
UNIVERSIDADE FEDERAL DO RIO GRANDE FURG … · (Fernão Capelo Gaivota). A Ivis Winievisck, meu...
98
i UNIVERSIDADE FEDERAL DO RIO GRANDE – FURG PROGRAMA DE PÓS- GRADUAÇÃO EM AQUICULTURA – PPGAq INSTITUTO DE OCEANOGRAFIA – IO RESPOSTAS ANTIOXIDANTES E DANO OXIDATIVO NO CAMARÃO Litopenaeus vannamei: EFEITOS DA SUPLEMENTAÇÃO COM ANTIOXIDANTE E USO DE TECNOLOGIA DE BIOFLOCOS ÁTILA CLIVEA DA SILVA MARTINS RIO GRANDE, RS AGOSTO, 2015 1
Transcript of UNIVERSIDADE FEDERAL DO RIO GRANDE FURG … · (Fernão Capelo Gaivota). A Ivis Winievisck, meu...

i
UNIVERSIDADE FEDERAL DO RIO GRANDE – FURG
PROGRAMA DE PÓS- GRADUAÇÃO EM AQUICULTURA – PPGAq
INSTITUTO DE OCEANOGRAFIA – IO
RESPOSTAS ANTIOXIDANTES E DANO OXIDATIVO NO CAMARÃO
Litopenaeus vannamei: EFEITOS DA SUPLEMENTAÇÃO COM
ANTIOXIDANTE E USO DE TECNOLOGIA DE BIOFLOCOS
ÁTILA CLIVEA DA SILVA MARTINS
RIO GRANDE, RS
AGOSTO, 2015
1

i
UNIVERSIDADE FEDERAL DO RIO GRANDE – FURG
PROGRAMA DE PÓS- GRADUAÇÃO EM AQUICULTURA – PPGAq
INSTITUTO DE OCEANOGRAFIA – IO
TESE DE DOUTORADO
RESPOSTAS ANTIOXIDANTES E DANO OXIDATIVO NO CAMARÃO
Litopenaeus vannamei: EFEITOS DA SUPLEMENTAÇÃO COM ANTIOXIDANTE E
USO DE TECNOLOGIA DE BIOFLOCOS
ÁTILA CLIVEA DA SILVA MARTINS
Orientador: Dr. José María Monserrat (FURG)
Coorientador: Dr. Wilson Wasielesky Jr. (FURG)
Rio Grande – RS – Brasil
Agosto, 2015
Tese apresentada ao Programa de Pós-
graduação em Aquicultura da
Universidade Federal do Rio Grande,
como requisito parcial à obtenção do título
de DOUTOR.

ii
1

ii
SUMÁRIO
DEDICATÓRIA................................................................................... v
AGRADECIMENTOS......................................................................... vii
RESUMO.............................................................................................. viii
ABSTRACT.......................................................................................... x
1 INTRODUÇÃO GERAL..................................................................... 1
2 HIPÓTESES......................................................................................... 10
3 OBJETIVOS......................................................................................... 10
3.1 OBJETIVO GERAL.............................................................................. 10
3.2 OBJETIVOS ESPECÍFICOS................................................................. 10
4 MATERIAL E MÉTODOS GERAL.................................................. 11
4.1 DESENHO EXPERIMENTAL.............................................................. 11
4.1.1 Preparação das nanocapsulas e cápsulas vazias................................ 14
4.1.2 Diluição do ácido lipóico em hidróxido de sódio............................... 14
4.2 ANÁLISE DA ÁGUA........................................................................... 15
4.3 COLETA E HOMOGENEIZAÇÃO DAS AMOSTRAS...................... 16
4.4 DETERMINAÇÃO DA ATIVIDADE DA ENZIMA GLUTATIONA
S-TRANSFERASE (GST) ....................................................................
16
4.5 DETERMINAÇÃO DA ATIVIDADE DA GLUTATIONA
REDUZIDA (GSH) ...............................................................................
17
4.6 DETERMINAÇÃO DOS NÍVEIS DE PEROXIDAÇÃO LIPÍDICA.. 17
4.7 DETERMINAÇÃO DA CAPACIDADE ANTIOXIDANTE TOTAL
CONTRA RADICAIS PEROXIL (ACAP) ..........................................
18
4.8 CONTAGEM DIFERENCIAL DE HEMÓCITOS................................ 19
4.9 ANÁLISE ESTATÍSTICA DOS RESULTADOS.................................. 19
REFERÊNCIAS................................................................................... 21
CAPÍTULO I: Antioxidant and oxidative damage responses in
different organs of Pacific white shrimp Litopenaeus vannamei
(Boone, 1931) reared in a biofloc technology system........................
28
ABSTRACT.......................................................................................... 29
1 INTRODUCTION................................................................................ 30
2 MATERIALS AND METHODS......................................................... 31

iii
2.1 DETERMINATION OF GST ACTIVITY............................................. 32
2.2 DETERMINATION OF TOTAL ANTIOXIDANT CAPACITY........... 32
2.3 DETERMINATION OF CONCENTRATION OF REDUCED
GLUTATHIONE (GSH)........................................................................
33
2.4 DETERMINATION OF LIPID PEROXIDATION............................... 33
2.5 DETERMINATION OF TOTAL AMMONIA....................................... 34
2.6 STATISTICAL ANALYSIS................................................................... 34
3 RESULTS.............................................................................................. 34
4 DISCUSSION....................................................................................... 38
REFERENCES..................................................................................... 41
CAPÍTULO II: Antioxidant effects of nanoencapsulated lipoic
acid in tissues and immune condition in hemolymph of shrimp
Pacific Litopenaeus vannamei (Boone, 1931).....................................
45
ABSTRACT.......................................................................................... 46
1 INTRODUCTION................................................................................ 47
2 MATERIALS AND METHODS......................................................... 50
2.1 DETERMINATION OF GLUTATHIONE S-TRANSFERASE
ACTIVITY (GST)..................................................................................
53
2.2 DETERMINATION OF CONCENTRATION OF REDUCED
GLUTATHIONE (GSH)........................................................................
53
2.3 DETERMINATION OF LIPID PEROXIDATION............................... 53
2.4 DETERMINATION OF TOTAL AMMONIA....................................... 54
2.5 DIFFERENTIAL HEMOCYTE COUNT (DHC) ................................. 54
2.6 STATISTICAL ANALYSIS................................................................... 55
3 RESULTS.............................................................................................. 55
4 DISCUSSION....................................................................................... 56
5 CONCLUSÃO...................................................................................... 61
REFERENCES..................................................................................... 62
CAPÍTULO III: Effects of lipoic acid in the total antioxidant
capacity in biofloc.................................................................................
71
ABSTRACT.......................................................................................... 72
1 INTRODUCTION................................................................................ 72

iv
2 MATERIALS AND METHODS......................................................... 74
2.1 EXPERIMENTAL DESIGN.................................................................. 74
2.2 DILUTION LIPOIC ACID IN SODIUM HYDROXIDE..................... 74
2.3 WATER QUALITY ANALYSIS............................................................ 75
2.4 COLLECTION AND HOMOGENIZATION OF SAMPLES............... 75
2.5 DETERMINATION OF TOTAL ANTIOXIDANT CAPACITY
AGAINST PEROXYL RADICAL (ACAP) .........................................
76
2.6 STATISTICAL ANALYSIS................................................................... 76
3 RESULTS.............................................................................................. 77
4 DISCUSSION....................................................................................... 80
5 CONCLUSION..................................................................................... 81
6 FUTURE PERSPECTIVE.................................................................. 81
REFERENCES..................................................................................... 82
CONCLUSÃO GERAL....................................................................... 85

v
DEDICATÓRIA
À minha mãe (Célia Santana) e a meu pai (Antônio Carlos Martins), entre erros e
acertos não permitiram que o amor incondicional provocasse vícios e dependências, me
deram autonomia, confiança e independência para traçar rumos, fazer escolhas, superar
frustações e cometer meus próprios erros e acertos. Em cada nova fase de minha vida
tivemos perdas e ganhos dos dois lados, porque o amor de pai e mãe é processo de
libertação permanente e esse vínculo não para de se transformar ao longo da vida.
Dando-me dando a certeza a cada dia de que eles, até quando puderem, estarão lá firmes
na concordância ou na divergência, no sucesso ou no fracasso, com o peito aberto para
um aconchego, abraço apertado e conforto em todas as horas. A natureza nos ajuda a
enxergar e compreender a importância das raízes. Quanto mais vigorosas são, maior e
mais forte é a árvore. Para voar alto e livre é preciso ter um terreno firme de onde
decolar. É tudo isso que meus pais representam para mim.
À minha vó (Maria Madalena Santana – em memória), que estava sempre preocupada
com as horas que estávamos sem comer, se andávamos descalços. A mulher que a seu
modo através de olhares e sorrisos conseguia dizer o quanto nos amava. Ainda tenho
vivo na memória nossos últimos momentos juntas. Presenciei alguns sorrisos, alguns
olhares atentos, e percebi, ela ainda estava ali, estava vendo e observando todos nós, por
trás daquela mulher fraca estava ela: A minha vó, a mesma de sempre!
À minha irmã (Carla Martins), que me ajudou a enfrentar o mundo com inteligência,
coragem e sabedoria. Inteligência para encontrar no mundo a oportunidade de mudança
e aprendizado. Coragem para aceitar mudanças. Sabedoria para sorrir, chorar, sem
perder a linha, sem perder o passo. Ensinou-me que algumas situações na vida servem
como cinzel que esculpe, que talha, que faz o bloco amorfo de mármore se transformar
em estátua, em obra de arte.
A Carlos Eduardo Winievisck, quem tem sorriso mais contagiante que já conheci.
Carrega consigo a alegria de viver, que ainda conserva um sotaque irresistivelmente
encantador. E quando estamos juntos a paz chega bem pertinho de mim, os problemas
que embalam os dias e que roubam as energias misteriosamente se escondem como se

vi
por aqueles instantes não mais existissem. Somos apenas nós diante de um cenário
escolhido, jogando palavras, formando conversas e confissões. Quem me trouxe os mais
lindos instantes e me completou com a magnitude de cada olhar. "Como tudo o que não
pode ser tocado com a mão e nem visto como os olhos, e ainda se torna mais forte. As
únicas coisas que importam são as feitas de verdade e alegria, não as de lata e vidro..."
(Fernão Capelo Gaivota).
A Ivis Winievisck, meu amigo foram tantas as vezes que você apareceu no momento
certo, e que mesmo sem querer dizia exatamente o que eu precisava ouvir, como se
pudesse me ler. Nos conhecemos sem que sejam necessárias as palavras. É tudo tão
grande que as diferenças se tornaram pequenos detalhes. Homem de frases lindas que
tanto respondi com brincadeiras tão sinceras, mas que hora ou outra me faz dizer
olhando nos olhos frases tão difíceis de serem ditas pessoalmente por mim. Que me
ensinou a arte de ver a mim mesmo com minhas forças e fraquezas, mas sem máscaras,
sem ilusões. A arte de perceber que as feridas cicatrizam sempre, e que ali a pele se
torna mais resistente.
Ao Prof. Dr. José María Monserrat, que foi meu orientador acadêmico e na vida pessoal
que me ensinou a ver que o que me acontece no presente não vai definir meu futuro nem
quem sou e sim a maneira como eu reajo a tudo que me acontece é o que vai definir
quem eu serei e quem eu quero ser.

vii
AGRADECIMENTOS
Ao meu co-orientador Prof. Dr. Wilson Wasielesky.
A CAPES pelo suporte financeiro através da concessão de bolsa de estudos.
A todos do grupo do Projeto Camarão.
A todos do grupo de Piscicultura Estuarina e Marinha.
Ao grupo EAOx (Espécies ativas de oxigênio), em especial a Josencler, Camilla Porto,
Juliana Artigas.
Agradeço a Jani (Ivanildo), pela ajuda no processo de qualificação do doutorado, pela
companhia na preparação e análise do terceiro trabalho e aos passeios de bicicleta.
Aos amigos que aqui fiz: Paula Maicá, Mércia, Vita, André Braga, Yorleys (Miss
Colômbia), Adriana (Shakira), Viviana Lisboa, Paola, Alain (Aladin), Cecilia (Ceci),
agradeço por todas as vezes que me fizeram fortes, que me fizeram sorrir que me
abraçaram no momento exato, que me ajudaram academicamente e assim renovaram
minhas energias para seguir sempre em frente.
Aos meus amigos de sempre: Thays, Gisa, Aline, Guto, Bernardo, Mila, cada um em seu
momento soberam me dar incentivo a continuar, lutar e superar a distância.

viii
RESUMO 1
Em aquicultura o aumento da intensificação está diretamente ligado a aplicação de 2
novas tecnologias que aumente o volume de produção ao mesmo tempo que causa o 3
mínimo de impacto ao meio ambiente que circunda a produção. Deste modo a 4
tecnologia de biofloco desponta como método no qual o tratamento de qualidade de 5
água é efetuado dentro do tanque de criação de modo que organismos fotoautotrofico 6
(microalgas), autotróficos (bactéria nitrificante) e heterotróficos (bactéria heterotrófica) 7
reciclam compostos nitrogenados que podem vir a ser tóxicos para o camarão 8
Litopenaeus vannamei, além de serem capazes de transformar amônia em biomassa 9
bacteriana que servirá como fonte proteica e lipídica para o camarão, podendo reduzir 10
custo com ração. Com aumento da densidade no sistema de criação faz-se necessário 11
aumentar a resistência bioquímica do animal criado, para tanto este trabalho apresenta 12
em três capítulos meios de suplementação com antioxidante ácido lipóico (AL) que 13
auxilia na resposta bioquímica antioxidante como mecanismo de melhorar o bem estar 14
do camarão Litopenaeus vannamei. Primeiramente, através de análises bioquímicas 15
como atividade da glutationa S-transferase (GST), concentração de glutationa reduzida 16
(GSH), capacidade antioxidante total contra radicais peroxil (ACAP) e níveis de 17
peroxidação lipídica (TBARS) em brânquias, hepatopâncreas e músculo de camarão, 18
observando-se que o biofloco induz aumento da atividade da GST em brânquias, 19
aumento da concentração de GSH em músculo, aumenta a capacidade antioxidante total 20
em músculo e reduz níveis de peroxidação lipídica em hepatopâncreas. O segundo 21
trabalho, aplicou-se ácido lipóico nanoencapsulado (NCLA) e cápsula vazia (NC) na 22
ração, em água clara e em água com biofloco no qual foi observado que NCLA induzida 23
aumento da atividade de GST nos hepatopâncreas. A concentração de GSH foi maior no 24
músculo do que em brânquias e hepatopâncreas. A capacidade antioxidante também 25
mostrou um padrão tecido-específico, tendo hepatopâncreas com maior capacidade 26
antioxidante nenhuma ação evidente do desempenho do NCLA contra os radicais 27
peroxil. Níveis de peroxidação lipídica foram menores no músculo, com acentuado 28
efeito do NCLA. Nos grupos com NCLA houve um aumento na porcentagem de 29
hemócitos granulares, células com maiores quantidades de componentes 30
imunocompetentes. No trabalho 3, foi observado que o AL é capaz de aumentar a 31
capacidade antioxidante no biofloco, analisado através da determinação da capacidade 32

ix
antioxidante total contra radicais peroxil (ACAP), principalmente para concentração de 33
10 µM (2.06 mg de AL in 1 L de água destilada). 34
35
Palavras chave: Biofloco, Litopenaus vannamei, antioxidante, ácido lipoíco, análises 36
bioquímicas, nanotecnologia, contagem diferencial de hemócitos. 37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56

x
ABSTRACT 57
In aquaculture, the increased intensification is directly linked to application of new 58
technologies to increase the volume of production while causing minimal impact to the 59
environment surrounding the production. Thus, the biofloc blunts as a new technology 60
method in which the treatment water quality is made inside the tank, so that creation of 61
photoautotrophic organisms (microalgae), autotrophs (nitrifying bacteria) and 62
heterotrophic (heterotrophic bacteria) recycle nitrogen compounds that may to be toxic 63
to shrimp Litopenaeus vannamei. These microorganisms are able to turn ammonia into 64
bacterial biomass that will serve as protein and lipid source for shrimp and may reduce 65
feed cost. With increased density in the build system it is necessary to increase the 66
resistance biochemistry of the animal created, therefore this work presents in four 67
chapters how of supplementation with the antioxidant lipoic acid (LA) can be assist in 68
antioxidant biochemical response as a mechanism to improve the wellness of shrimp 69
Litopenaeus vannamei. Firstly, by biochemical analyzes as gluthatione S-transferase 70
(GST) activity, reduced gluthatione (GSH) concentration, total antioxidant capacity 71
against peroxyl radicals (ACAP) and levels of lipid peroxidation (TBARS) in gills, 72
hepatopancreas and shrimp muscle, observing that the biofloc induces increased in GST 73
activity in gills, increased GSH concentration in muscle, increases the total antioxidant 74
capacity in muscle and reduces lipid peroxidation levels in hepatopancreas. The second 75
work was applied nanoencapsulated lipoic acid (NCLA) and empty nanocapsules (CN) 76
in feed in clean water (SW) and biofloc (BFT) in which it was observed that NCLA 77
induced increased GST activity in hepatopancreas. The GSH concentration was higher 78
in muscle than in gills and hepatopancreas. The antioxidant activity also showed a 79
specific pattern of tissue having higher antioxidant capacity in hepatopancreas, without 80
no obvious action of NCLA in performance against peroxyl radicals. Lipid peroxidation 81
levels were lower in the muscle, with marked effect of NCLA. In groups with NCLA 82
there was an increase in the percentage of granular hemocytes, cells with higher 83
amounts of immunocompetents components. In the work 3, it was observed that LA can 84
increase the antioxidant capacity on biofloc, analyzed by determining the total 85
antioxidant capacity against peroxyl radicals (ACAP), especially for concentration 10 86
µM (2.06 mg AL in 1 L of water distilled). 87

xi
Keywords: Biofloc, Litopenaus vannamei, antioxidant, lipoic acid, biochemistry, 88
nanotechnology, differential count of hemocytes. 89
90

1
1 INTRODUÇÃO GERAL 91
Várias pesquisas no setor de aquicultura são destinadas ao aumento da 92
intensificação da produção. Estes esforços englobam maximizar as atividades na relação 93
aquicultura e ambiente externo, como também o aumentar a sustentabilidade 94
(econômica, social e ambiental). Ainda deve ser considerada a relação entre aquicultura 95
e ambiente interno, que corresponde a potencializar o crescimento e sobrevivência do 96
camarão, em menores ciclos de produção e com rendimento e qualidade do produto 97
final. A carcinicultura se tornou uma grande atividade aquícola e sua expansão em todo 98
mundo tem aumentado a preocupação ambiental em torno desta atividade (Xu et al. 99
2013). Tendo o camarão branco do Pacífico, Litopenaeus vannamei (Boone 1931), a 100
espécie com maior volume de produção (Kim et al. 2014) por apresentar rápido 101
crescimento, maior índice de sobrevivência e ser tolerante a alta densidade e estocagem 102
(Xu et al. 2012). 103
A produção mundial cresceu a uma taxa de 15,1% entre 2000 e 2008, que se 104
deve a intensificação da produção de Litopenaeus vannamei na China, Tailândia e 105
Indonésia (FAO 2012). Em 2014, a produção mundial de camarão em cativeiro 106
aumentou para 3.680.404 toneladas, um aumento de 7% em relação aos 3.436.918 de 107
toneladas produzidas em 2013, com base em estimativas obtidas de fontes oficiais e na 108
sua ausência de fontes oficiais, as estimativas fornecidas por fontes da indústria, a 109
produção brasileira se estabilizou entre os anos de 2013 e 2014 em 90 mil toneladas 110
(Shrimp News International 2015). 111
O camarão branco Litopenaeus vannamei é a espécie de camarão mais 112
comercializada em muitas partes do mundo e sua produção em sistemas intensivos de 113
biofloco com pouca ou nenhuma troca de água tem demostrado ser uma prática 114
sustentável (Avnimelech 2012; Xu e Pan 2014), especialmente em aquicultura intensiva 115
que está ligada com a poluição da água por um excesso de materiais orgânicos e 116
nutrientes que são susceptíveis de causar efeitos tóxicos agudos e riscos ambientais de 117
longo prazo (Piedrahita 2003). Em um sistema aquícola convencional o método mais 118
comum para lidar com este tipo de poluição tem sido a substituição contínua da água do 119
tanque com água fresca externo (Gutierrez-Wing e Malone, 2006). No entanto, o 120

2
volume de água necessário para pequeno a médio tanque aquícola pode chegar a várias 121
centenas de metros cúbicos por dia. 122
Uma nova abordagem é uso da tecnologia de biofloco (BFT), que é a formação e 123
estimulação de um microecossistema que incluem microalga, bactérias autotróficas, 124
bactérias heterotróficas, detritos orgânicos e inorgânicos. Desta forma, processos de 125
renovação de água neste sistema é mínima ou zero, havendo, portanto, reutilização da 126
água e alguns riscos, como a introdução de agentes patogénicos, escapamento de 127
espécies exóticas e descarga de águas residuais (poluição) são reduzidos ou mesmo 128
eliminados (Ray 2012). 129
Estes microrganismos (biofloco) tem como funções principais: (i) manutenção 130
da qualidade da água, pela absorção de compostos nitrogenados e sua transformação em 131
proteína microbiana e (ii) nutrição que aumenta viabilidade econômica da produção, 132
reduzindo a conversão alimentar e uma diminuição dos custos de alimentação 133
(Emerenciano 2013). Os macroagregados (biofloco) é fonte natural rica em lipídeos e 134
proteínas, disponível in situ 24 horas por dia (Avnimelech 2007). Na coluna de água 135
ocorre uma complexa interação entre matéria orgânica, substrato físico e grande 136
variedade de microrganismos, como fitoplâncton, bactérias livres e aderidas, agregados 137
de partículas de matéria orgânica e herbívoros, como os rotíferos, ciliados e flagelados 138
protozoários e copépodes (Ray 2010). 139
Esta produtividade natural tem um papel importante na reciclagem de nutrientes 140
e na manutenção da qualidade de água. Bactérias autotróficas fazem a conversam de 141
amônia a nitrito e depois convertem nitrito a nitrato e, as bactérias heterotróficas 142
conseguem compostos nitrogenados e transformam em proteína microbiana. As 143
bactérias autotróficas são mais eficientes nesta conversão, porém o processo é feito de 144
forma lenta, e as bactérias heterotróficas tem crescimento mais rápido e, portanto, 145
retiram de forma mais rápida os nitrogenados e transformam em proteína microbiana, 146
por isso há a manipulação da taxa de carbono e nitrogênio na proporção 20:1 para 147
favorecer crescimento e domínio de bactérias heterotróficas (Avnimelech 1999). 148
O consumo de biofloco por camarão ou peixe tem demonstrado inúmeros 149
benefícios tais como a melhoria da taxa de crescimento, diminuição da taxa de 150
conversão alimentar e os custos associados em alimentos para animais (Buford et al. 151

