
UNIVERSIDADE CEUMA-UNICEUMA PRÓ-REITORIA DE PÓS … Jos... · 5 AGRADECIMENTOS Ao único que é...
98
UNIVERSIDADE CEUMA-UNICEUMA PRÓ-REITORIA DE PÓS-GRADUAÇÃO, PESQUISA E EXTENSÃO PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA PARASITÁRIA SAULO JOSÉ FIGUEIREDO MENDES EFEITO IMUNOMODULADOR DO CINAMALDEÍDO NO TRATAMENTO DA SEPSE INDUZIDA EM CAMUNDONGOS São Luís 2015
Transcript of UNIVERSIDADE CEUMA-UNICEUMA PRÓ-REITORIA DE PÓS … Jos... · 5 AGRADECIMENTOS Ao único que é...

1
UNIVERSIDADE CEUMA-UNICEUMA
PRÓ-REITORIA DE PÓS-GRADUAÇÃO, PESQUISA E EXTENSÃO
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA PARASITÁRIA
SAULO JOSÉ FIGUEIREDO MENDES
EFEITO IMUNOMODULADOR DO CINAMALDEÍDO NO TRATAMENTO DA
SEPSE INDUZIDA EM CAMUNDONGOS
São Luís
2015

2
SAULO JOSÉ FIGUEIREDO MENDES
Efeito imunomodulador do cinamaldeído no tratamento da sepse induzida em
camundongos
Dissertação apresentada ao Programa de Pós-Graduação em Biologia Parasitária como parte dos requisitos para a obtenção do título de Mestre em Biologia Parasitária. Orientador: Prof(a). Dr(a). – Elizabeth Soares Fernandes Co-orientador: Prof. Dr. – Marcos Augusto Grigolin Grisotto. Prof. Dr. – Robson da Costa.
São Luís
2015

3
SAULO JOSÉ FIGUEIREDO MENDES
Efeito imunomodulador do cinamaldeído no tratamento da sepse induzida em
camundongos
Dissertação apresentada ao Programa de Pós-Graduação em Biologia Parasitária como parte dos requisitos para a obtenção do título de Mestre em Biologia Parasitária.
A Comissão julgadora da Defesa do Trabalho Final de Mestrado em
Biologia Parasitária, em sessão pública realizada no dia / / ,
considerou o candidato:
( ) APROVADO ( ) REPROVADO
1) Examinador __________________________________
2) Examinador ___________________________________
3) Examinador ___________________________________
4) Presidente (Orientador)__________________________________

4
A minha mãe, mulher de oração, pelo seu
apoio, intercessão, dedicação e amor
incondicional.

5
AGRADECIMENTOS
Ao único que é digno de receber a honra e a glória, a força e o poder, ao rei eterno e
imortal, invisível, mas real. Coroamos a ti, ó rei JESUS!
A minha Mãe, minha companheira, pelo apoio, conselhos, oração, exortação e amor
incondicional. Obrigado é muito pouco, presente não é tudo. Mas, o reconhecimento,
isso! Sim, é pra valer. Te amo!
Ao meu Pai, exemplo de seriedade, honestidade, respeito e profissionalismo.
Obrigado por tudo.
Ao meu irmão Sólon Uchôa Lago Netto, pelo apoio em todos os momentos da minha
vida. Meu muito Obrigado!
Aos meus familiares pelas orações e incentivo.
A minha orientadora, Dr (a). Elizabeth Soares Fernandes, pelo exemplo de paciência
e dedicação na orientação ao trabalho. Obrigado pelas oportunidades a mim
concedidas, por muitas vezes ser como uma mãe, pelos conselhos, preocupação e
palavras de incentivo. Fica pequeno apenas descrever algumas palavras diante da
minha tamanha admiração por seu profissionalismo, competência e inteligência
afinada. Minha eterna gratidão.
Ao meu co-orientador, Professor Dr. Marcos Augusto Grigolin Grisotto, por ter me
ajudado em todos os momentos da pesquisa.
Ao Professor, Dr. José Luiz Martins do Nascimento, por abrir as portas do seu
laboratório na UFPA para que eu pudesse realizar alguns experimentos.
A Fundação de Amparo a Pesquisa do Maranhão (FAPEMA) pela bolsa de mestrado
concedida e pelo financiamento do projeto assim como à CAPES e CNPq.
A Universidade Ceuma, por disponibilizar toda a estrutura e equipamentos
necessários para a realização deste trabalho.
Aos funcionários do biotério da Universidade Ceuma, em especial a pessoa do Seu
Edson pelo apoio e amizade nessa jornada.
Aos colegas da turma XI do mestrado de Biologia Parasitária, pelo convívio divertido
e harmonioso em que o companheirismo foi soberano a qualquer diferença nesta
caminhada.
A todos os amigos de laboratório de Imunologia que colaboraram para a realização
deste trabalho.

6
“Que darei eu ao Senhor, por todos os
benefícios que me tem feito?”
Salmos 116:12

7
RESUMO
A sepse pode ser definida como um conjunto de reações do organismo causada
pela resposta inflamatória sistêmica descontrolada do indivíduo em decorrência de
uma infecção. Na medicina tradicional chinesa, a canela é indicada como
analgésico, antipirético e para mialgia. Os principais óleos essenciais da casca da
canela são o eugenol e o cinamaldeido (CNA). O receptor TRPA1 foi identificado na
ultima década como o alvo farmacológico do CNA no organismo. Assim, o objetivo
deste trabalho foi avaliar o potencial terapêutico do cinamaldeído na resposta
inflamatória causada pela sepse induzida por lipopolissacarídeo em camundongos.
A sepse foi causada por administração de LPS e foi usado o pré-tratamento com
CNA e/ou HC-030031. Foram avaliados parâmetros locais, circulantes e sistêmicos
da sepse. A administração de CNA atenua a falência hepática além de alterar o
perfil de células inflamatórias circulantes e também presentes no lavado peritoneal
de animais sépticos, ainda o CNA promove o aumento da viabilidade de células
inflamatórias peritoneais como as Ly6C+ e CD8+ o qual é acompanhado por uma
redução no número absoluto de células PMNs. O CNA ainda, promove a diminuição
nos níveis de NO no lavado peritoneal, diminuição dos níveis circulantes de IL-1β e
aumento dos níveis circulantes de IL-10. O antagonismo do TRPA1 pelo tratamento
com HC-030031 é capaz de reverter alguns dos efeitos do CNA, como, o perfil de
células inflamatórias circulantes e do lavado peritoneal (Ly6C+ e CD8+), além de
reduzir os níveis circulantes de IL-10. Em conjunto, os dados do presente trabalho
sugerem que o CNA é capaz de modular a sepse por mecanismos dependentes e
independentes da ativação do TRPA1.
Palavras–Chaves: Sepse, cinamaldeído, TRPA1.

8
ABSTRACT
Sepsis can be defined as a set of body reactions caused by uncontrolled systemic
inflammatory response of the individual as a result of an infection. In traditional
Chinese medicine, cinnamon is indicated as an analgesic, antipyretic and myalgia.
The main essential oils of cinnamon bark are eugenol and cinnamaldehyde (CNA).
The TRPA1 receptor was identified in the last decade as the pharmacological target
of the CNA in the body. The objective of this study was to evaluate the therapeutic
potential of cinnamaldehyde in the inflammatory response to sepsis induced by
lipopolysaccharide in mice. Sepsis is caused by administration of LPS and was used
pre-treatment with CNA and/or HC-030031. We evaluated local parameters, current
and systemic sepsis. The administration of CNA attenuates liver failure in addition to
changing the profile of circulating inflammatory cells and also present in peritoneal
fluid of septic animals, although the CNA promotes increased viability of peritoneal
inflammatory cells such as Ly6C+ and CD8+ which is accompanied by a reduction in
absolute number of PMN cells. The CNA also promotes the reduction of NO levels in
peritoneal lavage, reduction of circulating levels of IL-1β and increased circulating
levels of IL-10. Antagonism of TRPA1 by treatment with HC-030031 is able to
reverse some of the effects of the CNA, as the profile of circulating inflammatory cells
and peritoneal lavage (Ly6C+ and CD8+), while reducing circulating levels of IL-10.
Taken together, the present data suggest that CNA can modulate sepsis by
mechanisms dependent and independent of TRPA1 activation.
Key Words: Sepsis, cinnamaldehyde, TRPA1.

9
LISTA DE FIGURAS
Figura 1: Relação entre SIRS, sepse, sepse grave e choque séptico ....................... 18
Figura 2: Score de severidade, peso e temperatura corporal em relação aos valores
basais de cada animal após 4-24 horas da injeção i.p de LPS (A) Escore de
severidade 24 horas após a sepse. (B) Porcentagem de redução do peso corporal
em relação ao basal. (C) Porcentagem de redução da temperatura corporal em
relação ao basal.. ...................................................................................................... 49
Figura 3: Marcadores bioquímicos de falência a órgãos dosados 4-24 horas após a
injeção i.p de LPS. (A) Dosagem de AST para verificar lesão tecidual hepática. (B)
Dosagem de creatinina para verificar dano renal. (C) Dosagem de Lipase para
verificar dano pancreático ......................................................................................... 51
Figura 4: Análise da celularidade do sangue após 4-24 horas da injeção i.p de LPS.
(A) Células Mononucleares; (B) PMNs ...................................................................... 53
Figura 5: Análise da celularidade do lavado peritoneal após 4-24 horas da injeção i.p
de LPS. (A) Mononucleares; (B) PMNs ..................................................................... 55
Figura 6: Efeito do CNA e CNA + HC em monócitos do lavado após 24h da injeção
i.p de LPS. (A) Macrófagos viáveis do lavado. (B) Painel representativo, dot-plots
para monócitos viáveis do lavado peritoneal. (C) Macrófagos Ly6Chigh. (D) Ly6Clow.
.................................................................................................................................. 57
Figura 7: Efeito do CNA e CNA + HC em linfócitos do lavado após 24h da injeção i.p
de LPS. (A) Painel representativo, dot-plots para linfócitos viáveis do lavado
peritoneal. (B) CD4+ viáveis no lavado. (B) CD8+ viáveis no lavado.. ....................... 58
Figura 8: Dosagem de nitrito + Nitrato (NOx) e H2O2 4-24 horas após a injeção i.p de
LPS. (A) Dosagem de NOx. (B) Dosagem de H2O2 ................................................... 60
Figura 9: Dosagens de citocinas plasmáticas após 4 horas da injeção i.p de LPS. (A)
TNFα (B) IL-1β. (C) IL-6. (D) IL-10.. .......................................................................... 62
Figura 10: Dosagens de citocinas plasmáticas após 24 horas da injeção i.p de LPS.
(A) TNFα (B) IL-1β. (C) IL-6. (D) IL-10. ..................................................................... 63
Figura 11: Dosagens de citocinas do lavado peritoneal 4 horas após a injeção i.p de
LPS. (A) TNFα. (B) IL-1β. (C) IL-6. (D) IL-10. ............................................................ 64
Figura 12: Dosagens de citocinas do lavado peritoneal 24 horas após a injeção i.p de
LPS. (A) TNFα. (B) IL-6. (C) IL-10. ........................................................................... 65
Figura 13: Viabilidade de macrófagos obtidos de monócitos do sangue periférico
humano, quando tratadas com LPS (100 ng/ml) na presença ou ausência de CNA
e/ou HC-030031. ....................................................................................................... 66

10
LISTA DE ABREVIATURAS
15d-PGJ2 15-Deoxi-delta (12,14) – Prostaglandina J2
4-HNE 4-Hidroxi-2-nonenal
ACCP Do inglês, American College of Chest Phisician
APCs Células Apresentadoras de Antígenos
AST Aspartato Amino Transferase
ATP Adenosina Trifosfato
CD11b Integrina M de Membrana Presente em Monócitos/Macrófagos
CD14 Marcador de Monócitos
CD18 β2-integrina
CGRP Do inglês, Calcitonin Gene-Related Peptide
CNA Cinamaldeído
COX-2 Cicloxigenase tipo 2
DMEM Meio de Eagle Modificado por Dulbecco
DMSO Dimetilsulfóxido
eNOS Óxido Nítrico Sintase Endolelial
EROS Espécies Reativas de Oxigênio
H2O2 Peróxido de Hidrogênio
ICAM Molécula de Adesão Intercelular
IFN- Interferon – gama
IL-10 Interleucina 10
IL-13 Interleucina 13
IL-1R1 Receptor de Interleucina 1 Tipo 1
IL-1R2 Receptor de Interleucina 1 Tipo 2
IL-1β Interleucina 1β
IL-2 Interleucina 2

11
IL-4 Interleucina 4
IL-6 Interleucina 6
IL-8 Interleucina 8
IRF3 Do inglês, Interferon Regulatory Fator 3
LBP Proteína Plasmática Ligadora de LPS
LPS Lipopolissacarídeo
MD2 Proteína Mielóide Diferenciadora 2
MSR Receptor de Macrófagos Scavenger
NADPH Do inglês, Nicotinamide Adenine Dinucleotide Phosphate-Oxidase
NF- κB Fator Nuclear κB
NK Células Natura Killer
nNOS Óxido Nítrico Neuronal
NO Óxido Nítrico
NOD Proteína Contendo Domínio de Oligomerização Nucleotídica
NOS Óxido Nitrico Sintase
NOSi Óxido Nítrico Sintase Induzida
ONOO- Peroxinitrito
PAMPs Padrões Moleculares Relacionados a Patógenos
PBS Tampão Fosfato Salino
PGE2 Prostaglandina E2
PI Iodeto de Propídio
PRRs Receptores de Reconhecimento Padrão
RNS Espécies Reativas de Nitrogênio
SCCM Do inglês, Society of Critical Care
SIRS Síndrome da Resposta Inflamatória Sistêmica
SP Substância P
TGF-β Fator de Crescimento Transformante Beta

12
TLR Receptores Semelhantes a Toll
TNFα Fator de Necrose Tumoral α
TRP Receptores de Potencial Transitório
TRPA1 Receptor de Potencial Transitório Ankyrin 1
TRPC Receptor de Potencial Transitório Canônica
TRPM Receptor de Potencial Transitório Melastina
TRPML Receptor de Potencial Transitório Mucolipina
TRPP Receptor de Potencial Transitório Policistina
TRPV Receptor de Potencial Transitório Vanilóide
UTI Unidades de Terapia Intensiva
VCAM-1 Molécula de Adesão da Célula Vascular

13
SUMÁRIO
1 INTRODUÇÃO ....................................................................................................... 15
1.1Sepse: conceito e epidemiologia ...................................................................... 15
1.2 Fisiopatologia da sepse .................................................................................... 20
1.3 Mediadores Inflamatórios ................................................................................. 23
1.4 O estresse oxidativo na sepse ......................................................................... 26
1.5 Modelos experimentais de sepse .................................................................... 29
1.5.1 Modelos de endotoxemia ................................................................................. 29
1.5.2 Ligação e perfuração do ceco (Do inglês, cecal ligation and puncture;CLP).... 30
1.5.3 Introdução de cateter no cólon ascendente (Do inglês, colon ascendens stent
peritonitis; CASP) ...................................................................................................... 31
1.5.4 Inóculo bacteriano ............................................................................................ 32
1.6 Potencial imunomodulador da canela ............................................................. 33
1.6 TRPA1 e cinamaldeído ...................................................................................... 36
1 OBJETIVOS ........................................................................................................... 39
2.1 Geral ................................................................................................................... 39
2.2 Específicos ........................................................................................................ 39
2 METODOLOGIA .................................................................................................... 40
3.1 Animais .............................................................................................................. 40
3.2 Drogas farmacológicas e reagentes ................................................................ 40
3.3 Tratamento farmacológico ............................................................................... 40
3.4 Indução da sepse e escore de severidade ...................................................... 41
3.5 Temperatura e peso .......................................................................................... 42
3.6 Avaliação da disfunção de órgãos vitais ........................................................ 42
3.7 Análise de células do Sangue e Lavado Peritoneal ....................................... 42
3.8 Dosagem de NOx ............................................................................................... 43
3.9 Dosagem de H2O2 .............................................................................................. 44

14
3.10 Viabilidade e Fenotipagem de células inflamatórias em animais com sepse
tardia 44
3.11 Dosagem de citocinas..................................................................................... 45
3.12 Cultura de macrófagos humanos .................................................................. 45
3.13 Análise Estatística ........................................................................................... 47
4 RESULTADOS ....................................................................................................... 48
4.1 Score de severidade, peso e temperatura corporal ....................................... 48
4.2 Marcadores bioquímicos de falência a órgãos ............................................... 50
4.3 Análise da celularidade do sangue e lavado peritoneal ................................ 52
4.4 População de Células no Lavado Peritoneal .................................................. 56
4.5 Avaliação da produção de NO e H2O2 .............................................................. 59
4.6 Avaliação de citocinas pró e anti-inflamatórias ............................................. 61
4.7 Viabilidade de macrófagos ............................................................................... 66
5 DISCUSSÃO .......................................................................................................... 67
6 CONCLUSÃO ........................................................................................................ 84
REFERÊNCIAS ......................................................................................................... 85

15
1 INTRODUÇÃO
1.1 Sepse: conceito e epidemiologia
A sepse pode ser definida como um conjunto de reações do organismo
causado pela resposta inflamatória sistêmica descontrolada do indivíduo em
decorrência de uma infecção, e que pode causar disfunção ou falência de um ou
mais órgãos, levando o paciente à morte. (WESTPHAL et al., 2009; PAULO et al.,
2003).
A palavra sepse origina-se do grego “σηψις” que significa “decomposição
da matéria orgânica animal ou vegetal na presença de bactérias”. Esta palavra foi
citada primeiramente por Hipócrates (460-370 AC), e foi encontrada também em
poemas de Homero (GEROULANOS; DOUKA, 2006). Sendo assim, o conceito da
sepse existe há mais de 2.700 anos. Ao longo do tempo, à medida que o estudo da
sepse foi se aprofundando, foram criadas teorias baseadas na noção de que as
bactérias causavam doenças. Hieronymus Fracastorius foi o primeiro a postular esta
hipótese em seu livro De contagionibus et contagiosis morbis et eorum curatione
(1546), de que doenças pudessem ocorrer devido a agentes contagiosos vivos,
criando assim, a doutrina do contagium vivum (BULLOCK, 1960). No entanto, por
mais de dois séculos, a doutrina de Fracastorius foi discutida apenas como
especulação, até que em 1675, Leeuwenhoek descobriu os “animalcula” (micróbios)
através do uso da microscopia, técnica estabelecida por Robert Hooke (1665) e que
deu origem à era da microbiologia (BULLOCK, 1960).
Em 30 de abril de 1878, foi apresentada à Academia de Ciências de Paris
por Pasteur, Joubert & Chamberland, a teoria dos germes. Neste trabalho, baseado

16
em suas experiências relativas ao carbúnculo e a septicemia pelo vibrião séptico,
Pasteur e colaboradores, desenvolveram a teoria microbiana da infecção, a qual
definiu que toda doença infecciosa tem como causa, um microorganismo (DOBELL,
1960).
O estabelecimento do conceito “microorganismos causadores de doença”,
foi acompanhado pela descoberta dos primeiros antibióticos. Em 1929, Alexander
Fleming descobriu uma substância chamada “penicilina” e publicou o primeiro
trabalho demonstrando suas propriedades antibacterianas, o primeiro agente anti-
bacteriano utilizado com sucesso no tratamento de infecções, como a sepse (FUNK
et al., 2009).
A maioria dos casos de sepse é devido a bactérias Gram-negativas (E.
coli, Klebsiella pneumoniae, Enterobacter sp, Pseudomonas aeruginosa e outras).
Staphylococcus aureus e Streptococcus pnemoniae e outras bactérias Gram-
positivas são responsáveis pelos casos remanescentes. Nos pacientes
imunossuprimidos, os fungos, bem como as bactérias, podem causar sepse
(QUENZER et al., 1994).
A heterogeneidade da população séptica levou a um consenso para uma
melhor definição da palavra, visto que vários agentes e estímulos podem
desencadear a sepse. Em 1991 em Chicago, o American College of Chest Phisician
e a Society of Critical Care Medicine (ACCP/SCCM) chefiadas pelo Dr. Roger Bone,
realizaram uma conferência com o intuito de oferecer uma definição prática e
universal da resposta inflamatória à infecção, sendo assim estabelecido, o termo
Síndrome da Resposta Inflamatória Sistêmica (SIRS), para descrever a reação
inflamatória desencadeada pelo organismo frente a qualquer agressão infecciosa ou
não infecciosa. Em humanos, a SIRS é definida quando dois ou mais dos seguintes

17
sintomas clínicos são diagnosticados: hipotermia ou hipertermia (temperatura
corporal inferior a 36ºC ou superior a 38ºC, respectivamente); taquicardia (> 90
batimentos por minuto); taquipnéia (> 20 movimentos respiratórios por minuto ou
PaCO2 < 32 mmHg); leucocitose (> 12.000/mm3 ou menor que 4.000/mm3 ou a
presença de mais de 10% de bastonetes) (BONE et al., 1996). Portanto, Sepse é a
resposta inflamatória sistêmica do organismo frente ao estímulo infeccioso, e suas
manifestações clínicas são similares à SIRS. Sepse grave significa sepse associada
à disfunção de órgãos, hipotensão ou hipoperfusão.
A perfusão anormal acarreta acidose lática, oligúria e ainda, alteração
aguda do estado mental incluindo a perda de função cognitiva. O Choque séptico é
caracterizado por sepse grave associada à hipotensão arterial persistente, mesmo
depois de adequada reposição volêmica. A figura 1 ilustra a relação entre os
conceitos apresentados acima. A terminologia “disfunção” caracteriza o fenômeno no
qual a função orgânica não é capaz de manter a homeostase (PAULO et al., 2003;
BONE et al., 1992).

18
Figura 1: Relação entre SIRS, sepse, sepse grave e choque séptico. A sepse é caracterizada por SIRS decorrente a uma infecção, podendo evoluir para sepse grave e choque séptico. No entanto, a SIRS também pode ocorrer em decorrência de processos não infecciosos como, por exemplo, traumas, queimaduras e pancreatites. Adaptado de Chest 101(6): 1644-55, 1992 e J Biomed Biotechnol. 2011; 2011: 986491.
Ainda hoje, existe uma grande dificuldade em se diagnosticar
rapidamente um paciente com sepse (CHRISTINA et al., 2013). Essa deficiência,
juntamente com a baixa disponibilidade de medicamentos eficazes no combate à
sepse, são realidades agravantes dessa síndrome. Atualmente, a terapia da sepse
se restringe na administração de antibióticos, de fluidos e drogas vasopressoras
(NOAH, 2014). No entanto, o aumento crescente no número de casos de sepse
causadas por bactérias resistentes a diferentes antibióticos é preocupante, e indica a
necessidade do estudo de novos produtos com potencial antibacteriano.
A sepse e suas complicações representam atualmente um grande desafio
para a medicina. Sua alta mortalidade, morbidade e incidência; geram consideráveis
custos financeiros, especialmente devido às suas complicações como choque
séptico e disfunção múltipla dos órgãos. A sua incidência foi estimada em 9% ao ano
nos Estados Unidos da América (EUA), com um custo anual de 17 bilhões de
Sepse
Sepse
Grave
Choque
Séptico
Resposta
Inflamatória
sistêmica
(SIRS)
Infecção

19
dólares (CORDIOLI et al., 2013; MARTIN et al., 2003). A taxa de mortalidade
permanece alta, variando de 50% a mais de 75% (ANGUS et al., 2001).
No Brasil, a sepse é a síndrome geradora de maiores custos aos setores
de saúde públicos e privados do país. Segundo Sogayar et al. (2008), o custo médio
de pacientes sépticos em UTIs no Brasil é de cerca de 23 mil reais, enquanto que o
custo diário por paciente gira em torno de 3 mil reais. Em 2006, Sales Júnior e
colaboradores realizaram um estudo epidemiológico no qual foram investigadas 75
UTIs distribuídas pelo país, sendo detectada uma taxa de incidência dos casos de
sepse de 16% nestas unidades. Dentre esses pacientes, a taxa de mortalidade geral
atingiu 46,6% sendo que destes, 16% eram pacientes com sepse, 34% com sepse
grave (sepse associada à disfunção orgânica, hipotensão ou hipoperfusão) e 65,3%
com choque séptico (sepse com hipotensão arterial persistente). Ainda, a média de
idade desses pacientes foi de 61 anos, sendo prevalentes, as infecções de origem
pulmonar (69%) e abdominal (21%).
Sugere-se que essa alta incidência possa estar relacionada à diferentes
fatores, como aumento da expectativa de vida (principalmente em idosos), aumento
do número de pacientes imunocomprometidos, uso irracional de antibióticos
(levando ao aumento da resistência microbiana) e aumento de procedimentos
invasivos, como cirurgias bariátricas, dentre outras (ANGUS et al., 2001; RANGEL-
FRAUSTO, 2005).
Apesar da alta incidência e mortalidade; existe ainda uma grande
necessidade de identificação dos diversos mecanismos envolvidos no
desencadeamento desta síndrome. Assim, a próxima seção discutirá os mecanismos
envolvidos na fisiopatologia da sepse.

