
UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA … · Lariza por terem se disponibilizado a...
145
UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO” FACULDADE DE CIÊNCIAS AGRONÔMICAS CAMPUS DE BOTUCATU FISIOLOGIA E METABOLISMO DA VIDEIRA CV. SYRAH NO SUBMÉDIO DO VALE DO SÃO FRANCISCO SOB TRÊS ESTRATÉGIAS DE IRRIGAÇÃO CAIO MÁRCIO GUIMARÃES SANTOS Tese apresentada à Faculdade de Ciências Agronômicas da UNESP - Campus de Botucatu, para obtenção do título de Doutor em Agronomia (Horticultura) BOTUCATU – SP Janeiro – 2012
Transcript of UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA … · Lariza por terem se disponibilizado a...

UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CAMPUS DE BOTUCATU
FISIOLOGIA E METABOLISMO DA VIDEIRA CV. SYRAH NO
SUBMÉDIO DO VALE DO SÃO FRANCISCO SOB TRÊS
ESTRATÉGIAS DE IRRIGAÇÃO
CAIO MÁRCIO GUIMARÃES SANTOS
Tese apresentada à Faculdade de Ciências
Agronômicas da UNESP - Campus de Botucatu,
para obtenção do título de Doutor em Agronomia
(Horticultura)
BOTUCATU – SP
Janeiro – 2012

UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CAMPUS DE BOTUCATU
FISIOLOGIA E METABOLISMO DA VIDEIRA CV. SYRAH NO
SUBMÉDIO DO VALE DO SÃO FRANCISCO SOB TRÊS
ESTRATÉGIAS DE IRRIGAÇÃO
CAIO MARCIO GUIMARÃES SANTOS
Orientador: Prof. Dr. João Domingos Rodrigues
Co-orientadora: Dra Bárbara França Dantas
Tese apresentada à Faculdade de Ciências
Agronômicas da Unesp - Campus de Botucatu,
para obtenção do título de Doutor em Agronomia
(Horticultura)
BOTUCATU – SP
Janeiro – 2012



III
DEDICATÓRIA
Ao amor divino de Jesus Cristo;
Ao amor incondicional de Laura Rêgo Guimarães Santos e José de Souza Santos;
Ao amor sem limites de Aida de Magalhães Rêgo (in memorian);
Ao amor pouco explorado de Rosalvo Amaral Guimarães (in memoriam) e Bernadina (in
memoriam);
Ao amor fraterno de Joara Márcia Guimarães Santos e Laiara Márcia Guimarães Santos;
Ao amor familiar próximo de tia Nenza e Vilma; Suze e Sandoval; Thaís e Mariel; dos
queridos sobrinhos: Lineker, Iasmin, Larissa, Maria Eduarda e Artur.
Ao amor familiar separado apenas pela distância física de todos os tios e primos que compõe a
grande família dos Guimarães e, em especial, às tias Sandra, Dilma (in memoriam) Vilma,
Ivanir e ao tio Bemvindo;
Aos amores que me deram força no passado, aos amores presentes e aos amores futuros a
serem construídos.
Sempre dê adeus ao homem, mas nunca ao seu legado!!!

IV
AGRADECIMENTOS
Ao Pai, ao Filho e ao Espírito Santo de Deus!
À Faculdade de Ciências Agronômicas - UNESP – Departamento de
Produção Vegetal/Horticultura por ter disponibilizado os recursos humanos e materiais.
À Embrapa Semiárido - CPATSA por ter permitido a pesquisa e dado
todo suporte estrutural e técnico necessário.
À CAPES pela concessão da bolsa de estudos durante o curso.
Ao Prof. Dr. João Domingos Rodrigues por ter confiado em mim e
creditado a sua orientação.
À amiga e mestranda da UFRB, Márcia Moura, por sua fundamental
ajuda na realização das atividades práticas do experimento, sendo a maior colaboradora na
conclusão do trabalho.
Ao Professor Doutor Carlos Ducatti pelos ensinamentos e aos
funcionários do Laboratório de Isótopos Estáveis Ambientais da UNESP-Botucatu pela
atenção concedida.
Ao pesquisador do CPATSA Dr. Luís Henrique Bassoi pelo convite
para participar no seu macroprojeto com o meu trabalho de tese e pelos recursos
disponibilizados durante a execução do mesmo.
À pesquisadora do CPATSA e co-orientadora Dra Bárbara França
Dantas por ter auxiliado na execução das análises laboratoriais e por suas correções na tese.
Aos pesquisadores doutores do CPATSA Giuliano Elias Pereira e
Maria Auxiliadora por terem disponibilizado os laboratórios sob sua responsabilidade para as
análises químicas.

V
Aos funcionários do CPATSA: Zizinho, Alberto, Clétis, Dora, Ana,
Helena, Hebert, Fábio, Raimundo, Hélio, entre outros do campo experimental de Bebedouro
por suas contribuições essenciais.
Aos bolsistas e estagiárias do laboratório de Controle Biológico e
Enologia pela solidariedade e disposição em ajudar.
Às minhas queridas ex- alunas do IF Sertão-PE, Nayra, Laiane e
Lariza por terem se disponibilizado a ajudar no que fosse preciso e sempre com alto astral.
Ao grande amigo e colega Reginaldo pela amizade, companheirismo e
todo apoio fornecido desde o início do curso.
A todos os demais colegas da pós-graduação, pela amizade construída,
especialmente: Marcão, Luchele, Luiza, Nelsão, Camila, Rigléia, Manoel, Edvar, Sidnei,
Luciana, Amaralina, Bárbara, Angela, Karol, Ane e Essione.
À tia Eliane e seu Darci pelo apoio residencial, emocional e moral na
reta final da defesa.
A todos os colegas professores do IF SERTÃO-PE que com palavras e
pensamentos positivos ajudaram a impulsionar e concluir essa nova etapa em minha vida
pessoal e profissional.

VI
SUMÁRIO
LISTA DE ABREVIATURAS E SÍMBOLOS...................................................................... VIII
LISTA DE FIGURAS.............................................................................................................. X
LISTA DE TABELAS............................................................................................................. XI
RESUMO................................................................................................................................... 14
SUMMARY................................................................................................................................ 16
1 INTRODUÇÃO...................................................................................................................... 18
2 REVISÃO DE LITERATURA............................................................................................ 21
2.1 VITIVINICULTURA............................................................................................................ 21
2.2 CARACTERÍSTICAS DA CULTIVAR SYRAH................................................................ 23
2.3 CARACTERÍSTICAS DO CULTIVO DA VIDEIRA......................................................... 24
2.4 ECOFISIOLOGIA DA VIDEIRA........................................................................................ 24
2.4.1 Fenologia........................................................................................................................... 30
2.4.2 Crescimento de Ramos....................................................................................................... 31
2.4.3 Relação Fonte/Dreno......................................................................................................... 32
2.5 METABOLISMO DA VIDEIRA......................................................................................... 34
2.5.1 Carboidratos...................................................................................................................... 34
2.5.2 Ácidos orgânicos................................................................................................................ 36
2.5.3 Compostos nitrogenados.................................................................................................... 37
2.6 CARACTERÍSTICAS DO FRUTO PARA PRODUÇÃO DE VINHO............................... 38
3 MATERIAL E MÉTODOS.................................................................................................. 40
3.1 CARACTERIZAÇÃO DA ÁREA EXPERIMENTAL........................................................ 40
3.2 CARACTERIZAÇÃO DO EXPERIMENTO...................................................................... 41
3.3 CARACTERÍSTICAS AVALIADAS.................................................................................. 46
3.3.1 Determinação do Potencial Hídrico Foliar de Base (Ψb)................................................. 46
3.3.2 Trocas Gasosas.................................................................................................................. 46
3.3.3 Índice SPAD de Clorofila................................................................................................... 47
3.4 ANÁLISES BIOQUÍMICAS................................................................................................ 47
3.4.1 Coleta e Armazenamento do Material Vegetal.................................................................. 47
3.4.2 Açúcares Solúveis Totais................................................................................................... 48
3.4.3 Açúcares Redutores............................................................................................................ 48
3.4.4 Determinação do Teor de Proteína solúvel Total.............................................................. 48
3.4.5 Atividade das Invertases (EC 3.2.1.26).............................................................................. 49
3.4.6 Atividade da enzima Nitrato Redutase (NR) (EC 1.6.6.3) “in vivo’’................................. 50
3.5 ANÁLISES DA QUALIDADE DOS FRUTOS................................................................... 50
4 RESULTADOS E DISCUSSÃO.......................................................................................... 52
4.1 POTENCIAL HÍDRICO FOLIAR DE BASE (Ψb)............................................................. 52

VII
4.2.TROCAS GASOSAS............................................................................................................ 61
4.2.1 Assimilação de CO2 (A)..................................................................................................... 61
4.2.2 Transpiração (E)................................................................................................................ 65
4.2.3 Condutância Estomática (gs)............................................................................................. 70
4.2.4 Eficiência no uso da água (A/E)........................................................................................ 75
4.2.5 Concentração interna de CO2 na câmara subestomática (Ci).......................................... 77
4.2.6 Eficiência de carboxilação in vivo da Rubisco (A/Ci)....................................................... 80
4.3 TEOR SPAD DE CLOROFILA........................................................................................... 83
4.4 ATIVIDADE DA ENZIMA NITRATO REDUTASE (NR)................................................ 85
4.5 PROTEÍNA SOLÚVEL TOTAL.......................................................................................... 89
4.6 AÇÚCAR SOLÚVEL TOTAL E AÇÚCAR REDUTOR.................................................... 93
4.7 INVERTASES: NEUTRA DO CITOSSOL (INC), ÁCIDA DO VACÚOLO (IAV) E
ÁCIDA DA PAREDE CELULAR (IAP)...................................................................................
98
4.8 ANÁLISES QUÍMICAS: ACIDEZ, pH e ºBRIX................................................................ 105
5 CONSIDERAÇÕES FINAIS............................................................................................... 112
6 CONCLUSÕES..................................................................................................................... 114
7 REFERÊNCIAS BIBLIOGRÁFICAS................................................................................ 116

VIII
LISTA DE ABREVIATURAS E SÍMBOLOS
A – taxa de assimilação líquida de CO2
A/Ci – eficiência momentânea da Rubisco
ABA – ácido abscísico
AFR – área foliar real
AFDP – área foliar da planta
AR – açúcares redutores
AST – açúcares solúveis totais
AT – acidez titulável
CV. – cultivar
Ci – concentração intracelular do CO2
CV – coeficiente de variação
DAP e Dapp – dias após a poda
DAT – dias após o tratamento
DFFFA – densidade de fluxo de fótons fotossinteticamente ativos
DI e ID – irrigação deficitária
DNS - dinitrosalicílico
DPV – défice de pressão de vapor
E - transpiração
EC – enzyme comission
En – transpiração noturna
ET - evapotranspiração
ETo – evapotranspiração de referência
ETc – evapotranspiração da cultura
EUA e WUE– eficiência no uso da água
FV – fator de variação
GL – graus de liberdade
gm – condutância do mesófilo
gn – condutância noturna
gs – condutância estomática

IX
IAP – invertase ácida da parede celular
IAV – invertase ácida do vacúolo
IDC e RDI – irrigação com défice controlado
INC – invertase neutra do citossol
IP- irrigação plena
IRC – índice relativo de clorofila
IRGA – Infrared Gas Analyzer
Kc – coeficiente da cultura
LED – diodo emissor de luz
MF – massa fresca
Mpa – mega pascal
NR – nitrato redutase
Pep-case – fosfoenol piruvato carboxilase
pH – potencial hidrogeniônico
PRD – irrigação parcial das raízes
RH – regimes hídricos
Rubisco – ribulose-1,5-bisfosfato carboxilase/oxigenase
SPAD – Soil Plant Analysis Development
SVSF – Submédio do Vale do São Francisco
Ψb – potencial hídrico foliar de base
SS – sólidos solúveis
Ψmd – potencial hídrico foliar ao meio-dia

X
LISTA DE FIGURAS
Figura 1. Temperatura (A), insolação (B), umidade (C) e radiação solar global (D) durante o
ciclo da videira cv. Syrah. Embrapa Semiárido, Petrolina-PE,
2010..........................................................................................................................43
Figura 2. Precipitação pluviométrica (mm) registrada durante o ciclo de produção da videira
cv. Syrah. Embrapa Semiárido, Petrolina-PE, 2010. Fonte: Goncalves
(2011)........................................................................................................................43
Figura 3. Armazenamento (mm; A, B, C) e variação do armazenamento (mm; D, E, F) da água
em diferentes profundidades do solo (15, 30, 45 e 60 cm) durante o ciclo de
produção da videira cv. Syrah, sob três estratégias de irrigação: irrigação plena (A e
D), irrigação com déficit controlado (B e E) e irrigação deficitária (C e F).
...................................................................................................................................44
Figura 4. Escala de Baggiolini para determinação das fases fenológicas da videira.................45
Figura 5. Valores médios da densidade de fluxo de fótons fotossinteticamente ativos (DFFFA,
μmol m-2
s-1
), medidos entre 8:00 e 9:30 horas da manhã ao longo do ciclo da
videira cv. Syrah, sob três estratégias de irrigação. Embrapa Semiárido, Petrolina-
PE, 2010....................................................................................................................65

XI
LISTA DE TABELAS
Tabela 1. Resumo da análise de variância (teste F) para o potencial hídrico foliar de base (Ψb,
MPa) em folhas de videira Syrah aos 56, 70, 86, 99 e 113 dias após a poda (DAP)
em função de três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................53
Tabela 2. Potencial hídrico foliar de base (Ψb, MPa) em folhas de videira cv. Syrah durante o
ciclo em função de três estratégias de irrigação (Irrigação com défice controlado-
IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina,
PE, 2010....................................................................................................................54
Tabela 3. Resumo da análise de variância (teste F) da taxa de assimilação de CO2 em folhas de
videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda (DAP) em função de
três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................62
Tabela 4. Taxa de assimilação de CO2 (μmol m-2
s-1
) em folhas de videira ‘Syrah’ ao longo do
ciclo após o início das três estratégias de irrigação (Irrigação com défice controlado-
IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina,
PE, 2010....................................................................................................................63
Tabela 5. Resumo da análise de variância (teste F) da taxa de transpiração em folhas de videira
‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda (DAP) em função de três
estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................66
Tabela 6. Taxa de transpiração (mmol m-2
s-1
) em folhas de videira ‘Syrah’ ao longo do ciclo
em função de três estratégias de irrigação (Irrigação com défice controlado-IDC,
Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................67
Tabela 7. Resumo da análise de variância (teste F) da condutância estomática em folhas de
videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda (DAP) em função de
três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................71
Tabela 8. Condutância estomática (mol m-2
s-1
) em folhas de Syrah ao longo do ciclo em
função de três estratégias de irrigação (Irrigação com défice controlado-IDC,
Irrigação deficitário-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................73
Tabela 9. Resumo da análise de variância (teste F) da eficiência do uso da água (assimilação
de CO2/transpiração) em folhas de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias
após a poda (DAP) em função de três estratégias de irrigação (EI). Embrapa
Semiárido – Petrolina, PE, 2010...............................................................................75
Tabela 10. Eficiência do uso da água (Assimilação de CO2 /Transpiração, μmol CO2 / mmol
H2O) em folhas de videira ‘Syrah’ ao longo do ciclo em função de três estratégias

XII
de irrigação (Irrigação com défice controlado-IDC, Irrigação deficitária-ID e
Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................76
Tabela 11. Resumo da análise de variância (teste F) da concentração interna de CO2 na câmara
subestomática em folhas de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a
poda (DAP) em função de três estratégias de irrigação (EI). Embrapa Semiárido –
Petrolina, PE, 2010....................................................................................................78
Tabela 12. Concentração interna de CO2 na câmara subestomática (μmol CO2 m-2
) em folhas de
videira ‘Syrah’ ao longo do ciclo em função de três estratégias de irrigação
(Irrigação com défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP).
Embrapa Semiárido – Petrolina, PE, 2010................................................................80
Tabela 13. Resumo da análise de variância (teste F) da eficiência de carboxilação in vivo da
enzima Rubisco em folhas de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após
a poda (DAP) em função de três estratégias de irrigação (EI). Embrapa Semiárido –
Petrolina, PE, 2010....................................................................................................81
Tabela 14. Eficiência de carboxilação in vivo da enzima Rubisco (A/Ci, μmol m-2
s-1
de CO2.
μmol mol-1
CO2) em folhas de videira ‘Syrah’ ao longo do ciclo em função de três
estratégias de irrigação (Irrigação com défice controlado-IDC, Irrigação deficitária-
ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE, 2010........................82
Tabela 15. Resumo da análise de variância (teste F) para o teor SPAD de clorofila em folhas de
videira ‘Syrah’ aos 54, 60, 73, 87 e 101 dias após a poda (DAP) em função de três
estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................83
Tabela 16. Teor SPAD da clorofila em folhas de videira ‘Syrah’ ao longo do ciclo em função de
três estratégias de irrigação (Irrigação com défice controlado-IDC, Irrigação
deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE,
2010..........................................................................................................................84
Tabela 17. Resumo da análise de variância (teste F) da atividade da enzima nitrato redutase em
folhas de videira ‘Syrah’ aos 54, 60, 73, 87 e 101 dias após a poda (DAP) em
função de três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................86
Tabela 18. Atividade da enzima nitrato redutase (NR, μmol de NO2 g MF h-1
) em folhas de
videira ‘Syrah’ ao longo do ciclo em função de três estratégias de irrigação
(Irrigação com défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP).
Embrapa Semiárido – Petrolina, PE, 2010................................................................87
Tabela 19. Resumo da análise de variância (teste F) do teor de proteína solúvel total em folhas
de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda (DAP) em função
de três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................90
Tabela 20. Teor de proteína solúvel total (μg g-1
MF) em folhas de videira ‘Syrah’ ao longo do
ciclo em função de três estratégias de irrigação (Irrigação com défice controlado-
IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina,
PE, 2010....................................................................................................................93
Tabela 21. Resumo da análise de variância (teste F) de açúcar solúvel total (AST) e açúcar
redutor (AR) em folhas de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a

XIII
poda (DAP) em função de três estratégias de irrigação (EI). Embrapa Semiárido –
Petrolina, PE, 2010....................................................................................................94
Tabela 22. Açúcar solúvel total (AST, μg g-1
de MF) em folhas de videira ‘Syrah’ ao longo do
ciclo em função de três estratégias de irrigação (Irrigação com défice controlado-
IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina,
PE, 2010....................................................................................................................95
Tabela 23. Açúcar redutor (AR, μg g-1
de MF) em folhas de videira ‘Syrah’ ao longo do ciclo
em função de três estratégias de irrigação (Irrigação com défice controlado-IDC,
Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................97
Tabela 24. Resumo da análise de variância (teste F) da atividade da invertase neutra do citossol
(INV), invertase ácida do vacúolo (IAV) e invertase ácida da parede (IAP) em
folhas de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda (DAP) em
função de três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE,
2010...........................................................................................................................99
Tabela 25. Atividade da enzima invertase neutra do citossol (INC, µmol g-1
MF h-1
) em folhas
de videira ‘Syrah’ ao longo do ciclo em função de três estratégias de irrigação
(Irrigação com défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP).
Embrapa Semiárido – Petrolina, PE, 2010..............................................................100
Tabela 26. Atividade da enzima invertase ácida do vacúolo (IAV, µmol g-1
MF h-1
) em folhas de
videira ‘Syrah’ ao longo do ciclo em função de três estratégias de irrigação
(Irrigação com défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP).
Embrapa Semiárido – Petrolina, PE, 2010..............................................................101
Tabela 27. Atividade da enzima invertase ácida da parede (IAP, µmol g-1
MF h-1
) em folhas de
videira ‘Syrah’ ao longo do ciclo em função de três estratégias de irrigação
(Irrigação com défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP).
Embrapa Semiárido – Petrolina, PE, 2010..............................................................104
Tabela 28. Resumo da análise de variância (teste F) para acidez titulável (AT), pH, teor de
sólidos solúveis (SS) e relação sólidos solúveis/acidez titulável (SS/AT) do mosto (g
L-1
) em função de três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina,
PE, 2010..................................................................................................................106
Tabela 29. Acidez titulável (AT), sólidos solúveis (SS) e relação acidez/sólidos solúveis
(SS/AT) do mosto (g L-1
) em função de três estratégias de irrigação (Irrigação com
défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa
Semiárido – Petrolina, PE, 2010.............................................................................108
Tabela 30. Potencial hidrogeniônico (pH) do mosto (g L-1
) em função de três estratégias de
irrigação (Irrigação com défice controlado-IDC, Irrigação deficitária-ID e Irrigação
plena-IP). Embrapa Semiárido – Petrolina, PE, 2010.............................................110

FISIOLOGIA E METABOLISMO DA VIDEIRA CV. SYRAH NO SUBMÉDIO DO VALE
DO SÃO FRANCISCO SOB TRÊS ESTRATÉGIAS DE IRRIGAÇÃO. Botucatu, 2012,
143p. Tese (Doutorado em Agronomia/Horticultura), Faculdade de Ciências Agronômicas,
Universidade Estadual Paulista.
Autor: CAIO MÁRCIO GUIMARÃES SANTOS
Orientador: JOÃO DOMINGOS RODRIGUES
Co-orientadora: BÁRBARA FRANÇA DANTAS
RESUMO
A vitivinicultura é uma atividade econômica, relativamente, recente no
Brasil e a região semiárida do Submédio do Vale do São Francisco, utilizando-se de irrigação
localizada, tem atraído muitos investidores internacionais para a produção de uvas e vinhos. O
presente trabalho teve o objetivo de avaliar a fisiologia e o metabolismo da videira ‘Syrah’,
sob três estratégias de irrigação (Irrigação com Défice Controlado-IDC, Irrigação Deficitária-
ID e Irrigação Plena-IP,) no Submédio do Vale do São Francisco. A pesquisa foi desenvolvida
na Embrapa Semiárido, especificamente, no Campo Experimental de Bebedouro, localizado
no município de Petrolina-PE. O delineamento experimental foi em blocos casualizados, em
três esquemas fatoriais: 3 x 5, três estratégias de irrigação e cinco avaliações para potencial
hídrico foliar de base (Ψb), teor SPAD da clorofila e atividade da enzima nitrato redutase
(NR); 3 x 6, três estratégias de irrigação e seis avaliações para trocas gasosas, açúcares
solúveis totais, açúcares redutores, proteína solúvel total e atividade das invertases e 3 x 3, três
estratégias de irrigação e três avaliações para as análises químicas de sólidos solúveis, acidez
titulável, relação sólidos solúveis/acidez titulável e pH, sendo nove repetições para cada
tratamento. A imposição da deficiência hídrica controlada a partir da fase fenológica de 'cacho
fechado' em videira cv. Syrah resultou em plantas ao longo do ciclo com maior eficiência no
uso da água e economia no recurso hídrico. O porta-enxerto ‘Paulsen 1103’ demonstra
tolerância e/ou resistência ao défice hídrico e a cultivar copa Syrah, comportamento

15
anisohídrico. A maior disponibilidade hídrica na irrigação plena promoveu as maiores taxas de
assimilação, transpiração e condutância estomática aos 73 e 87 dias após a poda. Além disso,
proporcionou maior atividade da redutase do nitrato ao final do ciclo. A menor densidade de
fluxo de fótons fotossinteticamente ativos aos 101 dias após a poda influenciou em maior
concentração interna de CO2 na câmara subestomática e menor eficiência na carboxilação in
vivo da enzima Rubisco. A manutenção de elevado teor SPAD da clorofila nas folhas entre 60
e 101 dias após a poda indica boas condições de funcionamento do aparelho fotossintético. A
irrigação deficitária conferiu maior atividade média da invertase ácida da parede ao final do
ciclo. A acidez titulável do mosto diminuiu e os açúcares solúveis totais, açúcares redutores,
sólidos solúveis e a relação sólidos solúveis/acidez titulável aumentaram para todas as
estratégias de irrigação ao final do ciclo.
_______________________________________
Palavras-Chave: Açúcar, nitrato redutase, défice hídrico, trocas gasosas, invertases.

PHYSIOLOGY AND METABOLISM OF GRAPEVINE CV. SYRAH IN SUBMEDIUM OF
THE SAN FRANCISCO VALLEY UNDER THREE IRRIGATION STRATEGIES.
Botucatu, 2012, 143p. Tese (Doutorado em Agronomia/Horticultura), Faculdade de Ciências
Agronômicas, Universidade Estadual Paulista.
Author: CAIO MÁRCIO GUIMARÃES SANTOS
Adviser: JOÃO DOMINGOS RODRIGUES
Co-adviser: BÁRBARA FRANÇA DANTAS
ABSTRACT
Viticulture is a relatively recent economic activity in Brazil and the semiarid region of the
Submedium of the San Francisco Valley, using drip irrigation, has attracted many international
investors to grapes and wine production. The study aimed to evaluate the physiology and
metabolism of grapevine ‘Syrah’ under three irrigation strategies (Regulated Deficit Irrigation-
RDI, Deficit Irrigation-DI and Full Irrigation-FI,) in the Submedium of the San Francisco
Valley. The research was conducted at Embrapa Semiarid, specifically in the Bebedouro
Experimental Field, located in Petrolina, Pernambuco-Brazil. The statistical design was in
randomized blocks, factorial in three regimens: 3 x 5, three irrigation strategies and five
evaluations of leaf water potential (Ψb), SPAD chlorophyll content and activity of the nitrate
reductase enzyme (NR); 3 x 6, three irrigation strategies and six evaluations for gas exchange,
soluble sugars, reducing sugars, total soluble protein and invertase activity, and 3 x 3, three
irrigation strategies and three evaluations for the chemical variables: soluble solids, titratable
acidity, soluble solids / titratable acidity and pH, nine replicates for each treatment. The
imposition of water deficit from the phenological phase of 'closed lock' in grapevine ‘Syrah’
results in the plant cycle with greater efficiency in water use and savings on water resources.
The rootstock ‘1103 Paulsen’ demonstrates tolerance and / or resistance to water deficit and
scion ‘Syrah’, anisohydric behavior. The greater water availability in the full irrigation

17
promotes the highest rates of assimilation, transpiration and stomatal conductance at 73 and 87
days after pruning. It also provides the highest activity of nitrate reductase at the end of the
cycle. The lower density of active photosynthetic photon flux at 101 days after pruning
influences in higher internal CO2 concentration in the substomatal chambers and lower
efficiency in vivo carboxylation enzyme Rubisco. Maintaining high levels of SPAD
chlorophyll in the leaves between 60 and 101 days after pruning indicates good working of the
photosynthetic apparatus. The deficit irrigation provides greater average activity of acid
invertase wall at the end of the cycle. The acidity of the wine is reduced and the total soluble
sugars, reducing sugars, soluble solids and soluble solids/titratable acidity increase for all
irrigation strategies at the end of the cycle.
_______________________________________
Keywords: Sugar, nitrate reductase, water deficit, gas exchange, invertases

18
1 INTRODUÇÃO
A posição geográfica, a diversidade edafoclimática, os recursos
hídricos, o uso de tecnologias e a expansão equilibrada das fronteiras agrícolas no Brasil,
possibilitam ao país gerar grandes riquezas no setor primário e se tornar uma das maiores
potências agrícolas mundiais do século XXI.
Entre os ramos do setor agrícola em grande expansão está a
fruticultura, com destaque para a região Nordeste, maior exportadora de frutas tropicais do
país e de algumas frutas de clima temperado, a exemplo da uva. A uva de mesa cultivada,
principalmente, no perímetro irrigado dos Estados de Pernambuco e Bahia é uma das frutas
com maior valor agregado na exportação, tendo em vista o uso de tecnologias modernas de
produção e a possibilidade de disponibilizar frutos no mercado internacional durante o período
da entressafra dos maiores produtores mundiais. Em 2010 a produção de uvas no Brasil para
consumo ‘in natura’ foi de 1.300.000 toneladas métricas, ocupando o quarto lugar entre os
maiores produtores mundiais dessa fruta (AGRIANUAL, 2011).
A viticultura tropical brasileira foi efetivamente desenvolvida a partir
da década de 1960, com o plantio de vinhedos comerciais de uva de mesa na região do SVSF,

19
no Nordeste Semiárido brasileiro, onde o uso de tecnologia permite obter alta produtividade e
um produto de qualidade superior, com amplas condições de acesso ao mercado internacional.
Na agricultura irrigada do SVSF, a cultura da videira destaca-se como
um exemplo de sucesso, apresentando na última década notável expansão da área colhida,
passando de 1.759 ha em 1990 (AGRIANUAL, 1997) para 11.034 ha em 2010
(AGRIANUAL, 2011) e a diversificação de atividades com a exploração da produção de
vinhos tintos e espumantes já se encontram estabelecidas, com uma produção relativamente
crescente e bem aceita no mercado interno e externo. Contudo, por ser uma atividade
relativamente recente nas condições semiáridas do Nordeste requer informações científicas
adequadas à realidade da região, no que tange à cultivar, produção, produtividade, qualidade
dos frutos e, consequentemente, dos vinhos.
Os resultados de produção, produtividade e qualidade de uma cultivar
estão diretamente relacionados com as condições de solo e clima do local de cultivo e o
manejo adotado. Dentro desse enfoque são numerosas as possíveis variáveis que influenciam
na composição da uva e, consequentemente, na qualidade potencial de seus subprodutos.
Análises fisiológicas de acompanhamento do crescimento e
desenvolvimento das plantas cultivadas facilitam a compreensão e indicam as possíveis
respostas das mesmas às condições externas (solo, água, clima e manejo adotado) às quais
estão submetidas.
Na região semiárida nordestina, na maioria das vezes, o fator limitante
ao crescimento e desenvolvimento vegetal é a ausência quase na sua totalidade do recurso
natural água e/ou a sua má distribuição. Neste contexto, o uso da técnica da irrigação se torna
imprescindível como fator de produção.
A irrigação racional nos cultivos agrícolas proporciona uma hidratação
adequada a nível celular, além de condicionar maior e melhor absorção de nutrientes
essenciais para um bom funcionamento do metabolismo primário e secundário das plantas.
Pesquisas com irrigação fornecem informações da evapotranspiração e
quantidade de água consumida pelas plantas durante o ciclo. Um maior ou menor consumo de
água pela videira está diretamente relacionado com seu estádio fenológico, com ênfase no
consumo durante a floração e produção de frutos.

20
O défice hídrico quando severo é uma das principais barreiras à
produtividade dos vinhedos, pois altera o metabolismo primário e secundário da videira, além
de reduzir a qualidade e a produtividade de frutos. No entanto, a videira é uma planta
relativamente tolerante ao estresse hídrico, onde um menor aporte de água dentro de um
intervalo de tolerância e durante o seu desenvolvimento pode conferir até mesmo
características bioquímicas de qualidade aos frutos, como aumento no teor de sólidos solúveis
e polifenóis.
A produção de certos fitoquímicos durante a formação, crescimento e
maturação das bagas da uva é influenciada grandemente, pelas condições edafoclimáticas,
sendo o fornecimento de água um dos fatores que mais interferem na qualidade do fruto.
A Syrah é uma das cultivares de Vitis vinifera mais plantadas na região
do SVSF para a produção de vinhos tintos finos de mesa. A determinação de informações
relacionadas a estratégias de irrigação e ao metabolismo de carboidratos necessário para essas
plantas durante o seu ciclo é de grande importância, pois influencia diretamente na
acumulação de monossacarídeos (glicose e frutose) que serão utilizados pelas leveduras
durante o processamento da uva para a produção do vinho.
Atualmente, há uma necessidade de aprimoramento das práticas
agronômicas que visem melhorar a qualidade da produção vinícola nordestina, uma vez que a
composição da uva é um fator determinante para a qualidade dos vinhos. Todavia, pesquisas
direcionadas à fisiologia e bioquímica de plantas em campo geram informações que facilitam
e auxiliam a tomada de decisão dos vitivinicultores no melhor manejo dos parreirais.
Considerando o exposto, o presente trabalho teve o objetivo de avaliar
a fisiologia e o metabolismo da videira cultivar Syrah sob três estratégias de irrigação (IDC-
Irrigação com Défice Controlado, ID- Irrigação Deficitária e IP-Irrigação Plena) nas condições
edafoclimáticas do Submédio do Vale do São Francisco.

21
2 REVISÃO DE LITERATURA
2.1 Vitivinicultura
A videira é uma planta pertencente à família Vitaceae, gênero Vitis,
possuindo inúmeras espécies, destacando-se a Vitis vinifera L. conhecida como produtora de
uvas finas, de origem européia (BOLIANI et al., 2008) e a mais difundida comercialmente
para a produção de frutos destinados ao consumo ‘in natura’ e para o processamento de
vinhos. É uma planta muito cultivada desde os tempos mais remotos antes de Cristo até a
atualidade, tendo sido citada nas escrituras sagradas da bíblia. A sua domesticação ocorreu há
cerca de 10.000 anos atrás, no Oriente Médio, mais precisamente na Região do Cáucaso, entre
o Mar Negro e o Mar Cáspio, a partir da espécie selvagem Vitis vinifera caucásica (OLMO,
1995; EBELER e THORNGATE, 2009)
A viticultura tem grande importância sócioeconômica e cultural para
diversos países, principalmente, devido ao uso dos frutos como matéria prima para a
fabricação de vinhos e outros derivados (YANG et al., 2009). Em 2009, os parreirais

22
ocuparam mais de 7,5 milhões de hectares atingindo a produção mundial de 67,5 milhões de
toneladas (FAO, 2011).
Na atualidade, a vitivinicultura tem se expandido para diversas regiões
no mundo, com muitos contrastes de clima e solo. Aliado a isso, destaca-se que a tradição
enológica não favorece a introdução de novos materiais genéticos (novas cultivares) e,
portanto, a parte que mais se pode alterar é no manejo do vinhedo. Nesse sentido, deve-se
sempre levar em consideração as relações de proporcionalidade que existem na planta de
videira, como: parte aérea x raiz e crescimento vegetativo (folhas e ramos) x produção
(cachos). Ambas as relações são coordenadas pela proporção entre fonte e dreno que existe
entre os diferentes tecidos de uma planta (SANTOS, 2006).
No Brasil a videira é cultivada em quase todas as regiões, tendo sua
produção concentrada nas regiões Sul, Sudeste e Nordeste. Na região Sul, a uva é destinada à
elaboração de vinhos e sucos; no Sudeste, além da elaboração de vinhos e sucos, as uvas são
consumidas ‘in natura’ e no Nordeste, a produção é quase totalmente destinada à mesa
(ELIAS, 2008).
A viticultura é uma atividade econômica de grande potencial produtivo
na região semiárida do Nordeste brasileiro, com pouca tradição no cultivo, quando comparado
às tradicionais regiões Sul e Sudeste. Na agricultura irrigada do Vale do São Francisco, a
cultura da videira destaca-se como um exemplo de sucesso, devido ao aporte tecnológico
empregado, conferindo alta produtividade e um produto de qualidade superior, com amplas
condições de acesso ao mercado internacional, podendo-se obter até mais de duas safras/ano, o
que é regulado pelas janelas de mercado.
O desenvolvimento da vitivinicultura no Brasil está baseada em grande
parte nos resultados de pesquisa que buscam aprimorar as práticas de cultivo, visando maiores
produtividades e melhorando a qualidade da uva produzida (MALAGI, 2011).
O Brasil é o quinto maior produtor de vinhos do hemisfério Sul. Os
vinhedos do Vale do São Francisco são os únicos do mundo com duas colheitas por ano, no
paralelo 9o
Sul, com planícies e clima extremamente seco, onde o cultivo da uva é possível
através de irrigação (WINES FROM BRASIL, 2009).
No SVSF, a vitivinicultura tem evoluído no sentido de aprimorar a
aplicação de práticas agronômicas que visem melhorar a qualidade da produção vinícola, uma

23
vez que a composição da uva é um fator determinante para a qualidade dos vinhos dessa
região caracterizados como jovens, frutados e aromáticos, denominados Vinhos do Sol, que
estão conquistando cada vez mais o mercado brasileiro e europeu. Além disso, as condições
climáticas dessa região caracterizam-se pela alta disponibilidade de radiação solar, intensidade
de luz solar e baixa precipitação pluvial, o que leva à necessidade da prática da irrigação para
suprir as necessidades hídricas das plantas (GONÇALVES, 2011).
2.2 Características da cultivar Syrah
A cultivar Syrah é uma das mais antigas das castas cultivadas.
Algumas referências sugerem que esta seria originária de Schiraz, na Pérsia, outras, que seria
nativa da Vila de Siracusa, na Sicília. Independentemente de sua origem, é cultivada na França
há muito tempo, principalmente no Vale do Rhône (FRAGA, 2010). A denominação ‘Syrah’ é
mais utilizada na França. Nos Estados Unidos, África do Sul, Austrália e Canadá é mais,
frequentemente, chamada ‘Shiraz’ (FAVERO, 2007). A planta caracteriza-se por possuir porte
semi-ereto e sua identificação é facilmente percebida devido à presença abundante de pelos de
coloração branca na extremidade do ramo. Apresenta folhas jovens de coloração verde clara e
folhas adultas penta lobadas, com seio peciolar aberto. Seus ramos são frágeis de coloração
verde clara e entrenós longos. Os cachos são médios, mas compactos e cilíndricos, de bagas
ovaladas e pequenas (ELIAS, 2008). É uma cultivar muito vigorosa, produtiva, respondendo
bem à poda curta em regiões quentes. Possui um curto período de maturação e revela-se
bastante sensível à podridão no final da maturação. Origina vinhos de coloração intensa,
aromáticos, finos e complexos, aptos ao envelhecimento e de grande qualidade (ENTAV,
1995 apud FAVERO, 2007).
No Brasil essa cultivar é pouco explorada na região Sul, maior
produtora de uvas e vinhos, em função da sua grande sensibilidade à podridão do cacho.
Entretanto, nas condições semiáridas do SVSF tem mostrado ótimo desempenho
(CAMARGO, 2009), sendo juntamente com a ‘Cabernet Sauvignon’ as castas mais cultivadas
e com bons resultados em produção e qualidade de vinhos tintos finos (MOURA et al., 2007) .

