
RIO REFERENTE AO PROCESSO AGRISUS 1082/12 · A macrofauna do solo é composta por invertebrados com...
23
RELATÓRIO REFERENTE AO PROCESSO AGRISUS 1082/12 Projeto n° 1082/12 Titulo da Pesquisa: “Influência da mudança do uso da terra nos atributos físicos e na fauna do solo em áreas de agricultura familiar”. Coordenador do Projeto: Miguel Cooper Instituição: Escola Superior de Agricultura “Luiz de Queiroz”. Avenida Pádua Dias, 11 – Piracicaba – SP, CEP: 13418-900. Telefone: (19) 34294100. E-mail: [email protected] Local da Pesquisa: Projeto de Assentamento Praialta Piranheira, Nova Ipixuna, Pará. Valor financiado pela Fundação Agrisus: R$ 5000,00 Vigência do projeto: 14/12/2012 a 01/07/2013. RESUMO O uso inadequado do solo pode resultar em modificações na sua qualidade física e biológica. O objetivo deste trabalho foi realizar uma avaliação da qualidade do solo em duas topossequências, uma de mata nativa e outra de pastagem em uma área de agricultura familiar no sudeste do Pará. A análise da qualidade física e biológica do solo nos diferentes usos será usada para nortear as ações de intervenções que poderão ser realizadas na região, bem como a alocação de eventuais recursos financeiros destinados ao assentamento. O presente relatório diz respeito à terceira viagem de campo realizada com apoio financeiro fornecido pela Agrisus. Esta viagem visou a realização das coletas referentes à macro e mesofauna do solo e de amostras para a determinação dos parâmetros de avaliação da qualidade físico-química do solo como análise da umidade, química e curva de retenção de água no solo. 1. INTRODUÇÃO A macrofauna do solo é composta por invertebrados com diâmetro corporal acima de 2 mm representado por minhocas, coleópteros, cupins, formigas, aranhas, quilópodas, diplópodas, dentre outros e são fundamentais para o funcionamento do solo e imprescindíveis para a ciclagem e mineralização de nutrientes, decomposição de compostos orgânicos (Lavelle, Bignell e Lepage, 1997; Thomas et al., 2004). Geralmente recebem a denominação de “engenheiros do solo” devido a sua capacidade de se mover pelo solo e construir estruturas organo-minerais, galerias, ninhos e câmaras (Lavelle, Bignell e Lepage, 1997; Jouquet et al., 2006). Essa movimentação é mais frequente e evidente na camada de 0-20 cm, criando uma macroporosidade capaz de impactar agregação, estrutura e propriedades hidráulicas do solo (Lavelle, Bignell e Lepage, 1997; Barros et al., 2001; Lavelle et al., 2001).
Transcript of RIO REFERENTE AO PROCESSO AGRISUS 1082/12 · A macrofauna do solo é composta por invertebrados com...

RELATÓRIO REFERENTE AO PROCESSO AGRISUS 1082/12
Projeto n° 1082/12
Titulo da Pesquisa: “Influência da mudança do uso da terra nos atributos físicos e na fauna do solo em áreas de agricultura familiar”.
Coordenador do Projeto: Miguel Cooper
Instituição: Escola Superior de Agricultura “Luiz de Queiroz”. Avenida Pádua Dias, 11 – Piracicaba – SP, CEP: 13418-900. Telefone: (19) 34294100. E-mail: [email protected]
Local da Pesquisa: Projeto de Assentamento Praialta Piranheira, Nova Ipixuna, Pará.
Valor financiado pela Fundação Agrisus: R$ 5000,00
Vigência do projeto: 14/12/2012 a 01/07/2013.
RESUMO
O uso inadequado do solo pode resultar em modificações na sua qualidade física e
biológica. O objetivo deste trabalho foi realizar uma avaliação da qualidade do solo em duas
topossequências, uma de mata nativa e outra de pastagem em uma área de agricultura
familiar no sudeste do Pará. A análise da qualidade física e biológica do solo nos diferentes
usos será usada para nortear as ações de intervenções que poderão ser realizadas na região,
bem como a alocação de eventuais recursos financeiros destinados ao assentamento. O
presente relatório diz respeito à terceira viagem de campo realizada com apoio financeiro
fornecido pela Agrisus. Esta viagem visou a realização das coletas referentes à macro e
mesofauna do solo e de amostras para a determinação dos parâmetros de avaliação da
qualidade físico-química do solo como análise da umidade, química e curva de retenção de
água no solo.
1. INTRODUÇÃO
A macrofauna do solo é composta por invertebrados com diâmetro corporal acima de
2 mm representado por minhocas, coleópteros, cupins, formigas, aranhas, quilópodas,
diplópodas, dentre outros e são fundamentais para o funcionamento do solo e imprescindíveis
para a ciclagem e mineralização de nutrientes, decomposição de compostos orgânicos (Lavelle,
Bignell e Lepage, 1997; Thomas et al., 2004). Geralmente recebem a denominação de
“engenheiros do solo” devido a sua capacidade de se mover pelo solo e construir estruturas
organo-minerais, galerias, ninhos e câmaras (Lavelle, Bignell e Lepage, 1997; Jouquet et al.,
2006). Essa movimentação é mais frequente e evidente na camada de 0-20 cm, criando uma
macroporosidade capaz de impactar agregação, estrutura e propriedades hidráulicas do solo
(Lavelle, Bignell e Lepage, 1997; Barros et al., 2001; Lavelle et al., 2001).

