
Relatório anual de monitoramento da ictiofauna do ... · oxigênio dissolvido (OD) da água dos...
31
1 Relatório anual de monitoramento da ictiofauna do reservatório da UHE Irapé (condicionantes 23 e 24) Biólogo Francisco Ricardo de Andrade Neto (CRBio 44968/04) Maio 2014
Transcript of Relatório anual de monitoramento da ictiofauna do ... · oxigênio dissolvido (OD) da água dos...

1
Relatório anual de monitoramento da ictiofauna do
reservatório da UHE Irapé
(condicionantes 23 e 24)
Biólogo Francisco Ricardo de Andrade Neto (CRBio 44968/04)
Maio 2014
2
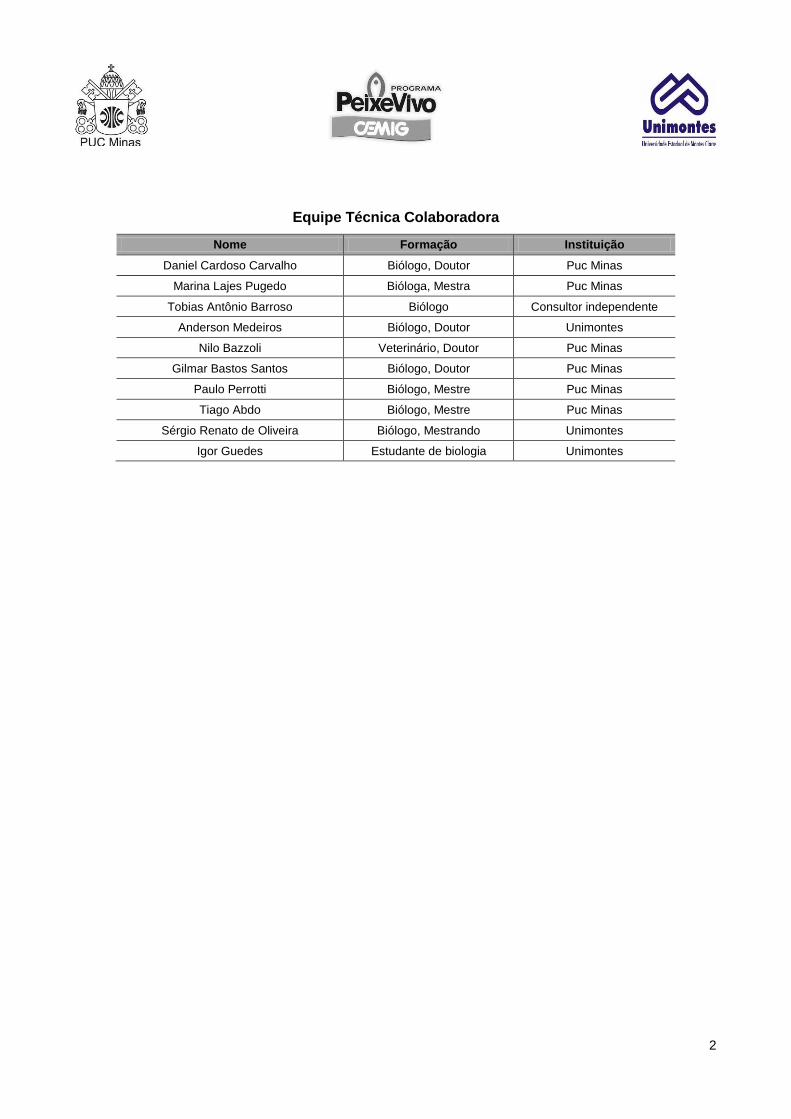
Equipe Técnica Colaboradora
Nome Formação Instituição
Daniel Cardoso Carvalho Biólogo, Doutor Puc Minas
Marina Lajes Pugedo Bióloga, Mestra Puc Minas
Tobias Antônio Barroso Biólogo Consultor independente
Anderson Medeiros Biólogo, Doutor Unimontes
Nilo Bazzoli Veterinário, Doutor Puc Minas
Gilmar Bastos Santos Biólogo, Doutor Puc Minas
Paulo Perrotti Biólogo, Mestre Puc Minas
Tiago Abdo Biólogo, Mestre Puc Minas
Sérgio Renato de Oliveira Biólogo, Mestrando Unimontes
Igor Guedes Estudante de biologia Unimontes

3
ÍNDICE
Apresentação .................................................................................................................................... 4 Introdução ........................................................................................................................................ 4 Objetivo ............................................................................................................................................ 5
Metodologia empregada nas coletas ................................................................................................ 5 Caracterização dos pontos de coleta ................................................................................................ 6 Operação do reservatório e caracterização limnológica .................................................................. 8 Composição da ictiofauna do reservatório da UHE Irapé ............................................................. 12 Caracterização das espécies capturadas no monitoramento ........................................................... 15
Prochilodus hartii ....................................................................................................................... 15 Leporinus elongatus ................................................................................................................... 17
Leporinus steindachneri ............................................................................................................. 18 Hypomasticus garmani ............................................................................................................... 19 Brycon devillei ........................................................................................................................... 20 Serrasalmus brandtii ................................................................................................................... 21
Moenkausia costae ..................................................................................................................... 25 Gênero Hoplias .......................................................................................................................... 26
Gênero Hypostomus ................................................................................................................... 27 Wertheimeria maculata .............................................................................................................. 29
Bibliografia .................................................................................................................................... 30

4
Apresentação
Em atendimento às condicionantes nos
23 e 24 da LO067/10, de 09/12/2010, referente ao
processo nº 00094/1994/006/2009, foi elaborado este relatório reportando os resultados obtidos no
monitoramento da ictiofauna do reservatório da UHE Irapé.
Introdução
A colonização de um reservatório recém-formado é um processo de duração variável onde se
observam alterações na composição da ictiofauna até que se atinja um certo “equilíbrio” (Agostinho
et al. 2007). Durante a colonização os peixes ocupam o novo ambiente e o sucesso no
estabelecimento depende da capacidade de cada espécie em se desenvolver e reproduzir em águas
com características lênticas.
Segundo Agostinho et al. (2007) fatores temporais e espaciais influenciam na colonização de
um reservatório. O fator temporal está relacionado às mudanças na produtividade primária do
reservatório ao longo do tempo. Nos primeiros anos do represamento existe alta concentração de
nutrientes provenientes da matéria orgânica submersa que será decomposta. Esses nutrientes
geralmente são disponibilizados a todos os níveis tróficos aumentando a produção. Nesse período a
pesca é mais elevada. Com o passar do tempo, reduz-se a matéria orgânica em decomposição e com
ela caem os níveis de nutrientes disponíveis entre os níveis tróficos e a pesca. Após esse período
atinge-se uma estabilidade que em geral fica acima da produtividade do antigo ambiente lótico e
abaixo do início da colonização.
O fator espacial foi identificado por Agostinho et al (1999) pela orientação longitudinal
(fluvial, transicional e lacustre), latitudinal (litorânea e pelágica) e vertical (epipelágica e
batipelágica). Nessa estratificação, maior diversidade e abundância é observada na região fluvial
litorânea enquanto esses parâmetros são menores em regiões batipelágicas.
O reservatório da Usina Hidrelétrica de Irapé tem nove anos desde a sua formação e está
passando por alterações na sua colonização por peixes, que vêm sendo acompanhadas desde 2009
até o presente. Não há informações sobre as características reprodutivas e a capacidade de
colonização das espécies de peixes da bacia do rio Jequitinhonha, o que dificulta qualquer
diagnóstico futuro sobre a comunidade de peixes no reservatório da UHIR. Neste trabalho foram