24
2.3 Características do Cultivo da Videira
Empregando-se tecnologias de produção (manejo da irrigação,
nutrição, poda e tratos fitossanitários) e dispondo de estudos do comportamento fenológico e
melhoramento genético, a região Nordeste do país vem se destacando na produção de uvas
para vinho e uvas finas de mesa para o consumo ‘in natura’, ambas voltadas tanto para o
mercado nacional como para o internacional.
Com relação às técnicas de manejo da videira, a escolha correta do
sistema de condução e a eficiência no uso da água (EUA) podem afetar significativamente o
crescimento vegetativo da videira, a produtividade do vinhedo e a qualidade da uva e do
vinho. As características do sistema de condução da planta exercem esse efeito em função da
altura e da largura do dossel vegetativo; da divisão do dossel em cortinas; do posicionamento
das gemas e dos frutos; da carga de gemas/ha e do espaçamento entre fileiras e entre plantas
(NORBERTO, 2006). Assim, a quantidade e a distribuição das folhas no espaço modificam o
microclima (temperatura das folhas, umidade, intensidade de radiação, etc) no interior do
dossel vegetativo (MIELE e MANDELLI, 2003).
Os efeitos da interação entre a forma do dossel vegetativo e os fatores
ambientais locais resultam no microclima que determinará alterações em alguns aspectos
fisiológicos da videira, podendo afetar significativamente o crescimento vegetativo, a
produtividade do vinhedo, bem como a qualidade da uva e do vinho (NORBERTO et al.,
2009).
2.4 Ecofisiologia da videira
O fator ambiental clima determina as condições para o crescimento da
planta e sua área de distribuição e impõe limites para sua sobrevivência (LARCHER, 2000).
A videira é uma planta adaptada a regiões de clima temperado, por
apresentar folhas decíduas e o seu cultivo está estabelecido em uma ampla faixa compreendida
entre as latitudes de 52ºN e 40ºS (PEDRO JUNIOR e SENTELHAS, 2003). No Brasil, a

25
videira vem sendo cultivada, com sucesso, desde regiões frias e úmidas do Rio Grande do Sul
até regiões quentes e secas do semiárido nordestino.
As condições climáticas influem na fenologia e fisiologia das plantas e
na produção e qualidade dos frutos. Portanto, as videiras cultivadas em clima semiárido
apresentam comportamento fisiológico e bioquímico totalmente distinto daquele das regiões
de clima subtropical e temperado, estando condicionada ao controle da irrigação e época de
poda.
Um dos principais problemas nos vinhedos irrigados para produção de
vinhos de qualidade é o aumento do vigor vegetativo, causando desequilíbrio entre área foliar
e produção de frutos, e quando a irrigação é excessiva resulta em elevado vigor dos ramos,
aumentando a competição por fotoassimilados entre frutos e ramos; alterando o microclima na
região dos cachos, comprometendo a síntese de compostos fenólicos; reduzindo a fertilidade
das gemas e dificultando os tratamentos fitossanitários (JACKSON e LOMBARD, 1993).
Dentre os vários recursos para controlar o vigor vegetativo, o que mais tem recebido atenção
nos últimos anos tem sido o manejo da irrigação, uma vez que a composição da uva é
fortemente influenciada pelo estado hídrico da videira (DRY e LOVEYS, 1998). Vinhedos
submetidos à irrigação plena durante todo o ciclo de produção resultam na produção de vinhos
de baixa qualidade. A imposição de estresse hídrico moderado à videira em determinadas fases
fenológicas tem impacto positivo sobre a concentração de açúcares, acidez, pH e compostos
fenólicos no mosto (DELOIRE et al., 2004), sendo estes últimos responsáveis pela cor, flavor,
corpo e estrutura dos vinhos tintos.
Para Larcher (2000) a ecofisiologia vegetal é a ciência que analisa os
processos e as respostas das plantas em função das mudanças nos fatores ambientais. As
pesquisas relacionadas à fisiologia vegetal e ao hábito de crescimento das plantas sob
diferentes condições possibilitam a implantação de sistemas de produção que atendam as
demandas específicas de cada espécie nos diferentes microclimas e tipos de solos
(LARCHER, 2000; BORGHEZAN, 2010).
O acúmulo de matéria seca por uma planta ou por uma cobertura
vegetal está diretamente relacionado com a captação de energia solar pelo aparelho
fotossintético e a quantidade de radiação fotossinteticamente ativa que incide sobre a cobertura
vegetal (FERNANDES, 2009). Além da luz, outros fatores como temperatura (NOGUEIRA,

26
1984; NORBERTO et al., 2009; KRIEDEMANN, 1977; 1968), umidade relativa
(SEPÚLVEDA e KLIEWER, 1986), disponibilidade hídrica (ALLEWELDT e RUHL, 1982;
COSTACURTA e ROSELLI, 1980; HIDALGO, 1993; WINKLER, 1970), idade da folha
(ALLEWELDT et al., 1982; KLOBET, 1984; KRIEDEMANN, 1968; 1977; KRIEDEMANN
et al., 1970), concentração de CO2 e O2 (KRIEDEMANN, 1968; 1977), relação fonte-dreno e
cultivar (FERNANDES, 2009) contribuem para explicar a variabilidade do comportamento
fotossintético.
A radiação influencia a abertura dos estômatos, órgãos responsáveis
pelas trocas gasosas da folha, bem como, pelo estímulo ao nível dos cloroplastos. A videira é
uma planta de sol que realiza a fixação do CO2 atmosférico segundo o metabolismo C3 (TAIZ
e ZEIGER, 2009). Este tipo de planta apresenta, na ausência de fatores limitantes, uma
resposta da fotossíntese caracterizada por uma primeira fase de aumentos proporcionais e uma
segunda fase de aumentos decrescentes, até atingir a taxa fotossintética máxima
correspondente ao ponto de saturação (FERNANDES, 2009).
Altas e baixas temperaturas, dependendo da intensidade e da duração,
impedem as atividades metabólicas, o crescimento e desenvolvimento e a produtividade das
plantas. O estresse pelo calor, primeiramente provoca distúrbios no aparelho fotossintético,
inativando enzimas termolábeis, desorganizando o metabolismo dos ácidos nucléicos e
proteínas e degenerando as biomembranas celulares. A baixa temperatura, inicialmente,
provoca a parada da corrente citoplasmática, atingindo em seguida, a fotossíntese, com lesões
nas biomembranas e interrupção do fornecimento de energia celular (LARCHER, 2000).
Ambos os estresses pelo calor ou pelo frio quando não reversíveis podem provocar a morte
celular.
A umidade relativa do ar está diretamente relacionada com a
evapotranspiração e com os aspectos fitossanitários da videira. Em condições de baixa
umidade relativa, como em regiões áridas e semiáridas, o fluxo transpiratório é intenso,
principalmente, nas horas mais quentes do dia, provocando o fechamento estomático como
forma de defesa das plantas contra a perda excessiva de água para a atmosfera. O vento, ao
influenciar as transferências de calor de matéria e de energia por convecção, tem também
papel importante nas perdas de água de uma cobertura vegetal (LOPES, 1994). Além disso,
favorece a proliferação de ácaros e oídio. Em condições de alta umidade relativa às plantas

27
apresentam desenvolvimento de ramos mais vigorosos, porém aumenta a incidência de
doenças fúngicas (MOTA et al., 1974; SENTELHAS, 1998). A umidade relativa ideal,
considerando o manejo fitossanitário, seria entre 62% e 68% (KISHINO e CARAMORI,
2007).
Entre os recursos naturais que as plantas precisam para o crescimento,
a água é o mais limitante para a produtividade agrícola, visto ser essencial aos diversos
processos metabólicos, principalmente, durante o período inicial de desenvolvimento (SOUSA
et al., 2001).
A videira é uma planta bastante resistente à seca, adaptando-se bem,
desde regiões com 200 mm (PEDRO JUNIOR e SENTELHAS, 2003), 250 mm (TONIETTO
e MANDELLI, 2003), 400 mm (WILLIAMS e MATTHEWS, 1990; EVANS et al., 1993;
ÁVILA NETO et al., 2000) a 1000 mm (PEDRO JUNIOR e SENTELHAS, 2003) 1200 mm
(WILLIAMS e MATTHEWS, 1990; EVANS et al., 1993; ÁVILA NETO et al., 2000) de
chuva durante o ciclo. A demanda hídrica da videira varia em função das fases do ciclo
fenológico, sendo que uma das características da sua capacidade de adaptação a regiões
relativamente áridas é a presença de um sistema radicular profundo (DOORENBOS e
KASSAN, 1994).
A irrigação tem a possibilidade de interferir de forma direta no estado
hídrico das videiras, tornando-a uma técnica desejável e ao mesmo tempo controversa
(KLIEWER et al., 1983; MATTHEWS e ANDERSON, 1988; GIRONA, 2005).
Irrigar a videira é promover um défice hídrico tolerado pelas plantas,
de modo que as mesmas mantenham seu metabolismo compatível com uma produção de
qualidade. Em várias regiões do mundo, os viticultores utilizam a irrigação em períodos de
seca, de forma a promover a estabilidade da produção e da sua qualidade. No entanto, a sua
utilização não é desprovida de inconvenientes; o maior problema resultante desta prática é o
excesso de vigor derivado do crescimento contínuo dos novos brotos, que tem repercussões na
composição dos frutos (MCCARTHY, 2002) e na sua qualidade fitossanitária. Assim, é
necessário estabelecer o equilíbrio entre o crescimento vegetativo e reprodutivo da videira
para tirar o máximo proveito da técnica da irrigação.
Para o melhor uso dos recursos hídricos, atuais e futuros, a irrigação
deficitária, definida como a aplicação de água abaixo das necessidades totais da cultura (ET,

28
evapotranspiração), constitui instrumento importante na redução do uso de água em culturas
irrigadas (FERERES e SORIANO, 2006), sem prejuízo ao processo produtivo.
A carência e/ou abundância hídrica geram estresses nas plantas, o que
pode resultar em uvas de baixa qualidade, com reflexos na produção e processamento das
mesmas. O equilíbrio entre esses dois extremos (abundância e carência) está na gestão de um
défice hídrico moderado em fases fenológicas com maior resposta à disponibilidade hídrica
(WINKLER et al., 1974; KOUNDOURAS et al., 1999). Desta forma, a utilização criteriosa da
irrigação poderá contribuir em resultados positivos concretos (ESCALONA et al., 1999;
CABRITA et al., 2001; GAUDILLERE et al., 2002).
O estado hídrico da planta tem sido reconhecido como um fator
determinante que influencia em todos os aspectos do crescimento e qualidade das uvas. Por
este motivo, a técnica do controle do estresse hídrico é agora utilizada de forma extensa para
manipular a uva para produzir vinho de qualidade (DRY et al., 2001). É conhecido que um
determinado nível de estresse hídrico pode surpreendentemente melhorar a qualidade da uva e,
portanto, do vinho (ONCINS et al., 2005).
Vários estudos realizados nas principais regiões vitivinícolas do
mundo, visando avaliar o impacto de estratégias de irrigação durante fases fenológicas da
videira, constataram que um estresse moderado nas plantas proporciona redução no
crescimento dos ramos e melhoria na composição das bagas, como o aumento na concentração
de açúcares, taninos e antocianinas (Mc CARTHY, 1997; 2002; WAMPLE, 2002;
PEDREIRA DOS SANTOS et al., 2007).
A irrigação deficitária é defendida pela maioria dos autores como a
melhor estratégia para controlar o estresse hídrico (CIFRE et al., 2005) e o vigor das videiras
(DRY et al., 2001; KANG e ZHANG, 2004; BRAVDO, 2005). O desenvolvimento de
estratégias de irrigação deficitárias tiveram como objetivo o aumento da eficiência do uso da
água pelas culturas (LOVEYS et al., 1997; DRY et al., 2001; MAROCO et al., 2002;
FLEXAS et al., 2004; GU et al., 2004; SOUZA et al., 2005), levando a importantes reduções
do uso da água na agricultura (STIKIC et al., 2003). A irrigação deficitária permite melhorar o
microclima ao nível dos cachos (BRAVDO, 2005) conseguindo-se ganhos em muitos dos
parâmetros qualitativos das bagas (DRY et al., 2001). Em estudos onde se compararam
diferentes estratégias de irrigação deficitária com uma testemunha irrigada plenamente, de

29
forma a manter sempre uma elevada condição hídrica, verificaram-se valores mais elevados
dos açúcares totais (SCHULTZ et al., 1996; LOPES et al., 2001), da concentração de
antocianinas (MATTHEWS et al., 1990; HAMMAN e DAMI, 2000; DRY et al., 2001), dos
compostos fenólicos (MATTHEWS e ANDERSON, 1988; DRY et al., 2001;
PETERLUNGER et al., 2005) e na razão película/polpa das bagas (ROBY et al., 2004) com a
irrigação deficitária.
As duas estratégias de irrigação mais utilizadas pelos países
produtores, no manejo do crescimento vegetativo e reprodutivo da videira, além de aumentar a
eficiência no uso da água são: irrigação parcial das raízes-PRD (COSTA et al., 2007; BIDON
et al., 2008; INTRIGLIOLO e CASTEL, 2009) e irrigação com défice controlado-RDI
(WAMPLE, 2002; KANG e ZHANG, 2004; PONI et al., 2005) . Essas duas estratégias de
irrigação apresentam alguns resultados diferentes em relação à produção e produtividade de
uva e vinho. Para Mc Carthy (2002) a PRD possui vantagens em relação à RDI por não
apresentar perda de rendimento de frutos e favorecer uma maior economia de água na
irrigação. No entanto, a RDI proporciona melhor controle no tamanho das bagas e na
qualidade final da uva e do vinho.
A RDI consiste na imposição de estresse hídrico às plantas através da
redução da quantidade de água aplicada na irrigação em determinado período durante o ciclo
de produção. No caso específico da videira, a irrigação é interrompida entre a fase da fixação
dos frutos e o pintor onde se conseguem melhores resultados no controle do vigor (DRY e
LOVEYS, 1998). Também em outras espécies cultivadas, como pereira (GOODWING e
BOLAND, 2002), pessegueiro e oliveira (FERNÁNDEZ et al., 2006), a RDI tem demonstrado
ser excelente técnica para alcançar bom equilíbrio entre o crescimento vegetativo e
reprodutivo.
Uvas produzidas com o manejo RDI apresentam aumento na acidez e
redução do pH, além de maior concentração de compostos fenólicos devido ao aumento da
razão entre película e polpa (DRY et al., 2001; WAMPLE et al., 2002).
Além de melhorias na qualidade da uva e dos vinhos, o uso da
irrigação com restrição hídrica representa economia de água utilizada na agricultura,
proporcionando a maximização da rentabilidade ao vinicultor. Além disso, atualmente existe

30
uma preocupação mundial quanto ao uso racional dos recursos hídricos, existindo uma pressão
sobre os produtores para que haja controle mais efetivo da irrigação (LOVEYS et al., 2004).
Portanto, o uso adequado dos recursos hídricos com défice controlado
nas videiras, além de contribuir na redução dos gastos e desperdício de água, aumenta a
concentração de fenóis que conferem melhores propriedades organolépticas e sensoriais ao
vinho, reduzindo também os riscos do processo de salinização nos solos do semiárido
nordestino muito susceptível a esse tipo de degradação.
2.4.1 Fenologia
É o estudo de eventos ou estádios de crescimento que se repetem
sazonalmente, e sua relação com vários fatores climáticos incluindo temperatura, radiação
solar e comprimento do dia. O propósito do estudo fenológico é descrever ou correlacionar a
duração dos estádios de crescimento com os fatores climáticos ou com outros eventos
fenotípicos. Graus-dia, crescimento em graus-dia e somatório de calor são variáveis usadas
para correlacionar a temperatura ambiente com o estádio de crescimento das plantas
(MULLINS et al., 2000).
O ciclo vegetativo da videira em regiões de clima temperado encontra-
se entre a brotação e a queda das folhas (dormência), enquanto o mesmo ciclo em regiões
tropicais se diferencia na não existência de queda das folhas e/ou um período de dormência
propriamente dito, apresentando apenas redução nas atividades metabólicas em função da
redução na disponibilidade hídrica às plantas, após a colheita dos frutos. Contudo, segundo o
modelo de Baillod e Baggiolini (1993), o ciclo é dividido em vários estádios fenológicos,
compreendendo 16 fases, designadas pelas letras A e P. No entanto, o ciclo vegetativo pode
ser dividido em três períodos principais, identificados quando 50% das gemas, das flores e das
bagas alcançam cada evento (LEEUWEN et al., 2004; DUCHÊNE e SCHNEIDER,2005;
LEBON et al., 2008). O primeiro período com intenso crescimento vegetativo seria a brotação,
onde ocorre o desenvolvimento dos ramos e folhas e das estruturas florais, a partir da
mobilização das reservas acumuladas nos ramos e raízes. O segundo período compreenderia o
intervalo entre a floração e a frutificação, caracterizado pela polinização, fixação dos frutos

31
(fruit set) e a formação das bagas. E o terceiro período resultaria na maturação, que inicia com
a mudança de cor das bagas, conhecida também pelo termo francês véraison e se estenderia
até a colheita da uva madura.
A avaliação dos estádios fenológicos é realizada visualmente na área
cultivada e propicia o conhecimento e a definição das épocas em que ocorrem as diversas
fases do período vegetativo e reprodutivo das videiras, o que pode facilitar na utilização das
práticas culturais. A caracterização fenológica por meio de estádios permite maior
detalhamento na descrição do ciclo da planta, em relação à utilização das fases, já que estas
podem ser demasiadamente distanciadas no tempo. Assim, torna-se possível utilizar a
fenologia para finalidades bem mais específicas, como em adubações de cobertura, em
tratamentos fitossanitários ou na observação de um evento importante qualquer (estresse
hídrico, térmico, etc), associado aos estádios bem definidos (ELIAS, 2008).
Segundo Leão (1999), o estudo da fenologia é de fundamental
importância para o planejamento das atividades a serem realizadas no vinhedo, bem como para
a previsão da data de colheita e comercialização.
2.4.2 Crescimento de Ramos
O crescimento é definido como aumento irreversível no tamanho das
plantas, e tem dois componentes: 1) aumento no tamanho das células já existentes e 2)
aumento no número de células pela divisão das células meristemáticas. No alongamento
celular ocorre a entrada de água na célula, resultante da diferença de potencial hídrico entre a
célula e os componentes ao seu redor. A propriedade mecânica dos tecidos celulares também
tem papel no processo de alongamento. No entanto, a regulação do crescimento é através da
ação hormonal, a qual, por sua vez, afeta os processos constituintes da divisão e alongamento
celular (MULLINS et al., 2000).
Em videira, o crescimento das estruturas vegetativas derivadas de
gemas compostas é quase exponencial na estação de crescimento. Em sequência à antese, a
taxa de crescimento vegetativo diminui e a curva de crescimento da parte aérea (folhas e
ramos ou hastes principais) sob condições de campo torna-se sigmóide (DE LA HARPE e

32
VISSOR, 1985; WILLIAMS, 1987). Este tipo de curva de crescimento ocorre se a variável de
tempo e o calendário de dias ou graus-dia for maior que 10oC. O aumento na massa seca de
ramos é quase linear até a frutificação, quando a maioria do incremento em massa seca da
videira é alocado para o desenvolvimento dos cachos.
O broto é a unidade primária de crescimento da videira e o principal
foco de muitas práticas culturais. Além disso, são também as hastes verdes crescentes que
surgem a partir de uma gema. Brotos primários surgem das gemas primárias e são
normalmente os ramos produtores de frutos. O eixo principal dos ramos consiste de tecidos de
sustentação e condução para transporte de água, nutrientes e produtos da fotossíntese.
Organizado ao longo do ramo em padrão regular estão as folhas, gavinhas, flores ou cachos de
frutos e gemas. Os ramos possuem muitos pontos de crescimento, mas o crescimento em
extensão ocorre a partir do broto apical. Novas folhas e gavinhas surgem a partir do ápice,
assim como cresce o ramo (HELLMAN, 2003).
O ramo é a haste longa e flexível que sai do tronco ou do braço. Braço
ou cordão é o ramo de mais de dois anos, ambos servem de condução à solução do solo
absorvida pela raiz e aos carboidratos elaborados pelas folhas, e funcionam também como
depósitos dos fotoassimilados. Broto é o nome dado ao ramo no início de seu
desenvolvimento. Sarmento ou bacelo é o ramo do ano maduro, lenhoso e sem folha, contendo
gemas dormentes no outono-inverno. Na extensão do sarmento encontram-se os entrenós, em
geral, mais curtos na base junto ao braço e separados por nós. No nó concentra-se maior
quantidade de amido. Sobre cada nó existe uma folha e, no mesmo lado desta, encontra-se
uma gema principal e outras duas secundárias (KISHINO, 2007).
2.4.3 Relação Fonte/Dreno
O acúmulo de matéria seca no órgão de interesse econômico é
controlado pela troca de metabólitos entre os tecidos fornecedores de fotoassimilados (a fonte)
e o órgão colhido (dreno) (PIMENTEL, 1998). A produtividade é influenciada por
características morfológicas e fisiológicas da fonte (órgãos fotossintetizantes) e do dreno
(órgãos consumidores dos metabólitos fotossintetizados, carboidratos, principalmente). Toda

33
produção de fitomassa depende da atividade fotossintética da fonte, porém a assimilação de
CO2 é apenas um dos muitos fatores que influenciam o crescimento e desenvolvimento vegetal
(FOYER e GALTIER, 1996).
Em fruteiras, o fruto é o dreno de importância econômica, de forma
que a relativa partição de matéria seca direcionada para o fruto irá determinar em parte, a sua
qualidade final. Contudo, o potencial da quantidade de fotoassimilados que podem ser
transportados para os drenos está diretamente ligado à atividade fotossintética de uma fonte
produtora de fotoassimilados (ZAMSKI, 1996).
A idade da folha é um componente importante para a expressão do
potencial genético de produção primária da videira. Kriedemann et al. (1970) e Poni et al.
(1994) estudando folhas de videira encontraram que a taxa fotossintética máxima de uma folha
totalmente expandida ocorria entre 30 e 40 dias após a brotação da gema, reduzindo em
seguida até a senescência. Em outro estudo, Intrieri et al. (2001) encontraram que entre 20 e
30 dias seria o tempo necessário para a completa expansão da lâmina foliar, alcançando o seu
máximo valor em área. A taxa de assimilação segue o mesmo padrão, sendo inicialmente
baixa (ponto de compensação próximo aos 8 dias) e entre os 20 e 30 dias atinge valores
máximos. No entanto, Schultz et al. (1996) relataram que folhas opostas ao cacho mantêm
altas taxas de fotossíntese até meses próximos à colheita.
Os compostos orgânicos (aminoácidos, ácidos orgânicos e açúcares)
formados com a fixação de CO2 atmosférico diferem com a idade da folha (RIBÉREAU-
GAYON, 1968). A relação difusiva da concentração de CO2 e O2 no mesofilo foliar de
videiras que são plantas com metabolismo C3 determina o seu rendimento fotossintético. O
enriquecimento em CO2 da atmosfera deverá beneficiar a fotossíntese dessas plantas,
diminuindo a importância da fotorrespiração (SAGE et al., 1990), assim como o seu uso de
água e nutrientes. No entanto, em condições de luz e temperatura estressantes, e uma maior
disponibilidade de O2 ao complexo enzimático Ribulose-1,5-bifosfato carboxilase/oxigenase
no ciclo de Calvin-Benson, pode provocar aumentos nas taxas fotorrespiratórias das plantas
diminuindo a produção de biomassa.
Nos tecidos fotossinteticamente ativos, como folhas maduras, a
produção de carboidratos é maior do que a sua necessidade para manutenção do metabolismo
e crescimento; desta forma, exportam excedentes, na forma de sacarose para tecidos que são

34
fotossinteticamente menos ativos ou inativos, como folhas jovens, raízes cachos ou ramos
(DANTAS et al., 2007a).
A maior dificuldade para melhorar a produtividade das videiras em
condições de campo é o estabelecimento de uma relação fonte-dreno adequada para cada
variedade e condições de cultivo (KRIEDERMANN, 1968). Os frutos são os maiores drenos
da videira, após o processo de frutificação (MULLINS, 1992).
2.5 Metabolismo da Videira
2.5.1 Carboidratos
Os carboidratos são sintetizados em órgãos fotossinteticamente ativos
(fonte) e transportados a órgãos menos ativos ou inativos (dreno) na forma de sacarose. A
hidrólise de sacarose no tecido de utilização (dreno) contribui para o descarregamento do
floema. Este fenômeno liga o metabolismo dos drenos com o transporte de sacarose e a
distribuição desta entre os vários drenos da planta. Entre as enzimas do metabolismo de
carbono, as invertases (EC 3.2.1.26) são usualmente classificadas de acordo com o seu ponto
isoelétrico, pH ótimo e localização celular. Sua atividade está inversamente relacionada à
concentração de sacarose, porém, positivamente correlacionada com as concentrações de
glicose e frutose em muitos tecidos vegetais. As invertases ácidas, localizadas no vacúolo e
parede celular, estão relacionadas com o mecanismo de translocação da sacarose do apoplasto
para o simplasto e as invertases neutras intracelulares e solúveis são envolvidas na
mobilização da sacarose do vacúolo para atender à demanda metabólica da célula (DANTAS
et al., 2005).
Tecidos fotossinteticamente ativos, como folhas maduras, produzem
mais carboidratos do que as mesmas necessitam para manterem seu metabolismo e
crescimento, exportando os fotoassimilados excedentes, na forma de sacarose, para tecidos
fotossinteticamente menos ativos ou inativos, como folhas jovens, raízes, cachos ou ramos. No

35
entanto, as relações fonte-dreno não são estáticas. Durante o crescimento vegetativo, a maioria
dos carboidratos é transportada para as raízes e folhas jovens, enquanto, após o florescimento,
os carboidratos são direcionados prioritariamente para os frutos, tubérculos e raízes de reserva
(ROITSCH et al., 2003).
De acordo com Palliotti e Cartechini (2001), os teores de glicose,
frutose e sacarose, bem como a atividade de enzimas do metabolismo de carboidratos em
folhas de videiras (Vitis vinifera L.), são influenciados por variações sazonais e fenológicas.
Tanto a expressão gênica quanto a atividade das invertases são influenciadas por uma
variedade de fatores intra e extracelulares (TYMOWSKA-LALANNE e KREIS, 1998), como
estímulos ambientais, estímulos hormonais e fases fenológicas (ROITSCH et al., 2003). Para
Pommer e Passos (2009) todas as formas de produtos de videira têm origem nos açúcares
produzidos nas folhas, através da fotossíntese e transportados tanto para os frutos, na época de
produção, como para troncos, raízes e folhas não expandidas na fase vegetativa.
Dantas et al. (2007a), avaliando a influência da posição do ramo e da
variação sazonal no teor de açúcares solúveis e insolúveis nas folhas da videira cultivar Syrah
definiram que os açúcares são produzidos em maior quantidade a partir da quinta folha
expandida e acumulados em folhas próximas aos cachos, principalmente, durante o início da
maturação, porém é fortemente influenciado pelas variações climáticas durante o ciclo.
As mudanças mais nítidas na composição das bagas acontecem durante
a segunda fase de crescimento ou fase de maturação. As bagas passam de um estado onde elas
são pequenas, duras, ácidas e com pouco açúcar para um estado onde elas são maiores,
macias, doces, menos ácidas e fortemente saborosas e coloridas (CONDE et al., 2007). O
sabor que se forma nos frutos é, principalmente, resultado do equilíbrio entre ácidos/açúcares
e a síntese de sabor, compostos aromáticos ou precursores ocorre neste momento. O
desenvolvimento destas características determinará em grande parte a qualidade do produto
final (BOSS e DAVIES, 2001).
A temperatura também exerce influência sobre a composição da uva:
quanto mais elevada for a temperatura da região de cultivo, dentro dos limites críticos, maior
será a concentração de açúcar e menor de ácido málico nos frutos, favorecendo as produções
de uva de mesa, passas e vinhos doces (COMBE et al., 1987).

36
O critério mais utilizado para definir a época correta da colheita é o
teor de açúcares, medidos em escala de graus Brix, que representa o teor de sólidos solúveis na
amostra, 90% dos quais são açúcares (GUERRA, 2002).
Chaves (1986) relata que os frutos de videira com a porção interior do
dossel bem exposta à luz solar tiveram aumento na concentração de açúcares, antocianinas e
fenólicos totais e decréscimo no nível de ácido málico, potássio e pH, quando comparados aos
frutos produzidos por videiras com pouca exposição da parte interior do dossel.
2.5.2 Ácidos Orgânicos
Os açúcares e os ácidos são os principais componentes da fração de
sólidos são conhecidos por serem os fatores mais importantes associados ao sabor da fruta. A
determinação da relação açúcares/ácidos é que melhor define o grau de maturação das uvas
(CARVALHO e CHITARRA, 1984). Os ácidos orgânicos, tartárico, málico e cítrico são os
principais responsáveis pelo sabor ácido da uva. Os ácidos tartárico e málico estão presentes
em maior quantidade dentro do vacúolo das células das bagas e são responsáveis por mais de
90% da acidez total. O acúmulo destes ácidos se dá durante o período de crescimento da uva,
por translocação das partes vegetativas ou por síntese nas próprias bagas (POSSNER e
KLIEWER, 1985). Os ácidos málico e tartárico se sobressaem ao cítrico, podendo variar em
função da cultivar. Formam-se tanto nas bagas verdes como nas folhas (GIOVANNINI, 2008).
O pH do mosto depende do tipo e da concentração dos ácidos orgânicos e da concentração de
cátions, especialmente de potássio (CHAMPAGNOL, 1986). O aumento gradual do pH
durante a maturação reflete a formação de sais ácidos às custas do ácido livre. A relação entre
sais ácidos e ácido livre é influenciada pela quantidade total de calor efetivo durante a
maturação (ELIAS, 2008).

37
2.5.3 Compostos Nitrogenados
O nitrogênio (N) é considerado nutriente essencial e participa da
constituição do vegetal, formando os aminoácidos, proteínas, enzimas, ácidos nucléicos,
clorofila, hormônios (TAIZ e ZEIGER, 2004), ATP, NADH e NADPH (BREDEMEIER e
MUNDSTOCK, 2000). Na videira, a disponibilidade de nitrogênio é quase sempre fator
limitante, influenciando o crescimento da planta mais do que qualquer outro nutriente.
O nitrogênio é encontrado no solo nas formas orgânica (proteínas,
aminoácidos, entre outras) e inorgânica (NH4+ e NO3
-). As principais formas absorvidas pelos
vegetais são NH4+ e NO3
-. No caso da videira, quase todo o nitrogênio é absorvido e
transportado até as folhas na forma de nitrato (NO3-), onde sofre redução para nitrito (NO2
-) e,
em seguida, para amônio (NH4+), na presença da enzima redutase do nitrato. A conversão do
nitrato a amônio ocorre em duas etapas e envolve oito elétrons. Na primeira ocorre a redução
do nitrato a nitrito pela enzima nitrato redutase (NR), com o consumo de dois elétrons e na
segunda etapa, o nitrito é convertido para amônia, pela nitrito redutase, com gasto de seis
elétrons (LEA, 1993).
Segundo vários autores, a nitrato redutase é extremamente sensível ao
estresse hídrico (MATOS et al., 2001), podendo provocar uma completa modificação do
metabolismo do nitrogênio nas videiras se não houver manejo adequado da irrigação e teor de
água na planta. A regulação da atividade da NR e, assim, de toda a via de assimilação de N,
durante o estresse hídrico, é muito complexa, pois envolve diversos fatores endógenos diretos
e indiretos, como a fotossíntese e seus produtos, incluindo alguns aminoácidos. A deficiência
de água no período reprodutivo da planta altera, portanto, todo o metabolismo da planta,
induzindo-a a modificar a distribuição de seu crescimento e morfologia, bem como a
composição do fruto na colheita (MATTHEWS e ANDERSON, 1988).
O nitrogênio participa de processos metabólicos e da regulação de
muitos processos fisiológicos da videira. Morinaga et al. (2003) observaram que a partição de
N e de fotoassimilados entre as raízes e os frutos foi influenciada pela maior carga de frutos
em videiras (Vitis labrusca var.Aki-queen), com 5 anos de idade. Uma maior carga de frutos
força a videira a acumular menos açúcares nas raízes e na parte aérea. Isto significa que o fruto
tem prioridade no acúmulo de fotoassimilados e na partição de N. Para Bates et al. (2002) as

38
raízes da videira são o maior reservatório de N total da planta e que este acúmulo ocorre entre
o período de florescimento e a colheita.
Os compostos nitrogenados assumem diversos papéis importantes no
que se refere à composição e qualidade do vinho. A composição de aminoácidos é de grande
importância na produção de vinhos, visto que age como uma fonte de nitrogênio para as
leveduras durante a fermentação. E é influenciada por diferentes fatores, como disponibilidade
de água para o transporte destas substâncias, a cultivar, condições climáticas e a fertilização
(SPONHOLZ, 1991). Do ponto de vista quantitativo, os principais aminoácidos presentes no
mosto e no vinho são: arginina, alanina, treonina, serina e ácido glutâmico e aspártico. Sua
quantificação proporciona uma indicação dos possíveis problemas de fermentação, assim
como a necessidade de suplementação com nitrogênio. O conteúdo total de nitrogênio no
mosto de uvas varia entre 60 e 2.400 mg/L (ZOECKLEIN et al., 2001).
Videiras deficientes em nitrogênio podem apresentar produção
reduzida, baixo conteúdo de nitrogênio total no suco e baixa sanidade. Baixos níveis de
nitrogênio total podem causar problemas durante o processo de fabricação do vinho,
provocando parada na fermentação ou fermentação prolongada devido ao inadequado
suprimento de nitrogênio para as leveduras. Por outro lado o excesso de nitrogênio nas
videiras torna-as muito vigorosas, prolongando o período de crescimento vegetativo e
retardando o amadurecimento da fruta (DAUDT et al., 1995).
No vinho, as quantidades de nitrogênio total são menores em relação
ao mosto de origem. Esta redução, muito significante no primeiro estádio de fermentação
ocorre devido à utilização do nitrogênio pelas leveduras, que o utilizam para sua multiplicação
e posterior fermentação (GARCIA e DAUDT, 1988).
2.6 Características do Fruto para produção de Vinho
Um cacho de uva maduro e sadio apresenta de 2% a 5% de engaço e
de 95% a 98% de baga ou grão. Estas variações podem ocorrer com o tipo de variedade da uva
e com o estado de sanidade da mesma (AQUARONE et al., 2001). A baga é formada de 6% a
12% de casca ou película, de 2% a 5% de sementes e de 85% a 92% de polpa. Na parte

39
exterior ficam localizadas as substâncias aromáticas características de cada variedade; as
antocianinas estão presentes na casca (contudo, em uvas tintureiras essas substâncias estão
presentes também na polpa) e os taninos de 0,5% a 2%, dependendo de cada variedade
(RIBÉREAU-GAYON e PEYNAUD, 1982). A polpa ou mosto constitui a parte principal da
baga. Os principais constituintes são (em g L-1
): 650 a 850 de água; 120 a 250 de açúcares
redutores (carboidratos); 6 a 14 de ácidos orgânicos; 2,5 a 3,5 de substâncias minerais; 0,5 a
1,0 de compostos nitrogenados e outros componentes que aparecem em quantidades mínimas
(AQUARONE et al., 2001).
O estado hídrico da planta influi diretamente no tamanho dos frutos
durante os estádios entre florada e início do pintor. A restrição hídrica modifica o crescimento
da baga (alongamento celular), diminuindo o volume celular (OJEDA et al., 2001). Isso pode
promover maior e melhor relação película/polpa. Os frutos sujeitos à carência hídrica
moderada apresentam maior quantidade dos compostos que estão presentes na película (tais
como taninos e antocianinas), estando estes, geralmente, associados a um melhor potencial
qualitativo (ROBY e MATTHEWS, 2004; MATTHEWS e KRIEDEMANN, 2006). O défice
hídrico moderado no período de maturação favorece o acúmulo de compostos fenólicos e
açúcares, além de proporcionar cachos mais soltos, aerados e sadios, sendo fatores de grande
importância para a elaboração de vinhos de qualidade (DELOIRE et al., 2004).

40
3 MATERIAL E MÉTODOS
3.1 Caracterização da Área Experimental
O estudo de campo foi desenvolvido na Embrapa Semiárido,
especificamente no Campo Experimental de Bebedouro, localizado no município de Petrolina-
PE (latitude 9º09’ Sul, longitude 40º22’ Oeste, e altitude média de 365,5 m), cujo clima
segundo Köeppen pode ser classificado como tipo BSh, que corresponde a uma região
semiárida muito quente. O solo da área em questão é classificado pelo Sistema Brasileiro de
Classificação de Solos como Argissolo Vermelho Amarelo Eutrófico Latossólico, textura
média (SILVA, 2005). As análises laboratoriais foram desenvolvidas no Laboratório de
Sementes e Fisiologia Vegetal da Embrapa Semiárido.

41
3.2 Caracterização do Experimento
A videira (Vitis vinifera L.) cultivar Syrah foi enxertada sobre o porta-
enxerto 1103 Paulsen, sendo que as mudas do porta-enxerto foram obtidas por meio de
estaquia. A implantação do vinhedo foi realizada em 13/03/2009 no espaçamento de 1 m entre
plantas e 3 m entre fileiras, ocupando uma área útil de 81 m2 e a condução das plantas foi feita
no sistema de espaldeira (com três fios de arame) em cordão bilateral e a poda de produção
adotada foi em esporão, deixando-se duas gemas por esporão e cinco a seis esporões por
planta. O sistema de irrigação utilizado foi o de gotejamento com emissores espaçados em 0,5
m na linha de plantas. Foram avaliadas três estratégias de irrigação durante o crescimento e
desenvolvimento das plantas a partir da fase fenológica de cacho fechado (‘L’) após a poda de
produção.
O manejo da área desde o plantio à colheita da primeira safra foi
realizado através de adubações, baseadas na análise de solo e foliar, capinas, amarrios de
ramos, controle de pragas e doenças, desbrota e desponte dos ramos, a uma altura de 20 cm
acima do último fio de arame.
Em 13/04/2010 foi feita a 1ª poda de produção das plantas e as fases
fenológicas foram acompanhadas de acordo com a escala de Baillod e Baggiolini (1993). A
partir da iniciação da fase fenológica ‘L’ em 28 de maio de 2010 (45 dias após a poda de
produção-DAP), teve início a aplicação dos tratamentos de irrigação: 1- Irrigação Plena (IP),
sem restrição de água às videiras durante todo o ciclo de produção, sendo a irrigação feita para
repor a lâmina correspondente à evapotranspiração da cultura (ETc); 2 - Irrigação Deficitária
(ID), onde a aplicação de água foi interrompida desde a fase fenológica de cacho fechado até a
colheita em 6 de agosto de 2010 (115 dias após a poda), na fase fenológica de cacho maduro;
3 - Irrigação com Défice Controlado (IDC), onde a aplicação de água foi interrompida desde a
fase fenológica de cacho fechado, mas a irrigação foi realizada aos 70, 71, 87, 90 e 111 dias,
respectivamente após a poda, para o aumento da umidade do solo na profundidade efetiva do
sistema radicular (0,6 m), de acordo com Silva (2005). O momento da irrigação foi decidido
conforme o monitoramento semanal da umidade do solo pela técnica de moderação de
nêutrons (sonda de nêutrons).

42
O manejo de irrigação foi realizado com base na evapotranspiração de
referência (ETo, mm) e em coeficientes de cultura (Kc) para videira de vinho cv. Syrah
(SILVA, 2005; BASSOI et al., 2007). Assim, o manejo de irrigação foi feito baseado na
estimativa da ETc (mm) obtida pelo produto ETo x Kc, para os diferentes estádios fenológicos
da cultura da videira. Registrou-se também, ao longo do ciclo da videira cv. Syrah na estação
meteorológica do campo experimental de Bebedouro, os dados climáticos de temperatura,
umidade, insolação, radiação solar global e precipitação (Figuras 1 e 2).
O monitoramento da dinâmica da água (Figura 3) do solo foi feito por
meio da técnica de moderação de nêutrons antes e/ou depois das irrigações, nas profundidades
de 0,15; 0,30; 0,45 e 0,60 m. Foram instalados 12 tubos de alumínio para acesso da sonda de
nêutrons, CPN Hydroprobe 503, em todos os tratamentos de irrigação.
O delineamento estatístico foi em blocos casualizados, em três
esquemas fatoriais: 3 x 5 para potencial hídrico foliar de base (Ψb), teor SPAD da clorofila e
atividade da enzima nitrato redutase (NR); 3 x 6 para trocas gasosas, açúcares solúveis totais,
açúcares redutores, proteína solúvel total e atividade das invertases; e 3 x 3 para as análises
químicas: sólidos solúveis, acidez titulável, relação sólidos solúveis / acidez titulável e pH,
sendo nove repetições para cada tratamento. Foram utilizadas três estratégias de irrigação (IP,
IDC e ID) e foram realizadas cinco (54, 60, 73, 87 e 101 DAP para o teor SPAD da clorofila e
NR; e 56, 70, 86, 99 e 113 DAP para o Ψb), seis (54, 60, 73, 87, 101 e 115 DAP) e três (98,
105 e 112 DAP) coletas ao longo do ciclo. Realizou-se a análise de variância e o teste F para
todas as medidas, comparando-se as médias pelo Tukey a 5% de probabilidade. As datas das
avaliações realizadas entre 54 e 112 DAP compreenderam o intervalo das fases fenológicas de
cacho fechado a cacho maduro (Figura 4).

