UNIVERSIDADE FEDERAL DO PARÁ PRÓ-REITORIA DE … · localidades da costa norte do Brasil: Santana...
15
UNIVERSIDADE FEDERAL DO PARÁ PRÓ-REITORIA DE PESQUISA E PÓS-GRADUAÇÃO DIRETORIA DE PESQUISA PROGRAMA INSTITUCIONAL DE BOLSAS DE INICIAÇÃO CIENTÍFICA RELATÓRIO TÉCNICO - CIENTÍFICO Período : Agosto/ 2014 a Julho/ 2015 ( ) PARCIAL ( X ) FINA IDENTIFICAÇÃO DO PROJETO Título do Projeto de Pesquisa (ao qual está vinculado o Plano de Trabalho): FILOGENIA MOLECULAR E CONECTIVIDADE GENÉTICA DE POPULAÇÕES DE PEIXES MARINHOS: UM ESTUDO NO ATLÂNTICO OCIDENTAL Nome do Orientador: MARIA IRACILDA DA CUNHA SAMPAIO Titulação do Orientador: DOUTORADO Faculdade: CIÊNCIAS BIOLÓGICAS Unidade: BRAGANÇA Laboratório: LABORATÓRIO DE GENÉTICA E BIOLOGIA MOLECULAR Título do Plano de Trabalho: DNA BARCODE DAS ESPÉCIES DE ELEOTRIDAE (TELEOSTEI: GOBIIFORMES) DA COSTA BRASILEIRA Nome do Bolsista: ÉRIKA MILA FERREIRA FERNANDES
Transcript of UNIVERSIDADE FEDERAL DO PARÁ PRÓ-REITORIA DE … · localidades da costa norte do Brasil: Santana...

UNIVERSIDADE FEDERAL DO PARÁ
PRÓ-REITORIA DE PESQUISA E PÓS-GRADUAÇÃO
DIRETORIA DE PESQUISA
PROGRAMA INSTITUCIONAL DE BOLSAS DE INICIAÇÃO CIENTÍFICA
RELATÓRIO TÉCNICO - CIENTÍFICO
Período : Agosto/ 2014 a Julho/ 2015
( ) PARCIAL
( X ) FINA
IDENTIFICAÇÃO DO PROJETO
Título do Projeto de Pesquisa (ao qual está vinculado o Plano de Trabalho):
FILOGENIA MOLECULAR E CONECTIVIDADE GENÉTICA DE POPULAÇÕES
DE PEIXES MARINHOS: UM ESTUDO NO ATLÂNTICO OCIDENTAL
Nome do Orientador: MARIA IRACILDA DA CUNHA SAMPAIO
Titulação do Orientador: DOUTORADO
Faculdade: CIÊNCIAS BIOLÓGICAS
Unidade: BRAGANÇA
Laboratório: LABORATÓRIO DE GENÉTICA E BIOLOGIA MOLECULAR
Título do Plano de Trabalho: DNA BARCODE DAS ESPÉCIES DE ELEOTRIDAE
(TELEOSTEI: GOBIIFORMES) DA COSTA BRASILEIRA
Nome do Bolsista: ÉRIKA MILA FERREIRA FERNANDES

Tipo de Bolsa: ( X ) PIBIC/CNPq
( ) PIBIC/UFPA
( ) PIBIC/INTERIOR
( ) PIBIC/FAPESPA
( ) PARD
( ) PARD – renovação
( ) Bolsistas PIBIC do edital CNPq 001/2007

