
recuperação de tório e terras raras via peróxido do resíduo ...

1
Artigo original/Original Article
Produção de peróxido de hidrogênio e atividade de enzimas antioxidantes de macrófagos peritoneais em resposta ao BCG (Bacilo de Calmette-guérin) em modelo murínico de desnutrição proteico-energéticaHydrogen peroxide production and activity of antioxidant enzymes of peritoneal macrophages in response to BCG (Bacillus Calmette-guérin) in a murine model of protein-energy malnutrition
OSMARINA DA PAIXÃO E SILVA1; RICARDO AMBRÓSIO FOCK2;
MARCELO MACEDO ROGERO3; PRIMAVERA
BORELLI21Universidade Federal do
Pará, Instituto de Ciências da Saúde, Faculdade
de Farmácia.2Universidade de São Paulo,
Faculdade de Ciências Farmacêuticas.
3Universidade de São Paulo, Faculdade
de Saúde Pública.Endereço para
correspondência:Primavera Borelli,
Laboratório de Hematologia Experimental, Departamento
de Análises Clínicas e Toxicológicas, Faculdade
de Ciências Farmacêuticas, Universidade de São Paulo,
Avenida Prof. Lineu Prestes, 580. São Paulo, SP
CEP 05508-900 Brasil.E-mail: [email protected] ou
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Hydrogen peroxide production and activity of antioxidant enzymes of peritoneal macrophages in response to BCG (Bacillus Calmette-guérin) in a murine model of protein-energy malnutrition. Nutrire: rev. Soc. Bras. Alim. Nutr. = J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
Protein-energy malnutrition (PEM) modifies resistance to infection, impairing a number of physiological processes, changing specifi c and nonspecifi c immune responses. Macrophages, which are directly involved in several aspects of immunity, may have their functions altered in the malnourishment condition, possibly playing a signifi cant role in the immune defi ciency observed in malnourished individuals. Two-month-old male Swiss mice were induced to PEM with a low-protein diet containing 4% protein as compared to 20% protein in the control diet. When the experimental group had lost about 20% of their original body weight the animals from both groups received intraperitoneal injections of 107 units of BCG (Bacillus Calmette-guérin) and after 7 days they were used in the experiments. We evaluated the complete blood count, peritoneal cellularity and hydrogen peroxide production, besides the activities of glutathione peroxidase, superoxide dismutase and catalase in peritoneal macrophages stimulated with BCG. Malnourished animals presented anemia, leukopenia and severe reduction of peritoneal cellularity. The production of hydrogen peroxide and the activity of glutathione peroxidase, superoxide dismutase and catalase were found to be signifi cantly lower in macrophages from malnourished animals. These fi ndings suggest that malnourished animals present a defi cient response to BCG. These fi ndings may be partly responsible for a decrease in the bactericidal and fungicidal activities observed in the malnourished mice. These data lead us to infer that the nutritional status interferes with the activation of macrophages and with the capacity to mount an immune response.
Keywords: Protein-Energy Malnutrition. Macrophages. Hydrogen Peroxide. Glutathione Peroxidase. Superoxide Dismutase. Catalase.
ABSTRACT
_13961e-NUTRIRE v36 n3.indd 1_13961e-NUTRIRE v36 n3.indd 1 02/02/12 16:2902/02/12 16:29

2
RESUMORESUMEN
La desnutrición proteico-energética modifi ca la resistencia a infecciones, alterando diversos procesos fi siológicos, mudando la capacidad de respuesta inmune, específi ca y no específi ca. Los macrófagos, células implicadas directamente en varios aspectos de la inmunidad, pueden tener sus funciones alteradas en condiciones de desnutrición desempeñando posiblemente un papel signifi cativo en la defi ciencia inmune observada en individuos desnutridos. En este estudio se utilizaron ratos Swiss machos, de 2 meses de edad, en los cuales fue inducida desnutrición proteico-energética por medio de una dieta que contenía 4% de proteína. El grupo control recibió una dieta estándar con 20% de proteína. Cuando el grupo experimental presentó una pérdida de 20% de su peso corporal original, se le administraron 107 unidades de BCG (Bacilo de Calmette-guérin), por vía intraperitoneal, y después de 7 días los animales fueron sacrifi cados para evaluación de diversos índices: hemograma, celularidad peritoneal, la producción de peróxido de hidrógeno y las actividades glutatión peroxidada, superóxido dismutasa, y catalasa en macrófagos peritoneales estimulados con BCG. Los animales subalimentados presentaron anemia, leucopenia y reducción de la celularidad peritoneal. La producción de peróxido de hidrógeno y la actividad de las enzimas glutatión peroxidada, superóxido dismutasa, y catalasa fue más baja en macrófagos de animales desnutridos. Los resultados sugieren que los ratos desnutridos presentan una respuesta defi ciente a BCG lo que explica en parte la disminución de la actividad bactericida y fungicida observada en animales desnutridos. Estos resultados permiten deducir que el estado nutricional interfi ere en la actividad de los macrófagos y en su capacidad de respuesta inmunológica.
Palabras clave: Desnutrición proteico-calórica. Macrófagos. Peróxido de hidrógeno. Glutatión peroxidada. Superóxido dismutasa. Catalasa.
