Tema: Direção da síntese de DNA, RNA e proteínas.
44
Seminário de Bioquímica II Prof. Dr. Júlio C. Borges Alunos: José Augusto M. Burgarelli Rafael da Fonseca Lameiro Seiti Inoue Venturini Tema: Direção da síntese de polímeros biomoleculares DNA mRNA Proteína
Transcript of Tema: Direção da síntese de DNA, RNA e proteínas.

Seminário de Bioquímica IIProf. Dr. Júlio C. Borges
Alunos:
José Augusto M. Burgarelli
Rafael da Fonseca Lameiro
Seiti Inoue Venturini
Tema: Direção da síntese de polímeros
biomolecularesDNA mRNA Proteína

TÓPICOS• Síntese de DNA: replicação
• Origem, direção e final da replicação
• Replicação reversa
• Síntese de RNA: transcrição
• Direção da transcrição de RNA
• Transcrição do RNA
• Comprovação da direção de transcrição
• Síntese de proteínas: tradução
• Direção da tradução (eucariotos)
• Direção da síntese proteica (eucariotos e procariotos)

SÍNTESE DE DNA


ORIGEM DA REPLICAÇÃO

ORIGEM DA REPLICAÇÃO
• John Cairns: estudos com DNA circular de E. coli. Durante a replicação há formação de um loop. Mostrou que ambas as extremidades do loop são replicantes: a síntese é bidirecional.

