
Relatório Final do Projecto PIDDAC nº 841 / 2002horta.0catch.com/aemh/relatorio final pidac...
39
Relatório Final do Projecto PIDDAC nº 841 / 2002 Refinamento das técnicas de inseminação artificial e produção in vitro de embriões em ovinos de raças nacionais, usando sémen congelado Período 2002 – 2004 Responsável: Carla Maria Ferreira Cruz Varanda Marques Equipa: JP Barbas, AEM Horta, MC Baptista, MI Vasques, RD Mascarenhas, RM Pereira, SC, Gonçalves Departamento de Reprodução Animal Estação Zootécnica Nacional Instituto Nacional de Investigação Agrária e das Pescas Vale de Santarém Março 2005
Transcript of Relatório Final do Projecto PIDDAC nº 841 / 2002horta.0catch.com/aemh/relatorio final pidac...

Relatório Final do Projecto PIDDAC nº 841 / 2002
Refinamento das técnicas de inseminação artificial e produção in vitro de embriões em ovinos de raças nacionais, usando sémen congelado
Período 2002 – 2004
Responsável: Carla Maria Ferreira Cruz Varanda Marques Equipa: JP Barbas, AEM Horta, MC Baptista, MI Vasques, RD Mascarenhas, RM Pereira, SC, Gonçalves
Departamento de Reprodução Animal Estação Zootécnica Nacional
Instituto Nacional de Investigação Agrária e das Pescas
Vale de Santarém Março 2005
- 1 -
Refinamento das técnicas de inseminação artificial e produção in vitro de embriões em ovinos de raças nacionais, usando sémen congelado
Relatório Final do Projecto PIDDAC nº 841/2002, EZN – INIAP.
CM Marques, JP Barbas, AEM Horta, MC Baptista, MI Vasques, RD Mascarenhas, RM Pereira,
SC Gonçalves Equipa de Investigação
Código Nome Categoria ETI Total
8365 António Eduardo Monteiro Horta Inv Coord. 0.60
14245 Carla Maria F. C. V. Marques Inv Aux. 0.75
29480 João Pedro S. S-C. Barbas Inv Aux. 0.75
47310 Maria da Conceição C. Baptista Inv Aux. 0.60
56429 Maria Irene A. M. R. Vasques Inv Aux. 0.60
72520 Ramiro Doutel Mascarenhas Inv Prin. 0.30
73650 Rosa Maria L. N. Pereira Inv Aux. 0.30
77240 Sandra Cavaco Gonçalves Inv Aux. 0.75
Localização
Este projecto foi realizado no Departamento de Reprodução Animal da Estação
Zootécnica Nacional, Fonte Boa, Vale de Santarém. As acções de inseminação artificial (IA) com sémen congelado foram realizadas em efectivos da Direcção Regional de Agricultura do Ribatejo e Oeste, no Posto Experimental de Pegões (PEP). Início: Janeiro de 2002 Fim: Dezembro de 2004
Cooperação com outras instituições As acções de congelação de sémen de carneiro, fertilização in vitro de oócitos,
electromiografia, efeito dos dilatadores cervicais e estudos de penetrabilidade cervical foram efectuadas na Estação Zootécnica Nacional, Fonte Boa, Vale de Santarém. As acções de inseminação artificial (IA) com sémen congelado foram realizadas em efectivos da Direcção Regional de Agricultura do Ribatejo e Oeste. Foi também estabelecida uma cooperação com a Faculdade de Medicina Veterinária de Lisboa para optimização das técnicas de avaliação seminal in vitro.

- 2 -
1.Objectivos
A IA com sémen congelado facilita o maneio reprodutivo dos efectivos, permite a concentração das parições nos períodos do ano mais favoráveis e é indispensável nos programas de melhoramento genético das nossas raças autóctones. Todavia, a sua utilização depende da identificação do período ideal de congelação do sémen de ovino, do domínio da técnica de congelação propriamente dita e da melhoria da penetrabilidade cervical com o “pistolet” no momento da IA, talvez o factor mais limitante da implementação da técnica da IA em ovinos. Assim, os principais objectivos deste projecto foram, em primeiro lugar, e após prévia identificação das melhores épocas para a congelação de sémen, prosseguir os trabalhos de congelação com o objectivo da sua utilização na IA em condições de rebanho. Pretendeu-se também avaliar a influência do período de congelação do sémen sobre os resultados da IA em ovelhas inseminadas na época tradicional de reprodução (Primavera).
Um dos principais factores limitantes da IA é a insuficiente abertura do canal cervical. Foram efectuados estudos da contractibilidade do canal cervical através da técnica da electromiografia, tentando identificar-se o sentido, frequência e intensidade das contracções durante o período do estro. Avaliou-se igualmente o efeito sobre estes parâmetros de agentes dilatadores cervicais, decorrendo paralelamente estudos sobre o efeito da administração de dilatadores cervicais, durante o período do estro, na penetrabilidade cervical no momento da IA. Seguidamente foram efectuados ensaios da sua utilização em efectivos inseminados com sémen refrigerado e congelado, para avaliação do seu efeito sobre os parâmetros reprodutivos.
Propôs-se determinar a capacidade fertilizante do sémen de carneiro congelado em duas épocas diferentes, através da tecnologia de fertilização (IVF) e produção de embriões (IVP) in vitro. O sémen foi congelado na época de Outono e época de Inverno, sendo utilizado na fertilização de oócitos maturados in vitro obtidos de fêmeas abatidas ao longo do ano. Sujeitou-se o sémen descongelado a capacitação in vitro, avaliando-se a sua mobilidade individual e o seu índice de aglutinação e mobilidade pós-swim up. Caracterizou-se a fertilidade pelas taxas de clivagem dos oócitos inseminados, de produção de blastócitos ao 7º dia de cultura e avaliação do desenvolvimento embrionário pós-descongelação, quer in vitro (cultura) quer in vivo, por transferência dos embriões para fêmeas sincronizadas. Os resultados sobre a capacitação dos espermatozóides (spz) e a taxa de fertilização in vitro, foram correlacionados com a fertilidade obtida in vivo. 2. Processamento do sémen fresco e congelado
As colheitas de sémen foram efectuadas nas épocas de Primavera, Outono e Inverno,
utilizando o método de recolha com vagina artificial. Após a recolha, o sémen é colocado num banho-maria mantido a 30 ºC, procedendo-se à sua imediata avaliação macroscópica (volume e cor) e microscópica, nomeadamente a mobilidade massal, a mobilidade individual (MI), classificada de acordo com a % de espermatozóides móveis com movimentos progressivos e rectilíneos, a % de espermatozóides vivos e mortos e a % de formas anormais referenciando a sua origem (cabeça, peça intermédia e cauda). A metodologia utilizada na avaliação destes parâmetros seminais encontra-se descrita detalhadamente em várias publicações (Barbas et al, 2001, 2002, 2004). Resumidamente, a mobilidade massal é avaliada numa gota de sémen fresco puro sobre platina aquecida, sendo classificada numa escala de 0-5 de acordo com o vigor dos movimentos de onda dos spz. A MI é classificada em valor percentual (0-100 %) numa gota de sémen diluído (1/100) em soro fisiológico. A % de spz vivos é determinada por contagem do número de spz não corados cerca de 3-4 horas após a realização de um esfregaço utilizando como corante vital a eosina-nigrosina. O mesmo esfregaço é utilizado para contagem do número de spz normais e anormais. Estes valores são expressos em valor percentual sendo as formas

- 3 -
anormais referenciadas à sua origem nomeadamente cabeça, peça intermédia (PI) e cauda. A observação dos esfregaços é realizada com objectiva de imersão utilizado 1000 aumentos. A concentração é determinada no sémen diluído (1/400) por contagem do número de spz mL-1, num espectrofotómetro calibrado para a espécie ovina utilizando um comprimento de onda 550 nanómetros. 2.1 . Processamento e congelação de sémen a) Recolha do sémen. O processo de recolha e avaliação do sémen foi anteriormente descrito.
Unicamente se submeteram ao processo de congelação os ejaculados que apresentaram MI ≥ 65 % e concentrações > 2 mil milhões de spz mL-1. O sémen recolhido foi processado e congelado de acordo com o protocolo descrito por Evans e Maxwell (1990).
b) Temperaturas de trabalho: A temperatura do laboratório de tecnologia do sémen é mantida estabilizada a 25 ºC. Depois da recolha do sémen, o tubo colector é colocado num banho-maria à temperatura de 30 Cº. O diluidor de congelação mantido no banho-Maria é adicionado ao sémen de modo a obter a concentração espermática desejada.
c) Diluição: Após a recolha do sémen determina-se a concentração espermática e o número de espermatozóides totais do ejaculado multiplicando o volume pela concentração espermática. O diluidor de congelação é adicionado numa só etapa (Evans e Maxwell, 1990), de modo a obter uma concentração de 1200 milhões de spz mL-1, pois pretende-se a obtenção de doses de sémen com 300 milhões de spz totais.
d) Preparação para a congelação: Adiciona-se o volume de diluidor de congelação (preparado de véspera) (Evans e Maxwell, 1990) necessário para acertar a concentração de espermatozóides por dose utilizada na IA (300 milhões). Para efectuar a diluição numa só etapa, o sémen é diluído à temperatura de 30 ºC com o diluidor de congelação que tem glicerol (6-8 %), o agente crioprotector mais adequado, e gema de ovo (3-4 %) actuando como protectora da integridade das membranas (Evans e Maxwell, 1990). O diluidor de congelação contém tris e ácido cítrico para tamponização do diluidor, glucose como substracto energético e uma associação de antibióticos (penicilina-estreptomicina) para inibir o desenvolvimento microbiano.
2.2 . Refrigeração do sémen destinado à congelação
Após a diluição do sémen efectuada a 30 ºC, o sémen é armazenado em palhinhas (0,25 ml) tipo Cassou, de cores diferentes, de acordo com o número de carneiros utilizados na mesma sessão de trabalho. Após o enchimento das palhinhas, estas são introduzidas dentro de uma proveta (inserida num copo com água à mesma temperatura) colocada numa bancada de refrigeração mantida a 4 ºC, onde decorre a refrigeração gradual do sémen durante um tempo aproximado de 3,5-4 horas. Decorrida a refrigeração, controla-se a % de espermatozóides móveis antes de iniciar o processo da congelação propriamente dito. Na congelação do sémen de carneiro e de bode o método de congelação em duas etapas não tem vantagens comparativamente à congelação realizada com uma só etapa, daí este método ser preferencial, devido à simplicidade e menor manuseamento do sémen antes da congelação (Evans e Maxwell,1990).
A congelação das palhinhas é efectuada utilizando um equipamento designado por “Floating Semen Freezing Rack” (Minutub GnbH & Co, KG, Germany). Resumidamente, este equipamento é composto por uma caixa de aço inoxidável com cerca de 1m2, um “rack” duplo do mesmo material onde são dispostas transversalmente as mini-palhinhas que está ligado a 4 flutuadores (2 de cada lado, unidos por uma barra transversal) que contêm várias aberturas permitindo a entrada do azoto líquido (vertido previamente na caixa metálica) e uma caixa de esferovite com cerca 95 x 49 x 35 cm onde é inserida esta estrutura.

