
UNIVERSIDADE FEDERAL DO RIO GRANDE DO SUL … · aumentada com a alimentação. Durante jejum...
37
UNIVERSIDADE FEDERAL DO RIO GRANDE DO SUL FACULDADE DE VETERINÁRIA COMISSÃO DE ESTÁGIO CONTROLE NEUROENDÓCRINO DA SACIEDADE PATRÍCIA RICK BARBOSA PORTO ALEGRE 2006
Transcript of UNIVERSIDADE FEDERAL DO RIO GRANDE DO SUL … · aumentada com a alimentação. Durante jejum...

UNIVERSIDADE FEDERAL DO RIO GRANDE DO SUL FACULDADE DE VETERINÁRIA
COMISSÃO DE ESTÁGIO
CONTROLE NEUROENDÓCRINO DA SACIEDADE
PATRÍCIA RICK BARBOSA
PORTO ALEGRE
2006

UNIVERSIDADE FEDERAL DO RIO GRANDE DO SUL FACULDADE DE VETERINÁRIA
COMISSÃO DE ESTÁGIO
CONTROLE NEUROENDÓCRINO DA SACIEDADE
PATRÍCIA RICK BARBOSA
Monografia apresentada à Faculdade de Veterinária como requisito parcial para a obtenção da Graduação em Medicina Veterinária Orientador: Prof. Félix Hilario Diaz González Co-orientadora: Simone Tostes de Oliveira
PORTO ALEGRE
2006

UNIVERSIDADE FEDERAL DO RIO GRANDE DO SUL FACULDADE DE VETERINÁRIA
COMISSÃO DE ESTÁGIO
CONTROLE NEUROENDÓCRINO DA SACIEDADE
Patrícia Rick Barbosa
_____________________________________
BANCA EXAMINADORA
______________________________________________ Prof.(ª) Nome Completo ______________________________________________ Prof.(ª) Nome Completo ______________________________________________ Prof.(ª) Nome Completo
______________________________________________ Orientador(a): Profº Félix Hilario Diaz González

RESUMO
Uma abundante literatura suporta a existência de um sistema homeostático que,
dinamicamente, ajusta a entrada de energia e o gasto energético para promover a estabilidade
da gordura corporal. Essa regulação é crucial para fisiologia e sobrevivência. O papel
principal do hipotálamo, no sistema nervoso central, tem sido no controle do apetite. Uma
variedade de hormônios e neurotransmissores está envolvida na modulação das vias neuronais
que estimulam ou inibem o apetite. Entre os peptídeos que fazem essa regulação estão os
orexígenos (estimulam o apetite) e os anorexígenos (inibem o apetite). Dentro do grupo de
neuropeptídeos que estimulam a ingestão de alimento estão o neuropeptídeo Y (NPY),
proteína relacionada Agouti (AGRP) e hormônio concentrador de melanina (MCH). E o grupo
formado por neuropeptídeos que inibem a ingestão de alimento contém a leptina, a
proopiomelanocortina (POMC) e a transcrição regulada pela cocaína e anfetamina (CART).
Peptídeos gastroentéricos (grelina, PYY3-36 e colecistocinina) também podem enviar sinais,
através do nervo vago, para o sistema nervoso central e inibir ou estimular a saciedade.
Palavras-chave: Hipotálamo, peptídeos, orexígenos, anorexígenos, NPY, MCH,
AGRP, leptina, grelina, CART, POMC, CCK, PYY3-36.

ABSTRACT
An abundant literature supports the existence of a homeostatic system that
dynamically adjusts energy intake and energy expendure to promote stability of body fat
mass. This regulation of energy homeostasis is critical for normal physiology and survival.
The role of the central nervous system, particularly the hypothalamus, has long been
recognized in the control of appetite. A variety of hormones and neurotransmitters are
involved in the modulation of neuronal pathways that stimulate or suppress appetite. Among
peptides that do this regulation are the orexígens (stimulate the appetite) and the anorexígens
(suppress the appetite). The neuropeptídes group includes the neuropeptides Y (NPY) that
stimulate the food intake, Agouti related protein (AGRP) and melanin concentrator hormone
(MCH). The groups of neuropeptides that suppress the food intake contain the leptin the
proopiomelanocortin (POMC) and the cocaine-amphetamine regulated transcript. The
gastroenteric peptides (ghhrelin, PYY3-36 and cholecystokinin) can send signs too, through
the vague nerve to the central nervous system and inhibit or stimulate the satiety.
Keywords: Appetite, stimulate, suppression, energy, control.

LISTA DE FIGURAS
Figura 1 Localização dos núcleos hipotalâmicos que controlam a saciedade. LH: núcleo
hipotalâmico lateral; PVN: núcleo hipotalâmico paraventricular; ARC: núcleo
arqueado; NST: núcleo trato solitário .................................................................................. 10
Figura 2 Concentração de leptina em animais obesos e em animais magros. ................... 16
Figura 3 Hormônios ativos gerados a partir do POMC e os receptores do peptídeo
melanocortina e seus respectivos locais de ação. ................................................................. 17
Figura 4 Via anorexígena do controle do apetite. ................................................................ 27
Figura 5 Interação da leptina com os demais peptídeos na diminuição e no aumento do
apetite. ...................................................................................................................................... 28
Figura 6 Atuação da grelina juntamente com a leptina para aumentar a ingesta
alimentar. ................................................................................................................................ 30

LISTA DE ABREVIATURAS, SÍMBOLOS E UNIDADES
% Porcentagem
RNA Ácido ribonucléico
NPY Neuropeptídeo Y
POMC Proopiomelanocortina
AGRP Proteína relacionada Agouti
MCH Hormônio concentrador de melanina
CART Transcrição regulada pela cocaína e anfetamina
SNC Sistema nervoso central
CCK Colecistocinina
PVN Núcleo paraventricular
ARC Núcleo arqueado
PYY3-36 Peptídeo YY3-36

SUMÁRIO
1 INTRODUÇÃO ..................................................................................................................... 9
2 NEUROPEPTÍDEOS ANOREXÍGENOS ........................................................................ 11
2.1 LEPTINA ........................................................................................................................... 11
2.2 POMC ................................................................................................................................. 16
2.3 CART ................................................................................................................................. 17
3 NEUROPEPTÍDEOS OREXÍGENOS .............................................................................. 19
3.1 NPY .................................................................................................................................... 19
3.2 AGRP ................................................................................................................................. 20
3.3 MCH ................................................................................................................................... 20
4 PEPTÍDEOS GASTROINTESTINAIS ............................................................................. 22
4.1 GRELINA ........................................................................................................................... 22
4.2 CCK .................................................................................................................................... 24
4.3 PYY3-36 ............................................................................................................................... 25
5 INTERAÇÃO ENTRE LEPTINA E DEMAIS PEPTÍDEOS NA REGULAÇÃO DO
APETITE ................................................................................................................................. 26
5.1 INTERAÇÃO ENTRE GRELINA E LEPTINA................................................................ 28
6 CONSIDERAÇÕES FINAIS .............................................................................................. 31
REFERÊNCIAS ..................................................................................................................... 32

9
1 INTRODUÇÃO
O peso corporal é regulado por uma interação complexa entre hormônios e
neuropeptídeos, sob controle principal de núcleos hipotalâmicos (RODRIGUES et al., 2003).
Entre os determinantes fisiológicos do controle do peso e do apetite estão fatores neuronais,
endócrinos, adipocitários e intestinais.
O hipotálamo é o órgão central que tem papel crítico na regulação a curto e a longo
prazo da ingestão da dieta via síntese de vários neuropeptídeos. Tem sido revelado, por vários
experimentos fisiológicos, que o centro da regulação do apetite está confinado no hipotálamo
ventromedial, hipotálamo lateral e núcleo arqueado (figura 1). Nessas regiões vários sinais
centrais e periféricos são integrados para afetar a expressão do complexo comportamento
alimentar e das funções neuroendócrinas da homeostase energética. O núcleo arqueado, que é
acessível aos hormônios circulantes, é considerado o lugar hipotalâmico primário na
regulação da ingesta alimentar.
Há dois grandes grupos de neuropeptídeos envolvidos nos processos, os anorexígenos
(inibem o apetite) e os orexígenos (aumentam o apetite). Entre os neuropeptídeos orexígenos
encontram-se o NPY, AGRP, MCH e entre os anorexígenos estão a leptina, POMC, CART.
Os neurônios que expressam esses neuropeptídeos interagem entre si e com sinais periféricos
(leptina, insulina, grelina e glicocorticóides), atuando na regulação do controle alimentar e no
balanço energético. A leptina e a insulina são hormônios secretados em proporção à massa
adiposa e atuam, perifericamente, estimulando o catabolismo. No SNC, a insulina e a leptina
interagem com receptores hipotalâmicos, favorecendo a saciedade.
Fatores intestinais também contribuem para modulação do apetite. O trato
gastrintestinal possui diferentes tipos de células secretoras de peptídeos que, combinados a
outros sinais, regula o processo digestivo e atuam no SNC para regulação da fome e da
saciedade. Os peptídeos intestinais, combinados a outros sinais, podem estimular (grelina) ou
inibir (colecistocinia, leptina) a ingestão alimentar. Todos atuam nos centros hipotalâmicos,
que são os grandes responsáveis pelo comportamento alimentar.

