
UNIVERSIDADE FEDERAL DE SERGIPE PRÓ-REITORIA DE PÓS ...contribuição dos receptores para cininas...
103
UNIVERSIDADE FEDERAL DE SERGIPE PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA MESTRADO EM CIÊNCIAS FISIOLÓGICAS THÁSSIO RICARDO RIBEIRO MESQUITA MODULAÇÃO DO SISTEMA CARDIOVASCULAR PELOS RECEPTORES PARA CININAS E SUA CONTRIBUIÇÃO PARA O STATUS REDOX CELULAR SÃO CRISTÓVÃO 2013
Transcript of UNIVERSIDADE FEDERAL DE SERGIPE PRÓ-REITORIA DE PÓS ...contribuição dos receptores para cininas...

UNIVERSIDADE FEDERAL DE SERGIPE PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
MESTRADO EM CIÊNCIAS FISIOLÓGICAS
THÁSSIO RICARDO RIBEIRO MESQUITA
MODULAÇÃO DO SISTEMA CARDIOVASCULAR PELOS RECEPTORES PARA CININAS E SUA CONTRIBUIÇÃO
PARA O STATUS REDOX CELULAR
SÃO CRISTÓVÃO 2013

i
THÁSSIO RICARDO RIBEIRO MESQUITA
MODULAÇÃO DO SISTEMA CARDIOVASCULAR PELOS RECEPTORES PARA CININAS E SUA
CONTRIBUIÇÃO PARA O STATUS REDOX CELULAR
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Fisiológicas da Universidade Federal de Sergipe como requisito à obtenção do grau de Mestre em Ciências Fisiológicas. Orientador: Drª. Sandra Lauton Santos Co-orientador: Dr. Luciano dos Santos Aggum Capettini
SÃO CRISTÓVÃO 2013

ii
FICHA CATALOGRÁFICA ELABORADA PELA BIBLIOTECA CENTR AL UNIVERSIDADE FEDERAL DE SERGIPE
Mesquita, Thássio Ricardo Ribeiro
M582m
O
Modulação do sistema cardiovascular pelos receptores para cininas e sua contribuição para o status redox celular / Thássio Ricardo Ribeiro Mesquita; orientadora Sandra Lauton Santos – São Cristóvão, 2013. xi, 85 f.: Il. Dissertação (mestrado em Ciências Fisiológicas) – Universidade Federal de Sergipe, 2013.
1. Sistema cardiovascular. 2. Sistema calicreína-cinina. 3. Espécies
reativas de oxigênio. 4. Espécies reativas de nitrogênio. I. Santos, Sandra Lauton, orient. II. Título.
CDU: 612.1

iii
THÁSSIO RICARDO RIBEIRO MESQUITA
MODULAÇÃO DO SISTEMA CARDIOVASCULAR
PELOS RECEPTORES PARA CININAS E SUA CONTRIBUIÇÃO PARA O STATUS REDOX CELULAR
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Fisiológicas da Universidade Federal de Sergipe como requisito à obtenção do grau de Mestre em Ciências Fisiológicas.
______________________________________________________ Orientadora: Drª. Sandra Lauton Santos
_____________________________________________________
Co-orientador: Dr. Luciano dos Santos Aggum Capettini
______________________________________________________ 1º Examinador: Drª. Silvia Carolina Guatimosim Fonseca
______________________________________________________
2º Examinador: Dr. Enilton Aparecido Camargo
______________________________________________________ 1º Examinador Suplente: Dr. Eduardo Antonio Conde Garcia
______________________________________________________
2º Examinador Suplente: Dr. Daniel Badauê Passos Junior

iv
Dedicatória Para as mulheres de minha vida, Minha Mãe, detentora de todo meu amor e admiração profunda. Minha Irmã, simplesmente amo. Minha Noiva, pelo amor, compreensão e dedicação ao nosso futuro. Ao meu Pai, ausência de corpo e presença de espírito diária.

v
AGRADECIMENTOS
Aos animais que doaram suas vidas inconscientemente em prol da evolução da ciência.
A toda minha família e amigos, que mesmo não sabendo ao certo o que eu fazia, e com
minha ausência, me compreendiam e apoiavam incondicionalmente.
À Drª. Sandra Lauton Santos, querida orientadora, que me ensinou mais do que
conhecimentos e técnicas, pela lição diária de compromisso com ensino e pesquisa. Agradeço
por provocar-me, também diariamente, que o limite é uma barreira que nós humanos
impomos e deve ser transposta, enfim, mas do que isto, a amizade foi e será um dos nossos
melhores bônus.
Ao Laboratório de Biofísica do Coração e seus membros, Dr. Eduardo Antônio Conde
Garcia, Drª. Carla Maria de Lins Vasconcelos, Drª. Evaleide Diniz de Oliveira, aos alunos de
mestrado, José Evaldo e os recentes mestres, Valéria Fernandes e Américo, aos alunos de
iniciação científica, Itamar Couto, Michel Santana, Jucilene Freitas e a todos que contribuíram
para o progresso de minha formação acadêmica.
Ao Dr. Jorge Luiz Pesquero, do Departamento de Fisiologia e Biofísica (ICB/UFMG)
e ao Dr. João Bosco Pesquero, do Departamento de Biofísica (UNIFESP), que gentilmente
forneceram os animais Knockouts para realização deste projeto.
À Drª. Silvia Carolina Fonseca Guatimosim e sua aluna de doutorado Aline Alves
Lara, do Departamento de Fisiologia e Biofísica (ICB – UFMG), pela valiosa contribuição
nos experimentos de microscopia confocal.
À Drª. Virgínia Soares Lemos, do Departamento de Fisiologia e Biofísica
(ICB/UFMG), Dr. Steyner França Côrtes e ao meu co-orientador, Dr. Luciano dos Santos
Aggum Capettini, do Departamento de Farmacologia (ICB/UFMG), por permitem livre
acesso aos seus respectivos laboratórios para realização de grande parte dos experimentos.
Aos membros da banca examinadora (PROASA, Qualificação e Defesa) pela
disponibilidade em contribuir com este trabalho.
Ao Programa de Pós-Graduação em Ciências Fisiológicas – PROCFIS, pela árdua
tarefa de implementar e consolidar este curso do qual orgulhosamente faço parte.
A CAPES-FAPITEC, pelo suporte durante todo período do mestrado.

vi
MODULAÇÃO DO SISTEMA CARDIOVASCULAR PELOS RECEPTORE S PARA CININAS E SUA CONTRIBUIÇÃO PARA O STATUS REDOX CELU LAR. Thássio
Ricardo Ribeiro Mesquita. São Cristovão, 2013
O sistema calicreína-cinina (SCC) é reconhecido por desempenhar funções que envolvem a manutenção da homeostase do sistema cardiovascular. O objetivo deste estudo foi avaliar a contribuição dos receptores para cininas na manutenção do status redox celular no sistema cardiovascular. Para isso, foi avaliada a função contrátil cardíaca (Langendorff) e vascular (reatividade vascular em banho de órgão isolado), além de medir, em coração e aorta torácica a produção de espécies reativas de oxigênio (ROS), nitrogênio (RNS), expressão de enzimas antioxidantes (SOD, CAT e GPx), assim como das que sintetizam o óxido nítrico (NO - eNOS e nNOS) e proteínas relacionadas com homeostase intracelular do cálcio (SERCA2a e NCX) em animais knockouts dos receptores B1 (B1
-/-) e B2 (B2-/-) para cininas. Para avaliar a
expressão proteica das enzimas antioxidantes, NOS (total e fosforilada), SERCA2a e NCX, foram realizados experimentos de western blot no coração. Foram também realizados experimentos de co-imunoprecipitação em pool de aorta torácica para identificação de proteínas complexadas aos receptores para cininas. Como resultados, identificamos redução na tensão sistólica cardíaca (B1
-/-: 53% e B2-/-: 52%), entretanto, não foram identificadas
alterações na tensão diastólica e frequência cardíaca, quando comparados ao grupo WT. Detectamos, em cardiomiócitos, aumento na geração do peróxido de hidrogênio (H2O2, B1
-/-: 75% e B2
-/-: 54%), radical superóxido (O2•-, B1
-/-: 40% e B2-/-: 41%) quando comparados ao
grupo WT. Estes resultados foram acompanhados pela redução na expressão de enzimas antioxidantes SOD (B1
-/-: 32% e B2-/-: 20%), CAT (B1
-/-: 34% e B2-/-: 39%) e GPx (B1
-/-: 29% e B2
-/-: 25%). Verificamos, ainda, aumento na produção do NO (B1-/-: 75% e B2
-/-: 36%) e sugerimos que esta alteração ocorre devido ao aumento na expressão e da atividade das NOS (eNOS: B1
-/-: 20% e B2-/-: 23% e nNOS: B1
-/-: 27% e B2-/-: 30%) e (peNOSSer1177: B1
-/-: 161% e B2
-/-: 154%; peNOSThr495: B1
-/-: 43% e B2-/-: 45%; pnNOS
Ser852: B1-/-: 25% e B2
-/-: 26%). Demonstramos ainda, redução da expressão da SERCA2a (B1
-/-: 41% e B2-/-: 46%) e aumento
na expressão do NCX (B1-/-: 16% e B2
-/-: 22%). Os resultados de Langendorff permitiram identificar elevada resistência coronariana nos grupos B1
-/- (38%) e B2-/- (38%), sugerimos que
a disfunção endotelial seja a principal causa dessa alteração. Confirmamos esta hipótese através de experimentos de reatividade vascular em aorta torácica, onde foi demonstrado aumento da resposta vasoconstritora (B1
-/-: 66% e B2-/-: 266%) e prejuízo na vasodilatadora
(B1-/-: 50% e B2
-/-: 56%), devido, principalmente, ao comprometimento da nNOS em aorta de animais dos grupos B1
-/- e B2-/-. Sugerimos que, em aorta, os receptores B1 e B2 estão
espontaneamente heterodimerizados com a nNOS e eNOS. Desta forma, propomos que a deleção dos receptores para cininas criam perturbações na atividade catalítica da nNOS favorecendo o estado de desacoplamento desta enzima e, consequentemente, comprometendo a biodisponibilidade do NO (B1
-/-: 30% e B2-/-: 20%) e gerando exacerbada de H2O2 (B1
-/-: 30% e B2
-/-: 52%) e O2•- (B1
-/-: 46% e B2-/-: 69%). Concluímos que os receptores para cininas
contribuem na homeostase da função contrátil do coração e aorta torácica, como também, regulam a produção de ROS e RNS em ambos os tecidos. Palavras Chave: Sistema Calicreína-Cinina; Sistema Cardiovascular; Espécies Reativas de
Oxigênio; Espécies Reativas de Nitrogênio.

vii
MODULATION OF CARDIOVASCULAR SYSTEM BY KININS RECEP TORS AND THEIR CONTRIBUTION TO CELLULAR REDOX STATUS. Thássi o Ricardo
Ribeiro Mesquita. São Cristovão, 2013 The kallikrein-kinin system (SCC) is known for performing functions that involve maintain homeostasis of cardiovascular system. The aim of this study was to evaluate the contribution of kinin receptors in maintaining the cellular redox status of the cardiovascular system. For this, the cardiac contractile function (Langendorff) and vascular (vascular reactivity in isolated organ bath) was evaluated, and measured, in heart and thoracic aorta the production of reactive oxygen species (ROS), nitrogen (RNS), expression of antioxidant enzymes (SOD, CAT and GPx), as well as those which synthesize nitric oxide (NO - eNOS and nNOS) and proteins related to intracellular calcium homeostasis (SERCA2a and NCX) in knockout animals B1 receptor (B1
-/-) and B2 (B2-/-) to kinins. To evaluate the protein expression of
antioxidant enzymes, NOS (total and phosphorylated), SERCA2a and NCX, western blot experiments were performed in heart. Were also performed, co-immunoprecipitation experiments in pool of the thoracic aorta to identify proteins complexed with kinins receptors. As results, were identified reduction in cardiac systolic tension (B1
-/-: 53% and B2-/-: 52%),
however, no changes in diastolic tension and heart rate were identified when compared to the WT group. We detected in cardiomyocytes increase of hydrogen peroxide (H2O2, B1
-/-: 75% and B2
-/-: 54%), superoxide radical generation (O2•-, B1
-/-: 40%, and B2-/-: 41%) compared to
the WT group. These findings were followed by reduction in expression of antioxidant enzymes SOD (B1
-/-: 32% and B2-/-: 20%), CAT (B1
-/-: 34% and B2-/-: 39%) and GPx (B1
-/-: 29% and B2
-/-: 25%). Also verified an increase in NO production (B1-/-: 75% and B2
-/-: 36%) and we suggest that this alteration occurred due increase in the expression and activity of NOS (eNOS: B1
-/-: 20% and B2-/-: 23% and nNOS: B1
-/-: 27% and B2-/-: 30%) and
(peNOSSer1177: B1-/-: 161% and B2
-/-: 154%; peNOSThr495: B1-/-: 43% and B2
-/-: 45%; pnNOSSer852: B1
-/-: 25% and B2-/-: 26%). We also demonstrated, reduction in expression of
SERCA2a (B1-/-: 41% and B2
-/-: 46%) and increases in expression of NCX (B1-/-: 16% and B2
-/-
: 22%). The Langendorff results allowed identifying high coronary resistance in groups B1-/-
(38%) and B2-/- (38%), suggesting, that endothelial dysfunction is the main cause of this
alteration. We confirm this hypothesis through vascular reactivity experiments in thoracic aorta, which was demonstrated increased vasoconstrictor response (B1
-/-: 66% and B2-/-:
266%) and impaired vasodilation (B1-/-: 50% and B2
-/-: 56%), mainly due to the involvement of nNOS in the aorta of animals in groups B1
-/- and B2-/-. We suggest that, in aorta, the B1 and
B2 receptors are in spontaneously heterodimerization with nNOS and eNOS. Thus, we propose that deletion of the kinin receptors creates disturbances in the catalytic activity of nNOS, favoring the uncoupling state of this enzyme, impairing bioavailability of NO (B1
-/-: 30% and B2
-/-: 20%) and exacerbated H2O2 generation (B1-/-: 30% and B2
-/-: 52%) and O2•- (B1
-
/-: 46% and B2-/-: 69%). We conclude that, kinin receptors contribute to homeostasis for
contractile function of the heart and thoracic aorta, but also regulate the production of ROS and RNS in both tissues. Key-words: Kallikrein-Kinin System, Cardiovascular System, Reactive Oxygen Species, Reactive Nitrogen Species.

viii
LISTA DE TABELAS E FIGURAS
Figura 1. Metabolismo peptídico do sistema calicreína-cinina...........................................03
Figura 2. Principais enzimas que metabolizam o peptídeo BK e seus respectivos sítios de
clivagem da molécula..............................................................................................................04
Figura 3. Representação esquemática da sequência de aminoácidos do receptor B1 em
humanos...................................................................................................................................05
Figura 4. Representação esquemática da sequência de aminoácidos do receptor B2 em
humanos...................................................................................................................................08
Figura 5. Registro típico de coração isolado de camundongo durante perfusão
retrógrada em sistema de Langendorff.................................................................................18
Figura 6. Genotipagem de camundongos knockouts para o receptor B1 para cininas (B1-/-
).................................................................................................................................................25
Figura 7. Genotipagem de camundongos knockouts para o receptor B2 para cininas (B2-/-
).................................................................................................................................................25
Figura 8. Variação da tensão sistólica em coração isolado de camundongos. Os registros
foram obtidos em corações de animais dos grupos selvagem (WT – Wild Type - n=5), e
knockouts para os receptores para cininas B1 (B1-/-- n=6) e B2 (B2
-/--
n=5)...........................................................................................................................................26
Figura 9. Variação da tensão diastólica em coração isolado de camundongos. Os
registros foram obtidos em corações de animais dos grupos selvagem (WT – Wild Type -
n=5), e knockouts para os receptores para cininas B1 (B1-/- - n=6) e B2 (B2
-/- -
n=5)...........................................................................................................................................27

ix
Figura 10. Variação da frequência cardíaca espontânea em coração isolado de animais
dos grupos selvagem (WT – Wild Type - n=4) e knockouts para os receptores para cininas
B1 (B1-/- - n=5) e B2 (B2
-/- - n=4)...............................................................................................27
Figura 11. Variação da pressão de perfusão em coração isolado de animais dos grupos
selvagem (WT – Wild Type - n=8) e knockouts para os receptores para cininas B1 (B1-/- -
n=6) e B2 (B2-/- - n=5)...............................................................................................................28
Figura 12. Produção basal do ânion superóxido em cardiomiócitos dos animais dos
grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1-/-)
e B2 (B2-/-)..................................................................................................................................29
Figura 13. Produção basal de peróxido de hidrogênio em cardiomiócitos dos animais dos
grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1-/-)
e B2 (B2-/-)..................................................................................................................................30
Figura 14. Produção basal de óxido nítrico em cardiomiócitos dos animais dos grupos
selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1-/-) e B2
(B2-/-)..........................................................................................................................................31
Figura 15. Determinação da expressão proteica da enzima Cu/Zn Superóxido Dismutase
(Cu/Zn SOD) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e
knockouts para os receptores para cininas B1 (B1-/-) e B2 (B2
-/-
).................................................................................................................................................32
Figura 16. Determinação da expressão proteica da enzima catalase em tecido cardíaco
dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para
cininas B1 (B1-/-) e B2 (B2
-/-)......................................................................................................33
Figura 17. Determinação da expressão proteica da enzima Glutationa Peroxidase (GPx)
em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para
os receptores para cininas B1 (B1-/-) e B2 (B2
-/-).....................................................................34

x
Figura 18. Determinação da expressão proteica da enzima Óxido Nítrico Sintase
endotelial (eNOS) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild
Type) e knockouts para os receptores para cininas B1 (B1-/-) e B2 (B2
-/-).............................35
Figura 19. Avaliação dos níveis de fosforilação da enzima Óxido Nítrico Sintase
endotelial (peNOS Ser1177) em tecido cardíaco dos animais dos grupos selvagem (WT –
Wild Type) e knockouts para os receptores para cininas B1 (B1-/-) e B2 (B2
-/-
).................................................................................................................................................36
Figura 20. Avaliação dos níveis de fosforilação da enzima Óxido Nítrico Sintase
endotelial fosforilada (peNOS Thr495) em tecido cardíaco dos animais dos grupos
selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1-/-) e B2
(B2-/-)..........................................................................................................................................37
Figura 21. Determinação da expressão proteica da enzima Óxido Nítrico Sintase
neuronal (nNOS) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type)
e knockouts para os receptores para cininas B1 (B1-/-) e B2 (B2
-/-)........................................38
Figura 22. Avaliação dos níveis de fosforilação da enzima Óxido Nítrico Sintase
neuronal fosforilada (pnNOS Ser852) em tecido cardíaco dos animais dos grupos selvagem
(WT – Wild Type) e knockouts para os receptores para cininas B1 (B1-/-) e B2 (B2
-/-
).................................................................................................................................................39
Figura 23. Determinação da expressão proteica do trocador sódio/cálcio (NCX) em
tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os
receptores para cininas B1 (B1-/-) e B2 (B2
-/-)..........................................................................40
Figura 24. Determinação da expressão proteica da Cálcio ATPase do Retículo
Sarcoplasmático (SERCA2a) em tecido cardíaco dos animais dos grupos selvagem (WT
– Wild Type) e knockouts para os receptores para cininas B1 (B1-/-) e B2 (B2
-/-
).................................................................................................................................................41

xi
Figura 25. Efeito vasorrelaxante induzido pela acetilcolina (ACh) em anéis de aorta de
camundongos dos grupos selvagem (WT – Wild Type - n=21) e knockouts para os
receptores para cininas B1 (B1-/- - n=10) e B2 (B2
-/- - n=13)..................................................42
Figura 26. Efeito vasoconstritor induzido pela fenilefrina (Phen) em anéis de aorta de
camundongos dos grupos selvagem (WT – Wild Type - n=8) e knockouts para os
receptores para cininas B1 (B1-/- - n=5) e B2 (B2
-/- - n=4).......................................................43
Figura 27. Efeito da inibição não seletiva das NOS pelo L-NNA (100 µmol.L-1) sobre a
resposta vasorrelaxante induzida pela acetilcolina (ACh) em anéis de aorta de
camundongos do grupo selvagem (WT – Wild Type - n=5 / L-NNA -
n=5)...........................................................................................................................................44
Figura 28. Efeito da inibição seletiva da nNOS sobre a resposta vasorrelaxante induzida
pela acetilcolina (ACh) em anéis de aorta de camundongos do grupo selvagem (WT –
Wild Type - n=8 / TRIM - n=8)...............................................................................................44
Figura 29. Efeito da inibição não seletiva das NOS pelo L-NNA (100 µmol.L-1) sobre a
resposta vasorrelaxante induzida pela acetilcolina (ACh) em anéis de aorta de
camundongos knockout para receptor B1 para cininas (B1-/- - n=8 / L-NNA -
n=8)...........................................................................................................................................45
Figura 30. Efeito da inibição não seletiva das NOS pelo L-NNA (100 µmol.L-1) sobre a
resposta vasorrelaxante induzida pela acetilcolina (ACh) em anéis de aorta de
camundongos knockout para receptor B2 para cininas (B2-/- - n=5 / L-NNA -
n=5)...........................................................................................................................................46
Figura 31. Efeito da inibição seletiva da nNOS pelo TRIM (300 µmol.L -1) sobre a
resposta vasorrelaxante induzida pela acetilcolina (ACh) em anéis de aorta de
camundongos knockout para receptor B1-/- para cininas (B1
-/- - n=5 / TRIM -
n=5)...........................................................................................................................................46

xii
Figura 32. Efeito da inibição seletiva da nNOS pelo TRIM (300 µmol.L -1) sobre a
resposta vasorrelaxante induzida pela acetilcolina (ACh) em anéis de aorta de
camundongos knockout para receptor B2 para cininas (B2-/- - n=5 / TRIM -
n=5)...........................................................................................................................................47
Figura 33. Produção basal de óxido nítrico em anéis de aorta torácica de camundongos
dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1
(B1-/-) e B2 (B2
-/-)........................................................................................................................48
Figura 34. Produção basal de peróxido de hidrogênio em anéis da aorta torácica de
camundongos dos grupos selvagem (WT – Wild Type) e knockouts para os receptores
para cininas B1 (B1-/-) e B2 (B2
-/-).............................................................................................49
Figura 35. Produção basal de ânion superóxido em anéis da aorta torácica de
camundongos dos grupos selvagem (WT – Wild Type) e knockouts para os receptores
para cininas B1 (B1-/-) e B2 (B2
-/-).............................................................................................50
Figura 36. Imunoprecipitação do complexo proteína-proteína nNOS/B1 e nNOS/B2 em
aorta torácica de camundongo selvagem (WT – Wild Type)...............................................51
Figura 37. Imunoprecipitação do complexo proteína-proteína eNOS/B1 e eNOS/B2 em
aorta torácica de camundongo selvagem (WT – Wild Type)...............................................51

xiii
LISTA DE ABREVIATURAS E SIGLAS
AA Ácido Araquidônico ACh Acetilcolina AP-1 Activator Protein-1 Akt Proteína Cinase B Ang II Angiotensina II ANOVA Análise de variância (Analyses of variance) B1R Receptor B1 B1
-/- Animais knockout para os receptores B1 de cininas B2R receptor B2 B2
-/- Animais knockout para os receptores B2 de cininas BH4 Tetrahidrobiopterina (cofator da NOS) BK Bradicinina BPM Batimentos por Minuto COX Ciclooxigenases cAMP AMP cíclico (adenosina monofosfato cíclico) cDNA DNA complementar (ácido desoxiribonucleico complementar) cGMP GMP cíclico (guanosina monofosfato cíclico) Cu/Zn SOD Cobre-Zinco Superoxido Dismutase DBK Des-Arg9-Bradicinina DAF-FM 4-amino5-metilamino-2,7difluoresceina diacetato DAG Diacilglicerol DCF 2',7'-Dichlorofluorescein DHE Dihydroethidium DLBK Des-Arg10 Lys-Bradicinina DNA Ácido desoxiribonucléico (Deoxyribose Nucleic Acid) DTT Ditiotreitol ECA Enzima Conversora de Angiotensina I EDHF Fator hiperpolarizante derivado do endotélio (Endothelium derived hiperpolarizing factor) EDTA Ácido etilenediaminotetraacético ELISA Enzyme-linked Immunosorbent Assay EMAX Efeito Máximo eNOS Óxido nítrico sintase endotelial; NOS III (endothelial nitric oxide synthase) EOX Epoxigenases e.p.m. Erro padrão da média EPN Endopeptidase Neutra RNS Espécies Reativas de Nitrogênio (reactive nitrogen species) ROS Espécies Reativas de Oxigênio (reactive oxygen species) ERK 1/2 Extracellular Signal-Regulated Kinases GPx Glutationa Peroxidase GRKs Quinases de Receptores Acoplados à Proteína G (G-Protein Coupled Receptor kinases) H2O2 Peróxido de hidrogênio HCl Ácido clorídrico HEPES N-2-Hidroetilpiperazina-N'-2-ácido etanosulfônico

xiv
(N-2-Hydroxyethylpiperazine-N'-2-ethanesulfonic Acid) H2DCFH-DA 2',7'-Dichlorodihydrofluorescein Diacetate HSP-90 Heat Shock Protein 90 HRP Horse radish peroxidase IL-1β Interleucina-1 beta iNOS Óxido nítrico sintase induzida ou indutivel; NOS II (inducible nitric oxide synthase) IP3 Inositol 1,4,5 Trifosfato JAK/STAT Janus Kinase K+ Íon potássio KATP Canal para potássio sensível a ATP KCa2+ Canal para potássio regulado por cálcio L-Arg L-Arginina L-NAME NG-nitro-L-arginina metil éster (NG-nitro-L-Arginine MethylEster) L-NNA NG-nitro-L-arginina (NG-Nitro-L-Arginine) LOX Lipooxigenases Lys-BK Calidina MAPK Mitogen-Activated Protein Kinase MnSOD Superóxido Dismutase Mitocondrial Manganês mtDNA DNA mitocondrial NF-κB Factor Nuclear Kappa B mRNA RNA mensageiro NADP Nicotinamida adenina difosfato NADPH Nicotinamida adenina difosfato reduzida NADPH oxidase Nicotine Adenine Dinucleotide Phosphate oxidase NCX Trocador sódio-cálcio nNOS Óxido nítrico sintase neuronal; NOS I (neuronal nitric oxide synthase) NO Óxido nítrico (Nitric Oxide) NOS Óxido nítrico sintase (nitric oxide synthase) O2
•- Ânion superóxido OD Densidade óptica (Optical Density) ODQ 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one (inibidor de guanilil ciclase) OH• Radical hidroxil OONO- Peroxinitrito pb Pares de base PBS Solução tamponada de potássio (Phosphate Buffer Solution) PCR Reação da polimerase em cadeia (Polymerase Chain Reaction) PGI2 Prostaglandina I2 (prostaciclina) pH Potencial hidrogeniônico Phen Fenilefrina Pi3-K Fosfatidilinositol-3-Cinase PIP2 Fosfatidilinositol 4,5- Bifosfato PKA Proteína cinase A PKC Proteína cinase C PKG Proteína cinase G PLA2 Fosfolipase A2 PLC Fosfolipase Cγ

xv
PLB Fosfolamban PLC β Fosfolipase Cβ PLI2 Prostaglandina I2
PMSF Fluoreto de Fenilmetilsilfonila RNA Ácido ribonucléico (RiboNucleic Acid) RPM Rotações por minuto RS Retículo sarcoplasmático RyR2 Receptor de rianodina SCC Sistema Calicreína-Cinina SDS-PAGE Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis SERCA2a Cálcio ATPase do retículo sarcoplasmático de subtipo 2a SOD Superóxido dismutase SNAP Nitroprussiato de sódio (sodium nitroprusside) SRA Sistema Renina Angiotensina TBE Tampão Tris - Ácido bórico - EDTA TEMED N,N,N’,N’-Tetramethylethylenediamine TRIM l-(2-Trifluoromethylphenyl)imidazole (inibidor seletivo da nNOS) u.a. Unidades arbitrárias (arbitraries units) WT Tipo selvagem – Wild type XO Xantina oxidase

xvi
SUMÁRIO
1 INTRODUÇÃO....................................................................................................................01
2 REVISÃO DA LITERATURA..........................................................................................02
2.1 Metabolismo de Cininas..........................................................................................02
2.2 Estrutura, Sinalização Intracelular do Receptor B1 e Sistema Cardiovascular.......04
2.3 Estrutura, Sinalização Intracelular do Receptor B2 e Sistema Cardiovascular.......08
2.4 Sinalização Das Espécies Reativas de Oxigênio e Nitrogênio em Cardiomiócito..11
3 OBJETIVOS.........................................................................................................................14
3.1 Objetivo Geral.........................................................................................................14
3.2 Objetivos Específicos..............................................................................................14
4 MATERIAIS E MÉTODOS...............................................................................................15
4.1 Grupos Experimentais.............................................................................................15
4.2 Genotipagem...........................................................................................................15
4.3 Procedimento Cirúrgico para Retirada do Coração................................................16
4.4 Estudo dos Parâmetros Cardíacos..........................................................................17
4.5 Obtenção de Células Cardíacas Isoladas................................................................18
4.6 Experimentos de Reatividade Vascular..................................................................18
4.6.1 Obtenção e Montagem dos Anéis de Aorta.............................................18
4.6.2 Avaliação da Integridade Endotelial........................................................19
4.6.3 Remoção do Endotélio Funcional............................................................20
4.6.4 Curva de Contração Induzida pela Fenilefrina.........................................20
4.6.5 Participação da Via das Óxido Nítrico Sintases na Resposta
Vasorrelaxante à Acetilcolina............................................................................20
4.7 Mensuração dos Níveis Intracelulares das Espécies Reativas de Oxigênio............20
4.8 Mensuração dos Níveis Intracelulares do Óxido Nítrico........................................21

xvii
4.9 Extração de Proteínas do Tecido.............................................................................22
4.10 Determinação da Concentração Total de Proteínas...............................................22
4.11 Determinação da Expressão Proteica por Western Blot........................................22
4.12 Co-imunoprecipitação...........................................................................................23
4.13 Análises Estatísticas..............................................................................................24
5 RESULTADOS ....................................................................................................................25
6 DISCUSSÃO.........................................................................................................................52
7 CONCLUSÃO......................................................................................................................66
8. CONSIDERAÇÕES FINAIS.............................................................................................66
9. REFERENCIAL BIBLIOGRÁFICO...............................................................................68

1
1. INTRODUÇÃO
Os peptídeos e os derivados metabólicos ativos do sistema calicreína-cinina
(SCC) exercem uma variedade de efeitos biológicos. Estes efeitos ocorrem através da ativação
da superfamília de receptores acoplados à proteína G, atuando sobre um complexo conjunto
de vias de sinalização intracelular. Estas vias regulam diversas funções fisiológicas e
fisiopatológicas. A ação das cininas é mediada após a ligação destes peptídeos em dois
receptores distintos, B1 (B1R) e B2 (B2R). A literatura clássica aponta que os B2R são
constitutivamente expressos em condições fisiológicas basais, enquanto, os B1R tem sua
expressão reduzida nestas mesmas condições, porém, tem aumento significativo em algumas
situações patológicas (MARCEAU et al., 1998).
Durante o estado inflamatório, como, asma (BARNES, 1992), alergia (POLOSA,
1993) e após traumas e lesões, as cininas promovem a migração de células do sangue para os
tecidos lesionados a fim de ativar vários componentes anti-inflamatórios, tais como:
mastócitos, fibroblastos, macrófagos e outras células do sistema imune (BHOOLA et al.,
1992).
No endotélio, as funções fisiológicas deste sistema promovem a liberação de
moléculas sinalizadoras, como óxido nítrico (NO), prostaciclinas e fator hiperpolarizante
derivado do endotélio (REGOLI e BARABÉ, 1980; TODA et al., 1987; SCHINI et al., 1990;
MOMBOULI e VANHOUTTE, 1995), produzindo, na circulação periférica, a vasodilatação e
aumento do fluxo sanguíneo através da redução do tônus da musculatura arterial lisa.
No coração, o SCC está envolvido em vias de sinalização intracelulares que
regulam a mobilização do cálcio e consequentemente regulação da maquinaria contrátil
(LAUTON-SANTOS, 2007; ROMAN-CAMPOS et al., 2010). Sua ação estende-se ainda
sobre o músculo liso visceral e a quase todos os órgãos controlados pelos sistemas nervoso
autônomo simpático (TOUSIGNANT et al., 1987) e parassimpático (LOPES e COUTURE,
1992).
A literatura aponta a relação entre o SCC e a produção de moléculas sinalizadoras
como as espécies reativas de oxigênio (ROS) (OLDENBURG et al., 2004) e nitrogênio (RNS)
(LAUTON-SANTOS, 2007; ROMAN-CAMPOS et al., 2010), propondo a participação
destas, em vias de sinalização intracelular que modulam a função cardiovascular durante o
estado fisiológico.

