossamento das raízes, devido a formação do aerênquima como resposta a
inundação, pode limitar a absorção de água e nutrientes quando a planta retorna às
condições de seca (Koncalová 1990 apud Crawford 1996). Entretanto, tendo o Pantanal
duas fases de forte estresse hídrico, ou pelo excesso ou pela falta, tais espécies perenes
além de tolerância à inundação devem possuir mecanismos de tolerância à dessecação
durante o período da seca no Pantanal.
O incremento de espécies entre os períodos ocorreu tanto nas isolinhas
inundáveis, quanto nos não inundáveis, indicando ciclos de vida altamente dependentes
da sazonalidade hídrica em ambos ambientes. Este incremento de espécies nos períodos
de cheia e vazante sugere elevada importância de diásporos viáveis no banco de
sementes, além de propágulos vegetativos, e foi particularmente importante nas
isolinhas em campos exóticos (inundáveis), extremamente pobres em espécies no
período de seca. Em relação ao banco de sementes Broke et al. (2003) afirmam que em
ambientes temporariamente inundáveis as plantas aquáticas depositam um grande
número de sementes, e que estas se mantêm dormentes e viáveis por um longo período
sem inundação, mantendo a resiliência destes ambientes. Nos experimentos realizados
por Broke et al. (2003), este banco de sementes não se exauriu após várias inundações.
Junk et al. (2006b) afirmam que há um rico banco de sementes no Pantanal e que a
ativação de somente parte do banco de sementes durante a fase úmida aumenta a
resiliência das comunidades herbáceas contra padrões irregulares de precipitação ou
períodos imprevisíveis de curta inundação, que pode levar muitas plântulas a morte.
Assim, os bancos de sementes e de propágulos vegetativos são importantes na
manutenção da resiliência da comunidade aquática durante o período de seca e vice-
versa. Este banco de sementes possui papel fundamental na recuperação de áreas
úmidas onde o regime hídrico fora alterado, devido a sua riqueza e longa viabilidade
(Middleton 2003).
De acordo com Elmqvist et al. (2003), a diversidade de espécies disponíveis e
suas diferentes estratégias são essenciais para a renovação e reorganização cíclica dos
ambientes sujeitos a distúrbios, como o pulso de inundação deve ser considerado.

97
Apesar do incremento de espécies entre os períodos, todos eles apresentaram um
padrão de diminuição da riqueza específica com o aumento da profundidade da
inundação, como também relatada por outros autores para o Pantanal, como Prado et
al.(1994), Pinder & Rosso (1998) e Fortney et al. (2004), que consideram apenas
espécies herbáceas e por Pott & Adámoli (1999), Adámoli & Pott (1999) e Nunes da
Cunha & Junk (2001), que consideram herbáceas e/ou lenhosas. Segundo Robertson et
al. (1984) e Mitsch & Gosselink (1986), de uma forma geral, ambientes
permanentemente inundados ou sujeitos a longos períodos de inundação têm baixa
diversidade, em virtude do menor conteúdo de oxigênio dos solos, além de outras
condições químicas, que limitam significativamente o número e os tipos de plantas que
podem sobreviver a tais condições. O aumento da riqueza e da variação da riqueza com
a profundidade, da estação seca para a vazante, sugere que a seca é o período que impõe
mais restrição à ocorrência de várias espécies e que espacialmente pequenas diferenças
na profundidade ou tempo de inundação promovem acentuada substituição de espécies,
pois, possivelmente, separa o conjunto de espécies preferencialmente de hábitats
terrestres e outro de espécies com tolerância e/ou adaptabilidade aos ambientes
inundáveis. Posteriormente, com a queda da riqueza em espécies e maior similaridade
entre os pares de isolinhas as quedas na diversidade beta são acentuadas. Como as
espécies dos ambientes inundáveis respondem de forma mais dinâmica aos ciclos de
seca e cheia, há uma maior influência do gradiente e substituição de espécies durante o
período de seca, pois as condições tornam-se mais contrastantes no gradiente, o que
mantém e restringe as espécies a determinados porções do gradiente.
Outro ponto importante foi que o efeito da sazonalidade hídrica sobre a
composição e cobertura das espécies se intensifica ao longo do gradiente de inundação e
também determina diferentes comunidades herbáceas em função do aumento desta
intensidade (profundidade e duração) de inundação. Schessl (1999) observou que nos
ambiente mais inundáveis do Pantanal de Poconé, há uma forte variação na composição
e cobertura de espécies promovida pela sazonalidade hídrica. Neste trabalho, isso
também pôde ser verificado, pois na classe 6 de profundidade a comunidade foi bastante
diferenciada entre as demais classes e com elevada dissimilaridade entre os períodos de
seca e cheia/vazante.
Portanto, pode-se dizer que com o aumento da intensidade da inundação a
estabilidade do ambiente tende a diminuir com acentuadas mudanças na composição e
cobertura, impondo maiores restrições às espécies, diminuindo a riqueza e selecionando

98
um grupo diferenciado de espécies, com adaptações à elevada profundidade e duração
da inundação. A resiliência, entretanto, é mantida produzindo uma mudança cíclica
sazonal nas comunidades herbáceas.
Neste estudo as espécies hemicriptófitas foram melhor representadas, seguidas
pelas terófitas e geófitas. Segundo Raunkiaer (1934) tais formas de vida indicam
adaptações a ambientes climaticamente desfavoráveis em que as gemas apresentam grau
de proteção superior às demais. Esses resultados são condizentes com os estudos Pott &
Adámoli (1999) para o Pantanal de Paiaguás, que também abrangeu diferentes tipos de
vegetação, entre áreas inundáveis e não inundáveis e florestadas e campestres, como
também, com o estudo de Schessl (1999), realizado na mesma sub-região que este
estudo, também incluindo diferentes tipos de vegetação, entretanto, com proporção
maior de terófitas. O que há de comum entre este estudo e os demais trabalhos é uma
maior representação de terófitas quando comparados ao Cerrado (Silva e Batalha 2008;
Meira-Neto et al. 2007; Batalha e Martins 2004; Costa et al. 2004; Batalha e Mantovani
2001; Warming 1892 apud Sarmiento e Monasterio 1983). Calculando a porcentagem
de formas de vida basicamente de espécies herbáceas, Pott e Adámoli (1999)
encontraram para o Pantanal de Paiaguás melhor representatividade das hemicriptófitas
com 57,41% das espécies, terófitas 21,53%, geófitas 10,05%, caméfitas 7,65%,
fanerófitas 1,43% (que inclui alguns subarbustos) e outros 1,91%. Schessl (1999)
encontrou para o Pantanal de Poconé uma porcentagem de hemicriptófitas (incluindo
lianas herbáceas) maior do que terófitas, entretanto de forma mais equitativa, com
35,34% e 33,53%, respectivamente, seguidas das geófitas 13,25%, caméfitas 9,84% e
outras 8,03%.
Assim, baseado neste e estes outros estudos realizados no Pantanal o espectro
para herbáceas pode variar da seguinte forma: as hemicriptófitas entre,
aproximadamente, 35 a 57%, terófitas ente 21 a 34%, geófitas entre 10 a 13% e
caméfitas entre 5 a 10% (Pott e Adámoli 1999; Schessl 1999; e este estudo).
Posteriormente, com os dados de espécies lenhosas, também coletados no mesmo
desenho amostral, será possível calcular o espectro mais completo. As proporções de
terófitas dos trabalhos realizados no Pantanal, portanto, são mais semelhantes às de
outras savanas Sul Americanas, que incluem savanas denominadas hipersazonais
(Sarmiento e Monasterio 1983; Batalha e Martins 2002), ou seja, savanas que
frequentemente ocorrem em áreas baixas ou outros sítios com problemas de drenagem e
que alternam anualmente períodos de escassez ou excesso de água (Sarmiento 1992).

99
No trabalho de Batalha e Martins (2002), que não inclui o Pantanal, o Cerrado não de se
agrupa aos demais sítios de savanas justamente pela ausência de terófitas. Períodos
desfavoráveis nestas savanas Sul Americanas incluem a seca e a cheia, fato que as
aproximam do Pantanal e, provavelmente, seleciona um contingente superior de
espécies terófitas, uma vez que neste estudo as terófitas obtiveram incremento durante o
ciclo hídrico. Apesar das terófitas estarem mais relacionadas a ambientes quentes e
secos (Batalha e Martins 2002), os distúrbios são citados como beneficiadores de
espécies anuais (Grime 1977; Belsky 1992), e no caso do Pantanal, bem como outras
savanas hipersazonais, a inundação pode ser considerada um distúrbio (White & Pickett
1985), sendo este fenômeno pouco presente no Cerrado.
Alguns trabalhos têm demonstrado que formas de vida respondem a gradientes
ambientais (Danin e Orshan 1990; Montalvo et al. 1991), como também, a distúrbios
(MacIntyre et al. 1995). De fato, as terófitas chegaram a representar 41% e as
hemicriptófitas 23% das espécies em um campo sujeito a inundação prolongada no
Pantanal de Poconé (Rebellato e Nunes da Cunha 2005). Schessl (1999) também
encontrou uma participação maior de terófitas em termos de cobertura para ambientes
com inundações mais pronunciadas. Já nos llanos Venezuelanos, por exemplo, a
importância das espécies terófitas varia com o tipo de habitat, podendo estas serem
comuns em ambientes secos, como dunas arenosas e afloramentos lateríticos, ou mesmo
em ambientes instáveis, com flutuações hídricas em áreas úmidas (Sarmiento e
Monasterio 1983), ou seja, extremos quanto ao grau de estresse hídrico.
Por ser o Pantanal dinâmico, há mudanças na participação das formas de vida
em relação aos diferentes períodos hidrológicos onde as gemas dos propágulos
vegetativos e/ou bancos de sementes das plantas estão mais ou menos ativos, ou seja,
vegetativamente visíveis no ambiente. Determinadas formas de vida, como as geófitas e
terófitas, foram bastante vinculadas ao período de cheia e vazante em termos de riqueza
e cobertura regional. E em relação ao pulso de inundação estas duas formas de vida
apresentam um maior número de espécies não só com adaptações à inundação, mas
também, com habilidades em crescer em ambientes mais profundos e, portanto,
tendência de aumento de cobertura nestes ambientes. Capon (2003) observou uma forte
colonização e elevada cobertura de gramíneas anuais e hidrófitas durante a inundação
promovida pelo banco de sementes, sendo significativamente maior a cobertura destas
nos ambientes com maior intensidade de inundação. Essas formas de vida, portanto,
caracterizam bem a flora aquática do Pantanal, que durante a cheia e vazante são

100
responsáveis pelo incremento de espécies confirmando a importância do banco de
semente e de propágulos vegetativos. Muitas das espécies terófitas e geófitas, portanto,
possuem estratégias de fuga à seca em função de se manterem no ambiente durante este
período na forma de sementes (terófitas) e ou rizomas, bulbos e tubérculos (geófitas).
As reservas de carboidratos nos órgãos subterrâneos das geófitas permitem que estas se
mantenham no ambiente durante todo o período de seca, com atividade e
desenvolvimento de partes aéreas durante o período de chuvoso, quando então,
florescem e frutificam ao longo da enchente e cheia (Larcher 1995). Uma grande
participação de terófitas exclusivas da vazante pode estar relacionada ao retorno de
melhores condições de oxigênio no solo, requerido pela maioria das espécies na geração
de energia para o processo de germinação (Lobo & Joly 1998).
Apesar de boa parte das hemicriptófitas serem duradouras e com maior
estabilidade de cobertura frente ao ciclo de cheia e seca, estas possuem quedas na
cobertura e riqueza no gradiente de inundação. Ou seja, ao longo do gradiente de
inundação a manutenção da cobertura de espécies perenes durante todo o período
hídrico é diminuída, o que ajuda a explicar as maiores variações em termos de
composição e cobertura em ambientes mais inundáveis. As gramíneas hemicriptófitas
com elevada cobertura regional durante os três períodos, Axonopus purpusii e Paspalum
lineare, por exemplo, são extremante limitadas na última classe de profundidade.
Entretanto, algumas macrófitas aquáticas são hemicriptófitas, exclusivas da cheia e
vazante, e possuem elevada cobertura em ambientes mais profundos, revelando que não
apenas a forma de vida, mas também a forma biológica e outras adaptações são
essenciais na seleção de espécies a ambientes com maiores profundidade. Segundo
Finlayson (2005), dentro da categorização de formas de vida e de crescimento há muitas
adaptações morfológicas e fisiológicas aos ambientes inundáveis, sendo a duração da
inundação o principal fator no estabelecimento da composição de espécies.
O fato da dependência de muitas espécies, com suas diferentes estratégias,
estarem relacionadas ao ciclo hídrico sazonal, reforça questões referentes a importância
da manutenção do regime hidrológico, como também, a importância de toda a sua
variabilidade espacial hídrica na planície. Ou seja, a conservação de diferentes tipos de
comunidades e diversidade de espécies requer a manutenção da variedade dos padrões
hidrológicos através da paisagem (Brook et al. 2003).

