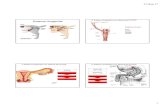
Sistema de Endomembranas
10
SISTEMA DE ENDOMEMBRANAS Engloba núcleo, retículo endoplasmático, complexo de Golgi, lisossomos e sistema de vesículas. São organelas que tiveram origem com a invaginação da membrana, por expansões e desprendimentos. NÚCLEO 1. É uma estrutura complexa com duas membranas em bicamadas lipídicas concêntricas, as membranas nucleares interna e externa, que têm composição semelhante à da membrana plasmática, com proteínas e fosfolipídios específicos. A externa é contínua ao retículo endoplasmático rugoso e compartilha as suas funções, como possuir ribossomos associados. As proteínas sintetizadas nos ribossomos da membrana nuclear externa são transportadas para o espaço entre as duas membranas, perinuclear, o qual é contínuo com o R.E. 2. As proteínas compõem estruturas complexas, os poros, compostos por mais de 100 proteínas. 3. As proteínas receptoras de lâminas, proteínas integrais, se ligam aos filamentos intermediários, lâminas nucleares. As principais dessas proteínas são LBP, LAP e emerina, que ficam na membrana interna e podem receber ajuda de proteínas adaptadoras para a associação com as lâminas. 4. As proteínas ancoradoras do poro contribuem para a estrutura do poro, como a gp210 e POM121. 5. Transporte no poro (tudo que entra e sai do núcleo é apenas pelos poros, diferente da membrana celular, que é por auxílio de proteínas ou por difusão): Passivo: moléculas de até 9 nm de diâmetro (também é facilitado por proteínas carreadoras específicas). Ativo (controlado por Ran GTP): moléculas entre 9 e 26 nm (mRNA, DNApol, RNApol...). O tráfego entre o núcleo e o citoplasma é feito pelos chamados COMPLEXOS DE PORO NUCLEAR. Oito filamentos complexos estão voltados para o citoplasma e estão envolvidos na atração das moléculas a serem
-
Upload
luis-rogerio-mueller -
Category
Documents
-
view
800 -
download
0
Transcript of Sistema de Endomembranas

SISTEMA DE ENDOMEMBRANAS
Engloba núcleo, retículo endoplasmático, complexo de Golgi, lisossomos e sistema de vesículas. São organelas que tiveram origem com a invaginação da membrana, por expansões e desprendimentos.
NÚCLEO
1. É uma estrutura complexa com duas membranas em bicamadas lipídicas concêntricas, as membranas nucleares interna e externa, que têm composição semelhante à da membrana plasmática, com proteínas e fosfolipídios específicos. A externa é contínua ao retículo endoplasmático rugoso e compartilha as suas funções, como possuir ribossomos associados. As proteínas sintetizadas nos ribossomos da membrana nuclear externa são transportadas para o espaço entre as duas membranas, perinuclear, o qual é contínuo com o R.E.
2. As proteínas compõem estruturas complexas, os poros, compostos por mais de 100 proteínas.
3. As proteínas receptoras de lâminas, proteínas integrais, se ligam aos filamentos intermediários, lâminas nucleares. As principais dessas proteínas são LBP, LAP e emerina, que ficam na membrana interna e podem receber ajuda de proteínas adaptadoras para a associação com as lâminas.
4. As proteínas ancoradoras do poro contribuem para a estrutura do poro, como a gp210 e POM121.
5. Transporte no poro (tudo que entra e sai do núcleo é apenas pelos poros, diferente da membrana celular, que é por auxílio de proteínas ou por difusão): Passivo: moléculas de até 9 nm de diâmetro (também é facilitado por
proteínas carreadoras específicas). Ativo (controlado por Ran GTP): moléculas entre 9 e 26 nm (mRNA, DNApol,
RNApol...).O tráfego entre o núcleo e o citoplasma é feito pelos chamados COMPLEXOS DE PORO NUCLEAR. Oito filamentos complexos estão voltados para o citoplasma e estão envolvidos na atração das moléculas a serem transportadas pelo poro. A classe mais abundante de proteínas do poro nuclear é conhecida como nucleoporinas (NUP’s).