20
1.2 Fisiopatologia da sepse
Conforme descrito anteriormente, a sepse é uma condição clínica
caracterizada por uma resposta inflamatória sistêmica e desencadeada por uma
infecção bacteriana ou outros agentes infecciosos (vírus, fungos e parasitas), sendo
a sepse de origem bacteriana, a mais frequente (PETER, 2012).
Diversos fatores estão implicados no desencadeamento e progressão da
sepse, sendo a resposta imune do hospedeiro frente à infecção, um de seus fatores
determinantes. De fato, na sepse, uma infecção que poderia ocorrer de forma auto-
limitada, permitindo assim, o restabelecimento da homeostasia do hospedeiro, torna-
se devastadora, e por muitas vezes, leva à morte independente do efeito direto do
microorganismo invasor, através do desbalanço da resposta imune do hospedeiro
(KUROSAWA, 2011).
A resposta imune do paciente frente à infecção bacteriana é
desencadeada através do reconhecimento de diferentes componentes bacterianos,
sendo o lipopolissacarídeo (LPS) derivado de bactérias gram-negativas, e o ácido
lipoteicóico derivado de bactérias gram-positivas, os principais componentes
responsáveis por ocasionar o processo inflamatório na sepse (GIRISH
RAMACHANDRAN, 2014). Estes produtos bacterianos são chamados de padrões
moleculares relacionados a patógenos (PAMPs) e são reconhecidos por receptores
de reconhecimento padrão (PRRs) expressos em células do sistema imune inato.
(PETER A, 2012; SIQUEIRA-BATISTA et al., 2011). Dentre os principais PRR,
podemos citar os receptores semelhantes a toll (TLR) presentes na membrana e
compartimentos intracelulares, moléculas de superfície, como o receptor scavenger
de macrófagos (MSR; do inglês Macrophage Scavenger Receptor), CD11b/CD18 e

21
canais iônicos, além dos receptores intracelulares NOD1 e NOD2 (proteína contendo
domínio de oligomerização nucleotídica) (COHEN, 2002; JIEUN et al., 2014). Mais
recentemente foi demonstrado que a família de receptores de potencial transitório
(TRP), especificamente o ankyrin 1 (TRPA1) é ativado pelo LPS (MESEGUER et al.,
2014). O TRPA1 é expresso em terminais nervosos periféricos e centrais
responsáveis pela nocicepção (KOIVISTO et al., 2014).
As endotoxinas de bactérias gram-negativas derivadas da parede celular
destas células e formadas principalmente por LPS estão entre os mais bem
estudados PAMPs. À medida que a infecção ocorre, com a proliferação bacteriana e
morte, esta endotoxina é liberada e rapidamente se difunde sistemicamente no
hospedeiro, atingindo diferentes órgãos e promovendo intensa resposta inflamatória
nos mesmos. O LPS inicialmente ligado a uma proteína plasmática ligadora de LPS
chamada proteína de ligação ao LPS (LBP; do inglês LPS-binding protein) é
transferido para o receptor CD14, encontrado na forma solúvel ou ligado à superfície
da célula por uma âncora de glicosilfosfatidilinositol (BOYSO et al., 2014). Uma vez
ligado ao CD14, o LPS se liga ao complexo TLR4-MD-2. A agregação do complexo
TLR4-MD-2 depois de ligado ao LPS leva a ativação de múltiplos componentes de
sinalização, incluindo o fator nuclear κB (NF-κB; do inglês nuclear fator κB) e fator
regulador de interferon (IRF3; do inglês interferon regulatory factor 3), ocasionando a
produção subsequente de citocinas pró-inflamatórias (PARK, 2013).
A ativação de TLRs, em parte pela ativação de fatores de transcrição
nuclear, leva à síntese e liberação de uma série de mediadores inflamatórios
incluindo citocinas, óxido nitrico (NO) e espécies reativas de oxigênio (EROS); assim
como o aumento da expressão de diversos receptores envolvidos no processo
imune e inflamatório (ASEHNOUNE et al., 2004; VICTOR et al., 2004; KAWAI e

22
AKIRA, 2005; MARCATO et al., 2008). Em seguida, ocorrem a migração e a
ativação de células imunes e inflamatórias, alterações da microvasculatura e
permeabilidade vascular e consequentemente, dano tecidual. Essas alterações
ocorrem rapidamente em resposta à infecção e se não controladas, resultam em
uma resposta inflamatória sistêmica caracterizada por falência de múltiplos órgãos,
colapso da vasculatura, regulação térmica, e ocasionalmente, morte (CROUSER et
al., 2008).
Ao invadir o hospedeiro, patógenos bacterianos são detectados por
células residentes que iniciam o combate ao microrganismo e a resposta
inflamatória. Os fagócitos, que incluem neutrófilos e macrófagos residentes ou
atraídos para o sitio inflamatório, são células cuja função primordial é identificar,
fagocitar e destruir esse patógeno (KUROSAWA et al., 2011). Ainda, células
adicionais são recrutadas para o sítio de infecção através da liberação de
mediadores inflamatórios como citocinas, quimiocinas, NO e EROS, para auxiliar a
erradicação dos patógenos invasores (ANDRADES et al., 2011). Os neutrófilos,
também chamados leucócitos polimorfonucleares (PMNs), possuem propriedades
antimicrobianas incluindo sua capacidade de liberar EROS, e são uma das primeiras
células inflamatórias a migrarem para o tecido infectado (RUDIGERA et al., 2008).
Durante a infecção bacteriana ocorre a interação entre células conhecidas
como apresentadoras de antígenos (APCs) e linfócitos. APCs são responsáveis por
capturar antígenos microbianos e outros antígenos, apresentá-los aos linfócitos e
fornecer sinais que estimulam a proliferação e diferenciação de células T. As
principais APCs são: Células dendríticas, macrófagos e células B. As APCs são
caracterizadas por ligar a imunidade inata à imunidade adquirida (KUROSAWA et

23
al., 2011). Monócitos e macrófagos também são capazes de fagocitar bactérias
(KUROSAWA et al., 2011).
O macrófago possui um papel determinante na sepse. Além de sua
capacidade fagocítica no tecido e relevância como APC, esta célula é capaz de
produzir uma série de mediadores inflamatórios capazes de regular a dimensão da
resposta inflamatória no tecido infectado. Estas células são capazes de
apresentarem fenótipos distintos caracterizados pela produção de grupos distintos
de mediadores inflamatórios, chamado de polarização. Assim, macrófagos com
fenótipo M1 possuem potencial pró-inflamatório e são produtores de IL-1β, Fator de
Necrose Tumoral (TNF), IL-6, enzima sintase induzida (NOSi) e NO. Macrófagos M2
por sua vez, estão associados a imunomodulação. Macrófagos M2 produzem fatores
de crescimento, enzimas de remodelação da matriz extracelular que promovem
reparação tecidual e citocinas anti-inflamatórias como o TGF-β e IL-10. O perfil M2
inclui pelo menos três subgrupos: M2a, induzido pela IL-4 ou IL-13; M2b, induzido
pelos complexos imunes e agonistas de TLR4 ou receptores de IL-1 e M2c, induzido
pela IL-10 e glicocorticoides (MARIE et al., 2008; FERNANDO, 2014).
1.3 Mediadores Inflamatórios
Como já mencionado, o reconhecimento de ligantes microbianos por PRR
leva à ativação de diversas vias de sinalização e, por fim, de fatores de transcrição,
que induzem a expressão de genes cujos produtos são importantes para o
desenvolvimento de respostas inflamatórias e antimicrobianas. Estes produtos finais
incluem citocinas pro-inflamatórias incluindo, TNF-α, IL-1β, IL-2, IL-6, IL-8, e IFN-γ e
citocinas anti-inflamatórias como IL-10, TGF-β e IL-4 induzem uma variedade de
respostas celulares (BOOMER et al., 2014). Estas respostas incluem, mas não se

24
limitam, ao aumento da atividade fagocítica, lesão endotelial vascular com
extravasamento capilar, síntese de proteínas de fase aguda pelo fígado, a
quimiotaxia de leucócitos para locais de infecção/inflamação e ativação do sistema
de coagulação. (BOOMER et al., 2014)
O TNF-α é considerado como uma citocina de importância crítica no
choque endotóxico e o dano tecidual. O TNF- α é produzido por macrófago, células
dendríticas e outros tipos celulares. Entre suas principais funçõs, destaca-se a
indução de quimiocinas como CXCLI e CCL2, a ativação de NOSi e de cicloxigenase
tipo 2 (COX-2), que catalisam a produção de NO e prostaglandina E2 (PGE2)
respectivamente. Ambos NO e PGE2 são vasodilatadores que podem causar a
redução da migração de neutrófilos para o sítio da infecção por inibição da ligação
leucócito-endotélio (BOSMANN et al., 2012). Além do aumento da produção de
macrófagos por células progenitoras, a promoção da ativação, diferenciação e
prolongamento da sobrevivência desta célula, o TNFα aumenta a expressão de
quimiocinas, moléculas de adesão em células inflamatórias (Integrinas) e no
endotélio vascular, como a molécula de adesão intercelular I (ICAM) e a molécula de
adesão da célula vascular I (VCAM-1), um subtipo chamado ICAM-3 expresso em
linfócitos também desempenha um importante papel na migração celular. O TNFα
também aumenta a adesão dependente de integrinas em neutrófilos, promovendo
seu acúmulo no tecido (SCHULTE et al., 2013).
Assim como o TNF, a IL-1β também é liberada principalmente por
macrófagos ativados, através de sinais de dois receptores distintos, denominados
receptor tipo I (IL-1R1) e tipo II (IL-1R2), sendo estes responsáveis por exercerem
efeitos similares nas células do sistema imune, tais como, hematopoiese e
extravasamento de células inflamatórias para o tecido (SCHULTE et al., 2013). IL-1β

25
e TNF-α atuam sinergicamente para induzir um estado parecido ao choque,
caracterizado por permeabilidade vascular, edema pulmonar e hemorragia. Estas
citocinas também têm sido identificadas por mediadores cruciais para o
desenvolvimento da febre e, portanto, pertencem a um grupo de citocinas
pirogênicas (SCHULTE et al., 2013).
A IL-6 é outra importante citocina pró-inflamatória produzida por uma
variedade de células, especialmente macrófagos, células dendríticas, linfócitos,
células endoteliais, fibroblastos e células da musculatura lisa em resposta a
estimulação com LPS, IL-1β e TNF-α (SCHELLER et al., 2006). A IL-6 apresenta
efeitos locais e sistêmicos, incluindo a indução da síntese hepática de diversos
outros mediadores inflamatórios, a estimulação da produção de neutrófilos na
medula óssea e a diferenciação de linfócitos T auxiliares (helper) produtores de IL-
17 (SCHULTE et al., 2013). A IL-6 ainda é uma importante reguladora da
homeostase em relação à temperatura corporal (CHAI et al., 1996). Assim, a
expressão e liberação de IL-6, vêm sendo relacionada com as alterações de
temperatura corporal decorrentes da sepse, i.e., febre e hipotermia (CHAI et al.,
1996). Apesar das suas propriedades pró-inflamatórias, a IL-6 também possui um
relevante papel como citocina anti-inflamatória. A IL-6 inibe a liberação de TNF e IL-
1 (SCHINDLER et al., 1990) e aumenta os níveis de circulação dos mediadores anti-
inflamatórios tais como IL-10 e TGF-β (PAPANICOLAOU et al., 1998). Assim, a IL-6
pode exercer papel tanto na febre como na hipotermia (RINCON, 2012).
A IL-10 é a citocina produzida por macrófagos, mas também, linfócitos T,
B e células NK (LATIFI et al., 2002). A IL-10 inibe a produção de diversos
mediadores pró-inflamatórios, tais como, IL-1, TNF e IL-12 (FIORENTINO et al.,
1991). Em modelo experimental murino a administração de IL-10, 30 minutos após o

26
estimulo com LPS, protegeu os animais contra a endotoxemia (HOWARD et al.,
1993). Em contraste a imunoneutralização desta citocina eleva os níveis circulantes
de TNFα e IL-6 em camundongos (HOWARD et al., 1992).
1.4 O estresse oxidativo na sepse
A falência de órgãos induzida pela sepse, a resposta inflamatória e
consequente estresse oxidativo, induzem modificações na mitocôndria na qual
resulta em sua disfunção e morte celular (GALLEY, 2011).
A membrana interna das mitocôndrias possui uma grande área de
superfície que é impermeável e contém enzimas responsáveis pela produção de
energia a partir do oxigênio. A produção de energia como o ATP ocorre através de
um fluxo de elétrons passando ao longo de cinco complexos moleculares da cadeia
de transporte de elétrons (GALLEY, 2011). A transferência de elétrons na cadeia
transportadora resulta em liberação de prótons, criando o potencial de membrana
mitocondrial. Como resultado, superóxido (O2-) é formado. Cerca de 1% do oxigênio
utilizado é convertido em EROS (GALLEY, 2011). A produção de EROS por
mitocôndrias é importante para a função celular normal e sua sobrevivência. Eles
participam da sinalização celular, homeostasia do cálcio e ferro (PATERSON et al.,
2000).
Em situação normal o sistema antioxidante é capaz de neutralizar a
formação de EROS pela mitocôndria, porem, quando há um desequilíbrio entre os
componentes pró-oxidantes e antioxidantes, ocorre o que chamamos de estresse
oxidativo (ANDRADES et al., 2011). Ao longo dos últimos 10-15 anos, tem havido
uma grande quantidade de estudos que descrevem o estresse oxidativo em
pacientes com sepse, com evidência de produção de EROS, danos associados, e a

27
depleção antioxidante. As respostas inflamatórias iniciadas por estresse oxidativo
ocorrem através da ativação via redox do NF-κB aumentando a liberação e
circulação de mediadores inflamatórios, incluindo citocinas e pentaxina-3, tem sido
reportados na sepse (PALOMERO et al., 2000; BERG et al., 2011; HILL et al., 2009).
A produção de EROS por células fagocíticas com o objetivo de destruir
patógenos é baseada no sistema primário de geração, conhecido como oxidase
fagocitária, mediada pela nicotinamida adenina dinucleótido fosfato (NADPH
oxidase), também conhecida como enzimas NOX (LETO et al., 2009).
É conhecido cerca de 7 familias NOX de mamíferos (NOX 1-5, DUOX 1 e
DUOX 2). Esta família de enzimas gera superóxido através da transferência de
elétrons do NADPH intracelular através da membrana, acoplando-os a oxigênio
molecular, formando o ânion superóxido, um radical livre reativo. Em um fagossoma,
o superóxido pode formar espontaneamente peróxido de hidrogênio, que irá passar
por subsequentes reações gerando diversas espécies reativas de oxigênio (ROS).
(LETO et al., 2009).
Como resultado, uma molécula de NADPH produz dois íons de
superóxido. No entanto, a estabilidade desta molécula pelos fagossomos é limitada,
favorecendo a detecção de seu produto mais estável, o peróxido de hidrogênio
(H2O2) (LETO et al., 2009).
A oxidase fagocitária é induzida e ativada por muitos estímulos, incluindo
IFN e sinais dos TLRs através do seu ligante o LPS, por exemplo (LETO et al.,
2009). O H2O2 é o EROS predominante detectável em células reconstituídas de
NOX – 4 (Martyn et al., 2006). Já a NOX- 1 tem sido relatada pelo seu papel na
regulação do tônus vascular baseado na angiotensina II (CHOI et al., 2008).

28
Além da produção de EROS ocorre também a formação de NO, que tem
um elétron desemparelhado e, portanto, um radical livre. Estes subprodutos são
chamados espécies reativas de nitrogênio (RNS) (PREDONZANI et al., 2015).
O NO endógeno é derivado de vias enzimáticas e não enzimáticas. A
formação do NO enzimático é catalisada por oxido nítrico sintase (NOS) através de
uma série de reações redox, com degradação de L-arginina para L-citrulina e NO na
presença de Oxigênio e NADPH. Três isoformas de NOS são reconhecidas:
endotelial (NOSe ou NOS3), neuronal (NOSn ou NOS1) e induzida (NOSi ou NOS2).
NOSn e NOSe são enzimas constitutivas que são controladas por cálcio intracelular
(Ca2+/calmodulina) (PARRATT, 1998). NOSi é induzido pela transcrição do gene,
independente do Ca2+, expressos por macrófagos e outros tecidos em resposta a
mediadores inflamatórios (LUIKING et al., 2010). A produção de NO é aumentada
durante a sepse por síntese de NOSi do tipo II de NOS. NO produzido por essa
isoforma foi originalmente identificado em macrófagos e contribui para a ação
citotóxica dessas células. O NO produzido por NOS endotelial (NOSe) na
vasculatura está envolvido na profunda vasodilatação do choque séptico. Além
disso, como resultado do estresse oxidativo a respiração celular está inibida e o
tecido torna-se incapaz de utilizar o oxigênio disponível, o que é chamado de hipóxia
metabólica (MONCADA et al., 2002).
Em altas concentrações, o NO torna-se potencialmente pro-inflamatório e
citotóxico pela reação com O2-, formando um produto toxico ONOO- (peroxinitrito),
que causa dano tecidual. O NO tem várias funções fisiológicas vitais, mas a
formação de RNS pode ter efeitos prejudiciais através da oxidação, nitrosilação, ou
nitração de diversos alvos celulares, incluindo proteínas e ácidos nucleicos
(CHANDRA et al., 2006).

29
A patogênese do dano mitocondrial como um resultado da sepse é
provavelmente uma série complexa de eventos. Ambos, NO e EROS combinados
com a liberação de uma variedade de mediadores inflamatórios podem levar a
influenciar direta ou indiretamente, na perda de função mitocondrial como
consequência do “burst” oxidativo (GALLEY, 2011).
1.5 Modelos experimentais de sepse
O uso de animais em modelos experimentais de sepse é essencial para
uma compreensão detalhada da sua fisiopatologia, além de mimetizar vários
eventos observados na patologia humana. Dessa forma, serão discutidos os
principais modelos experimentais de sepse.
1.5.1 Modelos de endotoxemia
A endotoxemia é comumente usada como modelo de sepse em animais
(FINK, 2008). A endotoxina mais comumente utilizada neste modelo é o LPS,
podendo este ser isolado de diferentes bactérias, e também por diferentes sorotipos
de uma mesma bactéria (STARR & SAITO, 2014). A endotoxemia pode variar de
acordo com a dose de endotoxina utilizada para a indução da sepse, bem como com
sua via de administração (FINK, 2014). As vias de administração mais utilizadas são
a injeção intravenosa ou intraperitoneal de endotoxina. De forma geral, uma vez
injetada, a endotoxina inicia uma resposta inflamatória sistêmica que se assemelha
em muitos aspectos com a sepse severa em humanos incluindo a produção
sistêmica e local de diversos mediadores inflamatórios como citocinas e quimiocinas,
a migração de células inflamatórias, coagulação intravascular disseminada e
disfunção vascular, além de falência de múltiplos órgãos decorrente da resposta

30
inflamatória sistêmica (WARD, 2012). Entretanto, percebe-se que o modelo
experimental de LPS difere em alguns pontos da sepse observada em humanos,
especialmente no que diz respeito ao curso temporal de produção de mediadores
inflamatórios, uma vez que o modelo de endotoxemia promove a rápida e alta
produção destas moléculas quando comparado a síndrome humana, na qual estes
eventos ocorrem de forma mais lenta (CAVAILLON et al., 2003; CHEN, 2014).
Assim, apesar das diferenças com a sepse humana, o modelo de endotoxemia pode
ser útil no entendimento do estudo da resposta inflamatória sistêmica e ao choque
séptico. Ressalta-se que quando comparado aos modelos a serem discutidos ainda
nesta seção, a endotoxemia apresenta benefícios importantes no que diz respeito às
diretrizes que regem o bem-estar do animal, uma vez que pode-se controlar a dose
de endotoxina e o tempo de avaliação,facilitando o estudo em um tempo pre-
mortalidade (RITTIRSCH et al., 2007).
1.5.2 Ligação e perfuração do ceco (Do inglês, cecal ligation and puncture;
CLP).
O modelo de CLP foi descrito inicialmente por Wichtermann et al., em
1980, e adaptado por Baker et al.,1983. Ele tem sido considerado o padrão ouro
para pesquisa em modelos de sepse, uma vez que é o que mais se assemelha ao
quadro de sepse em humanos, decorrente de traumas com perfurações das alças
intestinais, apendicite, colite ou peritonite pós-operatória (BENJAMIM, 2001). Após a
perfuração da parede intestinal, ocorre à liberação gradativa do conteúdo cólico (rico
principalmente em bactérias gram positivas e negativas) para a cavidade peritoneal,
induzindo um quadro de peritonite, o qual pode evoluir para sepse e choque séptico

31
(DEJAGER et al., 2011). Ainda, neste modelo, mediadores inflamatórios como
citocinas, apresentam um decurso temporal mais próximo a síndrome humana.
Levando em consideração que a maioria dos pacientes com relato de sepse clínica e
choque endotoxico são crianças e doentes idosos, esse modelo normalmente
emprega animais adultos o que pode não traduzir de forma apropriada estes
parâmetros clínicos (ANGUS et al., 2001). Ainda, este modelo requer intervenção
cirúrgica o que pode acarretar perda de animais decorrente do procedimento
cirúrgico (DEJAGER et al., 2011). Ainda, por se tratar de um procedimento invasivo,
requer a administração de agentes analgésicos, o que pode interferir com a resposta
inflamatória sistêmica, e consequente obtenção de resultados neste modelo. Apesar
de suas limitações, a utilização do modelo CLP para o estudo da sepse, apresenta
grande relevância para a compreensão da evolução da doença humana.
1.5.3 Introdução de cateter no cólon ascendente (Do inglês, colon ascendens
stent peritonitis; CASP)
O modelo de CASP têm sido usado para induzir septicemia em ratos. Um
cateter é inserido dentro do cólon ascendente ocasionando o vazamento intermitente
do conteúdo intestinal (por exemplo, depois de uma grande cirurgia abdominal)
levando a uma invasão bacteriana da cavidade peritoneal seguida por falência de
órgãos, choque séptico e morte (BENJAMIM, 2001). A letalidade observada em
CASP é geralmente uma consequência da falência múltipla de órgãos o que é
realmente reminicente de relatórios clínicos em mortes por sepse (ENTLEUTNER et
al., 2006) O Modelo CASP foi mostrado por ocasional uma disfunção de órgãos
semelhante à observada em pacientes sépticos (BARTHLEN et al., 1999). No

32
entanto, como contraponto, neste modelo a resposta anti-inflamatória de citocinas
ocorre simultaneamente com a resposta pró-inflamatória (ZANTL et al., 1998). Lesão
pulmonar aguda, insuficiência renal e disfunção de células da medula óssea foram
relatadas após CASP (BARTHLEN et al., 1999).
Da mesma forma que o modelo de CLP, o modelo de CASP requer
intervenção cirúrgica e a administração de agentes analgésicos, fatores estes, que
devem ser considerados na escolha do modelo animal a ser estudado a fim de se
reduzir o n experimental, diminuir prejuízos ao animal e também interferências
experimentais.
1.5.4 Inóculo bacteriano
Alguns modelos de sepse empregam a administração de bactérias vivas
por via intravenosa ou intraperitoneal (Hardaway et al., 1996; Fink, 2014). Este
modelo tem a relevância de mimetizar infecções pós-lesão observadas em
ambientes clínicos (CHEN et al., 2014). A Escherichia coli é bactéria a mais
comumente empregada neste modelo. Em mamíferos, diferentes concentrações de
E. coli diferentes respostas em decorrência da infecção (Parker & Watkins, 2001).
Como vantagens, este modelo pode mimetizar uma sepse clínica extrema, como o
visto em meningococcemia em bacteremia pneumocócica (Fink & Heard, 1990).
Ainda, ele permite o estudo dos efeitos agudos de intervenções em curtos períodos
de tempos, reduzindo as despesas em pesquisa e os efeitos deletérios sobre os
animais. Por outro lado, a administração de bactérias vivas pode levar a um imediato
colapso cardiovascular e morte rápida, principalmente quando por via intravenosa; o
que não ocorre na sepse humana (Garrido et al., 2004). Entretanto este modelo

33
apresenta grande valor. A taxa de mortalidade depende das espécies de animais
utilizados e o número de bactérias administradas, de modo que este modelo se torna
mais controlável e reprodutível do que o modelo de CLP ou CASP (Rittirsch et al.,
2007).
1.6 Potencial imunomodulador da canela
O uso histórico de plantas medicinais e seus constituintes mostram que
eles fazem parte da evolução humana e foram os primeiros recursos terapêuticos
utilizados pelos povos. Nas referências históricas sobre plantas medicinais,
podemos verificar que existem relatos de seu uso em praticamente todas as antigas
civilizações (DING, 1987). A primeira referência escrita sobre o uso de plantas como
remédios é encontrada na obra chinesa Pen Ts’ao (“A Grande Fitoterapia”), de Shen
Nung, que remonta a 2800 a.C (Tomazzoni et al., 2006). A OMS acredita que,
atualmente, a prática do uso de plantas medicinais é tida como a principal opção
terapêutica de aproximadamente 80% da população mundial (OMS, 2010). O
mercado mundial de fitoterápicos movimenta cerca de US$ 22 bilhões por ano
(PINTO et al., 2002). Em 2000 o setor faturou US$ 6,6 bilhões nos EUA e US$ 8,5
bilhões na Europa (PINTO et al., 2002). No Brasil estima-se que o comércio de
fitoterápicos seja da ordem de 5% do mercado total de medicamentos, avaliado em
mais de US$ 400 milhões (PINTO et al., 2002). Considerando essas informações,
observa-se que a natureza pode auxiliar no tratamento de diversas doenças
fornecendo diversos componentes químicos ativos úteis também, contra infecções
causadas por bactérias, vírus e fungos, ao mesmo tempo em que oferece suporte
terapêutico para múltiplos órgãos e sistemas do corpo (MCKENZIE et al., 2010).