10
Figura 1. Localização dos núcleos hipotalâmicos que controlam a saciedade. LH: núcleo hipotalâmico lateral; PVN: núcleo hipotalâmico paraventricular; ARC: núcleo arqueado; NST: núcleo trato solitário

11
2 NEUROPEPTÍDEOS ANOREXÍGENOS
Os neuropeptídeos anorexígenos são aqueles que inibem o apetite. Entre eles estão a
leptina, de grande importância na regulação do apetite, o proopiomelanocortina (POMC) e a
transcrição regulada pela cocaína e anfetamina (CART).
2.1 Leptina
A leptina é um hormônio de 16 kilodalton que foi descoberto em 1994 pelo grupo do
Dr. Friedman, da Universidade Columbia de Nova York. (RODRIGUES et al., 2003). Esta
proteína foi descoberta através de investigações sobre obesidade feitas, com duas cepas de
camundongos (ob/ob e db/db). Harvey, em 1959, fez experimentos de parabiose unindo
sistemas circulatórios de ratos magros e obesos havendo troca de 1% de fluxo sangüíneo entre
eles. Com isso, ele pôde verificar que o aumento do tecido gorduroso produziu um fator
circulante que em contato com o camundongo magro atuou induzindo a saciedade. Após
vários anos estudando com estes modelos experimentais ficou evidente que a cepa ob/ob não
produzia o fator circulante presente no tecido adiposo da cepa db/db. Duas décadas depois
(1994), o Dr. Friedman clonou o gene deste fator, que se denominou proteína ob ou leptina,
do grego leptos, isto é, magro (ZHANG et al., 1994) .
Esta proteína tem uma interrelação com o tecido adiposo, sistema nervoso e órgãos
periféricos (AHIMA & Osei, 2004) sendo sintetizada, principalmente, no tecido adiposo
branco (RODRIGUES et al., 2003), mas também ocorre, em menor grau, em outros órgãos:
epitélio gástrico, placenta, tecido adiposo marrom (BADO et al., 1998 e CINTI et al., 1997) e
no músculo esquelético. A literatura também descreve uma nova síntese de leptina no cérebro,
sugerindo uma ação parácrina ou autócrina (MORASH et al., 1999). Em um estudo feito por
Tsunoda et al. (2003) foi verificado que as células principais do estômago, em caninos,
sintetizam e secretam leptina (fase gástrica) em resposta à secreção de CCK. A expressão
deste gene no estômago é estimulada pela alimentação, CCK e gastrina, sugerindo um papel
na regulação do balanço energético (BADO et al., 1998). Outro trabalho sobre a síntese de
leptina feito por KOCHAN et al. (2006) demonstrou que em aves migratórias, esta proteína
também é produzida no fígado, além do tecido adiposo. Este achado levantou a possibilidade

12
de que a leptina das aves migratórias possa sinalizar o status de reservas energéticas durante
vôos migratórios.
Este hormônio, produzido principalmente pelos adipócitos, informa ao hipotálamo o
tamanho das reservas de gordura, sendo que sua elevação leva à redução da ingesta alimentar
e aumento do gasto energético (LUSTING, 2001), sendo assim um hormônio anorexígeno e
catabólico. A leptina e a insulina são hormônios catabólicos, sendo secretados de acordo com
a adiposidade, sendo assim, sua concentração está diminuída com a restrição calórica e
aumentada com a alimentação.
Durante jejum prolongado, este hormônio pode atuar como um moderador
neuroendócrino, pois a privação alimentar promove hiperatividade do eixo hipotálamo-
hiposário-adrenal, redução da fertilidade, diminuição do metabolismo basal e de repouso,
redução da atividade motora e queda dos hormônios tireoidianos, tendo, essas modificações o
intuito de garantir e prolongar o suprimento energético até que o organismo volte a ter,
novamente, alimento disponível. Com isso, pode-se verificar que a queda nos níveis
plasmáticos de leptina durante o jejum tem como função sinalizar ao cérebro ajustes
neurohormonais para garantir um metabolismo mais eficiente (VOLANTE et al., 1995).
Segundo FREDERICH et al. (1995), “a descoberta que mutações no gene da leptina
causa severa obesidade em roedores, sugere que a função desta proteína seja evitar a
obesidade durante consumo excessivo de alimento”. A descoberta desta proteína foi um
marco no conhecimento molecular do controle da gordura corporal em mamíferos (ZHANG
et al., 1995). Além da função de controle do peso corporal, ela tem muitas outras funções,
como:
• modulação no sistema imunológico
• sinal bioquímico que informa ao cérebro que as reservas energéticas são suficientes
para sustentar o início da puberdade e a reprodução.
• Regula glicose e lipídeos em roedores selvagens, em parte, através da estimulação da
gliconeogênese e aumento da lipólise;
• Ação localizada no estômago influencia a saciedade, através da regulação da
colecistocinina e gastrina;
• Regula metabolismo do músculo esquelético, hematopoiese, angiogênese, cicatrização
de feridas e desenvolvimento cerebral.

13
Hoje se sabe que a leptina é regulada por diversos hormônios e fatores. Em humanos,
vários fatores fisiológicos influenciam a síntese aguda de leptina levando a oscilações na sua
concentração (HARDIE et al., 1996). Esta pode estar diminuída, por exemplo, no jejum, na
exposição ao frio, na presença de testosterona ou agonistas ß-adrenérgicos. E pode ter sua
concentração aumentada por vários fatores como: excesso de ingestão alimentar, obesidade,
insulina, glicocorticóides, infecção aguda, entre outros (AHIMA & OSEI, 2004). Também se
descobriu que esta proteína é secretada em pulsos ao longo do dia, conforme foi demonstrado
por LICINIO et al. (1997) em humanos. Nestes, o padrão pulsátil e o seu período de secreção
ocorre durante a noite e as concentrações plasmáticas são pouco influenciadas pelas refeições,
o que influencia as concentrações plasmáticas é a massa total de tecido adiposo no organismo
(NEGRÃO et al., 2000). No cão, conforme trabalho de ISHIOKA et al. (2003), ocorre
variação sérica diurna da leptina de acordo com a refeição, sendo que esta tem valor mais
baixo uma hora antes da refeição (9:00) e valor mais alto oito horas após refeição (18:00).
Em humanos a secreção de leptina é inversamente proporcional à secreção de ACTH e
cortisol, tendo também um efeito supressor sobre a função tireoidiana (LICINIO et al., 1997)
e positivamente relacionado com gonadotropinas, estradiol e tireotropina (LICINIO et al.,
1998).
O RNAm e a proteína aumentam várias horas após a alimentação. Os efeitos da
nutrição são mediados, em parte, pela insulina, mostrado por uma estimulação direta da
síntese e secreção da leptina pelos adipócitos na presença de insulina. Em humanos e
roedores, a elevação de leptina pós prandial segue o pico de secreção da insulina. Ao
contrário, deficiência de insulina resulta em rápida redução da leptina.
Em humanos há diferenças entre sexos, de acordo com ELBERS et al. (1997). Assim,
as mulheres têm valores mais altos de leptina, devido à estimulação pelo estrógeno, e os
homens têm supressão deste peptídeo pela testosterona. Em roedores, a leptina do macho é
mais elevada, mas as razões não estão ainda bem esclarecidas. Em cães, segundo ISHIOKA et
al. (2006), a concentração plasmática de leptina não é influenciada por idade, sexo ou raça,
mas tem um aumento significativo de sua concentração em cães com escore corporal alto
(obesos), sugerindo esta proteína como um bom indicador da adiposidade nessa espécie
animal, assim como já é conhecido em humanos e roedores.
Após a descoberta e caracterização deste hormônio adipocitário, pesquisadores foram
em busca de seu receptor isolando, principalmente o RNA do receptor da leptina (Ob-R) do
plexo coróide do camundongo. Estudos in situ mostraram que a leptina se liga com alta