1. INTRODUÇÃO
A família Eleotridae está inserida na subordem Gobioidei, dentro da ordem
Gobiiformes (Wiley& Johnson; 2010). São peixes distribuídos em regiões tropicais,
subtropicais e temperadas de todo o mundo (Nelson, 1976), e podem ser encontrados
em ambientes marinhos, salobros e de água doce, sendo a maioria com hábito estuarino
residente (Thacker & Hardman, 2005; Thacker et al. 2009). Os membros desta família
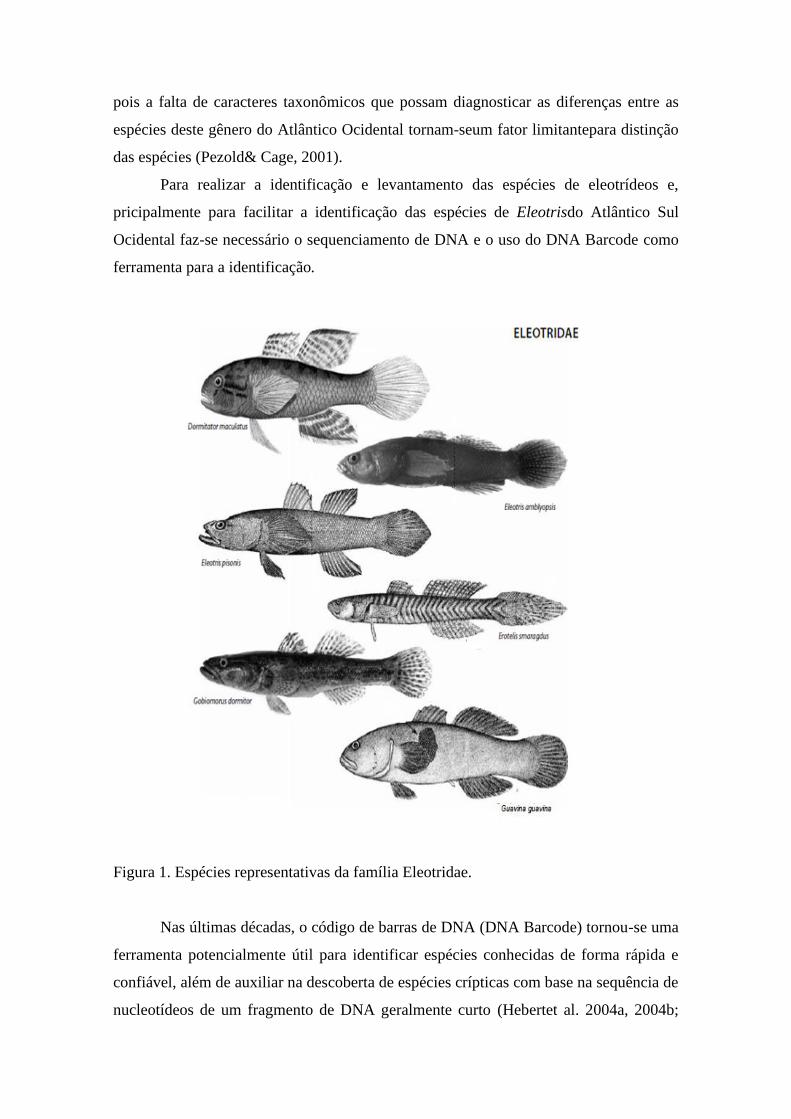
(Figura 1) usualmente são de tamanho pequeno a médio, e compreendem cerca de 130
espécies distribuídas em 23 gêneros (Eschmeyer, 2008). No Brasil, são popularmente
conhecidos como amorés, enquanto que o termo popular em inglês é sleepers
(dorminhocos), pois muitas vezes ficam imóveis no substrato.
São peixes omnívoros, que se alimentam principalmente de plantas, sedimentos,
pequenos invertebrados e outros peixes, incluindo até peixes da mesma família
(Winemiller & Ponwith 1998; Keith et al. 2000). De uma forma geral, estas espécies
apresentam os seguintes caracteres diagnósticos para identificação: duas nadadeiras
dorsais separadas, a primeira com VI a X espinhos flexíveis, a segunda com I espinho
flexível e 6-15 raios macios; nadadeira anal com I espinho flexível e 6-12 raios macios;
nadadeiras pélvicas sempre separadas, com I espinho e 5 raios macios; coloração
variável, e linha lateral ausente (FAO).
Os levantamentos de trabalhos sobre composição da ictiofauna referem-se à
Guavina guavina (Valenciennes 1837), Erotelis smaragdus Valenciennes 1837,
Dormitator maculatus (Bloch 1792) e Eleotris pisonis (Gmelin 1789) como as únicas
espécies de Eleotridae distribuídas na costa brasileira (Oliveira, 1972; Haimovici &
Klippel, 1999; Barletta et al. 2000; Camargo & Isaac, 2002; Barletta et al. 2005;
Barletta & Blaber, 2007; Andrade-Tubino et al. 2008; Bastos et al. 2013; Di Dario et al.
2013; Gonçalves et al. 2013).
No atlântico ocidental são atualmente reconhecidas três espécies de Eleotris:
Eleotris amblyopsis (Cope, 1871) está distribuído desde à Carolina do Norte, Golfo do
México e Caribe até a Guiana Francesa; Eleotris perninger ( Cope,1871) distribuída no
Caribe, mas também foi identificada no Rio de Janeiro; Eleotris pisonis, com
distribuição desde a Carolina do Sul, Golfo do México, Caribe até o Sul do Brasil
(Pezold & Cage, 2001).
A morfologia de algumas espécies encontradas nas Américas tem sido utilizada
de forma satisfatória para identificação, com exceção das espécies do gênero Eleotris,
pois a falta de caracteres taxonômicos que possam diagnosticar as diferenças entre as
espécies deste gênero do Atlântico Ocidental tornam-seum fator limitantepara distinção
das espécies (Pezold& Cage, 2001).
Para realizar a identificação e levantamento das espécies de eleotrídeos e,
pricipalmente para facilitar a identificação das espécies de Eleotrisdo Atlântico Sul
Ocidental faz-se necessário o sequenciamento de DNA e o uso do DNA Barcode como
ferramenta para a identificação.
Figura 1. Espécies representativas da família Eleotridae.
Nas últimas décadas, o código de barras de DNA (DNA Barcode) tornou-se uma
ferramenta potencialmente útil para identificar espécies conhecidas de forma rápida e
confiável, além de auxiliar na descoberta de espécies crípticas com base na sequência de
nucleotídeos de um fragmento de DNA geralmente curto (Hebertet al. 2004a, 2004b;