34
O Gênero Cinnamon é uma árvore aromática pertencente à família
Lauraceae, uma das famílias mais estudadas, a qual compreende cerca de 250
espécies e é conhecida popularmente como canela. Usos tradicionais da canela têm
sido registrados em toda Ásia, África e Europa (SHEN et al., 2012). Na medicina
tradicional chinesa, a canela é indicada como analgésico, antipirético e para mialgia.
Neste contexto, a casca da canela possui um papel importante uma vez que contém
uma grande quantidade de óleo essencial, o qual sugere-se que seja o principal
componente responsável por estes efeitos (SHEN et al., 2012 ).
Os principais óleos essenciais da casca da canela são o eugenol e o
cinamaldeido (CNA). Dados indicam que a casca contém 45% ~ 65% de CNA e 12%
~ 18% de eugenol (Cheng, 1983). Como composto majoritário da casca da canela, o
CNA tem sido extensivamente estudado nas ultimas décadas e indicado pelo seu
potencial imunomodulador.
O primeiro relato deste potencial foi descrito em 1998, em um estudo
realizado por Koh e colaboradores, o qual demonstrou que o CNA é capaz de alterar
a resposta imune in vitro, por promover o aumento da diferenciação de linfócitos
humanos CD4+/CD8+ positivos em CD4+ ou CD8+ positivos. Embora este potencial
imunomodulador tenha sido sugerido para o CNA, estudos direcionados ao
entendimento de seu mecanismo de acão somente ocorreram na ultima década,
sendo estes em sua maioria, estudos in vitro. Estes estudos indicaram que parte dos
efeitos imunomoduladores do CNA deve-se ao seu potencial de inibição da ativação
do NF-κB. Em 2007, sugeriu-se que o CNA inibe a ativação deste fator de
transcrição, por regular diferentes vias de sinalização, incluído MAPKinases. Esta
inibição acarreta subsequente inibição de vias inflamatórias importantes na sepse
como a atividade da iNOS e COX-2 (KIM et al., 2007). Ainda, foi sugerido que o

35
derivado do CNA (2’-hidroxi cinamaldeído) inibe a produção de NO e a ativação do
NF-κB induzida por LPS em macrofagos RAW 264.7 (LEE et al., 2005; KIM et al.,
2010). Similarmente, o CNA inibiu a liberação de IL-1β and TNFα por macrófagos
murinos (RAW 264.7), humanos e monócitos (THP-1) estimulados com LPS (CHAO
et al., 2008). Os mesmos autores demonstraram ainda, que a incubação de CNA
causa uma redução da liberação de EROS induzidas pelo LPS em macrófagos
J774A.1, sem no entanto, interferir com o reconhecimento do LPS pelos macrófagos
ou com a expressão de TLR4 e CD14. Alem disso, o CNA parece suprimir a
resposta pelo TLR4 por inibir a oligomerizacao deste receptor (YOUN et al., 2008).
Os efeitos imunomoduladores do CNA também foram avaliados in vivo.
Em um estudo realizado por Lin e colaboradores (2011), o tratamento repetido com
CNA demonstrou ser capaz de inibir a concentração sérica de citocinas (IL-2, IL-4 e
IL-10) em camundongos BALBc sensibilizados com ovalbumina. Ainda, o CNA é
capaz de inibir a produção de NO, TNF e PGE2 associada ao acúmulo de edema e
células polimorfonucleares induzidos pela injeção de carragenina na pata de
camundongos (LIAO et al., 2012), e regular a apoptose por inibição da ativação do
NF-κB in vivo (KWON et al., 2010).
Todas essas evidências abrem um leque de possibilidades acerca do
papel do CNA na regulação da resposta inflamatoria frente a doenças infecciosas,
como, por exemplo, a sepse; uma vez que este composto pode modular diversas
vias envolvidas nesta sindrome. Ainda, é importante a identificação e caracterização
do alvo farmacológico para este composto no organismo, além do entendimento da
relevância patofisiológica deste alvo, estes fatores vêm abrindo novas perspectivas
para o tratamento de doenças de cunho inflamatório. Assim, a próxima secao
introduzirá este alvo farmacológico. Em seguida, as relações entre o CNA e seu alvo

36
farmacológico, assim como suas possíveis implicações para a sepse, serão
discutidas.
1.6 TRPA1 e cinamaldeído
O receptor TRPA1 foi identificado na ultima década como o alvo
farmacológico do CNA no organismo, sendo o CNA hoje considerado como um
agonista exógeno para este receptor (BANDELL et al., 2004). O TRPA1 faz parte de
uma família de receptores originalmente identificados como expressos nas fibras
sensoriais do tipo C e Aδ e, quando estimulados, promovem a liberação dos
neuropeptídeos substância P (SP) e peptídeo relacionado ao gene da calcitonina
(Calcitonin Gene-Related Peptide; CGRP) (JORDT et al., 2004), considerados
preditores de letalidade em pacientes com sepse (BODKIN & FERNANDES, 2013).
A família de receptores TRP engloba mais de 30 membros distribuídos em
7 subfamilias: Canônica (TRPC; inclue TRPC1-5), Melastatina (TRPM; inclue
TRPM1-8), Vanilóide (TRPV; inclue TRPV1-6), Policistina (TRPP; inclue TRPP1-5),
Mucolipina (TRPML; inclue TRPML1-3), Anquirina (TRPA; inclue TRPA1-5) e
NOMPC (TRPN; encontrados em Drosophila sp, sapos, truta e vermes) (PEDERSEN
et al., 2005). Como característica comum, todos os receptores TRP sao permeáveis
a cátions, particularmente o Ca+2 e podem ser ativados por diversos agonistas
endógenos (incluindo mudanças na temperatura, pH, estímulos mecânicos,
hormônios, espécies reativas de oxigênio e produtos oxidados e outros mediadores
inflamatórios; e exógenos (como produtos derivados de plantas, fumaça de cigarro,
poluição, substâncias quimicas, etc (HUANG et al., 2006).
Recentemente, a importância dos receptores TRP na sepse, começou a
ser especulado, sendo o TRPV1 recentemente identificado como uma molécula

37
protetora na sepse bacteriana (CLARK et al., 2007; FERNANDES et al. 2012).
Assim, é importante ressaltar que além de serem expressos em neurônios, os
receptores TRP são também encontrados numa série de células e tecidos não-
neuronais, incluindo células inflamatórias e células da musculatura lisa da aorta e
microvasculatura (CLAPHAM, 2003). O TRPA1 possui um importante papel no
estabelecimento de processos dolorosos e inflamatórios e é encontrado em cerca de
50% das fibras sensoriais positivas para o TRPV1 além de tecidos não-neuronais
(FERNANDES et al., 2012). O TRPA1 pode ser ativado por diversos agonistas
endógenos e exógenos. Os ativadores endógenos do TRPA1 são frequentemente
compostos eletrofílicos como espécies reativas de oxigênio e mediadores lipídicos
como a 15-Deoxi-delta-prostaglandina J2 (15d-PGJ2) e 4-hidroxi-2-nonenal (4-HNE);
produzidos durante a sepse (TREVISANI et al., 2007). Embora não existam relatos
diretos do envolvimento do receptor TRPA1 na sepse, um papel immunosupressor
tem sido sugerido tanto para seus agonistas endógenos quanto exógenos
(ORENGO et al., 2008).
Da mesma forma como sugerido para o CNA, um efeito imunosupressor
foi sugerido para os agonistas endógenos do TRPA1. O 4-HNE, um produto de
peroxidação lipídica, quando incubado com monócitos humanos, inibe a
diferenciação destas celulas em células dendríticas, além de suprimir a ligação do
LPS ao TLR4 (YANG et al., 2003). 4-HNE também causa redução da ativação do
NF-κB em monócitos humanos THP-1 na presença de Chlamydia pneumonia, e
previne a produção de citocinas (TNFα, IL-1β, IL-6 e IL-8) por monócitos e
macrófagos humanos tratados com LPS (DONATH et al., 2002)
Recentemente, um estudo realizado por Meseguer e colaboradores
(2014), demonstrou que o LPS pode se ligar diretamente aos receptores TRPA1,

38
ativando-os, podendo também atuar de forma sinérgica com outros agonistas
endógenos para este receptor (4-HNE e produtos da peroxidação lipídica). Assim, a
co-aplicação em baixas doses de HNE e LPS, ativa o TRPA1 de forma potente
(MESEGUER et al., 2014). Embora o cinamaldeido tenha sido sugerido como um
agonista seletivo para este receptor, esta indicação provem de modelos
experimentais de dor e inflamação (EID et al., 2008; MIURA et al., 2013). No
entanto, a contribuição deste receptor para as ações imunomoduladoras do
cinamaldeido na sepse, nunca foram investigadas.
Ainda, com a sugestão de que o TRPA1 pode funcionar como um sensor
de endotoxina (MESEGUER et al., 2014), adiciona mais uma camada de
complexidade ao entendimento das ações imunomoduladoras do CNA. Portanto,
com potenciais ações anti-inflamatórias do CNA e a escassez de estudos sobre a
participação do TRPA1 na sepse, este trabalho se propõe em avaliar a ação deste
óleo essencial na sepse.

39
1 OBJETIVOS
2.1 Geral
Avaliar o potencial terapêutico do cinamaldeído na resposta inflamatória
causada pela sepse induzida por lipopolissacarídeo em camundongos.
2.2 Específicos
Avaliar o efeito do cinamaldeído em marcadores bioquímicos relacionados
ao dano tecidual e alterações na temperatura e peso corporal em animais com
sepse induzida pela injeção intraperitoneal de LPS;
Avaliar o efeito do cinamaldeído na migração de células para o peritôneo
em camundongos com sepse induzida por LPS;
Estudar o efeito do cinamaldeído sobre a produção de citocinas
plasmáticas e peritoneais em camundongos com sepse induzida por injeção
intraperitoneal de LPS;
Estudar a capacidade de produção de NO e EROS por células
inflamatórias presentes no peritôneo de camundongos tratados com cinamaldeído e
com sepse induzida por LPS.
Avaliar o efeito do cinamaldeído sobre a viabilidade de macrófagos
derivados a partir de monócitos obtidos do sangue periférico humano, e estimulados
com LPS.

40
2 METODOLOGIA
3.1 Animais
Foram usados camundongos Swiss mantidos em ambiente
climaticamente controlado (temperatura e ciclo claro12h /escuro12h) com comida e
água ad libitum. Em todo o estudo foram usados camundongos machos e fêmeas de
3 meses de idade fornecidos pelo biotério da Universidade Ceuma.
Todos os procedimentos foram conduzidos de acordo com a lei
11.794/2008 que regulamenta procedimentos científicos em animais e apreciado
pelo Comitê de ética em animais (CEUA) da Universidade Ceuma (Protocolo
273/12).
3.2 Drogas farmacológicas e reagentes
Os seguintes reagentes utilizados foram obtidos da Sigma-Aldrich:
DMEM, HC-030031, LPS, azul de tripan, PBS, MTT e DMSO. Reagentes obtidos da
RenyLab: Líquido de Turk. Reagentes obtidos da Alfa-Aesa: Cinamaldeído.
3.3 Tratamento farmacológico
Para avaliar o efeito agudo do tratamento com CNA na sepse, os animais
receberam uma administração única deste óleo essencial (250 mg/kg; n=5-8), por
via oral, 1h antes da indução da sepse.

41
Animais tratados com veículo (10% DMSO em salina) foram utilizados
como controle (n=8). Em outro grupo experimental, os animais receberam o
antagonista seletivo para o receptor TRPA1, HC-030031, (30 mg/kg; n=5-8) 5 min
antes do CNA por via subcutânea.
3.4 Indução da sepse e escore de severidade
A sepse foi induzida por administração intraperitoneal de salina (0,9%)
contendo LPS (E. coli, sorotipo 111:B4; 11,25 milhões EU/kg), uma hora depois do
esquema de tratamento em camundongos não anestesiados. Após 4 e 24h da
indução da sepse, os animais foram anestesiados com uma mistura de cetamina e
xilazina (75 mg/kg e 1 mg/kg, respectivamente) e então exanguinados para coleta de
sangue e posterior separação do plasma.
Em seguida, a cavidade peritoneal foi lavada com 2 ml de PBS estéril
gelado e o lavado peritoneal foi coletado.
Em um grupo separado de experimentos a severidade da sepse foi
avaliada 24 horas após a injeção de LPS. Para isto, parâmetros comportamentais
foram observados e um escore foi atribuído para cada um dos seguintes parâmetros:
comportamento de limpeza (1- Limpeza normal, 2- limpe za reduzida e 3- sem
limpeza), mortilidade (1- motilidade normal, 2- parcialmente reduzido, 3- pobre
motilidade, 4- sem motilidade), piloereção (1- ausência, 2-presença), secreção
ocular (1- ausência, 2-presença). A soma das pontuações atribuídas a cada um dos
parâmetros para cada animal foi tomado como índice de gravidade, com os escores
mais altos correspondentes a piora da doença. Para comparação, o escore basal foi
quantificado para cada animal para todos os grupos de camundongos.

42
3.5 Temperatura e peso
Os animais foram pesados com uma balança de precisão (FILIZOLA- MF-
6/1), antes do tratamento farmacológico e depois de 4 e 24h da indução da sepse. A
temperatura dos animais foi mensurada, em graus centígrados, antes do esquema
de tratamento e após 4 e 24h da injeção de LPS. Para isso foi utilizado um
termômetro digital e vaselina sólida, a haste do termômetro foi introduzida 1cm no
reto do animal.
3.6 Avaliação da disfunção de órgãos vitais
A disfunção do fígado, rins e pâncreas foi avaliada por quantificação dos
níveis plasmáticos de aspartato aminotransferase (AST), creatinina e lipase com
reagentes Labtest. Os resultados foram expressos em UI/L (lipase e AST) e nmol/l
(creatinina). Foi utilizado um analisador automático LABMAX-240/Labtest no
laboratório de bioquímica da Universidade Ceuma, São Luís-MA.
3.7 Análise de células do Sangue e Lavado Peritoneal
Um total aproximado de 1 ml de sangue foi coletado e processado no
analisador automático (SD20 – Labtest diagnóstica). Em seguida, foi feito a
contagem total (x106) a partir de 20µl de sangue e 0,4 de líquido de Turk (diluição
1:20) e analisadas na câmara de Neubauer utilizando-se de microscópio óptico
comum. A contagem diferencial de leucócitos foi realizada em esfregaço sanguíneos
corados com o corante Panótico. O plasma foi obtido por centrifugação do sangue
(Centrifuga refrigerada, CT-500R) a 1500 rpm por 15 min, separado em eppendorfs
para quantificação de citocinas plasmáticas e marcadores de falência.

43
Enquanto ao lavado, uma alíquota foi analisada em câmara
hemocitométrica tipo Neubauer, 20µl de lavado e 0,2 µ de Turk (diluição 1:10).
(x106). Outra alíquota de 20µl do exsudato peritoneal foi utilizada para determinar a
população de leucócitos através de análise de esfregaço corado com Kit Panótico
rápido. Ainda, as amostras foram analisadas em SDH20-Labtest automatizada, para
confirmação dos esfregaços. O restante do lavado foi dividido em 2 alíquotas, sendo
uma delas centrifugada (Centrifuga refrigerada, CT-500R) a 1500 rpm por 10 min,
para obtenção do sobrenadante e análise de citocinas e NO; e a outra utilizada para
análise de citometria de fluxo e dosagem de H2O2.
3.8 Dosagem de NOx
O Conteúdo de NO2 -/NO3
– foi mensurado pelo ensaio de Griess, um
indicador da produção de NO, obtidos do lavado peritoneal de animais submetidos
ao esquema de tratamento farmacológico. NO3 – foi reduzido para nitrito (NO2
-) por
incubação de 80µl de amostra com 20 µl de 1U/ml nitrato redutase e 10 µl de 1mM
NADPH por 30 min a 37ºC em placa de 96 poços. Após, 100 µl de reagente de
Griess (5% v/v H3PO4 contendo 1% de sulfanilamida/0,1% de dihidrocloreto de
naftileno) foi adicionado e incubado por 15 min a 37ºC. As absorbâncias foram
determinadas com filtro de 550 nm e mensuradas usando espectrofotômetro (MB-
580/Heales). Após as subtrações das leituras (com e sem reagente), cada amostra
foi comparada com a curva padrão de nitrito (0-300µM) e expressos como % da
produção de NOx em µM em relação ao controle (veículo).

44
3.9 Dosagem de H2O2
A produção de H2O2 por células inflamatórias peritoneais foi mensurada
usando o kit de ensaio H2O2/ peroxidase (Sigma-Aldrich). O ensaio foi realizado em
placa de 96 poços, de acordo com as instruções do fabricante. Para isto, as células
do lavado foram ajustadas para 25,000/poço e incubadas com 46µl do tampão de
ensaio, 2µl do substrato fluorescente de peroxidase e 2ul do substrato de H2O2, em
seguida a placa foi incubada a 37ºC no escuro, após 3 min a absorbância inicial foi
determinada com um filtro de 570nm, medindo a cada 10 min até que uma das
amostras apresente leitura similar ou maior do que o maior ponto da curva padrão.
As leituras das absorbâncias foram feitas na presença e na ausência de Amplex
Red, e foram comparadas com uma curva padrão de H2O2 (0-40 µM). Os resultados
foram expressos pela diferença entre as amostras incubadas na presença e
ausência do Amplex Red como % da produção de H2O2 em µM em relação ao
controle (veículo).
3.10 Viabilidade e Fenotipagem de células inflamatórias em animais com sepse
tardia
A viabilidade do lavado peritoneal, CD4+, CD8+ e Ly6C+ foi avaliada
usando iodeto de propideo (PI-). Para este fim, foram preparadas suspensões de
células únicas. As células também foram coradas em azul de tripan (Sigma-Aldrich,
Brasil) para avaliar a viabilidade. Células (5 x 105), foram lavadas, ressuspendidas
em tampão de citometria de fluxo (2% de soro fetal bovino Invitrogen, Brasil), em
solução salina com fosfato-PBS (Sigma-Aldrich, Brasil), e coradas com anticorpos
monoclonais diretamente conjugados (BD Biosciences ou eBioscciences, Brasil):
anti-Ly-6C FITC (clone AL-21), anti-CD4 APC (clone RM4-5), anti-CD8 PE (clone 53-

45
6.7). Todas as coletas de dados de células do lavado foram realizadas utilizando o
equipamento BD Accuri (BD Immunocytometry Systems) equipado com um laser de
488 nm e outro de 640 nm e os dados foram analisados usando o software FlowJo
(Tree Star, Ashland, OR) ou BD Accuri C6 softwaare (BD Immunocytometry
Systems). Para a estimativa da porcentagem do número de células, as viáveis foram
consideradas PI-, sendo os resultados expressos em termos de células viáveis. A
estratégia do Gate: Debris (FSC-A vs. SSC-A) e doublets (FSC-H vs. FSC-A) foram
excluídos e a discriminação de vivas/mortas foi determinada usando PI, as células
PI- foram consideradas viáveis. Células CD4+ e CD8+, bem como monócitos e
granulócitos foram estimados baseados em marcadores fenotípicos e/ou na
distribuição de granulosidade (FSC-A vs. SSC-A). Os resultados são expressos na
porcentagem do número total de células viáveis.
3.11 Dosagem de citocinas
As citocinas (IL-1, IL-6, IL-10 e TNF) foram quantificadas no plasma e no
lavado peritoneal dos animais em estudo, através do kit CBA (Cytometric Bead
Array) utilizando citometria de fluxo, conforme as especificações do fabricante. Os
resultados foram expressos em picogramas/mililitro (pg/ml).
3.12 Cultura de macrófagos humanos
O sangue foi coletado de 6 voluntários saudáveis, entre 20 e 35 anos, não
fumantes, sem alergia relatada a canela, pacientes que não apresentam artrite
reumatóide ou qualquer outra doença de cunho inflamatório de curso crônico,
pacientes que não tenham passado por infecções recentes, e pacientes que não
tenham feito uso de medicamentos anti-inflamatórios nos últimos 15 dias que

46
antecederam a coleta de amostras. Ainda, o projeto foi apreciado e aprovado sob o
número de protocolo 204.347.
Células mononucleares do sangue periférico foram isolados a partir do
sangue total usando Ficoll-Paque, por centrifugação e separação celular. As células
obtidas foram contadas em câmara de Neubauer. As células foram plaqueadas
(2x105 em placas de 96 poços) com DMEM (Sigma-Aldrich) contendo soro bovino
fetal (FBS) 10% e incubadas a 37ºC em estufa de CO2 durante 7 dias para a
diferenciação em macrófago. No sétimo dia as células foram estimuladas com LPS
de E. coli, sorotipo 111:B4 (100 ng/ml), CNA (1 e 10 µg/ml) e HC (10 µM) por 24
horas. O teste de viabilidade celular foi conduzido pelo teste do MTT ((3-(4,5-
dimetiltiazol-2yl)-2,5- difenil brometo de tetrazolina), é um ensaio colorimétrico para a
avaliação da viabilidade celular, citotóxidade ou atividade citostática, as enzimas
oxirredutase celulares são capazes de reduzir o corante MTT ao seu formazano
insolúvel, o qual tem uma cor púrpura. Brevemente, as drogas foram retiradas dos
poços e lavada 3x com PBS, em seguida foi acrescentado 100 µl da solução de MTT
(diluído em DMEM contendo 10% de FBS) em cada poço e incubado por 2 horas ao
abriga da luz. Após a incubação foi retirado a solução dos poços e acrescentado 200
µl de DMSO em cada poço, em seguida foi realizado a leitura por espectrofotometria
a 570nm.

47
3.13 Análise Estatística
Todos os valores foram expressos em média ± SEM com exceção dos
experimentos para detecção de citocinas, os quais encontram-se expressos como
mediana (mínimo – máximo). A porcentagem de inibição foi calculada como media +
SEM para cada experimento individual. A comparação entre os grupos foi realizada
através do teste de normalidade de Shapiro-Wilk, seguido de análise de variância
(ANOVA) e pelo teste de Bonferroni para comparações múltiplas ou Kruskal-Wallis
seguido de Dunns, no software estatístico GraphPad Prism 5.0. Níveis de P<0,05
foram considerados significantes.