101
CONCLUSÕES
• A sazonalidade hídrica anual influencia regionalmente a riqueza, a diversidade, a
composição, a cobertura de espécies e formas de vida, entretanto, mantendo as
mesmas famílias dominantes;
• A riqueza e a diversidade alfa variaram entre os períodos, sendo significativamente
mais elevadas durante os períodos de cheia e vazante em relação ao período de seca;
• Poucas espécies possuem elevada cobertura regional mantida durante os três
períodos, entre elas, Axonopus purpusii, Brachiaria humidicola e Paspalum lineare;
durante a cheia Eleocharis minima tem a mais elevada cobertura regional;
• Hemicriptófitas, terófitas e geófitas são as formas de vida dominantes;
• Muitas espécies hemicriptófitas permanecem relativamente estáveis frente às
mudanças hídricas sazonais, sendo estas compostas principalmente por gramíneas;
• Durante o período de cheia e vazante há uma maior participação na riqueza e
cobertura de terófitas e geófitas;
• O gradiente de inundação tem efeitos negativos e significativos sobre a riqueza em
espécies nos períodos de seca, cheia e vazante e, portanto, a hipótese de elevada
riqueza em níveis intermediários de inundação não foi sustentada;
• A substituição de espécies é elevada na transição entre os ambientes não inundáveis
e pouco inundáveis, tendendo a diminuir ao longo do gradiente de inundação,
principalmente, durante os períodos de cheia e vazante devido a queda na riqueza e
maior similaridade entre os pares de isolinhas comparadas;
• Há uma forte variação na cobertura das espécies nos ambientes inundáveis em
relação aos não inundáveis, de forma mais pronunciada nas classes de maior
profundidade;
• As hemicriptófitas são dominantes ao longo de todo o gradiente de inundação, com
quedas significativas na riqueza neste gradiente em todos os períodos, tendo geófitas
queda significativa na riqueza apenas durante o período de seca;
• Terófitas e geófitas tiveram tendências ao aumento de cobertura ao longo do
gradiente de inundação, entretanto, hemicriptófitas são sempre dominantes;
• Muitas espécies são dependentes do ciclo hídrico sazonal, reforçando as questões
referentes a importância da manutenção do regime hidrológico, como também, de
toda a sua variabilidade espacial na planície;

102
REFERÊNCIAS CITADAS
Adámoli, J. 1987. Fisiografia do Pantanal. Pp. 15-22. In: Recursos forrageiros nativos
do Pantanal Mato-Grossense. Embrapa/Cernagem, Brasília-DF. Documento 8. 339p.
Adámoli, J. & Pott, A. 1999. A. Estudo fitossociológico e ecológico do Pantanal de
Paiaguás. Pp.215-225. In: Anais do II Simpósio sobre Recursos Naturais e Sócio-
econômicos do Pantanal, Manejo e Conservação. Empresa Brasileira de Pesquisa
Agropecuária (Embrapa-CPAP), Corumbá. 537p.
Alvarenga, S. M. M., Brasil, A. E., Pinheiro, R. e Kux, H. J. H. 1984. Pp 90-183.
Estudo geomorfológico aplicado a Bacia do Alto Paraguai e Pantanais Mato-grossenses.
Boletim Técnico do Projeto RADAMBRASIL (Série Geomorfologia,1). 187p.
Amaral Filho, Z. P. 1986. Solos do Pantanal Mato-Grossense. Pp 91-104. In: Simpósio
sobre recursos naturais e sócio-econômico do pantanal. Empresa Brasileira de Pesquisa
Agropecuária (Embrapa-CPAP), Brasília. 265 p.
Batalha, M. A. and Mantovani, W. 2001. Floristic composition of the cerrado in the Pé-
de-Gigante Reserve (Santa Rita do Passa Quatro, southeastern Brazil). Acta botanica
brasilica 15(3):289-304.
Batalha, M. A. and Martins, F. R. 2002. The vascular flora of Cerrado in Emas National
Park (Goiás, Central Brazil). Sida 20(1):295-311.
Batalha, M. A.; Martins, F. R. 2004. Floristic, frequency, and vegetation life-form
spectra of a cerrado site. Brazilian Journal of Biology 64(2): 201-209.
Belsky, A.J., 1992. Effects of grazing, competition, disturbance and fire on species
composition and diversity in grassland communities. Journal of Vegetation Science 3:
187-200.

103
Brock, M. A., Nielsen, D. L., Shiel, R. J, Green, J. D. and Langley, J. D. 2003. Drought
and aquatic community resilience: the role of eggs and seeds in sediments of temporary
wetlands. Freshwater Biology 48:1207–1218.
Browm, K. Jr. 1986. Zoogeografia da região do Pantanal Matogrossense. Pp.137-178.
In: Anais do I Simpósio sobre Recursos Naturais e Sócio-econômicos do Pantanal.
Empresa Brasileira de Pesquisa Agropecuária (Embrapa-CPAP), Brasília.
Bullock, J. 1996. Plants. In: Ecological census techniques. W.J. Sutherland (ed). Pp.
111-138. Cambridge University Press, Cambridge. 336p.
Capon, S. J. 2003. Plant community responses to wetting and drying in a large arid
floodplain. River research and applications 19:509-520.
Casanova, M. T. and Brock, M. A. 2000. How do depth, duration and frequency of
flooding influence the establishment of wetland plant communities? Plant Ecology 147
(2):237 250.
Chapin III, F. S.; Autumn, K.; Pugnaire, F. 1993. Evolution of suites of traits in response to
environmental stress. The American Naturalist 142(supplement):78-92.
Colwell, R. K. 2005. EstimateS. Statistical estimation of species and shared species
from samples. Version 7.5. Persistent URL<purl.oclc.org/estimates>.
Connell, J. H. 1978. Diversity in tropical rainforest and coral reefs. Science 199:1302-
1310.
Costa, I. R., Araújo, F. S. e Lima-Verde, L. W. 2004. Flora e aspectos auto-ecológicos
de um encrave de cerrado na chapada do Araripe, Nordeste do Brasil. Acta botanica
brasilica 18(4): 759-770
Danin, A. and Orshan, G. 1990. The distribution of Raunkiaer life forms in Israel in
relation to the environment. Journal of Vegetation Science 1: 41-48.

104
Dias-Filho, M. B. 2005. Opções forrageiras para áreas sujeitas a inundação ou
alagamento temporário. Pp.71-93. In: 22o Simpósio sobre manejo de pastagem. Teoria e
prática da produção animal em pastagens. Pedreira, C. G. S.; Moura, J. C. de; Da Silva
S. C.; Faria, V. P. (Eds.). Piracicaba: FEALQ.
Elmqvist, T., Folke, C., Nyström, M., Peterson, G., Bengtsson, J., Walker, B. and
Norberg, J. 2003. Response diversity, ecosystem change, and resilience. Frontiers in
Ecology and Environment 1(9): 488–494
Fantim–Cruz, I. 2008. Dinâmica da Inundação em Meso-Escala na Planície do Rio
Cuiabá, Pantanal, Brasil. Dissertação de Mestrado apresentada ao Programa de Pós-
graduação em Ecologia e Conservação da Biodiversidade - Universidade Federal de
Mato Grosso - Instituto de ciências biológicas, Cuiabá.
Finlayson, C. M. 2005. Plant ecology of Australia’s tropical floodplain wetlands: A
Review. Annals of Botany 96:541–555.
Fortney, R. F., Benedict, M., Gottgens, J. F., Walters, T. L., Leady, B. S. and Rentch, J.
2004. Aquatic plants community composition and distribution along an inundation
gradient at two ecologically-distinct sites in the Pantanal region of Brasil. Wetlands
Ecology and Management 12:575-585.
Franco, M. S. M. e Pinheiro, R. 1982. Geomorfologia. Pp.161-224. In: Brasil.
Ministério das Minas e Energia. Departamento Nacional da Produção Mineral. Projeto
Radam-brasil. Folha SE.21 Corumbá. Rio de Janeiro.
Grime, J. P. 1977. Evidence for the existence of three primary strategies in plants and its
relevance to ecological and evolutionary theory. American Naturalist 111:1169-1195.
Hamilton, S. K., Sippel, S. J., Melack, J. M. 1996. Inundation patterns in the Pantanal
wetland of South America determined form passive microwave remote sensing.
Archives of Hydrobiology 137:1-23.

105
Heckman, C. W. 1997. Description of a dinamic ecotone in the Pantanal of Mato
Grosso, Brazil. Ecotropica 3:1-7.
Junk, W.J. 2005. Flood pulsing and linkages between terrestrial, aquatic and wetland
system. Verh. Internat. Verein. Limnol 29: 11-38.
Junk, W. J., Bayley, P. B. & Sparks, R. S. 1989. The flood pulse concept in river –
floodplain systems. In: Proc Int Large River Symp (LARS). D. P. Dodge (ed). Canadian
Journal of Fishers and Aquatic 106: 110-127.
Junk, W.J. and Piedade, M.T.F. 1993. Herbaceous plants of the Amazon floodplain near
Manaus: Species diversity and adaptations to the flood pulse. Amazoniana
XII(3/4):467-484.
Junk, W. J e Da Silva, C.J. 1999. O conceito do pulso de inundação e suas implicações
para o Pantanal de Mato Grosso. Pp.17-28. In: Anais do II Simpósio sobre Recursos
Naturais e Sócio-econômicos do Pantanal, Manejo e Conservação. Empresa Brasileira de
Pesquisa Agropecuária (Embrapa-CPAP), Corumbá. 537p.
Junk, W.J., Brown, M., Campbell, I. C., Finlayson, M, Gopal, B, Ramberg, L. and
Warner, B. G. 2006a. The comparative biodiversity of seven globally important
wetlands: a synthesis. Aquatic Science 68: 400–414.
Junk, W. J., Nunes da Cunha, C. N., Wantzen, K. M., Petermann, P., strüssmann, C.,
marques, M. I. & Adis, J. 2006b. Biodiversity and its conservation in the Pantanal of
Mato Grosso, Brazil. Aquatic Science 68:278–309.
Kent, M. & Coker, P. 1994. Vegetation description and analysis. A practical approach.
Chichester, UK. John Willey. 363p.
Köppen, G. W. 1948. Climatologia. Fundo de Cultura Econômica. México. Pp. 242-
248.

106
Kovach, W.L. 2007. Multivariate statistical package, version 3.13 for windows, Kovach
Computing Services 85 Nant-y-Felin Petrueth, Isle of Anlesey, LL75 8uy Wales, UK.
Larcher, W. (1995) “Physiological Plant Ecology”. Springer, Berlin.
Lenssen, J., Menting, F., van der Putten, W. and Blom , K. 1999. Control of plant
species richness and zonation of functional groups along a freshwater flooding gradient.
Oikos 86(3):523-534.
Lobo, P. C. & Joly, C. A. 1998. Tolerance to hypoxia and anoxia in neotropical tree
species. Pp.137-156. In: Ecophysiological strategies of xerophytic and amphious plants
in the neotropics. F. R. Scarano; A. C. Franco (Eds.). Series Oecologia Brasiliensis,
Vol. IV. PPGE-UFRJ. Rio de Janeiro, Brazil.
Loureiro, R. L., Souza Lima, J. P., Fronzar, P. C. Vegetação. Pp. 329-372. In: Brasil:
Ministério das Minas e Energia. Departamento Nacional da Produção Mineral. Projeto
Radam-brasil. Folha SE.21 Corumbá. Rio de Janeiro. (Levantamento de Recursos
Naturais, 27).
Magnusson, W. E., Lima, A. P., Luizão, R., Luizão,F., Costa, F. R. C., Castilho, C. V
and Kinupp, V. F. 2005. Rapeld: A modification of the Gentry method for biodiversity
surveys in long-term ecological research sites. Biota Neotropica 5(2): 1-6.
Margurran, A.E. 1988. Ecological diversity and its measurement. Princeton University,
Princeton. 179p.
McCune, B. & Mefford, M.J. 1999. PC-ORD. Multivariate analysis of ecological data,
version 4.01. Oregon Glaneden Beach, MjM Software Desing.
MacIntyre, S., Lavorel, S. and Tremont, R. M. 1995. Plant life-history attributes: their
relationship to disturbance response in herbaceous vegetation. Journal of Ecology 83:
31-44.

107
Meira-Neto, J. A. A., Martins, F. R. e Valente, J. E. 2007. Composição florística e
espectro biológico na estação ecológica de santa bárbara, estado de São Paulo, Brasil.
Revista Árvore 31(5):907-922.
Middleton, B. A. 2003. Soil seed banks and the potential restoration of forested
wetlands after farming. Journal of Applied Ecology 40(6): 1025-1034.
Mitsch, W. J. and Gosselink, J. G. 1986. Wetlands. Van Nostrand Reinhold, New York,
USA. 539p.
Montalvo, J., Casado, M. A., Levassor, C. and Pineda, F. D. 1991. Adaptation of
ecological systems: compositional patterns of species and morphological and functional traits.
Journal of Vegetation Science 2: 655-666.
Nunes da Cunha, C. 1998. Comunidades arbustivo-arbóreas de Capões e Diques
marginais no Pantanal de Poconé-MT: Caracterização e análise de gradiente. Tese
apresentada ao Programa de Pós-graduação em Ecologia e Recursos Naturais,
Universidade de São Carlos (UFSCAR).
Nunes da Cunha, C. & Junk, W.J. 1999. Composição florística de capões e cordilheiras:
localização das espécies lenhosas quanto ao gradiente de inundação no Pantanal de
Poconé, MT-Brasil. Pp.387-405. In: Anais do II Simpósio sobre Recursos Naturais e
Sócio-econômicos do Pantanal, Manejo e Conservação. Empresa Brasileira de Pesquisa
Agropecuária (Embrapa-CPAP), Corumbá. 537p.
Nunes da Cunha, C. & Junk, W.J. 2001. Distribuition of wood plant communities along
the flood gradient in the Pantanal of Poconé, Mato Grosso, Brazil. International Journal
of Ecology and Environmental 27:63-70.
Nunes da Cunha, C. & Junk, W.J. 2004. Year-to-year changes in water level drive the
invasion of Vochysia divergens in Pantanal grasslands. Applied Vegetation Science
7:103-110.

108
Nunes da Cunha, C., Rawiel, C. P., Wantzen, K. M., Junk, W. J. & Lemes do Prado, A.
2006. Mapping and characterization of vegetation units by means of Landsat imagery
and management recommendations for the Pantanal of Mato Grosso (Brazil), north of
Poconé. Amazoniana XIX(1): 1-32.
Pinder, L. and Rosso, S. 1998. Classification and ordination of plant formations in the
Pantanal of Brazil. Plant Ecology 136: 151–165.
Pinho, J. B. 2005. Comunidades de aves, biologia reprodutiva e dieta alimentar de aves
de sub-bosque em quatro áreas fitofisionomias diferentes no Pantanal de Mato Grosso.
Tese de doutorado apresentada ao Programa de Pós-graduação em Ecologia,
Conservação e Manejo da Vida Silvestre. Universidade Federal de Minas Gerais
(UFMG), Instituto de biociência, Belo Horizonte.
Pollock, M. M., Naiman, R. J. and Hanley, T. A. 1998. Plant species richness in riparian
wetlands - A test of biodiversity theory. Ecology 79:94-105.
Pott, A. & Adámoli, J. 1999. Unidades de vegetação do Pantanal dos Paiaguás. Pp. 183-
202. In: Anais do II Simpósio sobre Recursos Naturais e Sócio-econômicos do Pantanal,
Manejo e Conservação. Empresa Brasileira de Pesquisa Agropecuária (Embrapa-
CPAP), Corumbá. 537p.
Pott, V.J. & Pott, A. 1997. Checklist das macrófitas aquáticas do Pantanal, Brasil. Acta
Botânica Brasilica 11(2): 215-227.
Pott, A. & Pott, V. J. 1999. Flora do Pantanal - Listagem Atual de Fanerógamas. Pp.
297-325. In: Simpósio sobre Recursos Naturais e Sócio-Econômicos do Pantanal:
Manejo e Conservação. Empresa Brasileira de Pesquisa Agropecuária (Embrapa-CPAP),
Corumbá. 537p.
Pott, V.J. & Pott, A. 2000. Plantas aquáticas do Pantanal. Empresa Brasileira de
Pesquisa Agropecuária (Embrapa-CPAP), Corumbá. 353p.