14
afinidade nesta região, sugerindo que este seja o local de expressão do seu receptor
(TARTAGLIA et al., 1995).
Até hoje, foram descritos seis isoformas do receptor da leptina: Ra, Rb, Rc, Rd, Re e
Rf. A isoforma Rb, ou forma longa, é encontrada em grande quantidade no hipotálamo,
especialmente no núcleo hipotalâmico arqueado, dorsomedial, ventromedial e ventral,
implicando variações no comportamento alimentar, na termogênese e na regulação hormonal
(ELMQUIST et al., 1998), sendo também encontrado em outros órgãos incluindo pâncreas,
rim, medula adrenal, placenta, ovários e tecido adiposo. Esta variante contém o “BOX” STAT
(transdutoras e ativadoras de sinal de transcrição), o qual não é encontrado nos outros
receptores da leptina (CHEN et al., 1996), sendo esta a forma mais competente em ativar as
vias de sinalização no interior da célula. A variante Re, receptor solúvel, codifica a proteína
mais curta, na qual falta o domínio transmembrana (TARTAGLIA, 1997). Os chamados
receptores curtos (Ra, Rc, Rd, Re e Rf) estão presente no plexo coróide, meninges e vasos
sangüíneos do parênquima cerebral. Essas localizações levantam a possibilidade de que estes
receptores são responsáveis pelo transporte da leptina através da barreira hemato-encefálica
(BJORBAEK et al., 1998).
Mutações nos receptores da leptina foram descobertas em ratos, levando estes a
hiperfagia, obesidade, aumentados níveis de glicocorticóides e hiperglicemia (CHUA et al.,
1996). Segundo RODRIGUES et al. (2003), estudos da relação liquórica/sérica da leptina na
obesidade humana sugerem que a resistência poderia resultar de um defeito no transporte da
leptina ao sistema nervoso central (SNC). A etiologia do defeito não está bem esclarecida,
podendo ser saturação ou problema intrínsico dos transportadores. Outra possibilidade seria a
mutação do gene ob, que é raro na espécie humana. Conforme estudo feito por CHEN et al.
(1996) foi observado que a obesidade severa em camundongos db/db pode ocorrer devido a
uma mutação no receptor da leptina, causando fenótipo igual ao observado em camundongos
ob/ob. A leptino-resistência também poderia ocorrer devido a um defeito pós-receptor,
levando à falha na ativação dos mediadores neuroendócrinos e reguladores do peso corporal
(FILOZOF et al., 2000).
A leptina circula parcialmente ligada a proteínas plasmáticas. Essa ligação está em
equilíbrio com a leptina livre, que é o hormônio bioativo. Uma relação leptina ligada/leptina
livre é elevada em obesos, na gestação e quando existe uma mutação no receptor desta
proteína (SINHA et al., 1996). Pouco é conhecido sobre a interação da leptina com as
proteínas transportadoras na corrente sangüínea. Sinha et al. (1996), trabalhando com leptina
marcada, verificaram que ela se liga a macromoléculas circulantes específicas, de maneira

15
reversível. Em indivíduos magros (21% ou menos de gordura corporal), 60% a 98% da leptina
total foi encontrada de forma ligada. Os estudos sugerem que, em indivíduos obesos, a
maioria da leptina circula na forma livre e, assim, os obesos seriam resistentes à leptina livre.
HOUSEKNECH et al. (1996) e SINHA et al. (1996) acreditam que a proteína ligadora no
plasma seja a forma solúvel do receptor da leptina.
Esta proteína age como um sinal aferente no cérebro que inibe a fome e aumenta o
gasto energético (AHIMA et al., 1999). Uma vez na circulação sangüínea, ela se liga
receptores específicos (Ob-Re) no cérebro, levando ao SNC um sinal de saciedade que reflete
a quantidade de energia em forma de gordura no organismo. As regiões hipotalâmicas densas
em seus receptores estabelecem comunicação entre si e mandam sinais para o sistema nervoso
autônomo e regiões corticais, fornecendo o substrato anatômico onde se dão as alterações
comportamentais e metabólicas causadas por ela (CANTERAS et al., 1994). Agindo por
intermédio destes receptores, a leptina modifica a expressão e a atividade de inúmeros
peptídeos hipotalâmicos que regulam o apetite e o gasto de energia. Além disso, ela sinaliza o
estado nutricional do organismo a outros sistemas fisiológicos, modulando a função de várias
glândulas alvo (NEGRÃO & LICINIO, 2000). Baseado na observação de roedores obesos
(leptino deficientes), estes desenvolvem hiperfagia e obesidade mórbida, que são reversíveis
com o tratamento com leptina (CAMPFIELD et al., 1995). A ação dominante é agir como um
sinal de inanição, pois a leptina diminui rapidamente durante o jejum e desencadeia um
aumento nos glicocorticóides, havendo também um decréscimo na concentração de tiroxina,
dos hormônios sexuais e do crescimento. A diminuição da termogênese durante o jejum e
logo após a hiperfagia é, em parte, devido a um declínio nos níveis de leptina (AHIMA et al.,
2000).
Apesar de vários estudos demonstrarem a correlação entre insulina e leptina,
adipócitos de ratos e humanos sugerem que o metabolismo da glicose, ao invés da
concentração de insulina, é o determinante primário da secreção de leptina (MORAN &
KINZIG, 2004). Segundo NEGRÃO & LICINIO, a leptina administrada a cepa ob/ob de
camundongos corrige a hiperglicemia e hiperinsulinemia, mas em dose inferiores às
necessárias para diminuir o peso, o que sugere que a ela interfere de modo direto no
metabolismo da glicose. Acredita-se também que a leptina esteja associada com a hiperfagia
vista em casos de diabetes mellitus descompesados, isto porque a hiperfagia foi abolida após
administração deste hormônio nestes casos.
O rim é o maior sítio de catabolismo da leptina, removendo 80% de toda leptina
plasmática. Entretanto, os níveis de leptina no plasma permanecem constantes, sugerindo que

16
este peptídeo seja secretado continuamente a partir dos adipócitos, sendo sua velocidade de
remoção igual à taxa de produção (SOARES & GUIMARÃES, 2001).
Figura 2 Concentração de leptina em animais obesos e em animais magros.
2.2 POMC
POMC é um pró-hormônio que gera peptídeos bioativos como ACTH, MSH (α, β, e γ)
e β-endorfina (figura 3). É expressado na hipófise, pele, sistema imunológico e no SNC (no
núcleo arqueado do hipotálamo e no núcleo trato solitário do tronco cerebral).
Peptídeos derivados do POMC têm ação através da ligação com receptores da
melanocortina (MC1R a MC5R) nos tecidos periféricos e no SNC. Os receptores envolvidos
na regulação do peso corporal são MC3R (modula gasto energético) e MC4R (modula
ingestão alimentar). O receptor MC3 está presente no núcleo arqueado e núcleo trato solitário,
placenta, intestino, timo e adipócitos. Nos adipócitos, este é importante no aumento do gasto
energético e na lipólise mediados pelo α-MSH. Já , o receptor MC4 é densamente encontrado
no hipotálamo e é ativado pelo α-MSH reduzindo a ingestão alimentar.

17
Em camundongos, a inativação do gene MC4R causa síndrome da obesidade com
hiperfagia, hiperinsulinemia, hiperglicemia e aumento do crescimento linear sem alterações
da função adrenal nem da capacidade reprodutiva. Este receptor também pode ter efeito sobre
a regulação do comportamento alimentar. Em estudo feito por BRANSON et al. (2003),
verificou-se que todos os indivíduos portadores de mutações no MC4R apresentavam um
transtorno de compulsão alimentar periódica, enquanto apenas 14,2% dos indivíduos obesos e
nenhum indivíduo com peso normal tiveram distúrbio alimentar.
Camundongos com “knockout” do gene POMC tratados com um agonista de α-MSH
perderam peso rapidamente em virtude da diminuição do apetite e do aumento da lipólise.
De acordo com XU et al. (2006), ablação do POMC em ratos exibiu uma progressiva
obesidade e adultos que tiveram um aumentado ganho de peso comparado com o grupo
controle, e a magnitude do efeito foi maior em fêmeas (aumentou 30% o ganho de peso) do
que em machos (aumentou 15% o ganho de peso).
Figura 3 Hormônios ativos gerados a partir do POMC e os receptores do peptídeo melanocortina e seus respectivos locais de ação.
2.3 CART
Transcrição regulada pela cocaína e anfetamina (CART) foi descoberta em 1995 por
Douglas et al. como um peptídeo anoréxico regulado pela leptina. Desde então, foi
demonstrado que o CART é um peptídeo neurotransmissor que tem papel em alguns
processos fisiológicos, incluindo homeostase do peso corporal e balanço energético, em várias
espécies de vertebrados (LARSEN & HUNTER, 2006).