A desnutrição proteico-energética (DPE) altera a capacidade de resistência à infecção, alterando diversos processos fisiológicos, mudando a capacidade de respostas imunes específi cas e não específi cas. Os macrófagos são células envolvidas diretamente em diversos aspectos da imunidade, podem ter suas funções alteradas em condições de desnutrição desempenhando, possivelmente, um papel signifi cativo na imunodefi ciência observada nesses indivíduos. Camundongos Swiss, machos, de dois meses de idade, foram induzidos a DPE com uma dieta contendo baixa concentração de proteína (4%) em comparação à dieta controle (20%). Quando o grupo experimental perdeu aproximadamente 20% de seu peso corpóreo original, estes foram considerados aptos aos experimentos. Animais de ambos os grupos receberam injeção intraperitoneal contendo 107 unidades de BCG (Bacilo de Calmette-guérin) e após 7 dias foram sacrificados e utilizados nos experimentos. Avaliamos o hemograma, a celularidade peritoneal assim como a produção de peróxido de hidrogênio e a atividade da glutationa peroxidase, superóxido dismutase e catalase em resposta ao BCG em macrófagos peritoneais. Os animais desnutridos apresentaram anemia, leucopenia e a redução severa da celularidade peritoneal. A produção de peróxido de hidrogênio e a atividade das enzimas glutationa peroxidase, superóxido dismutase e catalase foi signifi cativamente menor nos macrófagos de animais desnutridos. Estes resultados sugerem que os animais desnutridos apresentem uma resposta defi ciente ao BCG, e que, em parte, podem explicar a diminuição nas atividades bactericidas e fungicidas observadas em animais desnutridos. Estes dados permitem deduzir que o estado nutricional interfere na ativação dos macrófagos e na sua capacidade de resposta imune.
Palavras-chave: Desnutrição proteico-calórica. Macrófagos. Peróxido de hidrogênio. Glutationa peroxidase. Superóxido dismutase. Catalase.
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 2_13961e-NUTRIRE v36 n3.indd 2 02/02/12 16:2902/02/12 16:29

3
INTRODUÇÃO
Estudos clínicos e epidemiológicos demonstram a associação entre desnutrição
e infecção. A DPE modifi ca os mecanismos específi cos e inespecífi cos de resistência
do organismo, modifi cando a resposta infl amatória. As relações entre desnutrição e
resposta inflamatória são complexas e ainda não estão totalmente esclarecidas
(CHANDRA, 2002; KEUSCH, 2003; SCRIMSHAW, 2010). Dados da literatura e do nosso
grupo evidenciam que a DPE compromete a migração celular, o espraiamento, a
fagocitose, a atividade bactericida e fungicida, bem como alterações na produção de
interleucina (IL)-1α, IL-6 e fator de necrose tumoral (TNF)-α (BORELLI et al., 1998;
FOCK et al., 2007).
Os macrófagos são classifi cados de acordo com o seu estado de ativação: em
residentes, infl amatórios ou ativados. Os macrófagos residentes representam células
teciduais que não foram expostas a substâncias estranhas, apresentando discreta atividade
funcional como, por exemplo, reduzida taxa de síntese de espécies reativas de oxigênio.
Quando macrófagos residentes são expostos à administração intraperitoneal de BCG,
estas células apresentam taxas elevadas de síntese de citocinas como IL-1 e TNF-α,
de ânion superóxido (O2•–) e peróxido de hidrogênio (H2O2), aumento da atividade
fagocítica, microbicida e tumoricida, atividades estas que caracterizam o estado ativado
dos macrófagos (BELLOSTA; BERNINI, 2005; LI et al., 2009).
Na eliminação de patógenos, por macrófagos, estão envolvidos processos
não oxidativos, que envolvem secreção de proteínas catiônicas com propriedades
antibiótico-like (defensina, catepsina G), lisozima, colagenase, elastase e proteínas
ácidas, e processos oxidativos, como a liberação de espécies reativas de nitrogênio
como óxido nítrico (NO) e de oxigênio como O2•–, H2O2, radical hidroxil (•OH) e
oxigênio singlete (1O2), eventos estes denominados de ‘burst” oxidativo (SCHLESINGER;
HORWITZ, 1991; YAMADA et al., 1987).
Para que esses eventos se iniciem, faz-se necessária à ativação do complexo
enzimático da NADPH oxidase. Este complexo catalisa a redução do oxigênio para
ânion superóxido usando NADPH proveniente da via das pentoses como doador
de elétrons. O O2•– produzido pode dismutar-se espontaneamente ou pela ação da
enzima superóxido dismutase, produzindo H2O2 e oxigênio (BABIOR, 1984). O H2O2
desempenha função fundamental no processo de morte microbiana e, por ser um forte
oxidante é capaz de degradar biomoléculas tanto in vivo como in vitro com potente
ação bacteriostática e bactericida (BALDECK; MARQUIS, 2008). Além disso, H2O2
pode gerar outros produtos de poder microbicida ainda mais potente como o •OH ou
formando hipohaleto oxidante(BABIOR, 1984; HALLIWELL, 1994).
Por outro lado, as células possuem um sistema de defesa antioxidante que
interage e inibe a ação da maioria dos radicais livres gerados. Em indivíduos saudáveis,
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 3_13961e-NUTRIRE v36 n3.indd 3 02/02/12 16:2902/02/12 16:29
4
esses radicais livres são controlados pela ação de enzimas antioxidantes (glutationa
peroxidase, superóxido dismutase e catalase) e por antioxidantes não enzimáticos
(ascorbato, tocoferóis e uratos) (SIES, 1993). Se houver um desequilíbrio neste sistema
podem ocorrer danos aos tecidos devido a alterações na integridade e funcionalidade
celular em função do estresse oxidativo (AERTS et al., 1995).
O objetivo deste trabalho foi avaliar a capacidade de produção de peróxido de
hidrogênio e a atividade de enzimas antioxidantes (glutationa peroxidase, superóxido
dismutase e catalase), em macrófagos peritoneais de camundongos inoculados com
BCG e submetidos à DPE.