- 4 -
O processo de congelação devido à acção dos vapores de azoto líquido pode ser dividido nas seguintes etapas:
a) As palhinhas refrigeradas a 4 ºC são dispostas transversalmente no “rack” metálico através de pinças metálicas mantidas à mesma temperatura. É possível regular a altura das palhinhas relativamente ao nível de azoto líquido (vertido previamente na caixa metálica), afinando a altura de fixação do rack nas caixas flutuadoras.
b) Na caixa de aço inoxidável colocada dentro da caixa de esferovite é vertido azoto líquido até este atingir no mínimo 7 cm de altura. Seguidamente o “rack”, manuseado através de dois “ganchos”, é disposto no interior da caixa metálica.
c) A caixa de esferovite é tapada. As palhinhas dispostas no “rack” encontram-se a uma altura de cerca de 4 cm relativamente ao nível do azoto líquido registando-se uma temperatura aproximada de -120 ºC. No decurso da congelação o azoto líquido vai penetrando pelas aberturas das caixas flutuadoras permitindo que no final do processo de congelação, que dura no mínimo 20 minutos, as palhinhas fiquem submersas no azoto.
d) Finalmente as palhinhas são cuidadosamente retiradas e armazenadas nos respectivos canisteres dos contentores onde permanecem até posterior utilização.
A apreciação da aptidão dos espermatozóides para suportar o processo de congelação/descongelação é realizada 24 horas mais tarde, retirando-se uma palhinha do contentor de armazenamento para um copo de vidro com água a 37-38 ºC durante 50 segundos. Seguidamente são cuidadosamente secas e o seu conteúdo é diluído em soro fisiológico (1 ml) contido em tubos de ensaio mantidos no banho-maria à mesma temperatura. Depois de uma breve agitação dos tubos num “vortex” é avaliado a % de espermatozóides móveis, realizando-se um esfregaço com eosina-nigrosina destinado a avaliar a % de spz vivos e de formas anormais. Só ficarão armazenados os ejaculados que apresentem pelo menos 40 % de MI, determinada após a descongelação de 1-2 palhinhas. A comparação destes parâmetros seminais no sémen fresco e descongelado constituem objectivos do nosso projecto. 3. Resultados 3.1. Variação das características seminais no sémen fresco e descongelado
Este trabalho foi realizado no Laboratório de Tecnologia do Sémen do Departamento de Reprodução da Estação Zootécnica Nacional, de Janeiro de 2002 até Dezembro de 2004. Nos primeiros 3 meses decorreu o treino de recolha de sémen dos carneiros de raça Saloia pertencentes ao Posto Experimental de Pegões (PEP) e a afinação das técnicas de recolha e processamento de sémen fresco que anteriormente foram descritas. Neste período foram igualmente ensaiados os diluidores destinados à congelação de sémen, designadamente a sua composição e concentração dos constituintes, e afinadas as técnicas de refrigeração e congelação propriamente ditas. Os carneiros foram mantidos num estábulo anexo ao Laboratório de Tecnologia do Sémen, sendo alimentados com fenos de aveia/ervilhaca, silagem de milho e concentrado comercial. As colheitas de sémen destinadas à execução deste projecto tiveram início em Março-Abril de 2002 e foram realizadas sempre que possível quinzenalmente durante as épocas de Primavera, Outono e Inverno. Neste ensaio, foram utilizados 5 carneiros de raça Saloia em bom estado sanitário e condição corporal, não sendo possível a utilização simultânea de todos os carneiros durante o período de ensaio, existindo em qualquer das estações avaliadas apenas 3 carneiros em estudo. Foram utilizados 83 ejaculados para avaliação estatística de resultados. Ejaculados que não tivessem boa qualidade para serem processados foram rejeitados. As variáveis independentes estudadas foram a época do ano, o factor carneiro e a interacção época x carneiro. No sémen fresco (SF) foram avaliados o volume (ml), a MI (%), a
- 5 -
concentração espermática, a % de spz vivos e normais e a % de anomalias da cabeça, PI e cauda, constiuindo as variáveis dependentes na análise estatística. Nos 5 carneiros utilizados e considerando as épocas de Primavera, Outono e Inverno, não foram apreciados estatisticamente os resultados quando o número de ejaculados por época do ano e por carneiro foram ≤ 3.
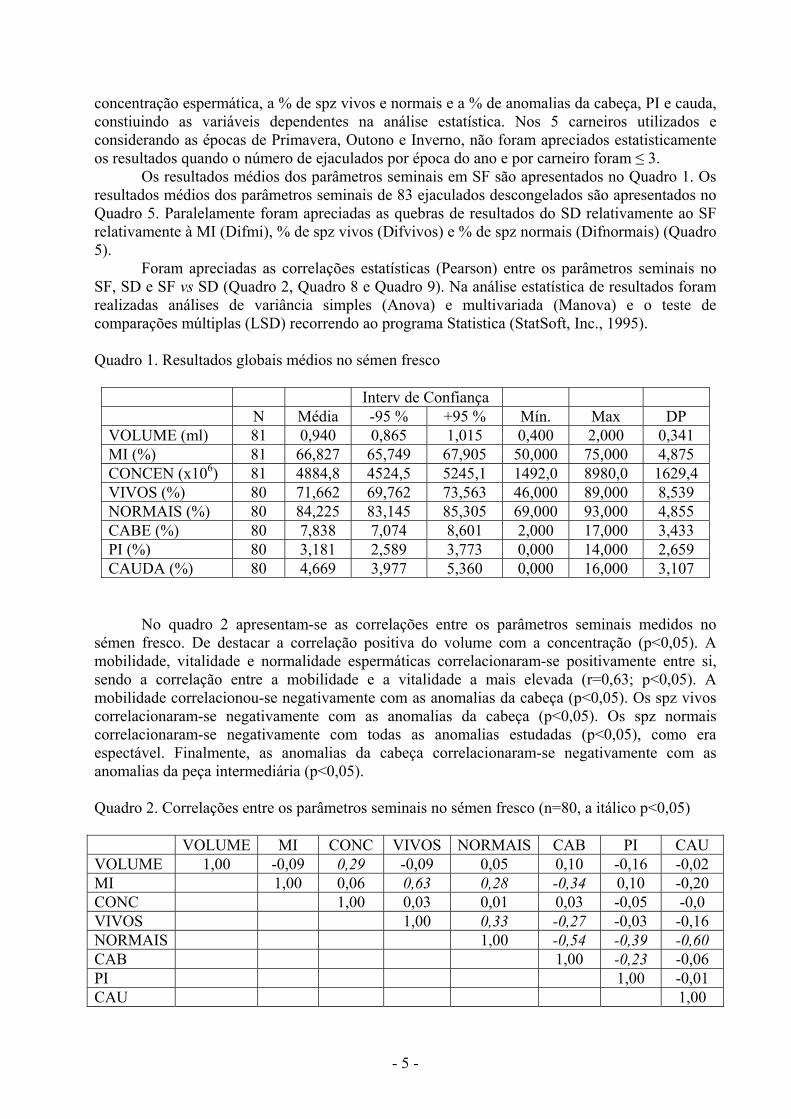
Os resultados médios dos parâmetros seminais em SF são apresentados no Quadro 1. Os resultados médios dos parâmetros seminais de 83 ejaculados descongelados são apresentados no Quadro 5. Paralelamente foram apreciadas as quebras de resultados do SD relativamente ao SF relativamente à MI (Difmi), % de spz vivos (Difvivos) e % de spz normais (Difnormais) (Quadro 5).
Foram apreciadas as correlações estatísticas (Pearson) entre os parâmetros seminais no SF, SD e SF vs SD (Quadro 2, Quadro 8 e Quadro 9). Na análise estatística de resultados foram realizadas análises de variância simples (Anova) e multivariada (Manova) e o teste de comparações múltiplas (LSD) recorrendo ao programa Statistica (StatSoft, Inc., 1995). Quadro 1. Resultados globais médios no sémen fresco
Interv de Confiança N Média -95 % +95 % Mín. Max DP
VOLUME (ml) 81 0,940 0,865 1,015 0,400 2,000 0,341 MI (%) 81 66,827 65,749 67,905 50,000 75,000 4,875 CONCEN (x106) 81 4884,8 4524,5 5245,1 1492,0 8980,0 1629,4 VIVOS (%) 80 71,662 69,762 73,563 46,000 89,000 8,539 NORMAIS (%) 80 84,225 83,145 85,305 69,000 93,000 4,855 CABE (%) 80 7,838 7,074 8,601 2,000 17,000 3,433 PI (%) 80 3,181 2,589 3,773 0,000 14,000 2,659 CAUDA (%) 80 4,669 3,977 5,360 0,000 16,000 3,107
No quadro 2 apresentam-se as correlações entre os parâmetros seminais medidos no
sémen fresco. De destacar a correlação positiva do volume com a concentração (p<0,05). A mobilidade, vitalidade e normalidade espermáticas correlacionaram-se positivamente entre si, sendo a correlação entre a mobilidade e a vitalidade a mais elevada (r=0,63; p<0,05). A mobilidade correlacionou-se negativamente com as anomalias da cabeça (p<0,05). Os spz vivos correlacionaram-se negativamente com as anomalias da cabeça (p<0,05). Os spz normais correlacionaram-se negativamente com todas as anomalias estudadas (p<0,05), como era espectável. Finalmente, as anomalias da cabeça correlacionaram-se negativamente com as anomalias da peça intermediária (p<0,05). Quadro 2. Correlações entre os parâmetros seminais no sémen fresco (n=80, a itálico p<0,05)
VOLUME MI CONC VIVOS NORMAIS CAB PI CAU VOLUME 1,00 -0,09 0,29 -0,09 0,05 0,10 -0,16 -0,02 MI 1,00 0,06 0,63 0,28 -0,34 0,10 -0,20 CONC 1,00 0,03 0,01 0,03 -0,05 -0,0 VIVOS 1,00 0,33 -0,27 -0,03 -0,16 NORMAIS 1,00 -0,54 -0,39 -0,60 CAB 1,00 -0,23 -0,06 PI 1,00 -0,01 CAU 1,00
- 6 -
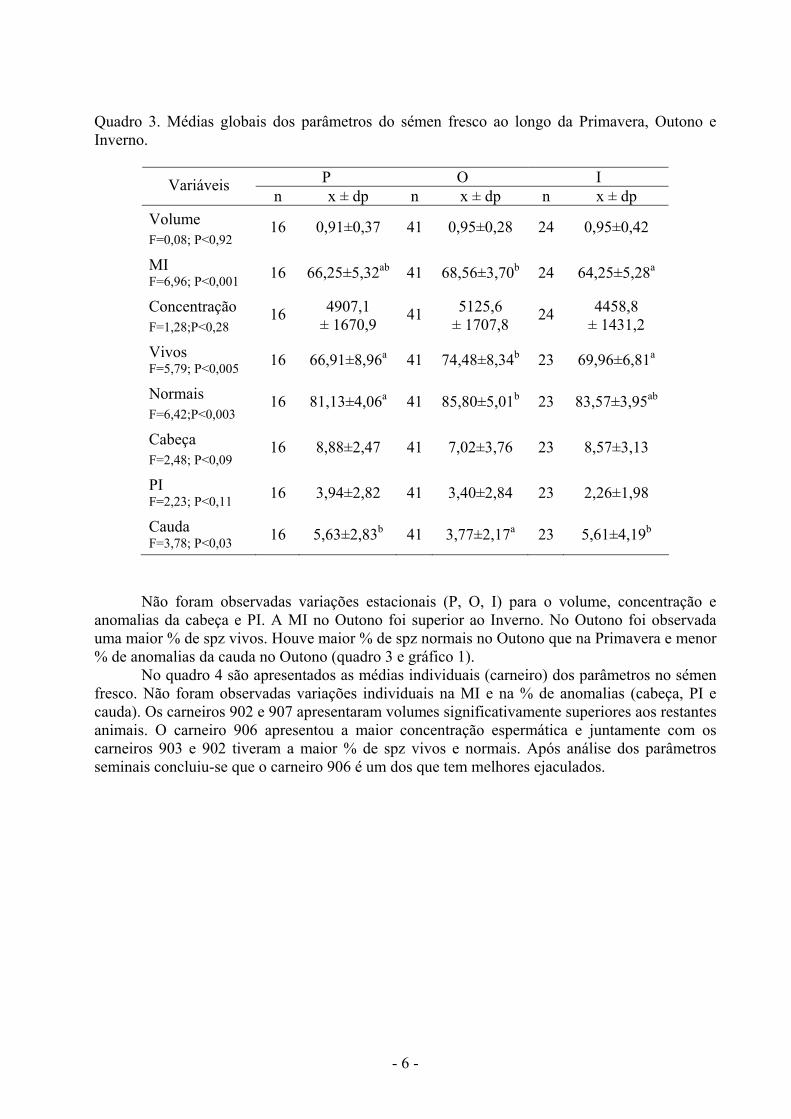
Quadro 3. Médias globais dos parâmetros do sémen fresco ao longo da Primavera, Outono e Inverno.
P O I Variáveis n x ± dp n x ± dp n x ± dp
Volume F=0,08; P<0,92
16 0,91±0,37 41 0,95±0,28 24 0,95±0,42
MI F=6,96; P<0,001 16 66,25±5,32ab 41 68,56±3,70b 24 64,25±5,28a
Concentração F=1,28;P<0,28
16 4907,1 ± 1670,9 41 5125,6
± 1707,8 24 4458,8 ± 1431,2
Vivos F=5,79; P<0,005 16 66,91±8,96a 41 74,48±8,34b 23 69,96±6,81a
Normais F=6,42;P<0,003
16 81,13±4,06a 41 85,80±5,01b 23 83,57±3,95ab
Cabeça F=2,48; P<0,09
16 8,88±2,47 41 7,02±3,76 23 8,57±3,13
PI F=2,23; P<0,11 16 3,94±2,82 41 3,40±2,84 23 2,26±1,98
Cauda F=3,78; P<0,03 16 5,63±2,83b 41 3,77±2,17a 23 5,61±4,19b
Não foram observadas variações estacionais (P, O, I) para o volume, concentração e anomalias da cabeça e PI. A MI no Outono foi superior ao Inverno. No Outono foi observada uma maior % de spz vivos. Houve maior % de spz normais no Outono que na Primavera e menor % de anomalias da cauda no Outono (quadro 3 e gráfico 1).
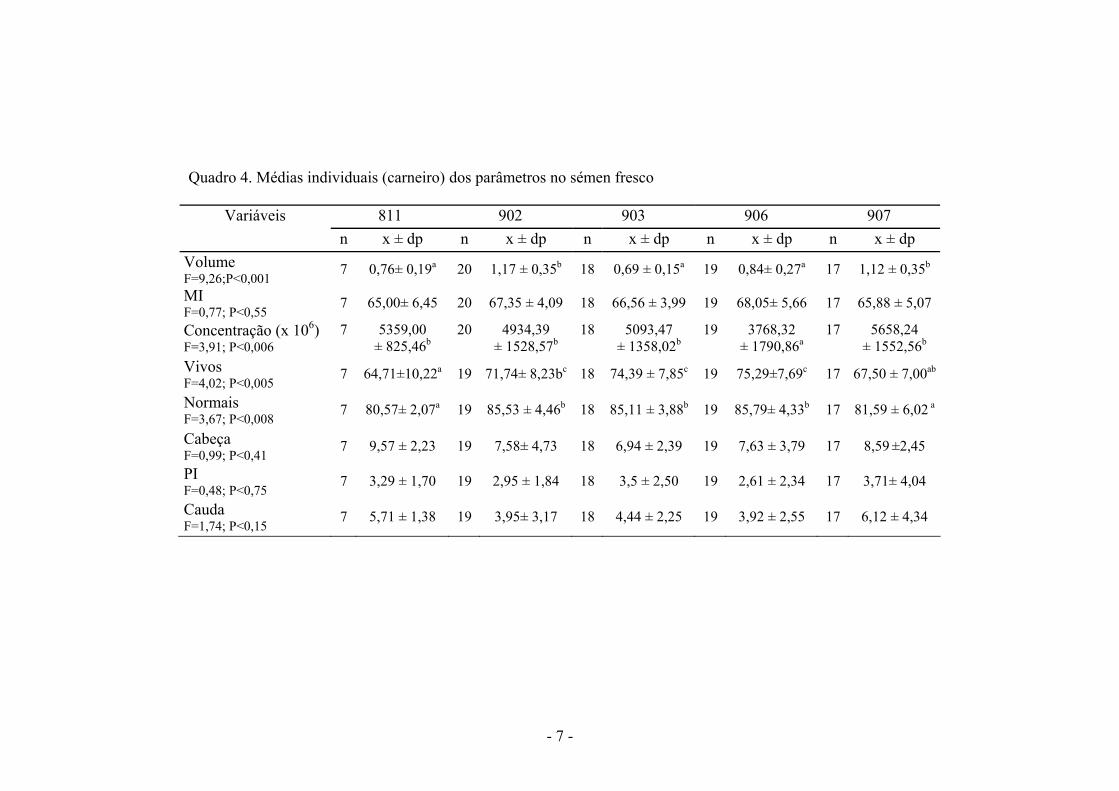
No quadro 4 são apresentados as médias individuais (carneiro) dos parâmetros no sémen fresco. Não foram observadas variações individuais na MI e na % de anomalias (cabeça, PI e cauda). Os carneiros 902 e 907 apresentaram volumes significativamente superiores aos restantes animais. O carneiro 906 apresentou a maior concentração espermática e juntamente com os carneiros 903 e 902 tiveram a maior % de spz vivos e normais. Após análise dos parâmetros seminais concluiu-se que o carneiro 906 é um dos que tem melhores ejaculados.
- 7 -
Quadro 4. Médias individuais (carneiro) dos parâmetros no sémen fresco
811 902 903 906 907 Variáveis n x ± dp n x ± dp n x ± dp n x ± dp n x ± dp
Volume F=9,26;P<0,001 7 0,76± 0,19a 20 1,17 ± 0,35b 18 0,69 ± 0,15a 19 0,84± 0,27a 17 1,12 ± 0,35b
MI F=0,77; P<0,55 7 65,00± 6,45 20 67,35 ± 4,09 18 66,56 ± 3,99 19 68,05± 5,66 17 65,88 ± 5,07
Concentração (x 106) F=3,91; P<0,006
7 5359,00 ± 825,46b
20 4934,39 ± 1528,57b
18 5093,47 ± 1358,02b
19 3768,32 ± 1790,86a
17 5658,24 ± 1552,56b
Vivos F=4,02; P<0,005 7 64,71±10,22a 19 71,74± 8,23bc 18 74,39 ± 7,85c 19 75,29±7,69c 17 67,50 ± 7,00ab
Normais F=3,67; P<0,008 7 80,57± 2,07a 19 85,53 ± 4,46b 18 85,11 ± 3,88b 19 85,79± 4,33b 17 81,59 ± 6,02 a
Cabeça F=0,99; P<0,41 7 9,57 ± 2,23 19 7,58± 4,73 18 6,94 ± 2,39 19 7,63 ± 3,79 17 8,59 ±2,45
PI F=0,48; P<0,75 7 3,29 ± 1,70 19 2,95 ± 1,84 18 3,5 ± 2,50 19 2,61 ± 2,34 17 3,71± 4,04
Cauda F=1,74; P<0,15 7 5,71 ± 1,38 19 3,95± 3,17 18 4,44 ± 2,25 19 3,92 ± 2,55 17 6,12 ± 4,34
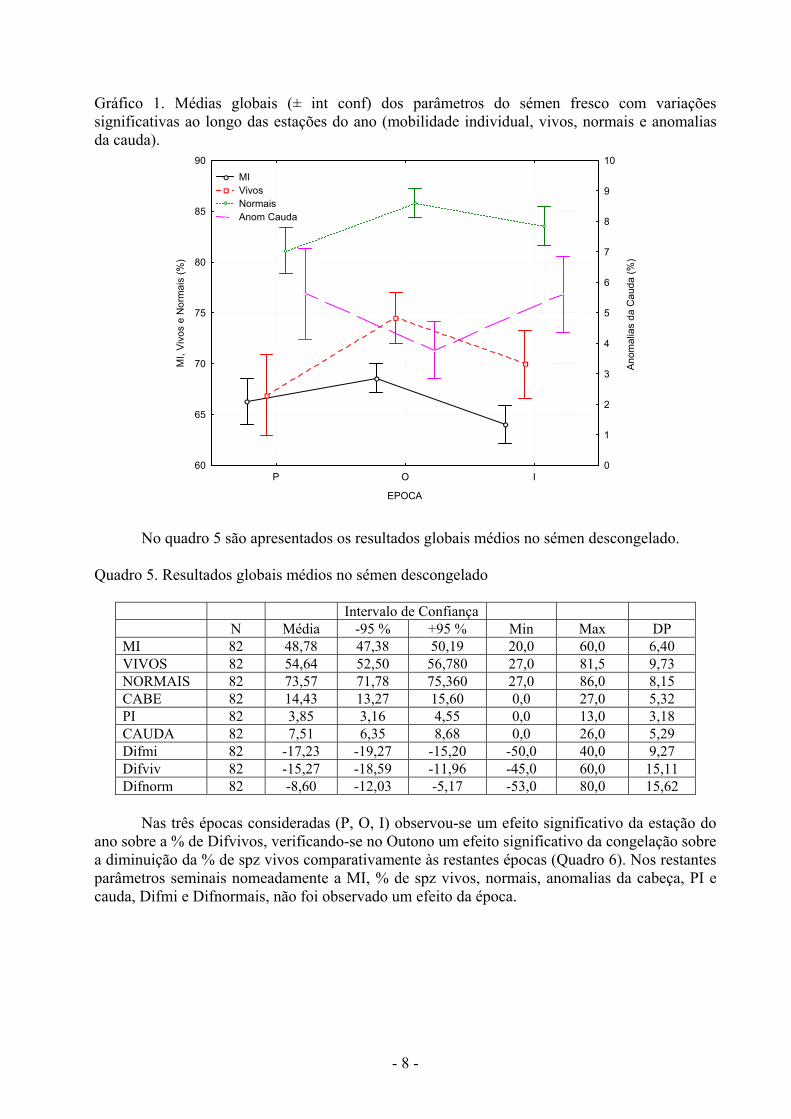
- 8 -
Gráfico 1. Médias globais (± int conf) dos parâmetros do sémen fresco com variações significativas ao longo das estações do ano (mobilidade individual, vivos, normais e anomalias da cauda).
P O I
EPOCA
60
65
70
75
80
85
90
MI,
Vivo
s e
Nor
mai
s (%
)
0
1
2
3
4
5
6
7
8
9
10
Anom
alia
s da
Cau
da (%
)
MI Vivos Normais Anom Cauda
No quadro 5 são apresentados os resultados globais médios no sémen descongelado. Quadro 5. Resultados globais médios no sémen descongelado
Intervalo de Confiança N Média -95 % +95 % Min Max DP
MI 82 48,78 47,38 50,19 20,0 60,0 6,40 VIVOS 82 54,64 52,50 56,780 27,0 81,5 9,73 NORMAIS 82 73,57 71,78 75,360 27,0 86,0 8,15 CABE 82 14,43 13,27 15,60 0,0 27,0 5,32 PI 82 3,85 3,16 4,55 0,0 13,0 3,18 CAUDA 82 7,51 6,35 8,68 0,0 26,0 5,29 Difmi 82 -17,23 -19,27 -15,20 -50,0 40,0 9,27 Difviv 82 -15,27 -18,59 -11,96 -45,0 60,0 15,11 Difnorm 82 -8,60 -12,03 -5,17 -53,0 80,0 15,62
Nas três épocas consideradas (P, O, I) observou-se um efeito significativo da estação do
ano sobre a % de Difvivos, verificando-se no Outono um efeito significativo da congelação sobre a diminuição da % de spz vivos comparativamente às restantes épocas (Quadro 6). Nos restantes parâmetros seminais nomeadamente a MI, % de spz vivos, normais, anomalias da cabeça, PI e cauda, Difmi e Difnormais, não foi observado um efeito da época.
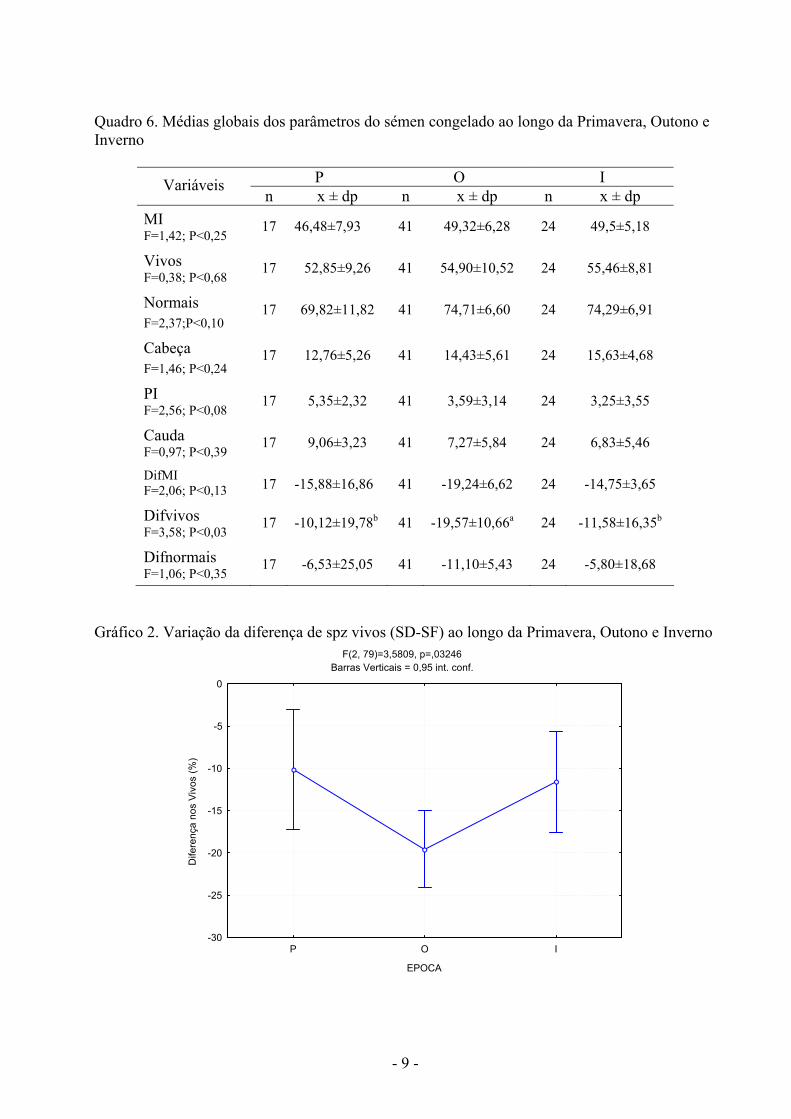
- 9 -
Quadro 6. Médias globais dos parâmetros do sémen congelado ao longo da Primavera, Outono e Inverno
P O I Variáveis n x ± dp n x ± dp n x ± dp
MI F=1,42; P<0,25 17 46,48±7,93 41 49,32±6,28 24 49,5±5,18
Vivos F=0,38; P<0,68 17 52,85±9,26 41 54,90±10,52 24 55,46±8,81
Normais F=2,37;P<0,10
17 69,82±11,82 41 74,71±6,60 24 74,29±6,91
Cabeça F=1,46; P<0,24
17 12,76±5,26 41 14,43±5,61 24 15,63±4,68
PI F=2,56; P<0,08 17 5,35±2,32 41 3,59±3,14 24 3,25±3,55
Cauda F=0,97; P<0,39 17 9,06±3,23 41 7,27±5,84 24 6,83±5,46
DifMI F=2,06; P<0,13 17 -15,88±16,86 41 -19,24±6,62 24 -14,75±3,65
Difvivos F=3,58; P<0,03 17 -10,12±19,78b 41 -19,57±10,66a 24 -11,58±16,35b
Difnormais F=1,06; P<0,35 17 -6,53±25,05 41 -11,10±5,43 24 -5,80±18,68
Gráfico 2. Variação da diferença de spz vivos (SD-SF) ao longo da Primavera, Outono e Inverno
F(2, 79)=3,5809, p=,03246Barras Verticais = 0,95 int. conf.
P O I
EPOCA
-30
-25
-20
-15
-10
-5
0
Dife
renç
a no
s Vi
vos
(%)
- 10 -
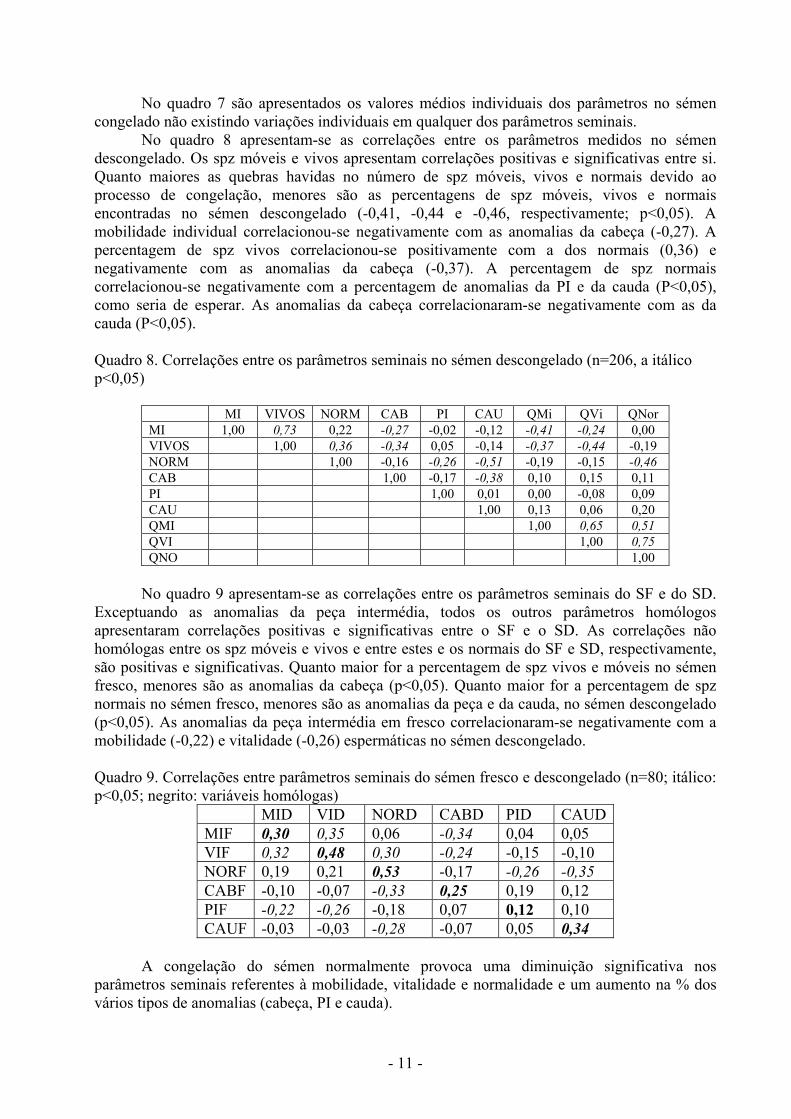
Quadro 7. Médias individuais (carneiro) dos parâmetros no sémen congelado
811 902 903 906 907 Variáveis n x ± dp n x ± dp n x ± dp n x ± dp n x ± dp
MI F=1,23; P<0,31 7 48,29±3,73 20 50,70±5,97 18 49,44±6,78 20 48,65± 5,60 17 46,18 ± 7,81
Vivos F=0,724; P<0,58 7 54,00±5,69 20 55,13± 10,85 18 54,81 ± 10,06 20 56,90±9,35 17 51,50±9,91
Normais F=1,37; P<0,25 7 70,71± 3,73 20 73,95 ± 6,49 18 75,92 ± 4,92 20 74,78± 12,54 17 70,41 ± 6,80
Cabeça F=1,11; P<0,36 7 15,00 ± 4,47 20 15,55± 5,49 18 14,28 ± 4,66 20 12,43±5,66 17 15,41 ± 5,55
PI F=2,14; P<0,08 7 6,43 ± 1,40 20 3,20 ± 3,05 18 2,78 ± 3,17 20 4,35± 3,30 17 4,12 ± 3,28
Cauda F=1,43; P<0,23 7 7,86 ± 1,86 20 7,30± 5,03 18 6,47 ± 5,84 20 6,38 ± 4,66 17 10,06 ±6,22
Difmi F=0,40; P<0,80 7 -16,71 ± 5,22 20 -16,65± 5,82 18 -17,11 ± 7,11 20 -16,00 ± 14,32 17 -19,71± 8,85
Difvivos F=0,64; P<0,63 7 -10,71± 8,44 20 -13,03 ± 19,34 18 -19,58 ± 11,88 20 -14,63 ± 19,51 17 -16,00 ± 6,74
Difnormais F=0,23; P<0,92 7 -9,86 ± 4,14 20 -7,30± 20,99 18 -9,19 ± 4,76 20 -6,73 ± 22,88 17 -11,18± 6,71
- 11 -
No quadro 7 são apresentados os valores médios individuais dos parâmetros no sémen congelado não existindo variações individuais em qualquer dos parâmetros seminais.
No quadro 8 apresentam-se as correlações entre os parâmetros medidos no sémen descongelado. Os spz móveis e vivos apresentam correlações positivas e significativas entre si. Quanto maiores as quebras havidas no número de spz móveis, vivos e normais devido ao processo de congelação, menores são as percentagens de spz móveis, vivos e normais encontradas no sémen descongelado (-0,41, -0,44 e -0,46, respectivamente; p<0,05). A mobilidade individual correlacionou-se negativamente com as anomalias da cabeça (-0,27). A percentagem de spz vivos correlacionou-se positivamente com a dos normais (0,36) e negativamente com as anomalias da cabeça (-0,37). A percentagem de spz normais correlacionou-se negativamente com a percentagem de anomalias da PI e da cauda (P<0,05), como seria de esperar. As anomalias da cabeça correlacionaram-se negativamente com as da cauda (P<0,05). Quadro 8. Correlações entre os parâmetros seminais no sémen descongelado (n=206, a itálico p<0,05)
MI VIVOS NORM CAB PI CAU QMi QVi QNor MI 1,00 0,73 0,22 -0,27 -0,02 -0,12 -0,41 -0,24 0,00 VIVOS 1,00 0,36 -0,34 0,05 -0,14 -0,37 -0,44 -0,19 NORM 1,00 -0,16 -0,26 -0,51 -0,19 -0,15 -0,46 CAB 1,00 -0,17 -0,38 0,10 0,15 0,11 PI 1,00 0,01 0,00 -0,08 0,09 CAU 1,00 0,13 0,06 0,20 QMI 1,00 0,65 0,51 QVI 1,00 0,75 QNO 1,00
No quadro 9 apresentam-se as correlações entre os parâmetros seminais do SF e do SD. Exceptuando as anomalias da peça intermédia, todos os outros parâmetros homólogos apresentaram correlações positivas e significativas entre o SF e o SD. As correlações não homólogas entre os spz móveis e vivos e entre estes e os normais do SF e SD, respectivamente, são positivas e significativas. Quanto maior for a percentagem de spz vivos e móveis no sémen fresco, menores são as anomalias da cabeça (p<0,05). Quanto maior for a percentagem de spz normais no sémen fresco, menores são as anomalias da peça e da cauda, no sémen descongelado (p<0,05). As anomalias da peça intermédia em fresco correlacionaram-se negativamente com a mobilidade (-0,22) e vitalidade (-0,26) espermáticas no sémen descongelado. Quadro 9. Correlações entre parâmetros seminais do sémen fresco e descongelado (n=80; itálico: p<0,05; negrito: variáveis homólogas)
MID VID NORD CABD PID CAUD MIF 0,30 0,35 0,06 -0,34 0,04 0,05 VIF 0,32 0,48 0,30 -0,24 -0,15 -0,10 NORF 0,19 0,21 0,53 -0,17 -0,26 -0,35 CABF -0,10 -0,07 -0,33 0,25 0,19 0,12 PIF -0,22 -0,26 -0,18 0,07 0,12 0,10 CAUF -0,03 -0,03 -0,28 -0,07 0,05 0,34
A congelação do sémen normalmente provoca uma diminuição significativa nos
parâmetros seminais referentes à mobilidade, vitalidade e normalidade e um aumento na % dos vários tipos de anomalias (cabeça, PI e cauda).