43
Figura 1. Temperatura (A), insolação (B), umidade (C) e radiação solar global (D) durante o
ciclo da videira cv. Syrah. Embrapa Semiárido, Petrolina-PE, 2010.
Figura 2. Precipitação pluviométrica (mm) registrada durante o ciclo de produção da videira
cv. Syrah. Embrapa Semiárido, Petrolina-PE, 2010. Fonte: Goncalves (2011).
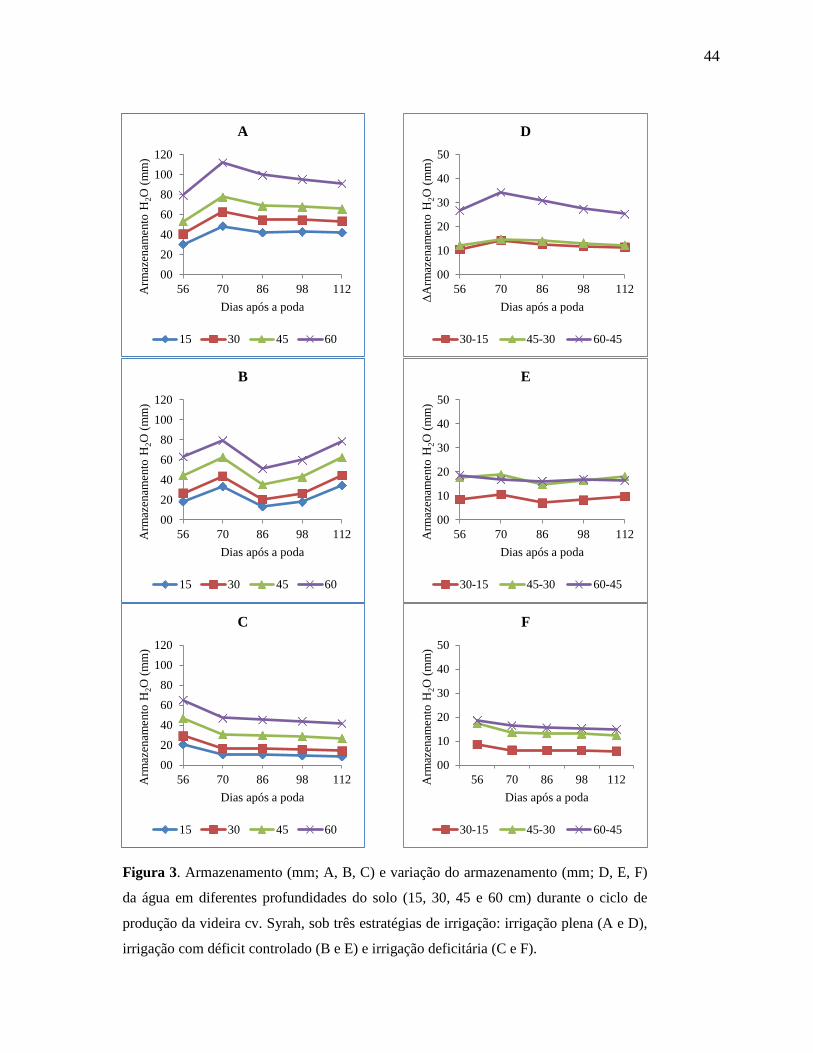
44
00
20
40
60
80
100
120
56 70 86 98 112Arm
azen
amento
H2O
(m
m)
Dias após a poda
A
15 30 45 60
00
20
40
60
80
100
120
56 70 86 98 112Arm
azen
amento
H2O
(m
m)
Dias após a poda
B
15 30 45 60
00
20
40
60
80
100
120
56 70 86 98 112Arm
azen
amento
H2O
(m
m)
Dias após a poda
C
15 30 45 60
00
10
20
30
40
50
56 70 86 98 112
ΔA
rmaz
enam
ento
H2O
(m
m)
Dias após a poda
D
30-15 45-30 60-45
00
10
20
30
40
50
56 70 86 98 112Arm
azen
amento
H2O
(m
m)
Dias após a poda
E
30-15 45-30 60-45
00
10
20
30
40
50
56 70 86 98 112Arm
azen
amento
H2O
(m
m)
Dias após a poda
F
30-15 45-30 60-45
Figura 3. Armazenamento (mm; A, B, C) e variação do armazenamento (mm; D, E, F)
da água em diferentes profundidades do solo (15, 30, 45 e 60 cm) durante o ciclo de
produção da videira cv. Syrah, sob três estratégias de irrigação: irrigação plena (A e D),
irrigação com déficit controlado (B e E) e irrigação deficitária (C e F).

45
Figura 4. Escala de Baggiolini para determinação das fases fenológicas da videira.

46
3.3 Características Avaliadas
3.3.1 Determinação do Potencial Hídrico Foliar de Base (Ψb)
O potencial hídrico foliar de base foi avaliado entre 3:00 e 4:30 horas
da madrugada, no período compreendido entre as fases fenológicas de cacho fechado e cacho
maduro. Foram coletadas amostras de folhas na porção mediana de ramos produtivos (três
folhas por planta) de diferentes plantas e utilizadas imediatamente após a coleta para a
medição do potencial hídrico foliar de base (Ψb, MPa), utilizando câmara de pressão de
Scholander (PMS InstrumentCo, model 1000) (SCHOLANDER et al., 1965) com nitrogênio
como gás inerte. As amostras foram coletadas, considerando-se os tratamentos: irrigação plena
(IP), irrigação deficitária (ID) e irrigação com défice controlado (IDC), aos 56, 70, 86, 99 e
113 dias, respectivamente, após a poda de produção, compreendendo o intervalo entre as fases
fenológicas de cacho fechado e cacho maduro.
3.3.2 Trocas gasosas
A taxa fotossintética líquida (mol m-2
s-1
) e a condutância estomática
(mol m-2
s-1
) foram determinadas pelo sistema portátil de medição das trocas gasosas modelo
LI-6400 XT (Licor Inc., NE, USA), analisador portátil de CO2 por infravermelho, em
diferentes dias, considerando as diferentes fases de crescimento traçando-se um gráfico
fotossintético de desempenho dos tratamentos nas condições edafoclimáticas e ecofisiológicas
estabelecidas. Em cada série foram mensuradas folhas adultas, sadias, situadas na porção
mediana do sarmento e exposta à densidade de fluxo de fótons fotossinteticamente ativos
saturante (> 1000 mmol m-2
s-1
). Avaliou-se uma planta por parcela, totalizando nove plantas
por tratamento com três repetições. As determinações de assimilação líquida de CO2 (A),
condutância estomática (gs), transpiração (E), concentração intracelular de CO2 (Ci), foram
feitas das 08:00 às 10:00 horas da manhã. A relação entre a quantidade de CO2 assimilado por
unidade de água perdida pela transpiração foi obtida a eficiência no uso da água (EUA) e da

47
relação entre a quantidade de CO2 assimilado por concentração interna de CO2 foi obtida a
eficiência de carboxilação in vivo da Ribulose-1,5-difosfato carboxilase/oxigenase (Rubisco).
3.3.3 Índice SPAD de clorofila
O índice SPAD foi determinado por aparelho portátil denominado
clorofilômetro, Chlorophyll Meter, modelo SPAD-502 (Soil and Plant Analysis Development)
que realiza medida instantânea e não destrutiva da folha, dando um valor de absorbância do
comprimento de onda na região do vermelho (pico em 650 nm), região de alta absorbância
pelas moléculas de clorofila. O aparelho possui dois LEDs (diodo emissor de luz)
posicionados na ponta do medidor, que emitem luz na faixa de 600 a 700 nm e na faixa de 860
a 1060 nm, em sequência, quando está fechado (GODOY, 2002). A luz passa pela janela de
emissão, sendo que a parte que atravessa a folha atinge um receptor (fotodiodo de silicone)
sendo convertida em sinais elétricos, amplificados e transformados em sinais digitais, usados
por um microprocessador para calcular os valores SPAD, que são mostrados num visor.
Foram amostradas dez folhas opostas ao cacho e completamente
expandidas para cada tratamento, sendo realizado aos 9, 15, 28, 42 e 56 dias após o início das
estratégias de irrigação, com os resultados expressos em Unidade SPAD.
3.4 Análises Bioquímicas
3.4.1 Coleta e armazenamento do material vegetal
Para a realização das análises bioquímicas de carboidratos: açúcares
solúveis totais e açúcares redutores em cada coleta (9, 15, 27, 42, 56 e 70 DAT (dias após o
início dos tratamentos)) para os três tratamentos (IP, IDC e ID), coletou-se sempre a folha
oposta ao cacho, pois segundo Schultz et al.,1996 esta possui alta capacidade fotossintética,

48
cujos teores de açúcares solúveis e insolúveis são produzidos em grande quantidade. Após a
coleta das folhas foi preparado o extrato, onde pesou-se 1 grama da amostra, colocou-se 5mL
de álcool etílico 80% e centrifugou-se a 3000 x g por 10 minutos, sendo repetido três vezes
este processo; em seguida, coletou-se o sobrenadante que foi congelado em freezer a -20ºC,
para posterior utilização para determinação de açúcares solúveis totais, açúcares redutores e
proteína solúvel total.
3.4.2 Açúcares solúveis totais
A quantificação dos teores de açúcares solúveis totais em μg g-1
de
matéria fresca (MF) foi realizada durante o ciclo por espectrofotometria no comprimento de
onda de 620 nm, utilizando-se curva padrão de glicose conforme metodologia descrita por
Morris (1948) e Yemm e Willis (1954), onde o reagente de antrona provocou reação
hidrolítica e desidratante sobre os carboidratos presentes nas folhas amostradas. As ligações
glicosídicas foram hidrolisadas e os açúcares simples desidratados formando furfural ou
hidroximetilfurfural. Essas substâncias se condensaram com a antrona (9,10-dihidro-9-
oxoantraceno) dando um produto de coloração azul petróleo (SILVA et al., 2003).
3.4.3 Açúcares redutores
Os açúcares redutores foram quantificados por espectrofotometria no
comprimento de onda de 540nm, utilizando-se curva padrão de glicose e pelo método do ácido
3-5-dinitrossalicílico (DNS) proposto por Miller (1959). Os resultados obtidos foram
expressos em μg g-1
MF.
3.4.4 Determinação do teor de proteína solúvel total

49
A determinação do teor de proteína solúvel total foi realizada de
acordo com o método proposto por Bradford (1976) utilizando o mesmo extrato obtido para a
determinação dos açúcares. Como curva padrão utilizou-se a caseína (1 mg mL-1
) e a leitura
foi feita por espectrofotometria no comprimento de 595 nm, sendo o teor de proteínas
expresso em μg g-1
MF.
3.4.5 Atividade das invertases (EC 3.2.1.26)
As atividades da invertase ácida do vacúolo (IAV), invertase ácida da
parede celular (IAP) e invertase neutra do citossol (INC) foram realizadas de acordo com
Nascimento et al. (1998) a partir da fase fenológica ‘L’, cacho fechado (BAILLOD e
BAGGIOLINI, 1993) durante o ciclo produtivo das plantas. Para as análises da atividade das
invertases foram coletadas folhas opostas ao cacho em diferentes plantas entre 8:00 e 9:00
horas da manhã, acondicionadas em sacos plásticos previamente identificados, armazenadas
em isopor, contendo gelo e levadas para o laboratório de Sementes e Fisiologia Vegetal da
Embrapa Semiárido, sendo armazenadas em freezer a -20oC até os ensaios enzimáticos. As
folhas congeladas foram maceradas em tampão de extração constituído de tampão fosfato de
potássio, 50 mmol L-1
, pH 7,0; 0,7% de mercaptoetanol e 6% de MnSO4 5mmol L-1
, na razão
de 1,0 g de material vegetal por 10 mL de tampão de extração, centrifugando a 15500 x g
durante 15 minutos. Coletou-se o sobrenadante para determinação da atividade da INC e IAV.
O precipitado resultante foi ressuspendido em 5mL da solução de extração, centrifugado
novamente e coletado o sobrenadante para determinação da atividade da IAP. Após a coleta
dos extratos enzimáticos, os mesmos foram congelados até a realização das análises. Os
ensaios das invertases foram realizados em meio de reação, constituído de 0,5 mL de extrato
enzimático; 2,5 mL de tampão fosfato de potássio 50 mmol L-1
, pH 7,0 para INC ou tampão
citrato de sódio 50 mmol L-1
, pH 5,0 para IAP e IAV e de 1,0 mL de sacarose 100 mmol L-1
,
como substrato. O meio de reação foi mantido durante 30 minutos em banho-maria a 35oC.
Foram retiradas alíquotas de 0,5mL para a determinação da atividade enzimática ao final de
cada tempo de reação (tempo 0= 10’ e tempo 1= 30’ após). Ao final do ensaio foi quantificado
o teor de açúcares redutores (AR) do meio de reação, pelo método do DNS. A atividade de

50
IAP, IAV e INC foi expressa pelo teor de AR em µmol, produzido pela hidrólise da sacarose,
em 1 g de folha por hora (µmol g-1
MF h-1
).
3.4.6 Atividade da enzima Nitrato Redutase (NR) (EC 1.6.6.3) ‘in vivo’
A atividade da NR in vivo foi estimada utilizando o método descrito
por Radin (1973) com modificações. Foram efetuadas coletas de folhas opostas ao cacho no
horário entre 8:00 e 9:00h da manhã. Retirou-se amostras de discos foliares (0,5 cm de
diâmetro) com perfurador, perfazendo 200mg de massa e incubou-se em frascos escuros de
plástico com o meio de reação (5mL de solução tampão fosfato (50mM), pH 7,5 + 1mL de
KNO3 (500mM) + 1mL de n-propanol 1% (v/v)). Em seguida, as amostras foram submetidas à
infiltração à vácuo durante 2 minutos com posterior transferência para banho-maria a 30oC,
sob agitação e ausência de luz. Após 1 hora retirou-se alíquotas de 1mL do meio de reação e
pipetou-se em tubos de ensaio com: 1mL de sulfanilamida (1% em HCl 1,5N), 1 mL de n-
naftil etileno diamina 0,02% (m/v) e 2 mL de tampão fosfato (50mM), pH 7,5. Agitou-se os
tubos e após 30 minutos efetuou-se as leituras de absorbâncias a 540nm. A atividade da NR
foi expressa em μmol de NO2 g MF h-1
.
3.5 Análises da qualidade dos frutos
Na colheita, em quatro plantas escolhidas aleatoriamente por estratégia
hídrica, foram coletadas 100 bagas do engaço e em diferentes posições do cacho (parte
inferior, mediana e basal). As bagas foram acondicionadas em sacos plásticos previamente
identificados, armazenadas em isopor, contendo gelo e levadas para o Laboratório de Enologia
da Embrapa Semiárido para análise. Em seguida foram pesadas em balança analítica digital e
colocadas em proveta graduada de 500 mL. As provetas foram aferidas com 300 mL de água.
A variação do volume de água foi considerada como sendo o volume de 100 bagas. Esta
metodologia foi aplicada em cada tratamento. Após a maceração das bagas, foi possível a
obtenção do volume do mosto das mesmas 100 bagas com proveta graduada. Além disso,

51
foram determinadas no mosto o teor de sólidos solúveis (SS), utilizando-se refratômetro
portátil marca ATAGO modelo Pocket PAL-1, acidez titulável - expressa em g L-1
de ácido
tartárico, conforme metodologia descrita por Pregnolatto e Pregnolatto (1985) e o pH que foi
determinado com pHmetro digital marca TECNAL.

52
4 RESULTADOS E DISCUSSÃO
4.1 Potencial hídrico foliar de base (Ψb)
Na Tabela 1 estão apresentados os dados referentes ao resumo da
análise de variância do potencial hídrico de base da folha (Ψb) aos 56, 70, 86, 99 e 113 dias
após a poda. Os fatores de avaliação: Estratégia de irrigação (EI), dias após a poda (DAP) e a
interação entre os fatores (EI x DAP) demonstraram-se significativos a p < 0,01 pelo teste F.
Isto permite afirmar que nessa característica, pelo menos uma estratégia hídrica foi diferente
estatisticamente, assim como houve variação entre as diferentes datas de coletas, enquanto que
a interação significativa mostra que o valor desta característica foi influenciado pela interação
EI x DAP. Desta forma, fez-se o desdobramento dos dias após a poda dentro de cada estratégia
de irrigação a fim de observar seus efeitos, isoladamente.

53
Tabela 1. Resumo da análise de variância (teste F) para o potencial hídrico foliar de base (Ψb,
MPa) em folhas de videira Syrah aos 56, 70, 86, 99 e 113 dias após a poda (DAP) em função
de três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE, 2010.
FV GL Quadrado Médio
EI 2 9,09**
DAP 4 6,47**
Bloco 2 0,32ns
EI x DAP 8 0,26**
Resíduo 28 0,32
CV (%) - 16,49
ns: não significativo; ** p<0,01.
Desde o desenvolvimento da câmara de pressão (SCHOLANDER et
al., 1965), a medição do potencial hídrico foliar de base tem sido utilizada e defendida por
muitos autores como uma ferramenta para avaliar a condição hídrica das plantas (JONES,
1990; CARBONNEAU, 1998; LOPES et al., 1999; CHONÉ et al., 2001; CIFRE et al., 2005;
SOUZA et el., 2003; 2005; MALHEIRO, 2005; DELOIRE et al., 2005; PATAKAS et al.,
2005; GIRONA et al., 2006; BAEZA et al., 2007). No entanto, outros autores preferem o Ψmd
(potencial do meio-dia), momento em que este atinge seus valores mais negativos
(DONOVAN et al., 2001; GAUDILLERE et al., 2002). No presente estudo foi utilizado
apenas o Ψb para monitorar o estado hídrico da videira.
Analisando os dados do Ψb em função dos tratamentos, não se observa
diferença significativa entre os mesmos aos 56 DAP (Tabela 2). Isso poderia estar relacionado
ao curto período de suspensão da irrigação (11 dias) nos tratamentos com os potenciais mais
negativos. No entanto, os valores de potencial hídrico foliar de base apresentados pelas
diferentes estratégias de irrigação são indicativos de que as plantas estavam sob défice hídrico
suave/leve a moderado/médio (DELOIRE et al., 2006; OJEDA, 2007), com menor intensidade
em valores absolutos no tratamento IP, em função do contínuo uso de lâminas de irrigação.

54
Tabela 2. Potencial hídrico foliar de base (Ψb, MPa) em folhas de videira cv. Syrah durante o
ciclo em função de três estratégias de irrigação (Irrigação com défice controlado-IDC,
Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE, 2010.
Dias após a poda Estratégias de irrigação
IDC ID IP
56 -0,433 Aab -0,399 Aab -0,347 Ab
70 -0,342 Bab -0,325 Ba -0,166 Aa
86 -0,299 Ba -0,300 Ba -0,129 Aa
99 -0,476 ABb -0,515 Bb -0,364 Ab
113 -0,371 ABab -0,422 Bab -0,262 Aab
CV (%) 16,49
Médias seguidas de mesma letra minúsculas na coluna e maiúsculas na linha, não diferem
entre si pelo teste Tukey a 5% de probabilidade.
A redução do conteúdo de água no solo afeta acentuadamente alguns
processos morfofisiológicos, enquanto outros são relativamente insensíveis (SANTOS e
CARLESSO, 1998). A extensão dos efeitos do défice hídrico em espécies vegetais depende da
sua intensidade, duração e da capacidade genética das plantas em responder às mudanças do
ambiente (CHAVES, 1991; CHAVES et al., 2010). Uma das primeiras mudanças metabólicas
que ocorrem nas plantas sob défice hídrico é a sinalização do hormônio vegetal ácido
abscísico (ABA). Segundo Zeevaart e Creelman (1988), o défice hídrico controlado promove
rápida redistribuição e acumulação do ABA nos tecidos da planta, o qual, ao ser transportado
para as folhas provoca o fechamento dos estômatos (LOVEYS, 1984; DAVIES e ZANG,
1991; MOUTINHO-PEREIRA et al., 2004; RODRIGUES et al., 2008) e, consequentemente, a
redução da transpiração, mas sem comprometer a produtividade da videira (LOVEYS et al.,
1998; ). A resposta mais proeminente das plantas ao défice hídrico e/ou sinalizadas pelo ABA
consiste no decréscimo da produção da área foliar, no fechamento dos estômatos, na
aceleração da senescência e na abscisão foliar (MCCREE e FERNÁNDEZ, 1989; TAIZ e
ZEIGER, 2009), bem como, na redução da síntese da parede celular e de membranas, da
divisão celular e síntese protéica (BURSSENS et al., 2000). Além disso, as plantas submetidas

55
ao défice hídrico podem modular o crescimento da raiz, formar barreiras apoplásticas,
compartimentalizar as vias de transporte de água e/ou regular a expressão e atividade das
aquaporinas (JOHANSSON et al., 2000; VERA-ESTRELLA et al., 2004; GALMÉS et al.,
2007; KALDENHOFF et al., 2008; LOVISOLO et al., 2008b; 2010; MAUREL et al., 2008;
VANDELEUR et al., 2009).
Desta forma na primeira avaliação realizada aos 56 DAP sugere-se que
as plantas das estratégias de irrigação IDC e ID já poderiam estar apresentando respostas
morfofisiológicas às condições de défice hídrico impostas pelo ambiente, ou seja, já poderiam
estar sob ação do ABA (Tabela 2).
Aos 70 DAP ocorreu aumento do Ψb em todas as três estratégias
hídricas (IP, IDC e ID) com destaque significativo apenas para a irrigação plena (IP) com o
valor de -0,16 MPa. Essas alterações podem ter sido influenciadas pela precipitação de 8,1
mm ocorrida três dias antes das leituras dos Ψb. Conforme Carbonneau (1998) e Assis e Lima
Filho (2000) consideram que a planta encontra-se sob boas condições hídricas, quando o Ψb
atinge até o valor mínimo de -0,2 MPa. Isso pode ser confirmado também na estratégia de
irrigação IP por ter registrado o maior armazenamento de água no solo (115,7 mm) a 60 cm de
profundidade (Figura 3) após o início dos tratamentos. Isso ocorreu em função da irrigação
fornecida frequentemente, somada à ocorrência de precipitações (26 mm foi a precipitação
acumulada do início dos tratamentos até a colheita).
A irrigação deficitária e a irrigação com défice controlado podem ter
aumentado seus potenciais hídricos foliares de base nas videiras, influenciados também pela
precipitação. No entanto, não se diferenciaram estatisticamente dos valores da leitura anterior.
Contudo, os valores lidos aos 70 e 86 DAP para esses dois tratamentos (Tabela 2) revelam
uma tendência crescente de seus Ψb, mas de acordo com seus valores, os mesmos continuam
provocando nas plantas características de défice suave, o que ainda assim, poderia resultar
também em sinalização química promovida pelo ABA, o qual desencadeia uma série de
reações e mecanismos morfofisiológicos de adaptação na videira. Estes podem ser ativados
e/ou desativados toda vez que as plantas entram ou saem do défice hídrico. Por outro lado,
aos 99 DAP os Ψb das plantas sob as três estratégias de irrigação, IDC, ID e IP tiveram queda
acentuada, alcançando valores da ordem de -0,47; -0,51 e -0,36 MPa, respectivamente,

56
enquadrando-se em estresse moderado a forte segundo Hsiao (1973) e Deloire et al. (2004;
2006).
Segundo Schultz (2003), as videiras apresentam diferentes níveis de
adaptação ao défice hídrico, dividindo-se em duas classes fisiológicas: isohídricas e
anisohídricas. As espécies e/ou variedades com comportamento isohídrico são capazes de
manter o controle rigoroso do potencial hídrico da planta sob uma gama de condições
ambientais, enquanto o potencial hídrico em plantas anisohídricas oscila em resposta às
condições ambientais (TARDIEU e SIMONNEAU, 1998). A videira ‘Syrah’, utilizada no
presente estudo, apresenta características anisohídricas (SCHULTZ, 2003), tendo
comportamento mais tolerante à manutenção da abertura estomática em condições
estressantes. A regulação estomática é a principal diferença entre as classes referidas,
atribuindo-se, geralmente, ao papel regulador ao ABA (LOVEYS, 1984). Além disso, a
condutividade hidráulica nos vasos xilemáticos também é afetada com a carência hídrica,
levando à diminuição do diâmetro dos vasos do xilema, uma vez que vasos menores, além de
reduzirem por si só as perdas de água, evitam também o processo de cavitação (LOVISOLO e
SHUBERT, 1998; ZUFFEREY et al., 2011).
Ao nível celular, em plantas sob défice hídrico, ocorre a expressão
gênica da família das aquaporinas, que desempenham importante papel no aumento da
permeabilidade celular (CHAVES et al., 2003; FOUQUET et al., 2008), resultante da
acumulação de ABA, representando uma forma de adaptação ao estresse. Vandeleur et al.
(2009) estudaram o envolvimento das aquaporinas na redução da condutividade hidráulica em
raízes de videira como forma de limitar as perdas de água em resposta ao défice hídrico,
evidenciando os diferentes níveis de variação diurna da permeabilidade das membranas em
função da variedade. As variedades anisohídricas quando sujeitas ao défice hídrico registraram
redução menos acentuada da condutividade hidráulica, apresentando maior expressão dos
genes responsáveis pela atividade das aquaporinas, em comparação com variedades
isohídricas.
Outro mecanismo celular de adaptação ao défice hídrico pelas videiras
é o ajustamento osmótico, que consiste num aumento de solutos nas células, diminuindo o
potencial osmótico e permitindo a manutenção da turgescência em condições de deficiência
hídrica (PATAKAS e NOITSAKIS, 1999).

57
Em videira o ajustamento osmótico ocorre por acúmulo de íons
inorgânicos (K+, Ca
2+, NO3
-), apresentando vantagem de baixo custo energético em relação à
acumulação de moléculas orgânicas, cuja síntese celular consome carbono que poderia ser
usado para o crescimento (PATAKAS et al., 2002). A manutenção da turgescência em
condições de deficiência hídrica pode também ser conseguida por alterações estruturais nas
paredes celulares das raízes (MOUTINHO-PEREIRA, 2000), aumentando sua rigidez. A
absorção de água e minerais pelas raízes e o seu envolvimento na regulação estomática fazem
do porta-enxerto o componente fundamental na definição do estado hídrico da videira, com
impacto não apenas nas relações hídricas da planta, mas também em outros processos
fundamentais como a fotossíntese, o crescimento da parte aérea e a produção (IACONO et al.,
1998; PARANYCHIANAKIS et al., 2004).
Para Chaves et al. (2010) a classificação das variedades de videira
como iso ou anisohídricas pode ser inadequado, pois as respostas dos estômatos para o défice
hídrico em uma variedade específica irá variar de acordo com a combinação particular com o
porta-enxerto, o clima (DPV e temperatura) e a intensidade e duração do défice hídrico. As
plantas sob défice hídrico prolongado podem desenvolver paredes celulares mais rígidas,
levando ao maior declínio no potencial hídrico ao meio-dia, característica de resposta
anisohídrica. Além disso, o ajuste osmótico pode contribuir para a manutenção dos estômatos
abertos em menores potenciais hídricos, permitindo melhor turgescência em resposta ao défice
hídrico imposto lentamente. Esta combinação de respostas interagirá com fatores estruturais
do enxerto, tais como a capacidade de condução hídrica de ramos e pecíolos.
Ainda aos 86 DAP as folhas analisadas atingiram os maiores valores
de Ψb analisados durante o ciclo, sendo -0,29; -0,30 e -0,12 MPa para IDC, ID e IP,
respectivamente. O tratamento IP apresentou maior Ψb, devido à ocorrência de precipitações e
contribuição da sua frequência de irrigação, enquanto os demais tratamentos, IDC e ID,
tiveram seus Ψb aumentados, provavelmente, em função da precipitação ocorrida quatro dias
antes das leituras, juntamente com possíveis mecanismos de adaptação morfofisiológica já,
previamente, ativados pelo ABA.
Aos 99 DAP a alteração significativa do Ψb na estratégia de irrigação
IP pode ter ocorrido em função do ajuste na lâmina de irrigação oferecida às plantas,
reduzindo-a em função dos frutos terem atingido a fase de maturação, com modificação do Kc

58
devido às exigências da fase fenológica da cultura, reduzindo a disponibilidade de água para
as plantas nessa fase. As plantas dos tratamentos com restrição hídrica nessa avaliação
apresentaram os mais negativos potenciais foliares de base, sem indicação visual de
murchamento e/ou comprometimento do aparato fotossintético, indicando assim, adaptação ao
estresse submetido. Esses resultados confirmam também a resistência do porta-enxerto
‘Paulsen 1103’ à seca (CARBONNEAU, 1985) e o comportamento anisohídrico da cultivar
copa Syrah (SCHULTZ, 2003; CHALMERS, 2007; ROGIERS et al., 2009).
Durante a noite com o fechamento estomático, o potencial hídrico das
plantas tende a se equilibrar com o potencial hídrico do solo. No entanto, há crescentes
evidências de que nem sempre os estômatos são fechados à noite e que a transpiração noturna
(En) pode afetar as relações hídricas das plantas durante o dia. A condutância noturna (gn) é
significativa em uma variada gama de espécies C3 e C4, incluindo os cultivos comerciais
(CAIRD et al., 2007; DAWSON et al., 2007; KAVANAGH et al., 2007) e a En pode exceder
em 30% os valores diurnos (CAIRD et al., 2007; DAWSON et al., 2007). Além disso, gn e En
podem contribuir para a vulnerabilidade das plantas ao défice hídrico. A En pode resultar
ainda em um incompleto equilíbrio ao amanhecer entre o potencial hídrico das plantas e do
solo (DONOVAN et al., 2001; KAVANAGH et al., 2007). Plantas anisohídricas,
normalmente, exibem menor controle estomatal sobre a demanda evaporativa e a umidade do
solo, permitindo maior flutuação no potencial hídrico foliar (FRANKS et al., 2007).
A relevância ecológica da En pode incluir maior absorção de nutrientes
(DONOVAN et al., 2001) ou a liberação do oxigênio dissolvido no tecido lenhoso (DAWSON
et al., 2007). Rogiers et al. (2009) estudaram a contribuição da En no comportamento
anisohídrico da variedade Semillon em relação a outras cultivares, concluindo que a
transpiração noturna contribuiu significativamente para os baixos valores de potencial hídrico
foliar encontrados na Semillon durante noites quentes e secas.
Desta forma, apesar de não ter sido medida a En, acredita-se que a
leitura dos potenciais hídricos foliares dos tratamentos ID e IDC aos 99 DAP possa ter sofrido
também influência da mesma.
Marinho et al. (2011) estudaram o potencial de água no solo e na folha
da videira variedade Sugarone sob défice hídrico no Submédio do Vale do São Francisco e
encontraram variações no potencial hídrico foliar de base (-0,10 a -0,27 MPa) na fase de

59
maturação das bagas, indicando que as plantas passaram por estresse suave a moderado e
concluíram que o uso do défice hídrico na região é uma estratégia sem muitos impactos nas
condições hídricas das videiras e com economia de água.
A última avaliação feita aos 113 DAP apresentou aumento no valor do
potencial hídrico foliar de base em todos os tratamentos. No entanto, só ocorreu diferença
significativa apenas entre as estratégias hídricas IP e ID. A estratégia de irrigação IP, com
regular disponibilidade hídrica durante todo o ciclo, pode ter tido acréscimo no valor do seu
potencial hídrico de base aos 113 DAP em função de uma modificação na lâmina de irrigação,
realizada aos 104 DAP para compensar a elevação da evapotranspiração da cultura (Etc). Essa
modificação pode ter contribuído também na manutenção de maior valor no potencial hídrico
de base nas videiras submetidas a esse tratamento. O tratamento IDC recebeu correção hídrica
dois dias antes das leituras dos potenciais, no entanto, não foi suficiente para diferenciá-lo do
tratamento com maior restrição hídrica (ID). A irrigação deficitária, mesmo sem suprimento
hídrico artificial (irrigação) entre as avaliações, não se diferenciou estatisticamente do
tratamento IDC. Assim, é possível que nas plantas sob esse tratamento possam ter ocorrido
alterações morfofisiológicas como aumento do seu sistema radicular, juntamente, com o
ajustamento osmótico e aumento da rigidez da parede celular das raízes, o que facilitaria a
absorção de água em maiores profundidades e tensões no solo, considerando que o porta-
enxerto ‘Paulsen 1103’ é resistente à seca (CARBONNEAU, 1985). Todavia, consegue
manter o potencial hídrico mais elevado em algumas cultivares copa de videira quando sob
deficiência hídrica no solo, como registrado por Souza et al. (2001) e no presente estudo. As
alterações citadas acima seriam alguns dos mecanismos de resistência e/ou tolerância à
deficiência hídrica. Essas respostas seriam sinalizadas pelo regulador vegetal ácido abscísico
que teria se acumulado nas folhas (KLIEWER, 1981), induzindo o fechamento estomático,
mas mantendo a absorção de água pelas raízes (TAIZ e ZEIGER, 2009).
A irrigação plena manteve o estado hídrico ótimo das plantas durante
todo o ciclo, para o Ψb variando de -0,12 a -0,36 MPa, valores estes muito abaixo das
condições consideradas estressantes (abaixo de -0,5 MPa) (HSIAO, 1973), o que permitiu às
plantas sob esse tratamento uma melhor recuperação do turgor celular quando comparadas às
plantas sob os tratamentos IDC (-0,29 a -0,476 MPa ) e ID (-0,30 a -0,51 MPa ). Deloire et al.
(2005) relatam que além do potencial hídrico ser considerado o método mais prático para

60
controlar o estado hídrico da videira, as diferentes variedades de plantas apresentam diferentes
potenciais hídricos foliar de base e a resposta da planta ao estresse hídrico vai depender da
situação atual em que se encontra e das condições anteriores pelas quais foram submetidas.
Os resultados de Ψb encontrados no presente experimento, discordam
dos encontrados por Pereira et al. (2006) e Souza et al. (2009) que trabalharam com diferentes
manejos de irrigação (Irrigação parcial das raízes-PRD e DI), diferentes copas e porta-enxertos
e não registraram potenciais hídricos foliares de base inferiores a -0,2 MPa durante suas
avaliações realizadas no SVSF.
Antes de iniciar as estratégias hídricas de irrigação plena (IP),
irrigação com défice controlado (IDC) e irrigação deficitária (ID), até 44 DAP o
armazenamento de água no solo nas camadas até 60 cm apresentava valores próximos e sem a
presença de deficiência hídrica.
Após o início das estratégias de irrigação, a quantidade de água
armazenada no solo nas camadas de 0,15 a 0,60 m de profundidade (Figura 3) apresentaram
variações em função das irrigações e ocorrência de precipitações, sendo a evapotranspiração e
a drenagem os principais depletores de água no perfil do solo. O tratamento IP apresentou a
maior variação do armazenamento de água em função da irrigação constante durante o ciclo.
Além da variação na disponibilidade hídrica, fatores intrínsecos do
solo e elementos meteorológicos podem ter interferido nas respostas fisiológicas e
bioquímicas das plantas. A presença na área experimental de fragipan, um horizonte
subsuperficial, aparentemente cimentado, denso e com consistência dura a muito dura, quando
seco, e, quando molhado, apresenta-se quebradiço (EMBRAPA, 1999), pode ter restringido
parcialmente a penetração de água e raízes no perfil do solo, provocando variações no Ψb das
folhas e no armazenamento de água no solo. As condições atmosféricas no momento das
coletas também podem ter influenciado as leituras fisiológicas e bioquímicas. É sabido que os
elementos climáticos: temperatura, radiação, insolação e umidade (Figura 1) são menos
limitantes ao crescimento e desenvolvimento das plantas no final do primeiro semestre no
Vale do Submédio do São Francisco, período em que foi executado o experimento, sendo
registradas condições climáticas favoráveis às plantas C3 como temperaturas mínimas e
máximas variando entre 19,8 a 31ºC (LUTTGE et al., 1996. citado por PIMENTEL, 1998),
respectivamente, menor incidência direta da radiação solar devido a períodos com

61
nebulosidades durante as avaliações, o que era de se esperar, visto coincidir com o período de
menor insolação e radiação solar nessa região, caracterizado como solstício de inverno. Isso
resultou em alguns períodos ao longo do ciclo das plantas em menor demanda evaporativa do
ar pelas folhas, o que facilitaria rápida recuperação da turgescência celular no período noturno
e antemanhã. Além disso, maior ou menor exigência hídrica devido à evolução das fases
fenológicas das plantas pode ter contribuído também para a variação do potencial hídrico
foliar de base.
A videira é uma planta com alguma adaptação à limitação da
disponibilidade hídrica do solo, devido ao seu sistema radicular profundo e aos mecanismos
fisiológicos desencadeados, traduzidos numa eficiente redução da condutância estomática e na
capacidade de ajustamento osmótico (RODRIGUES et al., 1993, PATAKAS e NOITSAKIS,
2001).
4.2 Trocas gasosas
4.2.1 Assimilação de CO2 (A)
A fotossíntese é um processo vital aos seres vivos dependentes de O2 e
metabólitos primários, envolvendo por um processo difusivo a entrada e saída de substâncias
químicas inorgânicas na forma gasosa (CO2, H2O e O2) entre os vegetais e o ambiente ao qual
estão inseridos, na presença da energia radiante (radiação solar fotossinteticamente ativa).
Além disso, envolve o consumo de nitrogênio para a síntese de enzimas carboxilativas
(Rubisco e Pep-case) e, também, no sítio fotossintético (cloroplastos) vegetal, reações
oxidorredutoras de transferência (e-); armazenamento de energia (ATP e NADPH) e reações
bioquímicas de construção de esqueletos carbônicos (triose fosfato), necessários às rotas do
metabolismo primário e secundário dos vegetais.
Os dados referentes ao resumo da análise de variância da taxa de
assimilação de CO2 aos 54, 60, 73, 87, 101 e 115 DAP encontram-se na Tabela 3. Ocorreram

62
diferenças significativas entre os tratamentos aplicados e nota-se também interação
significativa entre os mesmos.
Tabela 3. Resumo da análise de variância (teste F) da taxa de assimilação de CO2 em folhas
de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda (DAP) em função de três
estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE, 2010.
FV GL Quadrado Médio
EI 2 46,94**
DAP 5 124,68**
Bloco 2 0,17ns
EI x DAP 10 8,35**
Resíduo 34 1,14
CV (%) - 8,79
ns: não significativo; ** p<0,01.
Aos 54 e 60 DAP não foram observadas diferenças significativas em
relação à A entre as estratégias de irrigação aplicadas e os diferentes dias de coleta após a poda
(Tabela 4). Aos 73 DAP foram registrados os maiores valores de fotossíntese durante o ciclo
apenas para o ID e IP, com diferença estatística entre as três estratégias, indicando que havia
água disponível no solo e passível abertura estomática. Dentre os tratamentos aplicados, o que
promoveu maior inserção carbônica no sítio fotossintético das folhas nessa coleta foi o IP,
sendo superior em 25,1% ao ID e 36,3% ao IDC. Esses maiores valores fotossintéticos podem
estar relacionados com a precipitação (Figura 2) ocorrida seis dias antes das leituras
fotossintéticas, a irrigação realizada nos tratamentos IP e IDC para suprir a evapotranspiração
diária nessa fase fenológica (início do pintor) e aos fatores ambientais registrados durante esta
coleta (Figura 1), principalmente temperatura média de 24ºC, umidade relativa do ar de 60% e
menor radiação solar global, sendo estas condições favoráveis ao metabolismo C3 de fixação
do CO2. Além disso, fatores metabólicos internos das folhas como maior maturidade e força

63
dos drenos, podem ter provocado aumento no consumo de carbono nessa fase fenológica,
consequentemente, maiores taxas fotossintéticas.
Tabela 4. Taxa de assimilação de CO2 (μmol m-2
s-1
) em folhas de videira ‘Syrah’ ao longo do
ciclo após o início das três estratégias de irrigação (Irrigação com défice controlado-IDC,
Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE, 2010.
Dias após a poda Estratégias de irrigação
IDC ID IP
54 11,49 Aa 12,67 Ab 13,09 Ac
60 11,76 Aa 12,11 Ab 13,45 Ac
73 14,06 Ca 16,52 Ba 22,07 Aa
87 13,21 Ba 13,28 Bb 17,07 Ab
101 7,34 Ab 7,72 Ac 6,97 Ad
115 8,32 Bb 6,58 Bc 11,49 Ac
CV (%) 8,79
Médias seguidas de mesma letra minúsculas na coluna e maiúsculas na linha, não diferem
entre si pelo teste Tukey a 5% de probabilidade.
As taxas fotossintéticas obtidas aos 73 DAP foram superiores às
encontradas para a cv. Syrah por alguns autores em diferentes estudos (REGINA e
AUDEGUIN, 2005; FAVERO, 2007), Prieto et al. (2010) encontraram valores próximos a 16
μmol m-2
s-1
quando submeteram as plantas a diferentes intensidades luminosas; no entanto,
outros estudos com outras variedades registraram valores fotossintéticos de até 20 μmol m-2
s-1
(GAMON e PEARCY, 1990 DURING, 1991; WILLIAMS, 1996), ou mesmo superiores
(CHENG e CHENG, 2003; MALAGI, 2011). Para os autores citados anteriormente, as taxas
fotossintéticas variam em função da cultivar, da fase fenológica, do sistema de condução e das
condições ambientais.