MATERIAL E MÉTODOS
RAÇÕES
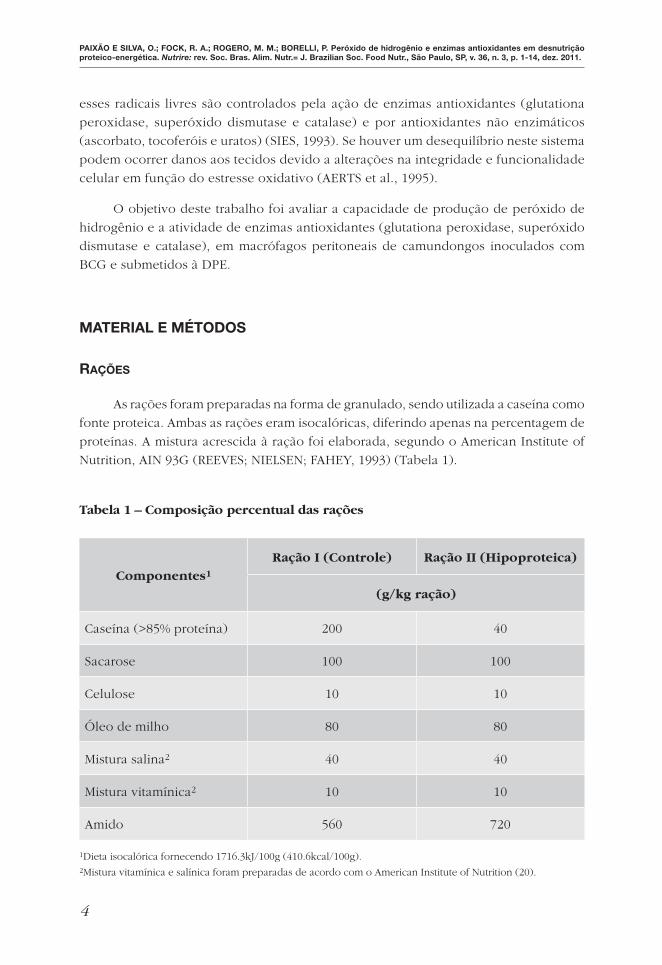
As rações foram preparadas na forma de granulado, sendo utilizada a caseína como
fonte proteica. Ambas as rações eram isocalóricas, diferindo apenas na percentagem de
proteínas. A mistura acrescida à ração foi elaborada, segundo o American Institute of
Nutrition, AIN 93G (REEVES; NIELSEN; FAHEY, 1993) (Tabela 1).
Tabela 1 – Composição percentual das rações
Componentes1
Ração I (Controle) Ração II (Hipoproteica)
(g/kg ração)
Caseína (>85% proteína) 200 40
Sacarose 100 100
Celulose 10 10
Óleo de milho 80 80
Mistura salina2 40 40
Mistura vitamínica2 10 10
Amido 560 720
1Dieta isocalórica fornecendo 1716.3kJ/100g (410.6kcal/100g).2Mistura vitamínica e salínica foram preparadas de acordo com o American Institute of Nutrition (20).
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 4_13961e-NUTRIRE v36 n3.indd 4 02/02/12 16:2902/02/12 16:29

5
ANIMAIS
Foram utilizados camundongos Swiss, machos, de 2 a 3 meses de idade, obtidos do
Biotério de Produção e Experimentação do Conjunto das Químicas da Universidade de São
Paulo. Os animais foram alocados em gaioleiros metabólicos, recebendo, ad libitum, ração
comercial e água durante o período de adaptação (21 dias), sendo mantidos sob temperatura
de 22-25°C e ciclo de luz de 12 horas (claro/escuro). Ao fi nal desse período, os animais
foram distribuídos em quatro grupos: controle (C), controle + BCG (C+BCG), desnutrido
(D) e desnutrido + BCG (D+BCG), sendo os grupos C e C+BCG alimentados com ração
contendo 20% de proteína (ração I), enquanto os grupos D e D+BCG foram alimentados
com ração contendo 4% de proteína (ração II) (Tabela 1). O peso corpóreo e o consumo
de ração foram determinados a cada 48 horas. A avaliação nutricional foi realizada pelo
controle do peso corpóreo, consumo de ração e determinação da concentração plasmática
de proteínas totais e de albumina. Este estudo foi aprovado pela Comissão de Ética no
Uso de Animais da Faculdade de Ciências Farmacêuticas da Universidade de São Paulo.
INDUÇÃO DA INFLAMAÇÃO
Quinze dias após o início do protocolo experimental de desnutrição os animais
dos grupos C+BCG e D+BCG foram inoculados, intraperitonealmente (i.p.), com 1mL
de suspensão de BCG contendo 107 unidades viáveis de bacilos. Os animais foram
sacrifi cados 7 dias após a administração de BCG para a coleta de macrófagos peritoneais
ativados. Os animais do grupo D+BCG apresentavam redução de aproximadamente 15%
do peso corpóreo no momento da inoculação do BCG.
DETERMINAÇÃO DO HEMOGRAMA E DAS PROTEÍNAS PLASMÁTICAS
Os animais foram anestesiados com Xilazina e Quetamina na dose de 50mg/Kg
de massa corporal e o sangue total foi obtido por punção cardíaca. Amostras coletadas
em tubos com o anticoagulante EDTA na concentração fi nal de 1mg/mL de sangue
(EDTA, Sigma Chemical Company, St. Louis, MO), foram utilizados para a realização
do hemograma empregando-se as metodologias hematológicas padrão (DACIE; LEWIS,
1995). Os leucócitos e eritrócitos foram contados em câmara de Neubauer e a análise
diferencial realizada em extensões sanguíneas coradas por soluções de May-Grunwald e
Giemsa (Sigma Chemical Co., St. Louis, MO). O plasma foi separado por centrifugação e
a dosagem de proteínas totais e albumina foram determinadas por metodologia padrão
(DOUMAS; WATSON; BIGGS, 1971; GORNALL; BARDAWILL; DAVID, 1949).
OBTENÇÃO DE MACRÓFAGOS PERITONEAIS
Após sete dias do estímulo inicial com BCG, animais de todos os quatro grupos
(C, D, C+BCG e D+BCG) foram anestesiados, exanguinados e a cavidade peritoneal
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 5_13961e-NUTRIRE v36 n3.indd 5 02/02/12 16:2902/02/12 16:29

6
lavada com 5mL de meio de cultura McCoy’s 5A (Sigma®), pH 7,4, estéril, aspirando-
se a seguir, o líquido peritoneal, o qual foi acondicionado em tubos plásticos, estéreis
e mantidos em banho de gelo até a realização dos ensaios, no prazo máximo de
30 minutos. A contagem das células presentes no lavado peritoneal foi realizada
em câmara de Neubauer e a viabilidade avaliada com solução de Azul de Trypan (0,1%).