6. As lâminas nucleares formam uma estrutura fibrosa que sustenta o envelope nuclear e se faz importante para a distribuição e a função dos cromossomos no núcleo, ao interagir com as proteínas receptoras e com os cromossomos na face em contato com o interior nuclear. São específicas para os tecidos, sendo que todos expressam pelo menos um tipo de lâmina B. A expressão gênica da lâmina depende do tipo de célula e do estágio de desenvolvimento. Elas adquirem uma mudança pós-traducional que as direcionam para o envelope nuclear. Esse processo envolve a adição enzimática de uma cauda de hidrocarboneto hidrofóbico a um motivo de aminoácidos. A – 7 tipos B – 3 tipos C – 2 tipos

7. IMPORTAÇÃO: Informações contidas na própria sequência de aminoácidos de uma proteína são responsáveis por definir o seu destino na célula, como a compatibilidade com a seletividade dos poros (sequências gênicas de cada proteína já indicam se ela é nuclear ou não). A sequência determinante para a passagem de uma proteína pelo poro chama-se SEQUÊNCIA CONSENSUAL (NLS – sinais de localização nuclear), rica em lisina e arginina. A IMPORTINA é responsável por reconhecer esta sequência e transportar a proteína para dentro do núcleo através dos poros. Dentro do núcleo, onde a Ran SEMPRE é fosforilada (ativada: GDP + Pᵢ GTP), esta compete pelo sítio da importina que reconhece a NLS, liberando a proteína no núcleo. O complexo Ran-importina vai para o citoplasma, onde a Ran realiza sua atividade GTPase para liberar a importina e voltar ao núcleo.
8. EXPORTAÇÃO: a EXPORTINA forma uma tríade com a Ran-GTP e com a molécula carreada, que é reconhecida pela sequência NES (sinal de exportação nuclear). Esse processo não é só utilizado para exportar mRNA, mas também proteínas que estão no núcleo e precisam ser recicladas.
9. As importinas, ao transportar uma proteína pelo poro, se ligam também à proteínas do próprio poro, cujo sítio de ligação são as repetições FG. O processo de passagem pelo poro é marcado por ligação, dissociação e religação a sequências FG adjacentes.
10. A Ran é um interruptor molecular que pode existir em dois estados conformacionais, dependendo se o GTP ou o GDP está ligado. A conversão entre os dois estados é desencadeada por duas proteínas reguladoras Ran-específicas: uma ativadora de GTPase, a GAP, citosólica, que aciona a hidrólise de GTP, convertendo Ran-GTP em Ran-GDP; e um fator de troca de guanina (GEF) nuclear, que promove a troca de GTP para GDP e, assim, converte Ran-GDP em Ran-GTP. Devido ao fato da Ran-GAP estar localizada no citosol e Ran-GEF estar no núcleo, o citosol contém principalmente Ran-GDP, e o núcleo, Ran GTP. Esse gradiente das duas formas dirige o transporte nuclear na direção apropriada.
11. Proteínas participantes de atividade no núcleo podem ter a NLS e a NES, e podem ser (des)ativadas por (des)fosforilação ou serem inibidas por proteínas citosólicas. Um sinal pode desencadear o processo de ativação dessas proteínas.
12. LAMINOPATIAS: mutação em genes das lâminas, da LAP, da LBP ou da emerina.a. Emerina mutada (EDMD): miopatia degenerativa, onde há contratura
dos músculos do calcanhar, cotovelos e pescoço. A progressão da doença leva a uma alteração cardíaca.
b. Mutação no gene da laminina A gera apenas uma leve alteração cardíaca.
RETÍCULO ENDOPLASMÁTICO
1. Sua distribuição na célula acontece de acordo com a função da mesma.2. O retículo endoplasmático liso tem muitas enzimas empenhadas no processo
de produção dos lipídios, não permitindo que haja espaço para os receptores de proteínas ribossomais.

3. Os tipos genéricos de receptores ribossomais são RIBOFORINA e TRANSLOCON.4. Os ribossomos não são fixos ao retículo endoplasmático rugoso.5. O retículo endoplasmático rugoso, geralmente, fica periférico.6. O retículo endoplasmático é ausente em células que sintetizam proteínas
citosólicas apenas para si mesmas (eritroblastos, células embrionárias, células neoplásicas de crescimento rápido). O R.E. é presente em células que sintetizam proteínas e não acumulam grânulos (fibroblastos e plasmócitos), que sintetizam proteínas e acumulam em grânulos (granulócitos, mastócitos, macrófagos) e em células que sintetizam proteínas e as secretam (células glandulares acinosa do pâncreas).
