RENAN MODESTO MONTEIRO - uenf.br · in EM Ca2+ e ∆pH, respectivamente. Os dados suportam a idéia...
68
NOVAS PROPRIEDADES DAS V H + -ATPases DE Saccharomyces cerevisiae: REGULAÇÃO DA ENZIMA PELOS CÁTIONS DIVALENTES CÁLCIO E MANGANÊS E PELA GLICOSE EXTRACELULAR RENAN MODESTO MONTEIRO UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE DARCY RIBEIRO – UENF CAMPOS DOS GOYTACAZES / RJ FEVEREIRO – 2010
Transcript of RENAN MODESTO MONTEIRO - uenf.br · in EM Ca2+ e ∆pH, respectivamente. Os dados suportam a idéia...

NOVAS PROPRIEDADES DAS V H+-ATPases DE
Saccharomyces cerevisiae: REGULAÇÃO DA ENZIMA PELOS
CÁTIONS DIVALENTES CÁLCIO E MANGANÊS E PELA GLICOSE
EXTRACELULAR
RENAN MODESTO MONTEIRO
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE
DARCY RIBEIRO – UENF
CAMPOS DOS GOYTACAZES / RJ
FEVEREIRO – 2010

i
NOVAS PROPRIEDADES DAS V H+-ATPases DE
Saccharomyces cerevisiae: REGULAÇÃO DA ENZIMA PELOS
CÁTIONS DIVALENTES CÁLCIO E MANGANÊS E PELA GLICOSE
EXTRACELULAR
RENAN MODESTO MONTEIRO
Orientador: Prof. Dr. Lev Alexandrovitch Okorokov
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE
DARCY RIBEIRO – UENF
CAMPOS DOS GOYTACAZES / RJ
FEVEREIRO – 2010
“Dissertação apresentada ao Centro de Biociências e
Biotecnologia da Universidade Estadual do Norte
Fluminense, como parte das exigências para obtenção
do título de Mestre em Biociências e Biotecnologia”

ii
NOVAS PROPRIEDADES DAS V H+-ATPases DE
Saccharomyces cerevisiae: REGULAÇÃO DA ENZIMA PELOS
CÁTIONS DIVALENTES CÁLCIO E MANGANÊS E PELA GLICOSE
EXTRACELULAR
RENAN MODESTO MONTEIRO
Aprovada em 22 de fevereiro de 2010 Comissão examinadora:
Prof. Dr. Alessandro Coutinho Ramos - UVV
Profa Dra. Anna Lvovna Okorokova Façanha – UENF
Dra. Flavia Emenegilda da Silva - UENF
Prof. Dr. Lev Alexandrovitch Okorokov - UENF
“Dissertação apresentada ao Centro de Biociências e
Biotecnologia da Universidade Estadual do Norte
Fluminense, como parte das exigências para obtenção
do título de Mestre em Biociências e Biotecnologia”

iii
Aos meus pais, Itamar e Luzia,
A minha irmã Karina,
A minha namorada Thatiane
por todo amor, carinho e compreensão.

iv
AGRADECIMENTOS
À Deus por ter me dado forças para superar todas as dificuldades.
Ao professor Lev, por ter aceitado me orientar; por sua amizade, ensinamentos,
paciência e grande dedicação.
Aos meus pais, Itamar e Luzia, por todo amor e por terem me apoiado e minha irmã
Karina, por todos os momentos bons e por toda ajuda e carinho.
A minha namorada Thatiane, por todos os momentos inesquecíveis que passamos
juntos, pelos conselhos e principalmente pelo carinho.
Ao prof. Alessandro, a profa. Anna e profa.Flavia por terem aceito o convite de
participar da banca.
Ao professor Carlos Eduardo por revisar a dissertação.
A todos os membros e amigos do grupo de trabalho: Camila, Flavia, Layz, Géssika,
Mariana pelo suporte e ótimo convívio.
Aos amigos do LFBM, Flávias, Ana Cristina, Ludmila, Viviane, André, Humberto,
Izabela, Érika, Suzana, Mariângela, Luiz, Luana pelo convívio e amizade.
Aos professores do LFBM Anna, Valdirene, João e Júlio pelo convívio e pelos
ensinamentos.
À UENF, CNPq e FAPERJ por terem financiado esse projeto e tornado possível a
realização do mesmo.

v
SUMÁRIO
ABREVIATURAS.............................................................................................. vii
RESUMO........................................................................................................... viii
ABSTRACT....................................................................................................... ix
1- REVISÃO BIBLIOGRÁFICA......................................................................... 01
1.1 - Homeostase iônica.............................................................................. 01
1.2 – H+-ATPase.......................................................................................... 01
1.3 - V H +-ATPases..................................................................................... 02
1.3.1 - Estrutura das V H+-ATPases........................................................ 05
1.3.2 - Diferentes formas de V H+-ATPases............................................. 09
1.3.3 - Regulação das V H+-ATPases...................................................... 10
1.3.3.a Regulação pela glicose extracelular............................................. 10
1.3.3.b Formação de ligações dissulfeto.................................................. 10
1.3.3.c Atividade de canais aniônicos........................................................ 11
1.3.3.d Outras formas de regulação........................................................... 12
1.3.4 - Inibidores de V H+-ATPases........................................................... 15
2- OBJETIVOS..................................................................................................... 16
2.1 - Objetivo principal................................................................................... 16
2.2 - Objetivos específicos............................................................................. 16
3- MATERIAL E MÉTODOS................................................................................. 17
3.1- Cepa de levedura................................................................................... 17
3.2- Meio de cultura e manutenção da cepa................................................. 17

vi
3.3- Preparo do pré-inóculo........................................................................... 17
3.4- Preparo da levedura para o isolamento................................................. 17
3.5- Isolamento de membranas e fracionamento subcelular........................ 18
3.5.1- Obtenção de células...................................................................... 18
3.5.2- Obtenção dos esferoplastos.......................................................... 18
3.5.3- Pré-incubação dos esferoplastos com glicose............................... 19
3.5.4- Obtenção de membranas totais..................................................... 19
3.5.5- Obtenção de membranas intracelulares fracionadas em
gradiente de densidade de sacarose........................................................ 20
3.6- Determinação do ∆pΗ (transporte de H+) (Okorokov & Lichko, 1983)..... 21
3.7- Determinação do conteúdo de proteína (Bradford, 1976)...................... 22
3.8- Determinação da hidrólise de ATP.......................................................... 23
4- RESULTADOS.................................................................................................. 24
4.1 Efeito do Ca2+ sobre o transporte de H+ realizado pela V H+-ATPase...... 24
4.2 Efeito do Mn2+ sobre o transporte de H+ realizado pela V H+-ATPase....... 26
4.3 Efeito do KNO3 sobre a atividade hidrolítica das V H+-ATPases................. 29
4.4 Efeito do KNO3 sobre o transporte de H+ mediado pela V H+-ATPase........ 34
4.5 Efeito da glicose sobre a ativação e o acoplamento da V H+-ATPase......... 37
5 – DICUSSÃO..................................................................................................... 42
6- CONCLUSÕES................................................................................................. 45
7 - REFERÊNCIAS BIBLIOGRÁFICAS............................................................... 47

vii
ABREVIATURAS
Abs Absorbância
ACMA 6-amino-6cloro-2-metoxiacridina
ATP Adenosina Trisfosfato
ATPase Adenosina trifosfatase
DO Densidade ótica
DTT Ditiotreitol
EM Potencial de membrana
Fmáx Amplitude máxima
g Gravidade
GDP Guanosina Difosfato
GDPase Guanosina Difosfatase
H+-ATPase ATPase translocadora de próton
MOPS Ácido 3-[N-morfolino] propano sulfônico
pH Potencial hidrogeniônico
P H+-ATPase ATPase translocadora de próton de membrana plasmática
RE Retículo endoplasmático
rpm Rotação por minuto
V H+-ATPase ATPase translocadora de próton vacuolar
Vo Velocidade inicial do transporte de H+
Vmáx Velocidade máxima
YEPD Yeast Extract Peptone Dextrose
∆ ∆ ∆ ∆ pH Gradiente de prótons através da membrana

viii
RESUMO
As V H+-ATPases são enzimas-chaves em diversos processos importantes,
como endocitose, tráfico de proteínas, transporte ativo e armazenamento de
metabólitos, fusão e fissão de membranas. O primeiro objetivo do nosso trabalho foi
comparar o efeito de íons Ca2+ e Mn2+ no transporte de H+ mediado por V H+-
ATPase das organelas da via de secretória em Saccharomyces cerevisiae após a
energização das membranas com glicose extracelular. A velocidade inicial (V0) do
transporte de H+ nessas membranas foi inibida diferentemente pelo Ca2+ e o Mn2+
estimulou seletivamente a V0. O efeito do Ca2+ e Mn2+ na formação do ∆pH em
vacúolos foi interpretada supondo que os cátions são transportados por trocadores
Me2+/H+ e o Ca2+ fecha o canal de ânions, enquanto o Mn2+ abre convertendo o ∆µH
in EMCa2+ e ∆pH, respectivamente. Os dados suportam a idéia da presença de
diferentes formas da enzima no RE, Golgi, vacúolos e envelope nuclear da levedura.
O segundo objetivo foi compreender como a V H+-ATPase pode ser regulada pela
glicose extracelular, uma vez que o complexo catalítico V1 não dissocia da
membrana. Nós constamos que: (i) os estados ativado e semi-ativado da enzima
mostram a mesma sensibilidade da inibição da ATPase pelo nitrato, sugerindo a
ligação idêntica do complexo V1 com a membrana; (ii) a velocidade inicial de
transporte de H+ foi inibida de forma de forma mais eficaz do que a atividade de
ATPase, apontando para a atividade de desacoplamento do nitrato, que é
significantemente maior para a enzima não ativada; (iii) a bomba ativada exibiu
maior acoplamento; (iv) a população das moléculas da enzima ativada de MT
consiste de sub-populações que mostra acoplamento diferente apresentando maior
resistência ao efeito de desacoplamento pelo nitrato. Nós sugerimos que uma
alteração bioquímica e a modulação do acoplamento da bomba são fatores chave do
seu regulamento, que não exige a dissociação do complexo V1 da membrana

ix
ABSTRACT
The V H+-ATPases are key enzymes in several important processes including
endocytosis, trafficking proteins, active transport and storage of metabolites, fusion
and fission of membranes. The first aim of our work was to compare the effect of ions
Ca2+ e Mn2+ on the H+ transport mediated by V H+-ATPases of the secretory pathway
organelles in Saccharomyces cerevisiae after energization of the membranes with
extracellular glucose. The initial velocity (V0) of H+ transport in those membranes was
inhibited by Ca2+ differently and Mn2+ selectively stimulated V0.. The effect of Ca2+
and Mn2+ in the formation of ∆pH in vacuoles was interpreted assuming that cations
are transported by Me2+/H+ exchangers and Ca2+ closes the anion channel while the
Mn2+ opens it converting ∆µH in EMCa2+ and ∆pH, respectively. The data support the
idea of the presence of different forms of the enzyme in the ER, Golgi, vacuoles and
nuclear envelope of yeast. The second goal was to understand how V H+-ATPase
can be regulated by extracellular glucose given that the V1 catalitic complex does not
dissociate from membranes.We found that: (i) semi-active and activated enzyme
states show the same ATPase sensitivity to inhibition by nitrate, suggesting the
identical binding of their V1 complexes to membrane; (ii) the initial velocities of the H+
transport were inhibited more effectively than the ATPase activity, pointing towards
the nitrate uncoupling action, which is significantly higher for the not activated
enzyme; (iii) the activated pump exhibited a higher coupling; (iv) the population of the
activated enzyme molecules of TM consist of the sub-populations showing the
different coupling presented higher resistance to the uncoupling effect of nitrate. We
suggest that a biochemical modification and the coupling modulation of the pump are
key factors of its regulation which does not require the dissociation of the V1 complex
from membrane.

1
1 – REVISÃO BIBLIOGRÁFICA
1.1 Homeostase iônica
Todos os processos da vida são realizados em condições ideais de íons: H+,
Ca2+, Mg2+, K+, Na+, Cl-, Pi entre outros. A homeostase iônica é um evento crítico
para diversos processos fisiológicos como biossíntese/degradação, enovelamento,
endereçamento de proteína, secreção protéica, fusão e divisão de membranas,
morfogênese de organelas e células, dinâmica de microtúbulos e divisão celular.
Desvios nessa homeostase podem causar doenças/ fenômenos patológicos tanto no
homem quanto em animais e plantas (Voet et al., 2000).
A homeostase iônica é mantida pela atividade coordenada de diferentes
transportadores. As proteínas transportadoras podem ser agrupadas em três
grandes classes: ATPases, carreadores e canais (Voet et al., 2000). Essa
classificação é baseada em diferentes formas de utilização da energia: ATPases que
são bombas protônicas primárias, que criam o gradiente protônico, utilizando
diretamente a energia da hidrólise do ATP, para transportar íons contra um gradiente
de concentração; e os transportadores secundários, que utilizam o gradiente
protônico para transportar íons, sendo eles os carreadores e os canais (Alberts et al.
1999).
1.2 H+-ATPase
As H+-ATPases, também chamadas bombas de H+, constituem uma grande
família de enzimas que transportam prótons através das membranas contra o
gradiente eletroquímico de H+ usando a energia da hidrólise do complexo ATP-Mg
(Nelson, 1992).
As H+-ATPases são enzimas eletrogênicas, ou seja, possuem a capacidade
de criar uma diferença de cargas elétricas através da membrana, sendo essa
diferença chamada de potencial de membrana (EM), assim como pode criar,
simultaneamente, um gradiente de concentração de prótons (∆pH) (Pedersen &
Carafoli, 1987).
Sabe-se da existência de três tipos de H+-ATPases: as H+-ATPases do tipo F
(F1Fo H+-ATPases), H+-ATPases do tipo P (P H+-ATPases) e as H+-ATPases do tipo
V (V H+-ATPases ou V1V0 H+-ATPases) (Pedersen & Carafoli, 1987).

2
As F1Fo H+-ATPases estão localizadas nas mitocôndrias, na membrana de
bactérias (B1Bo H+-ATPases) e cloroplastos (C1Co H
+-ATPases), as H+ ATPases do
tipo F são inibidas por azida e oligomicina. Estas enzimas consiste de um setor
catalítico F1, composto das subunidades α, β, γ, δ, ε e o setor da membrana, F0
responsável pelo transporte de prótons é composto pelas subunidades a, b, c (Futai
et al., 1989; Boyer, 1997). Funcionalmente, essas H+-ATPases são ATP sintases,
pois são responsáveis pela síntese de ATP. Foi mostrado que, in vitro, elas também
podem hidrolisar ATP (Pedersen & Carafoli, 1987). Já as ATPases do tipo P e V
utilizam a energia da hidrólise do ATP para criar o gradiente de H+ transmembranar
(Pedersen & Carafoli, 1987).
As P H+-ATPases são enzimas chaves da membrana plasmática de
leveduras, fungos filamentosos e plantas (Serrano, 1983; Pedersen & Carafoli,
1987). Esta enzima fornece energia do gradiente de prótons para transportar íons e
metabólitos através da membrana plasmática e possui um papel fundamental na
manutenção do pH intracelular (Gouffeau & Slayman, 1981; Axelsen & Palmgreen,
1998; Scarborough, 1999). Essa enzimas são inibidas por ortovanadato, um análogo
do fosfato inorgânico (Goffeau & Slayman, 1981; Bowman et al., 1986; Pedersen &
Carafoli, 1987).
As ATPases do tipo P são divididas em cinco grupos, baseando-se na
seletividade de íons diferentes que transportam. Além da afinidade aos íons, elas
também se diferenciam pela estrutura primária (Catty et al., 1997; Axelseen &
Palmgreen, 1998).
1.3 V H +-ATPases
As V H+-ATPases possuem como função primária acoplar a hidrólise de ATP
ao transporte de H+ através das membranas biológicas. A força próton motriz gerada
pelas V H+-ATPases em organelas de células eucarióticas é usada em vários
processos secundários na célula (Nelson & Harvey, 1999). Elas são importantes
para o transporte secundário de vários solutos, dirigindo uma variedade de sistemas
auxiliares de transporte ativo através dos co-transportadores e trocadores de H+
(Beyenbach & Wieczorek, 2006).
As V H+-ATPases foram primeiramente descritas em estudos sobre a
captação de catecolamina em grânulos de cromafina. Nestes estudos foi
demonstrado que uma ATPase energizava essas membranas por uma absorção de