A identifi cação dos tipos celulares foi realizada em lâminas obtidas por citocentrifugação
de alíquotas da suspensão celular, coradas pelas técnicas de May-Grunwald e Giemsa
e esterase inespecífi ca.
PRODUÇÃO DE PERÓXIDO DE HIDROGÊNIO
Para avaliação da produção de peróxido de hidrogênio em macrófagos peritoneais
ativados foi utilizado o método de Pick e Mizel (1981). Em 1mL de solução contendo
NaCl 140mM (Merck Chemical® S.A), KH2PO4 1mM (Merck Chemical®, Alemanha),
Glicose 5mM (Merck Chemical®, Alemanha), 0,56mM vermelho de fenol (Merck
Chemical®, Alemanha), 20UI/mL solução de peroxidase raíz forte tipo VI (Sigma Chemical
Company®, EUA) e de superóxido dismutase 2mg/mL (Sigma Chemical Company®,
EUA) adicionou-se 1x106 macrófagos peritoneais. Paralelamente, foi feita uma curva
padrão (0-7µL) de peróxido de hidrogênio (Honeywell Riedel-de Haën®, Alemanha)
1mM. As amostras foram incubadas em estufa a 37°C, com atmosfera de 5% de CO2 por
1 hora. Após esse período, a reação foi interrompida pela adição em todos os poços de
10µL de NaOH 1M (Merck Chemical®, Alemanha). O total de peróxido de hidrogênio
formado foi quantifi cado espectrofotometricamente em leitor de ELISA (SLT Spectra®,
SLT Instruments, Áustria) em fi ltro com comprimento de onda de 620nm. As amostras
foram realizadas em quadruplicata.
ATIVIDADE DA GLUTATIONA PEROXIDASE
Foi utilizado o método descrito por Wendel (1981), para a avaliação da atividade
da glutationa peroxidase em macrófagos peritoneais ativados. Em 1mL de solução
contendo fosfato de sódio 143mM (Merck Chemical®, Alemanha), EDTA 6,3mM (Merck
Chemical®, Alemanha), NaN3 0,25M (Merck Chemical®, Alemanha), NADPH 0,12mM
(Merck Chemical®, Alemanha), Glutationa redutase (GSH-R) 0,25U/mL (Sigma Chemical
Company®, EUA), Glutationa (GSH) 1mM (Sigma Chemical Company®, EUA), tert-Butil
Hidroperóxido (TBH) 1,2mM (Merck Chemical®, Alemanha) adicionou-se 1x106 de
macrófagos peritoneais previamente lisados por sonicação em um sonicador Heat System
equipado com microponta, por 6 ciclos de 20 segundos. As amostras foram incubadas
a 37°C durante 10 minutos para ativar a Glutationa peroxidase (GSH-Px) com a GSH. A
reação foi iniciada pela adição de TBH 1,2mM e o consumo de NADPH foi monitorado
durante 6 minutos em leitor de ELISA (SLT Spectra®, SLT Instruments, Áustria). As amostras
foram processadas em triplicata. Para os cálculos da atividade enzimática foram utilizados
o valor de 6,22x103 M-1 cm-1 como coefi ciente de extinção molar.
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 6_13961e-NUTRIRE v36 n3.indd 6 02/02/12 16:2902/02/12 16:29

7
ATIVIDADE DA SUPERÓXIDO DISMUTASE
Ao avaliar a atividade da superóxido dismutase em macrófagos peritoneais ativados
foi utilizado o método de McCord e Fridovich (1969). Em 1mL de solução contendo fosfato
de sódio 50mM (Merck Chemical®, Alemanha), EDTA 1mM (Merck Chemical®, Alemanha),
xantina sódica 500mM (Sigma Chemical Company®, EUA), citocromo C 1µM (Sigma
Chemical Company®, EUA) adicionou-se 1x106 macrófagos peritoneais previamente
lisados por sonicação em um sonicador Heat System equipado com microponta, por 6
ciclos de 20 segundos. A atividade da enzima superóxido dismutase foi determinada pelo
consumo de citocromo C em leitor de ELISA (SLT Spectra®, SLT Instruments, Áustria).
As amostras foram processadas em triplicata. A atividade enzimática foi calculada,
considerando-se que 1 unidade da enzima é capaz de produzir uma inibição de 50%
da reação.
ATIVIDADE DA CATALASE
Para avaliação da atividade catalase a partir de macrófagos peritoneais ativados, foi
utilizado o método descrito por Volkovová et al. (1996). Em 1mL de solução contendo
tampão Tris-HCl 1 M (Merck Chemical®, Alemanha), EDTA 5mM (Merck Chemical®,
Alemanha), e H2O2 10mM, pH 8,0 adicionou-se 1x106 macrófagos peritoneais previamente
lisados por sonicação em um sonicador Heat System equipado com microponta, por
6 ciclos de 20 segundos. O consumo de H2O2 avaliado espectrofotometricamente em
leitor de ELISA (SLT Spectra®, SLT Instruments, Áustria) a 230nm a 30°C. As amostras
foram processadas em triplicata. Os cálculos foram baseados no coefi ciente de extinção
molar da água oxigenada 0,071 M-1 cm-1.
ANÁLISE ESTATÍSTICA
Os resultados foram submetidos à análise estatística (one-way ANOVA, p ≤ 0,05) e
comparados pelo teste múltiplo de Tukey, utilizando-se o programa estatístico Graphpad
Prism®. Os dados foram expressos como média e desvio padrão.
RESULTADOS
CONSUMO DE RAÇÃO, PESO CORPÓREO E PROTEÍNAS PLASMÁTICAS
Os animais dos grupos D e D+BCG apresentaram signifi cativa redução do consumo
de ração quando comparados com os animais controles (Tabela 2). Os animais do grupo
D apresentaram perda de peso corpóreo, sendo esta perda mais acentuada nos animais
D+BCG. Por outro lado, os animais dos grupos C e C+BCG ganharam peso ao longo
ensaio (Tabela 2).