64
As taxas fotossintéticas se mantiveram altas até os 87 DAP,
caracterizando a fase de maturação, onde há grande demanda por fotoassimilados pelos
principais órgãos drenos (frutos) e, consequentemente, intensa atividade metabólica das
folhas. Além disso, as altas taxas fotossintéticas também podem estar relacionadas com a
maturidade das folhas. Resultados semelhantes foram obtidos por Intrieri (1992) e Regina
(1995) que relataram altas atividades fotossintéticas em folhas de videiras durante a maturação
e com idade superior a 120 dias. Nessa fase fenológica a estratégia de irrigação IP apresentou
taxa fotossintética 22% superior aos tratamentos IDC e ID.
Para Regina (1995) o estímulo gerado pela presença de frutos, aumenta
a taxa de assimilação do carbono na folha fonte. Sharkey (1985) citado por Regina (1995)
credita esse estímulo à relação existente entre a taxa de utilização das trioses fosfato e à
atividade da enzima Rubisco. Ele considera que, em situações de baixa produção de frutos, a
taxa reduzida de utilização das trioses fosfato ocasiona um decréscimo na atividade da enzima
Rubisco. O efeito estimulador do fruto na fotossíntese das folhas fonte é particularmente
importante no caso de plantas novas e com pequenas quantidades de reservas nos troncos e
raízes (LOVEYS e KRIEDEMANN, 1973; KRIEDEMANN, 1977; CHAVES, 1986;
BERTAMINI et al, 1992; WILLIAMS, 1996), como é o caso do presente experimento,
levando ao maior incremento fotossintético na necessidade de compor reservas para ciclos
consecutivos. Desta forma, os altos valores de fotossíntese registrados no presente estudo
podem estar relacionados com as características mencionadas, assim como, a necessidade das
plantas em compor reservas para os próximos ciclos.
Aos 101 DAP não houve diferença significativa entre as estratégias de
irrigação (Tabela 4). Entretanto, ocorreu redução nas taxas fotossintéticas, alcançando valores
de 6,97; 7,72 e 7,34 μmol m-2
s-1
nas respectivas estratégias hídricas IP, ID e IDC. O que
equivaleu a valores da ordem e 31,58; 46,73 e 52,20% dos maiores valores de A atingidos aos
73 DAP. Essa resposta pode estar relacionada à maior redução na disponibilidade de água para
as plantas, nessa fase em todos os tratamentos, conforme os valores do Ψb aos 99 DAP
(Tabela 2) e a menor densidade de fluxo de fótons fotossinteticamente ativos (DFFFA) nessa
avaliação (Figura 5).
A última avaliação ocorreu no dia da colheita 115 DAP e pode ser
notada alta atividade fotossintética nas folhas, com destaque para o tratamento IP (11,49 μmol

65
m-2
s-1
) que manteve elevados valores de assimilação de CO2 durante todo o período analisado.
Esse fato pode estar relacionado com a melhor hidratação das folhas durante todo o ciclo
(Tabela 2), levando à melhor relação média das trocas gasosas ao longo do ciclo (Tabela 4) e,
consequentemente, da fotossíntese.
Figura 5. Valores médios da densidade de fluxo de fótons
fotossinteticamente ativos (DFFFA, μmol m-2
s-1
), medidos entre 8:00 e
9:30 horas da manhã ao longo do ciclo da videira cv. Syrah, sob três
estratégias de irrigação. Embrapa Semiárido, Petrolina-PE, 2010.
4.2.2 Transpiração (E)
Os dados referentes ao resumo da análise de variância dos valores de
transpiração (mmol m-2
s-1
) aos 54, 60, 87, 101 e 115 dias após a poda estão apresentados na
Tabela 5. Ocorreram diferenças significativas para os tratamentos e os diferentes dias de
avaliação após a poda, assim como interação significativa entre os mesmos, para os quais
foram feitos os devidos desdobramentos, tendo sido avaliado os seus respectivos efeitos
isoladamente.

66
Tabela 5. Resumo da análise de variância (teste F) da taxa de transpiração em folhas de
videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda (DAP) em função de três
estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE, 2010.
FV GL Quadrado Médio
EI 2 16,74**
DAP 5 45,53**
Bloco 2 2,25ns
EI x DAP 10 2,19**
Resíduo 34 0,36
CV (%) - 12,25
ns: não significativo; ** p<0,01.
Uma das primeiras reações da planta a uma situação de baixa
disponibilidade hídrica consiste na redução da abertura estomática (LOVEYS, 1984; CIFRE et
al., 2005), consequentemente, reduzindo o processo de arrefecimento das folhas. As condições
que afetam a transpiração, principalmente, o potencial hídrico do ar, dos tecidos e do solo, a
condutividade e densidade estomática e a resistência ao movimento da água do solo para a
planta e nos tecidos da planta, assumem importante papel na definição do estado hídrico da
planta (GIL e PSZCZOŁKOWSKI, 2007).

67
Tabela 6. Taxa de transpiração (mmol m-2
s-1
) em folhas de videira ‘Syrah’ ao longo do ciclo
em função de três estratégias de irrigação (Irrigação com défice controlado-IDC, Irrigação
deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE, 2010.
Dias após a poda Estratégias de irrigação
IDC ID IP
54 4,23 Abc 5,05 Ab 5,31 Ac
60 3,06 Acd 3,10 Ac 3,56 Ade
73 6,17 Ca 8,34 Ba 10,24 Aa
87 4,70 Cab 6,97 Ba 8,33 Ab
101 2,51 Ad 2,71 Ac 3,16 Ae
115 3,09 Bcd 2,59 Bc 4,70 Acd
CV (%) 12,25
Médias seguidas de mesma letra minúsculas na coluna e maiúsculas na linha, não diferem entre si pelo
teste Tukey a 5% de probabilidade.
A transpiração nas folhas nesse estudo seguiu o mesmo padrão
estatístico observado para a assimilação de CO2 (Tabela 6).
Apesar de serem fenômenos distintos, a transpiração e a fotossíntese
estão interligados, principalmente, pelos estômatos das folhas que são a conexão da planta
com o ar atmosférico, e pelos quais as plantas realizam suas trocas gasosas.
Aos 54 e 60 DAP as taxas transpiratórias não foram influenciadas
pelos diferentes tratamentos, indicando que mesmo com a suspensão da irrigação nos
tratamentos IDC e ID, o fluxo transpiratório não diferenciou do tratamento com irrigação
plena (IP).
As maiores taxas transpiratórias neste estudo ocorreram aos 73 DAP
para o tratamento IP; aos 73 e 87 DAP para o tratamento ID e IDC. Resultados parecidos
podem ser encontrados também nas taxas fotossintéticas nessas datas e/ou intervalos (Tabela
4). O tratamento IP apresentou estatisticamente, as maiores médias transpiratórias e
fotossintéticas aos 73 DAP em relação aos tratamentos com restrição hídrica (IDC e ID)
durante o ciclo. As variações no conteúdo hídrico das folhas registradas logo ao amanhecer

68
(Tabela 2) e durante o ciclo indicaram a influência dos tratamentos no comportamento
fisiológico da videira. Os valores das taxas de transpiração aos 73 e 87 DAP para as três
estratégias de irrigação estão dentro do intervalo obtido por Souza et al. (2009) (6 a 13 mmol
m-2
s-1
) durante estudos no SVSF.
Aos 101 DAP não houve diferença significativa entre os tratamentos e
ocorreu queda acentuada no fluxo transpiratório para todas as estratégias hídricas em estudo.
Esta redução pode estar relacionada, principalmente, a carência hídrica no solo, além de menor
intensidade da radiação solar e temperatura do ar medidas nessa avaliação. A nebulosidade do
período promoveu, consequentemente, menor densidade no fluxo de fótons
fotossinteticamente ativos (Figura 5) disponível às plantas através da radiação solar, assim
como menor temperatura do ar e, consequentemente, menor défice de pressão de vapor (DPV).
Todavia o ambiente atmosférico não era estressante às plantas. Diferentemente, o ambiente
edáfico pode ter afetado de forma expressiva a taxa transpiratória, com redução de 62; 61,1 e
46,6% para os tratamentos IP, IDC e ID respectivamente. As plantas aos 101 DAP com base
nas leituras de Ψb (Tabela 2) feitas dois dias antes se encontravam sob défice moderado (IP e
IDC) a severo (ID). Logo, esta situação de estresse, aumentou as resistências internas da
planta (mesofílica e estomática) à perda de água na forma de vapor para o ambiente, o que,
consequentemente, afetou também as taxas fotossintéticas nesse dia. Desta forma, é possível
que nessa avaliação os estômatos estivessem impondo maior resistência à sua abertura em
função das condições adversas do ambiente edáfico como elevada tensão de água.
A taxa de transpiração (E) está relacionada com a perda de água na
forma de vapor, portanto, esses resultados estatísticos podem indicar maior fechamento
estomático nessa avaliação (101 DAP), para todas as estratégias hídricas trabalhadas em
relação à avaliação anterior. Com isso, as plantas evitariam maiores perdas de vapor d’água
para a atmosfera, o que poderia gerar problemas como a cavitação no fluxo contínuo solo-
planta-atmosfera. A capacidade de adaptação da videira a condições de falta d’água resulta em
grande parte da sua capacidade para regular as perdas por transpiração, através de aumento da
resistência estomática (CLIMACO et al., 1991; CHAVES et al., 2009; 2010). Em condições
de défice hídrico as raízes sintetizam sinais químicos, sob a forma de acido abscísico (ABA),
que são transportados até as folhas, conduzindo à redução da abertura estomática (LOVEYS,
1984; CHAVES et al., 2009; 2010). As perdas de água são reduzidas, assim como a entrada de

69
CO2. A limitação da fotossíntese resultante de condições de défice hídrico tem origem,
sobretudo na regulação da abertura dos estômatos, mas existem também limitações não
estomáticas que ocorrem ao nível do mesofilo clorofiliano (FLEXAS et al., 2002; CIFRE et
al., 2005; SOUZA et al., 2005a). Sob baixa a moderada disponibilidade hídrica a manutenção
das atividades das enzimas do ciclo de Calvim e a máxima taxa de carboxilação e o transporte
de elétrons ocorrem normalmente (SOUZA et al., 2005a). No entanto, se a deficiência hídrica
se intensifica ocorre declínio, principalmente, no transporte de elétrons, (MAROCO et al.,
2002; SOUZA et al., 2005a) possivelmente, resultando na redução da produção de ATP
(LAWLOR e TEZARA, 2009), levantando a hipótese de que espécies reativas de oxigênio
(ROS) produzidas sob condições de baixo CO2 e excesso de luz podem induzir danos
oxidativos na ATPase do cloroplasto.
A transpiração e a fotossíntese estão, portanto, intrínsicamente
relacionadas. Se por um lado o complexo mecanismo que regula as trocas gasosas das plantas
é importante na adaptação às condições de aridez, através da redução da transpiração, este
mesmo mecanismo poderá conduzir também à menor taxa fotossintética, com consequências
no crescimento e produção (KRAUTER, 2001). A solução para este problema passa por
melhor eficiência do uso da água pelas plantas, geralmente, avaliada pela razão entre a
fotossíntese e a transpiração (A/E) ou entre a fotossíntese e a condutância estomática (A/gs)
(DURING, 1994; FLEXAS et al., 1998).
A irrigação deficitária e a irrigação com défice controlado
apresentaram as menores taxas transpiratórias e fotossintéticas aos 115 DAP, evidenciando a
importância da disponibilidade hídrica no solo no favorecimento das trocas gasosas. Nesse
momento já havia sido restabelecido aumento na lâmina de irrigação segundo a ETc da cultura
na fase final do ciclo fenológico e o tratamento IP aumentou em valor significativo suas taxas
fotossintéticas e transpiratórias comparando-as à avaliação anterior (101 DAP). Esse resultado
pode ter sido influenciado, muito provavelmente, pelo retorno de uma maior incidência de
radiação solar, conforme a Figura 5, o que gerou também aumento na temperatura do ar e no
DPV, gerando maior efluxo do vapor d’água e influxo do CO2, especificamente, na câmara
subestomática nas folhas. Ainda nessa avaliação não houve diferença significativa para os Ψb
entre 99 e 113 DAP, no entanto, não se pode descartar a hipótese de que uma menor tensão da
água no solo registrada pelo Ψb aos 115 DAP possa ter contribuído nessa resposta do

70
tratamento IP, isso porque as atividades fisiológicas de trocas gasosas ocorrentes na parte
aérea são completamente dependentes de sinalizações químicas e/ou suprimentos hídricos
fornecidos pelas raízes. O tratamento ID e IDC não diferenciaram estatisticamente entre si e
nem entre as avaliações aos 101 e 115 DAP no que se refere às taxas transpiratórias e
apresentaram os menores valores de assimilação de CO2 e transpiração, indicando maior
fechamento estomático devido à baixa disponibilidade hídrica no solo e ao aumento no défice
de pressão de vapor do ar, provocado pelo aumento da radiação solar, insolação e temperatura
(Figura 1) sendo estes alguns dos fatores ambientais preconizados por Jones (1992), que
influenciam na resistência estomática. Contudo, a irrigação deficitária demonstrou maior
eficiência intrínseca no uso da água em relação aos demais tratamentos aos 115 DAP.
Quando as videiras são submetidas ao défice hídrico moderado as
taxas fotossintéticas, geralmente, declinam com o menor potencial hídrico foliar de base do
que a condutância estomática. Como consequência, a eficiência intrínseca do uso da água
(A/gs ou WUE) é, geralmente, maior em videiras sob défice de irrigação (défice hídrico leve a
moderado) do que sob condições bem irrigadas. Isto é refletido em menor uso da água e maior
WUE pelas culturas, importante objetivo das estratégias de irrigação com défice em vinhedos
(GAUDILLERE et al., 2002; CHAVES et al., 2004; SOUZA et al., 2005b) cultivados,
principalmente, em regiões semiáridas.
As maiores perdas de água na forma de vapor durante o ciclo foram
registradas no tratamento IP, assim como as maiores taxas fotossintéticas. Isto já era esperado,
partindo do princípio de que uma folha em seu estado hídrico ótimo apresenta estômatos com
o máximo de abertura dos ostíolos e, consequentemente, com maior volume de trocas gasosas
com o ambiente.
4.2.3 Condutância Estomática (gs)
Os dados referentes ao resumo da análise de variância dos valores de
condutância estomática (mol H2O m-2
s-1
) aos 54, 60, 87, 101 e 115 dias após a poda (DAP)
estão apresentados na Tabela 7. Houve resposta significativa tanto para as diferentes
estratégias hídricas quanto para as seis avaliações após a poda.

71
A condutância estomática respondeu, semelhantemente, à fotossíntese
e à transpiração, desde que são fatores intrinsecamente relacionados.
Os estômatos são orifícios presentes nas partes aéreas
fotossintetizantes, principalmente, na lâmina foliar, compostos por células guarda e ostíolo por
onde ocorre a maior parte das trocas gasosas nos vegetais. A sua abertura e o fechamento são
influenciados por muitos fatores, destacando a luminosidade e a variação da turgescência em
células guarda (ALQUINI et al., 2006).
Tabela 7. Resumo da análise de variância (teste F) da condutância estomática em folhas de
videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda (DAP) em função de três
estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE, 2010.
FV GL Quadrado Médio
EI 2 0,0523**
DAP 5 0,1796**
Bloco 2 0,0010ns
EI x DAP 10 0,0072ns
Resíduo 34 0,0069
CV (%) - 24,75
ns: não significativo; ** p<0,01.
Os valores médios da gs obtidos das folhas aos 54 e 60 DAP não
diferenciaram estatisticamente (Tabela 8), apresentando comportamento parecido com os
obtidos para a taxa de assimilação, onde não houve diferença significativa entre as estratégias
de irrigação utilizadas nessas duas avaliações.
Flexas et al. (2004) consideram a gs um parâmetro integrador dos
vários sinais associados à resposta da planta ao défice hídrico. A razão A/gs (eficiência
intrínseca do uso da água) permitirá avaliar o tipo de limitações envolvidas na resposta da
fotossíntese à carência hídrica (NAOR e WAMPLE, 1995; FLEXAS et al., 2004). De acordo
com Kindermann e Smart (1971) as limitações estomáticas ocorrem quando o potencial

72
hídrico na folha atinge valores entre -1,3 a -1,6 MPa. O início da ocorrência de limitações
metabólicas da fotossíntese é verificada quando os valores de condutância estomática situam-
se entre 0,15> gs > 0,05 mol H2O m-2
s-1
, caracterizado também como nível médio de carência
hídrica (FLEXAS et al., 2002; 2004; MEDRANO et al., 2002; CIFRE et al., 2005).
Os valores de potenciais citados acima não foram mensurados no
presente experimento, pois não se avaliou o potencial hídrico das plantas ao longo do dia e nas
horas mais quentes (maior défice de pressão de vapor). Entretanto, em função de um valor
mais ou menos negativo nas leituras do Ψb realizadas na madrugada posterior às condições
climáticas mais estressantes, facilitaria a interpretação de que aquelas plantas poderiam ter
atingido os seus limites estomáticos e/ou metabólicos de uma forma mais ou menos severa.
Alguns autores defendem que se deve irrigar a videira apenas quando a condutância
estomática decresce abaixo do nível médio de carência hídrica, uma vez que este nível permite
uma razão A/gs máxima, rápida recuperação da fotossíntese após a irrigação e melhor
qualidade das uvas, com perdas reduzidas de rendimento comparativamente a irrigação
intensiva (FLEXAS et al., 2002; MEDRANO et al., 2002; CIFRE et al., 2005).

73
Tabela 8. Condutância estomática (mol m-2
s-1
) em folhas de Syrah ao longo do ciclo em
função de três estratégias de irrigação (Irrigação com défice controlado-IDC, Irrigação
deficitário-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE, 2010.
Dias após a poda Estratégias de irrigação
IDC ID IP Média
54 0,38 0,38 0,41 0,39 b
60 0,32 0,33 0,35 0,33 bc
73 0,37 0,50 0,65 0,51 a
87 0,35 0,41 0,51 0,42 ab
101 0,22 0,26 0,30 0,26 c
115 0,09 0,07 0,15 0,10 d
Média 0,28 B 0,32 B 0,39 A
CV (%) 24,75
Médias seguidas de mesma letra minúsculas na coluna e maiúsculas na linha, não diferem
entre si pelo teste Tukey a 5% de probabilidade.
O valor médio da gs ao longo do ciclo para o tratamento IP (0,39 mol
m-2
s-1
) foi superior estatisticamente aos tratamentos IDC e ID. Essa resposta pode estar
associada com a frequência da irrigação, onde as folhas coletados no tratamento IP
apresentavam melhor estado hídrico, o que pode ter resultado em maior abertura estomática,
favorecendo a gs em relação aos tratamentos IDC e ID.
Aos 73 DAP as folhas marcadas para análise das trocas gasosas
apresentaram os maiores valores de condutância estomática, diferenciando estatisticamente
das folhas coletadas anteriormente (56 e 60 DAP), e aos 101 e 115 DAP. No entanto, essa
média do resultado de condutância estomática não diferiu da 4ª coleta feita aos 87 DAP
(Tabela 8). Os valores de gs registrados ao longo do ciclo estão de acordo com os resultados
obtidos para fotossíntese e transpiração, indicando boa interação entre esses três processos.
Esse resultado expressivo, principalmente, obtido na 3ª coleta (73 DAP) pode ter sido
influenciado por uma maior hidratação da zona radicular em função de precipitação e irrigação
localizada, ocasionando, consequentemente, maior absorção e transporte de água até a parte

74
aérea das plantas, mantendo um estado hídrico ótimo das folhas, garantindo maiores taxas de
E e A. Isto pode ser verificado conforme a Figura 2 e com base na leitura do Ψb feita aos 70
DAP (Tabela 2). Além disso, os fatores climáticos no momento da coleta não eram
estressantes, sendo registrada temperatura em torno de 24ºC, umidade relativa do ar de 60%,
baixa insolação e radiação solar (Figura 1), típico de ambientes favoráveis ao metabolismo C3.
Assim, com o baixo défice de pressão de vapor do ar e uma relativamente elevada gs aos 73
DAP, as plantas puderam expressar o seu potencial fotossintético, carboxilando com mais
eficiência o CO2 atmosférico e o convertendo em metabólitos primários.
A gs varia durante o dia, alcançando os maiores valores nas primeiras
horas da manhã, quando as condições atmosféricas são favoráveis às trocas gasosas. Em geral,
diminui durante o dia conforme aumenta a diferença de pressão de vapor (DPV) entre a folha e
a atmosfera (SCHULZE e HALL, 1982), havendo, contudo, diferenças significativas entre
espécies.
A partir dos 87 DAP até a realização da colheita houve tendência
decrescente da gs para todos os DAPs posteriores, indicando a presença de défice hídrico
suave a moderado, o que pode ser confirmado com as leituras de potencial hídrico foliar de
base presentes na Tabela 2. O estresse hídrico mesmo leve pode influenciar o fechamento
estomático, o que acarreta redução da gs, E e, consequentemente, da A, influenciando
diretamente na produção de frutos.
Na quinta avaliação aos 101 DAP o défice hídrico registrado pelos
potenciais de base aos 99 DAP pode ter afetado a condutância estomática, cujo valor
mensurado foi 0,26; o que sinaliza limitação estomática leve (FLEXAS et al., 2002; 2004;
MEDRANO et al., 2002; CIFRE et al., 2005). Os valores de gs obtidos durante o ciclo estão
de acordo com os obtidos por Souza et al. (2001) que trabalharam com mudas de videiras com
dois porta-enxertos sob deficiência hídrica e corroboram também com valores obtidos por
Souza et al. (2009) em trabalhos realizados na mesma região do presente trabalho.
Aos 115 DAP o valor médio da gs (0,10 mols H2O m-2
s-1
), apresentou
redução de 80,4 e 76,2 % em relação aos 73 e 87 DAP respectivamente, onde apresentaram os
maiores valores das trocas gasosas. O valor baixo da gs na última avaliação é equivalente a
situação de défice hídrico médio e se aproxima dos valores obtidos por Cerqueira (2011)
trabalhando com deficiência hídrica e estresse por frio em videiras ‘Thompson Seedles’ em

75
ambiente controlado. Este autor encontrou valores de gs da ordem de 0,09 mols H2O m-2
s-1
para os tratamentos com deficiência hídrica e 0,07 mols H2O m-2
s-1
para os tratamentos com
frio. Isso indica que são muitos os fatores internos e ambientais envolvidos com a abertura e o
fechamento estomático como: luminosidade, concentração de CO2, tensão de água,
temperatura e défice de pressão de vapor, umidade do ar, vento, nutrição mineral, ritmo
circadiano e hormônios vegetais (MARRENCO e LOPES, 2009).
4.2.4 Eficiência no uso da água (A/E)
A eficiência no uso da água é a relação entre o carbono assimilado e o
volume de água transpirado. Apenas pequena proporção da água que circula através da planta
é utilizada nos processos metabólicos, sendo a maior parte perdida para a atmosfera na forma
de vapor d’água no processo da transpiração.
Encontram-se na Tabela 9 os dados referentes ao resumo da análise de
variância e a significância dos tratamentos utilizados para a eficiência no uso da água (EUA)
aos 54, 60, 87, 101 e 115 dias após a poda (DAP).
Tabela 9. Resumo da análise de variância (teste F) da eficiência do uso da água (assimilação
de CO2/transpiração) em folhas de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a
poda (DAP) em função de três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina,
PE, 2010.
FV GL Quadrado Médio
EI 2 0,7501**
DAP 5 3,4808**
Bloco 2 1,1099**
EI x DAP 10 0,1421ns
Resíduo 34 0,1149
CV (%) - 12,56
ns: não significativo; ** p<0,01.
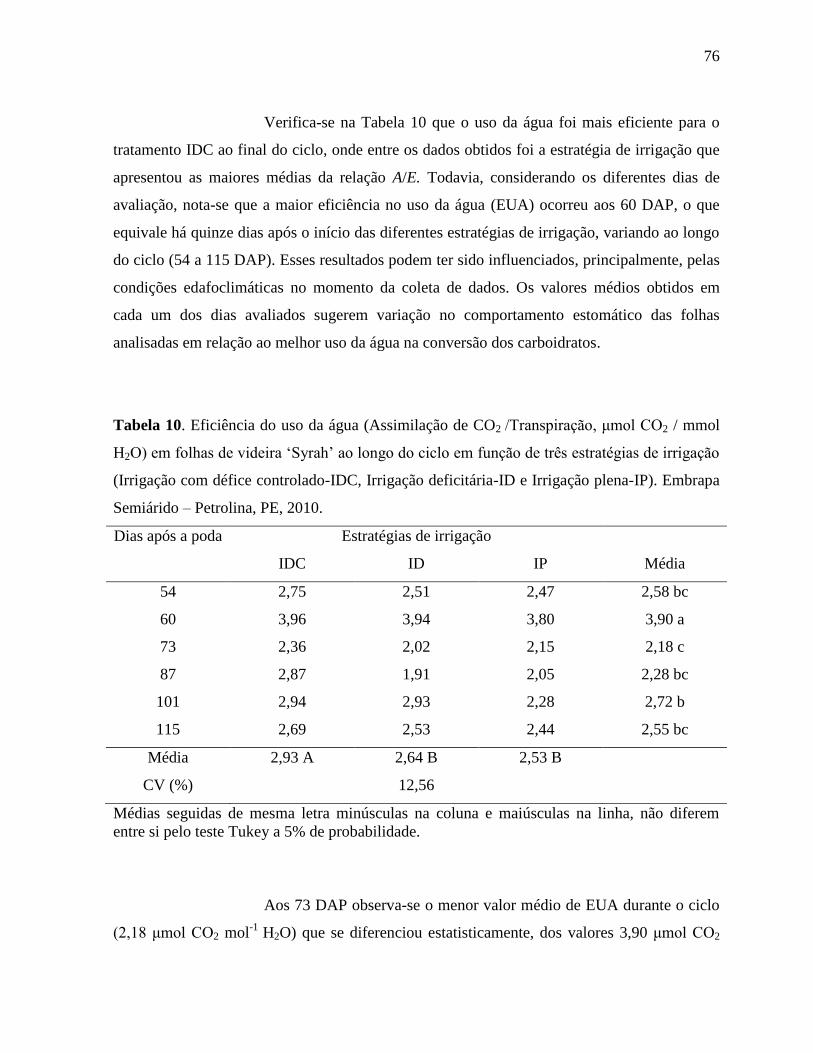
76
Verifica-se na Tabela 10 que o uso da água foi mais eficiente para o
tratamento IDC ao final do ciclo, onde entre os dados obtidos foi a estratégia de irrigação que
apresentou as maiores médias da relação A/E. Todavia, considerando os diferentes dias de
avaliação, nota-se que a maior eficiência no uso da água (EUA) ocorreu aos 60 DAP, o que
equivale há quinze dias após o início das diferentes estratégias de irrigação, variando ao longo
do ciclo (54 a 115 DAP). Esses resultados podem ter sido influenciados, principalmente, pelas
condições edafoclimáticas no momento da coleta de dados. Os valores médios obtidos em
cada um dos dias avaliados sugerem variação no comportamento estomático das folhas
analisadas em relação ao melhor uso da água na conversão dos carboidratos.
Tabela 10. Eficiência do uso da água (Assimilação de CO2 /Transpiração, μmol CO2 / mmol
H2O) em folhas de videira ‘Syrah’ ao longo do ciclo em função de três estratégias de irrigação
(Irrigação com défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa
Semiárido – Petrolina, PE, 2010.
Dias após a poda Estratégias de irrigação
IDC ID IP Média
54 2,75 2,51 2,47 2,58 bc
60 3,96 3,94 3,80 3,90 a
73 2,36 2,02 2,15 2,18 c
87 2,87 1,91 2,05 2,28 bc
101 2,94 2,93 2,28 2,72 b
115 2,69 2,53 2,44 2,55 bc
Média 2,93 A 2,64 B 2,53 B
CV (%) 12,56
Médias seguidas de mesma letra minúsculas na coluna e maiúsculas na linha, não diferem
entre si pelo teste Tukey a 5% de probabilidade.
Aos 73 DAP observa-se o menor valor médio de EUA durante o ciclo
(2,18 μmol CO2 mol-1
H2O) que se diferenciou estatisticamente, dos valores 3,90 μmol CO2

77
mol-1
H2O e 2,72 μmol CO2 mol-1
H2O nos 60 e 101 DAP, respectivamente. O menor valor
médio registrado na EUA aos 73 DAP pode estar relacionado com as maiores taxas de A, E e
gs registradas nessa avaliação. As condições climáticas no momento dessa terceira coleta eram
satisfatórias ao metabolismo de plantas C3 como a videira, assim conduzindo a elevadas taxas
de assimilação, condicionando a planta a maiores perdas de água por transpiração e,
consequentemente, maior condutância estomática.
É sabido que em condições semiáridas o uso de estratégias de irrigação
que disponibilizam maior volume de água para as plantas, em sua grande maioria tendem à
apresentar os maiores valores de E e gs como pode ser constatado por diversos autores
(MALHEIRO, 2005; SOUZA et al., 2005a; CHAVES et al., 2007; REYNOLDS et al., 2007;
INTRIGLIOLO e CASTEL, 2009) e, também, por esse estudo. O tratamento IP apresentou os
maiores valores de A, E e gs (Tabelas 4, 6 e 8), entretanto, obteve ao final do ciclo, os menores
valores médios da relação A/E, não diferindo do tratamento ID, caracterizando-os com menor
eficiência no uso da água para conversão em carboidratos. Para o presente estudo, essas
menores médias significativas obtidas para IP e ID poderiam estar relacionadas às suas
maiores médias significativas das taxas transpiratórias alcançadas ao longo do ciclo, sendo
respectivamente, 5,88 e 4,79 mmol m-2
s-1
. O tratamento IDC, contrariamente aos anteriores,
apresentou menor média significativa das taxas transpiratórias (3,96 mmol m-2
s-1
),
influenciando assim, na sua maior EUA registrada ao final do ciclo.
Desta forma, está caracterizado nesse experimento e já bastante
confirmado por muitas pesquisas (MEDRANO et al., 2002; FLEXAS et al., 2002; PATAKAS
et al., 2003; SOUZA et al., 2003; CIFRE et al., 2005; SOUZA et al., 2005a,b; PAYAN et al.,
2006;) que a melhor gestão da água pela planta vai depender do montante de recursos hídricos
disponibilizados pelo sistema de irrigação, sendo que altas taxas de E e gs, geralmente,
condicionam ao termo instituído por Escalona et al. (1997) "consumo de luxo". Assim plantas
sob deficiência hídrica controlada apresentam maior eficiência no uso da água.
4.2.5 Concentração interna de CO2 na câmara subestomática (Ci)

78
A difusão do dióxido de carbono até o sítio de carboxilação da Rubisco
no interior da folha é uma sucessão de rompimentos de resistências impostas pela camada
limítrofe de ar junto à superfície foliar, resistência estomática e resistência nos espaços
intercelulares.
Na Tabela 11 encontram-se o resumo da análise de variância e a
significância dos tratamentos utilizados e sua interação das médias da concentração
intracelular de CO2 na câmara subestomática aos 54, 60, 87, 101 e 115 dias após a poda.
Houve resposta significativa somente para as diferentes datas das avaliações realizadas.
Tabela 11. Resumo da análise de variância (teste F) da concentração interna de CO2 na
câmara subestomática em folhas de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a
poda (DAP) em função de três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina,
PE, 2010.
FV GL Quadrado Médio
EI 2 42,58ns
DAP 5 10238,39**
Bloco 2 87,83ns
EI x DAP 10 134,43ns
Resíduo 34 173,81
CV (%) - 4,12
ns: não significativo; ** p<0,01.
Os valores médios da pressão parcial interna de CO2 (Ci) para as
quatro primeiras avaliações entre 54 e 87 DAP, não diferiram estatisticamente entre si (Tabela
12). Porém, aos 101 DAP, sendo 14 dias após a última avaliação, verificou-se as maiores
médias significativa para Ci, alcançando 357,88 μmol CO2 m-2
. Esta resposta diferenciada aos
101 DAP pode ter sido influenciada, principalmente, por um défice hídrico momentâneo
durante essa avaliação que pode ter reduzido a abertura estomática. Isso pode ser confirmado
de acordo com o valor de Ψb, obtido dois dias antes das leituras das trocas gasosas,

79
alcançando -0,451 MPa, sendo o valor médio mais negativo registrado durante o ciclo. Essa
resposta ainda pode ser confirmada com os baixos valores médios de DFFFA (288,8 μmol m-2
s-1
), A, E, gs e do menor valor significativo de carboxilação in vivo da enzima Rubisco (0,026
μmol m-2
s-1
de CO2. μmol mol-1
CO2) aos 101 DAP, quando comparado à avaliação anterior
(87 DAP).
Aos 115 DAP a Ci apresentou o menor valor médio durante o ciclo,
257,12 μmol CO2 m-2
. Essa resposta pode ter sido influenciada pelas condições
edafoclimáticas momentâneas mensuradas nessa coleta, onde provavelmente, as plantas ainda
estariam sob défice hídrico, com valores de Ψb próximos ao registrado aos 113 DAP (-0,351
MPa). Isso pode ter provocado maior resistência estomática e menor disponibilidade de água
às raízes, além de distúrbios metabólicos iniciais reversíveis no aparato bioquímico de fixação
do carbono. Além disso, a permanência dessas condições durante a leitura pode ter
influenciado os valores de trocas gasosas, principalmente, os valores de gs (0,10 mol m-2
s-1
),
indicando que as plantas estariam sob carência hídrica média (FLEXAS et al., 2002;
MEDRANO et al., 2002; CIFRE et al., 2005), caracterizada por limitações estomáticas e início
de limitações não estomáticas, a nível do mesofilo clorofiliano.
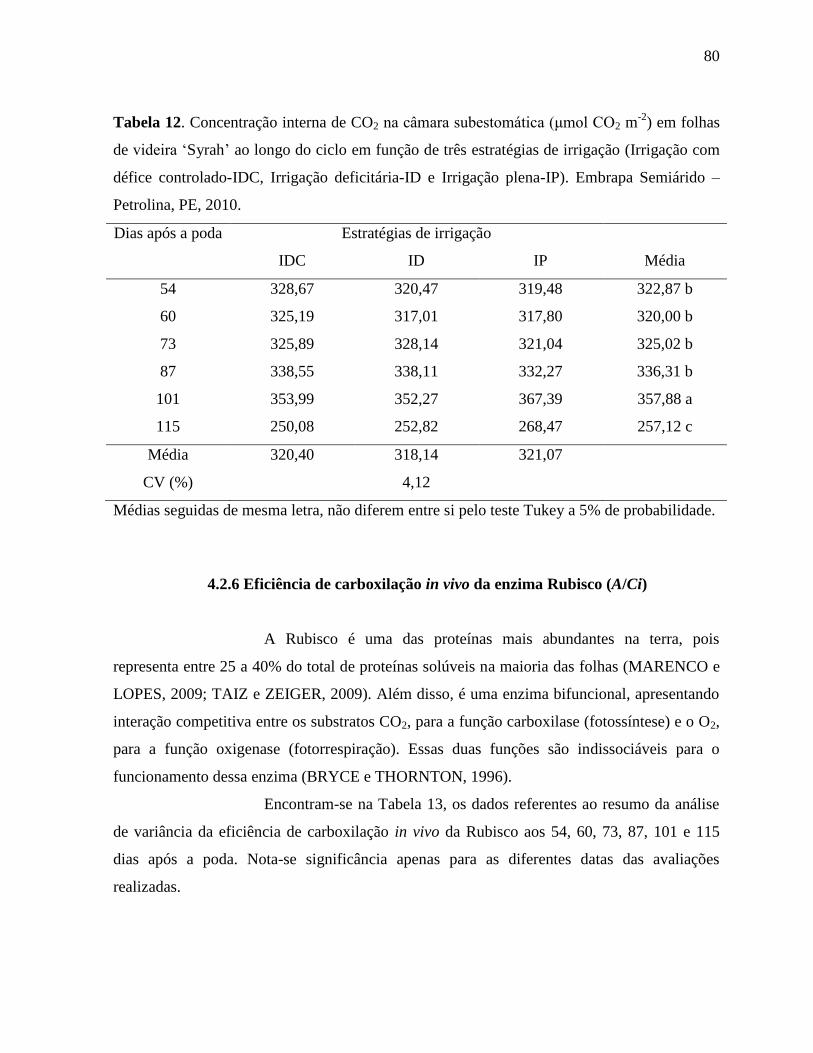
80
Tabela 12. Concentração interna de CO2 na câmara subestomática (μmol CO2 m-2
) em folhas
de videira ‘Syrah’ ao longo do ciclo em função de três estratégias de irrigação (Irrigação com
défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido –
Petrolina, PE, 2010.
Dias após a poda Estratégias de irrigação
IDC ID IP Média
54 328,67 320,47 319,48 322,87 b
60 325,19 317,01 317,80 320,00 b
73 325,89 328,14 321,04 325,02 b
87 338,55 338,11 332,27 336,31 b
101 353,99 352,27 367,39 357,88 a
115 250,08 252,82 268,47 257,12 c
Média 320,40 318,14 321,07
CV (%) 4,12
Médias seguidas de mesma letra, não diferem entre si pelo teste Tukey a 5% de probabilidade.
4.2.6 Eficiência de carboxilação in vivo da enzima Rubisco (A/Ci)
A Rubisco é uma das proteínas mais abundantes na terra, pois
representa entre 25 a 40% do total de proteínas solúveis na maioria das folhas (MARENCO e
LOPES, 2009; TAIZ e ZEIGER, 2009). Além disso, é uma enzima bifuncional, apresentando
interação competitiva entre os substratos CO2, para a função carboxilase (fotossíntese) e o O2,
para a função oxigenase (fotorrespiração). Essas duas funções são indissociáveis para o
funcionamento dessa enzima (BRYCE e THORNTON, 1996).
Encontram-se na Tabela 13, os dados referentes ao resumo da análise
de variância da eficiência de carboxilação in vivo da Rubisco aos 54, 60, 73, 87, 101 e 115
dias após a poda. Nota-se significância apenas para as diferentes datas das avaliações
realizadas.

