
Teorias de crescimento populacional segundo dr Ivomboa
26
Teorias de crescimento populacional Tema
-
Upload
ivaristo-americo -
Category
Education
-
view
573 -
download
3
Transcript of Teorias de crescimento populacional segundo dr Ivomboa

Teorias de crescimento populacional
Tema

Introdução
As teorias de crescimento da população são teorias que vem para
explicar como uma determinada população pode crescer
positivamente ou negativamente, sendo influenciados pelos
factores limitantes, assim como não num determinado
ecossistema.
Deste modo define-se “teoria como sendo uma hipótese que foi
completamente comprovada utilizando-se métodos científicos
validos” (Berryman, 2003).
Estas teorias também explicam o crescimento das populações,
quanto a densidade, ao seu nível trófico, as relações
interspecificas, massa, reprodução, a relação com as leis da
física, da matemática, etc.

Categorias de Crescimento Populacional
Existem categorias que explicam resumidamente as teorias de
crescimento populacional. Essas categorias podem ser:
Baseada nas leis de alguns cientistas;
Tamanho ou massa corporal;
Factores limitantes;
Factores bioticos;
Interações populacionais;
Feedback negativos;
Tempos de geração.
etc.

Teoria Maltusiana
A teoria Maltusiana descreve a situação padrão das populações, como se comportam na ausência de quaisquer factores perturbadores.
Aqui as taxas de nascimento e morte são constantes, a população irá crescer (ou cair) em progressão geométrica.
A teoria exerce uma função na ecologia semelhante à primeira lei de Newton da física.
Aristóteles afirmou que o estado normal de todos os objectos era o repouso e que o movimento ocorria apenas quando uma força era aplicada a um objecto.
Newton, entretanto mostrou que o movimento uniforme era o estado normal e que o movimento não uniforme e o repouso geralmente ocorriam apenas quando uma força era aplicada a um objecto.

Teoria Maltusiana
A teoria Maltusiana acabou afirmando que o estado normal de
uma população não é o repouso (isto é, uma população
constante), mas o movimento (isto é, crescimento ou declínio
exponencial), e que, quando as populações não crescem ou
declinam exponencialmente, é por causa de uma força externa
(algo no ambiente) que altera os índices de nascimentos e de
mortes.
Esta força externa (ambiental) pode ser um factor abiótico ou
biótico como “o grau de aglomeramento conespecífico e
densidades de todas as outras espécies na comunidade que
poderiam interagir.
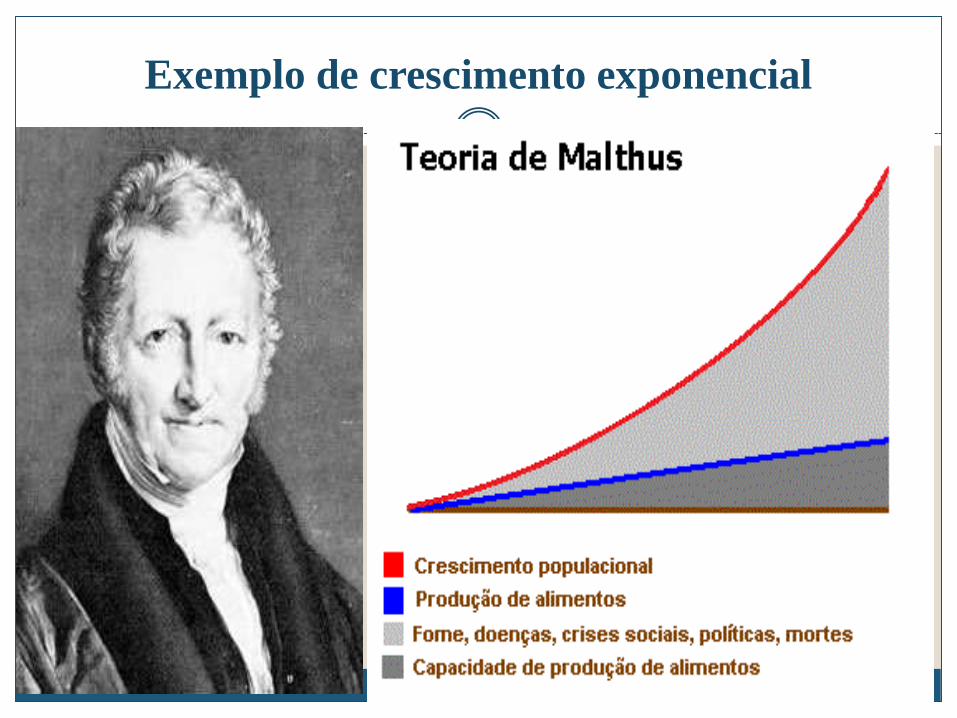
Exemplo de crescimento exponencial

Teoria de Allee
A teoria de Allee, dita que existe um relacionamento
positivo entre a aptidão individual e os números ou
densidade dos conespecíficos.
Entretanto “à medida que o número de indivíduos de uma
população aumenta, ou à medida que a densidade
populacional aumenta, a sobrevivência e a produção
também crescem”. (Berryman, 1999).
Um bom exemplo ocorre quando os animais se agregam
em grupos para protecção e, desta forma, diluem a
ameaça que cada indivíduo sofre de ser atacado por um
predador.