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 7_13961e-NUTRIRE v36 n3.indd 7 02/02/12 16:2902/02/12 16:29

8
Tabela 2 – Consumo de proteína, variação de peso corpóreo, dosagem de proteínas e albumina plasmática, hemograma e contagem de células do exsudato peritoneal de camundongos controles (C) e Desnutridos (D) inoculados ou não com BCG
ParâmetrosC
(n=9)
C + BCG
(n=11)
D
(n=10)
D + BCG
(n=12)
Consumo de ração(g/dia/animal)
3,70 ± 0,30a 3,40 ± 0,31a 2,75 ± 0,20b 2,25 ± 0,20b
Consumo de proteína(g/dia/animal)
0,74 ± 0,11a 0,68 ± 0,13a 0,11± 0,05b 0,09 ± 0,04b
Variação de peso corpóreo (%)
8,57 ± 1,01a 8,28 ± 0,19a -13,4 ± 2,01b -37,5 ± 2,30c
Proteína plasmática(g/dL)
7,08 ± 1,10a 6,91± 1,80a 4,30 ± 0,81b 3,70± 1,01b
Albumina (g/dL) 3,91± 0,22a 3,30 ± 0,41a 2,42 ± 0,41b 2,10 ± 0,22b
Hemoglobina (g/dL) 11,41 ± 0,74a 10,11 ± 1,62a 6,22 ± 0,72b 5,11 ± 0,83b
Eritrócitos(x 106/mm3)
7,11 ±0,71a 6,90± 0,60a 5,43± 0,22b 5,12 ± 0,21b
Leucócitos totais (/mm3)
3245 ± 227a 12980 ± 2230b 1470 ± 65c 2130 ± 120d
Polimorfonucleares(/mm3)
621,1 ± 15,1a 9201,2 ± 1721,1b 186,2 ± 4,7c 301,1 ± 12,0d
Linfócitos (/mm3) 2410 ± 75a 4210 ± 1012b 1370± 40c 1407 ± 65d
Monócitos (/mm3) 91,1 ± 4,4a 580,0 ± 130,1b 10,10± 0,51c 8,11± 0,52c
Células totais peritoneais(x 106/mL)
2,70 ± 0,43a 3,92 ± 0,81b 1,61 ± 0,32c 1,91 ± 0,52a,c
Macrófagos peritoneais(x 106/mL)
2,01± 0,42a 3,31 ± 0,71b 1,22± 0,22c 1,70 ± 0,33a,c
Granulócitos peritoneais (x 106/mL)
0,10± 0,04a 0,40± 0,09b 0,05 ± 0,03a 0,20 ± 0,09a
Mastócitos peritoneais (x 106/mL)
0,10 ± 0,01 0,10 ± 0,02 0,04 ± 0,02 0,06 ± 0,02
Valores da média ± o desvio padrão. Médias com letras sobrescritas diferentes indicam diferença estatisticamente signifi cativa entre os grupos (p≤0,05). n = número de animais avaliados por grupo.
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 8_13961e-NUTRIRE v36 n3.indd 8 02/02/12 16:2902/02/12 16:29

9
HEMOGRAMA
Os animais dos grupos D e D+BCG apresentaram anemia com diminuição da
concentração de hemoglobina e do número de eritrócitos. Os animais do grupo D
apresentaram leucopenia com depleção do número de polimorfonucleares, linfócitos
e monócitos quando comparados aos respectivos controles (Tabela 2). Apesar de
os animais do grupo D+BCG apresentar discreto aumento do número de leucócitos
circulantes, esse aumento não foi tão evidente como nos animais do grupo C+BCG que
apresentaram acentuada leucocitose acompanhada de aumento do número de células
polimorfonucleares.
CELULARIDADE DO EXSUDATO PERITONEAL
Observamos signifi cativa redução da celularidade no exsudato peritoneal dos
animais do grupo D em relação aos animais do grupo C e C+BCG (Tabela 2). Animais
do grupo C+BCG apresentaram aumento do número de células no exsudato peritoneal
com aumento do número de células macrofágicas quando comparado com os demais
grupos. A comparação entre o número de células no exsudato peritoneal dos grupos D
e D+BCG não apresentou diferenças signifi cativas.
PRODUÇÃO DE PERÓXIDO DE HIDROGÊNIO
Os macrófagos peritoneais obtidos de animais do grupo C+BCG apresentaram
maior produção de peróxido de hidrogênio quando comparados ao grupo D+BCG. Não
observamos diferenças estatisticamente signifi cativas na produção de H2O2 quando
comparamos os grupos C e D (Tabela 3).
Tabela 3 – Produção de peróxido de hidrogênio e atividade das enzimas antioxidantes de células do exsudato peritoneal de camundongos controles (C) e desnutridos (D) inoculados ou não com BCG
ParâmetrosC
(n=9)C + BCG(n=11)
D(n=10)
D + BCG(n=12)
Produção de peróxido de hidrogênio (nM/1x106 células/mL)
0,32± 0,02a 1,72± 0,04b 0,25 ± 0,05a 0,85 ± 0,05c
Atividade da glutationa peroxidase (U/1x106 células/mL)
2,64 ± 0,07a 2,44 ± 0,08a 2,11 ± 0,09b 1,92 ± 0,07b
Atividade da superóxido dismutase (U/1x106 células/mL)
2,70 ± 0,07a 2,53 ± 0,08a 2,23 ± 0,07b 1,85 ± 0,09c
Atividade da catalase(U/1x106 células/mL)
342,1 ± 9,1a 320,1 ± 8,1a 276,0 ± 9,3b 227,1 ± 7,1c
Valores da média ± o desvio padrão. Médias com letras sobrescritas diferentes indicam diferença estatisticamente signifi cativa entre os grupos (p≤0,05). n = número de animais avaliados por grupo.
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 9_13961e-NUTRIRE v36 n3.indd 9 02/02/12 16:2902/02/12 16:29

10
GLUTATIONA PEROXIDASE
A atividade da glutationa peroxidase em macrófagos peritoniais obtidos de animais
do grupo C e C+BCG, apresentou valores maiores quando comparada às dos grupos D
e D+BCG, respectivamente. A atividade da glutationa peroxidase, nos animais do grupo
D+BCG, foi signifi cativamente menor do que as dos grupos C e C+BCG (Tabela 3).