81
Tabela 13. Resumo da análise de variância (teste F) da eficiência de carboxilação in vivo da
enzima Rubisco em folhas de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda
(DAP) em função de três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE,
2010.
FV GL Quadrado Médio
EI 2 0,00068ns
DAP 5 0,00114*
Bloco 2 0,00023ns
EI x DAP 10 0,00033ns
Resíduo 34 0,00035
CV (%) - 45,15
ns: não significativo; * p<0,05.
Os resultados da eficiência de carboxilação in vivo da Rubisco tiveram
comportamento parecido com o da taxa de assimilação de CO2.
Aos 73 dias após a poda ocorreu a maior eficiência na carboxilação in
vivo da Rubisco entre as seis diferentes datas de coleta (Tabela14), assim como, a maior taxa
de assimilação líquida de CO2 (Tabela 4). Porém, as taxas de carboxilação nessa data não
diferiram estatisticamente dos 54 e 87 DAP como nas taxas de assimilação de CO2 (dados não
mostrados). Essa resposta estaria diretamente relacionada, com condições ambientais
favoráveis ao processo fotossintético, como: disponibilidade de luz, água e CO2 registrados no
momento dessa coleta. As plantas aos 73 DAP podem ter apresentado maior controle
estomático e maior condutância mesofílica o que pode ter favorecido a maior eficiência da
carboxilação. Mudanças na condutância do mesofilo (gm) podem estar ligadas a alterações
físicas na estrutura dos espaços intercelulares devido ao encolhimento da folha (LAWLOR e
CORNIC, 2002) ou a alterações na bioquímica (Conversão do bicarbonato a CO2) e/ou
permeabilidade da membrana (aquaporinas). Flexas et al. (2006) mostraram que a gm
responde muito rapidamente (em poucos minutos) à dessecação, após o corte do pecíolo da
folha no ar e pode ser induzida pela aplicação exógena de ácido abscísico (ABA) em plantas
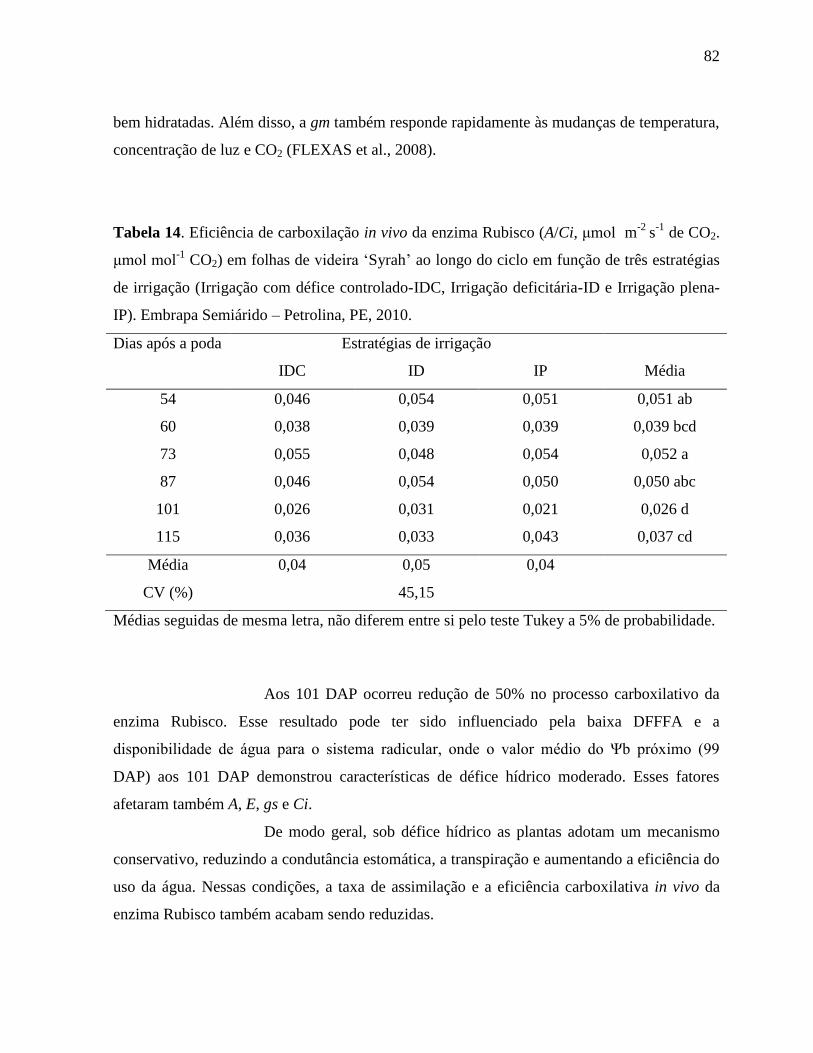
82
bem hidratadas. Além disso, a gm também responde rapidamente às mudanças de temperatura,
concentração de luz e CO2 (FLEXAS et al., 2008).
Tabela 14. Eficiência de carboxilação in vivo da enzima Rubisco (A/Ci, μmol m-2
s-1
de CO2.
μmol mol-1
CO2) em folhas de videira ‘Syrah’ ao longo do ciclo em função de três estratégias
de irrigação (Irrigação com défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-
IP). Embrapa Semiárido – Petrolina, PE, 2010.
Dias após a poda Estratégias de irrigação
IDC ID IP Média
54 0,046 0,054 0,051 0,051 ab
60 0,038 0,039 0,039 0,039 bcd
73 0,055 0,048 0,054 0,052 a
87 0,046 0,054 0,050 0,050 abc
101 0,026 0,031 0,021 0,026 d
115 0,036 0,033 0,043 0,037 cd
Média 0,04 0,05 0,04
CV (%) 45,15
Médias seguidas de mesma letra, não diferem entre si pelo teste Tukey a 5% de probabilidade.
Aos 101 DAP ocorreu redução de 50% no processo carboxilativo da
enzima Rubisco. Esse resultado pode ter sido influenciado pela baixa DFFFA e a
disponibilidade de água para o sistema radicular, onde o valor médio do Ψb próximo (99
DAP) aos 101 DAP demonstrou características de défice hídrico moderado. Esses fatores
afetaram também A, E, gs e Ci.
De modo geral, sob défice hídrico as plantas adotam um mecanismo
conservativo, reduzindo a condutância estomática, a transpiração e aumentando a eficiência do
uso da água. Nessas condições, a taxa de assimilação e a eficiência carboxilativa in vivo da
enzima Rubisco também acabam sendo reduzidas.

83
4.3 Teor SPAD de Clorofila
A quantificação de clorofilas presentes nas folhas é indicativo do poder
de assimilação de CO2 das plantas que estão submetidos aos fatores bióticos e abióticos do
ambiente. O método laboratorial tradicional de quantificação de clorofilas é o de Arnon
(1949), de fácil realização e precisão. No entanto, realiza avaliação destrutiva do tecido foliar,
além de ser relativamente demorado e dispendioso. Assim, o uso de equipamentos portáteis
como o clorofilômetro (SPAD-502, Minolta), devidamente calibrado, apresenta resultados
práticos, rápidos e não destrutivo das folhas, além de possuir baixo custo de manutenção.
Godoy et al. (2008) afirmaram que esse equipamento possibilita a obtenção do índice relativo
da clorofila (IRC), baseando-se na intensidade da coloração verde das folhas, o qual se
correlaciona com o teor de clorofila e de N.
Os dados referentes ao resumo da análise de variância para teor SPAD
de clorofila avaliados aos 54, 60, 73, 87 e 101 dias após a poda, encontram-se na Tabela 15.
Houve diferenças estatísticas significativas entre as estratégias hídricas aplicadas e os
diferentes dias de coletas após a poda.
Tabela 15. Resumo da análise de variância (teste F) para o teor SPAD de clorofila em folhas
de videira ‘Syrah’ aos 54, 60, 73, 87 e 101 dias após a poda (DAP) em função de três
estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE, 2010.
FV GL Quadrado Médio
EI 2 21,01*
DAP 4 88,83**
Bloco 2 17,52*
EI x DAP 8 4,03ns
Resíduo 28 4,26
CV (%) - 5,07
ns: não significativo (p> 0,05); * p<0,05; ** p<0,01.

84
As diferentes estratégias de irrigação apresentaram comportamento
variável ao longo do ciclo na média do Teor SPAD da clorofila, assim como, diferenças
estatísticas significativas (p<0,05) ao final do ciclo (Tabela 16).
Tabela 16. Teor SPAD da clorofila em folhas de videira ‘Syrah’ ao longo do ciclo em função
de três estratégias de irrigação (Irrigação com défice controlado-IDC, Irrigação deficitária-ID
e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE, 2010.
Dias após a poda Estratégias de irrigação
IDC ID IP Média
54 34,79 36,16 34,67 35,21 b
60 39,13 40,87 43,27 41,09 a
73 41,60 41,57 43,43 42,20 a
87 41,07 42,37 43,93 42,46 a
101 42,17 40,90 44,90 42,36 a
Média 39,75 B 40,37 AB 42,04 A
CV (%) 5,07
Médias seguidas de mesma letra minúsculas na coluna e maiúsculas na linha, não diferem
entre si pelo teste Tukey a 5% de probabilidade.
O tratamento IP apresentou a maior média do índice SPAD (42,04),
não diferindo estatisticamente do tratamento ID (40,37). As menores médias do índice SPAD
foram obtidas nas plantas submetidas ao tratamento IDC (39,75). Esses valores obtidos estão
dentro da normalidade, partindo-se do princípio de que as maiores diferenças consideráveis
entre os tratamentos em relação ao teor de clorofila ocorreriam se as plantas estivessem
submetidas ao défice hídrico severo (PIMENTEL, 2004), em que alterações fisiológicas e
bioquímicas influenciariam na redução desse pigmento, coisa que não aconteceu nesse
experimento.

85
Na literatura se encontram alguns estudos com uso do índice SPAD na
avaliação do teor de clorofila em videiras (RUPP e TRANKLE, 1995; RUPP et al., 1999;
STEELE, 2008; AMARANTE et al., 2009), porém, poucos ainda relacionando-o ao estresse
hídrico (FANIZZA et al., 1991), sendo mais utilizado na correlação com o estado nutricional
das plantas (PORRO et al., 2001; TECCHIO et al., 2011).
As leituras do teor SPAD realizadas aos 54 DAP apresentaram o
menor valor médio ao longo do ciclo (35,21). Este resultado pode ter sido influenciado pela
menor maturidade das folhas analisadas.
De acordo com os valores de índice SPAD obtidos e a relação que têm
com o teor de clorofila nas folhas, todos os demais dias após a primeira coleta (54 DAP)
apresentaram aumentos significativos (p<0,01) nesse índice, sem diferenciarem
estatisticamente, entre si. Desta forma, pode-se inferir também que houve incremento no teor
de clorofila nas folhas e este teor durou até próximo à colheita (101 DAP), indicando ainda
que as folhas não se encontravam senescentes. Os valores obtidos nesse experimento para
todas as três estratégias de irrigação estão dentro dos encontrados por Fanizza et al. (1991)
para videiras não estressadas. Muitos autores recomendam o uso do clorofilômetro para
diferentes tipos de cultivo, inclusive para o da videira, desde que devidamente calibrado.
4.4 Atividade da enzima Nitrato Redutase (NR)
Os dados referentes ao resumo da análise de variância da atividade da
NR aos 54, 60, 73, 87 e 101 dias após a poda, encontram-se na Tabela 17, onde foram
verificadas respostas significativas para as diferentes estratégias hídricas, as diferentes datas
de coleta e interação significativa entre eles.
O nitrato (NO3-) é a principal forma de nitrogênio inorgânico
disponível para as plantas e a nitrato redutase é a primeira enzima da rota de assimilação do N-
inorgânico, possuindo papel de extrema importância no metabolismo vegetal (DELÚ-FILHO
et al., 1998).
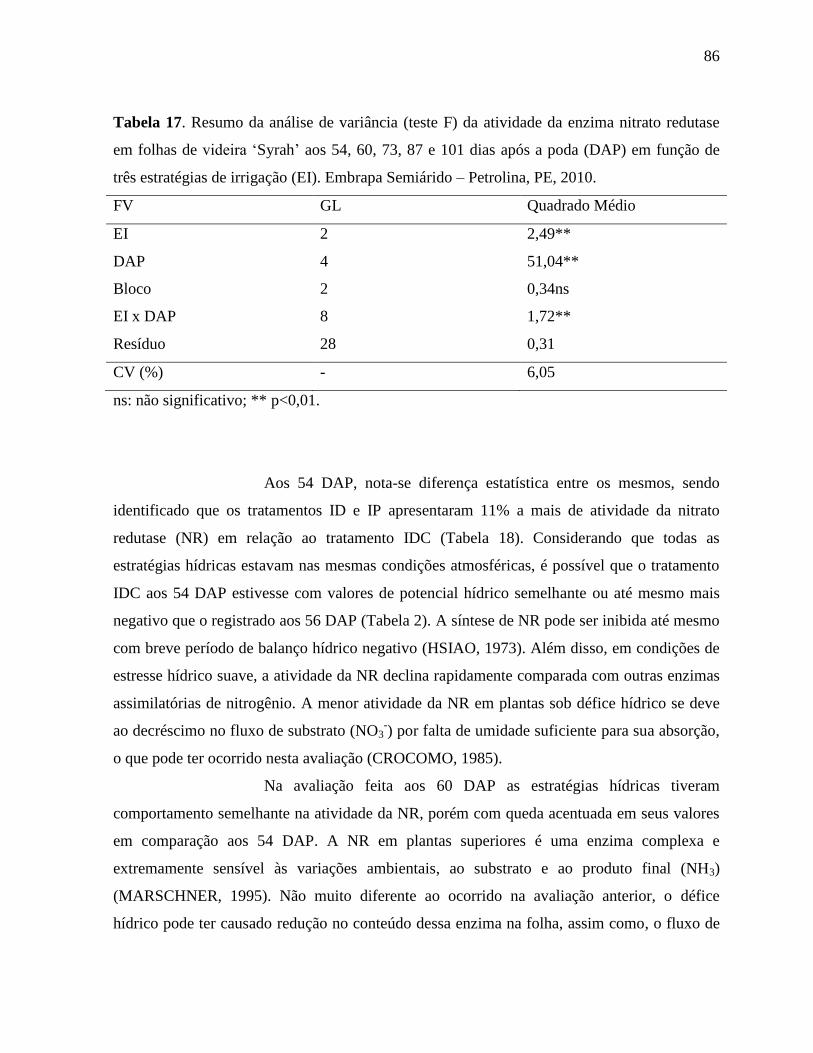
86
Tabela 17. Resumo da análise de variância (teste F) da atividade da enzima nitrato redutase
em folhas de videira ‘Syrah’ aos 54, 60, 73, 87 e 101 dias após a poda (DAP) em função de
três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE, 2010.
FV GL Quadrado Médio
EI 2 2,49**
DAP 4 51,04**
Bloco 2 0,34ns
EI x DAP 8 1,72**
Resíduo 28 0,31
CV (%) - 6,05
ns: não significativo; ** p<0,01.
Aos 54 DAP, nota-se diferença estatística entre os mesmos, sendo
identificado que os tratamentos ID e IP apresentaram 11% a mais de atividade da nitrato
redutase (NR) em relação ao tratamento IDC (Tabela 18). Considerando que todas as
estratégias hídricas estavam nas mesmas condições atmosféricas, é possível que o tratamento
IDC aos 54 DAP estivesse com valores de potencial hídrico semelhante ou até mesmo mais
negativo que o registrado aos 56 DAP (Tabela 2). A síntese de NR pode ser inibida até mesmo
com breve período de balanço hídrico negativo (HSIAO, 1973). Além disso, em condições de
estresse hídrico suave, a atividade da NR declina rapidamente comparada com outras enzimas
assimilatórias de nitrogênio. A menor atividade da NR em plantas sob défice hídrico se deve
ao decréscimo no fluxo de substrato (NO3-) por falta de umidade suficiente para sua absorção,
o que pode ter ocorrido nesta avaliação (CROCOMO, 1985).
Na avaliação feita aos 60 DAP as estratégias hídricas tiveram
comportamento semelhante na atividade da NR, porém com queda acentuada em seus valores
em comparação aos 54 DAP. A NR em plantas superiores é uma enzima complexa e
extremamente sensível às variações ambientais, ao substrato e ao produto final (NH3)
(MARSCHNER, 1995). Não muito diferente ao ocorrido na avaliação anterior, o défice
hídrico pode ter causado redução no conteúdo dessa enzima na folha, assim como, o fluxo de

87
nitrato das raízes até as folhas. A atividade da NR é induzida pelo seu substrato (NO3-), além
de possuir meia-vida curta, de poucas horas, a sua atividade varia diurnamente e com os
fatores ambientais que afetam o fluxo de nitrato para os sítios de indução e assimilação
(BEEVERS e HAGEMAN, 1983; SOLOMONSON e BARBER, 1990; HOFF et al., 1992).
Apesar de não ter sido mensurado, mas considerando que a nitrato redutase é uma enzima cuja
síntese e atividade são induzidas pelo substrato (BEEVERS e HAGEMAN, 1969) é possível
também que o nitrato interno (substrato) tenha influenciado na atividade da enzima em todos
os tratamentos.
Tabela 18. Atividade da enzima nitrato redutase (NR, μmol de NO2 g MF h-1
) em folhas de
videira ‘Syrah’ ao longo do ciclo em função de três estratégias de irrigação (Irrigação com
défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido –
Petrolina, PE, 2010.
Dias após a poda Estratégias de irrigação
IDC ID IP
54 12,25 Ba 13,74 Aa 13,74 Aa
60 8,41 Ab 8,65 Ab 8,17 Ac
73 8,14 Ab 6,80 Bc 7,64 ABc
87 8,48 Bb 9,06 ABb 9,87 Ab
101 7,33 Bb 6,41 Bc 8,74 Abc
Média 8,92 B 8,93B 9,63 A
CV (%) 6,05
Médias seguidas de mesma letra minúsculas na coluna e maiúsculas na linha, não diferem
entre si pelo teste Tukey a 5% de probabilidade.
Aos setenta e três dias após a poda notou-se diferença significativa
entre as estratégias hídricas, obtendo-se superioridade na atividade da enzima para o
tratamento IDC em relação ao tratamento ID. Esse resultado pode estar vinculado às condições
atmosféricas no momento da coleta, em que o tempo estava nublado e a insolação registrada

88
no dia foi relativamente baixa e oscilante, com apenas 3,7 horas. A luz e o teor de NO3- são
fatores importantes na indução e manutenção da atividade da NR nos tecidos vegetais. A luz
(LILLO, 1994) e o substrato (HEWITT et al., 1976) estimulam à síntese de novo da NR,
enquanto que o défice moderado, da ordem de -0,8 a -2,0 MPa , pode reduzir sua produção em
20%, chegando a 50% quando a planta sofre estresse intenso (HSIAO, 1979). O potencial
hídrico foliar no presente estudo não foi acompanhado ao longo do dia; no entanto, com os
dados de evapotranspiração da cultura registrados pela estação meteorológica do campo
experimental de Bebedouro, juntamente com o seu potencial hídrico foliar de base, sugere-se
que as plantas podem ter atingido valores próximos aos preconizados por Hsiao (1979), o que
pode ter influenciado na menor atividade da NR registrada no tratamento IDC aos 54 DAP e
ID aos 73 e 101 DAP. Além disso, o tratamento ID dispunha de menor quantidade de água no
solo (28 dias sem irrigação) disponível, o que dificulta também o transporte do nitrato das
raízes até o citossol das células foliares, gerando menor valor da atividade da NR nessas
avaliações. Em plantas superiores, a redução do NO3- ocorre, principalmente, nas células do
mesofilo, onde a nitrato redutase se localiza no citoplasma e a nitrito redutase no cloroplasto.
Tal redução ocorre também em raízes e outros tecidos não fotossintetizantes, como em
tremoço, onde a ferrodoxina funciona como doadora de elétrons. Essa redução de NO3- a NH3
ocorre, preferencialmente, em tecidos fotossintetizantes devido à grande demanda de elétrons,
que são fornecidos pelos fotossistemas, na presença de luz. Assim, a assimilação de NO3-
depende da fotossíntese para fornecimento de elétrons, pelo fotossistema I, para a redução do
NO3-, competindo com o CO2 pelo poder redutor (CLARK, 1991).
Quando as plantas foram avaliadas aos 87 DAP notou-se que o
tratamento ID não se distinguiu estatisticamente dos demais tratamentos, no entanto, o
tratamento IP diferiu estatisticamente do IDC, apresentando 14% a mais de atividade da
enzima nitrato redutase. Essa superioridade do tratamento IP em relação ao IDC pode ser
devido a sua maior disponibilidade de água aos processos fisiológicos e metabólicos das
plantas em estudo. A atividade da NR é aumentada, rapidamente, pela presença do substrato
inicial, o NO3-, pelo aumento da luz e temperatura, sendo reduzida pelo acúmulo do produto
final, o NH3, pelo estresse hídrico ou térmico e pela obscuridade (KRAMER e BOYER,
1995). O aumento da atividade do sistema é devido à síntese de novo das enzimas e a redução
pela sua degradação, sendo que sua atividade é máxima quando a expansão foliar é máxima,

89
assim como para a atividade fotossintética, decrescendo após, com a redução dessa atividade
(GUPTA e BEEVERS, 1985).
Na última avaliação da atividade da nitrato redutase (101 DAP) pode-
se perceber superioridade de 19 e 36% da estratégia de irrigação IP em relação às demais
estratégias hídricas, IDC e ID, respectivamente. A constância na manutenção da lâmina de
água às plantas pode ter contribuído para o bom desempenho do primeiro. Os tratamentos ID e
IDC apresentaram as menores atividade da nitrato redutase, indicando que a falta do recurso
água afeta a síntese e o funcionamento da enzima, além de afetar o deslocamento do seu
substrato (NO3-) do solo para o citossol das células foliares, onde serão metabolizados.
Conforme os dados apresentados na Tabela 18, pode-se observar que a
atividade da nitrato redutase diminui com o tempo de avaliação. Isto pode estar relacionado
com a idade da folha que é outro fator que influencia a redução do nitrato. Segundo Marrenco
e Lopes (2009) a atividade da NR aumenta com a ontogenia da folha, atingindo o pico de
maior atividade quando a taxa de expansão foliar é máxima. A partir desse momento, a
atividade da NR começa a diminuir, mostrando menor atividade em folhas completamente
expandidas. Isso pode ser verificado em todos os tratamentos ao longo do ciclo.
Os resultados aqui obtidos discordam do encontrados por Pereira et al.
(2006) que trabalharam com videira no SVSF e não registraram efeito dos diferentes manejos
de irrigação (RDI e PRD) utilizados e dos diferentes porta-enxertos na atividade da nitrato
redutase.
4.5 Proteína solúvel total
Na Tabela 19 encontram-se os dados referentes ao resumo da análise
de variância para o teor de proteína solúvel total (μg g-1
MF) aos 54, 60, 73, 87, 101 e 115 dias
após a poda. Os resultados mostram que houve resposta significativa entre os diferentes dias
após a poda.
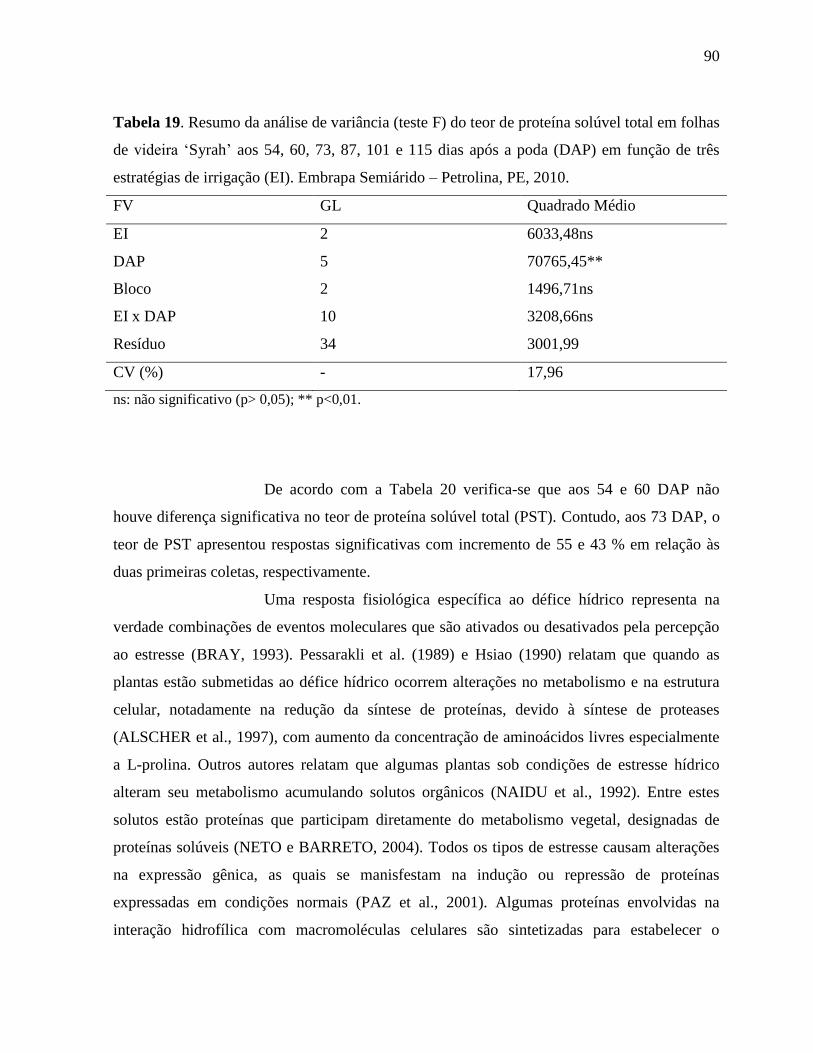
90
Tabela 19. Resumo da análise de variância (teste F) do teor de proteína solúvel total em folhas
de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda (DAP) em função de três
estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE, 2010.
FV GL Quadrado Médio
EI 2 6033,48ns
DAP 5 70765,45**
Bloco 2 1496,71ns
EI x DAP 10 3208,66ns
Resíduo 34 3001,99
CV (%) - 17,96
ns: não significativo (p> 0,05); ** p<0,01.
De acordo com a Tabela 20 verifica-se que aos 54 e 60 DAP não
houve diferença significativa no teor de proteína solúvel total (PST). Contudo, aos 73 DAP, o
teor de PST apresentou respostas significativas com incremento de 55 e 43 % em relação às
duas primeiras coletas, respectivamente.
Uma resposta fisiológica específica ao défice hídrico representa na
verdade combinações de eventos moleculares que são ativados ou desativados pela percepção
ao estresse (BRAY, 1993). Pessarakli et al. (1989) e Hsiao (1990) relatam que quando as
plantas estão submetidas ao défice hídrico ocorrem alterações no metabolismo e na estrutura
celular, notadamente na redução da síntese de proteínas, devido à síntese de proteases
(ALSCHER et al., 1997), com aumento da concentração de aminoácidos livres especialmente
a L-prolina. Outros autores relatam que algumas plantas sob condições de estresse hídrico
alteram seu metabolismo acumulando solutos orgânicos (NAIDU et al., 1992). Entre estes
solutos estão proteínas que participam diretamente do metabolismo vegetal, designadas de
proteínas solúveis (NETO e BARRETO, 2004). Todos os tipos de estresse causam alterações
na expressão gênica, as quais se manisfestam na indução ou repressão de proteínas
expressadas em condições normais (PAZ et al., 2001). Algumas proteínas envolvidas na
interação hidrofílica com macromoléculas celulares são sintetizadas para estabelecer o

91
metabolismo e agir na recuperação dos danos causados pelo estresse hídrico (LOBATO et al.,
2008).
O défice hídrico pode produzir efeitos no metabolismo das plantas,
levando ao acúmulo ou perda de metabólitos como carboidratos, ácidos orgânicos, ácido
abscísico (ABA) e aminoácidos, além de alterar a atividade de enzimas e síntese de proteínas
(MARANVILLE e PAULSEN, 1970). O acúmulo de metabólitos em plantas (ajustamento
osmótico) sob défice hídrico proporciona redução do potencial osmótico mediante aumento
líquido na concentração de solutos intracelulares (CHAVES, 1991). Esse ajustamento pode
auxiliar a planta a manter o turgor, sustentando, dessa maneira, o influxo de água para o
crescimento celular (PREMACHANDRA et al., 1992).
Santos et al. (2010) concordando com as informações dos autores
citados no parágrafo anterior, observaram aumento de proteínas em genótipos de feijão sob
deficiência hídrica e atribuíram à alteração do potencial osmótico da célula para evitar a perda
de água para o meio. A síntese de osmólitos, incluindo prolina, é amplamente usada por
plantas para estabilizar as membranas e manter a conformação das proteínas sob baixo
potencial hídrico (EFEOGLU et al., 2008).
A síntese e o acúmulo de solutos orgânicos e/ou inorgânicos variam
entre espécies vegetais, assim como em diferentes cultivares da mesma espécie. A prolina é
também conhecida por estar envolvida em reduzir danos nos tilacóides das membranas, agindo
como um eliminador de radicais livres (REDDY et al., 2004), além de marcador bioquímico
entre espécies e para genótipos dentro da mesma espécie sob estresse hídrico (SANTOS et al.,
2010).
O resultado obtido aos 73 DAP indicou que nessa avaliação a
produção de compostos nitrogenados foi significativamente, mais eficiente que todas as outras
datas de coletas. O Ψb mais próximo dessa avaliação, aos 70 DAP (-0,243 MPa) indicou que
as plantas estavam sob défice hídrico leve. Segundo Kirschbaum (1987, 1988) trabalhando
com Eucalyptus pauciflora sugeriu que a recuperação de plantas sob défice hídrico requer a
síntese de novo de proteínas fotossintéticas. Nessa mesma vertente, Bogeat-Triboulot et al.
(2007) mostraram que a recuperação ao défice hídrico para plantas de Populus euphratica foi
acompanhada pelo aumento de algumas proteínas fotossintéticas, particularmente a Rubisco.
Acredita-se que plantas no presente estudo aos 73 DAP tenham tido comportamento

92
semelhante ao das duas espécies vegetais citadas anteriormente. Uma maior síntese de
proteínas fotossintéticas pode ser confirmada com a geração de maior atividade fotossintética.
Isto ocorreu aos 73 DAP com a média da taxa de assimilação líquida de 17,53 μmol m-2
s-1
(dados não apresentados), registrando incremento fotossintético de aproximadamente 29 % em
relação à coleta anterior. Este resultado também condiz com a fase fenológica, uma vez que a
partir dessa data já se verificava início do ‘pintor’ que é uma fase de grande requisito
metabólico para as folhas.
De acordo com a Tabela 20, o teor de PST nas folhas em relação às
diferentes estratégias de irrigação aumentou até o início da fase fenológica de ‘pintor’ (73
DAP), com tendência decrescente após essa fase. Este resultado está relacionado com os
maiores valores de assimilação de CO2 obtidos durante essa avaliação. Pois, entre as proteínas
solúveis totais presentes nas folhas, a Rubisco é uma das mais expressivas. O aumento no teor
de proteínas do complexo fotossintético pode justificar as altas taxas significativas de
fotossíntese registradas para todas as estratégias hídricas nessa avaliação.
A partir dos 87 até os 115 DAP, mesmo com as variações no Ψb
(Tabela 2), não houve diferença significativa no teor de PST nessas coletas. Alguns autores
afirmam que o teor de proteínas pode aumentar com a síntese de novo (KIRSCHBAUM, 1987;
1988; BOGEAT-TRIBOULOT et al., 2007) ou diminuir pela ação de proteases ou proteólise
(DEBOUBA et al., 2006; KALA e GODORA, 2011) em função do défice hídrico. Além disso,
o consumo ou produção de proteínas vai estar relacionado ao nível de estresse, sendo a
expressão gênica o fator determinante para a recuperação ao estresse de uma forma menos
dispendiosa à manutenção do crescimento e desenvolvimento celular.

93
Tabela 20. Teor de proteína solúvel total (μg g-1
MF) em folhas de videira ‘Syrah’ ao longo
do ciclo em função de três estratégias de irrigação (Irrigação com défice controlado-IDC,
Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE, 2010.
Dias após a poda Estratégia de irrigação
IDC ID IP Média
54 168,66 244,13 226,64 213,14 c
60 281,22 283,92 248,47 271,20 bc
73 443,54 550,00 429,69 474,41 a
87 309,27 270,07 255,40 278,25 bc
101 294,49 297,42 269,72 287,21 bc
115 315,77 302,53 300,56 306,29 b
Média 302,16 324,68 288,41
CV (%) 17,96
Médias seguidas de mesma letra, não diferem entre si pelo teste Tukey a 5% de probabilidade.
4.6 Açúcar solúvel total e açúcar redutor
Os teores de glicose, frutose e sacarose, bem como a atividade de
enzimas do metabolismo de carboidratos em folhas de videiras, são influenciados por
variações sazonais e fenológicas das videiras (PALLIOTTI e CARTECHINI, 2001).
Os açúcares provenientes da fotossíntese agem como substrato para o
metabolismo energético e biossíntese de carboidratos, fornecendo condições de crescimento e
desenvolvimento aos tecidos dreno. Além disso, os açúcares podem funcionar como
mensageiros secundários assegurando que a planta continue a se desenvolver, mesmo após
estresses bióticos ou abióticos (HAMMOND e WHITE, 2008).
Encontram-se na Tabela 21, os dados referentes ao resumo da análise
de variância de açúcar solúvel total e açúcar redutor em folhas de videira ‘Syrah’ aos 54, 60,
73, 87, 101 e 115 dias após a poda. Notou-se significância apenas para os diferentes dias de
coleta de dados após a poda. As plantas das três estratégias hídricas apresentaram aumento

94
semelhante no teor de açúcares solúveis totais e redutores em suas folhas na fase de maturação
de frutos ao longo das avaliações. Esse aumento gradual nos açúcares era esperado como já
definido por muitos autores (MULLINS et al., 2000; JAIN et al., 2002; DANTAS et al., 2005;
2007a; 2007b; LIMA FILHO et al., 2009) em relação ao crescimento e desenvolvimento dos
frutos da videira em função do acúmulo de fotoassimilados provenientes das folhas.
Tabela 21. Resumo da análise de variância (teste F) de açúcar solúvel total (AST) e açúcar
redutor (AR) em folhas de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda
(DAP) em função de três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE,
2010.
FV GL Quadrado Médio
AST AR
EI 2 19277,97ns 668,45ns
DAP 5 969547,47** 17403,81**
Bloco 2 43,048,90ns 783,37ns
EI x DAP 10 16953,63ns 320,12ns
Resíduo 34 28118,71 426,90
CV (%) - 9,66 11,52
ns: não significativo (p> 0,05); ** p<0,01.
As diferentes estratégias hídricas estudadas não diferenciaram
estatisticamente entre si. Outros autores (BASSOI et al., 2007; DANTAS et al., 2007b;
SOUZA et al., 2009) avaliando na mesma região diferentes estratégias de irrigação e porta-
enxertos não encontraram influências dos diferentes manejos nas características avaliadas.
Desta forma, as restrições hídricas impostas às plantas através dos tratamentos IDC e ID não
foram limitantes à produção de açúcares pelas folhas.
Os maiores valores médios de açúcar solúvel total (AST) nas folhas da
videira foram obtidos nas duas últimas coletas (101 e 115 DAP) coincidindo com a fase de
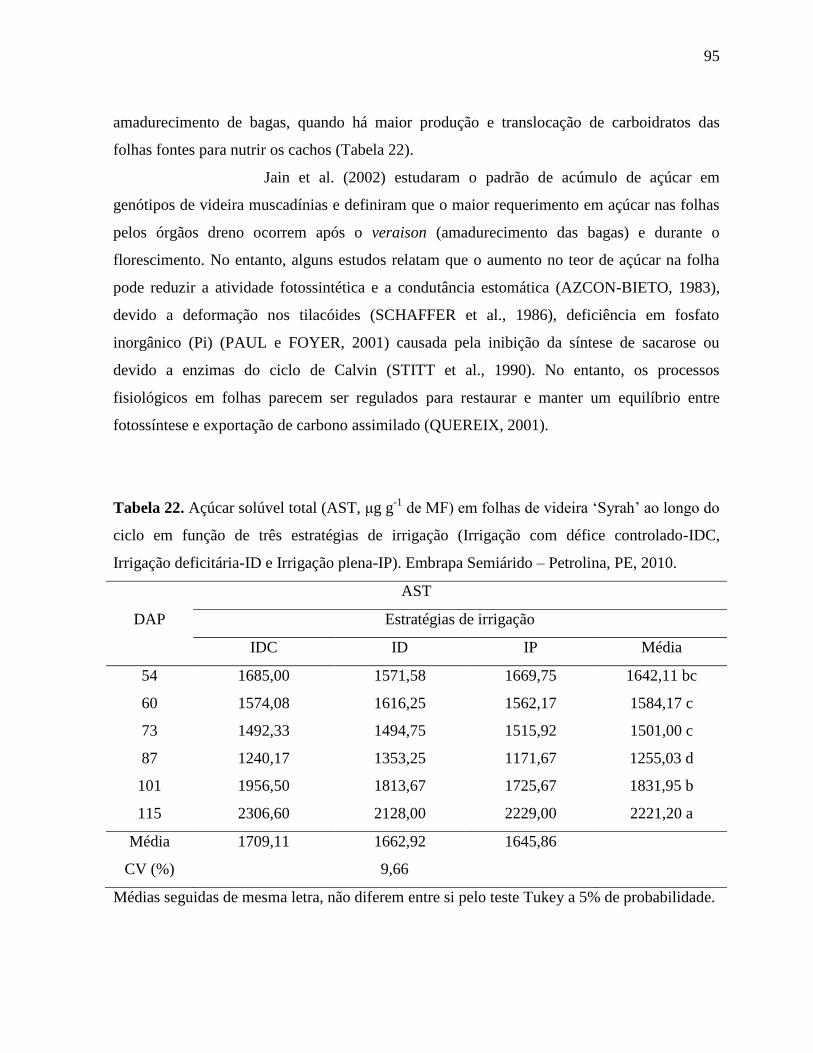
95
amadurecimento de bagas, quando há maior produção e translocação de carboidratos das
folhas fontes para nutrir os cachos (Tabela 22).
Jain et al. (2002) estudaram o padrão de acúmulo de açúcar em
genótipos de videira muscadínias e definiram que o maior requerimento em açúcar nas folhas
pelos órgãos dreno ocorrem após o veraison (amadurecimento das bagas) e durante o
florescimento. No entanto, alguns estudos relatam que o aumento no teor de açúcar na folha
pode reduzir a atividade fotossintética e a condutância estomática (AZCON-BIETO, 1983),
devido a deformação nos tilacóides (SCHAFFER et al., 1986), deficiência em fosfato
inorgânico (Pi) (PAUL e FOYER, 2001) causada pela inibição da síntese de sacarose ou
devido a enzimas do ciclo de Calvin (STITT et al., 1990). No entanto, os processos
fisiológicos em folhas parecem ser regulados para restaurar e manter um equilíbrio entre
fotossíntese e exportação de carbono assimilado (QUEREIX, 2001).
Tabela 22. Açúcar solúvel total (AST, μg g-1
de MF) em folhas de videira ‘Syrah’ ao longo do
ciclo em função de três estratégias de irrigação (Irrigação com défice controlado-IDC,
Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE, 2010.
AST
DAP Estratégias de irrigação
IDC ID IP Média
54 1685,00 1571,58 1669,75 1642,11 bc
60 1574,08 1616,25 1562,17 1584,17 c
73 1492,33 1494,75 1515,92 1501,00 c
87 1240,17 1353,25 1171,67 1255,03 d
101 1956,50 1813,67 1725,67 1831,95 b
115 2306,60 2128,00 2229,00 2221,20 a
Média 1709,11 1662,92 1645,86
CV (%) 9,66
Médias seguidas de mesma letra, não diferem entre si pelo teste Tukey a 5% de probabilidade.

96
Durante a fase de maturação, a sacarose é convertida em
monossacarídeos que vão se acumulando progressivamente nas bagas (ROBINSON e
DAVIES, 2000; BOSS e DAVIES, 2001; VINE et al., 2002; ALI et al., 2010). A relação entre
a fotossíntese e a disponibilidade hídrica foi variável no presente estudo, descartando a
hipótese de que quanto maior a disponibilidade de água maior a produção e qualidade de
frutos para o processamento. No entanto, esperava-se que o tratamento IP tivesse maior
influência na concentração de açúcares nas folhas e/ou frutos, considerando suas maiores taxas
fotossintéticas, mas isso não foi verificado. Confirmando assim, a importância do uso do
défice hídrico nos atributos de qualidade em videiras, sendo um dos assuntos mais pesquisados
em vinhedos em todas as regiões do globo terrestre com características semiáridas a áridas.
Todavia, a importância do uso da deficiência hídrica nas plantas vem como uma forma de
estimulá-las a metabolizarem com o máximo de eficiência e economia dos recursos hídricos
não limitantes.
Na irrigação com défice hídrico controlado a quantidade de açúcar
solúvel total variou de 1685 μg g-1
MF aos 54 DAP a 2306 μg g-1
MF no dia da colheita, o que
representou incremento ao longo do ciclo de 27%. No entanto, comparando o resultado obtido
aos 87 DAP com o da colheita se observa incrementos de até 46% entre essas datas. Isso
indica que o teor de açúcares solúveis totais nas folhas ao longo do ciclo é muito variável, pois
dependerá da fase fenológica da planta, o seu estado nutricional e as condições atmosféricas.
A irrigação plena promoveu aumentos de até 47% na quantidade de
açúcar solúvel total nas folhas durante o ciclo, variando de 1171,67 μg g-1
MF (87 DAP) a
2229 μg g-1
MF (115 DAP). A irrigação deficitária promoveu incrementos menores (36%) em
relação aos outros tratamentos.
Os principais açúcares redutores presentes nas folhas e frutos da
videira são a glicose e a frutose, encontrados, principalmente, na fase de maturação.
De acordo com a Tabela 23, observa-se aumento nos valores médios
de açúcares redutores nas folhas ao longo do ciclo em todas as três estratégias hídricas
estudadas, com seu ápice na última avaliação realizada aos 115 DAP.

