
UNIVERSIDADE FEDERAL DOS VALES DO JEQUITINHONHA … · universidade federal dos vales do...
27
UNIVERSIDADE FEDERAL DOS VALES DO JEQUITINHONHA E MUCURI FACULDADE DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE DEPARTAMENTO DE EDUCAÇÃO FÍSICA EFEITOS DO TREINAMENTO FÍSICO AERÓBICO SOBRE A FREQUÊNCIA CARDÍACA E PRESSÃO ARTERIAL DE CAMUNDONGOS KNOCKOUT PARA O RECEPTOR MAS Liliane Vanessa Costa Pereira Diamantina 2012/2
-
Upload
trinhtuong -
Category
Documents
-
view
230 -
download
0
Transcript of UNIVERSIDADE FEDERAL DOS VALES DO JEQUITINHONHA … · universidade federal dos vales do...

UNIVERSIDADE FEDERAL DOS VALES DO JEQUITINHONHA E MUCURI
FACULDADE DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE
DEPARTAMENTO DE EDUCAÇÃO FÍSICA
EFEITOS DO TREINAMENTO FÍSICO AERÓBICO SOBRE A FREQUÊNCIA
CARDÍACA E PRESSÃO ARTERIAL DE CAMUNDONGOS KNOCKOUT PARA O
RECEPTOR MAS
Liliane Vanessa Costa Pereira
Diamantina
2012/2

UNIVERSIDADE FEDERAL DOS VALES DO JEQUITINHONHA E MUCURI
FACULDADE DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE
EFEITOS DO TREINAMENTO FÍSICO AERÓBICO SOBRE A FREQUÊNCIA
CARDÍACA E PRESSÃO ARTERIAL DE CAMUNDONGOS KNOCKOUT PARA O
RECEPTOR MAS
Liliane Vanessa Costa Pereira
Orientador:
Prof. Dr. Marco Fabrício Dias Peixoto
Trabalho de Conclusão de Curso apresentado ao
Departamento de Educação Física, como parte
dos requisitos exigidos para a conclusão do curso.
Diamantina
2012/2

EFEITOS DO TREINAMENTO FÍSICO AERÓBICO SOBRE A FREQUÊNCIA
CARDÍACA E PRESSÃO ARTERIAL DE CAMUNDONGOS KNOCKOUT PARA O
RECEPTOR MAS
Liliane Vanessa Costa Pereira
Orientador: Prof. Dr. Marco Fabrício Dias Peixoto
Trabalho de Conclusão de Curso apresentado ao
Departamento de Educação Física, como parte
dos requisitos exigidos para a conclusão do curso.
APROVADO em 26 /03 /2013
_______________________________________
Profº Ms. Fernando Joaquim Gripp Lopes
________________________________________
Profº Dr. Flávio de Castro Magalhães
______________________________________
Profº Dr. Marco Fabrício Dias Peixoto – UFVJM

AGRADECIMENTO
A Deus e a Virgem Maria por me guiarem;
A minha Mãe e meu Pai por serem a base de tudo;
Aos meus irmãos, avós, segunda mãe, padrinho, tios e tias, primos pela companhia, incentivo
e amor;
A Filipe pela compreensão, carinho e amor.
Aos amigos e colegas de sala pelos anos de convivência e confidências;
Ao AMIGO e coorientador, Dirceu, pelos vários momentos de aprendizagem e descontração;
Ao Prof. Marco Fabrício por todos os saberes mediados e, ainda, pela confiança e serenidade;
A FAPEMIG por subsidiar o desenvolvimento do meu projeto de pesquisa;
Muito obrigada!

RESUMO
As doenças cardiovasculares são responsáveis por um número significativo de mortes
constituindo um problema a nível global. O treinamento físico aeróbico (TFA) constitui uma
ferramenta não farmacológica no combate às doenças cardiovasculares, devido às adaptações
promovidas ao sistema cardiovascular, como hipertrofia cardíaca fisiológica, bradicardia e
redução dos níveis pressóricos. O Sistema Renina-Angiotensina (SRA) é considerado um dos
principais sistemas envolvidos na homeostasia cardiovascular tendo como principais
peptídeos ativos a Angiotensina II (Ang II) que, via AT1, é um potente hipertensivo e exerce,
na maioria das vezes, efeitos deletérios ao SCV; e a Angiotensina 1-7 (Ang 1-7), que via Mas,
exerce efeitos cardioprotetores e antagônicos a Ang II. Estudos recentes apontam o eixo Ang-
(1-7)/Mas como mediador nas adaptações cardiovasculares do TFA. O presente estudo
objetivou investigar os efeitos do treinamento físico sobre a frequência cardíaca (FC) e
pressão arterial (PA) de camundongos knockout para o receptor Mas. Foram utilizados 33
camundongos C57/BL6 machos sendo 17 animais selvagens (WT) e 16 animais knockout
para o Mas (KO) distribuídos em quatro grupos experimentais: sedentário selvagem (SWT);
treinado selvagem (TWT); sedentário knockout (SKO) e treinado knockout (TKO). O
protocolo de treinamento em piscina teve a duração de 10 semanas. A PA e FC foram
mensuradas por pletismografia de cauda ao final do período experimental. Após dez semanas
de treinamento físico aeróbico as adaptações cardiovasculares do TFA não foram observadas
nos animais knockout para o receptor Mas. Para os valores de FC (SWT: 622,9 ± 68,29; TWT:
478,0 ± 26,60*; SKO: 541,1 ± 70,94; TKO: 500,9 ± 44,83) e PAS (SWT: 132,0 ± 7,15; TWT:
117,0 ± 12,30*; SKO: 138,7 ± 10,20; TKO: 130,8 ± 7,18) as diferenças foram encontradas
apenas entre os animais selvagens treinados e sedentários. Isoladamente, estes achados
sugerem que, na ausência do eixo Ang(1-7)/Mas, as adaptações cardiovasculares ao
treinamento físico aeróbio não são encontradas.
Palavras chave: treinamento físico, Angiotensina 1-7, receptor Mas

SUMÁRIO
1. INTRODUÇÃO...................................................................................................................06
1.1 Justificativa.....................................................................................................................07
1.2 Objetivo...........................................................................................................................07
2. REVISÃO DE LITERATURA...........................................................................................08
2.1 Adaptações cardiovasculares ao treinamento físico aeróbico..........................................09
2.2 Sistema Renina-Angiotensina e treinamento físico........................................................10
2.3 Animais Mas-/-.................................................................................................................10
3. METODOLOGIA.................................................................................................................11
3.1 Grupos Experimentais.......................................................................................................11
3.2 Protocolo do treinamento em piscina................................................................................11
3.3 Avaliação da frequência cardíaca e pressão arterial.........................................................13
3.4 Análise estatística.............................................................................................................13
4- RESULTADOS.....................................................................................................................14
5 - DISCUSSÃO.........................................................................................................................15
6- CONCLUSÃO.......................................................................................................................18
REFERÊNCIAS........................................................................................................................19