SUPERÓXIDO DISMUTASE
A atividade da superóxido dismutase em macrófagos peritoniais de animais dos
grupos C e C+BCG, apresentou valores maiores quando comparada às dos grupos D e
D+BCG, respectivamente. A comparação entre os grupos D e D+BCG também apresentou
diferenças estatisticamente signifi cativas (p≤0,05) (Tabela 3).
CATALASE
A atividade da catalase em macrófagos peritoniais dos animais do grupo C e
C+BCG apresentou valores maiores quando comparada às dos grupos D e D+BCG,
respectivamente. Estes apresentaram a menor atividade de catalase entre todos os grupos
estudados. Na comparação entre os grupos D e D+BCG também não houve diferenças
signifi cativas (Tabela 3).
DISCUSSÃO
Alterações na dieta e no estado nutricional podem infl uenciar a resposta infl amatória
e diminuir a capacidade de resposta a infecções. Neste estudo, avaliamos a capacidade
de produção de peróxido de hidrogênio e atividade de enzimas relacionadas ao sistema
antioxidante de macrófagos peritoneais de camundongos, inoculados ou não com BCG,
em um modelo de DPE.
A resposta infl amatória está intimamente relacionada com o processo de reparo
que, em geral, tem a fi nalidade de eliminar, diluir ou imobilizar o agente agressor
e, independente da sua duração, defl agra uma série de processos que, tanto quanto
possível, reconstituem o tecido lesado (CHANDRA, 2002; KEUSCH, 2003; SCRIMSHAW,
2010). Os estados carenciais alteram os mecanismos de defesa do indivíduo, sendo a
desnutrição ainda um importante problema em saúde à medida que observamos as
graves consequências da desnutrição sobre o organismo, como alterações metabólicas,
neurológicas e modifi cações do sistema linfo-hemopoético que podem induzir à maior
predisposição a agentes infecciosos (FOCK et al., 2007; FOCK et al., 2010; POWANDA;
BEISEL, 2003).
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 10_13961e-NUTRIRE v36 n3.indd 10 02/02/12 16:2902/02/12 16:29

11
Neste trabalho, os animais alimentados com ração hipoproteica consumiram menor
quantidade de ração e, portanto, de proteínas, em relação aos controles. A redução da
ingestão de ração hipoproteica pode ser uma tentativa de atenuar o balanço nitrogenado
negativo instalado no organismo. A anorexia observada, aparentemente paradoxal, pode
ser explicada por modifi cações hipotalâmicas induzidas provavelmente por citocinas
proinfl amatórias como TNF-α e IL-1 (FRAYN, 1986; SCRIMSHAW, 2003) e ainda por
alterações das concentrações plasmáticas de aminoácidos de cadeia ramifi cada, que
podem modifi car a ação de neurotransmissores, comportamento que poderia ser um
processo adaptativo, visando evitar maiores alterações no balanço nitrogenado (YOUNG;
MARCHINI, 1990).
Após a indução do processo infl amatório/infeccioso pela administração de BCG,
a redução do consumo de alimento tornou-se ainda mais evidente, possivelmente
devido à ativação de mecanismos fi siológicos adaptativos, como a redução da taxa
do metabolismo basal, utilização mais efi ciente de energia, redução na renovação de
proteínas celulares (CATCHATOURIAN; ECKERLING; FRIED, 1980). Também observamos
que animais submetidos a um estado de desnutrição apresentam anemia, dados esses
que vêm de encontro com outros dados do grupo, em que demonstram que essa anemia
é decorrente de uma defi ciência na produção de precursores hematopoéticos e não de
menor disponibilidade de ferro (BORELLI et al., 2007).
As mudanças tanto na imunidade celular como humoral são signifi cativamente
moduladas pelo estado nutricional (CHANDRA, 2002). Frente a estímulos
inflamatórios/infecciosos, macrófagos de animais desnutridos não respondem
com a mesma intensidade ou da mesma maneira que macrófagos oriundos dos
animais nutridos, possibilitando ou permitindo o desenvolvimento dos processos
infl amatórios e/ou infecciosos (BORELLI et al., 1998; FOCK et al., 2007; FOCK et
al., 2010; SCRIMSHAW, 2010). A desnutrição modifi ca a resposta do organismo ao
patógeno, podendo ocorrer alterações na produção de anticorpos, na atividade
fagocitária, diminuição da migração e mobilização celular para os focos infl amatórios
(BORELLI; MARIANO; BOROJEVIC, 1995), bem como diminuição da síntese de citocinas
proinfl amatórias (FOCK et al., 2007).
O peróxido de hidrogênio desempenha função fundamental na morte
microbiana por ser um forte oxidante capaz de degradar biomoléculas, in vitro e in
vivo, podendo agir como potente agente bacteriostático (BALDECK; MARQUIS, 2008).
Porém, sua maior importância reside no seu potencial em gerar outras espécies reativas
de oxigênio com capacidade ainda maior (BALDECK; MARQUIS, 2008; HERMES-
LIMA; STOREY, 1993). Assim, sabendo-se que a formação de peróxido de hidrogênio
desempenha importante função no processo de resposta a agentes infecciosos, nossos
resultados corroboram com a literatura, nos quais encontramos, em macrófagos
peritoneais de animais desnutridos, menor capacidade em sintetizar peróxido de
hidrogênio quando estimulados.
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 11_13961e-NUTRIRE v36 n3.indd 11 02/02/12 16:2902/02/12 16:29

12
A maioria dos radicais livres formados é removida pela ação dos sistemas superóxido
dismutase, catalase e glutationa peroxidase (AERTES et al.,1995; HALLIWELL, 1994).
Em circunstâncias em que o superóxido e peróxido de hidrogênio não são removidos
sufi cientemente rápidos, eles podem reagir entre si e produzir o radical hidroxil que
iniciará uma cascata de peroxidação lipídica causando danos teciduais (BABIOR, 1984).