3
2004; Wasielesky et al. 2006). A melhoria do crescimento tem sido atribuída a bactérias 152
e algas como componentes nutricionais, pelo qual até 30% da ração comercial pode ser 153
reduzido devido ao consumo de biofloco pelo camarão (Buford et al. 2004). E o uso de 154
biofloco pode ser uma alternativa para substituir o uso de proteínas alternativas como a 155
farinha de peixe (Azim 2008). 156
Estudo tem demostrado que além do uso manutenção da qualidade de água e 157
como fonte de proteína e lipídio, o biofloco também tem o efeito de manter o equilíbrio 158
das funções fisiológicas como sistema antioxidante, que é essencial para a manutenção 159
do bem estar do camarão e assim garantir crescimento e sobrevivência satisfatórios 160
(Castex et al. 2010; Xu e Pan 2014; Martins et al. 2014, 2015) em um ambiente de 161
criação. Estudos efetuados com Litopenaues vannamei indicaram que o biofloco pode 162
aumentar o estado antioxidante do camarão, com causa provável do biofloco ser rico em 163
microrganismos naturais e compostos bioativos de natureza antioxidante (Ju et al. 2008; 164
Xu e Pan 2013; Martins et al. 2015). 165
A espécie Litopenaeus vannamei vem largamente sendo afetada por doenças 166
(Kim et al. 2014). Muitas doenças são agravadas pela alteração do equilíbrio 167
bioquímico celular e pouco se conhece sobre os benéficos bioquímico/fisiológicos do 168
biofloco e seus efeitos no camarão. O desequilíbrio bioquímico, em espécies aeróbicas, 169
pode ocorrer quando há maior produção de espécies reativas de oxigênio (ERO), que 170
são produtos intermediários da redução parcial dos quatro elétrons do oxigênio 171
resultando em água (H2O), ânion superóxido (O2•-), radical hidroxila (OH•) e espécies 172
não radicalar como o peróxido de hidrogênio (H2O2) (Abele e Pintarulo 2004). Por 173
definição redução é a perda de oxigênio ou ganho de elétrons, desta forma o O2 sofre 174
redução tetravalente com ganho de quatro elétrons e formando H2O (Gutteridge e 175
Halliwell 2010) e neste processo os produtos intermediários reativos (O2•-, H2O2, OH•) 176
podem ser nocivos quando o sistema antioxidante não é capaz de controlá-los, situação 177
que pode derivar em estresse oxidativo (Sies 1985; Abele e Pintarulo 2004). 178
O estresse oxidativo é um estado de desbalanço entre a produção intra e 179
extracelular de ERO e o sistema antioxidante, resultando em dano oxidativo de muitos 180
tipos de moléculas como lipídios, proteínas e DNA. Desta forma, a necessidade de 181
prevenir, interceptar ou retardar as ações das ERO, no decorrer da evolução, acarretou 182

4
no desenvolvimento de defesas antioxidantes, comumente divididas em enzimáticas e 183
não-enzimáticas (Anderson 1998; Dickinson e Forman 2002). O sistema antioxidante 184
enzimático é o primeiro mecanismo de defesa celular e é composto por superóxido 185
dismutase (SOD), catalase (CAT), glutationa peroxidase (GPx) e glutationa-S-186
transferase (GST) que são moléculas de maior peso molecular, somado a estes há a ação 187
de outros compostos antioxiantes como vitaminas A, C e E que têm menor peso 188
molecular (Hellou et al. 2012). Dentre as defesas antioxidantes não-enzimáticas o 189
tripeptídeo glutationa (γ-L-glutamil–L-cisteinil–glicina) é considerado a primeira ação 190
de defesa contra ERO (Anderson 1998; Dickinson e Forman 2002) e está presente nos 191
organismos nas formas reduzidas (GSH) e oxidada (GSSG) (Hellou et al. 2012). Além 192
deste, inclui-se também α-tocoferol, carotenóides e flavonoides (Barreiros et al. 2006) e 193
o ácido lipóico (AL) que é um dos focos desta Tese. 194
Acredita-se que as mitocôndrias consumam 90% do oxigênio celular em células 195
átonas (jovens) e são os principais locais de produção de ERO em células aeróbicas 196
(Lenaz 1998; Abele e Pintarulo 2004). Na redução univalente, o O2 (oxigênio) é 197
convertido a O2•- (radical superóxido), que por ação da enzima superóxido dismutase 198
(SOD) é convertido em H2O2 (peróxido de hidrogênio), que é um ERO mas não é um 199
radical livre, este composto é difundo livremente através da mitocôndria de forma 200
espontânea (Abele e Pintarulo 2004). Caso o H2O2 não seja decomposto 201
enzimaticamente, este pode ser convertido em OH• (radical hidroxila), que tem curta 202
vida por ser altamente reativo (Halliwell e Gutteridge 1985; Abele e Pintarulo 2004). 203
Alguns componentes (poluentes) podem ser difíceis de oxidar e, portanto, é necessário 204
que o H2O2 seja ativado por catalizadores (ferro, cobre, manganês), sendo mais comum 205
utilizar o ferro como catalizador que quando reage com H2O2 caracteriza a reação de 206
Fenton que requer pH ácido e produz radicais hidroxila (OH•) que são altamente 207
reativos que degradam poluentes orgânicos (Wang et al. 2012). O H2O2 é convertido em 208
água e oxigênio, cuja reação é catalisada pela enzima catalase (CAT) ou é utilizado para 209
oxidar substratos, como por exemplo peroxidases como glutationa peroxidase (GPx) 210
(Hellou et al. 2012). 211
Um estado redox pró-oxidante é caracterizado com uma queda relativa na 212
proporção de glutationa reduzida (GSH/GSSG) e da relação NADH/NAD (Abele e 213

5
Pintarulo 2004). A glutationa é uma molécula produzido naturalmente pelo fígado, 214
também é encontrada em frutas, verduras e carnes, sendo uma combinação de três 215
blocos de proteína ou aminoácidos (tripéptido – cisteína, glicina e glutamina) que 216
contém um grupo químico de enxofre (SH), que atua como atrativo para moléculas que 217
podem causar dano ao organismo como espécies reativas de oxigênio e xenofibiótico 218
(Nuttall et al. 1998, Huber et al. 2008). Normalmente a glutationa é reciclada no corpo, 219
exceto quando há sobrecarga com muito estresse oxidativo ou muitas toxinas, a 220
glutationa se esgota e diminui a proteção contra os radicais livres ou toxinas (Nuttall et 221
al. 1998, Huber et al. 2008). A rede antioxidante é composta por vários componentes 222
que incluem vitaminas, minerais e produtos químicos especiais chamados tióis 223
(glutationa e o ácido alfa-lipóico) (Huber et al. 2008). 224
A glutationa é um antioxidante intracelular que tem a capacidade de maximizar a 225
atividade de todos os outros antioxidantes, incluindo vitaminas C, vitamina E e ácido 226
lipóico, removendo toxinas das células e protegendo contra os efeitos nocivos da 227
radiação, produtos químicos e poluentes ambientais (Schafer e Buettner 2001; 228
Dickinson e Forman 2002; Huber et al. 2008). O AL é uma molécula com características 229
hidro e lipossolúvel, com múltiplos efeitos benéficos em doenças como diabetes, 230
Alzheimer e hipertensão, o que mostra seu grande potencial biomédico (Packer et al. 231
1995). O AL e sua forma reduzida, o ácido dihidrolipóico (DHLA), preenchem todos os 232
critérios avaliados na análise do potencial antioxidante de um composto: quelam metais, 233
são varredores de ERO, participam da reciclagem de outras moléculas antioxidantes e 234
do reparo de moléculas danificadas pelo estresse oxidativo (Packer et al. 1995). O ácido 235
lipóico a princípio foi classificado como vitamina, contudo posteriormente foi 236
constatado que este composto é sintetizado em células animais (Carreau 1979), atuando 237
como cofator em complexos multi-enzimáticos que catalisam reações de 238
descarboxilação oxidativa no ciclo de Krebs (Packer et al. 1995). 239
Um antioxidante pode ser definido como qualquer substância que quando 240
presente em baixas concentrações em relação ao substrato oxidável (que causa dano), 241
atrasa significativamente ou impede a ação danosa do referido substrato (Halliwell e 242
Getturidge 1995). Nas últimas décadas o ácido lipóico tem recebido atenção devido a 243
sua função antioxidante em organismos aquáticos (Monserrat et al. 2008). O papel do 244

6
ácido lipóico no reestabelecimento dos níveis de ácido ascórbico em pacu (Piaractus 245
mesopotamicus) foi importante do ponto de vista bioquímico e fisiológico, o pacu, 246
assim como outros teleósteos, não sintetiza o ácido ascórbico, sendo este, portanto, uma 247
vitamina. A deficiência dela pode induzir uma redução de crescimento e maior 248
suscetibilidade a vários tipos de doenças (Terjesen et al. 2004; Trattner et al. 2007). Em 249
truta arco-íris (Oncorhynchus mykiss) e carpas (Cyprinus carpio) a deficiência de 250
vitamina C provoca anorexia, lordose, escoliose, hemorragias, deformações em 251
brânquias, exoftalmia (Trattner et al. 2007). 252
A vitamina C quela o ferro e o reduz a Fe2+, subsequentemente, o Fe2+ pode 253
transferir um elétron ao oxigênio ou para outro ERO e induzir o estresse oxidativo, 254
porém em quantidades equimolares de ferro e vitamina C, ácido lipóico é capaz de 255
competir com a vitamina C para a quelação e, consequentemente, haver proteção contra 256
a peroxidação de lípidos (Biewenga et al. 1997). O ácido lipóico tem sido referido como 257
um antioxidante universal por atuar tanto na membrana quanto na fase aquosa das 258
células, proporcionando proteção a membrana, devido a sua interação com os 259
antioxidantes vitamina C e glutationa, as quais por sua vez podem reciclar a vitamina E 260
(Flora 2009). As propriedades do ácido lipóico incluem também a capacidade de varrer 261
ERO, além de regenerar antioxidantes endógenos (Packer et al. 1995; Flora 2009; 262
Külkamp-Guerreiro et al. 2009). 263
O trabalho de Amado et al. (2011) avaliou o efeito quimioprotetor do ácido 264
lipóico contra a toxidade de microcistina em carpa Cyprinus carpo. Estes autores 265
observaram o tempo necessário para indução na expressão de genes que codificam três 266
classes da glutationa-S-transferase (alfa, mu e pi). Os resultados constataram que o AL 267
foi eficaz em promover aumento na transcrição de genes da GST no fígado após duas 268
injeções de AL dadas com intervalo de 24 h. Os dados sugerem que AL pode ser útil 269
como agente quimioprotetor contra indução toxica da microcistina, estimulando a 270
desintoxicação através do incremento da atividade da GST (cérebro) ou por meio da 271
reversão da inibição da GST (fígado). 272
O trabalho de Monserrat et al. (2008), considerou os efeitos do AL em diferentes 273
órgãos (brânquias, cérebro, músculo e fígado) no peixe Corudoras paleatus 274
(Callychthyidae). O AL, na dose de 70 mg/kg de massa corporal foi adicionada na 275

7
ração, alimentados diariamente (1% do peso). Os resultados mostraram redução da 276
concentração de espécies reativas de oxigênio no cérebro e aumento da atividade do 277
glutamato-cisteína ligase (GCL) no cérebro e no fígado do mesmo grupo experimental. 278
A GCL é uma enzima que controla a velocidade da síntese de GSH. Organismos 279
suplementados com AL apresentaram maior atividade da glutationa-S-transferase no 280
cérebro, indicando que o AL melhora a capacidade de desintoxicação nas reações de 281
fase II. Foi observado também notável redução da oxidação de proteínas no músculo e 282
no fígado dos peixes suplementados com AL, indicando que o tratamento foi eficaz na 283
redução de parâmetros de estresse oxidativo. 284
O organismo do camarão, como qualquer organismo aeróbico, é susceptível a 285
estresse oxidativo, resultado da ação de ERO que podem ser provenientes do meio 286
ambiente (exógenas) ou gerada no próprio organismo (endógenas). No camarão 287
Litopenaeus vannanmei, foi verificado o efeito de 3 doses de AL (35, 70 e 140 mg de 288
AL por 1 kg de ração), que foi suplementada na ração durante 45 dias e análises de 289
espécies reativas de oxigênio foi efetuada em brânquias e hepatopâncreas. Constatou-se 290
que das 3 doses aplicadas a dose de 70 mg/kg foi a que melhor resultou no aumento da 291
atividade antioxidante, principalmente em brânquias (Martins et al. 2014). 292
A utilização do ácido lipóico é promissora, contudo esta substância é lábil e sem 293
estabilidade química e, portanto, sujeito a degradação por ação térmica, fotoquímica, 294
meio ácido e oxidação. Sendo assim, existem pesquisas desenvolvendo um complexo de 295
ácido lipóico e ciclodextrina, que apresentam maior estabilidade à temperatura e a luz, o 296
que reduz a dispersão do AL em água, aumenta a biodisponibilidade e reduz seu odor, 297
no entanto, estes complexos apresentaram ampla distribuição de tamanho e aumento de 298
tamanho das partículas após duas semanas de armazenamento à temperatura ambiente 299
(Külkamp-Guerreiro et al. 2009). A alternativa ainda pouco explorada para a estabili-300
zação do ácido lipoico é o emprego de nanocápsulas poliméricas, as quais consistem em 301
sistemas vesiculares nanoestruturados carreadores de substâncias que apresentam 302
diâmetros entre 200 e 300 nm e baixa polidispersão, e que se destacam devido às suas 303
potencialidades no controle da liberação de substâncias e à capacidade de aumentar a 304
estabilidade do composto, tanto no armazenamento quanto nos fluídos biológicos 305
(Külkamp-Guerreiro et al. 2009). O termo nanotecnologia foi inserido em 1974, para 306

8
descrever a manipulação de partículas de menos de um micrômetro. Em particular, o 307
processo de nanoencapsulado é eficiente no desenvolvimento de produtos funcionais e 308
pode auxiliar a combater a perda de funcionalidade dos antioxidantes ou produtos 309
bioativos durante o processamento ou armazenamento, geração de maus odores e 310
sabores, entre outros problemas (Quintanilla-Carvajal et al. 2010). 311
A utilização de nanocápsula é descrito para proteção de diferentes sistemas 312
aplicados em fármacos e cosméticos, especialmente em substâncias que degradam em 313
temperaturas acima de 40 ºC ou são sensíveis à oxidação em presença de água, por 314
variação de pH ou por efeito de luz ultravioleta (Müller et al. 2004, Külkamp-Guerreiro 315
et al. 2009). A membrana polimérica da nanocápsula possui efeito protetor de 316
substâncias contra danos causados por agentes externos, prevenindo a degradação. 317
(Bauchemal et al. 2006, Weiss-Angeli et al. 2008, Külkamp-Guerreiro et al. 2009). Para 318
este estudo, a análise das condições do sistema antioxidante do biofloco e do camarão 319
Litopenaues vannamei envolveram as análises da atividade da glutationa S-transferase 320
(GST), concentração da glutationa reduzida (GSH), níveis de peroxidação lipídica 321
(TBARS), análise da capacidade antioxidante contra radicais peroxil (ACAP) e 322
contagem diferencial de hemócitos para análise do estado imune. 323
A glutationa-S-transferase é uma enzima multifuncional que está envolvida na 324
desintoxicação de xenobióticos, oferecendo proteção contra danos oxidativos e efetúa 325
também transporte intracelular de hormônios, metabólitos endógenos e exógenos de 326
produtos químicos em diversos organismos. Assim a GST é componente importante de 327
várias vias de desintoxicação e tolerância ao estresse, uma vez que protege contra lesões 328
induzidas por substâncias químicas ambientais (Zhou et al. 2009). 329
A glutationa (GSH) é um antioxidante muito importante na preservação do 330
estado redox celular, na defesa contra ERO e detoxificação de xenobióticos. Está 331
molécula é um tripeptídeo composto por ácido glutâmico, cisteína e glicina, que sob 332
condições normais e niveis de cisteína adequados a taxa limitante para sua sintese é 333
determinada pela atividade da enzima glutamato cisteína ligase (GCL) (White et al. 334
2003). 335
Quando em condições normais ou anormais a produção de ERO ultrapassa a 336
proteção endógena de enzimas específicas e vitaminas antioxidantes ocorre dano 337

9
celular, fenômeno este chamado de estresse oxidativo (Oakes e Van Der Kraak 2003). 338
As EROs atacam radicais livres como ácidos graxos poliinsaturados (PUFAs) que são 339
um substrato rico em elétrons (Esterbauer 1996; Oakes e Van Der Kraak 2003). Este 340
procedimento então, é realizado pela quantificação de compostos como o 341
malondialdeído (MDA), que é um subproduto da peroxidação lipídica (Janero 1990; 342
Oakes e Van Der Kraak 2003). Portanto, a reação do MDA com o ácido 2-tiobarbitúrico 343
(TBA), ensaio este conhecido como TBARS, é um dos mais amplamente utilizados 344
como estimadores de estresse oxidativo que analise os níveis de peroxidação lipídica 345
dos tecidos (Liu et al. 1997; Oakes e Van Der Kraak 2003). 346
Embora seja importante medir a eficência de antioxidante individuais para 347
combater a produção de oxiradicais, faz-se necessário compreender a resistência dos 348
tecidos a toxidade causada por ERO e não somente medir um numero limitado de 349
antioxidantes (Amado et al. 2009). Por isto usa-se a análise da capacidade antioxidante 350
contra radicais peroxil (ACAP), que é um método simples, rápido e confiável na 351
detecção de ERO por fluorometria, se valendo do 2,7´ diclorofluresceína diacetato 352
(H2DCF-DA) como substrato o qual, após sua deacetilação, que irá a interagir com 353
radicas peroxil que são gerados pela decomposição térmica a 37°C do 2,2- azobis 354
(2metilpropianoamidina) dihidrocloreto (ABAP). Uma queda na fluorescência nestas 355
condições é interpretada como um efeito antioxidante da amostra, através da 356
interceptação ou redução dos peroxi radicais, gerando um menor sinal de fluorescência 357
emitida pela reação entre EROs e H2DCF (Amado et al. 2009). 358
359
360
361
362
363
364
365
366
367

10
2 HIPÓTESES 368
- Camarões criados em sistema de biofloco terão sua maior capacidade antioxidante, em 369
função dos antioxidantes presentes no biofloco. 370
- O ácido lipóico aumentará a capacidade antioxidante e diminuir o dano oxidativo no 371
camarão branco do pacífico Litopenaus vannemei, visto os efeitos já descritos deste 372
antioxidante em espécies aquáticas. 373
- O ácido lipóico aumentará a competência antioxidante do biofloco, vista suas 374
caraterísticas descritas e definidas como de “antioxidante ideal”. 375
376
3 OBJETIVOS 377
3.1 OBJETIVO GERAL 378
Avaliar se a suplementação com ácido lipoíco na ração e no biofloco altera do 379
estado antioxidante do camarão L. vannemei e do biofloco através de análise 380
bioquímicas que contemplem respostas antioxidantes, de detoxificação e de dano 381
oxidativo. 382
383
3.2 OBJETIVOS ESPECÍFICOS 384
- Avaliar os efeitos antioxidantes do biofloco em brânquia, hepatopâncreas, músculo do 385
camarão L. vannemei por meio da determinação a atividade da glutationa S-transferase 386
(GST) e glutationa reduzida (GSH), bem como avaliar o dano oxidativo dos tecidos pela 387
análise dos níveis de peroxidação lipídica (TBARS) e capacidade antioxidante total 388
contra radicais peroxil (ACAP). 389
390
- Avaliar o estado redox dos tecidos (brânquia, hepatopâncreas, músculo) após 391
suplementação do ácido lipóico (AL) na ração do camarão L. vannemei por meio da 392
determinação a atividade da glutationa S-transferase (GST) e glutationa reduzida 393
(GSH), bem como avaliar o dano oxidativo dos tecidos pela análise dos níveis de 394
peroxidação lipídica (TBARS) e contagem diferencial de hemócitos (CDH). 395

11
- Avaliar os efeitos do ácido lipóico no biofloco através da análise da capacidade 396
antioxidante total contra radicais peroxil, capacidade dos compostos em atuar como 397
varredores de espécies reativas de oxigênio (ERO). 398
399
4 METODOLOGIA GERAL 400
4.1 DESENHO EXPERIMENTAL 401
Os camarões utilizados foram juvenis da espécie Litopenaeus vannamei, 402
provenientes dos viveiros da Estação Marinha de Aquacultura (EMA), Universidade 403
Federal do Rio Grande – FURG. Os animais foram aclimatados em 2 tanques de fibra de 404
vidro (1000 L) com volume útil de 800 L, 150 animais em cada tanque foram estocados 405
a aclimatados durante 25 dias (6 a 30 de maio). 406
Após estes dias, foram submetidos a biometria e transferidos para tanques de 407
plástico (70 L) com volume útil de 50 L. Foram separados para primeira publicação em 408
tratamentos: (i) água clara e (ii) biofloco (Figura 1). Para o segundo manuscrito os 409
tratamentos foram: (i) água clara/nanocapsula vazia, (ii) água clara/AL 410
nanoencapsulado, (iii) biofloco/nanocapsula vazia e (iv) biofloco/AL nanoencapsulado 411
(Figura 2). Os camarões foram estocados a uma densidade de 15 camarões por tanque 412
(cerca de 300 camarões por m3) e passaram 7 dias aclimatados nessa condição (31 de 413
maio a 6 de junho), a partir de então foram ofertadas as respectivas rações. Nos tanques 414
com água clara os animais apresentaram peso médio inicial de 5,91 ± 0,07 g, nos 415
tanques com biofloco o peso médio inicial foi de 5,01 ± 0,05 g e quando houve aumento 416
de amônia acima de 1 mg/L foi adicionado melaço. 417
418
Figura 1 – Desenho experimental trabalho 1. 419

12
420
Figura 2 – Desenho experimental trabalho 2. 421
422
A ração comercial SUPRA® (35% de proteína bruta) foi pesada, em seguida 423
trituradas, misturadas a respectivas soluções (Tabela 1) para obtenção de uma massa 424
homogenia, passada em seringa de 5 mL e produto foi colocado em estufa a 50°C e 425
depois peleitizada. A dieta foi administrada três vezes ao dia (8:00, 15:00 e 22:00 h) a 426
uma taxa alimentar de 3% da biomassa do camarão. O experimento teve duração de 30 427
dias (7 de junho a 6 de julho), nos quais nos tanques com água clara houve renovação 428
de 80-90 % de água a intervalo de 1 dia e nos tanques com biofloco não houve 429
renovação de água. 430
431
Tabela 1 – Quantidades de ração e solução em cada tratamento. 432
Tratamento Ração (g) Solução (mL)
Água clara 500 200 (água destilada)
Biofloco 500 200 (água destilada)
Água clara/cápsula vazia 500 200 (cápsula vazia)
Água clara/AL nanoencapsulado 500 200 (AL nanoencapsulado)
Biofloco/cápsula vazia 500 200 (cápsula vazia)
Biofloco//AL nanoencapsulado 500 200 (AL nanoencapsulado)
433