2
A literatura ainda é inconclusiva sobre a participação de ROS e RNS como
moléculas sinalizadoras do SCC e ainda, mais distante, que estas moléculas sejam capazes de
regular a dinâmica de cálcio intracelular. Contudo, este trabalho propõe avaliar a participação
dos receptores para cininas na produção de ROS e RNS no sistema cardiovascular, para tentar
contribuir na produção de novos conhecimentos sobre estas vias de sinalização e sua
influência na manutenção da homeostase cardíaca e vascular.
2. REVISÃO DA LITERATURA
2.1 Metabolismo de Cininas
O SCC é constituído por um componente plasmático e outro tecidual. O
nonapeptídeo bradicinina (BK) e o decapeptídeo calidina (Lys-BK) são os principais
peptídeos derivados da proteólise de seu precursor inativo, o cininogênio, através da ação
catalítica das enzimas cininogenases. A principal e mais potente cininogenase é a calicreína,
existindo nas formas plasmática e tecidual (Figura 1). Ambos os peptídeos produzidos pela
ação dessas enzimas são agonistas do receptor B2, tendo, porém, afinidades diferentes para o
mesmo. A cinina liberada pela calicreína plasmática humana, de rato ou camundongo é a BK,
entretanto, as calicreínas teciduais em humano produzem Lys-BK (BHOOLA et al., 1992).
No plasma, a BK pode, ainda, ser sintetizada através de outras cininogenases, como por
exemplo, a tripsina, uropepsina ou plasmina. Nos rins, a calicreína tecidual é a principal
cininogenase (CARRETERO e SCICLI, 1988). Estas enzimas diferem, dentre outros
aspectos, em sua massa molecular e no tipo de cininas sintetizadas (MARCEAU et al., 1998).
A BK apresenta meia vida curta, aproximadamente 15 segundos, e seus níveis
circulantes são relativamente baixos, variando entre 0,2–7,1 pM (NUSSBERGER et al, 1999;
MOREAU et al, 2005). Os peptídeos BK e Lys-BK são rapidamente hidrolisados pelas
enzimas cininases I e cininases II (Figura 1), as primeiras, compreendem as carboxipeptidases
M (membrana) e N (plasmática), e as cininases II compreendem a enzima conversora de
angiotensina I (ECA) e a endopeptidase neutra (EPN) (Figura 1). A carboxipeptidase N é
produzida no fígado e secretada na circulação, esta enzima é responsável pela degradação de
cerca de 90% da BK no plasma. A carboxipeptidase M é uma enzima de membrana
plasmática, encontrada no rim humano, pulmão e placenta (SKIDGEL e ERDÖS, 1998). Em
decorrência de sua localização na superfície celular, ancorada ao B1R, esta enzima
desempenha um importante papel no reconhecimento e sensibilização da interação agonista-
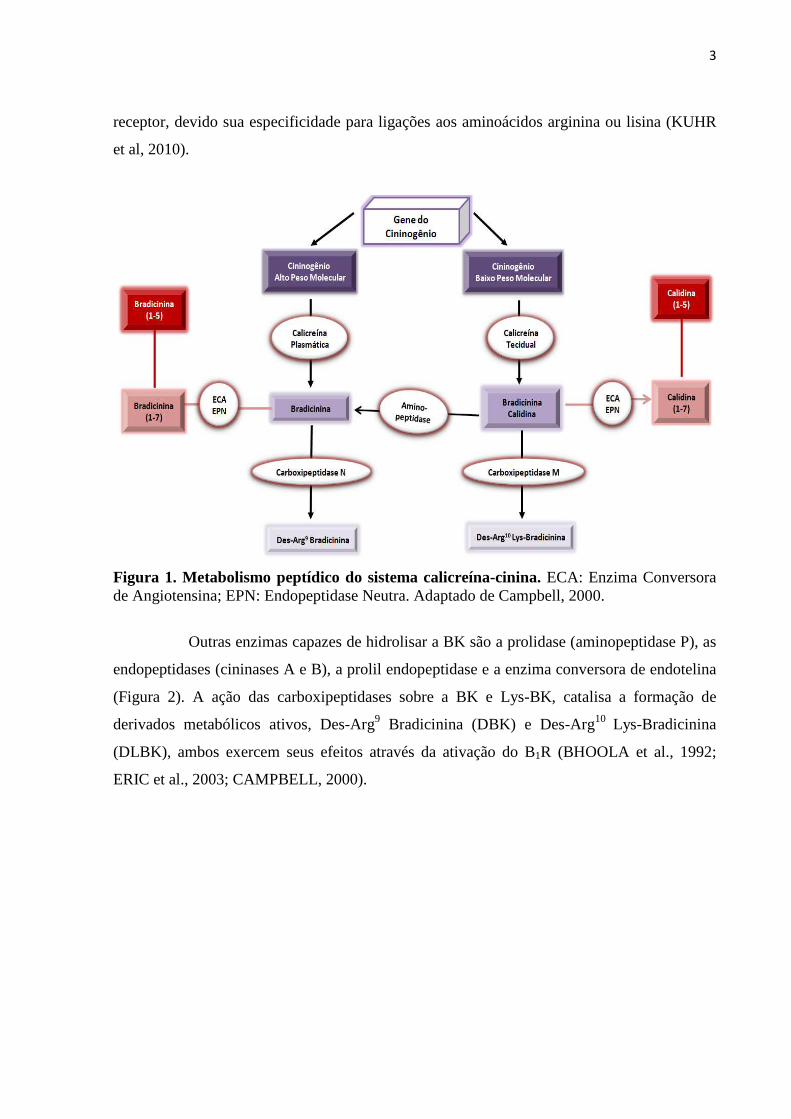
3
receptor, devido sua especificidade para ligações aos aminoácidos arginina ou lisina (KUHR
et al, 2010).
Figura 1. Metabolismo peptídico do sistema calicreína-cinina. ECA: Enzima Conversora de Angiotensina; EPN: Endopeptidase Neutra. Adaptado de Campbell, 2000.
Outras enzimas capazes de hidrolisar a BK são a prolidase (aminopeptidase P), as
endopeptidases (cininases A e B), a prolil endopeptidase e a enzima conversora de endotelina
(Figura 2). A ação das carboxipeptidases sobre a BK e Lys-BK, catalisa a formação de
derivados metabólicos ativos, Des-Arg9 Bradicinina (DBK) e Des-Arg10 Lys-Bradicinina
(DLBK), ambos exercem seus efeitos através da ativação do B1R (BHOOLA et al., 1992;
ERIC et al., 2003; CAMPBELL, 2000).

4
Figura 2. Principais enzimas que metabolizam o peptídeo BK e seus respectivos sítios de clivagem da molécula. ECA: Enzima Conversora de Angiotensina. Adaptado de Campbell, 2000.
2.2 Estrutura, Sinalização Intracelular do Receptor B1 e Sistema Cardiovascular
A descrição molecular do B1R iniciou-se com o sequenciamento em humanos
(MENKE et al., 1994), e posteriormente, em camundongos (PESQUERO et al., 1996) e ratos
(NI et al., 1999; JONES et al., 1999). A partir dos estudos destes receptores em humanos,
ficou conhecido que o B1R é composto por 334 aminoácidos e possui massa molecular de
aproximadamente 40 kDa, como representado na Figura 3 (PESQUERO et al., 1996;
MARCEAU et al., 1998).

5
Figura 3. Representação esquemática da sequência de aminoácidos do receptor B1 em humanos (Adaptado de LEEB-LUDBERG et al., 2005). AI-1: alça intracelular 1; AI-3: alça intracelular 2; AI-3: alça intracelular 3; AE-1: alça extracelular 1; AE-2: alça extracelular 2; AE-3: alça extracelular 3.
As características estruturais dos B1Rs os define como pertencentes a classe de
receptores metabotrópicos, acoplados à superfamília de proteínas G, exibindo uma arquitetura
molecular com 7 alfa-hélices transmembranares. A sinalização intracelular desencadeada pelo
B1R assemelha-se, em muitos aspectos, à do B2R, com a participação de proteínas Gq e Gi
acopladas ao receptor. Entretanto, a cascata de segundos mensageiros intracelulares associada
à sinalização do B1R envolve a ativação da fosfolipase C (PLC), a partir da hidrólise de um
fosfolipídeo da membrana celular, o fosfatidilinositol 4,5-bifosfato (PIP2) em inositol 1,4,5 -
trifosfato (IP3) e diacilglicerol (DAG). Estes segundos mensageiros estão envolvidos em
múltiplas funções celulares, como na ativação da proteína quinase C (PKC), que está
envolvida no aumento da influxo de cálcio (Ca2+), liberação de ácido araquidônico (AA),
produção de eicosaóides, e ativação da óxido nítrico sintase endotelial (eNOS) com
consequente produção do NO (TSUTSUI et al., 2000).

6
A partir de ensaios de binding em tecidos humanos (HESS et al., 1992;
SCHNECK et al., 1994) e em preparações in vivo (BHOOLA et al., 1992; LINZ et al., 1995)
foi demonstrado que os B1Rs e B2Rs diferem significativamente, no que se refere às respostas
induzidas pelas diferentes cininas. O B2R é ativado por BK e Lys-BK, e o B1R é mais sensível
aos peptídeos metabólitos gerados pela ação da cininase I e II, DBK e DLBK (MARCEAU et
al., 1998).
A afinidade das diferentes cininas aos seus respectivos receptores deve-se à
presença de um resíduo de lisina na posição 118 da porção C-terminal, localizada no terceiro
domínio transmembrânico do B1R. Este resíduo repele cargas positivas da BK e Lys-BK que
contém resíduos de arginina na posição 9, tornando o B1R seletivo para a DBK e ao DLBK
(MARCEAU et al., 1998).
O desenvolvimento de antagonistas seletivos tem sido crucial para definir as
respostas farmacológicas dos subtipos de receptores para cininas, entretanto, ainda no século
XXI se busca desenvolver antagonistas mais seletivos e potentes para o B1R. O primeiro
protótipo dos antagonistas de B1R foi o [Leu8] des-Arg9-BK (REGOLI et al., 1977), o qual
posteriormente, foi demonstrado não ser biologicamente ativo em bioensaios contráteis em
útero de rata, em íleo de cobaia ou em ensaios in vivo (REGOLI e BARABÉ, 1980).
Até o momento, os principais antagonistas para o B1R são de natureza peptídica.
Por apresentarem elevada seletividade para o B1R, são criticamente importantes para as
investigações de eventos fisiopatológicos e aplicações terapêuticas. Entretanto, peptídeos
sofrem várias desvantagens como potenciais agentes terapêuticos, tais como instabilidade
metabólica, baixa absorção oral, rápida eliminação, curta duração da ação e, em alguns casos,
dependendo da concentração, atividade como agonista parcial (MARCEAU et al., 1998).
Dentre os novos antagonistas, destaca-se o des-arg10 HOE 140, por exibir elevada
afinidade para o B1R em todas as espécies estudadas (ratos, camundongos, coelho e cobaia),
em ensaios de funcionalidade in vitro (RHALEB, GOBEIL e REGOLI, 1992) e in vivo
(LAGNEUX e RIBUOT, 1997), entretanto, alta concentrações e prolongada exposição à este,
é capaz de produz derivados metabólitos residuais que, dentre alguns efeitos, antagonizam o
B2R.
Os receptores para cininas apresentam diferentes padrões de expressão em
condições fisiológicas basais. Classicamente, é conhecido que o B2R é abundantemente
expresso, de forma constitutiva, em diferentes tecidos, incluindo endotélio vascular, músculo
liso vascular e cardiomiócitos (FELIPE et al., 2007). Os B1Rs são, geralmente, pouco
expressos em tecidos saudáveis, mas rapidamente expressos em situações de lesão tecidual e

7
inflamação (MARCEAU et al., 1998). Entretanto, artigos mais recentes demonstraram que
ambos os receptores para cininas desempenham funções no controle da homeostase
cardiovascular em estado fisiológico e fisiopatológico (LAUTON-SANTOS et al., 2007;
FELIPE et al., 2007).
Vários estudos revelam a expressão do B1R em fibras simpáticas do miocárdio
(FOUCART et al., 1997), em fibras do sistema de condução do coração (GOUIN et al., 1996;
FERNANDES et al., 2006), e sabe-se que sua ativação é capaz de prolongar a duração do
potencial de ação cardíaco em ratos (GOUIN et al., 1996). O significado fisiológico da
presença e sinalização do B1R em tecido cardíaco pode ser ainda mais complexa. Estudos tem
demonstrado relação destes receptores com o reparo tecidual (TSCHÖPE et al., 2000) e
redução da área infartada após lesão por isquemia no miocárdio (LAGNEUX, ADAM,
LAMONTAGNE, 2003).
Os B1R são regulados por uma série de citocinas presentes no processo
inflamatório, e outros fatores, tais como proteína quinase ativada por mitógeno (do inglês
mitogen-activated protein kinase - MAPK) (LARRIVÉ et al., 1998) e o fator nuclear kappa B
(do inglês nuclear factor kappa B - NF-κB) (SCHANSTRA et al., 1998; NI et al., 1998).
Alguns autores sugerem que o B2R esteja relacionado à resposta inflamatória
aguda, enquanto o B1R, à processos inflamatórios crônicos ou prolongados (McLEAN et al.,
1999). Esta hipótese foi levantada devido ao B2R apresentar a capacidade de regulação
negativa, rapidamente internalizando e indisponibilizando ambos, o receptor e a proteína G
(AUSTIN et al., 1997; WEERD e LEEB-LUNDBERG, 1997).
Por outro lado, o B1R exibe dessensibilização bastante limitada. Foi demonstrado
o importante papel de alguns aminoácidos de sua cauda C-terminal citoplasmática no
processo de internalização do receptor. Observou-se que resíduos de aminoácidos serina e
treonina, nesta referida localização, são necessários para o mecanismo de dessensibilização e
internalização do receptor, via quinases, acopladas à proteína G (do inglês G-protein coupled
receptor kinases - GRKs) (BLAUKAT et al.,1999; FAUSSNER et al., 1998).
Além disso, o mecanismo de ativação do B1R também diferere do B2R, já que, em
células humanas, a fosforilação do B1R é inexistente na presença ou ausência do agonista
(BLAUKAT et al., 1999). Esta diferença no mecanismo de ativação dos receptores para
cininas certamente contribui para uma estimulação mais duradoura por parte do B1R e, por
conseguinte, das distintas vias de sinalizações intracelulares associadas aos mesmos, em
diferentes tipos celulares (LEEB-LUNDBERG et al., 2005).

8
2.3 Estrutura, Sinalização Intracelular do Receptor B2 e Sistema Cardiovascular
A codificação molecular, o sequenciamento e a clonagem dos genes que
expressam os receptores para cininas iniciaram-se com a clonagem do B2R em ratos
(McEACHERN et al. 1991) e em diferentes espécies de mamíferos, inclusive o homem
(Figura 4) (HESS et al. 1994).
Figura 4. Representação esquemática da sequência de aminoácidos do receptor B2 em humanos (Adaptado de LEEB-LUDBERG et al., 2005). AI-1: alça intracelular 1; AI-3: alça intracelular 2; AI-3: alça intracelular 3; AE-1: alça extracelular 1; AE-2: alça extracelular 2; AE-3: alça extracelular 3.
A clonagem de cDNA do B2R em camundongos, ratos e humanos revelou grande
similaridade desta molécula nas diferentes espécies. O cDNA de camundongos é 92%
idêntico ao de ratos e 84% ao de humanos (McEACHERN et al., 1991; HESS et al., 1994;
PAQUET et al., 1999). Todavia, a semelhança entre as sequências de aminoácidos entre estes
dois receptores para cininas, em humanos, é relativamente baixa, apresentando apenas 36% de
homologia (LEEB-LUNDBERG et al., 2005).

9
Em camundongos e ratos, o B2R é constituído por 366 aminoácidos e apresenta
uma massa molecular de aproximadamente 69 kDa (FIGUEROA et al., 2001) e, ainda, baixa
homologia (30%) ao B1R (PESQUERO et al., 1996).
Devido à sua elevada expressão em diferentes tecidos, a maioria dos efeitos in
vivo, das cininas são atribuídos ao B2R, incluindo broncoconstrição, hipotensão, reações
inflamatórias agudas, dor e hiperalgesia (STERANKA e BURCH, 1991; HALL, 1992). As
respostas fisiológicas das cininas mediadas pelo B2R, são caracterizados por sua elevada
afinidade pela BK e por sua sensibilidade, mesmo a baixas concentrações do antagonista
sintético seletivo HOE 140 (REGOLI et al., 1998). A ativação dos B2R exercem importantes
efeitos em numerosas preparações in vivo, ex vivo e in vitro, incluindo o trato gastrointestinal,
cardiovascular, genito-urinário, respiratório, neural e em tecidos oculares (revisto por HALL,
1992).
A ativação do B2R e o consequente acionamento dos mecanismos de transdução
de sinal, envolvem uma complexa cascata de sinalização intracelular que se inicia com a
ligação do agonista ao seu receptor na membrana celular. A ligação do agonista ao B2R ativa
a enzima PLC, entretanto, é descrito sua interação com várias proteínas G, como Gαi
(EWALD et al, 1989, LINDER et al, 1990), Gαs (LIEBMANN et al., 1996), e Gα12/13
(GOHLA et al., 1999). A resposta resultante desta ação promove a síntese de DAG e IP3 que,
respectivamente, conduzem à ativação da PKC e à mobilização de Ca2+ a partir do retículo
sarcoplasmático.
A ativação do receptor pode, ainda, promover o influxo de Ca2+ do meio
extracelular através de canais catiônicos não seletivos. A modulação direta de canais iônicos é
também descrita e parece criar um complexo fenômeno de hiperpolarização envolvendo a
abertura de canais para potássio (OLDENBURG et al., 2004). Devido ao seu grande espectro
de distribuição em vários tecidos, os B2R estão ainda envolvidos em vias de produção do NO,
da adenosina monofosfato cíclica (cAMP), liberação de AA, produção de eicosanóides e
prostaglandina I2 (PGI2) (FIELD et al., 1994; YANO et al.,1985; FASOLATO et al., 1988).
Não somente a ativação do B1R, mas também, a do B2R, está associada com a
translocação de fatores de transcrição nuclear, que regulam múltiplas funções celulares, como
a produção de citocinas e apoptose envolvidas na lesão tecidual após reperfusão de região
isquêmica-cardíaca (XIE et al., 2000). A BK é capaz de ativar o NF-κB com subsequente
ativação da fosfatidilinositol-3-quinase/proteína quinase B (Pi3-K/Akt) (XIE et al., 2000),
bem como, RhoA quinase (PAN et al., 1998).

10
Em culturas de células de músculo liso, a BK estimula aumento da expressão de
interleucina 8 (IL-8), por ativação de vias de sinalização dependentes ou não de prostanóides
(ZHU et al., 2003). No acionamento independente dos prostanóides, envolve a ativação de
NF-κB, enquanto que a ativação dependente de prostanóides envolve o fator de transcrição
proteína ativadora 1 (activator protein-1 AP-1: c-jun/c-fos), ativação da quinase regulada por
sinal extracelular (extracellular-signal-regulated kinases - ERK 1/2) (EL-DAHR et al., 1998)
e espécies reativas de oxigênio (ROS) (GREENE,VELARDE e JAFFA, 2000).
Em cultura de células endoteliais, a BK é eficaz na ativação do complexo
enzimático da eNOS e concomitante produção de NO, através de mecanismos dependentes do
aumento da concentração do Ca2+ intracelular (BUSSE e FLEMING, 1995). Nestas células, a
BK promove, também, a fosforilação da MAPK (FLEMING, FISSLHALTER e BUSSE,
1995), PLCγ (FLEMING, FISSLHALTER e BUSSE, 1996), proteína de choque térmico 90
(do inglês heat shock protein 90 - Hsp90) (HARRIS et al., 2000), Janus quinase (JAK/STAT)
(JU et al., 2000) e inclusive do próprio B2R (LEEB-LUNDBERG et al. 2005).
A difusão do NO para a camada muscular promove, dentre outras respostas, a
hiperpolarização da membrana das células musculares lisas, através da abertura de canais para
K+, promovendo, por fim, a vasodilatação (BUSSE e FLEMING, 1995). A resposta
vasorelaxante à BK e DBK é controversa, e, fatores como concentração do agonista e a
espécie de animal utilizada experimentalmente, se apresentam como determinantes das
diferentes respostas apresentadas.
Neste sentido, Gobeil et al. (1996a), em ensaio in vitro, com tiras de veia
umbilical humana, demonstraram que a BK foi capaz de promover a contração do músculo
liso, sendo sugerido a participação de autacóides e a produção de prostanóides na resposta
vasoconstritora (REGOLI e BARABÉ, 1980). Recentemente, foi demonstrado, em
experimento de reatividade vascular utilizando anéis de aorta abdominal de camundongos,
que a BK e DBK são vasoconstritores capazes de promover contrações de 51% e 30%,
respectivamente (FELIPE et al., 2007).
Com a disponibilização do HOE 140, muito se pode conhecer acerca das vias de
sinalização das cininas, bem como, a discriminação entre as mesmas, e, ainda, a importância
das respostas produzidas em diversas situações fisiopatológicas. O HOE 140, também
conhecido como Icatibant, é um potente antagonista competitivo e seletivo para o B2R, e,
apresenta uma alta afinidade pelos B2R na espécie humana, coelho, cobaia, rato e
camundongo. Esta substância apresenta uma ação prolongada e, devido à sua capacidade de
proteção contra degradação enzimática, impulsionou o conhecimento sobre as funções

11
cardiovasculares dos B2R (HOCK et al., 1991; GOBEIL et al., 1996). Em preparações in
vitro, o bloqueio de B2R é totalmente reversível, todavia, após sua remoção da solução, a
recuperação da resposta ao agonista é lenta (HOCK et al., 1991).
Experimentos realizados em humanos, com a infusão intravenosa de BK, em
voluntários saudáveis, resultou em diminuição da pressão arterial, aumento nos batimentos
cardíacos e diminuição na resistência periférica, entretanto, essas alterações hemodinâmicas
foram antagonizadas pelo uso de HOE-140. Estes estudos, envolvendo antagonistas em
humanos, sugerem um importante papel fisiológico das cininas na manutenção da pressão
arterial, uma vez que o SCC, juntamente com o sistema renina-angiotensina (SRA), estão
constitutivamente expressos no coração, no endotélio vascular, rins e diversos outros órgãos
que desempenham a regulação e manutenção da pressão arterial (BERTRAND, 2004).
As diferenças entre os mecanismos de ativação, assim como, na dessensibilização
e internalização dos receptores para cininas apontam a complexidade molecular das vias de
sinalização celular que envolve o SCC (LEEB-LUNDBERG et al., 2005).
2.4 Sinalização Das Espécies Reativas de Oxigênio e Nitrogênio em Cardiomiócito
As espécies reativas derivadas tanto, do oxigênio, quanto, do nitrogênio (RNS),
são produzidas em todos os sistemas biológicos e podem participar de complexas vias de
sinalização celular, em estados fisiológicos normais, participando de processos vitais para a
célula. Por outro lado, estas espécies, são potencialmente danosas e aparecem, também,
descritas como causa/consequência de diversas patologias e processo de envelhecimento
(SANTOS et al., 2011).
A sinalização promovida por essas espécies reativas compreende a transmissão e
amplificação de sinais, geralmente, envolvendo modificações pós-traducionais, de
redução/oxidação (redox) em proteínas. As ROS e RNS, em cardiomiócitos e outras células,
são geradas por diversas fontes, dentre as quais, as mais importantes são: as mitocôndrias, a
nicotinamida adenina dinucleotídeo fostato-hidrogênio oxidases da família Nox (NADPH
oxidase), xantina oxidase (XO) e as óxido nítrico sintases (NOS) (TZIOMALOS e HARE,
2009; DUNCAN et al., 2005). A produção dessas moléculas, por estas diversas fontes, pode
aumentar diversas vezes em diferentes contextos celulares (por exemplo, estado patológico)
ou por estimulação específica (por exemplo, NADPH oxidases).
As ROS e RNS são classificadas como oxidantes radicalares ̠ dentre os quais, o
radical ânion superóxido (O2•-) e o NO, e os oxidantes não radicalares ˗ peróxido de
hidrogênio (H2O2) e o peroxinitrito (ONOO-). Suas ações dependem do balanço entre a taxa

12
de produção e remoção das mesmas. Este equilíbrio será então dependente de fatores como a
concentração celular de antioxidantes enzimáticos, não enzimáticos e cofatores (AUGUSTO
et al, 2002). Entretanto, a reduzida capacidade antioxidante e elevada síntese destas
substâncias reativas são capazes de formar espécies oxidantes ainda mais reativas, tais como,
o radical hidroxila (OH•), e, posteriormente, caracterizar a condição de estresse oxidativo
(WINTERBOURN, 2008).
Os papéis fisiológicos e patológicos do NO, em cardiomiócitos, foram reunidos e
sugeridos em importantes revisões (SEDDON, SHAH e CASADEI, 2007; MASSION e
BALLIGAND, 2007; KASS et al., 2007). Vários autores revelaram que a redução da
biodisponibilidade de NO devido à sua reação com o O2•-, pode modular vias de sinalizações
intracelulares, devido ao aumento da geração do ONOO- (PACHER, BECKMAN e
LIAUDET, 2007).
Mesmo em baixas concentrações, moléculas, tais como, o OH• (concentração
subnanomolar) podem causar oxidações prejudiciais em componentes celulares, gerando
disfunção cardíaca mal adaptativa (CAVE et al., 2006). Sabe-se, também, que o estresse
oxidativo ocorre, muitas vezes, pela inadequada concentração de antioxidantes endógenos,
que contribui para o desenvolvimento da insuficiência cardíaca. Esta condição, pode ainda,
modular a ativação de cascatas de sinalização que desregulam a homeostase de proteínas
relacionadas ao controle do íon Ca2+ dentro da célula (CAVE et al., 2006).
As alterações na homeostase do Ca2+ intracelular, podem ocorrer devido
modificações promovidas por ROS e RNS em proteínas do miocárdio, tais como, oxidação de
tióis na cisteína, hidroxilação da prolina e arginina, e nitrosilação de tirosina. Essas
modificações podem afetar a conformação, estabilidade e atividade de receptores celulares,
transportadores iônicos (bombas, trocadores e canais), quinases, fosfatases, caspases,
translocases (GTPases), fatores de transcrição, proteínas contráteis e estruturais (AUGUSTO
et al, 2002; WINTERBOURN, 2008).
No coração, as atividades de proteínas que são alvo de modulação por
modificações redox são particularmente importantes para a manutenção fisiológica da função
cardíaca. Dentre estes alvos potenciais estão a proteína quinase A (PKA) (BRENNAN et al.,
2006), a proteína quinase G (PKG) (BURGOYNE et al., 2007), o receptor de rianodina
(RyR2) (XU et al., 1998), a pequena proteína G (Ras) (KUSTER et al., 2006) e a classe II das
histonas desacetilases (HDACs) (AGO et al., 2008).
Por outra via, o aumento exacerbado da geração de ROS mitocondrial na
hipertrofia cardíaca dilatada pode induzir a danos ao DNA mitocondrial (mtDNA), como por

13
exemplo, tornando-o mais susceptível ao dano oxidativo devido à falta de proteção das
histonas mitocondriais (IDE et al., 2001).
Neste mesmo sentido, evidências comprovam que o aumento substancial da
geração de ROS, desenvolvida em camundongos knockout para a superóxido dismutase
mitocondrial manganês (MnSOD), provoca o desenvolvimento de uma grave cardiomiopatia
dilatada (LI et al., 1995).
Em contraste, camundongos transgênicos apresentando expressão aumentada
(overexpression) da glutationa peroxidase (GPx) (SHIOMI et al., 2004) mostraram uma
atenuação significativa de efeitos adversos do remodelamento ventricular esquerdo após
infarto agudo do miocárdio. Da mesma forma, os ratos com exacerbada expressão da catalase
mitocondrial, curiosamente apresentaram tempo de vida prolongado, com função cardíaca
melhorada (SCHRINER et al., 2005), avaliada por ecocardiograma. Observou-se uma redução
da ativação de vias de transcrição nuclear, relacionadas ao desenvolvimento e progressão da
hipertrofia cardíaca, provocada, pela senescência (DAI et al., 2009).
A literatura claramente demonstra que a sinalização tanto das ROS, quanto das
RNS, é essencial para a manutenção da homeostase dos cardiomiócitos e está centralmente
envolvida nos mecanismos de sinalização celular que constitui a base integrada das respostas
contráteis cardíacas (SANTOS et al., 2011).
Santos et al. (2011) preveem que vários aspectos fundamentais devem ser
considerados nesta homeostase, incluindo: (1) a regulação por ROS e RNS da sinalização em
compartimentos específicos do cardiomiócito, (2) mecanismos que regulam e integram a
sinalização entre os compartimentos celulares, (3) como procede a integração da geração de
ROS com agentes antioxidantes e processos metabólicos (4) determinação das funções
prejudiciais e/ou de proteção através das modificações redox/nitroso, e (5) identificação dos
principais alvos moleculares susceptíveis às modificações redox/nitroso. Desta forma, uma
melhor compreensão deste complexo sistema poderá permitir o desenvolvimento de
estratégias terapêuticas mais específicas para as diversas doenças cardiovasculares.