Ward et al. 2005). Este fragmento consiste numa região padronizada do Citocromo
Oxidase subunidade I (COI) do DNA mitocondrial (mtDNA) (Figura 2), a qual foi
proposta pela primeira vez por Hebert et al. (2003).
A costa norte do Brasil, a qual inclui os estados do Amapá, Pará e Maranhão,
abriga um vasto número de espécies, no entanto, reflete uma grande área pouco
conhecida e amostrada, e abriga o menor número de documentos a respeito da
composição da ictiofauna marinho-estuarina (Marceniuketal. 2013). Os eleotrídeos
fazem parte desta fauna ainda com informações resumidas.
Figura 2. Esquema da molécula de DNA mitocondrial, mostrando a localização do gene
Citocromo Oxidase subunidade I (COI) utilizado para DNA Barcode.
Neste trabalho, o sequenciamento do DNA barcode será empregado para
realizara identificação e levantamento das espécies da família Eleotridae que ocorrem
na costa do Brasil.
JUSTIFICATIVA
As espécies de eleotrídeos na costa brasileira ainda são pouco conhecidas sob
aspectos biológicos, ecológicos e genéticos, uma vez que as abordagens sobre estes
peixes são apenas resultados de estudos sobre a ocorrência, densidade e frequência em

ambientes estuarinos. De um ponto de vista genético e morfológico, é importante
conhecer quais espécies ocorrem na costa brasileira, e como estas estão relacionadas
com os ambientes que vivem.
OBJETIVOS
Objetivo Geral
Identificar as espécies da família Eleotridae coletadas na região costeira do
Brasil através do método do DNA barcode.
Objetivos Específicos
Coletar espécimes de peixes Eleotridaeem estuários da costa brasileira;
Identificar morfologicamente os espécimes coletados com base em
literatura especializada;
Realizar procedimentos laboratoriais para estudos genéticos nos peixes;
Gerar sequencias de DNA dos marcadores COI, e Zic1 para identificação
dos espécimes;
Realizar comparações entre as sequencias geradas neste trabalho e as
sequencias provenientes do GenBank;
Realizar análises das sequencias para contruir árvores filogenéticas e
matriz de distância nucleotídica entre os espécimes coletados.
MATERIAL E METODOS
Área de amostragem
Para este estudo, as espécies de Eleotridae foram coletados em sete diferentes
localidades da costa norte do Brasil: Santana (Amapá), Curuçá (Pará), Ilha de Marajó
(Pará), Ilha de Mosqueiro (Pará),Bragança (Pará), Estuário do Rio Benevente (Espiríto
Santo) e Delta do Parnaíba (Piauí) (Figura 3).