5
acompanhadas as alterações na composição da ictiofauna do reservatório de Irapé entre os anos de
2011 a 2014, com maior atenção voltada para o envelhecimento do reservatório, a introdução de
espécies exóticas e caracterização das espécies mais abundantes no monitoramento.
Objetivo
O objetivo deste trabalho foi cumprir as condicionantes 23 e 24 do licenciamento ambiental
da Usina Hidrelétrica de Irapé, que prevêm a continuidade do programa de monitoramento da
ictiofauna, em duas campanhas anuais, uma no período da piracema (outubro a fevereiro) e outra
fora deste período (maio a agosto), bem como a apresentação de relatórios anuais com os resultados
das campanhas e a caracterização das principais espécies que ocorrem no local.
Metodologia empregada nas coletas
As coletas foram realizadas no reservatório da UHE Irapé em diferentes pontos descritos na
próxima seção do texto. Entre abril de 2011 e abril de 2014, o reservatório foi amostrado em 96 dias.
Os intervalos entre as coletas foram irregulares, porém os 12 meses do ano foram contemplados nas
amostragens (Figura 1). Foi padronizado um esforço de 530 m2 de redes de espera entre as malhas
2.6, 3, 4, 5, 6, 7, 8, 10, 12, 14 e 16 cm entre nós opostos, detalhado na Tabela 1. Em algumas coletas
foi utilizado o esforço apenas com as redes de malha 8 e 10 cm.
As redes foram armadas por volta das 17 horas e retiradas às 7, totalizando 14 horas de
exposição. Os peixes capturados foram separados por malha, identificados, numerados e fixados em
formalina 10%. Parte dos indivíduos de cada espécie foram parcialmente dissecados e tiveram
fragmentos de gônadas fixados em líquido de Bouin por 24 horas. Algumas fêmeas maduras tiveram
fragmentos de ovários fixados em líquido de Gilson para posterior análise da fecundidade. O estádio
de maturação gonadal foi determinado macroscopicamente para a maior parte dos indivíduos de
acordo comas classes: repouso, em maturação/ maduro e desovado/ espermeado.
Durante as coletas foi utilizado um disco de Secchi e uma sonda multiparâmetros
Yellowsprings para medição de algumas variáveis físicas e químicas da água do reservatório da UHE
Irapé: profundidade do disco de Secchi (cm), oxigênio dissolvido (mg/L), temperatura (oC) e
condutividade (µS/s). As medidas foram feitas durante o dia, entre as 10 e 16 horas.

6
Rede M2.6 M3 M4 M5 M6 M7 M8 M10 M12 M14 M16
Área (m2) 30 45 45 45 45 53 60 48 51 51 57
Tabela 1: Esforço de pesca empregado nas coletas de peixes no reservatório de Irapé
Figura 1: Nível do reservatório da UHE Irapé entre janeiro de 2011 e abril de 2014.
▲ indicam os dias em que houve coleta deste projeto.
Caracterização dos pontos de coleta
O monitoramendo da ictiofauna do reservatório de Irapé possui quatro pontos de coleta,
distribuídos espacialmente de acordo com a Figura 2 e descritos a seguir:
Figura 2: Distribução dos pontos de coleta no reservatório da UHE Irapé.

7
IR2 (S16º46’25.8” W42º38’47.8” ) – Rio Jequitinhonha, na zona lacustre do reservatório. As
coletas são feitas na região litorânea, desde as proximidades da barragem até próximo à concluência
com o rio Itacambiruçu (Figura 3).
Figura IR2: Trecho do reservatório no ponto 3.
IR3 (S16º43’44.3” W42º40’49.1”) – Rio Itacambiruçu, nos primeiros quilômetros a montante da
confluência com o rio Jequitinhonha. As coletas são feitas na região litorânea, em grotas de rios
intermitentes que drenam pela chapada (Figura 4).
Figura 4: Armação de rede na região litorânea do ponto IR3.
IR8 (S16º44’06.9” W42º34’34.1”) – Rio Jequitinhonha, na zona lacustre do reservatório. As coletas
são feitas na região pelágica. As redes são armadas nas bóias que delimitam a área de segurança e
estão dispostas transversalmente ao reservatório (Figura 5).

8
Figura 5: Armação de redes no ponto IR8 (região pelágica).
IR9 (S16o32'59.12" W42
o46'34.66") – Rio Itacambiruçu no final do reservatório, onde existe uma
influência do ambiente lótico. Zona fluvial. Nesse ponto, a distância entre as margens do
reservatório é mais curta e a profundidade é menor. Os paliteiros de buritis são mais comuns, assim
como os bancos de areia causados pela deposição de silte trazidos pelo rio (Figura 6).
Figura 6: Trecho do reservatório no ponto IR9.
Operação do reservatório e caracterização limnológica
Dados históricos do nível e variação do reservatório da UHE Irapé entre janeiro de 2011 e abril
de 2014 foram obtidos junto à CEMIG, totalizando 1214 dias. Nesse período o reservatório variou
19.71 m entre as cotas 488.32 e 508.03 m (Figura 1). A cota média foi 499.85 m, e de forma geral o
9
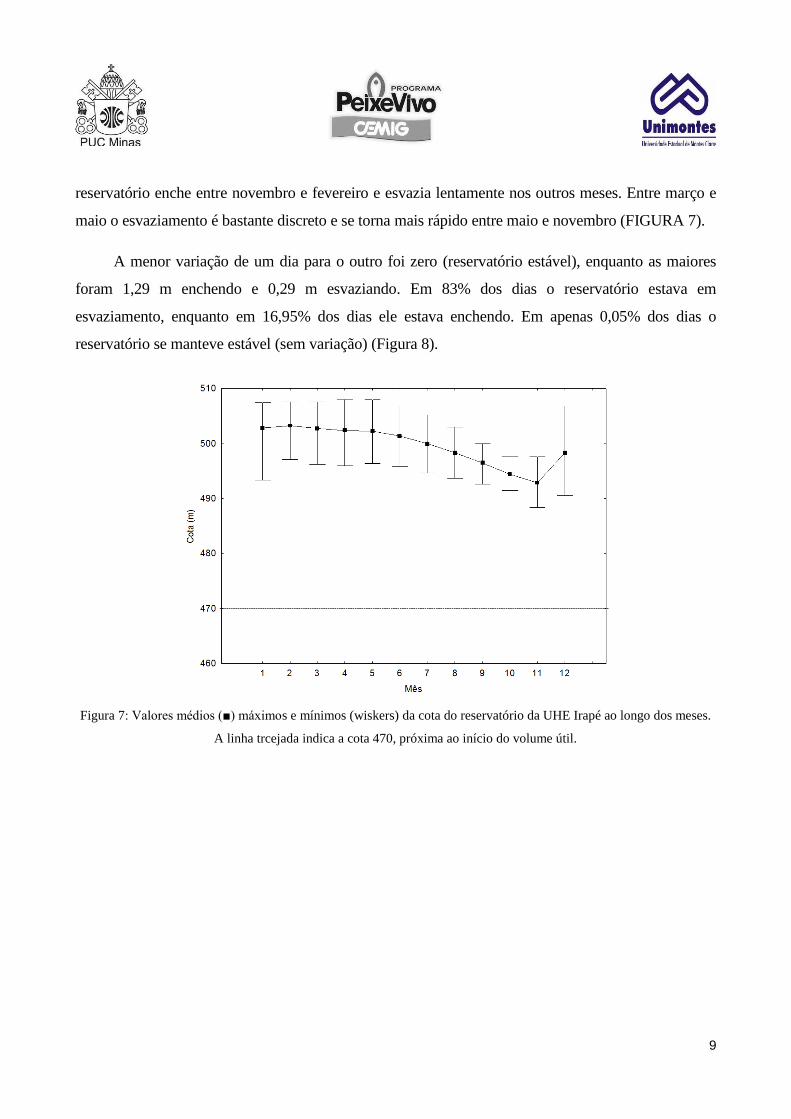
reservatório enche entre novembro e fevereiro e esvazia lentamente nos outros meses. Entre março e
maio o esvaziamento é bastante discreto e se torna mais rápido entre maio e novembro (FIGURA 7).
A menor variação de um dia para o outro foi zero (reservatório estável), enquanto as maiores
foram 1,29 m enchendo e 0,29 m esvaziando. Em 83% dos dias o reservatório estava em
esvaziamento, enquanto em 16,95% dos dias ele estava enchendo. Em apenas 0,05% dos dias o
reservatório se manteve estável (sem variação) (Figura 8).
Figura 7: Valores médios (■) máximos e mínimos (wiskers) da cota do reservatório da UHE Irapé ao longo dos meses.
A linha trcejada indica a cota 470, próxima ao início do volume útil.