18
O CART é sintetizado como uma grande molécula, sendo um precursor inativo do
peptídeo (pró-CART) e requer processos endoproteolíticos para gerar moléculas menores, as
formas ativas. Para isso são necessários pró-hormônio convertases (PCs) específicas (PC2 e
PC1/3) (STEIN et al., 2006). Este peptídeo (CART) é sintetizado em duas formas em ratos
(116 e 129 aminoácidos), mas somente a forma com 116 aminoácidos é encontrada em
humanos (MATTERI, 2001).
Estes peptídeos estão distribuídos no SNC e são encontrados em altos níveis no núcleo
arqueado e núcleo paraventricular. Ele também é encontrado em uma variedade de tecidos
periféricos incluindo neurônios ganglionares simpáticos, glândula adrenal, intestino, pâncreas
e sangue (VICENTIC, 2006). No pâncreas ele age, diretamente, inibindo a liberação de
insulina (MATTERI, 2001).
Resultados obtidos de vários estudos comportamentais e bioquímicos sugeriram a
existência de múltiplos receptores para o CART. No entanto, a identificação destes ainda não
foi possível (VICENTIC et al., 2006).
O peptídeo CART é um fator de saciedade e está associado às ações de dois
importantes reguladores do apetite, a leptina e o NPY. A administração
intracerebroventricular de peptídeos CART inibem a ingesta normal, a alimentação induzida
pela fome e a ingesta estimulada por NPY em ratos (MATTERI, 2001). Em um estudo feito
por KRISTENSEN et al. (1998), mostrou-se que a administração periférica de leptina em
ratos obesos estimula a expressão de mRNA do CART.
Segundo VICENTIC (2006), a expressão destes peptídeos em ratos e macacos rhesus
exibe um distinto ritmo diurno que está correlacionado com ritmo diário de corticosterona e
com a alimentação. No rato, a adrenalectomia siginifica diminuição nos seus níveis
sangüíneos e abolição no ritmo diário. Administração direta de corticosterona aumenta níveis
sangüíneos de CART. Isto sugere que a glândula adrenal pode ser a origem do CART no
sangue e que glicocorticóides podem ter o papel na regeneração do ritmo diurno deste
peptídeo.

19
3 NEUROPEPTÍDEOS OREXÍGENOS
Os neuropeptídeos orexígenos são aqueles que estimulam o apetite. Entre eles estão o
neuropeptídeo Y (NPY), a proteína relacionada Agouti (AGRP) e o hormônio concentrador de
melanina (MCH).
3.1 NPY
NPY é um neuropeptídeo com 36 aminoácidos encontrado tanto perifericamente como
no SNC (MATTERI, 2001). É um dos mais abundantes neuropeptídeos encontrado no cérebro
de mamíferos e age como potente estimulador do apetite (300). A administração central de
NPY induz hiperfagia, hipotermia, hiperinsulinemia e diminuição do gasto energético.
Também altera o metabolismo de carboidratos e lipídeos, induzindo a lipólise e utilização de
ácidos graxos e triglicerídeos (TACHIBANA et al., 2006).
Este neuropeptídeo está localizado em três áreas distintas: núcleo arqueado, núcleo
paraventricular e periventricular, e tem um importante papel no sistema orexígeno de
aumentar a ingesta alimentar, diminuir o gasto energético e aumentar a lipogênese em animais
(RODRIGUES et al., 2003).
Interessantemente, ratos sem NPY não são magros ou hipofágicos, refletindo métodos
compensatórios por outros peptídeos reguladores para manter a ingesta alimentar e
homeostase energética.
SINDELAR et al. (2006) investigaram o papel do peptídeo orexígeno NPY, em
respostas adaptativas para hipoglicemia induzida pela insulina e concluiram que a sinalização
deste peptídeo é requerida para alimentação hiperfágica, mas não para respostas
neuroendócrinas para hipoglicemias, pois tanto ratos NPY + quanto ratos NPY – produziram
glucagon e corticosterona.
Existem, pelo menos, seis subtipos de receptores (Y1 a Y6). Os receptores Y1 e Y5
estão envolvidos no controle do apetite. Vários subtipos de receptores foram clonados em
suínos (Y1, Y2, Y4, Y5 e Y6), em ovelhas (Y1, Y2 e Y3) e bovinos (Y2). Ambos
antagonistas Y1 e Y5 inibem a alimentação, no entanto, existem evidências de que o receptor
Y1 tem um papel predominante no controle da alimentação. Receptor Y1 tem papel de

20
sinalização na regulação de várias funções comportamentais e fisiológicas, incluindo o
comportamento alimentar e balanço energético, secreção do hormônio sexual e excitabilidade
neuronal. Expressão deste receptor é regulada pela atividade neuronal e por hormônios
periféricos. No hipotálamo, sua expressão é modulada por mudanças no balanço energético
que pode ser induzido por uma ampla variedade de condições, como jejum, prenhez,
hipofagia, obesidade (EVA et al., 2006). Camundogos deficientes em receptores Y1 e Y5
desenvolvem obesidade, limitando o papel de antagonistas destes receptores no tratamento da
obesidade.
3.2 AGRP
Proteína relacionada Agouti (AGRP) é uma proteína com 132 aminoácidos
encontrada, primeiramente, no núcleo arqueado. É estrutural e funcionalmente similar à
agouti, uma proteína produzida na pele, que clareia a cor do tegumento ( MATTERI, 2001).
Esta proteína é um potente antagonista do MC3R e MC4R e é um componente no
processo metabólico que regula alimentação e peso corporal. Sendo um antagonista destes
receptores, ela antagoniza os efeitos anorexígenos do hormônio estimulador de malanócitos
(α-MSH) (NEGRÃO & LICINIO, 2000).
Injeção intracerebroventricular de AGRP estimula a ingesta alimentar. Este peptídeo
encontra-se elevado no jejum e em animais obesos. A supressão da alimentação produzida
pela leptina pode ser bloqueada pela AGRP.
Em suínos jovens, altos níveis de RNAm de AGRP foram encontrados no hipotálamo
e pituitária, baixos níveis no timo, adrenal e testículos e sinais fracamente detectáveis de
AGRP na gordura, músculo, baço e fígado (MATTERI, 2001).
AGRP, em contraste com NPY, apresenta ação prolongada, tendo um potencial
terapêutico nas doenças que cursam com anorexia e emagrecimento.
3.3 MCH
Hormônio concentrador de melanina (MCH) é um peptídeo cíclico com 19
aminoácidos isolado pela primeira vez da glândula pituitária de salmão. Este hormônio é
assim denominado devido à habilidade de clarear a cor da pele pela concentração de

21
melanócitos com grânulos contendo melanina. Este hormônio foi descoberto em áreas do
cérebro de mamíferos associadas ao controle do apetite, no hipotálamo lateral. Neurônios
produtores de MCH no hipotálamo lateral se projetam aos centros do olfato e outras áreas do
córtex cerebral, sendo inibidos pelos neurônios produtores de MSH e estimulados pelos
neurônios NPY/AGRP.
Administração intracerebroventricular de MCH estimula potencialmente a alimentação
em ratos. Ratos deficientes em MCH são hipofágicos, magros e tem elevada taxa metabólica.
A redução da expressão de MCH induzida pela leptina poderia contribuir para reduzir
alimentação em animais normais. O efeito estimulatório de MCH na ingesta alimentar pode
ser bloqueada por inibidores do apetite, como por exemplo, a leptina, α-MSH.
Em ratos, a expressão do receptor do MCH é elevada durante jejum e no genótipo
ob/ob e inibida pelo tratamento com leptina.
Em tecidos humanos, mRNA do MCH tem sido reportado no timo, tecido
adiposo marrom, duodeno e testículos. Em tecidos periféricos de ratos, o MCH é encontrado no
estômago, intestino e testículos.

22
4 PEPTÍDEOS GASTRINTESTINAIS
Além dos peptídeos do SNC que atuam na regulação do apetite, também existem
sinais periféricos que tem como função a regulação da homeostase energética e da ingesta
alimentar, inibindo ou estimulando o apetite. A grelina, um peptídeo gástrico, tem como
função estimular o apetite. Em contrapartida, a colecistocinina (CCK) é um peptídeo intestinal
anorexígeno que envia sinais ao SNC para inibir o apetite. Outro peptídeo entérico citado é o
PYY3-36 que, assim como CCK, também tem função anorexígena.
4.1 Grelina
Descoberto em 1995 como um ligante endógeno do receptor secretagogo do hormônio
do crescimento (ORR & DAVY, 2005). Recentemente, foi identificado como um potente
liberador do hormônio do crescimento e como um peptídeo orexígeno (ALLIA et al., 2002).
A grelina é um hormônio orexígeno produzido principalmente na mucosa gástrica,
mas também tem expressão no intestino, hipotálamo e testículo (WIERUP, 2002). Em ratos e
humanos, é produzida por células gástricas e pela pituitária, placenta e tumores
gastroenteropancreáticos. Foi encontrado também em células endócrinas de pulmões fetais,
sendo esta uma origem adicional da circulação da grelina (ALLIA et al., 2002). Estudo feito
por WIERUP et al. (2002) identificou uma proeminente população de células endócrinas que
expressam grelina no pâncreas fetal, mas essa expressão é muito pequena comparada com a
do estômago. Adultos têm baixa concentração de grelina pancreática.
Este hormônio tem muitas funções, como estimulação da secreção do hormônio do
crescimento, incluindo efeitos na musculatura esquelética, no sistema cardiovascular, função
pancreática endócrina e metabolismo da glicose, aumenta mobilidade gástrica e secreção de
ácido gástrico, tem atividade também na secreção lactotrófica e corticotrófica (ORR &
DAVY, 2005). A grelina também aparece envolvida na regulação do sono, do cortisol e da
prolactina em humanos, sendo um fator que induz e/ou mantém o sono, podendo estar
aumentada após períodos de privação do sono (SCHÜSLLER et al., 2006).
Este peptídeo é um mediador fisiológico da alimentação e, provavelmente, tem uma
função na regulação do crescimento pela estimulação alimentar e liberação de GH. Sendo um

