
USO DE HABITAT, ORÇAMENTO TEMPORAL E INFLUÊNCIA DA … · 2017-10-19 · caracterizar o uso do...
90
1 KARLLA DE MORAES SOUZA LIMA COLLAÇO USO DE HABITAT, ORÇAMENTO TEMPORAL E INFLUÊNCIA DA MARÉ E DA GEOMORFOLOGIA DO FUNDO SOBRE O COMPORTAMENTO DO BOTO Sotalia guianensis van BÉNÉDEN, 1864 (CETACEA: DELPHINIDAE) NO PORTO DE MACEIÓ – ALAGOAS, BRASIL. Natal / RN Dezembro – 2008
Transcript of USO DE HABITAT, ORÇAMENTO TEMPORAL E INFLUÊNCIA DA … · 2017-10-19 · caracterizar o uso do...

1
KARLLA DE MORAES SOUZA LIMA COLLAÇO
USO DE HABITAT, ORÇAMENTO TEMPORAL E INFLUÊNCIA
DA MARÉ E DA GEOMORFOLOGIA DO FUNDO SOBRE O
COMPORTAMENTO DO BOTO Sotalia guianensis van BÉNÉDEN,
1864 (CETACEA: DELPHINIDAE) NO PORTO DE MACEIÓ –
ALAGOAS, BRASIL.
Natal / RN
Dezembro – 2008

2
KARLLA DE MORAES SOUZA LIMA COLLAÇO
USO DE HABITAT, ORÇAMENTO TEMPORAL E INFLUÊNCIA DA
MARÉ E DA GEOMORFOLOGIA DO FUNDO, SOBRE O
COMPORTAMENTO DO BOTO Sotalia guianensis van BÉNÉDEN,
1864 (CETACEA: DELPHINIDAE) NO PORTO DE MACEIÓ –
ALAGOAS, BRASIL.
Dissertação apresentada ao Programa de Pós-Graduação em Psicobiologia (Mestrado) do Departamento de Fisiologia da Universidade Federal do Rio Grande do Norte como parte dos requisitos para a obtenção do título de Mestre.
Orientador: Prof. Dr. Flavio José de Lima Silva
Natal / RN
Dezembro - 2008

3
Divisão de Serviços Técnicos
Catalogação da Publicação na Fonte. UFRN / Biblioteca Central Zila Mamede
Collaço, Karlla de Moraes Souza Lima. Uso de habitat, orçamento temporal e influência da maré da geomorfologia do fundo sobre o comportamento do boto Sotalia guianensis Van Bénéden, 1864 (Cetacea: Delphinidae) no Porto de Maceió - Alagoas, Brasil / Karlla de Moraes Souza Lima Collaço. – Natal, RN, 2008. 90 f.
Orientador: Flávio José de Lima Silva.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Psicobiologia.
1. Sotalia guianensis – Dissertação. 2. Orçamento temporal – Dissertação. 3. Uso de habitat – Dissertação. I. Silva, Flávio José de Lima. II. Universidade Federal do Rio Grande do Norte. III. Título.
RN/UF/BCZM CDU 599.53(043.3)

4

5
“A VIDA É BREVE,
A CIÊNCIA É DURADOURA,
A OPORTUNIDADE É ARDILOSA,
A EXPERIMENTAÇÃO É PERIGOSA,
O JULGAMENTO É DIFÍCIL.”
Hipócrates