13
Para o terceiro trabalho, foi realizado a suplementação do ácido lípoico na água 434
com biofloco. Neste caso foram utilizados 15 tanques de plásticos (70 L), com volume 435
útil de 50 L cada. Os tratamentos foram fixados em: Controle (0 mg de AL/L de 436
biofloco), Concentração de 1,25 µM (0,26 mg/L), Concentração de 2,5 µM (0,51 mg/L), 437
Concentração de 5 µM (1,03 mg/L) e Concentração de 10 µM (2,06 mg/L); todos em 438
triplicada (Figura 2). 439
Figura 2 – Desenho experimental do trabalho 3. 440
441
O primeiro trabalho, comparando água clara e biofloco foi publicado na revista 442
Marine and Freshwater Behaviour and Physiology, com título “Antioxidant and 443
oxidative damage responses in different organs of Pacific white shrimp Litopenaeus 444
vannamei (Boone 1931) reared in a biofloc technology system”. O segundo trabalho 445
comparando tratamentos com nanocapsula vazia e ácido lipóico nanoencapsulado foi 446
submetido a revista Comparative Biochemistry and Physiology e é intitulado: 447
“Antioxidant effects of nanoencapsulated lipoic acid in tissues and immune condition in 448
hemolymph of shrimp Pacific Litopenaeus vannamei (Boone, 1931)”. O terceiro 449
trabalho, com título de “Effects of lipoic acid in the total antioxidant capacity in 450
biofloc”, será submetido a revista Aquaculture. 451
452
453

14
4.1.1 Preparação das Nanocapsulas e Nanocapsulas vazias 454
Foi realizado segundo Longaray-Garcia et al. (2013), no qual a suspensão de 455
nanocápsulas com ácido lipóico foram preparados pelo método de precipitação do 456
polímero pré-formado. O ácido lipóico foi pesado (70 mg) e dissolvido na fase orgânica 457
composta por triglicéridos caprílico (0,33 mL), monoestearato de sorbitano (76,6 mg), 458
poli (ξ-caprolactona) (100 mg), acetona (26,7 mL) e butil-hidroxi-tolueno (BHT) (0,01 459
g). A fase orgânica foi injetada em fase aquosa contendo polissorbato 80 (76,6 mg), 460
diazolidinil ureia (0,01 g) e água Milli-Q (53,3 mL), através de um funil e mantida sob 461
agitação magnética moderada durante 10 min. A suspensão foi preparada protegida da 462
luz, e os solventes foram evaporados em evaporador rotativo (Bu chi R-114) a uma 463
temperatura de aproximadamente 30 °C até um volume final de 10 mL, para dar uma 464
concentração final de 70 mg mL-1. As suspensões foram nanocápsulas por espessão 465
com o emulsionante de silicone DC RM2051® (4 g) e Unistab S69® (0,5g). Somente 466
suspensão foi utilizada para o tratamento com nanocapsula vazia. 467
468
4.1.2 Diluição do Ácido Lipóico em Hidróxido de Sódio 469
As concentrações fixadas de α-ácido lipóico sintético (> 99% pureza, Sigma-470
Aldrich), foram dissolvidas individualmente para cada tanque, em solução de hidróxido 471
de sódio e água destilada para garantir a diluição total do ácido lipóico seguindo a 472
metodologia de Amado et al. (2011). Após a diluição o pH foi ajustado para 7,90. As 473
concentrações utilizadas foram adicionadas a cada 24 horas (Tabela 2) 474
475
476
477
478
479

15
Tabela 2: Preparação de solução de ácido lipóico, NaOH (hidróxido de sódio; 2 mM), 480
ddH2O (água destilada), AL (ácido lipóico). A partir do cálculo geral de 300 mL de 481
NaOH para 1g de AL e 1000 mL de H2Odd para 80 g de NaOH. 482
Tratamento NaOH (g) ddH2O (mL) AL (g)
Controle
1,25 µM
2,5 µM
5 µM
10 µM
0
0,62
1,22
2,47
4,94
0
7,8
15,3
30,9
61,8
0
0,26
0,51
1,03
2,06
483
4.2 ANÁLISE DA ÁGUA 484
Diariamente os parâmetros físicos e químicos da água foram monitorados, 485
incluindo oxigênio dissolvido (mg/L) e temperatura da água (°C) com multiparâmetro 486
YSI, salinidade com refratômetro ótico (Atago 103, ±1 ppt), pH com eléctrodo Mettler 487
Toledo FEP20 – FiveEasy Plus™. As determinações de alcalinidade (mg de CaCO3/L) e 488
nitrato (mg/L) foram feitas pelo método APHA (1985) e as de amônia total (NH3 + 489
NH4+ mg/L; UNESCO 1983) e nitrito (mg/L) de acordo com Benderschneider e 490
Robinson (1952) (Trabalho 1, 2 e 3). Quando a amônia foi superior a 1 mg/L foi 491
adicionado melaço de cana de açúcar como fonte de carbono para ajustar a relação C/N 492
de 20:1, segundo métodos de Ebeling et al. (2006) e Avnimelech (1999), nos quais 493
determinaram que 6 g de carbono é necessário para converter 1 g de nitrogênio 494
amoniacal total em biomassa bacteriana. 495
Os sólidos sedimentáveis (material particulado orgânico e inorgânico) foram 496
determinados pela sedimentação dos sólidos em cones Imhoff, onde coloca-se 1 L de 497
água e deixa-se repousar por 1 h, momento o qual são lidos os sólidos suspensos em 498
mL/L, caracterizado neste estudo como biomassa do biofloco (Tovar e Erazo 2009) 499
(Trabalho 3). 500
501
502

16
4.3 COLETA E HOMOGENEIZAÇÃO DAS AMOSTRAS 503
Para o trabalho 1 e 2, após 30 dias, foi efetuada a biometria final e os camarões 504
foram colocados em água com gelo para serem eutanaziado. Depois foram armazenados 505
em ultrafreezer (-80 oC) para no dia seguinte ser retirado brânquias, hepatopâncreas e 506
músculo. As amostras foram homogeneizadas (1:5, peso/volume) em solução tampão 507
de crustáceos (pH 7,2), contendo Tris-base (20 mM), EDTA (1 mM), MgCl2 (0,05 508
mM); DTT (Ditiotreitol – 1 mM); Sacarose (5 mM), KCl (Cloreto de potássio – 1 mM), 509
dissolvidos em água Milli Q. Posteriormente os extratos foram centrifugados a 9000 x 510
g, durante 30 minutos, em temperatura de 4 ºC e retirado o sobrenadante que foi 511
congelado a -80 ºC (Trabalho 1 e 2). 512
Para o trabalho 3, as amostras foram coletadas após leitura do cone Imhoff, no 513
qual a água foi retirada por sifonamento, e o floco colocados em falcon de 50 mL e 514
armazenado em gelo, para depois serem transferidos para eppendorf de 2 mL e serem 515
centrifugadas a 800 x g, a 4 °C, por 10 minutos e armazenado a -80°C. Para 516
homogeneização, as amostras de biofloco foram pesadas em eppendorff, adicionados 517
metanol (100%) sobre a relação 1000mg/1000µL, homogeneizadas e agitadas durante 3 518
horas, em seguida centrifugadas por 10 minutos a 10000 rpm e 4°C, então retirado o 519
sobrenadante que foram utilizados para determinar a capacidade antioxidante contra 520
radicais peroxil. 521
522
4.4 DETERMINAÇÃO DA ATIVIDADE DA ENZIMA GLUTATIONA S-523
TRANSFERASE (GST) 524
O ensaio da atividade da GST seguiu o método de Habig e Jakobi (1981). Neste 525
processo a absorbância é gerada pela conjugação de 1 mM de GSH (glutationa reduzida, 526
Sigma-Aldrich) com 1 mM de CDNB (1-chloro-2,4-dinitrobenzene, Sigma-Aldrich), 527
em absorbância de 340 nm, a 25 °C. Para leitura foi adicionado em microplaca 528
transparente de fundo chato, 15 μL do sobrenadante em 235 μL de meio de reação 529
(tampão fosfato 0,1 M e CNDB 50 mM, pH 7,0), mais 10 μL de GSH 25 mM. Nas 530
amostras de branco foi utilizado 15 μL tampão de homogeneização de crustáceo. As 531
leituras foram em espectrofluorímetro com leitora de placas (Víctor 2, Perkin Elmer). A 532

17
atividade específica da GST foi expressa em nanomoles de produto CDNB-GSH por 533
minuto por mg de proteína. 534
535
4.5 DETERMINAÇÃO DA ATIVIDADE DA GLUTATIONA REDUZIDA (GSH) 536
Foi efetuado pelo método de White et al. (2003) que mensura a concentração de 537
GSH pela sua reação com NDA (2,3 naftalenedicarboxialdeido), gerando um complexo 538
fluorescente (GSH-NDA) que detectado a 485 nm de excitação e 530 nm de emissão. 539
Para o ensaio primeiro foi feita uma curva padrão em diferentes diluições de GSH (40, 540
20, 10, 5 e 2,5 µM). Depois foram adicionados a microplaca transparente de fundo 541
cônico 25 μL de sobrenadante e 25 μL de tampão de homogeneização para o branco, 25 542
μL de ácido sulfosalicílico (200 mM) e incubado por 20 minutos. Em seguida a placa 543
foi centrifugada a 2500 rpm por 5 minutos. Foram então transferidos para microplaca 544
branca 20 μL de sobrenadante e 180 μL de solução de reação (Tris-base 50 mM, NaOH 545
500 mM e NDA 10 mM). As leituras foram feitas em espectrofluorímetro com leitora de 546
placas (Víctor 2, Perkin Elmer), em temperatura ambiente. A concentração foi expressa 547
em µomoles de GSH por mg de proteína. 548
549
4.6 DETERMINAÇÃO DOS NÍVEIS DE PEROXIDAÇÃO LIPÍDICA 550
A determinação de dano oxidativo segui o protocolo descrito por Oakes & Van 551
der Kraak (2003), este método envolve a reação do malondialdeído (MDA), um 552
subproduto da peroxidação lipídica (Hermes-Lima, 2004), com o ácido tiobarbitúrico 553
(TBA) sob condições de alta temperatura e acidez, gerando um cromógeno que é 554
quantificado por fluorometria. Para o ensaio foi feito a curva padrão com 6,25; 3,125; 555
1,5625; 0,78; 0,39; 0,195; 0,0975; 0,04887; 0,0243 e 0,0121 nmol de TMP (1,1,3,3-556
tetramethoxypropano). Em tubos de vidro (em duplicata) foi colocado 10 µL de amostra 557
e 41,2 µL de tampão de homogeneização de crustáceo nos brancos. Depois 20 µL de 558
solução estoque de BHT (hidroxitoluenobutilado, 1,407 mM), apenas nos tudos das 559
amostras. Em seguida 150 µL de solução de ácido acético 20%, 150 µL da solução de 560
TBA 0,8%; 50 µL de água MilliQ, 20 µL de SDS 8,1%. A mistura foi vortexada e 561
colocada em banho-maria a 95 ºC por 30 minutos. Logo após os tubos foram esfriados 562

18
por 10 minutos a temperatura ambiente, para então adicionar 100 µL de água MilliQ. O 563
contudo dos tubos foi transferido para eppendors de 1,5 mL e adicionou-se 500 µL de n-564
butanol, sendo logo vortexado e centrifugado a 3.000 x g por 10 minutos a 15 ºC. 565
Finalmente foi removida 150 µL da fase orgânica (sobrenadante) e transferida a 566
microplacas brancas. As leituras são feitas em fluorímetro, com comprimento de 567
excitação de 520 nm e emissão de 580 nm (Víctor 2, Perkin Elmer). Os resultados 568
foram expressos em nmol de TMP (Acros Organics) por mg de tecido fresco. 569
570
4.7 DETERMINAÇÃO DA CAPACIDADE ANTIOXIDANTE TOTAL CONTRA 571
RADICAIS PEROXIL (ACAP) 572
Foi efetuada de acordo com o protocolo de Amado et al. (2009), a dosagem tem 573
início com fixação a concentração de proteína em 2 mg/ml das amostras. Para o trabalho 574
3, não houve a fixação da concentração de proteína. Depois é adicionado a microplaca 575
branca 127,5 µL de tampão de reação, composto por 0,3575 g de ácido etanosulfônico 576
4.2-hidroxietil piperazina-1 (HEPES), 0,7455 g de cloreto de potássio (KCl), 0,0102 g 577
de cloreto de magnésio (MgCl2) dissolvidos em 50 ml de água Milli Q com pH ajustado 578
em 7,2; 10 µL de extrato de tecido; 7,5 µL de água MilliQ para as amostras sem ABAP 579
(2,2- azobis 2metilpropianoamidina dihidrocloreto) ou 7,5 µL de solução de ABAP para 580
amostras com ABAP (gerador de radicais peroxil) e 10 µL de solução de H2DCF-DA 581
(diacetato de ´2,7 diclorofluresceína). A leitura é realizada no tempo zero e depois a 582
cada 5 minutos até completar 30 minutos em fluorímetro de placas (Víctor 2, Perkin 583
Elmer) utilizando comprimento de onda de 530 nm de emissão e 485 nm de excitação, a 584
37 °C, temperatura que favorece a termólise do ABAP. Este método quantifica a 585
capacidade que o tecido possui em neutralizar as ERO geradas pela decomposição do 586
ABAP, incluindo as defesas antioxidantes enzimáticas e/ou não enzimáticas, através do 587
cálculo da área relativa. A área relativa apresenta uma relação inversa com a capacidade 588
antioxidante, onde menores valores da área relativa indicam uma maior capacidade 589
antioxidante e vice-versa. Para o trabalho 3, as leituras foram feitas em fluorímetro de 590
placas (FILTERMAX F5, Multi-mode microplate reader) utilizando comprimento de 591
onda de 530 nm de emissão e 485 nm de excitação, a 37 °C e o cálculo da área relativa 592
foi realizado segundo Monserrat et al. (2014). 593

19
4.8 CONTAGEM DIFERENCIAL DE HEMÓCITOS 594
A hemolinfa foi recolhido por punção cardíaca utilizando uma seringa de 3 mL 595
contendo solução anticoagulante de crustáceo, contendo NaCl (450 mM), glucose (100 596
mM), citrato de sódio (30 mM), ácido cítrico (23 mM), EDTA (20 mM) diluídos em 597
água MilliQ, com pH fixado em 7,4. Depois, um esfregaço de hemolinfa foi feita em 598
lâminas de vidro que foram imersas em metanol durante 5 minutos e coradas com May-599
Grunwald-Giemsa. Em seguida, hemócitos foram quantificados por microscópio de 600
lente ocular Integrando Disc 1, 25 pontos-G49 (Carl Zeiss), seguindo a metodologia de 601
Weibel (1980). 602
603
4.9 ANÁLISE ESTATÍSTICA DOS RESULTADOS 604
No primeiro trabalho os dados foram expressos em média ± desvio padrão. Cada 605
variável (atividade GST, capacidade antioxidante total, concentração de GSH, 606
peroxidação lipídica) foi analisada através de bi-fatorial ANOVA, sendo os fatores o 607
tratamento (água clara e biofloco) e órgãos (brânquias, hepatopâncreas e músculos). 608
Anteriormente, os pressupostos de normalidade e homogeneidade de variância foram 609
analisados e transformações matemáticas aplicada se a menos uma suposição foi 610
violado. Comparações de médias foram feitas usando a teste Newman-Keuls ou 611
contrastes ortogonais. Foram analisados os parâmetros físico e químicos da água por 612
meio do teste t de Student para variâncias desiguais (Zar, 1984). Em todos os casos foi 613
utilizado um nível de significância de 5%. 614
Para o segundo trabalho, os dados foram expressos em média ± desvio padrão. 615
Cada variável de cada órgão, incluindo a hemolinfa (atividade GST, peroxidação 616
lipídica, capacidade antioxidante total e contagem total de hemócitos) foi analisada 617
através de bi-fatorial ANOVA, sendo os fatores os tratamentos (NC ou NCLA) e 618
condição de criação (SW ou BFT). Anteriormente foram analisados os pressupostos de 619
normalidade e homogeneidade de variância. Comparações de médias foram feitas 620
usando o teste de Newman Keuls. Parâmetros físico e químicos da água foram 621
analisados pelo teste t de Student para variâncias desiguais (Zar 1984). Em todos os 622
casos foi utilizado um nível de significância de 5%. 623

20
Para o trabalho 3, os dados foram expressos em média ± desvio padrão. Com 624
análise feita por meio de bi-fatorial ANOVA (diferentes concentrações de ácido lipóico 625
e tempo). Anteriormente, os pressupostos de normalidade e homogeneidade de variância 626
foram analisados. Comparações de médias foram feitas usando a teste Newman-Keuls. 627
Em todos os casos foi utilizado um nível de significância de 5%. 628
629
630
631
632
633
634
635
636
637
638
639
640
641
642
643
644
645
646
647
648

21
REFERÊNCIAS 649
Abele, D, S Pintarulo. 2004. Formation of reactive species and induction of antioxidant 650
defence systems in polar and temperate marine invertebrates and fish. 651
Comparative Biochemistry and Physiology, Part A. 138: 405–415. 652
APHA (American Public Health Association). 1998. StandardMethods for the 653
Examination of Water and Wastewater, 20st edition. Washington, DC. 1193 pp. 654
Amado, LL, ML Garcia, PB Ramos, RF Freitas, B zafalon, JLR Ferreira, JS Yunes, JM 655
Monserrat. 2009. A method to measure total antioxidant capacity against peroxyl 656
radicals in aquatic organisms: Application to evaluate microcystins toxicity. 657
Science of the Total Environment. 407: 2115–2123. 658
Amado, LL, ML Garcia, TCB Pereira, JS Yunes, MR Bogo, JM Monserrat. 2011. 659
Chemoprotection of lipoic acid against microcystin-induced toxicosis in 660
common carp (Cyprinus carpoi, Cyprinidae). Comparative Biochemistry and 661
Physiology. Part C 154: 146–153. 662
Anderson, ME. 1998. Glutathione: an overview of biosynthesis and modulation. 663
Chemical Biology International. 111/112: 1–14. 664
Avnimelech, Y. 1999. Carbon and nitrogen ratio as a control element in aquaculture 665
systems. Aquaculture. 176: 227–235. 666
Avnimelech Y. 2007. Feeding with microbial flocs by tilapia in minimal discharge 667
bioflocs technology ponds. Aquaculture. 264:140–147. 668
Avnimelech, Y. 2012. Biofloc Technology – A Practical Guide Book. 2nd ed. The World 669
Aquaculture Society, Baton Rouge, United States. 670
Azim, ME, DC Little. 2008. The biofloc technology (BFT) in indoor tanks: Water 671
quality, biofloc composition, and growth and welfare of Nile tilapia 672
(Oreochromis niloticus). Aquaculture. 283:29–35. 673
Barreiros, ALBS, JM David, JP David. 2006. Estresse oxidativo: relação entre geração 674
de espécies reativas e defesa do organismo. Química Nova. 29: 113–123. 675

22
Bauchemal, K., S. Briançon, F. Cauenne, H. Fessi, M. Tayakout. 2006. Stability Studies 676
on Colloidal Suspensiond of Polyurethane Nanocapsules. Journal of 677
Nanoscience and Nanotechnology. 6: 3187 – 3192. 678
Benderschneider, K, RJ Robinson. 1952. A new spectrophotometric method for 679
determination of nitrate in sea water. Journal of Marine Research. 1: 69–87. 680
Biewenga, GPh, GRMM Haenen, A Bast. 1997. Review: The pharmacology of the 681
antioxidant lipoic acid. General Pharmacology: The Vascular System. 29: 315–682
331. 683
Burford, MA, PJ Thompson, RP McIntosh, RH Bauman, DC Pearson. 2004. The 684
contribution of flocculated material to shrimp (Litopenaeus vannamei) nutrition 685
in a highintensity, zero-exchange system. Aquaculture. 232:525–537. 686
Carreau, JP. 1979. Biosynthesis of lipoic acid via unsaturated fatty acids. Methods in 687
Enzymology. 62: 152–158. 688
Castex, M, P Lemaire, N Wabete, L Chim. 2009. Effect of dietary probiotic Pediococcus 689
acidilactici on antioxidant defences and oxidative stress status of shrimp 690
Litopenaeus stylirostris. Aquaculture. 294: 306−313. 691
Dickinson, DA, HJ Forman. 2002. Cellular glutathione and cellular metabolism. 692
Biochemical Pharmacology. 64: 1019–1026. 693
Ebeling, JM, MB Timmons, JJ Bisogni. 2006. Engineering analysis of the stoichiometry 694
of photoautotrophic, autotrophic, and heterotrophic control of ammonia-nitrogen 695
in aquaculture in aquaculture production systems. Aquaculture. 257: 346–358. 696
Emerenciano, M., G. Gaxiola, G. Cuzon. 2013. Biofloc Technology (BFT): A Review 697
for Aquaculture Application and Animal Food Industry, Biomass Now - 698
Cultivation and Utilization, Dr. Miodrag Darko Matovic (Ed.), ISBN: 978-953-699
51-1106-1, InTech. Chapter 12, p. 301–327. 700
Esterbauer, H. 1996. Estimation of peroxidative damage. A critical review. Pathologie 701
Biologie. 44: 25–28. 702

23
Flora, SJS. 2009. Structural, chemical and biological aspects of antioxidants for 703
strategies against metal and metalloid exposure. Oxidative Medicine and 704
Cellular Longevity. 4: 191–206. 705
Guidot, DM, JM McCord. 1999. Walking the tightrope: the balance between 706
mitochondrial generation and scavenging of superoxide anion. In: Cadenas, E., 707
Packer, L. (Eds.), Understanding the Process of Aging. Marcel Dekker. New 708
York. pp. 17– 38. 709
Gutierrez-Wing, MT, RF Malone. 2006. Biological filters in aquaculture: trends and 710
research directions for freshwater and marine applications. Aquacultural 711
Engineering. 34 163–171. 712
Gutteridge, JMC, B Halliwell. 2010. Antioxidants: Molecules, medicines, and myths. 713
Biochemical and Biophysical Research Communications. 393: 561–564. 714
Habig, NH, WB Jakoby. 1981. Assays for differentiation of glutathione-S-transferase. 715
Methods in Enzymology. 77: 398–405. 716
Halliwell, B, JMC Gutteridge. 1985. Lipid peroxidation: a radical chain reaction. In: 717
Halliwell, B., Gutteridge, J.M.C. (Eds.), Free Radicals in Biology and Medicine, 718
(2nd ed.). Clarendon Press, Oxford, pp. 188–276. 719
Halliwell, B, JMC Getturidge. 1995. Definition and measurement of antioxidants in 720
biological systems. Free Radical Biology & Medicine. 18: 125–126. 721
Hellou, J, NW Ross; TW Moon. 2012. Glutathione, glutathione S-transferase, and 722
glutathione conjugates, complementary markers of oxidative stress in aquatic 723
biota. Environmental Science and Pollution Research. 19: 2007–2023. 724
Hermes-Lima, M. 2004. Oxygen in biology and biochemistry: Role of free radicals. In: 725
Free radicals and oxidizing agents in biology. Chaper 12. KB Storey (Ed.). Wiley 726
Canada. 727
Huber, PC, A De Fátima, WP Almeida. 2008. Revisão: Glutationa e Enzimas 728
Relacionadas: Papel biológico e importância em processos patológicos. Química 729
Nova. 31: 1170 – 1179. 730