3
H+ dependente de ATP e que o gradiente protônico impulsiona o acúmulo de
catecolaminas pela troca de prótons. Logo depois foi demonstrado que uma bomba
de prótons similar operava em vacúolos de fungos e plantas (Nelson e Harvey,
1999). Foram denominadas ATPases do tipo V por terem sido caracterizadas
inicialmente nas membranas vacuolares (Pedersen & Carafoli, 1987; Rea et al.,
1987; Bowman & Bowman, 1988; Barkla & Pantoja,1996).
As V H+-ATPases são encontradas em membranas intracelulares, incluindo
endossomos, lisossomos, grânulos de cromafina, vesículas sinápticas, vesículas
cobertas de clatrina, Golgi, vesículas secretórias e vacúolo de plantas e eucariotos
inferiores (Stevens & Forgac, 1997; Forgac, 1998). Também são encontradas em
membranas plasmáticas de células animais especializadas como as células renais,
células epididimais, macrófagos, osteoclastos, osteoblastos e células tumorais
(Klionsky et al., 1990; Merzendorfer et al., 1997; Nelson. & Harvay, 1999; Wieczorek
et al., 1999; Beyenbach & Wieczorek, 2006).
O bombeamento de prótons realizado pelas V H+-ATPases é de vital
importância para vários processos intra e intercelulares, dentre eles a endocitose
mediada por receptores, o tráfego de proteínas, neurotransmissões, o transporte
ativo e armazenamento de metabólitos (Finbow & Harrison, 1997; Stevens & Forgac,
1997; Nelson & Harvey, 1999; Futai et al., 2000; Nishi & Forgac, 2002). Além do
importante papel na regulação do pH do citosol e na acidificação dos lisossomos e
vacúolos de plantas e fungos, criando o pH ótimo para o funcionamento das
hidrolases, incluindo proteases, glicosidases, lípases, nucleases e fosfatases
(Beyenbach & Wieczorek, 2006).
A acidificação de compartimentos dependente da V H+-ATPase é crucial para
a entrada de alguns vírus e bactérias dentro da célula. A hemaglutinina, proteína
capsidial do vírus da gripe, por exemplo, atua como um meio de fusão dependente
do pH, mediando a fusão entre a membrana viral e a membrana endossomal, isso
facilita a liberação do material genético do vírus no citoplasma da célula hospedeira
(White, 1992). Da mesma forma, as toxinas, tais como a toxina da difteria e a toxina
do antraz, contêm subunidades formadoras de poros que, em resposta ao baixo pH
endossomal após a sua interiorização, facilita a inserção da parte citotóxica no
citoplasma (Kevin et al., 2008).
Nos endossomos de células renais de hamster, as V H+-ATPases promovem
um ambiente ácido que serve como um sinal que ativa a liberação de ligantes

4
internos (lipoproteínas de baixa densidade e insulina) de seus receptores
(receptores que foram internalizados). Esta liberação permite que os receptores
reciclem para superfície celular, onde eles podem ser reutilizados, enquanto os
ligantes são direcionados para os lisossomos para a degradação (Claque, 1994).
São também encontradas no Plasmodium falciparum (agente causador da
malária) e Toxoplasma gondii (agente causador da toxoplasmose), em suas
membranas plasmáticas e vacúolos onde está envolvida na energização de
transportes secundários de várias substâncias (Moriyama et al., 2003; Beyenbach &
Wieczorek, 2006).
Em plantas, V H+-ATPases estão localizadas no tonoplasto e em outros
compartimentos intracelulares, no RE e no Golgi (Oberbeck et al., 1994). No
tonoplasto as V H+-ATPases correspondem a 6,5-35% do total de proteína, variando
de acordo com a espécie. A V H+-ATPase é indispensável para o crescimento de
plantas sob condições normais devido ao seu papel na energização do transporte de
solutos, endereçamento de proteínas, homeostase de íons/metabólitos,
fusão/divisão de vesículas de membranas (Clague et al., 1994; Matsouka et al.,
1997; Nelson e Harvey, 1999; Forgac, 1999; Ratajczak, 2000; Futai et al., 2000).
Sob condições de estresse como salinidade, seca, frio, estresse ácido, anoxia e
excesso de metais pesados no solo a sobrevivência da célula depende da
manutenção ou ajuste da atividade da V-ATPase (Dietz et al., 2001).
Na membrana plasmática do intestino da lagarta do tabaco Manduca sexta, o
trocador de K+/H+, utiliza o gradiente eletroquímico de H+, criado por V H+-ATPases,
para transporte do K+ (Wieczorek, 1991).
Em leveduras, as V H+-ATPases foram descritas não apenas nas membranas
vacuolares (Ohsumi e Anraku, 1981) mas também em todas as organelas da via
secretória (Okorokov, 1997; Okorokov et al, 2001; Samarão, 2003; Teodoro, 2004) e
nas vesículas secretórias (Alves, 2006). Elas são essenciais para o acúmulo de
aminoácidos básicos como arginina, ácidos orgânicos, purinas, pirimidinas, S-
adenosilmetionina, Ca2+, Mg2+, Mn2+, Zn2+ e fosfato inorgânico bem como para
energizar o balanço osmótico (Okorokov et al., 1980; Oshumi & Anraku, 1983;
Okorokov & Lichko, 1983; Okorokov et al., 1985; Harvey, 1992).
Em células animais as V H+-ATPases podem desempenhar um importante
papel na membrana plasmática de células especializadas. Estão envolvidas na
maturação e armazenamento de esperma no lúmem epididimal (Breton et al., 1996;

5
Beyenbach & Wieczorek, 2006). Em células renais as V H+-ATPases são
responsáveis pela secreção de íons de H+ nas células do ducto coletor, túbulos
proximais e segmentos dos néfrons (Gluck, 1992; Gluck et al., 1996), enquanto que
nos macrófagos e neutrófilos estas estão envolvidas na homeostase do H+ (Swallow
et al., 1993).
Em céluas tumorais as V H+-ATPases são direcionadas para a membrana
plasmática onde elas criam um ambiente extracelular acidífero, o qual é necessário
para a metástase tumoral (Zaguilan, 1993).
As V H+-ATPases desempenham um papel importante também nos
osteoclastos, os quais são responsáveis pela reabsorção óssea (Toyomura et al.,
2003). O meio ácido promovido por essas enzimas dissolve a matriz óssea e ativa
proteases que participam na reabsorção óssea. Pacientes com defeitos genéticos na
isoforma da V H+-ATPase expressa na membrana plasmática de osteoclastos
desenvolvem um distúrbio conhecido como osteopetrose (Frattini et al., 2000), que é
caracterizado por um aumento na densidade óssea e defeitos de desenvolvimento,
devido à impossibilidade de remodelação óssea (Frattini et al., 2000).
As V H+-ATPases também estão presentes em células do cérebro, nas
vesículas sinápticas (Nelson, 1991). Em vesículas sinápticas, estão envolvidas na
transdução de sinal (Graham et al., 2000; Bowman & Bowman, 2002; Inoue &
Forgac, 2005; Beyenbach & Wieczorek, 2006).
No homem uma doença neurodegenerativa autossômica progressiva infantil
(doença de Batten) com incidência de um para 12500 nascimentos vivos e com
aproximadamente 440.000 casos notificados nos Estados Unidos esta envolvida
com problemas na atividade de H+-ATPases. Há evidência de que essa doença
promove o aumento da atividade da V H+-ATPase na membrana plasmática.
(Chattopadhyay et al., 2000; Padilla-Lo´ Pez & Pearce, 2006).
1.3.1 Estrutura das V H+-ATPases
Muitos estudos vêm sendo desenvolvidos na tentativa de elucidar as
propriedades estruturais, funcionais e regulatórias dessa enzima (Nishi & Forgac,
2002; Beyenbach & Wieczorek, 2006). Na segunda metade dos anos oitenta, a
purificação da V H+-ATPase de células animais, de plantas e de fungos revelou a
composição das subunidades dessa enzima (Uchida et al., 1985; Okorokov, et al.,

6
1985; Xie & Stone, 1986; Bowman et al., 1986; Randall & Sze, 1986; Arai et al.,
1987; Moriyama e Nelson, 1987).
Nelson e Harvey (1999) baseados em estudos em que foi mostrada a
similaridade entre F-ATPase e V-ATPase sugeriram atribuir cada subunidade da V-
ATPase a uma das 4 partes da “máquina mecânoquímica” de Paul Boyer (proposta
para a F-ATPase): (1) unidade catalítica, (2) talo, (3) gancho e (4) turbina. Essas
nomenclaturas dadas para cada parte esta relacionada à forma como a enzima
trabalha, sendo considerada um “bio-motor”, onde o ATP é o combustível e
transporte de H+ é a ação. O setor catalítico tem a função de ligar e hidrolisar ATP,
promovendo uma rotação momentânea do talo, que por sua vez irá rodar a turbina, o
talo também tem a função de manter o setor catalítico no lugar. A turbina tem a
função de conduzir H+ através da membrana e o gancho previne a rotação da
unidade catalítica (figura 1).

7
A V H+-ATPase é dividida em dois domínios: o domínio catalítico hidrofílico
(V1) com aproximadamente 570 KDa e o domínio integral de membrana hidrofóbico
(V0) com aproximadamente 260 KDa (figura 1).
O complexo V1 é usualmente descrito como um hexâmero constituído de três
cópias de cada uma das subunidades “A” e “B” (alternadas) e cópias simples das
demais subunidades (Forgac, 1999a; Graham et al., 2000) (figura 1). Sabe-se que
tanto a subunidade “A” como a “B” são constituintes da unidade catalítica; a
subunidade “A” está envolvida na hidrólise de ATP e apresenta similaridade de
Figura 1 – Estrutura e mecanismo da V-ATPase. Modelo baseado na “máquina mecanoquí-
mica” de Paul Boyer (proposta para F-ATPase). (A) Disposição das subunidades da V-ATPase,
destacando os domínios V1 (amarelo) e V0 (verde). (B) Mecanismo de rotação da V-ATPase, as
subunidades rotatórias são destacadas em azul, enquanto as estacionárias são mostradas em
laranja. A hidrólise de ATP causa mudanças conformacionais na subunidade A, que dirige a
rotação do rotor (seta vermelha). (C) Mecanismo do transporte de prótons através de V0. Prótons
(pontos vermelhos) entram no complexo V0 através de um canal hemi citoplasmático (Cipriano,
2008).
Citoplasma
Lúmen ou meio extracelular
Citoplasma
Lúmen ou meio extracelular

8
seqüências com a subunidade catalítica “β” da F –ATPase (Finbow & Harrison,
1997). Supõe-se que a subunidade catalítica “A” tenha atividade hidrolítica, desde
que os complexos V1 e Vo encontrem-se associados. A subunidade “B”, com função
reguladora, possui homologia com a subunidade “α” de F–ATPase. Essa subunidade
possui um sítio de ligação de ATP não catalítico. A subunidade “B” pode contribuir
com importantes resíduos para a função catalítica da subunidade “A” (Nelson &
Harvey, 1999).
O canal de prótons, através da membrana, é formado pelo setor Vo que
contém cinco diferentes tipos de subunidades: a, c, c’, c” e d, com massa molecular
de 100 KDa, 16 KDa, 17 KDa, 19-23 KDa e 36-38 KDa, respectivamente. Três
dessas subunidades, c, c’ e c”, são proteolipídeos altamente hidrofóbicos (Moriyama
e Nelson, 1989; Kane, 1992; Finbow e Harrison, 1997; Forgac, 1998; Nelson e
Harvey, 1999; Forgac, 1999a; Graham et al., 2000; Ratajczak, 2000; Maeshima,
2001) (figura 1).
Foi proposto que o domínio V0 das V H+-ATPases, com base no estudo da
fusão de vacúolos homotípicos de leveduras, participa ativamente na fusão de
membranas, independente do seu papel na acidificação promovida pela enzima. É
sugerido que V0 catalisa diretamente a mistura das duas bicamada lipídica em
virtude da sua composição altamente hidrofóbica (Peters et al. 2001).
O domínio V1 é composto pelas subunidades A e B da cabeça, que tem
massa molecular de 67 a 73 KDa e 55 a 60 KDa, respectivamente, e as subunidades
do talo, C (40 a 45 KDa), D (32 a 33 KDa), E (28 a 32 KDa), F (13 a 14 KDa), G (12
a 16 KDa) e H (51 a 54 KDa) (Figura 1).
A subunidade D, constituinte do talo, parece ter um papel importante no
acoplamento do transporte de H+ e hidrólise de ATP (Xu & Forgac, 2000). A
subunidade C é um polipeptídeo encontrado no talo, ela ata monômeros de actina,
durante o processo de desassociação dos complexos da V H+-ATPases (Vo e V1) em
resposta a ausência de glicose (importante mecanismo de regulação da atividade da
V H+-ATPases), a subunidade C é liberada, sugerindo que esta possa ter um papel
importante neste processo (Nelson et al., 1990). A subunidade H parece ser
importante na regulação da atividade enzimática e na comunicação da V H+-
ATPases com outros processo celulares (Finbow & Harrison, 1997; Parra et al.,
2000).

9
Acredita-se que a subunidade c (proteolipídica), constituinte da turbina, seja a
principal subunidade envolvida na translocação de prótons através da membrana
(Hirata et al., 1997). A subunidade d, também constituinte da turbina, está associada
ao complexo Vo da enzima e possivelmente tem função na associação dos dois
complexos da enzima (Vo e V1) (Bauerle et al., 1993).
A ausência da subunidade F em leveduras impede a associação do complexo
Vo ao V1 (Graham et al., 1994; Finbow & Harrison, 1997; Ratajaczak, 2000).
1.3.2 Diferentes formas de V H+-ATPases
Foi mostrado por nosso grupo que existem formas funcionalmente ativa em
cada organela da via secretória, e que em um mesmo isolamento, a atividade da
enzima pode ser diferente em cada organela (Samarão et al., 2009).
Samarão e colaboradores (2009) mostraram através de estequiometria que
em levedura a razão entre as subunidades A e B de vacúolos é de
aproximadamente 1:1, e que em membranas mais densas que o vacúolo essa razão
muda, sendo mais alta. Eles imaginaram que modificações pós-translacionais da
subunidade A e da subunidade B pode alterar a imunoreatividade e promover o
aumento da estequiometria B:A alterando a velocidade inicial (Vo) de formação do
∆pH nos diferentes compartimentos da via secretória. A modificação parece ser mais
provável na subunidade A, pois sua imunoresposta foi mais irregular no RE e no
Golgi. Contudo a diferença na razão estequiométrica de 1:1 entre as subunidades A
e B pode ser causada por outro motivo como a ligação de diferentes subunidades do
complexo V1 com microtúbulos, enzimas glicolíticas como aldolase e proteínas do
complexo RAVE. Por exemplo, microtúbulos poderiam se ligar às subunidades B
livres, bem como a enzima, todas as subunidades B dissociariam desses complexos,
na presença de SDS, aumentando o número total de subunidades B em comparação
com a subunidades A (Samarão et al., 2009).
Outra diferença esta relacionada a sensibilidade da enzima ao inibidor
bafilomicina, onde foi encontrado que em vesículas de membranas do RE a V H+-
ATPase apresentou maior resistência aquele inibidor (inibição de 50 % foi obtida
com 4,4 nM), quando comparada com vesículas de membranas do vacúolo e do
Golgi (inibição de 50 % foi obtida com 0,03 nM) (Samarão et al., 2009).