6
AGRADECIMENTOS
Sempre é muito difícil ao final de mais uma etapa da vida agradecer a todos os que
contribuíram para o nosso crescimento. Afinal em dois anos e meio quantas pessoas
passam por nossa vida... No meu caso são bem mais que dois anos e meio. Quando saí de
Maceió para ter uma conversa com o meu na época “possível” orientador em Natal, não
imaginei que minha vida pudesse mudar tanto! Desde então (maio de 2005) tenho muito o
que agradecer.
Em primeiríssimo lugar meu orientador, Dr. Flávio Lima, que apostou em mim e
acreditou que eu poderia desenvolver uma pesquisa no meu Estado, tão carente de ciência.
Obrigada pela confiança, compreensão, pelos conselhos e por me lembrar que “Pós-
graduação é antes de tudo sacrifício”!
Ao senhor Berenaldo Lessa e ao Porto de Maceió por todo o apoio ao longo do
projeto, antes mesmo da minha pesquisa. Obrigada pela disponibilidade, confiança e boa
vontade de tornar a pesquisa com esses animais possível.
A MAMMALIA – Conservação & Pesquisa, pela bicicleta doada e que foi
fundamental para a coleta de dados.
A Liisão que é uma irmã! Quanta coisa agüentamos! Que calor!!! E como foi bom
ter alguém para estar ao meu lado, para estudar e dividir expectativas! Obrigada fia!
As meninas de Noronha, pela acolhida! Paty, Lica e Aninha. A Rose Emília (PPC)
não esqueci que você foi a primeira a me receber e sentar comigo para me mostrar às
atividades do grupo. Obrigada pela força e pela bibliografia.
A “Zé”, que foi o primeiro e grande incentivador do meu “largar tudo” para
realizar um sonho! Obrigada por acreditar em mim desde sempre! Certamente sem teu
incentivo eu não teria ido.
A família Santos Carvalho! D. Marisa, Ismael, Belinha, Juca, Neide, Branca e
Gabriel. Vocês me ofereceram sua casa, sua amizade e carinho sem pedir nada em troca, e
quanto tive que ir embora vocês simplesmente me apoiaram. Obrigada por me mostrar
um pedacinho do paraíso! Parte desta conquista devo a vocês.
A todo o pessoal da UFRN! Aos professores: Arrilton, Fátima, Fívia, Emília,
Daniel, Teresa, e todos os outros sempre disponíveis. A turma de mestrado de 2006 e todos
das turmas anteriores que de alguma forma estiveram presentes.
A todos os membros do PPC, Carol, Luísa, Priscila, Kelly, Sandra, Mariana, Lídio,
Rosana, Fernando, Fagner, pelas experiências, ensinamentos, pelas festas! Aprendi muito
com vocês, Obrigada!
Ao amigo LINDO! Manu!! Que conquista foi tua amizade pra mim! Valeu!!

7
Ao meu compadre Bira, obrigada por dividir a vida comigo por um ano, pela
amizade eterna. Não poderia ter tido melhor companheiro de república.
A minha família, fonte de tudo! Por vocês sou capaz de mover o mundo!
Tios: Márcia, Cláudio, Marli, pela ajuda inesperada! Valeu muito! Marisa pelo
exemplo.
Vó linda!!!!! Minha fonte de sabedoria e inspiração!
Mãe... O que dizer de você? Seu apoio foi a base da minha coragem. Você é a
minha GAIA!
Meus irmãos! Apesar dos pesares... A vocês TUDO o que fui, sou e serei!
Lourdes Maria, amor incondicional, te amo!!! JB, a felicidade que caiu do telhado!
Aline, obrigada pela força e por me chamar de volta à terra de vez em quando!
Você sem querer mudou nossas vidas.
Meu Pai...Um dia quem sabe você acredite. És meu Desafio! Do seu jeito você está
aí. Obrigada.
Meu sogro Armando Collaço e minha sogra Palmira Collaço ou devo dizer Pai e
Mãe, obrigada pela acolhida, pela força, pelo cuidado comigo. Nunca vou esquecer o que
vocês fizeram e fazem até hoje.
Meu esposo Bruno Collaço. Que história, não é?! O que é o destino? Ou Deus? Ele
nos deu várias chances e nós fingimos que não vimos, então ele resolveu nos obrigar a
viver na mesma casa. De amigos, confidentes e conselheiros a marido e mulher. Obrigada
por ser quem você é! Por acreditar, por sonhar, pela generosidade, pelo amor, pela
dedicação, pela batalha juntos e pelo JB. Por cuidar de mim quando o porto me deixava
doente. Você foi a melhor aquisição desse mestrado!
Aos amigos: Rogério, Danda, Aninha e Júlia: Incentivadores! Jú, Dada, Lígia.
Vocês foram essenciais! Chinha!! O que seria de mim sem você e o bebê “mala”. Vocês
tornaram tranqüila a parte mais difícil!
Aos professores e amigos: Gabriel Le Campion, obrigada pelo mar! Iracilda Lima,
obrigada pela ciência!
Aos personagens principais dessa jornada de vida, Os Golfinhos! Eles são a
materialização do sentido de ser Bióloga. Se não fosse a fascinação por essas criaturas
mágicas e maravilhosas nada disso teria acontecido.
E por fim e mais importante a Deus! Essa força universal que nos move! A energia
da vida que liga tudo que existe! OBRIGADA!

8
SUMÁRIO
RESUMO x
ABSTRACT xi
INTRODUÇÃO GERAL 12
ARTIGO 1 – Uso do Habitat: Influência da Geomorfologia do Fundo, Tráfego de embarcações e da
Maré sobre o Comportamento do Boto (Sotalia guianensis Van BÉNÉDEN, 1864) no Porto de Maceió
– Alagoas. 16
RESUMO 17
INTRODUÇÃO 18
MATERIAL E MÉTODOS 20
Área de Estudo 20
Coleta dos Dados 20
Categorias Comportamentais e encontros com embarcações 22
Análise dos Dados 24
RESULTADOS 25
DISCUSSÃO 28
AGRADECIMENTOS 36
LITERATURA CITADA 37
LISTA DE FIGURAS 46
FIGURAS 47
ANEXO- Normas para Publicação 49
ARTIGO 2 – Orçamento temporal do boto (Sotalia guianensis) no Porto de Maceió – Alagoas, Brasil. 54
RESUMO 55
INTRODUÇÃO 56
MATERIAL E MÉTODOS 58
Área de Estudo 58