14
3. OBJETIVOS
3.1 Objetivo Geral
O propósito deste estudo foi avaliar a contribuição dos receptores para cininas na
manutenção do status redox celular no sistema cardiovascular.
3.2 Objetivos Específicos
• Avaliar a participação dos receptores para cininas em parâmetros contráteis do
coração isolado de camundongos;
• Mensurar a contribuição dos receptores para cininas na produção de espécies reativas
de oxigênio (radical ânion superóxido, peróxido de hidrogênio) e da espécie reativa de
nitrogênio (óxido nítrico);
• Verificar a participação da sinalização para cininas na expressão de enzimas
antioxidantes (superóxido dismutase, catalase e glutationa peroxidase) e proteínas
relacionadas com a homeostase do cálcio intracelular (bomba cálcio ATPase do retículo
sarcoplasmático e trocador sódio/cálcio);
• Estudar as vias implicadas nas respostas vasorrelaxantes e vasoconstritoras associadas
aos receptores para cininas;

15
4. MATERIAIS E MÉTODOS
4.1 Grupos Experimentais
Foram utilizados para realização desses experimentos, camundongos da linhagem
C57BL6/J, machos, idades entre 12 a 16 semanas e pesando de 25 a 30 g. Para o
delineamento experimental os animais foram distribuídos nos seguintes grupos: grupo
controle (wild type - WT) e camundongos knockouts do receptor B1 para cininas (B1-/-), e B2
(B2-/-). Os animais foram mantidos em temperatura de 22 ± 2 ºC, em ciclo claro/escuro de 12
horas e acesso ad libidum à ração e água. Os camundongos transgênicos utilizados neste
estudo foram fornecidos pelos pesquisadores Dr. Jorge Luiz Pesquero (ICB/UFMG) e Dr.
João Bosco Pesquero (UNIFESP).
Todos os procedimentos experimentais foram conduzidos de acordo com o guia
de cuidados e usos de animais de laboratório e aprovado pelo Comitê de Ética de
Experimentação Animal da UFMG (protocolo # 078/05 e 236/09 – CETEA).
4.2 Genotipagem
A genotipagem dos camundongos knockout para os receptores de cininas foi
realizada pela técnica de reação em cadeia da polimerase (PCR). O DNA genômico foi
extraído de um fragmento de 0,5 cm da cauda dos animais e digerido em 80 µL do tampão de
extração contendo: 100 mM Tris-HCl, 200 mM NaCl, 2% SDS e pH ajustado para 8,5,
enriquecido com 8 µL de enzima proteinase K (10 mg/mL), por um período de 14 horas à 55
ºC sobre agitação de 550 rpm (Orbit M60, Labnet®®, USA). Após esse período, o
homogenato foi aquecido à 95ºC (ThermoBlock IT-2002, BIOPLUS®, USA) por 10 minutos e
então adicionado 700 µL de tampão Tris-HCl 10 mM contendo EDTA 1 mM, pH ajustado
para 8,0 e enriquecido com 5 µL de RNAse A (4 mg/mL). Em seguida as amostras foram
novamente incubadas à 37ºC por 25 minutos sob agitação de 550 rpm, posteriormente
centrifugadas a 10.000xg (Neofuge 15R, Heal Force®, China) por 10 minutos. O sobrenadante
foi separado por decantação e utilizado na PCR.
Para síntese do cDNA do B1R foram utilizados os oligonucleotídeos
TGAAGCTGTGAGCTCTTTG e GCTACCCAGATGAGCAGGCA e para o B2R
TGTCCTCAGCGTGTTCTTCC e GGTCCTGAACACCAACATGG. Os produtos da PCR
(550 pb para B1R e 361 pb para B2R) foram separados em gel de poliacrilamida 5%.
Paralelamente era feito a PCR para amplificar parte do gene da neomicina, presente nos
animais selvagens. Os oligonucleotídeos para a neomicina foram os seguintes:

16
CTTGGGTGGAGAGGCTATTC e AGGTGAGATGACAGGAGATC e o produto um
amplicon de 280 pb. Após a corrida, os géis foram fixados em solução contendo: etanol 10%
e ácido acético 0,5% por 3 minutos, sob suave agitação. Em seguida, o gel foi corado em
solução de nitrato de prata 10% por 7 minutos e lavado com água destilada por 2 vezes. O gel
foi colocado em solução reveladora (3% NaOH e 0,3% formaldeído) até as bandas
aparecerem. Após o descarte da solução reveladora, o gel foi novamente transferido para a
solução fixadora para interromper a reação. Os géis foram fotodocumentados para
interpretação dos resultados.
4.3 Procedimento Cirúrgico para Retirada do Coração
Os animais receberam injeção intraperitonial de 200 U.I. de heparina diluída em
200 µL de solução salina. Após 10 minutos, os camundongos foram sacrificados por
decapitação, e em seguida, procedeu-se a toracotomia. Neste procedimento, as cartilagens
costocondrais foram abertas em ambos os lados e o esterno rebatido para cima. Uma vez
exposta a cavidade torácica, seccionava-se sucessivamente a veia cava inferior, a veia cava
superior, as artérias aorta e pulmonar, a traqueia, o esôfago bem como parte dos pulmões,
retirando-se assim o coração em bloco. Depois de retirado, o coração foi colocado em um
béquer contendo solução de Krebs-Ringer contendo (mM): 118,4 NaCl; 4,7 KCl; 26,5
NaHCO3; 1,8 CaCl2.2H2O; 11,7 glicose; 1,2 KH2PO4; 1,2 MgSO4.7H2O ajustada para pH 7,4,
oxigenada com mistura carbogênica (95% de O2 e 5% de CO2; White Martins®, Brasil) e em
temperatura de aproximadamente 4ºC. Depois de transferido para uma placa de petri, os
excedentes de tecido pulmonar e vascular, da traqueia e do esôfago que acompanhavam o
coração, foram removidos.
Em seguida, a aorta ascendente foi seccionada com uma tesoura de microcirurgia
na altura de sua primeira ramificação (tronco braquiocefálico) e fixada com fio de sutura em
uma agulha de aço inoxidável modificada, acoplada a uma seringa de 1 mL contendo solução
de Krebs-Ringer (LAUTON-SANTOS, 2007; OLIVEIRA et al., 2007).
Após a fixação da aorta na agulha, este conjunto foi removido da seringa e fixado
em uma coluna de vidro já contendo a solução de Krebs-Ringer. O tempo ótimo para
realização deste procedimento era sempre igual ou inferior a 5 minutos. Caso o tempo de
preparação excedesse este tempo o coração era descartado (LAUTON-SANTOS, 2007).

17
4.4 Estudo dos Parâmetros Cardíacos
O sistema de Langendorff com fluxo constante de perfusão foi utilizado para o
estudo de parâmetros fisiológicos cardíacos do órgão isolado.
Os registros das medidas da força de contração e pressão de perfusão eram
obtidos por transdutores acoplados a uma placa conversora (A/D), conectada ao computador.
Para a transformação dos registros analógicos obtidos, em sinais digitais, e a aquisição dos
mesmos, utilizou-se o equipamento Biopac Systems, modelo M100A-CE e o software
Acqknowledge 3.5.7.22.
Para a perfusão do coração utilizou-se solução de Krebs-Ringer, oxigenada com
mistura carbogênica (95% de O2 e 5% de CO2; White Martins®, Brasil) e aquecida a 37°C por
uma jaqueta de água acoplada a um reservatório com um termostato e uma bomba peristáltica
(Watson Marlon M-501U) para manter um sistema de circulação. Manteve-se, nos
experimentos, um fluxo constante entre 2,3 e 2,6 mL por minuto.
A pressão de perfusão era captada por um transdutor de pressão (TP-200, Nihon
Khoden) previamente calibrado utilizando-se uma coluna de mercúrio, ligado através de um
fino tubo de polietileno à extremidade inferior da coluna de vidro com a solução de Krebs-
Ringer.
Para averiguação das tensões sistólica e diastólica, foi colocado no ápice do
coração, um anzol ligado a um fio que passava por um sistema de roldana e era fixado a um
transdutor de força (TSD 125C, Biopac Systems, USA), previamente calibrado por um peso
padrão de 10 gramas. Impôs-se uma tensão diastólica de 0,5 grama durante o período de
estabilização (primeiros 20 minutos). Após este período as medidas foram coletadas por mais
20 minutos sem qualquer ajuste na tensão (LAUTON-SANTOS, 2007). A frequência cardíaca
foi obtida a partir dos registros de tensão sistólica (frequência espontânea cardíaca), expressa
em batimentos por minuto (BPM). Finalizado o experimento, o coração foi rapidamente
removido do sistema e seu peso úmido foi mensurado em balança analítica (Mettler Toledo®,
Mod. XS 205) para posterior normalização de seu respectivo registro de tensão sistólica e
diastólica.
A figura 5 representa o perfil típico do registro mecânico da contração de corações
isolados de camundongos durante perfusão retrógrada em sistema de Langendorff. As análises
dos dados foram feitas usando-se os software Acqknowledge 3.5.7 e Prism 5.1. (GraphPad,
San Diego, CA, USA).

18
Figura 5. Registro típico de coração isolado de camundongo durante perfusão
retrógrada em sistema de Langendorff.
4.5 Obtenção de Células Cardíacas Isoladas
O coração foi obtido conforme o protocolo já mencionado. Para a obtenção dos
cardiomiócitos ventriculares isolados, foi utilizado o procedimento padrão para a dissociação
enzimática do tecido, como descrito por Mitra e Morad (1985). Basicamente, uma solução
extracelular contendo baixa concentração de Ca2+ e adicionada de 1 mg/mL de colagenase
tipo II (Worthington, USA), foi perfundida retrogradamente com o uso de um sistema de
Langerdorff de pressão constante por 5 à 10 minutos. Após este período, o tecido foi cortado
em pequenos fragmentos e prosseguiu-se com uma delicada agitação mecânica para facilitar o
isolamento dos cardiomiócitos. O Ca2+ extracelular foi restaurado gradativamente para 1,8
mM. As células ventriculares obtidas ficaram armazenadas em solução Tyrode contendo
(mM): 140 NaCl; 5 KCl; 0,5 MgCl2.7H2O; 1,8 CaCl2; 5 Glicose; 5 HEPES, pH 7,4.
4.6 Experimentos de Reatividade Vascular
4.6.1 Obtenção e Montagem dos Anéis de Aorta
O procedimento para montagem dos anéis de aorta foi realizado de acordo com
Capettini et al. (2011). Os animais foram sacrificados por decapitação. Após esse
procedimento, a aorta torácica foi cuidadosamente removida, retirando-se o excesso de tecido

19
adiposo e conjuntivo. Transferiu-se o material biológico para uma placa de petri com solução
nutridora de Krebs-Henseileit contendo (mM): 118 NaCl; 4,7 KCl; 25 NaHCO3; 2,5
CaCl2.2H2O; 11 glicose; 1,2 KH2PO4; 1,2 MgSO4.7H2O.
A artéria foi cortada em anéis de aproximadamente 2-3 mm e duas hastes
metálicas triangulares foram cuidadosamente passadas através do lúmen do vaso. Um dos
triângulos foi fixado à parede da cuba, e o outro foi conectado verticalmente a um transdutor
de tensão isométrica. Para obtenção do registro, esses anéis foram acondicionados em cubas
para órgão isolado com volume de 10 mL, contendo solução de Krebs-Henseileit, aerada
continuamente com mistura carbogênica (95% de O2 e 5% de CO2; White Martins®, Brasil),
mantida à temperatura de 37±1ºC e pH de 7,4. Após a montagem, os anéis aórticos foram
submetidos a uma tensão de repouso de 0,5 gramas, continuamente regulada na primeira hora
do experimento durante a estabilização da preparação. Neste período, a solução nutridora foi
trocada a cada 15 minutos, objetivando-se, dessa forma, evitar o acúmulo de metabólitos,
como também a restauração dos níveis de glicose necessários ao metabolismo vascular.
Os registros experimentais foram obtidos a partir de transdutores de tensão
isométrica (World Precision Instruments, Inc., USA), conectados a um amplificador (Modelo
TBM-4; World Precision Instruments, Inc., USA), sendo este, acoplado a um computador
equipado com uma placa conversora analógico-digital (AD16JR; World Precision
Instruments, Inc. USA). O programa WinDaq Data Acquisition (Dataq® Instruments, USA)
foi utilizado para aquisição dos dados, e, análise dos registros experimentais foi realizada no
Prism 5.1. (GraphPad, San Diego, CA, USA).
4.6.2 Avaliação da Integridade Endotelial
Após o período de estabilização, os anéis foram contraídos com o agonista α1-
adrenérgico, fenilefrina (Phen; 3X10-7 M; Sigma®, USA), e, observando-se um platô no
registro de contração, a presença de endotélio funcional vascular foi verificada através da
administração do agonista muscarínico, acetilcolina (ACh; 10-5 M; Sigma®, USA). Após
atingir o relaxamento máximo, a solução nutridora de Krebs-Henseileit foi renovada (por três
vezes). O procedimento anteriormente descrito foi repetido após 30 minutos. A integridade
funcional do endotélio foi considerada apenas nos anéis onde a ACh induziu > 70% de
relaxamento após contração com Phen, e, os anéis que não apresentaram este comportamento,
foram utilizados somente para construção de curvas de contração.

20
4.6.3 Remoção do Endotélio Funcional
Quando necessário, o endotélio foi removido mecanicamente com o auxílio de
uma haste metálica friccionada de forma delicada ao lúmen vascular. A ausência de endotélio
funcional foi confirmada pela ineficácia da ACh (10-5 M) em relaxar os vasos pré-contraídos
com Phen (3X10-7 M).
4.6.4 Curva de Contração Induzida pela Fenilefrina
Para realização desses experimentos foram utilizados anéis provenientes da aorta
torácica obtidos, como anteriormente descrito. Após o período de estabilização e teste
funcional do endotélio vascular, foram construídas curvas de contração concentração-resposta
cumulativas em resposta à Phen (10-9 - 10-4 M).
4.6.5 Participação da Via das Óxido Nítrico Sintases na Resposta Vasorrelaxante
à Acetilcolina
Após o período de estabilização e teste funcional do endotélio vascular, em anéis
aórticos pré-contraídos com concentração submáxima de Phen (10-7 M), foram construídas
curvas concentração-resposta à ACh (10-9 - 10-4 M), com a finalidade de se estudar a
participação das NOS na resposta vasorrelaxante induzida pela ACh.
Após lavagens de 30 minutos com solução nutridora, estes anéis de aorta foram
pré-incubados por 20 minutos com inibidores das NOS, NG-Nitro-L-Arginine (L-NNA, 10-6
M) ou l-(2 Trifluoromethylphenyl)imidazole (TRIM, 3x10-4 M). O L-NNA na concentração
utilizada é inibidor não-seletivo das NOS. O TRIM é inibidor seletivo da nNOS nesta
concentração. Em seguida, na presença dos inibidores, curvas concentração-resposta
cumulativas a ACh foram novamente construídas nos vasos pré-contraídos com Phen. Os
resultados obtidos foram comparados com as respostas vasorrelaxantes na ausência das drogas
inibidoras acima citadas.
4.7 Mensuração dos Níveis Intracelulares das Espécies Reativas de Oxigênio
Para a detecção dos níveis intracelulares do O2•- e do H2O2, os cardiomiócitos
ventriculares isolados e anéis de aorta torácica provenientes, ambos, dos animais em estudo,
foram obtidos como previamente descrito.
As células isoladas e os anéis de aorta foram carregados com 10 µM da sonda
fluorescente sensível ao O2•-, dihidroetídio (DHE), Calbiochem®, por 30 minutos à 37ºC. Para
a detecção do H2O2 foi utilizado 0,1 µM da sonda fluorescente sensível ao mesmo, 2,7-

21
diclorodihidrofluoresceína-diacetato (H2DCFH-DA), Invitrogen™|Molecular Probes®, por 15
minutos à 37ºC. Esta última é desacetilada intracelularmente por esterases não específicas, e,
pode ser oxidada pelo H2O2 intracelular, formando o composto 2,7-diclorofluoresceína (DCF)
cuja emissão de fluorescência pode ser correlacionada aos níveis de H2O2 intracelulares.
As amostras foram incubadas com as sondas fluorescentes em um solução de
Tyrode e então lavadas no mesmo tampão para remover o excesso das mesmas. Acrescenta-se
que a sonda DCF não é específica para o H2O2 detectando também a presença do OH-.
Para medir, nas células cardíacas a fluorescência das ROS, os miócitos foram
excitados no comprimento de onda de 488 nm e a luz emitida com comprimento de onda de
515 nm foi detectada com auxílio do microscópio confocal (Zeiss 510 Meta, GmbH, Jena,
Germany) (Cemel/ICB-UFMG) com a objetiva de imersão em óleo (63×).
Para a execução destes experimentos na aorta torácica, anéis com
aproximadamente 2 mm de dimensão, previamente carregados com as sondas, acima citadas,
foram incluídos em meio crioprotetor, Tissue-Tek O.C.T. (SAKURA Finetechnical®, Japan) e
mantidas à -20ºC. Posteriormente foram realizados cortes em criostato (CM 1850, Leica®)
com espessura de 10 µm. Os cortes dos anéis foram transferidos para lâminas histológicas, e,
para aquisição das imagens fluorescentes, foi utilizado microscópio de fluorescência (IX2-
ICB, Olympus®, USA) acoplado a câmera digital (XM-10, Olympus®, USA) com excitação e
emissão em comprimentos de onda semelhante ao utilizado para detecção em células
cardíacas.
O software do ImageJ 1.38x (NIH) foi utilizado para o processamento das
imagens capturadas. A análise das imagens foi realizada no Prism 5.1. (GraphPad, San Diego,
CA, USA). As imagens de microscopias confocal e fluorescência foram analisadas de acordo
com a intensidade de fluorescência por área. Os resultados obtidos foram expressos em
unidades arbitrárias (u.a.).
4.8 Mensuração dos Níveis Intracelulares do Óxido Nítrico
Para detecção dos níveis intracelulares de NO, os cardiomiócitos isolados e anéis
de aorta torácica foram processados como acima descrito, porém, carregados com 10 µM da
sonda fluorescente sensível ao NO, 4-amino-5-metilamino-2’,7’-difluorofluoresceína
diacetato (DAF-FM), Invitrogen™|Molecular Probes®, por 30 minutos à 37ºC. As imagens
foram capturadas e as análises processadas como descrito anteriormente.

22
4.9 Extração de Proteínas do Tecido
Após o sacrifício dos animais por decapitação, os ventrículos foram rapidamente
removidos e imersos em uma solução salina tamponada com Tris-Base (TBS) contendo
(mM): 50 Tris-Base; 150 NaCl, pH 7,4 e em seguida removidos os excesso de sangue, tecido
adiposo e conjuntivo. Posteriormente, em banho de gelo, os tecidos foram homogeneizados
(Dremel 300®) na presença de tampão de lise contendo (mM): 150 NaCl; 50 Tris 50; 5
EDTA.2Na; 1 MgCl2, pH 8,0 acrescido de 100 mM Dithiothreitol, 1% Nonidet P40, 0,3% de
Triton X-100, 0,5% de SDS, cocktail de inibidores de proteases (SigmaFast®, Sigma), 100
mM PMSF e de inibidores de fosfatases, 10 mM Na4P2O7, 10 mM Na3VO4 e 100 mM NaF.
Os tecidos foram homogeneizados a uma proporção de 100 mg/mL de tampão de
lise. Após o processamento, as amostras foram centrifugadas a 15.000 rpm (Neofuge 15R,
HEAL FORCE®, China) por 15 minutos à 4ºC. O sobrenadante foi aliquotado e congelado a
-80ºC para posterior utilização.
4.10 Determinação da Concentração Total de Proteínas
A concentração total de proteínas foi determinada pelo método de Lowry et al.
(1951). Resumidamente, as amostras foram diluídas 10x e adicionado NaOH 0,5 M. Após 15
de incubação, adicionou-se em cada amostra uma solução composta por 2% Na2CO3, 1%
CuSO4 e 2% KNaC4H4O6·4H2O, na proporção de 100:1:1. Ao final, foi adicionado o reagente
de folin diluído 2x. Após incubação por 30 minutos, foi realizada a leitura no comprimento de
onda de 630 nm em espectrofotômetro de placa (ELx800, BIOTEK Instruments®, USA). Para
confecção da curva de proteína padrão, utilizou-se albumina bovina (Sigma-Aldrich®) em
várias concentrações.
4.11 Determinação da Expressão Proteica por Western Blot
As amostras foram diluídas em tampão da amostra (Tris HCl/SDS pH=6,8, 3%
Glycerol, 1% SDS, 0,6% β-mercaptoetanol e 0,1% Azul de Bromofenol) e aquecidas à 98°C
por 5 minutos. Para separação, foram aplicados 60 µg de proteínas em gel de SDS-PAGE (do
inglês sodium dodecyl sulfate polyacrylamide gel electrophoresis) à 7,5% ou 10%.
Após serem separadas no gel de poliacrilamida, as proteínas foram transferidas
para uma membrana de nitrocelulose (Millipore®, USA) com poros de 0,44 µm. A qualidade
da transferência foi monitorada através da coloração da membrana com solução de Ponceau
0,3%. A membrana foi então lavada em solução salina tamponada com Tris-Base acrescido
com 0,05% de Tween 20% (TBS-T) e colocada por 1 à 2 horas em solução de bloqueio (4%

23
de albumina em TBS-T). Após o bloqueio, a membrana foi incubada em temperatura de 6-
8ºC, com o anticorpo primário específico diluído em 1% de albumina em TBS-T, overnight.
Os seguintes anticorpos primários foram utilizados: anti-nNOS / anti-eNOS / anti-NCX / anti-
GPx (1:1000; policlonal feito em coelho; Santa Cruz Biotechnology Inc - Santa Cruz, CA,
USA) e anti-pnNOS Ser852 / anti-peNOS Ser1177 / anti-peNOS Thr495 / anti-SOD-1 / anti-Catalase
/ anti-SERCA2a / anti-GAPDH (1:1000; policlonal feito em cabra; Santa Cruz Biotechnology
Inc - Santa Cruz, CA, USA).
Em seguida, a membrana foi lavada três vezes com TBS-T durante 5 minutos e
incubada por 2 horas com o anticorpo secundário conjugado à peroxidase (HRP) (1:5000,
anti-goat IgG-HRP ou anti-rabbit IgG-HRP, Sigma, St. Louis, MO) diluído em 1% de
albumina em TBS-T. Após o período de incubação, a membrana foi novamente lavada por
mais três vezes em TBS-T durante 5 minutos. As bandas proteicas foram detectadas por uma
reação de quimioluminescência (Luminata strong™ - Western HRP substrate, Merck-
Millipore, Darmstad, Germany) e a intensidade das mesmas foi avaliada por análise
densitométrica através do software ImageJ 1.38x (NIH).
Para os experimentos acima citados, foram utilizados o sistema Mini Protean III-
Tetracell e Mini Trans-Blot Electrophoretic Transfer Cell (BIORAD®, CA, USA).
4.12 Co-imunoprecipitação
Para se avaliar a interação entre os receptores B1, B2 e nNOS, foram realizados
experimentos de co-imunoprecipitação. Para este ensaio, 100 µg dos lisados proteicos
provenientes do pool de aortas torácicas de 3 camundongos do grupo WT foram incubados
com 1 µg do anticorpo anti-nNOS ou anti-eNOS por 2 horas à 4 °C, após este período, o
complexo amostra-anticorpo foi conjugada com beads de proteína A/G-sepharose, overnight à
4 °C.
Posteriormente, as amostras foram lavadas com tampão de imunoprecipitação
contendo (mM): 25 Tris-Base, 150 NaCl, pH 7,2 e centrifugadas por 3 minutos à 8000 rpm
(Neofuge 15R, HEAL FORCE®, China). O precipitado foi ressupendido em tampão de
eluição (200mM Glicina, pH 3,0) para desligamento do complexo imunoprecipitado das
beads.
Em seguida, foi adicionado o tampão de neutralização (1 M Tris-Base, pH 9,0) e
centrifugado por 1 minuto à 6.000 rpm. O sobrenadante foi aliquotado e submetido ao
protocolo de western blot, como previamente descrito. Para detecção do complexo de
interação proteína-proteína, foram executados experimentos com anticorpos primários que

24
reconhecem os receptores B1 e B2 para cininas, anti-B1 e anti-B2 (ambos, 1:1000; policlonal
feito em coelho; Santa Cruz Biotechnology Inc - Santa Cruz, CA, USA).
4.13 Análises Estatísticas
Para tratamento estatístico, as análises dos resultados obtidos foram processadas
em software Prism 5.1 (GraphPad, San Diego, CA, USA).
Os dados de coração isolado, tensão sistólica e diastólica, foram representados em
g/g ± e.p.m., a frequência cardíaca em BPM ± e.p.m. e a pressão de perfusão em mmHg ±
e.p.m., sendo adotado a análise de variância ANOVA two-way para medidas paramétricas,
seguidas do pós-teste de BONFERRONI para comparação entres os grupos.
Os resultados de contração e relaxamento vascular foram representados como o
percentual de reposta aos agonistas, Phen e ACh. As curvas concentração-resposta foram
analisadas ponto-a-ponto, através da análise de variância ANOVA two-way para medidas
paramétricas, seguidas do pós-teste de BONFERRONI, para comparação entres os grupos. O
efeito final máximo (Emax) foi a concentração da droga que produziu a resposta máxima de
contração ou relaxamento.
Os resultados de expressão proteica obtidos através do western blot foram
normalizados pelo resultado da proteína normalizadora (GAPDH) de cada amostra ou eNOS e
nNOS, em alguns casos, sendo expressos em u.a., média ± e.p.m., e, adotada a análise de
variância ANOVA one-way, seguidas do pós-teste de BONFERRONI, para comparação entres
os grupos.
As imagens de microscopias confocal e fluorescência foram analisadas de acordo
com a intensidade de fluorescência por área, sendo expressas em u.a., média ± e.p.m., e,
adotada a análise de variância ANOVA one-way, seguidas do pós-teste de BONFERRONI
para comparação entres os grupos.
Para efeito estatístico, foram considerados significativos os valores que
apresentavam p<0,05.