7. COMPOSIÇÃO QUÍMICA:a. Glicose-6-fosfatase: participa da glicogenólise, portanto, está presente
no músculo e nos hepatócitos.b. Glicosiltransferases OST (complexo oligossacaril-transferase): faz a
adição de monossacarídeos em lipídios e proteínas.c. Citocromo P450 e citocromo b₅: transporte de elétrons importante para
a síntese de lipídeos (colesterol e hormônios esteroides) processo de detoxificação.
d. Chaperonas (“scafolds”): durante a síntese de proteína, as chaperonas se ligam à estrutura primária para lhe dar a estrutura terciária funcional e final (adicionam ligações dissulfeto) e realizam também o controle de qualidade de proteínas. Elas atuam em processos de febre.Ex.: calnexina, calreticulina, dissulfeto-isomerase, Bip e HSP.
8. FUNÇÕES:a. Síntese de proteínas (de secreção e de composição de membrana
plasmática), participam:i. PRS
ii. Receptor de PRSiii. Sec61 – complexo de 3 proteínas que compõem a porção central
da riboforina ou transloconiv. TRAMv. TRAP
vi. Peptidase-sinal (cliva a sequência-sinal)vii. Bip – liga a Sec61, funciona como rolha.
Toda proteína de síntese no R.E. fará parte de algum componente do sistema de endomembranas ou da membrana plasmática.Todas as proteínas produzidas no citoplasma que serão secretadas ou irão compor a membrana apresentam uma sequência sinal que é reconhecida pelo PRS. Essa ligação desativa a produção de proteína pelo ribossomo. A PRS liga-se, então, ao seu receptor, que vai fazer com que a Bip se desligue da Sec61, para que o ribossomo se ligue ao translocon e sua maquinaria seja reativada. A peptidase é responsável por clivar o peptídeo sinal, que vai para o citoplasma, e a proteína vai para o lúmen do R.E. A sequência-sinal pode ser clivada pela peptidase-sinal ou pode ser extensão interna de aminoácidos e permanecer parte da proteína.Se o destino da proteína for compor membrana, ela inicalmente ficará ligada à membrana do R.E. por ter a parte hidrofóbica. A parte externa,

receptor, de uma proteína que vai compor membrana, em sua formação, fica voltada para o lúmen do R.E. (lado - ).Proteína solta dentro do retículo é expulsa da célula.
b. Síntese de lipídios – colesterol e fosfolipídios: as enzimas estão inclusas na membrana do retículo. São usados glicerol e ácidos graxos ligados à CoA.
c. Detoxificação: função do retículo contra herbicidas, desfolheantes, conservantes e corantes alimentares, medicamentos, dejetos industriais. Órgãos: fígado, rins, pele, pulmões. Funciona também fisiologicamente. Necessidade de solubilizar as toxinas para destruí-las, feito pelos citocromos (realiza redução e oxidação das substâncias).
d. Glicogenólise: manutenção da glicemia pelo fígado e contribui para a contração dos músculos estriados.
e. Regulação de Ca²⁺: o cálcio é um segundo mensageiro, amplificador de sinal, ou seja, sua concentração tem que estar sempre bem controlada, pois ativa muitas proteínas, como as caspases, que são ativadoras de apoptose.
9. CARACTERÍSTICAS:a. Proteínas solúveis (lumiais) – sequência marcadora KDEL ou HDEL: não
seguem para o complexo de Golgi, caso isso ocorra, são reconhecidas por receptores de lá e retornam ao R.E.
b. Proteínas transmembranares – sequência marcadora KKXX ou KXKXX: são mantidas no R.E. Ex.: Enzimas do processo de síntese dos lipídios.
10. DOENÇA:a. Crigler – Najjar, provocada pela deficiência da glicuronil-transferase, um
dos sistemas que atuam para alterar a molécula xenobiótica, tornando-a mais hidrofílica e permitindo assim, sua eliminação, ou seja, essas enzimas permitem que a bilirrubina solúvel seja secretada pelas células hepáticas na forma de bile. O acúmulo de bilirrubina no sangue leva a icterícia. Tratamento: administração de Barbitúricos, que estimulam a síntese dessa enzima, além de aumentar as regiões de retículo liso nas células hepáticas, devido à perda dos ribossomos adsorvidos em sua membrana. O aumento do R.E. liso contribui com a redução do efeito de certos medicamentos após uso prolongado.