- 12 -
Gráfico 3. Interacção entre a época e o tipo de sémen, fresco e descongelado, para a MI, Vivos e Normais
Interacção Época x SémenBarras Verticais 0,95 int conf
Sémen Fresco
P O I35
40
45
50
55
60
65
70
75
80
85
90
95P
erce
ntag
em
Sémen Descongelado
P O I
MI VIVOS NORMAIS
MI: F[2;157] = 3,0; p<0,05
No gráfico 3 são representadas as interacções época x tipo de sémen para a MI, % spz
vivos e normais, verificando-se uma interacção significativa com a MI (F=3,00; P<0,05). No gráfico 4 são apresentadas as interacções época x tipo de sémen para os vários tipos de anomalias. Não foi observado um efeito significativo do tipo de sémen (época x tipo de sémen) na % de anomalias da PI. Gráfico 4. Interacção entre a época e o tipo de sémen, para as anomalias da cabeça, PI e cauda
Interacção Época x SémenBarras Verticais 0,95 int conf
Sémen Fresco
P O I-2
0
2
4
6
8
10
12
14
16
18
20
Per
cent
agem
Sémen Descongelado
P O I
CAB PI CAUDA
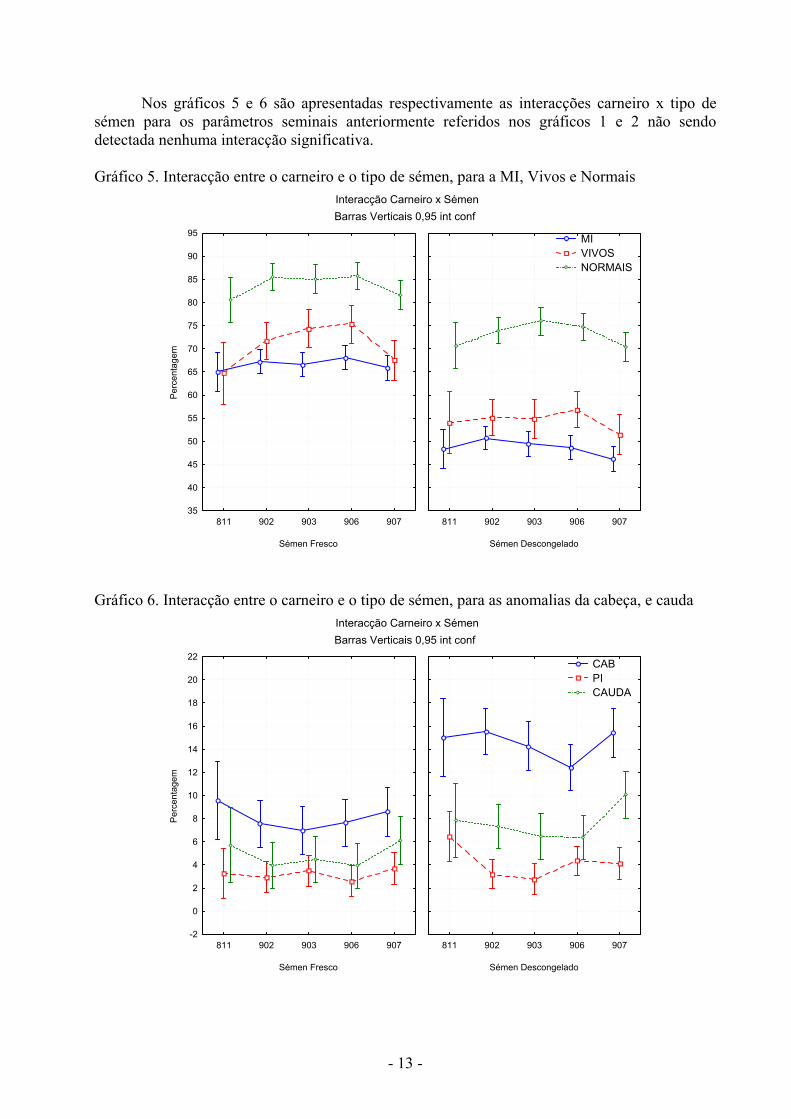
- 13 -
Nos gráficos 5 e 6 são apresentadas respectivamente as interacções carneiro x tipo de sémen para os parâmetros seminais anteriormente referidos nos gráficos 1 e 2 não sendo detectada nenhuma interacção significativa. Gráfico 5. Interacção entre o carneiro e o tipo de sémen, para a MI, Vivos e Normais
Interacção Carneiro x SémenBarras Verticais 0,95 int conf
Sémen Fresco
811 902 903 906 90735
40
45
50
55
60
65
70
75
80
85
90
95
Per
cent
agem
Sémen Descongelado
811 902 903 906 907
MI VIVOS NORMAIS
Gráfico 6. Interacção entre o carneiro e o tipo de sémen, para as anomalias da cabeça, e cauda
Interacção Carneiro x SémenBarras Verticais 0,95 int conf
Sémen Fresco
811 902 903 906 907-2
0
2
4
6
8
10
12
14
16
18
20
22
Per
cent
agem
Sémen Descongelado
811 902 903 906 907
CAB PI CAUDA