48
4 RESULTADOS
4.1 Score de severidade, peso e temperatura corporal
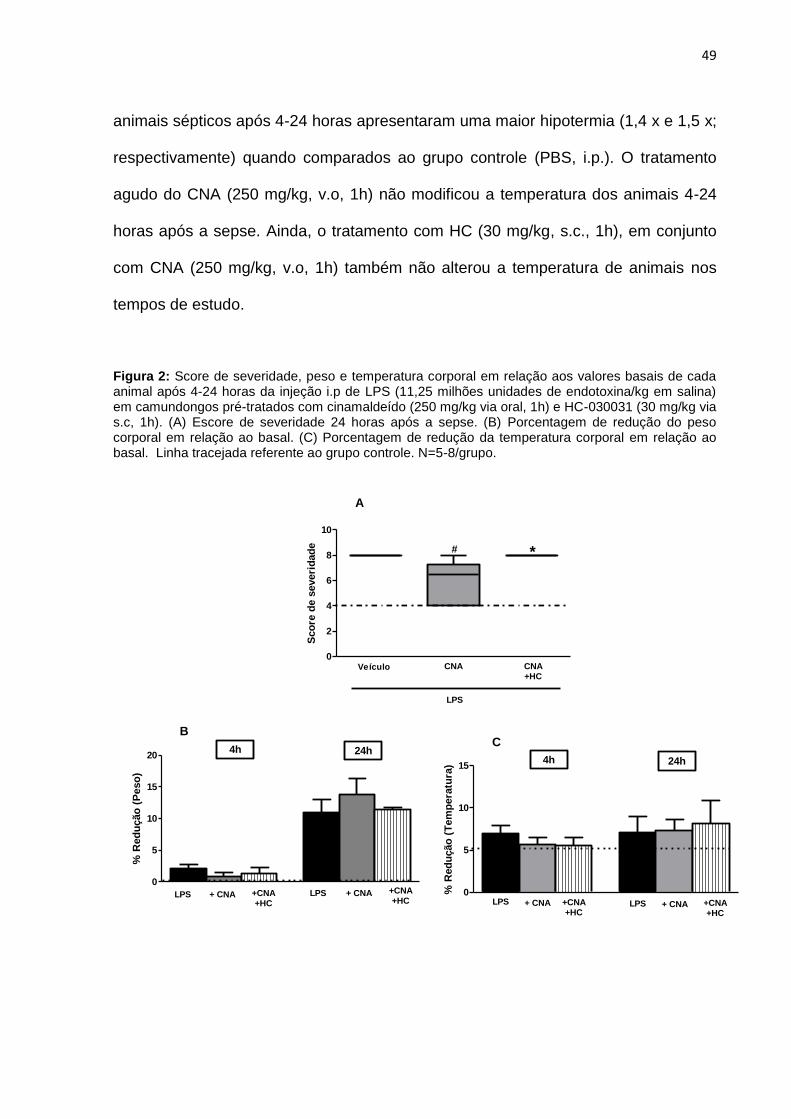
A figura 2A mostra as variações na severidade da sepse em animais
tratados com LPS (24h). Animais sépticos apresentam um escore de severidade
maior (mediana 8.0 (8.0-8.0)) quando comparado aos valores basais (4.0 (4.0-4.0)).
O pré-tratamento com CNA (250 mg/kg v.o, 1h) melhorou significativamente a
severidade da sepse, com animais que apresentaram pontuações mais baixas do
que em animais sépticos (mediana 6.5 (4.0-8.0)). Estes efeitos foram bloqueados
com o tratamento do HC (30 mg/kg s.c, 1h) (mediana 8.0 (8.0-8.0)).
Parâmetros fisiológicos como peso e temperatura corporal também foram
avaliados como alterações sistêmicas da sepse. Assim, foram realizadas medidas
basais e 4-24 horas após o LPS e os gráficos foram expostos em porcentagem de
redução. Os resultados da Figura 2B demonstram que o tratamento com LPS (4-
24h) causou maior perda de peso (2,2 x e 10,9 x respectivamente) quando
comparado ao grupo controle (PBS via intraperitoneal). O pré-tratamento com CNA
(250 mg/kg via oral, 1h), administrado 1h antes do LPS, não foi capaz de alterar o
peso dos animais 4 horas após a sepse, em 24 horas é possível visualizar um
aumento na perda de peso desses animais, porém, não foi significativo. O uso do
antagonista seletivo de receptores TRPA1, HC-030031 (30 mg/kg via s.c, 1h), junto
com o CNA (250 mg/kg v.o, 1h) também não alterou o peso dos animais em 4-24
horas após a sepse.
Na figura 2C está representada a temperatura corporal de animais
submetidos aos esquemas de tratamento. Portanto, é possível visualizar que os
49
animais sépticos após 4-24 horas apresentaram uma maior hipotermia (1,4 x e 1,5 x;
respectivamente) quando comparados ao grupo controle (PBS, i.p.). O tratamento
agudo do CNA (250 mg/kg, v.o, 1h) não modificou a temperatura dos animais 4-24
horas após a sepse. Ainda, o tratamento com HC (30 mg/kg, s.c., 1h), em conjunto
com CNA (250 mg/kg, v.o, 1h) também não alterou a temperatura de animais nos
tempos de estudo.
Figura 2: Score de severidade, peso e temperatura corporal em relação aos valores basais de cada animal após 4-24 horas da injeção i.p de LPS (11,25 milhões unidades de endotoxina/kg em salina) em camundongos pré-tratados com cinamaldeído (250 mg/kg via oral, 1h) e HC-030031 (30 mg/kg via s.c, 1h). (A) Escore de severidade 24 horas após a sepse. (B) Porcentagem de redução do peso corporal em relação ao basal. (C) Porcentagem de redução da temperatura corporal em relação ao basal. Linha tracejada referente ao grupo controle. N=5-8/grupo.
0
2
4
6
8
10
#
Veículo CNA CNA+HC
LPS
*
A
Sco
re d
e s
everi
dad
e
0
5
10
15
20
LPS + CNA +CNA+HC
LPS + CNA +CNA+HC
4h 24h
B
% R
ed
ução
(P
eso
)
0
5
10
15
LPS + CNA +CNA+HC
LPS + CNA +CNA+HC
4h 24h
C
% R
ed
ução
(T
em
pera
tura
)

50
4.2 Marcadores bioquímicos de falência a órgãos
Os marcadores de danos a órgãos foram avaliados 4-24 horas após a
indução da sepse e mensurados os níveis plasmáticos de AST (aspartato
aminotransferase), lipase e creatinina indicadores de falha no coração/fígado,
pancreática e renal respectivamente.
De acordo com a figura 3A, a enzima AST quantificada 4 horas após a
sepse em animais tratados com LPS não tiveram alteração nos seus níveis em
relação ao grupo controle. Da mesma forma, o grupo pré-tratado com CNA (250
mg/kg v.o, 1h) ou CNA (250 mg/kg v.o, 1h) junto com HC (30mg/kg via s.c, 1h) não
verificou-se alteração nos seus níveis enzimáticos. Analisando os níveis de AST 24
horas após a injeção i.p. de LPS observou-se um aumento (1,28 x) nos animais
tratados com LPS em relação ao grupo controle. O pré-tratamento com CNA (250
mg/kg v.o, 1h) diminuiu (24% ± 2%) seus níveis enzimáticos, porém, não foi de
forma significativa. Quando os animais foram tratados com CNA em conjunto com
HC (30mg/kg via s.c, 1h), observa-se uma diminuição quando comparado ao grupo
LPS (29% ± 3%) (Figura 3A).
Em relação aos níveis de creatinina, observa-se um aumento (36 x) 4
horas após a indução da sepse quando os animais foram tratados com LPS em
comparação ao grupo controle (PBS i.p).
O pré-tratamento com CNA (250 mg/kg v.o, 1h) 4 horas após a sepse,
diminuiu os níveis de creatinina (60,42% ± 4%) quando comparado ao grupo LPS. O
uso do HC-030031(30mg/kg via s.c, 1h) junto com CNA (250 mg/kg v.o, 1h) diminuiu
os níveis desta enzima (79% ± 3%) comparado ao tratamento com LPS (Figura 3B).
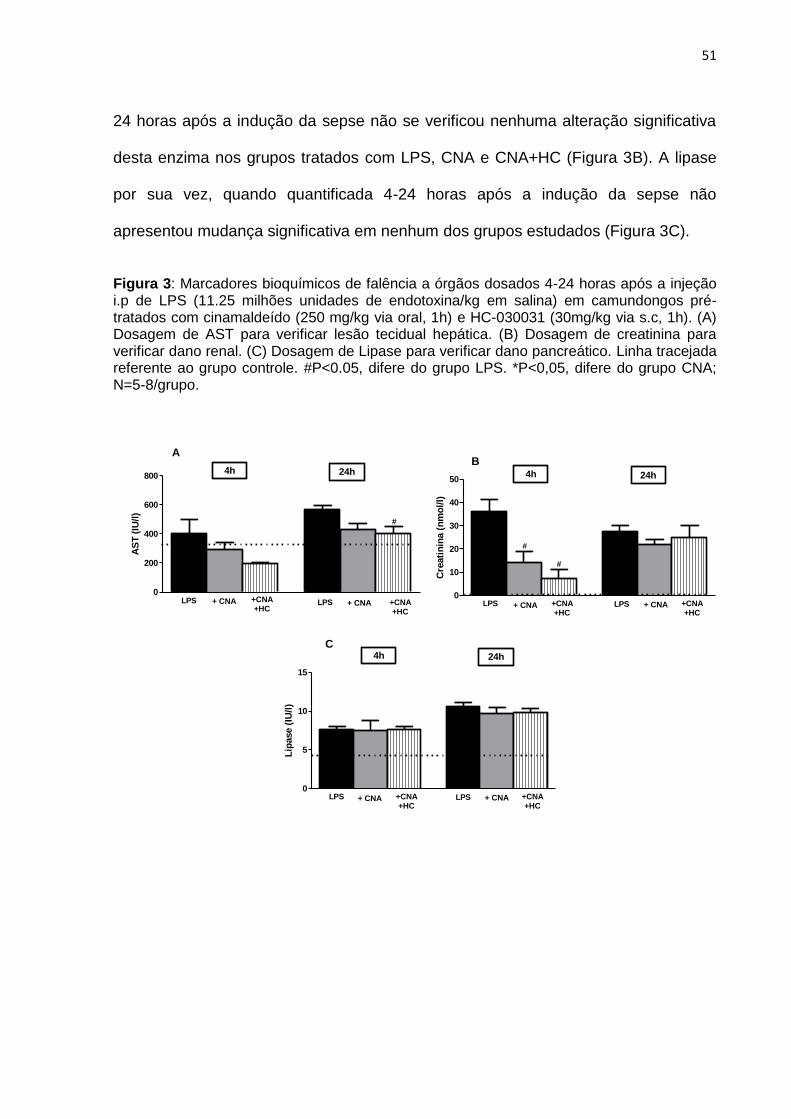
51
24 horas após a indução da sepse não se verificou nenhuma alteração significativa
desta enzima nos grupos tratados com LPS, CNA e CNA+HC (Figura 3B). A lipase
por sua vez, quando quantificada 4-24 horas após a indução da sepse não
apresentou mudança significativa em nenhum dos grupos estudados (Figura 3C).
Figura 3: Marcadores bioquímicos de falência a órgãos dosados 4-24 horas após a injeção i.p de LPS (11.25 milhões unidades de endotoxina/kg em salina) em camundongos pré-tratados com cinamaldeído (250 mg/kg via oral, 1h) e HC-030031 (30mg/kg via s.c, 1h). (A) Dosagem de AST para verificar lesão tecidual hepática. (B) Dosagem de creatinina para verificar dano renal. (C) Dosagem de Lipase para verificar dano pancreático. Linha tracejada referente ao grupo controle. #P<0.05, difere do grupo LPS. *P<0,05, difere do grupo CNA; N=5-8/grupo.
0
200
400
600
8004h 24h
#
LPS + CNA +CNA+HC
LPS + CNA +CNA+HC
A
AS
T (
IU/l
)
0
10
20
30
40
504h 24h
#
#
LPS + CNA +CNA+HC
LPS + CNA +CNA+HC
B
Cre
ati
nin
a (
nm
ol/
l)
0
5
10
15
4h 24h
LPS + CNA +CNA+HC
LPS + CNA +CNA+HC
C
Lip
ase (
IU/l
)

52
4.3 Análise da celularidade do sangue e lavado peritoneal
Células inflamatórias do sangue e lavado foram quantificadas para
estimar a influência da sepse e dos tratamentos utilizados na migração celular. Em
animais sépticos e pré-tratados com CNA (250 mg/kg v.o, 1h) não observa-se
alteração no quantitativo de células mononucleares do sangue 4 horas após a
sepse, no entanto, com o pré-tratamento do CNA (250 mg/kg v.o, 1h) em conjunto
com o HC (30 mg/kg via s.c, 1h), observa-se um aumento dessas células circulantes
(3,36 x) e (2,44 x) quando comparado ao grupo LPS e CNA respectivamente. (Figura
4A).
Em modelo de sepse tardia (24h) pode-se notar um aumento dessas
células quando os animais foram tratados com LPS comparado ao grupo controle
(3,46 x). O tratamento agudo com CNA (250 mg/kg v.o, 1h) reduziu a população de
mononuclear (28,4% ± 0,63%) quando comparado com animais sépticos. Com o uso
do antagonista seletivo dos receptores TRPA1, HC (30 mg/kg via s.c, 1h) em
conjunto com o CNA (250 mg/kg v.o, 1h), observa-se um aumento (1,77 x) dos
mononucleares do sangue comparado ao grupo que recebeu apenas CNA (250
mg/kg v.o, 1h), 24 horas após a indução da sepse (Figura 4A).
Com relação às células segmentadas do sangue, não houve diferença
entre os grupos de tratamentos 4-24 horas após a sepse (Figura 4B).
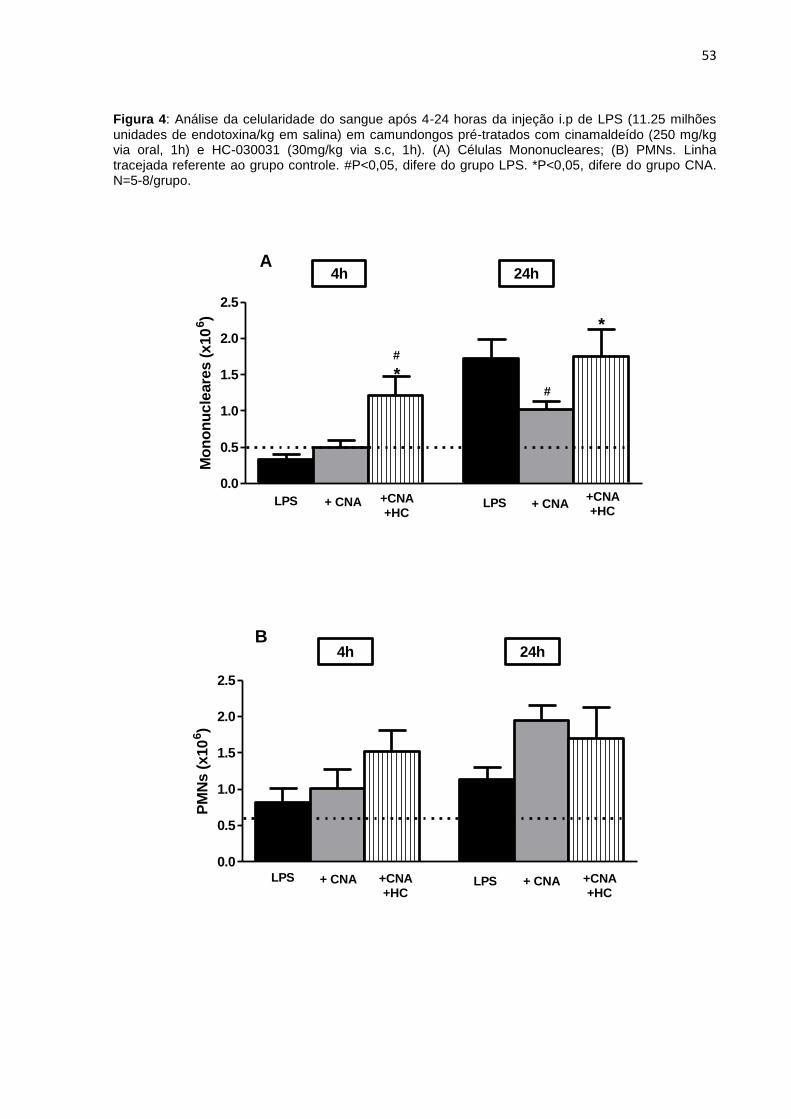
53
Figura 4: Análise da celularidade do sangue após 4-24 horas da injeção i.p de LPS (11.25 milhões unidades de endotoxina/kg em salina) em camundongos pré-tratados com cinamaldeído (250 mg/kg via oral, 1h) e HC-030031 (30mg/kg via s.c, 1h). (A) Células Mononucleares; (B) PMNs. Linha tracejada referente ao grupo controle. #P<0,05, difere do grupo LPS. *P<0,05, difere do grupo CNA. N=5-8/grupo.
0.0
0.5
1.0
1.5
2.0
2.5
LPS + CNA +CNA+HC
LPS + CNA+CNA+HC
4h 24h
#
*
#
*
A
Mo
no
nu
cle
are
s (
x10
6)
0.0
0.5
1.0
1.5
2.0
2.5
LPS + CNA +CNA+HC
LPS + CNA +CNA+HC
4h 24hB
PM
Ns
(x
10
6)

54
O perfil de migração celular para o peritônio também foi avaliado. De
acordo com a figura 5A não houve alteração nas células mononucleares de animais
que foram submetidos aos esquemas de tratamento 4 horas após a sepse.
Analisando o grupo tratado com LPS 24 horas após a sepse, observa-se que houve
uma diminuição dessas células quando comparado ao grupo controle (PBS i.p.)
(28% ± 0,50%). Os animais tratados 1h antes com CNA (250 mg/kg v.o, 1h), quando
avaliados no modelo de sepse tardia, aumentaram a celularidade de mononucleares
(1,30 x) no lavado quando comparado ao grupo tratado com LPS (24h). O pré-
tratamento de CNA (250 mg/kg v.o, 1h) em conjunto com o antagonista seletivo dos
receptores TRPA1, HC (30 mg/kg via s.c, 1h) aumentou significativamente (2,3 x)
estas células quando comparado ao grupo LPS (Figura 5A).
Conforme ilustra a figura 5B houve um aumento perfil de migração de
células segmentadas 4-24 horas após a indução da sepse. Entretanto, em 24 horas
o pré-tratamento com CNA (250 mg/kg v.o, 1h) aumentou essas células (3,73 x)
quando comparado com o grupo LPS (24h). Já o tratamento em conjunto do CNA
(250 mg/kg v.o, 1h) com HC (30 mg/kg via s.c, 1h) diminuiu o número de PMNs
(66% ± 1,02%) quando comparado ao grupo tratado apenas com CNA (250 mg/kg
v.o, 1h) (Figura 5B).
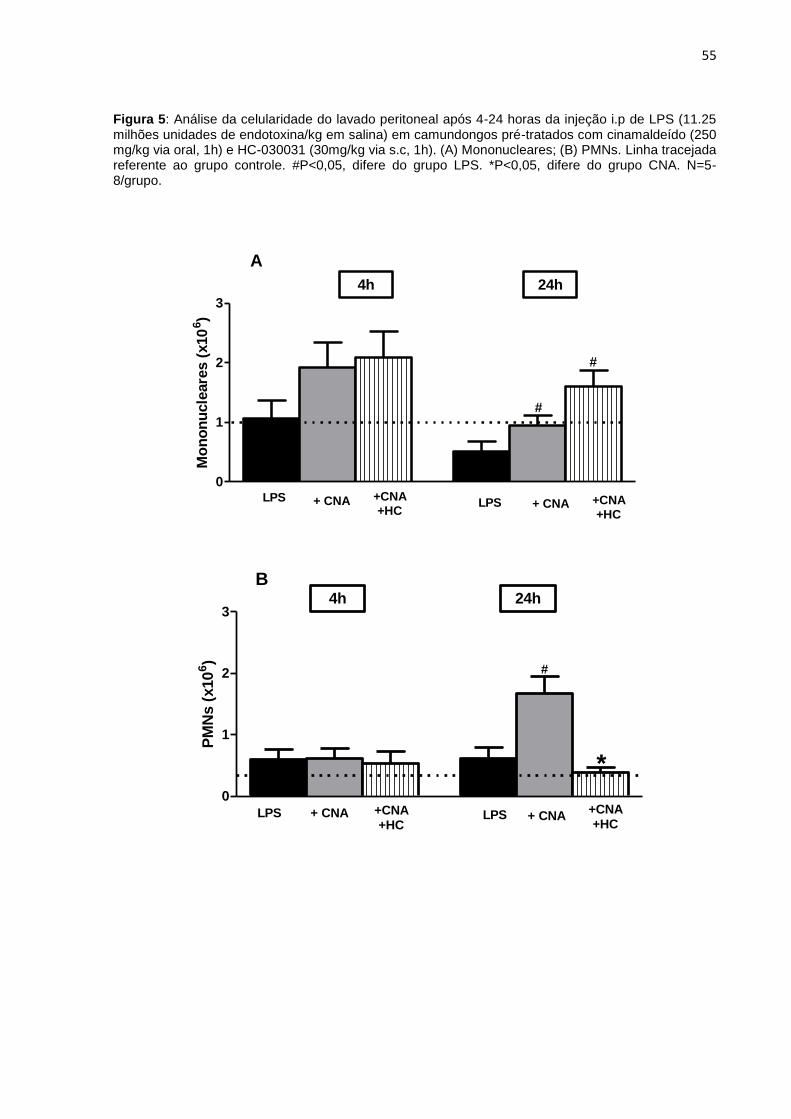
55
Figura 5: Análise da celularidade do lavado peritoneal após 4-24 horas da injeção i.p de LPS (11.25 milhões unidades de endotoxina/kg em salina) em camundongos pré-tratados com cinamaldeído (250 mg/kg via oral, 1h) e HC-030031 (30mg/kg via s.c, 1h). (A) Mononucleares; (B) PMNs. Linha tracejada referente ao grupo controle. #P<0,05, difere do grupo LPS. *P<0,05, difere do grupo CNA. N=5-8/grupo.
0
1
2
3
LPS + CNA +CNA+HC
LPS + CNA +CNA+HC
4h 24h
#
A
#
Mo
no
nu
cle
are
s (
x10
6)
0
1
2
3
LPS + CNA +CNA+HC
LPS + CNA+CNA+HC
4h 24h
#
B
PM
Ns (
x10
6)
*

56
4.4 População de Células no Lavado Peritoneal
As populações de células do lavado peritoneal (Ly6Chigh, Ly6Clow, CD4+ e
CD8+) foram analisadas 24 horas após a indução da sepse.
A análise da população de macrófagos peritoneais demonstra que o pré-
tratamento do CNA em animais sépticos aumentou o número dessas células (3,3 x)
(Figura 6A). A avaliação de macrófagos Ly6C+ demonstrou um aumento no número
de ambas populações Ly6Chigh (2,5 x) e Ly6Clow (3,5 x) em camundongos sépticos e
tratados com CNA (Figura 6B, 6C e 6D). O tratamento em conjunto com o
antagonista do receptor TRPA1, HC-030031, reduziu (~39,5%) a população de
macrófagos peritoneais bem como o número de macrófagos Ly6Chigh e Ly6Clow
(Figura 6B, 6C e 6D).