25
5. RESULTADOS
Inicialmente, amostras do DNA dos animais knockouts para os receptores de
cininas foram coletadas, a fim de confirmarmos a deleção genética completa dos B1R e B2R.
Os resultados apresentados na figura 6 confirmaram que os animais do grupo B1-/- não
apresentam a expressão do B1R, mas expressam o B2R. Já os resultados ilustrados na figura 7,
confirmaram que os animais do grupo B2-/- não apresentavam a expressão do B2R, mas
expressavam o B1R.
Figura 6.Genotipagem de camundongos knockouts para o receptor B1 para cininas (B1-/-).
Figura 7.Genotipagem de camundongos knockouts para o receptor B2 para cininas (B2-/-).
Após a confirmação da ausência dos receptores para cininas nos animais
knockouts, propusemos avaliar os mecanismos contráteis e fisiológicos do coração isolado,
através da técnica da perfusão retrógada em sistema de Langendorff. Na figura 8 estão

26
apresentados os resultados dos registros da variação da força desenvolvida pelo coração dos
camundongos durante a sístole, denominada como tensão sistólica.
0 5 10 15 20
1
2
3
4
B2-/-
B1-/-
WT
******
Tempo (min)
Ten
são
Sis
tólic
a (g
/g)
Figura 8. Variação da tensão sistólica em coração isolado de camundongos. Os registros foram obtidos em corações de animais dos grupos selvagem (WT – Wild Type - n=5), e knockouts para os receptores para cininas B1 (B1
-/-- n=6) e B2 (B2-/-- n=5). Os resultados
estão representados em média ± e.p.m. (***p<0,001, entre WT x B1-/- / WT x B2
-/- ).
As medidas de tensão sistólica revelaram que os animais knockouts para ambos os
receptores de cininas apresentam um comprometimento dos mecanismos envolvidos no
desenvolvimento da força máxima contrátil. Quando comparada a média dos registros obtidos
dos corações dos animais WT (3,04 ± 0,07 g/g), aos valores apresentados pelos animais
deficientes em receptores para cininas B1-/- (1,43 ± 0,08 g/g) e B2
-/- (1,47 ± 0,03 g/g), foi
detectado uma redução significativa (p<0,001) da tensão sistólica cardíaca durante o período
de 20 minutos. Esta diminuição, em relação aos animais selvagens, representou uma redução
de 53% em animais B1-/- e 52% em B2
-/-.
Na figura 9 são apresentados os resultados dos registros da variação da força,
desenvolvida pelo coração isolado, durante a diástole, denominado como tensão diastólica
(WT: 0,41 ± 0,01 g/g; B1-/-: 0,37 ± 0,01 g/g e B2
-/-: 0,44 ± 0,01 g/g). Já, na figura 10, são
apresentados os valores médios da frequência cardíaca espontânea do coração durante o
período de 20 minutos (WT: 326 ± 2 BPM; B1-/-: 321 ± 3 BPM e B2
-/-: 318 ± 1 BPM). Os

27
resultados encontrados em ambas as medidas, tensão diastólica e frequência cardíaca, não
diferem, de forma significativa (p>0,05) entre os grupos avaliados.
0 5 10 15 20
0.3
0.4
0.5
0.6
B2-/-
WTB1
-/-
Tempo (min)
Ten
são
Dia
stól
ica
(g/g
)
Figura 9. Variação da tensão diastólica em coração isolado de camundongos. Os registros foram obtidos em corações de animais dos grupos selvagem (WT – Wild Type - n=5), e knockouts para os receptores para cininas B1 (B1
-/- - n=6) e B2 (B2-/-- n=5). Os
resultados estão representados em média ± e.p.m.
0 5 10 15 20
250
300
350
400B2
-/-
WTB1
-/-
Tempo (min)
Fre
quên
cia
Car
díac
a (B
PM
)
Figura 10. Variação da frequência cardíaca espontânea em coração isolado de animais dos grupos selvagem (WT – Wild Type - n=4) e knockouts para os receptores para cininas B1 (B1
-/- - n=5) e B2 (B2-/- - n=4). Os resultados estão representados em média ± e.p.m.

28
Outro parâmetro fisiológico analisado foi a resistência coronariana, avaliada
através da medida da pressão de perfusão. A figura 11 mostra os valores médios dos registros
da pressão de perfusão do coração, obtidos em sistema de Langendorff durante 20 minutos.
Os resultados encontrados mostraram uma elevada pressão de perfusão nos animais dos
grupos B1-/-(173 ± 2,4 mmHg) e B2
-/- (173 ± 0,6 mmHg), que apresentaram alteração
significativa (p<0,001) quando comparados com os animais do grupo WT (125 ± 1,6 mmHg),
representando, desta forma, um aumento de 38% em ambos os grupos.
0 5 10 15 20
100
120
140
160
180
200 B2-/-
B1-/-
WT
******
Tempo (min)
Pre
ssão
de
Per
fusã
o (m
mH
g)
Figura 11. Variação da pressão de perfusão em coração isolado de animais dos grupos selvagem (WT – Wild Type - n=8) e knockouts para os receptores para cininas B1 (B1
-/- - n=6) e B2 (B2
-/- - n=5). Os resultados estão representados em média ± e.p.m. (***p<0,001, entre WT x B1
-/- / WT x B2-/-).
Com base no fato de que a função contrátil cardíaca é controlada tanto por ROS
quanto por RNS, e, que, a sinalização das cininas, em pontos específicos, pode influenciar a
produção de tais espécies (SANTOS et al, 2011), sugerimos que a deleção genética dos
receptores para cininas possa criar perturbações no metabolismo oxidativo e,
consequentemente, alterar a geração das ROS e RNS nas células cardíacas. Portanto, para este

29
tipo de análise, foram executados experimentos com cardiomiócitos isolados dos ventrículos
dos animais em estudo. Estas células foram carregadas com sondas sensíveis às ROS (DHE e
DCFH) e às RNS (DAF). Os resultados apresentados na figura 12 mostraram que os animais
com deleção dos receptores para cininas geram, de forma significativa (p<0,001), uma maior
quantidade de O2•-, B1
-/- (483 ± 13 u.a.; 40% maior) e B2-/- (484 ± 9 u.a.; 41% maior), quando
comparados aos animais do grupo controle (343 ± 9 u.a.).
WT B1 -/- B2 -/-
0
200
400
600
n=95 n=65 n=85
*** ***
DH
E F
luor
escê
ncia
(u.a
.)
Figura 12. Produção basal do ânion superóxido em cardiomiócitos dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2
-/-). No painel superior estão apresentadas as figuras representativas (A) WT, (B) B1-/-,
(C) B2-/-. Barra de escala: 10µm. Os resultados estão representados em média ± e.p.m.
(***p<0,001, entre WT x B1-/- e WT x B2
-/-).
Os dados obtidos e mostrados na figura 13, reforçam os resultados das medidas de
O2•- apresentados na figura 12. Assim, demonstramos que os animais com deleção dos
receptores para cininas geram, de forma significativa (p<0,001), uma maior quantidade de
H2O2 (B1-/-: 482 ± 19 u.a.; 75% maior e B2
-/-: 426 ± 13 u.a.; 54% maior), quando comparados
aos animais WT (275 ± 11 u.a.).

30
WT B1 -/- B2
-/-
0
200
400
600
n=130 n=50 n=60
******
DC
F F
luor
escê
ncia
(u.a
.)
Figura 13. Produção basal de peróxido de hidrogênio em cardiomiócitos dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2
-/-). No painel superior estão apresentadas as figuras representativas (A) WT, (B) B1-/-,
(C) B2-/-. Barra de escala: 10µm. Os resultados estão representados em média ± e.p.m.
(***p<0,001, entre WT x B1-/- e WT x B2
-/-).
Uma vez mensuradas as ROS, o passo seguinte foi avaliar a produção de RNS. Na
figura 14 estão apresentados os resultados das medidas da produção intracelular de NO em
cardiomiócitos isolados. Foi detectado aumento significativo (p<0,001) na produção de NO
nos animais do grupo B1-/- (568 ± 14 u.a.; 75% maior) e B2
-/- (441 ± 12 u.a.; 36% maior),
quando comparados aos animais WT (323 ± 8 u.a.). Foi ainda detectado aumento significativo
(p<0,001) na produção de NO nos animais do grupo B1-/- quando comparados com o grupo
B2-/-.
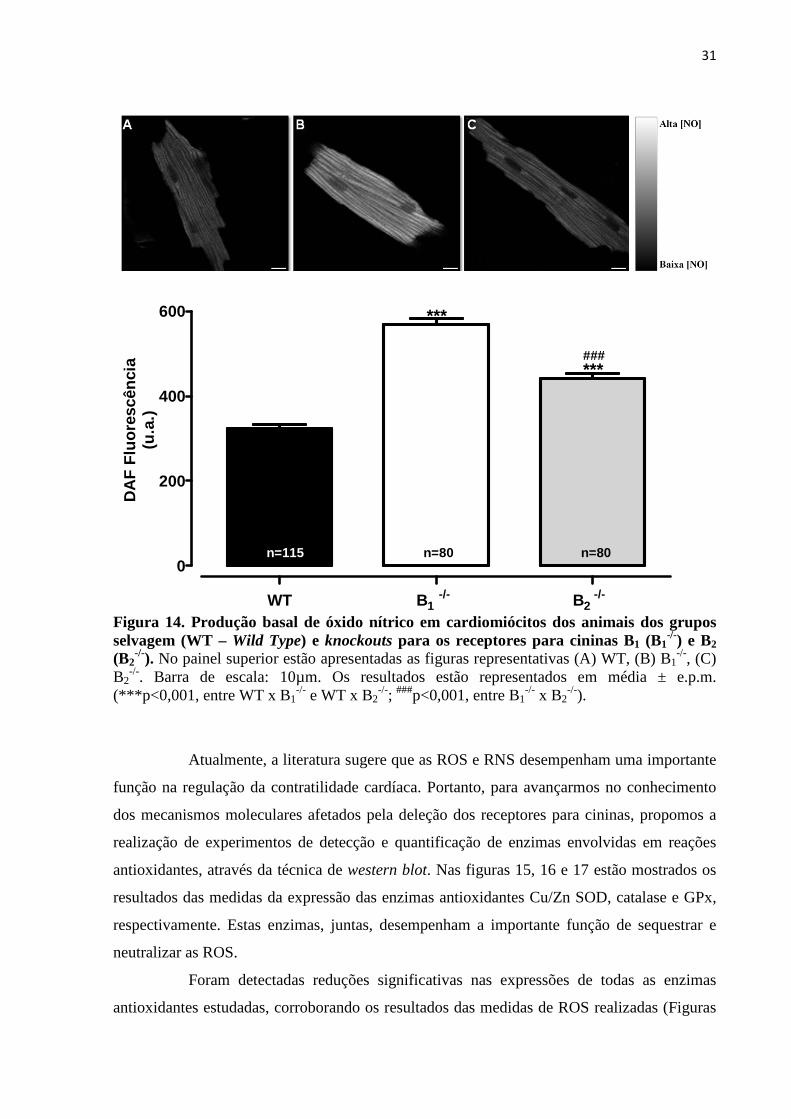
31
WT B1 -/- B2 -/-
0
200
400
600
n=115 n=80 n=80
***
***###
DA
F F
luor
escê
ncia
(u.a
.)
Figura 14. Produção basal de óxido nítrico em cardiomiócitos dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2
-/-). No painel superior estão apresentadas as figuras representativas (A) WT, (B) B1-/-, (C)
B2-/-. Barra de escala: 10µm. Os resultados estão representados em média ± e.p.m.
(***p<0,001, entre WT x B1-/- e WT x B2
-/-; ###p<0,001, entre B1-/- x B2
-/-).
Atualmente, a literatura sugere que as ROS e RNS desempenham uma importante
função na regulação da contratilidade cardíaca. Portanto, para avançarmos no conhecimento
dos mecanismos moleculares afetados pela deleção dos receptores para cininas, propomos a
realização de experimentos de detecção e quantificação de enzimas envolvidas em reações
antioxidantes, através da técnica de western blot. Nas figuras 15, 16 e 17 estão mostrados os
resultados das medidas da expressão das enzimas antioxidantes Cu/Zn SOD, catalase e GPx,
respectivamente. Estas enzimas, juntas, desempenham a importante função de sequestrar e
neutralizar as ROS.
Foram detectadas reduções significativas nas expressões de todas as enzimas
antioxidantes estudadas, corroborando os resultados das medidas de ROS realizadas (Figuras

32
12 e 13). Na mensuração da expressão da enzima Cu/Zn SOD (Figura 15) detectou-se uma
significativa redução de 32% e 20%, nos corações dos animais dos grupos B1-/- e B2
-/-,
respectivamente, quando comparados aos animais WT.
Figura 15. Determinação da expressão proteica da enzima Cu/Zn Superóxido Dismutase (Cu/Zn SOD) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). Os resultados estão
representados em média ± e.p.m. (**p<0,01 entre WT x B1-/- e *p<0,05 entre WT x B2
-/-).
Na determinação da expressão da catalase (Figura 16) também foi detectado uma
significativa redução de cerca de 34% e 39% nos corações dos animais dos grupos B1-/- e B2
-/-,
respectivamente, quando comparados aos animais WT.

33
Figura 16. Determinação da expressão proteica da enzima catalase em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). Os resultados estão representados em média ± e.p.m. (*p<0,05
entre WT x B1-/- e WT x B2
-/-).
Somando-se aos resultados anteriores, a expressão da GPx (Figura 17) apresentou
uma redução significativa, de cerca de 29% e 15% nos corações dos animais dos grupos B1-/- e
B2-/-, respectivamente, quando comparados aos WT.

34
Figura 17. Determinação da expressão proteica da enzima Glutationa Peroxidase (GPx) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). Os resultados estão representados em média
± e.p.m. (**p<0,01 entre WT x B1-/- e *p<0,05 entre WT x B2
-/-).
Para tentarmos justificar a origem do aumento na produção do NO, avaliamos a
expressão total e a atividade das enzimas NOS constitutivas, isoformas eNOS (Figuras 18, 19
e 20) e nNOS (Figuras 21 e 22). Estas enzimas são reconhecidamente expressas no tecido
cardíaco, e a atividade de ambas promovem modulações em vias de sinalização intracelular
que regulam, dentre outros processos, a contração cardíaca (KUSTER et al., 2006).
Na figura 18 está representada a medida da expressão total da eNOS, na qual foi
detectada um aumento significativo (p<0,05), cerca de 20% e 23% nos corações dos animais
dos grupos B1-/- e B2
-/-, respectivamente, quando comparados aos animais WT.
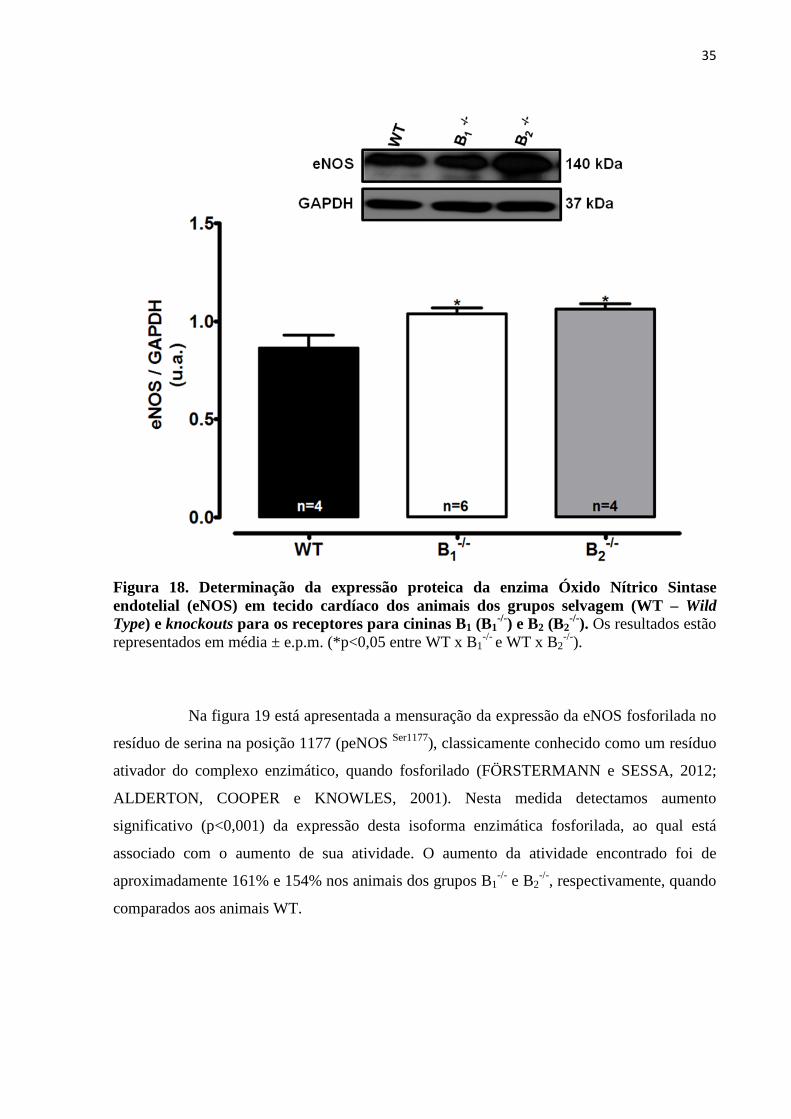
35
Figura 18. Determinação da expressão proteica da enzima Óxido Nítrico Sintase endotelial (eNOS) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). Os resultados estão
representados em média ± e.p.m. (*p<0,05 entre WT x B1-/- e WT x B2
-/-).
Na figura 19 está apresentada a mensuração da expressão da eNOS fosforilada no
resíduo de serina na posição 1177 (peNOS Ser1177), classicamente conhecido como um resíduo
ativador do complexo enzimático, quando fosforilado (FÖRSTERMANN e SESSA, 2012;
ALDERTON, COOPER e KNOWLES, 2001). Nesta medida detectamos aumento
significativo (p<0,001) da expressão desta isoforma enzimática fosforilada, ao qual está
associado com o aumento de sua atividade. O aumento da atividade encontrado foi de
aproximadamente 161% e 154% nos animais dos grupos B1-/- e B2
-/-, respectivamente, quando
comparados aos animais WT.

36
Figura 19. Avaliação dos níveis de fosforilação da enzima Óxido Nítrico Sintase endotelial (peNOS Ser1177) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). Os resultados
estão representados em média ± e.p.m. (***p<0,001 entre WT x B1-/- e WT x B2
-/-).
Reforçando esses resultados (Figura 19), os dados apresentados na figura 20
mostram a expressão da eNOS fosforilada no resíduo de treonina na posição 495 (peNOS Thr495), conhecido por inativar a eNOS, quando fosforilado (FÖRSTERMANN e SESSA,
2012; ALDERTON, COOPER e KNOWLES, 2001). Detectamos uma redução significativa
(p<0,05) de aproximadamente 44% e 45% na expressão da peNOS Thr495, nos corações dos
animais dos grupos B1-/- e B2
-/-, respectivamente, quando comparados aos animais WT.

37
Figura 20. Avaliação dos níveis de fosforilação da enzima Óxido Nítrico Sintase endotelial (peNOS Thr495) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). Os resultados
estão representados em média ± e.p.m. (*p<0,05 entre WT x B1-/- e WT x B2
-/-).
Na figura 21 esta mostrada a medida da expressão total da nNOS, detectamos um
aumento significativo (p<0,05), aproximadamente 27% e 31%, nos corações dos animais dos
grupos B1-/- e B2
-/-, respectivamente, quando comparados aos animais WT.
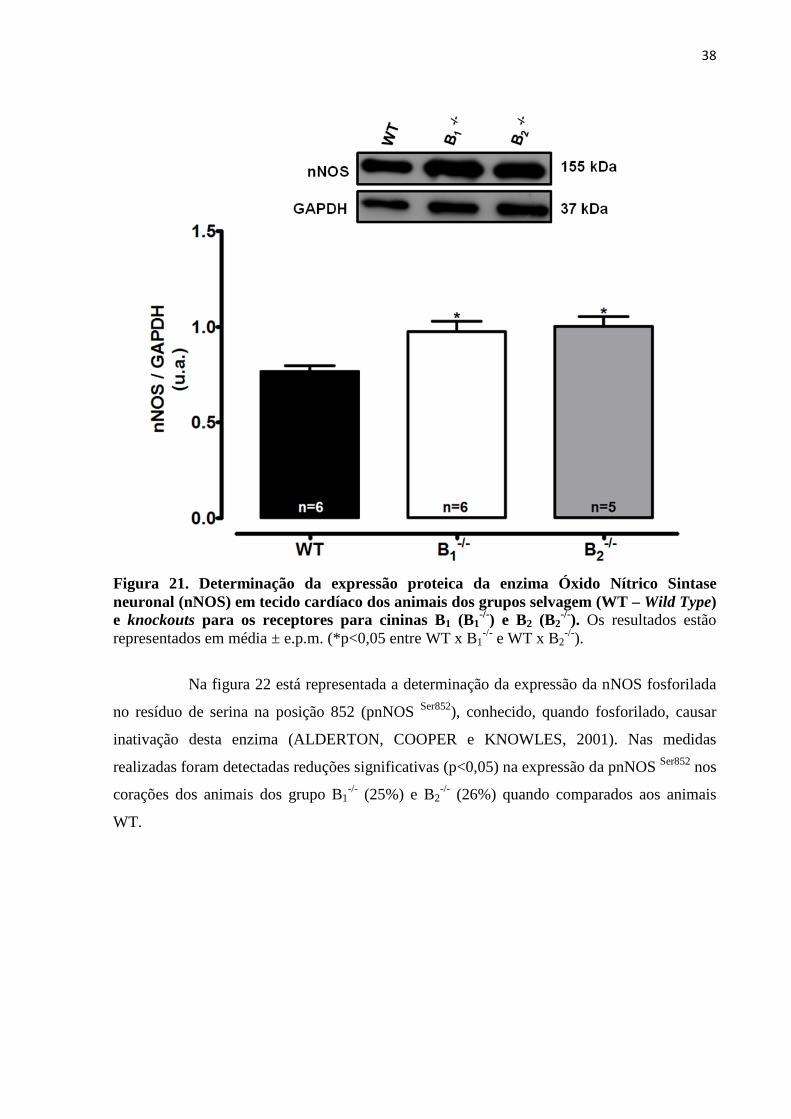
38
Figura 21. Determinação da expressão proteica da enzima Óxido Nítrico Sintase neuronal (nNOS) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). Os resultados estão
representados em média ± e.p.m. (*p<0,05 entre WT x B1-/- e WT x B2
-/-).
Na figura 22 está representada a determinação da expressão da nNOS fosforilada
no resíduo de serina na posição 852 (pnNOS Ser852), conhecido, quando fosforilado, causar
inativação desta enzima (ALDERTON, COOPER e KNOWLES, 2001). Nas medidas
realizadas foram detectadas reduções significativas (p<0,05) na expressão da pnNOS Ser852 nos
corações dos animais dos grupo B1-/- (25%) e B2
-/- (26%) quando comparados aos animais
WT.
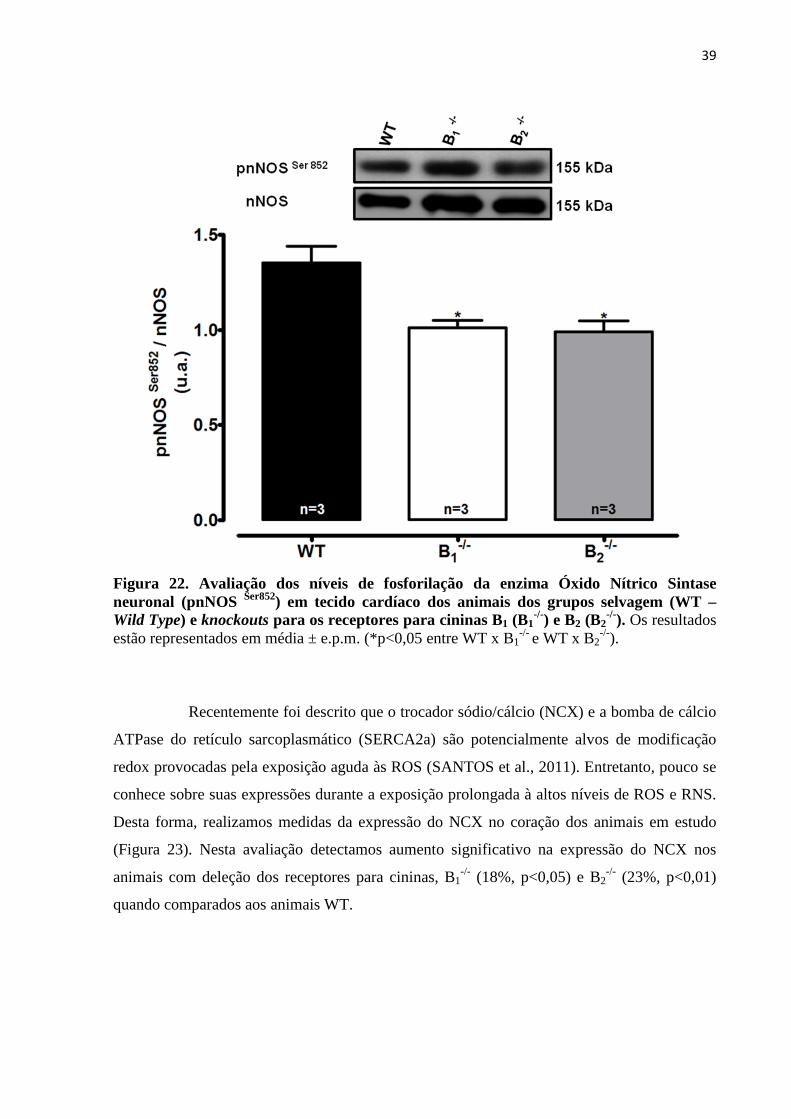
39
Figura 22. Avaliação dos níveis de fosforilação da enzima Óxido Nítrico Sintase neuronal (pnNOS Ser852) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). Os resultados
estão representados em média ± e.p.m. (*p<0,05 entre WT x B1-/- e WT x B2
-/-).
Recentemente foi descrito que o trocador sódio/cálcio (NCX) e a bomba de cálcio
ATPase do retículo sarcoplasmático (SERCA2a) são potencialmente alvos de modificação
redox provocadas pela exposição aguda às ROS (SANTOS et al., 2011). Entretanto, pouco se
conhece sobre suas expressões durante a exposição prolongada à altos níveis de ROS e RNS.
Desta forma, realizamos medidas da expressão do NCX no coração dos animais em estudo
(Figura 23). Nesta avaliação detectamos aumento significativo na expressão do NCX nos
animais com deleção dos receptores para cininas, B1-/- (18%, p<0,05) e B2
-/- (23%, p<0,01)
quando comparados aos animais WT.

40
Figura 23. Determinação da expressão proteica do trocador sódio/cálcio (NCX) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). Os resultados estão representados em média ±
e.p.m. (* p<0,05 entre WT x B1-/- e ** p<0,01 entre WT x B2
-/-).
Na figura 24 está apresentada a medida da expressão da SERCA2a, na qual foi
encontrada uma significativa (p<0,01) redução de 42% e 46%, na expressão desta bomba de
Ca2+ nos corações dos animais com deleção dos receptores para cininas, B1-/- e B2
-/-,
respectivamente, quando comparados aos animais WT.
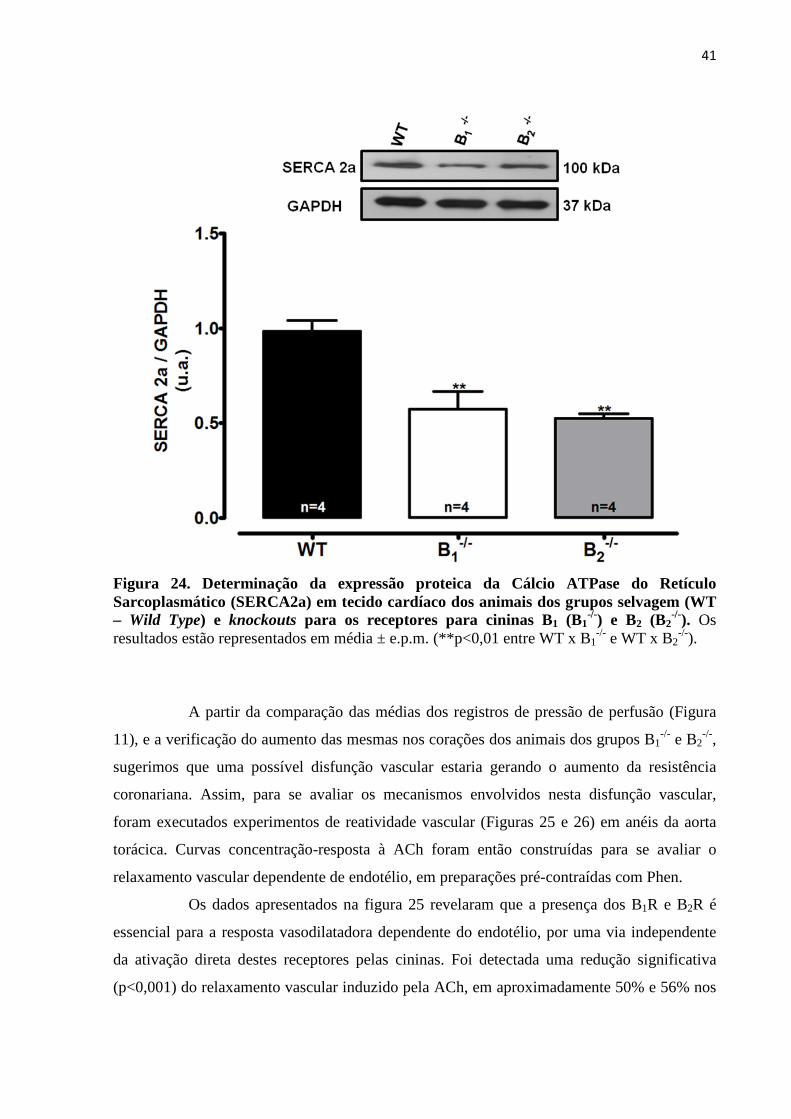
41
Figura 24. Determinação da expressão proteica da Cálcio ATPase do Retículo Sarcoplasmático (SERCA2a) em tecido cardíaco dos animais dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). Os
resultados estão representados em média ± e.p.m. (**p<0,01 entre WT x B1-/- e WT x B2
-/-).
A partir da comparação das médias dos registros de pressão de perfusão (Figura
11), e a verificação do aumento das mesmas nos corações dos animais dos grupos B1-/- e B2
-/-,
sugerimos que uma possível disfunção vascular estaria gerando o aumento da resistência
coronariana. Assim, para se avaliar os mecanismos envolvidos nesta disfunção vascular,
foram executados experimentos de reatividade vascular (Figuras 25 e 26) em anéis da aorta
torácica. Curvas concentração-resposta à ACh foram então construídas para se avaliar o
relaxamento vascular dependente de endotélio, em preparações pré-contraídas com Phen.
Os dados apresentados na figura 25 revelaram que a presença dos B1R e B2R é
essencial para a resposta vasodilatadora dependente do endotélio, por uma via independente
da ativação direta destes receptores pelas cininas. Foi detectada uma redução significativa
(p<0,001) do relaxamento vascular induzido pela ACh, em aproximadamente 50% e 56% nos

42
anéis de aorta obtidos dos animais dos grupos B1-/- (EMAX 48,4 ± 5,9%) e B2
-/- (EMAX 41,7 ±
4,9%), respectivamente, quando comparados aos animais WT (EMAX 95,3 ± 1,2%).
-9 -8 -7 -6 -5 -4
-25
0
25
50
75
100
WTB1
-/-
B2-/-
***
log [ACh] M
Rel
axam
ento
(%
)
Figura 25. Efeito vasorrelaxante induzido pela acetilcolina (ACh) em anéis de aorta de camundongos dos grupos selvagem (WT – Wild Type - n=21) e knockouts para os receptores para cininas B1 (B1
-/- - n=10) e B2 (B2-/- - n=13). Os resultados estão
representados em média ± e.p.m. (***p<0,001, entre WT x B1-/- e WT x B2
-/-).
A disfunção endotelial clássica, caracterizada por redução da vasodilatação
dependente de endotélio, é comumente acompanhada por um aumento da resposta contrátil
vascular à agonistas. Para aprofundar o entendimento das origens da disfunção endotelial
apresentada nos animais com deleção de receptores para cininas, foram construídas curvas de
contração-resposta à Phen. Os resultados apresentados na figura 26 revelaram um aumento
significativo (p<0,001) na tensão desenvolvida durante a contração pela aorta dos animais dos
grupos B1-/- (EMAX 5,1 ± 0,9 mN.mm-1) e B2
-/- (EMAX 11,3 ± 0,2 mN.mm-1), representando,
desta forma, um aumento de 66% e 266% dos respectivos grupos, quando comparados ao
grupo WT (EMAX 3,0 ± 0,2 mN.mm-1).