23
hormônio centralmente ativo, pois existe uma ligação entre estômago, hipotálamo e pituitária
(TSCHOP, 2000), sua ação principal é aumentar no jejum, alterando a ingestão calórica e/ou o
status nutricional. TSCHOP et al. (2000) demonstraram que a administração
intracerebroventricular de grelina em ratos gerou um aumento, dose dependente, na ingestão
alimentar e no ganho de peso, pela diminuição da utilização da gordura nesses animais.
GUALILLO et al. (2002) determinaram os níveis de grelina e seu mRNA gástrico
durante 21 dias de restrição alimentar crônica e também em ratos em gestação. Como
resultado, demonstraram que a restrição alimentar aumentou os níveis de grelina e o mRNA
desta, e que durante a gestação, os níveis plasmáticos de grelina aumentaram, particularmente,
durante o final da gestação (19 e 21 dias). Estes achados sugerem que o aumento da grelina
pode ter um papel de mediador das respostas fisiológicas na desnutrição.
CAMINOS et al. (2002) avaliaram em ratos se algumas alterações na homeostase
energética presente nas disordens da função da tireóide e deficiência de GH poderiam ser
mediadas pela grelina e concluiram que a expressão do gene da grelina é influenciada pelos
hormônios da tireóide e pelo GH, pois em ratos hipotireoideos houve um aumento nos níveis
de mRNA da grelina gástrica, ocorrendo o oposto em ratos hipertireoideos. Neste mesmo
modelo experimental com deficiência de GH, foi detectado uma diminuição nos níveis de
mRNA da grelina gástrica.
As concentrações de grelina respondem a flutuações agudas e crônicas do balanço
energético com aumento pré-prandial e durante balanço energético negativo, e diminui pós-
prandialmente e sob condições de balanço energético positivo (ORR & DAVY, 2005).
A grelina é secretada em episódios. Em ratos saciados, episódios secretórios de grelina
consistem em pulsos de baixa amplitude e regular freqüência (2 episódios/hora). Já, em
balanço energético negativo, após privação alimentar, há pulsos de alta amplitude e acelerada
freqüência (3 episódios/hora) (KALRA et al., 2005).
De acordo com GAYLE et al. (2006) há uma diferença nos mecanismos orexígenos e
anorexígenos em ratos conforme o sexo. Em seu experimento foram exploradas respostas
alimentares e hormonais para restrição alimentar de 12 horas ou para estímulo inflamatório
em ratos. Na restrição alimentar, as fêmeas apresentaram maior ingestão alimentar após jejum
do que os machos (40% vs 10%), tendo as fêmeas maior liberação de grelina quando
comparado com o sexo oposto (aumento de 141% nas fêmeas e 65% nos machos). Já, em
resposta ao estímulo inflamatório, ambos os sexos demonstraram similar redução do consumo
alimentar (90).

24
De acordo com CUMMINGS et al. (2006), a supressão de grelina pós-prandial não é
mediada pelos nutrientes no estômago ou duodeno e sim pelo aumento da osmolaridade pós
ingesta no intestino delgado (sinalizado, provavelmente, por nervos entéricos), bem como por
picos de insulina. Conseqüentemente, ingestão de lipídeos suprime pobremente este
enteropeptídeo comparado com outros macronutrientes.
4.2 CCK
CCK é um peptídeo intestinal que foi descoberto na década de 1970 (WOODS et al.,
1998). Tem uma variedade de efeitos no trato gastrointestinal que maximiza a absorção de
nutrientes. Atua como sinal de saciedade, atribuída, principalmente, à sua liberação em
resposta à presença de gordura e proteína (KONTUREK et al., 2004). Este efeito de saciedade
foi demonstrado, primeiramente, em 1973, quando foi visto que a administração de CCK em
ratos reduz, dose dependente, o tamanho da refeição (ORR & DAVY, 2005). Além de inibir a
ingestão alimentar, também induz a liberação de secreção pancreática, biliar e contração
vesicular (KONTUREK et al., 2004).
Este peptídeo é secretado por células enteroendócrinas classificadas como células I,
encontradas na mucosa do duodeno e jejuno (ORR & DAVY, 2005). Também é encontrado
em altas concentrações em várias partes do cérebro onde tem diversos papéis incluindo
memorização, saciedade e dor. O núcleo dorsomedial tem sido implicado na regulação central
da ingesta alimentar e saciedade (KOBELT, 2006). Pode agir também perifericamente pela
estimulação parácrina de aferentes vagais (MATTERI, 2001).
CCK inibe a ingesta diretamente e indiretamente. Ambas vias são mediadas via
receptores CCKA expressados no nervo vago, que projeta para o núcleo trato solitário, e
células musculares circulares do esfíncter pilórico (MORAN & KINZIG, 2004). CCK ativa
neurônios POMC no núcleo trato solitário, e o MC4R é requerido para inibição da ingesta
alimentar pelo CCK (FAN et al., 2004). Inibição indireta é adquirida através da ativação dos
receptores CCKA localizados nas células musculares do esfíncter pilórico, que causa
contração pilórica, inibição do fluxo transpilórico, e diminuição na taxa do esvaziamento
gástrico (MORAN & KINZIG, 2004). Há também evidência de que CCK pode inibir ingesta
pela supressão do NPY no hipotálamo dorsomedial.

25
Liberação gastrointestinal de CCK é mediada por macronutrientes específicos, sendo
que a gordura e a proteína produzem maior concentração pós-prandial de CCK do que
carboidratos.
4.3 PYY3-36
PYY3-36 foi isolado em 1980 de tecido intestinal de suíno e em 1993 foi administrado
perifericamente e se observou a diminuição do apetite, sendo então um peptídeo anorexígeno.
Há duas formas endógenas deste peptídeo: PYY1-36 e PYY3-36. Ambas as formas são
biologicamente ativas, mas somente o PYY3-36 demonstra alta afinidade para inibir Y2R, onde
age como um agonista. Altas concentrações de PYY1-36 são observadas durante o jejum,
enquanto as concentrações de PYY3-36 estão aumentadas após alimentação. A concentração de
PYY aumenta após 15 minutos da ingesta alimentar e continua elevada por 5 horas, com pico
após, aproximadamente, 2 horas.
O maior sítio de produção de PYY são as células endócrinas L do trato gastrointestinal
distal, e são secretados de acordo com a quantidade de energia ingerida.
Este enteropeptídeo é um potente redutor do esvaziamento gástrico. Pesquisas recentes
sugerem que PYY3-36 inibe o apetite através da habilidade de reduzir a secreção de NPY no
núcleo arqueado. PYY3-36 é um seletivo agonista do inibitório Y2R, que regula a secreção de
NPY. Ele diminui a concentração de RNAm do NPY e aumenta a concentração de RNAm do
POMC no núcleo arqueado.