24
Ju, ZY, I Forster, L Conquest, W Dominy. 2008. Enhanced growth effects on shrimp 731
(Litopenaeus vannamei) from inclusion of whole shrimp floc or floc fractions to 732
a formulated diet. Aquaculture Nutrition. 14, 533−543. 733
Kim, S-K, Z Pang, H-C Seo, Y-R Cho, T Samocha, I-K Jang. 2014. Effect of bioflocs on 734
growth and immune activity of Pacific white shrimp, Litopenaeus vannamei 735
postlarvae. Aquaculture Research. 45: 362–371. 736
Külkamp-Guerreiro, IC, KPSS Guterres, AR Pohlmann. 2009. Estabilização da ácido 737
lipóico via encapsulamento em nanocápsulas poliméricas planejadas para 738
aplicação cutânea. Química Nova. 8: 2078–2084. 739
Lnaz, G. 1998. Role of mitochondria in oxidative stress and ageing. Biochimica et 740
Biphysica Acta. 1366: 53– 67. 741
Liu, J, HC Yeo, SJ Daniger, BN Ames. 1997. Assay of aldehydes from lipid 742
peroxidation: gas chromatography – mass spectrometry compared with 743
thiobarbituric acid. Analytical Biochemistry: Methods in the Biological Sciences. 744
245: 161–166. 745
Livingstone, D. 2001. Contaminant-stimulated reactive oxygen species production and 746
oxidative damage in aquatic organisms. Marine Pollution Bulletin. 42: 656–666. 747
Martins, ÁCS, JA Flores, W Wasiliesky Jr, J Zanette, EG Primel, SS Caldas; JM 748
Monserrat. 2014. Modulation of antioxidant and detoxification responses 749
induced by lipoic acid in the Pacific White shrimp Litopenaeus vannamei 750
(Boone, 1931) subjected to hypoxia and reoxygenation. Marine and Freshwater 751
Behaviour and Physiology. 2014. 47: 335–348. 752
Monserrat, JM, JV Lima, JLR Ferreira, D Acosta, ML Garcia, PB Ramos, TB Moraes, 753
LC Santos, LL Amado. 2008. Modulation of antioxidant and detoxification 754
responses mediated by lipoic acid in the fish Corydoras paleatus 755
(Callychthyidae). Comparative Biochemisty and Physiology, Part C. 148: 287–756
292. 757

25
Müller, CR, SE Haas, VL Bassani, SS Guterres, H Fessi, MC Peralba, AR Pihlman. 758
2004. Degradação e estabilização do diclofenaco em nanocápsulas poliméricas. 759
Química Nova. 27: 555 – 560. 760
Nuttall S, U Martin, A Sinclair, M Kendall. 1998. Glutathione: in sickness and in health. 761
The Lancet. 351: 645 – 646. 762
Oakes, KD, GJ Van der Kraak. 2003. Utility of the TBARS assay in detecting oxidative 763
stress in white sucker (Catostomus commersoni) populations exposed to pulp 764
mill efluente. Aquatic Toxicology. 63: 447–463. 765
Packer, L, EH Wiltf, HJ Tritschler. 1995. Alpha-lipoic acid as a biological antioxidant. 766
Free Radical Biology Medicine, 19: 227–250. 767
Piedrahita, R.H., 2003. Reducing the potential environmental impact of tank 768
aquaculture effluents through intensification and recirculation. Aquaculture. 226: 769
35–44. 770
Quintanilla-Carvajal, MX, BH Camacho-Díaz, LS Meraz-Torres, JJ Chanona-Pérez, L 771
Alamilla-Beltrán, A Jimenéz-Aparicio, GF Gutiérrez-Lopes. 2010. 772
Nanoencapsulation: A new trend in food engineering processing. Food 773
Engineering Reviews. 2: 39–50. 774
Ray, AJ. 2012. Biofloc technology for super-intensive shrimp culture. In: Avnimelech 775
Y, editor. Biofloc Technology - a practical guide book, 2nd ed., The World 776
Aquaculture Society, Baton Rouge, Louisiana, USA. pp. 167-188. 777
Ray, AJ, G Seaborn, JW Leffler, SB Wilde, A Lawson, CL Browdy. 2010. 778
Characterization of microbial communities in minimal-exchange, intensive 779
aquaculture systems and the effects of suspended solids management. 780
Aquaculture 310:130–138. 781
Schafer, FQ, GR Buettner. 2001. Redox environment of the cell as viewed through the 782
redox state of the glutathione disulfide/glutathione couple. Free Radical Biology 783
and Medicine. 30: 1191–1212. 784

26
Sies, H. 1985. Oxidative stress: Introductory remarks. In: Sies, H. (Ed.), Oxidative 785
Stress. Academic Press, London and Harcourt Brace Jovanovich Publishers. 786
New York, pp. 1–8. 787
Shrimp News International. 2015. In: The World – Estimated Farmed Shrimp 788
Production in 2013 and 2014. Published: February 23, 2015. Accessed: July 15, 789
2015<https://www.shrimpnews.com/FreeReportsFolder/NewsReportsFolder/Wo790
rldProductionIn2013And2014.html >. 791
Storey, KB. 1996. Oxidative stress: animal adaptations in nature. Brazilian Journal of 792
Medical and Biological Research. 29: 1715–1733. 793
Terjesen, BF, K Park, MB Tesser, MC Portella, Y Zhang, K Dabrowski. 2004. Lipoic 794
acid and ascorbic acid affect plasma free amino acids selectively in the teleost 795
fish pacu (Piaractus mesopotamicus). American Society for Nutritional 796
Sciences. 134: 2930–2934. 797
Tovar, SAO, EC Erazo. 2009. Analisis de las Caracteristicas Fisico-Quimicas de Aguas 798
y Suelos de Cultivos Acuicolas Intensivos y Superintensivos. Ceniacua-799
Colciencias. Primera Edición. Bogotá. 2009. 82 p. 800
Trattner, S, J Pickova, KH Park, J Rinchard, K Dabrowski. 2007. Effects of α-lipoic and 801
ascorbic acid on the muscle and brain fatty acids and antioxidant profile of the 802
South American pacu Piaractus mesopotamicus. Aquaculture. 273: 158–164. 803
UNESCO. 1983. Chemical methods for use in marine environmental monitoring. 804
Manual and Guides 12. Intergovernmental Oceanographic Commission, Paris, 805
France. 806
Vianna, DR, G Bubols, G Meirelles, BV Silva, AL Da Rocha, M Lanznaster, JM 807
Monserrat, SC Garcia, G Von Poser, VL Eifler-Lima. 2012. Evaluation of the 808
antioxidant capacity of synthesized coumarins. International Journal of 809
Molecular Sciences. 13: 7260–7270. 810
Wasielesky W.Jr, Atwood H, Stokes A, Browdy CL (2006) Effect of natural production 811
in a zero exchange suspended microbial floc based super-intensive culture 812
system for white shrimp Litopenaeus vannamei. Aquaculture 258:396–403. 813

27
Weiss-Angeli, V, FS Poletto, LR Zancan, F Baldasso, AR Pohlmann, SS Guterres. 2008. 814
Nanocapsules of octyl methxycinnamate containing quercetin delayed the 815
photodegradation of both components under ultraviolet A radiation. Journal of 816
Biomedical Nanotechnology. 4: 80 – 89. 817
White, CC, H Viernes, CM Krejsa, D Botta, TJ Kavanagh. 2003. Fluorescence-based 818
microtiter plate assay for glutamate–cysteine ligase activity. Analytical 819
Biochemistry. 318: 175–180. 820
Xu, W-J; Pan L-Q. 2012. Effects of bioflocs on growth performance, digestive enzyme 821
activity and body composition of juvenile Litopenaeus vannamei in zero-water 822
exchange tanks manipulating C/N ratio in feed. Aquaculture. 356–357: 147–152. 823
Xu, WJ, LQ Pan. 2013. Enhancement of immune response and antioxidant status of 824
Litopenaeus vannamei juvenile in biofloc-based culture tanks manipulating high 825
C/N ratio of feed input. Aquaculture. 412−413: 117−124. 826
Xu, WJ, LQ Pan. 2014. Evaluation of dietary protein level on selected parameters of 827
immune and antioxidant systems, and growth performance of juvenile 828
Litopenaeus vannamei reared in zero-water exchange biofloc-based culture 829
tanks. Aquaculture. 426–427: 181–188. 830
Zar, JH, 1984. Biostatistical analysis. New Jersey: Prentice Hall. 736 p. 831
Zhou, J, W Wang, A Wang, W He, Q Zhou, Y Liu, J Xu. 2009. Glutathione S-transferase 832
in the white shrimp Litopenaeus vannamei: Characterization and regulation 833
under pH stress. Comparative Biochemistry and Physiology, Part C. 150: 224–834
230. 835
836
837
838
839
840

28
CAPÍTULO I 841
842
843
Antioxidant and oxidative damage responses in different 844
organs of Pacific white shrimp Litopenaeus vannamei (Boone, 845
1931) reared in a biofloc technology system 846
847
848
849
850
Átila Clivea da Silva Martins, Juliana Artigas Flores, Camilla 851
Porto, Wilson Wasielesky Junior & José Maria Monserrat 852
853
854
855
856
857
858
859
860
Marine and Freshwater Behaviour and Physiology, 2015. Vol. 48, 861
No. 4, 279–288. 862
863
864
865
866
867
868

29
ABSTRACT 869
Shrimp (Litopenaeus vannamei) reared in a conventional seawater (SW) aquarium 870
system SW were compared with those raised in a biofloc technology (BFT) system. 871
After 30 days, the L. vannamei shrimp were euthanized and samples of gills, 872
hepatopancreas and muscle were dissected. Statistical analysis was performed using bi-873
factorial ANOVA, with the factors of the treatment (shrimp reared in SW or in a BFT 874
system) and organs (gills, hepatopancreas and muscles). No differences (p>0.05) in 875
glutathione-S-transferase activity were observed between shrimp reared in SW and 876
shrimp reared in BFT (p< 0.05). Glutathione levels were lower (p<0.05) in the gills and 877
hepatopancreas of shrimp reared in the BFT system, suggesting changes in the 878
antioxidant composition of these organs. Lipid peroxidation levels were higher in the 879
hepatopancreas than in muscle in shrimp reared in SW (p<0.05) and this difference was 880
not observed in organisms reared in the BFT system (p>0.05). Overall, the results 881
showed that rearing shrimp in the BFT system altered both their antioxidant and 882
oxidative damage responses. This indicates that some microbial communities in BFT 883
systems can influence the redox state of L. vannamei. 884
885
Keywords: Litopenaeus vannamei; BFT; bioflocs; antioxidant responses; lipid 886
peroxidation; total antioxidant capacity. 887
888
889
890
891
892
893
894
895
896

30
1 INTRODUCTION 897
Among many aquatic species that are cultured in the world, the white shrimp 898
Litopenaeus vannamei has a number of advantages. For example, the species has a high 899
tolerance to variations in temperature, salinity, pH, and dissolved oxygen, possesses in 900
efficient feed conversion ratio, is resistant to several types of disease and is tolerant to 901
crowding (ABCC 2010). 902
Aquacultural activity can have some negative environmental outcomes such as 903
the release of toxic nitrogen compounds like ammonia and nitrite (Xu et al. 2013). The 904
use of bioflocs technology (BFT) systems has provided a useful alternative in which the 905
presence of aerobic and heterotrophic organisms together with vigorous aeration allows 906
for the rearing of aquatic organisms at a high density without the need for constant 907
water exchange (Furtado et al. 2011; Kim et al. 2014). Recent studies have shown that 908
the benefits of BFT systems are not restricted to the reduction of the need for water 909
exchanges. Additionally, the microorganism communities are beneficial for the 910
organisms being cultured. Xu et al. (2013) showed that BFT improved activity in 911
digestive enzymes such as proteases, amylase and lipase in L. vannamei. These authors 912
proposed that the high digestive enzyme activity could be related to exogenous enzymes 913
released from microorganisms lysed in the bioflocs. Improvement in weight gain, 914
specific growth rate and the feed conversion rate of L. vannamei reared in a BFT system 915
were reported by Xu and Pan (2012). These results led the authors to postulate that 916
bioflocs are a food supplement that improves digestion and growth. Kim et al. (2014) 917
observed augmented gene expression in the prophenol oxidase system in white shrimp 918
reared in BFT, which reinforces the idea that several biological benefits are linked by 919
this rearing strategy. 920
When considering bioflocs as an additional food source (Xu et al. 2013), it is 921
important to note that they contain several biomolecules including carotenoids, amino 922
sugars and phytosterols, among others (Ju et al. 2008). Molecules such as carotenoids 923
are known to possess antioxidant properties (Hermes-Lima 2004; Halliwell & 924
Gutteridge 2007). It is thus possible that BFT may improve not only digestive, imune 925
and growth parameters, but also antioxidant responses. Several conditions can generate 926
oxidative stress in aquatic organisms, and some of these conditions are important for 927

31
aquaculture practices. Crustaceans exhibit daily variations in their enzymatic 928
antioxidant system (Fanjul-Moles et al. 2003; Maciel et al. 2004). This means that they 929
may be more susceptible to oxidative stress during certain periods. Infection by the 930
white spot virus was also shown to reduce the antioxidant competence of 931
Fenneropenaeus indicus (Mohankumar & Ramasamy 2006) and the crustacean immune 932
response itself promotes a pro-oxidant condition (Cornet et al. 2007). 933
Taking into account the properties of BFT systems that led Xu et al. (2003) to 934
consider providing a natural food supplement to organisms such as L. vannamei, this 935
study aimed to evaluate the effect of a BFT system on the antioxidant and oxidative 936
damage responses of this species and to record other beneficial changes elicited by the 937
use of this technology. To address this aim we compared the antioxidant response of the 938
shrimp L. vannamei reared in a SW system (without biofloc) and in a BFT system. 939
940
2 MATERIALS AND METHODS 941
This study employed juvenile shrimp (L. vannamei) from the nursery sector of 942
the Aquaculture Marine Station at the Federal University of Rio Grande – FURG 943
(Brazil). During acclimation period (seven days) shrimp were dispersed in six plastic 944
tanks with 50 L of sea water (SW) each. Measured physico–chemical parameters of SW 945
were: 7.79 ± 0.15 (pH), 6.5 ± 0.12 mg/L (dissolved oxygen), 26.26 ± 0.38 °C 946
(temperature), 31.00 ± 0.01 (salinity), 0.78 ± 0.47 mg/L (ammonia), 0.07 ± 0.04 mg/L 947
(nitrite) and 0.39 ± 0.53 mg/L (nitrite). Organisms were stocked at a density of 15 948
shrimp per tank (approximately 210 shrimp per m3). The diet consisted of commercial 949
food with 35% crude protein supplied three times daily (08:00, 15:00 and 22:00 h) at a 950
feeding rate of 3% of shrimp biomass. When the concentration of total ammonia 951
exceeded 1 mg/L, molasses was added to the bioflocs (Avnimelech 1999). The 952
experiment lasted for 30 days and no water exchanges were performed in the BFT 953
system treatment. In SW, 80–90% of the water was replaced every 48 h. 954
During the experiment, the physical and chemical parameters of water, including 955
dissolved oxygen (mg/L), water temperature (°C), salinity and pH, were measured daily 956
using an YSI multiparameter apparatus (Yellow Springs Instruments, model 55). Shrimp 957

32
from both treatments were weighed at the end of the experiment, frozen, and the gills, 958
hepatopancreas and muscle from the second abdominal somite were dissected. 959
Organ and tissue samples were homogenized (1:5, w/v) in a buffer, previously 960
used for crustacean species (Pinho et al. 2005; de Souza et al. 2014). The composition 961
was Tris-Base (20 mM), EDTA (1 mM), dithiothreitol (1 mM; Sigma), sucrose (500 962
mM) and KCl (150 mM) dissolved in Milli Q water with the pH adjusted to 7.2. The 963
homogenates were centrifuged (9000 × g, 30 min at 4 °C), and the supernatants were 964
kept and divided into four aliquots for subsequent analysis of glutathione-S-transferase 965
(GST) activity, levels of lipid peroxidation (TBARS assay), levels of reduced 966
glutathione (GSH) and total antioxidant capacity against peroxyl radicals. 967
968
2.1 DETERMINATION OF GST ACTIVITY 969
GST was measured according to the protocol described by Habig et al. (1974). 970
Briefly, in the assay the conjugation of 1 mM 1-chloro-2,4-dinitrobenzene (CDNB, 971
Sigma) with 1 mM reduced glutathione (GSH, Sigma) in the presence of 10 μl of 972
sample homogenates (source of GST enzyme) generated a CDNB-GSH complex. The 973
absorbance of this complex was measured at 340 nm using a spectrofluorometer with a 974
microplate reader (Victor 2, Perkin Elmer). 975
976
2.2 DETERMINATION OF TOTAL ANTIOXIDANT CAPACITY 977
Total antioxidant capacity against peroxyl radicals was measured by determining 978
the reactive oxygen species in aliquot homogenates according to the protocol of Amado 979
et al. (2009). Peroxyl radicals were generated through thermal decomposition, at 37 °C 980
of 2,2′-azobis 2 methylpropionamidine dihydrochloride (ABAP; Aldrich). The probe 981
dichlorofluorescein diacetate (H2DCF-DA, Invitrogen; 40 μM) was added to a white 982
ELISA 96-well microplate, which was immediately placed in a fluorometer with a 983
microplate reader (Victor 2, Perkin Elmer) heated at 37 °C. The fluorescence was read 984
(excitation: 485 nm; emission: 520 nm) every 5 min over a 30-min period, and the 985
relative area was calculated according to Amado et al. (2009). According to these 986
authors, a high relative area means a low antioxidant capacity indicating a low capacity 987

33
of antioxidants present in the biological sample to degrade or intercept the peroxyl 988
radicals. 989
990
2.3 DETERMINATION OF CONCENTRATION OF REDUCED GLUTATHIONE 991
(GSH) 992
The procedure was based on that described by White et al. (2003) with 993
modifications proposed by da Rocha et al. (2009). The substrate 2,3 naphthalene 994
dicarboxialdehyde (NDA, Invitrogen) reacts with GSH to form a fluorogenic 995
compound. Aliquots of organ homogenates were transferred to white ELISA 96-well 996
microplate. The NDA solution was subsequently added to all wells and, after 30 min of 997
incubation, the fluorescence intensity of the NDA–GSH complex was read at excitation 998
and emission wavelengths of 485 and 530 nm, respectively, using a plate reader 999
fluorometer (Victor 2, Perkin Elmer). A standard curve of GSH was prepared to express 1000
the data in terms of μM equivalents of GSH per mg of total proteins. 1001
1002
2.4 DETERMINATION OF LIPID PEROXIDATION 1003
Lipid peroxidation was determined by a TBARS (thiobarbituric acid reactive 1004
substances) assay following Oakes and Van der Kraak (2003), adapted to microplate by 1005
Da Rocha et al. (2009). Aliquots (10 μl) of sample homogenates were added to glass 1006
tubes, together with thiobarbituric acid (0.8%, Sigma), acetic acid (20%), Milli Q water 1007
and sodium dodecyl sulfate (8.1%). The mixture was incubated at 95 °C over 30 min 1008
and, after cooling, n-butanol was added following a centrifugation step (3000 × g for 10 1009
min at 15 °C). The n-butanol phase was pipetted to a white ELISA 96-well microplate 1010
and the fluorescence read (excitation: 520 nm; emission: 580 nm) in a fluorometer with 1011
a microplate reader (Victor 2, Perkin Elmer). The standard employed was 1,1,3,3-1012
tetramethoxypropane (TMP, Across Organics). The results were expressed as nmol 1013
equivalents of TMP per mg of fresh tissue. 1014
1015
1016

34
2.5 DETERMINATION OF TOTAL AMMONIA 1017
Total ammonia determination (NH3+ NH4+) follows the UNESCO (1983) 1018
method in which ammonia in basic pH (between 10.8 and 11.5) reacts with bleaching 1019
solution, forming monochloramine. In the presence of phenol and an excess of 1020
hypochlorite and the addition of sodium nitroprusside as a catalyst, indophenol blue is 1021
formed, a molecule that absorbs at 630 nm. 1022
1023
2.6 STATISTICAL ANALYSIS 1024
Data were expressed as the mean ± standard deviation. Each variable (GST 1025
activity, lipid peroxidation, GSH levels and total antioxidant capacity) was analyzed 1026
through bifactorial ANOVA, with factors composed of the treatment (shrimp reared in 1027
SW or BFT and organs (gills, hepatopancreas and muscles). Prior to application of 1028
ANOVA the assumptions of normality and homogeneity of variance were tested and 1029
mathematical transformations were applied if at least one assumption was violated. 1030
Means comparisons were done using the Newman–Keuls test or orthogonal contrasts. 1031
Water physico–chemical parameters were analyzed using Student’s t test for unequal 1032
variances (Zar 1984). In all cases, a significance level of 5% was used. 1033
1034
3 RESULTS 1035
The average initial weight, final weight and average weight gain were not 1036
significantly different (p>0.05) for SW and BFT reared shrimp. In both cases, survival 1037
was 100% (Table 1). Several differences in physico–chemical parameters between SW 1038
and water from BFT system were observed (Table 2). Water from the BFT system 1039
showed significantly higher values for pH, salinity and nitrate (p<0.05) compared with 1040
SW whereas oxygen concentration and nitrite levels were lower (p<0.05). 1041
1042
1043
1044

35
Table 1. Mean concentration (±1 standard error) of morphological and physiological 1045
parameters measured in shrimp L. vannamei maintained in a SW or in a BFT system. 1046
1047
Note: No significant differences were detected between treatments (p>0.05). Number of 1048
samples analyzed in each treatment: 45. 1049
1050
No significant differences were observed in GST activity between treatments 1051
(p>0.05 and Table 3) although GST activity in the gills showed an upward trend in BFT 1052
shrimp. When comparing the different organs, the gills showed the highest GST activity, 1053
followed by the muscle and the hepatopancreas (p<0.05). 1054
The muscles of BFT shrimp showed significantly higher total antioxidant 1055
capacity (low relative area) when compared with SW shrimp (high relative area) 1056
(p<0.05 and Table 4). In SW shrimp the hepatopancreas had the highest antioxidant 1057
capacity (p<0.05) when compared with the muscle and gills, which presented similar 1058
values (p>0.05). BFT shrimp showed differences in every organ: highest antioxidant 1059
capacity in the hepatopancreas, followed by the muscle with the lowest mean value in 1060
the gills (p<0.05 and Table 4). 1061
A significant GSH reduction was observed in the gills and hepatopancreas of 1062
BFT shrimp (p<0.05) indicating that glutathione levels were affected by the rearing 1063
environment. In each system, the differences in GSH content among the different organs 1064
were the same: muscle showed the highest level, followed by the hepatopancreas and 1065
then the gills (p<0.05 and Table 5A). 1066
1067
1068
1069

36
Table 2. Mean concentration (±1 standard error) of the physico-chemical parameters 1070
measured in SW or in a BFT system. 1071
1072
Note: Asterisks (*) indicate significant differences (p<0.05) between the experimental 1073
groups after performing Student’s t-test assuming unequal variances. Number of 1074
samples analyzed in each treatment: 27–186. 1075
1076
Table 3. Mean activity (±1 standard error) of GST (nmol CDNB-GSH 1077
conjugate/min/mg of proteins) in the gills, hepatopancreas and muscle of shrimp L. 1078
vannamei reared in SW or in a BFT system. 1079
1080
Note: Different letters indicate significant differences (p<0.05) between experimental 1081
groups after performing a Student–Newman–Keuls post hoc test. Number of samples 1082
analyzed in each treatment: 15. GSH: reduced glutathione. CDNB: 1-chloro-2,4-1083
dinitrobenzene. 1084
1085
Table 4. Mean values (±1 standard error) of the total antioxidant capacity against 1086
peroxyl radicals (expressed in relative area) in the gills, hepatopancreas and muscle of 1087
L. vannamei reared in SW or in a BFT system. 1088