Figura 3. Localização geral do Brasil à esquerda, e à direita área de localização da costa
norte do Brasil onde os espécimes da Família Eleotridae foram amostrados para este
estudo. Legenda: 1) Santana-Amapá; 2) Ilha do Marajó-Pará; 3) Ilha de Mosqueiro-
Pará; 4) Bragança-Pará; 5) Curuçá-Pará, 6) Delta do Parnaíba; 7) Estuário do Rio
Benevente - Espírito Santo.
Os espécimes foram coletados de forma manual durante a maré vazante, ou com
o auxílio de rede de espera nos furos localizados nas regiões de manguezal.
Posteriormente, os espécimes foram levados ao laboratório, acondicionados em
álcool 90%, e então, foram realizadas identificações morfológicas com base em
literatura específica (FAO, Pezold& Cage, 2001).
Extração de DNA, amplificação e sequenciamento
O DNA total de cada espécime foi isolado a partir do protocolo proposto pelo
DNA Kit WizardGenomicPurification. Foram sequenciadas as regiões parciais do gene
CO1 (LCO1490 5’-GGT CAA CAA ATC ATA AAG ATA TTG G-3’ - HCO2198 5’-

TAA ACT TCA GGG TGA CCA AAA AAT CA-3’) (Folmer et al. 1994 e
respectivamente foram sequenciadas as regiões dos genes o Zic 1 (zic1_F9 5'-GGA
CGC AGG ACC GCA RTA YC-3' - zic1_R967 5' CTG TGT GTG TCC TTT TGT
GRA TYT T -3'). As amplificações foram realizadas para um volume final de 25 µl
contendo 4 µl de DNTP (1,25mM), 2,5 µl de solução tampão (10X), 1 µl de MgCl2 (25
Mm), 0,25 µl de cada iniciador (200 ng/µl), 1-1,5 µl de DNA genômico (100 ng/µl), 1U
de Taq DNA Polimerase (5U/µl) e água purificada para completar o volume final de
reação.
O protocolo de PCR utilizado foi: 1 ciclo de 4 minutos a 94 °C; 35 ciclos de 40 s
a 94°C, 30 s a 52 - 65 °C, e 1 minuto a 72 °C; 1 ciclo de 2 minutos a 72 °C; e hold
a4°C.
Os produtos da amplificação foram purificados com a enzima ExoSAP-IT, e em
seguida, os produtos purificados foram submetidos às reações de sequenciamento,
utilizando Big Dye Terminator Cycle Sequencing Kit (Applied Biosystems) no
sequenciador automático ABI 3500 (AppliedBiosystems).
Foram obtidas do GenBank, sequências de COI de eleotrídeos que ocorrem na
costa ocidental do Atlântico Sul: Eleotris amblyopsis (AY722154), G. guavina
(JQ840516.1) e D. maculatus (JQ840473.1). Devido a sequencia de E.pisonis não estar
disponível no GenBank para o gene COI, foi obtida da internet a sequencia de ZIC 1
para esta espécie (KF415953.1) proveniente do trabalho de Agorreta et al. (2013)
(Tabela 2).
Foram também inseridas sequencias de COI do GenBank de outras espécies de
Eleotris do oeste do Indo-Pacífico (Eleotris fusca) e do Hawaii (Eleotris sandwicensis),
bem como da espécie Eleotris picta coletada no estado de Sinaloa (México),
Microphilypnus sp. coletada na bacia Amazônica para complementar as análises dos
dados mostradas na Tabela 2. A espécie Odontobutis obscura foi escolhida como grupo
externo.
Análises das sequencias
As sequencias foram alinhadas automaticamente no ClustalW (Thompson et al.,
1994), e editadas no programa Bioedit 7.0.9 (Hall, 1999).
O programa MEGA 6.0 (Tamura et al. 2007) foi utilizados para calcular as
divergências entre as linhagens sob o modelo de Kimura 2-parameter (K2P) (Kimura