A porosidade é diretamente influenciada pela diversidade de populações dos
invertebrados, os quais definirão o tipo e tamanho de poros que formados (Lavelle et al.,
2001). As alterações provocadas pela fauna do solo podem ser tanto positivas quanto
negativas, dependendo da espécie e da quantidade de indivíduos presentes. A dominância de
cupins pode tanto aumentar quanto reduzir a porosidade, dependendo da espécie presente
(Garnier-Sillam, Braudeau e Tessier, 1991), as formigas são responsáveis por modificações nas
propriedades do solo, dependendo da densidade e tamanho dos indivíduos (Rosengren et al.,
1987; Woodell e King, 1991), já as minhocas atuam no solo criando canais que facilitam o
transporte de água e gases, a mistura de material mineral e orgânico e o acúmulo de excretas
(Brown et al., 2000).
O cálculo da diversidade de espécies da fauna do solo é feito através da relação entre
o número de espécies presentes (riqueza) e a distribuição do número de indivíduos entre as
espécies (Walker, 1989) e pode ser usada como indicador da qualidade do solo uma vez que a
fauna edáfica é extremamente sensível às alterações ocorridas nos agroecossistemas (Baretta
et al., 2003). A qualidade de um solo está relacionada à saúde e produtividade dos
ecossistemas e pode ser determinada através da utilização de indicadores como abundância,
diversidade e estrutura das comunidades de fauna edáfica (Lavelle, Bignell e Lepage, 1997;
Pankhurst et al., 1997; Ferreira, Tavares Filho e Ferreira, 2010). Elevadas frequências de
indivíduos podem ser consideradas indicadores de degradação do solo enquanto a presença
de determinadas espécies podem ser indicadores de boa qualidade do solo (Rovedder et al.,
2004).
As práticas de uso e manejo do solo podem alterar as comunidades de
macroinvertebrados do solo modificando o ambiente físico e reduzindo a entrada de material
orgânico no solo (Decaëns et al., 1994). A substituição das áreas de mata nativa por áreas de
cultivo ou pastagem resultam na modificação da fauna do solo e consequentemente da
morfologia e funcionamento hídrico do solo em períodos relativamente curtos (meses a anos)
(Mando e Miedema, 1997; Chauvel et al., 1999; Lavelle et al., 2001; Analy De Oliveira e
Merlim, 2005; Silva et al., 2006; Ferreira, Tavares Filho e Ferreira, 2010).
A mudança do uso do solo através da retirada da vegetação nativa e implantação de
pastagens é cada vez mais frequente, sendo estimado que o Brasil possua atualmente cerca de
172 milhões de hectares de pastagens, sendo cerca de 61 milhões localizados na Amazônia
Legal, ocupando 80% da área total desmatada nessa região (IBGE, INPE).
A retirada da vegetação natural para implantação de pastagens para criação de gado
foi incentivada a partir da década de 1970 através de uma política de incentivo fiscal do
governo federal, se expandindo pela região de maneira rápida e ocupando áreas dos grandes e

pequenos agricultores (Michelotti; Fearnside, 2005). Nas décadas seguintes observa-se o
aumento do número de agricultores ligados à criação de bovinos (Carvalho Machado, 2003),
principalmente devido ao aumento do número de assentamentos rurais e expansão da
reforma agrária na Amazônia (Tavares, 2003) .
Dentre as regiões de maior expansão do desmatamento destaca-se o Estado do Pará,
que apresentou nos últimos 8 anos a maior taxa de desmatamento dentre os estados
constituintes da Amazônia Legal, com cerca de 14441 km2 (Inpe).
A expansão da pecuária e a mudança do uso do solo nos assentamentos rurais da
região Sudeste do Pará são responsáveis pela compactação do solo, exaustão de nutrientes,
perda de produtividade, mudanças no regime hidrológico, perda de biodiversidade e emissão
de gases do efeito estufa (Fearnside, 2005). Essas mudanças reduzem a qualidade e
produtividade das pastagens acarretando no abandono destas terras e abertura de novas
áreas.
Sabendo que a conservação da fauna do solo é importante para auxiliar na
manutenção da produtividade e das propriedades físicas do solo (Mathieu et al., 2004) o
presente projeto pretende realizar uma análise quali-quantitativa da fauna edáfica objetivando
a criação de índices de qualidade do solo baseados nas características físicas e de fauna do
solo.
A criação deste índice de qualidade deve ser útil para que o agricultor consiga avaliar
por conta própria o nível de degradação das suas áreas produtivas e perceber que a adoção de
práticas conservacionistas pode auxiliar na manutenção da produtividade e assim manter a
população assentada em seus lotes.
2. Objetivos
2.1. Objetivo geral
Neste contexto, visando suprir a necessidade de mais pesquisas na área o presente
projeto pretende conhecer e quantificar a fauna do solo nas diferentes posições topográficas,
épocas do ano e usos do solo para entender como a mudança do uso do solo atua sobre a
fauna edáfica amazônica. Ao final também espera-se identificar os fatores que poderiam ser
considerados como índices de qualidade do solo baseados nas propriedades físicas e fauna do
solo.
O presente projeto é parte do projeto “Influência do uso da terra sobre o
funcionamento do solo e na sustentabilidade dos sistemas agroextrativistas da Amazônia
Oriental”, e recebeu apoio financeiro da Fundação Agrisus para a primeira, segunda e terceira

viagens de campo realizadas em março de 2011, julho de 2012 e janeiro de 2013,
respectivamente.
Durante a primeira viagem de campo (projeto 779/11) foram escolhidos os transectos
a ser trabalhados e os locais de abertura das trincheiras. Foram abertas três trincheiras em
cada uma das topossequências sendo 3 sob mata nativa e 3 pastagem. Em cada uma das
trincheiras foram coletadas amostras deformadas e indeformadas para caracterização do solo.
A segunda viagem de campo (projeto 964/12) teve como objetivo a caracterização da
fauna e das condições físicas, químicas e físico-hídricas do solo no período seco (julho 2012).
Nesta viagem foram coletadas amostras para avaliação da macro e mesofauna do solo, para
caracterização física (umidade), físico-hídrica (curva de retenção) e química do solo. Também
foram coletadas amostras para determinação da massa seca vegetal.
O presente relatório apresentará os dados referentes à terceira viagem de campo
(projeto 1082/12), a qual teve como objetivo a caracterização da fauna e das condições físicas,
químicas e físico-hídricas do solo para o período úmido (janeiro 2013). Durante esta viagem
foram coletadas amostras para avaliação da macro e mesofauna do solo e para caracterização
física (umidade), físico-hídrica (curva de retenção) e química do solo. Também foram coletadas
amostras para determinação da massa seca vegetal.
2. MATERIAL E MÉTODOS
O presente trabalho foi desenvolvido no Projeto de Assentamento Agroextrativista
Praialta Piranheira (PAE), situado no município de nova Ipixuna, localizado a 573 km da capital
Belém, no Estado do Pará.
A escolha da área foi motivada por ser uma área declivosa, com cerca de 1/3 da
propriedade coberta por pastagem e 2/3 coberta por mata nativa. Foram estabelecidas duas
topossequências, uma sob mata nativa e outra sob pastagem. Nestas topossequências foram
retiradas amostras para análise de macrofauna, mesofauna, química de rotina e umidade do
solo, além da realização de tradagens e descrições para confecção da análise estrutural da
área.