109
Pott, A. & Pott, V. J. 2009. Vegetação do Pantanal: fitogeografia e dinâmica. Pp.1065-
1076. In: Anais 2º Simpósio de Geotecnologias no Pantanal. Embrapa Informática
Agropecuária/INPE, Corumbá.
Prado, A. L., Heckman, C. W. and Martins, F. R. 1994. The seasonal succession of
biotic communities in wetlands of the tropical wet-and-dry climatic zone: II. The
Aquatic Macrophyte Vegetation in the Pantanal of Mato Grosso, Brazil. Internationale
Revue gesamten Hydrobiologie 79(4): 569-589.
Prance, G.T. and Schaller, G.B. 1982. Preliminary study of some vegetation types of the
Pantanal, Mato Grosso, Brazil. Brittonia 34: 228-251.
Raunkier, C. 1934. The life forms of plants and statistical plant geography. Clarendon
Press, Oxford. 200p.
Rebellato, L. e Nunes da Cunha, C. 2005. Efeito do “fluxo sazonal mínimo da
inundação” sobre a composição e estrutura de um campo inundável no Pantanal de
Poconé, MT, Brasil. Acta Botanica Brasilica 19: 789-799.
Robertson, P. A., Mackenzie, M. D. & Elliot, L. F. 1984. Gradient analysis and
classification of the woody vegetation for 4 sites in southern Illinois and adjacent
Missouri. Vegetatio 58: 87–104.
Sarmiento, G. and Monastério, M. 1983. Life forms and fenology. Pp.79-108. In:
Ecosystems of the Word: Tropical Savannas. F. Bourliére (ed.). Elsevier. Amsterdam.
730p.
Schessl, M. 1999. Floristic composition and structure of floodplain vegetation in
Northern Pantanal of Mato Grosso, Brasil. Phyton (Horn, Austria) 39(2):303-3.
Schneider, A. A. 2007. A flora naturalizada no estado do Rio Grande do Sul, Brasil:
herbáceas subespontâneas. Biociências, Porto Alegre, 15(2):257-268.

110
Silva, J. S. V. e Abdon, M. M. 1998. Delimitação do Pantanal brasileiro e suas sub-
regiões. Pesquisa agropecuária brasileira, Brasília-DF 33: 1703-1711.
Silva, I. A. & Batalha, M. A. 2008. Species convergence into life-forms in a
hyperseasonal cerrado in central Brazil. Brazilian Journal of Biology 68: 631-637.
Silva, M. P., Mauro, R., Mourão, G. E. & Coutinho, M. 2000. Distribuição e
quantificação de classes de vegetação do Pantanal através de levantamento aéreo.
Revista Brasileira de Botânica 23(2): 143-152.
Southal, E. J., Dale, M. P. and Kent, M. 2003. Spatial and temporal analysis of
vegetation mosaics for conservation: poor fen communities in a Cornish valley mire.
Journal of Biogeography 30:1427–1443.
Townsend, P. A. 2001. Relationships between vegetation patterns and hydroperiod on
the roanoke river floodplain, North Carolina. Plant Ecology 156(1): 43-58.
van der Valk, A. G. 1981. Succession in wetlands: a Gleasonian approach. Ecology
62(3): 689-696.
Vartapetian, B. B. and Jackson, M. B. 1997. Plant adaptations to anaerobic stress. _/
Annal of Botany 79 (Suppl. A): 3-20.
White, P. S. and Pickett, S. T. A. 1985. Natural disturbance and patch dynamics: An
introduction. Pp. 2-8. In: The ecology of natural disturbance and patch dynamics. S.T.A.
Pickett & P.S. White (eds). San Diego, Academic Press. 472p.
Zeilhofer, P. & Schessl, M. 1999. Relationship between vegetation and environmental
conditions in the northern Pantanal of Mato Grosso, Brazil. Journal of Biogeography
27: 59-168.

111
Capítulo 3 - Padrões de distribuição, riqueza e substituição de poáceas do Pantanal de Poconé-MT
RESUMO – No Pantanal, a família Poaceae possui maior riqueza entre as herbáceas e
grande importância econômica, podendo a variação hidrotopográfica afetar a
distribuição de suas espécies. Este trabalho realizado no Pantanal de Poconé objetivou
responder as seguintes questões: quais são as espécies com maior cobertura na região,
qual a influência da variação sazonal e microtopográfica na riqueza, cobertura,
distribuição, substituição de espécies, formas de vida e rotas fotossintéticas das
gramíneas? Os dados foram coletados utilizando-se o sistema RAPELD e metodologia
da parcela pontual para amostragem em 30 isolinhas de 250 m, realizadas nos períodos
de seca, cheia e vazante. No total foram amostradas 59 espécies, sendo três espécies
regionalmente dominantes e pouco variáveis em cobertura entre os três períodos, entre
elas: Axonopus purpusii, Brachiaria humidicola e Paspalum lineare. A distribuição e a
cobertura das gramíneas foram influenciadas pelo gradiente de inundação, assim como,
a riqueza em espécies, que diminuiu continuamente ao longo do gradiente de inundação.
Variações na diversidade beta foram notadas no espaço e no tempo. O número de
espécies de gramíneas anuais apesar de baixo foi superior ao que normalmente se
encontra para o Cerrado, com maior cobertura durante a seca nas isolinhas sujeitas a
maior profundidade de inundação. Espécies C4 estão presentes em maior número e
dominam em cobertura todo o gradiente. As espécies C3 também ocorrem ao longo de
todo gradiente com baixa cobertura e de forma mais equitativa.. Entretanto, a cobertura
de C3 aumenta em ambiente de maior profundidade durante a seca e cheia. A cobertura
média entre os períodos destas espécies C3 também revelou tendência de aumento nos
extremos, mais secos/sombreados e mais inundáveis. Assim, foi possível concluir que
esta família se distribui por todo o gradiente de inundação, mas que este afeta a
distribuição e a riqueza em espécies de gramíneas, de acordo com suas tolerâncias a
diferentes intensidades de estresse hídrico.
Palavras-chaves: Pantanal, gramíneas, gradiente de inundação, riqueza, diversidade
beta, forma de vida e rota fotossintética

112
Chapter 3 – Distribution, richness and turnover patterns of Poaceas in the Pantanal of Poconé, Mato Grosso
ABSTRACT – In the Pantanal, the Poaceae family has the greatest richness among
herbaceous plants and great economic importance. The hydro-topographic variable can
affect the distribution of its species. This study, made in the Pantanal of Poconé, aimed
to answer questions about which species are the most widespread in the region, what
influence seasonal changes may have and the microtopographics of the richness,
coverage, distribution, turnover, life forms and photosynthetic paths of grasses. Data
were collected using the RAPELD system and the point sampling method along 30
isolines 250m long, during the dry, flood and drain periods. In total, 59 species were
sampled, three of which are regionally dominant and showed little variation in coverage
over the three periods. These are: Axonopus purpusii, Brachiaria humidicola and
Paspalum lineare. The distribution and coverage of grasses were influenced by flood
gradient as well as species richness which diminished continuously along the flood
gradient. The beta diversity showed spatial and temporal changes in the species
composition. The number of annual grasses, despite being low, was higher than that
normally found in the Cerrado region and had greater coverage during the dry period
along the isolines subject to greater flooding depths. The average coverage over the
periods of the C3 species also showed a tendency to be higher in extreme conditions of
dryness/shade and flooding. It is thus possible to conclude that this family is distributed
along the whole flood gradient, but this affects the distribution and richness of grass
species as it favours species with more adaptable characteristics that can tolerate the
different stress intensities of this gradient.
Key words: Pantanal, gramineae, flood gradient, beta diversity, life form and
photosynthetic path

113
INTRODUÇÃO
As áreas savânicas e campestres, com elevada dominância de gramíneas, cobrem
cerca de 1/3 da superfície da Terra (Shantz 1954). A família Poaceae é extremamente
importante na produção de alimentos através de espécies domesticadas, como também,
para forragem na criação de herbívoros domésticos (Kellogg 2001; Jacobs et al. 1999).
Acredita-se que as primeiras gramíneas ocupavam bordas florestais ou ambientes
sombreados. Posteriormente, os membros desta família proliferaram em diversos tipos
de habitats (Kellogg 1998), devido a diversas adaptações morfofisiológicas que
contribuíram para o seu sucesso ecológico (Sarmiento 1992). Características
importantes originadas após o surgimento das gramíneas foram: a tolerância à
dessecação e a capacidade de crescer e prosperar em hábitas secos e abertos (Kellogg
2001). Nas savanas, seu domínio varia com o tipo de fisionomia ou densidade arbórea.
Os diferentes tipos de savanas (sazonal, hipersazonal e semi-sazonal) são
fisionomicamente semelhantes, sendo dominadas por espécies de gramíneas perenes,
entretanto, variando em composição, nas características morfofuncionais e dominância
em relação às rotas fotossintéticas de suas espécies (Killeen e Hinz 1992a; Sarmiento
1992; Medina e Motta 1990).
O Pantanal é uma savana hipersazonal, de acordo com Sarmiento (1983). Para
esta extensa planície inundável estão listadas, atualmente, 1903 espécies vegetais
superiores, sendo 1148 herbáceas, entre espécies terrestres, anfíbias e aquáticas (Pott &
Pott 1999; Junk et al. 2006). Este maior contingente de herbáceas ocorre devido a
predominância de áreas inundáveis, com formações aquáticas, campestres e savânicas
(Pott e Pott 1999). A família Poaceae é citada como a mais rica entre as herbáceas
registradas no Pantanal, com 212 espécies (Pott e Pott, 1999), sendo responsável por
90% da forragem (Allem & Valls 1987). Dentre os gêneros com maior número de
espécies estão Axonopus, Panicum e Paspalum (Pott e Pott 1999). Apesar desta
florística variada não há registros de endemismos, sendo este aspecto aparentemente um
atributo normal da vegetação de grandes planícies sedimentares, em especial, aquelas
com inundações periódicas (Valls 1994). A ausência de endemismos pode ser atribuída
à origem recente do ecossistema pantaneiro (Brown 1986) e à falta de isolamento (Junk
et al. 2006).

114
Em termos de composição florística de gramíneas, o Pantanal estaria mais
relacionado com outras áreas de savanas sul americanas do que o Cerrado do Brasil
Central, principalmente, por este apresentar latossolos com ótima drenagem na sua
maior parte, sendo edaficamente bastante distinto do Pantanal, que apresenta drenagem
deficiente relacionando-o de forma mais estreita às outras regiões de savanas, ditas
hipersazonais (Allem & Valls 1987).
A riqueza e a qualidade das gramíneas nativas, principalmente nas áreas
campestres, favoreceram a ocupação e introdução de herbívoros domésticos há cerca de
250 anos na região pantaneira (Mazza et al. 1994). Desde então, a criação de bovinos de
corte em sistemas extensivos, tem sido principal atividade econômica da região.
Segundo Allem e Valls (1987), além da riqueza em gramíneas forrageiras do Pantanal,
este abriga um conjunto mais nobre de espécies quando comparadas com o Cerrado
brasileiro e as outras savanas americanas, ocorrendo contínuas formações campestres
dominadas por uma só espécie pouco esclerófila e palatável. Killeen e Hinz (1992b)
argumentam que a riqueza em gramíneas no Pantanal é mais elevada, principalmente,
devido à sua variada microtopografia que cria diferentes micro-habitats, com distintos
regimes hídricos, cada um suportando diferenciadas espécies. Entretanto, nos últimos
anos tem havido crescente pressão para torná-lo competitivo. Entre as principais
modificações está a conversão de áreas florestadas em pastagens cultivadas,
principalmente aquelas localizadas sobre cordilheiras, livres da inundação (Silva et al.
1999). Para todo o Pantanal do Mato Grosso, estima-se que 6% da vegetação natural da
planície pantaneira fora suprimida, principalmente, para a introdução de gramíneas
exóticas, com diversos impactos sobre a fauna nativa devido a perda de habitats (Harris
et al. 2005). Um grande aumento no desmatamento de áreas não inundáveis ocupadas
por florestas deciduais e semi-deciduais ocorreu a partir de 1974, quando uma seqüência
de anos mais úmidos, com inundações mais pronunciadas, tornou grandes extensões de
campos nativos inundáveis menos disponíveis no ano (Silva et al. 1999). Campos
nativos sazonalmente inundáveis também têm sido convertidos em pastagens cultivadas
exóticas, principalmente, em áreas com domínio de gramíneas mais esclerófilas e pouco
palatáveis. Algumas espécies exóticas são adaptadas a tal variação hídrica sazonal,
como é o caso de Brachiaria humidicola (Dias-Filho & Carvalho 2000), destacando-se
em termos de área plantada (Santos et al. 2005). Segundo Harris et al. (2005) muitas
aves migratórias dependem dos campos nativos para manutenção de seu ciclo de vida.