1980). Uma árvore do gene CO1 sob o métodode Agrupamento de Vizinhos foi
construídas no MEGA 6.0 com modelo K2P, e a análise de bootstrap foi implementada
utilizando 1000 réplicas.
Foi gerada uma árvore concatenada com as sequências do gene mitocondrial
COI e de dois nucleares ZIC1 e Rodopsina com a finalidade de mostrar a
verossimilhança da diferença entre as linhagens.Foi utilizada a sequência do gene
ZIC1da espécie de Eleotris pisonis, essa sequência foi comparada com os outros
espécimes de Eleotris.
As relações filogenéticas foram avaliadas utilizando inferência Bayesiana (BI)
realizadas no MrBayes 3.1, utilizando os modelos HKY+G e parâmetrospreviamente
definidos no jModeltest(Posada, 2008). Quatro cadeias Markovianas foram utilizadas
para amostrar a probabilidade espacial em duas corridas simultâneas para 10000000
gerações, e 10% das árvores iniciais serão descartadas como burn-in. Para avaliar se as
corridas representaram com precisão a distribuiçãode probabilidade a posteriori de
todos os parâmetros, o Effective Sample Size (ESS) foi avaliado utilizando o software
Tracer 1.5.
RESULTADOS
Um total de 623 pb do gene COI, 418 pb do gene Rodopsina e 797 pb de ZIC 1
foram obtidos para as análises de todos os espécimes de Eleotridae obtidos para este
estudo.
Nas localidades amostradas, foram capturados um total de 27 espécimes
divididos em três gêneros e quatro espécies Eleotris sp1, Eleotris, sp2, Eleotris sp3, G.
guavina e D. maculatus. A Tabela 1 mostra as localidades com as respectivas espécies e
número de indivíduos capturados.
Tabela 1. Localidades e número de espécimes capturados para este estudo.
Localidade N Espécie Latitude Longitude
Santana - Amapá 3 Eleotris sp1 -0,057078 -51,186195
Mosqueiro - Pará 1 Eleotris sp1 -1,065246 -48,332710
Curuçá - Pará 4 Eleotris sp2 -0,730991 -47,844739
Ilha do Marajó - Pará 2 Eleotris sp2 -0,334248 -48,385857
Bragança - Pará 2 Eleotris sp2 -0,844745 -46,600207
Bragança - Pará 3 Guavina guavina -0,844745 -46,600207

Bragança - Pará 4 Dormitator maculatus -0,844745 -46,600207
Luiz Correa - Piauí 2 Eleotris sp3 -2,774285 -41,848885
Estuário do Rio Benevente- Espírito Santo 6 Eleotris sp3 -2,0634338 -40,691454
Na topologia de CO1, as espécies G. guavina e D. dormitator formaram clados
com 100% de similaridas com suas espécies correspondentes provenientes do GenBank.
Nas topologias que foram obtidas, os espécimes do gênero Eleotris amostrados na costa
brasileira formaram três diferentes linhagens genéticas as quais foram altamente
suportadas por valores de bootstrap.
Na topologia de CO1, os espécimes de Santana e Ilha de Mosqueiro formaram a
Linhagem Norte 1, e apresentaram uma divergência nucleotídica de 10,7% com a
espécie E. amblyopsis e 13,3% com as amostras que correspondem a seu grupo-irmão,
formado por espécimes de Bragança, Curuça e Ilha do Marajó (Linhagem Norte 2).
As espécimes da Linhagem Norte 2 formaram um clado com a espécie
E.amblyopsiscoletada no Panamá, com uma média de divergência genética de 6,5%.
Os espécimes coletadas no Espírito Santo e Piauí formaram um terceiro o clado
Linhagem Nordeste-Sudeste,mostrando uma divergência nucleotídica de 15,3% com E.
amblyopsis, 15,8% com a linhagem Norte 1, e 18,2% com a linhagem Norte 2 (Figura 4
e Tabela 2) .