6
1. Introdução
As Doenças cardiovasculares (DCVs) são responsáveis por um número significativo
de mortes nos países desenvolvidos e em desenvolvimento constituindo um problema a nível
global (LAURENTI et al. 2000). Mudanças no estilo de vida a partir da prática regular de
exercícios físicos e adoção de hábitos alimentares saudáveis constituem as principais
ferramentas não-farmacológicas no combate e prevenção das DCVs.
Já é bem documentado na literatura que o treinamento físico aeróbico promove
adaptações benéficas ao sistema cardiovascular, dentre estas adaptações destacam-se a
bradicardia e redução dos níveis pressóricos (RONDON E BRUM, 2003). Em conjunto, estas
alterações promovem uma redução da sobrecarga cardíaca reduzindo os riscos do surgimento
das doenças cardíacas tais como, cardiomiopatias, infarto do miocárdio e insuficiência
cardíaca.
O Sistema Renina-Angiotensina (SRA), tem sido considerado um dos principais
sistemas envolvidos na homeostasia cardiovascular sendo composto por dois principais
peptídeos biologicamente ativos: A Angiotensina II (Ang-II) e a Angiotensina 1-7 (Ang 1-7).
O primeiro exerce efeitos, na maioria das vezes, deletérios ao sistema cardiovascular tais
como a indução de arritmias cardíacas (FERREIRA et al., 2001), fibrose, hipertrofia cardíaca
(SADOSHIMA et al., 1993), além de aumento da pressão arterial. Já o segundo, a Ang-(1-7),
na maioria das vezes, exerce efeitos cardioprotetores e antagônicos à Ang II, tais como,
redução da incidência das arritmias cardíacas, inibição da fibrose, da hipertrofia cardíaca e
redução dos níveis pressóricos.
Apesar do conhecimento dos efeitos cardioprotetores da Ang 1-7, até recentemente
não se conhecia o receptor endógeno pelo qual a Ang-(1-7) exercia seus efeitos. Em 2003,
Santos et al. mostraram que o receptor Mas é o principal receptor mediador das ações da
Ang-(1-7). Nos últimos anos, os efeitos cardioprotetores da Ang-1-7 têm sido confirmados
com a utilização de animais Mas-/- que apresentam diversas disfunções metabólicas e
cardíacas, tais como dislipidemias (SANTOS, 2008) e redução da contratilidade cardíaca
(DIAS-PEIXOTO et al., 2008).
Estudos recentes mostram que o eixo Ang-(1-7)/Mas está envolvido com as
adaptações cardiovasculares ao treinamento. De uma forma geral, os estudos indicam que o

7
treinamento físico é acompanhado por uma redução da atividade ou expressão do eixo
ECA/Ang-(II)/ATI (FERNANDES et al., 2011) e/ou aumento da atividade ou expressão do
eixo ECA2/Ang-(1-7)/Mas (FILHO et al., 2008).
Sabendo-se, portanto que o SRA está envolvido com as adaptações do treinamento e
que animais Mas-/- apresentam disfunção metabólica e cardiovascular, levantamos a hipótese
inicial de que animais Mas-/- não responderiam de forma adequada ao estímulo do
treinamento físico, ou seja, a presença do receptor Mas seria importante para a geração das
adaptações cardiovasculares clássicas do treinamento físico.
1.1 Justificativa
Atualmente, já é bem compreendido que o treinamento físico aeróbico (TFA) promove
adaptações cardiovasculares como a bradicardia de repouso e redução de níveis pressóricos.
Estas adaptações levam a uma redução da sobrecarga cardíaca, que por sua vez, previne o
surgimento das doenças cardiovasculares (DCVs).
Neste contexto, o SRA tem tido destaque dentre os sistemas hormonais envolvidos nas
adaptações cardiovasculares ao TFA, de forma especial para este trabalho, o eixo Ang 1-7 /
receptor Mas.
A relevância deste trabalho se dá pela escassez de estudos na literatura científica que
investigam a participação do eixo Ang-(1-7) / receptor Mas nas adaptações cardiovasculares
ao TFA.
1.2 Objetivo
Avaliar os efeitos do treinamento físico aeróbico sobre a frequência cardíaca e pressão
arterial de camundongos knockout para o receptor Mas.

8
2. Revisão de Literatura
2.1 Adaptações cardiovasculares ao TFA
A prática de atividade física sistematizada é denominada de exercício físico e pode ser
classificados como resistido ou aeróbico. Este último é o de nosso maior interesse e se
caracteriza por atividades de longa duração e de baixa a moderada intensidade, que realizados
de forma crônica promove adaptações ao sistema cardiovascular.
Neste sentido, o treinamento físico aeróbico promove diversas alterações anatômicas e
hemodinâmicas (RONDON E BRUM, 2003) que culminam em adaptações cardiovasculares,
dentre estas as adaptações cardíacas de hipertrofia fisiológica (BLOMQVIST & SALTIN,
1983) e bradicardia (NEGRÃO et al.,1992; GAVA et al., 1995), além das adaptações
vasculares e metabólicas como a redução da pressão arterial de repouso (WHELTON et al.,
2002).
A hipertrofia cardíaca fisiológica ocorre especialmente a uma sobrecarga de volume
que ocorre ao longo do período do TFA gerando um aumento das câmeras ventriculares,
especialmente do ventrículo esquerdo, e espessamento das paredes ventriculares com a
inserção de sarcômeros em série (WAKATSUKI et al., 2004).
A bradicardia de repouso é um marcador de eficiência do TFA e sua ocorrência pode
ser explicada por três mecanismos: 1) aumento do tônus vagal para o coração (MEDEIROS et
al., 2004); 2) diminuição do tônus simpático para coração (GAVA et al., 1995); 3) diminuição
da FC intrínseca (NEGRÃO et al., 1992).
Já os principais mecanismos que explicam a redução da pressão arterial de repouso
após o TFA são: 1)redução do débito cardíaco de repouso (VERÁS-SILVA et al., 1997;
2)redução do tônus simpático (SILVA et al., 1997) e 3) a maior produção de substâncias
vasodilatadoras (CLAUDINO et al., 2004; WOODMAN et al., 2005).
Alterações metabólicas também são observadas com o treinamento físico aeróbico,
como aumento das enzimas oxidativas do ciclo de Krebs e cadeia respiratória (SALTIN E
GOLLNICK, 1983; BOOTH E THOMASON, 1991;). Maughan et al. (2000), corroborando,
observou um aumento no tamanho e número de mitocôndrias e sua capacidade em gerar ATP
pela fosforilação oxidativa. Em conjunto, estas adaptações fazem parte de um mecanismo de