115
As aves migratórias e nômades são ávidas consumidoras de sementes de capins nativos
do Pantanal, mas não há informação sobre consumo de espécies introduzidas, como as
braquiárias (Nunes e Tomas 2008).
Em áreas úmidas o regime hídrico, com variações na duração, amplitude e
freqüência de inundações, é determinante na distribuição e riqueza em espécies (Capon
2003; Southal et al. 2003; Townsend 2001; Casanova & Broke 2000; Lenssen et al.
1999; Robertson et al. 1984). No Pantanal, os gradientes hidrotopográfico e de solo
influenciam espacialmente a distribuição de espécies, dando origem a diferentes
comunidades vegetais (Nunes da Cunha e Junk 2001; Zeilhofer e Schessl 1999; Pinder e
Rosso 1998). Por outro lado, Rebellato & Nunes da Cunha (2005) relatam que os ciclos
anuais de inundações e secas promovem de forma temporal uma sucessão cíclica de
espécies aquáticas e terrestres nos campos inundáveis. As poáceas se distribuem por
todos os tipos de habitats do Pantanal, entretanto, dominam os ambientes com cotas
mais baixas do mesorelevo, ou seja, mais inundáveis, onde também são encontrados os
melhores campos nativos, do ponto de vista da pecuária (Pott 1988). As diversas
espécies desta família apresentam habitats preferenciais relacionados ao regime hídrico
e solo (Killeen e Hinz 1992b; Allem e Valls 1987; Pott 1986). O conhecimento de
diferentes comunidades distribuídas ao longo do gradiente hidrotopográfico, assim
como, o padrão comportamental dos animais domésticos frente a esta variabilidade de
habitats, podem auxiliar práticas de manejo sustentáveis, sendo essenciais na previsão
de uso de recurso e distribuição de impacto sobre a vegetação pelos animais (Santos et
al. 2001).
Portanto, considerando que em áreas úmidas a variação hidrotopográfica afeta o
estabelecimento de espécies vegetais, estimulando ou inibindo a germinação sementes
ou rebrota de determinadas espécies (Broke & Britton 1995), estando assim, as espécies
agrupadas de acordo com suas habilidades em tolerar ou responder à dinâmica de cheias
e secas (Casanova & Broke 2000), este trabalho pretende responder as seguintes
perguntas:
1) Como varia a diversidade de espécies de gramíneas entre os períodos de seca, cheia e
vazante?
2) Quais são as espécies de gramíneas com maior cobertura regional?

116
3) Como variam a composição e coberturas de espécies de gramíneas ao longo do
gradiente de inundação?
4) Como varia a diversidade beta ao longo do gradiente de inundação? A diversidade
beta, por exemplo, aumentará conforme aumenta o tempo de inundação neste gradiente?
5) Existe alguma influencia da vegetação local sobre a distribuição das espécies?
6) Como varia a riqueza em espécies de gramíneas ao longo do gradiente de inundação?
Esta seria mais elevada em cotas altimétricas intermediárias de inundação, de acordo
com a hipótese do distúrbio intermediário?
7) A cobertura das diferentes formas de vida de Raunkiaer e rotas fotossintéticas C3 e
C4 são influenciadas pelo gradiente de inundação?
MATERIAL E MÉTODOS
Características gerais e área de estudo – O relevo da Bacia do Alto Paraguai possui
cotas altimétricas que oscilam entre 80 a 1200 metros, distribuídas por três regiões
fisiográficas: dos planaltos (250 a 1200 metros), das depressões (150 a 250 metros) e
dos pantanais (80 a 150 metros) (Alvarenga et al. 1984). Hidrologicamente é
representada, principalmente, pelos rios Paraguai, Branco, Nabilieque, Miranda,
Aquidauana, Negro, Taquari, Cuiabá, Itiquira, Bento Gomes, São Lourenço e afluentes
(Amaral Filho 1986).
Geomorfologicamente, a região dos pantanais é representada pelas unidades
planícies e pantanais mato-grossenses, onde ocorrem os maiores fenômenos das
enchentes de toda a Bacia do Alto Paraguai (Alvarenga et al. 1984). Essas planícies
deprimidas e inundáveis recebem o nome de Pantanal e se localizam na parte central da
América do Sul, entre longitudes de 16º e 22º e latitudes de 55º e 58º. A declividade é
inexpressiva: o gradiente topográfico varia de 0,3 a 0,5 cm/km no sentido leste-oeste e
0,03 a 0,15 cm/km no sentido norte-sul. Esta região tem grande expressão espacial com
configuração de um enorme anfiteatro voltado para o oeste (Franco & Pinheiro 1982).
A tendência para inundações periódicas não está ligada à pluviosidade, que é
considerada baixa, mas sim à topografia plana, cotas altimétricas baixas e
predominância de solos hidromórficos, que influenciam na drenagem (Amaral Filho
1984). Entretanto, a heterogeneidade morfológica da região determina áreas em que a
inundação ocorre de forma distinta no tocante a altura, época e duração das inundações
(Adámoli 1987). Outra característica importante é que esta vasta superfície rebaixada é

117
recoberta por sedimentos quaternários, oriundos de diferentes rios que formam a bacia
do rio Paraguai (Adámoli 1987), sendo sua litologia constituída por sedimentos aluviais
da formação Pantanal, que associados à dinâmica do regime de alagamento, provocam
grande variação nos solos. Na parte norte predominam os solos com horizonte
subsuperficial de textura mais argilosa, na parte central, solos mais arenosos, e na parte
sul solos argilosos (Amaral Filho 1984). A heterogeneidade interna, promovida pelos
diferentes tipos de solos e regime hídrico, aliada a seu posicionamento fitogeográfico,
levaram a diferentes sistemas de divisão do Pantanal em sub-regiões. Silva & Abdon
(1998) delimitam e quantificam o Pantanal em 138.183 km2 e o dividem em 11 sub-
regiões de acordo com as suas características bióticas e abióticas.
A tendência para inundações periódicas não está ligada apenas à pluviosidade,
que é considerada baixa, mas sim à topografia plana, cotas altimétricas baixas e
predominância de solos hidromórficos que influenciam na drenagem (Amaral Filho
1986). Entretanto, a heterogeneidade topográfica da região determina áreas em que a
inundação ocorre de forma distinta no tocante a altura, época e duração das inundações
(Adámoli 1987). Outra característica é que esta vasta superfície rebaixada é recoberta
por sedimentos quaternários, oriundos de diferentes rios que formam a bacia do rio
Paraguai (Adámoli 1987), sendo sua litologia constituída por sedimentos aluviais da
formação Pantanal, que associados à dinâmica do regime de alagamento, provocam
grande variação nos solos. Ao norte predominam os solos com horizonte subsuperficial
de textura mais argilosa, na parte central, solos mais arenosos, e ao sul, solos argilosos
(Amaral Filho 1986). A heterogeneidade promovida pelos diferentes tipos de solos e
regime hídrico, que determinam diferenças fitogeográficas, levaram à divisão do
Pantanal em sub-regiões. Silva & Abdon (1998), por exemplo, delimitam o Pantanal em
138,183 km2 e o dividem em 11 sub-regiões.
Em termos forrageiros os fatores edáfico e regime hídrico têm forte influência
sobre a distribuição de espécies e na maior ou menor expressão de certos grupos ou
associações de espécies nos diferentes pantanais. Os pantanais da Nhecolândia e
Paiaguás, por exemplo, são essencialmente arenosos, apresentando grandes formações
campestres com a associação de gramíneas psamófilas, como Axonopus purpusii e
Reimarochloa brasiliensis, Ichnanthus procurrens, Reimarochloa acuta, Mesosetum
spp. Digitaria fuscescens, entre outras. Mas para exemplificar o efeito do regime hídrico
é interessante considerar o Pantanal de Aquidauana que é mais alto, sendo que na sua
parte arenosa esta associação não ocorre, uma vez que R. brasiliensis é mais exigente

118
em umidade do que A. purpusii, que domina as grandes porções de solos arenosos.
Entre os pantanais essencialmente argilosos estão o de Poconé e grande parte de
Nabileque e Miranda. Axonopus compressus (umbrófila), Hemarthria altissima,
Paspalum almum, Paspalum virgatum, Paspalum plicatum, Panicum laxum, Setaria
geniculata são gramíneas representativas do Pantanal de Poconé; Paspalum virgatum,
Paspalum plicatum, Panicum laxum, Hymenachne amplexicaulis, Leptochloa virgata
são expressivas em Nabileque, enquanto que Hemarthria altissima, Hymenachne
amplexicaulis, Paspalum almum, Panicum laxum e Leersia hexandra ocorrem em
Miranda. As espécies dos pantanais arenosos são mal representadas nos pantanais
argilosos, ocorrendo de forma mais esparsa. Entretanto, para o Pantanal de Poconé,
Reimarochloa brasiliensis é abundante em campos limpos, fato que também ocorre no
Pantanal de Barão de Melgaço, em sua porção argilosa. Outros pantanais como o de
Abobral, Aquidauana, Cáceres e Barão de Melgaço possuem solos ora argilosos ora
arenosos, sendo este último mais comum nestes pantanais. Miranda, Nabileque e
Poconé possuem maior proporção de solos siltosos ou argilosos e retém forragem verde
durante parte da estação seca, o que não ocorre nos pantanais arenosos e áreas do
Cerrado (Allen e Valls 1987).
As características edáficas e regime hídrico de cada Pantanal, portanto, refletem
na expressão das espécies forrageiras. Além disso, cada sub-região possui variações
internas em seu mesorelevo os quais também terão influência, em menor escala, na
distribuição e abundância de espécies de gramíneas.
A área em que foram realizadas as coletas engloba três fazendas da sub-região
do Pantanal de Poconé (Pantanal Norte), sendo estas, Fazendas Nossa Senhora da
Aparecida, São Carlos e Canoas, próximas à localidade do Pirizal, nordeste do Pantanal
de Poconé, município de Nossa Senhora do Livramento (Oeste: 56º 21', Leste: 56º 18',
Norte: 16º 19', Sul: 16º 22') (Fig. 1).
Os solos são argilosos em grande parte, com grau de fertilidade superior ao dos
outros pantanais (Adámoli 1987). Possui aportes aluviais originários quase que
totalmente do rio Cuiabá com transporte e deposição de sedimentos variados (argilosos,
siltosos e arenosos) derivados de rochas como filitos e xistos, principalmente. Apresenta
uma grande variedade de tipos de solos, predominando associações como planossolo
solódico eutrófico, laterita hidromórfica solódica eutrófica e solonetz solodizado
eutrófico, e imensas áreas de hidromórfico cinzento eutrófico, vertissolos, laterita
hidromórfica distrófica e planossolo distrófico (Amaral 1987). Ao norte, apresenta

119
extensa faixa com fraca inundação, onde as cotas altimétricas excedem 130 metros. Em
direção ao sul as cotas altimétricas decrescem para 110-100 metros, onde as áreas são
fortemente inundadas (Franco e Pinheiro 1982). Na presente área de estudo altimetria
ficou entre, aproximadamente, 115 e 112 metros.
O clima é do tipo AW-quente e úmido (Köppen 1948), com precipitação média
de 1.250 mm ao ano e a média anual de temperatura de 25,8 oC. No Pantanal Norte duas
estações são observadas durante o ano, sendo uma seca no período de maio a setembro e
a outra chuvosa, de outubro a abril (Nunes da Cunha & Junk 2004). Prado et al. (1994)
dividiram as diferentes fases ao longo do ano de três em três meses no Pantanal Norte,
sendo considerada: enchente (outubro, novembro e dezembro), cheia (janeiro, fevereiro
e março), vazante (abril, maio e junho) e seca (julho, agosto e setembro). No Pantanal
Norte a inundação é do tipo monomodal ocorrendo juntamente com a estação chuvosa
(outubro a abril) (Hamilton et al. 1996).
Para o Pantanal de Poconé a Savana (Cerrado) é a classe de vegetação que cobre a
maior parte da região (41,02%), seguida pela classe denominada de sistema de transição
(ex. florestas semi-sempre verdes sazonalmente inundáveis) (14.7%), florestas sazonais
semi-deciduais (6.23%), sistemas secundários (3.29%), corpos d’água (0.80%) e florestas
sazonais decíduas (0.40%) (Nunes da Cunha et al. 2006).

120
Figura 1- Área de estudo, fazendas Nossa Senhora Aparecida, São Carlos e Canoas,
próximas à localidade do Pirizal, município de Nossa Senhora do Livramento, sub-
região do Pantanal de Poconé-MT (Pinho 2005).
Desenho amostral – Este trabalho foi realizado utilizando-se do método RAPELD.
Este é uma modificação do método de Gentry (1982), desenvolvido para inventários
rápidos da biodiversidade em sítios de pesquisa ecológica de longa duração na
Amazônia (Magnusson et al. 2005), implementado no Pantanal de Poconé. Na área de
estudo foram instalados 30 isolinhas de 250 metros sistematicamente distribuídas em
um sistema de seis trilhas de 5 km cada orientadas no sentido norte-sul, ocupando uma
área de 25 Km2. Cada trilha possui uma identificação (A, B, C, D, E, F) onde foram

121
instaladas 5 isolinhas (Fig. 2), orientadas no sentido leste-oeste. A primeira isolinha de
cada uma das trilhas inicia-se a partir de 500 metros do início das mesmas. Assim, por
exemplo, a isolinha A1 está situada a 500 metros do início da trilha A. Cada isolinha
dista 1 Km uma da outra e foram implantadas em diferentes cotas altimétricas. A
marcação das mesmas foi feita com piquetes de ferro fixos no solo a cada 10 m, sendo a
altimetria do ponto inicial mantida ao longo dos 250 metros.
ABCDEF
500
1000
1500
2000
3000
2500
3500
4000
4500
5000
Figura 2- Imagem de satélite mostrando a área de estudo (5 km x 5 km) com sistemas de
6 trilhas (apenas Norte-sul) e esquema com distribuição das isolinhas para cada trilha,
Pantanal de Poconé-MT.
Amostragem de gramíneas – As coletas foram realizadas durante três fases
hidrológicas do Pantanal: na seca (setembro/outubro de 2006, fim da seca), na cheia
(fevereiro/março de 2007) e na vazante (maio de 2007). A amostragem das gramíneas
foi realizada pelo método da parcela pontual (Bullock 1996). Ao longo dos 250 metros
de cada isolinha foram distribuídas parcelas pontuais distantes 1 metro uma da outra. A
parcela pontual é considerada uma haste de metal fina posicionada verticalmente, onde
qualquer morfoespécie de gramínea que a toca é registrada. Um exemplar de cada morfo
espécie foi coletado, prensado e seco em estufa, sendo posteriormente enviado a
especialistas para identificação. Futuramente as exsicatas serão depositadas no Herbário
Central da Universidade Federal de Mato Grosso (UFMT) e Universidade Federal de
Minas Gerais (UFMG). As formas de vida das espécies identificadas foram
determinadas segundo Raunkier (1934), seguindo as classificações feitas nas listagens
florísticas de Schessl (1999), Meira-Neto et al. (2007) e, principalmente, Pott e Adámoli
1 km 5 km
Norte
Sul
Leste Oeste
1 km
F E D C B A
5 km