26
5 INTERAÇÃO ENTRE LEPTINA E DEMAIS PEPTÍDEOS NA REGULAÇÃO DO
APETITE
Apesar do progresso, pouco é conhecido sobre como a ação hipotalâmica da leptina
influencia o comportamento alimentar. Verificou-se que ela age de modo conpíscuo com
quatro neuropeptídeos produzidos no núcleo arqueado: NPY, AGRP, POMC e CART.
O efeito central da leptina é restringir a ingestão calórica pela supressão da síntese,
liberação e ação do NPY no eixo núcleo arqueado-núcleo paraventricular (KALRA et al.,
2005). A literatura afirma que os neurônios do núcleo arqueado são alvos para ação central da
leptina na homeostase energética. A sinalização da leptina no núcelo arqueado é a chave
determinante da resposta do cérebro para a entrada do sinal da saciedade.
Dentro do núcleo arqueado são descritas duas subpopulações neuronais que expressam
receptores de leptina, regulando a ingesta alimentar e que tem projeções para o núcleo
paraventricular e lateral. Uma subpopulação é composta por neurônios orexígenos, que
contêm NPY e AGRP e são inibidos pela leptina e a outra subpopulação são os neurônios
anorexígenos que expressam POMC e CART e são estimulados pela leptina. Portanto, as
propriedades anorexígenas da leptina são desencadeadas através da interação com
NPY/AGRP e POMC/CART no núcleo arqueado e suas subseqüentes influências nos
neurônios alvos de várias regiões do cérebro, envolvidas na regulação da ingestão alimentar.
A concentração de leptina a curto prazo é influenciada pela ingestão alimentar (ORR
& DAVY, 2005), sendo sua concentração pós-prandial influenciada pelos macronutrientes da
refeição, ou seja, alimentos com muito carboidrato e pouca gordura produzem maior aumento
nos níveis de leptina pós-prandial quando comparado com dieta com altos níveis de gordura e
baixo de carboidrato.
Durante o jejum, a diminuição da leptina estimula a expressão do NPY e AGRP, e
causa supressão do CART e POMC (figura 4 e figura 5). Este aumento na concentração de
NPY estimula a ingestão alimentar e ganho de peso (AHIMA et al, 2000), sendo este
neuropeptídeo um sensor do decréscimo na concentração plasmática da leptina. Leptina
aumenta diretamente a atividade dos neurônios POMC, causando mais freqüentes potenciais
de ação, e diminui a atividade dos neurônios NPY. A leptina aumenta o tônus excitatório e
diminui o tônus inibitório sobre os neurônios POMC e tem efeitos opostos nos neurônios
NPY.
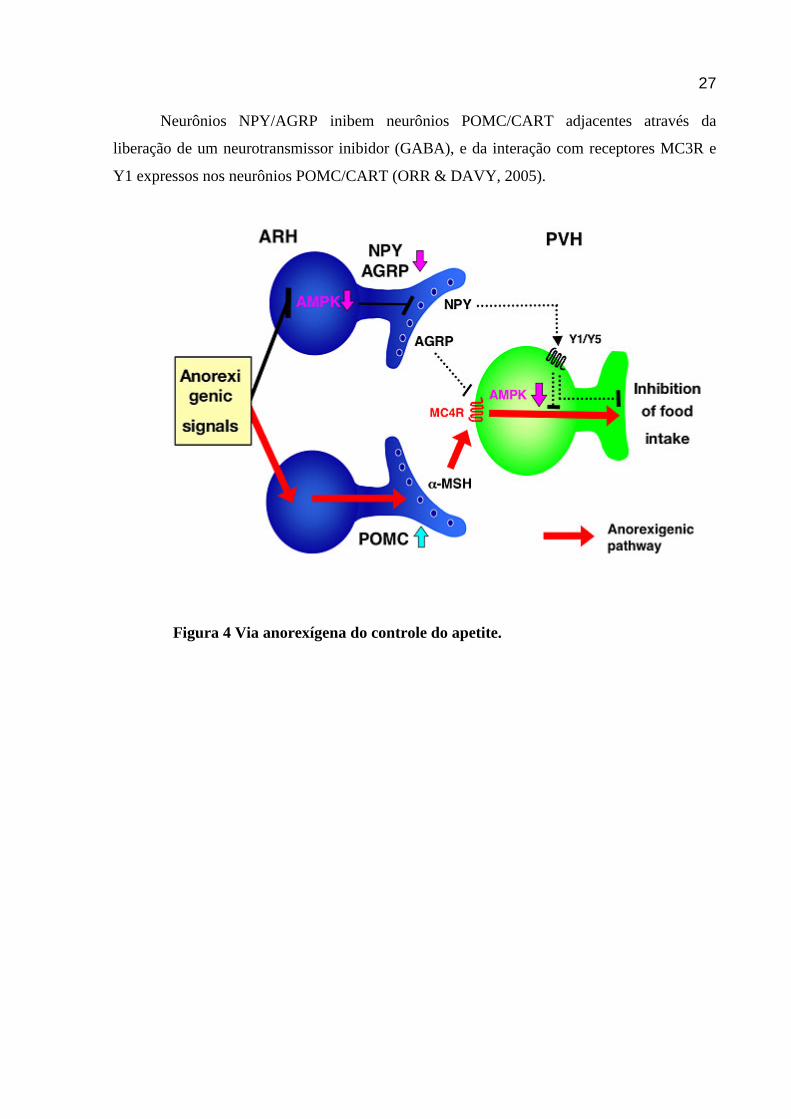
27
Neurônios NPY/AGRP inibem neurônios POMC/CART adjacentes através da
liberação de um neurotransmissor inibidor (GABA), e da interação com receptores MC3R e
Y1 expressos nos neurônios POMC/CART (ORR & DAVY, 2005).
Figura 4 Via anorexígena do controle do apetite.

28
Figura 5 Interação da leptina com os demais peptídeos na diminuição e no aumento do apetite.
5.1 Interação entre grelina e leptina
A grelina está envolvida na regulação hipotalâmica da homeostase energética. Ela
aumenta a expressão do gene NPY e bloqueia a redução da alimentação, induzida pela leptina,
deduzindo que há uma interação competitiva entre grelina e leptina na regulação alimentar
(NAKAZATO et al., 2001). Os dois hormônios, anorexígeno (leptina) e orexígeno (grelina),
são os principais sinais aferentes para a ativação da rede de regulação do apetite (KALRA,
2005).
Grelina promove a produção de neuropeptídeos orexígenos (NPY e AGRP), no núcleo
arqueado do hipotálamo, e ativa os neurônios que produzem esses neuropeptídeos, resultando
em um aumento na ingestão calórica e ganho de peso, liberação de ácido gástrico e secreção
de proteínas plasmáticas através da ativação vagal e neuronal intrapancreática,
respectivamente (YOSHIHARA et al., 2002). Esta proteína gástrica atravessa a barreira

29
hematoencefálica e, em conjunto com a grelina produzida localmente no hipotálamo, ocupam
o complexo de NPY e grupos no núcleo paraventricular e núcleo arqueado, eixo que desperta
o apetite (KALRA et al., 2005).
Grelina faz aumentar a ingesta alimentar, primariamente, através da interação com
NPY/AGRP no núcleo arqueado (KAMEGAI et al., 2001). Ela aumenta os níveis de RNAm
de NPY e AGRP no núcleo arqueado.
A leptina é o primeiro sinal que aumenta a termogênese e restringe os efeitos
orexígenos da grelina em alvos periféricos e centrais, sendo, portanto, este um regulador da
secreção de grelina. Segundo KAMEGAI et al. (2004), a insulina e a leptina inibem, dose
dependentemente, a secreção de grelina, enquanto o glucagon aumenta a grelina plasmática
dose dependente.
O aumento da grelina anteriormente à refeição dispara uma seqüência de eventos na
regulação do apetite. O início da síntese, no núcleo arqueado, é seguido pela liberação no
núcleo paraventricular de NPY, AGRP e GABA, que ativam receptores Y1/Y5 e GABA e
bloqueiam receptores MC4. Esta seqüência induz o apetite (118).A estimulação do apetite
pela grelina é propagada pela sinalização dinâmica do NPY no eixo núcleo arqueado - núcleo
paraventricular (KALRA et al., 2004).
Grelina está reduzida na obesidade e elevada na anorexia e em pacientes magros. Em
contraste, a leptina está diminuída nestas situações. A relação inversa entre grelina e leptina
no plasma é dependente da quantidade de gordura corporal assim como do status nutricional
(figura 6).
Uma análise eletrofisiológica revelou que a grelina aumenta a maioria das células
hipotalâmicas que são inibidas pela leptina. Então, os efeitos opostos da leptina e grelina na
alimentação podem ser mediadaos através de alvos neuronais similares no núcleo arqueado
(COWLEY et al., 2003).
CHEN et al. (1996) evidenciaram que a grelina periférica atua através do NPY/AGRP
hipotalâmico e neurônios POMC para estimular a alimentação. Grelina apenas funciona na
regulação da alimentação se NPY e AGRP estiverem presentes.
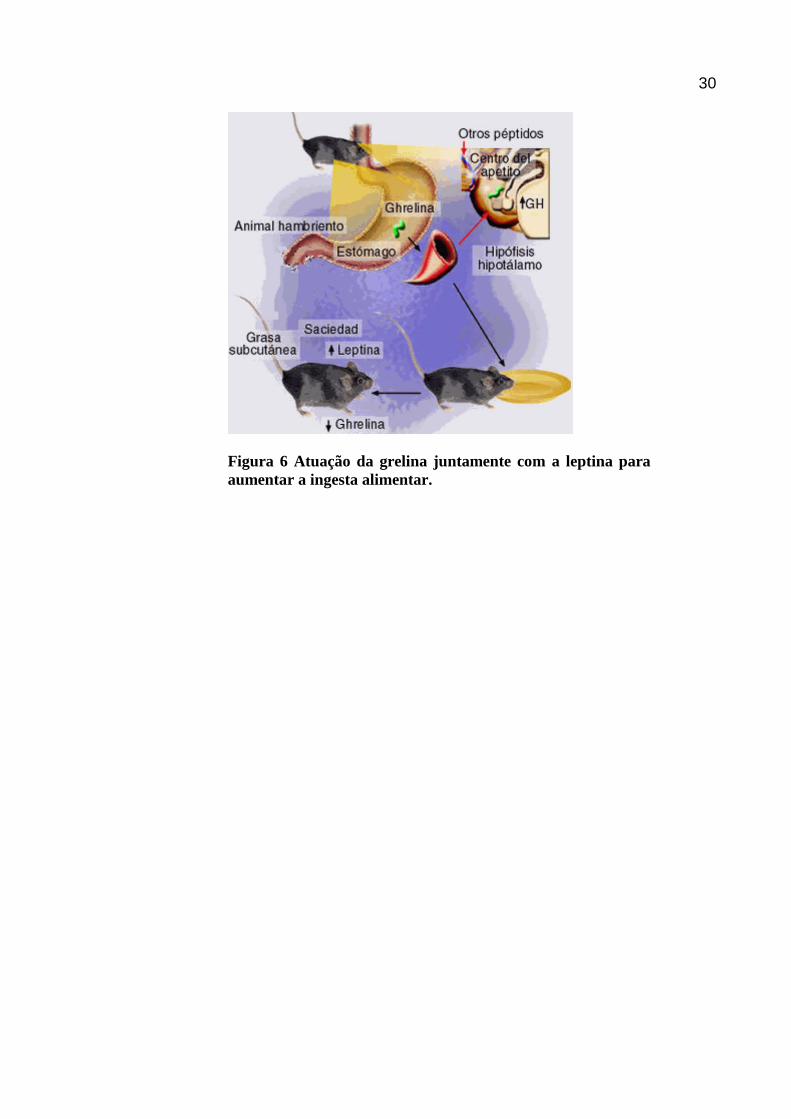
30
Figura 6 Atuação da grelina juntamente com a leptina para aumentar a ingesta alimentar.