- 14 -
3.2. Variação das características seminais em função da época e do carneiro (Interacções)
a) Sémen fresco Anteriormente apresentámos a variação estacional da qualidade dos ejaculados frescos e
descongelados provenientes de 5 carneiros em três épocas do ano, nomeadamente Primavera, Outono e Inverno. Neste trabalho pretendemos igualmente avaliar a qualidade dos ejaculados recolhidos em duas épocas do ano nomeadamente no Outono (O) e Inverno (I) e respectiva aptidão à congelação-descongelação.
A análise multivariada para avaliar a interacção época (O e I) x carneiro não foi possível ser realizada em todos os carneiros, sendo excluído um carneiro (801) da avaliação dos resultados, pois neste animal só foi possível recolher sémen no Inverno.
Quadro 10. Efeitos específicos da época (2 épocas x 4 carneiros)
O I Variáveis
n x ± dp n X ± dp MI F=13,09; P<0,0006
41 68,56 ± 3,70 a 24 64,25 ± 5,28 b
Concentração F=9,83; P<0,003
41 5125,57 ± 1707,76 a
24 4458,75 ±1431,23 b
PI F=3,97; P<0,05
41 3,40± 2,84 a 23 2,26± 1,98 b
Foram observados efeitos específicos significativos da época sobre a MI, concentração e
% de anomalias da PI. No Outono foram registadas maiores mobilidades individuais (MI), concentrações e anomalias da PI do que no Inverno (Quadro 10). Quadro 11. Efeitos específicos do carneiro (4 carneiros x 2 épocas)
902 903 906 907 Variáveis n x ± dp n x ± dp n x ± dp n x ± dp
Volume F=12,11; P<0,0001
20 1,17 ± 0,35 c 18 0,69± 0,15 a 16 0,91 ± 0,23 b 11 1,02 ± 0,38 bc
Concentração F=6,83; P<0,003
20 4934,39 ± 1528,57 ab
18 5093,47 ± 1358,02 ab
16 4036,13 ± 1787,93 a
11 5655,46 ± 1670,90 b
Foram observadas variações individuais significativas relativamente ao volume e
concentração (Quadro 11). Os carneiros 902 e 907 tiveram os volumes mais elevados. As concentrações foram idênticas em três carneiros, tendo o carneiro 906 concentrações inferiores.
No sémen fresco foram observadas interacções significativas época x carneiro para o volume e concentração (Quadro 12).
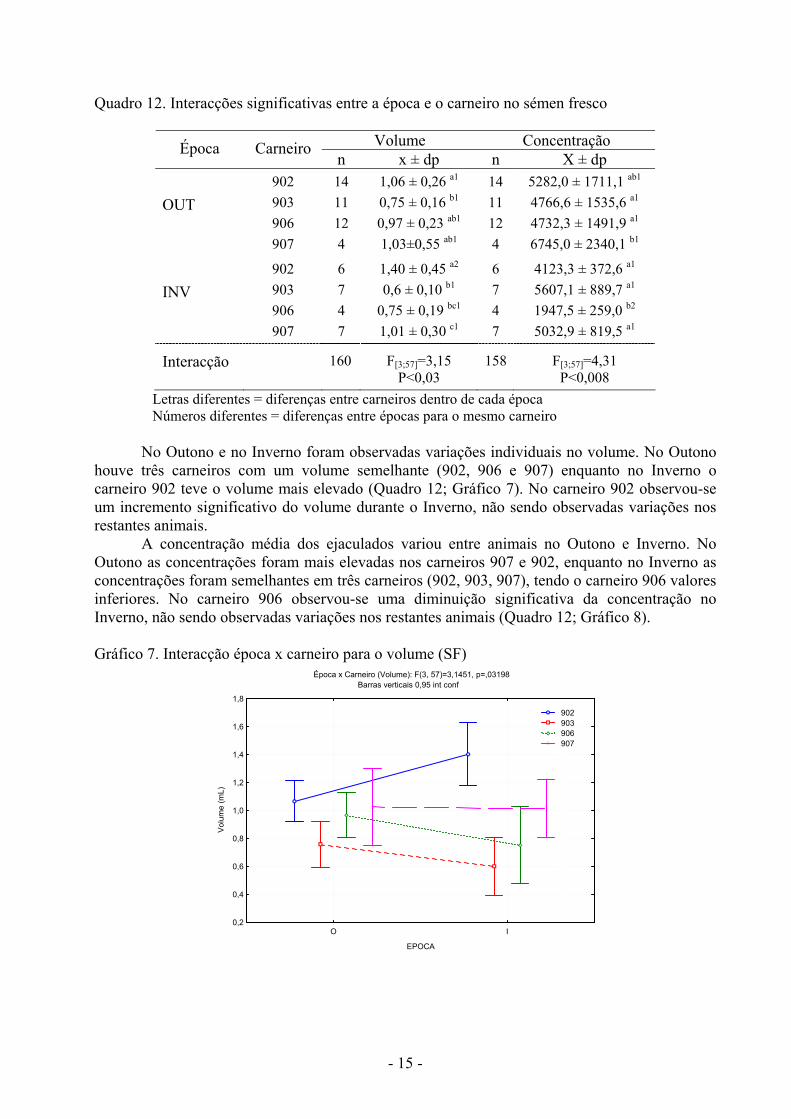
- 15 -
Quadro 12. Interacções significativas entre a época e o carneiro no sémen fresco
Volume Concentração Época Carneiro n x ± dp n X ± dp
OUT
902 903 906 907
14 11 12 4
1,06 ± 0,26 a1 0,75 ± 0,16 b1 0,97 ± 0,23 ab1 1,03±0,55 ab1
14 11 12 4
5282,0 ± 1711,1 ab1 4766,6 ± 1535,6 a1 4732,3 ± 1491,9 a1 6745,0 ± 2340,1 b1
INV
902 903 906 907
6 7 4 7
1,40 ± 0,45 a2 0,6 ± 0,10 b1
0,75 ± 0,19 bc1 1,01 ± 0,30 c1
6 7 4 7
4123,3 ± 372,6 a1 5607,1 ± 889,7 a1 1947,5 ± 259,0 b2 5032,9 ± 819,5 a1
Interacção 160 F[3;57]=3,15 P<0,03
158 F[3;57]=4,31 P<0,008
Letras diferentes = diferenças entre carneiros dentro de cada época Números diferentes = diferenças entre épocas para o mesmo carneiro
No Outono e no Inverno foram observadas variações individuais no volume. No Outono
houve três carneiros com um volume semelhante (902, 906 e 907) enquanto no Inverno o carneiro 902 teve o volume mais elevado (Quadro 12; Gráfico 7). No carneiro 902 observou-se um incremento significativo do volume durante o Inverno, não sendo observadas variações nos restantes animais.
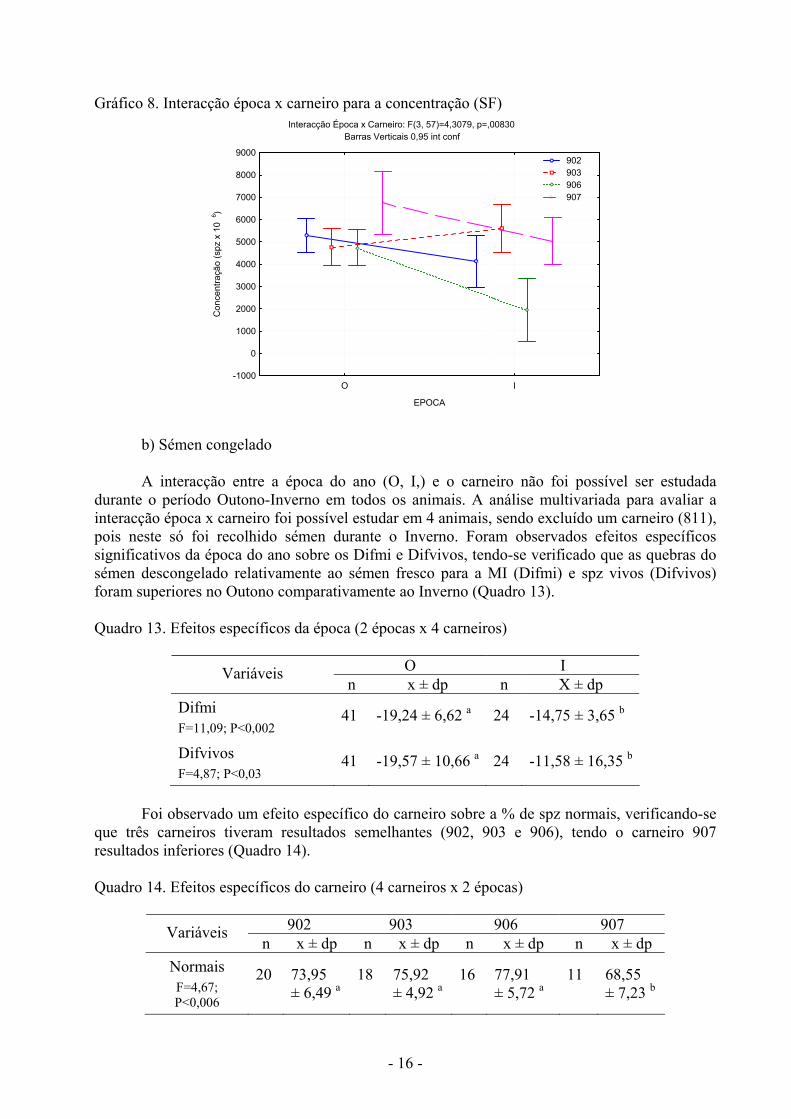
A concentração média dos ejaculados variou entre animais no Outono e Inverno. No Outono as concentrações foram mais elevadas nos carneiros 907 e 902, enquanto no Inverno as concentrações foram semelhantes em três carneiros (902, 903, 907), tendo o carneiro 906 valores inferiores. No carneiro 906 observou-se uma diminuição significativa da concentração no Inverno, não sendo observadas variações nos restantes animais (Quadro 12; Gráfico 8). Gráfico 7. Interacção época x carneiro para o volume (SF)
Época x Carneiro (Volume): F(3, 57)=3,1451, p=,03198Barras verticais 0,95 int conf
O I
EPOCA
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
Vol
ume
(mL)
902 903 906 907
- 16 -
Gráfico 8. Interacção época x carneiro para a concentração (SF) Interacção Época x Carneiro: F(3, 57)=4,3079, p=,00830
Barras Verticais 0,95 int conf
O I
EPOCA
-1000
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
Con
cent
raçã
o (s
pz x
10
6 )
902 903 906 907
b) Sémen congelado A interacção entre a época do ano (O, I,) e o carneiro não foi possível ser estudada
durante o período Outono-Inverno em todos os animais. A análise multivariada para avaliar a interacção época x carneiro foi possível estudar em 4 animais, sendo excluído um carneiro (811), pois neste só foi recolhido sémen durante o Inverno. Foram observados efeitos específicos significativos da época do ano sobre os Difmi e Difvivos, tendo-se verificado que as quebras do sémen descongelado relativamente ao sémen fresco para a MI (Difmi) e spz vivos (Difvivos) foram superiores no Outono comparativamente ao Inverno (Quadro 13).
Quadro 13. Efeitos específicos da época (2 épocas x 4 carneiros)
O I Variáveis
n x ± dp n X ± dp Difmi F=11,09; P<0,002
41 -19,24 ± 6,62 a 24 -14,75 ± 3,65 b
Difvivos F=4,87; P<0,03
41 -19,57 ± 10,66 a 24 -11,58 ± 16,35 b
Foi observado um efeito específico do carneiro sobre a % de spz normais, verificando-se
que três carneiros tiveram resultados semelhantes (902, 903 e 906), tendo o carneiro 907 resultados inferiores (Quadro 14). Quadro 14. Efeitos específicos do carneiro (4 carneiros x 2 épocas)
902 903 906 907 Variáveis n x ± dp n x ± dp n x ± dp n x ± dp
Normais F=4,67; P<0,006
20 73,95 ± 6,49 a
18 75,92 ± 4,92 a
16 77,91 ± 5,72 a
11 68,55 ± 7,23 b

- 17 -
Não foram observadas interacções significativas entre as épocas (Outono e Inverno) e os
carneiros (902, 903, 906 e 907) para os parâmetros seminais no sémen congelado. 4. Inseminação Artificial de Ovinos Saloios com Sémen Congelado
Existindo um protocolo de colaboração entre o Posto Experimental de Pegões (PEP) e a Estação Zootécnica Nacional, decorreram nos anos de 2002, 2003 e 2004 ensaios de IA com sémen congelado em ovinos de raça Saloia. Os carneiros Saloios foram utilizados no estudo de avaliação de ejaculados em fresco e congelados em duas épocas do ano, designadamente no Outono e no Inverno e na produção de doses de sémen congelado para inseminação artificial por via cervical no Posto Experimental de Pegões. Foram inseminadas 440 ovelhas nos anos de 2002 a 2004 com sémen congelado em duas épocas do ano, nomeadamente Outono e Inverno. No total foram utilizados 5 carneiros, sendo possível congelar sémen em 4 animais nas duas épocas de congelação, havendo 1 carneiro (811) em que só foi possível congelar sémen na época de Inverno (ano de 2002). As IA decorreram no mês de Abril, sendo inseminadas nos anos de 2002, 2003 e 2004 respectivamente 98, 175 e 167 ovelhas. Nos carneiros utilizados foram comparadas as épocas de congelação de sémen (Outono vs Inverno) relativamente à fertilidade, fecundidade e prolificidade nas ovelhas inseminadas.
4.1. Sincronização do estro e IA com sémen congelado
A sincronização do estro nas ovelhas inseminadas foi efectuada com esponjas vaginais
impregnadas com 40 mg de FGA (acetato de fluorogestona) durante 12 dias. No momento da retirada das esponjas vaginais (Rev) foram administradas 500 UI (i.m) de eCG (equine corionic gonadotrophin). As ovelhas apresentaram um bom estado sanitário e condição corporal tendo um comportamento reprodutivo normal na época de reprodução anterior.
Foram utilizados para IA ejaculados contendo pelo menos 40 % de spz móveis cinco minutos após a descongelação. O sémen destinado à IA foi transportado no respectivo contentor com azoto líquido. Após a descongelação realizada num banho-maria a 37 ºC durante 50 segundos, as palhinhas foram cuidadosamente secas, recortada a extremidade selada com álcool polivinílico e introduzidas no respectivo “pistolet”, procedendo-se de imediato à IA das ovelhas previamente sincronizadas.
A época de IA decorreu na época reprodutiva tradicional (Primavera) entre 30 de Março e 30 de Abril. Esta foi realizada em 440 ovelhas por via cervical entre as 54-56 horas após a remoção das esponjas vaginais (Rev) sem prévia detecção do estro utilizando duas doses de sémen por ovelha (600 milhões de spz totais). Aos 14 dias após a IA foram introduzidos no efectivo carneiros do PEP durante 45 dias, permitindo a cobrição de animais que não tivessem ficado gestantes após a IA. Foram avaliados os parâmetros reprodutivos da fertilidade, fecundidade e prolificidade, por reprodutor e por época de congelação de sémen (Quadro 15).
No Outono não foram observadas variações significativas entre carneiros relativamente à fertilidade e fecundidade. As ovelhas inseminadas com os carneiros 902 e 907 tiveram prolificidades mais elevadas.
No Inverno foram observadas variações individuais relativamente à fertilidade e fecundidade, tendo os carneiros 907 e 906 melhores resultados. Não foram observadas variações estacionais entre carneiros homólogos relativamente aos parâmetros reprodutivos avaliados (Quadro 15).

- 18 -
Quadro 15. Interacções significativas entre a época e o carneiro no sémen congelado
Época Carneiros IA Partos Crias Fertil Fecund Prolif
OUT
902 903 906 907
57 43 20 44
10 6 1 8
14 6 1 11
17,54 1* 13,95 1 5,0 1** 18,18 1
24,56 1
13,95 1 5,00 1** 25,00 1
140 b1 100 a1* 100 ab1
137,5 ab1
INV
811 902 903 906 907
30 64 51 26 105
2 5 5 4 25
3 8 7 4 35
6,70 a
7,81 a1* 9,81 a1
15,38 ab1** 23,81 b1
10,00 a 12,50 a1 13,73 a1
15,39 ab1** 33,33 b1
150 ab1 160 b
140 ab1* 100 a
140 ab1 Letras diferentes = diferenças entre carneiros dentro de cada época Números diferentes = diferenças entre épocas para o mesmo carneiro * P=0,10 (dentro da mesma coluna); ** P=0,26 (dentro da mesma coluna)
Num total de 440 ovelhas inseminadas com 5 carneiros, não foram observadas diferenças significativas entre épocas (Outono e Inverno) relativamente à fertilidade (15,24 % vs 14,85 %), fecundidade (19,61 % vs 20,65 %) e prolificidade (128 % vs 139 %).
Nos carneiros utilizados na IA com sémen congelado foram observadas correlações positivas nos parâmetros seminais da MI, % de spz vivos e normais existindo uma correlação positiva altamente significativa entre a MI e a % de spz vivos. Não foram encontradas correlações significativas entre estas características dos ejaculados com a fertilidade, fecundidade e a prolificidade das ovelhas inseminadas. 5. Estudos sobre a fertilidade in vitro com sémen descongelado
Na espécie ovina, poucos estudos estão descritos sobre a correlação entre os testes de fertilização in vitro e a fertilidade in vivo após a inseminação artificial. Numa primeira fase deste projecto propôs-se determinar a capacidade fertilizante do sémen de carneiros congelado em duas épocas diferentes, Outono e Inverno, através da tecnologia de fertilização (IVF) e produção de embriões in vitro (IVP). O sémen, após a descongelação, foi utilizado na fertilização de oócitos maturados in vitro recolhidos dos ovários de fêmeas abatidas ao longo do ano. O objectivo último seria, após os testes de fertilidade in vitro, prever a capacidade fecundante do sémen anteriormente à sua aplicação na inseminação artificial na ovelha.
As baixas taxas de desenvolvimento embrionário in vitro obtidos nesta espécie com o nosso sistema de IVP, levaram-nos a contactar, em Janeiro de 2004, a equipa francesa de produção in vitro de embriões em pequenos ruminantes, do Departamento de Fisiologia da Reprodução e dos Comportamentos, INRA, Nouzilly, chefiada pelo Professor Yves Cognié. A sua colaboradora, Dra. Nati Poulin, veio a Portugal demonstrar a sua técnologia de produção in vitro de embriões ovinos e caprinos. A implantação desta nova técnica, aliada à morosidade na recepção dos financiamentos dos projectos PIDDAC no início do ano de 2004, com consequente atraso na aquisição de novos reagentes e na resolução de problemas técnicos relacionados com a incubadora de 5% de CO2 e 5% de O2, não nos permitiram congelar um número suficiente de embriões para prosseguir no último ano com a respectiva transferência para fêmeas receptoras sincronizadas.