Golden e Ramdath (1987) sugeriram que um mecanismo de reparo e uma
proteção inadequada na desnutrição, podem causar a injúria mediada por radicais livres
e o desequilíbrio entre a produção de radicais livres e as respectivas neutralizações
poderiam ser decorrentes da diminuição das defesas antioxidantes que ocorrem durante
a desnutrição. A detoxifi cação de superóxido e peróxido de hidrogênio são catalisadas
pelas enzimas superóxido dismutase e catalase, respectivamente. O sistema glutationa
peroxidase também decompõe peróxidos orgânicos e inorgânicos, utilizando tripeptídio
glutationa como cossubstrato. Em nosso modelo, observamos que macrófagos peritoneais
de animais desnutridos, apresentam redução da atividade das enzimas antioxidantes
(glutationa peroxidase, catalase e superóxido dismutase) em comparação aos macrófagos
dos animais do grupo controle.
A glutationa peroxidase utiliza selênio como cofator, enquanto a superóxido
dismutase utiliza cobre, zinco e manganês (ZUO et al., 2006). Como nossos dados
demonstram que animais dos grupos D e D+BCG apresentaram redução da ingestão de
ração poderia existir também uma possível redução dos níveis desses minerais, o que
poderia explicar, ao menos em parte, a signifi cativa redução da atividade das enzimas
glutationa peroxidase e superóxido dismutase nos macrófagos peritoneais de animais
dos grupos desnutridos em comparação com seus controles.
A redução signifi cativa da ingestão de ração e de proteína verifi cada nos grupos D
e D+BCG em relação aos seus respectivos grupos controles pode favorecer a redução da
concentração plasmática do aminoácido glutamina (CUNHA et al., 2003). Este aminoácido
atua como precursor de glutamato em macrófagos peritoneais, sendo o glutamato um
dos aminoácidos envolvidos na síntese da glutationa (NEWSHOLME, 2001). Desse modo,
a redução da atividade da glutationa peroxidase pode estar relacionada à redução da
concentração do seu substrato (glutationa), fato esse relacionado com o consumo de
uma dieta hipoproteica.
CONCLUSÕES
Os resultados obtidos no presente estudo revelam que a desnutrição proteico-
energética altera mecanismos de detoxicação de radicais livres e reduz a capacidade de
produção de peróxido de hidrogênio em macrófagos estimulados com BCG e que, em
parte, pode explicar a diminuição nas atividades bactericidas e fungicidas observadas
em animais desnutridos.
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 12_13961e-NUTRIRE v36 n3.indd 12 02/02/12 16:2902/02/12 16:29

13
REFERÊNCIAS/REFERENCES
AERTS, C.; WALLAERT, B.; GOSSET, P.; VOISIN, C. Relationship between oxygen-induced alveolar macrophage injury and cell antioxidant defence. J Appl Toxicol., v. 15, n. 1, p. 53-58, Jan-Feb 1995.
BABIOR, B. M. Oxidants from phagocytes: agents of defense and destruction. Blood, v. 64, n. 5, p. 959-966, Nov 1984.
BALDECK, J. D.; MARQUIS, R. E. Targets for hydrogen-peroxide-induced damage to suspension and biofi lm cells of Streptococcus mutans. Can J Microbiol . , v. 54, n. 10, p. 868-875, Oct 2008.
BELLOSTA, S.; BERNINI, F. Modulation of macrophage function and metabolism. Handb Exp Pharmacol., n. 170, p. 665-695, 2005.
BORELLI, P.; BLATT, S.; PEREIRA, J.; DE MAURINO, B. B.; TSUJITA, M.; DE SOUZA, A. C.; XAVIER, J. G.; FOCK, R. A. Reduction of erythroid progenitors in protein-energy malnutrition. Br J Nutr., v. 97, n. 2, p. 307-314, Feb 2007.
BORELLI, P.; MARIANO, M.; BOROJEVIC, R. Protein malnutrition: effect on myeloid cell production and mobilization into infl ammatory reactions in mice. Nutr Res., v. 15, n. 10, p. 1477-1485, Oct 1995.
BORELLI, P.; SOUZA, I. P.; BOROJEVIC, R.; DAGLI, M. L. Z.; KANG, H. C. Protein malnutrition: some aspects of the in vitro adhesion of peritoneal mouse macrophages. Ann Nutr Metab., v. 42, n. 6, p. 367–373, 1998.
CATCHATOURIAN, R.; ECKERLING, G.; FRIED, W. Effect of short term protein deprivation on hemopoietic functions of healthy volunteers. Blood, v. 55, n. 4, p. 625–628, Apr 1980.
CHANDRA, R. K. Nutrition and the immune system from birth to old age. Eur J Clin Nutr., v. 56, p. S73-S76, Aug 2002. Supplement 3.
CUNHA, W. D.; FRIEDLER, G.; VAISBERG, M.; EGAMI, M. I . ; COSTA ROSA, L. F. Immunosuppression in undernourished rats: the effect of glutamine supplementation. Clin Nutr., v. 22, n. 5, p. 453-457, Oct 2003.
DACIE, J. V.; LEWIS, S. M. Practical Haematology. Edinburgh, England: Churchill Livingstone, 1995. p. 57-58.
DOUMAS, B. T.; WATSON, W. A.; BIGGS, H. G. Albumin standards and the measurement of serum albumin with bromcresol green. Clin Chim Acta., v. 31, n. 1, p. 87-96, Jan 1971.
FOCK, R. A.; ROGERO, M. M.; VINOLO, M. A.; CURI, R.; BORGES, M. C.; BORELLI, P. Effects of protein-energy malnutrition on NF-kappaB signalling in murine peritoneal macrophages. Inflammation, v. 33, n. 2, p. 101-109, Apr 2010.
FOCK, R. A.; VINOLO, M. A.; DE MOURA SÁ ROCHA, V.; DE SÁ ROCHA, L. C.; BORELLI, P. Protein-energy malnutrition decreases the expression of TLR-4/MD-2 and CD14 receptors in peritoneal macrophages and reduces the synthesis of TNF-alpha in response to lipopolysaccharide (LPS) in mice. Cytokine, v. 40, n. 2, p. 105-114, Nov 2007.