97
Tabela 23. Açúcar redutor (AR, μg g-1
de MF) em folhas de videira ‘Syrah’ ao longo do ciclo
em função de três estratégias de irrigação (Irrigação com défice controlado-IDC, Irrigação
deficitária-ID e Irrigação plena-IP). Embrapa Semiárido – Petrolina, PE, 2010.
AR
DAP Estratégias de irrigação
IDC ID IP Média
54 185,32 177,61 181,49 181,51 b
60 168,02 175,91 166,09 170,01 b
73 157,69 162,93 156,94 159,19 bc
87 140,21 152,16 120,84 137,74 c
101 174,01 163,93 153,42 163,79 bc
115 284,94 248,82 258,78 264,18 a
Média 185,03 180,22 172,93
CV (%) 11,52
Médias seguidas de mesma letra, não diferem entre si pelo teste Tukey a 5% de probabilidade.
As diferentes disponibilidades de água às plantas durante o ciclo não
apresentaram diferenças significativas em cada uma das avaliações realizadas até 115 DAP.
Contudo, quando se analisou as estratégias hídricas separadamente nas seis datas distintas,
notou-se que os mesmos influenciaram no maior ou menor volume de açúcares redutores nas
folhas. Isso era esperado, partindo do princípio de que com o avanço do ciclo fenológico das
plantas o volume de carboidratos requisitados pelos diferentes órgãos vegetais aumenta o que
demanda maior consumo hídrico.
A disponibilidade hídrica fornecida pelos tratamentos IDC, ID e IP
entre 54 e 115 DAP contribuíram com cerca de 35, 29 e 30%, respectivamente, no aumento do
volume de açúcares redutores nas folhas ao longo do ciclo. Notou-se também, que os maiores
incrementos no conteúdo de açúcares redutores ocorreram entre os 87 e 115 DAP nas três
estratégias de irrigação, com acréscimos de 53, 51 e 40% na IP, IDC e ID, respectivamente.

98
Esses resultados aparentemente discordantes refletem a complexidade
dos mecanismos de ação da disponibilidade hídrica em relação à concentração de açúcares. O
tratamento IP ao final do ciclo apresentou conteúdos de açúcares semelhantes aos demais
tratamentos com restrições hídricas, podendo boa parte dos seus fotoassimilados terem sido
desviados para a atividade vegetativa e/ou armazenamento nos ramos, raízes e braços durante
o período de pós-veraison, desviando assim, os açúcares que se acumulariam nas bagas
(ACEVEDO et al., 2005; KELLER, 2005; KOUNDOURAS et al., 2006). Essa afirmação
ainda pode ser confirmada por não ter havido diferença no teor de sólidos solúveis (Tabela 28)
entre as estratégias hídricas na colheita e um maior crescimento vegetativo para o tratamento
IP foi visualizado na área experimental. Este resultado confirma a importância do uso de
técnicas com défice hídrico, onde pode-se obter bons resultados de qualidade na produção com
economia e eficiência no uso da água.
O período de 54 a 87 DAP coincidiu com as maiores taxas de
fotossíntese, consequentemente, maior exportação de carboidratos produzidos para os
principais drenos (meristema apical, cacho e ramos). Desta forma, era de se esperar menor
acúmulo nas folhas durante esse período. Esses resultados estão de acordo com os obtidos por
Dantas et al. (2005; 2007a), onde o acúmulo de açúcares nas folhas aumenta durante o início
da maturação, porém, é fortemente influenciado pelas variações climáticas.
Desta forma, o défice hídrico suave a moderado pode influenciar na
arquitetura da planta, reduzindo o seu porte e/ou crescimento dos ramos, promovendo melhor
aproveitamento das trioses-P geradas no ciclo de Calvin para constituir os carboidratos de
reserva e estruturais, além de compostos nitrogenados essenciais para a constituição e ativação
enzimática.
4.7 Invertases: neutra do citossol (INC), ácida do vacúolo (IAV) e ácida da parede
celular (IAP)
As invertases são isoenzimas responsáveis pela hidrólise da sacarose,
principal carboidrato translocável nas plantas superiores, em D-glicose e D-frutose que
desempenham papel fundamental no metabolismo primário do carbono (BARRATT et al.,
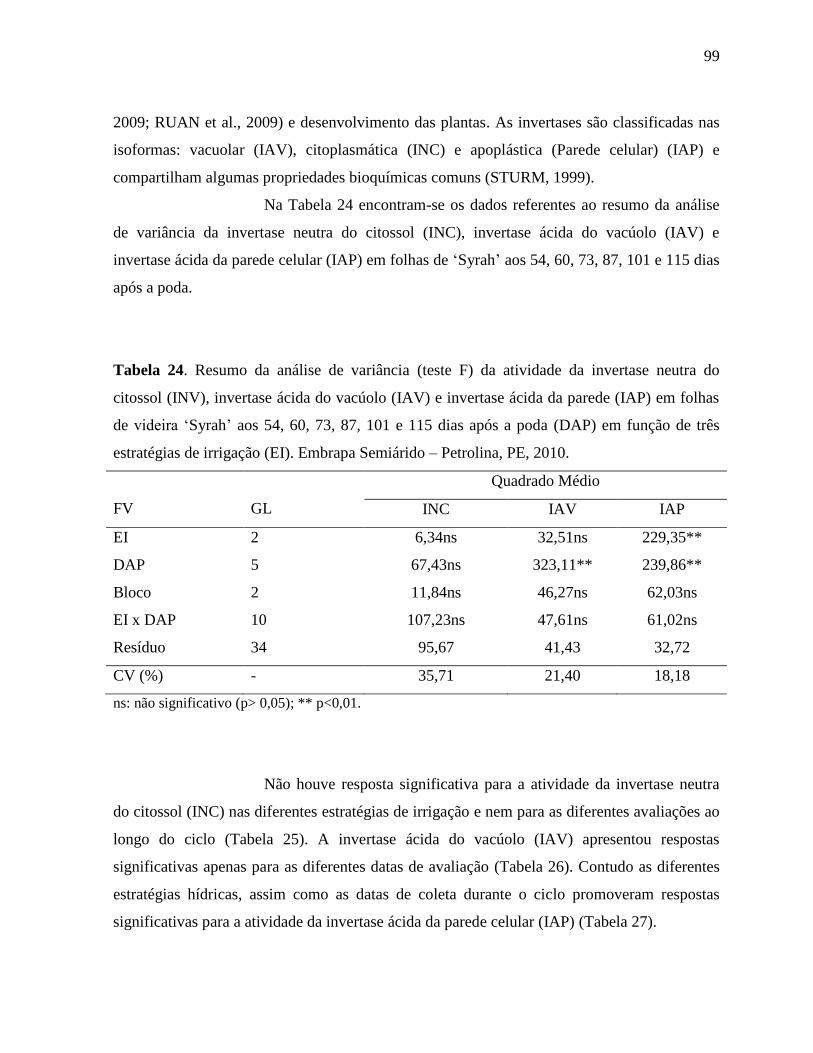
99
2009; RUAN et al., 2009) e desenvolvimento das plantas. As invertases são classificadas nas
isoformas: vacuolar (IAV), citoplasmática (INC) e apoplástica (Parede celular) (IAP) e
compartilham algumas propriedades bioquímicas comuns (STURM, 1999).
Na Tabela 24 encontram-se os dados referentes ao resumo da análise
de variância da invertase neutra do citossol (INC), invertase ácida do vacúolo (IAV) e
invertase ácida da parede celular (IAP) em folhas de ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias
após a poda.
Tabela 24. Resumo da análise de variância (teste F) da atividade da invertase neutra do
citossol (INV), invertase ácida do vacúolo (IAV) e invertase ácida da parede (IAP) em folhas
de videira ‘Syrah’ aos 54, 60, 73, 87, 101 e 115 dias após a poda (DAP) em função de três
estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE, 2010.
FV
GL
Quadrado Médio
INC IAV IAP
EI 2 6,34ns 32,51ns 229,35**
DAP 5 67,43ns 323,11** 239,86**
Bloco 2 11,84ns 46,27ns 62,03ns
EI x DAP 10 107,23ns 47,61ns 61,02ns
Resíduo 34 95,67 41,43 32,72
CV (%) - 35,71 21,40 18,18
ns: não significativo (p> 0,05); ** p<0,01.
Não houve resposta significativa para a atividade da invertase neutra
do citossol (INC) nas diferentes estratégias de irrigação e nem para as diferentes avaliações ao
longo do ciclo (Tabela 25). A invertase ácida do vacúolo (IAV) apresentou respostas
significativas apenas para as diferentes datas de avaliação (Tabela 26). Contudo as diferentes
estratégias hídricas, assim como as datas de coleta durante o ciclo promoveram respostas
significativas para a atividade da invertase ácida da parede celular (IAP) (Tabela 27).

100
Tabela 25. Atividade da enzima invertase neutra do citossol (INC, µmol g-1
MF h-1
) em folhas
de videira ‘Syrah’ ao longo do ciclo em função de três estratégias de irrigação (Irrigação com
défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido –
Petrolina, PE, 2010.
INC
DAP Estratégias de irrigação
IDC ID IP Média
54 24,03 23,85 20,58 22,82
60 23,39 31,09 30,73 28,40
73 29,54 26,23 28,14 27,97
87 17,95 24,75 33,99 25,56
101 37,66 31,07 21,73 30,15
115 32,48 23,51 32,37 29,45
Média 27,50 26,75 27,92
CV (%) 35,71
Diferentemente da IAV e IAP poucas informações sobre o papel da
INC nas plantas estão disponíveis na literatura. Alguns pesquisadores atribuem isso à sua alta
instabilidade e baixa atividade (RUAN et al., 2003). Entretanto, estudos recentes revelam que
a INC possa compensar a perda da atividade da sacarose sintase em mutantes de Arabidopsis
(BARRATT et al., 2009); no entanto, em drenos fortes como sementes de milho (CHOUREY
et al., 1998) e fibras de algodão (RUAN et al., 2003) isso não foi verificado.
De acordo com a Tabela 26 os maiores valores médios significativos
(p<0,01) para a atividade da IAV foram obtidos na colheita aos 115 DAP, com incrementos de
41 e 33% em relação aos 60 e 87 DAP. Porém, a atividade da enzima aos 115 DAP não se
diferenciou estatisticamente das coletas aos 54 e 101 DAP. Esse comportamento oscilante da
atividade da enzima é esperado, pois os processos bioquímicos são dinâmicos e podem ser
afetados, instantaneamente, por muitos fatores endógenos e ambientais. No entanto, esses
resultados podem indicar que durante o período de avaliação, a maior requisição da atividade
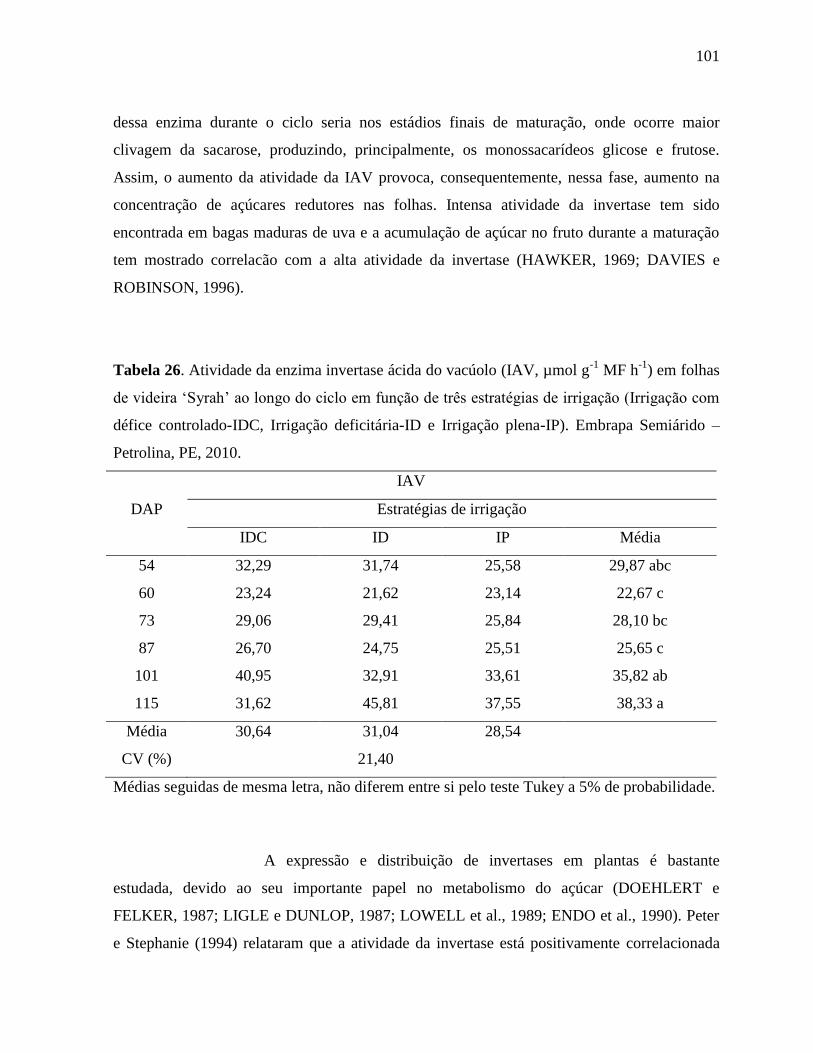
101
dessa enzima durante o ciclo seria nos estádios finais de maturação, onde ocorre maior
clivagem da sacarose, produzindo, principalmente, os monossacarídeos glicose e frutose.
Assim, o aumento da atividade da IAV provoca, consequentemente, nessa fase, aumento na
concentração de açúcares redutores nas folhas. Intensa atividade da invertase tem sido
encontrada em bagas maduras de uva e a acumulação de açúcar no fruto durante a maturação
tem mostrado correlacão com a alta atividade da invertase (HAWKER, 1969; DAVIES e
ROBINSON, 1996).
Tabela 26. Atividade da enzima invertase ácida do vacúolo (IAV, µmol g-1
MF h-1
) em folhas
de videira ‘Syrah’ ao longo do ciclo em função de três estratégias de irrigação (Irrigação com
défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido –
Petrolina, PE, 2010.
IAV
DAP Estratégias de irrigação
IDC ID IP Média
54 32,29 31,74 25,58 29,87 abc
60 23,24 21,62 23,14 22,67 c
73 29,06 29,41 25,84 28,10 bc
87 26,70 24,75 25,51 25,65 c
101 40,95 32,91 33,61 35,82 ab
115 31,62 45,81 37,55 38,33 a
Média 30,64 31,04 28,54
CV (%) 21,40
Médias seguidas de mesma letra, não diferem entre si pelo teste Tukey a 5% de probabilidade.
A expressão e distribuição de invertases em plantas é bastante
estudada, devido ao seu importante papel no metabolismo do açúcar (DOEHLERT e
FELKER, 1987; LIGLE e DUNLOP, 1987; LOWELL et al., 1989; ENDO et al., 1990). Peter
e Stephanie (1994) relataram que a atividade da invertase está positivamente correlacionada

102
com o acúmulo de açúcares redutores em tomates. Em estudos anteriores realizados por
Arnold (1965a; 1965b) em uvas brancas notou que a invertase se encontrava sob duas formas:
a ligada e a solúvel. Atualmente, é sabido que as plantas apresentam três isoformas de
invertases, a IAV, IAP e INC, todas com diferentes localizações nas células, mas, com
propriedades bioquímicas comuns (STURM, 1999).
Geralmente, acredita-se que IAV possa controlar a expansão celular
pela regulação osmótica (ROITSCH e GONZÁLEZ, 2004; SERGEEVA et al., 2006). Isto é
conseguido pela hidrólise da sacarose em duas moléculas de monossacarídeos, duplicando
assim, na contribuição osmótica, facilitando o influxo de água para impulsionar a expansão
das células. Essa pode ser também uma das alternativas das plantas sob défice hídrico, para
regular seu potencial hídrico interno. Isto pode ter ocorrido com maior frequência nos
tratamentos IDC e ID durante o ciclo.
A IAV hidrolisa a sacarose no vacúolo e sua atividade tem alta
correlação com o acúmulo de hexoses em vários órgãos, além de controlar a composição do
açúcar, como registrado para tomate (KLANN et al., 1996), uva (DAVIES e ROBINSON,
1996) e tubérculo de batata (GREINER et al., 1999). Alta expressão ou atividade da IAV tem
sido relatada na expansão de tecidos de tubérculo de batata (ROSS et al., 1994), raízes de
cenoura (TANG et al., 1999), ovários de milho (ANDERSEN et al., 2002) e raízes e
hipocótilos de Arabidopsis (SERGEEVA, 2006). A invertase não tem apenas o papel de suprir
em carbono as plantas, mas também na sinalização do açúcar e no desenvolvimento (RUAN et
al., 2010).
Conforme a Tabela 27 verificou-se que os valores médios da atividade
da IAP para as estratégias de irrigação estabelecidas se diferenciaram estatisticamente ao final
do ciclo, com destaque para a irrigação deficitária que apresentou 13 e 20% a mais de
atividade da enzima em relação às estratégias IDC e IP na devida ordem. Além disso, no
tratamento ID após o pico (50 µmol g-1
MF h-1
) de atividade, iniciou-se nas avaliações
posteriores, queda progressiva até a colheita. As invertases possuem limitado envolvimento no
crescimento radicular, mas atuam na osmorregulação, expansão celular e canalização da
sacarose no metabolismo (TANG et al., 1999). Este fato poderia caracterizar a maior atividade
da IAP aos 60 DAP na estratégia de irrigação ID em relação aos demais tratamentos. Durante
o descarregamento do floema nos drenos, a sacarose pode ser transportada pela via apoplástica

103
(através da matriz da parede celular) ou simplástica (através dos plasmodesmos) sendo a IAP
uma das mais ativas. Alguns trabalhos relatam a importância da IAP no desenvolvimento
floral (GOETZ et al., 2001; OLIVER et al., 2007; ZANOR et al., 2009), de sementes
(PATRICK e OFFLER, 2001; WEBER, et al., 2005; WANG et al., 2008; JIN et al., 2009) e
frutos (FRIDMANN et al., 2004; HAYES et al., 2007; ZANOR et al., 2009; JIN et al., 2009).
Além disso, algumas pesquisas relatam que a alta atividade da IAP é um componente essencial
para o atraso da senescência foliar mediada pelo hormônio vegetal citocinina (LARA et al.,
2004). Isso pode ter ocorrido no presente experimento, em função dos valores médios da
atividade da IAP aos 54 e aos 115 DAP não terem se diferenciado estatisticamente. Todavia
estudos recentes de Jin et al. (2009) revelaram que a redução da atividade da IAP pode levar a
uma baixa relação sacarose/hexose no apoplasto foliar podendo provocar ou permitir a
senescência, devido a expressão do gene da protease-CYS induzido pelo ABA.

104
Tabela 27. Atividade da enzima invertase ácida da parede (IAP, µmol g-1
MF h-1
) em folhas
de videira ‘Syrah’ ao longo do ciclo em função de três estratégias de irrigação (Irrigação com
défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido –
Petrolina, PE, 2010.
IAP
DAP Estratégias de irrigação
IDC ID IP Média
54 27,19 25,74 21,88 24,94 c
60 29,95 50,00 27,89 35,95 ab
73 40,66 39,47 36,62 38,92 a
87 29,12 33,79 28,72 30,54 bc
101 26,81 30,31 26,27 27,80 c
115 29,90 33,00 29,05 30,65 bc
Média 30,64 B 35,39 A 28,40 B
CV (%) 18,18
Médias seguidas de mesma letra minúsculas na coluna e maiúsculas na linha, não diferem
entre si pelo teste Tukey a 5% de probabilidade.
O tratamento IP teve comportamento parecido ao anterior, onde após
ter registrado no decorrer do ciclo o menor valor da atividade da enzima IAP (21,88 µmol g-1
MF h-1
) aos 54 DAP, apresentou comportamento variável nos demais dias de avaliação,
obtendo o seu pico (36,62 µmol g-1
MF h-1
) aos 73 DAP, na qual a data coincidiu com
intervalos de maiores potenciais hídricos foliares de base (Tabela 2) e taxas de assimilação
para esse tratamento (Tabela 4).
O processo de acumulação de açúcares em bagas de uva ainda não está
completamente elucidado e, embora, a invertase solúvel possa ter importante papel na
acumulação de hexoses no vacúolo, a sua síntese não parece estar relacionada a esse acúmulo
(DAVIES e ROBINSON, 1996; DREIER et al., 1998). Outros mecanismos devem estar
envolvidos na regulação do acúmulo de açúcares, tais como a diferença de potencial hídrico
entre a fonte e o dreno o que favorece o movimento da seiva do floema para as bagas (LANG

105
e DURING, 1991). Além disso, o carregamento de açúcares no floema pode ser afetado pelo
estresse hídrico (WANG et al., 2003).
Os resultados obtidos no presente estudo concordam com os obtidos
por Dantas et al. (2005). Além disso, a maior atividade média da IAP na estratégia ID ao final
do ciclo em relação à IDC e IP caracteriza maior transporte de sacarose fotossintética pela via
apoplástica das células.
Desta forma, as diferentes invertases (INC, IAV e IAP) presentes nas
células foliares, além de se diferenciarem na localização celular também se diferenciam na
atividade ao longo do ciclo metabólico das plantas, quando submetidas às diferentes
estratégias hídricas.
4.8 Análises químicas: Acidez, pH e sólidos solúveis
Todos os frutos produzidos pelos vegetais são o resultado do manejo
adotado nas várias etapas de crescimento e desenvolvimento das plantas, levando-se em
consideração as características edafoclimáticas e ecofisiológicas em que se encontram e a
resposta bioquímica das plantas ao manejo adotado.
O acompanhamento da evolução da maturação das bagas é de grande
importância para definir o balanço entre teor de sólidos solúveis (SS) e acidez titulável (AT),
no intuito de determinar o momento da colheita e a qualidade das uvas para o processamento,
sendo que a melhor relação SS/AT ainda confere ao vinho equilíbrio gustativo determinante
em sua qualidade geral.
Os dados referentes ao resumo da análise de variância das análises
químicas: acidez titulável (AT), pH, teor de sólidos solúveis (SS) e relação SS/AT realizadas
em bagas aos 98, 105 e 112 DAP estão apresentados na Tabela 28. Em relação ao aspecto
qualitativo as variáveis AT, SS e a relação SS/AT demonstraram-se significativos (p<0,01)
pelo teste F, para o fator de avaliação dias após a poda. Ainda na Tabela 28 nota-se também
que para a variável pH foram verificadas respostas significativas (p<0,01) para as diferentes
estratégias de irrigação (EI), os diferentes dias após a poda (DAP) e interação significativa
entre eles (p<0,05).

106
Tabela 28. Resumo da análise de variância (teste F) para acidez titulável (AT), pH, teor de
sólidos solúveis (SS) e relação sólidos solúveis/acidez titulável (SS/AT) do mosto (g L-1
) em
função de três estratégias de irrigação (EI). Embrapa Semiárido – Petrolina, PE, 2010.
FV
GL
Quadrado médio
AT pH SS SS/AT
EI 2 1.630833ns 0.005926** 0.347037ns 0.015016ns
DAP 2 40.175833** 0.173704** 40.645926** 1.909836**
Bloco 2 0.035833ns 0.000370ns 0.671481ns 0.005902ns
EI x DAP 4 1.068333ns 0.002593* 0.320926ns 0.026114ns
Resíduo 16 0.613333 0.000787 0.558148 0.009609
CV (%) 6,88 0,92 4,21 6,05 ns: não significativo (p> 0,05); * p<0,05; ** p<0,01
Os frutos da videira são compostos por carboidratos, ácidos orgânicos
e compostos fenólicos que são dependentes do estado hídrico da planta, do microclima dos
cachos e da arquitetura do dossel vegetativo (OJEDA et al., 2002).
Os principais ácidos orgânicos das uvas são os ácidos tartárico e
málico, que juntos representam mais de 90% da acidez total, o ácido cítrico (5 a 10%) e em
menor concentração os ácidos: ascórbico, fosfórico, succínico, fumárico, acético, glicólico,
láctico, aconítico, químico, chiquímico e mandélico (KANELLIS e ROUBELAKIS-
ANGELAKIS, 1993).
A acidez condiciona a estabilidade biológica, a cor e as características
gustativas dos vinhos. A redução da acidez durante o período de maturação ocorre,
principalmente, devido à diluição dos ácidos, ocorrida pela entrada de água nas bagas e pela
oxidação do ácido málico e ácido tartárico. Durante o processo de maturação, a concentração
de ácido tartárico nas bagas reduz muito pouco, enquanto que os níveis de ácido málico
aumentam até a mudança de cor e a partir desta fase reduzem de forma drástica (COOMBE,
1992; OLLAT et al., 2002).
De acordo com os dados da Tabela 29, o avanço do ciclo proporcionou
redução significativa e progressiva da acidez titulável, considerando-se os diferentes dias após

107
a poda (98, 105 e 112). Isso pode ser indicativo de que os frutos estavam sob bom índice de
maturação, mesmo tendo sido colhidos precocemente (10 dias antes da data prevista, 125
DAP). Durante o desenvolvimento dos frutos ocorre aumento no teor de ácido tartárico e ácido
málico até a fase de ‘pintor’, com posterior redução. Ao contrário do que ocorre com os
açúcares durante o desenvolvimento da baga, o teor dos ácidos vai diminuindo à medida que a
uva vai amadurecendo (OLLAT et al., 2002).
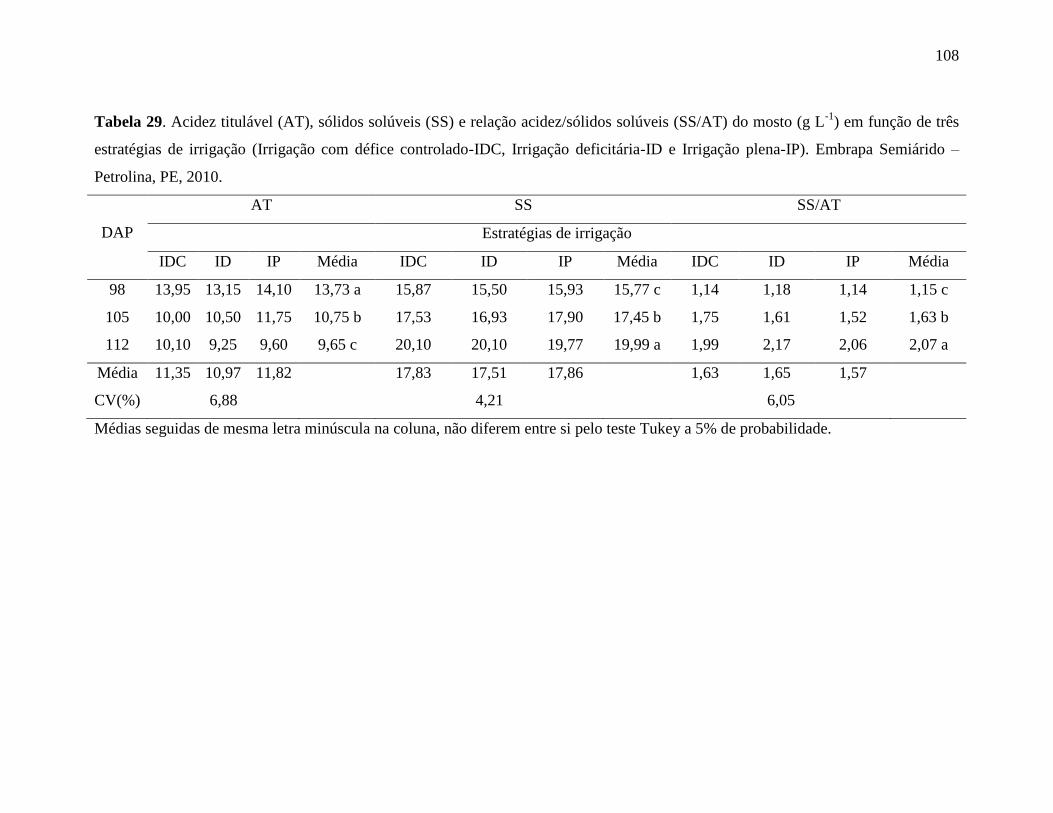
108
Tabela 29. Acidez titulável (AT), sólidos solúveis (SS) e relação acidez/sólidos solúveis (SS/AT) do mosto (g L-1
) em função de três
estratégias de irrigação (Irrigação com défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP). Embrapa Semiárido –
Petrolina, PE, 2010.
DAP
AT SS SS/AT
Estratégias de irrigação
IDC ID IP Média IDC ID IP Média IDC ID IP Média
98 13,95 13,15 14,10 13,73 a 15,87 15,50 15,93 15,77 c 1,14 1,18 1,14 1,15 c
105 10,00 10,50 11,75 10,75 b 17,53 16,93 17,90 17,45 b 1,75 1,61 1,52 1,63 b
112 10,10 9,25 9,60 9,65 c 20,10 20,10 19,77 19,99 a 1,99 2,17 2,06 2,07 a
Média 11,35 10,97 11,82 17,83 17,51 17,86 1,63 1,65 1,57
CV(%) 6,88 4,21 6,05
Médias seguidas de mesma letra minúscula na coluna, não diferem entre si pelo teste Tukey a 5% de probabilidade.

109
No presente trabalho os resultados obtidos concordam com os obtidos
por Freeman et al. (1980) que constataram redução nas características de qualidade da uva
‘Shiraz’ quando submetidas ao estresse hídrico. Segundo Esteban et al. (2002) condições de
défice hídrico favorecem o catabolismo do ácido málico, reduzindo assim, a acidez do mosto.
Para Conde et al. (2007) o aumento na temperatura das bagas, aumenta também o consumo de
energia e para manter a produção energética a célula recorre ao ácido málico armazenado
através do aumento de atividade da enzima málica. Além disso, a concentração mineral,
principalmente de Potássio, afeta a metabolização dos ácidos orgânicos, a relação
tartárico/málico e o pH (OLLAT et al., 2002).
Para Coombe (1987) a temperatura está relacionada com a acidez total
do mosto, pois quanto mais elevada for a temperatura, dentro dos limites críticos para cultura,
maior será a concentração de açúcares e menor de ácido málico nos frutos, já que o mesmo é
utilizado como substrato da respiração durante o amadurecimento. Ruffner et al. (1983),
atribui ao decréscimo na acidez titulável, a diluição da concentração de ácidos por aumento do
volume da baga, ativação da degradação, a inibição da síntese e transformação destes ácidos
em açúcares. Conde et al. (2007) relataram que a concentração ideal de ácidos orgânicos na
uva madura deve variar entre 0,65 e 0,85 g 100 mL-1
(90-110 meq/l). Entretanto, as condições
climáticas afetam o metabolismo dos ácidos orgânicos durante a maturação, regulando a taxa
de transformação do ácido málico em fontes de carbono para a respiração celular (JACKSON
e LOMBARD, 1993) ou a sua acumulação sob a forma de frutose e glicose (CONDE et al.,
2007). O aumento na quantidade de água aplicada à cultura pode interferir no aumento da
acidez total do mosto, contribuindo para a perda da qualidade no processamento da uva
(BEVILAQUA, 1995). Os valores de acidez obtidos no presente experimento estão bem acima
dos encontrados por outros autores (SANTOS e KAYE, 2009; FERNANDES, 2009). Esse
fato pode ser explicado pela coleta precoce dos frutos (ainda com elevada acidez titulável)
para as avaliações químicas, em função da antecipação da colheita devido ao ataque de
pássaros e insetos.
Os sólidos solúveis (SS) e a relação sólidos solúveis/acidez titulável
(SS/AT) tiveram comportamento inverso significativo ao da acidez, aumentando os seus
valores com o avanço do ciclo (Tabela 29). Esse comportamento é esperado, pois caracteriza a
evolução da maturação de bagas. O acúmulo de sólidos solúveis nas bagas ocorre lentamente

110
do crescimento herbáceo até a fase de ‘pintor’. A partir da qual, ocorre aumento no acúmulo
de SS até o ponto de máxima maturação (HIDALGO, 1993).
Os valores médios de SS obtidos ao final do ciclo (112 DAP)
19,99ºBrix foram superiores ao mínimo necessário (18ºBrix) para a formação de 10,7ºGL de
álcool potencial, que é a quantidade mínima de álcool necessária para a conservação do vinho
(RIBÉREAU-GAYON et al., 2004). Todavia, os valores de SS obtidos nessa safra de inverno
foram superiores ao valor médio de 16,5ºBrix, observado durante quatro safras de verão por
Souza et al. (2002) na mesma região de estudo.
As diferentes estratégias de irrigação e os diferentes dias após a poda
influenciaram os valores de pH do mosto (Tabela 30).
Tabela 30. Potencial hidrogeniônico (pH) do mosto (g L-1
) em função de três estratégias de
irrigação (Irrigação com défice controlado-IDC, Irrigação deficitária-ID e Irrigação plena-IP).
Embrapa Semiárido – Petrolina, PE, 2010.
DAP
pH
Estratégias de irrigação
IDC ID IP
98 2,90 Ac
3,00 Bb
3,20 Aa
2,93 Ac
3,10 Ab
3,20 Aa
2,93 Ac
3,10 Ab
3,20 Aa
105
112
CV(%) 0,92
Médias seguidas de mesma letra maiúscula na linha, não diferem entre si pelo teste Tukey a
5% de probabilidade.
O balanço entre o pH e a acidez titulável do mosto é importante no
processo de vinificação e na determinação das qualidades organolépticas do vinho.
Geralmente, mostos com baixo pH são menos propícios a oxidações microbianas indesejáveis
durante a fermentação, além de apresentar melhor balanço na coloração e expressão das
características do fruto. Durante o período de amadurecimento do fruto ocorre aumento no teor

111
de sólidos solúveis e pH, assim como, redução da acidez titulável, em decorrência,
principalmente, da redução do ácido málico (ESTEBAN et al., 2002). Os mesmos autores
concordam que o valor de pH é dependente, além da acidez titulável, das concentrações
relativas de ácidos málico e tartárico e do grau de formação de sais ácidos, que, por sua vez,
depende do conteúdo de potássio na baga. No presente estudo foi constatado que
independentemente da estratégia hídrica, os valores médios de pH aumentaram a partir de 98
DAP até à colheita (Tabela 30). Geralmente, videiras que se desenvolvem em regiões tropicais
com clima quente, tendem a produzir frutos com elevado teor de SS e baixos níveis de AT,
comparados à videira cultivada em clima frio. Além disso, há tendência para bagas expostas
ao sol terem pH mais elevado e níveis mais baixos de AT comparadas a bagas sombreadas.
Contudo, o défice hídrico também provoca aumento no pH do mosto (WILLIAMS e
MATTHEWS, 1990). No entanto, comparando os tratamentos ID e IP, que foram os dois
tratamentos com as características hídricas mais extremas durante o presente experimento, não
se constatou o resultado proposto pelos autores acima.
Aos 105 DAP nota-se menor valor de pH para o tratamento IDC em
relação ao ID e IP. Os ácidos orgânicos encontrados nos tecidos vegetais podem estar na
forma livre ou esterificada (metila, propila, hexila, etc) e os ácidos fracos livres, na presença
de seus sais de potássio, apresentam pequena variação no pH em função do equilíbrio
estabelecido no sistema (ELIAS, 2008). Na célula, esses ácidos encontram-se associados com
seus sais de potássio e constituem sistema tampões, que tem importante papel, particularmente
na regulação da atividade enzimática (CHITARRA e CHITARRA, 2005).
Desta forma, no presente estudo o aumento gradual do pH para os
tratamentos ID e IP nas três avaliações durante a maturação reflete maior formação de sais
ácidos às custas do ácido livre em relação ao tratamento IDC, sendo a relação entre sais ácidos
e ácido livre influenciada pela quantidade total de calor efetivo durante a maturação.

112
5 CONSIDERAÇÕES FINAIS
O Submédio do Vale do São Francisco é uma região, relativamente,
nova no cultivo comercial de videiras para os mais diversos fins. Assim, para que os genótipos
utilizados nessa região possam expressar fenotipicamente a sua capacidade produtiva, requer
do vitivinicultor o uso de técnicas de manejo adequadas para a realidade local. Entre as
técnicas utilizadas no ciclo produtivo, o fornecimento de água através de sistemas de irrigação
é fator limitante ao cultivo de espécies vegetais em regiões com clima semiárido. Ainda mais
importante do que ter o recurso hídrico é saber a sua qualidade e como utilizá-lo, por isso, são
testadas algumas estratégias de irrigação, buscando o uso mais criterioso e eficiente da água,
com menor gasto e geração de impactos ambientais como a salinização e desertificação. Além
disso, o uso de porta-enxertos com características de resistência à seca como o testado na
presente pesquisa é de grande importância por ser outra possibilidade de ajuste no consumo de
água pelas plantas. A melhor estratégia de irrigação em regiões semiáridas para uvas de vinho
não é aquela que leva a altas cargas produtivas com grande consumo hídrico, mas aquela em
que o menor consumo hídrico conduz à planta a maior exploração do seu potencial genético de

113
adaptação a ambientes desfavoráveis ao seu crescimento e desenvolvimento, atribuindo
qualidade aos frutos. Portanto, o uso de estratégias de irrigação que gerem estresse hídrico
temporário às plantas podem trazer benefícios ao produtor que economizará com o menor
volume de água aplicado e maior produção de metabólitos secundários que conferem
qualidade no processamento dos frutos.
Como sugestão para futuros trabalhos, recomenda-se, além da
utilização de diferentes estratégias de irrigação, realizar acompanhamento da safra no final do
segundo semestre e início do primeiro que é a época mais estressante em termos de
fornecimento de energia solar para as plantas. Com isso, avaliar durante o ciclo, características
não abordadas neste estudo como: taxas transpiratórias noturnas, potencial hídrico foliar ao
meio-dia, taxas respiratórias, peroxidação lipídica, fluorescência da clorofila a, atividade das
bombas de prótons, evolução da maturação dos frutos quando do 'pintor', atividade de enzimas
antioxidantes, solutos osmorreguladores como aminoácidos e quantificação de íons
inorgânicos como cálcio, potássio e sódio.
Desta forma, com maior volume de informações, se obteria abordagem
mais clara e abrangente dos efeitos do défice hídrico, durante o ciclo das videiras no semiárido
nordestino, facilitando a escolha do vitivinicultor na escolha da melhor estratégia hídrica a ser
utilizado, levando-se em consideração a qualidade dos frutos para o processamento e a sua
relação custo/benefício.

114
6 CONCLUSÕES
Nas condições e localização em que este estudo foi realizado, conclui-
se que:
A imposição da deficiência hídrica controlada a partir da fase
fenológica de 'cacho fechado' em videira cv. Syrah resulta em plantas ao longo do ciclo com
maior eficiência no uso da água e economia no recurso hídrico.
O porta-enxerto ‘Paulsen 1103’ demonstra tolerância e/ou resistência
ao défice hídrico e a cultivar copa Syrah comportamento anisohídrico.
A maior disponibilidade hídrica na irrigação plena promove as maiores
taxas de assimilação, transpiração e condutância estomática aos 73 e 87 dias após a poda.
Além disso, proporciona maior atividade da redutase do nitrato ao final do ciclo.
A menor densidade de fluxo de fótons fotossinteticamente ativo aos
101 dias após a poda influencia em maior concentração interna de CO2 na câmara
subestomática e menor eficiência na carboxilação in vivo da enzima Rubisco.

115
A manutenção de elevado teor SPAD da clorofila nas folhas entre 60 e
101 dias após a poda indica boas condições de funcionamento do aparelho fotossintético.
A irrigação deficitária confere maior atividade média da invertase
ácida da parede ao final do ciclo.
A acidez titulável do mosto diminuiu e os açúcares solúveis totais,
açúcares redutores, sólidos solúveis e a relação sólidos solúveis/acidez titulável aumentaram
para todas as estratégias de irrigação ao final do ciclo.