Figura 4: Topologia de Agrupamento de Vizinhos mostrando as espécies de Eleotridae e
as três diferentes linhagens de Eleotris capturados na costa brasileira.
Tabela 2: Matriz de divergência nucleotídica (K2P) do gene CO1, entre as espéciesde
Eleotridae e os diferentes grupos (linhagens) de Eleotris coletados na costa do Brasil
(marcadas em cinza).
1 2 3 4 5 6 7 8
1 Linhagem Norte 1
2 Linhagem Norte 2 13,3
3 Linhagem Nordeste - Sudeste 15,8 18,2
4 Eleotris amblyopsis 10,7 6,5 15,3
5 Guavina guavina 22,7 19,3 26,6 21,8
6 Guavina guavina GenBank 22,6 19,5 26,4 21,3 0,4
7 Dormitator maculatus 24,1 19,0 23,9 20,6 15,0 15,0
8 Dormitator maculatus GenBank 23,8 18,4 24,0 20,3 14,8 14,7 0,5
A topologia concatenada de Inferência Bayesiana com o marcador mitocondrial
COI e o nuclear ZIC1, mostraram a separação das três linhagens (Norte 1, Norte 2 e

Nordeste-Sudeste) (Figura 5), suportadas por altos valores de probabilidade. A matriz
de divergência nucleotídica baseada no gene Zic 1 é mostrada na Tabela 3.
Figura 5: Árvore concatenada de Inferência Bayesiana, com os marcadores CO1 e Zic 1,
mostrando a separação entre a espécie E.pisonis (GenBank) e as linhagens de Eleotris
capturadas na costa brasileira.
Tabela 3: Matriz de distância nucleotídica (distância p) do gene Zic1 mostrando as
divergências entre a espécie Eleotris pisonis e as linhagens de Eleotris coletadas na
costa do Brasil (marcadas em cinza).
1 2 3 4 5 6 7
1 Linhagem Norte 1 - 161
2 Linhagem Norte 2 - 92 0,4
3 Linhagem Nordeste Sudeste - 286 0,4 0,0
4 Eleotris pisonis (GenBank) 2,0 1,6 1,6
5 Eleotris picta - 2880 2,0 1,6 1,6 0,4
6 Eleotris picta (GenBank) 2,0 1,6 1,6 0,4 0,0
7 Odontobutis obscura 6,5 6,1 6,1 6,9 7,1 7,1
DISCUSSÃO
Segundo (Hebert et al. 2003), o gene mitocondrial, citocromo oxidase
subunidade I (CO1), pode servir como o núcleo de um sistema global para
bioidentificação de espécies animais.
No presente estudo observou-se através das análises moleculares que o gene

CO1 foi eficaz para o reconhecimento das espécies de Eleotridae coletadas na costa do
Brasil, pois as divergências entre as linhagens foram muito altas quando comparadas ao
valor interespecífico de 2% proposto pelo DNA Barcode de Herbert et al. (2003).
Estas análises revelaram três desconhecidas linhagens evolucionárias dos
Eleotriscoletados neste estudo, com valores de distância observados que variaram entre
6,2% e 19%,e que equivalem aos valores que separam as espécies como E. amblyopsis,
E. pisonis, E. picta e E. fusca.
A topologia de CO1 foi exata em mostrar que tanto a Linhagem Norte 1
(Santana e Ilha de Mosqueiro), a Linhagem Norte 2 (Curuçá, Bragança e Ilha do
marajó) e a Linhagem Nordeste-Sudeste (Espírito Santo) até então são desconhecidas,
devido ao fato de mostrarem alta divergência genética (>2%) com a espécie E.
amblyopsis.
Foi gerada a topologia concatenada de Inferência Bayesiana, utilizando o gene
mitocondrial CO1 e o gene nuclear Zic 1, os quais mostrou a separação das três
linhagens (Linhagem Norte1, Linhagem Norte 2 e Linhagem Nordeste-Sudeste), e a
diferença com a típica espécie conhecida na costa brasileira E. pisonis, diferenças
suportadas por altos valores de probabilidade.
Verificou-se que os três morfotipos do presente estudo foram segregados de
acordo como tipo de ambiente onde foram coletados; os Eleotris da linhagem Norte 1,
foram coletados em ambientes de várzea, com baixa salinidade influenciada pelas
descargas do rio Amazonas e Pará, respectivamente. Os Eleotris da linhagem Norte 2,
foram capturados em manguezais de macromaré super úmidos típicos das localidades de
Bragança, Curuçá e Ilha de Marajó, enquanto que os Eleotris pertencentes à linhagem
Nordeste-Sudeste dos estados do Piauí e Espírito Santo correspondem a ambientes de
maguezais tropicais.
Segundo Marceniuket al (2013) a respeito da conservação e conhecimento dos
peixes marinhos e estuarinos da costa norte do Brasil, os principais estudos com tais
peixes são provenientes das região nordeste e sudeste; logo, a grande e rica diversidade
marinha da costa norte brasileira pode evidenciar uma fauna desconhecida, ainda que
trabalhos estejam sendo produzidos para o conhecimento a respeito da composiçãoe
distribuição espaço-temporal dos peixes marinhos-estuarinos (Barletta et al., 2000;
Barletta - Bergan et al., 2002; Camargo & Isaac, 2002; Barletta etal., 2003, Giarrizzo et
al., 2006).
Os peixes que pertencem à família Eleotridae mesmo sendo citados como