10
1.3.3 Regulação das V H+-ATPases
Existem diferentes formas de regulação das V H+-ATPases. Entre elas estão
o controle da atividade enzimática pela glicose extracelular (Kane, 1995), a formação
de ligações dissulfeto entre resíduos de cisteina (Forgac, 1999b), a atividade de
canais aniônico para a modulação da atividade da enzima além de outros
mecanismos.
1.3.3.a Regulação pela glicose extracelular
Um dos mecanismos de regulação da V H+-ATPase é a associação/
desassociação reversível dos complexos V1 e V0. O transporte de H+ feito pela V H+-
ATPase necessita de uma associação funcional dos complexos V1 e V0. A
subunidade C, do complexo V1, pode ser perdida durante o isolamento das
membranas das células não pré-incubadas com glicose (Kane, 1995) (figura 2A). É
importante destacar que para avaliação de forma associada da enzima, Kane e
outros autores, usaram a capacidade do detergente E12C9 de solubilizar junto com
as subunidades do complexo V1, as subunidades do complexo V0, principalmente a
subunidade “a”. O baixo conteúdo desta subunidade (20-30%) detectado depois da
solubilização das membranas das células incubadas sem glicose, foi interpretada
como indicação de que a maioria dos complexos V1 dissociou do complexo V0 e que
somente 20-30% não foram dissociados (Kane, 1995). A possibilidade que o
complexo V1 da enzima não ativada pela glicose extracelular ainda pode estar ligada
com V0 à membrana mais fracamente, é proposta pelo nosso grupo de trabalho onde
foi avaliado o conteúdo de subunidades A e B do complexo V1 em membranas
isoladas dos esferoplastos incubados com e sem glicose. Foi mostrado que a forte
estimulação do transporte de prótons pela glicose extracelular não necessita de um
aumento significativo do número de complexos V1. Também foi observado que as
membranas de vacúolo isoladas de esferoplastos não pré-incubadas com glicose
podem perder a atividade do transporte de H+, ou ter essa atividade baixa, mas tem
quase o mesmo conteúdo de subunidades A e B em comparação com membranas
de Golgi e/ou RE nos quais ainda apresentam atividade significativa.
1.3.3.b Formação de ligações dissulfeto
Outro mecanismo proposto para regular a atividade de V H+-ATPase in vivo
envolve a formação de ligações disulfeto reversíveis entre resíduos de cisteina no

11
sítio catalítico das V H+-ATPases. Uma ligação disulfeto inibitória pode ser formada
entre o Cys 254 e Cys 532 na subunidade A bovina, essa ligação leva à inativação
reversível da enzima (figura 2B). Em concordância com essa afirmação, a mutação
do resíduo correspondente ao Cys 254, em levedura leva a uma enzima que é
resistente à oxidação. O interessante é que uma mutação na cisteina na via
biosintética de leveduras pode levar a um defeito na acidificação vacuolar e esse
defeito pode ser corrigido pela mutação em Cys261Val, sugerindo que a formação
de ligações disulfeto possam causar a inibição da atividade de V H+-ATPase in vivo
(Forgac, 1999b).
1.3.3.c Atividade de canais aniônicos
O transporte de H+ pela V H+-ATPase é um processo eletrogênico
necessitando de um co-transporte de ânions ou efluxo de H+ em troca com outro
cátion como Ca2+, Na+ ou K+ para dissipar o potencial de membrana formado pelo H+
durante o transporte deste íon (Kakinuma et al., 1981; Okorokov & Lichko, 1983; Arai
et al., 1989). As bombas eletrogenicas protônicas são inibidas pelo produto do
processo que elas catalisam, ou seja, tanto pelo potencial elétrico da membrana
formado pelo íons H+(EMH+) quanto pela diferença de potencial químico de H+, ∆pH.
Sendo que o efeito inibitório pelo EMH+ é mais significativo comparando com o efeito
do ∆pH. Importante também lembrar que a conversão de EMH+ em EM formado por
outro cátion, por exemplo K+, Na+ ou Ca2+ em resultado a troca de H+ com esses
cátions ativa a bomba protônica, ou seja a libera do controle negativo pelo EMH+
(Okorokov & Lichko, 1983; Okorokov et al., 1985). A dissipação é realizada por
ânions que entram através de um canal de Cl-, e a sua atividade é controlada por
fosforilação dependente de proteína kinase A (figura 2C). A defosforilação do canal
de Cl- pela proteína fosfatase diminui a sua condutância e a acidificação dependente
de ATP, enquanto que a fosforilação pela proteína kinase A aumenta as duas
atividades (Mulberg et al., 1991).
O conhecimento da importância do canal aniônico para a modulação da
atividade de V H+-ATPase foi proposta para explicar os efeitos do Ca2+, Mn2+ e Mg2+
sobre a atividade de V H+-ATPase e formação de ∆µH+ em membranas de vacúolos
inteiros (Okorokov et al., 1985). Sugeriu-se que o Ca2+ poderia fechar o canal
aniônico. A troca do Ca2+ com o H+ (trocador de Ca2+/H+) estimulou a hidrolise de
ATP pela V H+-ATPase em 60% e aumentou o EM feito pelo Ca2+ em 3 vezes,

12
simultaneamente a V0 e Fmax da formação de ∆pH foram diminuídas por causa de
rápida troca de prótons com Ca2+. O Mn2+ apresentou um efeito contrário indicando
que este pode abrir o canal aniônico, mas não pode ser transportado rapidamente
pelo trocador Mn2+/H+. O Mn2+ evitou a formação do EM e transformou este em ∆pH.
O Mg2+ pode abrir parcialmente o canal aniônico e estimular a formação de ∆pH e de
EM. Assim, cátions bivalentes como Ca2+, Mg2+ e Mn2+, na levedura S.
carlsbergensis, podem regular a atividade da V H+-ATPase e modificar a
contribuição de ∆pH e potencial de membrana em ∆µH+ e com isso regular as
atividades de transportadores secundários os quais usam o ∆pH (para trocar Ca2+,
arginina, lisina ou Zn2+) e EM (para co-transportar citrato e α-ketoglutarato)(Okorokov
et al., 1985). O gradiente eletroquímico (∆µH+) é a força impulsora resultante da
combinação entre o gradiente de concentração e o gradiente elétrico para cada
soluto carregado através da membrana (Alberts et al. 1999).
1.3.3.d Outras formas de regulação
A atividade das V H+-ATPase pode ser modulada pela variação da eficiência
de acoplamento entre a hidrólise de ATP e o transporte de H+, sendo acoplamento
atribuído às diferentes formas da enzima. Esta diferença na eficiência do
acoplamento foi proposta para explicar a diferença no pH das diferentes organelas
(Nishi & Forgac, 2002; Forgac, 2007; Nelson, 2003; Kawasaki-Nishi et al., 2001)
(figura 2D).
Foi proposto também que alguns lipídios específicos podem afetar a atividade
das V H+-ATPase. Mais especificamente, foi mostrado em levedura que
esfingolipídeos com um grupo acil C26 são necessários para a geração do complexo
V1 com atividade ATPásica (Chen et al., 2004). Curiosamente, vários inibidores de V
H+-ATPase, mostraram incorporar-se à bicamada lipídica e afetar a flexibilidade
estrutural da enzima (Dixon et al. 2004; Dixon et al. 2008).
As V H+-ATPases podem ser inativadas a 0 ºC, sendo que nessa temperatura
na presença de Mg2+ e ATP a atividade de ATPase pode cair 50% em apenas 10
min. Enquanto que em temperatura ambiente a mesma diminuição da atividade é
obtida em 60 min. Foi mostrado que a inativação por baixa temperatura da V H+-
ATPase resulta na dissociação do complexo V1 periférico da membrana (Moriyama
& Nelson, 1989).

13
Alguns íons como o Mg2+, Ca2+ e Mn2+ causam mudanças na eficiência no
acoplamento do transporte de prótons e hidrólise de ATP. O aumento na
concentração de Mg2+ livre até 1,5 mM estimulou a formação de ∆pH e de potencial
de membrana em 6,6 e 2,9 vezes, respectivamente (Okorokov et al., 1985). A
hidrólise de ATP foi estimulada em 10%. Isso significa que o Mg2+ livre funciona
como fator de acoplamento do transporte de H+ e hidrólise de ATP, mas o
mecanismo do acoplamento é desconhecido (Okorokov et al., 1985).
Altas concentrações de ATP causam diminuição no acoplamento funcional
entre o transporte de H+ e a hidrólise de ATP pela V H+-ATPase, onde a hidrólise do
ATP continua a aumentar mas o transporte de H+ chega ao platô e começa a
diminuir (Arai et al., 1989).
A proteólise também causa diminuição no acoplamento, uma vez que o
tratamento com baixas concentrações de tripisina promove uma perda na atividade
de transporte de H+, mesmo apresentando 50% da hidrólise de ATP (Adachi et al.,
1990).

14
Figura 2 – Regulação das V H+-ATPases. (A) dissociação reversível da enzima promovida pela glicose extracelular, após o esgotamento da glicose, o complexo V1 se dissocia do complexo V0. (B) a formação de ligações dissulfeto reversíveis entre os resíduos Cys-254 e Cys-532 da subunidade A bloqueia o sítio catalítico em uma conformação que é incapaz de hidrolisar ATP. (C) alterações no potencial de membrana. (D) mudança na eficiência de acoplamento da bomba. (Fonte - Kevin et al., 2008 com modificações)

15
1.3.4 - Inibidores de V H+-ATPases
Os inibidores mais estudados das V H+-ATPases identificados até agora são
os antibióticos Bafilomicina A1 (Bowman & Bowman, 1988) e Concanamicina A
(Dröse et al., 1993).
A Bafilomicina foi o primeiro inibidor específico de V H+-ATPase identificado
(Bowman et al., 1988), alguns autores afirmam que a Bafilomicina A1 se liga ao setor
Vo da V H+-ATPase. Zhang et al. (1994) sugerem que a Bafilomicina A1 impede a
atividade do transporte de prótons, ligando-se a subunidade “a” (100 KDa). Contudo,
Bowman e Bowman (2002) mostraram que o local de ligação da Bafilomicina A1 ao
setor Vo é na subunidade “c” (16 KDa).
A bafilomicina A1 inibe a atividade da V H+ -ATPase de diferentes organismos
em concentrações muito baixas (nanomolares). A atividade da P H+-ATPase também
pode ser inibida por este antibiótico em concentrações mil vezes maiores (Bowman
& Bowman., 1988).
A concanamicina A, outro inibidor das V H+-ATPases, é mais efetivo que a
Bafilomicina A1. A concentração ótima para inibição da V H+-ATPase está em torno
de um décimo da concentração de Bafilomicina A1 (Dröse & Altendorf, 1996;
Matsuoka et al., 1997). Em células vegetais a inibição ótima de V H+-ATPase por
Concanamicina A é obtida com concentrações em torno de 1 nM (Dröse & Altendorf,
1996; Matsuoka et al., 1997).
O ânion caotrópico nitrato em altas concentrações, aproximadamente 100mM
em baixa temperatura ( 0 ºC), libera cinco polipeptídeos do complexo catalítico V1
inibindo as V H+-ATPases, podendo chegar a 70% de inibição nesses casos,
contudo na maior parte das condições a inibição é reversível, ou seja depois da
retirada do agente a maior parte da atividade da enzima é restaurada (Moriyama &
Nelson, 1989a).

16
2 – OBJETIVOS
2.1 - Objetivo principal
Avaliar a regulação da V H+-ATPase por meio da análise dos efeitos de
diferentes concentrações de Ca2+ e Mn2+ sobre o transporte de H+ pelas V H+-
ATPases através de membranas de diferentes organelas da via secretória e também
o efeito de diferentes concentrações de KNO3 sobre a atividade de transporte de H+
e hidrólise de ATP das V H+-ATPases em MT de Saccharomyces cerevisiae.
2.2 - Objetivos específicos
• Verificar se os efeitos do Mn2+ e Ca2+ sobre a velocidade inicial (V0) e a
amplitude máxima (Fmax) da formação do ∆pH pelas V H+-ATPases de
membranas do RE, Golgi, vacúolo e envelope nuclear energizadas pela
glicose in vivo são seletivas ou não.
• Estudar o efeito de diferentes concentrações de KNO3 na hidrólise de ATP em
membranas totais energizadas ou não pela glicose extracelular, para saber se
a atividade ATPásica é inibida diferente por esse ânion caotrópico.
• Avaliar o efeito de diferentes concentrações de KNO3 sobre a velocidade
inicial e a amplitude máxima de formação do ∆pH pela V H+-ATPase em
membranas totais energizadas ou nao pela glicose in vivo, para saber se
essas atividades são inibidas de forma diferenciada por esse ânion.

17
3 – MATERIAL & MÉTODOS
3.1 - Cepa de levedura
Levedura Saccharomyces cerevisiae cepa “wild type” X-2180, genótipo (MATa
SUC2 mal mal gal2 CUP1). Essa cepa foi, generosamente, doada ao Prof. Ludwig
Lehle (Regensburg, Alemanha) pelo Prof. Hans Rudolph (Institut für Biochemie der
Universitat Stuttgart, Alemanha).
3.2 - Meio de cultura e manutenção da cepa
Foi utilizado o meio de cultura YEPD contendo: 1% de extrato de levedura
(Difco ou Oxoid); 2% de glicose (Vetec); 2% de bactopeptona (Difco ou Oxoid).
O meio YEPD sólido contém adicionalmente 2% de ágar (Vetec). Os meios
foram autoclavados a 0,5 atm (marcados no manômetro), 110 ºC por 30 min. O meio
sólido foi vazado em placas de Petri estéreis e levado a estufa a 37 ºC durante 24
horas para a realização do controle de esterilidade. Após este procedimento,
algumas colônias de S. cerevisiae (X-2180) foram retiradas com o auxílio de uma
alça bacteriológica, do meio YEPD sólido de estocagem e semeadas em um novo
meio YEPD sólido.
3.3 - Preparo do pré-inóculo
Algumas colônias de levedura foram retiradas do meio sólido YEPD e
colocadas em 40 mL do meio YEPD líquido, de forma que a densidade ótica (DO)
inicial do pré-inóculo no espectrofotômetro no comprimento de onda (λ) de 600 nm
ficasse em torno de 0,1. As células cresceram a 30 ºC sob agitação constante de
250 rpm até a fase estacionária, aproximadamente por 24 horas.
3.4 - Preparo da levedura para o isolamento
O volume do pré-inóculo a ser adicionado em 200 mL do meio YEPD
(Erlenmeyers de 1000) foi calculado, considerando o tempo de geração da cepa X-
2180, que é de 2 horas, de acordo com a curva de crescimento. De forma que, após
aproximadamente 17 horas a 30 ºC em agitação de 250 rpm, as células crescessem

18
até o meio da fase logarítimica. Após o crescimento celular, foi realizado o
isolamento das vesículas de membranas de S. cerevisiae.
3.5 - Isolamento de membranas e fracionamento subcelular
O isolamento foi feito de acordo com o método descrito por Okorokov & Lehle
(1998) com modificações. Todos os procedimentos foram realizados no gelo.
Soluções, tubos, rotores e centrífugas foram previamente resfriados.
3.5.1 - Obtenção de células
As células de S. cerevisiae X-2180 foram inoculadas em meio YEPD e
colocadas para crescer no shaker (250 rpm a 30 ºC) até o meio da fase logarítimica,
no caso da levedura X-2180 até aproximadamente 5 DO(600 nm/mL). A cultura de
suspensão celular foi centrifugada a 4000 x g por 6 min. a 4 ºC. Em seguida foi feito
o descarte do meio e determinado o peso úmido das células.
3.5.2 - Obtenção dos esferoplastos
Para cada 1 g de peso úmido de células foi adicionado 5 mL de tampão de
esferoplastos (Sorbitol 1,2 M, Tris 10 mM, pH 7.4), 1 mg do complexo de enzimas
líticas (liticase) e 12 µL β−mercaptoetanol (concentração final 30 mM). Essa
suspensão celular em tampão de esferoplastos foi incubada a 37 ºC sob fraca
agitação.
O monitoramento cinético da hidrólise da parede celular foi realizado
espectrofotometricamente no λ de 600 nm, misturando 10 µL dessa suspensão
celular em 990 µL H2O. Esse monitoramento foi feito a cada 5 ou 10 min a partir do
tempo 0 (zero) de incubação até o tempo máximo de 50 min ou até que a
absorbância chegasse ao valor entre 20 a 10% do valor inicial.
O tubo onde continha a suspensão de esferoplastos foi transferido para o gelo
e a reação de hidrólise da parede celular foi paralisada pela adição do Tampão de
parada (“stop solution”), nas concentrações finais de 10 mM de Tris-HCl pH 7,4 e 1
mM de EDTA (solução estoque 20 vezes concentrado: Sorbitol 1,2 M, Tris-HCl 200
mM pH 7,4, EDTA 20 mM), concentração final de Benzamidina 1 mM (200 mM
estoque) e concentração final de PMSF 1 mM (200 mM estoque).

19
A suspensão de esferoplastos (~15 mL) foi adicionada sobre 30 mL da
solução de Sorbitol 1,4M, Tris-HCl 50 mM pH 7,4 no tubo de centrífuga. Isto foi feito
com o auxílio de uma pipeta, devagar e inclinadamente evitando a mistura da
suspensão de esferoplastos com a solução colchão. Esse material foi centrifugado a
4000 x g por 6 min a 4°C. Esse passo do procedimento foi realizado com o objetivo
de eliminar os resíduos de enzimas líticas.
O sobrenadante foi cuidadosamente descartado e as paredes dos tubos
foram secas com papel filtro para evitar que possíveis enzimas líticas atuem nos
esferoplastos.
3.5.3 - Pré-incubação dos esferoplastos com glicose
Os esferoplastos foram incubados a 30 ºC por 10 min com 20 mL de solução
de incubação com glicose (glicose 100 mM, Sorbitol 1,2 M, MgSO4 3 mM, KH2SO4
10 mM, Tris-HCl 10 mM pH 7,2). Após a centrifugação os esferoplastos foram
centrifugados a 4000 x g por 6 min a 4 °C.
3.5.4 - Obtenção de membranas totais
O sedimento obtido das duas versões foram ressuspensos em 20 mL de
tampão de lise (sacarose 12,5% MOPS-KOH 20 mM pH 7,6), DTT 1 mM, 1 µg/mL,
coquetel de inibidores de proteases (solução estoque do coquetel de inibidores de
proteases é composto de quimiostatina, pepstatina, antipaina, leupeptina e
aprotinina na concentração de 1 mg/mL cada um), benzamidina 1 mM (200 mM
estoque), PMSF 1 mM (200 mM estoque).
A ressuspensão foi homogeneizada, em homogeneizador de vidro com pistilo
de teflon (“potter”), com 21 ciclos completos (“strokes”). O homogeneizado foi
centrifugado a 4000 x g por 6 min a 4 °C.
O sedimento foi desprezado e o sobrenadante (suspensão de membranas
totais) foi transferido para o tubo da ultracentífuga e centrifugada a 45000 x g por 45
min a 4 °C.