Figura 1. Esquema ilustrativo da localização da área
Figura 2. Representação da topossequência escolhida para as coletas e análises
2.1. Macrofauna
As coletas para análise da macrofauna do solo foram realizadas segundo metodologia
proposta por Anderson e Ingram (1993) que consiste na retirada de monólitos de 0,25 x 0,25 x
0,30 m de profundidade. Os monólitos retirados são separados em quatro camadas sendo
estas serapilheira, 0-0,1 m, 0,1-0,2 m e 0,2-0,3 m. Foram retirados 5 repetições em cada uma
das posições do relevo (topo – T1, ombro – T2 e sopé – T3) totalizando 60 amostras para a
topossequência sob mata nativa e 60 amostras para a topossequência sob pastagem.
Após a retirada dos monólitos a extração e separação da macrofuna foi realizada
através de separação manual (“handsorting”). As minhocas foram fixadas em solução de

formol 4% e outros invertebrados foram acondicionados em frascos de acrílico e conservados
em solução de álcool 70%.
No presente momento as minhocas e outros invertebrados encontrados estão em
processo de análise na Universidad Nacional de Rio Cuarto, Argentina.
Figura 3. Coleta da liteira Figura 4. Coleta dos monólitos Figura 5. Separação manual da macrofauna
2.2. Mesofauna
A coleta da mesofauna foi realizada com ajuda de anéis volumétricos de 0,05 m de
altura e 0,05 m de diâmetro. As amostras foram coletadas nas diferentes posições do relevo
(T1, T2 e T3) em cinco repetições. Cada uma das repetições foi separada em 4 estratos, sendo
a liteira, 0 - 0,05 m; 0,1 - 0,15 m; 0,2 - 0,25 m, totalizando 90 amostras para a topossequência
sob mata nativa e 90 amostras para a topossequência sob pastagem.
A extremidade do cilindro orientada em relação à superfície foi coberta com tule de
malha 2 mm para permitir a passagem da mesofauna e em seguida as amostras foram
colocadas em equipamento de Berlese para a extração da mesofauna. O equipamento foi
mantido ligado durante 11 dias e a mesofauna retirada foi acondicionada em frascos contendo
solução de álcool 70%.
Figura 6. Coleta dos anéis para mesofauna
Figura 7. Equipamento de Berlese
Figura 8. Frasco com álcool 70% para coleta de mesofauna

2.3. Massa seca
As amostras de liteira foram acondicionadas em sacos de papel e levados à estufa com
ventilação forçada a 60° C até que atingissem peso constante. Após obtenção de peso
constante as amostras foram pesadas para determinação da massa seca.
2.4. Umidade do solo
A determinação da umidade do solo no momento da amostragem foi realizada nas
amostras deformadas de solo coletadas nas profundidades de 0 - 0,1 m; 0,1 – 0,2 m e 0,2 – 0,3
m.
As amostras foram acondicionadas em recipientes metálicos e pesadas úmidas. Em
seguida estas foram levadas à estufa com ventilação forçada e temperatura constante de 105°
C por 48h. Após a secagem o cálculo da umidade foi realizado segundo fórmula a seguir
% = 100 � � − � �� �
Onde U é o teor de umidade, Pu é o peso do solo úmido e Ps o peso do solo seco.
2.5. Análises químicas
Foram coletadas amostras deformadas de solo nas profundidades 0 – 0,1; 0,1 – 0,2 e
0,2 – 0,3 m para a realização de análises químicas.
As análises de pH (água e KCl), K, P, Al, H+Al, Ca, Mg e matéria orgânica foram
realizadas de acordo com Raij et al (1987) e foram calculados os valores de soma de bases,
m%, CTC e V%.
2.6. Curva de retenção de água no solo
Após a extração da mesofauna as amostras foram utilizadas para determinação da
curva de retenção de água no solo. Foram coletadas amostras indeformadas em anéis
cilíndricos de aço inox (100 cm3) e a retenção foi determinada em câmaras de Richards com
placa porosa, com base na umidade gravimétrica para os potenciais de 1, 3, 6, 8, 10, 33, 70,
100 e 500 kPa. O potencial de 1500 kPa foi determinado com auxílio do aparelho WP4.
Os pontos da curva de retenção foram ajustados segundo o modelo van Genuchten
(1980) com ajuda do programa RetC.
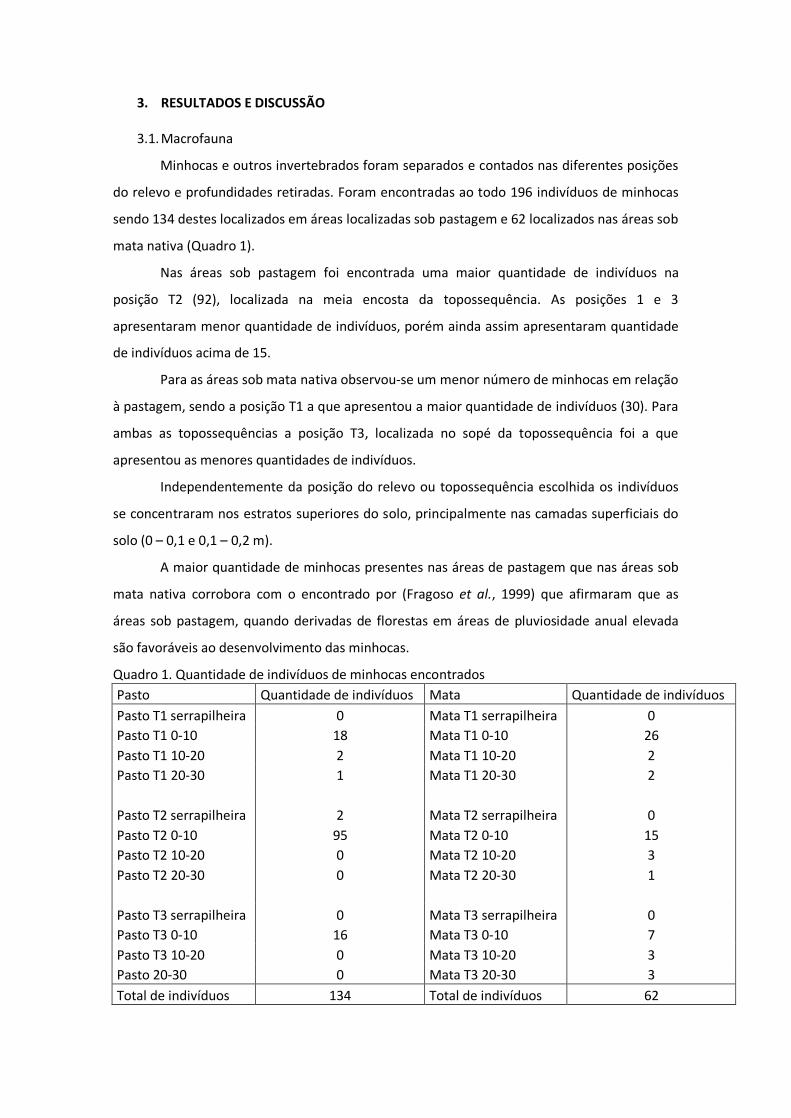
3. RESULTADOS E DISCUSSÃO 3.1. Macrofauna
Minhocas e outros invertebrados foram separados e contados nas diferentes posições
do relevo e profundidades retiradas. Foram encontradas ao todo 196 indivíduos de minhocas
sendo 134 destes localizados em áreas localizadas sob pastagem e 62 localizados nas áreas sob
mata nativa (Quadro 1).
Nas áreas sob pastagem foi encontrada uma maior quantidade de indivíduos na
posição T2 (92), localizada na meia encosta da topossequência. As posições 1 e 3
apresentaram menor quantidade de indivíduos, porém ainda assim apresentaram quantidade
de indivíduos acima de 15.
Para as áreas sob mata nativa observou-se um menor número de minhocas em relação
à pastagem, sendo a posição T1 a que apresentou a maior quantidade de indivíduos (30). Para
ambas as topossequências a posição T3, localizada no sopé da topossequência foi a que
apresentou as menores quantidades de indivíduos.
Independentemente da posição do relevo ou topossequência escolhida os indivíduos
se concentraram nos estratos superiores do solo, principalmente nas camadas superficiais do
solo (0 – 0,1 e 0,1 – 0,2 m).
A maior quantidade de minhocas presentes nas áreas de pastagem que nas áreas sob
mata nativa corrobora com o encontrado por (Fragoso et al., 1999) que afirmaram que as
áreas sob pastagem, quando derivadas de florestas em áreas de pluviosidade anual elevada
são favoráveis ao desenvolvimento das minhocas.
Quadro 1. Quantidade de indivíduos de minhocas encontrados Pasto Quantidade de indivíduos Mata Quantidade de indivíduos Pasto T1 serrapilheira 0 Mata T1 serrapilheira 0 Pasto T1 0-10 18 Mata T1 0-10 26 Pasto T1 10-20 2 Mata T1 10-20 2 Pasto T1 20-30 1 Mata T1 20-30 2
Pasto T2 serrapilheira 2 Mata T2 serrapilheira 0 Pasto T2 0-10 95 Mata T2 0-10 15 Pasto T2 10-20 0 Mata T2 10-20 3 Pasto T2 20-30 0 Mata T2 20-30 1
Pasto T3 serrapilheira 0 Mata T3 serrapilheira 0 Pasto T3 0-10 16 Mata T3 0-10 7 Pasto T3 10-20 0 Mata T3 10-20 3 Pasto 20-30 0 Mata T3 20-30 3 Total de indivíduos 134 Total de indivíduos 62