43
-9 -8 -7 -6 -5 -4
0
2
4
6
8
10
12 WTB1
-/-
B2-/-
***
log [Phen] M
Con
traç
ão (
mN
.mm
-1)
Figura 26. Efeito vasoconstritor induzido pela fenilefrina (Phen) em anéis de aorta de camundongos dos grupos selvagem (WT – Wild Type - n=8) e knockouts para os receptores para cininas B1 (B1
-/- - n=5) e B2 (B2-/- - n=4). Os resultados estão representados
em média ± e.p.m. (***p<0,001, entre WT x B1-/- e WT x B2
-/-).
É bastante consolidado o conhecimento sobre o papel das NOS sobre a função
vascular. Portanto, a fim de avançarmos no conhecimento dos mecanismos moleculares
implicados na disfunção vascular encontrada, propusemos avaliar a participação destas
enzimas no processo de relaxamento vascular destes animais em estudo.
Os resultados apresentados nas figuras 27 e 28 mostram, respectivamente, o
relaxamento vascular induzido pela ACh na presença ou ausência do inibidor não seletivo
para NOS (L-NNA) e do inibidor seletivo para nNOS (TRIM) em anéis de aortas de animais
do grupo WT. A inibição não-seletiva pelo L-NNA (Figura 27) praticamente aboliu o efeito
vasodilatador da ACh (EMAX 92,5 ± 4,4% para 10,8 ± 8,7% ), enquanto a inibição seletiva da
nNOS pelo TRIM (Figura 28) reduziu, pela metade, este efeito (EMAX 91,8 ± 4,0% para 42,7 ±
6,5%).

44
-9 -8 -7 -6 -5 -4
-25
0
25
50
75
100WT L-NNA
***
log [ACh] M
Rel
axam
ento
(%
)
Figura 27. Efeito da inibição não seletiva das NOS pelo L-NNA (100 µmol.L-1) sobre a resposta vasorrelaxante induzida pela acetilcolina (ACh) em anéis de aorta de camundongos do grupo selvagem (WT – Wild Type - n=5 / L-NNA - n=5). Os resultados estão representados em média ± e.p.m. (***p<0,001, entre WT x L-NNA).
-9 -8 -7 -6 -5 -4
-25
0
25
50
75
100
WTTRIM
***
log [ACh] M
Rel
axam
ento
(%
)
Figura 28. Efeito da inibição seletiva da nNOS sobre a resposta vasorrelaxante induzida pela acetilcolina (ACh) em anéis de aorta de camundongos do grupo selvagem (WT – Wild Type - n=8 / TRIM - n=8). Os resultados estão representados em média ± e.p.m. (***p<0,001, entre WT x TRIM).

45
Os experimentos de reatividade vascular conduzidos nos animais knockouts para
receptores para cininas demonstraram que a inibição não seletiva das NOS pelo L-NNA abole
a vasodilatação induzida pela ACh, tanto nos animais B1-/- (Figura 29, EMAX 51,4 ± 6,6% para
EMAX -18,0 ± 2,7%), quanto nos B2-/- (Figura 30, EMAX 51,8 ± 5,1% para EMAX -10,4 ± 4,0%).
Entretanto, em ambos os grupos, a presença do inibidor seletivo para nNOS não foi capaz de
reduzir o relaxamento induzido pela ACh, diferentemente do ocorrido nos animais WT,
sugerindo que nos animais B1-/- (Figura 31, EMAX 69,8 ± 3,7% para EMAX 58,0 ± 6,6%) e B2
-/-
(Figura 32, EMAX 56,7 ± 2,1% para EMAX 53,5 ± 1,7%) há um comprometimento da via da
nNOS.
-9 -8 -7 -6 -5 -4
-25
0
25
50
75
100
B1-/-
L-NNA
***
log [ACh] M
Rel
axam
ento
(%
)
Figura 29. Efeito da inibição não seletiva das NOS pelo L-NNA (100 µmol.L-1) sobre a resposta vasorrelaxante induzida pela acetilcolina (ACh) em anéis de aorta de camundongos knockout para receptor B1 para cininas (B1
-/- - n=8 / L-NNA - n=8).Os resultados estão representados em média ± e.p.m. (***p<0,001, entre B1
-/- x L-NNA).

46
-9 -8 -7 -6 -5 -4
-25
0
25
50
75
100
B2-/-
L-NNA
***
log [ACh] M
Rel
axam
ento
(%
)
Figura 30. Efeito da inibição não seletiva das NOS pelo L-NNA (100 µmol.L-1) sobre a resposta vasorrelaxante induzida pela acetilcolina (ACh) em anéis de aorta de camundongos knockout para receptor B2 para cininas (B2
-/- - n=5 / L-NNA - n=5). Os resultados estão representados em média ± e.p.m. (***p<0,001, entre B2
-/- x L-NNA).
-9 -8 -7 -6 -5 -4
-25
0
25
50
75
100B1
-/-
TRIM
log [ACh] M
Rel
axam
ento
(%
)
Figura 31. Efeito da inibição seletiva da nNOS pelo TRIM (300 µmol.L -1) sobre a resposta vasorrelaxante induzida pela acetilcolina (ACh) em anéis de aorta de camundongos knockout para receptor B1 para cininas (B1
-/- - n=5 / TRIM - n=5). Os resultados estão representados em média ± e.p.m., não apresentaram diferenças significativa (p>0,05).
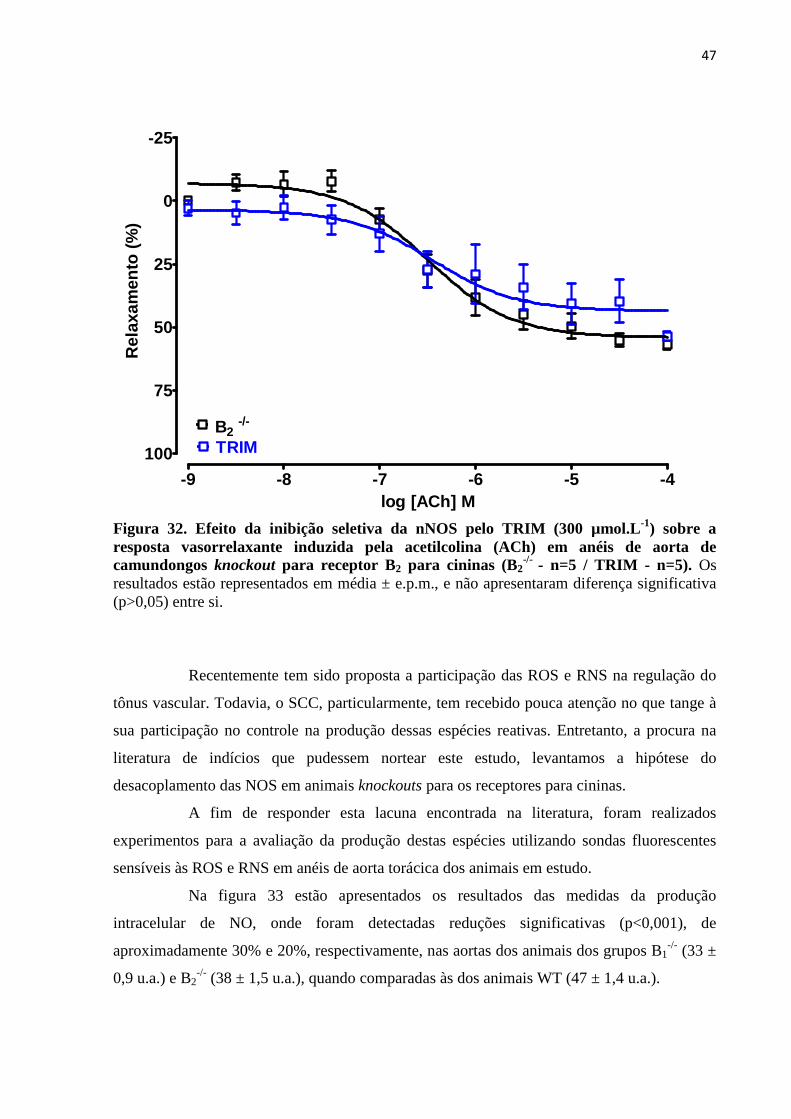
47
-9 -8 -7 -6 -5 -4
-25
0
25
50
75
100
B2-/-
TRIM
log [ACh] M
Rel
axam
ento
(%
)
Figura 32. Efeito da inibição seletiva da nNOS pelo TRIM (300 µmol.L -1) sobre a resposta vasorrelaxante induzida pela acetilcolina (ACh) em anéis de aorta de camundongos knockout para receptor B2 para cininas (B2
-/- - n=5 / TRIM - n=5). Os resultados estão representados em média ± e.p.m., e não apresentaram diferença significativa (p>0,05) entre si.
Recentemente tem sido proposta a participação das ROS e RNS na regulação do
tônus vascular. Todavia, o SCC, particularmente, tem recebido pouca atenção no que tange à
sua participação no controle na produção dessas espécies reativas. Entretanto, a procura na
literatura de indícios que pudessem nortear este estudo, levantamos a hipótese do
desacoplamento das NOS em animais knockouts para os receptores para cininas.
A fim de responder esta lacuna encontrada na literatura, foram realizados
experimentos para a avaliação da produção destas espécies utilizando sondas fluorescentes
sensíveis às ROS e RNS em anéis de aorta torácica dos animais em estudo.
Na figura 33 estão apresentados os resultados das medidas da produção
intracelular de NO, onde foram detectadas reduções significativas (p<0,001), de
aproximadamente 30% e 20%, respectivamente, nas aortas dos animais dos grupos B1-/- (33 ±
0,9 u.a.) e B2-/- (38 ± 1,5 u.a.), quando comparadas às dos animais WT (47 ± 1,4 u.a.).
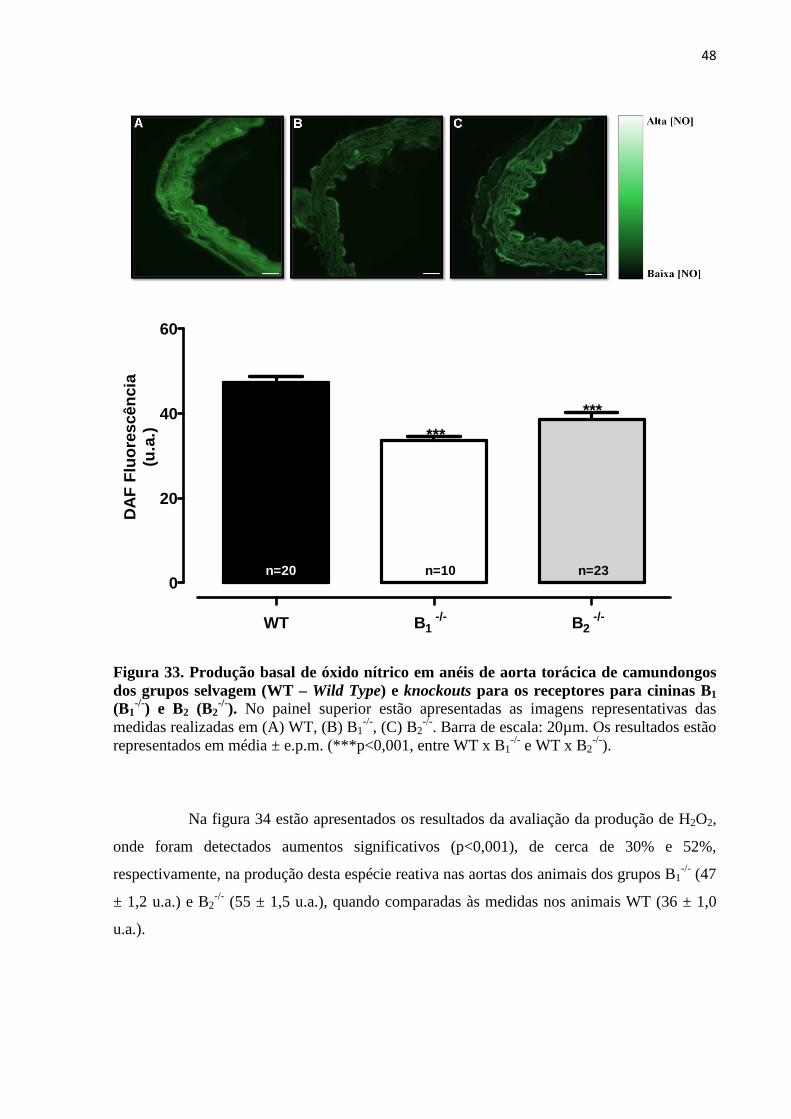
48
WT B1 -/- B2
-/-
0
20
40
60
***
***
n=20 n=10 n=23
DA
F F
luor
escê
ncia
(u.a
.)
Figura 33. Produção basal de óxido nítrico em anéis de aorta torácica de camundongos dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). No painel superior estão apresentadas as imagens representativas das
medidas realizadas em (A) WT, (B) B1-/-, (C) B2
-/-. Barra de escala: 20µm. Os resultados estão representados em média ± e.p.m. (***p<0,001, entre WT x B1
-/- e WT x B2-/-).
Na figura 34 estão apresentados os resultados da avaliação da produção de H2O2,
onde foram detectados aumentos significativos (p<0,001), de cerca de 30% e 52%,
respectivamente, na produção desta espécie reativa nas aortas dos animais dos grupos B1-/- (47
± 1,2 u.a.) e B2-/- (55 ± 1,5 u.a.), quando comparadas às medidas nos animais WT (36 ± 1,0
u.a.).
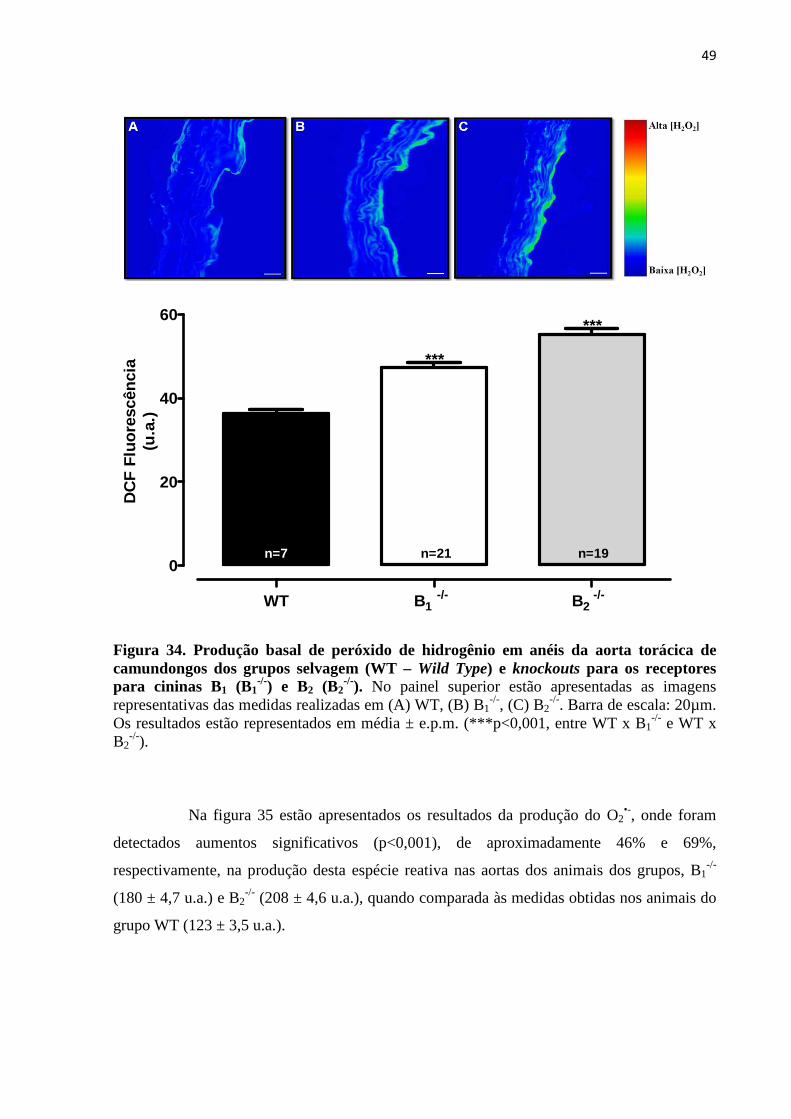
49
WT B1 -/- B2 -/-
0
20
40
60
***
***
n=7 n=21 n=19
DC
F F
luor
escê
ncia
(u.a
.)
Figura 34. Produção basal de peróxido de hidrogênio em anéis da aorta torácica de camundongos dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). No painel superior estão apresentadas as imagens
representativas das medidas realizadas em (A) WT, (B) B1-/-, (C) B2
-/-. Barra de escala: 20µm. Os resultados estão representados em média ± e.p.m. (***p<0,001, entre WT x B1
-/- e WT x B2
-/-).
Na figura 35 estão apresentados os resultados da produção do O2•-, onde foram
detectados aumentos significativos (p<0,001), de aproximadamente 46% e 69%,
respectivamente, na produção desta espécie reativa nas aortas dos animais dos grupos, B1-/-
(180 ± 4,7 u.a.) e B2-/- (208 ± 4,6 u.a.), quando comparada às medidas obtidas nos animais do
grupo WT (123 ± 3,5 u.a.).

50
WT B1 -/- B2
-/-
0
50
100
150
200
250
***
***
n=17 n=11 n=17
DH
E F
luor
escê
ncia
(u.a
.)
Figura 35. Produção basal de ânion superóxido em anéis da aorta torácica de camundongos dos grupos selvagem (WT – Wild Type) e knockouts para os receptores para cininas B1 (B1
-/-) e B2 (B2-/-). No painel superior estão apresentadas imagens
representativas das medidas realizadas em (A) WT, (B) B1-/-, (C) B2
-/-. Barra de escala: 20µm. Os resultados estão representados em média ± e.p.m. (***p<0,001, entre WT x B1
-/- e WT x B2
-/- ).
Com base nos resultados obtidos nos experimentos de reatividade vascular
(Figuras 25 a 32), propusemos que o comprometimento do relaxamento vascular, encontrado
nos animais dos grupos B1-/- e B2
-/-, seja devido ao desacoplamento catalítico da nNOS, e,
ainda, que este fenômeno esteja envolvido na produção de elevadas quantidades de ROS
tecidual identificadas nesses grupos.
A literatura aponta a importância da interação proteína-proteína na regulação do
funcionamento de receptores e enzimas (JU et al., 1998; GOLSER et al., 2000). Desta forma,
para elucidarmos a existência de interação física dos receptores para cininas com a nNOS
(Figura 36) e eNOS (Figura 37), executamos o experimento de co-imunoprecipitação de
proteínas (Figura 36 e 37). Neste ensaio, foram detectados que ambos B1R e B2R, estão

51
espontaneamente heterodimerizados formando um complexo de interação com as enzimas
nNOS e eNOS.
Figura 36. Imunoprecipitação do complexo proteína-proteína nNOS/B1 e nNOS/B2 em aorta torácica de camundongo selvagem (WT – Wild Type). As imagens são representativas de dois experimentos independentes onde foram usados pools de amostras de proteínas. No experimento foi imunoprecipitado (IP) a enzima nNOS e em seguida a identificação por western blot (WB) dos receptores B1 ou B2 para cininas.
Figura 37. Imunoprecipitação do complexo proteína-proteína eNOS/B1 e eNOS/B2 em aorta torácica de camundongo selvagem (WT – Wild Type). As imagens são representativas de dois experimentos independentes onde foram usados pools de amostras de proteínas. No experimento foi imunoprecipitado (IP) a enzima eNOS e em seguida a identificação por western blot (WB) dos receptores B1 ou B2 para cininas.

52
6. DISCUSSÃO
Os resultados apresentados nesta dissertação propõem a participação do SCC na
regulação contrátil do sistema cardiovascular através da sinalização envolvendo as ROS e
RNS. Para este estudo foram utilizados animais transgênicos para investigar a influência do
SCC (via receptores para cininas) na resposta contrátil cardíaca e função vascular.
O primeiro passo deste trabalho, foi confirmar, em animais knockouts, a
modificação genética. Desta forma, os animais foram previamente genotipados e comprovada
a deleção dos genes que expressam os B1R ou B2R para cininas nestes grupos avaliados
(Figuras 6 e 7).
Nolly et al. (1994) detectaram a expressão de mRNAs da calicreína em
cardiomiócitos atriais e ventriculares de ratos. Campbell, Kladis e Duncan (1993) e Lagneux
et al.(1998) mensuraram, no coração, a presença tecidual da BK e seus derivados peptídicos.
Estes estudos demonstraram a contínua produção de cininas no coração isolado, no entanto, a
isquemia parece ser um potente estímulo para uma maior liberação local destes peptídeos
(BAUMGARTEN et al., 1993) e consequente aumento da expressão do B1R e B2R no coração
(TSCHÖPE et al., 2000a; TSCHÖPE et al., 2000b). Em conjunto, estes resultados, associados
à dados da literatura, demonstraram a expressão, no coração, de todos os componentes do
SCC de forma fisiológica e a importância da sinalização deste sistema em situações de injúria.
Os efeitos benéficos cardíacos apresentados pelas cininas foram observados após
administração local de BK em miocárdio isquêmico de suínos (LINZ, WIEMER e
SCHOLKENS, 1996; TIO et al., 1991). Tais efeitos cardioprotetores são também
identificados através do aumento na expressão da calicreína em ratos transgênicos (SILVA et
al., 2000; PINTO et al., 2000), essas respostas, são semelhantes às obtidas com a inibição da
ECA (MARTORANA et al., 1990; ERTL et al., 1982).
Tem sido demonstrado, em estudos experimentais e clínicos, que os efeitos
cardioprotetores da inibição da ECA são mediados pela ativação dos B2R (HORNIG,
KOHLER e DREXLER, 1997; SILVESTRE et al., 2001; LIU et al., 1997). Entretanto,
diversas evidências indicam a importância da sinalização dos B1R nas ações envolvidas na
inibição desta enzima (WITHEROW et al., 2001; IGNJATOVIC et al., 2002; MARIN-
CASTANO et al., 2002).
Tschöpe et al. (2004) demonstraram que o B1R apresenta potencial cardioprotetor
e propõe, a partir de observações anteriores, que suas ações envolvem a modulação de
liberação da noradrenalina (FOUCART et al., 1997), e efeito antiarrítmico após reperfusão

53
(CHAHINE et al., 1993). Adicionalmente, a ativação do B1R também envolve o aumento na
produção das prostaglandinas, NO ˗ gerando a vasodilatação das artérias coronarianas (SU et
al., 2000) ̠ e efeitos antiproliferativos, estes últimos, suprimindo o fator de crescimento
fibroblastos e consequentemente a síntese de colágeno (KIM et al., 1999).
As funções cardioprotetoras da BK intermediada pelo B2R são classicamente
conhecidas, dentre as quais, sua ação anti-hipertrófica tem recebido bastante atenção
(EMANUELI et al., 1999). Tem sido proposto que a BK impede a hipertrofia de
cardiomiócitos isolados, através da liberação do NO associada à elevação de GMP cíclico
(cGMP) (ROSENKRANZ, DUSTING e RITCHIE, 2000).
Corroborando as observações anteriores, Rosenkranz et al. (2002) demonstraram,
em modelo de hipertrofia induzida por Ang II, que a BK (100 nmol), exerce um importante
papel anti-hipertrófico, sugerindo uma ação semelhante ao inibidor da ECA, ramiprilat (100
nmol), e ao doador de NO, nitroprussiato de sódio (3 µmol). Estes efeitos foram mediados
através da liberação de cGMP, que modula diferentes vias de sinalização intracelulares como,
a inibição da MAPK (SUHASINI et al., 1998) e endotelina-1 (GRAY et al., 1998).
Alternativamente, outros autores sugerem que a BK dispara mecanismos
adicionais independentes da sinalização por cGMP, incluindo, a ativação de receptores
purinérgicos do tipo P2 (ANNING et al., 1999), supressão do metabolismo mitocondrial
(STUMPE, DECKING e SCHRADER, 2001), disparando a produção de ROS (MASSOUDY,
BECKER e GERLACH, 1995), inibição da proliferação de células endoteliais (HELLER et
al., 1999), ativação direta de canais para Ca2+ dependente de voltagem (BOLOTINA et al.,
1994) e regulação direta da bomba Na+/K+ cardíaca (HOOL et al., 1995).
A literatura tem evidenciado a participação do SCC na função cardíaca
(LAUTON-SANTOS, et al., 2007; ROMAN-CAMPOS et al., 2010; EMANUELI et al.,
1999). A primeira parte de nossos resultados (Figura 8 – 11) não são inovadores, uma vez que
já foram previamente publicados (LAUTON-SANTOS, et al., 2007). Entretanto, estes
resultados tiveram papel essencial no acompanhamento do fenótipo dos animais descrito
naqueles estudos, os quais serviriam de base para o nosso trabalho.
Em nossos estudos, detectamos a participação do B1R e B2R para cininas na
manutenção da homeostase fisiológica do sistema cardiovascular (Figuras 8 – 11 e 25 – 32).
Esta afirmação originou-se de observações experimentais nas quais as deleções gênicas destes
receptores causaram modificações nas vias de sinalização intracelulares que repercutiram no
comprometimento da função sistólica cardíaca, com redução significativa da mesma (Figura

54
8). Apesar disto, nenhuma modificação foi detectada na tensão diastólica ou frequência
cardíaca espontânea durante seus registros em coração isolado (Figuras 9 e 10).
Nossos resultados apresentados na figura 8 reproduziu a observação da redução da
tensão sistólica verificada anteriormente (LAUTON-SANTOS, 2007) e estão de acordo com a
literatura onde foi demonstrado que cardiomiócitos de animais B1-/- apresentam uma redução
de 9% no pico do transiente de Ca2+, mas não apresentam alterações na taxa de recaptação
deste íon pela SERCA2a e, também, a corrente que flui pelo canal para cálcio do tipo-L não
estava alterada. Em nossa análise, estes resultados sugerem que as alterações funcionais
apresentadas pelo coração isolado podem ter ocorrido por modificações na atividade do RyR2
ou ainda, no conteúdo de Ca2+ do retículo sarcoplasmático (LAUTON-SANTOS et al., 2007).
De forma similar, nossos resultados dos registros do coração dos animais B2-/-,
também indicaram reduções na tensão sistólica (Figura 8), estes dados são corroborados pela
literatura, na qual, Roman-Campos et al. (2010) detectaram modificações em cardiomiócitos
ventriculares de animais B2-/- apresentando redução de 61% no encurtamento celular, avaliado
em sistema desenvolvido para detecção da velocidade e amplitude de contração de células
quando estimuladas eletricamente ou farmacologicamente. Detectaram, ainda, redução na
densidade de corrente através do canal retificador para potássio e no canal para Ca2+ do tipo-L
com concomitante prejuízo, cerca de 17%, no pico do transiente de Ca2+ intracelular.
Duka et al. (2008) demonstraram evidências de que a deleção do B1R aumenta a
expressão de mRNA do B2R, sugerindo existir um mecanismo de intercomunicação e
regulação entre os receptores para cininas. Estes autores sugerem que esta seja uma resposta
compensatória, no entanto, a regulação positiva dos receptores é insuficiente para criar efeito
cardioprotetor satisfatório, enfatizando, desta forma, a importância da presença de ambos
receptores de cininas para efetivamente produzir efeito cardioprotetor.
A contribuição dos B1Rs para a manutenção da função cardíaca sobre o estado
fisiológico e fisiopatológico tem sido pouco explorada. Bouchard, Chouinard e Lamontagne
(1998) demonstraram que o pré-condicionamento isquêmico protege a função endotelial, e
que o B1R, em parte, participa deste efeito protetor. Um mecanismo molecular possivelmente
implicado neste processo envolve a regulação positiva destes receptores através da
retroalimentação produzida pelo seu agonista, o DBK. Este peptídeo, de fato, representa um
poderoso sinal para indução de sua exponencial produção durante a isquemia cardíaca
(SCHANSTRA et al., 1998). Esta sinalização rápida ocorre através da transcrição gênica
envolvendo a proteína p38MAPK (CLERK et al., 1998) e o NF-κB (CHANDRAZEKAR e

55
FREEMAN, 1997) levando a expressão de novos B1R (LARRIVÉE et al., 1998; NI, CHAO,
CHAO, 1998).
Contraditoriamente, Xu et al. (2009) demonstraram que o B1R não parece
desempenhar papel essencial na hemodinâmica e na função cardíaca, em condições normais,
ou durante o desenvolvimento de insuficiência cardíaca. No entanto, podem estar envolvidos
na manutenção da estrutura cardíaca e integridade morfológica, já que camundongos B1-/-
apresentaram aumento da massa ventricular esquerda e da dimensão desta câmara em
condições basais, porém sem prejuízos na função cardíaca (TSCHÖPE et al, 2000).
Embora a deleção genética dos componentes do SCC, calicreína tecidual e o B2R,
torne susceptível o crescimento celular exagerado, Trabold et al. (2002) demonstraram um
fenótipo cardíaco normal em fêmeas de 3 a 5 meses de idade, deficiente em B2R, sugerindo
que períodos mais longos de tempo são necessários para a mutação resultar em significativos
efeitos cardíacos. Além disso, os autores acima citados propuseram que a influência protetora
dos hormônios sexuais femininos como o estrogênio ˗ que exerce ação anti-hipertrófica direta
e ação antiapoptótica em cardiomiócitos ˗ poderia ter prevenido o remodelamento ventricular
em animais B2-/- (GROHE et al, 1997). Estas contradições observadas podem ser devido à
diferenças metodológicas usadas pelos diversos autores.
Mais evidências da participação do SCC na função fisiológica cardíaca tem sido
apontadas em estudos com animais deficientes no gene que expressa o B2R ou a calicreína
tecidual, que mostraram ativação desequilibrada do sistema renina-angiotensina (SRA)
(EMANUELI et al., 1999; MENETON et al., 2001). Essas respostas também foram descritas
por Madeddu et al. (2000) que observaram que o bloqueio do receptor de angiotensina
(AngII) AT1 impede o remodelamento cardíaco hipertrófico de animais B2-/-.
Além disso, evidências sugerem a participação de receptores envolvidos no
controle hemodinâmico e na sinalização que contribui para a geração do estresse oxidativo e,
consequentemente, a reprogramação gênica e a hipertrofia dos miócitos cardíacos (IZUMIYA
et al., 2003; HIGUCHI et al., 2003; MARKOU, HADZOPOULOU-CLADARAS e LAZOU,
2004).
Em cultura de cardiomiócitos, a Ang II é capaz de estimular a produção de ROS e
fatores de crescimento que levam ao aumento da expressão ou da atividade de vários
reguladores críticos da hipertrofia cardíaca, tais como, MAPKs e o NF-κB (WENZEL et al.,
2001). A ativação de NF-κB foi também demonstrada desempenhar um papel crítico na
produção de ROS durante a hipertrofia cardíaca (PURCELL et al., 2001; HIGUCHI et al.,
2002).