20
3.5.5 - Obtenção de membranas intracelulares fracionadas em gradiente
de densidade de sacarose
O sedimento (membranas totais) foi ressuspenso em, aproximadamente, 1,5
mL (dependendo do conteúdo de membranas) de solução de resuspensão (10 mL
do Tampão de lise, 1,17g de glicerol, 10 µL de coquetel de inibidores de proteases (1
mg/mL), 25 µL benzamidina 200 mM, 25 µL PMSF 200 mM e 4 µL DTT 500 mM.
Esse volume foi adicionado gradativamente, geralmente de 300 em 300 µL
misturando bem, até que o volume final determinado fosse atingido. A ressuspensão
de membranas totais foi colocada no homogeneizador de vidro com o pistilo de
teflon (“potter”) e homogeneizada com 7 ciclos completos (“strokes”).
Foi colocado 1,2 mL de ressuspensão de membranas totais sobre um
gradiente descontínuo e simplificado de densidade de sacarose com as
concentrações de 25, 38 e 50% (cada concentração de sacarose foi preparada
peso/peso (p/p) em tampão MOPS-NaOH 10 mM, MgCl2 1 mM, pH 7,2),
complementado com glicerol afim de que todas as bandas formadas por diferentes
concentrações de sacarose tivessem a mesma osmolariade que a de maior
concentração (50%). Essas concentrações foram utilizada afim de se obter bandas
enriquecidas com vacúolo (25%), Golgi (38%), RE/MP (50%) e sedimento
(Membranas mais densa que RE, provavelmente RE/EN/MP) foi ressuspenso em
sacarose 50%. O gradiente foi centrifugado a 140000 x g por 2 horas e 45 minutos a
4 °C. O volume de membranas totais restante foi aliquotado em tubos cônicos de 1,5
mL (“eppendorfs”), congelado em nitrogênio líquido e armazenado no freezer a -70 oC até o momento de ser analisado.
As vesículas de membranas, separadas em gradiente descontínuo e
simplificado de densidade de sacarose, foram fracionadas pela retirada das bandas
que se encontravam acima da concentração de sacarose, com o auxílio de uma
pipeta, retirando apenas as bandas enriquecidas com as membranas citadas
anteriormente. Após a distribuição das bandas em tubos cônicos (“eppendorfs”),
sendo colocado 100 µL de membrana em cada tubo, as frações de vesículas de
membranas foram congeladas em nitrogênio líquido e armazenadas no freezer a -70
°C até o momento da análise.

21
3.6 - Determinação do ∆pΗ (Η (Η (Η (transporte de H+)))) (Okorokov & Lichko, 1983)
Foi registrado a formação do ∆pΗ através das membranas das organelas da
via secretória, a partir do transporte de H+ realizado pela V H+-- ATPases ou P H+--
ATPases.
Para tal, foi utilizado tampão de incubação contendo KCl 20 mM, MgSO4 2,5
mM, solução de MOPS-KOH (sacarose 12,5 %, MOPS-KOH 20 mM, pH 7,4) em
quantidade suficiente para complementar 2,0 mL de volume final, 3 µL de ACMA 1
mM (dissolvido em etanol) e suspensão de vesículas de membranas com volume
variando entre 10 a 30 µL, dependendo da atividade apresentada, de modo que a
amplitude máxima (Fmáx ) não ultrapasse o valor de 50%.
Após três minutos de incubação a 30 oC (ou o tempo necessário para
estabilização da fluorescência) foi acrescentado ATP 1 mM pH 7,2 e foi observado a
extinção da fluorescência da cinética enzimática no espectrofluorímetro, nos λ de
excitação 415 nm e emissão em 485 nm com abertura de 5 nm/10 nm. Após um
tempo aproximado de 600 seg, ou o suficiente para atingir o aquilíbrio entre reação.
Foi usado NH4Cl para desfazer o gradiente protônico estabelecido.
Para calcularmos a densidade relativa de fluorescência utilizaremos a formula
(1) e para determinarmos a V0 utilizaremos a formula (2) (esquema 1) onde:
(1) ∆Fmáx = Feq / Fmáx * 100;
(2) V0 = [F0 / (Fmáx * T)] * 100
V0: velocidade inicial de formação do ∆pH;
F0: Fluorescência dependente de V0, num tempo “T”, determinada pela extrapolação
de uma reta tangente à maior inclinação inicial para o eixo do tempo;
Fmáx: fluorescência máxima (total);
T: tempo em minutos;
Feq: fluorescência de equilíbrio, determinado como fluorescência do platô que reflete
o equilíbrio entre o influxo e o efluxo de H+ nas vesículas.

22
Na presença de ATP a fluorescência diminui demonstrando o transporte de
prótons e a conseqüente formação do gradiente de H+ das vesículas de membranas.
Ao atingir a fluorescência máxima e o equilíbrio, adiciona-se NH4Cl, que desfaz o
gradiente protônico.
3.7 - Determinação do conteúdo de proteína (Bradford, 1976):
Transferem-se 28 mg de Comassie Brilliant Blue G 90% para um Becker,
devidamente envolvido com papel alumínio a fim de evitar a entrada de luz,
contendo 12 mL de etanol absoluto. Deixa-se sob agitação por 1 hora a temperatura
ambiente (25-30 °C). Foi adicionado 25 mL de ácido ortofosfórico (85%) e após
homogeneização completa-se o volume para 250 mL com água destilada em balão
volumétrico. Filtra-se a solução em papel de filtro por três vezes. Armazena-se em
um vidro âmbar.
Para determinação da curva padrão é utilizada uma solução de albumina de
soro bovino na concentração 1 mg/mL, armazenada a 0°C . Medem-se volumes de 2
µL a 20 µL da solução de albumina 1mg/mL, completa-se para quantidade suficiente
Esquema 1. Determinação da velocidade inicial e amplitude máxima do transporte de H+ por membranas de S. cerevisiae, cepa X-2180. Velocidade inicial (V0=[F0/(Ftotal*T)]*100); Amplitude máxima (∆Fmáx=(Feq/Ftotal)*100).

23
de 100 µL com água destilada e adiciona-se 1 mL da solução de Bradford, em
intervalos de 30 em 30 seg. Aguarda-se exatamente 10 min, à temperatura ambiente
e procede-se as leituras em espectrofotômetro no λ de 595 nm obedecendo ao
mesmo intervalo utilizado anteriormente.
Determina-se o conteúdo de proteína utilizando-se entre 5 e 15 µL de
suspensão de membranas, completando-se o volume para 100 µl com água
destilada e adicionando-se 1 mL da solução de Bradford em intervalos de 30 seg.
Aguardam-se 10 min à temperatura ambiente e procede-se as leituras em
espectofotômetro no λ de 595 nm obedecendo ao mesmo intervalo de tempo
utilizado anteriormente. Para os valores do conteúdo de proteína que ficaram fora da
faixa de linearidade, ou seja, nas extremidades da curva padrão, os seguintes
procedimentos foram realizados: (1) ou foi aumentado o conteúdo de membranas (2)
ou foram diluidas as membranas com água destilada, a fim de que essas leituras
permanecessem, preferencialmente, entre os valores médios na curva padrão.
3.8 - Determinação da hidrólise de ATP
A atividade ATPásica foi medida pelo aumento da quantidade de fosfato
inorgânico (Pi) resultante da hidrólise de ATP.
O preparo dos ensaios de hidrólise de ATP (ATPase total) foi feita sob
refrigeração (gelo). Nos tubos de ensaio foram colocados 10 µL de suspensão de
vesículas de membranas e 230 µL da solução D (MES-KOH 30 mM, MgSO4 3,75
mM, molibdato de amônio 262 µM pH 6,5 – para obter as concentrações finais de
MES 23 mM, MgSO4 2,1 mM e molibdato de amônio 192 µM), 10 µL de ATP-NaOH
50 mM pH 7,2 (ATP 1,62 mM concentração final) e 50 µL de água destilada
necessária para completar 300 µL. O meio de reação foi incubado a 30 °C por 30
min. Após o tempo de incubação os tubos foram colocados no gelo novamente, foi
completado o volume da reação de hidrólise para 1 mL com água destilada gelada,
afim de parar a reação, e adicionou-se 2 mL da solução C (mistura 100:1 da solução
A e a solução B, que deve ser preparada próximo à adição desta na reação (solução A:
0,5% molibdato de amônio, 0,5% SDS, 2% H2SO4; solução B: 10% ácido ascórbico). A
solução foi incubada por 10 min, em banho maria a 30 °C, seguida imediatamente
da leitura da absorbância no espectrofotômetro no λ de 750 nm.
24
4 – RESULTADOS
4.1 Efeito do Ca2+ sobre o transporte de H+ realizado pela V H+-ATPase
Analisamos o efeito do Ca2+ sobre a atividade das V H+-ATPases em relação a
formação do ∆pH em populações de vesículas de membranas ativadas por glicose,
derivadas das organelas da via secretória e separadas em gradiente de densidade
de sacarose (Figuras 3 e 4). Sendo que nessas análises foram usadas as
concentrações: 0,1; 0,5 e 1 mM de Ca2+.
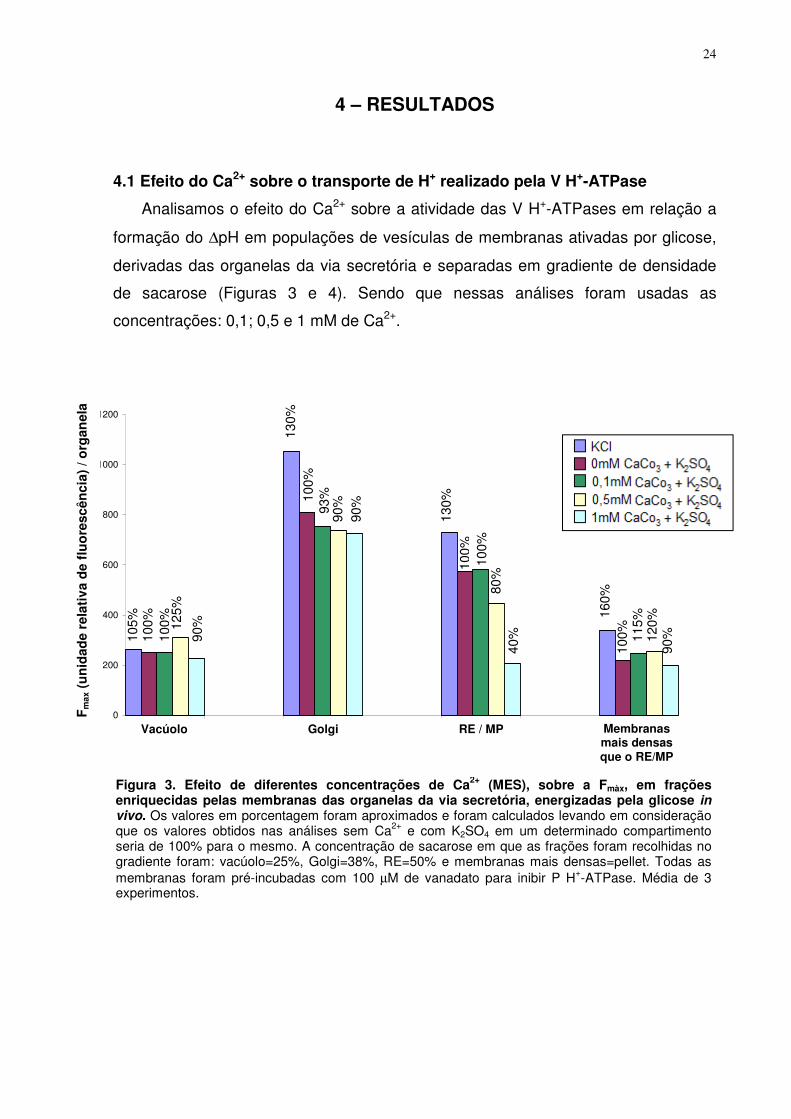
Figura 3. Efeito de diferentes concentrações de Ca2+ (MES), sobre a Fmàx, em frações enriquecidas pelas membranas das organelas da via secretória, energizadas pela glicose in vivo. Os valores em porcentagem foram aproximados e foram calculados levando em consideração que os valores obtidos nas análises sem Ca2+ e com K2SO4 em um determinado compartimento seria de 100% para o mesmo. A concentração de sacarose em que as frações foram recolhidas no gradiente foram: vacúolo=25%, Golgi=38%, RE=50% e membranas mais densas=pellet. Todas as membranas foram pré-incubadas com 100 µM de vanadato para inibir P H+-ATPase. Média de 3 experimentos.
0
200
400
600
800
1000
1200
vacúolo Golgi RE / MP membranasmais densasque o RE/MP
Fm
áx/m
g d
e p
rote
ina
KCl
0mM de Ca + K2SO4
0,1mM de Ca + K2SO4
0,5mM de Ca + K2SO4
1mM de Ca + K2SO4 130%
125%
40%
80
% 10
0%
120%
115%
100%
90%
130%
Fm
ax (
un
idad
e re
lati
va d
e fl
uo
resc
ênci
a) /
org
anel
a
160%
90%
93%
100%
100%
90%
100%
90
%
100%
105%
Vacúolo Golgi RE / MP Membranas mais densas que o RE/MP