9
adaptações cardiovasculares benéficas na prevenção e combate às doenças cardiovasculares
(DCVs).
2.2 Sistema Renina-Angiotensina e Treinamento Físico
Na visão clássica, o SRA é composto por enzimas e peptídeos que levam a formação
da Angiotensina II (Ang II) e as ações desta se dão por mediação dos receptores AT1 e AT2.
Via receptor AT1, a Ang II exerce efeitos hipertensivos e deletérios no remodelamento
cardíaco, contribuindo no processo de hipertrofia patológica (SENBONMATSU et al., 2003),
arritmias (FERREIRA et al., 2001) e fibrose (SADOSHIMA et al., 1993).
Na visão moderna, o SRA é visto como um complexo sistema hormonal composto por
enzimas e peptídeos vasoativos hipertensivos e anti-hipertensivos que agem de modo
endócrino, parácrino e autócrino (SIMÕES-E-SILVA et al., 1997; SANTOS et al., 2000) e
tem como principal produto final a Ang 1-7. A sua síntese consiste na conversão da Ang I,
pela ECA2, em Angiontensina 1-9, e posteriormente em Ang 1-7, pela ECA (principal via de
inativação da bradicinina) ou outras peptidases (DONOGHUE et al., 2000) ou pela conversão
direta, pela ECA2, de Ang II em Ang 1-7 (VICKERS et al., 2002). Esta tem como principal
ligante endógeno o Mas e exerce, na maioria das vezes, ações cardioprotetoras e antagônicas a
Ang II/ AT1.
Vários estudos elucidam os efeitos da Ang 1-7 via receptor Mas sobre o sistema
cardiovascular, dentre eles a inibição da proliferação de células musculares lisas vasculares
(FREEMAN et al., 1996), inibição do crescimento dos cardiomiócitos (TALLANT et al.,
1999), efeito antitrombótico (KUCHAREWICZ et al., 2000) e antiarrítmico (FERREIRA et
al., 2001), indução de vasodilatação coronariana (BROSNIHAN et al., 1996), capacidade de
potencialização do efeito hipotensor da bradicinina (BK) e sua contribuição para os efeitos
inibidores da ECA (PAULA et al., 2004; FERNANDES et al., 2005);
Em condições de exercício agudo, o papel fundamental do SRA na homeostase, é bem
descrito. Warren et al. (2001) mostraram que o SRA atua no controle da PA e que tanto a
inibição da ECA quanto o bloqueio do AT1 são capazes de reduzir a pressão arterial média
(PAM) durante o exercício.
No entanto, a participação do SRA nas adaptações cardiovasculares crônicas do TFA
bem como a mediação da Ang-1-7 e do receptor Mas ainda não são bem compreendidas. De
forma interessante, nos últimos quatro anos, estudos indicam que a Ang 1-7 e seu receptor

10
Mas, têm papel importante no desenvolvimento das adaptações cardiovasculares decorrentes
do TFA.
Filho et al. (2008) mostraram um aumento na relação Ang 1-7/Ang II e na expressão
do Mas, no ventrículo esquerdo, em ratos espontaneamente hipertensos submetidos ao TFA.
Esta relação indica que, em animais treinados, as ações da Ang 1-7 sobrepõem as ações Ang
II, favorecendo as ações benéficas da Ang-(1-7) sobre o sistema cardiovascular. Corroborando,
em um estudo com ratos obesos submetidos ao TFA, Barreti (2010) observou um aumento da
atividade e concentração da ECA2 no ventrículo esquerdo em ambos os grupos treinados,
independentemente da obesidade, o que sugere que o exercício físico, por si só, foi eficiente
em aumentar a ECA2. De maneira concernente, Fernandes et al. (2011) observou, após o
treinamento físico de natação, uma redução da ECA cardíaca e níveis de Ang II e aumento
nos níveis de Ang 1-7)
2.3 Animais Mas-/-
Para possibilitar a investigação das ações de tal receptor, um modelo experimental,
knockout Mas (Mas -/-) foi desenvolvido, a partir da engenharia genética. A partir da deleção
gênica o animal Mas-/- não expressa a proteína Mas e, portanto a Ang 1-7 não pode exercer
seus efeitos neste organismo. Camundongos Mas -/-, quando comparados a animais selvagens,
apresentam disfunções no sistema cardiovascular como redução na contratilidade cardíaca
(DIAS-PEIXOTO et al., 2008), prejuízo na função cardíaca in vitro, aumento da pressão de
perfusão coronária e diminuição na fração de encurtamento (SANTOS et al., 2011); e também
disfunções no sistema endócrino como hiperglicemia e aumento dos lípedes circulantes
(SANTOS et al., 2008), além de ausência do efeito antidiurético, em animais com sobrecarga
hídrica (SANTOS et al., 2003).

11
3. Metodologia
3.1 Grupos Experimentais
No estudo foram utilizados camundongos machos, C57/BL6 selvagem (Mas+/+) e
knockout para o receptor Mas (Mas-/-), com idade de 12 semanas. Os grupos experimentais
foram divididos em 1) Selvagem sedentário – 9 camundongos Mas+/+ que não realizaram
treinamento físico; 2) Selvagem treinado – 7 camundongos Mas+/+ que realizaram
treinamento físico; 3) Knockout sedentário – 8 camundongos Mas-/- que não realizaram
treinamento físico; 4) Knockout treinado – 8 camundongos Mas-/- que realizaram treinamento
físico.
Os animais foram fornecidos pelo Dr. Robson Augusto dos Santos (Departamento de
Fisiologia e Biofísica, UFMG). Os animais foram mantidos, manipulados e sacrificados
conforme recomendações do “Guia de Manutenção e Uso de Animais de Laboratório” e de
acordo com o Comitê de Ética em Experimentação Animal (CETEA) da UFMG. A presente
proposta foi aprovada pelo referido comitê de ética (protocolo de número 036/10).
3.2 Protocolo do exercício em piscina
Aclimatação ao biotério e ao exercício – assim que os animais chegaram ao laboratório
ocorreu, durante uma semana, a aclimatação às condições do biotério e ao ciclo invertido (12
horas claro-escuro). Neste mesmo período, os animais foram aclimatados ao exercício: os
animais eram mantidos na piscina por 10 minutos diários, sem sobrecarga, até o final da
semana (temperatura da água 31 ± 1º C).
Determinação da intensidade máxima do exercício em piscina – consistiu em um protocolo
de carga progressiva até exaustão, realizado 48 horas após o período de aclimatação. Antes da
realização do teste os animais foram pesados para que, a partir da massa corporal dos mesmos,
fossem confeccionadas sobrecargas correspondentes a 2% da massa corporal. O teste foi
realizado individualmente, na mesma raia, com água na temperatura de 31 ± 1º C. A partir do
momento em que o animal, com uma sobrecarga de 2% afixada na cauda, era colocado na
água, o cronômetro era disparado. O teste foi de caráter contínuo, porque não houve