10
Figura 8: Variação diária de nível do reservatório da UHE Irapé, por mês.
Valores médios indicados por ■ e os máximos e mínimos pelos wiskers.
Linha tracejada indica variação= 0 de um dia para outro.
Em cinza o período em que houve variação positiva.
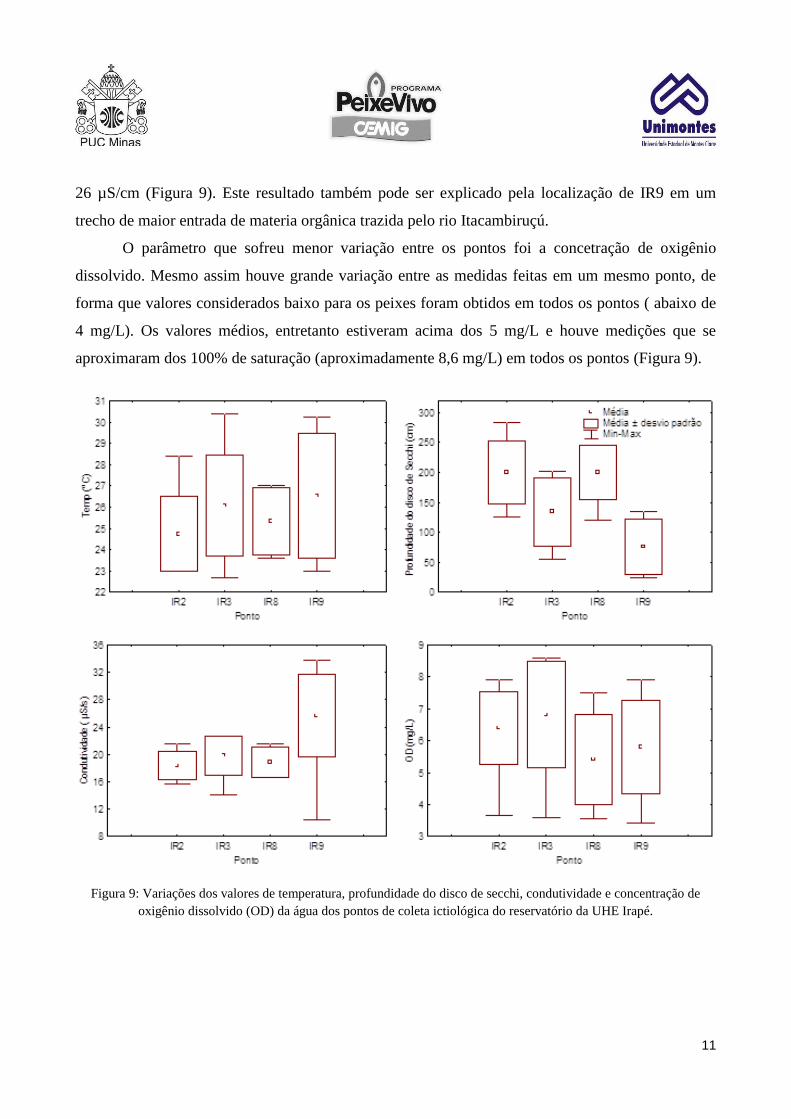
Os parâmetros físicos e químicos da água do reservatório apresentaram grande variação
entre os pontos de amostragem e até mesmo em um mesmo ponto houve variação temporal. A
temperatura, um dos parâmetros mais importantes para a reprodução de peixes (Sato et al. 2005)
teve maior variação nos pontos IR3 e IR9, localizados no rio Itacambiruçú. Nesse pontos a variação
foi de 22,5 a 30,5 ºC, com valores médios entre 26 e 26,5ºC em IR3 e IR9, respectivamente. Em
IR2 e IR8, próximos entre si e localizados no rio Jequitinhonha, a variação foi menor 23 a 28,5 ºC e
a média entre 24,5 e 25,5 em IR2 e IR8, respectivamente (Figura 9).
A profundidade do disco de Secchi, que é uma medida da transparência da água também
seguiu uma tendência similar, com IR3 e IR9 apresentando águas menos transparentes em relação a
IR2 e IR8 (Figura 9). Os menores valores foram obtidos em IR9, o que é de se esperar devido a
localização do ponto, próximo ao trecho lótico e num local onde há mais materia em suspensão.
Nos pontos IR2 e IR8 a maior parte deste material já se sedimentou, fazendo com que a água fique
mais transparente, mesmo nos períodos de chuva.
A condutividade da água mostrou um padrão bastante diferente, com IR2, IR3 e IR8
sofrendo menores variações, entre 13 e 23 µS/cm e IR9 variando entre 10 e 34 µS/cm com média de
11
26 µS/cm (Figura 9). Este resultado também pode ser explicado pela localização de IR9 em um
trecho de maior entrada de materia orgânica trazida pelo rio Itacambiruçú.
O parâmetro que sofreu menor variação entre os pontos foi a concetração de oxigênio
dissolvido. Mesmo assim houve grande variação entre as medidas feitas em um mesmo ponto, de
forma que valores considerados baixo para os peixes foram obtidos em todos os pontos ( abaixo de
4 mg/L). Os valores médios, entretanto estiveram acima dos 5 mg/L e houve medições que se
aproximaram dos 100% de saturação (aproximadamente 8,6 mg/L) em todos os pontos (Figura 9).
Figura 9: Variações dos valores de temperatura, profundidade do disco de secchi, condutividade e concentração de
oxigênio dissolvido (OD) da água dos pontos de coleta ictiológica do reservatório da UHE Irapé.
12
Composição da ictiofauna do reservatório da UHE Irapé
Utilizando-se a equação da relação espécie-área de Welcomme (1985) o número estimado de
espécies para a bacia do Jequitinhonha está próximo de 80. No entanto, foram capturadas 20
espécies de peixes no reservatório da UHE Irapé entre 2011 e 2014, sendo que duas foram
introduzidas na bacia (M. costae e S. brandtii). Este é um valor baixo e frequentemente observado,
uma vez que reservatórios tropicais apresentam menos espécies em relação aos rios aos quais
pertencem (Araujo & Santos, 2001).
Os Characiformes são o grupo mais rico em número de espécies (60%), seguidos pelos
Siluriformes (25%). Juntamente com Gymnotiformes com uma espécie capturada, observa-se que
90% da ictiofauna do reservatório pertence aos Otophysi. Este grupo representa 70% da ictiofauna
do reservatório de Lajes (Araujo & Santos, 2001) e 94% em Juramento (Silva et al., 2006).
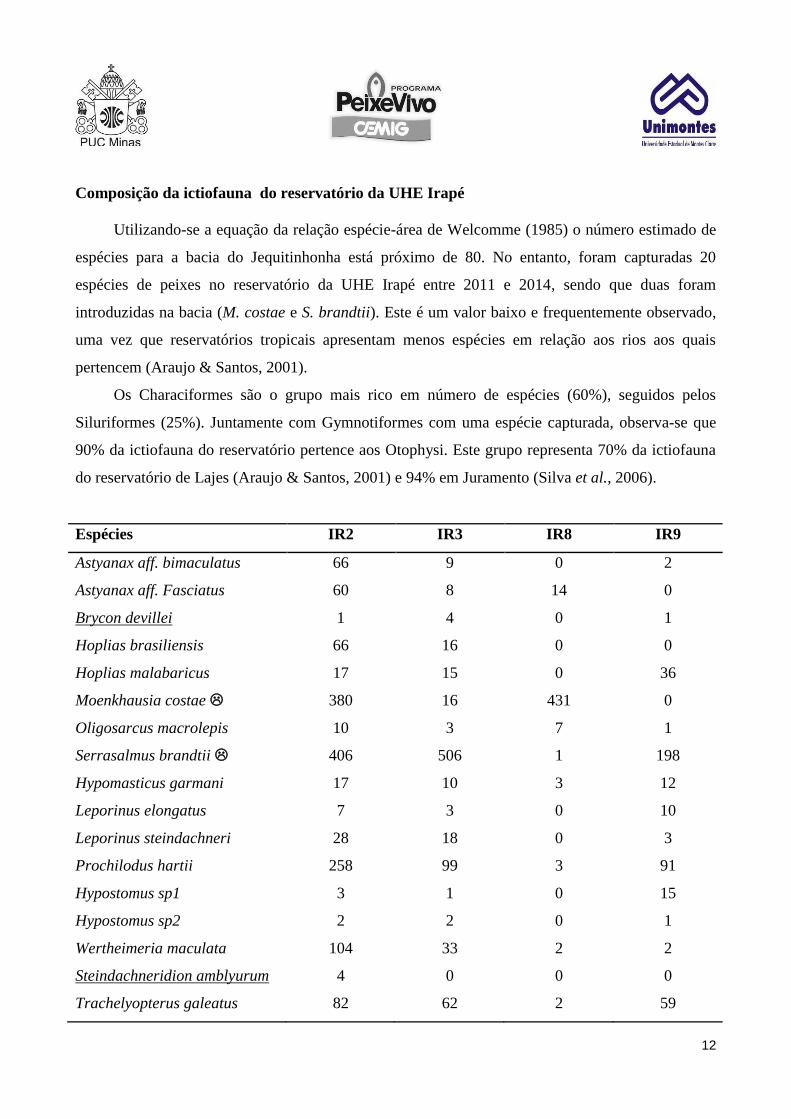
Espécies IR2 IR3 IR8 IR9
Astyanax aff. bimaculatus 66 9 0 2
Astyanax aff. Fasciatus 60 8 14 0
Brycon devillei 1 4 0 1
Hoplias brasiliensis 66 16 0 0
Hoplias malabaricus 17 15 0 36
Moenkhausia costae 380 16 431 0
Oligosarcus macrolepis 10 3 7 1
Serrasalmus brandtii 406 506 1 198
Hypomasticus garmani 17 10 3 12
Leporinus elongatus 7 3 0 10
Leporinus steindachneri 28 18 0 3
Prochilodus hartii 258 99 3 91
Hypostomus sp1 3 1 0 15
Hypostomus sp2 2 2 0 1
Wertheimeria maculata 104 33 2 2
Steindachneridion amblyurum 4 0 0 0
Trachelyopterus galeatus 82 62 2 59