COMPLEXO DE GOLGI
1. CARACTERÍSTICAS:a. Possui redes e cisternas, sendo que as redes possuem
intercomunicações.b. O pH das cisternas tende a se acidificar da rede cis para a trans.c. As proteínas contidas em vesículas passam por todas as cisternas.d. Células com maior taxa metabólica apresentam um maior número de
cisternas.e. Rede cis: próxima do R.E., vesículas cis se fusionam com o complexo.
Rede trans: saída de vesículas.

2. FUNÇÕES:a. Glicosilação, sulfatação e fosfatação.b. N-acetilglicosamina fosfotransferase: adiciona N-acetilglicosamina
fosfato ao C-6 de uma das manoses dos oligossacarídeos hidrolases.N-acetilglicosamina glicosidase: remove a N-acetilglicosamina, mas deixa o fosfato em C-6 da manose.
c. Na formação do lisossomo, a manose-6-fosfato das enzimas se liga ao MPR (Manose Phosphate Receptor) para agrupá-las, logo, todas as enzimas lisossomais possuem a manose-6-fosfato. Isso ocorre numa cisterna de pH 6,5, conferindo o mesmo ao lisossomo. O pH 6,5 é o ideial para a ligação da MPR à manose-6-fosfato, que se dissociam em pH 6, já dentro do lisossomo.
LISOSSOMO
1. Quando o lisossomo se forma, as suas enzimas ficam inativas. No caso de endocitose, o lisossomo funde-se ao endossomo primário, originando o endossomo secundário. A bomba de prótons originalmente presente no lisossomo vai bombear prótons H⁺ para o interior do endossomo, gerando o pH necessário para ativar as enzimas lisossômicas.
2. FUNÇÕES:a. Digestão celular:
i. Pinocitose específicaii. Pinocitose regulada (rafts) – conjunto de receptores que
acumulam substâncias específicas para formarem vesículasiii. Fagocitose
b. Autofagia: ligada ao R.E. Uma lamela do R.E. engloba a estrutura a ser degradada, funcionando posteriormente com o lisossomo.
c. Transcitose: por causa da sinalização, o lisossomo não reconhece o endossomo, e ele não segue a via digestiva.Ex.: IgA no leite materno e no lúmen do intestino.
3. DOENÇAS:a. Tay-Sachsb. Gaucherc. Niemann-Pickd. Doença de células I (de inclusão)e. Hipercolesterolemia familiar
OBS: CHAPERONAS NA FEBRE, EMERINA E LAMININA A NA MIOPATIA (EMBRIOGÊNESE), COMO OCORRE O PROCESSO DE DETOXIFICAÇÃO? QUAL A PARTICIPAÇÃO DA P450 E SUA REDUTASE? QUE SÃO OS BARBITÚRICOS? QUAIS SUAS FUNÇÕES? COMO AGEM?
ANOTAÇÕES THE CELL:1. As proteínas podem se mover entre os compartimentos por três caminhos,
definido pela sua sequência-sinal:c. Transporte mediado (poro);

d. Transporte transmembrana (citosol lúmen do R.E. / mitocôndria)e. Transporte vesicular (move proteínas entre compartimentos
topologicamente equivalentes).2. Áreas do R.E. liso a partir das quais vesículas carregando proteínas recém-
sintetizadas e lipídeos se desprendem para transporte até o aparelho de Golgi são chamadas de R.E. transicional, sendo proeminente em células especializadas no metabolismo de lipídios, como o colesterol gerando hormônios.
3. O hepatócito possui um R.E. liso abundante, que é o principal sítio de produção de lipoproteínas por apresentar as enzimas responsáveis por isso. O R.E. liso também contém enzimas que catalisam uma série de reações para destoxificar substâncias lipossolúveis e vários compostos danosos produzidos pelo metabolismo. As reações de destoxificação são realizadas por citocromos, como a família do citocromo P450, que catalisam uma série de reações nas quais substâncias insolúveis em água, ou metabólitos que, de outra forma, poderiam ser acumulados em níveis tóxicos, são transformadas em solúveis em água o suficiente para deixarem a célula e serem excretadas na urina.