56
Baseado nas observações do comprometimento da função cardíaca e na
importância de sua regulação pelas ROS (SANTOS et al., 2011), resolvemos investigar o
status redox nos animais deficientes de receptores para cininas.
Nossos resultados de mensuração intracelular das ROS em cardiomiócitos
isolados podem ser visualizados nas figuras 12 e 13, os quais comprovaram que os animais
pertencentes aos grupos B1-/- e B2
-/- apresentam um significativo aumento na produção destas
espécies reativas. Em particular, demonstramos que a produção de O2•- (Figura 12) teve
aumento substancial em ambos os grupos, B1-/- e B2
-/-, acompanhado pela baixa expressão da
enzima Cu/Zn SOD (Figura 15), que catalisa a dismutação do O2•- em H2O2.
Apesar da alta reatividade (velocidade de reação: 109 M-1s-1) (HUIE e
PADMAJA, 1993), o O2•- desempenha uma variedade de efeitos biológicos. Estes efeitos
podem produzir prejuízos, não somente, pela alteração da sua velocidade de reação química,
mas também, através da produção de outras ROS, como o H2O2 que pode reagir com o NO
gerando um radical ainda mais reativo, o peroxinitrito (ONOO-) (CZAPSKI e GOLDSTEIN,
1995).
Em sequência, executamos experimentos para medidas de H2O2 (Figura 13), onde
encontramos o aumento significativo da produção desta espécie. Este resultado foi reforçado
pela verificação da expressão reduzida da enzima catalase (Figura 16) e GPx (Figura 19), que
decompõem a referida espécie reativa em água (H2O) e oxigênio (O2). Estes achados
reforçaram nossa hipótese da ocorrência de estresse oxidativo na ausência dos receptores para
cininas.
Oldenburg et al. (2004) demonstraram que cardiomiócitos ventriculares produzem
grandes quantidades de ROS quando estimulados com BK, sendo proposto, a participação dos
canais mitocondriais para potássio sensíveis ao ATP (mitoKATP) na produção desta resposta.
Corroborando esses resultados, Sasaki et al. (2000), demonstraram em cardiomiócitos
ventriculares de coelhos, que o doador de NO (SNAP), provoca a abertura dos canais de
mitoKATP, através da ativação do cGMP, e, consequentemente, fosforilação da PKG e geração
das ROS.
Rakhit et al. (2000), utilizando coração de ratos submetidos ao pré-
condicionamento isquêmico, identificaram que o inibidor da guanilato ciclase (ODQ),
bloqueou os efeitos cardioprotetores produzido pelo SNAP, e concluíram a dependência do
cGMP na cardioproteção produzida pelo NO. Portanto, estes dados são consistentes e
demonstram a participação das RNS em eventos cardioprotetores.

57
O SCC é classicamente conhecido por apresentar ação cardioprotetora através da
ativação de vias intracelulares que produzem o NO, demonstrando melhora da função
ventricular após isquemia cardioplégica (OLDENBURG et al., 2004). Tem sido demonstrado
que a sinalização envolvendo as cininas, é um importante mecanismo de proteção em eventos
isquêmicos e de pré-condicionamento (GOTO et al., 1995, PAN et al., 2000, WALL,
SHEEHY e HARTMAN, 1994) e, a ativação do B2R sugere ser o gatilho fisiológico desta
cardioproteção (GOTO et al., 1995, WALL, SHEEHY e HARTMAN, 1994).
A partir das evidências que revelam a participação do NO em vias de sinalização
intracelulares iniciada pelo SCC, realizamos a medida da produção intracelular de NO em
células cardíacas ventriculares (Figura 14). Foi detectado aumento significativo da
fluorescência, sendo esta medida, indicativo de que os cardiomiócitos dos animais
pertencentes aos grupos B1-/- e B2
-/- tem produção aumentada do NO. Esses achados foram
previamente descritos, e, comprovam que animais deficientes de B1R e B2R para cininas
apresentam distúrbio na síntese desta biomolécula (LAUTON-SANTOS et al. 2007 e
ROMAN-CAMPOS et al. 2010).
Para buscarmos a gênese da produção desequilibrada do NO (Figura 14),
realizamos experimentos para mensurar a expressão proteica das principais enzimas que a
sintetizam no coração, eNOS e nNOS, conhecidas por apresentarem expressão constitutiva
neste tecido (Figuras 18 e 21). Em nossos resultados, identificamos aumentos significativos
tanto na expressão total de eNOS (Figura 18), quanto na de nNOS (Figura 21) nos corações
dos animais dos grupos B1-/- e B2
-/-. Portanto, sugerimos que nossos resultados anteriores, os
quais mostraram o aumento de NO intracelular (Figura 14), podem ser explicados devido ao
aumento na expressão destas enzimas.
Lauton-Santos et al. (2007) demonstraram que animais B1-/- apresentam aumento
na expressão do mRNA de nNOS e iNOS, não sendo detectado alterações na expressão do
mRNA da eNOS. Entretanto, Duka et al. (2008) demonstraram que ambos grupos, B1-/- e B2
-/-,
apresentam aumento na expressão do gene que codifica a síntese de eNOS e, posteriormente,
Roman-Campos et al. (2010) verificou aumento na expressão proteica da eNOS, mas, com
redução da expressão da nNOS em coração de animais B2-/-. A controvérsia de alguns de
nossos resultados com dados da literatura, pode ser justificada pelas diferentes técnicas
metodológicas utilizadas nos estudos citados.
Demonstramos, ainda, que além destas enzimas estarem com expressão
aumentada, ambas, eNOS e nNOS, apresentaram aumento significativo em suas atividades
nos grupos de animais B1-/- e B2
-/ (Figura 19, 20 e 22). A atividade da eNOS foi medida

58
através da fosforilação da serina1177, onde verificamos o aumento significativo da expressão
da peNOSSer1177, sendo este, um sítio de ativação desta enzima (Figura 19) (FÖRSTERMANN
e SESSA, 2012; ALDERTON, COOPER e KNOWLES, 2001). De forma similar,
comprovamos este resultado uma vez que demonstramos a diminuição da fosforilação no
resíduo de treonina495, peNOS Thr495, sítio de inativação da enzima (Figura 20)
(FÖRSTERMANN e SESSA, 2012; ALDERTON, COOPER e KNOWLES, 2001).
A atividade da nNOS foi também verificada por ensaios similares (Figura 22).
Identificamos diminuição significativa na expressão da forma fosforilada desta enzima, no
resíduo serina852, pnNOSSer852, a fosforilação deste resíduo, na referida enzima, está
relacionado ao seu estado inativado (ALDERTON, COOPER e KNOWLES, 2001).
Nosso conjunto de resultados demonstrou, de forma convincente, que o aumento
da produção do NO em cardiomiócitos com deleção de receptores para cininas (Figuras 14),
ocorre devido, não somente, ao aumento da expressão das enzimas eNOS e nNOS, como,
também, pelo aumento na atividade das mesmas (Figuras 18 - 22).
Vários autores sugerem que a produção de ROS dependente de cininas é iniciada
pela produção do NO, que irá conduzir efeitos diretos e indiretos em nível subcelular. Estes
efeitos envolvem modificações complexas na estrutura e atividade de algumas proteínas.
Dentre estas modificações, pode ser evidenciado a abertura de canais mitoKATP e
consequentemente, geração de ROS (OLDENBURG et al., 2002; LAGNEUX et al. 2003;
LOCHNER et al., 2000; VEGH, SZEKERES e PARRATT, 1991) que podem levar à ativação
de cascatas de sinalização via modificações redox.
Dentre as principais modificações redox em proteínas, os resíduos de cisteínas
representam um alvo molecular importante. Dentre as proteínas, receptores e bombas
relacionados à homeostase do cálcio que podem sofrer essas modificações, destaca-se a
SERCA2a, conhecida por ser modificada pela oxidação da cisteína ou nitração de tirosina,
entretanto, vários outros resíduos aminoácidos podem ser modificados oxidativamente
(SCHÖNEICH e SHAROV, 2006; ADACHI et al., 2004; XU et al., 2006; SHAROV et al.,
2006). Estas modificações, nesta bomba, incluem a nitração de tirosinas294 e 295 (XU et al.,
2006), S-glutationação e oxidação de cisteínas669 e 674 (ADACHI et al., 2004; SHAROV et al.,
2006). Sugere-se que os baixos níveis de ROS podem aumentar a atividade da SERCA2a,
enquanto, níveis mais elevados, podem causar inativação como resultado das irreversíveis
modificações oxidativas (SCHÖNEICH e SHAROV, 2006; SHAROV et al., 2006).
As alterações causadas pelo ambiente redox celular podem também culminar em
alteração da translocação de fatores de transcrição celular, levando, assim, a alterações na

59
expressão gênica de proteínas (SANTOS et al., 2011). Para conhecermos acerca da
participação das ROS e RNS sobre a regulação das expressões de proteínas que estão
relacionadas com a homeostase de cálcio no coração, foram realizados medidas da expressão
da SERCA2a (Figura 24) e NCX (Figura 23).
Nossos resultados, revelaram que corações de animais B1-/- e B2
-/- apresentam
reduções na expressão da SERCA2a (Figura 24), e aumento na expressão do NCX (Figura
23). Juntos, estes dados justificam, em parte, nossos resultados de contratilidade sistólica
cardíaca diminuída (Figura 8-10). Entretanto, Lauton-Santos et al. (2007) e Romam-Campos
et al. (2010) não detectaram alterações no tempo de recaptação do Ca2+ em cardiomiócitos de
animais B1-/- e B2
-/-, indicando não haver alteração na atividade da SERCA2a. Esta
discordância dos nossos resultados pode ser devido a utilização de métodos indiretos para
medida da atividade desta bomba utilizada nos trabalhos citados acima.
A literatura tem associado redução no transiente de Ca2+ em cardiomiócitos ao
estresse oxidativo provocado, dente outras ROS, pelo H2O2 (LI et al., 2006). Esta molécula
pode estar envolvida na diminuição na atividade da SERCA2a e aumento da atividade do
NCX. Em nossos resultados detectamos redução da expressão da SERCA2a (Figura 24) e
aumento na expressão do NCX (Figura 23), que juntos desempenham a função de reduzir o
conteúdo de Ca2+ no citosol (KUSTER et al., 2010).
Tocchetti et al. (2007) relataram efeito inotrópico positivo envolvendo exposição
aguda de cardiomiócitos ao nitroxil, sendo detectado, aumento na recaptação do Ca2+ para o
retículo sarcoplasmático. As modificações oxidativas irreversíveis, envolvendo exposição
prolongadas às ROS e RNS, produzem alterações na atividade da SERCA2a e NCX
contribuindo para a anormalidade no transiente de Ca2+ e déficit da função contrátil. Assim,
estes resultados reforçam nossa hipótese da participação das ROS e RNS na regulação da
homeostase do Ca2+ intracelular, visto que, como previamente descrito, em cardiomiócitos de
animais B1-/- e B2
-/- apresentam redução no pico de transiente de Ca2+ (LAUTON-SANTOS et
al., 2007 e ROMAN-CAMPOS et al., 2010).
Finalizamos a discussão dos nossos resultados obtidos em coração propondo a
participação das ROS (Figuras 12-13, 15-17) e RNS (Figuras 14 e 18-22) como biomoléculas
sinalizadoras do SCC, sendo estas moléculas, capazes de criar modificações redox e nitroso
e/ou capazes de modular a expressão de proteínas que estão relacionadas com a homeostase
de Ca2+ intracelular em cardiomiócitos ventriculares (Figuras 23 e 24) e, portanto, regulando a
função contrátil cardíaca (Figuras 8-10).

60
Durante o experimento de registro da função contrátil cardíaca no sistema de
Langendorff, foram simultaneamente adquiridos os resultados da pressão de perfusão (Figura
11), onde, foram detectados uma pressão de perfusão aumentada nos grupos B1-/- e B2
-/-, sendo
esta, uma medida do aumento da resistência coronariana. Esses resultados comprovam dados
da literatura que apontam a participação do SCC na função vascular (LOIOLA et al., 2011).
Ainda hoje é bastante controversa a vasodilatação, dependente do endotélio,
produzida pelos peptídeos derivados do SCC, na circulação coronária de ratos (McLEAN et
al., 1999), bois (DRUMMOND, 1995), porcos (PRUNEAU et al., 1996), coelhos
(PRUNEAU e BELICHARD, 1993) e humanos (DRUMMOND et al., 1995) sendo
demonstrado, que a vasodilatação não envolve a ativação das ciclo-oxigenases e geração de
prostaciclinas, mas sim, da geração endógena do NO.
Contrariamente, tem sido demonstrado em alguns estudos que a ativação de B1R
pode causar constrição em aorta de coelho (LEVESQUE et al., 1995; BAWOLAK et al.,
2008), veia mesentérica de coelho (REGOLI, MARCEAU e BARABE, 1978), veia renal de
porcos (RIZZI et al., 1997) e em veia e artéria umbilical de humanos (GOBEIL et al., 1996a;
ABBAS et al., 1998).
Foi demonstrado em anéis da aorta abdominal de camundongos da linhagem
C57JBL/6, que a ativação do B2R e B1R é capaz de produzir vasoconstrição de 51% e 30%,
respectivamente (FELIPE et al, 2007). No entanto, se propõe que a vasocontrição identificada
é geralmente independente do endotélio, e, envolve a ativação de PKC e influxo de
Ca2+(McLEAN et al., 1999). Portanto, avaliar a função vascular dos componentes do SCC
parece ser uma tarefa ainda mais complexa, que envolve critérios experimentais como a
escolha da espécie animal e/ou leito vascular estudado.
A contração no músculo liso vascular é desencadeada por uma série de estímulos
extracelulares, como a ligação de substâncias químicas, hormônios, neurotransmissores
peptídeos aos seus receptores na membrana plasmática ou através da ativação de receptores
ou canais sensíveis a distensão mecânica (BOLOTINA et al., 1994).
A Phen, é classicamente conhecida por produzir uma potente vasoconstrição,
atuando como agonista de receptores adrenérgicos α1, sendo, sua resposta contrátil,
dependente da mobilização Ca2+. Nossos resultados de reatividade vascular (Figura 26)
demonstraram a participação do SCC na resposta contrátil do músculo liso vascular.
Demonstramos que a aorta torácica quando estimulada com Phen produziu uma amplificada
vasoconstrição em animais dos grupos B1-/- e B2
-/- quando comparados com WT. Esses

61
resultados nos sugeriram que os receptores para cininas participam da mobilização de Ca2+
intracelular.
Partindo deste pressuposto, e tentando esclarecer os resultados acima observados,
executamos ensaios experimentais onde verificamos, ainda, um significativo prejuízo no
relaxamento vascular induzido pela ACh (Figura 25). Sabe-se que a ativação do receptor
muscarínico M3 produz o NO, considerado o principal fator relaxante derivado do endotélio
(FURCHGOTT e ZAWADZKI, 1980). Desta forma, a fim de avaliar a participação do NO
nestas respostas, medimos na condição basal a produção do NO em anéis de aorta torácica
(Figura 33). Detectamos uma reduzida biodisponibilidade de NO neste leito vascular
justificando, assim, o reduzido relaxamento vascular nos grupos B1-/- e B2
-/-.
O comprometimento na produção e biodisponibilidade do NO em animais
knockout para os receptores de cininas foi recentemente proposto por Loiola et al. (2011), que
também demonstraram uma reduzida resposta vasorrelaxante em artéria mesentérica,
comprovando experimentalmente a baixa biodisponibilidade do NO tecidual e sistêmico.
Entretanto, os resultados de ensaios enzimáticos da atividade das NOS constitutivas
demonstraram aumento significativo, e estes autores propuseram um funcionamento
desacoplado das NOS.
As NOS são heme proteínas com arquitetura molecular homodimérica durante seu
estado ativado. Em sua estrutura, os monômeros homólogos das NOS são constituídos por um
domínio C-terminal transportador de elétrons denominado domínio redutor, ligante à
calmodulina, e um domínio N-terminal ou oxidante, que forma o sítio catalítico para síntese
do NO (FÖRSTERMANN, 2000).
A síntese do NO ocorre através da transferência de elétrons iniciada no domínio
redutor, onde se encontram os sítios de ligação para aceptar os elétrons doados pelo NADPH,
que doa elétrons para os cofatores, flavina-adenina dinucleotídeo (FAD) e flavina
mononucleotídeo (FMN), que transferem esses elétrons para o domínio redutor. Entretanto,
entre os domínios redutor e oxidante, encontra-se o domínio de ligação à calmodulina, que,
nas isoformas eNOS e nNOS, este sítio desempenha a importante função de autoinibição
enzimática (FENG, 2012).
No domínio oxidante, os elétrons são transferidos para o átomo de ferro do
grupamento heme, permitindo a ligação do oxigênio ao sítio de ligação do cofator
tetraidrobiopterina (BH4), ao substrato, a L-arginina (FLEMING e BUSSE, 1999). A reação
catalítica das NOS constitutivas envolve dois estágios de oxidação: a hidroxilação da L-
arginina em NG-hidroxi-L-arginina, seguida da oxidação desta molécula formando L-citrulina

62
e NO (ANDREW e MAYER, 1999). Na reação de síntese do NO, a contribuição relativa dos
cofatores, como ferro heme, BH4 e L-arginina, tem recebido bastante atenção, sendo descrito
que a baixa biodisponibilidade desses componentes é capaz de provocar o desacoplamento da
atividade enzimática das NOS (VASQUEZ-VIVAR et al., 1998; CAI e HARRISON, 2000).
Durante o estado de desacoplamento, as NOS permanecem ativas e são capazes de
receber e transferir elétrons entre seus domínios redutor e oxidante. Entretanto, a ausência do
cofator BH4 e/ou do substrato L-arginina, gera a redução do grupo heme, que, gradativamente,
doa elétrons ao O2, e consequentemente, produz o O2•- (SATOH et al., 2005). A literatura tem
proposto que durante o estado de desacoplamento não ocorre diminuição da síntese do NO,
mas aumento de seu consumo em outras reações, como por exemplo, reação com O2•-, e,
consequentemente, reduzindo sua biodisponibilidade (MONCADA e HIGGS, 1993).
Sugere-se que o principal componente deste desacoplamento seja a ausência de
BH4, que está envolvido na manutenção da dimerização dos momômeros das NOS
(FÖRSTERMANN e SESSA, 2012). Além disso, sugere-se também, que o BH4 regula a
atividade do complexo Ferro(I)-O2 (presente no grupamento heme) permitindo o correto
posicionamento do grupamento para a oxidação da NG-hidroxi-L-arginina, e
consequentemente, síntese do NO (ALDERTON, COOPER e KNOWLES, 2001;
FÖRSTERMANN e SESSA, 2012).
Em condição fisiológica, as NOS constitutivas desempenham sua atividade sobre
estado acoplado, onde a isoforma eNOS é capaz de produzir somente NO e a nNOS produz
baixas concentrações de O2•-, H2O2 e NO (VASQUEZ-VIVAR et al., 1998; POU et al., 1992).
Entretanto, como previamente descrito, sobre determinadas condições patológicas ou baixa
biodisponibilidade dos substratos e/ou cofatores, a atividade de ambas, eNOS e nNOS, é
drasticamente alterada passando a atuar sob estado desacoplado, produzindo grandes
concentrações de O2•-, H2O2 e NO (VASQUEZ-VIVAR et al., 1998; POU et al., 1992).
Conhecendo a atividade das NOS constitutivas e com os indícios de que sua
atividade esteja desacoplada, sugerimos a ocorrência de disfunção endotelial. Portanto,
realizamos experimentos para mensurar a produção tecidual das ROS, H2O2 (Figura 34) e O2•-
(Figura 35), onde detectamos elevada produção dessas espécies, fortalecendo nossa proposta
que a deleção genética cria perturbações na homeostase catalítica das NOS.
Esta bem estabelecida a participação conjunta das isoformas eNOS e nNOS na
resposta vasorrelaxante induzida pela ACh. A deleção da isoforma nNOS atenua a resposta
vasodilatadora em aorta de camundongos (NANGLE, COTTER e CAMERON, 2004).

63
Confirmando esta proposta, Capettini et al. (2008) descreveram a participação do eixo
nNOS/H2O2/NO no relaxamento vascular.
A fim de avançarmos no conhecimento, propomos avaliar a contribuição relativa
das NOS constitutivas no relaxamento vascular, sendo assim, nossos resultados no grupo WT
demonstraram, através do experimento de reatividade vascular, a contribuição significativa
das NOS (Figura 27) e, particularmente, da nNOS (Figura 28), no relaxamento induzido pela
ACh.
Para fortalecermos nossa hipótese da participação dos receptores para cininas na
modulação da atividade enzimática das NOS, propomos que a deleção genética do B1R ou
B2R, resulta em distúrbio na produção do NO e geração exacerbada das ROS.
Os resultados de relaxamento vascular induzido pela ACh, na presença de
inibidores não seletivos para NOS (Figuras 29 e 30) e seletivos para nNOS (Figuras 31 e 32),
revelaram que a deleção dos receptores para cininas criam perturbações sobre a via de
sinalização da nNOS, visto que, o relaxamento não apresentou alteração na presença de seu
respectivo inibidor. Por outro lado, a inibição não seletiva das NOS foi capaz de abolir a
resposta vasorrelaxante nos grupos B1-/- e B2
-/-.
Interessantemente, nossos resultados se assemelham ao estudo de Li et al. (2004)
que após infusão crônica de Ang II foi induzido hipertensão arterial, sendo detectado,
desequilíbrio entre a geração de ROS e de moléculas antioxidantes, atribuindo a condição de
estresse oxidativo como fator determinante para uma menor sensibilidade e resposta
vasodilatadora dependente do endotélio.
É clássico o conhecimento das ações da Ang II via receptor AT1 atuando na
resposta inflamatória e alterando a permeabilidade vascular, extravasamento leucocitário e
reparo tecidual (TRACEY, 2002). Este peptídeo é reconhecido como um importante mediador
inflamatório em vários tecidos, incluindo leitos vasculares (SUZUKI et al., 2003), e
produzindo apoptose e remodelamento por fibrose (TOUYZ et al., 2003).
A Ang II participa, ainda, de processos inflamatórios através da geração de ROS e
RNS, como O2•-, OH•, H2O2, NO e o ONOO-. Entretanto, em células vasculares, a principal
fonte de geração de ROS é proveniente da enzima NADPH oxidase (nicotine adenine
dinucleotidephosphate oxidase) que permanece constitutivamente ativa na camada adventícia
e músculo liso vascular, catalisando a constante produção de O2•- (LASSEGUE et al., 2003).
Duka et al. (2001) revelaram que a infusão de Ang II aumentou a expressão de
B2R na aorta dos ratos. Entretanto, Fernandes et al. (2006) demonstraram, semelhantemente,

64
que a hipertensão renovascular provoca aumento na produção sistêmica de Ang II e
consequente aumento na expressão de mRNA do B1R.
Ceravolo et al. (2007) demonstraram também, em aorta de ratos, que a infusão de
Ang II induz o aumento da geração do O2•- e, concomitantemente, produz aumento na
atividade e translocamento nuclear do NF-κB neste leito vascular. Interessantemente, o
tratamento com apocinina, inibidor da NADPH oxidase, reduziu a geração de O2•-, a atividade
e translocamento do NF-κB, como também reduziu a expressão de B1R na aorta de ratos,
sugerindo uma relação de dependência do O2•- com a expressão deste receptor para cininas.
Estes dados revelaram a íntima comunicação e interação entre os SRA e SCC,
onde ambos, desempenham importantes funções na regulação do sistema vascular. Abdalla et
al. (2005) demonstraram, através de experimentos em cultura de células, que os B2R e AT1
estão espontaneamente heterodimerizados. As consequências desta heterodimerização
B2R/AT1 foi descrita em modelo de pré-eclampsia e em células mesenquimais de ratos
espontaneamente hipertensos (Spontaneous hypertensive rats - SHR) onde a interação entre
estes receptores é responsável pelo aumento da sensibilidade, potência e eficácia da resposta à
Ang II, entretanto, com redução da potência e eficácia à BK (ABDALLA et al., 2001;
ABDALLA et al., 2005).
A literatura aponta as interações proteína-proteína como importantes mecanismos
moleculares na sinalização intracelular desencadeada por receptores. Estas interações entre os
receptores para cininas foram recentemente demonstradas em células humanas, onde os B1R e
B2R foram identificados espontaneamente heterodimerizados quando co-expressados em
cultura de células HEK293 transfectadas, sendo detectado, através de co-imunoprecipitação e
co-localização na membrana plasmática por microscopia de imunoelétrons (KANG et al.,
2005). Os autores sugeriram que este é o mecanismo de intercomunicação entre os receptores
para cininas, onde, o estímulo prolongado do B2R, gera a heterodimerização destes receptores
aos B1R, e o conseguinte direcionamento dos B2R para vias de degradação proteolítica. Desta
forma, o B1R passa assumir funções inicialmente desempenhadas pelo B2R (KANG et al.,
2005).
É conhecida a interação física dos receptores de cininas com diversas proteínas G
(MATHIS e LEEB-LUNDBERG, 1991). A ativação do B2R em células endoteliais estimula a
produção do NO através da ativação mediante aumento da concentração de Ca2+ intracelular e
fosforilação da NOS constitutiva (BUSSE e FLEMING, 1995; VENEMA, 2002), entretanto,
este não é o único mecanismo regulador das NOS.

65
Ju et al. (1998) e Golser et al. (2000) detectaram, em células endoteliais, a
interação física do B2R/eNOS e B2R /nNOS através de resíduos de aminoácidos localizados
no 4º domínio intracelular da porção C-terminal do receptor. Experimentalmente, a
estimulação com BK causou dissociação do complexo B2R/eNOS, sugerindo que, a interação
física entre estas proteínas é de natureza inibitória.
Nossos resultados demonstraram, de forma inédita, através da técnica de co-
imunoprecipitação, que em aorta torácica de camundongos, o complexo enzimático da eNOS
e nNOS se encontram heterodimerizados com os B1R e B2R (Figuras 36 e 37). Sendo estes,
fortes indícios, e, que nos fizeram sugerir que a deleção genética dos receptores para cininas
criam perturbações na homeostase catalítica das NOS, principalmente da isoforma nNOS,
gerando seu funcionamento desacoplado, comprovado pela ineficiência do relaxamento
vascular e exacerbada produção de ROS.

66
7. CONCLUSÃO
Concluímos, ao final deste trabalho, que a deleção genética do receptor B1 ou B2
para cininas gera o aumento na produção de ROS e RNS em consequência da diminuída
expressão das enzimas antioxidantes e das que sintetizam o NO. Portanto, nossa hipótese
sugere que o status redox resultante desta ausência de sinalização pelos receptores para
cininas regulam os mecanismos que controlam a função celular do sistema cardiovascular.
Sugerimos ainda, que esta modulação da homeostase celular ocorre pela interação destes
receptores para cininas, entre si, e com as NOS.
8. CONSIDERAÇÕES FINAIS
A participação do SCC na regulação da função cardiovascular é ainda mais
complexa que a apresentada pela literatura atual. Comprovamos a participação deste sistema
na regulação do metabolismo celular, tanto por estar envolvido em vias de sinalização
intracelular que promovem a produção de ROS e RNS, como, também regulando a expressão
de enzimas antioxidantes.
Sugerimos que as alterações cardíacas e vasculares, identificadas neste trabalho
estejam relacionadas aos seguintes fatores:
(1) dependência de ambos os receptores para cininas para a manutenção fisiológica da função
cardiovascular;
(2) a remoção do B1R ou B2R para cininas amplifica as ações do SRA e consequentemente a
atividade da Ang II;
(3) propomos que a ausência de B1R ou B2R e consequente potencialização do SRA são
fatores determinantes para o desenvolvimento do estresse oxidativo identificado em
cardiomiócitos dos grupos B1-/- e B2
-/-;
(4) com base nas evidências da literatura que apontam a participação das ROS e RNS em
modificações redox/nitroso em proteínas relacionadas com a homeostase do Ca2+ intracelular,
sugerimos que o estado de estresse oxidativo nos grupos B1-/- e B2
-/- causou o
comprometimento na função sistólica cardíaca através de oxidações e/ou nitrosilações que
repercutiram na redução da expressão da SERCA2a e aumento na expressão do NCX;

67
(5) com relação a resistência coronariana aumentada nos grupos B1-/- e B2
-/-, sugerimos que a
disfunção endotelial destas artérias seja a principal causa dessa alteração, confirmadas através
do comprometimento nos mecanismos vasodilatadores e vasoconstritores;
(6) apesar dos fortes indícios no comprometimento do eixo nNOS/NO demonstrado na
reatividade vascular, confirmamos que ambos receptores para cininas, B1 ou B2, estão
heterodimerizados com a nNOS e eNOS, portanto, a deleção dos receptores criam
perturbações na biodisponibilidade do NO intracelular e, consequentemente, alterações no
relaxamento vascular induzido pela ACh;
(7) adicionalmente, sugerimos que a deleção dos receptores modifica a atividade catalítica da
nNOS favorecendo o estado de desacoplamento, e consequentemente, uma exacerbada
produção de ROS, identificado em aorta de camundongos B1-/- e B2
-/-.