57
Figura 6: Efeito do CNA e CNA + HC em monócitos do lavado após 24h da injeção i.p de LPS. Debris (FSC-A vs. SSC-A) e doublets (FSC-H vs. FSC-A) foram excluídos e a discriminação de vivas/mortas foi determinada usando PI, células PI- são consideradas como viáveis. Células CD4+ e CD8+, bem como monócitos e granulócitos foram estimados baseados em marcadores fenotípicos e/ou na distribuição de granulosidade (FSC-A vs. SSC-A). (A) Macrófagos viáveis do lavado. (B) Painel representativo, dot-plots para monócitos viáveis do lavado peritoneal. (C) Macrófagos Ly6Chigh. (D) Ly6Clow. Os dados são expressos como média ± SD, n=5 por grupo. *P<0,05 comparados com os respectivos grupos controles.
0
100
200
300
400
LPS + CNA + CNA+HC
*
#
A
Macro
fag
os P
EL
F (
x10
3)
0
20
40
60
80
LPS + CNA + CNA+HC
C
Macro
fag
os L
y6C
hig
h (
x10
3)
0
100
200
300
400
*
#
LPS + CNA + CNA+HC
D
Macro
fag
os L
y6C
low
(x10
3)
B

58
Não houve alteração significativa no número de células CD4+ e CD8+
obtidas do lavado peritoneal 24 horas após a sepse. (Figura 7A -C).
Figura 7: Efeito do CNA e CNA + HC em linfócitos do lavado após 24h da injeção i.p de LPS. Debris (FSC-A vs. SSC-A) e doublets (FSC-H vs. FSC-A) foram excluídos e a discriminação de vivas/mortas foi determinada usando PI, células PI- são consideradas como viáveis. Células CD4+ e CD8+, bem como monócitos e granulócitos foram estimados baseados em marcadores fenotípicos e/ou na distribuição de granulosidade (FSC-A vs. SSC-A). (A) Painel representativo, dot-plots para linfócitos viáveis do lavado peritoneal. (B) CD4+ viáveis no lavado. (C) CD8+ viáveis no lavado. Os dados são expressos como média ± SD, n=5 por grupo. *P<0,05 comparados com os respectivos grupos controles.
0
50
100
150
LPS + CNA + CNA+HC
CD
4+ (
x10
3)
B
0
20
40
60
80
LPS + CNA + CNA+HC
CD
8+
(x10
3)
C
A

59
4.5 Avaliação da produção de NO e H2O2
Foi quantificada a produção das enzimas óxido nítrico e peróxido de
hidrogênio pelas células do lavado peritoneal. Como podem ser observados na
figura 8A os níveis de NO avaliados 4-24 horas após a sepse aumentaram (2,2 x e
2.7 x respectivamente) em animais tratados com LPS em relação ao grupo controle.
Apesar da visível diminuição desta enzima 4-24 em animais pré-tratados com CNA
(250 mg/kg v.o, 1h) em relação ao grupo LPS, este não foi significativo (23% ± 1% e
41% ± 2% respectivamente). O tratamento do CNA (250 mg/kg v.o, 1h) em conjunto
com HC (30mg/kg via s.c, 1h) não alterou NO 4-24 (Figura 8A). Os níveis de H2O2
dosados 4 horas após a sepse também tiveram aumentados (1,92 x) quando os
animais foram tratados com LPS em relação ao grupo controle. O tratamento agudo
com CNA (250 mg/kg v,o, 1h) não alterou a enzima H2O2 4-24 horas, da mesma
forma, quando combinado com o antagonista HC (30mg/kg via s.c, 1h) não houve
mudança (Figura 8B).
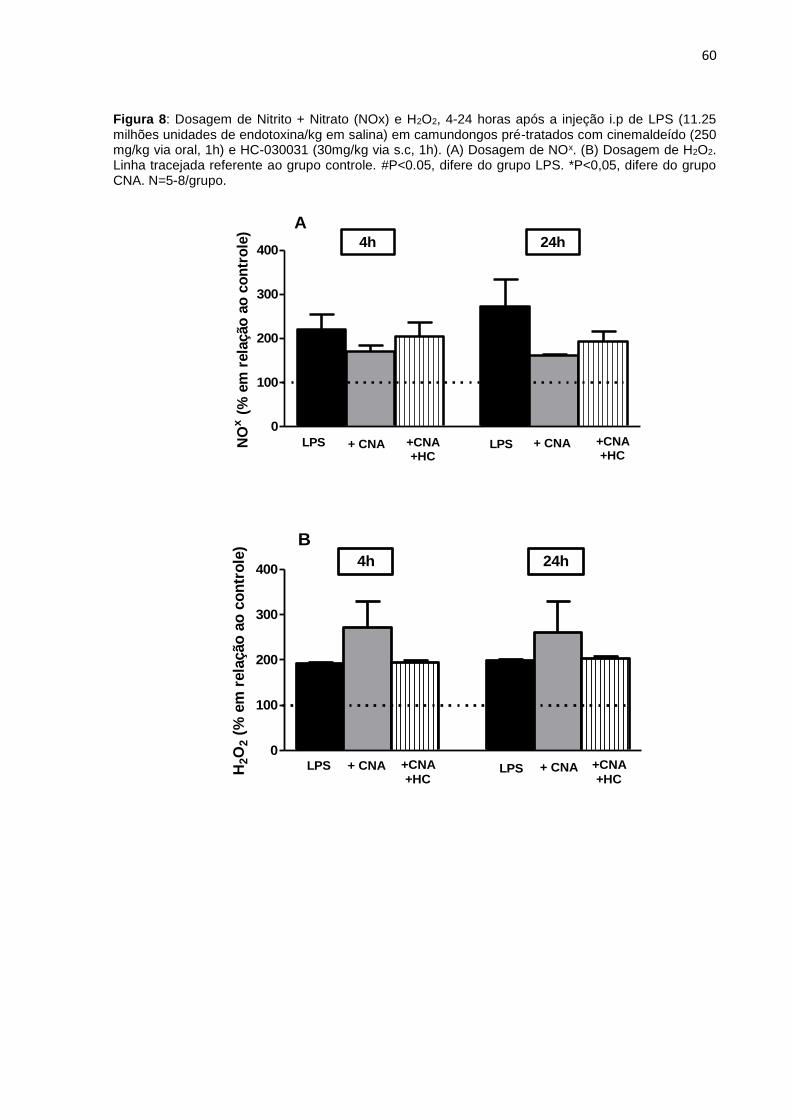
60
Figura 8: Dosagem de Nitrito + Nitrato (NOx) e H2O2, 4-24 horas após a injeção i.p de LPS (11.25 milhões unidades de endotoxina/kg em salina) em camundongos pré-tratados com cinemaldeído (250 mg/kg via oral, 1h) e HC-030031 (30mg/kg via s.c, 1h). (A) Dosagem de NOx. (B) Dosagem de H2O2. Linha tracejada referente ao grupo controle. #P<0.05, difere do grupo LPS. *P<0,05, difere do grupo CNA. N=5-8/grupo.
0
100
200
300
400
LPS + CNA +CNA+HC
LPS + CNA +CNA+HC
4h 24h
AN
Ox (
% e
m r
ela
ção
ao
co
ntr
ole
)
0
100
200
300
400
LPS + CNA +CNA+HC
LPS + CNA +CNA+HC
4h 24h
B
H2O
2 (
% e
m r
ela
ção
ao
co
ntr
ole
)

61
4.6 Avaliação de citocinas pró e anti-inflamatórias
Conforme foi exposto nesse trabalho, as citocinas desempenham um
papel principal na fisiopatologia da sepse. Dentre estas citocinas, TNF-α e IL-6 são
as mais extensivamente estudadas, ambas são capazes de ativar alvos celulares e
induzir a produção de mais mediadores inflamatórios, nesse sentido sabe-se que
seus níveis estão aumentados na sepse (WIERSINGA et al., 2014).
Foi quantificada a presença de citocinas plasmáticas e no lavado
peritoneal. Conforme ilustra a figura 9, a citocina pró-inflamatória TNF-α plasmática
avaliada 4 horas após a sepse tem seus níveis elevados em animais tratados com
LPS (Figura 9A). O tratamento agudo com CNA (250 mg/kg v.o, 1h) não alterou os
níveis de TNF-α, de igual modo, quando os animais foram pré-tratados com CNA
(250 mg/kg v.o, 1h) em conjunto com o antagonista seletivo dos receptores TRPA1,
HC (30mg/kg via s.c, 1h) não houve alteração significativa (Figura 9A). Já em 24
horas após a sepse, percebe-se que o pré-tratamento com CNA (250 mg/kg v.o, 1h)
reduziu os níveis plasmáticos de TNF-α de forma significativa (Figura 10A). Em
relação aos níveis de IL-1 no plasma, verificou-se uma diminuição desta citocina
quando os animais foram pré-tratados com CNA (250 mg/kg v.o, 1h) 4 horas após a
sepse. O uso do antagonista HC (30mg/kg via s.c, 1h) em cojunto com o CNA (250
mg/kg v.o, 1h) não alterou a IL-1 plasmática (Figura 9B). Em 24 horas a IL-1 não foi
detectada. O tratamento agudo com CNA (250 mg/kg v.o, 1h) não alterou os níveis
de IL-6 no plasma 4 horas após a sepse, todavia, quando administrado
conjuntamente com HC (30mg/kg via s.c, 1h) houve uma diminuição, comparado ao
grupo LPS (Figura 9C). Em 24 horas após a sepse não houve nenhuma alteração
significativa no perfil de IL-6 (Figura 10B).
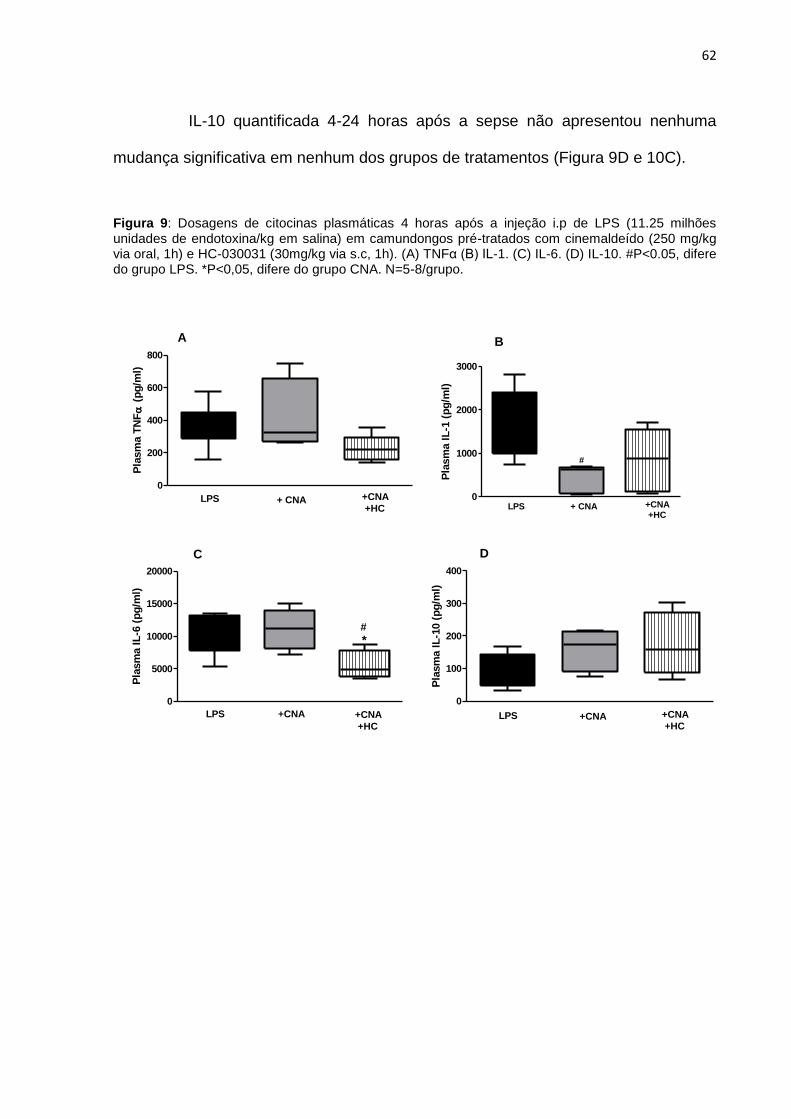
62
IL-10 quantificada 4-24 horas após a sepse não apresentou nenhuma
mudança significativa em nenhum dos grupos de tratamentos (Figura 9D e 10C).
Figura 9: Dosagens de citocinas plasmáticas 4 horas após a injeção i.p de LPS (11.25 milhões unidades de endotoxina/kg em salina) em camundongos pré-tratados com cinemaldeído (250 mg/kg via oral, 1h) e HC-030031 (30mg/kg via s.c, 1h). (A) TNFα (B) IL-1. (C) IL-6. (D) IL-10. #P<0.05, difere do grupo LPS. *P<0,05, difere do grupo CNA. N=5-8/grupo.
0
200
400
600
800
LPS + CNA +CNA
+HC
A
Pla
sm
a T
NF
(p
g/m
l)
0
1000
2000
3000
LPS + CNA +CNA+HC
B
#
Pla
sm
a I
L-1
(p
g/m
l)
0
5000
10000
15000
20000
LPS +CNA +CNA
+HC
C
*#
Pla
sm
a I
L-6
(p
g/m
l)
0
100
200
300
400
LPS +CNA +CNA
+HC
D
Pla
sm
a I
L-1
0 (
pg
/ml)

63
Figura 10: Dosagens de citocinas plasmáticas 24 horas após a injeção i.p de LPS (11.25 milhões unidades de endotoxina/kg em salina) em camundongos pré-tratados com cinemaldeído (250 mg/kg via oral, 1h) e HC-030031 (30mg/kg via s.c, 1h). (A) TNFα (B) IL-1. (C) IL-6. (D) IL-10. #P<0.05, difere do grupo LPS. *P<0,05, difere do grupo CNA. N=5-8/grupo.
0
50
100
150
LPS + CNA + CNA
+ HC
A
#
Pla
sm
a T
NF
(p
g/m
l)
0
100
200
300
400
LPS + CNA + CNA
+ HC
B
Pla
sm
a I
L-6
(p
g/m
l)
0
20
40
60
LPS + CNA + CNA
+ HC
C
Pla
sm
a I
L-1
0 (
pg
/ml)
Quantificou-se também a liberação de citocinas no exsudato peritoneal.
Conforme ilustra a figura 11 e 12, as citocinas dosadas 4-24 horas após a sepse não
sofreram mudanças significativas.
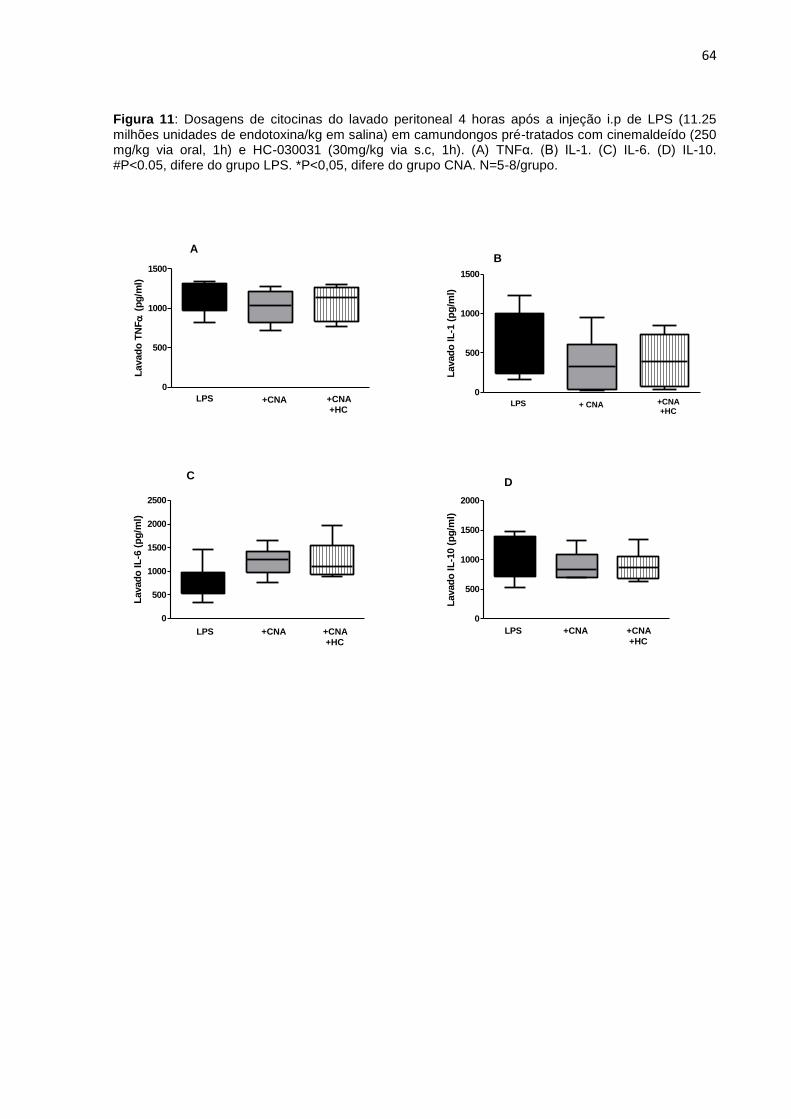
64
Figura 11: Dosagens de citocinas do lavado peritoneal 4 horas após a injeção i.p de LPS (11.25 milhões unidades de endotoxina/kg em salina) em camundongos pré-tratados com cinemaldeído (250 mg/kg via oral, 1h) e HC-030031 (30mg/kg via s.c, 1h). (A) TNFα. (B) IL-1. (C) IL-6. (D) IL-10. #P<0.05, difere do grupo LPS. *P<0,05, difere do grupo CNA. N=5-8/grupo.
0
500
1000
1500
LPS +CNA +CNA
+HC
A
Lavad
o T
NF
(p
g/m
l)
0
500
1000
1500
LPS + CNA +CNA+HC
B
Lavad
o I
L-1
(p
g/m
l)
0
500
1000
1500
2000
2500
LPS +CNA +CNA
+HC
C
Lavad
o I
L-6
(p
g/m
l)
0
500
1000
1500
2000
LPS +CNA +CNA
+HC
D
Lavad
o I
L-1
0 (
pg
/ml)
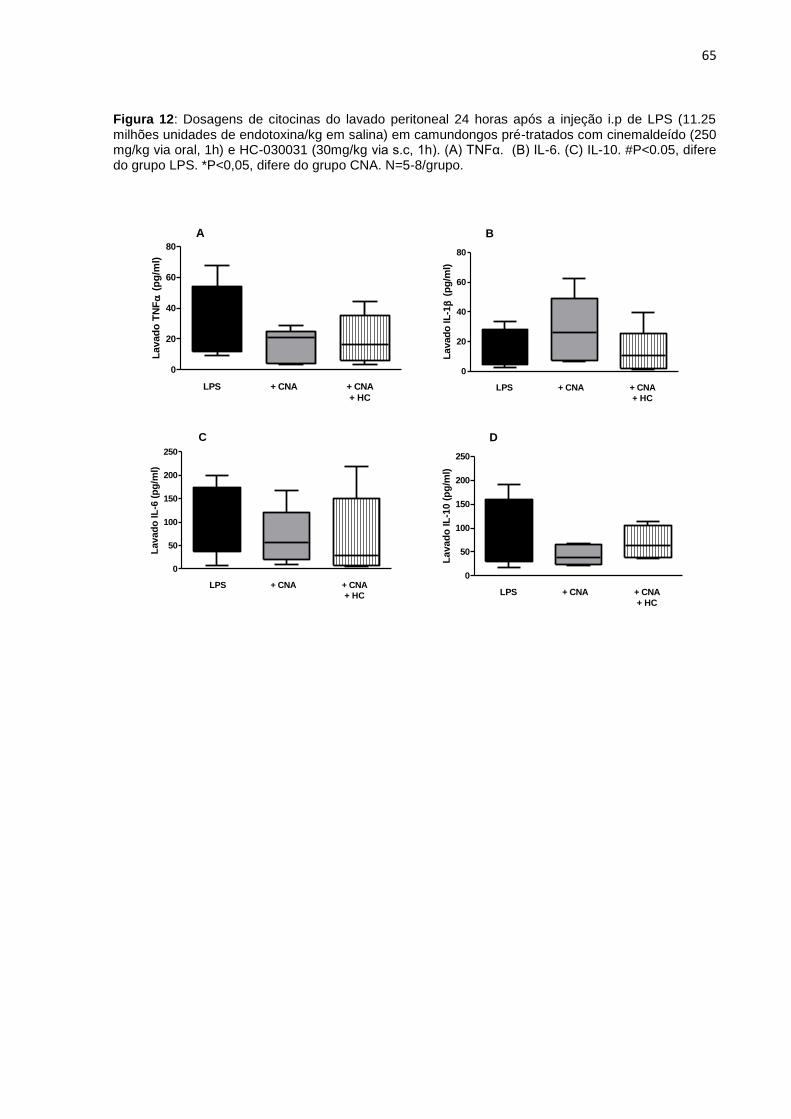
65
Figura 12: Dosagens de citocinas do lavado peritoneal 24 horas após a injeção i.p de LPS (11.25 milhões unidades de endotoxina/kg em salina) em camundongos pré-tratados com cinemaldeído (250 mg/kg via oral, 1h) e HC-030031 (30mg/kg via s.c, 1h). (A) TNFα. (B) IL-6. (C) IL-10. #P<0.05, difere do grupo LPS. *P<0,05, difere do grupo CNA. N=5-8/grupo.
0
20
40
60
80
LPS + CNA + CNA
+ HC
A
Lavad
o T
NF
(p
g/m
l)
0
20
40
60
80
LPS + CNA + CNA
+ HC
B
Lavad
o IL
-1
(p
g/m
l)
0
50
100
150
200
250
LPS + CNA + CNA
+ HC
C
Lavad
o IL
-6 (
pg
/ml)
0
50
100
150
200
250
LPS + CNA + CNA
+ HC
DL
avad
o IL
-10 (
pg
/ml)
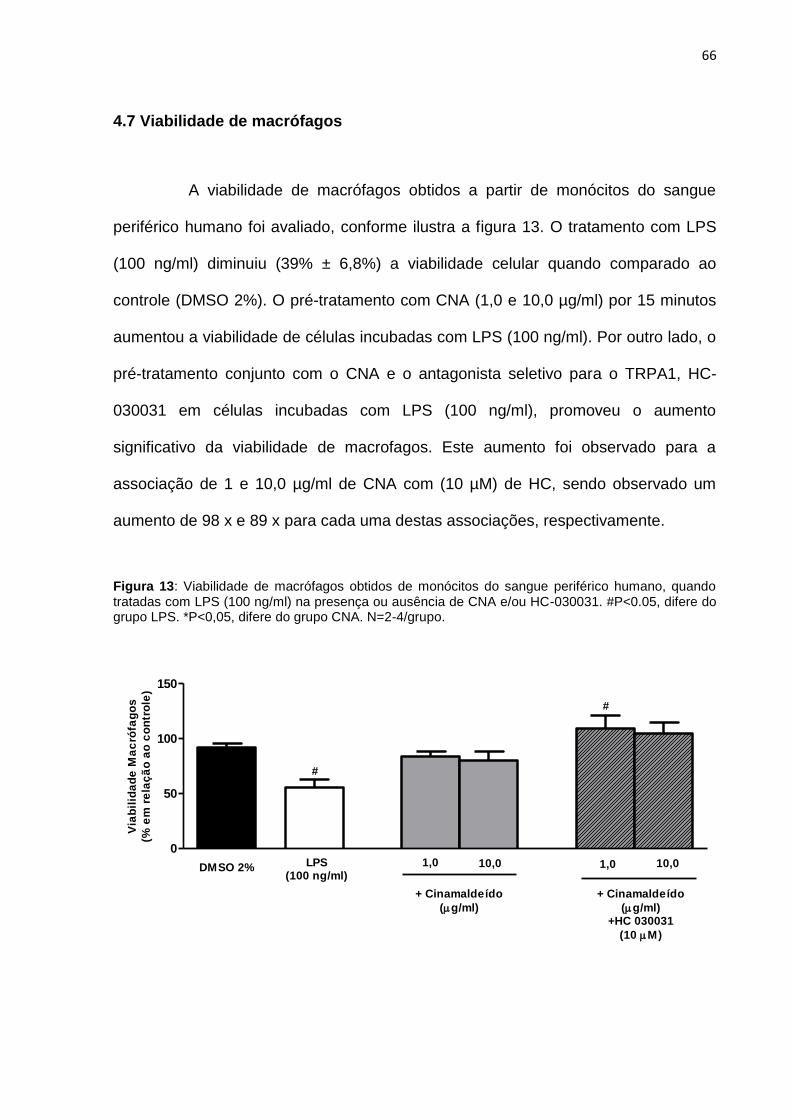
66
4.7 Viabilidade de macrófagos
A viabilidade de macrófagos obtidos a partir de monócitos do sangue
periférico humano foi avaliado, conforme ilustra a figura 13. O tratamento com LPS
(100 ng/ml) diminuiu (39% ± 6,8%) a viabilidade celular quando comparado ao
controle (DMSO 2%). O pré-tratamento com CNA (1,0 e 10,0 µg/ml) por 15 minutos
aumentou a viabilidade de células incubadas com LPS (100 ng/ml). Por outro lado, o
pré-tratamento conjunto com o CNA e o antagonista seletivo para o TRPA1, HC-
030031 em células incubadas com LPS (100 ng/ml), promoveu o aumento
significativo da viabilidade de macrofagos. Este aumento foi observado para a
associação de 1 e 10,0 µg/ml de CNA com (10 µM) de HC, sendo observado um
aumento de 98 x e 89 x para cada uma destas associações, respectivamente.
Figura 13: Viabilidade de macrófagos obtidos de monócitos do sangue periférico humano, quando tratadas com LPS (100 ng/ml) na presença ou ausência de CNA e/ou HC-030031. #P<0.05, difere do grupo LPS. *P<0,05, difere do grupo CNA. N=2-4/grupo.
0
50
100
150
DMSO 2% LPS(100 ng/ml)
1,0 10,0 1,0 10,0
+ Cinamaldeído
(g/ml)
+ Cinamaldeído
(g/ml)+HC 030031
(10 M)
#
#
Via
bil
ida
de
Ma
cró
fag
os
(% e
m r
ela
çã
o a
o c
on
tro
le)