25
Em relação à Vo de formação do ∆pH pelas V H+-ATPases, verificamos que
em todos os compartimentos ela foi inibida de forma dependente ao aumento da
concentração de Ca2+. A diminuição da V0 em nosso experimento foi de
aproximadamente: 1,7 vezes para as vesículas de membranas vacuolares, 2 vezes
para o Golgi, 50 vezes para o RE e de 1,4 vezes para vesículas de membranas mais
densas, sendo esses resultados encontrados na presença de 1 mM de Ca2+. Esse
efeito é observado também quando a concentração de Ca2+ foi 0,5 mM e em ambas
concentrações ele foi mais evidente em membranas do RE (tabela 1). Sendo que no
vacúolo e no Golgi por 0,5 mM foi próxima a obtida com 1 mM de Ca2+ (tabela 1).
Figura 4. Efeito de diferentes concentrações de Ca2+ (MES), na Vo de formação do ∆∆∆∆pH, em frações enriquecidas pelas membranas das organelas da via secretória, energizadas pela glicose in vivo. Os valores em porcentagem foram aproximados e foram calculados levando em consideração que os valores obtidos nas análises sem Ca2+ e com K2SO4 em um determinado compartimento seria de 100% para o mesmo. A concentração de sacarose em que as frações foram recolhidas no gradiente foram: vacúolo=25%, Golgi=38%, RE=50% e membranas mais densas=pellet. Todas as membranas foram pré-incubadas com 100 µM de vanadato para inibir P H+-ATPase. Média de 3 experimentos.
0
200
400
600
800
1000
1200
vacúolo Golgi RE / MP membranasmais densasque o RE/MP
Vo
/mg
de
pro
tein
a
KCl
0mM de Ca + K2SO4
0,1mM de Ca + K2SO4
0,5mM de Ca + K2SO4
1mM de Ca + K2SO4
290%
50%
150%
70%
60%
100%
55%
55%
10%
100%
90
%
155%
70%
70%
150%
100%
Vacúolo Golgi RE / MP Membranas mais densas que o RE/MP
100%
100%
2%
Vo (
∆∆ ∆∆p
H /
min
.)
100%
26
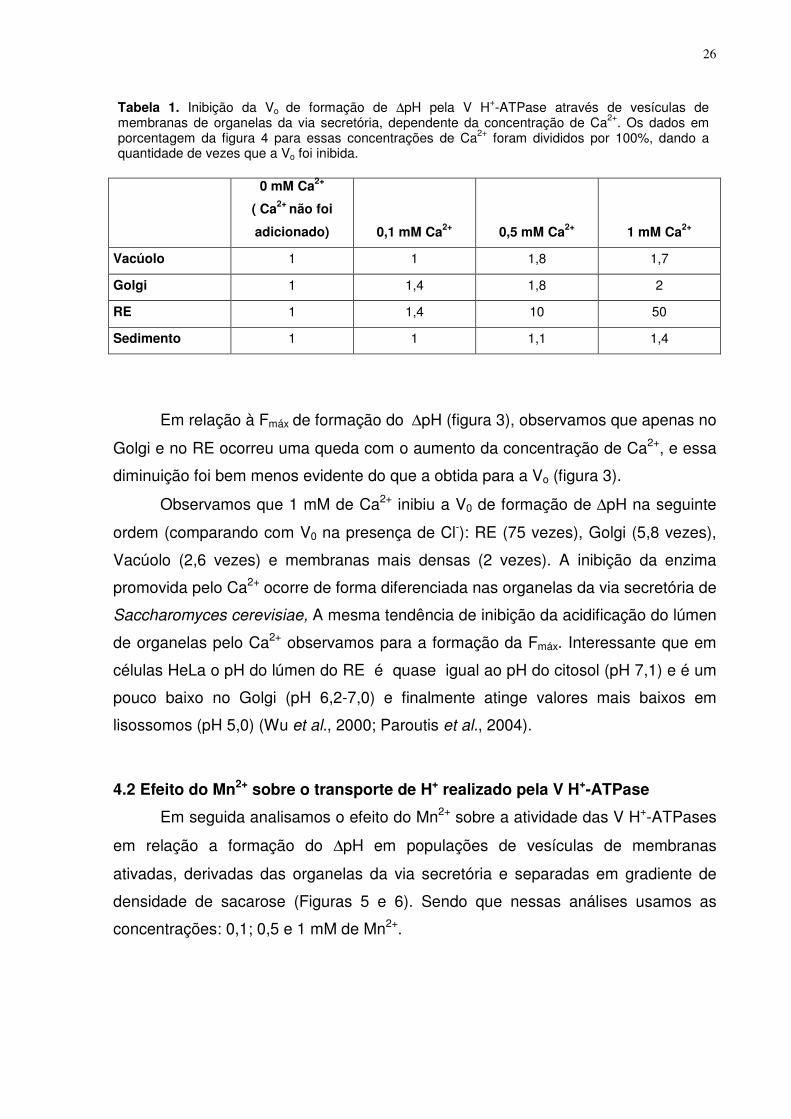
Em relação à Fmáx de formação do ∆pH (figura 3), observamos que apenas no
Golgi e no RE ocorreu uma queda com o aumento da concentração de Ca2+, e essa
diminuição foi bem menos evidente do que a obtida para a Vo (figura 3).
Observamos que 1 mM de Ca2+ inibiu a V0 de formação de ∆pH na seguinte
ordem (comparando com V0 na presença de Cl-): RE (75 vezes), Golgi (5,8 vezes),
Vacúolo (2,6 vezes) e membranas mais densas (2 vezes). A inibição da enzima
promovida pelo Ca2+ ocorre de forma diferenciada nas organelas da via secretória de
Saccharomyces cerevisiae, A mesma tendência de inibição da acidificação do lúmen
de organelas pelo Ca2+ observamos para a formação da Fmáx. Interessante que em
células HeLa o pH do lúmen do RE é quase igual ao pH do citosol (pH 7,1) e é um
pouco baixo no Golgi (pH 6,2-7,0) e finalmente atinge valores mais baixos em
lisossomos (pH 5,0) (Wu et al., 2000; Paroutis et al., 2004).
4.2 Efeito do Mn2+ sobre o transporte de H+ realizado pela V H+-ATPase
Em seguida analisamos o efeito do Mn2+ sobre a atividade das V H+-ATPases
em relação a formação do ∆pH em populações de vesículas de membranas
ativadas, derivadas das organelas da via secretória e separadas em gradiente de
densidade de sacarose (Figuras 5 e 6). Sendo que nessas análises usamos as
concentrações: 0,1; 0,5 e 1 mM de Mn2+.
0 mM Ca2+
( Ca2+ não foi
adicionado) 0,1 mM Ca2+ 0,5 mM Ca2+ 1 mM Ca2+
Vacúolo 1 1 1,8 1,7
Golgi 1 1,4 1,8 2
RE 1 1,4 10 50
Sedimento 1 1 1,1 1,4
Tabela 1. Inibição da Vo de formação de ∆pH pela V H+-ATPase através de vesículas de membranas de organelas da via secretória, dependente da concentração de Ca2+. Os dados em porcentagem da figura 4 para essas concentrações de Ca2+ foram divididos por 100%, dando a quantidade de vezes que a Vo foi inibida.

27
0
200
400
600
800
1000
1200
1400
vacúolo Golgi RE / MP membranasmais densasque o RE/MP
Fm
áx/m
g d
e p
rote
ina
KCl0mM de Mn + K2SO40,1mM de Mn + K2SO40,5mM de Mn + K2SO41mM de Mn + K2SO4
Fm
ax (
un
idad
e re
lati
va d
e fl
uo
resc
ênci
a) /
org
anel
a
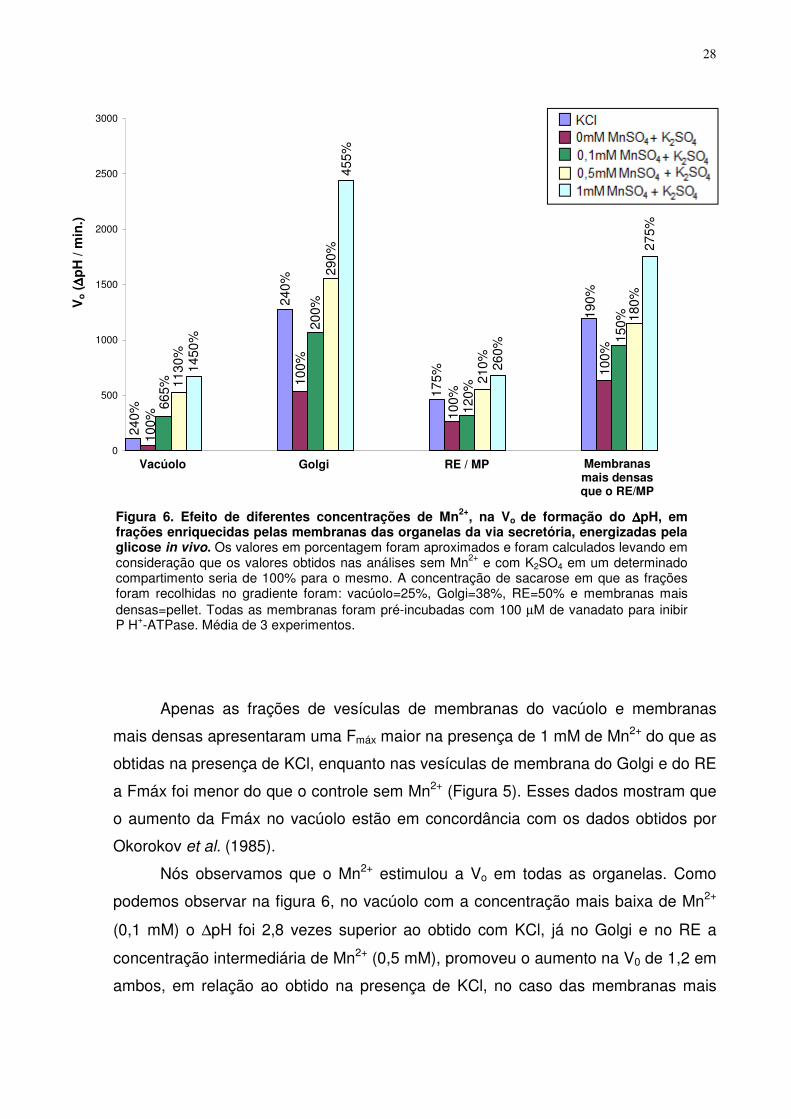
Figura 5. Efeito de diferentes concentrações de Mn2+, sobre a Fmàx, em frações enriquecidas pelas membranas das organelas da via secretória, energizadas pela glicose in vivo. Os valores em porcentagem foram aproximados e foram calculados levando em consideração que os valores obtidos nas análises sem Mn2+ e com K2SO4 em um determinado compartimento seria de 100% para o mesmo. A concentração de sacarose em que as frações foram recolhidas no gradiente foram: vacúolo=25%, Golgi=38%, RE=50% e membranas mais densas=pellet. Todas as membranas foram pré-incubadas com 100 µM de vanadato para inibir P H+-ATPase. Média de 3 experimentos.
100%
11
5%
150%
130%
10
0%
80%
80%
100%
65
%
70%
100%
80
% 10
0%
Vacúolo Golgi RE / MP Membranas mais densas que o RE/MP
115%
150%
75%
130%
60
%
120%
120%
28
Apenas as frações de vesículas de membranas do vacúolo e membranas
mais densas apresentaram uma Fmáx maior na presença de 1 mM de Mn2+ do que as
obtidas na presença de KCl, enquanto nas vesículas de membrana do Golgi e do RE
a Fmáx foi menor do que o controle sem Mn2+ (Figura 5). Esses dados mostram que
o aumento da Fmáx no vacúolo estão em concordância com os dados obtidos por
Okorokov et al. (1985).
Nós observamos que o Mn2+ estimulou a Vo em todas as organelas. Como
podemos observar na figura 6, no vacúolo com a concentração mais baixa de Mn2+
(0,1 mM) o ∆pH foi 2,8 vezes superior ao obtido com KCl, já no Golgi e no RE a
concentração intermediária de Mn2+ (0,5 mM), promoveu o aumento na V0 de 1,2 em
ambos, em relação ao obtido na presença de KCl, no caso das membranas mais
0
500
1000
1500
2000
2500
3000
vacúolo Golgi RE / MP Membranasmais densasque o RE/MP
Vo
/mg
de
pro
tein
a
KCl0mM de MnSO4 + K2SO40,1mM de MnSO4 + K2SO40,5mM de MnSO4 + K2SO41mM de MnSO4 + K2SO4
Figura 6. Efeito de diferentes concentrações de Mn2+, na Vo de formação do ∆∆∆∆pH, em frações enriquecidas pelas membranas das organelas da via secretória, energizadas pela glicose in vivo. Os valores em porcentagem foram aproximados e foram calculados levando em consideração que os valores obtidos nas análises sem Mn2+ e com K2SO4 em um determinado compartimento seria de 100% para o mesmo. A concentração de sacarose em que as frações foram recolhidas no gradiente foram: vacúolo=25%, Golgi=38%, RE=50% e membranas mais densas=pellet. Todas as membranas foram pré-incubadas com 100 µM de vanadato para inibir P H+-ATPase. Média de 3 experimentos.
100%
1450
%
240%
290%
100%
260%
150%
27
5%
120%
210%
175%
Vo (
∆∆ ∆∆p
H /
min
.)
200%
Golgi RE / MP Membranas mais densas que o RE/MP
455%
180%
Vacúolo
665%
11
30%
100%
240%
190%
100%

29
densas, a concentração intermediária de Mn2+ não estimulou a V0 em comparação
com a obtida na presença de KCl.
Assim como o observado para o Ca2+ o Mn2+ também apresenta efeito
diferenciado nas diferentes organelas da via secretória. Fortalecendo a idéia da
presença de diferentes formas da V H+-ATPases nas diferentes organelas da via
secretória.
A estimulação da Vo foi mais efetiva em membranas do vacúolo,
apresentando um aumento de 14,5 vezes quando adicionado 1 mM de Mn2+, a
segunda maior estimulação foi no Golgi. É interessante notar que o RE mostrou a
menor estimulação da Vo por Mn2+, apenas 2,6 vezes, enquanto na presença de
Ca2+ ele apresentou a maior inibição (50 vezes), como descrito anteriormente.
4.3 Efeito do KNO3 sobre a atividade hidrolítica das V H+-ATPases
A V H+-ATPase pode ser inativada, pelo nitrato e outros anions caotropicos
na presença de Mg2+ e ATP em baixa temperatura (Moriyama & Nelson,
1989a).Nestas condiçoes a enzima perde as 5 polipeptideos de complexo catalitico
V1 e simultaneqamente as atividades de ATPase e transporte de H+. A dependência
desta inibição pele temperatura baixa (‘’cold inactivation’’) foi achada com
membranas e enzima isoladas de celulas animais (Moriyama & Nelson, 1989a). A
enzima de levedura tambem perde as 5 polipeptideos na presença de nitrato, Mg2+ e
0 mM Mn2+
(Mn2+ não foi
adicionado)
0,1 mM Mn2+ 0,5 mM Mn2+ 1 mM Mn2+
Vacúolo 1 6,7 11,3 14,5
Golgi 1 2 2,9 4,6
RE / MP 1 1,2 2,1 2,6
Membranas mais
densas que o RE/MP
1 1,5 1,8 2,8
Tabela 2. Estimulação da Vo, do transporte de H+ através de membranas de organelas da via secretória, de acordo com o aumento da concentração de Mn2+ extravesicular. Os dados em porcentagem da figura 10 para as devidas concentrações de Mn2+ foram divididos por 100%, dando a quantidade de vezes que a Vo foi estimulada.

30
ATP mas independente da temperatura de tratamento das membranas (Kane et al.
1989).
Para verificar se a enzima foi inativada pelo KNO3 a 0 ºC, analisamos primeiro
a hidrólise de ATP variando as concentrações de KNO3 em membranas totais
incubadas ou não com glicose usando 1,7 mM de ATP, sendo também feita a pré-
incubação no gelo por 30 min com Mg2+ (Figura 7).
Verificamos que tanto em membranas energizadas com glicose como em não
energizadas a hidrólise de ATP diminuiu de acordo com o aumento da concentração
de KNO3 e a inibição de 50% foi obtida com aproximadamente 50 mM de KNO3 para
as duas versões (Figura 7). Mas é importante destacar que não ocorreu diferença
entre a enzima ativada e não ativada pela glicose extracelular. Isto pode significar
que a integridade da estrutura da enzima que determina a atividade hidrolitica em
ambos os estados, ativada e não ativada, é muito similar.
No experimento seguinte analisamos novamente o efeito do KNO3 sobre a
hidrólise de ATP em MT incubadas ou não com glicose, sendo também feita a pré-
Figura 7. Hidrólise de ATP pela V H+-ATPase de MT. As MT foram pré-incubadas a 0 ºC por 30 min. na presença do tampão de incubação, contendo Mg2+, KCl e ATP (foi usado 1,7 mM de ATP), a hidrólise foi feita incubando 30 min. a 30 ºC). Isolamento do dia 16/06/08, onde a ativação foi: V0=4,4x, Fmáx=7x e ATPase=2,6x; 100% da atividade de V H+-ATPase correspondem a atividade inibida por 200mM de KNO3.

31
incubação no gelo por 30 min., mas aumentamos o número de concentrações de
KNO3 usadas, afim de obtermos um melhor “perfil” de ação do KNO3. Aumentamos
também a concentração de ATP para 5 mM afim de identificarmos uma melhor razão
entre o conteúdo de proteína e a concentração de ATP (Figura 8).
Na figura 8 também não notamos diferença em relação ao efeito inibitorio do
KNO3 sobre a enzima ativada ou não. A inibição de 50% da bomba foi obtida com
aproximadamente 100 mM de KNO3 para as ambas as versões.
Figura 8. Hidrólise de ATP pela V H+-ATPase de MT. As MT foram pré-incubadas a 0 ºC por 30 min. na presença do tampão de incubação, contendo Mg2+, KCl e ATP (foi usado 5 mM de ATP, a hidrólise foi feita incubando 30 min. a 30 ºC). Isolamento do dia 13/08/08, onde a ativação foi : V0=2,2x, Fmáx=1,6x e ATPase=1,1x; 100% da atividade de V H+-ATPase correspondem a atividade inibida por 200mM de KNO3.