116
7. REFERÊNCIAS
ACEVEDO, C. et al. Efecto de diferentes niveles de agua aplicada en post-cuaja y en post-
pinta sobre la calidad del vino cv. Cabernet Sauvignon. Agricultura Tecnica, Santiago de
Chile, v. 65, n. 4, p. 397-410, 2005.
AGRIANUAL. Anuário estatístico da agricultura brasileira. São Paulo: FNP, 1997. p. 424-
435, 1997.
AGRIANUAL. Anuário estatístico da agricultura brasileira. São Paulo: FNP, 2011. p. 505-
520, 2011.
ALI, K. et al. Metabolic constituents of grapevine and grape-derived products.
Phytochemistry Reviews, Dordrecht, v. 9, p. 357-378, 2010.
ALLEWELDT, G.; RUHL, E. Untersuchungenzum gaswechsel der rebeI I.
einflußlanganhaltender bodentrockenheit auf die leistungsfähigkeitverschierdener rebsorten.
Vitis, Siebeldingen, v. 21, p. 313-324, 1982.
ALLEWELDT, G.; EIBACH, R.; RÜHL, E. Investigations on gas-exchange in grapevine. I.
Influence of temperature, leaf age, and daytime on net photosynthesis and transpiration. Vitis,
Siebeldingen, v. 16, p. 93-100, 1982.
ALQUINI, Y. et al. Epiderme. In: GLORIA, B. A.; GUERREIRO, S. M. C. (Eds.). Anatomia
vegetal. Viçosa, MG: UFV, 2006. 438 p.

117
ALSCHER, R. G.; DONAHUE, J. L.; CRAMER, C. L. Reactive oxygen species and
antioxidants: relationships in green cells. Physiologia Plantarum, Malden, v. 100, p. 224-233,
1997.
AMARANTE, C. V. T. et al. Quantificação da área e do teor de clorofilas em folhas de plantas
jovens de videira ‘Cabernet Sauvignon’ mediante métodos não destrutivos. Revista Brasileira
de Fruticultura, Jaboticabal, v. 31, n. 5, p. 680-686, 2009.
ANDERSEN, M. N. et al. Soluble invertase expression is an early target of drought stress
during the critical, abortion-sensitive phase of young ovary development in maize. Plant
Physiology, Minneapolis, v. 130, p. 591-604, 2002.
AQUARONE, E. et al. Biotecnologia industrial: biotecnologia na produção de alimentos.
São Paulo: Edgard Blücher, 2001. v. 4. 523 p.
ARNON, D. I. Copper enzymes in isolated chloroplasts: polyphenoloxidases in Beta vulgaris.
Plant Physiology, Maryland, v. 24, p. 1-15, 1949.
ARNOLD, W. N. β-fructofuranoside from grape berries. Biochimica et Biophysica Acta,
Amsterdam, v. 110, p.134-147, 1965a.
ARNOLD, W. N. β-fructofuranoside from grape berries II. Solubilization of bound fraction.
Biochimica et Biophysica Acta, Amsterdam, v. 128, p. 124-129, 1965b.
ASSIS, J. S. de; LIMA FILHO, J. M. P. Aspectos fisiológicos da videira irrigada. In: SOUZA
LEÃO, P. C. de; SOARES, J. M. (Eds.). A viticultura no semiárido. Petrolina: Embrapa
Semiárido, 2000. p. 129-146.
ÁVILA NETTO, J. et al. Exigências hídricas da videira na região do submédio São Francisco.
Pesquisa Agropecuária Brasileira, Brasília, DF, v. 35, n. 8, p. 1559-1566, 2000.
AZCON-BIETO, J. Inhibition of photosynthesis by carbohydrates in wheat leaves. Plant
Physiology, Minneapolis, v. 73, p. 681-686, 1983.
BAEZA, P. et al. Water relations between leaf water potential, photosynthesis and agronomic
vine response as a tool for establishing thresholds in irrigation scheduling. Scientia
Horticulturae, Amsterdam, v. 114, p. 151-158, 2007.
BAILLOD, M.; BAGGIOLINI, M. Lesstadesrepères de lavigne. Revue Suisse de Viticulture,
Arboriculture et Horticulture, Nyon, v. 25, n. 1, p. 7-9, 1993.
BARRATT, D. H. et al. Normal growth of Arabidopsis requires cytosolic invertase but not
sucrose synthase. Proceedings of the National Academy of Sciences of the United States of
America, Washington, DC, v. 106, p. 13124-13129, 2009.
BASSOI, L. H. et al. Preliminary results of a long term experiment about RDI and PRD
irrigation strategies in winegrape in São Francisco Valley, Brazil. Acta Horticulturae,
Leuven, v. 754, p. 275-282, 2007.

118
BATES, T. R.; DUNST, R. M.; JOY, P. Seasonal dry matter, starch, and nutrient distribution
in Concord grapevine roots. HortScience, Alexandria, v. 37, p. 313-316, 2002.
BEEVERS, L.; HAGELMAN, R. H. Nitrate reduction in higher plant. Annual Review of
Plant Physiology, Palo Alto, v. 20, p. 495-522, 1969.
BEEVERS, L.; HAGEMAN, R. H. Inorganic plant nutrition. In: LAUCHI, A.; BIELESKI,
R. L. (Eds.). Berlin: Springer-Verlag, 1983. 351 p.
BERTAMINI, M. et al. Influenza del la disposizione dei germogli e del la rimozione foglia
resullosviluppo vegetativo e la forzadei sinks; effetti sulla efficienza fotosintetica in Vitis
vinifera L: cv. 'Chardonnay' In: INTERNATIONAL SYMPOSIUM GRAPEVINE
PHISIOLOGY, 4., 1992, Torino. Proceedings... Torino: Fondazione Giovanni Dalmasso,
1992. p. 57-62.
BEVILAQUA, G. P. Avaliações físico-químicas durante a maturação de videiras cultivadas no
Rio Grande do Sul. Revista Brasileira de Agrociências, Pelotas, v. 1, n. 3, p. 151-156, 1995.
BINDON, K.; DRY, P.; LOVEYS, B. Influence of partial rootzone drying on the composition
and accumulation of anthocyanins in grape berries (Vitis vinifera cv. Cabernet Sauvignon).
Australian Journal of Grape and Wine Research, Adelaide, v. 4, n. 2, p. 91-103, 2008.
BISSON, L. F. Influence of nitrogen on yeast and fermentation of grapes. In:
INTERNATIONAL SYMPOSIUM ON NITROGEN IN GRAPES AND WINE, 1991, Seattle.
Proceedings… Washington, DC: American Society for Enology and Viticulture, 1991. p. 78-
89.
BOGEAT-TRIBOULOT, M. B. et al. Gradual soil water depletion results in reversible
changes ofgene expression, protein profiles, ecophysiology, and growth performancein
Populus euphratica, a poplar growing in arid regions. Plant Physiology, Minneapolis, v. 143,
p. 876-892, 2007.
BOLIANI, A. C.; FRACARO, A. A.; CORRÊA, L. de S. Uvas rústicas de mesa: cultivo e
processamento em regiões tropicais. Jales: [s.n.], 2008. 368 p.
BORGHEZAN, M. Comportamento ecofisiológico da videira (Vitisvinifera L.) cultivada
em São Joaquim, Santa Catarina: área foliar, crescimento vegetativo, composição da uva e
qualidade sensorial dos vinhos. 2010. 226 f. Tese (Doutorado em Ciências/Recursos
Genéticos)-Universidade Federal de Santa Catarina, Florianópolis, 2010.
BOSS, P. K.; DAVIES, C. Molecular biology of sugar and anthocyanin accumulation in grape
berries. In: ROUBELAKIS-ANGELAKIS, K. A. (Ed.). Molecular biology and
biotechnology of the grapevine. Dordrecht: Kluwer Academic, 2001. p. 1-33.
BRADFORD, M. M. A rapid and sensitive method for quatification of microgram quantities
of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, New York,
v. 72, p. 248-251, 1976.

119
BRAVDO, B. Physiological mechanisms involved in the production of non-hydraulic root
signals by partial rootzone drying: a review. Acta Horticulturae, Leuven, v. 689, p. 267-276,
2005.
BRAY, E. A. Molecular responses to water deficit. Plant Physiology, Minneapolis, v. 103, p.
1035-1040, 1993.
BREDEMEIER, C.; MUNDSTOCK, C. M. Regulação da absorção e assimilação do
nitrogênio nas plantas. Ciência Rural, Santa Maria, v. 30, n. 2, p. 365-372, 2000.
BRYCE, J. H.; THORNTON, J. M. Respiration and growth metabolism. In: ZAMSKI, E.;
SCHAFFER, A. A. Photoassimilate distribution in plants and crops: source-sink
relationships. New York: Marcel Dekker, 1996. p. 115-156.
BURSSENS, S. et al. Expression of cell cycle regulatory genes and morphological alterations
in response to salt stress in Arabidopsis thaliana. Planta, Berlin, v. 211, p. 632-640, 2000.
CABRITA, M. J. et al. Importância da rega na maturação de uvas da casta Aragones
(Vitisvinifera L.) na região do Alentejo. In: SIMPÓSIO DE VITIVINICULTURA DO
ALENTEJO, 5., 2001, Évora. Actas... Évora: CVRVV, 2001. p. 299-306.
CAIRD, M. A.; RICHARDS, J. H.; DONOVAN, L. A. Night-time stomatal conductance and
transpiration in C3 and C4 plants. Plant Physiology, Minneapolis, v. 143, p. 4-10, 2007.
CAMARGO, U. A. Variedades de uva. In: GUERRA, C. G. et al. Conhecendo o essencial
sobre uvas e vinhos. Bento Gonçalves: Embrapa Uva e Vinho, 2009. 69 p.
CARBONNEAU, A. The early selection of grapevine rootstocks for resistance to drought
conditions. American Journal of Enology and Viticulture, Davis, v. 36, p. 195-198, 1985.
CARBONNEAU, A. Aspects qualitatifs, 258-276. In: TIERCELIN, J. R. (Ed.).
Traited’irrigation. Paris: Tec & Doc. Lavosier, 1998. 1011 p.
CARVALHO, V. D. de.; CHITARRA, M. I. F. Aspectos qualitativos da uva. Informe
Agropecuário, Belo Horizonte, v. 10, n. 117, p. 75-79, 1984.
CERQUEIRA, R. C. Influência do frio e do déficit hídrico nas características fisiológicas e
bioquímicas em videiras (Vitisvinifera L.) cv. Thompson seedless. 2011. 122 f. Tese
(Doutorado em Agronomia/Horticultura)-Faculdade de Ciências Agronômicas, Universidade
Estadual Paulista, Botucatu, 2011.
CHALMERS, Y. M. Influence of sustained deficit irrigation on physiology and phenolic
compounds in wine grapes and wine. 2007. 200 f. Thesis (Doctor of Philosophy)-School of
Agriculture and Wine, Adelaide University, Adelaide, 2007.
CHAMPAGNOL, F. L. Acidité dês môuts et dês vins. Revue Française d´Oenologue.
Montpellier, v. 26, n. 1044, p. 26-57, 1986.

120
CHAVES, M. M. Fotossíntese e repartição dos produtos de assimilação em (Vitisvinifera
L.). 1986. 220 f. Tese (Doutorado)-Universidade Técnica de Lisboa, Lisboa, 1986.
CHAVES, M. M. Effects of water deficits on carbon assimilation. Journal of Experimental
Botany, Oxford, v. 42, n. 234, p. 1-16, 1991.
CHAVES, M. M.; MAROCO, J. P.; PEREIRA, J. S. Understanding plant responses to
drought: from genes to the whole plant. Functional Plant Biology, Victoria, v. 30, p. 239-
264, 2003.
CHAVES, M. M.; OLIVEIRA, M. M. Mechanisms underlying plant resilience to water
deficits: prospects for water-saving agriculture. Journal of Experimental Botany, Oxford, v.
55, p. 2365-2384, 2004.
CHAVES, M. M. et al. Deficit irrigation in grapevine improves water use efficiency while
controlling vigour and production quality. The Annals of Applied Biology, Oxford, v. 150, p.
237-252, 2007.
CHAVES, M. M.; FLEXAS. J.; PINHEIRO, C. Photosynthesis under drought and salt Stress:
regulation mechanisms from whole plant to cell. Annals of Botany, London, v. 103, p. 551-
560, 2009.
CHAVES, M. M. et al. Grapevine under déficit irrigation: hints from physiological and
molecular data. Annals of Botany, London, v. 105, p. 661-676, 2010.
CHEN, L. S.; CHENG, L. Carbon assimilation and carbohydrate metabolism of 'concord'
grape (Vitislabrusca L.) leaves in response to nitrogen supply. Journal of American Society
of Horticultural Science, Mount Vernon, v. 128, n. 5, p.7 54-760, 2003.
CHOUREY, P. S. et al. Genetic evidence that the two isozymes of sucrose synthase present in
developing maize endosperm are critical, one for cell wall integrity and the other for starch
biosynthesis. Molecular & General Genetics, Heidelberg, v. 259, p. 88-96, 1998.
CHITARRA, M. I. F.; CHITARRA, A. B. Pós-colheita de frutos e hortaliças: fisiologia e
manuseio. 2. ed. Lavras: UFLA, 2005. 785 p.
CHONÉ, X. et al. Stem water potential is a sensitive indicator of grapevine water status.
Annals of Botany, London, v. 87, p. 477-483, 2001.
CIFRE, J. et al. Physiological tools for irrigation scheduling in grapevine (Vitisvinifera L.).
Agriculture, Ecosystems & Environment, Amsterdam, v. 106, p. 159-170, 2005.
CLARK, R. B. Physiology of cereals for mineral nutrient uptake, use, and efficiency. In:
Crops as enhancers of nutrient use. DUNCAN, R. R.; BALIGAR, V. C. (Eds.). San Diego:
Academic Press, 1991. p. 131-209.

121
CLIMACO, P.; ABRANTES, M. L.; CASTRO, R. Influence du regime hydriquesur la
production et la qualite du mout cv. Vital. Revue Viticulture and Oenology, Siebeldingen, v.
4, p. 183-188, 1991.
CONDE, C. et al. Biochemical changes throughout grape berry development and fruit and
wine quality. Food, London, v. 1, p. 1-22, 2007.
COOMBE, B. C. Influence of temperature on composition and quality of grapes. Acta
Horticulturae, Wangeningen, v. 206, p. 23-35, 1987.
COOMBE, B. G. Research on development and ripening of the grape berry. American
Jounal of Enology and Viticulture, Davis, v. 43, p. 101-110, 1992.
COSTA, J. M.; ORTUÑO, M. F.; CHAVES, M. M. Deficit irrigation as strategy tosave water:
physiology and potential application to horticulture. Journal of Integrative Plant Biology,
Oxford, v. 49, p. 1421-1434, 2007.
COSTACURTA, A.; ROSELLI, G. Critéresclimatiquesetedaphiques pour l’etablissement des
vignobles. Bulletin de l’O.I.V., Paris, v. 53, n. 596, p. 783-786, 1980.
CROCOMO, O. J. Assimilação de nitrogêniopelasplantas. In: FERRI, M. G. (Ed.). Fisiologia
vegetal. São Paulo: EPU, 1985. p. 181-209.
DANTAS, B. F. et al. Atividade de invertases em cultivares-copa x porta-enxertos durante a
formação de videiras no vale do São Francisco. In: CONGRESSO BRASILEIRO DE
FRUTICULTURA, 18., 2004, Florianópolis. Anais... Florianópolis: EPAGRI, 2004. 1 CD-
ROM.
DANTAS, B. F. et al. Foliar carbohydrates content and invertase activity in vines at São
Francisco River Valley - Brazil. Revista Brasileira de Fruticultura, Jaboticabal, v. 27, n. 2,
p. 198-202, 2005.
DANTAS, B. F.; RIBEIRO, L. de. S.; PEREIRA, M. S. Teor de açúcares solúveis e insolúveis
em folhas de videiras, cv. syrah, em diferentes posições no ramo e épocas do ano. Revista
Brasileira de Fruticultura, Jaboticabal, v. 29, n. 1, p. 042-47, 2007a.
DANTAS, B. F. et al. Foliar carbohydrate content and invertase activity of 'Syrah' and
'Moacato Canelli' vines subjected to partial rootzone drying and regulated deficit irrigation.
Acta Horticulturae, Leuven, v. 754, p. 301-308, 2007b.
DAUDT, C. E.; BOEIRA, L. S.; PEREIRA, C. N. Influência da adubação nitrogenada e de
linhagens de leveduras nos teores de vinhos de Vitis vinífera cv. Cabernet Sauvignon. Ciência
e Tecnologia de Alimentos, Campinas, v. 15, n. 2, p. 166-169, 1995.
DAVIES, C.; ROBINSON, S. P. Sugar accumulation in grape berries: cloning of two putative
vacuolar invertase DNAs and their expression in grapevine tissues. Plant Physiology,
Minneapolis, v. 111, p. 275-283, 1996.

122
DAVIES, W. J.; ZHANG, J. Root signals and the growth and development of plants in drying
soil. Annual Review of Plant Physiology and Plant Molecular Biology, Palo Alto, v. 42, p.
55-76, 1991.
DAWSON, T. E. et al. Night-time transpiration in woody plants from contrasting
ecosystems.Tree Physiology, Oxford, v. 27, p. 561-575, 2007.
DE LA HARPE, A. C.; VISSER, J. H. Growth characteristics of Vitisvinifera L. cv. Cape
Riesling. South African Journal of Enology and Viticulture, Dennesig, v. 6, p. 1-6, 1985.
DEBOUBA, M. et al. NaCl stress effects on enzymesinvolved in nitrogen assimilation
pathway in tomato Lycopersiconesculentumseedlings. Journal of Plant Physiology, Stuttgart,
v. 163, p. 1247-1258, 2006.
DELOIRE, A. et al. Vine and water: a short review. Journal International des Sciences de la
Vigne et du Vin, Bordeaux, v. 38, n. 1, p. 1-13, 2004.
DELOIRE, A. et al. Grapevine responses to Terroir: a global approach. Journal International
des Sciences de la Vigne et du Vin, Bordeaux, v. 39, n. 4, p. 149-162, 2005.
DELOIRE, A. et al. A. Influence de l'etat hidrique de lavig ne sur le style de vin. Revue
française d'oenologie, v. 215, p. 11-15, 2006.
DELÚ FILHO, N. et al. Redução do nitrato e assimilação do amônio em plantas jovens de
seringueira cultivadas sob níveis crescentes de nitrato. Revista Brasileira de Fisiologia
Vegetal, Brasília, DF, v. 10, n. 3, p. 185-190, 1998.
DOEHLERT, D. C.; FELKER, F. C. Characterization and distribution of invertase activity in
developing maize (Zea mays) Kernels. Physiologia Plantarum, Malden, v. 70, p. 51-57,
1987.
DONOVAN, L. A.; LINTON, M. J.; RICHARDS, J. H. Predawn plant water potential does
not necessarily equilibrate with soil water potential under well-watered conditions. Oecologia,
Berlin, v. 129, p. 328-335, 2001.
DOORENBOS, J.; KASSAM, A. H. Efeito da água no rendimento das culturas. Campina
Grande: UFPB, 1994. 306 p.
DREIER, L. P.; HUNTER, J. J.; RUFFNER, H. P. Invertase activity, grape berry development
and cell compartmentation. Plant Physiology & Biochemistry, New Delhi, v. 36, p. 865-872,
1998.
DRY, P. R.; LOVEYS, B. R. Factors influencing grapevine vigour and the potential for
control with partial rootzone drying. Australian Journal of Grape and Wine Research,
Adelaide, v. 4, p. 140-148, 1998.

123
DRY, P. R. et al. Strategic irrigation management in Australian vineyards. Journal
International des Sciences de la Vigne et du Vin, Bordeaux, v. 35, n. 3, p. 129-139, 2001.
DUCHÊNE, E.; SCHNEIDER, C. Grapevine and climatic changes: a glance at the situation in
Alsace. Agronomy for Sustainable Development, Paris v. 25, p. 93-95, 2005.
DURING, H. Determination of the photosynthetic capacity of grapevine leaves. Vitis,
Siebeldingen, v. 30, p. 49-56, 1991.
DURING, H. Photosynthesis of ungrafted and grafted grapevines: effects of rootstock
genotype and plant age. American Journal of Enology and Viticulture, Davis, v. 45, n. 3, p.
297-299, 1994.
EBELER, S. E.; THORNGATE, J. H. Wine chemistry and flavour: looking into the crystal
glass. Journal of Agricultural and Food Chemistry, Easton, v. 57, n. 18, p. 8098-8108,
2009.
EFEOGLU, B.; EKMEKÇI, Y.; ÇIÇEK, N. Physiological responses of three maize cultivars
to drought stress and recovery. South African Journal of Botany, Pretoria, v. 75, p. 34-42,
2008.
ELIAS, H. H. de S. Caracterização física, química e bioquímica de cultivares de videira
durante a maturação. 2008. 74 f. Tese (Doutorado em Ciência dos Alimentos)-Universidade
Federal de Lavras, Lavras, 2008.
EMBRAPA. Centro Nacional de Pesquisa de Solos. Sistema brasileiro de classificação de
solos. Brasília, DF: Embrapa 1999. 412 p.
ENDO, M. et al. Sizeandlevels of mRNA for acid invertase in ripe tomato fruit. Plant Cell
Physiology, Oxford, v. 31, p. 655-659, 1990.
ESCALONA, J.M.; FLEXAS, J.; MEDRANO, H. Comparison of heat balance and gas
exchange methods to measure transpiration in irrigated and water stressed grapevines. Acta
Horticulturae, Hague, v. 526, p. 145-155, 1997.
ESCALONA, J. M.; FLEXAS, J.; MEDRANO, H. Stomatal and non-stomatal limitations of
photosynthesis under water stress in field-grown grapevines. Australian Journal Plant
Physiology, Melbourne, v. 26, p. 421-433, 1999.
ESTEBAN, M. A.; VILLANUEVA, M. J.; LISSARRAGUE, J. R. Relationships between
different berry components in Tempranillo (Vitisvinifera L) grapes from irrigated and
nonirrigated vines during ripening. Journal of the Science of Food and Agriculture,
London, v. 82, n. 10, p. 1136-1146, 2002.
EVANS, R. G. et al. Water use of Vitis vinifera grapes in Washington. Agricultural Water
Management, Amsterdam, v. 23, p. 109-124, 1993.

124
FANIZZA, G.; RICCIARD, L; BAGNULO, C. Leaf greenness measurements to evaluate
water stressed genotypes in Vitisvinifera. Euphytica, Wageningen, v. 55, p. 27-31, 1991.
FAVERO, A. C. Viabilidade de produção de videira ‘Syrah’ em ciclos de verão e inverno
no Sul de Minas Gerais. 2007. 112 f. Dissertação (Mestrado em Agronomia/Fitotecnia)-
Universidade Federal de Lavras, Lavras, 2007.
FOOD AND AGRICULTURE ORGANIZATION. Produção e área cultivada de uvas no
mundo. Disponível em:
<http://faostat.fao.org/site/567/DesktopDefault.aspx?PageID=567#ancor>. Acesso em: 12
nov. 2011.
FERNANDES, P. A. de C. Comportamento agronômico e enológico das castas Touriga
Nacional e Syrah em seis regiões portuguesas. 2009. 92 f. Dissertação (Mestrado em
Viticultura e Enologia)-Instituto Superior de Agronomia, Universidade Técnica de Lisboa,
Lisboa, 2009.
FERERES, E.; SORIANO, M. A. Deficit Irrigation for reducing agricultural water use.
Journal of Experimental Botany, Oxford, v. 58, p. 147-159, 2006.
FERNÁNDEZ, J. E. et al. Water relations and gas Exchange in olive trees Ander regulated
déficit irrigation and partial rootzone drying. Plant and Soil, Dordrecht, v. 284, p. 273-291,
2006.
FLEXAS, J.; ESCALONA, J. M.; MEDRANO, H. Down-regulation of photosynthesis by
drought under field conditions in grapevine leaves. Australian Journal of Plant Physiology,
Melbourne, v. 25, p. 893-900, 1998.
FLEXAS, J. et al. Effects of drought on photosynthesis in grapevines under field conditions:
an evaluation of stomatal and mesophyll limitations. Functional Plant Biology, Victoria, v.
29, p. 461-471, 2002.
FLEXAS, J. et al. Understanding down-regulation of photosynthesis under water stress: future
prospects and searching for physiological tools for irrigation management. Annals of Applied
Biology, Warwick, v. 144, p. 273-283, 2004.
FLEXAS, J. et al. Decreased Rubisco activity during water stress is not induced by decreased
relative water content but related to conditions of low stomatal conductance and chloroplast
CO2 concentration. New Phytologist, Cambridge, v. 172, p. 73-82, 2006.
FLEXAS, J. et al. Mesophyll condutance to CO2: current knowledge and future prospects.
Plant Cell & Environment, Oxford, v. 31, p. 602-612, 2008.
FOUQUET, R. et al. Identification of grapevine aquaporins and expression analysis in
developing berries. Plant Cell Reports, Berlin, v. 27, n. 9, p. 1541-1550, 2008.

125
FOYER, C. H.; GALTIER, N. Source-sink interaction and communication in leaves. In:
ZAMSKI, E.; SCAFFER, A. A. (Eds.). Source-sink relations. New York: Marcel Dekker,
1996. p. 311-340.
FRAGA, K. de F. Aminas bioativas durante a maturação de uvas Syrah produzidas em
diferentes regiões e sistemas de condução. 2010. 92 f. Dissertação (Mestrado em Ciência de
Alimentos)-Faculdade de Farmácia, Universidade Federal de Minas Gerias, Belo Horizonte,
2010.
FRANCIS, F. J. Anthocyanins and betalains: composition and applications. Cereal Food
World, Saint Paul, v. 45, n. 5, p. 208-213, 2000.
FRANKS, P. J.; DRAKE, P. L.; FROEND, R. H. Anisohydric but isohydrodynamic:
seasonally constant plant water potential gradient explained by a stomatal control mechanism
incorporating variable plant hydraulic conductance. Plant, Cell and Environment, Oxford, v.
30, p. 19-30, 2007.
FREEMAN, B. M.; LEE, T. H. TURKINGTON, C. R. Interaction of irrigation and pruning
level on grape and wine quality of shiraz vines. American Journal of Enology and
Viticulture, Davis, v. 31, p. 124-135, 1980.
FRIDMAN, E. et al. Zooming in on a quantitative trait for tomato yield using interspecific
introgressions. Science, Washington, DC, v. 305, p. 1786-1789, 2004.
GALMÉS, J. et al. Aquaporin expression in response to different water stress intensities and
recovery in Richter-110 (Vitis sp.): relationship with ecophysiological status. Planta, Berlin,
v. 226, n. 3, p. 671-681, 2007.
GAMON, J. A.; PEARCY, R. W. Photoinhibition in Vitiscalifornica: interactive effects of
sunlight, temperature and water status. Plant, Cell and Environment, Oxford, v. 13, p. 267-
275, 1990.
GARCIA, N. G.; DAUDT, C. E.; Utilização de nitrogênio durante a fermentação de mostos.
Revista do Centro de Ciências Rurais, Santa Maria, v. 18, n. 2, p. 141-147, 1988.
GAUDILLERE, J. P.; VAN LEEUWEN, C.; OLLAT, N. Carbon isotope composition of
sugars in grapevine, an integrated indicator of vineyard water status. Journal of
Experimental Botany, Oxford, v. 53, n. 369, p. 757-763, 2002.
GIL, G. F.; PSZCZOŁKOWSKI, P. Viticultura: fundamentos para optimizer produccion y
calidad. Santiago de Chile: Ediciones Universidad Catolica de Chile, 2007. 535 p.
GIOVANNINI, E. Produção de uvas para vinho, suco e mesa. 3. ed. Porto Alegre:
Renascença, 2008. 368 p.
GIRONA, J. Analisis de la repuesta de la vina a la aplicacion de la tecnica de partial root
zone drying (PRD). Toulouse: Station Regionale ITV Midi-Pyrénées, 2005, p. 18-24.

126
GIRONA, J. et al. The use of midday leaf water potential for scheduling deficit irrigation in
vineyards. Irrigation Science, New York, v. 24, p. 115-127, 2006.
GODOY, L. J. G. Manejo do nitrogênio em coberturana cultura do milho (Zea mays L.)
em solo arenoso baseado no índice relativo de clorofila. 2002. 94 f. Dissertação (Mestrado
em Agronomia/Agricultura)-Faculdade de Ciências Agronômicas, Universidade Estadual
Paulista, Botucatu, 2002.
GODOY, L. J. G. et al. Índice relativo de clorofila e o estado nutricional em nitrogênio
durante o ciclo do cafeeiro fertirrigado. Revista Brasileira de Ciência do Solo, Campinas, v.
52, p. 217-226, 2008.
GOETZ, M. et al. Induction of male sterility in plants by metabolic engineering of the
carbohydrate supply. Proceedings of the National Academy of Sciences of the United
States of America, Washington, DC, v. 98, p. 6522-6527, 2001.
GONÇALVES, S. de O. Influência de manejos de irrigação sobre aspectos de ecofisiologia
e de produção da videira cv. Syrah/paulsen 1103. 2011. 54 f. Dissertação (Mestrado em
Agronomia/Irrigação e Drenagem)-Faculdade de Ciências Agronômicas, Universidade
Estadual Paulista, Botucatu, 2011.
GOODWING, I.; BOLAND, A. M. Scheduling deficit irrigation of fruit trees for optimizing
water use efficiency. In: FOOD AND AGRICULTURE ORGANIZATION. Deficit irrigation
practices. Rome: FAO, 2002. p. 67-77.
GREINER, S. et al. Ectopic expression of a tobacco invertase inhibitor homolog prevents
cold-induced sweetening of potato tubers. Nature Biotechnology, New York, v. 17, p. 708-
711, 1999.
GU, S. L. et al. Effects of irrigation amount on water relations, vegetative growth, yield and
fruit composition of Sauvignon blanc grapevines under partial rootzone drying and
conventional irrigation in the San Joaquin Valley of California, USA. Journal of
Horticultural Science & Biotechnology, Ashford, v. 79, n. 1, p. 26-33, 2004.
GUERRA, C. C. Maturação da uva e condução da vinificação para a elaboração de vinhos
finos. In: SIMPÓSIO MINEIRO DE VITICULTURA E ENOLOGIA, 1., 2002. Andradas.
Anais... Caldas: EPAMIG-FECD, 2002. p. 179-192.
GUPTA, S. C.; BEEVERS, L. Regulation of nitrite reduction. In: HARPER, J. E.;
SCHRADER, L. E.; HOWELL, R. W. (Eds.). Exploitation of physiological and genetic
variability to enhance crop productivity. Rockville: American Society of Plant Physiology,
1985. p. 1-11.
HAMMAN, R. A.; DAMI, I. E. Effects of irrigation on wine, grape growth and fruit quality.
HortTechnology, Alexandria, v. 10, n. 1, p. 162-168, 2000.
HAMMOND, J. P.; WHITE, P. J. Sucrose transport in the phloem: integrating root responses
to phosphorus starvation. Journal of Experimental Botany, Oxford, v. 59, p. 93-109, 2008.

127
HAYES, M. A.; DAVIES, C.; DRY, I. B. Isolation, functional characterization, and
expression analysis of grapevine (Vitisvinifera L.) hexose transporters: differential roles in
sink and source tissues. Journal of Experimental Botany, Oxford, v. 58, p. 1985-1997, 2007.
HAWKER, J. S. Changes in the activities of enzymes concerned with sugar metabolism
during the development of grape berries. Phytochemistry, New York, v. 8, p. 9-17, 1969.
HELLMAN, E. W. Grapevine structure and function. In: HELLMAN, E. W. (Ed.). Oregon
viticulture. Corvallis: Oregon State University Press, 2003. p. 5-19.
HEWITT, E. J.; HUCKLESBY, D. P.; NORTON, B. A. Nitrate metabolism. In: BONNER, J.;
VARNER, J. E. (Ed.). Plant biochemistry. New York: Academic Press, 1976. v. 20. p. 633-
681.
HIDALGO, L. Tratado de viticulture general. Madrid: Mundi, 1993. 983 p.
HOFF, T.; STUMMANN, B. M.; HENNINGSEN, K. W. Structure, function and regulation of
nitrate reductase in higher plants. Physiologia Plantarum, Malden, v. 84, p. 616-624, 1992.
HSIAO, T. C. Plant responses to water stress. Annual Review of Plant Physiology, Palo
Alto, v. 24, p. 519-570, 1973.
HSIAO, T. C. Plant responses to water deficits, efficiency, and drought resistance.
Agricultural Meteorology, Amsterdam, v. 14, p. 59-84, 1979.
HSIAO, T. C. Measurements of plant water status. In: STEWART, B. A.; NIELSEN, D. R.
(Eds.). Irrigation of agricultural crops. Madison: American Society of Agronomy, 1990. p.
244-280.
IACONO, F.; BUCCELLA, A.; PETERLUNGERB, E. Water stress and rootstock influence
on leaf gas exchange of grafted and ungrafted grapevines. Scientia Horticulturae,
Amsterdam, v. 75, p. 27- 39, 1998.
INTRIERI, C. et al. Leaf age, leaf position and photosynthesis in potted grapevines. Advances
in Horticultural Science, Firenze, v. 6, p. 23-27, 1992.
INTRIERI, C. et al. Vine performance and leaf physiology of conventionally and minimally
pruned Sangiovese grapevines. Vitis, Siebeldingen, v. 40, n. 3, p. 123-130, 2001.
INTRIGLIOLO, D. S.; CASTEL, J. R. Response of Vitisvinifera cv. ‘Tempranillo’ to partial
rootzone drying in the field: water relations, growth, yield and fruit and wine quality.
Agriculture Water Management, Amsterdam, v. 96, p. 282-292, 2009.
JACKSON, D. I.; LOMBARD, P. B. Environmental and management practices affecting
grape composition and wine quality: a review. American Journal of Enology and
Viticulture, Davis, v. 44, p. 409-430, 1993.

128
JAIN, A. K. et al. Variation in the sugar accumulation pattern of muscadine grape genotypes.
Proceedings of the Florida State Horticultural Society, Goldenrod, v. 115, p. 329-336,
2002.
JIN, Y.; NI, D. A.; RUAN, Y. L. Post translational elevation of cell wall invertase activity by
silencing its inhibitor in tomato delays leaf senescence and increases seed weight and fruit
hexose level. The Plant Cell, Rockville, v. 21, p. 2072-2089, 2009.
JOHANSSON, I. et al. The role of aquaporins in cellular and whole plant water
balance.Biochimica and Biophysica Acta Biomembranes, Amsterdam, v. 1465, p. 324-342,
2000.
JONES, J. A. Valuable information from accurate crop estimation. Australian Grape Grow
Winemak, Adelaide, v. 316, p. 59-60, 1990.
JONES, H. G. Plants and microclimate: a quantitative approach to environmental plant
physiology. 2. ed. Cambridge: Cambridge University Press, 1992. 428 p.
KALA, S.; GODORA, A. K. Effect of moisture stress on leaf total proteins, proline and free
amino acid content in commercial cultivars of Ziziphus mauritiana. Journal of Scientific
Research Banaras Hindu University, Uttar Pradesh, v. 55, p. 65-69, 2011.
KALDENHOFF, R. et al. Aquaporins and plant water balance. Plant Cell & Environment,
Oxford, v. 31, p. 658-666, 2008.
KANELLIS, A. K.; ROUBELAKIS-ANGELAKIS, K. A. Grape. In: SEGMOUR, G.;
TAYLOR, J. TUCKER, G. Biochemistry of fruit ripening. London: Chapman & Hall, 1993.
cap. 6. p. 189-234.
KANG, S.; ZHANG, J. Controlled alternate partial root-zone irrigation: its physiological
consequences and impact on water use efficiency. Journal of Experimental Botany, Oxford,
v.55, p. 2437-2446, 2004.
KAVANAGH, K. L.; PANGLE, R.; SCHOTZKO, A. D. Nocturnal transpiration causing
disequilibrium between soil and stem predawn water potential in mixed conifer forests of
Idaho. Tree Physiology, Oxford, v. 27, p. 621-629, 2007.
KELLER, M. Estratégias de irrigação de uvas brancas e tintas. Enovitis, Lisboa, n. 3, p. 16-
20, 2005.
KINDERMANN, P. E.; SMART, R. E. Effect of irradiance, temperature and leaf water
potential on photosynthesis of vine leaves. Photosynthetica, Prague, v. 5, p. 6-15, 1971.
KIRSCHBAUM, M. U. F. Water-stress in Eucalyptus pauciflora: comparison of effects on
stomatal conductance with effects on the mesophyllcapacity for photosynthesis, and
investigation of a possible involvementof photoinhibition. Planta, Berlin, v. 171, p. 466-473,
1987.

129
KIRSCHBAUM, M. U. F. Recovery of photosynthesis from water stress inEucalyptus
pauciflora: a process in two stages. Plant Cell & Environment, Oxford, v. 11, p. 685-694,
1988.
KISHINO, A. Y. Características da planta. In: KISHINO, A. Y.; CARVALHO, S. L. C.;
ROBERT, S. F. (Orgs.). Viticultura tropical: o sistema de produção do Paraná. Londrina:
IAPAR, 2007. v. 1, p. 87-140.
KISHINO, A. Y.; CARAMORI, P. H. Fatores climáticos e o desenvolvimento da videira. In:
KISHINO, A. Y.; CARVALHO, S. L. C.; ROBERTO, S. R. Viticultura tropical: o sistema
de produção no Paraná. Londrina: IAPAR, 2007. cap. 3. p. 59-86.
KLANN, E. M.; HALL, B.; BENNETT, A. B. Antisense acid invertase (TIV1) gene alters
soluble sugar composition and size in transgenic tomato fruit. Plant Physiology, Minneapolis,
v. 112, p. 1321-1330, 1996.
KLIEWER, W. M. Grapevine physiology: how does a grapevine make sugar? Berkeley:
University of California, 1981. 13 p.
KLIEWER, W. M.; FREEDMAN, B. M.; HOSSOM, C. Effect of irrigation, crop level and
potassium fertilization on Carignane vines. I. Degree of water stress and effect on growth and
yield. American Journal of Enology and Viticulture, Davis, v. 27, p. 55-61, 1983.
KLOBET, W. Influence of light and temperature on vine performance in cool climates and
applications to vineyard management. In: INTERNATIONAL SYMPOSIUM ON COOL
CLIMATE VITICULTURE AND ENOLOGY, 1984, Oregon. Proceedings… Oregon:
Oregon State University, 1984. p.139-157.
KOUNDOURAS, S. et al. Influence de l’alimentation en eau sur la croissance de la vigne, la
maturation des raisins et les caracteristiques des vins en zone mediterraneenne (exemple de
Nemee, Grece, cepage Saint-Georges, 1997). Journal International des Sciences de la
Vigne et du Vin, Bordeaux, v. 33, p. 149-160, 1999.
KOUNDOURAS, S. et al. Influence of vineyard location and vine water status on fruit
maturation of nonirrigated cv. agiorgitiko (Vitisvinifera l.). Effects on wine phenolic and
aroma components. Journal of Agricultural and Food Chemistry, Easton, v. 54, p. 5077-
5086, 2006.
KRAUTER, C. Environmental factors that influence grapevine water requirements. In:
CENTRAL COAST VITICULTURE AND ENOLOGY ISSUES CONFERENCE, 2001,
Fresno. Proceedings... Fresno: CATI, 2001. p. 44-49.
KRIEDEMANN, P. E. Photosynthesis in vine leaves as a function of light intensity,
temperature, and leaf age. Vitis, Siebeldingen, v. 7, p. 213-220, 1968.