37
1089
Note: Different letters indicate significant differences (p<0.05) between experimental 1090
groups after performing a Student–Newman–Keuls post hoc test. Number of samples 1091
analyzed in each treatment: 10–15. 1092
1093
Table 5. Mean concentration (±1 standard error) of: (A) reduced glutathione (GSH) (μM 1094
of GSH/mg of total proteins) and (B) thiobarbituric reactive substances (nmol of 1095
TMP/mg of tissue) in the gills, hepatopancreas and muscle of L. vannamei reared in SW 1096
or in a BFT system. 1097
Note: Different letters indicate significant differences (p<0.05) between experimental 1098
groups after performing a Student–Newman–Keuls post hoc test. Number of samples 1099
analyzed in each treatment: 10–15. TMP: 1,1,3,3-tetramethoxypropane, the standard 1100
employed in TBARS assay. 1101
Oxidative damage measured in terms of TBARS was similar between the organs 1102
of BFT shrimp and SW shrimp (p>0.05 and Table 5B). The TBARS concentration in 1103
each organ was, however, different depending on the rearing environment. In SW 1104
shrimp, the highest TBARS concentration was observed in the hepatopancreas, followed 1105
by the gills, with the lowest TBARS concentration observed in the muscle (p<0.05 and 1106
Table 5B). In BFT shrimp no differences in TBARS levels were detected between 1107
organs (p>0.05 and Table 5B). 1108

38
4 DISCUSSION 1109
Nitrate levels can vary from 4.52 to 26 to mg/L during the rearing of commercial 1110
shrimp (Muir et al. 1991) and can reach 500 mg/L in recirculation systems (Pierce et al. 1111
1993). In studies with juvenile Penaeus monodon, however, the highest safe nitrate level 1112
at a salinity of 35 was reported to be 158 mg/L (Tsai & Chen 2002). Lower nitrate levels 1113
were observed during the present study in both SW, where 80–90% of water was 1114
renewed every 48 h and in BFT, where no water renewal was performed. 1115
Safe nitrite levels were estimated for L. vannamei to be 15.2 mg/L (Lin & Chen 1116
2003), a value much higher than those registered in both our treatments. The study of 1117
Zhang et al. (2006) into oxygen tolerance determined that in L. vannamei, the lethal 1118
levels were below 0.5 mg/L of dissolved oxygen. This shows again that the measured 1119
O2 levels in both our treatments were far below a level that could be considered 1120
stressful. The lower O2 concentration observed here with BFT was also reported in a 1121
previous study from our group (Emerenciano et al. 2012) and is consistent with the high 1122
density of aerobic organisms. 1123
The measured parameters of water quality (temperature, salinity, pH, dissolved 1124
oxygen, ammonia, nitrite and nitrates) in a BFT system and in SW in the study of 1125
Wasielesky et al. (2006) were close to those registered in the present study. In summary, 1126
according to values cited in the literature the water quality parameters in our study were 1127
within acceptable limits for the survival and growth of L. vannamei. 1128
Different organs of aquatic species differ in antioxidant competence and 1129
oxidative damage, as found by Monserrat et al. (2008) in the fish Corydoras paleatus, 1130
where, for example, GSH levels were higher in the muscle compared with the brain or 1131
gills. In L. vannamei shrimp, Lobato et al. (2013) found higher TBARS levels in the 1132
hepatopancreas compared to the muscle. In the present study, some differences were 1133
observed between the gills, hepatopancreas and muscle of L. vannamei. For example, 1134
higher GST activity was observed in the gills of shrimp reared in both systems. Some 1135
GSTs possess peroxidase activity, catalyzing the degradation of organic hydroperoxides 1136
(Halliweel & Gutteridge 2007), and this most likely explains why the gills have 1137
moderate TBARS levels even when the total antioxidant capacity is lower than that in 1138
the other organs. The differences among the organs in terms of lipid peroxidation for 1139

39
SW reared shrimp were not observed in BFT reared shrimp which suggests that the 1140
microbial community of this system could be influencing the redox state of L. vannamei 1141
resulting in the organs presenting similar levels of oxidative damage expressed in terms 1142
of TBARS levels. 1143
Other evidence of the positive influence of BFT on the antioxidant system of L. 1144
vannamei included the higher total antioxidant competence against peroxyl radicals 1145
(lower relative area) in the muscle of shrimp reared in the BFT system compared with 1146
SW reared shrimp. The observed reduction in GSH levels in the gills and 1147
hepatopancreas of shrimp reared in BFT deserves careful consideration. These results a 1148
priori can be interpreted as a pro-oxidant action induced by BFT, lowering the 1149
concentration of the antioxidant. However, the total antioxidant capacity in these two 1150
organs was similar in shrimp reared in both systems, and the TBARS levels in the gills 1151
and hepatopancreas were also similar between shrimp reared in the two systems. It is 1152
possible that rearing the shrimp in the BFT system altered their antioxidant 1153
composition. According to Regoli and Winston (1999) and Regoli (2000), antioxidants 1154
such as GSH, ascorbic acid, uric acid and α-tocopherol accounts for almost 70% of 1155
antioxidant capacity against peroxyl radicals. In this sense, a reduction in GSH content 1156
can be compensated by other antioxidants, leaving the total antioxidant capacity 1157
unchanged. 1158
Several examples exist in nature that show how biological interactions modify 1159
the antioxidant defense system. Moraes et al. (2006) showed that the mucus secretion of 1160
the polychaete Laeonereis acuta is colonized by bacteria communities that confer 1161
antioxidant properties. In the present study, the results indicated altered antioxidant 1162
responses and ameliorated lipid peroxidation in shrimps maintained in the BFT system. 1163
Future studies will need to consider the influence of microbial communities with 1164
different species compositions on the antioxidant competence of cultured organisms. It 1165
will also be important to learn how a microbial community of a BFT system can vary its 1166
antioxidant properties such as by adding exogenous antioxidants to the system. 1167
1168
1169

40
Acknowledgements 1170
J.M. Monserrat and Wilson Wasielesky Junior receive productivity research fellowships 1171
from the Brazilian agency CNPq (process numbers PQ 307880/2013-3 and PQ 1172
310993/2013-0, respectively). J.A. Artigas and C. Porto received undergraduate 1173
fellowship from FAPERGS. The authors appreciate the commentaries and suggestions 1174
of the three anonymous reviewers that helped to improve the focus and clarity of the 1175
manuscript. The financial support of PROPESP from FURG for English revision is also 1176
acknowledged. 1177
1178
Disclosure statement 1179
No potential conflict of interest was reported by the authors. 1180
1181
ORCID: José Maria Monserrat http://orcid.org/0000-0002-9463-3049 1182
1183
1184
1185
1186
1187
1188
1189
1190
1191
1192
1193
1194

41
REFERENCES 1195
ABCC. 2010. Análise da produção e do mercado interno e externo do camarão cultivado 1196
[Analysis of production and domestic and foreign markets of farmed shrimp]. 1197
Revista da ABCC. 1:18–23. 1198
Amado LL, Garcia ML, Ramos PB, Freitas RF, Zafalon B, Ferreira JLR, Yunes JS, 1199
Monserrat JM. 2009. A method to measure total antioxidant capacity against 1200
peroxyl radicals in aquatic organisms: application to evaluate microcystins 1201
toxicity. Sci Total Environ. 407:2115–2123. 1202
Avnimelech Y. 1999. Carbon/nitrogen ratio as a control element in aquaculture systems. 1203
Aquaculture. 176:227–235. 1204
Cornet S, Biard C, Moret Y. 2007. Is there a role for antioxidant carotenoids in limiting 1205
selfharming immune response in invertebrates? Biol Lett. 3:284–288. 1206
Da Rocha AM, Salomão de Feitas DP, Burns M, Vieira JP, de la Torre FR, Monserrat 1207
JM. 2009. Seasonal and organ variations in antioxidant capacity, detoxifying 1208
competence and oxidative damage in freshwater and estuarine fishes from 1209
Southern Brazil. Comp Biochem Physiol C. 150:512–520. 1210
Emerenciano M, Ballester ELC, Cavalli RO, Wasielesky W. 2012. Biofloc technology 1211
application as a food source in a limited water exchange nursery system for pink 1212
shrimp Farfantepenaeus brasiliensis (Latreille, 1817). Aquaculture Res. 43:447–1213
457. 1214
Fanjul-Moles ML, Durán-Lizarraga ME, Gonsebatt ME, Prieto-Sagredo J. 2003. The 1215
effect of photoperiod and light irradiance on the antioxidant circadian system of 1216
two species of crayfish from different latitudes: Procambarus clarkii and P. 1217
digueti. Photochem Photobiol. 77:210–218. 1218
Furtado PS, Poersch LH, Wasielesky W Jr. 2011. Effect of calcium hydroxide, carbonate 1219
and sodium bicarbonate on water quality and zootechnical performance of 1220
shrimp Litopenaeus vannamei reared in bio-flocs technology (BFT) systems. 1221
Aquaculture. 321:130–135. 1222

42
Habig WH, Pabst MJ, Jakoby WB. 1974. Glutathione S-transferases: the first enzymatic 1223
step in mercapturic acid formation. J Biol Chem. 249:7130–7139. 1224
Halliwell B, Gutteridge JMC. 2007. Free radicals in biology and medicine. 4th ed. New 1225
York, NY: Oxford University Press. 1226
Hermes-Lima M. 2004. Oxygen in biology and biochemistry: role of free radicals. In: 1227
Storey KB, editor. Free radicals and oxidizing agents in biology. Chapter 12. 1228
Canada: Wiley. 1229
Ju ZY, Forster I, Conquest L, Dominy W. 2008. Enhanced growth effects on shrimp 1230
(Litopenaeus vannamei) from inclusion of whole shrimp floc or floc fractions to 1231
a formulated diet. Aquaculture Nutr. 14:533–543. 1232
Kim S-K, Pang Z, Seo H-C, Cho Y-R, Samocha T, Jang I-K. 2014. Effect of bioflocs on 1233
growth and immune activity of Pacific white shrimp, Litopenaeus vannamei 1234
postlarvae. Aquaculture Res. 45:362–371. 1235
Lin Y-C, Chen J.-C. 2003. Acute toxicity of nitrite on Litopenaeus vannamei (Boone) 1236
juveniles at different salinity levels. Aquaculture. 224:193–201. 1237
Lobato RO, Nunes SM, Wasielesky W Jr, Fattorini D, Regoli F, Monserrat JM, Ventura-1238
Lima J. 2013. The role of lipoic acid in the protection of metallic pollutants 1239
effects in the shrimp Litopenaeus vannamei (Crustacea, Decapoda). Comp 1240
Biochem Physiol A. 165:491–497. 1241
Maciel FE, Rosa CE, Santos EA, Monserrat JM, Nery LEM. 2004. Daily variations in 1242
oxygen consumption, antioxidant defenses, and lipid peroxidation in the gills 1243
and hepatopancreas of an estuarine crab. Can J Zool. 82:1871–1877. 1244
Mohankumar K, Ramasamy P. 2006. White spot syndrome virus infection decreases the 1245
activity of antioxidant enzymes in Fenneropenaeus indicus. Virus Res. 115:69–1246
75. 1247
Monserrat JM, Lima JV, Ferreira JLR, Acosta D, Garcia ML, Ramos PB, Moraes TB, 1248
Santos LC, Amado LL. 2008. Modulation of antioxidant and detoxification 1249
responses mediated by lipoic acid in the fish Corydoras paleatus 1250
(Callychthyidae). Comp Biochem Physiol C. 148:287–292. 1251

43
Moraes TB, Ribas Ferreira JL, da Rosa CE, Sandrini JZ, Votto AP, Trindade GS, 1252
Geracitano LA, Abreu PC, Monserrat JM. 2006. Antioxidant properties of the 1253
mucus secreted by Laeonereis acuta (Polychaeta, Nereididae): a defense against 1254
environmental pro-oxidants? Comp Biochem Physiol C. 142:293–300. 1255
Muir PR, Sutton DC, Owens L. 1991. Nitrate toxicity to Penaeus monodon protozoea. 1256
Mar Biol. 108:67–71. 1257
Oakes KD, Van der Kraak GJ. 2003. Utility of TBARS assay in detecting oxidative 1258
stress in white sucker (Catostomus commersoni) populations exposed to pulp 1259
mill effluent. Aquat Toxicol. 63:447–463. 1260
Pierce RH, Weeks JM, Prappas JM. 1993. Nitrate toxicity to five species of marine fish. 1261
J. World Aquacult Soc. 24:105–107. 1262
Pinho GL, Moura da Rosa C, Maciel FE, Bianchini A, Yunes JS, Proença LAO, 1263
Monserrat JM. 2005. Antioxidant responses and oxidative stress after 1264
microcystin exposure in the hepatopancreas of an estuarine crab species. 1265
Ecotoxicol Environ Saf. 61:353–360. 1266
Regoli F. 2000. Total oxyradical scavenging capacity (TOSC) in polluted and 1267
translocated mussels: a predictive biomarker of oxidative stress. Aquat Toxicol. 1268
50:351–361. 1269
Regoli F, Winston GW. 1999. Quantification of total oxidant scavenging capacity of 1270
antioxidants for peroxynitrite, peroxyl radicals, and hydroxyl radicals. Toxicol 1271
Appl Pharmacol. 156:96–105. 1272
Souza DM, Martins ÁC, Jensen L, Wasielesky W Jr, Monserrat JM, Garcia LO. 2014. 1273
Effect of temperature on antioxidant enzymatic activity in the Pacific white 1274
shrimp Litopenaeus vannamei in a BFT (Biofloc technology) system. Mar 1275
Freshw Behav Physiol. 47:1–10. 1276
Tsai S-J, Chen J-C. 2002. Acute toxicity of nitrate on Penaeus monodon juveniles at 1277
diferente salinity levels. Aquaculture. 213:163–170. 1278
UNESCO. 1983. Chemical methods for use in marine environmental monitoring. Paris: 1279
Paris, Intergovernmental Oceanographic Commission; p. 53. 1280

44
Wang C, Liu H., Sun Z. 2012. Review Article: Heterogeneous Photo-Fenton Reaction 1281
Catalyzed by Nanosized Iron Oxides for Water Treatment. International Journal 1282
of Photoenergy: 1 – 10. 1283
Wasielesky Jr W, Atwood H, Stokes A, Browdy CL. 2006. Effect of natural production 1284
in a zero exchange suspended microbial floc based super-intensive culture 1285
system for white shrimp Litopenaeus vannamei. Aquaculture. 258:396–403. 1286
White CC, Viernes H, Krejsa CM, Botta D, Kavanagh TJ. 2003. Fluorescence-based 1287
microtiter plate assay for glutamate-cysteine ligase activity. Anal Biochem. 1288
318:175–180. 1289
Xu W-J, Pan L-Q. 2012. Effects of bioflocs on growth performance, digestive enzyme 1290
activity and body composition of juvenile Litopenaeus vannamei in zero-water 1291
exchange tanks manipulating C/N ratio in feed. Aquaculture. 356–357:147–152. 1292
Xu W-J, Pan LQ, Sun XH, Huang J. 2013. Effects of bioflocs on water quality, and 1293
survival, growth and digestive enzyme activities of Litopenaeus vannamei 1294
(Boone) in zero-water exchange culture tanks. Aquaculture Res. 44:1093–1102. 1295
Zar JH. 1984. Biostatistical analysis. New Jersey (NJ): Prentice Hall. 1296
Zhang P, Zhang X, Li J, Huang G. 2006. The effect of body weight, temperature, 1297
salinity, pH, light intensity and feeding condition on lethal DO levels of whiteleg 1298
shrimp Litopenaeus vannamei. Aquaculture. 256:579–587. 1299
1300
1301
1302
1303
1304
1305
1306
1307

45
CAPÍTULO II 1308
1309
1310
Antioxidant effects of nanoencapsulated lipoic acid in tissues 1311
and differential hemocyte count in hemolymph of shrimp 1312
Pacific Litopenaeus vannamei (Boone, 1931) created in biofloc 1313
system 1314
1315
1316
Átila Clivea da Silva Martins, Juliana Artigas Flores, Camilla 1317
Porto, Luis Alberto Romano, Wilson Wasielesky Junior, Sergiane 1318
S. Caldas, Ednei G. Primel, Irene Clemes Külkamp-Guerreiro, 1319
José Maria Monserrat 1320
1321
1322
Submetido a: 1323
Comparative Biochemistry and Physiology - Part A: Molecular & 1324
Integrative Physiology 1325
1326
1327
1328

46
ABSTRACT 1329
One of the limiting factors for the intensification of production in aquaculture is the 1330
welfare of reared organisms. In this study, we evaluated antioxidant and oxidative 1331
damage responses and immune parameters in shrimp Litopenaeus vannamei reared in 1332
saline water (SW) and in biofloc system (BFT). Animals were fed with ration 1333
supplemented with nanocapsules containing the antioxidant lipoic acid (NCLA) or with 1334
empty nanocapsules (NC). Oxidative stress analysis was evaluated through 1335
measurement of glutathione S-transferase activity (GST), reduced glutathione (GSH) 1336
concentration and lipid peroxidation (TBARS) in gills, hepatopancreas and muscle. 1337
Immune parameters were determined through differential hemocyte count in the 1338
hemolymph. There was an increase in the average final weight of shrimps treated with 1339
NCLA (p< 0.05). In gills, increased GST activity (p< 0.05) was verified in shrimps 1340
reared in SW in both treatments (WNC and NCLA). In the same organ, higher GSH 1341
levels were observed in shrimps maintained in BFT (p< 0.05), and decreased levels of 1342
lipid peroxidation was observed in shrimps supplemented with NCLA (p< 0.05). In 1343
hepatopancreas, both in SW and BFT, NCLA induced an increase of GST activity and 1344
GSH concentration, the lipid peroxidation was increased in BFT and NCLA treatment. 1345
The muscle was low levels of GST activity, there was no significant difference between 1346
treatments in GSH concentration, and there was an increased lipid peroxidation. There 1347
was a decrease in the percentage of hyaline hemocytes and increased granular 1348
hemocytes in all treatments. The nanoencapsulated lipoic acid had tissue-specific action, 1349
with increased antioxidant activity in gills and hepatopancreas. 1350
1351

47
Keywords: Biofloc System, Nanotechnology, Shrimp, Antioxidants supplements, 1352
Antioxidant defenses, Oxidative damage. 1353
1354
1. INTRODUCTION 1355
Intensification of production in carciniculture demand priority attention in welfare of 1356
reared organisms, as well as an integrated water management to avoid or minimize 1357
pollution of water bodies (i.e.: release of nitrogenous compounds) and a greater control 1358
over infection of pathogens (Xie and Yu, 2007; Moss et al., 2012; Xu and Pan, 2013) 1359
These actions are particularly important for the rearing of the white shrimp Litopenaeus 1360
vannamei, one of the most marketed aquatic organisms worldwide (Xu and Pan, 2013). 1361
Biofloc technology systems (BFT) appears as a sustainable solution that enables the 1362
improvement of water quality with the removal of toxic nitrogen (ammonia, nitrite), 1363
highest growth performance through food supplement and digestive stimulant by 1364
biofloc organisms (Xu and Pan, 2013). BFT are rich in natural microorganisms and 1365
bioactive compounds, forming a microbial culture through heterotrophic aggregates of 1366
organic particulate in suspended matter (Ray et al., 2010; Xu and Pan, 2013). 1367
Animal welfare is compromised by several factors, including reactive oxygen species 1368
(ROS) that are generated as oxygen metabolism products in aerobic organisms (Vergely 1369
et al., 2003; Rochette et al., 2013). However, conditions of oxidative stress are 1370
generated when exist a redox imbalance between pro-oxidants and antioxidants causing 1371
damage to macromolecules. Several conditions can lead to a scenario of oxidative 1372
stress, including factors that simultaneously increase ROS generation and reduce 1373
antioxidant activity (Jones, 2006; Oudot et al., 2006; Rochette et al., 2013). To prevent 1374

48
or ameliorate this imbalance, organisms have developed defense mechanisms like 1375
antioxidants. 1376
An antioxidant (enzymatic and non-enzymatic) is any substance that, when present in 1377
low concentrations compared to an oxidable substrate, significantly delays or inhibits 1378
oxidation of this substrate (Halliwell and Gutteridge, 1995; Rochette et al., 2013). 1379
Lipoic acid (LA), also known as thioctic acid, is a naturally compound synthesized in 1380
small amounts by plants and animals. LA makes covalent bonds with specific proteins, 1381
acting as co-factor for mitochondrial enzyme complexes (Carreau, 1979; Reed, 2001). 1382
In addition, LA neutralize ROS, chelates metals and regenerates antioxidants (Rochette 1383
et al., 2013; Monserrat et al., 2014). 1384
LA is a hydro and liposoluble antioxidant molecule, extensively distributed in cell 1385
membranes, cytosol, and extracellular spaces. When supplemented in the diet may 1386
accumulate in several tissues and a fraction is reduced to DHLA (dihydrolipoic acid), 1387
where the amount of this reduction is tissue-specific (Rochette et al., 2013; Monserrat et 1388
al., 2014). LA administered in the diet can reduce oxidative stress and restore levels of 1389
other antioxidants that may be at low levels (Monserrat et al., 2008; Martins et al., 1390
2014). 1391
However LA is an unstable compound, and being subject to degradation by thermal and 1392
photochemical degradation, acidic oxidation and thus the use of polymeric nanocapsules 1393
(lipid) for the protection and slow release of the antioxidant has been considered and 1394
applied in aquatic organisms as carp Cyprinus carpio (Longaray-Garcia et al., 2013). 1395
Nanocapsules are nanostructured vesicular systems carriers of with diameters between 1396
200 and 300 nm and low polydispersity, which stand out due to their capabilities in 1397

49
controlling the release of substances and the ability to increase the stability of the 1398
compound, both in storage and in the biological fluid (Külkamp-Guerreiro et al., 2009). 1399
The cellular stress leads to the activation of defense mechanisms resulting in increase in 1400
the detoxification capacity (Kültz, 2005; Roncalli et al., 2015). One of the detoxification 1401
enzymes belongs to the glutathione S-transferase (GST) family, proteins of 200-250 1402
amino acids that are activated in response to oxidative damage and/or exposure to a 1403
variety of xenobiotics (Frova, 2006; Sheehan et al., 2001; Roncalli et al., 2015). GST 1404
enzymes catalyze the conjugation of reduced glutathione (GSH) with hydrophobic 1405
molecules, enhancing their solubility and further excretion (Ranson and Hemingway, 1406
2005; Roncalli et al., 2015). 1407
Reduced glutathione (GSH) is an intracellular antioxidant, which assists in 1408
detoxification and elimination of ROS and maintains the cellular redox equilibrium 1409
(Rochette et al., 2013). Studies in rats suggest that LA can increase GSH synthesis in 1410
aged rats via increased expression of γ- glutamate cysteine ligase (GCL), the limiting 1411
enzyme for GSH generation (Suh et al., 2004a). 1412
According to Amado et al. (2009), it is also necessary to evaluate oxidative damage and 1413
the ability of each tissue in neutralizing reactive oxygen species. Therefore, non-specific 1414
responses as thiobarbituric acid reactive substances (TBARS) have been measured in 1415
this work. Measurement of lipid peroxidation through TBARS assay offers information 1416
about phospholipid peroxidation of cell membranes (Gorbi et al., 2008; Diaz-Jaramillo 1417
et al., 2013). 1418
The shrimp immune defense is performed by hemocytes present in the hemolymph and 1419
the measurement of number and kind of hemocytes may indicate the shrimp welfare 1420