67
5 DISCUSSÃO
Foram avaliados diversos parâmetros sistêmicos da sepse, como perda
de peso, alterações na temperatura corpórea e marcadores de falência de órgãos
vitais ao funcionamento do organismo, como fígado, pâncreas e rins. Os dados
obtidos neste trabalho demonstram que animais sépticos sofrem perda de peso e
hipotermia quando comparado com os valores basais, que foram associados com a
severidade da doença, caracterizada por redução da mobilidade, presença de
piloereção e secreção ocular, quando comparados com animais saudáveis. Este
trabalho demonstra pela primeira vez que o CNA é capaz de diminuir a severidade
da sepse e que este é um efeito bloqueado pelo tratamento com o antagonista do
receptor TRPA1, HC-030031, sugerindo que o CNA pode ser protetor na sepse por
mecanismos dependentes da ativação deste receptor. Em contraste, a perda de
peso e hipotermia permaneceu inalterada nestes animais, quando comparados com
camundongos sépticos e tratados com veículo. Curiosamente, a melhora na
severidade em animais pré-tratados com CNA foi abolida em animais pré-tratados
com o antagonista HC-030031. Granger et al., 2014 estudando a morbidade
causada pela sepse induzida por ligadura e punção cecal em camundongos BALB/c,
relatou a perda de peso já nas primeiras 24 horas após a infecção. Similarmente, em
um estudo realizado por Sordi, 2013 foi demonstrado que a inoculação intra-traqueal
de bactérias (K. pneumoniae) promoveu a perda de peso corporal (8%) nas
primeiras 24 horas. A perda de peso durante a sepse encontra-se relacionada à
diminuição da ingestão de alimentos e presença de diarreia. Ainda, ocorre perda da
auto-regulação endocrinometabólica. Assim, diversas moléculas importantes ao
balanço energético do organismo, são afetadas. Por exemplo, a ingestão de glicose

68
torna-se incapaz de reduzir a neoglicogêse e a lipólise, resultando em um
desequilíbrio no balanço anabolismo-catabolismo e aumento sérico da uréia,
creatinina, lipase, amônia e ácido úrico, que são excretados, em excesso, na urina
(FILHO et al., 2001).
A sepse é caracterizada pela SRIS decorrente de infecção e um dos
critérios clínicos estabelecidos em humanos é a temperatura inferior a 36ºC
(hipotermia) ou superior a 38ºC (febre), esta variabilidade de temperatura pode ser
explicada pelas citocinas liberadas durante a SRIS. Granger, 2014 avaliando a
temperatura corporal em animais sépticos também encontrou uma hipotermia até 24
horas após a sepse. A sepse e o choque séptico estão associados a mudanças no
equilíbrio da temperatura corporal. A febre é uma resposta esperada durante a
sepse e possui efeitos benéficos quando não se manifesta de modo exacerbado.
Estes efeitos incluem: a) impedir o crescimento de bactérias; b) ativação de células
T; c) produção de citocinas pelas células mononucleares (FUHONG et al., 2005). A
hipotermia, porém, pode ocorrer nos casos de choques sépticos e acredita-se que é
um agravante no prognóstico do paciente (THEILEN et al., 2007). Vários autores
sugerem que a hipotermia nos casos de infecção grave indica a impossibilidade do
organismo de combater o agente agressor. Porém, outros estudos sugerem que a
hipotermia é uma resposta controlada do corpo que muda o ponto de regulação do
hipotálamo, estimulando a diminuição da produção e aumentando a perda de calor.
Diante do exposto os mecanismos de proteção e os efeitos deletérios ainda não são
totalmente compreendidos, principalmente nos casos de infeção (SCAMMELL et al.,
1996).
Os resultados do presente estudo demonstram que o tratamento agudo
com CNA não afeta o peso dos animais com sepse (4-24 h). Em contraste, Wang,

69
2015 sugeriu que o pré-tratamento com óleo da canela (50 mg/kg) protege contra
danos na mucosa do intestino delgado em animais tratados com LPS. Ainda, o
mesmo estudo relatou o aumento da ingestão de comida e a diminuição da
incidência de diarreias em animais sépticos tratados com este óleo. No entanto,
seus estudos foram conduzidos em modelo suínos e o pré-tratamento com o óleo da
canela foi de forma crônica o que diferencia do modelo usado no presente trabalho.
Por outro lado, Camacho et al, 2015 relatou que o tratamento agudo com CNA (250
mg/kg) diminui a ingestão de comida por camundongos (C57BL6/J), por 2 horas
após o tratamento, efeito este, sugerido por estar associado a diminuição de grelina
(Hormônio responsável pela sensação de fome) nestes animais. Ainda, o mesmo
estudo sugeriu que este efeito é dependente da ativação do TRPA1. Nossos dados
indicam que a perda de peso em animais sépticos é independente da ativação deste
receptor. As discrepâncias entre os dados obtidos no presente trabalho e o de
Camacho et al. (2015), podem estar relacionadas aos modelos utilizados (i.e., sepse
x naive), o emprego ou não de jejum prévio de 23h, podendo o esvaziamento
gástrico interferir com a absorção do CNA e consequente ativação do TRPA1,
quando seus efeitos estão sendo avaliados em modelo de sepse inicial (4h).
O CNA e outros agonistas têm sido sugeridos como indutores da
termogênese por mecanismos dependentes da ativação do TRPA1, sem, entretanto,
induzirem a difusão de calor (MASAMOTO et al., 2009). Ainda, em um estudo
recente, Andersson et al. (2011) relataram que o TRPA1 é um importante mediador
das ações analgésicas do acetaminofeno embora sua participação nos efeitos
antipiréticos desta droga, ainda não tenha sido confirmada. Embora os dados da
literatura sugiram ações de regulação da temperatura corporal, tanto positivas
quanto negativas, os dados do presente estudo demonstram que o pré-tratamento

70
com CNA não apresenta efeitos sobre a hipotermia em camundongos com sepse
induzida por LPS. De fato, evidências têm sugerido que o uso de antagonistas
seletivos para TRPA1 não alteram a temperatura corporal em camundongos (CHEN
et al., 2011). Estes dados corroboram com os dados obtidos no presente estudo.
Enquanto que, as diferenças entre o presente estudo e os achados de Masamoto et
al., (2009) estão relacionadas aos modelos empregados (i.e., sepse x naive).
A redução de perfusão tecidual e consequente falência de órgãos são
características da evolução clínica da sepse, desde seu desencadeamento até que a
mesma se torne severa. Embora os mecanismos precisos que levam à esta
disfunção não estejam completamente elucidados, sabe-se que a falta de oxigênio
nos tecidos ocorre em decorrência da queda do fluxo sanguíneo nos capilares e da
redução do débito cardíaco, contribuindo de forma deletéria para o funcionamento
de órgãos (HENKIN et al., 2009). O que determina a falência de órgãos vitais, no
entanto, permanece ainda não esclarecido. Os dados do presente estudo e diversas
evidências demonstram que a injeção de LPS causa aumento plasmático de AST e
consequente dano ao fígado em camundongos (FERNANDES et al., 2012;
BAYRAKTAR et al., 2015). Dados obtidos em estudos anteriores demonstram que o
aumento de AST causado pelo LPS é diminuído pelo pré-tratamento com o óleo
essencial da Canela (50 mg/kg) em modelos suínos (WANG et al., 2015). Tung,
2011 também sugeriu uma proteção de CNA (170 mg/kg) contra o dano hepático
induzido pelo LPS em camundongos BALBc, indicando que essa atividade pode ser
por propriedades anti-inflamatórias do CNA. Em conjunto, estas evidências sugerem
um papel protetor para o CNA, quando avaliado em relação ao dano hepático
decorrente da sepse. Em nossos resultados observou-se uma diminuição nos níveis
de AST 4-24 horas após a sepse quando os animais foram pré-tratados com CNA,

71
entretanto esse achado não foi estatisticamente significativo. Ainda, observou-se
que o co-tratamento com HC-030031 é capaz de diminuir os níveis de AST
plasmático de forma significativa. A competição por ligação no TRPA1 pode
direcionar a resposta do LPS via TLR-4 e NOD, o que pode ter reduzido o dano
hepático em camundongos pré-tratados com CNA e HC, no entanto, não se pode
descartar a ideia de que a ação do CNA em reduzir o dano hepático possa ser via
TRPA1.
A hipoperfusão tecidual (com ou sem hipotensão) e a lesão inflamatória
resultam em um dano isquêmico e disfunção tubular renal. Há oligúria progressiva
com queda da taxa de filtração glomerular e elevação da creatinina (JÚNIOR et al.,
1998). A insuficiência renal aguda, associada a sepse, determina um prognóstico
extremamente reservado, com taxas de mortalidade de até 80% (AKAMINE et al.,
1994). Os dados do presente trabalho demonstram que o pré-tratamento com CNA
reduz os níveis creatinina em modelo de sepse inicial (4h), mas não tardia,
sugerindo um efeito protetor aos rins. Esses achados são corroborados por dados
publicados anteriormente. De fato, Sayed et al. (2013) demonstraram que
camundongos com nefrotoxicidade induzida por cisplatina e níveis de creatinina
elevados, apresentam uma melhora na função renal com o pré-tratamento de CNA
(40 mg/kg, v.o.). O mesmo estudo sugere que este efeito encontra-se relacionado as
propriedades antioxidantes do CNA (SAYED et al., 2013). Por outro lado, os dados
do presente estudo demonstram que o CNA promove aumento da produção de H2O2
em animais sépticos, não podendo relacionar a proteção renal observada, com as
propriedades antioxidantes descritas anteriormente para este composto. É possível
que no choque séptico, o CNA possa regular diferentes vias envolvidas na proteção
renal, que contrabalanceiem os efeitos deletérios do aumento da produção de EROS

72
por este composto, como modulação da distribuição de fases do ciclo celular,
protegendo as células renais de efeitos tóxicos como, por exemplo, o causado pelo
LPS (PATRA et al., 2012). Ainda, a co-administração do CNA com o antagonista
seletivo dos receptores TRPA1, HC-030031, promoveu uma diminuição ainda mais
acentuada dos níveis de creatinina em animais sépticos. Este efeito sugere que uma
potencial competição pelo sitio de ligação ao TRPA1 possa estar ocorrendo entre
LPS, CNA e HC-030031.
A lipase sérica é largamente utilizada como marcador de inflamação e
pode estar elevada em algumas situações que mimetizam pancreatite (BYRNE et al.,
2002). A patogênese da pancreatite se estabelece a partir de uma exposição a um
fator casual, o qual desencadeia uma série de eventos que culminam com a
inflamação do pâncreas. A sepse e outros fatores podem desencadear uma
pancreatite aguda muito em função da inflamação pancreática causada pela SRIS
(CARNEIRO et al., 2004).
Os níveis de lipase no presente estudo mostram um dano pancreático
bem acentuado 4-24 horas após a sepse. Estudos têm sugerido o aumento dessa
enzima em animais sépticos (FERNANDES et al., 2012) e em humanos com
pancreatite causada pelo choque séptico (PEZZILLI et al., 2011). Recentemente
mostrou-se que o TRPA1 está envolvido na liberação de insulina por células β-
pancreáticas (CAO et al., 2002). No entanto, nem o pré-tratamento com CNA nem
em conjunto com HC alterou os níveis dessa enzima, sugerindo que não há
participação de TRPA1 na regulação do dano pancreático na sepse murina induzida
pelo LPS.
Para um melhor entendimento sobre o impacto do pré-tratamento utilizado
e o modelo experimental, avaliou-se as alterações inflamatórias circulantes e locais

73
(peritônio), como a produção de mediadores inflamatórios e alterações em
populações celulares. Em decorrência da sepse, é característica a alteração das
populações de células inflamatórias circulantes, assim como daquelas no sítio inicial
da infecção (i.e., células mononucleares e polimorfonucleares); como parte da
resposta imune inata do hospedeiro frente à infecção (WIERSINGA et al., 2011). De
fato, a patogênese da sepse está relacionada a uma disfunção generalizada da
resposta imune/inflamatória, sobretudo no âmbito celular, a qual implica no aumento
da morbidade dos pacientes acometidos (LUAN et al., 2013). No presente estudo,
foram avaliadas as populações de células inflamatórias em amostras de sangue
periférico e de lavado peritoneal. Nossos resultados mostram que em 24 horas após
a sepse ocorre um aumento acentuado do número de células mononucleares no
sangue. De fato, com o agravamento da infecção e a migração de células
inflamatórias para o sitio inicial de infecção, mais células são recrutadas da medula
óssea, ocasionando o aumento do nível circulante destas células (COWAN et al.,
2015). Ainda, endotoxinas liberadas pelas bactérias, promovem a liberação de
diferentes mediadores inflamatórios, principalmente por monócitos e macrófagos e
consequentemente a ativação e recrutamento dessas células para o sítio infeccioso
(BOOMER et al., 2014).
Células segmentadas também desempenham um importante papel na
erradicação ao agente causador. A contagem de neutrófilos totais no sangue pode
cair com a sepse aguda, em resposta, esses neutrófilos são cada vez liberados a
partir da medula óssea, estimulados pela presença do foco infeccioso, a liberação de
endotoxinas, a produção de citocinas e a ativação da cascata da coagulação
ativando assim os leucócitos polimorfonucleares e ampliando a resposta inflamatória
(RUDIGER et al., 2008). A estimulação destas células aumenta o consumo de

74
oxigênio, através do aumento da atividade de NADPH-oxidase que produz formas
ativas de oxigênio, incluindo os radicais peróxido e superóxido que são essenciais
durante a sepse no combate as bactérias fagocitadas (JÚNIOR et al., 1998).
O aumento dessas células na corrente sanguínea está associada a
evolução do processo infeccioso no tecido. As citocinas e quimiocinas que são
secretadas no sítio da infecção têm múltiplos efeitos sobre as células endoteliais
vasculares, os leucócitos e a medula óssea, que, juntos, aumentam a concentração
local de células que podem combater os patógenos e reparar o tecido (LUAN et al.,
2014). Houve também alterações em células do peritônio, os dados do presente
estudo demonstram que o LPS promove uma redução significativa do número de
células mononucleares peritoniais 24h após a sua injeção. Dados anteriores
sugerem que esta redução pode estar relacionada ao aumento da expressão e
ativação de mecanismos de apoptose celular desencadeados pelo LPS (SOLDATO
et al., 2007) ou ao fato destas células migrarem para órgãos linfoides secundários a
fim de apresentarem antígenos e consequentemente ativarem respostas especificas
ao LPS (PLATZER et al., 2014). De fato, a sepse é uma síndrome altamente
complexa, com múltiplas vias de sinalização que medeiam a produção de
mediadores inflamatórios, e também podem promover a ativação da apoptose e
acelerar eventos inflamatórios (LUAN et al., 2014). Por outro lado, o número de
PMNs peritoneais permanece similar em 4h e 24h após o LPS, enquanto que o
numero de PMNs sanguíneos encontra-se mais elevado em 24h quando comparado
com 4h após a indução da sepse, sugerindo que estas células continuam sendo
recrutadas em 24h e que as mesmas estejam exercendo um papel de relevância na
resposta ao LPS no modelo utilizado. Desempenhando funções de fagocitose,
liberação de EROS, através da oxidase fagocitária, além de enzimas proteolíticas

75
nos fagolisossomos que destroem microorganismos, como, por exemplo, a elastase
(KRUGER et al., 2015).
Evidências sugerem que o CNA é capaz de modular a migração celular
em diferentes modelos de inflamação (WALTER et al., 2014; LIAO et al., 2008;
HONG et al., 2012). Neste trabalho avaliou-se o efeito do tratamento agudo com
CNA no número de leucócitos e na migração de células inflamatórias para o sítio
primário de infecção (peritônio). Os dados do presente trabalho demonstram que o
CNA reduz o número de células mononucleares circulantes, e ao mesmo tempo,
aumenta o numero de células mononucleares no peritônio. Como citado
anteriormente, o efeito deletério do LPS tem sido relatado durante o choque
endotóxico, induzindo apoptose em células como consequência da liberação do
TNF-α e a produção de NO liberado por macrófagos (XAUS et al., 2000). Kim, 2010
mostrou que o CNA protege células da citotoxicidade induzida por LPS, em grande
parte por supressão da produção de NO. Em contraste, em um estudo in vitro
realizado por Liao, 2008, demonstrou-se que a incubação do CNA (50 ou 100 µM)
em co-cultura de monócitos e células endoteliais estimulados com TNFα, diminui a
expressão de moléculas de adesão (ICAM-1 e VCAM-1), sugerindo um efeito
inibidor da migração de monócitos por este composto. As diferenças entre estes
achados e os do presente trabalho, podem estar relacionados à diferenças no
estímulo inflamatório (i.e., LPS x TNF), além de diferenças sistemáticas entre os
diferentes modelos inflamatórios utilizados nos estudos (i.e.; in vivo x in vitro). A
princípio, os dados do presente estudo sugeriram que o CNA promove recrutamento
de células mononucleares sanguíneas de forma dependente da ativação do TRPA1
e que o recrutamento e/ou sobrevivência destas células na cavidade peritoneal
parece ser independente da ativação deste receptor. No entanto, a partir da analise

76
fenotípica da população de células mononucleares, observou-se que o TRPA1 é um
importante regulador da atividade do CNA sobre macrófagos, mas não linfócitos do
tipo CD4+ ou CD8+.
De forma interessante, a análise fenotípica de macrófagos peritoneais
demonstrou que o CNA aumenta o número de células Ly6Chigh e Ly6Clow. Ly6C é
uma glicoproteína expressa principalmente em células hematopoiéticas, as quais
migram para o sitio inflamatório na presença de um estimulo. Monócitos Ly6Chigh e
Ly6Clow se diferenciam em macrófagos e adptam ao perfil M1 e M2, respectivamente
(TACKE et al., 2014). Ainda, os dados do presente trabalho demonstram que a
capacidade regulatória positiva do CNA sobre ambas estas populações de
macrófagos é bloqueada pelo antagonismo do receptor TRPA1. Estas são as
primeiras evidencias de que o receptor TRPA1 pode influenciar o perfil de
diferenciação e o número de células inflamatórias, em especial, os macrófagos.
Ainda, em 24h após a sepse, observou-se um aumento de células segmentadas no
lavado peritoneal de camundongos pré-tratados com CNA, o qual pode ser revertido
pelo antagonismo do receptor TRPA1, sugerindo mais uma vez, a importância da
participação deste receptor para as ações do CNA sobre as células inflamatórias.
Sugere-se que este efeito do CNA sobre as populações de células inflamatórias está
diretamente relacionado com o efeito protetor deste composto na severidade da
sepse. De fato, estas células inflamatórias são capazes de produzir uma série de
mediadores inflamatórios na tentativa de erradicar a infecção, os quais podem ser
benéficos ou deletérios dependendo do estagio de evolução desta síndrome, bem
como de sua severidade (CHAUDHRY, et al., 2013).
Dentre este mediadores inflamatórios, destacam-se as EROS. As suas
propriedades pró-inflamatórias incluem dano às células endoteliais, formação de

77
fatores quimiotáticos, recrutamento de neutrófilos, dano ao DNA e formação de
peroxinitrito (STECKERT et al., 2014). O peroxinitrito possui vários efeitos citotóxicos
e pró-inflamatórios independentes que levam a dano celular irreversível, como
evidenciado no choque séptico (BERG et al., 2011). O “burst” oxidativo presente no
início da sepse é comandado pela ação de neutrófilos e macrófagos. Estas células
fazem parte do sistema imune inato e são a primeira linha de defesa do hospedeiro
contra patógenos. Sendo assim, eles liberam diferentes EROS, como, por exemplo,
O2- e NO.
O NO é produzido pelas enzimas NOS, principalmente, pela isoforma
induzida (NOSi), sua expressão pode ser induzida após a ativação de células como
o musculo liso, macrófago e célula endotelial, por bactérias, LPS e citocinas pró-
inflamatórias (IL-1 e TNF) (CHANDRA et al., 2006). Existe um número substancial de
trabalhos indicando que o NO é um dos principais efetores na sepse e no choque
séptico (BEISHUIZEN et al., 1998; TITHERADGE, 1999; PARRATT, 1998; FEIHL et
al., 2001; FLEMING et al., 1991). Embora o NO seja uma importante ferramenta do
hospedeiro contra infecções, a liberação em excesso pode causar efeitos
cardiovasculares indesejáveis, como diminuição do tônus vascular e efeitos
citotóxicos através da reação com ONOO- e dióxido de nitrogênio (NO2) (PARRATT,
1998).
Nesse sentido, nossos resultados mostram uma grande liberação de NO
em células peritoneais de camundongos tratados com LPS. Fernandes, 2012
também encontrou um aumento desses mediadores em animais sépticos.
Recentemente Lee, 2015 mostrou um aumento de nitrato/nitrito em animais sépicos,
contudo, o pré-tratamento do óleo essencial de Cinnamomum osmophloeum
Kanehira reduziu estes níveis com uma dose de 6,5 mg/kg. Trabalhos apontam um

78
efeito supressor do NO pelo CNA. Macrófagos murinos estimulados com LPS
tiveram uma diminuição na produção de NO quando incubados com concentrações
de CNA >1 mg/ml (WALTER et al., 2014). Ainda, Kim, 2010 mostrou uma forte
supressão de NO em células (RAW264.7) tratadas com CNA. Trabalhos têm
sugerido que estes efeitos do CNA sobre a produção de NO ocorrem através da
supressão do NF-kB (KIM et al., 2010) e/ou inibição da atividade da NOSi (PANNEE,
2014). No presente estudo não houve uma ação inibitória estatisticamente
significante do CNA sobre a produção deste mediador. A falta de trabalhos avaliando
o potencial do CNA em modelos experimentais de sepse limita o melhor
entendimento acerca dos efeitos do CNA e consequente participação da ativação do
TRPA1 na produção de NO.
Também foi avaliada a liberação de H2O2 no lavado peritoneal. O H2O2 é
um pró-oxidante estável e possui um papel importante na fisiopatologia da sepse
(ANDRADES et al., 2011). O H2O2 teve seus níveis elevados em camundongos
tratados com LPS. Fernandes, 2012 utilizando modelo de sepse experimental
também descreveu um aumento dos níveis de H2O2 no lavado peritoneal em animais
sépticos. O aumento de H2O2 faz parte do arsenal liberado pelas células
inflamatórias durante o combate ao agente agressor e é um importante agente
microbicida (ANDRADES et al., 2011). Tem sido relatada a ativação do receptor
TLR4 por LPS dependente da ativação de PI3K, sugerindo que, quando o TLR4 é
ativado recruta o complexo NADPH oxidase, este complexo é, então, capaz de gerar
EROS sob a forma de peróxido de hidrogênio (NGKELO et al., 2012).
Nossos resultados mostram que o pré-tratamento de CNA não alterou os
níveis de H2O2, apesar de um aumento visível, porém, não significativo. Liao, 2010
relata a influencia do CNA em células endoteliais no sistema redox da célula,