12
necessidade de retirar o animal da água quando era adicionada a sobrecarga, a cada três
minutos, até a exaustão. O estado de exaustão foi caracterizado pela mudança no padrão do
nado do animal, o qual passava a „cavalgar‟ e, logo era retirado da piscina. A capacidade
aeróbica máxima foi definida como a maior sobrecarga alcançada durante o teste.
Treinamento na piscina - Os camundongos foram submetidos a diferentes tempos e
intensidades de treinamento, durante as dez semanas de experimento conforme protocolo de
treinamento em piscina (Almeida e et al., 2009). Os animais foram pesados no início de cada
semana, às segundas-feiras pela manhã, antes de iniciarem o exercício (pesagem extra de cada
animal foi realizada nos dias dos testes). Os animais treinaram de segunda à sexta-feira no
período da manhã. Objetivando evitar qualquer diferença em relação à natação dos animais na
piscina, as raias foram alternadas a cada dia de treinamento e os testes de exaustão eram
considerados como dia de treinamento. Após a primeira semana (quatro dias de adaptação e
um teste de carga máxima no quinto dia) os animais descansaram 72 horas e, na segunda
semana, foi iniciado o protocolo de treinamento físico propriamente dito. Nessa semana os
animais treinaram numa frequência de 5 vezes, com um volume de 40 minutos diários a uma
intensidade de 50% da carga máxima (CM). Na terceira semana a intensidade foi mantida e
houve um acréscimo de 10 minutos no volume diário de exercício. Na 4ª semana os animais
treinaram 5 vezes, por 40 minutos a 60% da CM. Na 5ª semana o volume se manteve, porém a
uma intensidade de 70% da CM. Na 6ª e 7ª semanas os animais se exercitaram 5vezes, por 40
minutos, a 80% da CM. Na 8ª, 9ª e 10ª semanas os animais se exercitaram 5 vezes, por 60
minutos, a 80% da CM. Após 72 horas do final do treinamento foi realizado um novo teste de
carga máxima.
SEMANA 2 E 3
50% CM 5X
40 -50 min
60% CM 5X
40 min
SEMANA 4 SEMANA 5 SEMANA 6-10
80% CM 5X
40-60 min
70% CM 5X
40 min
SEMANA 1
ADAPTAÇÃO
72h 72h
Figura 1: Esquema ilustrativo do protocolo de treinamento físico em piscina

13
3.3 Avaliação da Frequência Cardíaca e Pressão Arterial
O registro da pressão arterial sistólica (PAS) e frequência cardíaca (FC), foram
realizados após o período de treinamento, antecedendo o sacrifício, pelo método não invasivo
da pletismografia caudal. As medidas foram realizadas utilizando o sistema CODA (Kent
Scientific, CODA - Torrington, CT, EUA). Inicialmente os animais passaram por uma
adaptação no sistema para minimizar o estresse durante a avaliação. Para realização da
medida os camundongos foram acondicionados em um tubo cilíndrico de acrílico, no qual
foram aquecidos e ventilados de maneira adequada para a realização das medidas. Para tal
procedimento, inseriu-se na cauda dos animais em um manguito de borracha adaptado à
região proximal da cauda e conectado ao esfignomanômetro para insuflar e desinsuflar em
intervalos fixos de aproximadamente 50 segundos. Próximo ao manguito foi acoplado um
transdutor de pulso (sensor) que captava os sinais, que eram enviados e registrados em
computador. Através dos pulsos de pressão registrados foram determinadas PAS e PAD e,
calculada a frequência cardíaca.
3.4 Análise estatística
Os dados são apresentados como a média ± desvio padrão. Para comparações de duas
variáveis foi utilizado o teste t student e, para comparação de mais de duas variáveis foi
utilizado o teste one way ANOVA. O nível de significância foi estabelecido para um *p< 0,05.
14
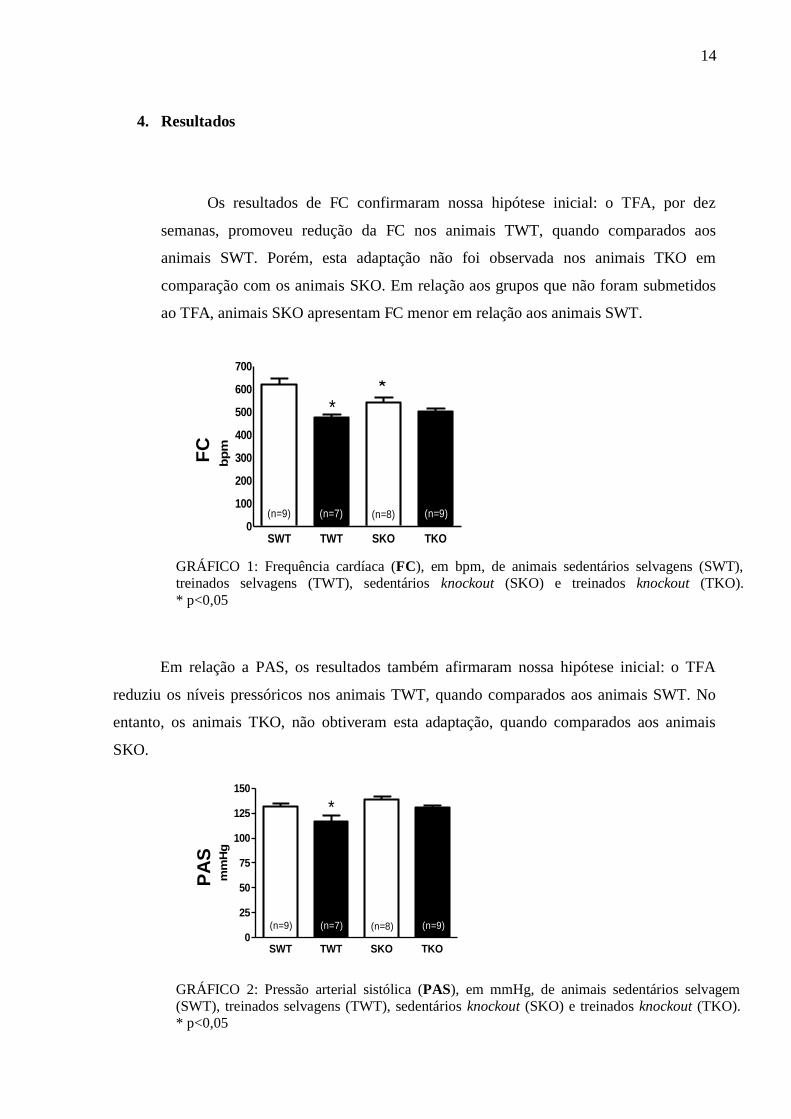
4. Resultados
Os resultados de FC confirmaram nossa hipótese inicial: o TFA, por dez
semanas, promoveu redução da FC nos animais TWT, quando comparados aos
animais SWT. Porém, esta adaptação não foi observada nos animais TKO em
comparação com os animais SKO. Em relação aos grupos que não foram submetidos
ao TFA, animais SKO apresentam FC menor em relação aos animais SWT.
Em relação a PAS, os resultados também afirmaram nossa hipótese inicial: o TFA
reduziu os níveis pressóricos nos animais TWT, quando comparados aos animais SWT. No
entanto, os animais TKO, não obtiveram esta adaptação, quando comparados aos animais
SKO.
SWT TWT SKO TKO0
100
200
300
400
500
600
700
(n=9) (n=8)(n=7) (n=9)
*
bp
m
SWT TWT SKO TKO0
25
50
75
100
125
150
(n=9) (n=8)(n=7) (n=9)
*
mm
Hg
GRÁFICO 2: Pressão arterial sistólica (PAS), em mmHg, de animais sedentários selvagem
(SWT), treinados selvagens (TWT), sedentários knockout (SKO) e treinados knockout (TKO).
* p<0,05
GRÁFICO 1: Frequência cardíaca (FC), em bpm, de animais sedentários selvagens (SWT),
treinados selvagens (TWT), sedentários knockout (SKO) e treinados knockout (TKO).
* p<0,05
FC
P
AS
*