ORIGEM DA REPLICAÇÃO
• Ross Inman: Mostrou que em E. coli os loops de replicação sempre começam em um ponto, denominado origem (oriC). Essa região consiste de 245 pb e é rica em A e T.
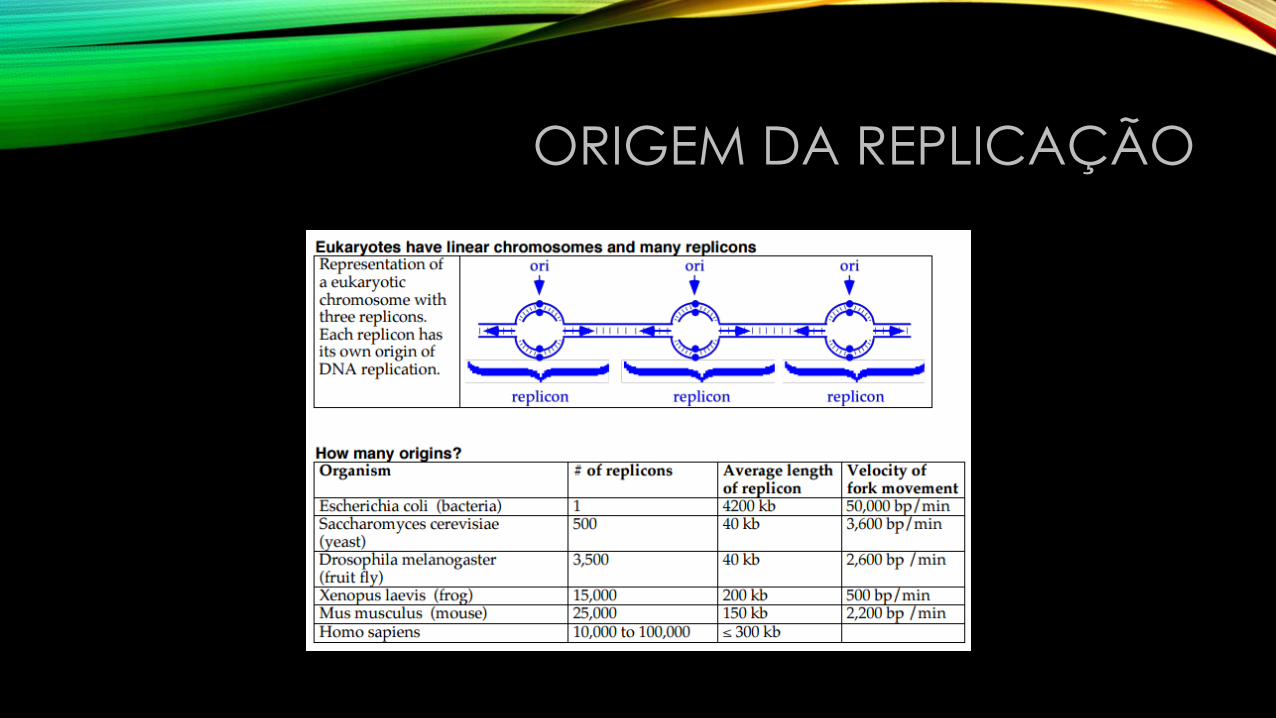
ORIGEM DA REPLICAÇÃO
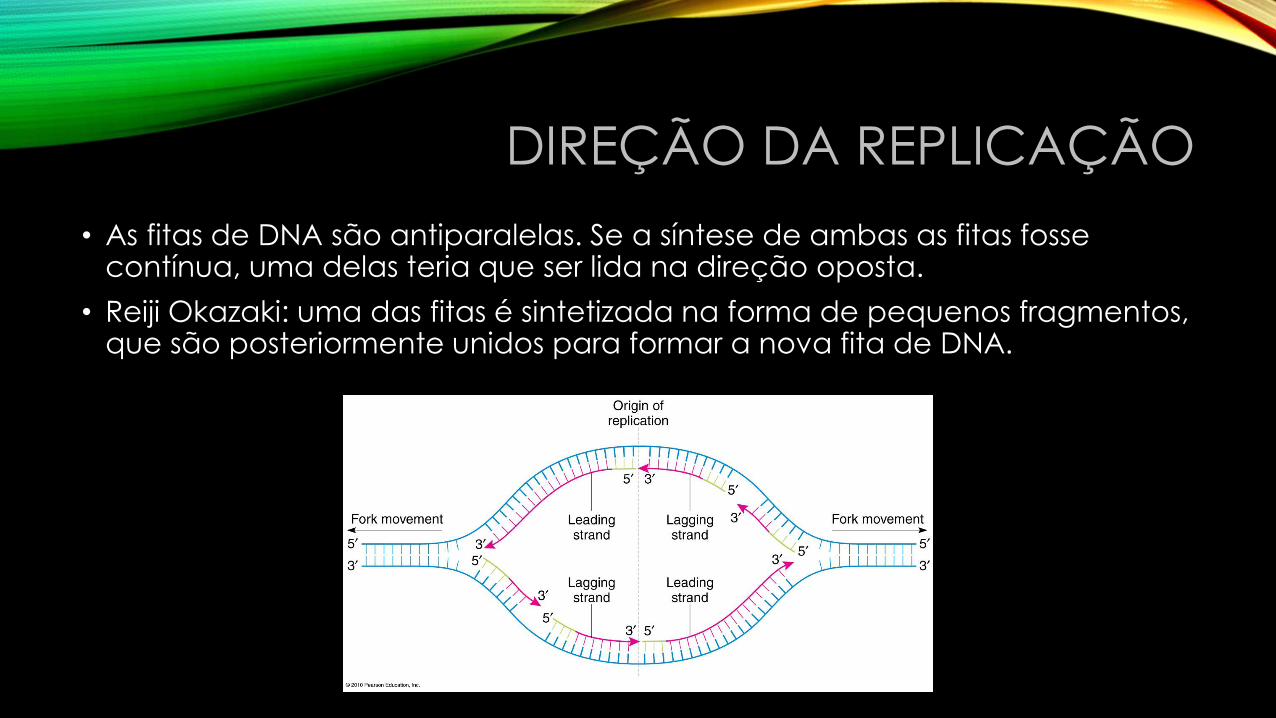
DIREÇÃO DA REPLICAÇÃO
• As fitas de DNA são antiparalelas. Se a síntese de ambas as fitas fosse contínua, uma delas teria que ser lida na direção oposta.
• Reiji Okazaki: uma das fitas é sintetizada na forma de pequenos fragmentos, que são posteriormente unidos para formar a nova fita de DNA.