50
(Perazzolo et al., 2002). Hyaline cells have the immune function perform phagocytosis, 1421
the semigranular cells perform encapsulation, phagocytosis (limited), storage and 1422
release of prophenoloxidase system and cytotoxic activity, and granule cells has the 1423
function of storing and releasing the prophenoloxidase system and cytotoxic activity 1424
(Johansson et al., 2000). In this study was the analysis of the immune response of the 1425
shrimp L. vannmaei by differential hemocyte count in the hemolymph, by hyaline and 1426
granular hemocyte percentage and contribute to the wellness paranorama in front of the 1427
use of nanoencapsulated lipoic acid (NCLA) or nanocapsule without lipoic acid (empty 1428
nanocapsule – NC) in shrimp. 1429
The objective of the study was therefore to determine the animal welfare conditions 1430
using NCLA in clean water and in a biofloc system, through biochemical analysis of 1431
antioxidant and oxidative damage parameters in gills, hepatopancreas and muscle, plus 1432
immune responses determined by differential hemocyte count in the hemolymph of 1433
Pacific white shrimp L. vannmaei. 1434
1435
2. MATERIAL AND METHODS 1436
It was used juvenile shrimp (L. vannamei) reared at the Marine Aquaculture Station 1437
(EMA) of the Federal University of Rio Grande – FURG (Brazil). The shrimps were 1438
stored at a density of 15 shrimp per tank (about 210 shrimp per m3), arranged in 12 1439
plastic tanks with 50 L (useful volume) each. The shrimps were submitted to four 1440
different treatments: (i) shrimps maintained in saline water and fed with ration 1441
supplemented with nanocapsules without lipoic acid (SW-NC), (ii) shrimps mantained 1442
in saline water and fed with ration supplemented with nanoencapsulated lipoic acid 1443

51
(SW-NCLA), (iii) shrimps maintained in water with bioflocs and fed with ration 1444
supplemented with nanocapsules without lipoic acid (BFT-NC, (iv) shrimps reared in 1445
water with bioflocs and fed with ration supplemented with nanoencapsulated lipoic acid 1446
(BFT-NCLA). In the two paragraphs below are described the methodologies used to 1447
prepare the nanocapsules suspensions and how they were included in the ration. 1448
Nanocapsules containing lipoic acid were prepared according to Longaray-Garcia et al. 1449
(2013). Briefly, lipoic acid (70 mg/ml) was dissolved in the organic phase composed of 1450
triglycerides of caprylic and capric acid, sorbitan monostearate, poly (-caprolactone), 1451
acetone and butylated hydroxytoluene. The organic phase was disposed over an aqueous 1452
phase in the dark that contained polysorbate 80, diazolidinilureia and Milli-Q water and 1453
stirred for 10 min. The formulation was evaporated at 35 °C until a final volume of 10 1454
ml, obtaining nanocapsules, which included lipoic acid (NCLA). Nanocapsules 1455
containing all the components cited above except the inclusion of lipoic acid were also 1456
prepared and represented the nanocapsules control (NC) (Külkamp-Guerreiro et al., 1457
2009). The mean diameter and polydispersion of suspended nanocapsules were 1458
measured through dynamic light scattering employing the equipment Zetasizer Nano 1459
series Nano-Zs (Malvern Instruments). Samples were diluted (1:500 v/v) in Milli-Q 1460
water, and the size distribution by intensity was analyzed in triplicate. Determination of 1461
lipoic acid in diet was performed through liquid chromatography with mass 1462
spectrometric detection, following the procedures described in Martins et al. (2014). 1463
Commercial diet SUPRA® with 35% crude protein was grinded and mixed with 1464
dispersions of NC or NCLA. The homogeneous mass was passed through a 5 mL 1465
syringe, and the obtained mass filaments were placed in oven at 50 °C, for 24 h before 1466
obtaining the pellets. The diet was provided three times a day (8:00, 15:00 and 22:00 h) 1467

52
at a feeding rate of 3% of shrimp biomass. The concentration of LA present in the ration 1468
was estimated in 68 ± 8.6 mg LA/Kg. 1469
Physical and chemical parameters of water were measured daily, including dissolved 1470
oxygen (mg/L), water temperature (°C), salinity and pH. Ammonia and nitrite was 1471
measured at intervals of one day and nitrate once a week. When the total ammonia 1472
concentration exceeded 1 mg/L, molasses were added to BFT water (Avnimelech, 1473
1999). The experiment lasted 30 days, where there was no water changes in BFT 1474
treatment. In the treatment of saline water, 80-90% of water was renewed every 48 h. 1475
Parameters of water quality presented significant differences (p< 0.05) for the 1476
concentration of dissolved oxygen, being lower at BFT. Nitrite concentration also was 1477
lower at BFT, although nitrate levels were augmented in BFT (Table 1). 1478
At the beginning and the end of the experiment, shrimps from the four treatment were 1479
weighed. At the end of the experiment, the hemolymph was collected from shrimp heart 1480
using a 3 mL syringe (see details in section 2.6). After, shrimps were frozen and in the 1481
next day the tissues gills, hepatopancreas and muscle from the second abdominal somite 1482
were dissected and stored in ultrafreezer at -80 oC. 1483
Organ samples were homogenized (1:5, w/v) in a buffer previously employed for 1484
crustacean species (Pinho et al., 2005; Sousa et al., 2014) composed of Tris-Base (20 1485
mM), EDTA (1 mM), dithiothreitol (1 mM; Sigma), sucrose (500 mM), KCl (150 mM) 1486
and dissolved in Milli-Q water, with pH adjusted to 7.2. Homogenates were centrifuged 1487
(9,000 x g, 30 min at 4°C) and the supernatants were kept and divided into three 1488
aliquots for later analysis of glutathione-S-transferase (GST) activity, levels of lipid 1489
peroxidation (TBARS assay) and levels of reduced glutathione (GSH). 1490

53
2.1. DETERMINATION OF GLUTATHIONE S-TRANSFERASE ACTIVITY (GST) 1491
Glutathione-S-transferases was measured according to the protocol of Habig et al. 1492
(1974). The conjugation of 1 mM of 1-chloro-2,4-dinitrobenzene (CDNB, Sigma) with 1493
1 mM of reduced glutathione (GSH, Sigma) in presence of 20 µL of sample 1494
homogenates (source of GST enzyme) generated a CDNB-GSH complex. The 1495
absorbance of this complex was measured at 340 nm in a spectrofluorometer with a 1496
microplate reader (Victor 2, Perkin Elmer). 1497
1498
2.2. DETERMINATION OF CONCENTRATION OF REDUCED GLUTATHIONE 1499
(GSH) 1500
The procedure was based on White et al. (2003). The substrate 2,3 naphthalene 1501
dicarboxialdehyde (NDA, Invitrogen) reacts with GSH to form a fluorogenic 1502
compound. Aliquots from each homogenates were transferred to white ELISA 96-well 1503
microplates. After, it was added to all wells the NDA solution, and after 30 min of 1504
incubation, the fluorescence intensity of the NDA-GSH complex was read at excitation 1505
and emission wavelength of 485 and 530 nm, respectively, using a plate reader 1506
fluorometer (Victor 2, Perkin Elmer). A standard curve of GSH was prepared to express 1507
the data in terms of µM of GSH/mg of total proteins. 1508
1509
2.4. DETERMINATION OF LIPID PEROXIDATION 1510
Lipid peroxidation was determined by TBARS (thiobarbituric acid reactive substances) 1511
assay following Oakes and Van der Kraak (2003). Aliquots (20 µl) of sample 1512
homogenates were added to glass tubes, allowing the reaction with thiobarbituric acid 1513

54
(0.8%, Sigma), 20% acetic acid, Milli Q water and 8.1 % of sodium dodecyl sulfate. 1514
The mixture was incubated at 95 oC during 30 min and after cooling it was added n-1515
buthanol. Following a centrifugation step (3,000 x g during 10 min at 15 oC), the n-1516
buthanol phase was pippeted to a white ELISA 96-well microplate and the fluorescence 1517
read (excitation: 520 nm; emission: 580 nm) in a fluorometer with a microplate reader 1518
(Victor 2, Perkin Elmer). As standard it was employed 1,1,3,3-tetramethoxypropane 1519
(TMP, Across Organics). The results were expressed as nmol equivalents of TMP per 1520
mg of fresh tissue. 1521
1522
2.5. DETERMINATION OF TOTAL AMMONIA 1523
Total ammonia determination (NH3 + NH4+) follows the UNESCO (1983) method in 1524
which ammonia in basic pH (between 10.8 and 11.5) reacts with a bleaching solution, 1525
forming monochloramine. In the presence of phenol and an excess of hypochlorite, 1526
addition of sodium nitroprusside as a catalyst generates indophenol blue, a molecule 1527
that absorbs at 630 nm. 1528
1529
2.6. DIFFERENTIAL HEMOCYTE COUNT (DHC) 1530
The hemolymph was collected by heart puncture using 3 mL syringe containing 1531
anticoagulant crustacean solution, containing NaCl (450 mM), glucose (100 mM), 1532
sodium citrate (30 mM), citric acid (23 mM), EDTA (20 mM) diluted in MilliQ water, 1533
with pH fixed at 7.4. After, a smear of hemolymph was made on glass slides that were 1534
fixed in metanol for 5 minutes a stained with May-Grunwald-Giemsa. Then, hemocytes 1535
were quantified by microscope eyepiece lens Integrating Disc 1, 25 points-G49 (Carl 1536
Zeiss), following methodology of Weibel (1980). 1537

55
2.7. STATISTICAL ANALYSIS 1538
Data were expressed as mean ± standard deviation. Each variable from each organ, 1539
including hemolymph (GST activity, lipid peroxidation, total antioxidant capacity and 1540
total hemocyte count) was analyzed through bi-factorial ANOVA, being the factors the 1541
treatments (NC or NCLA) and rearing condition (SW or BFT). Previously the 1542
assumptions of normality and homogeneity of variance were analyzed. Means 1543
comparisons were done using the Newman-Keuls test. Water physic-chemical 1544
parameters were analyzed using Student t test for unequal variances (Zar 1984). In all 1545
cases it was used a significance level of 5%. 1546
1547
3. RESULTS 1548
The initial weight was not significantly different between treatments (p> 0.05), but after 1549
30 days of experiment, shrimps that were supplemented with NCLA showed higher 1550
weight, both in SW and BFT, although the maximum increase was observed in BFT-1551
NCLA treatment. In every case survival was of 100% (Table 2). 1552
The GST activity was higher in gills of shrimp of SW-NC in SW and lower in BFT-NC 1553
when compared with the other two treatments (p< 0.05) (Figure 1a). In hepatopancreas 1554
the highest GST activity was observed in BFT-NCLA and the minimum in the treatment 1555
SW-NC (p< 0.05). In muscle, there was no significant difference in GST activity 1556
between treatments (p> 0.05) (Figure 1a). 1557
In gills, the peak of GSH concentration was observed in treatment BFT-NCLA, being 1558
different from BFT-NC treatment (p <0.05), whereas no differences were observed in 1559
SW (Figure 1b). In hepatopancreas, treatment with NCLA induced higher GSH levels 1560

56
(p< 0.05) both in SW and in BFT. However, the BFT-NCLA showed the highest GSH 1561
level (Figure 1b). Muscle was the organ that presented highest GSH levels, although no 1562
influence of treatments was verified (p> 0.05) (Figure 1b). 1563
In gills, shrimps from SW-NC presented the highest TBARS levels, an effect that was 1564
lowered in SW-NCLA treatment (p< 0.05). Shrimps maintained in BFT presented the 1565
lowest TBARS levels, both for NC and NCLA (p< 0.05; Figure. 1c). In hepatopancreas 1566
a significant (p< 0.05) peak of TBARS was registered in BFT-NCLA, being different 1567
from the rest of the treatments (Figure 1c). In muscle, the treatment SW-NC showed the 1568
highest TBARS levels, being different from the other treatments (p< 0.05) (Figure 1c). 1569
Hemocytes counts were different in the four treatments. The percentage of hyaline 1570
hemocytes was highest in SW-NC treatment, and lowest in BFT-NCLA. Values for SW-1571
NCLA and BFT-NC remained intermediate between those registered in SW-NC and 1572
BFT-NCLA (p< 0.05; Figure 2). An opposite response was observed for percentage of 1573
granular hemocytes, being highest for BFT-NCLA and lowest for SW-NC treatment, 1574
where SW-NCLA and BFT-NC responses remained intermediate (p< 0.05; Figure 2). 1575
1576
4. DISCUSSION 1577
Water quality parameters were in accordance with proposed parameters for the 1578
Litopenaeus vannamei species proposed by Samocha et al. (2010) and Ray et al. (2014). 1579
These parameters were also in according with previous experiment Martins et al. (2015) 1580
with saline water and biofloc, except that salinity in such biofloc experiment was 33. 1581
Ammonia and nitrite concentrations remained below of the levels considered toxic for 1582
this species (Lin and Chen, 2001; Lin and Chen, 2003). The low concentration of nitrite 1583

57
and high concentration of nitrate in BFT indicate heterotrophic bacteria activity in the 1584
conversion of nitrogen products (Ebeling et al., 2006). There was an increase in the 1585
average final weight of the treatments with LA, indicating that this zootechnical 1586
parameter is improved with supplementation of LA for Litopenaueus vannamei shrimp 1587
in clean water and biofloc, as previously reported by Martins et al. (2014) for the same 1588
species reared in saline water when supplemented with lipoic acid at doses higher than 1589
190 mg LA/kg of ration. 1590
Shrimps as any other aerobic organism, possess an integrated and coordinated 1591
enzymatic and non-enzymatic antioxidant systems that maintain the redox equilibrium 1592
of the organism (Ren et al., 2014). Under this context, it is expected that lipoic acid 1593
supplementation should aid in the establishment of the redox system and thus for the 1594
organism welfare. 1595
Previous studies have showed several beneficial effects of lipoic acid in aquatic 1596
organisms (for a review: Kütter et al., 2014), although the responses are strongly organ-1597
dependent (Monserrat et al., 2008; Kütter et al., 2013). Some of the positive effects 1598
include: high GST activity, low concentration in thiobarbituric reactive substances in 1599
different organs, higher levels of GSH and of the limiting enzyme in their synthesis 1600
(glutamate cysteine ligase) (Monserrat et al., 2008; Martins et al., 2014; Monserrat et 1601
al., 2014; Enamorado et al., 2015). The lower GST activity observed in SW-NCLA 1602
when compared with SW-NC fits to the previous observation of Monserrat et al. (2014), 1603
where LA also reduced GST activity in gills of Jenynsia multidentata. These authors 1604
postulated that high levels of the antioxidant in these organs should favor a reductive 1605
redox state that should inhibit the up-regulation of genes related to antioxidant and 1606
detoxification processes. This idea is also supported by the results of Kütter et al. 1607

58
(2013), where in liver of pompano fish Trachinotus marginatus, high LA doses (40 and 1608
60 mg LA/kg of body weight) lowered GST activity and at a lower one (20 mg LA/kg 1609
of body weight) it was registered a significant increase of enzyme activity. 1610
By the other side, the hepatopancreas presented high GST activity in shrimps reared in 1611
SW and BF and submitted to NCLA, a result similar to that found by Kütter et al. 1612
(2013) with low doses of free LA. A previous study of Lobato et al. (2013) also showed 1613
a similar result in the same species, L. vannamei, even when the free LA dose employed 1614
by those authors was higher than in present study. The comparison of GST results in 1615
gills and hepatopancreas suggest that the biodistribution and entry of LA in the different 1616
organs should determine strong or moderate reductive redox state and this should 1617
influence GST activity. The tripeptide-reduced glutathione (GSH) is used in conjugation 1618
reactions catalyzed by GST and also acts as a first barrier against reactive oxygen 1619
species (ROS). In this way, increased levels of this antioxidant in hepatopancreas of 1620
shrimps from experimental groups SW-NCLA and BFT-NCLA indicate the antioxidant 1621
responses triggered by LA. Up-regulation of genes that code for both modulatory and 1622
catalytic sub-units of glutamate cysteine ligase (the limiting enzyme for GSH synthesis) 1623
by LA has been described in rats (Suh et al., 2004). In gills, the same effect was 1624
observed only in shrimps reared in BFT indicating that in this case some components, 1625
including antioxidants present in bioflocs as reported by Xu et al. (2013), should aid to 1626
obtain a better antioxidant response that in saline water. Also Martins et al. (2015) 1627
reported that shrimps reared in BFT presented different antioxidant responses than 1628
shrimps maintained in saline water. In the case of muscle, the so much higher GSH 1629
levels indicate that the antioxidant system is expressed near its maximum and few 1630

59
regulations can occur even for the antioxidants present in the bioflocs or by the 1631
exogenous LA offered through ration. 1632
In terms of oxidative damage, the antioxidant treatment with LA was effective only in 1633
SW, where a reduction of TBARS levels were observed in gills and muscle, as 1634
previously reported by Lobato et al. (2013) in muscle of the same species L. vannamei. 1635
Coincident with this previous study no effect was observed in hepatopancreas. The 1636
results in BFT were quite different, since a pro-oxidant condition was verified in 1637
hepatopancreas when exposed to NCLA. The antioxidant/pro-oxidant duality of LA has 1638
been recognized by several authors like Çakatay et al. (2005), Atukeren et al. (2010), 1639
and Valdecantos et al. (2010). The pro-oxidant condition of LA is related to its ability to 1640
reduce Fe3+ to Fe2+, favoring the generation of hydroxyl radical and thus the triggering 1641
of lipid peroxidation (Goralska et al., 2003). The reduced form of lipoic acid, 1642
dihydrolipoic acid (DHLA) possess the ability to remove iron from proteins like ferritin, 1643
again favoring the Fenton reaction and hydroxyl radical generation (Çakatay et al., 1644
2005). Several enzymes, including glutathione reductase that employs GSH as reductive 1645
power, favor the reduction of LA to DHLA (for a review: Kütter et al., 2014). Note that 1646
hepatopancreas GSH levels are higher in shrimps from BFT-NCLA (824.75±34.64 µM 1647
of GSH mg-1 of total proteins) than the values presented by shrimps from SW-NCLA 1648
group (526.36 µM of GSH mg-1 of total proteins). Thus, a greater reductive power in 1649
shrimps from BFT-NCLA group should favor the generation of DHLA and thus 1650
promoting iron release from proteins. 1651
On granular hemocytes (GH) are found degradative and coagulation enzymes, 1652
antimicrobial peptides, apopitotics factors and prophenoloxidase system, these have 1653
higher amount immunoefetoras and immunoregulatory molecules are considered more 1654

60
immunocompetent (Barracco et al., 2008). Some immune responses are associated with 1655
the production of reactive oxygen species (ROS) and are highly microbicides (Bachère 1656
et al., 1995; Muñoz et al., 2000; Campa-Cordova et al., 2002). The process begins when 1657
stimulation leads to an increase in oxygen consumption, whose reduction, catalyzed by 1658
NADPH-oxidase bound to membrane yields superoxide anion (Campa-Cordova et al., 1659
2002). The antioxidant superoxide dismutase (SOD) converts this metabolite 1660
microbicide in hydrogen peroxide which passes freely through membranes (Campa-1661
Cordova et al., 2002). The antioxidant catalase (CAT) and glutathione peroxidase (GPx) 1662
remove hydrogen peroxide from the cells (Campa-Córdova et al., 2002). A significant 1663
quantity passes into the extracellular environment and can cause damage to cells 1664
(Warner, 1994; Campa-Cordova et al., 2002). To avoid this damage, cells and organisms 1665
use three defense strategies; one involves low molecular weight antioxidants such as 1666
ascorbic acid, α-tocopherol and glutathione, which can directly interact with ROS and 1667
neutralizing those (Campa-Cordova et al., 2002). The other two involve a variety of 1668
enzymes that metabolize ROS (SOD, catalase, glutathione peroxidase) or 1669
macromolecular damage repair as nucleic acids, proteins and lipids (DNA repair 1670
enzymes, proteases, lipases, etc.) caused by ROS (Warner, 1994; Campa-Cordova et al., 1671
2002). 1672
In NCLA treatments was increased in HH that is explained by the fact in this 1673
study were not exposed to any pathogen so no requiring activation of components 1674
associated with phagocytosis and/or coagulation. Thus, there is an increased percentage 1675
of GH with a greatest amount of immunoefetoras cell components and 1676
immunoregulatory to protect the shrimp against possible pathogenic attacks. There little 1677
research with differential hemocyte count, being more found total hemocyte count, and 1678
other study with compounds supplemented becomes delicate comparisons by the 1679
difference in composition and functionality of the compound. In conclusion, NCLA 1680
treatments improved the immune response by increasing the number of granular 1681
hemocytes. 1682
1683
1684

61
5. CONCLUSION 1685
The GST activity was tissue-specific, with greater activity in gills. However, the NLA 1686
induced increase in GST activity in the hepatopancreas. Likewise that the GST activity 1687
the GSH concentration is tissue-specific with greater performance in the muscle 1688
although not presenting significant difference between treatments had higher 1689
concentration in this tissue has direct antioxidant activity and not as a substrate for 1690
detoxification activity. Antioxidant capacity also showed a standard tissue-specific, with 1691
higher antioxidant capacity hepatopancreas no evident action of the performance of 1692
NLA against peroxyl radicals. Lipid peroxidation levels were lower in muscle, with 1693
obvious performance NLA action. It can say that the groups with NLA there was an 1694
increase in the percentage of granular hemocytes, cells with higher amounts of 1695
components immunocompetents, but cannot say that the NLA is a compound with 1696
immunostimulatory behavior. 1697
1698
Acknowledgments. José Maria Monserrat and Wilson Wasielesky Junior receive 1699
productivity research fellowships from the Brazilian agency CNPq (process numbers 1700
PQ 307880/2013-3 and PQ 310993/2013-0, respectively). 1701
1702
1703
1704
1705
1706
1707
1708