31
6 CONSIDERAÇÕES FINAIS
A regulação fisiológica do apetite é uma área em grande expansão na pesquisa e de
grande importância tanto para animais domésticos quanto para seres humanos, pois o índice
de obesidade tem crescido cada vez mais no mundo. É estimado que 24 a 30% da população
pet nos EUA esteja obesa.
A importância da ingestão alimentar em animais domésticos é inquestionável. No
entanto, existem poucos estudos de controle do apetite nestas espécies quando comparado a
roedores de laboratório. Para animais de produção, por exemplo, essa regulação pode ter
grande significado para aumentar o ganho de peso do animal, melhorar os índices de
reprodução e aumentar a eficiência da taxa de crescimento e lactação. Já, em animais de
companhia, como cães e gatos, e humanos a obesidade vem se tornando um grande problema
e a descoberta de peptídeos que controlam o apetite e os seus mecanismos de ação podem
gerar novas drogas para controlar este problema que traz sérios riscos à saúde e que pode
levar à morte.

32
REFERÊNCIAS
AHIMA, Rexford et.al. Distinct physiologic and neuronal responses to decreased leptin and mild hyperleptinemia. The Endocrinology Society, USA [s.d], vol, 140, n. 11.
AHIMA, Rexford S. & OSEI, Suzette Y. Leptin signaling. Physiology & Behavior 81 (2004) 223-241.
AHIMA, Rexford S. et.al. Leptin regulation of neuroendocrine systems.: Frontiers in Neuroendocrinology. July, 200, vol. 21, Issue 3, p. 263-307.
AHIMA, RS. et.al. Role os leptin in the neuroendicrine response to fasting. Nature, 1996 Jul 18;382(6588):250-2.
AILHAUD, Gerard. Adipose tissue as a secretory organ: from adipogenesis to the metabolic syndrome. Disponível em: www.sciencedirect.com acesso em 06/10/2006.
ARORA, Sarika & ANUBHUTI. Role of neuropeptides in appetite regulation and obesity – A review. Neuropeptides. Article in press, correct proof – Note to users.
BADO, A. et.al. The stomach is a source of leptin. Nature, 1998 Aug 20;394(6695):790-3.
BJORBEK, Christian. et.al. Expression of leptin receptor isoforms in rat brain microvessels. Journal of The Endocrinology Society, september, 2006, Vol. 139, number 8.
BOGHOSSIAN, S. et.al. Increased leptin expression in the dorsal vagal complex suppresses adiposity without affecting energy intake and metabolic hormones. Obesity (Silver Spring). 2006 Jun;14(6):1003-9.
BRANSON, R., POTOCZA, N., KRAL, J.G., LENTES, K.U., HOEHE, M.R., HORBER, F.F. Binge eating as a major phenotype of melanocortin 4 receptor gene mutations. N. Engla. J. Med. 2003 Mar; 348 (12): 1096-103.
CAMINOS, JE. et.al. Influence of tryroid status and growth hormone deficiency on ghrelin. Eur J Endocrinol 2002 Jul;147(1): 159-63.
CAMPFIELD, LA.et.al. Recombinant mouse OB protein: evidence for a peripheral signal linking adiposity and central neural nteworks. Science. 1995 Jul 28;269(5223):546-9.
CHEN, H. et.al. Evidence that the diabetes gene encodes the leptin receptor: identification of a mutation in the leptin receptor gene in db/db mice. Cell. 1996 Feb 9;84(3):491-5.

33
CHUA, SC Jr. et.al. Phenotypes of mouse diabetes and rat fatty due to mutations in the OB (leptin) receptor. Science. 1996 Feb 16;271(5251):994-6.
CINTI, Saverio. et.al. immunohistochemical localization of leptin and uncooupling protein in white and brown adipose tissue. Endocrinology, September, 2006, Vol. 138, number 2.
COWLEY MA. The distribution and mechanism of action of ghrelin in the CNS demonstrates a novel hypothalamic circuit regulating energy homeostasis. Regul Pept. 2004 Jun 15;119(1-2):77-81.
COWLEY, MA. et.al. The distribution and mechanism of action of ghrelin in the CNS demonstrates a novel hypothalamic circuit regulating energy homeostasis. Neuron, 2002 Feb 20;37(4):550-3.
COWLEY, MA. et.al. The distribution and mechanism of action of ghrelin in the CNS demonstrates a novel hypothalamic circuit regulating energy homeostasis. Neuron. 2003 Feb 20;37(4):550-3.
COWLEY, MA. Hypothalamic melanocortin neurons integrate signals of energy state. Neuron. 2003 Feb 20;37(4):649-61.
CUMMINGS, David E. Ghrelin and the short – and long-term regulation of appetite and body weight. Physiology & Behavior. 30 august 2006, vol. 89, Issue 1, p. 71-84.
CUPPLES, W.A. Physiological regulation of good intake. Disponível em: www.ajpregu.org acesso em: 17/10/2006.
ELMQUIST, JK.et.al. Distributions of leptin receptor mRNA isoforms in the rat brain. J Comp Neurol. 1998 Jun 15;395(4):535-47.
EMILSSON, V. et.al. Expression of the functional leptin receptor mRNA in pancreatic isltes and direct inhibitory action of leptin on insulin scretion. Diabetes. 1997 Feb;46(2):313-16.
EVA, Carola et.al. Physiology and gene regulation of brain NPY Y1 receptor.: Frontires in Neuroendocrinology. September, 2006, vol 27, Issue 3, p. 308-339.
FAN, W. Cholecystokinin-mediated suppression of feeding involves the brainstein melanocortin system. Nat Neurosci. 2004 Apr;7(4):335-6. EPUB 2004 Mar 14.
FANTUZZI, Giamilia. Adipose tissue, adipokines, and inflammation. American Academy of Allergy, Asthma and Imunologyi. 10.1016/j.jaci.2005.02.023.
FILOZOF, C.M. et al. Low plasma leptin comcentration and low rates of fat oxidation in weight – stable post – obese subjects. Obese Res. 2000 May; 8 (3): 205-10.

34
GAYLE, Dave A. Gender-specific orexigenic and anorexigenic mechanisms in rats. Life Sciences. 13 September 2006, vol. 79, Issue 16, p. 1531-1536.
GUALILLO, O. et.al. Effect of food restriction on ghrelin in normal-cycling female rats and in pregnancy. Obes Res 2002 Jul; 10(7): 682-687.
HALAAS, JL.et.al. Weight-reducing effects of the plasma protein encoded by the obese gene. Science. 1995 Jul 28;269(5223):543-6.
HOUSEKNECHT, KL. et.al. Evidence for leptin binding to proteins in serum of rodents and humans: modulation with obesity. Diabetes. 1996 Nov;45(11):1638-43.
HOUSEKNECHT, K.L.; PORTOCARRERO, C.P. Leptin and its receptors: regulators of whole-body energy homeostasis. Domest Anim Endocrinol. 1998 Nov;156(6):457-75.
ISHIOKA, K. et.al. Diurnal variations of serum leptin hit2hit2 in dogs: effects of fasting and re-feeding. Endocrinology, dói:10.1210/em??.2005-1464.
ISHIOKA, K. et.al. Plasma leptin concentration in dogs: effects of body condition score, age, gender and breeds. Research in Veterinary Science (2006), dói:10.1016/j.rvsc.2006.06.002.
JOLANDA, M. et.al. Reversal of the sex diffrence in serum leptin levels upon cross-sex hormone administration in transsexuals. Journal of Clinical Endocrinology and Metabolism, September, 2005, vol. 82, number10.
KALRA, Satya P. Circumventing leptin resistence for weight control. PNAS 2001 Apr, vol. 98, number 8, 4279-4281.
KALRA, Satya P. et.al. Neuropeptide Y secretion increase in the paraventricular nucleus in association with increased apptide for food. Proc. Natl. Acad. Sci, USA. Vol. 88, pp. 10931-10935, December 1991. Neurobiology.
KALRA, Satya P. et.al. Stimulation of appetite by ghrelin is regulated by leptin restraint: peripheral and central sites of action. Disponível em: http://jn,nutrition.org acesso em: september/2006.
KALRA, SP & KALRA, PS. Neuropeptide Y: a physiological orexigen modulated by the feedback action of ghrelin and leptin. Endocrine. 2003 Oct; 22(1):49-56.
KAMEGAI, J. et.al. Effects of insulin, leptin, and glucagon on ghrelin secretion from isolated perfused rat stomach. Brain Mol Brain Res 1992 Sep;15(1-2):15-8.
KAMEGAI, J. et.al. Effects of insulin, leptin, and glucagon on ghrelin scretion from isolated perfused rat stomach. Regul Petpt. 2004 Jun 15;119(1-2):77-81.