DIREÇÃO DA REPLICAÇÃO
• Mecanismo de elongação da cadeia: Reação de desidratação que une o grupo fosfato do dNTP ao grupo hidroxila do monômero terminal da cadeia.
• O nucleófilo é o 3’-OH do nucleotídeo da terminação 3’ do DNA, que faz o ataque ao α-fósforo do 5’-desoxinucleotídeo trifosfato (dNTP). Grupo de saída = pirofosfato inorgânico (PPi), posteriormente clivado.
• Nesse mecanismo, dois cátions metálicos divalentes (Mg2+) facilitam a transferência do par de elétrons do OH livre ao α-fosfato.

DIREÇÃO DA REPLICAÇÃO
• Mecanismo da síntese. O grupo OH livre do carbono 3’ é o ponto de elongação da cadeia.


FINAL DA REPLICAÇÃO
• Durante a elongação da cadeia, a polimerase realiza uma função de checagem (proofreading). Caso o último nucleotídeo adicionado não corresponda ao determinado pela fita molde, ele é removido. No caso da síntese no sentido 5’3’, esse não é o final da replicação.
• A enzima DNA ligase deve unir os fragmentos de Okazaki. Isso é feito às custas de ATP.

INVERSÃO DO SENTIDO DA REPLICAÇÃO
• Parece ter havido uma preferência evolutiva pela síntese 5’3’.

INVERSÃO DO SENTIDO DA REPLICAÇÃO
• Primeira hipótese: acaso. No início haviam proto formas de vida que realizavam replicação em ambos os sentidos. Parte dessa população foi eliminada, restando apenas uma fração de espécies que se reproduziram, evoluíram e deram origem aos outros seres vivos.
• Segunda hipótese: impossibilidade química. A reação no sentido inverso simplesmente não pode ser realizada pelas enzimas.
• A segunda hipótese é facilmente descartada: a catálise promovida pela
polimerase envolve apenas a hidroxila 3’ e o α-fosfato, não importando em
que espécies eles se localizam (DNA ou dNTP).
De que maneiras a síntese no sentido 3’5’ seria possível?

INVERSÃO DO SENTIDO DA REPLICAÇÃO
1.Usando como substratos nucleotídeos com 3’-trifosfato. Essa situação não pode ocorrer: não são comumente observados nucleotídeos desse tipo porque se decompõem rapidamente.

INVERSÃO DO SENTIDO DA REPLICAÇÃO
2. Envolvendo o ataque do OH 3’ do dNTP de entrada ao α-fósforo 5’ da cadeia polinucleotídica. Para que esse mecanismo fosse possível, a extremidade da cadeia polinucleotídica precisaria apresentar na extremidade um nucleotídeo com di ou trifosfato.

INVERSÃO DO SENTIDO DA REPLICAÇÃO
• Uma vez que a síntese é possível, deve ser formulada uma terceira hipótese: deve haver alguma vantagem evolutiva no sentido 5’3’.
• Revisão (proofreading): a retirada de um nucleotídeo inserido incorretamente não prejudica a síntese 5’3’, mas interromperia a 3’5’.
• Além do mecanismo de correção, é importante ressaltar que trifosfatos se
hidrolisam espontaneamente, o que também comprometeria a síntese.


INVERSÃO DO SENTIDO DA REPLICAÇÃO
• Simulação de competição entre espécies com polimerases 5’3’ (forward) e 3’5’ (reverse) em diferentes temperaturas.
• Esperado: em temperaturas maiores, as reverse devem ser desfavorecidas (hidrólise mais frequente do trifosfato do DNA, favorecendo polimerases mais rápidas que ocasionariam maior taxa de mutação e, consequentemente, menor fidelidade de replicação).
• De fato observa-se que em ambientes competitivos e com temperaturas maiores, a replicação no sentido 5’3’ é a estratégia evolutivamente estável.

SÍNTESE DE RNA

DIREÇÃO DA SÍNTESE DE RNA
• Sentido da direção da transcrição:
5´ → 3´
• A RNA polimerase faz a leitura da fita molde no sentido 3´ → 5´ acabando por sintetizar um RNA complementar semelhante ao da fita complementar
de DNA. A diferença é que no RNA a base nitrogenada timina (T) é substituída pela uracila (U).