Teoria de Allee
O maior número de conespecíficos beneficia a população porque
aumenta a diluição ou saturação do predador e assim
proporciona baixas densidades de crescimento populacionais no
caso de um decresço na de uma certa população.
Por exemplo, quando o tamanho da população de uma planta
polinizada por insectos torna-se baixo, ou se um pequeno número
de indivíduos floresce durante um ano, menos sementes serão
produzidas por planta porque os insectos polinizadores terão mais
dificuldade para encontrar poucas flores.
Pelo facto de as pequenas populações apresentarem uma menor
reprodução ou sobrevivência, a de Allee é de interesse especial
aos ecologistas que trabalham com espécies ameaçadas de
extinção.

Teoria de Allen

Teoria de Verhulst
O crescimento populacional pode atingir um limite superior acima do qual a densidade populacional não pode mais aumentar. Entretanto vários factores diferentes podem limitar uma população tais como: predadores, doenças, níveis de recursos e a competição.
Esta teoria, preocupa-se apenas com um factor: a competição intra-específica (a competição entre os membros da mesma espécie).
Pelo facto de os organismos que limitam a população também serem membros da população (auto-limitação populacional).
Assim sendo a Teoria de Verhulst propõe que, “em algum momento, o índice de crescimento per capita de uma população é limitado directa e imediatamente por sua própria densidade, através do processo de competição intra-específica” (Berryman, 1999).

Teoria de Verhulst
Os mecanismos de competição intra-específica, que aumentam
com a densidade populacional crescente e actuam para finalmente
limitar o crescimento da população:
A agressão intra-específica;
A territorialidade;
o canibalismo;
Entre outros.
Esses mecanismos aumentam com a crescente densidade
populacional porque os indivíduos lutam para ocupar a quantidade
insuficiente de espaço disponível no momento, espaço este
necessário para reunir recursos, esconder-se ou escapar dos
inimigos.

Competição intra-específica

Teoria de Lotka-Volterra
Os organismos interagem com outras espécies e com o ambiente
físico de várias maneiras.
Essas interacções às vezes incluem “feedbacks negativos”.
Um exemplo de feedback negativo é quando um aumento na
população de uma espécie de presa gera um aumento na
população de seus predadores (através do aumento da
reprodução), e isso, por sua vez, retroalimenta para reduzir a
população de presas através do aumento da mortalidade devido à
predação.

O aumento populacional de leões pode levar a um feed
negativos de populações de Zebras

Teoria de Liebig
Uma dada população geralmente irá interagir com diferentes espécies de presas e predadores, através das teia alimentares, também mesmo os factores abióticos distintos fora da teia alimentar são suficientemente nem para fazer o controle do crescimento de uma dada população.
A experiência mostra que apenas uma ou duas outras espécies dominam a estrutura de feedback de uma população em qualquer dado momento e local.
A identidade dessas espécies dominantes pode mudar com o tempo e o local, mas o número de espécies que limitam uma dada população (isto é, controlam activamente sua dinâmica) é geralmente apenas um ou dois.

Teoria de Liebig
A teoria de Liebig, em sua forma moderna, expressa esta ideia. Propõe que, entre todos os factores bióticos ou abióticos que controlam uma dada população, um deve ser limitante (isto é, activo, controlador da dinâmica).
Os atrasos produzidos por este factor limitante geralmente duram por uma ou duas gerações (Berryman, 2003).
Tudo que existe na natureza está relacionado, portanto, uma mudança na abundância de um organismo afectará a abundância de todos os outros.
Embora seja verdade isso pode induzir a erro, porque novas pesquisas demonstram que apenas algumas das várias espécies nas interacções relativamente poucas serrão limitadas o seu crescimento populacional em um determinado tempo e local (Berryman, 2003).