- 19 -
5.1. Metodologia a) Colheita de ovários e aspiração de oócitos
Os ovários foram transportados até ao laboratório numa solução fosfato-salina tamponizada de Dulbecco (PBS) a 37ºC a que se adicionaram 0,05 mg mL-1 de sulfato de kanamicina e 0,15 % (p/v) de albumina sérica bovina (BSA; Fracção V). Aspiraram-se os oócitos com agulha 18G e seringa de 5 ml a partir de folículos entre 2 a 5 mm de diâmetro, utilizando-se meio de aspiração adaptado para ovinos, constituído por TCM 199 (Tissue Culture Medium 199, com sais de Earle, L-glutamina e 25 mM mL-1 de Hepes), suplementado com 5% de soro de ovelha em cio superovulada (SOCS, v/v, confeccionado no próprio laboratório), 2% de uma solução antibiótica ml-1 (v/v), correspondente a 100 UI mL-1 de penicilina e 100 µg ml-1 de estreptomicina, 50 µg mL-1 de heparina e 0,4 % de albumina sérica bovina (BSA). Estas duas últimas substâncias foram adicionadas ao meio inicialmente utilizado para bovinos, para vencer a alta adesividade encontrada entre as células do cumulus dos vários complexos cumulus-oócitos (COC) ovinos, obtidos após a aspiração. Os oócitos foram depois observados à lupa e seleccionados para outro meio de selecção, semelhante ao meio de recolha mas sem heparina e BSA.
b) Maturação in vitro de oócitos ovinos
Apenas se seleccionaram para a fase de maturação os oócitos que apresentaram citoplasma uniformemente granuloso e sem sinais de atrésia nas células do cumulus e da granulosa que os envolvem. A maturação foi realizada em placas de Petri contendo meio TCM-199, a que se adicionaram 10% de soro de ovelha em cio superovulada (SOCS) e 2% da solução antibiótica já referida anteriormente, em estufa com 5% de CO2 e ar saturado de humidade a 39º C, durante 22 horas. Adicionaram-se a este meio células da granulosa em suspensão, retiradas de uma diluição final obtida para a formação de monocamadas para suporte do desenvolvimento de embriões, na concentração de 3 a 5 x 106 células mL-1 de meio de cultura (Lu et al., 1987). No nosso sistema de produção in vitro e durante o primeiro ano do projecto, ao meio de maturação adicionou-se 10 µg mL-1 de FSH (hormona foliculo-estimulante), permitindo uma maior expansão das células do cumulus durante este processo. Durante o ano de 2003, incluímos também no referido meio 5µg ml-1 de LH (hormona luteinizante), uma vez que a utilização simultânea das duas hormonas, FSH e LH, é apontada por alguns autores como aumentando significativamente a taxa de maturação dos oócitos e a produção posterior de blastócitos, diminuindo simultaneamente a sua taxa de degenerescência (Wang et al., 1998).
No nosso sistema, é relevante o número de oócitos obtidos a partir de fêmeas não adultas. Em 2004 foi sugerido pela Dra. Nati Poulin a suplementação do meio definido TCM 199, utilizado na maturação, por estradiol (E2, 200 ng mL-1), cisteamina (100 µM, De Matos et al., 1999; Cognié et al., 2002), precursor da síntese do glutatião, e um factor de crescimento, EGF (Epidermal Growing Factor, 10ng mL-1), estes últimos responsáveis pelo aumento da competência nuclear e citoplasmática de oócitos pré-púberes em ovinos e caprinos (Cognié et al., 2003). O glutatião é também responsável, depois da fecundação, pela descondensação da cabeça do espermatozóide e pela sua transformação em pronúcleo masculino. c) Descongelação e capacitação dos espermatozóides
Previamente à fase de fertilização dos oócitos maturados in vitro, o sémen congelado foi sujeito a descongelação e capacitação in vitro pela técnica de swim up (Crozet et al., 1987).

- 20 -
Imediatamente após a descongelação procedeu-se à avaliação microscópica da sua mobilidade individual (%MI pós-descongelação ou %MIPD). Segundo a metodologia do swim up, utilizada entre 2002 e 2003 no nosso laboratório, o sémen dos carneiros descongelado foi depositado no fundo de tubos de vidro que continham meio de capacitação próprio para ovinos (meio básico definido de Brackett modificado, tamponizado com 10 mM de HEPES, suplementado com 4,16 mM de NaHCO3, 122,5 mM de NaCl e 2,25 mM de CaCl2, adicionado de 20% de soro de ovelha em cio superovulada). Colocou-se o sémen assim preparado em estufa incubadora a 39ºC, com 5% de CO2 e máxima humidade, durante 1 hora. Durante este período, os espermatozóides (spz) mais activos progridem até à superfície do líquido de capacitação, sendo o sobrenadante retirado, depositado em tubos de vidro de fundo cónico, graduados e esterilizados, procedendo-se então à sua centrifugação a 500 g durante 5 minutos, para lavagem e concentração dos espermatozóides, antes da contagem em câmara de Neubauer e posterior fertilização. Após a capacitação in vitro, classificou-se de novo o sémen quanto à sua mobilidade individual (%MISU), à sua concentração por mililitro (CONC ou 1x106spz mL-1) e à sua capacidade de aglutinação (% CAP), correspondente ao número de espermatozóides aglutinados numa contagem total de 100.
De acordo com Cognié et al. (2004), foi aconselhada a alteração da estratégia de capacitação do sémen de carneiro. A centrifugação do sémen com gradientes de densidade (Percoll, 45%/90%) parece superior a outros procedimentos quando se utiliza sémen descongelado para fertilização in vitro. Iremos de futuro adoptar este método, segundo o qual, 1 ml de Percoll a 90% (19 ml de Percoll a 100% com 1 ml de cloreto de sódio a 9%) será colocado suavemente, sem misturar, no fundo de um tubo cónico de 15 ml contendo já 1 ml de Percoll a 45% Este último gradiente prepara-se adicionando 500µl de Percoll a 90% com 500 µl de meio SOF (Synthetic Oviductal Fluid ou fluido tubárico sintético, Takahashi e First, 1992) sem BSA, suplementado com 2,35 mg HEPES mL-1. O conteúdo de duas palhinhas de sémen descongelado é então depositado directamente na superfície deste gradiente, previamente equilibrado durante 1 hora em estufa com 5% de CO2 a 39ºC. Após a centrifugação durante 10 minutos a 900 g, rejeita-se o sobrenadante, determina-se a concentração do eluato na câmara de Neubauer e calcula-se a diluição de modo a obter uma concentração de 1x107 spz mL-1 no meio de capacitação. O meio de capacitação é constituído por SOF sem BSA suplementado com 10% de soro de ovelha em cio, 10 µg mL-1 de heparina e 4 µL mL-1 de gentamicina. Os espermatozóides incubam durante 15 a 30 minutos neste meio em estufa a 39ºC com 5% de CO2 e máxima humidade.
d) Fertilização in vitro
A fertilização realizou-se, nos anos de 2002 e 2003, em microgotas, imersas em óleo
mineral, de 40 µl de meio de fertilização (meio definido de Brackett modificado, de constituição idêntica à do meio para capacitação já descrito anteriormente, apenas suplementado com 7,75 mM de lactato de cálcio), em estufa incubadora a 39ºC, com 5% de CO2 e máxima humidade. Este processo realizou-se durante 22 horas, contendo cada microgota 10 oócitos e espermatozóides na proporção de 1 x 106 spz mL-1. Após este período, transferiram-se os presumíveis zigotos para placas de cultura, contendo monocamadas das células da granulosa ovinas em gotas de 100 µL de meio de desenvolvimento embrionário (meio TCM-199, suplementado com 10% de soro de ovelha em cio superovulada e 2% de uma solução antibiótica correspondente a 100 UI mL-1 de penicilina e 100 µg mL-1 de estreptomicina), submersas em óleo mineral. As monocamadas de células da granulosa ovina possuem já um período de cultura de 48 horas. A taxa de clivagem (% clivagem)foi determinada às 48 horas após a inseminação correspondendo à razão entre o número de zigotos que se encontram na fase de duas ou mais células e o total de oócitos inseminados.

- 21 -
Os resultados levaram-nos a adoptar, a partir de 2004, outra estratégia para a fase de fertilização do processo de IVP. Assim, futuramente, os oócitos maturados deverão ser desnudados das células do cumulus por pipetagens sucessivas em meio SOF com BSA, adicionado de 10% de soro de ovelha em cio e 4 µl mL-1 de gentamicina. Após a passagem por quatro banhos sucessivos neste mesmo meio, os oócitos lavados serão colocados em grupos de 40 a 50 dentro de poços com 450 µl de meio de fertilização, constituído por meio SOF sem BSA com 10% soro de ovelha em cio e gentamicina. A este meio adicionam-se 50 µl da diluição de espermatozóides preparada como é referenciado na alínea anterior, de modo a obter uma concentração final de 1x106 spz mL-1. As gotas para a fertilização in vitro, realizada em estufa a 39ºC com 5% CO2 e máxima humidade, estão cobertas com 350 µl de óleo mineral. Para Cognié et al. (2003), após 18 horas de contacto com os espermatozóides, os possíveis zigotos serão de novo lavados por pipetagens sucessivas com meio SOF e BSA sem gentamicina, sendo então colocados em grupos de 25 embriões no interior de gotas de 25 µl do mesmo meio, cobertas por 700 µl de óleo mineral, onde continuarão o seu desenvolvimento em estufa a 39ºC com 5% CO2, 5% de O2 e 90% de N2. e) Desenvolvimento embrionário in vitro
Durante o ano de 2002, utilizaram-se monocamadas de células da granulosa ovinas como
suporte ao desenvolvimento embrionário até ao 7º dia após a fertilização, dia programado para a possível congelação de embriões. A taxa de embriões (% Emb D7) foi determinada pela razão entre o número de embriões nos estádios de mórula, jovem blastócito, blastócito e blastócito expandido observados ao dia 7 de desenvolvimento e o número de embriões clivados às 48 horas após a fertilização in vitro.
Para a preparação das monocamadas células da granulosa (CG) procedeu-se à aspiração dos folículos ováricos de 2 a 5 mm de diâmetro, para obtenção dos complexos cumulus-oócitos e das células da granulosa. Diluiu-se o líquido folicular recolhido em meio de lavagem constituído por TCM 199, adicionado de 5% de soro de ovelha em cio superovulada (SOCS) e antibióticos. Após a rejeição dos oócitos por estereomicroscopia, as células da granulosa remanescentes da diluição anterior foram concentradas por centrifugação a 100g durante 5minutos. Rejeitou-se o sobrenadante e refez-se a suspensão celular com meio de cultura de oócitos, semelhante ao meio de lavagem das células referido anteriormente. Foi determinada a concentração e viabilidade celulares desta suspensão na câmara de Neubauer, retirando-se uma alíquota de 20 µl do sobrenadante e juntando-a a 20 µl de corante Azul de Tripan a 0,4% (concentração de 1:1). Diluiu-se de novo a suspensão para se obter a concentração de 1x106 CG mL-1 em gotas de 100 µl submersas em óleo mineral. As placas assim preparadas colocaram-se em estufa incubadora a 39ºC, com 5% de CO2 e saturada de humidade, recebendo os presumíveis zigotos às 48 horas de cultura. Estes continuaram aqui o seu desenvolvimento até ao 7/8 dia após a fertilização.
O sistema de desenvolvimento de embriões ovinos em SOF com aminoácidos e BSA na ausência de soro e células somáticas, em incubadora a 39ºC com 5% de CO2, 5% de O2, 90% de N2 e máxima humidade, está de um modo geral adoptado em todos os laboratórios de produção de embriões ovinos in vitro. No ano de 2003, os zigotos foram cultivados num sistema semi-definido utilizando o meio SOF modificado com citrato de sódio, suplementado com 10% de SOCS. No entanto, progressos recentes defendem a utilização de meios sequenciais onde os seus constituintes mudam de acordo com as exigências do embrião. Assim, alguns laboratórios adicionam 10% de soro de ovelha em cio ao meio SOF apenas entre os dias 2 e 3 dias após a fertilização (Cognié et al,, 2003), para promover uma maior viabilidade após a transferência dos embriões IVP ovinos. Em projectos futuros pretendemos seguir esta metodologia.
- 22 -
5.2. Resultados
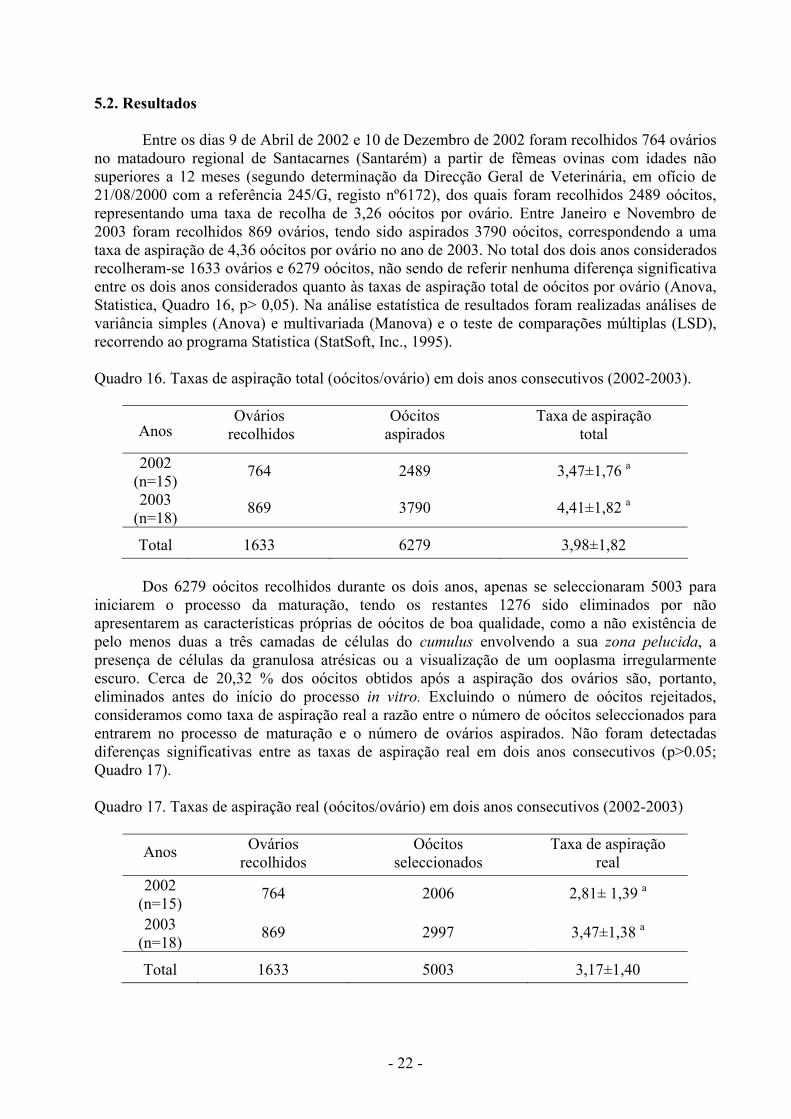
Entre os dias 9 de Abril de 2002 e 10 de Dezembro de 2002 foram recolhidos 764 ovários no matadouro regional de Santacarnes (Santarém) a partir de fêmeas ovinas com idades não superiores a 12 meses (segundo determinação da Direcção Geral de Veterinária, em ofício de 21/08/2000 com a referência 245/G, registo nº6172), dos quais foram recolhidos 2489 oócitos, representando uma taxa de recolha de 3,26 oócitos por ovário. Entre Janeiro e Novembro de 2003 foram recolhidos 869 ovários, tendo sido aspirados 3790 oócitos, correspondendo a uma taxa de aspiração de 4,36 oócitos por ovário no ano de 2003. No total dos dois anos considerados recolheram-se 1633 ovários e 6279 oócitos, não sendo de referir nenhuma diferença significativa entre os dois anos considerados quanto às taxas de aspiração total de oócitos por ovário (Anova, Statistica, Quadro 16, p> 0,05). Na análise estatística de resultados foram realizadas análises de variância simples (Anova) e multivariada (Manova) e o teste de comparações múltiplas (LSD), recorrendo ao programa Statistica (StatSoft, Inc., 1995). Quadro 16. Taxas de aspiração total (oócitos/ovário) em dois anos consecutivos (2002-2003).
Anos
Ovários recolhidos
Oócitos aspirados
Taxa de aspiração total
2002 (n=15) 764 2489 3,47±1,76 a
2003 (n=18) 869 3790 4,41±1,82 a
Total 1633 6279 3,98±1,82
Dos 6279 oócitos recolhidos durante os dois anos, apenas se seleccionaram 5003 para iniciarem o processo da maturação, tendo os restantes 1276 sido eliminados por não apresentarem as características próprias de oócitos de boa qualidade, como a não existência de pelo menos duas a três camadas de células do cumulus envolvendo a sua zona pelucida, a presença de células da granulosa atrésicas ou a visualização de um ooplasma irregularmente escuro. Cerca de 20,32 % dos oócitos obtidos após a aspiração dos ovários são, portanto, eliminados antes do início do processo in vitro. Excluindo o número de oócitos rejeitados, consideramos como taxa de aspiração real a razão entre o número de oócitos seleccionados para entrarem no processo de maturação e o número de ovários aspirados. Não foram detectadas diferenças significativas entre as taxas de aspiração real em dois anos consecutivos (p>0.05; Quadro 17).
Quadro 17. Taxas de aspiração real (oócitos/ovário) em dois anos consecutivos (2002-2003)
Anos Ovários recolhidos
Oócitos seleccionados
Taxa de aspiração real
2002 (n=15) 764 2006 2,81± 1,39 a
2003 (n=18) 869 2997 3,47±1,38 a
Total 1633 5003 3,17±1,40