DIREÇÃO DA SÍNTESE DE RNA

DIREÇÃO DA SÍNTESE DE RNA
Fita de DNA não molde
(codificadora)
Fita de DNA molde
Transcrito de RNA

DIREÇÃO DA SÍNTESE DE RNA
Alongamento da cadeia de RNA pela RNA-polimerase

DIREÇÃO DA SÍNTESE DE RNA
A reação não toma sentido contrário ao movimento 5´ → 3´ em momento algum mas pode parar quando:
• Um nucleotídeo recém adicionado na extremidade 3´ é adicionado de forma a parear indevidamente no processo de transcrição.
• A remoção é feita por reversão direta da reação da RNA polimerase;
• Mas esse mecanismo não é bem elucidado.

DIREÇÃO DA SÍNTESE DE RNA
Como se descobriu que a síntese de RNA é feita no sentido 5´ → 3´ ?
• R: Através de marcação do fosfato γ do GTP.
• Observação da taxa da radiação que é incorporado ao RNA;
αβγ

DIREÇÃO DA SÍNTESE DE RNA

DIREÇÃO DA SÍNTESE DE RNA• Outras evidências do crescimento 5´ → 3´
Adenosina TrifosfatoAntibiótico contra bactérias

DIREÇÃO DA SÍNTESE DE RNA
A informação a ser codificada no RNA tem início e fim:
• As sequências regulatórias marcam o início e o fim da transcrição.
• São sequências específicas de DNA denominadas promotores.
• As sequências de consenso, o espaçamento entre elas e sua distância até o sítio de transcrição determinam o quão eficientemente a RNA polimerase se liga a essas e inicia a transcrição.
• Mutações nos pares de base dessa região consenso afetam a transcrição.
• Caso não existissem pontos de início da transcrição, ou seja de forma aleatória, isso traria um grande desperdício de energia.

SÍNTESE DE PROTEÍNAS

POR QUE A LEITURA DA TRADUÇÃO OCORRE NO SENTIDO 5` 3`???

DIREÇÃO DA TRADUÇÃO (EUCARIOTOS)
CAP (ou quepe) e Cauda Poli(A) (Eucariotos)

DIREÇÃO DA TRADUÇÃO (EUCARIOTOS)
O que é o CAP?

Complexo proteico que envolve o CAP
DIREÇÃO DA TRADUÇÃO (EUCARIOTOS)

Estrutura do ribossomo
DIREÇÃO DA TRADUÇÃO (EUCARIOTOS)

Interação: ribossomo – complexo proteico
DIREÇÃO DA TRADUÇÃO (EUCARIOTOS)