122
(1999). Posteriormente, as espécies foram classificadas segundo as rotas fotossintéticas
C3 e C4 (Grass Phylogeny Working Group (GPWG) 2001; Killeen e Hinz 1992;
Zuloaga 1986; Brown 1977).
Análises dos dados – As contagens das espécies nas parcelas pontuais permitiram
estimar a riqueza, cobertura, diversidade e variação de diversidade entre isolinhas e
períodos de seca, cheia e vazante. O tempo de inundação nas isolinhas foi altamente
correlacionada com a profundidade da inundação (r2 = 0,913, P < 0.001, n = 30). A
profundidade da inundação variou entre 0 cm e 63 cm (média = 23 cm, dp = 16,14). O
tempo de inundação variou entre 0 e 200 dias (média = 90,26 dias, dp = 50,30). Os
dados de profundidade e duração da inundação foram coletados entre os anos de 2006 e
2007 por Fantin-Cruz (2008, dissertação de mestrado).
Curvas das coberturas relativas das espécies, ranqueadas da mais abundante para
a menos abundante - uma representação visual da diversidade - foram comparadas entre
os três períodos. Para isso, foram utilizados os valores de cobertura regional de cada
espécie em cada período.
O número de parcelas pontuais em que cada espécie ocorreu foi utilizado para o
cálculo de sua cobertura na isolinha (número de parcelas pontuais na qual a espécie
ocorreu / 250). Outro cálculo realizado foi o de cobertura regional das espécies (soma
do número de parcelas pontuais nos quais a espécie ocorreu / total de parcelas pontuais
(= total de ocorrências / 30 x 250). A cobertura por diferentes formas de vida e rotas C3
e C4 seguiu o mesmo cálculo da cobertura regional, sendo feito da seguinte forma:
soma do número de parcelas pontuais na qual a forma de vida ou rota fotossintética
ocorreu / 30 x 250. Para avaliarmos o efeito da profundidade da inundação na cobertura
das formas de vida e rotas fotossintéticas, as isolinhas foram agrupados em classes de
profundidade de 9 cm (totalizando seis classes), sendo o cálculo para cobertura
realizado da seguinte forma: soma do número de parcelas pontuais na qual a forma de
vida ou rota fotossintética ocorreu em cada classe de profundidade / total de isolinhas
daquela classe de profundidade x 250. Estes cálculos foram feitos para os períodos de
seca, cheia e vazante.
Um ordenamento das espécies de gramíneas de acordo com suas coberturas em
cada uma das isolinhas dispostos segundo o gradiente de duração de inundação foi feito
para cada período. Para tanto, utilizou-se o programa COMUNIDATA desenvolvido
por Dias (2006).

123
As relações entre riqueza em poáceas, profundidade e períodos amostrados
(seca, cheia e vazante) foram investigadas através de uma Análise de Covariância
(ANCOVA), utilizando o modelo: Y(riqueza em poáceas) = constante + classes de
profundidade (covariável) + período (fator). O teste de Kolmogov-Smirnov foi
realizado para verificar se a distribuição dos resíduos era normal, utilizando-se para
tanto os valores de probabilidades de Lilliefors. Para esta análise utilizou-se o pacote
estatístico SYSTAT 10.2.
O cálculo da beta diversidade (β) foi realizado através do índice βT (beta
turnover) sendo βT = [g(H) + l(H)] / 2αm, onde g(H) é o número de espécies novas
adicionadas e l(H) o número de espécies perdidas ao longo do gradiente, sendo αm a
média da diversidade alfa das 30 isolinhas. O βT (beta turnover) é indicado para
amostras que podem ser ordenadas ao longo de um gradiente dominante, sendo
estimado entre pares de amostras adjacentes ou não (Wilson e Shmida 1984). A
diversidade beta foi analisada de diferentes formas. Na primeira, a diversidade beta foi
analisada entre pares de isolinhas adjacentes ordenados em um gradiente de tempo de
inundação. Na segunda, foram comparados os pares de isolinhas formadas sempre entre
uma cota fixa (formada pelas 3 isolinhas não inundáveis) com as demais 27 isolinhas
ordenados num gradiente de tempo de inundação. Na terceira, foram comparados os
pares de isolinhas adjacentes ordenados em um gradiente de tempo de inundação, onde
os valores de βT , posteriormente, foram ordenados em relação às diferenças de tempo
da inundação entre os pares formados, com o ajuste de uma parábola. E na quarta, os
períodos foram ordenados na seqüência seca-cheia, cheia-vazante e vazante-seca e βT
foi calculada. Para a segunda forma de análise este índice permitiu testar a hipótese de
que as mudanças na composição das espécies de gramíneas se acentuam ao longo do
gradiente de tempo de inundação e que essas mudanças variam entre os períodos
amostrados. A correlação de Spearman foi usada para correlacionar βT com os valores
de tempo de inundação.
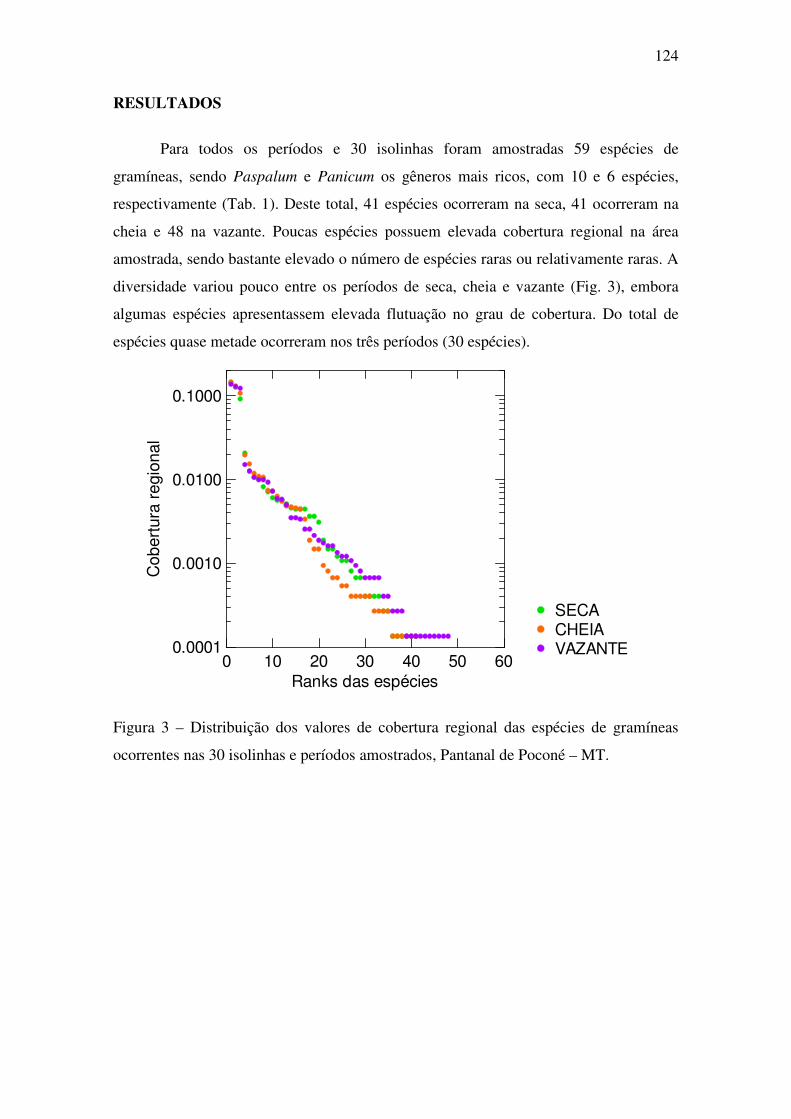
124
RESULTADOS
Para todos os períodos e 30 isolinhas foram amostradas 59 espécies de
gramíneas, sendo Paspalum e Panicum os gêneros mais ricos, com 10 e 6 espécies,
respectivamente (Tab. 1). Deste total, 41 espécies ocorreram na seca, 41 ocorreram na
cheia e 48 na vazante. Poucas espécies possuem elevada cobertura regional na área
amostrada, sendo bastante elevado o número de espécies raras ou relativamente raras. A
diversidade variou pouco entre os períodos de seca, cheia e vazante (Fig. 3), embora
algumas espécies apresentassem elevada flutuação no grau de cobertura. Do total de
espécies quase metade ocorreram nos três períodos (30 espécies).
0 10 20 30 40 50 60
Ranks das espécies
0.0001
0.0010
0.0100
0.1000
Co
ber t
ura
re
gio
nal
VAZANTECHEIASECA
Figura 3 – Distribuição dos valores de cobertura regional das espécies de gramíneas
ocorrentes nas 30 isolinhas e períodos amostrados, Pantanal de Poconé – MT.

125
Tabela 1 – Listagem das espécies de gramíneas amostradas nas 30 isolinhas e períodos
(seca, cheia e vazante) e respectivos valores de cobertura regional, formas de vida, rota
fotossintética e forma de crescimento, Pantanal de Poconé-MT.
Espécies Seca Cheia Vazante Formas de
vida Rota
fotossintética Andropogon bicornis L. 0.001867 0.00093333 0.0012 Hemi C4 Andropogon hypogynus Hack. 0.0044 0.0044 0.00493333 Hemi C4 Andropogon selloanus (Hack.) Hack.
0.0044 0.0072 0.00586667 Hemi C4
Aristida cappilacea Lam. 0 0.00026667 0.00213333 Tero C4 Aristida cf. riparia Trin. 0 0 0.00026667 Hemi C4 Aristida sp. 0.0004 0 0 Hemi C4* Axonopus cf. brasiliensis (Spreng.) Kuhlm.
0 0 0.00013333 Hemi C4
Axonopus chrysoblepharis (Lag.) Chase
0.000267 0.00026667 0.0008 Hemi C4
Axonopus compressus (Sw.) P. Beauv.
0.0004 0.0004 0.0004 Hemi C4
Axonopus leptostachyus (Flüggé) Hitchc.
0.0036 0.00146667 0.00173333 Hemi C4
Axonopus purpusii (Mez) Chase 0.144133 0.10626667 0.13586667 Hemi C4 Brachiaria humidicola (Rendle) Schweick.
0.0908 0.1428 0.12186667 Hemi C4
Coelorachis aurita (Steud.) A. Camus
0 0.00013333 0.00013333 Hemi C4
Digitaria ciliaris (Retz.) Koeler 0 0 0.00013333 Tero C4 Digitaria sp.3 0.0004 0.00013333 0.00013333 Ni C4* Digitaria sp.4 0 0 0.00013333 Ni C4* Echinolaena gracilis Swallen 0.000267 0.00186667 0.00186667 Hemi C3 Elyonurus muticus (Spreng.) Kuntze
0.0124 0.0152 0.01253333 Hemi C4
Eragrostis articulata (Schrank) Nees
0.000667 0 0 Tero C4
Eragrostis rufescens Schrad. ex Schult.
0.000667 0.00026667 0.00066667 Tero C4
Ichnanthus procurrens (Nees ex Trin.) Swallen
0.007067 0.00466667 0.00346667 Hemi C3
Imperata brasiliensis Trin. 0.0044 0.00546667 0.00573333 Hemi C4 Imperata tenuis Hack. 0.000133 0.0004 0.00026667 Hemi C4 Lasiacis sorghoidea (Desv. ex Ham.) Hitchc. & Chase
0 0 0.00013333 Hemi C3
Leersia hexandra Sw. 0.000133 0 0 Hemi C3 Leptochloa virgata (L.) P. Beauv. 0.0004 0 0 Hemi C4 Leptocoryphium lanatum (Kunth) Nees
0.0036 0.00453333 0.00346667 Hemi C4
Luziola fragilis Swallen 0 0.00333333 0.00066667 Hemi C3 Mesosetum ansatum (Trin.) Kuhlm. 0.004533 0.0108 0.00253333 Hemi C4 Mesosetum chaseae Luces 0.001067 0.0004 0 Hemi C3 Mesosetum loliiforme (Hochst. ex Steud.) Chase
0.008133 0.01053333 0.00986667 Hemi C4
Otachyrium versicolor (Döll) Henrard
0 0.00013333 0.0016 Hemi C3
Panicum dichotomiflorum Michx. 0.000133 0 0 Tero C4
continua….

126
continuação Tab. 1 Panicum discrepans Döll, atual Cyphonanthus discrepans (Döll) Zuloaga & Morrone
0.001467 0 0 Tero C4
Panicum laxum Sw. 0.005067 0.01173333 0.0092 Hemi C3 Panicum pilosum Sw. 0.001067 0.0008 0.00066667 Hemi C3 Panicum stenodes Griseb. 0.01 0.01946667 0.01493333 Hemi C4 Panicum trichanthum Nees 0 0.0004 0.00013333 Tero C3 Paratheria prostrata Griseb. 0.000133 0 0 Tero C3 Paspalum carinatum Humb. & Bonpl. ex Flüggé
0.003067 0.00066667 0.00106667 Hemi C4
Paspalum delicatum Swallen 0 0 0.00013333 Tero C4 Paspalum gardnerianum Nees 0.000133 0 0 Tero C4 Paspalum lineare Trin. 0.124933 0.12973333 0.1276 Hemi C4 Paspalum multicaule Poir. 0 0.00026667 0.00093333 Tero C4 Paspalum plicatulum Michx. 0.0056 0.0048 0.01053333 Hemi C4 Paspalum sp.1 0.001467 0.00146667 0.0012 Hemi C4* Paspalum sp.2 0 0.00053333 0.0016 Hemi C4* Paspalum stellatum Humb. & Bonpl. ex Flüggé
0 0.00066667 0.00333333 Hemi C4
Paspalum subciliatum Chase 0.0012 0 0 Hemi C4 Poaceae21 0 0 0.00066667 Ni * Poaceae31 0 0 0.00013333 Ni * Poaceae9 0.000133 0 0 Hemi * Reimarochloa brasiliensis (Spreng.) Hitchc.
0.020533 0.00013333 0.00026667 Tero C4
Sacciolepis myuros (Lam.) Chase 0 0.0004 0.00133333 Tero C3 Schizachyrium microstachyum (Desv. ex Ham.) Roseng.
0.0008 0.00053333 0.0004 Hemi C4
Schizachyrium tenerum Nees 0.0056 0.00733333 0.00986667 Hemi C4 Setaria geniculata P. Beauv. 0.010533 0.00013333 0.00253333 Tero C4 Setaria vulpiseta (Lam.) Roem. & Schult.
0 0.00013333 0.00013333 Hemi C4
Sorghastrum setosum (Griseb.) Hitchc.
0.006 0.00626667 0.0072 Hemi C4
* Espécies que foram classificadas em C3 e C4 de acordo com os gêneros indicados por Vall (1994)

127
Cobertura regional de gramíneas – Nos três períodos analisados três espécies de
gramíneas obtiveram elevada cobertura regional: Axonopus purpusii (0,144, 0,106 e
0,135), Paspalum lineare (0,124, 0,129 e 0,127) e Brachiaria humidicola (0,908, 0,142
e 0,121), respectivamente para a seca, a cheia e a vazante (Tab. 2). Axonopus purpusii é
a espécie que também possui a mais elevada freqüência de ocorrência nas 30 isolinhas
amostradas, com ocorrência em 83,3% das isolinhas na seca, 80,0% na cheia e 83,3% na
vazante. As demais 56 espécies possuem coberturas muito mais baixas em relação a
estas três. Entre estas, aquelas que possuem cobertura um pouco mais elevada se
mantêm no ambiente durante os três períodos. Algumas delas apresentando variações
maiores na cobertura regional, como é o caso de Ichinathus procurrens, Mesosetum
ansatum, Panicum laxum, Paspalum plicatulum Panicum stenodes, mas ainda assim
com certa estabilidade. As gramíneas Reimarochloa brasiliensis e Setaria geniculata,
tiveram uma forte variação na cobertura entre os períodos, sendo mais características do
período de seca (Fig. 4).