15
5. Discussão
O objetivo deste estudo foi investigar os efeitos do treinamento físico aeróbico (TFA)
sobre a frequência cardíaca (FC) e pressão arterial (PA) de camundongos knockout para o
receptor Mas (Mas -/-). Como esperado, o TFA promoveu adaptações nos animais treinados
selvagens (TWT) que apresentaram redução da frequência cardíaca e da pressão arterial
sistólica. No entanto, o TFA não promoveu essas adaptações cardiovasculares nos animais
treinados Mas -/- (TKO) sugerindo que o receptor Mas está envolvido nas adaptações
cardiovasculares ao TFA.
Nos últimos cinco anos, alguns estudos sugerem que o SRA, especialmente o eixo
Ang 1-7/Mas, participa das adaptações cardiovasculares ao TFA tanto em animais sadios
como acometidos por patologias. Em um estudo com ratos obesos e não obesos submetidos ao
TFA, Barreti et al. (2010) observaram um aumento da atividade e concentração da ECA2
(principal enzima envolvida na formação da Ang-(1-7)) no ventrículo esquerdo em ambos os
grupos treinados. Filho et al. (2008) realizaram um protocolo de TFA em piscina com ratos
Wistar hipertensos (SHR) e observaram que estes animais desenvolveram hipertrofia cardíaca
fisiológica associada com aumento na concentração de Ang-(1-7) e na expressão do Mas no
ventrículo esquerdo (VE). Corroborando com estes resultados, Fernandes et al. (2011)
também associaram o desenvolvimento de hipertrofia cardíaca fisiológica em ratos Wistar
normotensos com a redução na atividade da ECA e aumento da atividade da ECA2 no
ventrículo esquerdo destes animais. Além disso, a redução na atividade da ECA foi
acompanhada pela redução nos níveis de Ang II e o aumento na atividade da ECA2 foi
seguido por um aumento na produção de Ang-(1-7) sugerindo que o TF aumenta a produção
de Ang 1-7, a partir da Ang II.
Dos estudos citados, dois observaram o desenvolvimento de hipertrofia cardíaca
seguida de aumento da expressão de Ang 1-7 e de Mas no VE (FILHO et al., 2008) ou
redução na atividade da ECA e Ang II e aumento na atividade da ECA 2 (FERNANDES et
al., 2011). Apesar destes estudos não terem avaliado os efeitos do TFA sobre a FC, a
hipertrofia cardíaca fisiológica é uma adaptação cardiovascular que pode estar envolvida com
a bradicardia, pois o aumento da cavidade ventricular esquerda pelo TFA aumenta o volume
de ejeção cardíaco permitindo que o coração possa bater em um ritmo mais lento para manter
o débito cardíaco (EKBLOM & HERMANSEN, 1968).

16
Apesar desta possibilidade, ainda não é bem compreendido a participação do eixo
Ang-(1-7) nas adaptações cardiovasculares ao treinamento. Neste contexto, a constatação de
que animais Mas-/- treinados não apresentam bradicardia e redução da pressão arterial
sistólica reforça a participação do receptor Mas nas adaptações ao TFA e traz mais
especulações a respeito dos possíveis mecanismos envolvidos nestas respostas.
As possíveis explicações para a constatação de que animais Mas-/- apresentam falha
no desenvolvimento da bradicardia e redução da pressão arterial sistólica em resposta ao TFA
se baseiam nas ações periféricas e centrais da Ang-(1-7). Já é bem documentado que a Ang-
(1-7) é um potente vasodilatador central e periférico. A Ang 1-7 é capaz de aumentar a
sensibilidade do barorreceptor (CAMPAGNOLE-SANTOS et al., 1992) e potencializar o
efeito vasodilatador da bradicinina (PAULA et al., 1995) e do óxido nítrico (NO)
(BROSNIHAN et al., 1996). No que diz respeito a bradicinina, a sua interação com a Ang 1-
7, foi mostrada in vivo (PAULA et al., 1995; LIMA et al., 1997), em cultura de células
(DEDDISH et al., 1998) e em vasos isolados (LI et al., 1997). Portanto, acreditamos que o
TFA promove aumento das concentrações teciduais e plasmáticas da Ang-(1-7) em animais
Mas-/-, porém o efeito vasodilatador da Ang-(1-7) está abolido nestes animais.
Já em relação aos efeitos da Ang-(1-7)/Mas sobre a FC, os resultados ainda são
controversos. Santos et al. (2004) mostraram que o aumento dos níveis plasmáticos de Ang-
(1-7) observado em ratos que superexpressam Ang-(1-7), produziu um aumento da FC basal
em condições in vivo e in vitro. Além disso, Santos et al. (2006) observaram que corações
isolados de camundongos Mas-/- apresentam redução na FC comparados a camundongos
selvagem (Mas+/+). Por outro lado, Braga et al. (2002) observaram que a infusão de Ang 1-7,
por sete dias, promoveu uma discreta bradicardia em ratos Wistar e Campagnole-Santos et al.
(1992) notaram uma melhora no controle reflexo da FC, em SHR, após a infusão, central ou
periférica, de Ang 1-7. Benter et al. (1995) mostrou que infusões intravenosas de Ang 1-7
inverteram os efeitos inibitórios da Ang II no controle reflexo da FC em ratos SHR Wistar
Kyoto. Em concordância, um estudo de Cangussu (2008), mostrou que a Ang-(1-7) pode
ativar centralmente os mecanismos de bradicardia e, estes efeitos, são mediados pelo receptor
Mas. Nos dois últimos estudos, microinjeções na medula ventrolateral caudal do bulbo
restauraram a bradicardia barorreflexa em ratos com hipertensão renovascular e o uso do
antagonista do receptor Mas, o A779, aboliu estas respostas.
Apesar destas discordâncias, a maioria dos estudos indicam que a Ang-(1-7) estimula
a bradicardia e os estudos que indicam efeitos de aumento da FC devem ser interpretados com