No presente momento as amostras de macrofauna referentes a outros exemplares que não sejam minhocas estão em processo de análise. As espécies foram separadas em grande grupos (Diplópodas, Chilopodas, Formigas, Cupins, Diplura, Dermáptera, etc) e quantificadas. Até o presente momento foram realizadas as amostras referentes à segunda coleta de campo (Projeto n° 964/12) e está em andamento a separação das amostras referentes à terceira coleta de campo (Projeto n° 1082/12).
Para as amostras retiradas em T1 percebe-se uma menor quantidade de macrofauna do pasto em consideração à mata em serapilheira (Figura 9). Além da maior quantidade de indivíduos nas amostras de serapilheira da mata também pode-se observar uma maior diversidade de espécies sendo que na pastagem foram encontrados indivíduos pertencentes somente a 2 grupos (Cupins ou térmitas e aranhas) e nas áreas de mata foram encontrados indivíduos pertencentes a 8 grupos diferentes (Coleóptera Formiga, Cupins, Dermáptera, Aranha, Quilópodes, Blatária e larvas).
Para as amostras em profundidade se percebe uma mudança nessa tendência, com maior número de indivíduos nas amostras de pasto, porém estes pertencentes a poucos grupos, sendo que nas áreas de mata percebe-se uma maior diversidade de indivíduos, apesar do pouco número de exemplares presentes.
Entretanto, esse comportamento não é observado para a profundidade de 10-20 cm onde as áreas de mata apresentam pequeno número de indivíduos e diversidade menor que nas áreas de pasto.

Figura 9. Distribuição dos indivíduos em grandes grupos de pastagem e mata para trincheira 1. NTI = Número total de indivíduos
Para a trincheira 2 percebe-se uma maior quantidade de indivíduos e também de
diversidade de grupos para as área de mata em relação à área de pasto (Figura 10). Em profundidade percebe-se uma quantidade maior de indivíduos na pastagem, porém uma maior diversidade de grupos encontrados nas áreas de mata, exceto para a profundidade de 20-30 cm que segue o mesmo padrão para serapilheira, com maior quantidade e diversidade de indivíduos.
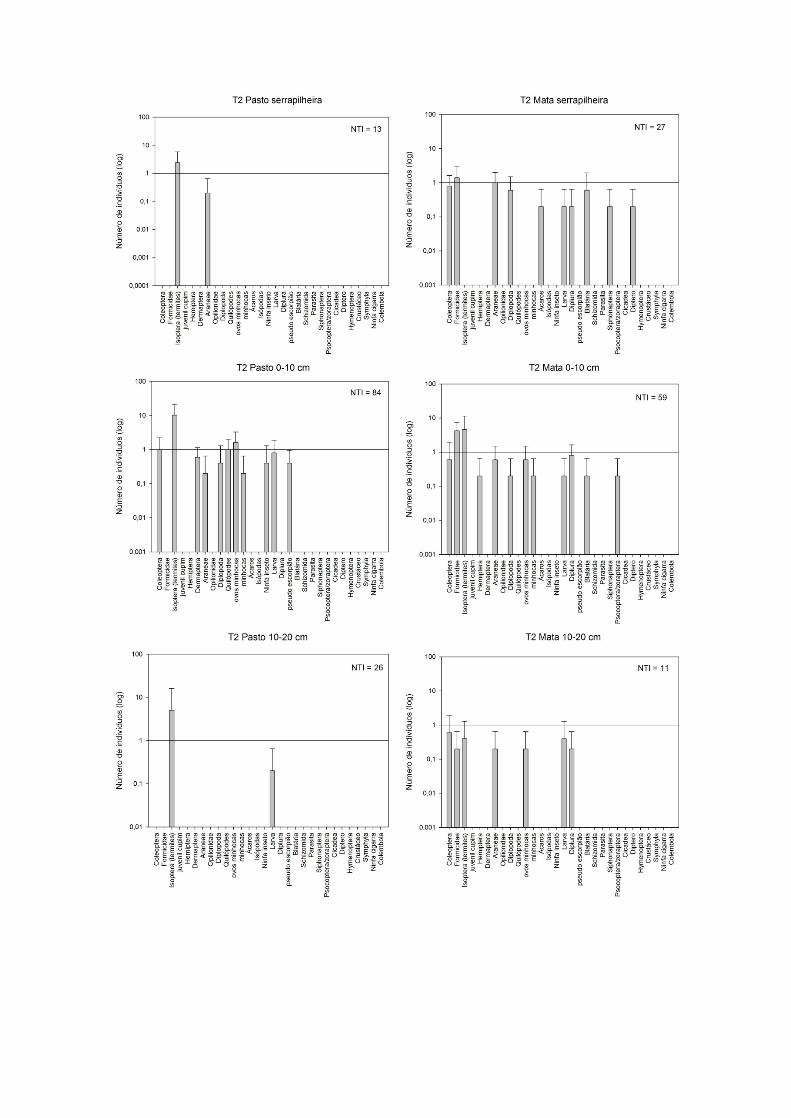