128
Figura 4 – Cobertura regional de 56 espécies entre os diferentes períodos, Pantanal de
Poconé – MT. cores branca = seca, cinza = cheia e preto = vazante. * espécies ocorrentes em
apenas um período.

129
Distribuição de gramíneas ao longo do gradiente de inundação – As gramíneas
apresentaram preferências quanto a sua ocorrência no gradiente de inundação. Nas
figuras 5.1, 5.2 e 5.3 observa-se que muitas espécies ocorrem preferencialmente em
ambientes livres da inundação ou pouco inundáveis, como Elionurus muticus, Imperata
brasiliensis, Paspalum carinatum, Axonopus chrysoblepharis, Axonopus compressus,
Setaria vulpiseta e Aristida capillacea. Outras espécies se localizam, praticamente, por
todo o gradiente nas isolinhas inundáveis, como Axonopus purpusii, Mesosetum
lolliforme, Andropogon selloanus, Paspalum lineare, Schizachyrium tenerum,
Ichnanthus procurrens, Mesosetum ansatum e Andropogon hypogynus, entretanto, a
maioria apresentando limitações entre os extremos do gradiente. Na cheia e na vazante a
cobertura de gramíneas localizadas nas isolinhas muito inundáveis é extremamente
baixa, sendo composta por Luziola fragilis, Panicum laxum e Brachiaria humidicola.
Estas isolinhas durante o período da seca têm elevada cobertura de espécies como
Brachiaria humidicola, Setaria geniculata e Reimarochloa brasiliensis, tendo estas
duas últimas ocorrências preferenciais nestes ambientes.

130
Figura 5.1 – Distribuição de valor de cobertura das 41 espécies ocorrentes nas 30 isolinhas
durante período de seca ao longo do gradiente de tempo da inundação, Pantanal de Poconé-MT.

131
Figura 5.2 - Distribuição de valor de cobertura das 41 espécies ocorrentes nas 30 isolinhas
durante período de cheia ao longo do gradiente de tempo da inundação, Pantanal de Poconé-
MT.

132
Figura 5.3 - Distribuição de valor de cobertura das 48 espécies ocorrentes nas 30 isolinhas
durante período de vazante ao longo do gradiente de tempo da inundação, Pantanal de Poconé-
MT.

133
Riqueza em gramíneas no gradiente de inundação – A riqueza em espécies de poáceas
diminui significativamente com o aumento da profundidade dos isolinhass (r2 = 0.839, p
< 0,001), sendo esta variação independente dos períodos (p = 0,348) (Fig. 6).
1 2 3 4 5 6
Classes de Profundidade
5
10
15
20
25
30
Riq
ueza e
m E
sp
écie
s d
e P
oá
ceas
Figura 6 – Relação entre a riqueza acumulada de gramíneas nas classes de profundidade
das 30 isolinhas agrupados a cada 9 cm de profundidade para os três períodos (círculos
azuis: seca; círculos verde: cheia; círculos alaranjados: vazante).
Diversidade Beta no gradiente – Nas figuras 7 (A, B, C D) podemos observar o
comportamento da βT de diferentes formas. A primeira análise (Fig.7A) mostra que os
valores de βT são mais elevados, principalmente, entre os isolinhass não inundáveis ou
pouco inundáveis, ou seja, nas cotas mais elevadas, sendo mais baixos entre as isolinhas
muito inundáveis. Na segunda forma de análise (Fig. 7B) observa-se que βT inicialmente
aumenta até certo ponto e novamente decai quando o tempo de inundação ultrapassa os
100 dias, para a cheia e vazante. A correlação de Spearman apenas é positiva e
significativa durante o período da seca (rs=0,5259), enquanto na cheia e vazante, a
relação não ocorre (rs=0,1329 e rs=0,045, respectivamente). Na terceira forma de
análise (Fig. 7C) observa-se há uma tendência de aumento de βT em relação ao aumento
das diferenças de tempo de inundação entre os pares, mas que, entretanto, quando muito
acentuadas as diferenças, ocorreram quedas. Entre os pares com diferença zero há uma

134
grande variação nos valores de βT. Entre períodos, βT é mais elevada entre os pares seca-
cheia e vazante-seca (Fig.7D).
Figura 7 – Diversidade beta (βT) comparando pares de isolinhas adjacentes (A), pares
formados entre uma cota fixa (composição de espécies das 3 isolinhas não inundáveis) e
as demais 27 isolinhas (B), pares de isolinhas adjacentes e ordenadas pelas diferenças de
tempo da inundação (C) e entre pares de períodos (D), Pantanal de Poconé-MT.
Formas de vida e rotas C3 e C4 - Cinqüenta e quatro, das 59 espécies, foram
classificadas segundo as formas de vida de Raunkier (1934), sendo 76% (41 espécies)
consideradas hemicriptófitas e 24% (13 espécies) como terófitas. Mesmo quando as
formas de vida são analisadas entre os diferentes períodos, as hemicriptófitas são as
dominantes, com 32 espécies na seca, 33 na cheia e 35 na vazante (Fig. 8). Nota-se,
entretanto, que existe uma tendência de aumento gradativo na cobertura de espécies
terófitas no sentido cheia-seca (Fig. 9). Especificamente para o período da seca, estas se
concentram nas cotas mais baixas do relevo (classe 6 de profundidade, sendo

135
compostas, principalmente, por Reimarochloa brasiliensis e Setaria geniculata (Figura
10), espécies que provavelmente iniciam seu ciclo no final da seca (setembro/outubro),
quando o ocorrem as primeiras chuvas, e o terminam quando inicia-se a cheia.
Quarenta e quatro espécies foram classificadas como C4, o que corresponde a
74,6% do total de espécies, 12 foram classificas como C3 (20,3%) e 3 não identificadas.
(5,08%). Em todos os períodos as espécies com rotas C4 estiveram presentes em maior
número. A fração de espécies C4/C3 variou entre períodos, sendo 33/7 (ou
80,49/17,07%) na seca, 32/9 (ou 78,05/21,95%) na cheia e 37/9 (ou 77,08/18,75%) na
vazante. (Fig. 8). As espécies C4 também são dominantes em cobertura em todos os
períodos (Fig. 9) e em todo o gradiente de inundação, com cobertura variando entre as
classes de profundidade. A cobertura das espécies C3 foi bastante equitativa entre as
classes de profundidade, com sutil aumento na última classe durante a seca e a cheia
(Fig. 10). Entretanto, a cobertura de espécies C3 é mais elevada nos extremos (classe 1 e
classe 6) quando é feita a média entre os períodos (Fig. 11).
0
10
20
30
40
50
60
70
80
90
Hemi Tero C3 C4
Porc
enta
gem
de e
spécie
s
Seca Cheia Vazante
Figura 8 – Porcentagens das formas de vida e rotas fotossintéticas das espécies
ocorrentes nas 30 isolinhas e diferentes períodos amostrados, Pantanal de Poconé – MT.

136
0
0.1
0.2
0.3
0.4
0.5
0.6
Hemi Tero C3 C4
Cobert
ura
regio
nal
Seca Cheia Vazante
Figura 9 – Valores de cobertura regional das formas de vida e rotas fotossintéticas das
30 isolinhas e diferentes períodos amostrados, Pantanal de Poconé – MT.

137
SECA
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
classe 1 classe 2 classe 3 classe 4 classe 5 classe 6
Valo
r d
e c
ob
ert
ura
Hemi Tero C3 C4
CHEIA
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
classe 1 classe 2 classe 3 classe 4 classe 5 classe 6
Valo
r d
e c
ob
ert
ura
Hemi Tero C3 C4
VAZANTE
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
classe 1 classe 2 classe 3 classe 4 classe 5 classe 6
Valo
r d
e c
ob
ert
ura
Hemi Tero C3 C4
Figura 10 – Valores de cobertura das formas de vida em 6 classes de profundidades das
30 isolinhas para os diferentes períodos amostrados, Pantanal de Poconé – MT.

138
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
classe 1 classe 2 classe 3 classe 4 classe 5 classe 6
Valo
r d
e c
ob
ert
ura
C4
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
Valo
r d
e c
ob
ert
ura
C3
C4 C3
Figura 11 – Médias das coberturas de espécies C3 e C4 entre os períodos de seca, cheia
e vazante das diferentes classes de profundidade, Pantanal de Poconé-MT
DISCUSSÃO
Schessl (1999) realizou um grande levantamento na região norte do Pantanal,
totalizando 104 espécies de gramíneas, sendo 34 comuns a este trabalho. Assim, o
número atual de espécies ou a riqueza em espécies de gramíneas para a região do
Pantanal de Poconé pode chegar a 129 espécies. Pott & Adamoli (1999) identificaram
64 espécies de gramíneas no Pantanal de Paiaguás, sendo 33 comuns a este trabalho.
Nestes três levantamentos, os gêneros Paspalum e Panicum apresentaram maior número
de espécies.
Das 57 espécies consideradas comuns entre diferentes formações savânicas da
América do Sul (Cerrado brasileiro, Venezuelanas, Colômbianas e das Guianas)
(Sarmiento 1983), 18 são comuns a este trabalho: Andropogon bicornis, Andropogon
hypogynus, Andropogon selloanus, Aristida capillacea, Axonopus leptostachyus,
Axonopus purpusii, Leersia hexandra, Leptocoryphium lanatum, Mesosetum chaseae,
Mesosetum lolliforme, Panicum laxum, Panicum stenodes, Paratheria prostrata,
Paspalum carinatum, Paspalum gardnerianun, Paspalum multicaule, Paspalum
plicatulum e Setaria geniculata. Muitas destas ocorreram com coberturas mais elevadas
neste estudo e em todos os períodos, provavelmente, porque apresentam uma maior
plasticidade, distribuindo-se com maior amplitude e resistindo bem aos ciclos de seca e
cheia. Alguns gêneros, entretanto, comuns em determinadas formações savânicas,
sequer são citados para o Pantanal, como Ctenium, Diectomis e Tristachya, gêneros

139
encontrados no Cerrado, e Eriochrysis, gênero encontrado nas savanas Colombianas e
Venezuelanas (Allem & Valls 1987; Sarmiento 1983).
Das espécies que apresentaram alta cobertura regional, Axonopus purpusii
(capim-mimoso), espécie típica das savanas continentais (ocorrendo do México até o
Paraguai, sul do Brasil e Argentina), perene, psamófila e que resiste à submersão
temporária, se destaca como importante para a pecuária, pois é considerada uma boa
forrageira (Comastri Filho 1984; Allem e Valls 1987). Entretanto, densas formações
monoespecíficas desta espécie não foram encontradas na área de estudo, como ocorre
nos pantanais de Nhecolândia, Paiaguás, Aquidauana, Abobral e Cáceres. Já Paspalum
lineare (fura-bucho), também nativa e com elevada cobertura, é considerada uma
espécie grosseira, que tem como defesa folhas pontiagudas, roliças e fibrosas, podendo
ser pastada na rebrota após queima. Sarmiento (1992) comenta que muitas gramíneas
das savanas neotropicais são altamente esclerófilas, em parte, devido aos tecidos
mecânicos de sustentação e vasculares de suas folhas, que as tornam rígidas e
pontiagudas, tornando-as pouco palatáveis aos pastejadores. A qualidade e a
palatabilidade das pastagens dominadas por estas espécies é aumentada pela queima.
Brachiaria humidicola é uma gramínea exótica bastante utilizada devido a sua boa
adaptação aos ambientes alagados e ao pisoteio (Santos et al. 2005; Dias-Filho &
Carvalho 2000). Durante a inundação observou-se que esta espécie cresce
acompanhando o nível da inundação e por isso consegue se manter em ambientes muito
inundáveis, uma vez que suas folhas não ficam submersas. Gramíneas exóticas estão
sendo cada vez mais utilizadas no lugar das espécies nativas, seja em ambientes
inundáveis como em ambientes não inundáveis, florestados e não florestados, fato que
pode homogeneizar todo o mosaico vegetacional que propicia a grande riqueza e
abundância em espécies animais, incluindo espécies globalmente ameaçadas como a
arara-azul (Anodorhynchus hyacinthinus) (Harris et al. 2005). Aves migratórias
granívoras, como os papa-capins (Emberizidae), com grande representação do gênero
Sporophila, também utilizam extensas áreas campestres do Pantanal para procura de
sementes de gramíneas nativas. (Nunes e Tomas 2008).
O tipo de solo e da inundação (nível, duração e origem pluvial e/ou fluvial)
determinam os grupos de espécies de gramíneas presentes (Pott 1988). Gramíneas como
Panicum laxum, Reimarochloa brasiliensis e Setaria geniculata, com menor cobertura
regional neste trabalho, são citadas como espécies com alto valor forrageiro e
preferenciais de ambientes muito alagáveis (Pott 1986). Schessl (1999) e Rebellato e