17
cautela. Os estudos de Santos et al (2004), por exemplo, observaram efeito taquicárdico da
Ang-(1-7), porém utilizaram animais transgênicos que tem níveis de Ang-(1-7) plasmáticos
em concentrações muito superiores aos níveis fisiológicos. Estudos destes mesmos autores,
em 2006, mostraram que animais Mas-/- apresentaram redução da FC comparados animais
Mas+/+. De Mello (2004) mostrou que a Ang 1-7 ativa a bomba de sódio, hiperpolarizando a
célula cardíaca reestabelecendo a condução do impulso durante a isquemia e reperfusão o que
indica que o efeito antiarritmogênico da Ang 1-7 (FERREIRA et al., 2001) poderia ser devido
a mudanças nas correntes iônicas. Estas observações sugerem que a Ang 1-7 poderia
realmente modular alguns componentes de correntes iônicas de marcapasso através da
estimulação do Mas efeitos estes abolidos nos animais Mas-/-.
Neste sentido, uma possível explicação pelo não desenvolvimento da bradicardia em
camundongos Mas-/- é que estes, naturalmente, já possuem uma FC menor. Entretanto, os
mecanismos envolvidos nas alterações de FC em camundongos Mas-/-ainda precisam ser
estabelecidos. Neste sentido acreditamos que em condições fisiológicas a Ang-(1-7) promove
redução da FC e, estes efeitos, são abolidos em animais Mas-/-.
Para confirmar a participação do eixo Ang-(1-7) /Mas nas adaptações cardiovasculares
ao TFA sugerimos que novos experimentos sejam realizados com animais que
superexpressam Ang-(1-7) para avaliar, por exemplo, se de forma oposta aos animais Mas-/-,
animais que superexpressam Ang-(1-7) apresentam uma resposta exacerbada das adaptações
cardiovasculares ao TFA, mais especificamente uma maior resposta bradicárdica e de redução
da pressão arterial.

18
6. Conclusão
Em conclusão, este estudo mostrou que, de forma oposta aos camundongos selvagem
(Mas+/+), camundongos knockout para o receptor Mas (Mas-/-) não apresentaram bradicardia
de repouso e redução de níveis pressóricos após o treinamento físico aeróbico, ou seja, o
treinamento físico aeróbico não foi eficiente em promover adaptações cardiovasculares nos
animais Mas -/-. Portanto, o receptor Mas é essencial para que ocorram adaptações
cardiovasculares ao treinamento físico aeróbico.

19
Referências Bibliográficas
ALMEIDA, PWM; GOMES-FILHO, A; FERREIRA, AJ; RODRIGUES, CEM; DIAS-
PEIXOTO, MF; RUSSO, RC; TEIXEIRA, MM; CASSALI, GD; FERREIRA, E; SANTOS,
IC; GARCIA, AMC; SILAMI-GARCIA, E; WISLOFF, U; PUSSIELD, GA. Swin training
suppresses tumor growth in mice. Journal of Applied Physiology; v. 107, p. 261-265, 2009.
BENTER IF, DIZ DI, FERRARIO CM. Pressor and reflex sensitivity is altered in
spontaneously hypertensive rats treated with angiotensin-(1-7).Hypertension; 26:1138-1144.
BRAGA AN, DA SILVA LEMOS M, DA SILVA JR, FONTES WR, DOS SANTOS RA.
Effects of angiotensins on day-night fluctuations and stress-induced changes in blood
pressure. Am J Physiol Regul Integr Comp Physiol 282: R1663–R1671, 2002.
BROSNIHAN, KB, LI, P, FERRARIO, CM. Angiotensin-(1-7) dilates canine coronary
arteries through kinins and nitric oxide. Hypertension 27, 523-528,1996.
BRUM PC, FORJAZ CLM, TINUCCI T, NEGRÃO CE. Adaptações agudas e crônicas do
exercício físico no sistema cardiovascular. Rev pau. Educ. Fís., São Paulo, v. 18, p.21-31,
ago 2004.
CAMPAGNOLE-SANTOS MJ, HERINGER SB, BATISTA EN, KHOSLA MC, SANTOS
RAS. Differential baroreceptor reflex modulation by centrally infused angiotensin
peptides. Am J Physiol. 1992;263:R89-R94.
CANGUSSU LM. Efeitos Cardiovasculares Induzidos por Antagonistas Angiotensinérgicos
no Bulbo Ventrolateral de Ratos com Hipertensão Renovascular. Dissertação de Mestrado
apresentada ao Instituto de Ciências Biológicas da Universidade Federal de Ouro Preto, Ouro
Preto, 2008.
CHOR D, FONSECA MJM, ANDRADE CR, WAISMANN W, LOTUFO PA. Doenças
cardiovasculares: panorama da mortalidade no Brasil. In: Minayo MC, editor. Os muitos
Brasis. Ed. Rio de Janeiro: Hucitec-Abrasco, 1999; 57-86.