68
9. REFERENCIAL BIBLIOGRÁFICO
ABBAS, F.; CLAYTON, J.K.; MARSHALL, K.M.; SENIOR, J. Characterisation of kinin receptors on the human isolated umbilical artery. Journal of Endocrinology. 156(2): 389–394, 1998. ABDALLA, S.; ABDEL-BASET, A.; LOTHER, H.; EL MASSIERY, A.; QUITTERER, U. Mesangial AT1/B2 receptor heterodimers contribute to angiotensin II hyperresponsiveness in experimental hypertension. Journal of Molecular Neuroscience. 26(2): 185-92, 2005. ABDALLA, S.; LOTHER, H.; EL MASSIERY, A.; QUITTERER, U. Increased AT(1) receptor heterodimers in preeclampsia mediate enhanced angiotensin II responsiveness. Nature Medicine. 7(9): 1003-9, 2001. ADACHI, T.; WEISBROD, R. M.; PIMENTEL, D. R.; YING, J.; SHAROV, V. S.; SCHÖNEICH, C.; COHEN, R. A. S-glutathiolation by peroxynitrite activates SERCA during arterial relaxation by nitric oxide. Nature Medicine. 10: 1200–1207, 2004. AGO, T.; LIU, T.; ZHAI, P.; CHEN, W.; LI, H.; MOLKENTIN, J. D.; VATNER, S. F.; SADOSHIMA, J. A redox-dependent pathway for regulating class II HDACs and cardiac hypertrophy. Cell. 133: 978–993, 2008. ALDERTON, W. K.; COOPER, C. E. e KNOWLES, R. G. Nitric oxide synthases: structure, function and inhibition. Biochemical Journal. 357: 593 - 615, 2001. ALFIE, M. E.; SIGMON, D. H.; POMPOSIELLO, S. I.; CARRETERO, O. A. Effect of high salt intake in mutant mice lacking bradykinin-B2 receptors. Hypertension. 29(1): 483-7. 1997. ANDREW, P. J.; MAYER, B. Enzymatic function of nitric oxide synthases. Cardiovascular Research. 43: 521-31, 1999. ANNING, P. B.; PRENDERGAST, B. D.; MACCARTHY, P. A.; SHAH, A. M.; BUSS, D. C.; LEWIS, M. J. ATP is involved in myocardial and vascular effects of exogenous bradykinin in ejecting guinea pig heart. American Journal of Physiology - Heart and Circulatory Physiology. 277: 818–825, 1999. ARAÚJO, R. C. Geração e caracterização do camundongo nocaute para o receptor B1 das cininas. [Tese] Universidade Federal de São Paulo, 2000.
ARAÚJO, R. C.; KETTRITZ, R.; FICHTNER, I.; PAIVA, A. C. M.; PESQUERO, J. B.; BADER, M. Alttered nutrophil homeostasis in Kinin B1 receptor-deficient mice. Biological chemistry. 382: 91-5. 2001 AUGUSTO, O.; BONINI, M. G.; AMANSO, A. M.; LINARES, E.; SANTOS, C. C.; MENEZES, S. L. Nitrogen dioxide and carbonate radical anion: two emerging radicals in biology. Free Radical Biology & Medicine. 32: 841–859, 2002.

69
AUSTIN, K. E.; FAUSSNER, A.; ROBINSON, H. E.; CHAKRAVARTY, S.; KYLE, D. J.; BATHON, J. M.; PROUD, D. Stable expression of the human kinin B1 receptor in Chinese hamster ovary cells. The Journal of Biological Chemistry. 272: 11420–11425, 1997. BARNES, P. J. Bradykinin and asthma. Thorax. 47: 979–983, 1992. BAUMGARTEN, C. R.; LINZ, W.; KUNKEL, G.; SCHOLKENS, B. A.; WIEMER, G. Ramiprilat increases bradykinin outflow from isolated rat hearts. British Journal of Pharmacology. 108(2): 293–5, 1993. BAWOLAK, M. T.; TOUZIN, K.; MOREAU, M. E.; DÉSORMEAUX, A.; ADAM, A.; MARCEAU, F. Cardiovascular expression of inflammatory signaling molecules, the kinin B1 receptor and COX2, in the rabbit: effects of LPS, anti-inflammatory and antihypertensive drugs. Regulatory Peptides. 146(1-3): 157-68, 2008. BERTRAND, M. E. Provision of cardiovascular protection by ACE inhibitors: a review of recent trials. Current Medical Research & Opinion. 20(10): 1559-69, 2004. BHOOLA, K. D.; FIGUEROA, C. D.; WORTHY, K. Bioregulation of kinins: kallikreins, Kininogens. Pharmacological Reviews. 44: 1-79, 1992. BLAUKAT, A.; HERZER, K.; SCHROEDER, C.; BACHMANN, M.; NASH, N.; MÜLLER-ESTERL, W. Overexpression and functional characterization of kinin receptors reveal subtype-specific phosphorylation. Biochemistry. 38: 1300–1309, 1999. BOLOTINA, V. M.; NAJIBI, S.; PALACINO, J. J.; PAGANO, P. J.; COHEN, R. A. Nitric oxide directly activates calcium-dependent potassium channels in vascular smooth muscle. Nature. 368: 850–853, 1994. BORKOWSKI, J. A.; RANSOM, R. W.; SEABROOK, G. R.; TRUMBAUER, M.; CHEN, H.; HILL, R. G.; STRADER, C. D.; HESS, J. F. Targeted disruption of a B2 bradykinin receptor gene in mice eliminates bradykinin action in smooth muscle and neurons. The Journal of Biological Chemistry. 270 (23): 13706-10. 1995. BOUCHARD, J. F.; CHOUINARD, J.; LAMONTAGNE, D. Role of kinins in the endothelial protective effect of ischemic preconditioning. British Journal of Pharmacology. 123 (3):413– 20, 1998. BRENNAN, J. P.; BARDSWELL, S. C.; BURGOYNE, J. R.; FULLER, W.; SCHRÖDER, E.; WAIT, R.; BEGUM, S.; KENTISH, J. C.; EATON, P. Oxidant-induced activation of type I protein kinase A is mediated by RI subunit interprotein disulfide bond formation. The Journal of Biological Chemistry. 281: 21827–21836, 2006. BURGOYNE, J. R.; MADHANI, M.; CUELLO, F.; CHARLES, R. L.; BRENNAN, J. P.; SCHRÖDER, E.; BROWNING, D. D.; EATON, P. Cysteine redox sensor in PKGIa enables oxidant induced activation. Science. 317:1393–1397, 2007. BUSSE, R.; FLEMING, I. Regulation and functional consequences of endothelial nitric oxide formation. Annals of Medicine. 27: 331–340, 1995.

70
CAI, H.; HARRISON, D. G. Endothelial dysfunction in cardiovascular diseases: the role of oxidant stress. Circulation Research. 87: 840-4, 2000. CAMPBELL, D. J. Towards understanding the kallikrein-kinin system: insights from measurement of kinin peptides. Brazilian Journal of Medical and Biological Research. 33: 665-77, 2000. CAMPBELL, D. J.; KLADIS, A.; DUNCAN, A. M. Bradykinin peptides in kidney, blood and other tissues of the rat. Hypertension. 21(2): 155–65, 1993. CAPETTINI, L. S. A.; CORTES, S. F.; SILVA, J. F.; ALVAREZ-LEITE, J. I.; LEMOS, V. S. Decreased production of nNOS-derived hydrogen peroxide contributes to endothelial dysfunction in atherosclerosis. British Journal of Pharmacology. 164 (6): 1738–1748, 2011. CAPETTINI, L. S. A.; CORTES, S. F.; GOMES, M. A.; SILVA, G. A. B.; PESQUERO, J. L.; Lopes, M. J.; Teixeira, M. M.; LEMOS, V. S. Neuronal nitric oxide synthase-derived hydrogen peroxide is a major endothelium-dependent relaxing factor. American Journal of Physiology. Heart and Circulatory Physiology. 295: 2503-2511, 2008. CARRETERO, O. A.; SCICLI, A. G. Kinins paracrine hormone. Kidney International. 26 (Suppl): 552-9, 1988. CAVE, A. C.; BREWER, A. C.; NARAYANAPANICKER, A.; RAY, R.; GRIEVE, D. J.; WALKER, S.; SHAH, A. M. NADPH oxidases in cardiovascular health and disease. Antioxidants & Redox Signaling. 8:691–728, 2006. CERAVOLO, G. S.; FERNANDES, L.; MUNHOZ, C. D.; FERNANDES, D. C.; TOSTES, R. C. A.; LAURINDO, F. R. M.; SCAVONE, C.; FORTES, Z. B.; CARVALHO. M. H. C. Angiotensin II Chronic Infusion Induces B1 Receptor Expression in Aorta of Rats. Hypertension. 50:756-761, 2007. CHAHINE, R.; ADAM, A.; YAMAGUCHI, N.; GASPO, R.; REGOLI, D.; NADEAU, R. Protective effects of bradykinin on the ischemic heart: implication of the B1 receptor. British Journal of Pharmacology. 108: 318– 22, 1993. CHANDRAZEKAR, B.; FREEMAN, G. L. Induction of nuclear factor-κB and activating protein 1 in postischemic myocardium. FEBS Letters. 401(1): 30–4,1997. CLERK, A.; FULLER, S.; MICHAEL, A.; SUGDEN, P. H. Stimulation of stress regulated mitogen activated protein kinases (SAPKs/ JNKs and p38-MAPK) in perfused rat hearts by oxidative and other stress. The Journal of Biological Chemistry. 273(2): 7228–34, 1998. CZAPSKI, G.; GOLDSTEIN S. The role of the reactions of NO with superoxide and oxygen in biological systems: a kinetic approach. Free Radical Biology & Medicine. 19(6): 785-94,1995. DAI, D. F.; SANTANA, L. F.; VERMULST, M.; TOMAZELA, D. M.; EMOND, M. J.; MACCOSS, M. J.; GOLLAHON, K.; MARTIN, G. M.; LOEB, L. A.; LADIGES, W. C.; RABINOVITCH, P. S. Overexpression of catalase targeted to mitochondria attenuates murine cardiac aging. Circulation. 119: 2789–2797, 2009.

71
DRUMMOND, G. R.; COCKS, T. M. Endothelium-dependent relaxation to the B1 kinin receptor agonist des-Arg9-bradykinin in human coronary arteries. British Journal of Pharmacology. 116(8): 3083-3085, 1995. DUKA, A.; KINTSURASHVILI, E.; DUKA, I.; ONA, D.; HOPKINS, T.A.; BADER, M.; GAVRAS, I.; GAVRAS, H. Angiotensin-converting enzyme inhibition after experimental myocardial infarct: role of the kinin B1 and B2 receptors. Hypertension. 51(5):1352-7, 2008 DUKA, I.; KINTSURASHVILI, E.; GAVRAS, I.; JOHNS, C.; BRESNAHAN, M.; GAVRAS, H. Vasoactive potential of the B1 bradykinin receptor in normotension and hypertension. Circulation Research. 88(3), 275-281, 2001. DUNCAN, J. G.; RAVI, R.; STULL, L. B.; MURPHY, A. M. Chronic xanthine oxidase inhibition prevents myofibrillar protein oxidation and preserves cardiac function in a transgenic mouse model of cardiomyopathy. American Journal of Physiology - Heart and Circulatory Physiology. 289: 1512–1518, 2005. EL-DAHR, S. S.; DIPP. S.; BARICOS, W. H. Bradykinin stimulates the ERK–Elk-1–Fos/AP-1 pathway in mesangial cells. American Journal of Physiology - Renal Physiology. 275: 343–352, 1998. EMANUELI, C.; MAESTRI, R.; CORRADI, D.; MARCHIONE, R.; MINASI, A.; TOZZI, M. G.; SALIS, M.B.; STRAINO, S.; CAPOGROSSI, M. C.; OLIVETTI, G.; MADEDDU, P. Dilated and failing cardiomyopathy in bradykinin B(2) receptor knockout mice. Circulation.100: 2359–2365, 1999. ERIC, J.; BKAILY, G.; BKAILY, G. B.; VOLKOV, L.; GABRA, B. H.; SIROIS, P. Des-Arg9-bradykinin increases intracellular Ca2+ in bronchoalveolar eosinophils from ovalbumin-sensitized and -challenged mice. European Journal of Pharmacology.15 (475):129-37, 2003. ERTL, G.; KLONER, R. A.; ALEXANDER, R. W.; BRAUNWALD, E. Limitation of experimental infarct size by angiotensin converting enzyme inhibition. Circulation.65:40– 8, 1982. EWALD, D. A.; PANG, I. H.; STERNWEIS, P. C.; MILLER, R. J. Differential G protein mediated coupling of neurotransmitter receptors to Ca2+ channels in rat dorsal root ganglion neurons in vitro. Neuron. 2: 1185–1193, 1989. FASOLATO, C., PANDIELLA, A.; MELDOLESI, J.; POZZAN, T. Generation of inositol phosphates, cytosolic Ca2+ and ionic fluxes in PC12 cells treated with bradykinin. The Journal of Biological Chemistry. 263: 17350–17359, 1988. FAUSSNER, A.; PROUD, D.; TOWNS, M.; BATHON, J. M. Influence of the cytosolic carboxyl termini of human B1 and B2 kinin receptors on receptor sequestration, ligand internalization, and signal transduction. The Journal of Biological Chemistry. 273 (5): 2617-23, 1998.

72
FELIPE, S. A.; RODRIGUES, E. S.; MARTIN, R. P.; PAIVA, A. C. M.; PESQUERO, J. B.; SHIMUT. S. I. Functional expression of kinin B1 and B2 receptors in mouse abdominal aorta. Brazilian Journal of Medical and Biological Research. 40: 649-655, 2007. FERNANDES, L.; CERAVOLO, G. S.; FORTES, Z. B.; TOSTES, R.; SANTOS, R. A. S.; SANTOS, J. A.; MORI, M. A. S.; PESQUERO, J. B.; CARVALHO M. H. C. Modulation of Kinin B1 receptor expression by endogenous angiotensin II in hypertensive rats. Peptides. 136 (1-3): 92-7, 2006. FIELD, J. L.; BUTT, S. K.; MORTON, I. K. M.; HALL, J. M. Bradykinin B2 receptors and coupling mechanins in the smooth muscle of the guinea-pig taenia caeci. British Journal of Pharmacology.113: 607-13, 1994. FIGUEROA, C. D.; MARCHANT, A.; NOVOA. U.; FORSTERMANN, U.; JARNAGIN, K.; SCHOLKENS, B.; MULLER-ESTERL, W. Differential Distribution of Bradykinin B(2) Receptors in the Rat and Human Cardiovascular System. Hypertension. 37(1): 110-120, 2001. FLEMING, I.; FISSLHALTER, B.; BUSSE, R. Calcium signaling in endothelial cells involves activation of tyrosine kinases and leads to activation of mitogen-activate protein kinases. Circulation Research. 76: 522–529, 1995. FLEMING, I.; FISSLTHALER, B.; BUSSE, R. Interdependence of calcium signaling and protein tyrosine phosphorylation in human endothelial cells. The Journal of Biological Chemistry. 271: 11009–11015, 1996. FLEMING, I .; RUDI, B. Signal transduction of eNOS activation. Cardiovascular Research. 43:532–541, 1999. FÖRSTERMANN, U. Regulation of Nitric Oxide Synthase Expression and Activity. In: MAYER, B., (Ed.). Nitric Oxide. Springer-Verlag Berlim Heidelberg, 71-91, 2000. FÖRSTERMANN, U. e SESSA, W. C. Nitric oxide synthases: regulation and function. European Heart Journal. 33 (7): 829-837, 2012. FOUCART, S.; GRODIN, L.; COUTURE, R.; NADEU, R. Modulation of noradrenaline release by B1 and B2 kinin receptor during metabolic anoxia in the rat isolated atria. Canadian Journal of Physiology and Pharmacology. (75): 639-645, 1997. FURCHGOTT, R. F.; ZAWADZKI, J. V. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature. 288(5789): 373-376, 1980. GOBEIL, F.; FILTEAU, C.; PHENG, L. H.; JUKIC, D.; NGUYEN-LE, X. K.; REGOLI, D. In vitro and in vivo characterization of bradykinin B2 receptors in the rabbit and the guinea pig. Canadian Journal of Physiology and Pharmacology. 74: 137-144, 1996. GOBEIL, F.; PHENG, L.H.; BADINI, I.; NGUYEN-LE, X.K.; PIZARD, A.; RIZZI, A.; BLOUIN, D.; REGOLI, D. Receptors for kinins in the human isolated umbilical vein. British Journal of Pharmacology. 118(2): 289–294, 1996a.

73
GOHLA, A. S.; OFFERMANNS, S.; WILKIE, T. M.; SCHULTZ, G. Differential involvement of Galpha12 and Galpha13 in receptor-mediated stress fiber formation. The Journal of Biological Chemistry. 274: 17901–17907, 1999. GOLSER, R.; GORREN, A. C.; LEBER, A.; ANDREW, P.; HABISCH, H. J.; WERNER, E. R.; SCHMIDT, K.; VENEMA, R. C.; MAYER, B. Interaction of endothelial and neuronal nitricoxide synthases with the bradykinin B2 receptor. Binding of an inhibitory peptide to the oxygenase domain blocks uncoupled NADPH oxidation. The Journal of Biological Chemistry. 275: 5291–5296, 2000. GOTO, M.; LIU, Y.; YANG, X. M.; ARDELL, J. L.; COHEN, M. V.; DOWNEY, J. M. Role of bradykinin in protection of ischemic preconditioning in rabbit hearts. Circulation Research. 77: 611–621, 1995. GOUIN, L.; CARDINAL, R.; ADAM, A.; DRAPEU, G.; NADEU, R. Kinin-induced prolongation of action-potential duration in right ventricular muscle from rat: involvement of B1 and B2 receptors. Journal of Cardiovascular Pharmacology. 28 (2): 333-343, 1996. GRAY, M. O.; LONG, C. S.; KALINYAK, J. E.; LI, H. T.; KARLINER, J. S. Angiotensin II stimulates cardiac myocyte hypertrophy via paracrine release of TGF-beta 1 and endothelin-1 from fibroblasts. Cardiovascular Research. 40: 352–363, 1998. GREENE, E. L.; VELARDE, V.; JAFFA, A. A. Role of reactive oxygen species in bradykinin-induced mitogen-activated protein kinase and c-fos induction in vascular cells. Hypertension. 35: 942–947, 2000. GROHE, C.; KAHLERT, S.; LOBBERT, K.; STIMPEL, M.; KARAS, R.H.; VETTER, H.; NEYSES, L. Cardiac myocytes and fibroblasts contain functional estrogen receptors. FEBS Letters. 416:107–112, 1997. HALL, J. M. Bradykinin receptors: Pharmacological properties and biological roles. Pharmacology & Therapeutics. 56: 131-90, 1992. HARRIS, M. B.; JU, H.; VENEMA, V. J.; BLACKSTONE, M.; VENEMA, R. C. Role of heat shock protein 90 in bradykinin-stimulated endothelial nitric oxide release. General Pharmacology.35: 165–170, 2000. HUIE, R.E.; PADMAJA, S. The reaction of no with superoxide. Free Radical Research Communication. 1993;18(4):195-9. HELLER, R.; POLACK, T.; GRABNER, R.; TILL, U. Nitric oxide inhibits proliferation of human endothelial cells via a mechanism independent of cGMP. Atherosclerosis. 144: 49–57, 1999. HESS, J. F.; BORKOWSKI, J. A.; MACNEIL, T.; STONESIFER, G. Y.; FRAHER, J.; STRADER, C. D.; ROSASOM, R. W. Differential pharmacology of cloned human and mouse B2 bradykinin receptors. Molecular Pharmacology. 45: 1-8, 1994.

74
HESS, J. F.; BORKOWSKY, J. A.; YOUNG, G. S.; STRADER, C. D.; RANSOM, R. W. Cloning and pharmacological characterization of a human bradykinin (BK-2) receptor. Biochemical and Biophysical Research Communications. 184: 260-268, 1992. HIGUCHI, Y.; OTSU, K.; NISHIDA, K.; HIROTANI, S.; NAKAYAMA, H.; YAMAGUCHI, O.; HIKOSO, S.; KASHIWASE, K.; TAKEDA, T.; WATANABE, T.; MANO, T.; MATSUMURA, Y.; UENO, H.; HORI, M. The small GTP-binding protein Rac1 induces cardiac myocyte hypertrophy through the activation of apoptosis signal-regulating kinase 1 and nuclear factor-kappa B. The Journal of Biological Chemistry. 278: 20770–7, 2003. HIGUCHI, Y.; OTSU, K.; NISHIDA, K.; HIROTANI, S.; NAKAYAMA, H.; YAMAGUCHI, O.; MATSUMURA, Y.; UENO, H.; TADA, M.; HORI, M. Involvement of reactive oxygen species-mediated NF-κB activation in TNF-alpha-induced cardiomyocyte hypertrophy. Journal of Molecular and Cellular Cardiology. 34: 233–40, 2002. HOCK, F. J.; WIRTH, K.; ALBUS, U.; LINZ, W.; GERHARDS, H. J.; WIEMER, G.; HENKE, S.; BREIPOHL, G.; KÖNIG, W.; KNOLLE, J.; SCHÖLKENS, B. A. HOE 140 a new potent and long acting bradykinin antagonist: in vitro studies. British Journal of Pharmacology. 102: 769-773, 1991. HOOL, L. C.; WHALLEY, D. W.; DOOHAN, M. M.; RASMUSSEN, H. H. Angiotensin converting enzyme inhibition, intracellular Na+, and Na(+)-K+ pumping in cardiac myocytes. American Journal of Physiology - Heart and Circulatory Physiology. 268: 366–375, 1995. HORNIG, B.; KOHLER, C.; DREXLER, H. Role of bradykinin in mediating vascular effects of angiotensin-converting enzyme inhibitors in humans. Circulation. 95:1115– 8, 1997. IDE, T.; TSUTSUI, H.; HAYASHIDANI, S.; KANG, D.; SUEMATSU, N.; NAKAMURA, K.; UTSUMI, H.; HAMASAKI, N.; TAKESHITA, A. Mitochondrial DNA damage and dysfunction associated with oxidative stress in failing hearts after myocardial infarction. Circulation Research. 88: 529–535, 2001. IGNJATOVIC, T.; TAN, F.; BROVKOVYCH, V.; SKIDGEL, R. A.; ERDOS, E.G. Novel mode of action of angiotensin I converting enzyme inhibitors: direct activation of bradykinin B1 receptor. The Journal of Biological Chemistry. 277: 16847–52, 2002. IZUMIYA, Y.; KIM, S.; IZUMI, Y.; YOSHIDA, K.; YOSHIYAMA, M.; MATSUZAWA, A.; ICHIJO, H.; IWAO, H. Apoptosis signal-regulating kinase 1 plays a pivotal role in angiotensin II-induced cardiac hypertrophy and remodeling. Circulation Research.93:874–83, 2003. JONES, C.; PHILLIPS, E.; DAVIS, C.; ARBUCKLEY, J.; YAQOOB, M.; BURGESS, G.M.; DOCHERTY, R.J.; WEEBB, M.; BEVAN, S.J.; MCINTYRE, P. Molecular characterization of cloned bradykinin B1 receptors from rat and human. European Journal of Pharmacology. 374: 423-33, 1999. JU, H.; VENEMA, V. J.; LIANG, H.; HARRIS, M. B.; ZOU, R.; VENEMA, R. C. Bradykinin activates the Janus-activated kinase/signal transducers and activators of

75
transcription (JAK/STAT) pathway in vascular endothelial cells: localization of JAK/STAT signalling proteins in plasmalemmal caveolae. Biochemical Journal. 351: 257–264, 2000. JU, H.; VENEMA, V. J.; MARRERO, M. B.; VENEMA, R. C. Inhibitory interactions of the bradykinin B2 receptor with endothelial nitric-oxide synthase. The Journal of Biological Chemistry. 273: 24025–24029, 1998. KANG, D. S.; GUSTAFSSON, C.; MÄRGELIN, M.; LEEB-LUNDBERG, L. M. F. B1 bradykinin receptor homo-oligomers in receptor cell surface expression and signaling: effects of receptor fragments. Molecular Pharmacology. 67: 309–318, 2005. KASS, D. A.; TAKIMOTO, E.; NAGAYAMA, T.; CHAMPION, H. C. Phosphodiesterase regulation of nitric oxide signaling. Cardiovascular Research. 75: 303–314, 2007. KIM, N. N.; VILLEGAS, S.; SUMMEROUR, S. R; VILLARREAL, F. J. Regulation of cardiac fibroblast extracellular matrix production by bradykinin and nitric oxide. Journal of Molecular and Cellular Cardiology. 31: 457– 66, 1999. KUHR, F.; LOWRY, J.; ZHANG, Y.; BROVKOVYCH, V.; SKIDGEL, R.A. Differential regulation of inducible and endothelial nitric oxide synthase by kinin B1 and B2 receptors. Neuropeptides. 44: 145–154, 2010. KUSTER, G. M.; LANCEL, S.; ZHANG, J.; COMMUNAL, C.; TRUCILLO, M. P.; LIM, C. C.; PFISTER, O.; WEINBERG, E. O.; COHEN, R. A.; LIAO, R.; SIWIK, D. A.; COLUCCI, W. S. Redox mediated reciprocal regulation of SERCA and Na+–Ca2+ exchanger contributes to sarcoplasmic reticulum Ca2+ depletion in cardiac myocytes. Free Radical Biology & Medicine. 48: 1182–1187, 2010. KUSTER, G. M.; SIWIK, D. A.; PIMENTEL, D. R.; COLUCCI, W. S. Role of reversible, thioredoxin-sensitive oxidative protein modifications in cardiac myocytes. Antioxidants & Redox Signaling. 8: 2153–2159, 2006. LAGNEUX, C.; ADAM, A.; LAMONTAGNE, D. A study of the mediators involved in the protection induced by exogenous kinins in the isolated rat heart. International Immunopharmacology. 3 (10): 1511-1518, 2003. LAGNEUX, C.; INNOCENTI-FRANCILLARD, P.; GODIN-RIBUOT, D.; BADER, M.; RIBUOT, C. Heat stress-induced B1 receptor synthesis in the rat: an ex vivo study. British Journal of Pharmacology.125(4): 812– 6, 1998. LAGNEUX, C.; RIBUOT, C. In vivo evidence for B1-receptor synthesis by heat stress in the rat. Bristh Journal of Pharmacology 121:1045–1046, 1997. LARRIVÉE, J. F.; BACHVAROV, D. R.; HOULE, F.; LANDRY, J.; HUOT, .J.; MARCEAU, F. Role of the mitogen-activated protein kinases in the expression of the kinin B1 receptors induced by tissue injury. The Journal of Immunology. 160(3): 1419– 26, 1998. LASSEGUE, B.; CLEMPUS, R. E. Vascular NAD(P)H oxidases: specific features, expression, and regulation. American Journal of Physiology - Regulatory, Integrative and Comparative Physiology. 285(2): 277-97, 2003.

76
LAUTON-SANTOS, S. Participação do receptor B1 para cininas no controle da função cardíaca em camundongos. Tese de Doutorado, Universidade Federal de Minas Gerais, 2007. LAUTON-SANTOS, S.; GUATIMOSIM, S.; CASTRO, C. H. ; OLIVEIRA, F. A. ; ALMEIDA, A. P. ; DIAS-PEIXOTO, M. F.; GOMES, M. A. ; PESSOA, P. ; PESQUERO, J. L. ; PESQUERO, J. B. ; BADER, M. ; CRUZ, J. S. Kinin B1 receptor participates in the control of cardiac function in mice. Life Sciences. 81: 814-822, 2007. LEEB-LUNDBERG, L. M. F.; MARCEAU, F.; MÜLLER-ESTERL, W.; PETTIBORE, J.; ZURAW, B. L. International Union of Pharmacology. XLV. Classification of the Kinin Receptor Family: from Molecular Mechanisms to Pathophysiological Consequences. Pharmacological Reviews. 57: 27-77, 2005. LEVESQUE, L.; LARRIVEE, J.F.; BACHVAROV, D.R.; RIOUX, F.; DRAPEAU, G.; MARCEAU, F. Regulation of kinin-induced contraction and DNA synthesis by inflammatory cytokines in the smooth muscle of the rabbit aorta. British Journal of Pharmacology. 116(1): 1673-9, 1995. LI, J. M.; SHAH, A. M. Endothelial cell superoxide generation: regulation and relevance for cardiovascular pathophysiology. American Journal of Physiology - Regulatory, Integrative and Comparative Physiology. 287(5): 1014-30, 2004. LI, S. Y.; YANG, X.; CEYLAN-ISIK, A. F.; DU, M.; SREEJAYAN, N.; REN, J. Cardiac contractile dysfunction in Lep/Lep obesity is accompanied by NADPH oxidase activation, oxidative modification of sarco(endo)plasmic reticulum Ca2+-ATPase and myosin heavy chain isozyme switch. Diabetologia. 49: 1434–1446, 2006. LI, Y.; HUANG, T. T.; CARLSON, E. J.; MELOV, S.; URSELL, P. C.; OLSON, J. L.; NOBLE, L. J.; YOSHIMURA, M. P.; BERGER, C.; CHAN, P. H.; WALLACE, D. C.; EPSTEIN, C. J. Dilated cardiomyopathy and neonatal lethality in mutant mice lacking manganese superoxide dismutase. Nature Genetics. 11: 376–381, 1995. LIEBMANN, C.; GRANESS, A.; LUDWIG, B.; ADOMEIT, A.; BOEHMER, A.; BOEHMER, F. D.; NURNBERG, B.; WETZKER, R. Dual bradykinin B2 receptor signalling in A431 human epidermoid carcinoma cells: activation of protein kinase C is counteracted by a GS-mediated stimulation of the cyclic AMP pathway. Biochemical Journal. 313: 109–118, 1996. LINDER, M. E.; EWALD, D. A.; MILLER, R. J.; GILMAN, A. G. Purification and characterization of Go alpha and three types of Gi alpha after expression in Escherichia coli. The Journal of Biological Chemistry. 265: 8243–8251, 1990. LINZ, W.; WIEMER, G.; GOHLKE, P.; UNGER, T.; SCHÖLKENS, B. A. Contribution of kinins to the cardiovascular actions of angiotensin-converting enzyme inhibitors. Pharmacological Reviews. 47 (1): 25-49, 1995. LINZ, W.; WIEMER, G.; SCHOLKENS, B.A. Role of kinins in the pathophysiology of myocardial ischemia. Diabetes. 45(1): 51– 7, 1996.