Figura 10. Distribuição dos indivíduos em grandes grupos de pastagem e mata para trincheira 2. NTI = Número total de indivíduos
Para a terceira trincheira, mais próxima ao curso d’água em todas as camadas amostradas (serapilheira, 0-10; 10-20 e 20-30 cm) tanto a quantidade de indivíduos quanto a diversidade de espécies encontradas foram maiores na área de mata que na área de pastagem (Figura 11).
Na maioria dos casos apresentados as áreas amostradas sob mata apresentaram maior diversidade, com a fauna encontrada distribuída em maior quantidade de grupos que no pasto. Além disso, percebe-se que em quase todas as amostras há uma predominância de insetos sociais (formigas e cupins) concordando com dados apresentados por (Manhães et al., 2013).
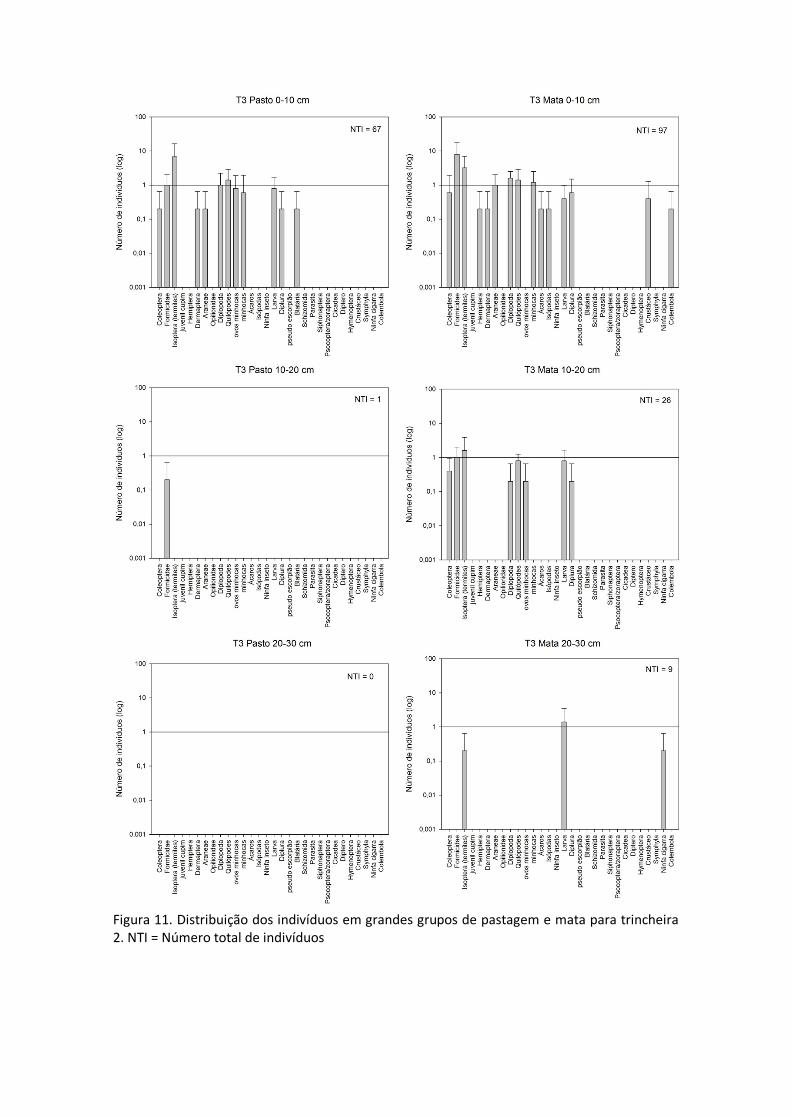
Figura 11. Distribuição dos indivíduos em grandes grupos de pastagem e mata para trincheira 2. NTI = Número total de indivíduos

3.2. Mesofauna
A extração da mesofauna foi realizada das amostras de serapilheira e dos anéis de solo
coletados com auxílio do equipamento de Berlese. Os indivíduos retirados foram armazenados
e ainda não foram realizadas as determinações para mesofauna.
3.3. Massa seca (MS)
A massa seca produzida nas áreas de pastagem foi menor que a produção de massa
seca nas áreas cobertas por mata nativa (Quadro 2). As diferenças observadas podem ser
atribuídas, principalmente, às diferenças quanto ao tipo de material produzido em cada uma
das topossequências analisadas. As áreas sob pastagem apresentam produção de material
pouco lignificado, homogêneo e com presença de material folhoso (Figura 12). Para as áreas
sob mata nativa a serapilheira é composta por material diversificado, com presença de folhas e
material mais lignificado como talos, troncos e gravetos (Figura 13).
A maior quantidade de MS da serapilheira não indica obrigatoriamente uma maior
cobertura do solo. Uma composição mais homogênea composta basicamente por material
folhoso pode garantir um espalhamento mais homogêneo do material vegetal sobre o solo,
oferecendo melhor cobertura que a serapilheira composta por material diversificado e
heterogêneo como, por exemplo, das áreas de mata nativa.
A composição predominantemente herbácea das pastagens, pobre em lignina e
facilmente decomponível é usada como fonte rápida de alimento pelos organismos
decompositores, oferecendo melhores condições à proliferação e desenvolvimento das
minhocas nas áreas localizadas sob pastagem (Barros et al., 2004)
Quadro 2. Produção de massa seca das áreas de pastagem e mata nativa
Massa Seca determinada para Macrofauna
Pasto MS t ha-1 Mata MS t ha-1 T1 16.04
T1 26.48
T2 15.22
T2 22.94 T3 27.11
T3 48.30