32
Resolvemos então analisar o efeito do KNO3 sem pré-incubação no gelo nas
mesmas condições, a fim de compararmos o efeito do KNO3 a 0 ºC (Figura 9).
Na figura 9 novamente não podemos notar diferença significativa, em relação
a inibição por KNO3 entre as formas ativada e não ativada da enzima, sendo que a
inibição de 50% foi obtida com aproximadamente 50 mM para cada estado da
enzima.
Nos experimentos seguintes o tempo da hidrólise foi alterado afim de
verificamos o tempo da incubação quando o hidrolise tem a linearidade (Figura 10).
A concentração de ATP foi diminuída para 1mM sendo essa concentração igual à
utilizada nos experimentos de transporte de H+. A mudança desses dois parâmetros
foi feita para podermos comparar melhor o efeito do KNO3 no transporte com a
hidrólise.
Figura 9. Hidrólise de ATP pela V H+-ATPase de MT. As MT não foram pré-incubadas (foi usado 5 mM de ATP, a hidrólise foi feita incubando 30 min. a 30 ºC). Isolamento do dia 13/08/08, onde a ativação foi: V0=2,2x, Fmáx=1,6x e ATPase=1,1x; 100% da atividade de V H+-ATPase correspondem a atividade inibida por 200mM de KNO3.
33
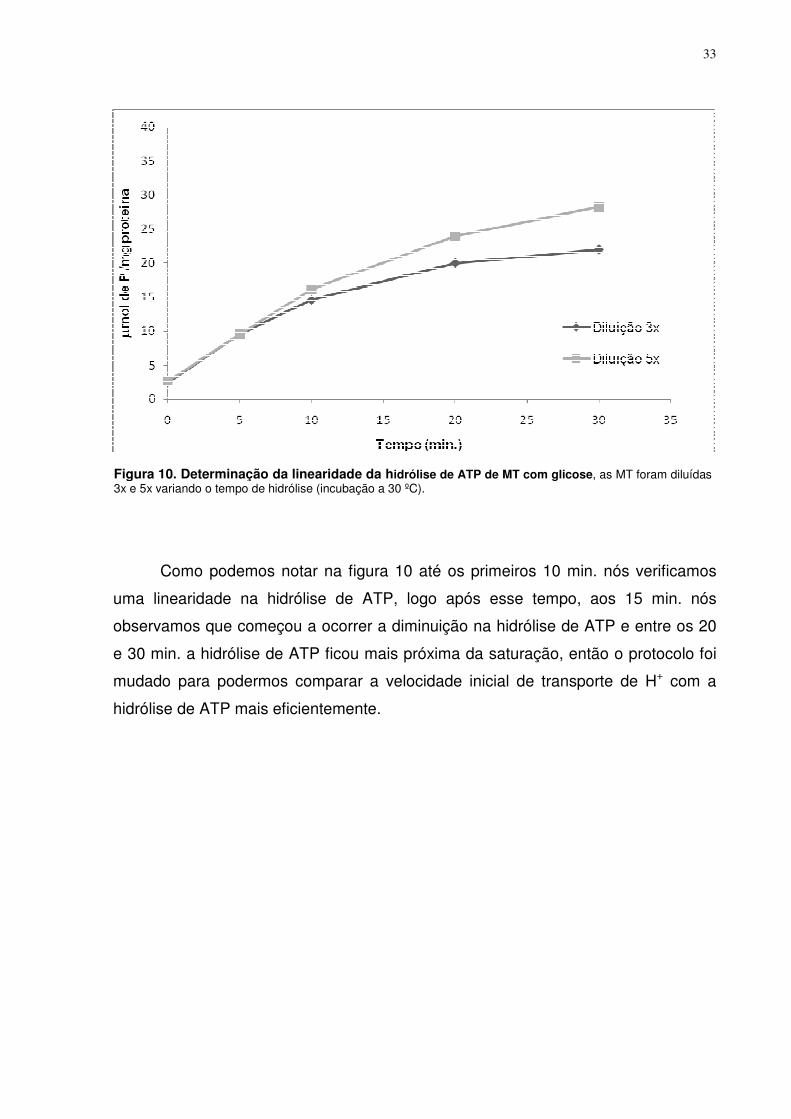
Como podemos notar na figura 10 até os primeiros 10 min. nós verificamos
uma linearidade na hidrólise de ATP, logo após esse tempo, aos 15 min. nós
observamos que começou a ocorrer a diminuição na hidrólise de ATP e entre os 20
e 30 min. a hidrólise de ATP ficou mais próxima da saturação, então o protocolo foi
mudado para podermos comparar a velocidade inicial de transporte de H+ com a
hidrólise de ATP mais eficientemente.
Figura 10. Determinação da linearidade da hidrólise de ATP de MT com glicose, as MT foram diluídas 3x e 5x variando o tempo de hidrólise (incubação a 30 ºC).

34
Analisamos então a hidrólise de ATP em MT, sendo usado 1mM de ATP sem
pré-incubação no gelo, e o tempo de hidrólise foi de 15 min. a 30 ºC (Figura 11).
Nos experimentos que deram origem a figura 11 também não ocorreu
diferença entre o efeito inibitório do KNO3, sobre as formas ativada e não ativada da
enzima e a inibição de 50% da enzima foi obtida com aproximadamente 50 mM de
KNO3.
Quando analisamos todos os nossos resultados de hidrólise de ATP podemos
notar que a inibição da bomba pelo nitrato não mostra diferença significativa entre a
enzima ativada e a não ativada, mesmo quando usamos altas concentrações de
ATP (5 mM) ou quando incubamos no gelo.
4.4 Efeito do KNO3 sobre o transporte de H+ mediado pela V H+-ATPase
Primeiramente analisamos o transporte de H+ da V H+-ATPase em
membranas totais, energizadas ou não pela glicose extracelular variando a
Figura 11. Hidrólise de ATP pela V H+-ATPase de MT, média de dois experimentos. As MT não foram pré-incubadas (foi usado 1 mM de ATP, a hidrólise foi feita incubando 15 min. a 30 ºC). Isolamento do dia 26/11/08, onde a ativação foi: V0=2,8x, Fmáx=2,4x e ATPase=1,9x; 100% da atividade de V H+-ATPase correspondem a atividade inibida por 200mM de KNO3.

35
concentração de KNO3, afim de sabermos se esse transporte de H+ pela bomba
ativada e não ativada é inibido de forma diferenciada pelo KNO3 no caso da bomba
energizada ou não pela glicose extracelular ou se a inibição do transporte de H+
ocorre de maneira similar a que achamos para ATPase.
Observamos que a Vo de transporte de H+ foi inibida pelo nitrato de maneira
diferente para os dois estados da enzima revelando a resistência mais alta da
enzima ativada (Figura 12). A inibição de 50% da Vo de formação do ∆pH pela
enzima não ativada foi obtida com aproximadamente 10 mM de KNO3 enquanto em
membranas energizadas pela glicose extracelular essa inibição foi atingida acima de
25 mM de KNO3. Observamos também que a enzima não ativada quase perdeu a
atividade de transporte de H+ na presença de 100mM de nitrato enquanto a enzima
ativada ainda apresenta aproximadamente 30% da V0 e 50% de “steady state” de
transporte de H+.
Figura 12. Efeito do KNO3 sobre a Vo de transporte de H+ mediado pela V H+-ATPase. Dados de três experimentos feitos com membranas totais energizadas ou não por glicose extracelular, as análises foram feitas na presença de 300µM de VO4
-3 para inibir a P H+- ATPase.As barras apresentam o erro padrão.

36
A Fmáx de formação do ∆pH também foi inibida com o aumento da
concentração de KNO3, mas a inibição da Fmáx foi menos efetiva do que a inibição da
Vo tanto em membranas energizadas quanto em não energizadas pela glicose. A
Fmáx de formação do ∆pH em membranas não ativadas foi inibida 50% com
aproximadamente 25 mM de KNO3 enquanto nas membranas ativadas essa inibição
foi obtida com 50 mM de KNO3 (Figura 13). Devem ser feitas novas análises de ∆pH
a fim de diminuir algumas variações obtidas na Fmáx diminuindo também a faixa de
erro do experimento.
Podemos observar que tanto a Vo quanto a Fmáx de formação do ∆pH foram
inibidas pelo KNO3 de forma menos eficiente na forma ativada da enzima,
confirmando assim o efeito da glicose sobre a enzima, deixando a mesma mais
estável. Sabendo que nitrato e um reagente caotrópico que causa a dissociação de
5 polipepitídeos do complexo V1 (Moriyama e Nelson, 1989a). Concluímos que a
enzima ativada pela glicose extracelular tem uma estrutura mais solida e sua
Figura 13. Efeito do KNO3 sobre a Fmáx de formação do ∆∆∆∆pH. Dados de três experimentos feitos com membranas totais energizadas ou não por glicose extracelular, as análises foram feitas na presença de 300µM de VO4
-3 para inibir a P H+-ATPase. As barras apresentam o erro padrão.

37
integridade necessária para transporte de H+ e acoplamento dos dois processos é
mais estável.
4.5 Efeito da glicose sobre a ativação e o acoplamento da V H+-ATPase
Comparando os resultados sobre o efeito inibitório do nitrato na hidrólise de
ATP e no transporte de H+, nós observamos que o transporte de H+ diminuiu mais
efetivamente do que a hidrólise de ATP, tanto para enzima ativada quanto para a
enzima não ativada. Isto evidencia que o nitrato desacopla os dois processos
catalizados pela bomba ou seja, o efeito que para nosso conhecimento nunca foi
revelado no caso de V H+-ATPases. Interessante, que a enzima ativada apresentou
mais alta resistência a esse desacoplamento causado pelo ânion caotrópico,
indicando que a ligação entre V1 e V0 foi mais estável para segurar o acoplamento
mais efetivo. Vale a pena ressaltar também que a ativação da V H+-ATPase pela
glicose causou não somente aumento igual da atividade de ATPase e transporte de
H+, mas aumentou também o acoplamento dos dois processos. Observamos que a
estimulação da V0 de transporte de H+ foi sempre acima da estimulação da atividade
hidrolítica da bomba. Isto significa que a glicose extracelular não só aumenta
igualmente a atividade ATPásica e de transporte de H+ mas aumenta mais
efetivamente o transporte, ou seja, estimula o acoplamento da enzima. Essa
propriedade da V H+-ATPase não pode ser explicada com uso do modelo atual de
dissociação/ re-assosiação de V1 (Kane, 1995). E mais,sabendo que o complexo V1
esta ligado com membrana e V0 não dissocia em ambos (Bernardes, 2006; Ribeiro
et al 2007) podemos concluir que essa estimulação de V0 pode ser atingida somente
por uma mudança de conformação da enzima.

38
Acoplamento 1,42 1,17 1,41
Como podemos notar na figura 14 o nitrato estimulou o acoplamento entre a
V0 de transporte de H+ e a hidrólise de ATP em membranas totais, chegando a
estimular quase 3 vezes quando usamos 75 mM de nitrato.
Estimulação Isolamento 1 Isolamento 2 Isolamento 3
ATPase 3,09 1,86 2,02
Fmáx 6,98 1,61 2,36
Vo 4,38 2,18 2,84
Tabela 3– Efeito da glicose sobre a estimulação da hidrólise de ATP, e da Fmáx e Vo de formação
do ∆pH nas membranas totais pré-incubadas com glicose de diferentes isolamentos de
membrana. No Isolamento 1 foi usado 1,7 mM de ATP e com pré-incubação no gelo na
verificação da hidrólise de ATP, nos demais foi usado 1 mM ATP e feito sem pré-incubação no
gelo. A unidade utilizada para o calculo de estimulação é vezes.
Figura 14. Efeito do nitrato na estimulação do acoplamento entre a V0 de transporte de H+ e hidrólise de ATP, sendo que a hidrólise foi feita sem pré-incubação no gelo e incubando 15 min. a 30 ºC e usado 1 mM ATP. A razão de acoplamento foi calculada fazendo: acoplamento enzima ativada/ acoplamento enzima não ativada, isso foi feito para cada concentração.

39
Outra observação interessante é que o aumento da concentração de nitrato
revelou o significante aumento da estimulação do transporte de H+, pela glicose
extracelular, mas não ocorreu mudança significativa na hidrólise de ATP (Figura 15).
Isto indica que as moléculas da V H+-ATPase ativada ainda resistentes a
desestabilização pelo nitrato tem o acoplamento ate mais comparando com as
moléculas de enzima não ativada (também resistentes) quando as suas atividades
hidrolíticas estão caindo igualmente (Fig.7-9,11). Como grau de estimulação da
hidrólise de ATP foi semelhante em todas as concentrações de nitrato, assumimos
que 1) a população de moléculas da enzima em MT não é homogênia, mas
apresentada de diferente acoplamento pelas moléculas e 2) a modulação da
eficiência do acoplamento da bomba por moléculas de glicose extracelular é o fator
chave da regulação das V H+-ATPases.
Figura 15. Ativação da V0 de transporte de H+ e da hidrólise de ATP pela glicose extracelular. A hidrólise de ATP foi realizada na presença de 1,7 mM ATP, com pré-incubação de 30 min. no gelo. Foi calculado a ativação fazendo: V0 ativada/ V0 não ativada e ATPase ativada/ ATPase não ativada, sendo isso feito para cada concentração.
Ati
vaçã
o d
a V
0 d
e tr
ansp
ort
e d
e H
+ e
hid
rólis
e d
e A
TP
(va
zes)

40
Acoplamento
(Vo / ATPase)
Isolamento 1 Isolamento 2 Isolamento 3
Não Ativada 13,4 18,1 10,4
Ativada 19 22,5 25,8
Estimulaçao 1,4 1,24 2,5 dos acoplamentos
Na figura 16 podemos notar que o aumento da concentração de nitrato
diminui o acoplamento entre a V0 de transporte de H+ e hidrólise de ATP, tanto na
enzima não ativada quanto na enzima ativada, mas a forma ativada da enzima é
mais resistente a essa diminuição no acoplamento.
Podemos notar que a estimulação do acoplamento aumenta de acordo com o
aumento da concentração de nitrato, chegando até quase três (fig. 16). Isso reforça
a idéia de que a enzima ativada é mais resistente ao efeito de dissociação do nitrato,
sendo assim mais estável.
Podemos notar que na enzima ativada o acoplamento caiu, ficou estável e
depois aumentou com o a concentração de KNO3 entre 100 e 150mM enquanto na
enzima não ativada ele diminui, ficou estável e caiu ate zero entre 100-150 mM de
nitrato ( figura 16). Importante que o acoplamento consequentemente aumentou.
Tabela 4 – Razão de acoplamento e estimulação do acoplamento pela glicose extracelular. Na hidrólise de ATP no Isolamento 1 foi usado 1,7 mM de ATP, com pré-incubação a 0 ºC e 30 min. de hidrólise, nos demais foi usado 1 mM ATP e feito sem pré-incubação a 0 ºC e o tempo de hidrólise foi de 15 min..
41
Os dados sobre a inibição de V H+-ATPase pelo nitrato levantam várias
questões, tais como: (i) qual é a origem/natureza das moleculas da enzima
mostrandos propriedades diferentes, (ii) será que o mesmo tipo de organela pode
possuir vários graus de ativação da bomba e (iii) quais são os pontos chaves para a
modulação das V H+-ATPases?
Figura 16. Efeito do nitrato na V0 de transporte de H+, acoplamento e razão de acoplamento de V H+ ATPases ativadas e não ativadas. O experimento de hidrólise foi feito na presença de 5 mM de ATP, sem pré-incubação no gelo e incubação de 30 min. a 30 ºC. A V0 está em mg/mL de proteína e o acoplamento está em vezes.

42
5 – DICUSSÃO
A inibição do trasnporte de H+ por Ca2+ em V H+-ATPases já havia sido
observado por Okorokov e colaboradores (1985) no caso de membranas de
vacúolos inteiros. O Ca2+ provavelmente pode ser considerado como um regulador
importante da acidificação do lúmen do RE. Com sua concentração elevada no
citosol e provavelmente junto com algumas proteínas moduladoras como
calmodulina, que pode diminuir a efetiva concentração de Ca2+, ele é capaz de
prevenir a acidificação do lúmen do RE. Este efeito do Ca2+ parece ser mais
significativo para o RE.
A mesma tendência de inibição da acidificação do lúmen de organelas pelo
Ca2+ observamos para a formação da Fmáx. Nossos dados mostram que
provavelmente há uma regulação diferenciada da acidificação do lúmen de
organelas da via secretória pelo Ca2+ e provavelmente pela(s) proteínas que ligam
com o Ca2+ que pode(m) prevenir essa acidificação com eficiência máxima no RE,
diminuindo então a partir do RE para o Golgi e depois para vacúolos. O efeito
diferenciado do Ca2+ nas organelas da via secretória pode nos indicar também essa
existência de formas diferentes da enzima (Samarao et al., 2009).
Em relação ao efeito do Mn2+ apenas as frações de vesículas de membranas
do vacúolo e membranas mais densas apresentaram uma Fmáx maior na presença
de 1 mM de Mn2+ do que as obtidas na presença de KCl, enquanto nas vesículas de
membrana do Golgi e do RE a Fmáx foi menor do que o controle sem Mn2+ (Figura
5). Esses dados sobre o aumento da Fmáx nas vesiculas de membranas vacúolares
estão em concordância com os dados obtidos para vacuolos inteiros por Okorokov
et al. (1985). Observamos também que o Mn2+ estimulou a Vo de transporte de H+ ,
em todas as organelas, o que também está em concordância com os dados
mostrados por Okorokov et al. (1985), onde eles observaram o aumento da Vo na
presença de Mn2+ em membranas de vacúolos inteiros.
O Ca2+ possui a propriedade de fechar o canal aniônico e esse fechamento do
canal aniônico junto com o funcionamento do trocador convertem o ∆µH+ em ∆µCa2+
(Okorokov et al., 1985), o que explica a inibição da formação do ∆pH.
Alternativamente Mn2+ abre o canal aniônico convertendo EMH+ em ∆pH (Okorokov et
al., 1985), estimulando assim a formação do ∆pH.