20
CLAUDINO MA, PRIVIERO FB, TEIXEIRA CE, DE NUCCI G, ANTUNES E, ZANESCO
A. Improvement in relaxation response in corpus cavernosum from trained rats. Urology.
2004;63(5):1004-8.
DE MELLO WC. Angiotensin (1-7) re-establishes impulse conduction in cardiac muscle
during ischaemia-reperfusion. The role of the sodium pump. J Renin Angiotensin
Aldosterone Syst. 2004;5:203–208.
DEDDISH PA, MARCIC B, JACKMAN HL, WANG HZ, SKIDGEL AR, ERDOS EG
(1998). NDomain-specific substrate and c-domain inhibition of angiotensin-converting
enzyme: angiotensin-(1-7) and keto-ACE. Hypertension, 31: 912-917.
DIAS-PEIXOTO MF, SANTOS RA, GOMES ER, ALVES MN, ALMEIDA PW, GRECO L,
ROSA M, FAULER B, BADER M, ALENINA N, GUATIMOSIM S. Molecular mechanisms
involved in the angiotensin-(1-7)/Mas signaling pathway in cardiomyocytes. Hypertension.
52(3):542-8, 2008.
DONOGHUE M, HSIEH F, BARONAS E, GODBOUT K, GOSSELIN M, STAGLIANO N,
DONOVAN M, WOOLF B, ROBISON K, JEYASSELAN R, BREITBART RE, ACTON S.
A novel angiotensin converting enzyme related carboxy peptidase (ACE 2) converts
angiotensin I to angiotensin 1-9. Cir Res, v.87, p.E1-E2, 2000.
EKBLOM B, ASTRAND PO, SALTIN B, STENBERG J, WALLSTROM B. Effect of
training on circulatory response to exercise. J Appl Physiol 1968;24: 518-28.
FERNANDES L, FORTES ZB, CASARINI DE, NIGRO D, TOSTES RC, SANTOS RA, DE
CARVALHO MH. Role of PGI2 and effects of ACE inhibition on the 74 bradykinin
potentiation by angiotensin-(1-7) in resistance vessels of SHR. Regul Pept. 2005 Apr
15;127(1-3):183-9.
FERNANDES, T; HASHIMOTO, NY; MAGALHÃES, FC; FERNANDES, DEC;
CARMONA, AK; KRIEGER, JE; PHILLIPS, I; OLIVEIRA, E. Aerobic Exercise Training–
Induced Left Ventricular Hypertrophy Involves Regulatory MicroRNAs, Decreased
Angiotensin-Converting Enzyme-Angiotensin II, and Synergistic Regulation of Angiotensin-

21
Converting Enzyme 2-Angiotensin (1-7). Hypertension. 2011;58:182-189; originally
published online June 27, 2011.
FERRARIO CM, BARNES KL, BLOCK CH, BROSNIHAN KB, DIZ DI, KHOSLA MC,
SANTOS RAS. Pathways of angiotensin formation and function in the brain. Hypertension
15: I13-I19, 1990. 2.
FERREIRA AJ, SANTOS RA, ALMEIDA AP. Angiotensin-(1-7): cardioprotective effect in
myocardial ischemia/reperfusion. Hypertension 38:665-8, 2001.
FILHO AG, FERREIRA AJ, SANTOS SH, NEVES SR, SILVA CAMARGOS ER, BECKER
LK, BELCHIOR HA, DIAS-PEIXOTO MF, PINHEIRO SV, SANTOS RA. Selective
increase of Angiotensin-(1-7) and its receptor in spontaneously hypertensive rat hearts
subjected to physical training. Exp Physiol. 93(5):589-98, 2008.
FREEMAN EJ, CHISOLM GM, FERRARIO CM, TALLANT EA. Angiotensin-(1-7)
inhibits vascular smooth muscle cell growth. Hypertension 28: 104-8, 1996.
GAVA, NS; VÉRAS-SILVA, AS; NEGRÃO, CE; KRIEGER, EM. Low-intensity exercise
training attenuates cardiac beta-adrenergic tone during exercise in spontaneously hypertensive
rats. Hypertension, Dallas, v.26, p.1129-33, 1995.
GRASSI G, SERAVALLE G, CALHOUN DA, BOLLA GB, GIANNATTASIO C,
MARABINI M. Mechanisms responsible for sympathetic activation by cigarette smoking in
humans. Circulation 1994;90(1):248-53.
KEMI OJ, LOENNECHEN JP, WISLØFF U. Intensity-controlled treadmill running in mice:
cardiac and skeletal muscle hypertrophy. J Appl Physiol. 2002;93:1301–1309.
KUCHAREWICZ, I; CHABIELSKA, E; PAWLAK, R; MATYS, T; ROLKOWSKI, R;
BUCZKO, W. The antithrombotic effect of angiotensin-(1-7) closely resembles that of
losartan. J Renin Angiotensin Aldosterone Syst.1 (3):268-272, 2000.

22
LAURENTI R, BUCHALLA CM, CARATIN CVS. Doença isquêmica do coração.
Internações, tempo de permanência e gastos. Brasil, 1993 a 1997. Arq Bras Cardiol
2000;74:6:483-7.
LI P, CHAPPELL MC, FERRARIO CM, BROSNIHAN KB. Angiotensin-(1-7) augments
bradykinin-induced vasodilation by competing with ACE and releasing nitric oxide.
Hypertension, 29 (Part 2): 394-400, 1997.
LIMA CV, PAULA RD, RESENDE FL, KHOSLA MC, SANTOS RAS. Potentiation of the
hypotensive effect of bradykinin by short-term infusion of angiotensin-(1-7) in normotensive
and hypertensive rats. Hypertension, 30 (Part 2): 542-548, 1997.
MEDEIROS A, OLIVEIRA EM, GIANOLLA R, CASARINI DE, NEGRÃO CE, BRUM
PC. Swimming training increases cardiac vagal effect and induces cardiac hypertrophy in rats.
Brazilian Journal of Medical and Biological Research, Ribeirão Preto, 2004.
NEGRÃO CE, MOREIRA ED, BRUM PC, DENADAI MLDR, KRIEGER EM. Vagal and
sympathetic controls of the heart rate during exercise in sedentary and trained rats. Braz J
Med Biol Res 1992;25:1045-52.
NEGRÃO CE, MOREIRA ED, SANTOS MC, FARAH VM & KRIEGER EM. Vagal
function impairment after exercise training. J Appl Physiol. 72, 1759-1753, 1992.
NEGRÃO, CE.; MOREIRA, ED.; SANTOS, MC.; FARAH, VM.; KRIEGER, EM. Vagal
function impairment after exercise training. Journal of Applied Physiology, Bethesda, v.72,
n.5, p.1749-53, 1992b.
NEGRÃO CE, RONDON MUPB. Exercício físico, hipertensão e controle barorreflexo da
pressão arterial. Rev Bras Hipertens 2001;8:89-95.
PAULA RD. Estudo dos mecanismos envolvidos na potenciação do efeito hipotensor da
bradicinina pela angiotensina-(1-7). 2004. Dissertação de mestrado em Fisiologia e
Farmacologia. Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais.