13
Gymnotus carapo 3 1 0 2
Cichlasoma facetum 0 0 0 5
Geophagus brasiliensis 0 1 0 0
Tabela 2: Espécies e número de indivíduos capturados nos quatro pontos de amostragem do reservatório da UHE Irapé.
Sublinhadas estão as espécies ameaçadas de extinção. indica espécie introduzida na bacia do Jequitinhonha.
Foi feita a avaliação da comunidade de peixes por ordenação baseada na abundância de cada
espécie, através de um teste nMDS (nonmetric multidimensional scaling) no programa Past, usando
a distância euclideana como medida de similaridade. Esta análise foi feita por ano (2011, 2012 e
2013) e por ponto (IR2, IR3, IR8 e IR9). A diferença entre os anos e pontos foi verificada pelo teste
ANOSIM (analysis of similarity) (Loures & Pompeu, 2012). M. costae corresponde numericamente
a cerca de 25% de todos os peixes capturados, porém a maior parte foi obtida em apenas três
coletas. Para evitar efeitos extremos dessa espécie nas análises, foi considerado apenas um
indíviduo (presença) nos casos citados de capturas muito maiores (acima de 79 indivíduos).
Foi observada diferença significativa entre os anos de coleta (Figura 10 A), indicando que a
comunidade sofreu alterações ao longo do tempo e também no espaço. Entre os anos verifica-se a
formação de três grupos onde 2011 está isolado e 2012 e 2013 têm pequena sobreposição. O stress
associado ao teste foi de 0,049. O teste ANOSIM forneceu um valor de R= 0,27 e p= 0,023. Esse
resultado possivelmente está relacionado à ocorrência de chuvas. 2011 foi o ano de maior
pluviosidade entre os três. 2012 as chuvas foram menos frequente e intensas e 2013 ainda menos. A
flutuação do reservatório, cujo enchimento está relacionado à ocorrência de chuvas fornece uma
idéia de como a ocorrência de chuvas foi baixa em 2013 (Figura 1), uma vez que o reservatório
ficou a mais de 10 metros abaixo dos anos anteriores e subsequente.
Considerando a abordagem por ponto de amostragem, são formados 4 grupos dos quais IR8
e IR9 estão bem definidos e IR2 e IR3 têm grande sobreposição (Figura 10 B). O stress associado
ao teste foi de 0,092. O teste ANOSIM forneceu um valor de R= 0,31 e p< 0,0001. IR8 pode ser
separado dos outros pontos pela sua localização na região pelágica. Na maior parte das coletas a
captura é zero. Uma vez que há maior entrada de nutrientes na região litorânea através das zonas
ripárias e à maior diversidade e estruturação dos habitats, espera-se que esta região seja menos
colonizada por organismos em relação à faixa litorânea (Agostinho et al 1999). O ponto IR9 é o
mais distante dos pontos em termos espaciais. Seu posicionamento isolado em dos demais pontos