- 23 -
Um dos factores responsáveis pela sazonalidade reprodutiva em ovelhas é a luminosidade. Os ovinos de climas frios e temperados utilizam o ciclo anual do fotoperíodo (horas luz/dia) para determinar mudanças sazonais na sua actividade sexual. Estes animais apresentam uma estação reprodutiva bem definida durante o ano, o qual se estende desde o final do Verão até ao início do Inverno, iniciando a sua ciclicidade à medida que a luminosidade diária diminui. Obedecem portanto a um fotoperíodo decrescente, sendo classificados como animais de dias curtos.
Assim, agrupámos as sessões de recolha dos ovários em duas épocas, a primeira abrangendo o período do solstício de Dezembro ao solstício de Junho, fase de fotoperíodo crescente, e a segunda de Junho a Dezembro, fase de fotoperíodo decrescente, verificámos no entanto não haver diferenças significativas entre as taxas de aspiração total e real de oócitos por ovário entre as duas épocas referidas (p> 0,05, Quadro 18). Quadro 18. Efeito do fotoperíodo sobre as taxas de aspiração total e real (oócitos/ovário) no conjunto dos dois anos (2002 e 2003).
Semestre Ovários recolhidos
Oócitos aspirados
T. de aspiração total
Oócitos seleccionados
T. de aspiração real
Dez-Jun (n=15) 705 2961 4,14± 1,74 a 2397 3,39±1,38 a
Jun-Dez (n=10) 928 3318 3,85±1,94 a 2606 2,99±1,43 a
No quadro 19 observámos que não se verificaram, de um ano para o seguinte, diferenças
significativas em relação à taxa de maturação dos oócitos ovinos (p>0,05). Quadro 19. Taxa de maturação (%) de oócitos ovinos em dois anos consecutivos (2002-2003)
Anos Oócitos a maturar
Oócitos maturados Maturação (%)
2002 (n=15) 2006 1614 80,38±8,2 a
2003 (n=18) 2416 a) 1859 78,62±11,1 a
Total 4422 3474 79,47±9,6 a) No ano de 2003, duas sessões no total de 581 oócitos seleccionados não completaram a maturação por contaminação por fungos (Aspergillus fumigatus), pelo que foram afastados daquele grupo.
Agrupando as sessões de maturação de oócitos em duas épocas, a primeira abrangendo o período do solstício de Dezembro ao solstício de Junho, fase de fotoperíodo crescente, e a segunda de Junho a Dezembro, fase de fotoperíodo decrescente, verificámos que a taxa de maturação dos oócitos no período de luminosidade decrescente é significativamente superior à taxa de maturação no período de luminosidade crescente, entre Dezembro e Junho (83,48% ± 5,67% vs. 74,61% ± 11,43%, respectivamente, p<0,05, Quadro 20).

- 24 -
Quadro 20. Influência da época do ano sobre as taxas de maturação de oócitos ovinos recolhidos nos dois anos (2002-2003).
Semestre Oócitos a maturar
Oócitos maturados
Maturação (%)
Dez-Jun (n=15) 2086 1526 74,61 ± 11,43 ª
Jun-Dez (n=18) 2336 1947 83,48 ± 5,67 b
Sobrescritos diferentes na mesma coluna indicam diferenças significativas (p<0,05)
Antes da fertilização in vitro, foi examinado à descongelação o sémen de dois carneiros de raça Serra da Estrela (nº 501 e 802), congelado em duas épocas diferentes, no Outono e no Inverno (n=18). Foram determinados os parâmetros da mobilidade imediatamente após a descongelação (%MIPD) e os parâmetros após swim up, como a mobilidade (%MISU), taxa de capacitação (%CAP ou número de spz aglutinados num total de 100) e a concentração (CONC, 1x106spz mL-1). Analisaram-se os efeitos da época, do carneiro e da interacção entre estes dois factores quanto aos parâmetros descritos (Quadro 21). A interacção entre os dois factores apenas foi significativa para a MIPD, (p=0,02, gráfico 9). Entre épocas, o sémen do carneiro 501 congelado no Outono apresenta taxas de mobilidade após descongelação significativamente superiores às obtidas quando o sémen do mesmo animal é congelado no Inverno, o que não acontece com o carneiro 802. Também na época de Outono e em relação ao mesmo parâmetro, o carneiro 501 é superior ao carneiro 802 (p <0,05, gráfico 10) enquanto na mesma época de Inverno os dois carneiros não são diferentes. Para a capacitação e na mesma época de congelação, quer de Outono quer de Inverno, verificámos que o carneiro 802 é superior ao carneiro 501 (p <0,05). Para a capacitação, não existem diferenças entre épocas para qualquer dos carneiros considerados, pelo que a interacção entre carneiro e época para este último parâmetro não é significativa. Quadro 21. Interacção época x carneiro para as taxas de motilidade após a descongelação (% MIPD), de motilidade após o swim up (% MISU), de capacitação (% CAP) e concentração do sémen (CONC x 106 spz mL-1) de dois carneiros Serra da Estrela em duas épocas diferentes (Outono e Inverno)
Efeito Carn N MIPD MISU CAP CONC 501 5 56,0 ± 5,5 b2 42,0 ± 4,5 a1 26,4 ± 4,1 b1 74,8 ± 36,31 a1 Out 802 13 36,9 ± 10,1 a1 35,8 ± 10,8 a1 37,5 ± 13,5 a1 74,13 ± 42,2 a1 501 5 42,0 ± 4,5 a1 34,0 ± 8,9 a1 24,8 ± 4,2 b1 69,8 ± 26,7 a1 Inv 802 13 40,8 ± 11,2 a1 41,2 ± 18,0 a1 39,2 ± 12,9 a1 75,9 ± 49,0 a1
Interacção 36 F[1,32]=6,3 p=0,02 NS NS NS
501 10 49,0 ± 8,8 a 38,0 ± 7,9 a 25,6 ± 4,0 a 72,3 ± 30,2 a Efeito Carneiro 802 26 38,9 ± 10,6 b 38,5 ± 14,8 a 38,4 ± 13,0 b 75,0 ± 45,1 a
Out 18 42,2 ± 12,5 a 37,5 ± 9,7 a 34,4 ± 12,6 a 74,3 ± 39,6 a Efeito Época Inv 18 41,1 ± 9,6 a 39,2 ± 16,1 a 35,2 ± 12,9 a 74,2 ± 43,7 a Letras diferentes na mesma coluna para cada efeito, indicam diferenças significativas (p < 0,05) Números diferentes na mesma coluna indicam diferenças significativas entre efeitos para animais homólogos (p < 0,05)

- 25 -
Gráfico 9. Efeito específico do carneiro sobre a percentagem de mobilidade pós-descongelação (% MIPD) e capacitação (%CAP) após o swim-up em sémen de carneiros Serra da Estrela
Efeito específico do Carneiro
(MIPD: F(1, 32)=8,16; p=0,007) (CAP:F(1, 32)=8,69; p=0,006)Barras Verticais 0,95 int conf
501 802
Carneiro
10
15
20
25
30
35
40
45
50
55
60P
erce
ntag
em MIPD CAP
Gráfico 10. Interacção época x carneiro na mobilidade pós-descongelação (%MIPD) em sémen de carneiros Serra da Estrela.
Época x Carneiro (MPD): F(1, 32)=6,3035, p=,01730Barras Verticais 0,95 int conf
Outono Inverno25
30
35
40
45
50
55
60
65
70
Mob
ilida
de p
ós-d
esco
ngel
ação
(%)
Carneiro 501 Carneiro 802
Descongelaram-se palhinhas de sémen de três carneiros da raça Saloia, congelado no Outono (carneiros 903 e 907, n=4) e Inverno (carneiros 811 e 907, n=15). Imediatamente após a descongelação avaliaram-se a percentagem de MIPD e após o swim-up as percentagens de MISU e de CAP e também a CONC (1x106 spz mL-1). No Outono, o carneiro 907 apresentou uma taxa de MIPD significativamente superior à respectiva taxa quando o seu sémen é congelado no Inverno (p< 0,01). Também no Outono, a percentagem de MISU no carneiro 903 foi superior à do carneiro 907 (p< 0,02, Quadro 22).
- 26 -
Quadro 22. Taxas de mobilidade após a descongelação (% MIPD), taxa de mobilidade após o swim up (% MISU), taxa de capacitação (% CAP) e concentração do sémen (CONC x 106 spz mL-1) de dois carneiros de raça Saloia em duas épocas diferentes (Outono e Inverno, 2003)
Efeito Carneiro N MIPD MISU CAP CONC 903 4 52,5 ± 15,0 56,3 ± 7,5 a 47,6 ± 16,2 208,0 ± 175,4 Out 907 4 56,3 ± 7,5 1 36,3 ± 11,1 b 47,6 ± 19,4 147,5 ± 158,7 811 15 40,5 ± 9,5 36,0 ± 10,4 48,2 ± 15,4 68,3 ± 22,0 Inv 907 15 41,3 ± 9,7 2 37,7 ± 10,8 46,6 ± 18,1 75,1 ± 30,3
Letras diferentes entre carneiros da mesma época representam diferenças significativas (p<0,02) Carneiro 907 entre épocas para a MIPD: p<0,01
O Quadro 23 apresenta as interacções entre as épocas (Outono e Inverno) e raças (Serra da Estrela e Saloio) para as diferentes taxas de avaliação do sémen antes da fertilização in vitro. As interacções são significativas para os parâmetros da mobilidade após a descongelação (%MIPD, p=0,03) e da concentração após o swim up (%CONC, p=0,001, Gráfico 11). A raça Saloia apresentou sémen com taxas de MIPD e de CONC superiores quando congelado no Outono (p<0,05), relativamente ao sémen da mesma raça congelado no Inverno. Dentro da mesma época, a raça Saloia foi, no Outono, significativamente superior à raça Serra da Estrela para a MIPD e a CONC e, no Inverno, significativamente superior quanto à capacitação.
Quadro 23. Interacção época (Outono e Inverno) x raça (Serra da Estrela e Saloio) para as taxas de mobilidade após a descongelação (% MIPD), de mobilidade após o swim up (% MISU), de capacitação (% CAP) e concentração (CONC x 106 spz mL-1) do sémen. Efeito Raça N MIPD MISU CAP CONC
Saloio 8 54,4 ± 11,2 a1 46,2 ± 13,8 a1 47,6 ± 16,5 a1 177,8 ± 158,2 a1 Out Serra 18 42,2 ± 12,5 b1 37,5 ± 9,7 a1 34,4 ± 12,6 a1 74,3 ± 39,6 b1 Saloio 30 40,9 ± 9,4 a2 36,8 ± 10,5 a1 47,4 ± 16,5 a1 71,7 ± 26,3 a2 Inv Serra 18 41,1 ± 9,6 a1 39,2 ± 16,1 a1 35,2 ± 12,9 b1 74,2 ± 43,6 a1
Interacção 74 F[1,70]=5,1 p=0,03 p>0,08 p>0,90 F[1,70]=11,5 p=0,001
Saloio 38 43,8 ± 11,1 a 38,8 ± 11,7 a 47,5 ± 16,3 a 94,0 ± 84,8 a Efeito Raça Serra 36 41,7 ± 11,0 b 38,3 ± 13,2 a 34,8 ± 12,6 b 74,3 ± 41,1 b
Out 26 46,0 ± 13,2 a 40,2 ± 11,6 a 38,5 ± 15,0 a 106,1 ± 102,2 a Efeito Época Inv 48 41,0 ± 9,4 b 37,7 ± 12,8 a 42,8 ± 16,3 a 72,6 ± 33,4 b Letras diferentes na mesma coluna para cada efeito, indicam diferenças significativas (p< 0,05) Números diferentes na mesma coluna indicam diferenças significativas entre efeitos para a mesma raça (p< 0,05)
- 27 -
Gráfico 11. Interacção entre raças (Serra da Estrela e Saloia) e épocas (Outono e Inverno) para a concentração (CONC, 1x106 spz mL-1) do sémen de carneiros após swim up.
Época x Raça: F(1, 70)=11,477, p=,00116Barras Verticais 0,95 int conf
Outono Inverno
Época
20
40
60
80
100
120
140
160
180
200
220
240C
once
ntra
ção
pós
swim
up
(spz
x 1
06 m
L-1)
Saloia Serra
Apresentamos em seguida os dados referentes à fertilização in vitro de oócitos ovinos com o sémen descongelado de carneiros Serra da Estrela (501: n=5, 802: n=10 sessões), recolhido em duas épocas diferentes, Outono e Inverno. Um total de 817 oócitos (carneiro 501=361 oócitos e carneiro 802=456 oócitos) foi inseminado com sémen congelado na época de Inverno, sujeito a descongelação e capacitação in vitro, de que resultaram 191 oócitos clivados, 46 pelo carneiro 501 e 145 pelo carneiro 802. Foram fertilizados 797 oócitos (carneiro 501=367; carneiro 802=430) com sémen congelado no Outono, de que resultaram 267 oócitos clivados, 73 pelo carneiro 501 e 194 pelo carneiro 802. O Quadro 24 revela os efeitos do carneiro e da época de congelação do sémen sobre as taxas de fertilização de oócitos ovinos (% de clivagens) e taxas de embriões ao dia 7 (% Emb D7) obtidas na raça Serra da Estrela. Não houve interacção significativa entre os dois factores quanto a estes parâmetros estudados. Para a mesma época de congelação de Outono, as taxas de clivagem do carneiro 802 foram superiores às do carneiro 501 (p=0,052), o mesmo não se verificando de Inverno. Para as taxas de produção de embriões e na mesma época de Inverno, o carneiro 802 mostrou-se superior ao 501 (p=0,036).
- 28 -
Quadro 24. Interacção entre carneiros e épocas de congelação do sémen para as taxas de clivagem (% clivagem) e de embriões (% Emb D7) na raça Serra da Estrela.
Efeito Carneiro N Clivagem Emb D7 501 5 21,9 ± 8,4 a1 2,5 ± 0,5 a1 Out 802 10 43,1 ± 22,8 b1 6,8 ± 3,0 a1 501 5 16,2 ± 11,3 a1 1,5 ± 0,8 a1 Inv 802 10 28,5 ± 21,0 a1 6,9 ± 4,8 b1
Interacção 30 NS NS
501 10 19,0 ± 9,8 a 2,0 ± 0,8 a Efeito Carneiro 802 20 35,8 ± 22,6 b 6,9 ± 3,9 b
Out 15 36,0 ± 21,5 a 5,7 ± 3,3 a Efeito Época Inv 15 24,4 ± 18,9 a 5,4 ± 4,8 a
Letras diferentes na mesma coluna para cada efeito, indicam diferenças significativas (p < 0,05) Números diferentes na mesma coluna indicam diferenças significativas entre efeitos para a mesma raça (p < 0,05)
No ano de 2003, foram inseminados 228 oócitos com sémen processado in vitro congelado no Outono (n=3), proveniente de carneiros de raça Saloia, cabendo um grupo de 118 oócitos ao carneiro 903 e um segundo grupo de 110 oócitos ao carneiro 907. Cerca de 48 horas após a fertilização, clivaram 52 e 31 oócitos, respectivamente para os grupos fertilizados pelo carneiro 903 e 907. Obtiveram-se 14 embriões ao dia 7 de cultura in vitro com o carneiro 903 e 9 embriões com o carneiro 907. No Inverno (n=12), foram inseminados 1518 oócitos (carneiro 907=770; carneiro 811=748 oócitos), tendo clivado 321 e 295, respectivamente para o carneiro 907 e 811. Este último produziu 51 embriões in vitro enquanto o carneiro 907 produziu 65, também no Inverno. Para as taxas de clivagem, não se manifestaram diferenças significativas no carneiro 907 entre épocas. Para as taxas de embriões, o carneiro 907 no Outono foi significativamente superior ao mesmo carneiro no Inverno (p=0,01, Quadro 25). Quadro 25. Efeitos do carneiro e da época de congelação do sémen sobre as taxas de clivagem (% Clivagem) e de embriões (% Emb D7) em carneiros de raça Saloia.
Efeito Carneiro N Clivagem Emb D7 903 3 44,3 ± 20,9 26,8 ± 13,7 Out 907 3 28,0 ± 7,8 38,1 ± 6,7 a 811 12 39,5 ± 17,4 21,6 ± 10,3 Inv 907 12 44,5 ± 16,2 20,3 ± 7,6 b
a vs b: p=0,01
No Quadro 26 estão referenciados os dados relativos à interacção entre as raças estudadas e a época de congelação do sémen para as taxas de clivagem e de embriões ao dia 7 após a fertilização in vitro. A interacção só foi significativa para a taxa de produção de embriões (p=0,04), a qual é afectada quer pela raça quer pela época de congelação do sémen. Para a raça Saloia, a taxa de embriões demonstrou ser significativamente superior quando se utilizou sémen de Outono relativamente ao sémen de Inverno (p=0,007). Para a raça Serra da Estrela não existem diferenças entre épocas. O sémen de raça Saloia congelado quer no Outono quer no Inverno produziu significativamente mais embriões que o sémen Serra da Estrela congelado nas