79
dividindo-o em dois estágios que são dependentes da duração do tratamento, no
primeiro momento, 1 hora após o tratamento com CNA (100µM), há diminuição de
glutationa (molécula antioxidante responsável pela metabolização do H2O2) e
consequente aumento de H2O2, no segundo estágio (9 horas após o tratamento com
CNA) a glutationa volta a níveis basais reagindo com radicais hidroxil. Portanto
sugere-se que o CNA possa estar envolvido na modulação redox através da
ativação de enzimas antioxidantes sendo esse mecanismo dependente da duração
do tratamento, acredita-se que o TRPA1 também possa estar envolvido nessa
modulação redox uma vez que tem sido demostrado que ROS também ativam
TRPA1 (ANDERSSON & BEVAN, 2008). Sendo assim, torna-se mais complexo o
entendimento sobre a ativação de TRPA1 uma vez que não houve alteração com o
pré-tratamento conjunto HC e CNA nos níveis de H2O2.
A ativação celular, ativação de sistemas séricos e dos leucócitos, contribui
para a consequente liberação de mediadores endógenos (citocinas, radicais reativos
do oxigênio e do nitrogênio e mediadores lipídicos). Dentre esses mediadores
endógenos, liberados, primeiramente, por células residentes e, posteriormente por
células recrutadas para o foco infeccioso, as citocinas desempenham um papel
primordial na resposta do hospedeiro (BENJAMIM, 2004). As mais extensivamente
citocinas estudadas na sepse são TNF e IL-1, ambas são capazes de ativar alvos
celulares e induzir a produção de mais mediadores inflamatórios. Outras citocinas
também são importantes na resposta do hospedeiro, a IL-6 e IL-10.
O TNFα é um mediador conhecido e muito importante na sepse, sendo
reportado em vários estudos, incluindo modelos murinos. A administração de LPS
resulta na produção e liberação de TNF na circulação sistêmica onde os picos de
concentrações deste mediador são encontrados em 60-90 minutos após (SCHULTE

80
et al., 2013). O TNF e IL-1 aumentam a expressão de moléculas de adesão,
proporcionam a aderência dos neutrófilos a integrina e promove seu extravasamento
no tecido. A IL-6 tem uma variedade de efeitos biológicos, incluindo a ativação de
linfócitos B e T, sistemas de coagulação e modulação da hematopoese (BORDEN &
CHIN, 1994). Efeitos protetores têm sido atribuídos a esta citocina em modelos de
endotoxemia (YOSHIZAWA et al., 1996). Ao contrário da IL-1 e do TNF, a
administração per se de IL-6 não produz um quadro de SRIS (SCHMARTZ et al.,
1991), sendo sua função chave, mecanismos de regulação da febre e mediação da
resposta de fase aguda (CHAI et al., 1996). Apesar de suas propriedades pro-
inflamatórias, a IL-6 também pode promover uma resposta anti-inflamatória, inibindo
a liberação de TNF e IL-1, e a ativação de células inflamatórias (SCHINDLER et al.,
1990; POLL et al., 1994).
Estudos in vitro tem reportado o efeito anti-inflamatório do CNA, reduzindo
a liberação de citocinas como TNF, IL-1 e IL-6 (PANNEE, 2014). Liao e
colaboradores mostrou que o CNA reduz os níveis de TNF induzido por LPS em
macrófagos murinos, assim como em modelo de edema de pata induzida por
carragenina. Estudos in vivo com o extrato da canela em modelos de endotoxemia,
relatam a diminuição sérica de TNFα com doses de 20 e 100 mg/kg (HONG et al.,
2012). Ainda, estudos com o óleo da canela demonstram uma redução nos níveis de
IL-1β e TNFα quando administrados em animais sépticos nas doses de 13 e 26
mg/kg (LEE et al., 2015). Nossos resultados corroboram com os achados acima,
visto que, o CNA reduziu os níveis de TNFα e IL-1β a nível sistêmico.
A participação dos receptores TRPA1 nas doenças inflamatórias ainda
permanece obscuro, alguns estudos indicam efeitos pro-inflamatórios e anti-
inflamatórios, ou sem influência (KUN et al., 2014). Engel, 2012 relata que a ativação

81
basal de TRPA1 reflete na ação pró-inflamatória do cólon através da liberação de
substancia P, em contraste, condições inflamatórias agudas propõe uma mudança
na função de TRPA1 no sentido de melhorar a inflamação. Bonet, 2013 trabalhando
com modelo de hiperalgesia induzida por carragenina relatou que o bloqueio de
TRPA1 via HC não alterou a expressão de citocinas. Contudo, nosso estudo mostra
que o uso conjunto do HC e CNA diminuiu IL-6 diferentemente das outras citocinas.
A IL-10 é produzida por muitos tipos de células imunes, tais como,
monócitos, macrófagos, linfócitos B e T e células NK (LATIFI et al., 2002). Estudos
funcionais revelam um efeito anti-inflamatório de IL-10, in vitro IL-10 suprime a
produção de IL-1, TNF e IL-6 (MALEFYT et al., 1991). Em modelos experimentais a
administração de IL-10 tem protegido contra efeitos letais em modelos de
endotoxemia (HOWARD et al., 1993). Pannee, 2014 mostrou um aumento de
mediadores anti-inflamatórios (IL-10 e TGF-β) in vitro quando as células foram
tratadas com CNA e LPS. Lin, 2011 também relata efeito modulador do CNA na
liberação de IL-10. No presente estudo, observa-se que não houve alteração na
produção de IL-10 quando os animais foram pré-tratados com CNA. As diferenças
observadas entre os achados do presente estudo e os autores citados acima podem
estar relacionado aos diferentes modelos inflamatórios utilizados nos estudos (i.e.; in
vivo x in vitro).
Como mencionado anteriormente, o LPS causa apoptose em vários tipos
celulares (PARK et al., 2013). Alterações na viabilidade celular podem refletir em
uma resposta imune não eficiente, podendo acarretar prejuízos na sepse. Estudos
relatam que a morte celular por apoptose contribui para a disfunção imune e falha
múltipla de órgãos observada na sepse, e que o bloqueio desta morte celular pode
melhorar a sobrevida de animais em modelos experimentais de sepse (AYALA et al.,

82
1998; HOTCHKISS et al., 1999). Experimentos in vitro realizados no presente estudo
sugerem que o CNA aumenta a viabilidade de macrófagos humanos estimulados
com LPS, embora este efeito não tenha sido significativo. De fato, foi sugerido
anteriormente que o CNA protege macrófagos da citotoxicidade induzida pelo LPS
por inibir a produção de NO via ativação de NF-B e por interferir com as fases do
ciclo celular (KIM et al., 2010; PATRA et al., 2012). Ainda, Walter, 2014 avaliando a
viabilidade de células cancerígenas, observou que o tratamento com CNA (1µg/ml) e
LPS (5 µg/ml) aumentou a viabilidade celular em células Jurkat E6-1 obtidas de
monócitos humanos, enquanto que, em outras linhagens celular essa concentração
não interferiu na viabilidade celular. Por outro lado, Pannee, 2014 analisou a
viabilidade de macrofagos J774A.1 tratados com CNA (1.25 -20 µg/ml) e LPS
(100ng/ml), de modo que não observou diferença na viabilidade celular em nenhuma
das concentrações estudadas de CNA. Sugere-se que as diferenças observadas
nestes estudos estão relacionadas as diferentes linhagens celulares assim como a
diferentes concentrações de LPS utilizadas nestes estudos.
A participação de TRPA1 na viabilidade celular ainda permanece não
esclarecida. Dados encontrados no presente trabalho indicam um aumento na
viabilidade celular quando as células foram pré-tratadas conjuntamente com CNA (1
µg/ml; 10 µg/ml) e HC (10 µM). Este resultado sugere mais uma vez que a
competição destes 3 compostos por sítios de ligação no TRPA1, possam deslocar a
ação do LPS para que a mesma se restrinja a receptores TLR-4 e NOD. Nossos
dados reforçam a complexidade desta síndrome e a dificuldade para controlar todos
os eventos patofisiológicos da doença através de um tratamento adequado, já que
no início do processo, a resposta inflamatória é intensa com o intuito de controlar a
infeção e mais tardiamente deve ser regulada para não prejudicar o paciente. O

83
CNA demonstra potenciais ações anti-inflamatórias que podem ajudar a controlar e
equilibrar esses eventos na sepse. O aumento/manutenção de alguns tipos celulares
como macrófagos e linfócitos no local da infeção, o efeito protetor frente ao dano
renal, a redução de NO embora não seja significativa, e a regulação de algumas
citocinas contribui de forma positiva no controle dessa síndrome.

84
6 CONCLUSÃO
O presente estudo demonstra que o CNA possui propriedades protetoras
na sepse. A administração de CNA atenua a falência hepática além de alterar o perfil
de células inflamatórias circulantes e também presentes no lavado peritoneal de
animais sépticos, ainda o CNA promove o aumento da viabilidade de células
inflamatórias peritoneais como as Ly6C+ o qual é acompanhado por um aumento no
número absoluto de células PMNs. O CNA ainda, promove a diminuição nos níveis
de NO no lavado peritoneal, diminuição dos níveis circulantes de IL-1β e TNF-α. O
antagonismo do TRPA1 pelo tratamento com HC-030031 é capaz de reverter alguns
dos efeitos do CNA, como, o perfil de células inflamatórias circulantes e do lavado
peritoneal (Ly6C+). Em conjunto, os dados do presente trabalho sugerem que o CNA
é capaz de modular a sepse por mecanismos dependentes e independentes da
ativação do TRPA1. Ressalta-se que este trabalho fornece os primeiros indícios de
que o TRPA1 pode representar uma molécula moduladora na sepse, além de ser de
grande relevância para os efeitos antiinflamatórios do CNA. No entanto, mais
estudos são necessários para melhor esclarecer o potencial antiinflamatório do CNA
e o real papel do TRPA1 na sepse.

85
REFERÊNCIAS
AKAMINE, N., FERNANDES JR.C.J., WEY, S. B., KNOBEL, E., KNOBEL, E. Choque séptico. Condutas no paciente grave. Editora Atheneu, São Paulo, p. 175-210, 1994.
ALEJANDRA G. GARRIDO, LUIZ FRANCISCO POLI DE FIGUEIREDO, MAURÍCIO ROCHA E SILVA. Experimental models of sepsis and septic shock: an overview. Acta Cirúrgica Brasileira - Vol 19 (2) 2004.
ANDERSSON, D.A., GENTRY, C., ALENMYR, L., KILLANDER, D., LEWIS, S. E., ANDERSSON, A., BUCHER, B., GALZI, J. L., STERNER, O., BEVAN, S., HÖGESTÄTT, E. D., ZYGMUNT, P. M. TRPA1 mediates spinal antinociception induced by acetaminophen and the cannabinoid Δ (9)-tetrahydrocannabiorcol. Nature Communications, p. 222-551, 2011.
ANDRADES, M.E., MORINA, A., SPASIĆ, S., SPASOJEVIĆ, I. Bench-to-bedside review: Sepsis - from the redox point of view. Critical Care. P. 215:230, 2011.
ANGUS, D. C., LINDE-ZWIRBLE, W. T., LIDICKER, J., et al. Epidemiology of severe sepsis in the United States: analysis of incidence, outcome, and associated costs of care. Critical Care Medicine, p. 1303-1310, 2001.
ARNALICH, F., GARCIA-PALOMERO, E., LOPEZ, J. Predictive value of NFkB activity and plasma cytokine levels in patients with sepsis. Infection and Immunity. p. 1942–1945, 2000.
ASEHNOUNE, K., STRASSHEIM, D., MITRA, S., KIM, J.Y., ABRAHAM, E. Involvement of Reactive Oxygen Species in Toll-Like Receptor 4-Dependent Activation of NF-B1. Journal of Immunology. P. 2522-2529, 2004.
AYALA, A., EVANS, T.A., CHAUDRY, I.H., Does hepatocellular injury in sepsis involve apoptosis? The journal of surgical Reseach. P. 165–173, 1998.
BANDELL, M., STORY, G.M., HWANG, S.W., VISWANATH, V., EID, S.R., PETRUS, M.J., EARLEY, T.J., PATAPOUTIAN, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron. P. 849-57, 2014.
BARTHLEN, W., ZANTL, N., PFEFFER, K., HEIDECKE, C. D., HOLZMANN, B., STADLER, J. Impact of experimental peritonitis on bone marrow cell function. Surgery V. 126, p. 41–47, 1999.
BASILE-FILHO, A., SUEN, V.M.M., MARTINS, M.A., COLETTO, F.A., MARSON, F. Monitorização da resposta orgânica ao trauma e a sepse. Medicina (Ribeirao Preto. Online), v. 34, n. 1, 2001.

86
BAYALA, B., BASSOLE, I.H.N., SCIFO, R., GNOULA, C., MOREL, L., LOBACCARO, J.A., SIMPORE, J. Anticancer activity of essential oils and their chemical components - a review. American Journal of Cancer Reseach. P. 591-607, 2014
BAYRAKTAR, O., TEKIN, N., AYDIN, O., AKYUZ, F., MUSMUL, A., BURUKOGLU, D. Effects of S-allyl cysteine on lung and liver tissue in a rat model of lipopolysaccharide-induced sepsis. Naunyn-Schmiedererg’s archives of pharmacology. P. 327-350, 2015.
BENJAMIM, C.F. Atualização sobre mediadores e modelos experimentais de sepse. Revista de Medicina de Riberão Preto. V. 34, p. 18-26, 2001.
BENJAMIM, C.F., LUNDY, S.K., LUKACS, N.W., HOGABOAM, C.M., KUNKEL, S.L. Reversal of long-term sepsis-induced immunosuppression by dendritic cells. Blood. P. 88-95, 2004.
BERG, R.M., MØLLER, K., BAILEY, D.M. Neuro-oxidative-nitrosative stress in sepsis. Journal of Cerebral Blood Flow and Metabolism. p. 1532-1544, 2011.
BODKIN, J.V., FERNANDES, E.S. TRPV1 and SP: key elements for sepsis outcome?. British Journal of Pharmacology. P. 1279–1292, 2013.
BONE, R.C., NEWTON, I. Sepsis, SIRS and CARS. Critical Care Medicine, v. 24, n. 7, p. 1125-1128, 1996.
BONET, I.J., FISCHER, L., PARADA, C.A., TAMBELI, C.H. The role of transient receptor potential A 1 (TRPA1) in the development and maintenance of carrageenan-induced hyperalgesia. Neuropharmacology. V. 65, 2013.
BOOMER, J.S., GREEN, J.M., HOTCHKISS, R.S. The changing immune system in sepsis: is individualized immuno-modulatory therapy the answer?. Virulence. P. 45-56, 2014.
BOOMER, J.S., JONATHAN, M. GREEN, HOTCHKISS, R.S. The changing immune system in sepsis Is individualized immuno-modulatory therapy the answer?. Virulence. P. 45–56, 2014.
BOSMANN, M., WARD, P.A. The inflammatory response in sepsis. Trends in Immunology. P.129-136, 2013.
BRUMATTI, G., SHERIDAN, C., MARTIN, S.J. Expression and purification of recombinant annexin V for the detection of membrane alterations on apoptotic cells. Methods. P. 235-240, 2008.
BULLOCK, W. The History of Bacteriology. London: University Press. 258 p, 1960.
BURDOCK, G.A. Review of the biological properties and toxicity of bee propolis (propolis). Food and Chemical Toxicology. P. 347–636, 1998.

87
BYRNE, M.F., MITCHELL, R.M., STIFFLER, H., JOWELL, P.S., BRANCH, M.S., PAPPAS, T.N. Extensive investigation of patients with mild elevations of serum amylase and/or lipase is ‘low yield’. Cancer Journal of Gastroenterology. P. 849-854, 2002.
BYUNG, H.U.N., LEE, K.Y.G., LEE, J., LEE, J.Y., CHO, J.Y. Regulatory Effect of Cinnamaldehyde on Monocyte/Macrophage-Mediated Inflammatory Responses. Mediators of inflammation. V. 2010, p. 529 – 359, 2010.
CAMACHO S, MICHLIG S, DE SENARCLENS-BEZENÇON C, MEYLAN J, MEYSTRE J, PEZZOLI M, MARKRAM H, LE COUTRE J. Anti-obesity and anti-hyperglycemic effects of cinnamaldehyde via altered ghrelin secretion and functional impact on food intake and gastric emptying. Scientific Reports. P. 529-540, 2015.
CAO, D. S. Expression of transient receptor potential ankyrin 1 (TRPA1) and its role in insulin release from rat pancreatic beta cells. Plos. One. 2012.
CARNEIRO, M.C., SIQUEIRA-BATISTA, R. O mosaico patogênico da pancreatite aguda grave. Rev. Col. Bras. P. 391-397, 2004.
CARSWELL, E. A., OLD, L. J., KASSEL, R. L., GREEN, S., FIORE, N., WILLIAMSON, B. (1975) An endotoxin-induced serum factor that causes necrosis of tumors. Proc. Natl. Acad. Sci. USA 72, 3666–3670.
CAVAILLON, J. M., ADIB-CONQUY, M., FITTING, C., ADRIE, C., PAYEN, D. (2003) Cytokine cascade in sepsis. Scand. J. Infect. Dis. 35, 535–544.
CHANDRA A, ENKHBAATAR P, NAKANO Y, TRABER LD, TRABER DL. Sepsis: emerging role of nitric oxide and selectins. Clinics (Sao Paulo). P. 71-76, 2006.
CHAO LK, HUA KF, HSU HY, CHENG SS, LIN IF, CHEN CJ, CHEN ST, CHANG ST. Cinnamaldehyde inhibits pro-inflammatory cytokines secretion from monocytes/macrophages through suppression of intracellular signaling. Food and Chemical Toxicology. P. 220-231, 2008.
CHAUDHRY, H., ZHOU, J., ZHONG, Y., ALI, M.M., MCGUIRE, F., NAGARKATTI, P.S., NAGARKATTI, M. Role of cytokines as a double-edged sword in sepsis. In vivo. P. 669-684, 2013.
CHEN, J., JOSHI, S.K., DIDOMENICO, S., PERNER, R.J., MIKUSA, J.P., GAUVIN, D.M., SEGRETI, J.A., ZHANG, X.F., NIFORATOS, W., BIANCHI, B.R., BAKER, S.J., ZHONG, C., SIMLER, G.H., MCDONALD, H.A., SCHMIDT, R.G., MCGARAUGHTY, S.P., CHU, K.L., FALTYNEK, C.R., KORT, M.E., REILLY, R.M., KYM, P.R. Selective blockade of TRPA1 channel attenuates pathological pain without altering noxious cold sensation or body temperature regulation. Pain. P. 1165-1172, 2011.
CHEN, P., STANOJCIC, M., JESCHKE, M.G. Differences between murine and human sepsis. The surgical clinics of Noth America. P. 1135-1149, 2014.

88
CHOI, H., LETO, T.L., HUNYADY, L., CATT, K.J., BAE, Y.S., AND RHEE, S.G. Mechanism of angiotensin II-induced superoxide production in cells reconstituted with angiotensin type 1 receptor and the components of NADPH oxidase. Journal of Biological Chemical. V. 283, p. 255–267, 2008.
CHRISTINA WACKER, ANNA PRKNO, FRANK M BRUNKHORT, PETER SCHLATTMANN. Procalcitonin as a diagnostic marker for sepsis: a systematic review and meta-analysis. Lancet infectious Disease. 2013; 13: 426-35.
CLAPHAM, D.E. TRP channels as cellular sensors. Nature. P. 517-524, 2003.
COHEN, J. The immunopathogenesis of sepsis. Nature. V. 420, p. 885-91, 2002.
COLIN, G., NARDI, O., POLITO, A., ABOAB, J., MAXIME, V., CLAIR, B., FRIEDMAN, D., ORLIKOWSKI, D., SHARSHAR, T., ANNANE, D. Masseter tissue oxygen saturation predicts normal central venous oxygen saturation during early goal-directed therapy and predicts mortality in patients with severe sepsis. Critical Care Medicine. P. 435-440, 2012.
CORDIOLI, R.L., CORDIOLI, E., NEGRINI, R., SILVA, E. Sepsis and pregnancy: do we know how to treat this situation? Revista Brasileira de Terapia Intensiva. P. 334-344, 2013.
COWAN, J.E, JENKINSON, W.E, ANDERSON, G. Thymus medulla fosters generation of natural Treg cells, invariant γδ T cells, and invariant NKT cells: what we learn from intrathymic migration. European journal of immunology. P. 652-660, 2015.
CROUSER, E., EXLINE, M., KNOELL, D., WEWERS, M.D. Sepsis: links between pathogen sensing and organ damage. Current Pharmaceutical Design. P. 1840-1852, 2008.
CRUZ-ORENGO, L., DHAKA, A., HEUERMANN, R.J., YOUNG, T.J., MONTANA, M.C. Cutaneous nociception evoked by 15-delta PGJ2 via activation of ion channel TRPA1. Molecular Pain. P. 4:30, 2008.
D. F. FIORENTINO, A. ZLOTNIK, T. R. MOSMANN, M. HOWARD, AND A. O’GARRA. IL-10 inhibits cytokine production by activated macrophages. The Journal of Immunology, vol. 147, p. 3815–3822, 1991.
DANIEL RITTIRSCH, L. MARCO HOESEL, AND PETER A. WARD. The disconnect between animal models of sepsis and human sepsis. Journal of Leukocyte Biology Volume 81, January 2007.
DANIEL RITTIRSCH, L. MARCO HOESEL, AND PETER A. WARD. The disconnect between animal models of sepsis and human sepsis. Journal of Leukocyte Biology. V. 81, 2007.

89
DEBORAH, J., STEARNS-KUROSAWA, MARCIN, F., OSUCHOWSKI, CATHERINE VALENTINE, SHINICHIRO KUROSAWA, AND DANIEL G. REMICK. The Pathogenesis of Sepsis. Revista anual de patologia. P. 19–48, 2001.
DEJAGER, L., PINHEIRO, I., DEJONCKHEERE, E., LIBERT, C. Cecal ligation and puncture: the gold standard model for polymicrobial sepsis?. Trends in Microbiology. Vol. 19, 2011.
DESNUES, B., AND MEGE, J. Macrophage Polarization in Bacterial Infections. Journal Immunol. P. 3733-3739, 2008. DING SF. Sialic acids and the cellular function of the hematopoietic system. P. 66-68, 1987. DOBELL, A.R., SANCHEZ, F.R., MURPHY, D.R. Experimental evaluation of ivalon as replacement for right ventricular myocardium. Archives of Surgery. P. 118-122, 1960. DOĞANYIĞIT, Z., KÜP, FÖ., SILICI, S., DENIZ, K., YAKAN, B., ATAYOGLU, T. Protective effects of propolis on female rats histopathological, biochemical and genotoxic changes during LPS induced endotoxemia. Phytomedicine. P. 632-639, 2013.
DONATH, B., FISCHER, C., PAGE, S., PREBECK, S., JILG, N., WEBER, M., DA COSTA, C., NEUMEIER, D., MIETHKE, T., BRAND, K. Chlamydia pneumoniae activates IKK/I kappa B-mediated signaling, which is inhibited by 4-HNE and following primary exposure. Atherosclerosis. P. 79-88, 2002.
DOREEN, E., WESCHE-SOLDATO, RYAN, Z., SWAN, C.C., AND ALFRED, A. The Apoptotic Pathway as a Therapeutic Target in Sepsis. Current Drug Targets. P. 493–500, 2007.
DOUGLAS, D., BANNERMAN, AND SIMEON, E. GOLDBLUM. Mechanisms of bacterial lipopolysaccharide-induced endothelial apoptosis. American Journal of Physiological Lung Cell and Molecular Physiological. P. 899–914, 2003.
EL-SAYED EL-SM, ABD EL-RAOUF OM, FAWZY HM, MANIE MF. Comparative study of the possible protective effects of cinnamic acid and cinnamaldehyde on cisplatin-induced nephrotoxicity in rats. Journal of Biochemical Molecular Toxicology. P. 508-514, 2013.
ENGEL, M.A., LEFFLER, A., NIEDERMIRTL, F., BABES, A., ZIMMERMANN, K., FILIPOVIĆ, M.R., IZYDORCZYK, I., EBERHARDT, M., KICHKO, T.I., MUELLER-TRIBBENSEE, S.M., KHALIL, M., SIKLOSI, N., NAU, C., IVANOVIĆ-BURMAZOVIĆ, I., NEUHUBER, W.L., BECKER, C., NEURATH, M.F., REEH, P.W. TRPA1 and substance P mediate colitis in mice. Gastroenterology. P. 1346-1358, 2011.
ENTLEUTNER, M., TRAEGER, T., WESTERHOLT, A., HOLZMANN, B., STIER, A., PFEFFER, K., MAIER, S., HEIDECKE, C. D. Impact of interleukin-12, oxidative burst, and iNOS on the survival of murine fecal peritonitis. International Journal of Colorectal Disease. V. 21, p. 64–70, 2006.