140
Nunes da Cunha (2005) relatam que os ambientes em que estas espécies ocorrem com
elevada cobertura passam por uma acentuada mudança hidrológica ao longo do ano,
promovendo mudanças drásticas na florística e cobertura de espécies, sendo estas
dominantes durante a seca, declinando durante a cheia, como também fora observado
neste trabalho. Já Luziola fragilis se torna abundante em algumas isolinhas muito
inundáveis durante a cheia e vazante, pois também seu crescimento acompanha ao nível
da inundação. Estas gramíneas sendo mais comuns em cotas mais baixas do
mesorelevo, ou seja, com longa duração da inundação, tiveram baixa cobertura regional,
pois poucas isolinhas localizaram-se nestas cotas. Já espécies como Aristida capillacea,
Axonopus brasiliensis, Eragrostis rufescens, Paspalum carinatum e Paspalum
multicaule são citadas como preferenciais de ambientes pouco inundáveis, enquanto que
Axonopus compressus, Eragrostis articulata, Lasiacis sorghoidea Paspalum
gardnerianum, Schizachyrium microstachyum e Setaria vulpiseta são típicas de
ambientes livres da inundação (Pott 1986), o que também foi verificado nesse estudo.
O regime hídrico influencia notavelmente a riqueza em espécies de poáceas. A
primeira classe de profundidade une ambientes não inundáveis com os pouco
inundáveis, fato que, provavelmente, resultou em maior número de espécies. Entretanto,
a queda na riqueza aumentou com a profundidade, indicando que boa parte das espécies
não tolera níveis maiores de umidade por tempo mais prolongado. Alguns trabalhos
realizados em áreas úmidas também demonstraram a tendência de queda na riqueza em
espécies em ambiente muito ou permanentemente inundáveis (Fortney et al. 2004;
Capon 2003; Lenssen et al. 1999; Pinder e Rosso 1998). A profundidade e/ou a duração
da inundação, portanto, podem influenciar a comunidade como um todo, assim como
famílias específicas como a das poáceas. A hipótese de que uma maior riqueza seria
encontrada em cotas altimétricas (ou profundidades) intermediárias não foi sustentada,
reforçando o fato de que as gramíneas que predominam nos campos do Pantanal são
oriundas de ambientes não sujeitos a inundações. Provavelmente, essas gramíneas se
mantêm nos campos pantaneiros devido à alternância dos períodos úmido e seco,
quando então as populações depauperadas pelo excesso de umidade podem se recuperar.
A maior parte das espécies de ervas e gramíneas (78%) é considerada terrestre,
entretanto, possuem impressionante plasticidade morfológica e fisiológica que as
permitem co-ocorrer durante períodos úmidos no Pantanal (Junk et al. 2006).
A substituição de espécies entre cotas com altimetrias elevadas e próximas é
elevado, provavelmente refletindo a maior riqueza em espécies nos ambientes livres de

141
inundação e pouco inundáveis, como também, a passagem entre os ambientes não
inundáveis e os ambientes inundáveis, aumentando a substituição de espécies. Por outro
lado, nas cotas mais baixas, valores de βT baixos indicariam menor riqueza e menor
substituição de espécies. Além da menor riqueza, maior similaridade devido as
condições mais estressantes ligadas à submersão que poderiam resultar em menor
variação em βT. Quando o conjunto das isolinhas não inundáveis (unidas) é utilizado na
comparação com as demais 27 isolinhas, os valores de βT inicialmente aumentam até
certo ponto e novamente decai, o que também estaria relacionada a menor riqueza em
espécies e menor substituição de espécies nestas isolinhas mais profundas, isso para os
períodos de cheia e vazante. Desta forma, a hipótese de que βT aumentaria com o tempo
de inundação é verificada apenas para o período da seca, onde a correlação de Spearman
foi positiva e significativa, pois neste período as cotas mais baixas (ou isolinhas com
maior tempo de inundação) possuem mais espécies quando comparadas aos períodos de
cheia e vazante, possivelmente porque nestes períodos as mesmas são eliminadas, dadas
as condições hídricas extremas da cheia e pós cheia. Por outro lado, βT é influenciada
pela vegetação que predomina nas diferentes isolinhas, mascarando parte da variação
prevista pelas diferenças nos tempos de inundação, o que poderia explicar a relação
parabólica entre βT e diferença no tempo de inundação. Os valores de βT são mais
elevados quando compara o período de seca com aqueles inundados (cheia) e/ou úmido
(vazante) devido, principalmente, a maior substituição de espécies entre estes períodos
dadas as condições hídricas muito contrastantes que propiciam os aparecimento de
novas espécies aumentando a riqueza e a dissimilaridade na maior parte das isolinhas
nestes períodos.
O padrão de ocorrência e de substituição de espécies, portanto, pode ser
influenciado pelo regime hídrico, como também verificado com outros grupos vegetais
(ex. Capon 2003; Southal et al. 2003; Townsend 2001; Casanova & Broke 2000).
Entretanto, determinadas formações vegetais localizadas em ambientes com
pronunciada inundação, são mais pobres em espécies, o que diminui o acréscimo de
espécies, com queda na diversidade beta.
Para as comunidades herbáceas as espécies hemicriptófitas estão entre as formas
de vida mais representativas do Cerrado (Silva e Batalha 2008; Meira-Neto et al. 2007;
Batalha e Martins 2004; Costa et al. 2004; Batalha e Mantovani 2001; Warming 1892
apud Sarmiento e Monasterio 1983). No Cerrado, as hemicriptófitas possuem uma
tendência de aumento no gradiente fisionômico do Cerrado que vai do cerradão ao

142
campo limpo, tendo as fanerófitas tendência contrária (Meira-Neto et al. 2007). As
porcentagens de espécies hemicriptófitas podem variar de 30 a 55,1%, tendo as espécies
anuais variação entre 0,7 a 6,7% (Batalha e Martins 2002). Em trabalhos realizados no
Cerrado, a família Poaceae é quase que exclusivamente representada por espécies
hemicriptófitas (Silva e Batalha 2008; Meira-Neto et al. 2007; Costa et al. 2004; Batalha
2001).
Quando se considera apenas a família Poaceae, as hemicriptófitas são as mais
bem representadas com 78.1% de ocorrência (seguidas pelas terófitas, com 20.3% e
geófitas, com 1.6%) no Pantanal de Paiaguás (Pott e Adámoli 1999) e também no
Pantanal de Poconé, em que as espécies hemicriptófitas chegam a 71,2% (seguidas pelas
terófitas, com 23,1% e geófitas, com 3,9%) (Schessl 1999), proporções estas
semelhantes aos do atual estudo. Este maior contingente de espécies de gramíneas
anuais no Pantanal pode estar relacionado à variabilidade ambiental que inclui
ambientes sujeitos à inundação sazonal e outros distúrbios. Regimes de distúrbios são
citados como beneficiadores de espécies anuais (Grime 1977; Belsky 1992). No caso do
Pantanal, além de distúrbios como o fogo e o pastejo, a inundação também pode ser
considerada um tipo de distúrbio sazonal, selecionando um maior contingente de ervas e
gramíneas anuais em relação ao Cerrado. No Cerrado mesmo quando se comparou
savanas sazonais com hipersazonais não houve um aumento significativo de ervas e
gramíneas anuais. Provavelmente, o distúrbio ocasionado pelo alagamento desta savana
hipersazonal estudada não seria tão severo a ponto de favorecer a estratégia terófita. As
variações hídricas no pantanal são variáveis, podendo ocorrer inundações extremas, o
que pode selecionar para alguns ambientes uma maior contribuição de espécies anuais.
Neste estudo, a contribuição de espécies anuais pode ter sido baixa devido a
pouca amostragem em ambientes com variações hídricas mais extremas. Entretanto, elas
estão relacionadas ao gradiente de inundação. Elas estão presentes no final do período
de seca, sendo a cobertura mais elevada nas áreas onde o nível de inundação é maior e
onde, provavelmente, germinam com as primeiras chuvas, crescem e se reproduzem
antes do nível de inundação se tornar muito elevado. Tais espécies não são encontradas
nestas cotas durante a cheia, provavelmente, pela maior profundidade e duração da
inundação que as eliminam.
Valls (1994) comenta que apesar do Pantanal se encontrar em uma área tropical,
possui um número significativo de espécies de gramíneas caracterizadas pela rota
fotossintética C3, muitas destas, forrageiras exponenciais. Killeen e Hinz (1992a)

143
demonstraram que a rota fotossintética está relacionada com a forma de crescimento,
sendo que as espécies C3 apresentam folhas largas, folhagem caulescente e menor
esclerificação, o que as tornam mais palatáveis. Alguns trabalhos demonstram a
tendência de aumento da abundância de espécies C3 em habitats mais úmidos ou
encharcáveis (Killeen e Hinz 1992a; Medina e Motta 1990). Isso pode explicar a
tendência de uma maior cobertura de espécies C3 na classe de maior profundidade de
inundação ou nas cotas altimétricas mais baixas, durante o período de seca e cheia,
como também a presença de campos mais nobres, do ponto de vista da pecuária, nas
cotas mais baixas do mesorelevo como apontado por Pott (1988). Medina e Motta
(1990) observaram que nas savanas sazonais há predominância de gramíneas C4, nas
savanas hipersazonais co-ocorrem espécies C3 e C4 e a dominância de espécies C3
ocorreria apenas nas savanas semi-sazonais. Como o Pantanal é considerado uma
savana hipersazonal as espécies com as diferentes rotas fotossintéticas co-ocorreram em
acordo com os autores acima, entretanto, com dominância da rota C4. Isto sugere que o
estresse causado pela inundação só seria mais notável nas cotas altimétricas mais
baixas, onde as espécies C3 seriam favorecidas devido ao longo período de submersão,
no entanto, insuficiente para deslocar o balanço C3/C4 em direção a maior cobertura
pelas espécies com rota C3. Nas demais cotas o estresse causado pela duração do
período seco prevaleceria, favorecendo a prevalência de espécies C4. Entretanto, as
gramíneas C3 também apresentam tendência de ocupar ambientes sombreados,
conforme também observado por Killeen e Hinz (1992a) e Klink e Joly (1989), o que
pode explicar coberturas médias mais elevadas nos extremos das classes (Classe 1 e
Classe 6).
CONCLUSÕES
Três espécies foram regionalmente dominantes nos três períodos: Axonopus
purpusii, Brachiaria humidicola e Paspalum lineare.
A distribuição das diferentes espécies de gramíneas é influenciada pelo gradiente
de inundação, reflexo de suas tolerâncias diferenciadas ao grau de saturação hídrica ou
secura do solo.
A riqueza em espécies não se revelou superior nos níveis intermediários de
inundação, diminuindo continuamente ao longo do gradiente de inundação, fato que,

144
provavelmente, está relacionado ao aumento do estresse hídrico que elimina um maior
número de espécies.
A diversidade beta revelou mudanças espaciais, assim como, temporais na
composição de gramíneas.
O número de espécies de gramíneas anuais apesar de baixo é superior ao que
normalmente se encontra para o Cerrado, tendo maior cobertura na classe de maior
profundidade, sujeita a um maior estresse hídrico. Provavelmente, em níveis de maior
profundidade a cobertura e a riqueza em espécies anuais podem aumentar.
Espécies C4 estão presentes em maior número e dominam todos os ambientes
em cobertura. As espécies C3 também ocorrem ao longo de todo gradiente de forma
discreta e equitativa. Entretanto, a cobertura de C3 aumenta na classe de maior
profundidade durante a seca e cheia, onde apenas uma espécie anual foi responsável por
este aumento. Provavelmente, em níveis de maior profundidade a cobertura e a riqueza
em espécies C3 podem aumentar A cobertura destas espécies também apresentaram
tendência a aumentar nos extremos das classes de profundidade.
Desta forma, é possível concluir que as espécies da família Poaceae estão
distribuídas por todos os ambientes relacionados neste estudo, mas que a variabilidade
ambiental do Pantanal afeta a distribuição e a riqueza em espécies de gramíneas que
possuem adaptações para enfrentar as diferentes intensidades de estresse do gradiente,
assim como, dos ciclos sazonais de secas e cheias.
REFERÊNCIAS CITADAS
Adámoli, J. 1987. Fisiografia do Pantanal. Pp. 15-22. In: Recursos forrageiros nativos
do Pantanal Mato-Grossense. Embrapa/Cernagem, Brasília-DF. Documento 8.
Adámoli, J. 1982. O Pantanal e suas relações fitogeográficas com os cerrados: discussão
sobre o conceito de complexo do Pantanal. Pp.109-119. In: Anais do 32° Congresso
nacional da Sociedade Botânica do Brasil, Teresina, Universidade Federal do Piauí.
Allem, A. C. & Valls, J. F. M. 1987. Recursos forrageiros nativos do Pantanal Mato-
Grossense. Embrapa/Cernagem, Brasília-DF. Documento 8. 339p.

145
Alvarenga, S. M. M., Brasil, A. E., Pinheiro, R. e Kux, H. J. H. 1984. Estrudo
geomorfológico aplicado a Bacia do Alto Paraguai e Pantanais Mato-grossenses.
Boletim Tecnico do Projeto RADAMBRASIL (Serie Geomorfologia,1). Pp 90-183.
Amaral Filho, Z. P. 1984. Solos do Pantanal Mato-Grossense. Pp 91-96. In: Simpósio
sobre recursos naturais e sócio-econômico do pantanal, 1, Corumbá. Anais. Brasília,
EMBRAPA-DDT.
Amaral, J. A. M. 1987. Solos do Pantanal. Pp. 19-21.In: Recursos forrageiros nativos do
Pantanal Mato-Grossense. Embrapa/Cernagem, Brasília-DF. Documento 8.
Batalha, M. A. and Martins, F. R. 2004. Floristic, frequency, and vegetation life-form
spectra of a cerrado site. Brazilian Journal of Biology.64(2):201-209, 2004.
Batalha MA and Martins FR. 2002. Life-form spectra of Brazilian cerrado sites. Flora
197: 452-460.
Batalha, M. A. and Mantovani, W. 2001. Floristic composition of the cerrado in the Pé-
de-Gigante Reserve (Santa Rita do Passa Quatro, southeastern Brazil). Acta bot. bras.
15(3):289-304.
Belsky, A.J., 1992. Effects of grazing, competition, disturbance and fire on species
composition and diversity in grassland communities. Journal of Vegetation Science
3:187-200.
Bond, W.J., Midgley, G.F. & Woodward, F.I. 2003. What controls South African
vegetation – climate or fire? South African Journal of Botany 69:1-13.
Brown, W. V. 1977. The Kranz Syndrome and its subtypes in grass systematics. Memoirs of the Torrey Botanical Club 23(3): 1-97.
Bullock, J. 1996. Plants. In: Ecological census techniques. W.J. Sutherland (ed). Pp.
111-138. Cambridge University Press, Cambridge. 336p.