Figura 12. Serrapilheira presente em área sob pastagem
Figura 13. Serrapilheira presente em área sob mata nativa.
3.4. Umidade do solo
A umidade do solo é um fator que pode contribuir favoravelmente para a quantidade
de organismos presente no solo, uma vez que a mesofauna do solo é extremamente
dependente da mesma, movimentando-se nos poros do solo e na interface solo-serrapilheira
(Coleman, 2004; Albuquerque et al., 2010).
Para esta coleta, realizada na época úmida pode-se perceber que os valores de
umidade nas áreas de mata nativa se mantiveram maiores que para as áreas de pastagem
(Quadro 3). Também percebe-se que as camadas superficiais apresentaram maiores valores de
umidade para ambas as topossequências, o que poderia contribuir para a concentração das
minhocas na profundidade de 0 – 0,1 m.
Quadro 3. Umidade média encontrada nas áreas de pastagem e mata nativa
Pasto Umidade (%) Mata Umidade (%) T1 0-10 cm 20
T1 0-10cm 29
T1 10-20 cm 16
T1 10-20cm 29 T1 20-30 cm 17 T1 20-30cm 30 T2 0-10 cm 25
T2 0-10cm 26
T2 10-20 cm 17
T2 10-20cm 26 T2 20-30 cm 18 T2 20-30cm 28 T3 0-10 cm 21
T3 0-10cm 32
T3 10-20 cm 16
T3 10-20cm 27 T3 20-30 cm 16 T3 20-30cm 26
3.5. Análises químicas
Os dados obtidos para as análises químicas nas áreas de pastagem e mata nativa estão
apresentados a seguir (Quadro 4).
O pH nas áreas sob pastagem apresentam-se em torno de 5 para todas as
profundidades analisadas, valores estes maiores que para as áreas de mata nativa que se
mantiveram em torno de 4 para todas as profundidades (Quadro 4).
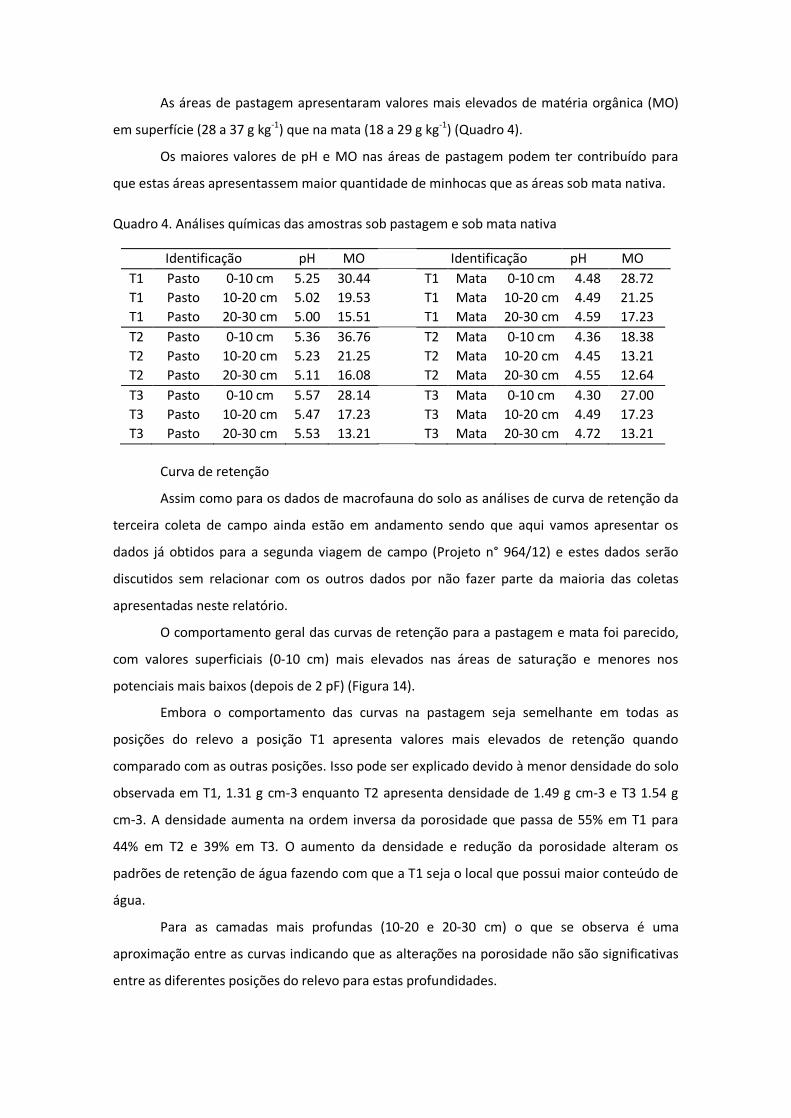
As áreas de pastagem apresentaram valores mais elevados de matéria orgânica (MO)
em superfície (28 a 37 g kg-1) que na mata (18 a 29 g kg-1) (Quadro 4).
Os maiores valores de pH e MO nas áreas de pastagem podem ter contribuído para
que estas áreas apresentassem maior quantidade de minhocas que as áreas sob mata nativa.
Quadro 4. Análises químicas das amostras sob pastagem e sob mata nativa
Identificação pH MO Identificação pH MO T1 Pasto 0-10 cm 5.25 30.44 T1 Mata 0-10 cm 4.48 28.72 T1 Pasto 10-20 cm 5.02 19.53 T1 Mata 10-20 cm 4.49 21.25 T1 Pasto 20-30 cm 5.00 15.51 T1 Mata 20-30 cm 4.59 17.23 T2 Pasto 0-10 cm 5.36 36.76 T2 Mata 0-10 cm 4.36 18.38 T2 Pasto 10-20 cm 5.23 21.25 T2 Mata 10-20 cm 4.45 13.21 T2 Pasto 20-30 cm 5.11 16.08 T2 Mata 20-30 cm 4.55 12.64 T3 Pasto 0-10 cm 5.57 28.14 T3 Mata 0-10 cm 4.30 27.00 T3 Pasto 10-20 cm 5.47 17.23 T3 Mata 10-20 cm 4.49 17.23 T3 Pasto 20-30 cm 5.53 13.21 T3 Mata 20-30 cm 4.72 13.21
Curva de retenção
Assim como para os dados de macrofauna do solo as análises de curva de retenção da
terceira coleta de campo ainda estão em andamento sendo que aqui vamos apresentar os
dados já obtidos para a segunda viagem de campo (Projeto n° 964/12) e estes dados serão
discutidos sem relacionar com os outros dados por não fazer parte da maioria das coletas
apresentadas neste relatório.
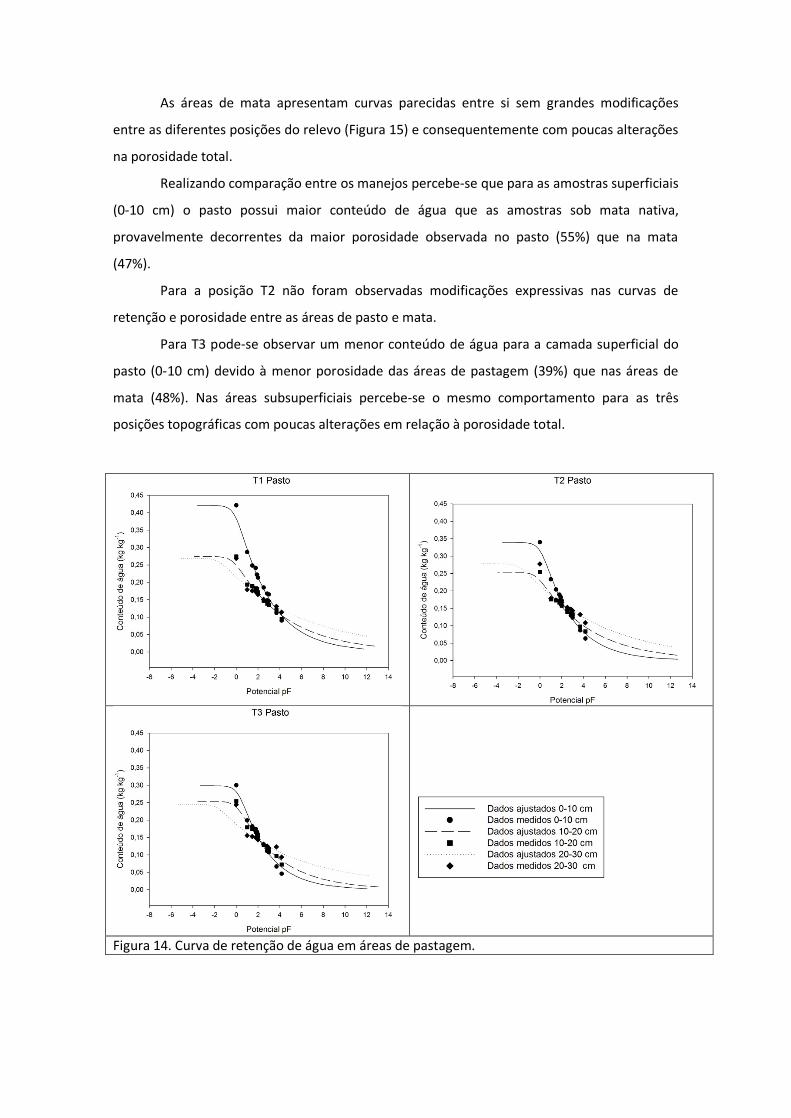
O comportamento geral das curvas de retenção para a pastagem e mata foi parecido,
com valores superficiais (0-10 cm) mais elevados nas áreas de saturação e menores nos
potenciais mais baixos (depois de 2 pF) (Figura 14).
Embora o comportamento das curvas na pastagem seja semelhante em todas as
posições do relevo a posição T1 apresenta valores mais elevados de retenção quando
comparado com as outras posições. Isso pode ser explicado devido à menor densidade do solo
observada em T1, 1.31 g cm-3 enquanto T2 apresenta densidade de 1.49 g cm-3 e T3 1.54 g
cm-3. A densidade aumenta na ordem inversa da porosidade que passa de 55% em T1 para
44% em T2 e 39% em T3. O aumento da densidade e redução da porosidade alteram os
padrões de retenção de água fazendo com que a T1 seja o local que possui maior conteúdo de
água.
Para as camadas mais profundas (10-20 e 20-30 cm) o que se observa é uma
aproximação entre as curvas indicando que as alterações na porosidade não são significativas
entre as diferentes posições do relevo para estas profundidades.
As áreas de mata apresentam curvas parecidas entre si sem grandes modificações
entre as diferentes posições do relevo (Figura 15) e consequentemente com poucas alterações
na porosidade total.
Realizando comparação entre os manejos percebe-se que para as amostras superficiais
(0-10 cm) o pasto possui maior conteúdo de água que as amostras sob mata nativa,
provavelmente decorrentes da maior porosidade observada no pasto (55%) que na mata
(47%).
Para a posição T2 não foram observadas modificações expressivas nas curvas de
retenção e porosidade entre as áreas de pasto e mata.
Para T3 pode-se observar um menor conteúdo de água para a camada superficial do
pasto (0-10 cm) devido à menor porosidade das áreas de pastagem (39%) que nas áreas de
mata (48%). Nas áreas subsuperficiais percebe-se o mesmo comportamento para as três
posições topográficas com poucas alterações em relação à porosidade total.
Figura 14. Curva de retenção de água em áreas de pastagem.