14
pode ser explicado pela distância e tipo de ambiente em que se encontra. O final do reservatório traz
características comuns a ambientes lênticos e lóticos que certamente influenciam na composição da
comunidade. Neste ponto foi observada maior variação na temperatura, menor profundidade do
disco de Secchi e maior variação da condutividade (Figura 9). Além disso maior parte das espécies
que exibiu atividade reprodutiva no reservatório foi coletada em IR9. Os pontos IR2 e IR3 tiveram
suas composições mais similares. Isto pode ser explicado pela maior proximidade entre os pontos e
a realização das coletas na região litorânea.
Figura 10: Ordenação multivariada (nMDS) da ictiofauna amostrada no reservatório de Irapé.
A – análise por anos. B análise por pontos de coleta.

15
Caracterização das espécies capturadas no monitoramento
Prochilodus hartii
Figura 11: A – Prochilodus hartii. B – macho com testículos maduros. C – fêmea com ovários desovados.
O monitoramento de peixes na área de influência da UHE Irapé é realizado em parceria com
a PUC Minas, principalmente nos estudos genéticos e da reprodução dos peixes da bacia. O
primeiro estudo realizado na região foi feito com a curimba, uma espécie importante para a pesca de
subsistência e comercial. Foi observado que a curimba não entra em maturação gonadal no
reservatório e imediatamente a jusante do barramento.
Nesses pontos os machos apresentavam testículos em repouso com lume dos túbulos
seminíferos ocluído e parede apenas com espermatogônias, ou testículos em maturação inicial com
lume dos túbulos seminíferos contendo pequena quantidade de espermatozoides (Figura 12 a). Nas
fêmeas os ovários estavam em repouso contendo ovócitos perinucleolares inicial (O1) e avançado
(O2). (d) ovário em maturação inicial contendo O1, O2 e ovócito pré vitelogênico (O3) com
característicos alvéolos corticais (Figura 12 b).
No trecho lótico do rio Itacambiruçu foram coletados machos com testículos em maturação
avançada contendo grande quantidade de espermatozóides no lúmen dos túbulos seminíferos

16
(Figura 13 a) e testículos espermiados com lúmen dos túbulos seminíferos vazios ou contendo
espermatozóides residuais (Figura 13 b). As fêmeas tinham ovários em maturação avançada
contendo numerosos ovócitos vitelogênicos (O4) (Figura 13 c) eovários desovado contendo O1, O2
e folículos pós ovulatórios (Figura 13 d).
Figura 12. testículos (a e b) e ovários (c e d) de P. hartii em repouso ou maturação inicial.
Figura 13. Testículos (a e b) e ovários (c e d) de P. hartii maduros e espermeado/ desovado.

17
Leporinus elongates
Figura 14: A – Leporinus elongatus. B – fêmea com um ovário desovado e outro repleto de ovócitos.
O antigo nome Leporinus crassilabris passou a sinônimo júnior de Leporinus elongatus
após revisão de Britski et al. (2012). A piapara é uma espécie de grande porte bastante importante
para a pesca local e alvo de peixamentos pela EPMM. No reservatório de Irapé foram capturados 38
indivíduos, com comprimento total variando de 201 a 360 mm e o peso de 167 a 450 g. Cerca de
75% das capturas ocorreram em IR9, inclusive com vários indivíduos em atividade reprodutiva.
Oito dos nove machos se encontravam em atividade reprodutiva, enquanto as fêmeas mais
comuns estavam em maturação (dez em dezoito). Uma fêmea foi considerada madura, quatro desovadas
e três estavam em repouso. Os estádios em maturação e maduro foram observados em outubro e
novembro, enquanto as desovadas foram coletadas em novembro e maio. Com relação às diferenças de
tamanho atribuídas ao sexo, cerca de ¾ das fêmeas eram maiores que os machos (Figura 15).

18
Figura 15: Diferenças no comprimento padrão entre os sexos em L. elongatus.
Leporinus steindachneri
Figura 16: A – Leporinus steindachneri. B – fêmea com ovários maduros.
O piau três pintas é um Anostomida de médio a grande porte (Comprimento padrão do
maior indivíduo capturado = 33,0 cm) que tem uma ampla distribuição entre as bacias do leste
brasileiro, como Jequitinhonha, Mucuri, Itanhem e Doce. Esta espécie apresenta características que
indicam a capacidade de reprodução no reservatório da UHE Irapé: presença de indivíduos jovens
no monitoramento e de fêmeas desovadas. Em relação aos sexos, as fêmeas são ligeiramente
F M
Sexo
140
160
180
200
220
240
260
280
300
320
340
360
Com
prim
ento
padrã
o (m
m)
Median 25%-75% Min-Max

19
maiores que os machos, mas esta diferença não é significativa (ANOVA = 0.40, p = 0.52). O
tamanho da primeira maturação desta espécie está próximo aos 21 cm para os dois sexos.
Hypomasticus garmani
Figura 17: A – Hypomasticus garmani. B – fêmea com ovários maduros. C – fêmea desovada.
Esta espécie conhecida popularmente como timburé é caracterizada pelo pequeno porte
(Comprimento padrão do maior indivíduo capturado = 21,4 cm), posição da boca subinferior e
única mancha negra no pedúnculo caudal. A espécie pertencia ao gênero Leporinus e foi
recentemente incluída em Hypomasticus juntamente com L. mormyrops e L. thayeri, também de
outras bacias costeiras do leste brasileiro (Sidlauskas & Vari, 2008).
Assim como L. steindachneri, H. garmani também apresentou fêmeas desovadas no reservatório da
UHE Irapé. Além disso, não foi verificada diferenças de tamanho corporal entre os sexos (ANOVA
= 1.61; p= 0.2).