62
REFERENCES 1709
Amado, L.L., Garcia, M.L., Ramos, P.B., Freitas, R.F., Zafalon, B., Ferreira J.L.R., 1710
Yunes, J.S., & Monserrat, J.M., 2009. A method to measure total antioxidant 1711
capacity against peroxyl radicals in aquatic organisms: Application to evaluate 1712
microcystins toxicity. Sci. Total Environ. 407, 2115–2123. 1713
Atukeren, P., Aydin, S., Uslu, E., Gumustas, K.M., & Çakatay, U., 2010. Redox 1714
homeostasis of albumin in relation to alfa-lipoic acid and dihydrolipoic acid. 1715
Oxid. Med. Cell Longev. 3, 206–213. 1716
Barracco, M.A., Ranzanipaiva, M.J.T., Takemoto, R.M., & Lizama, M.A.P., 2004. 1717
Mecanismos de resistência à doença em crustáceos. Sanidade de Organismos 1718
Aquáticos. Ed. Varela, p. 49–72. 1719
Bachère, E., Mialhe, E., Nöel, D., Boulo, V., Morvan, A., & Rodriguez, J., 1995. 1720
Knowledge and research prospects in marine mollusk and crustacean 1721
immunology. Aquaculture. 132, 17–32. 1722
Çakatay, U., Kayali, R., Sivas, A., & Tekeli, F., 2005. Prooxidant activities of alfa-lipoic 1723
acid on oxidative protein damage in the aging rat heart muscle. Arch Gerontol 1724
Geriatr. 40, 231–240. 1725
Campa-Córdova, A.I., Hernández-Saavedra, N.Y., & Ascencio, F., 2002. Superoxide 1726
dismutase as modulator of immune function in American white shrimp 1727
(Litopenaeus vannamei). Comp. Biochem. Physiol. C. 133, 557–565. 1728
Carreau, J.P., 1979. Biosynthesis of lipoic acid via unsaturated fatty acids. Method 1729
Enzymol. 62, 152–158. 1730
Ebeling, J.M., Timmons, M.B., & Bisogni, J.J., 2006. Engineering analysis of the stoi-1731
chiometry of photoautotrophic, autotrophic, and heterotrophic removal of 1732
ammonia–nitrogen in aquaculture systems. Aquaculture. 257, 346–358. 1733
Enamorado, A.M., Martins, A.C., Flores J.A., Tesser, M.B., Caldas, S.S., Primel, E.G., 1734
& Monserrat, J.M., 2015. Biochemical responses over time in common carp 1735
Cyprinus carpio (Teleostei, Cyprinidae) during fed supplementation with α-1736
lipoic acid. Comp. Biochem. Physiol. A. 188, 9–16. 1737

63
Frova, C., 2006. Glutathione transferases in the genomics era: new insights and 1738
perspectives. Biomol. Eng. 23, 149–169. 1739
Goralska, M., Dackor, R., Holley, B., & McGahan, M.C., 2003. Alpha lipoic acid 1740
changes iron uptake and storage in lens epithelial cells. Exp. Eye Res. 76, 241–1741
248. 1742
Habig, W.H., Pabst, M.J., & Jakoby, W.B., 1974. Glutathione S-transferases: The first 1743
enzymatic step in mercapturic acid formation. J. Biol. Chem. 249, 7130–7139. 1744
Halliwell, B., Gutteridge, J.M., 1995. The definition and measurement of antioxidants in 1745
biological systems. Free Radic. Biol. Med. 18, 125–126. 1746
Johansson, M.W., Keyser, P., Sritunyalucksana, K., & Söderhäll, K., 2000. Crustacean 1747
haemocytes and haematopoiesis. Aquaculture. 191, 45–52. 1748
Jones, D.P., 2006. Redefining oxidative stress. Antioxid Redox Signal. 8, 1865–1879. 1749
Ju, Z.Y., Forster, I., Conquest, L., & Dominy, W., 2008a. Enhanced growth effects on 1750
shrimp (Litopenaeus vannamei) from inclusion of whole shrimp floc or floc 1751
fractions to a formulated diet. Aquacul. Nutr. 14, 533–543. 1752
Külkamp-Guerreiro, I.C., Guterres, S.S., & Pohlmann, A.R., 2009. Estabilização da 1753
ácido lipóico via encapsulamento em nanocápsulas poliméricas planejadas para 1754
aplicação cutânea. Quím. Nova. 8: 2078–2084. 1755
Kütter, M.T., Monserrat, J.M., Primel, E.G., Caldas, S.S., & Tesser, M.B., 2013. Effects 1756
of dietary α-lipoic acid on growth, body composition and antioxidant status in 1757
the Plata pompano Trachinotus marginatus (Pisces, Carangidae). Aquaculture. 1758
368–369, 29–35. 1759
Kütter, M.T., Romano, L.A., Ventura-Lima, J., Tesser, M.B., & Monserrat, J.M., 2014. 1760
Antioxidant and toxicological effects elicited by alpha-lipoic acid in aquatic 1761
organisms. Compa Biochem. Physiol C. 162, 70–76. 1762
Kültz, D., 2005. Molecular and evolutionary basis of the cellulr stress response. Annu. 1763
Rev. Physiol. 67, 225–257. 1764

64
Lin, Y.C., & Chen, J.C., 2001. Acute toxicity of ammonia on Litopenaeus vannamei 1765
(Boone) juveniles at different salinity levels. J. Exp. Mar. Biol. Ecol. 259, 109–1766
119. 1767
Lin, Y.C., & Chen, J.C., 2003. Acute toxicity of nitrite on Litopenaeus vannamei 1768
(Boone) juveniles at different salinity levels. Aquaculture. 224, 193–201. 1769
Lobato, R.O., Nunes, S.M., Wasielesky Jr., W., Fattorini, D., Regoli, F., Monserrat, 1770
J.M., & Ventura-Lima, J., 2013. The role of lipoic acid in the protection against 1771
of metallic pollutant effects in the shrimp Litopenaeus vannamei (Crustacea, 1772
Decapoda). Comp. Biochem. Physiol A. 165, 491–497. 1773
Longaray-Garcia, M., Flores, J.A., Külkamp-Guerreiro, I.C., Guterres, S.S., Carneiro, 1774
T.C. Brandão, M.R.B., & Monserrat, J.M., 2013. Modulation of antioxidant and 1775
detoxifying capacity in fish Cyprinus carpio (Cyprinidae) after treatment with 1776
nanocapsules containing lipoic acid. Comp. Biochem. Physiol. A. 165, 468–475. 1777
Martins, Á.C.S., Flores, J.A., Wasielesky Jr, W., Zanette, J., Primel E.G., Caldas, S.S., & 1778
Monserrat, J.M., 2014. Modulation of antioxidant and detoxification responses 1779
induced by lipoic acid in the Pacific white shrimp Litopenaeus vannamei 1780
(Boone, 1931) submitted to hypoxia and re-oxygenation. Mar. Freshw. Behav. 1781
Phy. 47, 335–348. 1782
Martins, Á.C.S., Flores, J.A., Porto, C., Wasielesky Jr, W., & Monserrat, J.M., 2015. 1783
Antioxidant and oxidative damage responses in different organs of Pacific white 1784
shrimp Litopenaeus vannamei (Boone, 1931) reared in a biofloc technology 1785
system. Mar. Freshw. Behav. Phy. 48, 279–288. 1786
Monserrat, J.M., Ventura-Lima, J., Ferreira, J.L.R., Acosta, D., Longaray-Garcia, M., 1787
Ramos, P.B., Moraes, T.B., Dos Santos, L.C., & Amado, L.L., 2008. Modulation 1788
of antioxidant and detoxification responses mediated by lipoic acid in the fish 1789
Corydoras paleatus (Callychthyidae). Comp. Biochem. Physiol. C. 148, 287–1790
292. 1791
Monserrat, J.M., Longaray-Garcia, M., Ventura-Lima, J., Gonzalez, M., Ballesteros, 1792
M.L., Miglioranza, K.S., Ame, M.V., & Wunderlin, D.A., 2014. Antioxidant, 1793
phase II and III responses induced by lipoic acid in the fish Jenynsia 1794

65
multidentata (Anablapidae) and its influence on endolsulfan accumulation and 1795
toxicity. Pestic. Biochem. Phys. 108, 8–15. 1796
Moss, S.M., Moss, D.R., Arce, S.M., Lightner, D.V., & Lotz, J.M., 2012. The role of 1797
selective breeding and biosecurity in the prevention of disease in penaeid shrimp 1798
aquaculture. J. Invertebr. Pathol. 110, 247–250. 1799
Muñoz, M., Cedeño, R., Rodríguez, J., Van der Knaap, W.P., Mialhe, E., & Bachère, E., 1800
2000. Measurement of reactive oxygen intermediate production in haemocytes 1801
of the penaeid shrimp, Penaeus vannamei. Aquaculture 191, 89–107. 1802
Oakes, K.D., & Van der Kraak, G.J., 2003. Utility of TBARS assay in detecting 1803
oxidative stress in white sucker (Catostomus commersoni) populations exposed 1804
to pulp mill effluent. Aquat Toxicol. 63, 447–463. 1805
Oudot, A., Martin, C., Busseuil, D., & Vergely, C., 2006. NADPH oxidases are in part 1806
responsible for increased cardiovascular superoxide production during aging. 1807
Free Radical Bio Med. 40, 2214–2222. 1808
Perazzolo, L.M., Gargioni, R., Ogliari, P., & Barracco, M.A., 2002. Evaluation of some 1809
hemato-immunological parameters in the shrimp Farfantepenaeus paulensis 1810
submitted to environmental and physiological stress. Aquaculture. 214, 19–33. 1811
Pinho, G.L., Da Rosa, C.M., Maciel, F.E., Bianchini, A., & Yunes, J.S., Proenca, L.A.O., 1812
& Monserrat, J.M., 2005. Antioxidant responses and oxidative stress after 1813
microcystin exposure in the hepatopancreas of an estuarine crab species. Ecotox. 1814
Environ. Safe. 61, 353–360. 1815
Ranson, H., & Hemingway, J., 2005. Mosquito glutathione transferases. Method 1816
Enzymol. 401, 226–241. 1817
Ray, A.J., Seaborn, G., Leffler, J.W., Wilde, S.B., Lawson, A., & Browdy, C.L., 2010. 1818
Characterization of microbial communities in minimal-exchange, intensive 1819
aquaculture systems and the effects of suspended solids management. 1820
Aquaculture. 310, 130–138. 1821
Reed, L.J., 2001. A trail of research from lipoic acid to alpha-keto acid dehydrogenase 1822
complexes. J. Biol. Chem. 276, 38329–38336. 1823

66
Ren, X., Pan, L., & Wang, L., 2014. Effect of florfenicol on selected parameters of 1824
immune and antioxidantes systems, and damage indexes of juvenile Litopenaeus 1825
vannamei following oral administration. Aquaculture. 432, 106–113. 1826
Repetto, M., Semprine, J., & Boveris, A., 2012. Lipid Peroxidation: Chemical 1827
mechanism, biological implications and analytical determination. Biochemistry, 1828
Genetics and Molecular Biology. INTECH. Chapter 1. p 3–30. 1829
Rochette, L., Ghibu, S., Richard, C., Zeller, M., Cottin, Y., & Vergely, C., 2013. Review: 1830
Direct and indirect antioxidant properties of α-lipoic acid and therapeutic 1831
potential. Mol. Nutr. Food Res. 57, 114–125. 1832
Roncalli, V., Cieslak, M.C., Passamaneck, Y., Christie, A.E., & Lenz, P.H., 2015. 1833
Glutathione S-Transferase (GST) gene diversity in the crustacean Canlaus 1834
finmarchicus– Contributors to cellular detoxification. PLOS ONE 10(5): 1835
e0123322. doi:10.1371/journal.pone.0123322. 1836
Samocha, T.M., Wilkenfeld J.S., Morris T.C., Correia E.S., Hanson T. 2010. Intensive 1837
raceways without water exchange analyzed for White shrimp culture. Global 1838
Aquaculture Advocate. 13:22–24. 1839
Sheehan, D., Foley, D.M., & Dowd, C.A., 2001. Structure, function and evolution of 1840
glutathione transferases: Implications for classification of non-mammalian 1841
members of an ancient enzyme superfamily. Biochem. J. 360, 1–16. 1842
Souza, D.M., Martins A.C., Jensen, L., Wasielesky Jr., W., Monserrat, J.M., & Garcia, 1843
L.O., 2014. Effect of temperature on antioxidant enzymatic activity in the 1844
Pacific white shrimp Litopenaeus vannamei in a BFT (Biofloc technology) 1845
system. Mar. Freshw. Behav. Phy. 47, 1–10. 1846
Suh, J.H., Wang, H., Liu, R.M., Liu, J., & Hagen, T.M., 2004a. (R)-alpha-lipoic acid 1847
reverses the age-related loss in GSH redox status in post-mitotic tissues: 1848
evidence for increased cysteine requirement for GSH synthesis. Arch. Biochem. 1849
Biophys. 423, 126–135. 1850
Suh, J.H., Swapna, V.S., Dixon B.M., Liu, H., Jaiswal, A.K., Liu, R-M., & Hagen, T.M., 1851
2004. Decline in transcriptional activity of Nrf-2 causes age-related loss of 1852

67
glutathione synthesis, which is reversible with lipoic acid. Proc. Natl. Acad. Sci 1853
USA. 101, 3381–3386. 1854
UNESCO, 1983. Chemical methods for use in marine environmental monitoring. Paris, 1855
Intergovernmental Oceanographic Commission. Paris, France. 53p. 1856
Valdecantos, M.P., Perez-Matute, P., Quintero, P., & Martinez, J.A., 2010. Vitamin C, 1857
resveratrol and lipoic acid actions on isolated rat liver mitochondria: All 1858
antioxidants but different. Redox Report. 15, 207–216. 1859
Vergely, C., Maupoil, V., Clermont, G., & Bril, A., 2003. Identification and 1860
quantification of free radicals during myocardial ischemia and reperfusion using 1861
electron paramagnetic resonance spectroscopy. Arch. Biochem. Biophys. 420, 1862
209–216. 1863
Warner, H.R., 1994. Superoxide dismutase, aging, and degenerative disease. Free Radic. 1864
Biol. Med. 3, 249–258. 1865
Weibel, E.R., 1980. Stereological Methods, 2. Academic, London 253–257. 1866
White, C.C., Viernes, H., Krejsa, C.M., Botta, D., & Kavanagh, T.J., 2003. Fluorescence 1867
based microtiter plate assay for glutamate-cysteine ligase activity. Anal. 1868
Biochem. 318, 175–180. 1869
Xie, B., Yu, K.J., 2007. Shrimp farming in China: Operating characteristics, 1870
environmental impact and perspectives. Ocean Coast. Manage. 50, 538–550. 1871
Xu, W-J., & Pan, L-Q., 2012. Effects of bioflocs on growth performance, digestive 1872
enzyme activity and body composition of juvenile Litopenaeus vannamei in 1873
zero-water exchange tanks manipulating C/N ratio in feed. Aquaculture. 356–1874
357, 147–152. 1875
Xu, W-J., Pan, L-Q., 2013. Enhancement of immune response and antioxidant status of 1876
Litopenaeus vannamei juvenile in biofloc-based culture tanks manipulating high 1877
C/N ratio of feed input. Aquaculture. 412–413, 117–124. 1878
Zar, J. H., 1984. Biostatistical analysis. New Jersey: Prentice Hall. 1879
1880

68
Table captions 1881
Table 1 – Parameters physical and chemical water quality on average function (± 1 1882
standard deviation). In saline water (SW) or in bioflocs system (BFS). Animals were fed 1883
with ration suplemented with nanocapsules containing lipoic acid (NCLA) or without 1884
the antioxidant (empty nanocapsules - NC). Asterisk (*) indicate significant differences 1885
(p <0.05) between groups. 1886
Parameter SW-NC SW-NCLA BFT-NC BFT-NCLA
Temperature °C (n=186) 27.01 (± 0,01) 27.01 (± 0,03) 27.03 (± 0,01) 27.01 (± 0,02)
[DO] (mg L-1) (n=186)* 6.45 (± 0.01) 6.51 (± 0.01) 5.56 (± 0.05) 5.61 (± 0.04)
Salinity (n=186) 31.00 (± 0.01) 31.00 (± 0.01) 31.98 (± 0.01) 31.96 (± 0.01)
pH (n=186) 7.89 (± 0.06) 7.94 (± 0.02) 8.00 (± 0.01) 7.82 (± 0.07)
Ammonia (mg L-1) (n=66) 1.72 (± 1.36) 1.71 (± 1.43) 1.84 (± 2.53) 1.90 (± 2.86)
Nitrite (mg L-1) (n=66)* 0.55 (± 0.75) 0.53 (± 0.69) 0.27 (± 0.39) 0.25 (± 0.33)
Nitrate (mg L-1) (n=30)* 3.64 (± 1.88) 0.18 (0.69) 84.60 (± 5.57) 80.90 (± 5.55)
1887
Table 2 – Average (± 1 standard deviation) initial and final weight in shrimp 1888
Litopenaeus vannamei reared in saline water (SW) or in bioflocs system (BFS). Animals 1889
were fed with ration suplemented with nanocapsules containing lipoic acid (NCLA) or 1890
without the antioxidant (white nanocapsules - WNC). Statistical differences between 1891
treatments were represented by different lower case letters (p<0.05) for both initial and 1892
final weight. Number of samples analyzed per treatment: 45. 1893
Treatments Average initial weight (g) Average final weight (g) Survival
SW-NC 5.90 (± 0.08)a 9.72 (± 0.06)c 100%
SW-NCLA
5.91 (± 0.07)a
11.85 (± 0.11)b
100%
BFT-NC
5.91 (± 0.04)a
9.43 (± 0.04)d
100%
BFT-NCLA
5.91 (± 0.01)a
12.34 (± 0,05)a
100%
1894
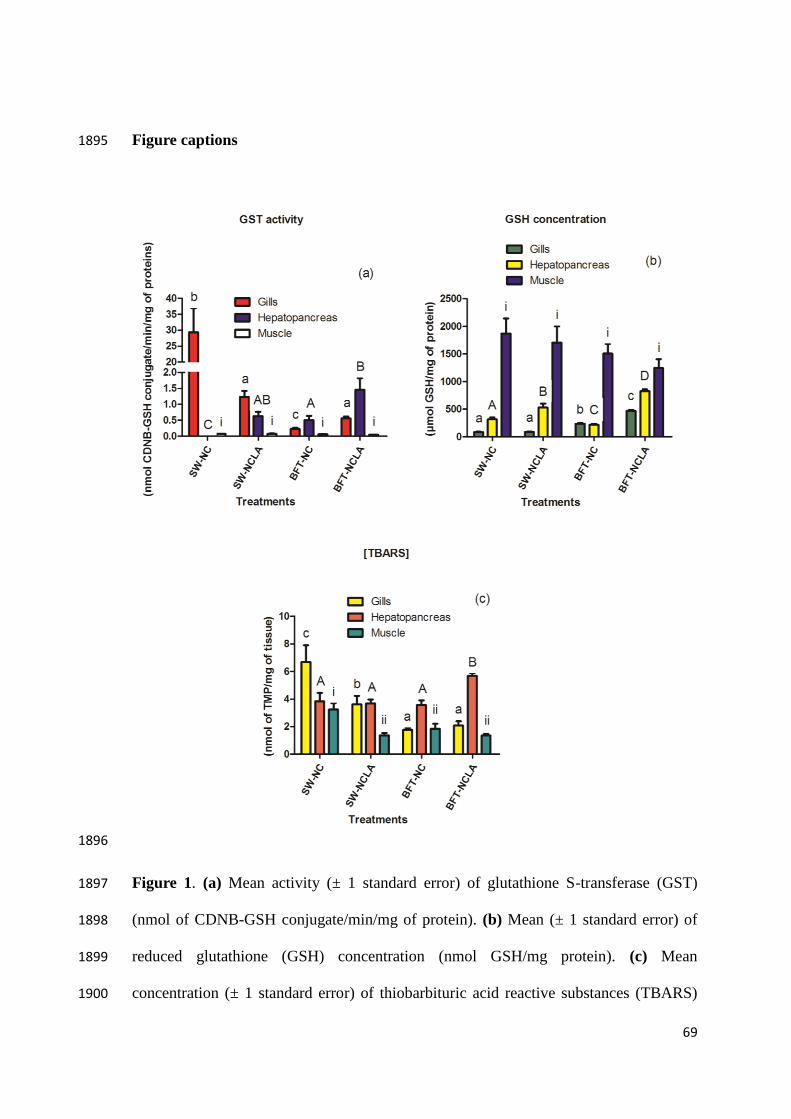
69
Figure captions 1895
1896
Figure 1. (a) Mean activity (± 1 standard error) of glutathione S-transferase (GST) 1897
(nmol of CDNB-GSH conjugate/min/mg of protein). (b) Mean (± 1 standard error) of 1898
reduced glutathione (GSH) concentration (nmol GSH/mg protein). (c) Mean 1899
concentration (± 1 standard error) of thiobarbituric acid reactive substances (TBARS) 1900

70
(TMP nmol/mg tissue). In all cases, measurements were performed in gills, 1901
hepatopancreas and muscle of L. vannamei reared in saline (SW) or in biofloc system 1902
(BFT) and exposed to empty nanocapsules or nanoencapsulated lipoid acid. Different 1903
letters indicate significant differences (p< 0.05) between the experimental groups. 1904
Similar letters (lowercase in gills; uppercase in hepatopancreas and Roman numerals in 1905
muscle) means absence of statistical differences (p> 0.05). Number of samples analyzed 1906
per treatment: 11 – 15. 1907
1908
1909
Figure 2. Mean (± 1 standard error) of percentage of hyaline and granular hemocytes in 1910
hemolymph from shrimps L. vannamei submitted to different experimental conditions in 1911
the same treatments. Shrimps were reared in saline water (SW) or in bioflocs system 1912
(BFT) and fed with ration supplemented with nanocapsules containing lipoic acid 1913
(NCLA) or empty nanocapsules (NC). Similar letters (lowercase in hyaline hemocytes 1914
and uppercase in granular hemocytes) means absence of statistical differences (p>0.05). 1915
Number of samples analyzed per treatment: 7. 1916

71
CAPÍTULO III 1917
1918
1919
Effects of lipoic acid in the total antioxidant capacity in 1920
biofloc 1921
1922
1923
1924
Átila Clivea da Silva Martins, Wilson Wasielesky Junior, José 1925
Maria Monserrat 1926
1927
1928
1929
Será submetido a revista Aquaculture 1930
1931
1932
1933
1934
1935
1936

72
ABSTRACT 1937
Several studies prove the effectiveness of biofloc in improving growth, survival and 1938
feed conversion to shrimp species Litopenaues vannamei, and the presence of bioactive 1939
compounds that help the immune and antioxidant defense. In this study, was observed 1940
the total antioxidant capacity of biofloc with application of antioxidant lipoic acid in 1941
different concentrations (control, [1,25μM], [2,5μM], [5μM] and [10μM]). Using the 1942
method of total antioxidant capacity against peroxyl radical (ACAP), for which was 1943
found increase in antioxidant capacity with the application of lipoic acid, especially in 1944
the treatment [10μM]. 1945
1946
Keywords: biofloc, bioactive compound, lipoic acid, total antioxidant capacity. 1947
1948
1 INTRODUCTION 1949
The biofloc Technology (BFT) is a production system with minimal water 1950
exchange, which includes the development of microorganisms aggregate belonging to 1951
different taxonomic groups (bacteria, micro-algae, protozoa and other) together with 1952
debris (organic and inorganic particles) that are suspended in the water column with 1953
constant aeration (Avnimelech, 2009; Cardona et al., 2015). The aggregates or biofloc 1954
of this microbial community in this system helps to maintain the water quality by 1955
setting of the recycling of nutrients (nitrogen compounds) and can be used as a source 1956
of proteins and lipids to aquatic organisms including fish and shrimp (Burford et al., 1957
2004; Avnimelech, 2009). 1958
From the ecology point of view, the biofloc can be understood as a 1959
microecosystem constituted by a great diversity of microorganisms (bacteria, 1960
phytoplankton, zooplankton), which develop at the expense of biological processes 1961
occurring in environment of raised. Thus, the nutritional value of biofloc is directly 1962
linked to the composition of the microbial communities and bioactive compounds 1963
present in these organisms. In the studies by Ju et al. (2008), Xu and Pan (2013) and 1964
Cardona et al., (2015), they proposed that animals raised in biofloc system showed an 1965