CURIOSIDADE: É possível o ribossomo acoplar-se em outro
local do mRNA que não ao complexo proteico do CAP ?
DIREÇÃO DA TRADUÇÃO (EUCARIOTOS)
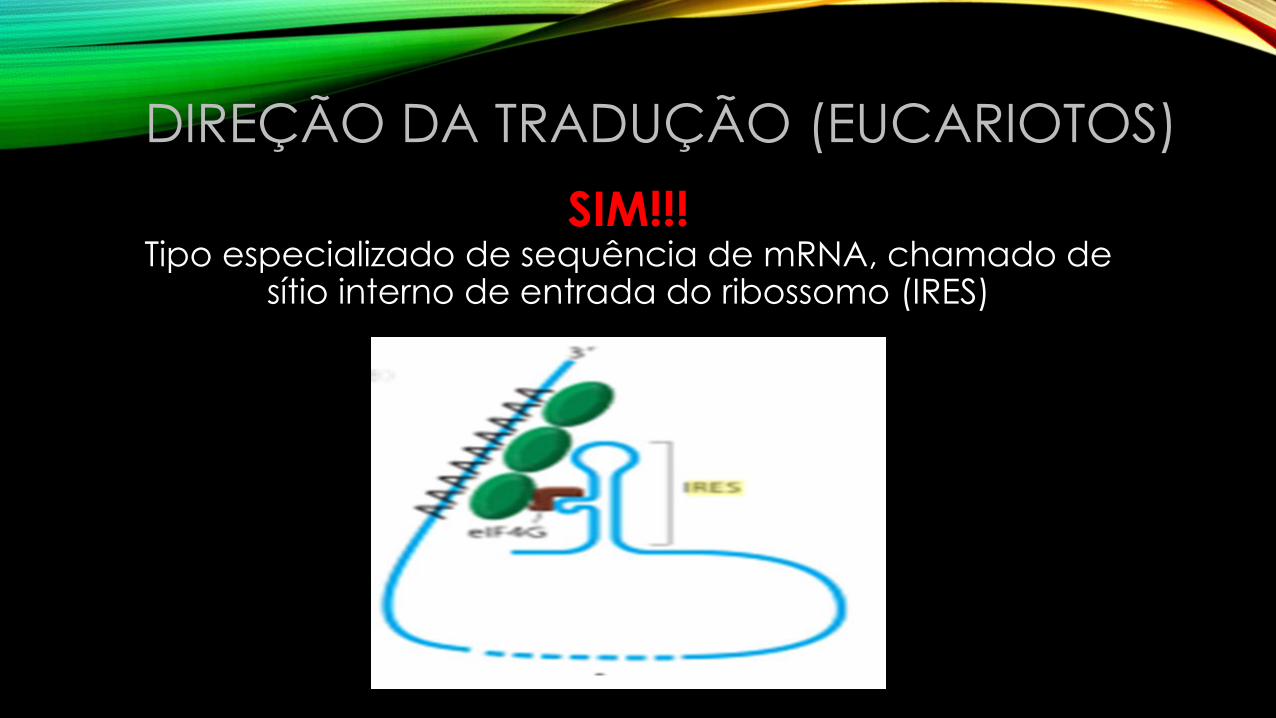
SIM!!!Tipo especializado de sequência de mRNA, chamado de
sítio interno de entrada do ribossomo (IRES)
DIREÇÃO DA TRADUÇÃO (EUCARIOTOS)

Agora que descobrimos o porquê do sentido 5` 3`; por que a proteína é sintetizada no sentido
N terminal C terminal?
DIREÇÃO DA SÍNTESE PROTEICA

Interação tRNA – aminoácido
Peptidil-transferase
DIREÇÃO DA SÍNTESE PROTEICA

Síntese de proteínas 5`3` (procariotos)
1- Subunidade ribossomal (30S) se liga a dois fatores de iniciação,
IF-1 e IF-3, o mRNA estão se liga a essa subunidade. O 5´AUG
iniciador é guiado até sua posição correta pela sequencia de
Shine-Dalgarno.
2- O complexo constituído da subunidade 30S do ribossomo, do
IF-3 e do mRNA se junta ao IF-2ligado ao GTP e ao fMet-RNA-tfmet
iniciador. O anticódon desse RNA-t pareia então corretamente
com o códon iniciador do mRNA.
3- Esse grande complexo se combina com a subunidade 50S do
ribossomo, simultaneamente, o GTP liberado ao IF-2 é hidrolisado
a GDP e Pi, os quais são liberados do complexo. Nesse ponto,
todos os três fatores de iniciação saem do ribossomo, dando
origem ao ribossomo funcional 70S.
DIREÇÃO DA SÍNTESE PROTEICA

REFERÊNCIAS
GENETICA ON LINE. AULA 4 BIOLOGIA – PROCESSAMENTO. Disponível em:
<http://aprendendogenetica.blogspot.com.br/2011/04/aula-4-biologia-processamento.html>. Acesso em 23 nov. 2015.
ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K.; WALTER, P. Biologia Molecular da Célula. São Paulo: Artmed, 2009.
1728 p.
VOET, D.; VOET, J, G. Bioquímica. São Paulo: Artmed, 2013. 1504 p.
NELSON, D, L.; COX, M, M. Princípios de Bioquímica de Lehniger. São Paulo: Artmed, 2014. 1328 p.
JACKMAN, J. E.; GOTT, J. M.; GRAY, M. W. Doing it in reverse: 3'-to-5' polymerization by the Thg1 superfamily. RNA, v. 18, n.5, p.
886 – 899, 2012.
BALLANCO, J.; MANSFIELD, M. L. A Model for the Evolution of Nucleotide Polymerase Directionality. PLoS One, v. 6, n. 4, 2011.

OBRIGADO !