90
FERNANDO, O., MARTINEZ, SIAMON, G. The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Reports. P. 6-13, 2014.
FINK MP, HEARD SO. Laboratory models of sepsis and septic shock. J Surg Res 1990;49:186-96.
FUHONG, S., NAM, D.N., ZHEN, W., YING, C., PETER, R., JEANLOUIS, V. Fever control in septic shock: beneficial or harmful? Shock. P. 516-520, 2005.
FUNK, D.J., PARRILLO, J.E., KUMAR, A. Sepsis and septic shock: a history. Critical of Care Clinical. P. 83-101, 2009.
GENDE, L.B.I., FLORIS, R., FRITZ, EGUARAS, M. J.. “Antimicrobial activity of cinnamon (Cinnamomum zeylanicum) essential oil and its main components against paenibacillus larvae from argentine”. Bulletin of Insectology, vol. 61, no. 1, pp. 1–4, 2008.
GEROULANOS, S., EVANGELIA, T., DOUKA, H. Historical perspective of the word “sepsis”. Intensive Care Medicine. V. 32, p 2077, 2006.
GRANGER JI, RATTI PL, DATTA SC, RAYMOND RM, OPP MR. Sepsis-induced morbidity in mice: effects on body temperature, body weight, cage activity, social behavior and cytokines in brain. Psychoneuroendocrinology. 2013, 1047-1057.
H. F. GALLEY. Oxidative stress and mitochondrial dysfunction in sepsis. British Journal of Anaesthesia. 107 (1): 57–64 (2011).
HALLIWELL B, GUTTERIDGE JMC. Free Radicals in Biology and Medicine. Ed 4. Clarendon Press, Oxford; (2006).
HARDAWAY RM, WILLIAMS CH, SUN Y. A new approach to the treatment of experimental septic shock. J Surg Res 1996;61:311-6.
HENKIN, C.S., COELHO, J.C., CHISS, M., MORAIS, P.R., DIAS, S.R. Sepse: uma visão atual. Scientia Medica. V. 19, p. 135-145, 2009. HILL AL, LOWES DA, WEBSTER NR, SHETH CC, GOW NA, GALLEY HF. Regulation of pentraxin-3 by antioxidants. Br J Anaesth. 2009; 103: 833–39.
HOWARD, M.T., MUCHAMUEL, S. ANDRADE, AND MENON, M. “Interleukin 10 protects mice from lethal endotoxemia”. Journal of Experimental Medicine. Vol. P. 1205–1208, 1993.
HOWARD, T.M., ANDRADE, S., MENON, S. Interleukin 10 protects mice from lethal endotoxemia. Journal of Experimental Medicine. Vol. 177, no. 4, p. 1205–1208, 1993.
HUANG J, ZHANG X, MCNAUGHTON PA. Modulation of temperature-sensitive TRP channels. Seminal Cell Dev Biol. 2006 Dec;17(6):638-45. Epub 2006 Nov 11.

91
J. SCHELLER AND S. ROSE-JOHN. Interleukin-6 and its receptor: frombench to bedside. MedicalMicrobiology and Immunology, vol. 195, no. 4, pp. 173–183, 2006.
JAMES R. PARRATT. Nitric oxide in sepsis and endotoxaemia. Journal of Antimicrobial Chemotherapy. (1998) 41, Suppl. A, 31–39.
JAVIER OVIEDO-BOYSO, ALEJANDRO BRAVO-PATIÑO, AND VÍCTOR M. BAIZABAL-AGUIRRE. Collaborative Action of Toll-Like and Nod-Like Receptors as Modulators of the Inflammatory Response to Pathogenic Bacteria. Mediators of Inflammation. Volume 2014, Article ID 432785, 16 pages
JORDT, S.E., BAUTISTA, D.M., CHUANG, H.H., MCKEMY, D.D., ZYGMUNT, P.M., HOGESTATT, E.D., MENG, I.D., JULIUS, D. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature. P. 260–265, 2004.
JÚNIOR, G.A.P., MARSON, F., ABEID, M., FÁTIMA, M., OSTINI, S.H.S. FISIOPATOLOGIA DA SEPSE E SUAS IMPLICAÇÕES TERAPÊUTICAS. Medicina, Ribeirão Preto. P. 349-362, 1998. JÚNIOR, J.A.L.S., DAVID, C.M., HATUM, R., SOUZA, P.C.S.P., JAPIASSÚ, A., PINHEIRO, C.T.S., FRIEDMAN, G., SILVA, O.B., DIAS, M.D., KOTERBA, E., DIAS, F.S., PIRAS, C. Sepse Brasil: Estudo Epidemiológico da Sepse em Unidades de Terapia Intensiva Brasileiras. Revista Brasileira Terapia Intensiva. Volume 18 - Número 1 - Janeiro/Março 2006. KOH, W.S., YOON, S.Y., KWON, B.M., JEONG, T.C., NAM, K.S., HAN, M.Y. Cinnamaldehyde inhibits lymphocyte proliferation and modulates T-cell differentiation. Int J Immunopharmacol. P.643-60, 1998.
KOIVISTO, A., CHAPMAN, H., JALAVA, N., KORJAMO, T., SAARNILEHTO, M., LINDSTEDT, K., PERTOVAARA, A. TRPA1: a transducer and amplifier of pain and inflammation. Basic Clinical Pharmacology Toxicology. 2014.
KRUGER, P., SAFFARZADEH, M., WEBER, A.N., RIEBER, N., RADSAK, M., VON BERNUTH, H., BENARAFA, C., ROOS, D., SKOKOWA, J., HARTL, D. Neutrophils: Between host defence, immune modulation, and tissue injury. Plos Pathogens. V. 12, 2015.
KWON, H.K., HWANG, J.S., SO J.S., LEE, C.G., SAHOO, A., RYU, J.H., JEON, W.K., KO, B.S., IM, C.R., LEE, S.H., PARK, Z.Y., IM, S.H. Cinnamon extract induces tumor cell death through inhibition of NFkappaB and AP1. BMC Cancer. 2010.
LAHTI, A., MAIBACH, H.I. An animal model for nonimmunologic contact urticaria. Toxicology and Applied Pharmacology. P. 219-224, 1984.
LANG, J.D., MATUTE-BELLO, G. Lymphocytes, apoptosis and sepsis: making the jump from mice to humans. Critical Care. 2009.
LATIFI, M.A., O’RIORDAN, LEVINE D.. Interleukin- 10 controls the onset of irreversible septic shock. Infection and Immunity, vol. 70, p. 4441–4446, 2002.

92
LIAO, J.C, DENG, J.S., CHIU, C.S., HOU, W.C., HUANG, S.S., SHIE, P.H., HUANG, G.J. Anti-Inflammatory Activities of Cinnamomum cassia Constituents In Vitro and In Vivo. Evidence-Based Complementary and Alternative Medicine. 2012. LIN, S.S., LU, T.M., CHAO, P.C., LAI, Y.Y., TSAI, H.T., CHEN, C.S., LEE, Y.P., CHEN, S.C., CHOU, M.C., YANG, C.C. In vivo cytokine modulatory effects of cinnamaldehyde, the major constituent of leaf essential oil from Cinnamomum osmophloeum Kaneh. Phytother Res. P. 1511-1518, 2011.
LUAN, Y., DONG, N., XIE, M., XIAO, X., YAO, Y. The Significance and Regulatory Mechanisms of Innate Immune Cells in the Development of Sepsis. Journal of interferon & cytokine research. V. 34, 2014.
MARTIN, G.S., MANNINO, D.M., EATON, S., MOSS, M. The epidemiology of sepsis in the United States from 1979 through 2000. The New England Jounal of Medicine. P. 1546-1554, 2003.
MARTYN, K.D., FREDERICK, L.M., VON LOEHNEYSEN, K., DINAUER, M.C., KNAUS, U.G. Functional analysis of Nox4 reveals unique characteristics compared to other NADPH oxidases. Cell Signal. P. 69–82, 2006.
MASAMOTO, Y., KAWABATA, F., FUSHIKI, T. Intragastric administration of TRPV1, TRPV3, TRPM8, and TRPA1 agonists modulates autonomic thermoregulation in different manners in mice. Biosci Biotechnol Biochem. P. 1021-1027, 2009.
MCKENZIE, C. Antibiotic dosing in critical illness. Journal Antimicrob Chemother, P. 25-32, 2011.
MESEGUER, V., YERANDDY, A., ALPIZAR, E.L., TAJADA, S., DENLINGER, B., FAJARDO, R., MANENSCHIJN, J.,N FERNANDEZ-PENA, C., TALAVERA, A., KICHKO, T., NAVIA, B., SANCHEZ, A., SENARIS, R., PREEH, P., PEREZ-GARCIA, M.T., LOPEZ-LOPEZ, J.G., VOETS, T., BELMONTE, C., TALAVERA, K., VIANA, F. TRPA1 channels mediate acute neurogenic inflammation and pain produced by bacterial endotoxins. Nature. 2014.
MIURA, S., TAKAHASHI, S., IMAGAWA, T., UCHIDA, K., SAITO, S., TOMINAGA, M., OHTA, M.. Involvement of TRPA1 activation in acute pain induced by cadmium in mice. Miura et AL. MOLECULAR PAIN. P. 9-7, 2013.
MONCADA, S., HIGGS, A. The L-arginine-nitric oxide pathway. The New England Journal of Medicine. P.2002–2012, 1993.
NGKELO, A., MEJA, K., YEADON, M., ADCOCK, I., KIRKHAM, P.A. LPS induced inflammatory responses in human peripheral blood mononuclear cells is mediated through NOX4 and Giα dependent PI-3kinase signalling. The Journal of Inflammation. 2012.
O’BRIEN J.M., NAEEM, A.A., ABEREGG, S.K. Sepsis. American Journal of Medicine. P. 1012-1022, 2007.

93
OPAL SM, COHEN J. Clinical gram-positive sepsis: does it fundamentally differ from gram-negative bacterial sepsis? Crit Care Med 1999;27:1608-16.
PARK, B., S. & LEE, J. Recognition of lipopolysaccharide pattern by TLR4 complexes. Experimental & Molecular Medicine. P. 45-66, 2003.
PARKER SJ, WATKINS PE. Experimental models of gram-negative sepsis. Br J Surg 2001;88:22-30.
PATERSON, R.L, GALLEY, H.F, DHILLON, J.K., WEBSTER, N.R. Increased NFkB activation in critically ill patients who die. Critical Care Medicine. P. 1047–51, 2000.
PATRA, K., BOSE, S., SARKAR, S., RAKSHIT, J., JANA, S., MUKHERJEE, A., ROY, A., MANDAL, D.P., BHATTACHARJEE, S. Amelioration of cyclophosphamide induced myelosuppression and oxidative stress by cinnamic acid. Chemmical Biology Interaction. P. 231-239, 2012.
PAULO, R.A., CARVALHO, E.A. Avanços no diagnóstico e tratamento da sepse. Jornal de Pediatria. 2003.
PEDERSEN, S.F., OWSIANIK, G., NILIUS, B. TRP channels: an overview. Cell Calcium. P. 233-252, 2005.
PETER, A. WARD: New approaches to the study of sepsis. EMBO medicina molecular. P. 1234–1243, 2012.
PINTO, C.A, SILVA, D.H.S., BOLZANI, U.S., LOPES, N.P., EPIFÂNIO, R.A. Produtos naturais: atualidade, desafios e perspectivas. Quimica Nova. P. 45-61, 2002.
PIPER, R.D., COOK, D.J., BONE, R.C., SIBBALD, W.J. Introducing critical appraisal to studies of animal models investigating novel therapies in sepsis. Critical Care Medicine. P. 2059-2070, 1996.
PLATZER, B., STOUT, M., FIEBIGER, E. Antigen cross-presentation of immune complexes. Frontiers in immunology. 2014.
PRABUSEENIVASAN, S.M., JAYAKUMAR, IGNACIMUTHU, S.. In vitro antibacterial activity of some plant essential oils. BMC Complementary and Alternative Medicine, vol. 6, 2006.
PREDONZANI, A., CALÌ, B., AGNELLINI, A.H., MOLON, B. Spotlights on immunological effects of reactive nitrogen species: When inflammation says nitric oxide. World jornal of experimental medicine. P. 64-76, 2015.
QUENZER, R. Infections in the critically ill. In: BONGARD FS & SUE DY. Current critical care diagnosis & treatment. Appleton & Lange, Connecticut. P. 131-155, 1994.

94
RAMACHANDRAN, R. Gram-positive and gram-negative bacterial toxins in sepsis: A brief review. Virulence. P., 213–218, 2014.
RANGEL-FRAUSTO MS1. Sepsis: still going strong. Archives Medical Research. P. 672-681, 2005.
REMICK, D. G., STRIETER, R. M., ESKANDARI, M. K., NGUYEN, D. T., GENORD, M. A., RAIFORD, C. L., KUNKEL, S. L. (1990) Role of tumor necrosis factor-α in lipopolysaccharide-induced pathologic alterations. Am. J. Pathol. 136, 49–60.
REMICK, D.G., NEWCOMB, D.E., GERALD, L.B., CALL, D.R., Comparison of the mortality and inflammatory response of two models of sepsis: lipopolysaccharide vs cecal ligation and puncture. Shock. P. 110-116, 2000.
RIEDEMANN, N.C, GUO, R.F, WARD, P.A. The enigma of sepsis. Journal Clinical Investigation. P. 460-467, 2003.
RONAN, M.G., BERG, K.M., BAILEY, D.M. Neuro-oxidative-nitrosative stress in sepsis. Journal of Cerebral Blood Flow & Metabolism. P. 1532–1544, 2011.
ROTH-WALTER, F., MOSKOVSKICH, A., GOMEZ-CASADO, C., DIAZ-PERALES, A., OIDA, K., SINGER, J., KINACIYAN, T., FUCHS, H., JENSEN-JAROLIM , E. Immune Suppressive Effect of Cinnamaldehyde Due to Inhibition of Proliferation and Induction of Apoptosis in Immune Cells: Implications in Cancer. Plos One. P. 1-14, 2014.
RUDIGERA, A., STOTZB, M., SINGER, M. Cellular processes in sepsis. Swiss medical wkly, p. 629–634, 2008.
SAMER R EID, ERIC D CROWN, ERIC L MOORE, HONGYU A LIANG, KAR- CHAN CHOONG, SHELLEY DIMA, DARRELL A HENZE, STEFANIE A KANE AND MARK O URBAN. HC-030031, a TRPA1 selective antagonist, attenuates inflammatory- and neuropathy-induced mechanical hypersensitivity. Molecular Pain 2008, 4:48 doi:10.1186/1744-8069-4-48.
SCAGLIONE, F. Pharmacokinetic/pharmacodynamic (PK/PD) considerations in the management of Gram-positive bacteraemia. International Journal of Antimicrobial Agents. P. 33-39, 2010.
SCAMMELL, T.E., ELMQUIST, J.K., SAPER, C.B. Inhibition of nitric oxide synthase produces hypothermia and depresses lipopolysaccharide fever. The American Journal of Physiology. P. 333-338, 1996.
SCHINDLER, R.J., MANCILLA, S., ENDRES, R., GHORBANI, S., C. CLARK, DINARELLO, C.D. Correlations and interactions in the production of interleukin-6 (IL-6), IL-1, and tumor necrosis factor (TNF) in human blood mononuclear cells: IL-6 suppresses IL-1 and TNF. Blood, vol. 75, p. 40–47, 1990.

95
SCHULTE, W., BERNHAGEN, J., BUCALA. R. Cytokines in Sepsis: Potent Immunoregulators and Potential Therapeutic Targets—An Updated View. Mediators of Inflammation. 16 pages, 2013.
SFORCIN, J.M., BANKOVA, V. Propolis: is there a potential for the development of new drugs? Journal Ethnopharmacol. P. 253–260, 2011.
SHAD, A. A., AHMAD, S., ULLAH, R., ABDEL-SALAM, H., HUSSAIN, H., SAEED, W. Phytochemical and Biological Activities of Four Wild Medicinal Plants. Scientific worl journal. V. 2014, p. 1-7, 2014.
SHEN, Y., JIA, L., HONMA, N., HOSONO, T., ARIGA, T., SEKI, Y. Beneficial Effects of Cinnamon on the Metabolic Syndrome, Inflammation, and Pain, and Mechanisms Underlying These Effects – A Review. Journal of Traditional and Complementary Medicine. V. 1. p. 27-32, 2012.
SHIH-CHIEH LEE, JIE-SHENG HSU, CHIEN-CHUN LI, KE-MING CHEN, CHENG-TZU LIU. Protective Effect of Leaf Essential Oil from Cinnamomum osmophloeum Kanehira on Endotoxin-Induced Intestinal Injury in Mice Associated with Suppressed Local Expression of Molecules in the Signaling Pathways of TLR4 and NLRP3. DOI:10.1371/journal.pone.0120700 March 20, 2015.
SOGAYAR, A.M., MACHADO, F.R., REA-NETO, A., DORNAS, A., GRION, C.M., LOBO, S.M., TURA, B.R., SILVA, C.L., CAL, R.G., BEER, I., MICHELS, V., SAFI, J., KAYATH, M., SILVA, E. A multicentre, prospective study to evaluate costs of septic patients in Brazilian intensive care units. Pharmacoeconomics. P. 425-434, 2008.
SONG, M., KELLUM, J. A. Interleukin-6. Critical care medicine. V. 33, p. 463– 465, 2005.
SORDI, R, MENEZES-DE-LIMA, O.J.R., HOREWICZ, V., SCHESCHOWITSCH, K., SANTOS, L.F., ASSREUY, J. Dual role of lipoxin A4 in pneumosepsis pathogenesis. Int Immunopharmacology. P. 283-292, 2013.
STARR, M., SAITO, H. Sepsis in Old Age: Review of Human and Animal Studies. Aging and Disease. V. 5, p. 126-136, 2014.
STECKERT, A.V., CASTRO, A.A., DAL-PIZZO, F., QUEVEDO, J. Sepsis in the Central Nervous System and Antioxidant Strategies with Nacetylcysteine, Vitamins and Statins. Current Neurovascular Research, 2014, Vol. 11, No. 1.
TACKE, F., ZIMMERMANN, H.W., Macrophage heterogeneity in liver injury and fibrosis. Journal of Hepatology. V. 60, 2014.
TAKIKAWA, O., OKU, T., USHIO, Y., YAMAMOTO, N., YONEDA, Y., TSUJI, J., SANCHEZ-BUENO, A., VERKHUSHA, V., YOSHIDA, R. Multiple expression of Ly-6C and accumulation of a Ly-6C pre-mRNA in activated macrophages involved in rejection of an allografted tumor. Biochemical Biophysical Research Communication. P. 247-253, 1996.

96
THE WORLD HEALTH REPORT 1996: Fighting Disease, Fostering Development. Geneva, World Health Organization, 1996.
THEILEN, H., RAGALLER, M. Therapy of hyperthermia and septic shock. Necessary or injurious? Anaesthesiology. P. 949-952, 2007.
THOMAS, L., LETO, S.M., HURT, D., UEYAMA, T. Targeting and Regulation of Reactive Oxygen Species Generation by Nox Family NADPH Oxidases. Antioxidants & redox signaling. V. 11, 2009.
TOMAZZONI, M.I., NEGRELLE, R.R.B., CENTA, M.L. Fitoterapia popular: a busca instrumental enquanto prática terapêutica. Texto Contexto Enferm. P. 115-121, 2006.
TREVISANI, M., SIEMENS, J., MATERAZZI, S., BAUTISTA, D.M., NASSINI, R. 4-Hydroxynonenal, na endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1. Proc. Natl. Acad. Sci. USA. P. 19–24, 2007.
TUNG, Y.T., HUANG, C.C., HO, S.T., KUO, Y.H., LIN, C.C., LIN, C.T., WU, JH. Bioactive phytochemicals of leaf essential oils of Cinnamomum osmophloeum prevent lipopolysaccharide/D-galactosamine (LPS/D-GalN)-induced acute hepatitis in mice. Journal of Agricultural Food Chemistry. P. 8117-8123, 2011.
VAN DER POLL, T. Immunotherapy of sepsis. Lancet Infect Dis. 165-174, 2001.
VICTOR, V.M.N., GUAYERBAS, D., GARROTE, M., DEL RIO, M., DE LA FUENTE. Modulation of murine macrophage function by N-acetylcysteine in a model of endotoxic shock. Biofactors. 2004 10:347.
WALLEY, K.R., LUKACS, N.W., STANDIFORD, T.J., STRIETER, R.M., KUNKEL, S.L. Balanced of inflammatory cytokines related to severity and mortality of murine sepsis. Infection and Immunity 64: 4733-4738, 1996.
WANG, L., HOU, Y., YI, D., DING, B., ZHAO, D., WANG, Z., ZHU, H., LIU, Y., GONG, J., ASSAAD, H.W.U.G. Beneficial roles of dietary oleum cinnamomi in alleviating intestinal injury. Frontiers in Bioscience. P. 814-828, 2015.
WARD, P.A. New approaches to the study of sepsis. EMBO Molecular Medicine. P. 1234–1243, 2012.
WEBER, S.U., SCHEWE, J.C., LEHMANN, L.E., MÜLLER, S., BOOK, M., KLASCHIK, S., HOEFT, A., STÜBER, F. Induction of Bim and Bid gene expression during accelerated apoptosis in severe sepsis. Critical Care. 2008.
WEINA, G.A.O., JIANQUAN, W.U., JINGYU, W.E.I, LINGLING, P.U., CHANGJIANG, G.U.O., YANG, J., YANG, M., LUO, H. Brazilian green propolis improves immune function in aged mice. Journal of Clinical Biochemical Nutrition. Vol. 55, 2014.

97
WESTPHAL, G.A., FEIJÓ, J., ANDRADE, P.S., TRINDADE, L., SUCHARD, C., MONTEIRO, M.A.G. Early detection strategy and mortality reduction in severe sepsis. Revista Brasileira de Terapia Intensiva. P. 113-123, 2009. WIERSINGA, W.J., LEOPOLD, S.J., CRANENDONK, D.R., POLL, T.V.D. Host innate immune responses to sepsis. Virulence. P. 36–44, 2014.
WORLD HEALTH ORGANIZATION. World Health Statistics. Who library cataloguing-in-publication data, 2010.
XAUS, J., COMALADA, M., VALLEDOR, A.F., LLOBERAS, J., LÓPEZ-SORIANO, F., ARGILÉS, J.M., BOGDAN, C., CELADA, A. LPS induces apoptosis in macrophages mostly through the autocrine production of TNF-alpha. Blood. P. 3823-3831, 2000.
YANG Y, SHARMA R, SHARMA A, AWASTHI S, AWASTHI YC. Lipid peroxidation and cell cycle signaling: 4-hydroxynonenal, a key molecule in stress mediated signaling. Acta Biochim Pol. 2003;50(2):319-36.
YOUN, H.S., LEE, J.K., CHOI, Y.J., SAITOH, S.I., MIYAKE, K., HWANG, D.H., LEE, J.Y. Cinnamaldehyde suppresses toll-like receptor 4 activation mediated through the inhibition of receptor oligomerization. Biochemical Pharmacology. P. 494-502, 2007.
YVETTE, C., LUIKING, P.K.J., ENGELEN, P., NICOLAAS, E.P. Regulation of nitric oxide production in health and disease. Current Opinion Clinical Nutrition Metabolic Care. P. 97–104, 2010.
Z.CHAI, S. GATTI, C. TONIATTI,V. POLI, AND T. BARTFAI. Interleukin (IL)-6 gene expression in the central nervous system is necessary for fever response to
lipopolysaccharide or IL-1𝛽: a study on IL- 6-deficient mice. Journal of Experimental. Vol. 183, p. 311–316, 1996.
ZANTL N, UEBE A, NEUMANN B, ET AL. Essential role of gamma interferon in survival of colon ascendens stent peritonitis, a novel murine model of abdominal sepsis. Infection and Immunity. P. 2300-2309, 1998.
ZEHAVA L. NOAH, MD. The History of Sepsis Management Over the Last 30 Years. VOL. 15, NO. 2, 2014.

98


