20
Brycon devillei
Figura 18: Exemplar de Brycon devillei (A). Ovários em maturação avançada (B).
A piabanha do Jequitinhonha (Figura 18- A) é uma espécie de grande porte, que ocorre nos
tributários e na calha principal das bacias dos rios Jequitinhonha e Doce. Entretanto, Lima (2001)
em sua revisão sobre o gênero Brycon indica a existência de duas espécies ainda não descritas
nessas bacias. Desta forma, o nome B. devillei foi utilizado na lista de espécies ameaçadas de forma
provisória (Vieira et al. 2008). As poucas informações sobre a piabanha do Jequitinhonha estão em
Godinho et al. (1999) e Azevedo et al. (2011) e abrangem sua dieta e comportamento social. Sobre
sua reprodução existe a informação da existência de uma estratégica periódica, semelhante a de
peixes de piracema.
Até o momento foram capturados 13 indivíduos no reservatório de Irapé. Entre eles, o
comprimento padrão variou de 150 a 346 mm e o peso entre 67 e 985 g. Alguns desses indivíduos
sobreviveram à captura e manipulação e foram mantidos vivos para serem transportados a Estação
de Piscicultura de Machado Mineiro (EPMM) para comporem o plantel de matrizes e reprodutores.
Doze peixes foram dissecados. Apenas uma fêmea capturada em agosto esta em repouso. Os outros
eram oito machos e três fêmeas em avançada maturação gonadal ou maduros (Figura 18 –B). Esses
peixes foram coletados em outubro, dezembro e janeiro, o que reforça a hipótese de uma estratégia

21
reprodutiva periódica provavelmente relacionada ao período chuvoso, como na maioria das espécies
de peixes Neotropicais.
É interessante que uma espécie de ambientes lóticos e ameaçada de extinção seja capturada
num ambiente lêntico e alterado como um reservatório. Pricipalmente em atividade reprodutiva.
Esse resultado levanta a possibilidade de que esses peixes entrem em maturação no reservatório e
utilizem regiões de grota onde há córregos intermitentes que transportam água e material alóctone
para o reservatório durante o período chuvoso. Mesmo que não haja reprodução, talvez essas
regiões tenham condições mais parecidas com as de um rio no período chuvoso, tais como variação
de nível, redução da transparência da água e aumento na temperatura.
Dentre os peixes capturados em atividade reprodutiva, os machos possuíam as nadadeiras
peitorais, pélvicas e anal com ganchos. Este é um fato comum ao gênero Brycon. Porém, dois
resultados inusitados foram (i) a ocorrência de ganchos também na nadadeira dorsal de machos e
(ii) a ocorrência de ganchos na dorsal fêmeas. Em apenas outras duas espécies de Brycon ganchos
na dorsal haviam sido observados em machos: B. coquenani da Venezuela e B. stolzmanni do Peru.
Já em fêmeas, apenas um indivíduo proveniente do rio Pardo tinha essa característica.
Serrasalmus brandtii
A pirambeba (S. brandtii) é nativa da bacia do São Francisco, onde é a espécie dominante
em ambientes lênticos (Silva et al. 2006). No rio Jequitinhonha, foi registrada pela primeira vez em
2005 antes do fechamento da UHE Irapé. No reservatório, começou a ser capturada em 2009 e a
partir de 2011 se tornou constante nas coletas. A partir daí sua participação em termos numéricos
vem se tornando maior a cada ano. Contabilizando-se todas as coletas realizadas entre 2011 e 2014
nos pontos IR2 e IR3 (Figura 19) verifica-se que em IR3, S. brandtii representava em média cerca
de 10% dos peixes capturados em 2011 e em 2013 já estava acima dos 40%. Em 2013 houve coleta
em que foi a única espécie capturada. Já em IR2 a situação é ainda mais complexa, pois sua
participação média já era superior a 20% em 2011 e cresceu de modo que em 2013 era próximo a
70%. Em 2012 e 2013 houve coletas em que só S. brandtii fora coletada. Para os dois pontos o
aumento ou no mínimo a alta participação é prevista para 2014, uma vez que até abril já se
observava valores próximos a 40% em IR3 e 70% em IR2.

22
Para avaliar os efeitos da presença e do aumento de S. brandtii no reservatório, analisamos
os peixes capturados em redes de emalhar quanto a presença de mutilações nas nadadeiras e no
corpo. Foram analisados 1524 peixes das 20 espécies capturadas no reservatório. Dentre os pontos,
IR2 foi o que apresentou maio relação entre não atacados/ atacados (Figura 20), o que é contrastante
com a participação de S. brandtii no ponto (Figura 19)., onde há maior infestação por S. brandtii. A
mutilação de nadadeiras também foi avliada na planície de inundação do alto rio Paraná nas décadas
de 1980 e 1990, onde Agostinho et al. (2003) observaram que nadadeiras de peixes correspondiam a
um dos mais importantes itens alimentares de duas espécies do gênero Serrasalmus. A dieta de S.
brandtii em ambientes lênticos do rio São Francisco, também mostrou alta frequência para esse
item alimentar (Pompeu, 1999).
Das 20 espécies analisadas todas tiveram algum ataque ás nadadeiras, sendo as mais
atacadas, Brycon devillei, Hypostomu sp1, Hypostomus sp2, Leporinus steindachneri e
principalmente Prochilodus hartii, espécie importante para a pesca e que vem sendo predada nas
redes de pesca e gerando prejuízos aos pescadores. Entre as espécies menos atacadas, está a própria
S. brandtii (Figura 21) .
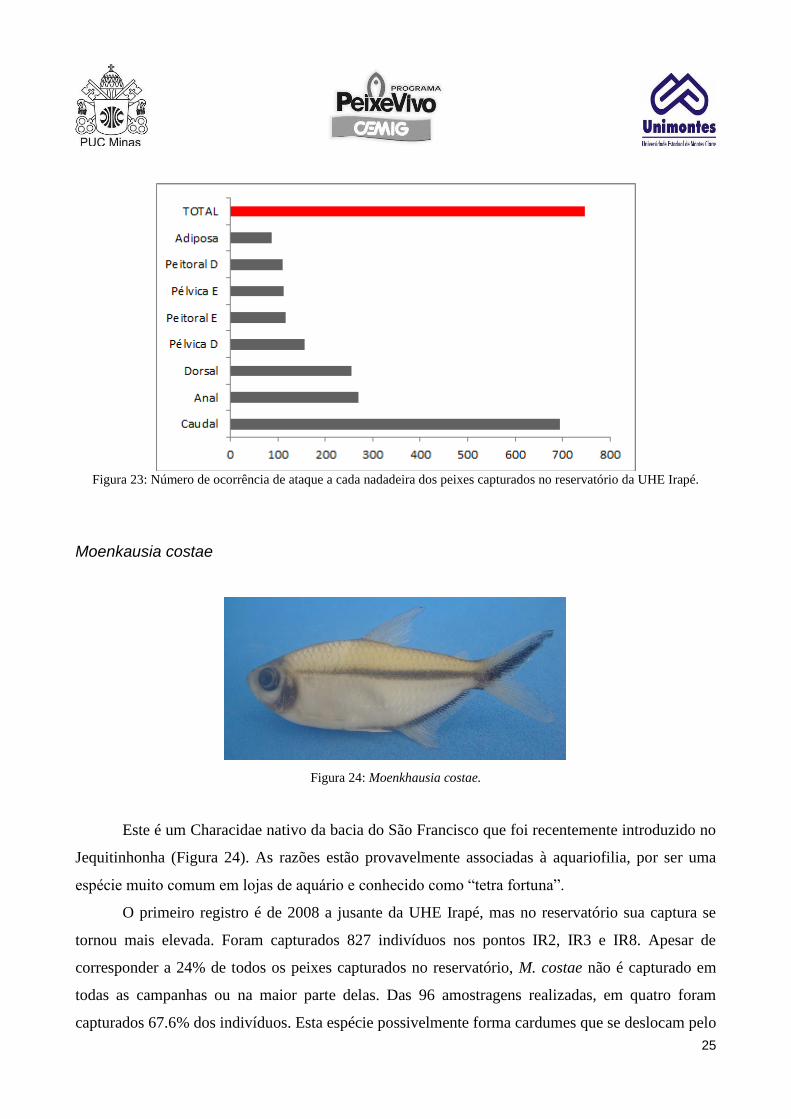
Entre os peixes atacados, observou-se que a nadadeira caudal é a mais atacada, com
frequência superior a 70% dos casos. Na sequencia aparecem anal e dorsal; as peitorais e pélvicas e
por fim a adiposa que tem menor área e está ausente em algumas espécies, o que pode explicar sua
baixa taxa de ataque (Figura 22). Esses resultados podem mostrar algumas características do
comportamento dos S. brandtii ao atacarem outros peixes. As espécies mais frequentemente
atacadas não são piscivoras e não são agressivas. As nadadeiras mais atacadas são as que têm maior
área e se localizam na região posterior do peixe, o que pode aumentar as chances de sucesso no
ataque, sem que o indivíduo perceba a aproximação do predador.