- 29 -
épocas correspondentes (p<0,001). Os efeitos específicos da época e da raça foram significativos relativamente à produção de embriões. Quadro 26. Interacção entre a raça (Saloia e Serra da Estrela) e a época de congelação (Outono e Inverno) do sémen para as taxas de clivagem (% Clivagem) e de embriões (% Emb D7)
Efeito Raça N Clivagem Emb D7 Saloio 6 36,1 ± 16,7 a1 31,3 ± 12,0 a1 Out Serra 24 36,0 ± 21,5 a1 5,7 ± 3,3 b1 Saloio 15 42,0 ± 16,6 a1 21,0 ± 8,9 a2 Inv Serra 15 24,4 ± 18,9 a1 5,4 ± 4,8 b1
Interacção 60 NS F[1,48]=4,3 p=0,04
Saloio 30 40,9 ± 16,5 a 22,7 ± 10,1 a Efeito Raça Serra 30 30,2 ± 20,7 a 5,6 ± 4,0 b
Out 21 36,1 ± 19,8 a 13,3 ± 13,7 a Efeito Época Inv 39 35,2 ± 19,3 a 16,1 ± 10,7 b
Letras diferentes na mesma coluna para cada efeito, indicam diferenças significativas (p < 0,05) Números diferentes na mesma coluna indicam diferenças significativas entre efeitos para a mesma raça (p < 0,05)
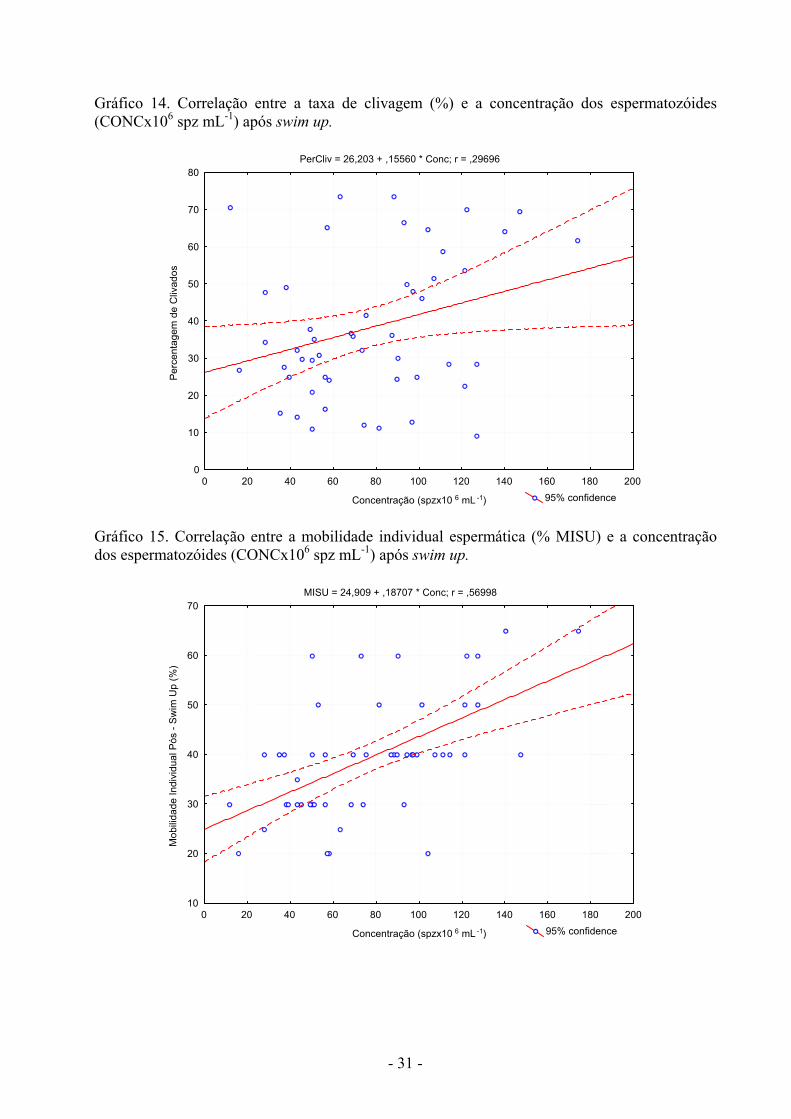
Estabeleceram-se as correlações entre os parâmetros de análise seminais após a descongelação (%MIPD) e após o swim up (%MISU, CONC, 1x106 spz mL-1, %CAP) e as taxas de clivagem (%) e de embriões em D7 (%) após a fertilização in vitro, para o total de raças e épocas (Quadro 27). As correlações positivas e significativas entre as taxas de clivagem e de embriões e a capacitação dos espermatozóides após swim up estão ilustradas nos gráficos 12, 13 e 14. No gráfico 15 apresenta-se a correlação entre a mobilidade após swim up e a concentração espermática. Quadro 27. Correlações entre os parâmetros de análise seminais pós-descongelação (%MIPD) e pós-swim up (%MISU, CONC, 1x106 spz mL-1, %CAP) e as taxas de clivagem (%) e de embriões em D7 (%) após a fertilização in vitro.
MIPD MISU Conc Capac PerCliv PerEmb MIPD 1,00 0,01 -0,08 -0,29 -0,41 0,13 MISU 1,00 0,57 0,25 -0,01 0,16 Conc 1,00 0,23 0,30 0,09 Capac 1,00 0,39 0,54 PerCliv 1,00 0,14 PerEmb 1,00
- 30 -
Gráfico 12. Correlação entre as taxas de clivagem (%) após a fertilização e a taxa de capacitação dos espermatozóides (%) após swim up.
PerCliv = 19,498 + ,43232 * Capac; r = ,39484
10 20 30 40 50 60 70 80
Capacitação
0
10
20
30
40
50
60
70
80
Perc
enta
gem
de
Cliv
agem
95% confidence
Gráfico 13. Correlação entre as taxas de embriões D7 (%) e a taxa de capacitação dos espermatozóides (%) após swim up.
PerEmb = -,4239 + ,36718 * Capac; r = ,54138
10 20 30 40 50 60 70 80
Capacitação
-10
0
10
20
30
40
50
60
Embr
iões
D7
(%)
95% confidence
- 31 -
Gráfico 14. Correlação entre a taxa de clivagem (%) e a concentração dos espermatozóides (CONCx106 spz mL-1) após swim up.
PerCliv = 26,203 + ,15560 * Conc; r = ,29696
0 20 40 60 80 100 120 140 160 180 200
Concentração (spzx10 6 mL -1)
0
10
20
30
40
50
60
70
80Pe
rcen
tage
m d
e C
livad
os
95% confidence
Gráfico 15. Correlação entre a mobilidade individual espermática (% MISU) e a concentração dos espermatozóides (CONCx106 spz mL-1) após swim up.
MISU = 24,909 + ,18707 * Conc; r = ,56998
0 20 40 60 80 100 120 140 160 180 200
Concentração (spzx10 6 mL -1)
10
20
30
40
50
60
70
Mob
ilida
de In
divi
dual
Pós
- Sw
im U
p (%
)
95% confidence

- 32 -
5.2.1. Fertilidade in vivo vs fertilidade in vitro
Não se observaram correlações significativas entre taxas de fertilidade in vivo com as taxas de clivagem e de embriões in vitro, quando se utilizaram dados de 3 carneiros da raça Saloia, a partir de sémen congelado nas mesmas épocas. 5.3. Conclusões
Pretendeu-se, para além de implementar em Portugal a técnica de produção in vitro de embriões ovinos (IVP), estabelecer uma possível correlação entre os parâmetros de fertilidade in vitro e os de fertilidade in vivo após a inseminação artificial em ovelhas, utilizando sémen congelado no Outono e no Inverno. O objectivo último seria, partindo dos testes de fertilização in vitro, prever a qualidade do sémen anteriormente à sua aplicação na inseminação artificial. Vários parâmetros foram estudados in vitro, como a taxa de aspiração total e real de oócitos por ovário, a taxa de maturação dos oócitos (% Maturação), a taxa de mobilidade (%MIPD) após a descongelação do sémen, as taxas de mobilidade (%MISU) e capacitação (%CAP) do sémen após o swim up assim como a sua concentração (CONC x 106 spz mL-1) e as taxas de fertilização in vitro, determinadas pelas taxas de clivagem (% Clivagem) dos oócitos e de produção de embriões (% Emb D7) ao dia 7 de desenvolvimento. Verificámos que a taxa de maturação dos oócitos quando obtidos no período de luminosidade decrescente é significativamente superior à mesma quando os oócitos são recolhidos no período de luminosidade crescente. As taxas de colecção de oócitos por ovário, quer total quer real, não são diferentes das apontadas por outros autores, que referem 1 a 2 complexos cumulus-oócitos de qualidade aceitável obtidos por ovelha adulta. A condição expressa pela Direcção Geral de Veterinária de apenas serem recolhidos órgãos genitais de ovelhas abatidas em matadouros com idade não superior a 12 meses, devido aos problemas das encefalopatias espongiformes transmissíveis que assolaram o nosso país, levou a que os oócitos obtidos proviessem muitas vezes de animais não adultos. Estes oócitos têm uma competência de desenvolvimento reduzido, facto que poderá ter sido responsável pelas fracas taxas de embriões ovinos produzidos. Em relação ao sémen, entre épocas diferentes, a raça Saloia apresentou uma taxa de mobilidade após a descongelação (%MIPD) e de concentração (CONC, 1x106 spz mL-1) após o swim up significativamente superiores no Outono (p <0,05), relativamente ao sémen na mesma raça congelado no Inverno. Dentro da mesma época, a raça Saloia foi, no Outono, significativamente superior quanto à MIPD e à CONC e, no Inverno, significativamente superior quanto à capacitação (%CAP) relativamente à raça Serra da Estrela. Embora as taxas de fertilização obtidas com o nosso primeiro sistema de produção de embriões ovinos in vitro sejam pouco elevadas nesta espécie, poderemos no entanto sugerir que a interacção entre a raça e a época de congelação do sémen foi significativa para a taxa de produção de embriões (p=0,04), o mesmo não acontecendo com a taxa de clivagem, em que não houve influência daqueles dois factores. Entre épocas diferentes, para a raça Saloia, a taxa de embriões mostrou ser significativamente superior quando se utiliza sémen de Outono relativamente ao sémen de Inverno (P<0,007). Para a raça Serra da Estrela não existem diferenças entre épocas. O sémen de raça Saloia congelado quer no Outono quer no Inverno produziu significativamente mais embriões que o sémen Serra da Estrela congelado nas épocas correspondentes. Foram verificadas correlações positivas e significativas entre as taxas de clivagem e a capacitação dos espermatozóides após swim up (p=0,39) e entre as taxas de clivagem e a concentração do sémen após swim up (p=0,30). Observou-se também também uma correlação positiva entre a taxa de embriões (% Emb) e a capacitação (p=0,54). No ano de 2003 o sistema de co-cultura de embriões com células somáticas da granulosa foi substituído por um sistema de cultura semi-definido, utilizando meio SOF (Synthetic oviductal fluid ou fluido sintético do oviduto) modificado, suplementado com 10 % de soro de ovelha em cio
- 33 -
superovulada, incubando numa atmosfera contendo 5% CO2, 5% O2, 90% N2, a 39ºC e com máxima humidade. Este factor poderá também ter contribuído para um ligeiro aumento da taxa de produção de embriões a partir deste ano. A qualidade dos embriões obtidos nos anos de 2003 e 2004 não foi, no entanto, suficiente para se proceder à respectiva congelação/descongelação e transferência para fêmeas receptoras sincronizadas, pelo que consideramos ser este um objectivo não atingido dentro do projecto. 6. Electromiografia e estudos de penetrabilidade cervical
Foi avaliada a actividade contráctil e mobilidade cervical (EMG) durante a fase folicular
do ciclo éstrico de 5 ovelhas. Em cada animal foi previamente implantado cirurgicamente, na camada de músculos lisos do cérvix uterino, um par de eléctrodos monopolares de Teflon, providos de agulha. Nos períodos de medição, os eléctrodos foram ligados a um sistema modular computorizado (Lablink V, Coulbourn Instruments, Allentown, USA).
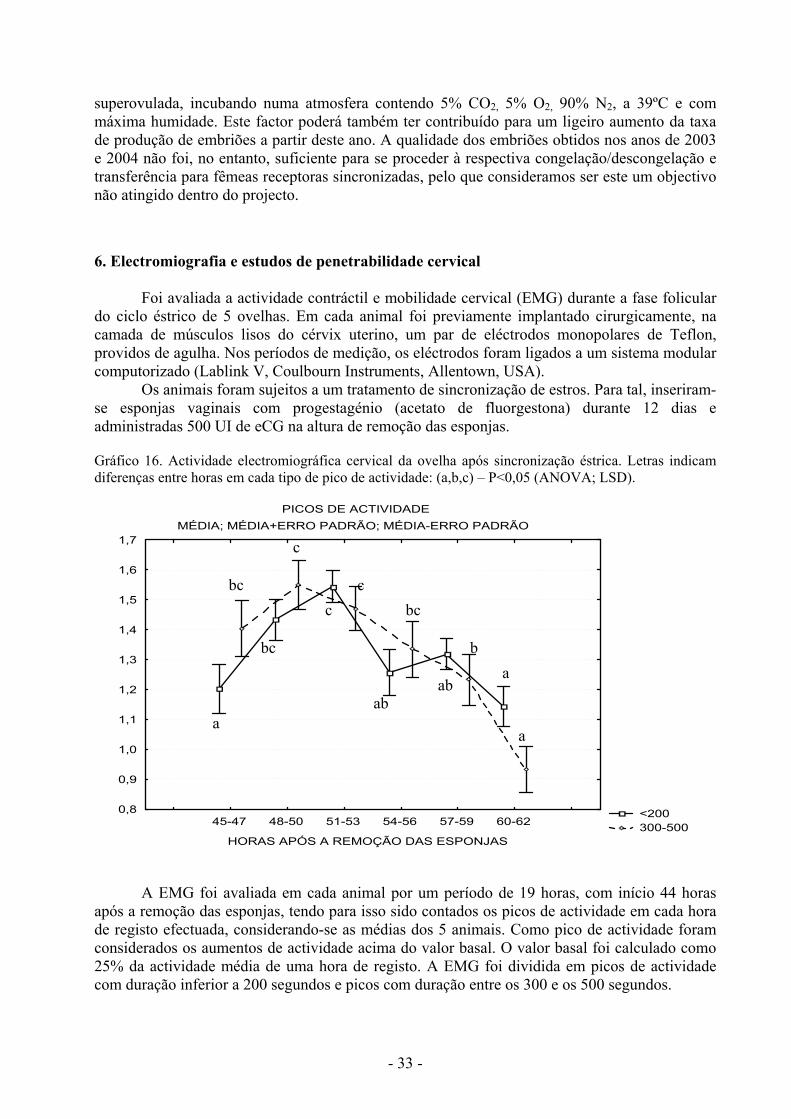
Os animais foram sujeitos a um tratamento de sincronização de estros. Para tal, inseriram-se esponjas vaginais com progestagénio (acetato de fluorgestona) durante 12 dias e administradas 500 UI de eCG na altura de remoção das esponjas. Gráfico 16. Actividade electromiográfica cervical da ovelha após sincronização éstrica. Letras indicam diferenças entre horas em cada tipo de pico de actividade: (a,b,c) – P<0,05 (ANOVA; LSD).
A EMG foi avaliada em cada animal por um período de 19 horas, com início 44 horas
após a remoção das esponjas, tendo para isso sido contados os picos de actividade em cada hora de registo efectuada, considerando-se as médias dos 5 animais. Como pico de actividade foram considerados os aumentos de actividade acima do valor basal. O valor basal foi calculado como 25% da actividade média de uma hora de registo. A EMG foi dividida em picos de actividade com duração inferior a 200 segundos e picos com duração entre os 300 e os 500 segundos.
PICOS DE ACTIVIDADEMÉDIA; MÉDIA+ERRO PADRÃO; MÉDIA-ERRO PADRÃO
<200 300-50045-47 48-50 51-53 54-56 57-59 60-62
HORAS APÓS A REMOÇÃO DAS ESPONJAS
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
a
bc
c
ab ab
a
bc
c
c bc
b
a