77
LIU, Y. H.; YANG, X. P.; SHAROV, V. G.; Nass, O.; Sabbah, H.N.; Peterson, E.; Carretero, O. A. Effects of angiotensin-converting enzyme inhibitors and angiotensin II type 1 receptor antagonists in rats with heart failure. Role of kinins and angiotensin II type 2 receptors. The Journal of Clinical Investigation. 99:1926–35, 1997. LOCHNER, A.; MARAIS, E.; GENADE, S.; MOOLMAN, J. A. Nitric oxide: a trigger for classic preconditioning?. American Journal of Physiology - Heart and Circulatory Physiology. 279(6): 2752-65, 2000. LOIOLA, R. A.; REIS, F. C. G.; KAWAMOTO, E. L.; SCAVONE, C., ABDALLA, D. S.; FERNANDES, L.; PESQUERO, J. B. Role of vascular Kinin B1 and B2 receptors in endothelial nitric oxide metabolism. Peptides. 32: 1700–1705, 2011. LOPES, P.; COUTURE, R. Cardiovascular responses elicited by intrathecal kinins in the conscious rat. European Journal of Pharmacology. 210: 137–147, 1992. MADEDDU, P.; EMANUELI, C.; MAESTRI, R.; SALIS, M. B.; MINASI, A.; CAPOGROSSI, M. C.; OLIVETTI, G. Angiotensin II type 1 receptor blockade prevents cardiac remodeling in bradykinin B2 receptor knockout mice. Hypertension. 35: 391–396, 2000. MARCEAU, F.; FRED HESS, J.; BACHVAROV, D.R. The B1 receptors for Kinins. Pharmacological Reviews. 50: 357-386, 1998. MARIN-CASTANO, M. E.; SCHANSTRA, J. P.; NEAU, E.; PRADDAUDE, F.; PECHER, C.; ADER, J.L.; GIROLAMI, J.P.; BASCANDS, J.L. Induction of functional bradykinin B(1)-receptors in normotensive rats and mice under chronic angiotensin-converting enzyme inhibitor treatment. Circulation. 105: 627– 32, 2002. MARKOU, T.; HADZOPOULOU-CLADARAS, M.; LAZOU, A. Phenylephrine induces activation of CREB in adult rat cardiac myocytes through MSK1 and PKA signaling pathways. Journal of Molecular and Cellular Cardiology. 37:1001–11, 2004. MARTORANA, P.A.; KETTENBACH, B.; BREIPOHL, G.; LINZ, W.; SCHÖLKENS, B.A. Reduction of infarct size by local angiotensin-converting enzyme inhibition is abolished by a bradykinin antagonist. European Journal Pharmacology.182(2): 395– 6, 1990. MASSION, P. B.; BALLIGAND, J. L. Relevance of nitric oxide for myocardial remodeling. Current Heart Failure Reports. 4:18–25, 2007. MASSOUDY, P.; BECKER, B. F.; GERLACH, E. Nitric oxide accounts for postischemic cardioprotection resulting from angiotensin-converting enzyme inhibition: indirect evidence for a radical scavenger effect in isolated guinea pig heart. Journal of Cardiovascular Pharmacology. 25: 440–447, 1995. MATHIS, S. A.; LEEB-LUNDBERG, L. M. F. Bradykinin recognizes different molecular forms of the B2 kinin receptor in the presence and absence of guanine nucleotides. Biochemical Journal. 276: 141–147,1991.

78
McEACHERN, A. E.; SHELTON, E. R.; BHAKTA, S.; OBERNOLTE, R.; BACH, C.; ZUPPAN, P.; FUJISAKI, J.; ALDERICH, R. W.; JARNAGIN, K. Expression cloning of rat B2 bradykinin receptor. Proceedings of the National Academy of Sciences. 88: 7724-7728, 1991. McLEAN, P. G.; PERRETTI, M.; AHLUWALIA, A. Inducible expression of kinin B1 receptor in the endotoxemic heart: mechanisms of des-Arg9bradykinin-induced coronary vasodilatation. British Journal of Pharmacology. 128 (2): 275-282, 1999. MENETON, P.; BLOCH-FAURE, M.; HAGEGE, A. A.; RUETTEN, H.; HUANG, W.; BERGAYA, S.; CEILER, D.; GEHRING, D.; MARTINS, I. I.; SALMON, G.; BOULANGER, C. M.; NUSSBERGER, J.; CROZATIER, B.; GASC, J. M.; HEUDES, D.; BRUNEVAL, P.; DOETSCHMAN, T.; MENARD, J.; ALHENC-GELAS, F. Cardiovascular abnormalities with normal blood pressure in tissue kallikrein–deficient mice. Proceedings of the National Academy of Sciences (USA). 98: 2634–2639, 2001. MENKE, J. G.; BORKOWSKI, J. A.; BIERILO, K. K.; MACNEIL, T.; DERRICK, A. W.; SCHNEC, K. A.; RANSOM, R. W.; STRADER, C. D.; LINEMEYYER, D. L.; HESS, F. J. Expression cloning of a human B1 Bradykinin receptor. The Journal of Biological Chemistry. 269: 21583-6, 1994. MITRA, R.; MORAD, M. A uniform enzymatic method for dissociation of myocytes from hearts and stomachs of vertebrates. American Journal ofPhysiology - Heart and Circulatory Physiology.249: 1056-1060, 1985. MOMBOULI, J. V.; VANHOUTTE, P. M. Kinins and endothelial control of vascular smooth muscle. Annual Review of Pharmacology and Toxicology. 35: 679– 705, 1995. MONCADA, S.; HIGGS, A. The L-arginine-nitric oxide pathway. The New England Journal of Medicine. 329: 2002-2012, 1993. MOREAU, M. E.; GARBACKI, N.; MOLINARO, G.; BROWN, N. J.; MARCEAU, F.; ADAM, A. The kallikrein- kinin system: current and future pharmacological targets. Journal of Pharmacological Sciences. 99: 6–38, 2005. MORI, M. A.; ARAUJO, R. C.; REIS, F. C.; SGAI, D. G.; FONSECA, R. G.; BARROS, C. C.; MERINO, V. F.; PASSADORE, M.; BARBOSA, A. M.; FERRARI, B.; CARAYON, P.; CASTRO, C. H.; SHIMUTA, S. I.; LUZ, J.; BASCANDS, J. L.; SCHANSTRA, J. P.; EVEN, P. C.; OLIVEIRA, S. M.; BADER, M.; PESQUERO, J. B. Kinin B1 receptor deficiency leads to leptin hypersensitivity and resistance to obesity. Diabetes, 10. 2008. NANGLE, M.R.; COTTER, M.A.; CAMERON N. E. An in vitro investigation of aorta and corpus cavernosum from eNOS and nNOS gene-deficient mice. Pflügers Archiv - European Journal of Physiology. 448: 139-145, 2004 NI, A.; CHAI, K. X.; CHAO, L.; CHAO, J. Molecular cloning and expression of rat bradykinin B1 receptors from rat and human. European Journal of Pharmacology. 374: 423-33, 1999.

79
NI, A.; CHAO, L.; CHAO, J. Transcription factor nuclear factor κB regulates the inducible expression of the human B1 receptor gene in inflammation. The Journal of Biological Chemistry. 273 (5): 2784-2791, 1998. NOLLY, H.; CARBINI, L. A.; SCICLI, G.; CARRETERO, O. A.; SCICLI, A. G. A local kallikrein kinin system is present in rat heart. Hypertension. 23(6): 919–23, 1994. NUSSBERGER, J.; CUGNO, M.; CICARDI, M.; AGOSTONI, A. Local bradykinin generation in hereditary angioedema. Journal of Allergy and Clinical Immunology. 104:1321–1322, 1999. OLDENBURG, O.; COHEN, M. V.; YELLON, D. M.; DOWNEY, J. M. Mitochondrial KATP channels: role in cardioprotection. Cardiovascular Research. 55: 429–437, 2002. OLDENBURG, O.; QIN, Q.;KRIEG, T.; YANG, X.; PHILIPP, S.; CRITZ, S. D.; COHEN, M. V.; DOWNEY, J. M. Bradykinin induces mitochondrial ROS generation via NO, cGMP, PKG, and mitoKATP channel opening and leads to cardioprotection. American Journal of Physiology - Heart and Circulatory Physiology. 286: 468–476, 2004. OLIVEIRA, F. A.; GUATIMOSIM, S.; CASTRO, C. H.; GALAN, D. T.; LAUTON-SANTOS, S.; RIBEIRO, A. M.; ALMEIDA, A. P.; CRUZ, J. S. Abolition of reperfusion-induced arrhythmias in hearts from thiamine-deficient rats. American Journal of Physiology. Heart and Circulatory Physiology. 293: 394-401, 2007. PACHER, P.; BECKMAN, J. S.; LIAUDET, L. Nitric oxide and peroxynitrite in health and disease. Physiological Reviews. 87: 315–324, 2007. PAN, H. L.; CHEN, S. R.; SCICLI, G. M.; CARRETERO, O. A. Cardiac interstitial bradykinin release during ischemia is enhanced by ischemic preconditioning American Journal of Physiology - Heart and Circulatory Physiology. 279: 116–121, 2000. PAN, Z. K.; YE, R. D., CHRISTIANSEN, S. C.; JAGELS, M. A.; BOKOCH, G. M.; ZURAW, B. L. Role of the Rho GTPase in bradykinin-stimulated nuclear factor-κB activation and IL-1β gene expression in cultured human epithelial cells. The Journal of Immunology. 160: 3038– 3045, 1998. PAQUET, J. L.; LUCCARINI, J. M.; FOUCHET, C.; DEFRÊNE, E.; LOILLIER, B.; ROBERT, C.; BÉLICHARD, P.; CREMERS, B.; PRUNEAU, D. Pharmacological characterization of the bradykinin B2 receptor: inter-species variability and dissociation between binding and functional responses. British Journal of Pharmacology. 126: 1083-1090, 1999. PESQUERO, J.B.; ARAÚJO, R.C.; HEPPNTALL, P.A.; TUCKY, C.L.; SILVA, J.A.; WALTHER, T.; OLIVEIRA, S. M.; PESQUERO, J.L.; PAIVA, A.C.M.; CALIXTO, J.B.; LEWIN, G.R.; BADER, M. Hypoalgesia and altered inflammatory responses in mice lacking kinin B1 receptors. Proceedings of the National Academy of Sciences. 97: 8140-5, 2000. PESQUERO, J. B.; PESQUERO, J. L.; OLIVEIRA, S. M.; ROSCHER, A. A.; METZGER, R.; GANTEN, D.; BADER M. Molecular Cloning and functional characterization of a mouse

80
bradykinin B1 gene. Biochemical and Biophysical Research Communications. 220: 219-25. 1996. PINTO, Y. M.; BADER, M.; PESQUERO, J. B.; TSCHÖPE, C.; SCHOLTENS, E.; van GILST, W. H.; BUIKEMA, H. Increased kallikrein expression protects against cardiac ischemia. The FASEB Journal. 14(13): 1861– 3, 2000. POLOSA, R. Role of the kallikrein pathway in allergic diseases. Allergy. 48: 217–225, 1993. POU, S.; POU, W. S.; BREDT, D. S.; SNYDER, S. H.; ROSEN, G. M. Mechanism of Superoxide Generation by Neuronal Nitric-oxide Synthase. The Journal of Biological Chemistry. 267: 24173–24176, 1992. PRUNEAU, D.; BELICHARD, P. Induction of bradykinin B1 receptor-mediated relaxation in the isolated rabbit carotid artery. European Journal of Pharmacology. 239(1-3): 63–67, 1993. PRUNEAU, D.; LUCCARINI, J. M.; DEFRENE, E.; PAQUET, J. L.; BELICHARD, P. Characterisation of bradykinin receptors from juvenile pig coronary artery. European Journal of Pharmacology. 297(1-2): 53–60, 1996. PURCELL, N. H.; TANG, G.; YU, C.; MERCURIO, F.; DIDONATO, J. A.; LIN, A. Activation of NF-κB is required for hypertrophic growth of primary rat neonatal ventricular cardiomyocytes. Proceedings of the National Academy of Sciences (USA). 98: 6668–73, 2001. RAKHIT, R. D.; EDWARDS, R. J.; MOCKRIDGE, J. W.; BAYDOUN, A. R.; WYATT, A. W.; MANN, G. E.; MARBER, M. S. Nitric oxide-induced cardioprotection in cultured rat ventricular myocytes. American Journal of Physiology - Heart and Circulatory Physiology. 278: 1211–1217, 2000. REGOLI, D.; ALLOGHO, S. N.; RIZZI, A.; GOBEIL, F. J. Bradykinin receptors and their antagonists. European Journal of Pharmacology. 348: 1–10.1998. REGOLI, D.; BARABÉ, J. Pharmacology of bradykinin and related kinins. Pharmacological Reviews. 32: 1–46, 1980. REGOLI, D.; BARABÉ, J.; PARK, W. K. Receptors for bradykinin in rabbit aortae. Canadian Journal of Physiology and Pharmacology. 55:855–867, 1977. REGOLI, D.; MARCEAU, F.; BARABE, J. De novo formation of vascular receptors for bradykinin. Canadian Journal of Physiology and Pharmacology.56(4): 674–677, 1978. RHALEB, N. E.; GOBEIL, F.; REGOLI, D. Non-selectivity of new bradykinin antagonists for B1 receptors. Life Science. 51: 125–129, 1992. RIZZI, A.; CALO, G.; AMADESI, S.; REGOLI, D. Kinin B1 and B2 receptors in pig vessels: characterization of two monoreceptor systems. Naunyn-Schmiedeberg's Archives of Pharmacology. 356(5): 662–670, 1997.

81
ROMAN-CAMPOS, D.; DUARTE, H. L.; GOMES, E. R. ; CASTRO, C. H.; GUATIMOSIM, S.; NATALI, A. J.; ALMEIDA, A. P.; PESQUERO, J. B.; PESQUERO, J. L.; CRUZ, J. S. Investigation of the cardiomyocyte dysfunction in bradykinin type 2 receptor knockout mice. Life Sciences. 87: 715-723, 2010. ROSENKRANZ, A. C.; DUSTING, G. J.; RITCHIE, R. H. Endothelial dysfunction limits the antihypertrophic action of bradykinin in rat cardiomyocytes. Journal of Molecular and Cellular Cardiology. 32: 1119–1126, 2000. ROSENKRANZ, A. C.; HOOD, S. G.; WOODS, R. L.; DUSTING , G. J.; RITCHIE, R. H. Acute Antihypertrophic Actions of Bradykinin in the Rat Heart: Importance of Cyclic GMP. Hypertension. 40: 498-503, 2002. SANTOS, C. X.; ANILKUMAR, N.; ZHANG, M.; BREWER, A.C.; SHAH, A.M. Redox signaling in cardiac myocytes. Free Radical Biology & Medicine. 50(7): 777-93, 2011. SASAKI, N.; SATO, T.; OHLER, A.; O'ROURKE, B.; MARBAN, E. Activation of mitochondrial ATP-dependent potassium channels by nitric oxide. Circulation. 101: 439-45, 2000. SATOH, M.; FUJIMOTO, S.; HARUNA, Y.; ARAKAWA, S.; HORIKE, H.; KOMAI, N.; SASAKI, T.; TSUJIOKA, K.; MAKINO, H.; KASHIHARA, N. NAD(P)H oxidase and uncouple nitric oxide synthase are major sources of glomerular superoxide in rats with experimental diabetic nephropathy. American Journal of Physiology: Renal Physiology. 288: 1144-1152, 2005. SCHANSTRA, J. P.; BATAILLÉ, E.; MARIN-CASTAÑO, M. E.; BARASCUD, Y.; HIRTZ, C.; PESQUERO, J. B.; PECHER, C.; GAUTHIER, F.; GIROLAMI, J.; BASCANDS, J. The B1-agonist [des-Arg10]-kallidin activates transcription factor NF-κB and induces homologous upregulation of the bradykinin B1-receptor in cultured human lung fibroblasts. The Journal of Clinical Investigation. 101 (10): 2080-2091, 1998. SCHINI, V. B.; BOULANGER, C.; REGOLI, D.; VANHOUTTE, P. M. Bradykinin stimulates the production of cyclic GMP via activation of B2 kinin receptors in cultured porcine aortic endothelial cells. Journal of Pharmacology and Experimental Therapeutics. 252: 581–585, 1990. SCHNECK, K. A.; HESS, J. F.; STONESIFER, G. Y.; RANSOM, R. W. Bradykinin B1 receptors in rabbit aorta smooth muscle cells in culture. European Journal of Pharmacology. 266 (3): 277-82, 1994. SCHÖNEICH, C.; SHAROV, V. S. Mass spectrometry of protein modifications by reactive oxygen and nitrogen species. Free Radical Biology & Medicine. 41: 1507–1520, 2006. SCHRINER, S. E.; LINFORD, N. J.; MARTIN, G. M.; TREUTING, P.; OGBURN, C. E.; EMOND, M.; COSKUN, P. E.; LADIGES, W.; WOLF, N.; VAN REMMEN, H.; WALLACE, D. C.; RABINOVITCH, P. S. Extension of murine life span by overexpression of catalase targeted to mitochondria. Science. 308: 1909–1911, 2005.

82
SEDDON, M.; SHAH, A. M.; CASADEI, B. Cardiomyocytes as effectors of nitric oxide signalling. Cardiovascular Research. 75:315–326, 2007. SHAROV, V. S.; DREMINA, E. S.; GALEVA, N. A.; WILLIAMS, T. D.; SCHÖNEICH, C. Quantitative mapping of oxidation-sensitive cysteine residues in SERCA in vivo and in vitro by HPLC-electrospray-tandem MS: selective protein oxidation during biological aging. Biochemical Journal. 394: 605–615, 2006. SHIOMI, T.; TSUTSUI, H.; MATSUSAKA, H.; MURAKAMI, K.; HAYASHIDANI, S.; IKEUCHI, M.; WEN, J.; KUBOTA, T.; UTSUMI, H.; TAKESHITA, A. Overexpression of glutathione peroxidase prevents left ventricular remodeling and failure after myocardial infarction in mice. Circulation. 109: 544–549, 2004. SILVA, J. A.; ARAUJO, R. C.; BALTATU, O.; OLIVEIRA, S.M.; TSCHOPE, C.; FINK, E.; HOFFMANN, S.; PLEHM, R. CHAI, K. X.; CHAO, L.; CHAO, J.; GANTEN, D.; PESQUERO, J. B.; BADER, M. Reduced cardiac hypertrophy and altered blood pressure control in transgenic rats with the human tissue kallikrein gene. The FASEB Journal. 14: 1858–1860, 2000. SILVESTRE, J. S.; BERGAYA, S.; TAMARAT, R.; DURIEZ, M.; BOULANGER, C. M.; LEVY, B. I. Proangiogenic effect of angiotensin-converting enzyme inhibition is mediated by the bradykinin B(2) receptor pathway. Circulation Research. 89:678– 83, 2001. SKIDGEL, R. A.; ERDÖS, E.G. Cellular carboxypeptidases. Immunological Reviews. 161: 129–141, 1998. STERANKA, L. R.; BURCH, R. M. Bradykinin antagonists in pain inflamation. Em: Bradykinin Antagonists: Bacic and Clinical Research. New York. Burch R M, Marcel Dekker Inc. 191-211, 1991. STUMPE, T.; DECKING, U. K.; SCHRADER, J. Nitric oxide reduces energy supply by direct action on the respiratory chain in isolated cardiomyocytes. American Journal of Physiology - Heart and Circulatory Physiology. 280: 2350–2356, 2001. SU, J.B.; HOUEL, R.; HELOIRE, F.; Barbe, F.; Beverelli, F.; Sambin, L.; Castaigne, A.; Berdeaux, A.; Crozatier, B.; Hittinger, L. Stimulation of bradykinin B(1) receptors induces vasodilation in conductance and resistance coronary vessels in conscious dogs: comparison with B(2) receptor stimulation. Circulation. 101: 1848– 53, 2000. SUHASINI, M.; LI, H.; LOHMANN, S. M.; BOSS, G. R.; PILZ, R. B. Cyclic-GMPdependent protein kinase inhibits the Ras/Mitogen-activated protein kinase pathway. Molecular and Cellular Biology. 18: 6983–6994, 1998. SUZUKI, Y.; RUIZ-ORTEGA, M.; LORENZO, O.; RUPEREZ, M.; ESTEBAN, V.; EGIDO, J. Inflammation and angiotensin II. The International Journal of Biochemistry & Cell Biology. 35(6): 881-900, 2003. TIO, R.; TOBÉ, T. J.; BEL, K. J.; LANGEN, C. D.; van GILST, W. H.; WESSELING, H. Beneficial effects of bradykinin on porcine ischemic myocardium. Basic Research in Cardiology. 86(2) :107–16, 1991.

83
TOCCHETTI, C. G.; WANG, W.; FROEHLICH, J. P.; HUKE, S.; AON, M. A.; WILSON, G. M.; DI BENEDETTO, G.; O'ROURKE, B.; GAO, W. D.; WINK, D. A.; TOSCANO, J. P.; ZACCOLO, M.; BERS, D. M.; VALDIVIA, H. H.; CHENG, H.; KASS, D. A.; PAOLOCCI, N. Nitroxyl improves cellular heart function by directly enhancing cardiac sarcoplasmic reticulum Ca2+ cycling. Circulation Research. 100: 96–104, 2007. TODA, M.; BIAN, K.; AKIBA, T.; OKAMURA, T. Heterogeneity in mechanisms of bradykinin action in canine isolated blood vessels. European Journal of Pharmacology. 135: 321–329, 1987. TOUSIGNANT, C.; DION, S.; DRAPEAU, G.; REGOLI, D. Characterization of pre- and post-junctional receptors for kinins and neurokinins in the rat vas deferens. Neuropeptides. 9: 333–343. 1987. TOUYZ, R. M.; TABET, F.; SCHIFFRIN, E. L. Redox-dependent signalling by angiotensin II and vascular remodelling in hypertension. Clinical and Experimental Pharmacology and Physiology. 30(11): 860-6, 2003. TRABOLD, F.; PONS, S.; HAGEGE, A. A.; BLOCH-FAURE, M.; ALHENC-GELAS, F.; GIUDICELLI, J. F.; RICHER-GIUDICELLI, C.; MENETON, P. Cardiovascular phenotypes of kinin B2 receptor- and tissue kallikrein-deficient mice. Hypertension. 40: 90–95, 2002. TRACEY, K. J. The inflammatory reflex. Nature. 420(6917): 853-9, 2002. TSCHÖPE, C.; HERINGER-WALTHER, S.; KOCH, M.; SPILLMANN, F.; WENDORF, M.; LEITNER, E.; SCHULTHEISS, H. P.; WALTHER, T. Upregulation of bradykinin B1-receptor expression after myocardial infarction. British Journal of Pharmacology. 129(8): 1537– 8, 2000. TSCHÖPE, C.; HERINGER-WALTHER, S.; KOCH, M.; SPILLMANN, F.; WENDORF, M.; HAUKE, D.; BADER, M.; SCHULTHEISS, H. P.; WALTHER, T. Myocardial bradykinin B2-receptor expression at different time points after induction of myocardial infarction. Journal of Hypertension. 18(2): 223–8, 2000a. TSCHÖPE, C.; HERINGER-WALTHER, S.; WALTHER, T. Regulation of the kinin receptors after induction of myocardial infarction: a mini-review. Brazilian Journal of Medical and Biological Research. 33 (6): 701-8, 2000b. TSCHÖPE, C.; SPILLMANN, F.; ALTMANN, C.; KOCH, M.; WESTERMANN, D.; DHAYAT, N.; DHAYAT, S.; BASCANDS, J.; GERA, L.; HOFFMANN, S.; SCHULTHEISS, H.; WALTHER, T. The bradykinin B1 receptor contributes to the cardioprotective effects of AT1 blockade after experimental myocardial infarction. Cardiovascular Research. 61: 559– 569, 2004. TSUTSUI, M.; ONOUE, H.; IIDA, Y.; SMITH, L.; O’BRIEN, T.; KATUSIC Z.S. B(1) and B(2) bradykinin receptors on adventitial fibroblasts of cerebral arteries are coupled to recombinant eNOS. American Journal of Physiology - Heart and Circulatory. 278(2): 367-72, 2000.

84
TZIOMALOS, K.; HARE, J. M. Role of xanthine oxidoreductase in cardiac nitroso redox imbalance. Frontiers in Bioscience. 14: 237–262, 2009. VASQUEZ-VIVAR, J.; KALYANARAMAN, B.; MARTASEK, P.; HOGG, N.; MASTERS, B.S.; KAROUI, H.; TORDO, P.; PRITCHARD, K. A. Superoxide generation by endothelial nitric oxide synthase: the influence of cofactors. Proceedings of the National Academy of Sciences (USA). 95(16): 9220-5, 1998. VEGH, A.; SZEKERES, L.; PARRATT, J. R. Local intracoronary infusions of bradykinin profoundly reduce the severity of ischaemia-induced arrhythmias in anaesthetized dogs. British Journal of Pharmacology.104: 294-5, 1991 VENEMA, R. C. Post-translational mechanisms of endothelial nitric oxide synthase regulation by bradykinin. International Immunopharmacology. 2: 1755–1762, 2002. WALL, T. M.; SHEEHY, R.; HARTMAN, J. C. Role of bradykinin in myocardial preconditioning. Journal of Pharmacology and Experimental Therapeutics 270: 681–689, 1994. WEERD, W.F.; LEEB-LUNDBERG, L.M. Bradykinin sequesters B2 bradykinin receptors and the receptor-coupled Ga subunits Gaq and Gai in caveolae in DDT1 MF-2 smooth muscle cells. The Journal of Biological Chemistry. 272:17858–17866, 1997 WENZEL, S.; TAIMOR, G.; PIPER, H.M. SCHLUTER, K.D. Redox-sensitive intermediates mediate angiotensin II-induced p38 MAP kinase activation, AP-1 binding activity, and TGF-beta expression in adult ventricular cardiomyocytes. The FASEB Journal. 15:2291–3, 2001 WINTERBOURN, C. C. Reconciling the chemistry and biology of reactive oxygen species. Nature Chemical Biology. 4: 278–286, 2008. WITHEROW, F. N.; HELMY, A.; WEBB, D. J.; FOX, K. A.; NEWBY, D. E. Bradykinin contributes to the vasodilator effects of chronic angiotensin-converting enzyme inhibition in patients with heart failure. Circulation. 30(104): 2177– 81, 2001. XIE, P.; BROWNING, D. D.; HAY, N.; MACKMAN, N.; YE, R. D. Activation of NF-κB by bradykinin through a Gαq- and Gβγ-dependent pathway that involves phosphoinositide 3-kinase and Akt. The Journal of Biological Chemistry. 275: 24907–24914, 2000. XU, J.; CARRETERO, O. A.; SHESELY, E. G.; RHALEB, N.; YANG, J. J.; BADER, M.; YANG, X. The kinin B1 receptor contributes to the cardioprotective effect of angiotensin-converting enzyme inhibitors and angiotensin receptor blockers in mice.Experimental Physiology. 94(3): 322-9, 2009. XU, L.; EU, J. P.; MEISSNER, G.; STAMLER, J. S. Activation of the cardiac calcium release channel (ryanodine receptor) by poly-S-nitrosylation. Science. 279: 234–237; 1998. XU, S.; YING, J.; JIANG, B.; GUO, W.; ADACHI, T.; SHAROV, V.; LAZAR, H.; MENZOIAN, J.; KNYUSHKO, T. V.; BIGELOW, D.; SCHÖNEICH, C.; COHEN, R. A. Detection of sequence specific tyrosine nitration of manganese SOD and SERCA in

85
cardiovascular disease and aging. American Journal of Physiology - Heart and Circulatory Physiology. 290: 2220–2227, 2006. YANO, K.; HIGASHIDA, H.; HATTORI, H.; NOZAWA, Y. Bradykinin-induced transient accumulation of inositol trisphosphate in neuron-like cell line NG108–15 cells. FEBS Letters. 181: 403–406,1985. ZHU, Y. M.; BRADBURY, D. A.; PANG, L.; KNOX, A. J. Transcriptional regulation of interleukin (IL)-8 by bradykinin in human airway smooth muscle cells involves prostanoid-dependent activation of AP-1 and nuclear factor (NF)-IL-6 and prostanoid- independent activation of NF-κB. The Journal of Biological Chemistry. 278: 29366–29375, 2003.


