FRAYN, K. N. Hormonal control of metabolism in trauma and sepsis. Clin Endocrinol., v. 24, n. 5, p. 577-599, May 1986.
GOLDEN, M. H.; RAMDATH, D. Free radicals in the pathogenesis of kwashiorkor. Proc Nutr Soc., v. 46, n. 1, p. 53-68, Feb 1987.
GORNALL, A. G.; BARDAWILL, C. J.; DAVID, M. M. Determination of serum proteins by means of the biuret reaction. J Biol Chem., v. 177, n. 2, p. 751–766, Feb 1949.
HALLIWELL, B. Free radicals, antioxidants, and human disease: curiosity, cause, or consequence? Lancet, v. 233, n. 8924, p. 721-724, Sept 1994.
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 13_13961e-NUTRIRE v36 n3.indd 13 02/02/12 16:2902/02/12 16:29

14
HERMES-LIMA, M.; STOREY, K. B. Antioxidant defenses in the tolerance of freezing and anoxia by garter snakes. Am J Physiol., v. 265, n. 3 pt2, p. R646-R652, Sept 1993.
KEUSCH, G. T. History of nutrition: malnutrition, infection and immunity. J Nutr., v. 133, n. 1, p. S336-S340, 2003.
LI, S.; SUN, Y.; LIANG, C. P.; THORP, E. B.; HAN, S.; JEHLE, A. W.; SARASWATHI, V.; PRIDGEN, B.; KANTER, J. E.; LI, R.; WELCH, C. L.; HASTY, A. H.; BORNFELDT, K. E.; BRESLOW, J. L.; TABAS, I.; TALL, A. R. Defective phagocytosis of apoptotic cells by macrophages in atherosclerotic lesions of ob/ob mice and reversal by a fish oil diet. Circ Res., v. 105, n. 11, p. 1072-1082, Nov 2009.
MCCORD, J. M.; FRIDOVICH, I. The utility of superoxide dismutase in studying free radical reactions. I. Radicals generated by the interaction of sulfi te, dimethyl sulfoxide, and oxygen. J Biol Chem., v. 244, n. 22, p. 6056-6063, Nov 1969.
NEWSHOLME, P. Why is L-glutamine metabolism important to cells of the immune system in health, postinjury, surgery or infection? J Nutr., v. 131, n. 9, p. S2515-S2522, Sept 2001. Supplement.
PICK, E.; MIZEL, D. Rapid microassays for the measurement of superoxide and hydrogen peroxide production by macrophages in culture using an automatic enzyme immunoassay reader. J Immunol Methods., v. 46, n. 2, p. 211-226, 1981.
POWANDA, M. C.; BEISEL, W. R. Metabolic effects of infection on protein and energy status. J Nutr., v. 133, n. 1, p. 322S-327S, Jan 2003.
REEVES, P. G.; NIELSEN, F. H.; FAHEY, G. C. AIN-93 purifi ed diets for laboratory rodents: fi nal report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the Ain-76a Rodent. Diet J Nutr., v. 123, n. 11, p. 1939–1951, Nov 1993.
SCHLESINGER, L. S. ; HORWITZ, M. A. Phagocytosis of Mycobacterium leprae by human monocyte-derived macrophages is mediated by complement receptors CR1 (CD35), CR3 (CD11b/CD18), and CR4 (CD11c/CD18) and IFN-gamma activation inhibits complement receptor function and phagocytosis of this bacterium. J Immunol., v. 147, n. 6, p. 1983-1994, Sept 1991.
SCRIMSHAW, N. S. Historical concepts of interactions, synergism and antagonism between nutrition and infection. J Nutr., v. 133, n. 1, p. 316S-321S, Jan 2003.
SCRIMSHAW, N. S. INCAP studies of nutrition and infection. Food Nutr Bull., v. 31, n. 1, p. 54-67, Mar 2010.
SIES, H. Strategies of antioxidant defense. Eur J Biochem., v. 215, n. 2, p. 213-219, Jul 1993.
VOLKOVOVÁ, K.; BENO, I.; STARUCHOVÁ, M.; BOBEK, P.; MEKINOVÁ, D.; TATARA, M. Antioxidative enzyme activity in the blood of healthy persons. Bratisl Lek Listy., v. 97, n. 3, p. 134-138, Mar 1996.
WENDEL, A. Glutathione peroxidase. Methods Enzymol., v. 77, p. 325-333, 1981.
YAMADA, Y.; SAITO, H.; TOMIOKA, H.; JIDOI, J. Relationship between the susceptibility of various bacteria to active oxygen species and to intracellular killing by macrophages. J Gen Microbiol., v. 133, n. 8, p. 2015-2021, Aug 1987.
YOUNG, V. R.; MARCHINI, J. S. Mechanisms and nutritional signifi cance of metabolic responses to altered intakes of protein and amino acids, with reference to nutritional adaptation in humans. Am J Clin Nutr., v. 51, n. 2, p. 270-289, Feb 1990.
ZUO, X. L.; CHEN, J. M.; ZHOU, X.; LI, X. Z.; MEI, G. Y. Levels of selenium, zinc, copper, and antioxidant enzyme activity in patients with leukemia. Biol Trace Elem Res., v. 114, n. 1-3, p. 41-53, Winter 2006.
Recebido para publicação em 22/09/10.Aprovado em 25/08/11.
PAIXÃO E SILVA, O.; FOCK, R. A.; ROGERO, M. M.; BORELLI, P. Peróxido de hidrogênio e enzimas antioxidantes em desnutrição proteico-energética. Nutrire: rev. Soc. Bras. Alim. Nutr.= J. Brazilian Soc. Food Nutr., São Paulo, SP, v. 36, n. 3, p. 1-14, dez. 2011.
_13961e-NUTRIRE v36 n3.indd 14_13961e-NUTRIRE v36 n3.indd 14 02/02/12 16:2902/02/12 16:29


