23
PAULA RD, LIMA CV, KHOSLA MC, SANTOS RAS. Angiotensin-(1-7) potentiates the
hypotensive effect of bradykinin in conscious rats. Hypertension, 26 (Part 6): 1154-1159,
1995.
RONDON MUPB, BRUM PC. Exercício físico como tratamento não farmacológico da
hipertensão arterial. Rev Bras Hipertens vol 10(2): abril/junho de 2003.
SADOSHIMA J, IZUMO S. Molecular characterisation of angiotensin II induced hypertrophy
of cardiac myocytes and hyperplasia of cardiac fibroblasts. Circ Res. 73:413–423, 1993.
SANTOS RAS, CAMPAGNOLE-SANTOS MJ, ANDRADE SP. Angiotensin-(1-7): an
update. Regul Pept, v.91, p.45-62. 2000.
SANTOS RAS, CASTRO CH, GAVA E, PINHEIRO SV, ALMEIDA AP, PAULA RD,
CRUZ JS, RAMOS AS, ROSA KT, IRIGOYEN MC, BADER M, ALENINA N, KITTEN
GT, FERREIRA AJ. Impairment of in vitro and in vivo heart function in angiotensin-(1–7)
receptor MAS knockout mice. Hypertension 47: 996–1002, 2006.
SANTOS RAS, FERREIRA AJ, NADU AP, BRAGA AN, ALMEIDA AP,
CAMPAGNOLE-SANTOS MJ, BALTATU O, ILIESCU R, REUDELHUBER TL, BADER
M. Expression of an angiotensin-(1–7)-producing fusion protein produces cardioprotective
effects in rats. Physiol Genomics 17: 292–299, 2004.
SANTOS RAS, SIMÕES E SILVA AC, MAGALDI AJ. Evidence for a physiological role of
Angiotensin-(1-7) in the control of hydroelectrolyte balance. Hypertension 27: 875-84,
1996.
SANTOS RAS, SIMOES E SILVA AC, MARIC C, SILVA DM, MACHADO RP, DE B, I,
HERINGER-WALTHER S, PINHEIRO SV, LOPES MT, BADER M, MENDES EP,
LEMOS VS, CAMPAGNOLE-SANTOS MJ, SCHULTHEISS HP, SPETH R, WALTHER T.
Angiotensin-(1-7) is an endogenous ligand for the G protein-coupled receptor Mas. Proc Natl
Acad Sci 100:8258- 63, 2003.

24
SANTOS RD, MARANHÃO RC, LUZ PL, LIMA JC, FILHO WS, AVEZUM A, et al. III
Diretrizes brasileiras sobre dislipidemias e diretrizes de prevenção da aterosclerose do
Departamento de Aterosclerose da Sociedade Brasileira de Cardiologia. Arq Bras Cardiol
2001;77(Suppl 3):1-191.
SANTOS SH, FERNANDES LR, MARIO EG, FERREIRA AV, POˆ RTO LC, ALVAREZ-
LEITE JI, BOTION LM, BADER M, ALENINA N, SANTOS RA. Mas deficiency in FVB/N
mice produces marked changes in lipid and glycemic metabolism. Diabetes. 2008;57:340 –
347.
SENBONMATSU T, ICHIHARA S, PRICE E, GAFFNEY FA, INAGAMI T. Evidence for
angiotensin II type 2 receptor-mediated cardiac myocyte enlargement during in vivo pressure
overload. J Clin Invest. 106: R25–R29, 2000.
SILVA GJJ, BRUM PC, NEGRÃO CE, KRIEGER EM. Acute and chronic effects of exercise
on baroreflexes in spontaneously hypertensive rats. Hypertension 1997; 30:714-9.
SIMÕES E SILVA AC, BARACHO NCV, PASSAGLIO KT, SANTOS RAS. Renal actions
of angiotensin-(1-7). Braz J Med Biol Res 30 (4): 503-13, 1997.
TALLANT EA, FERRARIO CM, GALLAGHER PE. Angiotensin-(1-7) inhibits growth of
cardiac myocytes through activation of the mas receptor. Am J Physiol Heart Circ Physiol
289:H1560-H1566, 2005.
VÉRAS-SILVA AS, MATTOS KC, GAVA NS, PC, NEGRÃO CE, KRIEGER EM. Low
intensity exercise training decreases cardiac output and hypertension inspontaneously
hypertensive rats. Am J Physiol: Heart Circ Physiol 1997;273: H2627-31.
VICKERS, C; HALES, P; KAUSHIK, V; DICK, L; GAVIN, J; TANG, J; GODBOUT, K;
PARSONS, T; BARONAS, E; HSIEH, F; ACTON, S; PATANE, M; NICHOLS, A;
TUMMINO, PJ. Hydrolisis of biological peptides by human angiotensin-converting enzyme-
relates carboxypeptidase (ACE2). JBC Papers, v. 277, n. 17, p. 14838–14843, 2002.
XU P, COSTA-GONCALVES AC, TODIRAS M, RABELO LA, SAMPAIO WO, MOURA
MM, SANTOS SS, LUFT FC, BADER M, GROSS V, ALENINA N, SANTOS RA.

25
Endothelial dysfunction and elevated blood pressure in MAS gene deleted mice.
Hypertension. 2008;51:574 –580.
WAKATSUKI T, SCHLESSINGER J, ELSON EL. The biochemical response of the heart to
hypertension and exercise. Trends Biochem Sci. 2004; 29 (11): 609-17.
WARREN, JH.; LEWIS, W.; WRAA, C.E.; STEBBINS, CL. Central and peripheral effects
of angiotensin II on the cardiovascular response to exercise. J. Cardiovasc. Pharmacol., v.
38, p. 693–705, 2001.
WHELTON SP, CHIN A, XIN X, HE J. Effect of aerobic exercise on blood pressure: A
meta-analysis of randomized, controlled trials. Ann Intern Med 2002;136:493-503.
WOODMAN CR, THOMPSON MA, TURK JR, LAUGHLIN MH. Endurance exercise
training improves endothelium-dependent relaxation in brachial arteries from
hypercholesterolemic male pigs. J Appl Physiol. 2005;99(4):1412-21.
.

26
AUTORIZAÇÃO
Eu autorizo a reprodução e/ou divulgação total ou parcial do presente trabalho, por qualquer
meio convencional ou eletrônico, desde que citada a fonte.
______________________________________________
Liliane Vanessa Costa Pereira
UNIVERSIDADE FEDERAL DOS VALES DO JEQUITINHONHA E MUCURI
Campus JK. Rodovia MG 367 – km 583, nº 5000 – Alto da Jacuba
Diamantina - MG


