43
Quando analisamos todos os nossos resultados de hidrólise de ATP podemos
notar que a inibição da bomba pelo nitrato não mostra alteração significativa entre a
enzima ativada e a não ativada, mesmo quando usamos altas concentrações de
ATP (5 mM) ou quando incubamos a 0 ºC. Como podemos notar parece que em
levedura a incubação no gelo não promove a inativação da enzima, isto é contrário
ao obtido por, Moriyama & Nelson 1989a, em V H+-ATPase de células animais, mas
esta de acordo com o obtido por Kane et al. 1989, também em levedura.
Podemos observar em relação ao efeito do KNO3 sobre o transporte de H+
que tanto a Vo quanto a Fmáx de formação do ∆pH são inibidas pelo KNO3 de forma
menos eficiente nas membranas pré-incubadas com glicose, confirmando assim o
efeito da glicose sobre a enzima, deixando a mesma mais estável. Sabendo que o
nitrato e um reagente caotrópico que causa a dissociação de 5 polipepitídeos do
complexo V1, concluímos que enzima ativada pela glicose extracelular possui as
ligações entre complexos V1 e V0 mais estáveis (Figura 17).
Comparando os resultados sobre a hidrólise de ATP e o transporte de H+, nós
observamos que o transporte de H+ diminuiu mais rapidamente do que a hidrólise de
ATP, tanto para enzima ativada quanto para a enzima não ativada. Isto evidencia
que o nitrato desacopla os dois processos catalizados pela bomba ou seja, o efeito
que para nosso conhecimento nunca foi revelado no caso de V H+-ATPases.
Acreditamos que existem pelo menos duas maneiras de explicar a natureza
heterogênea das moléculas da V H+-ATPase. Primeiro é a existência de formas
Figura 17. Efeito sugerido da glicose extracelular sobre a atividade das V H+-ATPases em Saccharomyces cerevisiae.

44
distintas da enzima nas organelas via secretória (Stevens & Forgac, 1997; Li &
Kane, 2009; Nelson & Harvey, 1999, Cipriano et al., 2008; Samarao et al.2009). O
fato de que as enzimas do RE e as formas da enzima do envelope nuclear tem um
baixo acoplamento, em comparação com o Golgi e o vacúolo (Samarão et al. 2009)
em conjunto com a nossa observação de que as moléculas de enzima com baixo
acoplamento foram inibidas já com concentração de 10mM de nitrato, pode apontar
para as enzimas destas organelas como bons candidatos para explicar a notável
diminuição do transporte de H+ nessas condições. A atividade residual, portanto,
pode ser devido às bombas do vacúolo e do Golgi.
Vale pena relembrar que a explicação atual da regulação da VH+-ATPase
pela glicose extracelular considera que somente o número de moléculas da enzima
na membrana determina a atividade da V H+-ATPase. Por exemplo, % da diminuição
da atividade hidrolítica da bomba foi explicada pela dissociação da % do complexo
V0 (Kane, 1995). Portanto, este modelo admite que as moléculas da enzima não
ativada tem as propriedades idênticas com as moléculas da enzima ativada. Isto é a
atividade de hidrolise de ATP, do transporte de H+ e o acoplamento. Também a igual
resistência destas atividades ao efeito inibitório do nitrato é logicamente esperável.
Mas foi demonstrado que o complexo V1 não dissocia da membrana e do complexo
V0 (Bernardes, 2006; Ribeiro et al. 2007, 2010) indicando que a base da hipótese
atual é errada. Entretanto, nossos dados sobre a resistência igual da atividade
hidrolítica ao efeito inibitório do nitrato (Figuras 7, 8, 9 e 11) estão em concordância
com a hipótese atual. Mas os dados sobre o transporte de H+ e acoplamento da V
H+-ATPase ativada e não ativada apresentam fortes evidências sobre uma grande
diferença entre os dois estados da enzima.Todos os dados juntos permitem concluir
que a regulação da enzima pela glicose extracelular não necessita da dissociação
do complexo V1, mas precisa da modificação bioquímica da bomba que modifica\
varia sua atividade de ATPase e ainda mais sua atividade de transporte de H+
modulando em fim o acoplamento. As variações do acoplamento da V H+ -ATPase
já foram registradas (Davis et al., 1994; Yabe et al.,1999; Kettner et al., 2003), mas
não foi conhecido que estas variações podem ser chaves para a regulação da
enzima pela glicose in vivo.

45
6 – CONCLUSÕES
1 - Ca2+ e Mn2+ mostraram efeitos contrários sobre a velocidade inicial de transporte
de H+ nas membranas das organelas da via secretória, ou seja, 1mM de Ca2+ inibiu
enquanto 1mM de Mn2+ estimulou a velocidade inicial de transporte de H+. Ambos os
cátions apresentam efeito diferenciado em diferentes organelas da via secretória
sobre o transporte de H+. O Ca2+ inibiu o transporte de H+ até 50 vezes em
membranas do RE, quando a inibição em vacúolos, Golgi e envelope nuclear variou
entre 1,4 e 2,2 vezes. O Mn2+ estimulou a velocidade inicial de transporte de H+ em
membranas do RE, envelope nuclear, Golgi e vacúolos até 2,6; 2,8; 4,5 e 14,5 vezes
respectivamente. Isso sugere a existência de diferentes formas de V H+-ATPase
e/ou de transportadores de Ca2+ e Mn2+ nestas organelas da via secretória.
2 - O nitrato, um ânion caotrópico que libera cinco polipeptídeos do complexo
cataítico V1 e inibe a atividade de hidrolise de ATP pelas V H+-ATPases, inibiu este
processo igualmente pela forma ativada e não ativada das V H+-ATPases de
levedura. Este fato revela que a força de ligação do complexo V1 com membrana é
idêntica em ambas formas da enzima .
3 - A velocidade inicial de transporte de H+ é inibida mais efetivamente em
comparação com a hidrolise de ATP independente do estado de energização da
bomba, revelando que o nitrato desacopla estes processos catalisados pela V H+-
ATPase.
4 - A glicose extracelular estimula a velocidade inicial de transporte de H+ mais
efetivamente que o hidrolise de ATP e portanto aumenta o acoplamento da bomba
ativada em comparação com a bomba semi-ativa. O acoplamento mais alto da V H+-
ATPase em resultado de sua ativação pela glicose in vivo não foi conhecido
anteriormente.
5 - A população de enzimas de membranas totais não é homogenia e é composta de
sub-populações de moléculas que possuem diferente acoplamento. Sub-populações
de moléculas da bomba com acoplamento máximo são mais resistentes ao
desacoplamento causado pelo nitrato, mostrando que a estrutura de suas moléculas
é mais solida em comparação com aquelas moléculas que apresentam um fraco
acoplamento.

46
6 – Concluímos que a modulação do acoplamento é um novo fator e chave da
regulação da V H+-ATPase de levedura pela glicose extracelular. Essa modulação
do acoplamento da bomba explica como ela está regulada em condições quando o
complexo catalítico V1 não dissocia da membrana (Bernandes, 2006; Ribeiro et al.
2007, 2010).

47
7 – REFERÊNCIAS BIBLIOGRÁFICAS
Adachi, I., Arai, H, Pimentel, R., Forgac, M. (1990). Proteolysis and orientation on
reconstitution of the coated vesicle próton pump. J. Biol. Chem. 265: 960-966.
Alberts, B., Bray, D., Johnson, A., Lewis, J., Raff, M., Roberts, K. & Walter, P. (1999).
Fundamentos da Biologia Celular: Uma introdução à biologia molecular da célula. 1
ed. Artmed, 1999.
Alves, O. T. (2006). Estudo dos transportadores de H+ e Ca2+ em vesículas
secretórias do mutante sec6-4 da levedura Saccharomyces cerevisiae. Monografia
apresentada ao Centro de Biociências e Biotecnologia, para obtenção do título de
biólogo, na Universidade Estadual do Norte Fluminense Darcy Ribeiro, Campos dos
Goytacases, Rio de Janeiro.
Arai, H., Berne, M., Terres, G., Terres, H., Puopolo, K., Forgac, M. (1987). Subunit
composition and ATP site labeling of the coated vesicle próton-translocating
adenosinetrisphosphatase. Biochem. 26(21):6632-8.
Barkla, B.J., Pantoja, O. (1996). Physiology of ion transport across the tonoplast of
higher plants Annu. Rev. Plant Phisiol. Plant Mol. Biol. 47:159-184.
Bauerle, C., HO, M. N., Lindorfer, M. A., Stevens, T. H. (1993). The Sccharomyces
cerevisiae VMA6 gene encodes the 36-KDa subunit of the vacuolar H+-ATPase
membrane sector. J. Biol. Chem. 268: 12749-12757.
Bernardes, N. R. (2006). “A nova hipótese de regulação da V H+-ATPase da via
secretória de levedura pela glicose extracelular.” Monografia apresentada ao Centro
de Biociências e Biotecnologia, para obtenção do título de biólogo, na Universidade
Estadual do Norte Fluminense Darcy Ribeiro, Campos dos Goytacases, Rio de
Janeiro.
Beyenbach, K.W & Wieczorek, H. (2006). The V-type H+-ATPase: molecular
structure and function, physiological roles and regulation. J. Experimental Biology
209: 577-589. (Review).

48
Bond, S. and Forgac, M. (2008). The Ras/cAMP/protein kinase A pathway regulates
glucose-dependent assembly of the vacuolar (H+)-ATPase in yeast. J.Biol.Chem.
283: 36513-21.
Bowman, E. J. (1983). Comparison of the vacuolar membrane ATPase of
Neurospora crassa with the mitochondrial and plasma-membrane ATPases J. Biol.
Chem., 258 (24): 5238-5244.
Bowman, E. J., Mandala, S., Taiz, L. And Bowman, B. J. (1986). Structural studies of
the vacuolar membrane ATPase from Neurospora crassa and comparison with the
tonoplast membrane ATPase from Zea mays. Proc. Natl. Acad. Sci. USA 83, 48-52.
Bowman, B.J., Bowman, E.J. (1988). Purification of vacuolar membranes,
mitochondria, and plasma membranes from Neurospora crassa and modes of
discriminating among the different H+-ATPases. Methods Enzymol., 157:562-73.
Bowman, E. J., Siebers, A. and Altendorf, K. (1988). Bafilomycins: a class of
inhibitors of membrane ATPases from microorganisms, animal cells, and plant cells.
Proc. Natl. Acad. Sci. USA 85, 7972-7976.
Bowman B. J., Dschida, W. J., Harris, T., Bowman, E. J. (1989). The vacuolar
ATPase of Neurospora crassa contains an F1-like structure. J. Biol. Chem., 264:
15606-15612.
Bowman, B.J., Bowman, E.J. (2002). Mutations in subunit C of the vacuolar ATPase
confer resistance to bafilomycin and identify a conserved antibiotic binding site. J Biol
Chem. 277(6): 3965-72.
Boyer, P.D. (1997). The ATP synthase-a splendid molecular machine. Annu Rev.
Biochem. 67: 717-749.
Bradford, M. M. (1976). A rapid and sensitive method for the quantization of
microgram quantitaties of protein utilizing the principle of dye binding. Analyt.
Biochem., 72: 248-254.
Breton, S., Brown, D. (2006). New insights into the regulation of V-ATPase-
dependent proton secretion. J Physiol Renal Physiol 292: 1–10.

49
Breton, S., Smith, P., Lui, B., Brown, D. (1996). Acidification of the male reproductive
tract by a proton pumping (H+)-ATPase. Natl. Med. 2: 470-473.
Chattopadhyay S., Muzaffar N. E., Sherman F. and Pearce D. A. (2000). The yeast
model for Batten Disease: Mutations in btn1, btn2 and hsp30 Alter pH Homeostasis.
J. of Bacteriology 182: 6418-6423.
Chen SH, Bubb MR, Yarmola EG, Zuo J, Jiang J, et al. (2004). Vacuolar H+-ATPase
binding to microfilaments: regulation in response to phosphatidylinositol 3-kinase
activity and detailed characterization of the actin-binding site in subunit B. J. Biol.
Chem. 279: 7988–7998.
Cipriano, D. J., Wang, Y., Bond, S., Hinton, A., Jefferies, KC., Qi, J. and Forgac, M.
(2008). Structure and Regulation of the Vacuolar ATPases, Biochim Biophys Acta.
1777: 599–604.
Clague, M., J., Urbe, S., Aniento, F., Gruenberg, J. (1994). Vacuolar ATPase activity
is required for endossomal carrier vesicle formation. J. Biol. Chem. 269: 21-24.
Davis, J. M., Hunt, I. and Sanders, D. (1994). Vacuolar H+-pumping ATPase
variable transport coupling ratio controlled by pH, Proc. Natl. Acad. Sci. USA 91:
8547-8551.
Dietz, K. J., Tavakoli, N., Kluge, C., Mimura, T., Sharma, S. S., Harris, G. C.,
Chardonnens, A. N., Golldack, D. (2001). Significance of the V-type ATPase for the
adaptation to stressful growth conditions and its regulation on the molecular and
biochemical level. J. Exp. Bot., 52: 1969-1980.
Dixon N, Pali T, Kee TP, Ball S, Harrison MA, et al. (2008) Interaction of spinlabeled
inhibitors of the vacuolar H+-ATPase with the transmembrane Vosector. Biophys. J.
94: 506–514.
Dixon N, Pali T, Kee TP, Marsh D (2004) Spin-labelled vacuolar-ATPase inhibitors in
lipid membranes. Biochim. Biophys. Acta 1665: 177–183.

50
Dröse, S., Bindseil, K.U., Bowman, E.J., Siebers, A.Zeeck,A., Altendorf, K.(1993).
Inhibitor effect of modified bafilomycins and concanamyains on P and V type
adenosinetrisphosphatases. Biochem. 32: 3902-3906.
Dröse, S., Altendorf, K., (1996). Bafilomycins and concanamyains as inhibitoris of V-
ATPases and P-ATPases. J.Exp. Biol. 200: 1-8.
Duby, G. and Boutry, M. (2009). The plant plasma membrane proton pump ATPase:
a highly regulated P-type ATPase with multiple physiological roles,Pflugers Arch –
Eur. J. Physiol. 457: 645–655.
Eraso, P., Mazón M. J. and Portillo, F. (2006). Yeast protein kinase Ptk2 localizes at
the plasma membrane and phosphorylates in vitro the C-terminal peptide of the H+-
ATPase. Biochi.Biophys.Acta-Biomembranes.1758: 164-170.
Finbow, M.E., Harrison, M.A. (1997). The vacuolar H+-ATPase: a universal proton
pump of eukaryotes. Biochem. J. 324: 697-712.
Forgac, M. (1998). Struture, function and regulation of the vacuolar (H+) ATPase.
FEBES Lett. 440 (3): 258-63.
Forgac, M. (1999a). Structure and properties of the vacuolar H+-ATPase. J. Ext.
Chem. 274(19): 12951-12954.
Forgac, M. (1999b). Struture, mechanism and regulation of the chaltrin-coated
vesicle na yeast vacuolar H+-ATPase. J. Exp. Biol. 203: 71-80.
Forgac M (2007). Vacuolar ATPases: rotary proton pumps in physiology and
pathophysiology. Nat. Rev. Mol. Cell Biol. 8: 917–929.
Futai, M., Noumi, T., Maeda, M. (1989) ATP synthase (H+-ATPase): Results by
combined biochemical and molecular biological approaches. Annu Rev. Biochem 58:
111-136.
Futai, M., Oka, T., Sun-Wada, G. H., Moriyama, Y., Kanazawa, H., Wada, Y. (2000).
Luminal acidification of diverse organelles by V H+-ATPase in animal cells J. Exp.
Biol. 203, 107-116.