Figura 15. Curva de retenção de água em áreas de mata nativa.
4. CONCLUSÕES
As áreas sob pasto apresentaram melhores condições de umidade e química do solo
favorecendo o aparecimento e desenvolvimento de maior quantidade de indivíduos da
macrofauna distribuídos em uma pequena variedade de grupos.
As áreas sob mata apresentam menor quantidade de indivíduos da macrofauna,
porém estes estão distribuídos em uma maior variedade de grupos.
As melhores condições químicas e físicas do solo sob pastagem podem ser atribuídas
ao manejo pouco intensivo da área, com pastejo de poucas unidades animal por hectare,
fazendo com que o pisoteio causado pelo gado seja minimizado e não prejudique a qualidade
física do solo.
Os autores gostariam de ressaltar que algumas análises de identificação da
macrofauna, cálculos de riqueza e diversidade de espécies e curva de retenção de água no solo
ainda estão em andamento. A conclusão destas ainda não pode ser concluída devido ao tempo
de andamento das análises ser maior que o tempo fornecido pela Agrisus entre as coletas de
campo e apresentação do presente relatório.

Sendo assim os autores ressaltam que não podemos tirar conclusões definitivas sobre
os dados coletados nas viagens de campo uma vez que as análises supracitadas tem o prazo de
finalização em dezembro de 2013 e todos os dados são imprescindíveis para que se possa
concluir de maneira mais consistente o presente trabalho.
5. COMPENSAÇÕES OFERECIDAS À FUNDAÇÃO AGRISUS
Os primeiros resultados provenientes do financiamento realizado pela Fundação
AGRISUS às primeira e segunda viagem de campo já podem ser conferidos em:
VIDAL, A. Caracterização morfológica e micromorfológica do solo em sistemas agroextrativistas
da Amazônia Oriental. Trabalho de Conclusão de Curso apresentado à Escola Superior de
Agricultura “Luiz de Queiroz”. 2012. 57p.
VIDAL, A. Caracterização físico-hídrica do solo em sistemas agroextrativistas da Amazônia
oriental. Trabalho referente ao Estágio Vivencial apresentado à Escola Superior de Agricultura
“Luiz de Queiroz”. 2012. 49p.
MACUL, M. S. Caracterização do funcionamento do solo em sistemas agroextrativistas da
Amazônia Oriental. In: XX Simpósio Internacional de Iniciação Científica (apresentação oral).
2012.
Castilho, S. C. P.; Cooper, M.; Bedano, J. C. “Influência da mudança de uso da terra nos
atributos físicos e fauna do solo em áreas de agricultura familiar”. In: III Congreso Nacional de
Ecología y Biología de Suelos (pôster), Argentina. 2013.
Com os dados coletados nas três viagens de campo todas financiadas pela Fundação
Agrisus ainda estão em andamento duas teses de doutoramento e dois trabalho de pós-
doutoramento, todos realizados na Escola Superior de Agricultura “Luiz de Queiroz”. Os
trabalhos de doutoramento e um dos trabalhos de pós-doutoramento tem previsão de
conclusão em 2014 e o outro trabalho de pós-doutoramento tem previsão para conclusão em
2015.