- 34 -
Os dois tipos de picos de actividade considerados apresentaram um padrão semelhante ao longo do período de observação, se bem que não coincidente no tempo. Nos picos de actividade menores de 200 segundos verificou-se um aumento do seu número entre as 48 e 50 horas após a remoção das esponjas, com o número máximo observado entre as 51 e as 53 horas. Quanto ao picos de actividade de duração entre os 300 e os 500 segundos, o seu número máximo foi observado entre as 48 e as 50 horas após a remoção das esponjas, mantendo-se até às 51-53 horas (Gráfico 16).
O período de actividade aumentada (48-53 horas após a remoção das esponjas) é coincidente com o momento de estro e cobrição /IA. O aumento de actividade da musculatura cervical no período coincidente com o estro e/ou IA parece sugerir o envolvimento do cérvix no mecanismo de transporte espermático. A sua modulação experimental poderá ser utilizada com o objectivo de facilitar o transporte espermático, aumentando a fertilidade após IA cervical. 7. Objectivos vs Realização A realização deste projecto não permitiu a realização total dos objectivos pretendidos. No entanto, um dos objectivos mais importantes, a implementação da técnica de congelação de sémen de carneiro, foi alcançado após várias afinações que esta técnica exigiu. Foi possível congelar sémen de carneiro nas duas épocas do ano, embora para alguns animais não fosse possível realizar colheitas de sémen em ambas as épocas por capricho dos animais, doenças, etc. A existência de maior número de carneiros para as colheitas de sémen e de maior número de ovelhas inseminadas permitiria uma melhor consolidação dos resultados. Ensaiou-se também a utilização de dilatadores cervicais durante o período do estro em estudos de contractilidade do canal cervical por electromiografia, embora não em condições de rebanho no momento da IA com sémen refrigerado e congelado. Prevê-se a sua administração em próximos ensaios de IA.
Relativamente à avaliação da capacidade do sémen descongelado para fertilização in vitro de oócitos ovinos, a recolha destes constituiu o problema nuclear devido à pouca disponibilidade de ovários nos matadouros regionais, agravado por determinações sanitárias que só autorizavam a recolha a partir de animais até ao ano de idade. Esta idade não permite a obtenção de oócitos com boa competência para o desenvolvimento posterior, o que se traduziu pelas baixas taxas de clivagem e produção obtidos in vitro. Foi apesar disso implementada a técnica de produção in vitro de embriões ovinos, sendo de realçar a excelente colaboração de uma investigadora francesa, a Dra. Nati Poulin, que nos os aconselhou uma nova metodologia para o desenvolvimento embrionário in vitro em ovinos. A implantação desta nova técnica, aliada à morosidade na recepção dos financiamentos dos projectos PIDDAC no início do ano de 2004, com consequente atraso na aquisição de novos reagentes e na resolução de problemas técnicos relacionados com a incubadora de 5% de CO2 e 5% de O2, não nos permitiram congelar um número suficiente de embriões para prosseguir no último ano com a respectiva transferência para fêmeas receptoras sincronizadas. 8. Cooperações
Com a realização deste projecto mantivemos colaborações com a Direcção Regional de Agricultura de Ribatejo e Oeste (DRARO), nomeadamente o PEP, que nos cedeu os carneiros destinados aos estudos da aptidão à congelação do sémen nas épocas de Outono e Inverno e à produção de doses seminais destinadas à IA com sémen congelado. As ovelhas inseminadas no decurso deste projecto são pertencentes ao PEP, tendo aí decorrido as inseminações. Foram realizadas cooperações com França, designadamente através da realização de um estágio no

- 35 -
Laboratório de Congelação de Sémen da Unidade Experimental de Inseminação Caprina e Ovina (INRA) sob a orientação do Professor Bernard Leboeuf. Foi igualmente possível receber a visita da Investigadora francesa, Dra. Nati Poulain, do Centro de Recherches de Tours, que tem prestado um excelente contributo na realização de ensaios de fertilização “in vitro” de oócitos ovinos com o objectivo de testar “in vitro” a fertilidade de carneiros das raças Saloia e Serra da Estrela. 9. Encargos financeiros
O orçamento programado (36 767 €) foi concretizado em 90,5%, permitindo a realização das acções de investigação previstas. O relatório financeiro foi elaborado pela Repartição Financeira da EZN. 10. Estágios
Este projecto permitiu a realização de 3 estágios intercalares do Curso de Medicina Veterinária da Universidade de Évora. No decurso deste projecto tivemos a oportunidade de realizar um estágio no Laboratório de Congelação de Sémen da Unidade Experimental de Inseminação Caprina e Ovina (INRA), sob a orientação do Professor Bernard Leboeuf. 11. Publicações Barbas, J.P., Baptista, C.C., Canas Serras, C., Marques, C.C., Cavaco-Gonçalves, S., Vasques,
M.I. e Horta, A.E.M. (2004). First results in Portugal of artificial insemination by cervical route using ram frozen semen on reproductive performance. 15th International Congress on Animal Reproduction Brasil, Porto Seguro, Brasil, 8-12 Agosto.
Barbas, J.P., Horta, A.E.M., Baptista, C.C. e Marques, C.C. (2003). Variação na capacidade fertilizante do sémen refrigerado e congelado em ovelhas de raça Saloia. Proceedings do IV Congresso Ibérico de Reprodução Animal, Federação Ibérica de Reprodução Animal (Publ.), 11-12 Julho, Las Palmas, Canarias, p. 63.
Barbas, J.P., Horta, A.E.M., Baptista, C.C., Marques, C.C e Canas Serras, C. (2003). Inseminação artificial de ovelhas Saloias com sémen refrigerado e congelado. Proceedings da XXIV Reunião de Primavera da Sociedade Portuguesa de Pastagens e Forragens, XLII Reunião da Sociedade Portuguesa de Ovinotecnia e Caprinotecnia, 9 a 11 de Abril, Palmela, Portugal, pp. 34-35.
Barbosa, A (2003). Produção Animal, Tecnologia de Sémen, Controlo do Ciclo Éstrico e Inseminação Artificial em Pequenos Ruminantes. Estágio intercalar para obtenção da licenciatura em Medicina Veterinária pela Universidade de Évora.
Cavaco Gonçalves, S., Marques, C.C., Horta, A.E.M, e Figueroa, J.P. (2005). Cervical electrical activity during oestrus in progestagen treated ewes. Submetido para publicação no Animal Reproduction Science.
Lopes, F. (2004). Tecnologia do Sémen e Inseminação Artificial em Pequenos Ruminantes. Estágio intercalar para obtenção da licenciatura em Medicina Veterinária pela Universidade de Évora.
Marques, C., Barbas, J., Horta, A., Baptista, M., Vasques, M., Mascarenhas, R., Pereira, R. E Gonçalves, S. (2005). Refinamento das técnicas de inseminação artificial e produção in

- 36 -
vitro de embriões em ovinos de raças nacionais, usando sémen congelado. Relatório Final do Projecto PIDDAC nº 841, EZN – INIAP.
Soudo, A. (2003). Reprodução Controlada em Pequenos Ruminantes: Tecnologia do sémen, Controlo do Ciclo Éstrico e Inseminação Artificial. Estágio intercalar para obtenção da licenciatura em Medicina Veterinária pela Universidade de Évora.
12. Referências bibliográficas Barbas, J., Baptista, C. e Horta, A. (2002). Comparação entre dois métodos de congelação de
sémen de carneiros Merino Regional e Serra da Estrela ao longo do ano. Livro de Actas, 100 anos da Sociedade Portuguesa de Ciências Veterinárias, 473-474, Oeiras, Taguspark, 10 a 12 de Outubro.
Barbas, J., Mascarenhas, R., Horta, A., Tavares, H., Cannas-Serra, C., Cardigos, L., Moura, R. e Cannas Simões, J.P. (2004). Influência do diluidor, época e exploração, nos resultados da IA em ovelhas de raça Saloia. Revista Portuguesa de Ciências Veterinárias, 99 (549): 59-63.
Barbas, J., Sousa, J., Ferreira, G., Baptista, C. e Horta, A. (2001). Variação anual das caracteristicas seminais de carneiros Merino Regional e Serra da Estrela em sémen fresco. X Congresso de Zootecnia, “Progressos Zootécnicos nos Países de Língua Oficial Portuguesa. EZN, Vale de Santarém. Revista Portuguesa de Zootecnia, 8(1): 312-323.
Cognié, Y., Baril, G., Poulin, N. e Mermillod, P. (2003). Current status of embryo technologies in sheep and goat. Theriogenology, 59:171-188.
Cognié, Y., Poulin, N. e Mermillod, P. (2002). Cysteamine improves in vitro goat oocyte maturation in defined medium. Proc.18Th Mtg assoc. Eur. Trans Emb (AETE), 154 (abstract).
Crozet, N., Huneau, D., De Smedt V., Theron, M-C., Szollozi, D., Torres, S. e Sevellec, C. (1987). In vitro fertilisation with normal development in sheep. Gamete Research, 16:159-170.
De Matos, DG, Furnus, C.C., Moses D.F., Martinez, A.G. e Matkovic, M. (1996). Stimulation of glutathione synthesis of in vitro matured bovine oocytes and its effects on embryo development and freezability. Molec. Reprod. Dev., 45: 451-457.
Evans, G. e Maxwell, W. (1990). Inseminacion Artificial de Ovejas e Cabras. Editorial Acribia, S. A. Zaragosa (España).
FAO. (1993). Manuel de formation pour l’ insemination artificielle chez les ovins et les caprins. Étude FAO Production Et Santé Animales, nº 83.
Ferreira, G , Sousa, J, Barbas, J. e Horta, A. (2001). Teste de endosmose (Host) em sémen de caprinos da raça Serrana. III Congresso Ibérico de Reprodução Animal. Fundação Cupertino de Miranda, Porto, 5 a 8 de Julho. Livro de Resumos, 559-604.
Lu, K.H.,Gordon, I., Boland, M.P. e Crosby, T.F. (1987). In vitro fertilization of bovine oocytes matured in vitro.Proceedings of the british Society of Animal Production (Winter Meeting), Paper 81.
Sousa, J., Barbas, J., Ferreira, G. e Horta, A. (2001). Variação anual das características seminais em bodes de raça Serrana. X Congresso de Zootecnia, “Progressos Zootécnicos nos Países de Língua Oficial Portuguesa. EZN, Vale de Santarém. Revista Portuguesa de Zootecnia, 8(1): 297-311.
StatSoft, Inc., 1995. Statistica for Windows (computer program manual). Tulsa, OK: Statsoft, Inc., 2325 East 13 th Street, Tulsa, OK, 74104, USA.
Wang, S., Liu, Y., Holyoak, A., Evans, R.C., Bunch,T.D. (1998). A protocol for in vitro maturation and fertilizatin of sheep ocytes. Small Ruminant Research, 29: 83-88.

- 37 -
13. Resumo
Pretendeu-se prosseguir os trabalhos de congelação de sémen com o objectivo da sua utilização na IA em condições de rebanho e avaliar a influência do período de congelação do sémen sobre os resultados da IA em ovelhas inseminadas na época tradicional de reprodução (Primavera). Foram efectuados estudos da contractibilidade do canal cervical através da técnica da electromiografia, avaliando-se igualmente o efeito de agentes dilatadores cervicais. Propôs-se também determinar a capacidade fertilizante do sémen de carneiro congelado em duas épocas diferentes, através da tecnologia de fertilização (IVF) e produção de embriões (IVP) in vitro.
As colheitas de sémen foram efectuadas nas épocas da Primavera, Outono e Inverno utilizando o método de recolha com vagina artificial. Unicamente foram submetidos ao processo de congelação os ejaculados que apresentaram MI ≥ 65 % e concentrações > 2 mil milhões de spz/ml. O sémen recolhido foi processado e congelado de acordo com o protocolo descrito por Evans e Maxwell (1990). Foram utilizados 83 ejaculados para avaliação estatística de resultados. A congelação das palhinhas foi efectuada utilizando um equipamento designado por “Floating Semen Freezing Rack” (Minutub GnbH & Co, KG, Germany). As variáveis independentes estudadas foram a época do ano, o factor carneiro e a interacção época x carneiro. No sémen fresco (SF) foram avaliados o volume (ml), a MI (%), a concentração espermática, a % de spz vivos e normais e a % de anomalias da cabeça, (PI) e cauda (variáveis dependentes na análise estatística).
No sémen fresco (SF) a mobilidade, vitalidade e normalidade espermáticas correlacionaram-se positivamente entre si, sendo a correlação entre a mobilidade e a vitalidade a mais elevada (r=0,63; p<0,05). A mobilidade correlacionou-se negativamente com as anomalias da cabeça (p<0,05). Os spz vivos correlacionaram-se negativamente com as anomalias da cabeça (p<0,05). Os spz normais correlacionaram-se negativamente com todas as anomalias estudadas (p<0,05). Não foram observadas variações estacionais (P, O, I) para o volume, concentração e anomalias da cabeça e PI. A MI no Outono foi superior ao Inverno. No Outono foi observada uma maior % de spz vivos. Houve maior % de spz normais no Outono que na Primavera e menor % de anomalias da cauda no Outono. Não foram observadas variações individuais na MI e na % de anomalias (cabeça, PI e cauda).
Para o sémen congelado (SD) e relativamente às três épocas consideradas (P, O, I) observou-se um efeito significativo da estação do ano sobre a % de Difvivos, verificando-se no Outono um efeito significativo da congelação sobre a diminuição da % de spz vivos comparativamente às restantes épocas. Nos restantes parâmetros seminais nomeadamente MI, % de spz vivos, normais, anomalias da cabeça, PI e cauda, Difmi e Difnormais não foi observado um efeito da época. No sémen congelado (SD) não foram observados variações individuais em nenhum dos parâmetros seminais avaliados. No SD os spz móveis e vivos apresentaram correlações positivas e significativas entre si. A mobilidade individual, correlacionou-se negativamente com as anomalias da cabeça (-0,27). A percentagem de spz vivos correlacionou-se positivamente com a dos normais (0,36) e negativamente com as anomalias da cabeça (-0,37). No SF vs SD foram observadas correlações positivas e significativas entre os parâmetros homólogos exceptuando as anomalias da PI.
A análise multivariada para avaliar a interacção época (O e I) x carneiro não foi possível ser realizada em todos os carneiros sendo excluído 1 carneiro (801) da avaliação dos resultados, pois neste animal só foi possível recolher sémen durante o Inverno. No SF foram observados efeitos específicos significativos da época sobre a MI, concentração e % de anomalias da PI. No Outono foram registadas maiores mobilidades individuais (MI), concentrações e anomalias da PI do que no Inverno. Verificaram-se variações entre carneiros no volume e concentração tal como

- 38 -
interacções significativas época x carneiro para estes parâmetros. Foram observados efeitos específicos significativos da época do ano sobre os Difmi e Difvivos, tendo-se verificado que as quebras do sémen descongelado relativamente ao sémen fresco para a MI (Difmi) e spz vivos (Difvivos) foram superiores no Outono comparativamente ao Inverno. No SD foi observado um efeito específico do carneiro sobre a % de spz normais não tendo sido observadas interacções significativas entre as épocas (Outono e Inverno) e os carneiros para qualquer dos parâmetros seminais avaliados no SD.
A IA foi realizada por via cervical (n=440) entre as 54-56 horas após a remoção das esponjas vaginais (Rev) sem prévia detecção do estro utilizando duas doses de sémen por ovelha contendo 600 milhões de spz totais. No Outono não foram observadas variações significativas entre carneiros relativamente à fertilidade e fecundidade, tendo-se registado diferenças relativamente à prolificidade. No Inverno foram observadas variações individuais relativamente à fertilidade e fecundidade. Num total de 440 ovelhas inseminadas com 5 carneiros Saloios não foram observadas diferenças significativas entre épocas (Outono e Inverno) relativamente à fertilidade (15,24 % vs 14,85 %), fecundidade (19,61 % vs 20,65 %) e prolificidade (128 % vs 139 %). Nos carneiros utilizados na IA com sémen congelado foram observadas correlações positivas nos parâmetros seminais MI, % de spz vivos e normais, existindo uma correlação positiva altamente significativa entre a MI e a % de spz vivos. Não foram encontradas correlações significativas entre estas características dos ejaculados com a fertilidade, fecundidade e a prolificidade nas ovelhas inseminadas.
Quanto aos testes de fertilidade in vitro, determinámos a taxa de mobilidade (%MIPD) após a descongelação do sémen, as taxas de mobilidade (%MISU) e capacitação (%CAP) do sémen após o swim up assim como a sua concentração (CONC, 1x106 spz mL-1) e as taxas de fertilização in vitro, determinadas pelas taxas de clivagem (% Clivagem) dos oócitos e de produção de embriões (% Emb D7) ao dia 7 de desenvolvimento. A raça Saloia apresentou uma taxa de mobilidad e do sémen após a descongelação (%MIPD) e de concentração (CONC, 1x106 spz mL-1) após o swim up significativamente superiores no Outono (p <0,05), relativamente ao sémen na mesma raça congelado no Inverno. Dentro da mesma época, a raça Saloia foi, no Outono, significativamente superior quanto à MIPD e à CONC e, no Inverno, significativamente superior quanto à capacitação (%CAP) relativamente à raça Serra da Estrela. Embora as taxas de fertilização obtidas com o nosso primeiro sistema de produção de embriões ovinos in vitro sejam pouco elevadas nesta espécie, poderemos no entanto sugerir que a interacção entre a raça e a época de congelação do sémen foi significativa para a taxa de produção de embriões (p=0,04), o mesmo não acontecendo com a taxa de clivagem, em que não houve influência daqueles dois factores. Entre épocas diferentes, para a raça Saloia, a taxa de embriões mostrou ser significativamente superior quando se utiliza sémen de Outono relativamente ao sémen de Inverno (P<0,007). Para a raça Serra da Estrela não existem diferenças entre épocas. O sémen de raça Saloia congelado quer no Outono quer no Inverno produziu significativamente mais embriões que o sémen Serra da Estrela congelado nas épocas correspondentes. Foram verificadas correlações positivas e significativas entre as taxas de clivagem e a capacitação dos espermatozóides após swim up (p=0,39) e entre as taxas de clivagem e a concentração do sémen após swim up (p=0,30). Observou-se também uma correlação positiva entre a taxa de embriões (% Emb) e a capacitação espermática (p=0,54).


