146
Capon, S. J. 2003. Plant community responses to wetting and drying in a large arid
floodplain. River research and applications 19:509-520.
Casanova, M. T. and Brock, M. A. 2000. How do depth, duration and frequency of
flooding influence the establishment of wetland plant communities? Plant Ecology 147
(2):237 250.
Comastri Filho, J. A. 1984. Pastagens nativas e cultivada no Pantanal
Matogrossense.Corumbá:EMBRAPA-UEPAE de Corumbá, (EMBRAPAUEPAE de
Corumbá. Circular Técnica, 13). 48p.
Costa, I. R., Araújo, F. S. e Lima-Verde, L. W. 2004. Flora e aspectos auto-ecológicos
de um encrave de cerrado na chapada do Araripe, Nordeste do Brasil. Acta botânica
brasílica 18(4): 759-770
Dias-Filho, M.B.; Carvalho, C.J.R. 2000. Physiological and morphological responses of
Brachiaria spp. to flooding. Pesquisa Agropecuária Brasileira 35:1959-1966
Fortney, R. F., Benedict, M., Gottgens, J. F., Walters, T. L., Leady, B. S. and Rentch, J.
2004. Aquatic plants community composition and distribution along an inundation
gradient at two ecologically-distinct sites in the Pantanal region of Brasil. Wetlands
Ecology and Management 12:575-585.
Franco, M. S. M. e Pinheiro, R. 1982. Geomorfologia. Pp.161-224. In: Brasil.
Ministério das Minas e Energia. Departamento Nacional da Produção Mineral. Projeto
Radam-Brasil. Folha SE.21 Corumbá. Rio de Janeiro.
Grass Phylogeny Working Group (GPWG). 2001. Phylogeny and subfamilial
classification of the grasses (Poaceae). Annals of the Missouri Botanical Garden 88(3):
373-457.
Grime, J.P., 1977. Evidence for the existence of three primary strategies in plants and its
relevance to ecological and evolutionary theory. Am. Nat. 111, 1169-1195.

147
Hamilton et al. 1996 Hamilton, S. K., Sippel, S. J., Melack, J. M. 1996. Inundation
patterns in the Pantanal wetland of South America determined form passive microwave
remote sensing. Archiv Hyd. 137: 1-23
Harris, M. B.; Arcangelo, C.; Pinto, E.C.T.; Camargo, G.; Ramos Neto, M.B.; Silva,
S.M. 2005. Estimativas de perda da área natural da Bacia do Alto Paraguai e Pantanal
Brasileiro. Relatório Técnico não publicado. Consevação Internacional, Campo Grande,
MS.
Jacobs, B. F., Kingston, J. D and Jacobs, L. L. 1999. The Origin of Grass-Dominated
Ecosystems. Annals of the Missouri Botanical Garden 86(2):590-643.
Junk, W. J., Nunes da Cunha, C. N., Wantzen, K. M., Petermann, P., strüssmann, C.,
marques, M. I. & Adis, J. 2006. Biodiversity and its conservation in the Pantanal of
Mato Grosso, Brazil. Aquatic Science 68:278–309.
Killeen, T. J. and Hinz, P. N. 1992a. Grasses of the Precambrian Shield Region in
Eastern Lowland Bolivia. I. Habitat Preferences. Journal of Tropical Ecology 8:389-
407.
Killeen, T. J. and Hinz, P. N. 1992b. Grasses of the Precambrian Shield region in
eastern lowland Bolivia. II. Life-formas and C3-C4 photosynthetic types. Journal of
Tropical Ecology 8:409-433.
Kellogg, E. A. 2001. Evolutionary History of the Grasses. Plant Physiology 125: 1198–
1205.
Klink, C. A. and Joly, C. A. 1989. Identification and distribution of C3 and C4 grasses
in open and shaded habitats in São Paulo State, Brazil. Biotropica 21(1):30-34.
Köppen, G. W. 1948. Climatologia. Fundo de Cultura Econômica. México, p. 242-248.

148
Lenssen, J., Menting, F., van der Putten, W. and Blom , K. 1999. Control of plant
species richness and zonation of functional groups along a freshwater flooding gradient.
Oikos 86(3):523-534.
Medina, E . & Motta, N . 1990. Metabolism and distribution of grasses in tropical
flooded savannas in Venezuela. Journal of Tropical Ecology 6: 77-89.
Meira-Neto, J. A. A., Martins, F. R. Valente, J. E. 2007. Composição florística e
espectro biológico na estação ecológica de santa bárbara, estado de São Paulo, Brasil.
Revista Árvore 31(5):907-922.
Menaut, J.C. 1983. The vegetation of African savannas. Pp.109-149. In: F. Bourliére
(ed.). Ecosystems of the Word: Tropical Savannas. Amsterdam, Elsevier
Mazza, M. C. M., Mazza, C. A. S., Sereno, J. R. B., Santos, S. A. and Pellegrin, A. O.
1994. Etnobiologia e conservação do bovino Pantaneiro. Corumbá: EMBRAPA- CPAP;
Brasília: EMBRAPA- SPI. 61p.
Nunes, A. P. &. Tomas, W. M 2008. Aves migratórias e nômades ocorrentes no
Pantanal. Empresa Brasileira de Pesquisa Agropecuária (Embrapa Pantanal), Corumbá.
24p.
Nunes da Cunha, C.; Rawiel, C. P.; Wantzen, K. M.; Junk, W. J. & Lemes do Prado, A.
2006. Mapping and characterization of vegetation units by means of Landsat imagery
and management recommendations for the Pantanal of Mato Grosso (Brazil), north of
Poconé. Amazoniana XIX(1):1-32.
Nunes da Cunha, C. and Junk, W.J. 2004. Year-to-year changes in water level drive the
invasion of Vochysia divergens in Pantanal grasslands. Applied Vegetation Science
7:103-110.
Nunes da Cunha, C. & Junk, W.J. 2001. Distribuition of Wood Plant Communities along
the Flood Gradient in the Pantanal of Poconé, Mato Grosso, Brazil. International Journal
of Ecology and Environmental 27:63-70.

149
Pinder, L. and Rosso, S. 1998. Classification and ordination of plant formations in the
Pantanal of Brazil. Plant Ecology 136:151–165.
Prado, A. L., Heckman, C. W. and Martins, F. R. 1994. The seasonal succession of
biotic communities in wetlands of the tropical wet-and-dry climatic zone: II. The
Aquatic Macrophyte Vegetation in the Pantanal of Mato Grosso, Brazil. Internationale
Revue gesamten Hydrobiologie 79(4): 569-589.
Pott, A. e Pott, V. J. 1999. Flora do Pantanal -Listagem Atual de Fanerógamas. Pp. 297-
325. In: Simpósio sobre Recursos Naturais e Sócio-Econômicos do Pantanal: Manejo e
Conservação. Empresa Brasileira de Pesquisa Agropecuária (Embrapa-CPAP),
Corumbá. 537p.
.
Pott, A. & Adámoli, J. 1999. Unidades de vegetação do Pantanal dos Paiaguás. Pp. 183-
202. In: Anais do II Simpósio sobre Recursos Naturais e Sócio-econômicos do Pantanal,
Manejo e Conservação. Empresa Brasileira de Pesquisa Agropecuária (Embrapa-
CPAP), Corumbá. 537p.
Pott, A. 1988. Pastagens no Pantanal. Empresa Brasileira de Pesquisa Agropecuária
(Embrapa-CPAP), Corumbá, documentos 7. 58p.
Pott, A. 1986. Pastagens no Pantanal. Pp. 413-421. In: Anais do Congresso Brasileiro e
Pastagens e 8o Simpósio Sobre Manejo de Pastagens. Piracicaba, Publicação Fundação
de Estudos Agrários Luiz de Queirós - ESALQ.
Raunkier, C. 1934. The life forms of plants and statistical plant geography. Clarendon
Press, Oxford. 200p.
Rebellato, L. e Nunes da Cunha, C. 2005. Efeito do “fluxo sazonal mínimo da
inundação” sobre a composição e estrutura de um campo inundável no Pantanal de
Poconé, MT, Brasil. Acta Botanica Brasilica 19: 789-799.

150
Robertson, P. A., Mackenzie, M. D. & Elliot, L. F. 1984. Gradient analysis and
classification of the woody vegetation for 4 sites in southern Illinois and adjacent
Missouri. Vegetatio 58: 87–104.
Santos, S. A., Crispim, S. M. A., Comastri-Filho, J. A., Pott, A. e Cardoso, E. L. 2005.
Substituição de pastagens nativas de baixo valor nutritivo por forrageiras de melhor
qualidade no Pantanal. Empresa Brasileira de Pesquisa Agropecuária (Embrapa-CPAP),
Circular técnica 62. 5p.
Santos, S. A., Costa, C., Chalita, L. V. A. S., Pott, A., Alvarez, J. M. and Ortiz, A. G.
2001. Grazing behavior and activity patterns by free-grazing cattle in the Pantanal
region. Proceendings of the XIX International Grasslands Congress.
Sarmiento, G. 1992. Adaptive strategies of perennial grasses in South American
savannas. Journal of Vegetation Science 3: 325-336.
Sarmiento, G. 1983. The savannas of Tropical America. Pp.245-288. In: Ecosystems of
the Word: Tropical Savannas. F. Bourliére (ed.). Amsterdam, Elsevier. 730p.
Sarmiento, G. and Monastério, M. 1983. Life forms and fenology. Pp.79-108. In:
Ecosystems of the Word: Tropical Savannas. F. Bourliére (ed.). Elsevier. Amsterdam.
730p.
Schessl, M. 1999. Floristic composition and structure of floodplain vegetation in
Northern Pantanal of Mato Grosso, Brasil. Phyton (Horn, Austria) 39(2):303-3.
Shantz, H. L. 1954. The place of grasslands in the earth's cover. Ecology 35(2): 143-
145.
Silva, I. A. and Batalha, M. A. 2008. Species convergence into life-forms in a
hyperseasonal cerrado in central Brazil. Brazilian Journal of Biology 68: 631-637.
Silva, M. P., Mauro, R., Mourão, G. and Coutinho, M. 1999. Conversion of forests and
woodlands to cultivated pastures in the wetland of Brazil. Ecotropicos 12(2):101-108.

151
Silva, J. S. V. e Abdon, M. M. 1998. Delimitação do Pantanal brasileiro e suas sub-
regiões. Pesquisa agropecuária brasileira, Brasília-DF 33: 1703-1711.
Southal, E. J., Dale, M. P. and Kent, M. 2003. Spatial and temporal analysis of
vegetation mosaics for conservation: poor fen communities in a Cornish valley mire.
Journal of Biogeography. 30:1427–1443.
Townsend, P. A. 2001. Relationships between vegetation patterns and hydroperiod on
the roanoke river floodplain, North Carolina. Plant Ecology 156(1): 43-58.
Valls, J.F.M. 1994. O espectro taxonômico das gramíneas do Pantanal. Pp.227-237. In:
Utilizacion y manejo de pastizales. J. P. Puignon (ed.). Montivideo: IICA-PROCISUR
(IICA - PROCISUR. Dialogo, 40).
Wilson, M. V. & A. Shmida. 1984. Measuring beta diversity with presence-absence
data. Journal of Ecology 72: 1055–1064.
Zeilhofer, P. & Schessl, M. 1999. Relationship between vegetation and environmental
conditions in the northern Pantanal of Mato Grosso, Brazil. Journal of Biogeography
27: 59-168.
Zuloaga, F.O. 1986. Systematics of New World Species of Panicum (Poaceae:
Paniceae). Pp. 287-306. In: Grass Systematics and Evolution. T. R. Soderstroms (ed.).
Washington: Smithsonian Institution Press.
152
ANEXO
(Capítulo 3)
Tabela – Sub-tipos de vegetação ocorrentes nas 30 isolinhas, duração da inundação (dias), profundidade da inundação (cm), classes de profundidade (9 cm), das 30 isolinhas amostradas, Pantanal de Poconé, Mato Grosso.
isolinhas Sub-tipos de vegetação Duração
(dias) Profundidade
(cm)
Classes de
profundidade
A0500 Floresta semi-sempreverde sazonalmente inundável
111 30 4
A1500 Floresta semi-sempreverde sazonalmente inundável
104 42 5
A2500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 17 2
A3500 Savana sazonalmente inundável com térmitas (campo de murundus)
104 21 3
A4500 Savana sazonalmente inundável com térmitas (campo de murundus)
104 25 3
B0500 Sistema secundário (campo exótico) 42 15 2
B1500 Interface: campo exótico/floresta sazonal semi-decídua sobre solo seco (cerradão)
0 0 1
B2500 Floresta semi-sempreverde sazonalmente inundável
154 50 6
B3500
Interface: Savana sazonalmente inundável com térmitas (campo de murundus)/ floresta sazonal semi-decídua sobre solo seco (cerradão)
6 7 1
B4500 Sistema secundário (campo exótico) 56 16 2
C0500 Sistema secundário (campo exótico) 89 23 3
C1500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 16 2
C2500 Savana sazonalmente inundável com térmitas (campo de murundus)
104 25 3
C3500 Savana sazonalmente inundável com térmitas (campo de murundus)
110 29 4
C4500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 14 2
D0500 Sistema secundário (campo exótico) 200 58 6
D1500 Campo limpo sazonalmente inundável 200 63 6
D2500 Floresta semi-sempreverde sazonalmente inundável
126 35 4
D3500 Floresta semi-sempreverde sazonalmente inundável
90 16 2
D4500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 26 3
E0500 Savana sazonalmente inundável com térmitas (campo de murundus)
125 44 5
E1500 floresta sazonal semi-decídua sobre solo seco (cerradão)
0 0 2
E2500 Interface: campo limpo sazonalmente inundável/ floresta sazonal semi-decídua sobre solo seco (cerradão)
90 26 3

153
E3500
Interface: Savana sazonalmente inundável com térmitas (campo de murundus)/ floresta sazonal semi-decídua sobre solo seco (cerradão)
23 7 1
E4500 Floresta semi-sempreverde sazonalmente inundável
125 39 5
F0500 Savana sazonalmente inundável com térmitas (campo de murundus)
95 42 5
F1500 floresta sazonal semi-decídua sobre solo seco (cerradão)
0 0 1
F2500 Sistema secundário (campo exótico) 110 18 2
F3500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 24 3
F4500 Savana sazonalmente inundável com térmitas (campo de murundus)
90 24 3

154