6. DEMONSTRAÇÕES FINANCEIRAS DOS RECURSOS DA AGRISUS
Valor solicitado: R$ 6700,00
Valor aprovado: R$ 5000,00 (prestação de contas já realizada)
Piracicaba, 21 de junho de 2013
_________________________
Miguel Cooper
Coordenador do projeto
7. REFERÊNCIAS BIBLIOGRÁFICAS
ALBUQUERQUE, M. P. et al. Fauna edáfica em sistema de plantio homogêneo, sistema agroflorestal e em mata nativa em dois municípios do Rio Grande do Sul, Brasil. Biociências (On-line), v. 17, n. 1, p. 59-66, 2010.
ANALY DE OLIVEIRA, M.; MERLIM, A. D. O. Macrofauna edáfica em ecossistemas preservados e degradados de araucária no Parque Estadual de Campos do Jordão, SP. 2005. Piracicaba.
BARETTA, D. et al. Fauna edáfica avaliada por armadilhas de catação manual afetada pelo manejo do solo na região oeste catarinense. Revista de Ciências Agroveterinárias, v. 2, n. 01, p. 97-106, 2003.
BARROS, E. et al. The role of macrofauna in the transformation and reversibility of soil structure of an oxisol in the process of forest to pasture conversion. Geoderma, v. 100, n. 1-2, p. 193-213, Mar 2001. ISSN 0016-7061. Disponível em: < <Go to ISI>://WOS:000167007200009 >.
______. Soil physical degradation and changes in macrofaunal communities in Central Amazon. Applied Soil Ecology, v. 26, n. 2, p. 157-168, Jun 2004. ISSN 0929-1393. Disponível em: < <Go to ISI>://WOS:000221601300009 >.
CHAUVEL, A. et al. Pasture damage by an Amazonian earthworm. Nature, v. 398, n. 6722, p. 32-33, Mar 1999. ISSN 0028-0836. Disponível em: < <Go to ISI>://WOS:000079033900036 >.
COLEMAN, D. C. Fundamentals of soil ecology. Amsterdam: Elsevier Academic Press, 2004. 386.

DECAËNS, T. et al. Impact of land management on soil macrofauna in the Oriental Llanos of Colombia. European Journal of Soil Biology, v. 30, n. 4, p. 157-168, 1994. ISSN 1164-5563.
FEARNSIDE, P. M. Desmatamento na Amazônia brasileira:história, índices e conseqüências. Megadiversidade. Belo Horizonte: Conservation International do Brasil. 1: 10 p. 2005.
FERREIRA, R. R. M.; TAVARES FILHO, J.; FERREIRA, V. M. Efeitos de sistemas de manejo de pastagens nas propriedades físicas do solo. Semina: Ciências Agrárias, v. 31, n. 4, p. 913-932, 2010. ISSN 1679-0359.
FRAGOSO, C. et al. Earthworm communities of tropical agroecossystems: origin, structure and influence of management practices. In: LAVELLE, P.;BRUSSAARD, L., et al (Ed.). Earthworm management in tropical agroecossystem. Wallingford, Oxon, UK New York, NY, USA: Wallingford, Oxon, UK CABI Pub. New York, NY, USA, 1999. p.27-55.
GARNIER-SILLAM, E.; BRAUDEAU, E.; TESSIER, D. Rôle des termites sur le spectre poral des sols forestiers tropicaux. Cas deThoracotermes macrothorax Sjöstedt (Termitinae) et deMacrotermes mülleri (Sjöstedt)(Macrotermitinae). Insectes Sociaux, v. 38, n. 4, p. 397-412, 1991. ISSN 0020-1812.
JOUQUET, P. et al. Soil invertebrates as ecosystem engineers: Intended and accidental effects on soil and feedback loops. Applied Soil Ecology, v. 32, n. 2, p. 153-164, Jun 2006. ISSN 0929-1393. Disponível em: < <Go to ISI>://WOS:000237904500001 >.
LAVELLE, P. et al. SOM management in the tropics: Why feeding the soil macrofauna? Nutrient Cycling in Agroecosystems, v. 61, n. 1-2, p. 53-61, 2001. ISSN 1385-1314. Disponível em: < <Go to ISI>://WOS:000172975900006 >.
LAVELLE, P.; BIGNELL, D.; LEPAGE, M. Soil function in a changing world: the role of invertebrate ecosystem engineers. Eur. J. Soil Biol., v. 33, n. 4, p. 159-193p., 1997.
MANDO, A.; MIEDEMA, R. Termite-induced change in soil structure after mulching degraded (crusted) soil in the Sahel. Applied Soil Ecology, v. 6, n. 3, p. 241-249, 11// 1997. ISSN 0929-1393. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0929139397000127 >.
MANHÃES, C. M. C. et al. Meso-and macrofauna in the soil and litter of leguminous trees in a degraded pasture in Brazil. Agroforestry Systems, p. 1-12, 2013. ISSN 0167-4366.
MATHIEU, J. et al. A multi-scale study of soil macrofauna biodiversity in Amazonian pastures. Biology and Fertility of Soils, v. 40, n. 5, p. 300-305, Oct 2004. ISSN 0178-2762. Disponível em: < <Go to ISI>://WOS:000224612600002 >.

MICHELOTTI, F. Luta pela Terra e Assentamentos no Sudeste do Pará. III Encontro da Rede de Estudos Rurais. Campina Grande: s.n. p.
PANKHURST, C. E. et al. Biodiversity of soil organisms as an indicator of soil health. Biological indicators of soil health., p. 297-324, 1997. ISSN 0851991580.
ROVEDDER, A. P. et al. Fauna edáfica em solo suscetível à arenização na região sudoeste do Rio Grande do Sul. Revista de Ciências Agroveterinárias, v. 3, n. 2, p. 87-96, 2004.
SILVA, R. F. et al. Macrofauna invertebrada do solo sob diferentes sistemas de produção em Latossolo da Região do Cerrado. Pesquisa Agropecuária Brasileira, v. 41, n. 4, p. 697-704, 2006.
THOMAS, F. et al. Soil macrofaunal communities along an abandoned rice field chronosequence in Northern Argentina. Applied Soil Ecology, v. 27, n. 1, p. 23-29, Sep 2004. ISSN 0929-1393. Disponível em: < <Go to ISI>://WOS:000223732500003 >.
WALKER, D. Diversity and stability. In: CHERRETT, J. M. (Ed.). Ecological concepts. Oxford: Blackwell Scientific Public, 1989. p.115-146.

about:blank
1 de 1 21/12/2012 17:20


