Teoria de Fenchel
O crescimento populacional exponencial é relacionado ao tamanho (massa) do corpo.
As espécies com maior tamanho corporal têm menores índices de crescimento populacional.
O índice máximo de reprodução cai com o tamanho do corpo a uma potência de aproximadamente 1/4 da massa corporal.
A expressão matemática é dada pela formula:
Onde:
r é o índice intrínseco de aumento natural da população;
a é uma constante que possui 3 valores diferentes (um para organismos unicelulares, um para heterotermos e outro para homeotermos)
W é o peso (massa) corporal médio do organismo.
r = aW-1/4

Teoria de Calder
Os períodos de oscilação nas populações de mamíferos herbívoros são relacionados ao tamanho do corpo (massa).
As espécies com maior tamanho corporal geralmente têm ciclos populacionais mais longos.
A duração do ciclo populacional aumenta com o aumento do tamanho do corpo a uma potência de aproximadamente 1/4 da massa corporal.
A expressão matemática é dada pela formula:
Onde:
t é o tempo médio do ciclo populacional;
a é uma constante;
W é o peso (massa) corporal médio do organismo.
t = aW1/4

Teoria de Calder
Antes da pesquisa de Calder, sabia-se que os pequenos mamíferos
herbívoros como os lemingues (Lemmus) e arganazes (Microtus)
tinham ciclos populacionais de 3 a 4 anos.
Enquanto os da lebre (Lepus americanus) com corpos maiores
eram de 8 a 10 anos, e os do ainda maior alce (Alces alces) e da
rena (Rangifer tarandus) eram de 20 a 40 anos.
Entretanto, antes de Calder, ninguém havia indicado a correlação
entre o grande tamanho corporal e os ciclos populacionais mais
longos.

Teoria de Damuth
A densidade populacional é relacionada ao tamanho (massa) do
corpo.
As espécies com maior tamanho corporal geralmente têm
densidades populacionais médias mais baixas.
A densidade média de uma população cai com o tamanho do
corpo a uma potência de aproximadamente 3/4 da massa corporal.
A teoria de Damuth é dada pela formula:
Onde:
d é a densidade média da população;
a é uma constante;
W é o peso (massa) corporal médio do organismo.
d = aW-3/4

Exemplo do Elefante
As populações de
elefantes são
relativamente
medias, devido a
relação tamanho
ou massa do corpo
de Damuth a uma
potência de
aproximadamente
3/4 de massa
corporal.

Teoria de tempo de geração
O tempo de geração é o período necessário para que um individuo possa crescer e amadurecer até a idade reprodutiva ( relacionado ao tamanho do corpo).
As espécies com maior tamanho corporal geralmente têm tempos de geração mais longos.
O tempo de geração aumenta com o tamanho do corpo a uma potência de aproximadamente 1/4 da massa corporal do organismo no momento da reprodução.
A teoria é expressa pela seguinte equação:
Onde:
g é o tempo de geração médio da população;
a é uma constante;
W é o peso (massa) corporal médio do organismo.
g = aW1/4

Teoria de Ginzburg
A transferência de qualidade do efeito materno influencia o crescimento populacional e em qualquer ponto do tempo depende não apenas do ambiente actual, mas também do ambiente da geração anterior (Ginzburg & Colyvan, 2004).
Acredita-se que a alteração ambiental do índice de crescimento populacional per capita ocorra pela modificação do índice de mudança deste índice de crescimento, e não pela alteração directa do índice de crescimento per capita.
A duração de um ciclo populacional (oscilação) é o resultado do efeito maternal e do crescimento populacional inercial, essas durações da população devem ser de duas, seis ou mais gerações (Ginzburg & Colyvan, 2004).

Teoria de Ginzburg
Os predadores ou os outros factores ambientais podem ser a causa do ciclo populacional.
Estes podem afectar a amplitude, o formato do ciclo e a duração do período do ciclo que pode ser é específico à espécie e não depende da causa.
Este período de ciclo específico da espécie é denominado eigenperiod.
A eigenperiod, explica por que espécies semelhantes têm períodos de ciclo similares, muito embora possam habitar em ambientes muito diferentes, mesmo sendo atacadas por predadores diferentes e em algumas situações como nas ilhas, onde não haver predadores.
Esta teoria explica por que os períodos de ciclo populacional de 3 a 5 gerações são desconhecidos na natureza.

Conclusão
Finalmente entende-se que as teorias de crescimento vem explicar como as diversas oscilações populacoes verificadas num dado ecossistemas.
Embora abundâncias máximas possam ocorrer simultaneamente em grandes áreas, assim como também algumas populações estão em via de extinção.
Portanto estas teorias vieram explicar e esclarecer o crescimento populacional baseando-se alguns cientes em teorias que desenvolveram em outaras ciencias.

Obrigado pela atenção
Deixamos este momento para duvidas e subsídios.
Fim
5o grupo
Boaventura Benzane
Graça Ana Banze
Ivaristo Américo Mboa Júnior
Miqueias Chauque
Quinito Maurício Mussacate


