73
increase in immune response and antioxidant status, probably for the intake of bioactive 1966
compounds present in these organisms. 1967
In an intensive rearing system may increase the action of bioactive compounds 1968
by exogenous addition of other compounds with bioactive properties to assist in the 1969
wellness of animals raised, in this sense lipoic acid (LA) is presented as a multifaceted 1970
bioactive agent. It is known that the physiological level, this compound chelates free 1971
radicals, induces increase in the reduced glutathione concentration and restores the 1972
levels of other antioxidants; in this way also assists in maintaining the intracellular 1973
redox state (Shay et al., 2009). 1974
The quantification of biofloc is monitored through the settleable solids in BTF 1975
system (Samocha et al., 2010; Ray et al., 2010; Schveitzera et al., 2013) and the 1976
presence of these solids over 10–14 mL L-1, may increase consumption of dissolved 1977
oxygen limiting their concentrations and may change other variables such as alkalinity 1978
and pH (Tovar and Erazo, 2009; Samocha et al., 2010; Ray et al., 2010). The 1979
measurement of settleable solids may be a staunchly useful to interpret the interactions 1980
among the AL and biological processes that lead to increased or decreased of the biofloc 1981
quantity. 1982
The application of AL in biofloc can increase the antioxidant capacity of biofloc, 1983
providing a means of reducing the damaging effects of reactive oxygen species (ROS) 1984
(Ghiselli et al., 2000). The ROS such as superoxide anion (O2·-); hydroxyl radical (OH·) 1985
and hydrogen peroxide (H2O2) are produced in cells of the organisms in normal or 1986
pathologic physiological conditions (Wajner et al., 2004), and can also be generated by 1987
cyclically dissolved oxygen and practice use of hydrogen peroxide to establish the 1988
oxygen levels in the water with biofloc by reduction or absence of mechanical aeration 1989
(Cardona et al., 2015). These ROS act on biomolecules that comprise cell membranes, 1990
proteins, DNA and RNA and the negative effects of this process are counteracted or 1991
mitigated by the action of endogenous or exogenous antioxidants (Barreiros et al., 1992
2006). 1993
An antioxidant may be defined as any substance that, when present at low 1994
concentrations compared with an oxidizable substrate, significantly delays or prevents 1995
oxidation of said substrate (Dickinson and Forman, 2002). The sum of the antioxidant 1996

74
activity of an antioxidant compound is known as total antioxidant capacity (Bartosz, 1997
2003). The term antioxidant activity is restricted to characterize the rate constant of a 1998
single antioxidant at a specific oxidant (generally free radical), whereas the antioxidant 1999
capacity should refer to the number of moles of oxidant eliminated by a given sample 2000
(Ghiselli et al., 2000). 2001
Therefore, the objective of this study is to evaluate the effects of lipoic acid in 2002
biofloc over time (8 days), by analyzing the total antioxidant capacity against peroxyl 2003
radicals. 2004
2005
2 MATERIAL AND METHODS 2006
2.1 EXPERIMENTAL DESIGN 2007
It was held at the Green House I (GHI), in Marine Aquaculture Station of the 2008
Federal University of Rio Grande - FURG (Brazil), from 26 October to 2 November of 2009
2014. In 15 tanks plastics (70 L), with a useful volume of 50 L each. Treatments were 2010
set at: Control (0 mg LA L-1 of biofloc), Concentration of 1.25 µM (0.26 mg L-1), 2011
Concentration of 2.5 µM (0.51 mg L-1), Concentration of 5 µM (1.03 mg L-1) and 2012
Concentration of 10 µM (2.06 mg L-1); all in triplicate. The collections of biofloc were 2013
made every 24 hours. After the collection of biofloc, were added the AL concentrations. 2014
2015
2.2 DILUTION LIPOIC ACID IN SODIUM HYDROXIDE 2016
The fixed concentrations of synthetic α-lipoic acid (> 99% purity, Sigma-2017
Aldrich) were dissolved individually for each tank, with sodium hydroxide solution and 2018
distilled water to ensure full dilution of lipoic acid following the methodology Amado et 2019
al. (2011). After dilution, the pH was adjusted to 7.5. The concentrations of LA were 2020
added every 24 hours (Table 1). 2021
2022
2023
2024

75
Table 1: Preparation of lipoic acid solution. NaOH (sodium hydroxide; 2mM), dwH2O 2025
(distilled water), LA (lipoic acid). From the overall calculation of 300 mL of NaOH to 1 2026
g of LA and 1000 mL of dwH2O to 80 g of NaOH. 2027
Treatment NaOH (g) dwH2O (mL) LA (g)
Control
1.25 µM
2.5 µM
5.0 µM
10.0 µM
0
0.62
1.22
2.47
4.94
0
7.8
15.3
30.9
61.8
0
0.26
0.51
1.03
2.06
2028
2.3 WATER QUALITY ANALYSIS 2029
Daily physical and chemical parameters of water were monitored, including 2030
dissolved oxygen (mg L-1) and temperature (°C) with YSI multiparameter, salinity with 2031
optical refractometer (Atago 103, ± 1 ppt), pH with electrode Mettler Toledo FEP20 - 2032
FiveEasy Plus™. Determination of alkalinity (mg CaCO3 L-1) was made by the method 2033
APHA (1985), and calculating the concentration of sodium bicarbonate was performed 2034
according Furtado et al. (2011). The analyzes of total ammonia (NH3 + NH4+ mg L-1) by 2035
method UNESCO (1983) and nitrite (mg L-1) according Benderschneider and Robinson 2036
(1952) (Figure 1). 2037
The settleable solids (organic and inorganic particulate matter) were determined 2038
by sedimentation of solids in Imhoff cones where placed 1 L of water and allowed to 2039
stand for 1 h, time which is made to read the settleable solids mL L-1 (Tovar and Erazo 2040
2009) (Figure 1). 2041
2042
2.4 COLLECTION AND HOMOGENIZATION OF SAMPLES 2043
Samples were collected after reading the Imhoff cone, in which the water was 2044
removed by siphoning and flake placed in falcon (50 mL) and stored on ice, and then 2045
they were transferred to eppendorf (2 mL) and centrifuged at 800 x g being the 4 °C for 2046
10 minutes and stored at -80 °C. For homogenization, the biofloc samples were weighed 2047

76
in eppendorff, added methanol (100%) on relationship 1000 mg 1000 μL-1, 2048
homogenized and stirred for 3 hours, then centrifuged for 10 minutes at 10000 rpm and 2049
4 °C, the supernatant removed and it used to determine the antioxidant capacity against 2050
peroxyl radicals. 2051
2052
2.5 DETERMINATION OF TOTAL ANTIOXIDANT CAPACITY AGAINST 2053
PEROXYL RADICAL (ACAP) 2054
It was conducted in accordance with the protocol Amado et al. (2009), but for 2055
this work was not the fixation of the protein concentration. After white microplate was 2056
added to 127.5 µL of a reaction buffer consisting of 0.3575 g of ethanesulfonic acid, 4.2 2057
hydroxyethyl-piperazine-1 (HEPES), 0.7455 g of potassium chloride (KCl), 0.0102 g 2058
magnesium chloride (MgCl2) dissolved in 50 mL of MilliQ water with pH adjusted to 2059
7.2; 10 µL tissue extract; 7.5 µL of water MilliQ to samples without ABAP (2,2 azobis 2060
2metilpropianoamidina dihidrocloreto) or 7.5 µL ABAP solution for samples with 2061
ABAP (peroxyl radical generator) and 10 µL of H2DCF-DA solution (diacetate '2,7 2062
diclorofluresceín). Reading is performed at time zero and then every 5 minutes to 2063
complete 30 minutes in fluorimeter plates (FilterMax F5, Multi-mode microplate 2064
reader) using a wavelength of 530 nm emission and 485 nm excitation, at 37 °C, the 2065
temperature that favors ABAP thermolysis. This method quantifies the ability of the 2066
tissue to neutralize the ROS generated by the decomposition of ABAP, including 2067
enzymatic antioxidant defenses and/or non-enzymatic, by calculating the relative area 2068
performed according Monserrat et al. (2014). The relative area has an inverse 2069
relationship with the antioxidant capacity, the area where lower values indicate a higher 2070
relative antioxidant capacity and vice-versa. 2071
2072
2.6 STATISTICAL ANALYSIS 2073
Data were expressed in mean ± standard deviation. On analysis by two-factor 2074
ANOVA (different concentrations of lipoic acid and time). Previously, the assumptions 2075
of normality and homogeneity of variance were analyzed. Mean comparisons were 2076
made using the Newman-Keuls test. In all cases, we used a 5% significance level. 2077

77
3 RESULTS 2078
The water quality parameters such as temperature got overall average of 28.48 ± 2079
0.09 °C. The dissolved oxygen was not significantly different (p>0.05) between 2080
treatments or over time, getting between minimum 7.68 ± 0,05 and maximum 7.99 ± 2081
0,02 mg L-1, both in control at time 0 and time 2, respectively. The pH showed changes 2082
in time 4, 5 and 6 in the control treatment, with values of 7.81 ± 0.04, 7.69 ± 0.04 e 7.62 2083
± 0.07 respectively and have minimum of 7.62 and maximum of 8.10 on 3 (time 3) both 2084
in the control treatment. The lowest salinity was measured on time 1 in the treatment 2085
[1.25 µM] with 26.27 ± 2.99 and from time 4 was in 30 for all treatments. For alkalinity, 2086
the minimum value of 126.67 ± 53.64 mg CaCO3 L-1 occurred at time 5 in the treatment 2087
[10 µM], with a maximum of 213.33 ± 38.33 at time 3 for the same treatment. The 2088
ammonia presents change in time 4 in treatment [5 µM], with maximum value of 0.41 ± 2089
0.32 mg L-1 and have minimum of 0.02 ± 0.00 mg L-1 at time 3 in treatment [10 µM]. 2090
The minimum levels of nitrite was among 0.03 ± 0,00 mg L-1 in the control treatment at 2091
times 2, 6 and 7 and maximum levels at time 5, 6 e 7 with value 0.34 ± 0.23, 0.31 ± 0.15 2092
and 0.34 ± 023 mg L-1 respectively in treatment [1.25 µM]. The settleable solids 2093
increased over time and showed significant differences (p>0.05) when compared to 2094
control, with a minimum value of 2.33 ± 0.33 mL L-1 at time 2 in the treatment control 2095
and up to 216.67 ± 16.67 mL L-1 in time 6 in treatment [1.25 µM]. 2096
For total antioxidant capacity, at time 0 was there is significant difference 2097
(p<0.05) between control and of the treatments 1.25 µM (0.26 mg L-1), 2.5 µM (0.51 2098
mg L-1), 5 µM (1.03 mg L-1) and 10 µM (2.06 mg L-1). At time 1, noted increase in total 2099
antioxidant capacity with significant difference (p<0.05) between control and of the 2.5 2100
µM (0.51 mg L-1), 5 µM (1.03 mg L-1) and 10 µM (2.06 mg L-1). At time 2, there was a 2101
significant difference (p<0.05) between control and the other treatments, following this 2102
pattern for the times 3, 4, 5, 6 and 7 (Figure 2). 2103
2104
2105
2106
2107

78
2108
Figure 1 – Mean concentration (±1 standard error) of the physico and chemical 2109
parameters. Number of samples analyzed in each treatment: 3 2110

79
2111
Figure 2 – Mean values (± 1 standard error) of total antioxidant capacity against peroxyl 2112
radicals (expressed in relative area) in biofloc. Different letters indicate significant 2113

80
differences (p<0.05) between experimental groups after performing Newman-Keuls 2114
post hoc test. Number of samples analyzed in each treatment: 9. 2115
2116
4 DISCUSSION 2117
Water quality parameters were monitored to comply with requirements physical 2118
and chemical of the water for species Litopenaues vannamei, following values proposed 2119
by Lin and Chen (2001; 2003), Ray et al. (2010) and Samocha et al. (2010), that 2120
recommended: (i) temperature should be between 28–30 °C, because low temperature 2121
decreases growth and high temperature cause stress; (ii) dissolved oxygen greater than 5 2122
mg L-1, reduces stress on the shrimp; (iii) pH in 7.5, preferably greater than 7; (iv) 2123
salinity greater than 10 with a tolerance of 4–35, high salinity help in buffering capacity 2124
of water; (v) alkalinity greater than 160 mg L L-1, assists in balancing the pH; (vi) 2125
ammonia in 3.35 mg L-1 with salinity of 25 and 3.95 mg L-1 in salinity 35; (vii) nitrite 2126
levels between 5–25 mg L-1, thresholds rise with increased salinity; (viii) settleable 2127
solids 10–14 mg L-1, increase in the floc concentration may result in anaerobic mean. 2128
For this work, on time 1 the salinity had decreased to 26.67 ± 2.99, but is still in the 2129
recommended range for L. vannamei. On time 5, when the alkalinity has reached 2130
minimum 126.67 ± 53.64 mg CaCO3 L-1, was added sodium bicarbonate to increase the 2131
alkalinity levels. On time 4, showed an increase of ammonia remaining with value of 2132
0.41 ± 0.32 mg L-1, below the recommended amount. On time 5, 6 and 7, an increase in 2133
nitrite levels getting the value of 0.34 ± 0.23 mg L-1; 0.31 ± 0.15 mg L-1 and 0.34 ± 0.23 2134
mg L-1, below the recommended values. At the end of the experiment, only settleable 2135
solids were beyond to standard recommended for shrimp L. vannamei, with a significant 2136
increase in treatments with LA, which may suggest that the AL induces increased 2137
amount of floc or the increase was due to the absence of shrimp in the tank, providing 2138
an imbalance between production and consumption floc. 2139
The total antioxidant capacity of biofloc increased since the first day of 2140
application of AL, highlighting to treatments [2.5 µM], [5 µM] and [10 µM]. On the 2141
second day of the experiment, the increase occurred in treatments [1.25 µM] and [10 2142
µM]. On the third day, the increase was observed in treatment [10 µM]. On the fourth 2143
day, increased total antioxidant capacity was observed [10 µM]. On the fifth day, the 2144

81
increase occurred in treatments [1.25 µM] [µM 5] and [10 µM]. Thus, AL induces 2145
increasing the capacity of floc cells to neutralize ROS. On day 6 and 7 was observed 2146
decrease in antioxidant capacity with respect to treatment with AL, suggesting that 2147
under experimental conditions the biofloc has limited capacity to absorb AL or increase 2148
the amount of floc, evidenced by the increase in settleable solids, reduces the 2149
concentration and the LA effects. For the control treatment, no significant change over 2150
time. 2151
2152
5 CONCLUSION 2153
The lipoic acid induces increase in total antioxidant capacity in biofloc, having 2154
the concentration of 10 µM AL per liter of water at most the total antioxidant capacity 2155
over time. 2156
2157
6 FUTURE PERSPECTIVE 2158
Lipoic acid can convey their bioactive properties without causing damage to 2159
biofloc, but these benefits can be apply to the community that makes up the biofloc or 2160
it benefits are community-specific? 2161
Once lipoic acid increases the total antioxidant capacity of the flake, the dose of 2162
10 µM of LA per liter of water can influence the nutritional composition of biofloc? 2163
2164
Acknowledgments. José M. Monserrat, Wilson Wasielesky Junior, are research 2165
fellowships from the Brazilian agency CNPq (process numbers PQ 307880/2013-3, PQ 2166
310993/2013-0, respectively). 2167
2168
2169
2170
2171

82
REFERENCES 2172
Amado, L.L., Garcia, M.L., Ramos, P.B., Freitas, R.F., Zafalon, B., Ferreira J.L.R., 2173
Yunes, J.S., Monserrat, J.M., 2009. A method to measure total antioxidant 2174
capacity against peroxyl radicals in aquatic organisms: Application to evaluate 2175
microcystins toxicity. Science of the Total Environment. 407, 2115–2123. 2176
Amado, L.L., Garcia, M.L., Pereira, T.C.B., Yunes, J.S., Bogo, M.R., Monserrat J.M., 2177
2011. Chemoprotection of lipoic acid against microcystin-induced toxicosis in 2178
common carp (Cyprinus carpio, Cyprinidae). Comparative Biochemistry and 2179
Physiology, Part C. 154, 146–153. 2180
Avnimelech, Y., 2009. Biofloc Technology - A practical guide Book. Baton Rouge, 2181
Louisiana, United States, The World Aquaculture Society. 2182
Barreiros, A.L.B.S., David, J.M., David, J.P., 2006. Estresse oxidativo: relação entre 2183
geração de espécies reativas e defesa do organismo. Química Nova. 29, 113-123. 2184
Bartosz, G., 2003. Total antioxidante capacity. Advances in Clinical Chemistry. 37, 219–2185
292. 2186
Burford, M.A., Thompson, P.J., McIntosh, P.R., Bauman, R.H., Pearson, D.C., 2004. 2187
The contribution of flocculated material to shrimp (Litopenaeus vannamei) 2188
nutrition in a high-intensity, zero exchange system. Aquaculture. 232, 525–537. 2189
Cardona, E., Lorgeoux B., Chim L., Goguenheim J., Le Delliou H., Cahu C., 2015. 2190
Biofloc contribution to antioxidant defence status, lipid nutrition and 2191
reproductive performance of broodstock of the shrimp Litopenaeus stylirostris: 2192
Consequences for the quality of eggs and larvae. Aquaculture. doi: 2193
10.1016/j.aquaculture.2015.08.003. 2194
Dickinson, D.A., Forman, H.J., 2002. Cellular glutathione and cellular metabolism. 2195
Biochemical Pharmacology. 64, 1019–1026. 2196
Ferreira, A.L.A., Matsubara, L.S., 1997. Radicais livres: conceitos, doenças 2197
relacionadas, sistema de defesa e estresse oxidativo. Departamento de Clínica 2198
Médica da Faculdade de Medicina de Botucatu, SP. Revista Associação Médica 2199
Brasileira. 43, 61–68. 2200

83
Furtado, P.S., Poersch, L.H., Wasielesky Jr, W., 2011. Effect of calcium hydroxide, 2201
carbonate and sodium bicarbonate on water quality and zootechnical 2202
performance of shrimp Litopenaeus vannamei reared in bio-flocs technology 2203
(BFT) systems. Aquaculture. 321, 130–135. 2204
Ghiselli, A., Serafini, M., Natella, F., Scaccini, C., 2000. Total antioxidant capacity as a 2205
tool to assess redox status: Critical view and experimental data. Free Radical 2206
Biology & Medicine. 29, 1106–1114. 2207
Ju Z., Forster, I., Conquest, L., Dominy, W., 2008. Enhanced growth effects on shrimp 2208
(Litopenaeus vannamei) from inclusion of whole shrimp floc or floc fractions to 2209
a formulated diet. Aquaculture Nutrition. 14, 533–543. 2210
Lin, Y-C., Chen, J-C., 2001. Acute toxicity of ammonia on Litopenaeus vannamei 2211
Boone juveniles at diferente salinity levels. Journal of Experimental Marine 2212
Biology and Ecology. 259, 109–119. 2213
Lin, Y-C., Chen, J-C., 2003. Acute toxicity of nitrite on Litopenaeus vannamei (Boone) 2214
juveniles at different salinity levels. Aquaculture 224, 193–201. 2215
Monserrat, JM, Garcia, M.L., Ventura-Lima, J., González, M., Ballesteros, M.L., 2216
Miglioranza, K.S., Amé, M.V., Wunderlin, D.A., 2014. Antioxidant, phase II and 2217
III responses induced by lipoic acid in the fish Jenynsia multidentata 2218
(Anablapidae) and its influence on endolsulfan accumulation and toxicity. 2219
Pesticide Biochemistry and Physiology. 108, 8–15. 2220
Packer, L., Wiltf, E.H., Tritschler, H.J., 1995. Alpha-lipoic acid as a biological 2221
antioxidant. Free Radical Biology Medicine. 19, 227–250. 2222
Ray, A.J., Seaborn, G., Leffler, J.W., Wilde, S.B., Lawson, A., Browdy, C.L., 2010. 2223
Characterization of microbial communities in minimal-exchange, intensive 2224
aquaculture systems and the effects of suspended solids management. 2225
Aquaculture. 310, 130–138. 2226
Samocha, T.M., Wilkenfeld, J.S., Morris, T.C., Correia, E.S., Hanson, T., 2010. 2227
Intensive raceways without water exchange analyzed for White shrimp culture. 2228
Global Aquaculture Advocate. 13, 22–24. 2229

84
Schveitzera, R., Arantesa, R., Costódio, P.F.S., Santo, C.M.E., Aranaa, L.V., Seiffert, 2230
W.Q., Andreatta, E.R., 2013. Effect of different biofloc levels on microbial 2231
activity, water quality and performance of Litopenaeus vannamei in a tank 2232
system operated with no water Exchange. Aquacultural Engineering. 56, 59–70. 2233
Shay, K.P., Moreau, R.F., Smith, E.J., Smith, A.R., Hagen, T.M., 2009. Review Alpha-2234
lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic 2235
potential. Biochimica et Biophysica Acta. 1790, 1149–1160. 2236
Tovar, S.A.O., Erazo, E.C., 2009. Analisis de las Caracteristicas Fisico-Quimicas de 2237
Aguas y Suelos de Cultivos Acuicolas Intensivos y Superintensivos. Ceniacua-2238
Colciencias. Primera Edición. Bogotá. 82 p. 2239
Wajner, M., Latini, A., Wyse, A.T.S., Dutra-Filho, C.S., 2004. The role of oxidative 2240
damage in the neuropathology of organics acidurias: insights from animal 2241
studies. Journal of Inherited Metabolic Disease. 27, 427–448. 2242
Xu, W.J., Pan, L.Q., 2013. Enhancement of immune response and antioxidant status of 2243
Litopenaeus vannamei juvenile in biofloc-based culture tanks manipulating high 2244
C/N ratio of feed input. Aquaculture. 412, 117–124. 2245
2246
2247
2248
2249
2250
2251
2252
2253
2254

85
CONCLUSÃO GERAL 2255
Os camarões criados em sistema de biofloco, em 30 dias de exposição, tiverem 2256
aumento na competência antioxidante, sendo a resposta tecido-específica para camarão 2257
branco do pacífico Litopenaus vannemei. Deste modo, algumas comunidades de 2258
microrganismo do sistema BFT influencia no estado redox de tecidos de camarão 2259
(brânquias, hepatopâncreas e músculo), como por exemplo: diminuição da concentração 2260
de glutaiona reduzida em brânquias e hepatopâncreas e níveis de peroxidação lipídica 2261
menores em hepatopâncreas. 2262
Com suplementação com ácido lipóico nanoencapsulado, aumento atividade 2263
gluationa S-transferase em hetaopâncreas, aumento da concentração de glutationa 2264
reduzida em músculo, maior capacidade antioxidante no hepatopâncreas e níveis de 2265
peroxidação lipídica menos em músculo. E aumento dos hemóctios granulares em 2266
camarões exposto ao ácido lipóico. E, portanto, o ácido lipóico foi capaz de aumentar 2267
capacidade antioxidante e diminuir o dano oxidativo em tecido do camarão branco do 2268
pacífico Litopenaus vannemei com ação antioxidante tecido-específica. 2269
O ácido lipóico foi capaz de aumentar a capacidade antioxidante no biofloco em 2270
concentração de [10 µM] (2,06 mg de AL/L de biofloco), com aumento gradual da 2271
capacidade antioxidante durante 5 dias. 2272
2273


