130
KRIEDEMANN, P. E. Vine leaf photosynthesis. In: INTERNATIONAL SYMPOSIUM ON
THE QUALITY OF THE VINTAGE, 1997, Capetown. Proceedings… Capetown: RSA,
1977. p. 67-87.
KRIEDEMANN, P. E.; KLIEWER, W. M.; HARRIS, J. M. Leaf age and photosynthesis in
VitisviniferaL. Vitis, Siebeldingen, v. 9, p. 97-104, 1970.
LANG, A.; DÜRING, H. Partitioning control by water potentialgradient: Evidence for
compartmentation breakdown in grapeberries. Journal of Experimental Botany, Oxford, v.
40, p. 1069-1078, 1991.
LARA, M. E. B. et al. Extracellular invertase is an essential component of cytokinin-mediated
delay of senescence. The Plant Cell, Rockville, v. 16, p. 1276-1287, 2004.
LARCHER, W. Ecofisiologia vegetal. São Carlos: C.H.B.A., 2000. 531 p.
LAWLOR, D. W.; CORNIC, G. Photosynthetic carbon assimilation and associated
metabolism in relation to water deficits in higher plants. Plant Cell & Environment, Oxford,
v. 25, p. 275-294, 2002.
LAWLOR, D. W.; TEZARA, W. Causes of decreased photosynthetic rate and metabolic
capacity in water-deficient leaf cells: a critical evaluation of mechanisms and integration of
processes. Annals of Botany, London, v. 103, p. 561-579, 2009.
LEA, P. J. Nitrogen metabolism. In: LEA, P. J.; LEEGOOD, R. C. Plant biochemistry and
molecular biology. Chichester: John Willey and Sons, 1993. cap. 7. p. 155-180.
LEÃO, P. C. S. Avaliação do comportamento fenológico e produtivo de seis cultivares de
uva sem sementes no Vale do Rio São Francisco. 1999. 120 f. Dissertação (Mestrado)-
Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal,
1999.
LEBON, G. et al. Sugars and Flowering in the grapevine (Vitis vinifera L.). Journal of
Experimental Botany, Oxford, v. 59, n. 10, p. 2565-2578, 2008.
LEEUWEN, C. et al. The influence of climate, soil and cultivar on terroir. American Journal
of Enology and Viticulture, Davis, v. 55, n. 3, p. 207-217, 2004.
LIGLE, S. E.; DUNLOP, J. R. Sucrose metabolism in netted muskmelon fruit during fruit
development. Plant Physiology, Minneapolis, v. 84, p. 386-389, 1987.
LILLO, C. Light regulation of nitrate reductase in greenleaves of higher plants: Mini Review
Physiologia Plantarum, Malden, v. 90, p. 616-620, 1994.
LIMA FILHO, J. M. P. et al. Aspectos fisiológicos. In: SOARES, J. M.; LEÃO, P. C. de S.
(Eds.). A vitivinicultura no semiárido brasileiro. Petrolina: Embrapa Semiárido, 2009. p.72-
108.

131
LOBATO, A. K. S. et al. Biochemical and physiological behavior of Vigna unguiculata (L.)
Walp. under stress duringthevegetativephase. Asian Journal of Plant Sciences, v. 7, p. 44-49,
2008.
LOPES, C. Influência do sistema de conducão no microclima do coberto, vigor e
produtividade da videira (Vitis vinífera L.). 1994. 205 f. Tese (Doutorado)-Instituto
Superior de Agronomia, Lisboa, 1994.
LOPES, C. M. A. Relationships between leaf water potential and photosynthetic activity of
field-grown grapevines under a mediterranean environment. Acta Horticulturae, Hague, v.
493, p. 287-292, 1999.
LOPES, C. et al. Rega da vinha no Alentejo. Dotações e época de aplicação. In: SIMPÓSIO
DE VITIVINICULTURTA DO ALENTEJO, 5., 2001, Évora. Actas... Évora: ATEVA/
CCRA, 2001. p. 99-105.
LOVEYS, B. R. Diurnal changes in water relations and abscisic acid in field grown Vitis
vinifera cultivars. III. The influence of xylem-derived abscisic acid on leaf gas exchange. New
Phytologist, Cambridge, v. 98, p. 563-573, 1984.
LOVEYS, B. R.; KRIEDEMANN, P. E. Rapid changes in ABA-like inhibitors following
alterations in vine leaf water potential. PhysiologiaPlantarum, Copenhagen, v. 28, p. 476-
479, 1973.
LOVEYS, B. R. et al. Progress in the development of partial rootzone drying. Australian
Grapegrower and Winemaker, Goodwood, v. 403, p. 18-20, 1997.
LOVEYS, B. et al. Partial rootzone drying stimulates stress responses in grapevine to improve
water use efficiency while maintaining crop yield and quality. Australian Grape Grower
and Winemaker, Goodwood, v. 414, p. 108-113, 1998.
LOVEYS, B. R.; STOLL, M.; DAVIES, W. J. Physiological approaches to enhance water use
efficiency in agriculture: exploiting plant signalling in novel irrigation practice. In: BACON,
M. A. (Ed.). Water use efficiency in plant biology. Lancaster: University of Lancaster, 2004.
p. 113-141.
LOVISOLO, C.; SCHUBERT, A. Effects of water stress on vessel size and xylem hydraulic
conductivity in Vitis vinifera L. Journal of Experimental Botany, Oxford, v. 49, n. 321, p.
693-700, 1998.
LOVISOLO, C. et al. Mercurial inhibition of root hydraulic conductance in Vitisspp.
rootstocks under water stress. Environmental and Experimental Botany, Elmsford, v. 63, p.
178-182, 2008.
LOWELL, C. A.; TOMLISON, P. T.; KOCH, K. E. Sucrose metabolizing enzymes in
transport tissues and adjacent sink structures in developing citrus fruit. Plant Physiology,
Minneapolis, v. 90, p. 1394-1402, 1989.

132
MALAGI, G. Respostas agronômicas e ecofisiológicas de videira, cultivar BRS Violeta,
influenciadas por sistemas de adubação. 2011. 176 f. Dissertação (Mestrado em
Agronomia/Produção vegetal)-Universidade Tecnológica Federal do Paraná, Pato Branco,
2011.
MALHEIRO, A. Microclimate, yield and water-use of vineyards in the Douro Region,
Portugal. Thesis (Doctor of Philosophy)-Cranfield University, Silsoe, 2005.
MANFROI, L. Avaliação do processo fermentativo e da composição de vinho Merlot
elaborado com diferentes espécies de Saccharomycesoenococcuse Lactobacillus. 2002. 119
f. Tese (Doutorado em Ciência e Tecnologia de Alimentos)-Departamento de Tecnologia de
Alimentos, Universidade Federal de Viçosa, Viçosa, MG, 2002.
MARANVILLE, J. E.; PAULSEN, G. M. Alteration of carbohydrate composition of corn (Zea
mays L.) seedlings during moisture stress. Agronomy Journal, Madison, v. 62, p. 605-608,
1970.
MARINHO, L. B. et al. P.Potencial de água no solo e na folha da videira "Sugraone" sob
déficit hídrico. Revista Brasileira de Engenharia Agrícola e Ambiental, Campina Grande,
v. 15, n. 11, p. 1115-1122, 2011.
MAROCO, J. P. et al. Limitations to leaf photosynthesis in field-grown grapevines under
drought - metabolic and modelling approaches. Functional Plant Biology, Victoria, v. 29, p.
451-459, 2002.
MARRENCO, A. R.; LOPES, N. F. Fisiologia vegetal. 3. ed.Viçosa, MG: UFV, 2009. 486 p.
MARSCHNER, H. Mineral nutrition of higher plants. 2. ed. New York: Academic Press,
1995. 889 p.
MATOS, N. N. et al. Nitrate reductase is partially correlated to transpirational flux in cashew
seedlings exposed to water and salt stress. In: CONGRESSO BRASILEIRO DE FISIOLOGIA
VEGETAL, 8., 2001, Ilhéus. Anais… Ilhéus: SBFV/CEPLAC, 2001.
MATTHEWS, M. A.; ANDERSON, M. M. Fruit ripening in Vitis vinifera L.: responses to
seasonal water deficits. American Journal of Enology and Viticulture, Davis, v. 39, n. 4, p.
313-320, 1988.
MATTHEWS, M. A.; KRIEDEMANN, P. E. Water deficit, yield, and berry size as factors for
composition and sensory attributes of red wine. In: PROCEEDINGS OF THE AUSTRALIAN
SOCIETY OF VITICULTURE AND OENOLOGY, 2006, Victoria. Proceedings...Victoria:
ASVO, 2006. 9 p.
MAUREL, C. et al. Plant aquaporins: membrane channels with multiple integrated functions.
Annual Review of Plant Biology, Palo Alto, v. 59, p. 595-624, 2008.
MAZZA, G. Anthocyanins in grape and grape products.Critical Review of Food science and
Nutrition, Boca Raton, v. 35, n. 4, p. 341-371, 1995.

133
MCCARTHY, M. G. The effect of transient water deficit on berry development of Shyraz
(Vitisvinifera L). Australian Journal of Grape and Wine Research, Adelaide, v. 3, p. 102-
108, 1997.
MCCARTHY, M. G. et al. Regulated deficit irrigation and partial rootzone drying as irrigation
management techniques for grapevines. In: FOOD AND AGRICULTURE
ORGANIZATION. Deficit Irrigation practices. Rome: FAO, 2002. p. 79-87. (Water
Reports, n. 22).
MCCREE, K. J.; FERNANDEZ, C. J. Simulation model for studying physiological water
stress responses in whole plants. Crop Science, Madison, v. 29, p. 353-360, 1989.
MEDRANO, H. et al. Regulation of photosynthesis of C3 plants in response to progressive
drought: the interest of stomatal conductance as a reference parameter. Annals of Botany,
London, v. 89, p. 895-905, 2002.
MIELE, A.; MANDELLI, F. Sistemas de condução. In: KUHN, G. B. (Org.). Uva para
processamento: produção. Brasília, DF: Embrapa Informação Tecnológica, 2003. p. 56-65.
(Série Frutas do Brasil, 34).
MILLER, G. L. Use of dinitrosalicylis acid reagent for determination of reducing sugars.
Analytical Chemistry, Washington, DC, v. 31, n. 4, p. 426-428, 1959.
MORINAGA, K. et al. Effects of fruit load on partitioning of 15
N and 13
C, respiration, and
growth of grapevine roots at different fruit stages. Scientia Horticulturae, Amsterdam, v. 97,
p. 239-253, 2003.
MORRIS, D. L. Quantitative determination of carbohydrates with drywood´santhrone reagent.
Science, Washington, DC, v. 107, n. 1, p. 254-255, 1948.
MOTA, F. S. et al. Zoneamento climático do Rio Grande do Sul para a videira europeia. In:
CONFERÊNCIA LATINOAMERICANA DEL VINO Y LA UVA, 6., 1974, Caxias do Sul.
Actas... Porto Alegre: OLAVU, 1974.
MOURA, M. S. B. et al. Exigência térmica e caracterização fenológica da videira Syrah no
Vale do Rio São Francisco. In: CONGRESSO BRASILEIRO DE AGROMETEOROLOGIA,
15., 2007, Aracaju. Anais... Aracaju: Sociedade Brasileira de Agrometeorologia, 2007. 1 CD-
ROM.
MOUTINHO-PEREIRA, J. M. Caracterização fisiológica e agronômica de diferentes
estratégias culturais para minimizar o stress estival em Vitis vinifera L. na região
demarcada do douro. 2000. 287 f. Tese (Doutorado)-Universidade de Trás-os-Montes e Alto
Douro, Vila Real, 2000.
MOUTINHO-PEREIRA, J. M. et al. Leaf gas exchange and water relations of grapevines
grown in three different conditions. Photosynthetica, Prague, v. 42, p. 81-86, 2004.
MULLINS, M. G.; BOUQUET, A.; WILLIAMS, L. E. In: MULLINS, M.G. (Ed.). Biology of
the grapevine. Cambridge: Cambridge University Press, 1992. 239 p.

134
MULLINS, M. G.; BOUQUET, A.; WILLIAMS, L. E. Biology of the grapevine. Cambridge:
University Press, 2000. 239 p.
MUÑOZ-ESPADA, A. C. et al. Anthocyanin quantification and radical scavering capacity of
Concord, Norton, and Marechal Foch grapes and wines, Journal of Agricultural and Food
Chemistry, Washington, DC, v. 52, n. 22, p. 6779-6786, 2004.
NACZK, M.; SHAHIDI, F. Extraction and analysis of phenolics in food. Journal of
Chromatography A, Amsterdam, v. 1054, n. 1-2, p. 95-111, 2004.
NAIDU, B. P.; PALEG, L. G.; JONES, G. P. Nitrogenous compatible solutes in drought
stressed Medicago spp. Phytochemistry, New York, v. 31, p. 1195-1193, 1992.
NAOR, A.; WAMPLE, R. L. A rapid field method for measuring net assimilation rate stomata
conductance relationship: a feasibility test using grapevine leaves. Scientia Horticulturae,
Amsterdam, v. 60, p. 287-292, 1995.
NASCIMENTO, R.; MOSQUIM, P. R.; ARAÚJO, E. F.; SANTANNA, R. Distribuição de
amido, açúcares solúveis e atividades de invertases em explantes de soja sob várias
concentrações de sacarose e diferentes fontes de nitrogênio. Revista Brasileira de Fisiologia
Vegetal, Brasília, DF, v. 10, n. 2, p. 125-130, 1998.
NETO, E. B.; BARRETO, L. P. Análises químicas de tecidos e produtos vegetais. Recife:
UFRPE, 2004. 148 p.
NOGUEIRA, D. J. P. O. O clima na viticultura. Informe Agropecuário, Belo Horizonte, v.
10, n. 117, p. 11-14, 1984.
NORBERTO, P. M. Sistemas de condução em videira: análises econômicas e
ecofisiológicas. 2006. 118 f. Tese (Doutorado em Agronomia)-Universidade Federal de
Lavras, Lavras, 2006.
NORBERTO, P. M. et al. Efeito do sistema de condução em algumas características
ecofisiológicas da videira (Vitislabrusca L.) Ciência Agrotecnológica, Lavras, v. 33, n. 3, p.
721-726, 2009.
OJEDA, H. A rega na vinha: estratégias e técnicas de monitorização. Pinhão: Vinideas, 2007.
Não publicado.
OJEDA, H.; DELOIRE, A.; CARBONNEAU, A. Influence of water deficits on grape berry
growth.Vitis, Siebeldingen, v. 40, n. 3, p. 141-145, 2001.
OJEDA, H. et al. Influence of pre and postveraison water deficit on synthesis and
concentration of skin phenolic compounds during berry growth of Vitis vinifera L., cv. Shiraz.
American Journal of Enology and Viticulture, Davis, v. 53, n. 4, p. 261-267, 2002.
OLIVER, S. N.; DENNIS, E. S.; DOLFERUS, R. ABA regulates apoplastic sugar transport
and is a potential signal for cold induced pollen sterility in rice. Plant and Cell Physiology,
Kyoto, v. 48, p. 1319-1330, 2007.

135
OLLAT, N. et al. Grape berry development: a review. Journal International des Sciences de
la Vigne et du Vin, Bordeaux, v. 36, p. 109-131, 2002.
OLMO, H. P. Grapes: Vitis, Muscadinia (Vitacea). In: SMART, J.; SIMMONDS, N. W.
Evolution of crop plants. Singapore: Longman Publishers, 1995. p. 485-490.
ONCINS, J. A. et al. Fitomonitorização: estratégias hídricas na vinha para um vinho de
qualidade – o caso do Chardonnay. In. CONGRESO IBERICO DE CIENCIAS
HORTÍCOLAS, 5.; CONGRESO IBEROAMERICANO DE CIENCIAS HORTICOLAS, 4.,
2005, Porto. Anais... Porto: Associação Portuguesa de Horticultura, 2005. p. 1-6.
ORLANDO, T. G. S. et al. Comportamento das cultivares Carbenet Sauvignon e Syrah em
diferentes porta-enxertos. Ciência Agrotecnológica, Lavras, v. 32, n. 3, p.749-755, 2008.
OUGH, C. S.; AMERINE, M. A. Methods for analysis of musts and wines. 2. ed. New
York: John Wiley & Sons, 1987. 341 p.
PALLIOTTI, A.; CARTECHINI, A. Developmental changes in gas exchange activity in
flowers, berries, and tendrils of field-grown Cabernet Sauvignon. American Journal of
Enology and Viticulture, Davis, v. 52, p. 317-323, 2001.
PARANYCHIANAKIS, N. V.; AGGELIDES, S.; ANGELAKIS, A. N. Influence of
rootstock, irrigation level and recycled water on growth and yield of Sultanina grapevines.
Agricultural Water Management, Amsterdam, v. 69, n. 1, p. 13-27, 2004.
PATAKAS, A.; NOITSAKIS, B. Mechanisms involved in diurnal changes of osmotic
potential in grapevines under drought conditions. Journal of Plant Physiology, Stuttgart, v.
154, p. 767-774, 1999.
PATAKAS, A.; NOITSAKIS, B. Leaf age effects on solute accumulation in water-stressed
grapevines. Journal of Plant Physiology, Stuttgart, v. 158, n. 1, p. 63-69, 2001.
PATAKAS, A.; KOFIDIS, G.; BOSABALIDIS, A. M. The relationships between CO2
transfer mesophyll resistance and photosynthetic efficiency in grapevine cultivars. Scientia
Hoticulturae, Amsterdam, v. 97, p. 255-263, 2003.
PATAKAS, A.; NOITSAKIS, B.; CHOUZOURI, A. Optimization of irrigation water use in
grapevines using the relationship between transpiration and plant water status. Agriculture,
Ecosystems & Environment, Amsterdam, v. 106, p. 253-259, 2005.
PATAKAS, A. et al. The role of organic solute and ion accumulation in osmotic adjustment in
drought-stressed grapevines. Plant Science, Shannon, v. 163, p. 361-367, 2002.
PATRICK, J. W.; OFFLER, C. E. Compartmentation of transport and transfer events in
developing seeds. Journal of Experimental Botany, Oxford, v. 52, p. 551-564, 2001.
PAUL, M. J.; FOYER, C. H. Sink regulation of photosynthesis: review article. Journal of
Experimental Botany, Oxford, v. 52, n. 360, p. 1383-1400, 2001.

136
PAYAN, J. C.; GARCIA DE CORTAZAR ATAURI, I.; SEGUIN, B. Ressource en eau et
changement climatique: uneproblematique? Gruissan: Les EntretiensVitivinicoles Rhone-
Méditerranée, 2006.
PAZ, N. E.; AGUIRRE, F. M.; ONTIVEROS, J. L. R. Efecto del déficit hídrico
enelpatrónelectroforético de proteínas totalesen dos variedades de maíz. Revista Fitotecnia
Mexicana, Montecillo, v. 24, n. 1, p. 121-128, 2001.
PEDREIRA DOS SANTOS, T. et al. Effect of deficit irrigation strategies on cluster
microclimate for improving composition of Moscatel field-grown grapevines. Scientia
Horticulturae, Amsterdam, v. 112, p. 321-330, 2007.
PEDRO JUNIOR, M. J.; SENTELHAS, P. C. Clima e produção. In: POMMER, V. C.
(Coord.). Uva: tecnologia de produção, pós-colheita, mercado. Porto Alegre: Cinco
Continentes, 2003. 778 p.
PEREIRA, M. S. et al. Potencial hídrico e atividade da redutase do nitrato em videiras
submetidas a diferentes manejos de irrigação e porta-enxertos. In: JORNADA DE
INICIAÇÃO CIENTÍFICA DA EMBRAPA SEMIÁRIDO, 1., 2006, Petrolina. Anais...
Petrolina: Embrapa Semiárido, 2006. p. 171-176. 2006.
PESSARAKLI, M.; HUBER, J. T.; TUCKER, T. C. Protein synthesis in green beans under
salt stress with two nitrogen sources. Journal of Plant Nutrition, New York, v. 12, n. 11, p.
1361-1377, 1989.
PETER, B.; STEPHANIE, D. Sugar accumulation in tomato and partial purification of buffer
insoluble invertase. Phytochemistry, New York, v. 36, p. 837-884, 1994.
PETERLUNGER, E.; SIVILOTTI, P.; COLUSSI, V. Water stress increased polyphenolic
quality in ‘merlot’ grapes. Acta Horticulturae, Hague, v. 689, p. 293-300, 2005.
PETERSEN, G. W. et al. Fragipans in Pennsylvania soils: a statistical study of laboratory data.
Soil Science Society of America Journal, Madison, v. 34, p. 719-722, 1970.
PIMENTEL, C. Metabolismo de carbono na agricultura tropical. Seropédica: Edur, 1998.
150 p.
PIMENTEL, C. A relação da planta com a água. Seropédica: EDUR, 2004. 191 p.
POMMER, C. V.; PASSOS, I. R. Fisiologia da videira: como produz açúcar uma videira.
Disponível em: <http://users.directnet.com.br/pommer/COMOPRODUZACUCAR.pdf>.
Acesso em: 03 out. 2009.
PONI, S.; INTRIERI, C.; SILVESTRONI, O. Interactions of leaf age, fruiting, and exogenous
cytokinins in Sangiovese grapevines under non-irrigated conditions. I. Gas exchange.
American Journal of Enology and Viticulture, Davis, v. 45, p. 71-78, 1994.

137
PONI, S.; BERNIZZONI, F.; REINOTTI, M. Whole-canopy and single leaf gas-exchange,
responses to partial rootzone drying in potted ‘Cabernet Sauvignon’ grapevines. Acta
Horticulturae, Hague, v. 689, p. 277-284, 2005.
PORRO, D. et al. Lo SPAD nella diagnosi dello stato nutrizionale della vite. Informatore
Agrario, Verona, v. 57, n. 26, p. 49-55, 2001.
POSSNER, D. R. E.; KLIEWER, W. M. The localization of acids, sugars, potassium and
calcium in developing grape berries.Vitis, Siebeldingen, v. 24, p. 229-240, 1985.
PREGNOLATTO, W.; PREGNOLATTO, N. P. Normas analíticas do Instituto Adolfo
Lutz: metodosquimicos e físicos para análise de alimentos. 3. ed. São Paulo: Instituto Adolfo
Lutz, v. 1, 1985.
PREMACHANDRA, G. S. et al. Osmotic adjustment and stomatal response to water deficits
in maize. Journal of Experimental Botany, Oxford, v. 43, n. 256, p.1451-1456, 1992.
PRIETO, J. A.; GALANT GIORDI, E.; PEREZ PEÑA, J. Modelling photosynthetic-light
response on Syrah leaves with different exposure. Vitis, Siebeldingen, v. 49, n. 3, p.145-146,
2010.
PROTAS, J. F. S.; CAMARGO, U. A.; MELLO, L. M. R. Viticultura brasileira: regiões
tradicionais e pólos emergentes. Informe Agropecuário, Belo Horizonte, v. 27, n. 234, p. 7-
15, 2006.
QUEREIX, A. Analyse et modélisation du fonctionnementstomatique de lavigne Vitis
vinifera L. 2001. 132 f. Thèse (Doctorat)-Université de Bordeaux, Bordeaux, 2001.
RADIN, J.W. In vivo assay of nitrate reductase in cotton leaf discs. Plant Physiology,
Minneapolis, v. 51, n. 2, p. 332-336, 1973.
ROBY, G.; MATTHEWS, M. A. Relative proportions of seed, skin and flesh, in ripe berries
from Cabernet Sauvignon grapevines grown in a vineyard either well irrigated or under water
deficit. Australian Journal of Grape and Wine Research, Adelaide, v. 10, p. 74-82, 2004.
ROBY, G. et al. Berry size and vine water deficits as factors in winegrape composition:
anthocyanins and tannins. Australian Journal of Grape and Wine Research, Adelaide, v.
10, p. 100-107, 2004.
RODRIGUES, M. L. et al. Osmotic adjustment in water stress grapevine leaves in relation to
carbon assimilation. Australian Journal of Plant Physiology, Adelaide, v. 20, p. 309-321,
1993.
RODRIGUES, M. L. et al. Hydraulic and chemical signalling in the regulation of stomatal
conductance and plant water use of field grapevines growing under deficit irrigation.
Functional Plant Biology, Victoria, v. 35, p. 565-579, 2008.
ROITSCH, T.; GONZÁLEZ, M. C. Function and regulation of plant invertases: sweet
sensations. Trends in Plant Science, London, v. 9, p. 606-613, 2004.

138
REDDY, A. R.; CHAITANYA, K. V.; VIVEKANANDAN, M. Drought-induced responses of
photosynthesis and antioxidant metabolism in higher plants. Journal of Plant Physiology,
Stuttgart, v. 161, p. 1189-1202, 2004.
REGINA, M. de A. Influência dos frutos sobre a fotossíntese das folhas em Vitis vinifera.
Revista Brasileira de Fisiologia Vegetal, Londrina, v. 7, n. 2, p. 165-169, 1995.
REGINA, M. A.; AUDEGUIN, L. Avaliação ecofisiológica de clones de videira cv. Syrah.
Ciência e Agrotecnologia, Lavras, v. 29, n. 4, p. 875-879, 2005.
REYNOLDS, A. et al. Influence of irrigation on vine performance, fruit composition, and
wine quality of Chardonnay in a cool, humid climate. American Journal of Enology and
Viticulture, Davis, v. 58, n. 2, p. 217-228, 2007.
RIBÉREAU-GAYON G. Etude des mecanismes de syntheseet de transformation de
l'acidemalique, de l'acidetartriqueet de l'acidecitrique chez Vitis vinifera L.
Phytochemistry, New York, v. 7, n. 9, p. 1471-1482, 1968.
RIBÉREAU-GAYON, J. et al. Traitéd´oenologie: science et techiques du vin. Tome I –
Analyse et contrôle des vins. 2. ed. Paris: Dunod, 1982. 671 p.
RIBÉREAU-GAYON, J. et al. Traité d'oenologie - 1. Microbiologie du vin: vinifications. 5.
ed. Paris: Dunod, 2004. 661p.
ROBINSON, S. P.; DAVIES, C. Molecular biology of grape berry ripening. Australian
Journal of Grape and Wine Research, Adelaide, v. 6, p. 175-188, 2000.
ROGIERS, S. Y. et al. Does night-time transpiration contribute to any sohidric behaviour in a
Vitisvinifera cultivar? Journal of Experimental Botany, Oxford, v. 60, n. 13, p. 3751-3763,
2009.
ROITSCH, T. et al. Extracellular invertases: metabolic enzyme and metabolic protein.
Journal of Experimental Botany, Oxford, v. 54, n. 382, p. 513-524, 2003.
ROSS, H. A. et al. Developmental changes in carbohydrate content and sucrose degrading
enzymes in tuberisingstolons of potato (Solanumtuberosum). Physiology Plantarum,
Copenhagen, v. 90, p. 748-756, 1994.
RUAN, Y. L.; LLEWELLYN, D. J.; FURBANK, R. T. Suppression of sucrose synthase
expression represses cotton fibre cell initiation, elongation and seed development. The Plant
Cell, Rockville, v. 15, p. 952-964, 2003.
RUAN, Y. L.; JIN, Y.; HUANG, J. Capping invertase activity by its inhibitor: roles and
implications in sugar signaling, carbon allocation, senescence and evolution. Plant Signalling
& Behavior, Austin, v. 4, p. 983-985, 2009.
RUAN, Y. L. et al. Sugar input, metabolism, and signaling mediated by invertase: roles in
development, yield potential, and response to drought and heat. Molecular Plant, Oxford, v.
3, n. 6, p. 942-955, 2010.

139
RUFFNER, H. P.; BREM, S.; MALIPIERO, U. The physiology of acid metabolism in grape
berry ripening. Acta Horticulturae, Hague, v. 139, p. 123-128, 1983.
RUPP, D.; TRANKLE, L. A non-destructive measurement method for chlorophyll in
grapevines. Mitteilungen Klosterneuburg. Rebe und Wein, Obstbau und
Fruechteverwertung, Stuttgart, v. 45, n. 5/6, p. 139-142, 1995.
RUPP, D.; TRANKLE, L.; FOX, R. Non-destructive measurement of chlorophyll in grapes -
evaluation of varietal influences and effects of sampling methods. Mitteilungen
klosterneuburg, Rebe und Wein, Obstbau und Fruechteverwertung, Stuttgart, v. 49, n. 3, p.
86-92, 1999.
SAGE, R. F.; SHARKEY, T. D.; PEARCY, R. W. The effect of leaf nitrogen and temperature
on the CO2 response of photosynthesis in the C3 dicot Chenopodium album L. Australian
Journal of Plant Physiology, Adelaide, v. 17, p. 135-148, 1990.
SANTOS, H. P. Aspectos ecofisiológicos na condução da videira e sua influência na
produtividade do vinhedo e na qualidade dos vinhos. Bento Gonçalves: Embrapa, 2006. 9
p. (Comunicado Técnico, 71).
SANTOS, R. F.; CARLESSO, R. Déficit hídrico e os processos morfológicos e fisiológicos
das plantas. Revista Brasileira de Engenharia Agrícola e Ambiental, Campina Grande, v. 2,
n. 3, p. 287-294, 1998.
SANTOS, A. O.; KAYE, O. Composição quali-quantitativa da produção de 'Syrah' cultivada
sob estresse hídrico transiente. Revista Brasileira de Engenharia Agrícola e Ambiental,
Campina Grande, v. 13, n. 3, p. 272-281, 2009.
SANTOS, C. F.; LIMA, G. P. P.; MORGADO, L. B. Tolerância e caracterização bioquímica
em feijão-caupi submetido a estresse hídrico na pré-floração. Naturalia, Rio Claro, v. 33, p.
34-44, 2010.
SCHAFFER, A. A. et al. Citrus leaf chlorosis induced by sink removal: starch, nitrogen and
chloroplast ultrastructure. Journal of Plant Physiology, Stuttgart, v. 124, p. 111-121, 1986.
SCHOLANDER, P. F. et al. Sappressure in vascular plants. Science, Washington, DC, v. 148,
p. 339-346, 1965.
SCHULTZ, H. R. Differences in hydraulic architecture account for nearisohydric and
anisohydricbehaviour of two field-grown Vitis vinifera L. cultivars during drought. Plant, Cell
& Environment, Oxford, v. 26, p. 1393-1405, 2003.
SCHULTZ, H. R.; KIEFER, W.; GRUPPE, W. Photosynthetic duration, carboxylation
efficiency and stomatal limitation of sun and shade leaves of different ages in field-grown
grapevine (Vitis vinifera L.).Vitis, Siebeldingen, v. 35, p. 169-179, 1996.
SCHULZE, E. D.; HALL, A. E. Estomatal responses, water loss and CO2 assimilation rates of
plant in contrasting environments. In: LANGE, O. L. et al. Physiological plant ecology II. v.
12B. Berlin: Springer-Verlag, 1982. p. 181-230.

140
SENTELHAS, P. C. Aspectos climáticos para a viticultura tropical. Informe Agropecuário,
Belo Horizonte, v. 19, n. 194, p. 9-14, 1998.
SEPÚLVEDA, G.; KLIEWER, W. M. Stomatal response ofthreegrape-vinecultivars (Vitis
vinifera L.) to high temperature. American Journal of Enology and Viticulture, Davis, v.
37, p. 44-52. 1986.
SERGEEVA, L. I. et al. Vacuolar invertase regulates elongation of Arabidopsis thaliana roots
as revealed by QTL and mutant analysis. Proceedings of the National Academy of Sciences
of the United States of America, Washington, DC, v. 103, p. 2994-2999, 2006.
SILVA, J. A. M. Irrigação lateralmente alternada e com déficit hídrico na videira cv.
Petite Syrah. 2005. 99 f. Dissertação (Mestrado em Irrigação)-Universidade Federal de
Viçosa, Viçosa, MG, 2005.
SILVA, R. N. et al. Comparison methods for the determination of reducers sugars and total in
honey. Ciência e Tecnologia de Alimentos, Campinas, v. 23, n. 3, 2003. Disponível em:
<http://www.scielo.br>. Acesso em: 18 ago. 2011.
SOLOMONSON, L. P.; BARBER, M. J. Assimilatory nitrate reductase: functional properties
and regulation. Annual Review of Plant Physiology and Plant Molecular Biology, Palo
Alto, v. 41, p. 225-253, 1990.
SOUZA, C. R.; SOARES, A. M.; REGINA, M. de A. Trocas gasosas de mudas de videira,
obtidas por dois porta-enxertos, submetidas à deficiência hídrica. Pesquisa Agropecuária
Brasileira, Brasília, DF, v. 36, n. 10, p. 1221-1230, 2001.
SOUZA,C. R. et al. indicação de cultivares de videira para o sul de Minas Gerais. In:
REGINA, M. A. (Coord.). Viticultura e enologia: atualizando conceitos. Caldas: EPAMIG-
FECD, 2002. p. 227-286.
SOUZA, C. R. et al. Partial rootzone drying: regulation of stomatal aperture and carbon
assimilation in field grown grapevines (Vitis vinifera). Functional Plant Biology, Victoria, v.
30, p. 653-662, 2003.
SOUZA, C. R. et al. Control of stomatal aperture and carbon uptake by deficit irrigation in
two grapevine cultivars. Agriculture, Ecosystems and Environment, Amsterdam, v. 106, p.
261-274, 2005a.
SOUZA, C. R. et al. Impact of deficit irrigation on water use efficiency and carbon isotope
composition (δ13C) of field-grown grapevines under Mediterranean climate. Journal of
Experimental Botany, Oxford, v. 56, n. 418, p. 2163-2172, 2005b.
SOUZA, C. R. et al. Water relations of field-grown grapevines in the São Francisco Valley,
Brazil, under different rootstocks and irrigations strategies. Scientia Agrícola, Piracicaba, v.
66, n. 4, p. 436-446, 2009.

141
SPONHOLZ, W. R. Nitrogen compounds in grapes, must, and wine. In: INTERNATIONAL
SYMPOSIUM ON NITROGEN IN GRAPES AND WINE, 1991, Seattle. Proceedings…
Washington, DC: American Society for Enology and Viticulture, 1991. p. 67-77.
STEELE, M. R.; GITELSON, A. A.; RUNDQUIST D. C. A comparison of two techniques for
nondestructive measurement of chlorophyll content in grapevine leaves. Agronomy Journal,
Madison, v. 10, n. 3, p. 779-782, 2008.
STIKIC, R. et al. Partial root drying (PRD): a new technique for growing plants that saves
water and improves the quality of fruit. Bulgarian Journal of Plant Physiology, Sofia,
Special Issue, p. 164-171, 2003.
STITT, M.; VON SCHAEWEN, A.; WILLMITZER, L. Sink regulation of photosynthetic
metabolism in transgenic tobacco plants expressing yeast invertase in their cell wall involves a
decrease of the Calvin cycle enzymes and an increase of glycolytic enzymes. Planta, Berlin,
v. 183, p. 40-50, 1990.
STURM, A. Invertases: primary structures, functions and roles in plant development and
sucrose partitioning. Plant Physiology, Minneapolis, v. 121, p. 1-7, 1999.
TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 3. ed. Porto Alegre: Artmed, 2004. 719 p.
TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 4. ed. Porto Alegre: Artmed, 2009. 820 p.
TANG, G. Q.; LUSCHER, M.; STURM, A. Antisense repression and vacuolar and cell wall
invertase in transgenic carrot alters early plant development and sucrose partitioning. The
Plant Cell, Rockville, v. 11, p. 177-189, 1999.
TARDIEU, F.; SIMONNEAU, T. Variability among species of stomatal control under
fluctuating soil water status and evaporative demand: modeling isohydric and
anisohydricbehaviours. Journal of Experimental Botany, Oxford, v. 49, p. 419-432, 1998.
TECCHIO, M. A. et al. Teores foliares de nutrientes, índice relativo de clorofila e teores de
nitrato e de potássio na seiva do pecíolo na videira 'niagara rosada'. Revista Brasileira de
Fruticultura, Jaboticabal, v. 33, n. 2, p. 649-659, 2011.
TONIETTO, J.; MANDELLI, F. Uvas viníferas para processamento em regiões de clima
temperado. Brasília, DF: Embrapa-SP4, 2003. Disponível em:
<http://www.cnpuv.embrapa.br/publica/sprod/UvasViniferasRegioesClimaTemperado/clima.h
tm>. Acesso em: 09 ago. 2011.
TYMOWSKA-LALANNE, Z.; KREIS, M. The plant invertases: physiology, biochemistry
and molecular biology. Advances in Botanical Research, London, v. 28, p. 71-117, 1998.
VANDELEUR, R. K. et al. The role of plasma membrane intrinsic protein aquaporins in water
transport through roots: diurnal and drought stress responses reveal different strategies
between isohydric and anisohydric cultivars of grapevine. Plant Physiology, Minneapolis, v.
149, p. 445-460, 2009.

142
VERA-ESTRELLA, R. et al. Novel regulation of aquaporins during osmotic stress. Plant
Physiology, Minneapolis, v. 135, p. 2318-2329, 2004.
VINE, R. P.; HARKNESS, E. M.; LINTON, S. J. Winemaking: from grape growing to
marketplace. Amsterdam: Springer, 2002.
WAMPLE, R. L. Regulated deficit irrigation as a water management strategy in Vitisvinifera
production. In: FOOD AND AGRICULTURE ORGANIZATION. Deficit irrigation
practices. Rome: FAO, 2002. p. 89-102.
WANG, A. P. et al. An in vivo experimental system sugar phloem unloading in ripening grape
berries during water deficiency stress. Annals of Botany, London, v. 92, p. 523-528, 2003.
WANG, E. et al. Control of rice grain-filling and yield by a gene with a potential signature of
domestication. Nature Genetics, New York, v. 40, p. 1370-1374, 2008.
WEBER, H.; BORISJUK, L.; WOBUS, U. Molecular physiology of legume seed
development. Annual Reviewof of Plant Biology, Palo Alto, v. 56, p. 253-279, 2005.
WILLIAMS, L. E. Grape. In: ZAMSKI, E. (Ed.). Photoassimilate distribution in plants and
crops: source–sink relationships. New York: Marcel Dekker, 1996. p. 851-881.
WILLIAMS, L. E. The effect of cyanamide on budbreak and vine development of Thompson
seedless grapevines in the San Joaquin Valley of California. Vitis, Siebeldingen, v. 26, p.107-
113, 1987.
WILLIAMS, L. E.; MATTHEWS, M. A. Grapevine. In: STEWART, B. A.; NIELSEN, D. R.
(Eds.). Irrigation of agricultural crops. Madison: American Society of Agronomy, 1990. p.
1019-1055. (Agronomy Monograph, 30).
WINES FROM BRASIL. Vinho brasileiro, reconhecimento internacional. Disponível em:
<http://www.winesfrombrazil.com/brasilvitivinicola.php>. Acesso em: 5 jun. 2009.
WINKLER, A. J. Viticultura. México: Compañia Editorial Continental, 1970. 792 p.
WINKLER, A. J. et al. General viticulture. Berkeley: University of California Press, 1974.
710 p.
YANG, J.; MARTINSON, T. E.; LIU, R. H. Phytochemical profilesand antioxidant activities
of wine grapes. Food Chemistry, London, v. 116, p. 332-339, 2009.
YEMM, E. W.; WILLIS, A. J. The estimation of carbohydrates in plants extracts by anthrone.
Biochemical Journal, Colchester, v. 57, n. 5, p. 508-514, 1954.
ZAMSKI, E. Anatomical and physiological characteristics of sink cells. In: ZAMSKI, E.;
SCHAFFER, A. A. (Eds.). Source-sink relations. New York: Marcel Dekker, 1996. p. 283-
310.
ZANOR, M. I. et al. RNA interference of LIN5 in Solanumly copersicum confirms its role in
controlling Brix content, uncovers the influence of sugars on the levels of fruit hormones and

143
demonstrates the importance of sucrose cleavage for normal fruit development and fertility.
Plant Physiology, Minneapolis, v. 150, p. 1204-1218, 2009.
ZEEVAART, Z. R.; CREELMAN, R. A. Metabolism and physiology of abscisic acid. Annual
Review of Plant Physiology, Palo Alto, v. 39, p. 439-473, 1988.
ZOECKLEIN, B. W. et al. Analyses and production of Wine. Zaragoza: Acribia, 2001. 613
p.
ZUFFEREY, V. et al. Diurnal cycles of embolism formation and repair in petioles of
grapevine (Vitis vinifera cv. Chasselas). Journal of Experimental Botany, Oxford, v. 62, n.
11, p. 3885-3894, 2011.


