ocorrentes nessas devidas áreas ainda é um grupo não conhecido em seus detalhes, tanto
pela sua taxonomia quanto pelos seus termos genéticos.
REFERÊNCIAS
Agorreta,,A., San Mauro, D., Schliewen, U., Van Tassell, J. L., Kovacic, M., Zardoya,
R., Ruber, L. (2013). Molecular phylogenetics of Gobioidei and phylogenetic placement
of European gobies. Molecular Phylogenetics and Evolution 69, 619-633.
Barletta, M., Saint-Paul, U., Barletta-Bergan, A., Ekau, W. & Schories, D. (2000).
Spatial and temporal distribution of Myrophis punctatus (Ophichthidae) and associated
fish fauna in a northernBrazilian intertidal mangrove forest. Hydrobiologia 426, 65-74.
Felsenstein, J. (1985). Confidence limits on phylogenies: an approach using the
bootstrap. Evolution39,783-791.
Hall, T. A. (1999). BioEdit: a user-friendly biological sequence alignment editor
andanalysis program for Windows 95/98/NT. Nucleic Acids Symposium Series 41, 95-
98.
Hebert, P. D. N., Cywinska, A., Ball, S. L. &deWaard, J. R. (2003).Biological
identifications through DNA barcodes.Proceedings of the Royal Society of London
B270, 313–321.
Kimura, M. (1980). A simple method for estimating evolutionary rate of base
substitutions through comparative studies of nucleotide sequences. Journal of
Molecular Evolution16, 111-120.
Marceniuk, A. P., Caires, R. A., Wosiacki, W. B. & Di Dario, F. (2013).Conhecimento
e conservação dos peixes marinhos e estuarinos (Chondrichthyes e Teleostei) da costa
norte do Brasil.Biota Neotropical 13, 251-259.

Perrone, E. C. & Vieira, F. (1991). Hábito alimentar de Eleotrispisonis (Teleostei:
Eleotrididae) ns região estuarina do rio Jocu, Espírito Santo, Brasil. RevistaBrasileira
de Biologia51, 867-872.
Pezold, F. L. & Cage, B. (2002). A review of the spinycheek sleepers, genus
Eleotris(Teleostei: Eleotridae), of the Western Hemisphere, with comparison to the
West African species. Tulane Studies in Zoology and Botany31, 19-63.
Posada, D. (2008).jModelTest: phylogenetic model averaging. Molecular Biology and
Evolution 25, 1253–1256.
Tamura, K., Stecher, G., Peterson, D., Filipski, A. & Kumar, S. (2013). MEGA6:
Molecular Evolutionary Genetics Analysis Version 6.0. Molecular Biology and
Evolution 30, 2725-2729.
Thacker, C. E. & Hardman, M. A. (2005). Molecular phylogeny of basal
gobioidWshes: Rhyacichthyidae, Odontobutidae, Xenisthmidae, Eleotridae (Teleostei:
Perciformes: Gobioidei). Molecular Phylogenetics and Evolution 37, 858-87.
Thacker, C. E. (2003). Molecular phylogeny of the gobioid fishes (Teleostei:
Perciformes: Gobioidei). Molecular Phylogenetics and Evolution26, 354-368.
Thompson, J. D., Higgins, D. G. & Gibson, T. J. (1994). CLUSTAL, W: improving the
sensitivity of progressive multiple sequence alignment through sequence weighting,
position specific gap penalties and weight matrix choice. Nucleic Acids Research22,
4673-4680.