23
Figura 20: Participação (%) de S. brandtii nas coletas realizadas em IR3 (superior) e IR2 (inferior) entre 2011 e 2014.

24
Figura 21: Relação entre peixes não atacados/ atacados nos pontos do reservatório da UHE Irapé
Figura 22: Relação entre peixes não atacados/ atacados por espécie capturada no reservatório da UHE Irapé.
25
Figura 23: Número de ocorrência de ataque a cada nadadeira dos peixes capturados no reservatório da UHE Irapé.
Moenkausia costae
Figura 24: Moenkhausia costae.
Este é um Characidae nativo da bacia do São Francisco que foi recentemente introduzido no
Jequitinhonha (Figura 24). As razões estão provavelmente associadas à aquariofilia, por ser uma
espécie muito comum em lojas de aquário e conhecido como “tetra fortuna”.
O primeiro registro é de 2008 a jusante da UHE Irapé, mas no reservatório sua captura se
tornou mais elevada. Foram capturados 827 indivíduos nos pontos IR2, IR3 e IR8. Apesar de
corresponder a 24% de todos os peixes capturados no reservatório, M. costae não é capturado em
todas as campanhas ou na maior parte delas. Das 96 amostragens realizadas, em quatro foram
capturados 67.6% dos indivíduos. Esta espécie possivelmente forma cardumes que se deslocam pelo

26
reservatório entre as regiões litorâneas e pelágicas. Isso explica eventos isolados de captura de um
grande número de indivíduos nos pontos IR3, IR2 e IR8.
Com exceção de quatro indivíduos capturados em redes M3, todos os outros foram capturados nas
M2.6. O comprimento total varia de 65 a 95 mm e o peso de 5.25 a 8.25 g. A maior parte dos peixes
é composta por fêmeas, fato similar ao observado por Hojo et al. (2004). É possível que o petrecho
M2.6 esteja influenciando na proporção sexual. Por serem menores os machos devem ser
capturados em redes de malhas ainda menores que M2.6, que é a menor utilizada neste trabalho. Ao
exame macroscópico os ovários foi possível verificar que M. costae se reproduz o ano todo com
desova parcelada, o que também foi visto por Hojo et al. (2004).
Gênero Hoplias
Figura 25: A – Hoplias brasiliensis. B – Sínfise mandibular em H. brasiliensis. C – Sínfise mandibular em H. malabaricus.

27
As espécies do gênero Hoplias capturadas no reservatório da UHE Irapé são H. malabaricus
(traira) e H. brasiliensis (trairão). Esta última pertencia ao complexo H. lacerdae que foi
recentemente desmembrado em cinco espécies por Oyakawa & Mattox (2009). Ambas podem ser
diferenciadas pelas margens mediais dos dentários, dispostas paralelamente em H. brasiliensis ou
convergindo em direção à sínfise mandibular em H. malabaricus (Figura 25 B e C).
Existe uma tendência a se acreditar que os indivíduos maiores pertencem a H. brasiliensis. Desta
forma foi feita um teste de ANOVA onde se verificou que H. brasiliensis são em média 4 cm
maiores que H. malabaricus (ANOVA = 17.90; p = 0,00004). Além dessas diferenças citadas, H.
brasiliensis parece ser menos frequente no reservatório em relação a H. malabaricus e o oposto
acontecendo em riachos e córregos.
Atividade reprodutiva foi observada para as duas espécies durante todo o ano. Já é
documentado que as traíras e trairões apresentam desova parcelada e apresentam cuidado parental
através da construção de ninhos.
Gênero Hypostomus
Figura 26: A – Hypostomus sp1. B – Fêmea com ovário maduro.
28
Não há nenhuma espécie do gênero Hypostomus descrita para a bacia do Jequitinhonha.
Entretanto, assim como acontece na bacia do rio Paraíba do Sul, ocorrem duas espécies, sendo uma
mais característica da calha principal (Hypostomus sp1) e outra mais comum em córregos de médio
e pequeno porte (Hypostomus sp2) (Mazzoni & Caramaschi, 1995). Apesar dessa tendência, existe
superposição na distribuição das espécies.
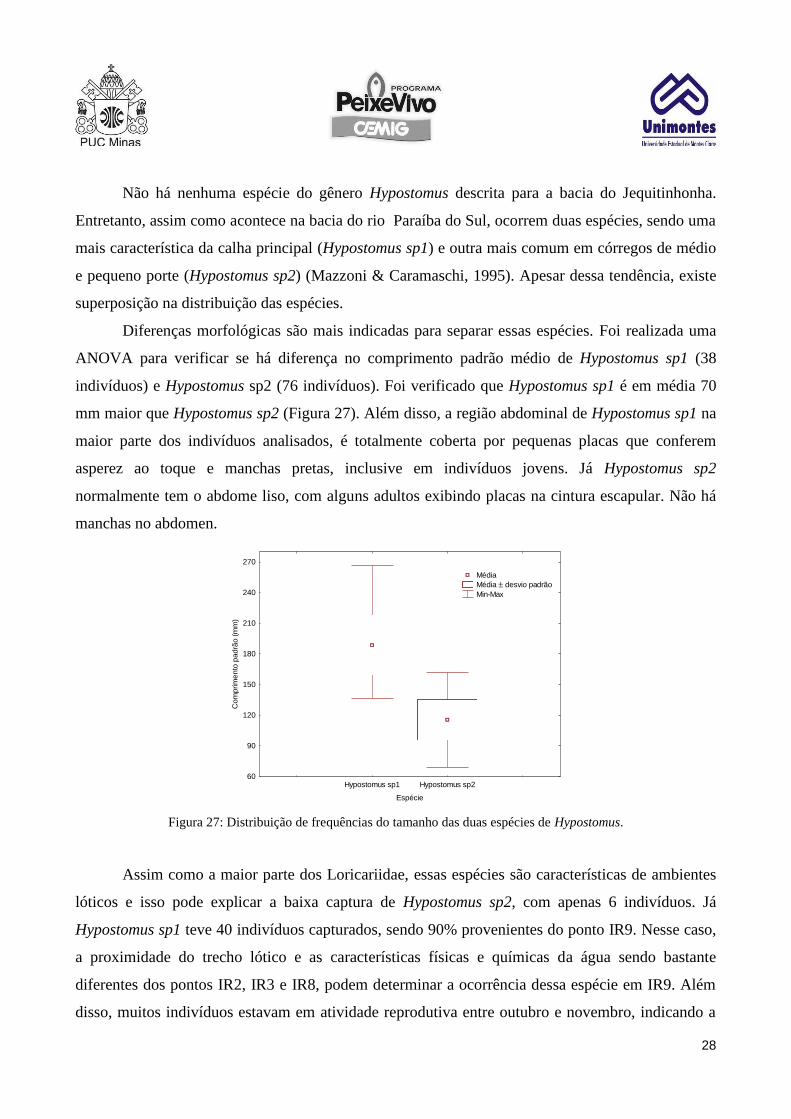
Diferenças morfológicas são mais indicadas para separar essas espécies. Foi realizada uma
ANOVA para verificar se há diferença no comprimento padrão médio de Hypostomus sp1 (38
indivíduos) e Hypostomus sp2 (76 indivíduos). Foi verificado que Hypostomus sp1 é em média 70
mm maior que Hypostomus sp2 (Figura 27). Além disso, a região abdominal de Hypostomus sp1 na
maior parte dos indivíduos analisados, é totalmente coberta por pequenas placas que conferem
asperez ao toque e manchas pretas, inclusive em indivíduos jovens. Já Hypostomus sp2
normalmente tem o abdome liso, com alguns adultos exibindo placas na cintura escapular. Não há
manchas no abdomen.
Figura 27: Distribuição de frequências do tamanho das duas espécies de Hypostomus.
Assim como a maior parte dos Loricariidae, essas espécies são características de ambientes
lóticos e isso pode explicar a baixa captura de Hypostomus sp2, com apenas 6 indivíduos. Já
Hypostomus sp1 teve 40 indivíduos capturados, sendo 90% provenientes do ponto IR9. Nesse caso,
a proximidade do trecho lótico e as características físicas e químicas da água sendo bastante
diferentes dos pontos IR2, IR3 e IR8, podem determinar a ocorrência dessa espécie em IR9. Além
disso, muitos indivíduos estavam em atividade reprodutiva entre outubro e novembro, indicando a
Hypostomus sp1 Hypostomus sp2
Espécie
60
90
120
150
180
210
240
270
Com
prim
ento
padrã
o (m
m)
Média
Média ± desvio padrão
Min-Max