35
KAMEGAI, Jun et.al. Chronic central infusion of ghrelin increases hypothalamic neuropeptide y and agouthi-related protein MRNA levels and body weight in rats. Diabetes, vol 50, november 2001.
KOBELT, Peter. Peripheral injection of CCK-8S induces fos expression in the dorsomedial hypothalamic nucleus in rats. Disponível em: http://www.sciencedirect.com/science acesso em 07/10/2006.
KOCHAN, Zdzislaw; KARBOWSKA, Joanna; MEISSNER, Wlodzimierz. Leptin is synthesized in the liver and adipose tissue of the dunlin (Calidris alpina). General and comparative endocrinology. September 2006, vol. 148, Issue 3, p. 336-339.
KONTUREK, S.J. et al. Brain-gut axis and its role in the control of food intake. J. Physiol. Pharmacol. 2004 Mar; 55: 137-54.
KRISTENSEN, P., et al. Hypothalamic CART is a new anoretic peptide regulated by leptin. Nature. 1998 May; 393 (6680): 72-6.
LARSEN, Philip J. & HUNTER, Richard G. The role of CART in body weight homeostasis. Peptides. August 2006, vol. 27, Issue 8, p. 1981-1986.
LICINIO, J. et.al. Human leptin levels are pulsatile and inversely related to pituitary-adrenal function. Nat Méd. 1997 May;3(5):575-9.
LICINIO, Julio. et.al. Sex differences in circulating human leptin pulse amplitude: clinical implications. Journal of Clinical Endocrinology and Matabolism. September, 2006, Vol 89, number 11.
LUSTING, R.H. The neuroendocrinology of obesity. Endocrinol Metab Clin North Am. 2001 Sep;30(3):765-85.
MATTERI, R.L. Overview of central targets for appetite regulation. J. Anim. Sci. 79(E. Suppl): E148-E158.
MORAN Timothy H. & KINZIG, Kimbewrly P. Gastrointestinal satiety signals. Am J Physiol Gastrointest Liver Physiol 286: G183-G188, 2004.
MORASH, Barbara. et.al. Leptin gene expression in the brain and pitutary gland. Journal of The Endocrinology Society, september, 2006, Vol. 140, number 12.
MORTON J. gregory et.al. Leptin action in the forebrain regulates the hindbrain response to satiety signals. The journal of clinical investigation, disponível em: www.jci.org March 2005, vol. 115, Number 3.
NAKAZATO, M. et. Al. A role for ghrelin in the central regulation of deeding. Nature 2001 jan 11; 409(6817): 194-8.

36
NEGRÃO, André B. & LICINIO, Julio. Leptina: o diálogo entre adipócitos e neurônios. Arq Bras Endocrinol Metab, vol 44, nº 3, junho 2000.
ORR, Jeb. & DAVY, Brenda RD. Dietary influences on peripheral hormones regulating energy intake: potential applications for weight management. Journal of the AMERICAN DIETETIC ASSOCIATION, July, 2005, vol. 105, number 7.
OTUKONYONG, EE. et.al. Central leptin differentially modulates ultradian secretory patterns of insulin, leptin and ghrelin independent of effects on food intake and body weight. Peptides. 2005 Dec;26(12):2559-66.
RODRIGUES, Adriane M; SUPLICY, Henrique L; RADOMINSKI, Rosana B. Controle neuroendócrino do peso corporal: implicações na gênese da obesidade. Arq Bras Endocrinol Metab, vol 47, nº 4, agosto 2003.
SCHÜSSLER, P. et.al. Noctural ghrelin, ACTH, GH and cortisol secretion after sleep deprivation in humans.: Psychoneuroendocrinology. September 2006, vol. 31, Issue 8, p. 915-923.
SINDELAR, Dana K. Neuropeptide Y is required for hyperphagic feeding in response to neuroglucopenia. Disponível em: www.endo.endojournals.og acesso em: 05/09/2006.
SINHA, Madhur K. et.al. Evidence of free and bound leptin in human circulation. The American Society for Clinical Investigation. Vol. 98, number 6, setember 1996, 1277-1282.
SOARES, M.A.M. & GUIMARÃES, S.E.F. O papel da leptina e de seus receptores no metabolismo da gordura. Disponível em: http://www.conferencia.uncnet.br/pork/seg/pal/anais01p2_soares_pt.pdf acesso em 6/08/2006.
STEIN, Jeffrey; STEINER, Donald F.; ARUNANGSU, Dey. Processing of cocaine – and amphetamine-regulated transcript (CART) precursor proteins by prohormone convertases (PCs) and its implications. Peptides. August 2006, vol. 27, Issue 8, p. 1919-1925.
TACHIBANA, Tetsuya et.al. Intracerebroventricular injection of neuropeptide y modifies carbohydrate and lipid metabolism in chicks. Disponível em: www.sciencedirect.com acesso em: 10/10/2006.
TARTAGLI, LA. et.al. Identification and expression cloning of a leptin receptor, OB-R. Cell. 1995 Dec 29;83(7):1263-71.
TARTAGLIZ, Louis A. The leptin receptor. The Journal of Biological Chemistry, Vol. 272, number 10, Issue of march 7, pp. 6093-6096, 1997.
TSCHOP, M.; SMILEY, DL.; HEIMAN, ML. Ghrelin induces adiposity in rodents. Nature 2000 oct 19; 407 (6806):908-13.

37
TSUNODA, Yasuhiro et.al. Cholecystokinin synthesizes and secretes leptin in isolated canine gastric chief cells. Biochemical and Biophysical Research Communications. 24 october 2003, vol. 310, Issue 3, p. 681-684.
UENO, Naohiko. et.al. Leptin modulates orexigenic effects of ghrelin and attenuates adiponectin and insulin levels and selectively the dark-phase feeding as revealed by central leptin gene therapy. Endocrinology. 2004 145(9):4176-4184.
VICENTIC, Aleksandra. CART peptide diurnal variations in blood and brain. Peptides. August 2006, vol. 27, Issue 8, p. 1942-1948.
VICENTIC, Aleksandra; LAKATOS, Anita; JONES, Douglas. The CARTS receptors: backgroun and recent advances. Peptides. August 2006, vol. 27, Issue 8, p. 1934-1937.
VICENTIC, Alksandra. CART peptide diurnal variations in blood and brain. Peptides. August 2006, vol. 27, Issue 8, p. 1942-1948.
VOLANTE, M. et.al. Ghrelin expression in fetal, infant and adult human lung. Histochem Cytochem 2002 Aug:50(8): 1013-21.
VRANG, Niels. Anatomy of hypothalamic CART neurons. Peptides. August 2006, vol. 27, Issue 8, p. 1970-1980.
WIERUP, N. et.al. The ghrelin cell: a novel developmentally regulated islet cell in the human pancreas. Regul pept 2002 Jul 15;107(1-3): 63-9.
WOODS, S.C. et al. Signals that regulate food intake and energy homeostasis. Science May 1998, 280 (5368):1378-83. Rewiew.
XU, Allison Wanting et.al. Effects of hypothalamic neurodegeneration on energy balance. Disponível em: www.plosbiology.org December 2006, vol. 3, Issue 12, E415.
XU, B. et.al. Daily changes in hypothalamic gene expression of neuropeptide Y, galanin, proopiomelanocortin, and adipocyte leptin gene expression ands secretion: effects of food restriction. The Endocrinology Society, USA [s.d], vol, 140, n. 6.
YOSHIHARA, F. et.al. Ghrelin: a novel peptide for growth hormone release and feeding regulation. Curr Opin Clin Nutr Metab Care 2002 Jul; 5(4): 391-5.
ZHANG, Y. et.al. Positional cloning of the mouse obese gene and its human homologue. Nature. 1995 Mar 9;374(6518):124.
ZHANG, Y. et.al. Positional cloning of the mouse obese gene and this human homologue. Nature. 1995 Mar 9;374(6518):124.


