4. No R.E., a bomba de Ca²⁺ o transporta do citosol para o lúmen do R.E. O armazenamento desse íon é facilitado pelas altas concentrações de proteínas que se ligam a ele no lúmen do R.E. Nas células musculares, o R.E. liso é modificado, chamando-se retículo sarcoplasmático. A liberação e recaptação de cálcio pelo retículo sarcoplasmático desencadeiam a contração e o relaxamento das miofibrilas.
5. A SRP é uma estrutura alongada que envolve a subunidade ribossomal maior, com uma extremidade ligada à sequência-sinal assim que o peptídeo emerge do ribossomo. A outra extremidade bloqueia o sítio de ligação do fator de alongamento na interface entre as subunidades maior e menor do ribossomo. Esse evento gera uma pausa na síntese proteica até que o ribossomo se ligue à membrana do R.E., garantindo que a proteína não seja liberada no citosol.
6. Ribossomos livres e os ligados à membrana do R.E. são estrutural e funcionalmente idênticos, a proteína que está sendo produzida em um dado momento que os difere.
7. O R.E. faz uma adição covalente de açúcares à proteína, sendo uam das suas principais funções biossintéticas. A maioria das proteínas solúveis e das proteínas ligadas à membrana que é sintetizada no R.E. são GLICOPROTEÍNAS. Os oligossacarídeos dentro da estrutura da proteína são por onde as chaperonas calreticulina e calnexinas reconhecem a molécula que precisa ser enovelada.
8. O aparelho de Golgi é um dos principais sítios de síntese de carboidratos, sendo que muitos dos oligossacarídeos são associados a proteínas e lipídios, bem como estação de classificação e de destinação para produtos do R.E.
9. Um subconjunto de oligossacarídeos serve como rótulos para direcionar proteínas específicas a vesículas que, então, as transportam para os lisossomos. Eles também podem ser reconhecidos de outras maneiras, tendo outros destinos.

10. O citocromo apresenta o íon ferro no grupo heme, que é responsável pela capacidade de transferência de elétrons, podendo alternar entre os estados de oxidação Fe²⁺ e Fe³⁺. O citocromo P450 é responsável pela transferência de elétrons do NADPH⁺ + H⁺ para o oxigênio e pela transferência de grupamento OH para o substrato. O citocromo P450, em suas reações de oxidação e redução na detoxificação, tornam substâncias lipossolúveis em hidrossolúveis. Essa alteração da substância, consequentemente, dificulta a difusão da substância pela membrana plasmática, que é hidrofóbica, assim como a reabsorção das substâncias nos túbulos renais, acelerando a eliminação das substâncias. Somando-se a isso, a alteração realizada pelo citocromo na substância pode modificá-la de tal forma que altera a interação dela com biorreceptores.
11. Os lisossomos são compartimentos definidios por membranas preenchidos por enzimas hidrolíticas, todas hidrolases ácidas, como proteases, nucleases, glicosidases, lipases, fosfolipases, que controlam a digestão intracelular de macromoléculas. As enzimas precisam de um pH em torno de 4,5 e 5 para uma ótima atividade. Uma H⁺ ATPase na membrana lisossomal utiliza a energia da hidrólise de ATP para bombear H⁺ para dentro do lisossomo, diminuindo o pH do seu lúmen.
12. CHAPERONAS: a síntese de HSP (proteínas de choque térmico) a partir do DNA é aumentada em caso de estresse térmico ou químico que acarreta em alteração da conformação correta das proteínas, agindo em resposta ao choque térmico (HSR). O fator de transcrição desse gene é ativado no caso de estresse. A HSP atua então como chaperona, garantindo que a montagem da estrutura final da proteína seja completa e correta, conferindo-lhe a estrutura terciária. Os níveis aumentados de proteínas do estresse dão a célula meios para: a) identificar e talvez facilitar redobramento de proteínas afetadas de modo adverso pelo estresse metabólico; b) identificar e fazer ligação com proteínas anormalmente dobradas, de modo que estas sejam marcadas e enviadas a um sistema proteolítico adequado, assim facilitando a eliminação das proteínas defeituosas; c) facilitar a síntese e a maturação de novas proteínas, que irão substituir aquelas destruídas no estresse metabólico.