29
utilização desse ambiente de transição lótico-lêntico e de uma possível estratégia periódica de
reprodução.
As fêmeas de Hypostomus sp1 produzem ovócitos grandes (Figura 26 B), característicos do
gênero e que indicam baixa fecundidade e presença de cuidado parental.
Wertheimeria maculata
Figura 28: A – Wertheimeria maculata. B – Fêmea com ovários maduros.
Espécie símbolo do rio Jequitinhonha, o roncador (W. maculata) é um Doradidae primitivo
que ocorre nas bacias dos rios Jequitinhonha e Pardo. Dentre as poucas referências sobre os peixes
do Jequitinhonha está o trabalho de Vono & Birindelli (2007), onde é sugerido o tipo de desova
parcelado, devido à captura de indivíduos maduros ao longo de todo ano. A dieta da espécie é do
tipo onivora e inclui itens alóctones, como frutos e insetos terrestres (Vono & Birindelli, 2007).

30
Bibliografia
Agostinho, A. A.; Gomes, L. C.; Pelicice, F. M. 2007. Ecologia e manejo dos recursos
pesqueiros em reservatórios do Brasil. Maringá: EDUEM. 501p.
Agostinho, A. A.; Miranda, L. E.; Bini, L. M.; Gomes, L. C.; Thomaz, S. M.; Suzuki, H. I.
1999. Patterns of colonization in neotropical reservoirs, and prognosis on aging. In: Tundisi, J. G.;
Straskraba, M. (ED). Theoretical reservoir ecology and its applications. p. 227-265.
Agostinho, C. S., Hahn, N. S., & Marques, E. E. (2003). Patterns of food resource use by two
congeneric species of piranhas (Serrasalmus) on the upper Paraná River floodplain. Brazilian
Journal of Biology, 63(2), 177-182.
Azevedo, P. G., Melo, R., & Young, R. J. 2011. Feeding and social behavior of the piabanha,
Brycon devillei (Castelnau, 1855)(Characidae: Bryconinae) in the wild, with a note on following
behavior. Neotropical Ichthyology, 9(4), 807-814.
Araújo, F. G., & Santos, L. N. 2001. Distribution of fish assemblages in Lajes reservoir, Rio
de Janeiro, Brazil. Brazilian Journal of Biology, 61(4), 563-576.
Britski, H. A., Birindelli, J. L. O., & Garavello, J. C. (2012). A new species of Leporinus
Agassiz, 1829 from the upper Rio Paraná basin (Characiformes, Anostomidae) with redescription of
L. elongatus Valenciennes, 1850 and L. obtusidens (Valenciennes, 1837). Papéis Avulsos de
Zoologia (São Paulo),52(37), 441-475.
Hojo, R. E. S., Santos, G. B., & Bazzoli, N. 2004. Reproductive biology of Moenkhausia
intermedia (Eigenmann)(Pisces, Characiformes) in Itumbiara Reservoir, Goiás, Brazil. Revista
Brasileira de Zoologia, 21(3), 519-524.
Loures, R. C., & Pompeu, P. S. 2012. Temporal variation in fish community in the tailrace at Três
Marias Hydroelectric Dam, São Francisco River, Brazil. Neotropical Ichthyology, 10(4), 731-740.
Mazzoni, R., & Caramaschi, E. P. 1995. Size structure, sex ratio and onset of sexual maturity
of two species of Hypostomus. Journal of Fish Biology,47(5), 841-849.
Pompeu, P. D. S. (1999). Dieta da pirambeba Serrasalmus brandtii Reinhardt (Teleostei,
Characidae) em quatro lagoas marginais do rio São Francisco, Brasil. Revista Brasileira de
Zoologia, 16, 19-26.
Sato, Y., Bazzoli, N., Rizzo, E., Boschi, M. B., & Miranda, M. O. (2005). Influence of the
Abaeté River on the reproductive success of the neotropical migratory teleost Prochilodus

31
argenteus in the São Francisco River, downstream from the Três Marias Dam, southeastern Brazil.
River Research and Applications, 21(8), 939-950.
Sidlauskas, B. L., & Vari, R. P. (2008). Phylogenetic relationships within the South American
fish family Anostomidae (Teleostei, Ostariophysi, Characiformes). Zoological Journal of the
Linnean Society, 154(1), 70-210.
Silva, A. R., Santos, G. B., & Ratton, T. 2006. Fish community structure of Juramento
reservoir, São Francisco River basin, Minas Gerais, Brazil. Revista Brasileira de Zoologia, 23(3),
832-840.
Vono, V., & Birindelli, J. L. (2007). Natural history of Wertheimeria maculata, a basal
doradid catfish endemic to eastern Brazil (Siluriformes: Doradidae). Ichthyological Exploration Of
Freshwaters, 18(2), 183.


