51
Frattini A, Orchard PJ, Sobacchi C, Giliani S, Abinun M, Mattsson JP, Keeling D. J.,
Andersson AK, Wallbrandt P, Zecca L, Notarangelo LD, Vezzoni P, Villa A. (2000).
Defects in TCIRG1 subunit of the vacuolar proton pump are responsible for a subset
of human autosomal recessive osteopetrosis. Nat. Genet. 25: 343–346.
Gluck, S. L. (1992). The structure and biochemistry of the vacuolar H+-ATPase in
proximal and distal urinary acidification. J. Bioenerg. Biomembr. 24: 231-360.
Gluck, S. L., Underhill, D. M., Lyori, M., Holliday, M., Kostrominova, T. Y., Lee, B. S.,
(1996). Physiology and Biochemistry of the kidney vacuolar H+-ATPase. Ann. Rev.
Physiol.58: 427-445.
Graham L. A., Hill, K. J., Stevens, T. H. (1994). VMA7 encodes a novel 14-KDa
subunit of the Sacharomyces cerevisiae vacuolar H+-ATPase complex. J. Biol. Chem.
269: 25974-25977.
Graham L.A., Powel B, Stevens T.H. (2000). Composition and assembly of the yeast
vacuolar H+-ATPase complex. J. Exp. Biol. 203: 61-70.
Harvey, W.R., (1992). The physiology of V-ATPases. J. Exp. Biol. 172, 1-17.
Holliday, L. S., Lu, M. Leei, B.S., Nelson, R.D., Solivan, S., Zhang, L. and Gluck, S.
L. (2000) The Amino-terminal Domain of the B Subunit of Vacuolar H1-ATPase
Contains a Filamentous Actin Binding Site. J.Biol.Chem. 275: 32331–32337.
Jones, P. C., Fillingame, R. H. (1998). Genetic fusions of subunit c in the F-0 sector
of H+-transporting ATP synthase – functional dimers and trimers and determination of
stoichiometry by cross-linking analysis. J. Biol. Chem. 273 (45): 29701-29705.
Kakinuma, Y., Ohsumi, Y., Anraku (1981). Properties of H+-translocating adenosine
triphsphate in vacuolar membranes of Saccharomyces cerevisiae. J. Biol. Chem.
256: 10859-10863.
Kane, P., Yamashiro, C., Stevens, T. (1989). Biochemical Characterizationo of the
Yeast Vacuolar H+-ATPase. J. Biol. Chem. 264: 19236-19244.
Kane, P. (1992). Biogenesis of the yeast vacuolar H+-ATPase. J. Exp. Biol., 172: 93-
103.

52
Kane, P., (1995). Disassembly and reassembly of the yeast vacuolar H-ATPase in
vivo, J. Biol. Chem. 270: 17025-17032.
Kawasaki-Nishi S, Bowers K, Nishi T, Forgac M, Stevens TH (2001) The amino-
terminal domain of the vacuolar proton-translocating ATPase a subunit controls
targeting and in vivo dissociation, and the carboxyl-terminal domain affects coupling
of proton transport and ATP hydrolysis. J. Biol. Chem. 276: 47411–47420.
Kettner, A.C., Bertl, G., Obermeyer, Slayman, C., Bihler H. (2003). Electrophysiolo-
gical analysis of the yeast V-type proton pump: Variable coupling ratio and proton
shunt. Bioph. J. 85: 3730–373.
Kevin C. Jefferies1, Daniel J. Cipriano1, and Michael Forgac (2008). Function,
structure and regulation of the vacuolar (H+)-ATPases. Arch Biochem Biophys.
476(1): 33–42.
Klionsky Et Al., Klionsky, D. J., Herman, P. K., Emr, S. D. (1990). The fungal vacuole:
composition, function and biogenesis. Microbiol. Ver. 54: 266-292
Lecchi, S., Allen, K. E., Pardo, J. P., Mason, A. B. and Slayman, C.W. (2005).
Conformational Changes of Yeast Plasma Membrane H+-ATPase during Activation
by Glucose: Role of Threonine-912 in the Carboxy-Terminal Tail, Biochemistry, 44:
16624–16632.
Lecchi, S., Nelson, C.J., Allen, K.E., Swaney, D.L., Thompson, K.L., Coon, J.J.,
Sussman, M.R., Slayman, C.W. (2007). Tandem phosphorylation of Ser-911 and Thr-
912 at the C terminus of yeast plasma membrane H+-ATPase leads to glucose-
dependent activation. J Biol Chem. 282: 35471-35481.
Li, S. C. and Kane, P.M. (2009). The yeast lysosome-like vacuole: Endpoint and
crossroads. Biochim.Biophys.Acta.1793: 650-663.
Maeshima, M. (2001). Tonoplast transporters: Organization and function. Annu. Rev.
Plant. Physiol. Plant. Mol. Biol. 52: 469-497.

53
Matsuoka, K., Higushi, T., Maeshima, M., Nakamura, K. (1997). A vacuolar-type H+-
ATPase in a nonvacuolar organelle is required for tha sorting of soluble vacuolar
protein precursors in tobacco cells. Plant Cell. 9: 533-546.
Merzendorfer, H., Gräf. R., Huss. M., Harvey. W.R., Wieczorek. H. (1997).
Regulation of proton-translocating V-ATPases. The journal of Experimental Biology.
200,225-235.
Monteiro, R. M. (2008). Efeito dos íons ca2+ e mn2+ sobre a atividade das V H+-
ATPases e no gradiente de h+ nas organelas da via secretória de Saccharomyces
cerevisiae Monografia apresentada ao Centro de Biociências e Biotecnologia, para
obtenção do grau de Bacharel em Ciências Biológicas, na Universidade Estadual do
Norte Fluminense Darcy Ribeiro, Campos dos Goytacases, Rio de Janeiro.
Moriyama, Y And Nelson, N. (1987). The purified ATPases from chromaffin granule
membranes is an anion-dependent protn pump. J. Biol. Chem. 262: 9175-9180.
Moriyama, Y. And Nelson, N. (1989a). Cold Inactivation of Vacuolar Proton-ATPases.
J. Biol. Chem. 264: 3577-3582.
Moriyama, Y. And Nelson, N. (1989b). H+-translocating ATPase in Golgi-apparatus-
charaterisation as vacuolar H+-ATPase and its subunit structures. J. Biol. Chem. 264:
18445-18450.
Moriyama, Y., Hayshi, M., Yatsushiro, S., Yamamoto, A. (2003). Vacuolar proton
pumps in malaria parasite cells. J. Bioenerg. Biomenbr. 35: 367-375.
Mulberg, A. E., Tulk, B. M., Forgac, M. (1991). Modulation of coated vesicle chloride
channel activity and acidification by reversible protein kinase A-dependent
phosphorilation. J. Biol. Chem. 266: 20590-20593.
Nelson, H., Mandiyan, S., Noumi, T., Moriyama, Y., Miedel, M.C., Nelson, N. (1990).
Molecular cloning of cDNA enconding the C subunit of H(+)-ATPase from bovine
chromaffin granules. J. Biol Chem. 265(33): 20390-20393.
Nelson N. (1992). The vacuolar H+-ATPases- one of the most fundamental ion
pumps in nature. J. Exp. Biol. 172: 19-27.

54
Nelson. N., Harvey, W.R. (1999). Vacuolar and plasma membrane
protonadenosinetriphosphatases. Physiol Rev.: 79(2): 361-289.
Nelson, N., Perzov, N.,Cohen, A. (2000). The cellular biology of proton-motive force
generation by V-ATPase. J. Exp. Biol. 203 (1): 89-95.
Nelson N (2003) A journey from mammals to yeast with vacuolar H+-ATPase (V-
ATPase). J. Bioenerg. Biomembr. 35: 281–289.
Nicolay, K., Schefers, W.A., Bruinenberg, P.M. and Kaptein, R. (1982). Phosphorus –
P31 nuclear magnetic resonance studies of intracellular pH,phosphate
compartmentation and phosphate transport in yeast. Arch. Microbiol 133: 83-89.
Nishi, T., Forgac, M. (2002). The vacuolar (H+)-ATPases: nature most versatile
proton pumps. Nature Reviews Molecular Cell Biol 3: 94-103.
Oberbeck, K., Drucker, M., Robinson, D. G. (1994). V-ATPase and pyrophosphatase
in endomembranes of maize roots. J. Exp. Bot. 45: 235-244.
Ohsumi, Y., Anraku, Y., (1981). Active-transport of basic-amino-acids driven by a
proton motive force in vacuolar membrane-vesicles of Saccharomyces cerevisiae. J.
Biol Chem 256: 2079-2082.
Ohsumi, Y., Anraku, Y., (1983). Calcium transport driven by a proton motive force en
vacuolar membrane vesicles of S. cerevisiae. J. Biol. Chem. 258: 5614-5617
Okorokov, L. A., Lichko, L. P., Kulaev, I. S. (1980). Vacuoles: main compartments of
potassium, magnesim and phosfate ions in Saccharomyces carsbergensis cells. J.
Bacteriol. 144(2): 661-665.
Okorokov, L. A. And Lichko, L. P.(1983). The identification of a proton pump on
vacuoles of the yeast Saccharomyces carsbergensis; ATPases is electrogenic H+
translocase. FEBS Letters. 155: 102-106.
Okorokov, L. A., Kulakovskaya T. V., Lichko, L. P. (1985), Biochemistry & function of
vacuolar adenosine-triphosphatase in fungi & plants springer verlag.

55
Okorokov, L.A., (1997). Diversity of Ca2+-transporters and Ca2+ store compartments
in yeast: possible role in protein targeting and in signal transduction. Folia Microbiol.
42: 244-245.
Okorokov, L. A., Lehle (1998). Ca2+-ATPases of Saccharomyces cerevisiae: diversity
and possible role in protein sortimg. FEMS Microbiol. Lett.. 162: 83-91.
Okorokov, L. A., Silva, F. E., Façanha, A. L. O. (2001). Ca2+ and H+ homeostasis in
fission yeast: a role of Ca2+/H+ exchange and distinct VH+ - ATPases of secretory
pathway organelles FEBSLett. 505:321-324.
Padilla-Lo´ Pez S. and Pearce D. A. (2006). Saccharomyces cerevisiae Lacking
Btn1p Modulate Vacuolar ATPase Activity to Regulate pH Imbalance in the Vacuole.
J. of Biological Chemistry 281: 10273–10280.
Parra, K. J, Keenan, K. L., Kane, P. M. (2000). The H subunit (Vma13p) of the yeast
V-ATPase inhibits the ATPase activity of cytosolic V1 complexes. J. Biol. Chem.
275(28): 21761-7.
Pedersen, P.L., Carafoli, E. (1987). Ion motive ATPase. I. Ubiquity, properties and
significance to cell function. Trends Biochem. Science. 12, 146-150.
Peters C, Bayer MJ, Bühler S, Andersen JS, Mann M, Mayer A. (2001). Trans-
complex formation by proteolipid channels in the terminal phase of membrane fusion.
Nature. 409: 581-588.
Randall, S. K. And Sze, H. (1986). Properties of the partially purifed tonoplast H+
pumping ATPase. From oat roots. J.Biol. Chem. 261, 1364-1371.
Ratajczak, R. (2000). Structure, function and regulation of the plant vacuolar H+
translocating ATPase. Biochim. Biophys. Acta 1465: 17-36.
Rea, P. A., Griffith, C.J., Sanders, D. (1987). Purification of the N,N’-
dicyclohexylcarbo-diimide-binding proteolipid of a higher plant tonoplast H+- ATPase.
J. Biol Chem. 25(30):14745-52.

56
Rein, J., Voss, M., Blenau, W., Walz, B. and Baumann, O. (2008). Hormone-induced
assembly and activation of V-ATPase in blowfly salivary glands is mediated by
protein kinase A. Am J Physiol Cell Physiol, 294: C56-C65.
Ribeiro, C. C., Bernardes, N. R., Freitas, F. P., Monteiro, R. M., Retamal, C. A.,
Teixeira, L. R. S., Samarao, S. S., Granato, Thais M., Silva, F. E., Façanha, A. R.,
Okorokova-Façanha, A. L., Okorokov, L. A. (2010). New insights into the glucose-
dependent regulation of the yeast V H+-ATPase in vivo: the coupling modulation
without the V1 dissociation from membrane. BBA Biomemb. Submitted.
Samarão, S.S.(2003). “Caracterização das atividades das V H+-ATPase nas
organelas de via secretória de levedura e regulação destas enzimas pela glicose
extracelular” Tese apresentada ao Centro de Biociências e Biotecnologia, para
obtenção do grau de Doutor em Ciências Biológicas, na Universidade Estadual do
Norte Fluminense Darcy Ribeiro, Campos dos Goytacases, Rio de Janeiro.
Samarão S.S., Teodoro C.E., Silva F.E., Ribeiro C.C., Granato T.M., Bernardes N.R.,
Retamal C.A., Façanha A.R., Okorokova-Façanha A.L., Okorokov L.A. (2009). V
H(+)-ATPase along the yeast secretory pathway: Energization of the ER and Golgi
membranes. Biochim Biophys Acta 1788(2): 303-13.
Serrano, R. (1983). In vivo glucose activation of the yeast plasma membrane
ATPase. FEBS Lett. 156(1): 11-14.
Serrano, R. (1993). Structure,function and regulation of plasma membrane H+-
ATPase, FEBS Lett. 325: 108-111.
Stevens, T. H., Forgac, M. (1997). Structure, function and regulation of the vacuolar
H+-ATPase. Annu. Ver. Cell. Dev. Biol. 13:779-808.
Swallow, C. J., Grinstein, S.,Sudsbury, R. A., Rotstein, O. D., (1993). Relative roles
of Na+/H+ exchange and vacuolar-type H+ ATPase in regulating cytoplasmic pH and
function in murine peritoneal macrophages. J. Cell. Physiol. 157: 453-460.
Teodoro, C. E. S. (2004). Bombas protonicas de organelas de via secretória:
comparação entra leveduras e plantas. Tese apresentada ao Centro de Biociências
e Biotecnologia, para obtenção do grau de Doutor em em Ciências Biológicas, na

57
Universidade Estadual do Norte Fluminense Darcy Ribeiro, Campos dos
Goytacases, Rio de Janeiro.
Toyomura T, Murata Y, Yamamoto A, Oka T, Sun-Wada GH, Wada Y, Futai M.
(2003). From lysosomes to the plasma membrane: localization of vacuolar-type H+ -
ATPase with the a3 isoform during osteoclast differentiation. J. Biol. Chem. 278:
22023–22030.
Uchida, E., Ohsumi, Y. And Anraku, Y. (1985). Purification and properties of H+-
translocating, Mg2+-adenosine triphosphstase from vacuolar membranes of
Saccharomyces cerevisiae. J. Biol. Chem. 260: 1090-1095.
Venema, K. and Palmgren, M.G. (1995). Metabolic modulation of transport coupling
ratio in yeast plasma membrane H+-ATPase. J.Biol.Chem. 270: 19659-19667.
Vitavska, O., Wieczorek, H. and Merzendorfer, H. (2003). A novel role for subunit C
in mediating binding of the H+-V-ATPase to the actin cytoskeleton J.Biol.Chem. 278:
18499–18505
Voet, D., Voet, J. G., Pratt, C. W. Fundamentos de Bioquímica. Porto Alegre: Artes
Médicas Sul Ltda., 2000.
White, J. M. (1992). Membrane fusion. Science 258: 917-924.
Wieczorek, H., Brown, D., Grinstein, S., Ehreufeld, J., Harvey, W. R. (1999). Animal
plasma membrane energization by proton-motive V-ATPases. BioEssays 21: 637-
648.
Wieczorek, H., Putzenlechner, M., Zeiske, W., Klein, U. (1991). A vacuolar type
proton pump energizes K+/H+ antiport in an animal plasma membrane. J. Biol.
Chem. 266: 15340-15347.
Wilkens, S., Zhang, Z., Zheng, Y. (2005). A structural model of the vacuolar ATPase
from transmission electron microscopy. Micron 36: 109-126.
Xie, X. S. And Stone, D. K. (1986). Isolation and reconstitution of the clathrincoated
vesicle proton translocating complex. J. Biol. Chem. 261: 2492-2495.

58
Xu, T., Forgac, M. (2000). Subunit D (Vma8p) of the yeast V-ATPase plays a role in
coupling of proton transport and ATP hydrolysis. J. Biol. Chem. 275: 22075-22081.
Yabe, I., Horiuchi, K., Nakahara, K., Hiyama, T., Yamanaka, T., Wang, P., Toda, K.,
Hirata, A. ,Ohsumi, Y., Hirata, R., Anraku, Y., and Kusaka, I. (1999) Patch clamp
studies on V-type ATPase of vacuolar membrane of haploid Saccharomyces
cerevisiae. J. Biol. Chem. 274: 34903–34910.
Zhang, J., Feng, Y., Forgac, M. (1994). Proton Conduction and Bafilomycin Binding
by the Vo Domain of the Coated Vesicle V-ATPase. J. Biol. Chem., 269: 23523-
25523