9
Coleta dos Dados 58
Categorias Comportamentais e Interação com embarcações 61
Análise dos Dados 62
RESULTADOS 62
DISCUSSÃO 68
AGRADECIMENTOS 73
REFERÊNCIAS 74
LEGENDAS DAS FIGURAS 78
ANEXO- Normas para Publicação 79
DISCUSSÃO GERAL 82
CONSIDERAÇÕES FINAIS 85
LITERATURA CITADA (INTRODUÇÃO E DISCUSSÃO GERAL) 87

10
RESUMO
Os fatores físicos e ambientais apresentados por cada hábitat e a ritimicidade dos
padrões comportamentais influenciam fortemente a ecologia e o comportamento dos
seres vivos em geral. Ao mesmo tempo tais fatores podem fornecer pistas sobre a
estrutura de uma população e seu equilíbrio ecológico. A estrutura organizacional,
ecológica e comportamental de uma espécie avaliada em regiões sabidamente em
equilíbrio quando comparada à mesma avaliação feita em áreas degradadas pode
fornecer um panorama claro de como as influências antrópicas agiram sobre estas
espécies e que medidas podem ser tomadas no intuito de mitigar os efeitos nocivos e
conservar a população. A região onde foi realizado esse estudo está, assim como a
maioria das áreas de porto, sujeita à intensa degradação física e ambiental. Com o
interesse emergente de mudança na qualidade desses ambientes por parte também das
próprias empresas que utilizam os serviços portuários, o estudo proposto visou
caracterizar o uso do habitat, a distribuição das atividades comportamentais realizadas
ao longo do dia e a influência da geomorfologia do fundo, profundidade e das variações
da maré sobre a expressão dos comportamentos de Sotalia guianensis na região do
Porto de Maceió – Alagoas. A partir dessas informações será possível estabelecer
parâmetros para comparação com outras populações de S. guianensis e delinear medidas
de conservação para a população ocorrente no Porto de Maceió – AL, servindo ainda
como base para ações de conservação futuramente executadas em outras regiões
portuárias.
Palavras-chave: Sotalia guianensis, orçamento temporal, orçamento de atividades.
x

11
ABSTRACT
The physical and environmental factors presented by each habitat and the rhythm of
behavior patterns strongly influence the ecology and behavior of the all living beings.
At same time this factors may provide clues about the structure of a population and its
ecological balance. The organizational structure, ecology and behavior of a species
appraised in a region if we know be in balance when compared to the same type of
appraisal made in a degraded area can provide a clear view of how the anthropogenic
influences acted on these species and what steps can be taken in order to mitigate the
effects and keep the population. The region where this study was conducted is, like
most areas of port, subject to intense physical and environmental degradation. With the
emerging interest of change in the quality of these environments also by the companies
themselves that use the port services, the proposed study aimed to characterize the use
of habitat, the distribution of behavioral activities carried out throughout the day and
influence of geomorphology of the bed, depth and variation of tide on the expression
of the behavior of Sotalia guianensis in the port of Maceio - Alagoas. From this
information will be possible establish parameters for comparison with other populations
of S. guianensis and establish conservation measures for the population occurring in the
port of Maceio - AL, serving also as a basis for conservation actions future performed
in other port regions.
Palavras-chave: Sotalia guianensis, time budget, activity budget.
xi

12
INTRODUÇÃO GERAL
Muitos estudos têm mostrado que a distribuição de uma população está
intimamente ligada ao que o habitat pode fornecer (Ballance, 1992; Hastie et al. 2004;
Rossi-Santos, 2006; Azevedo et al. 2007; Araújo et al. 2007). No entanto, apenas a
informação sobre a distribuição não revela a função ecológica da área utilizada. Essa
função somente é determinada quando os fatores comportamento e tipo de habitat são
analisados em conjunto (Grigg e Markowitz, 1997; Hastie et al. 2004).
Segundo Ballance (1992), cada habitat é composto por um mosaico de partes
que diferem umas das outras física e biologicamente. Alguns locais oferecem uma
maior proteção a alguns elementos como predação, outros fornecem uma grande
concentração de alimento. Em função dessa heterogeneidade encontramos padrões
específicos na maneira como os animais se distribuem e no modo como utilizam os
nichos que ocupam em cada habitat (Hanson e Defran, 1993; Daura-Jorge et al. 2005;
Daura-Jorge et al. 2007)
Delphinideos apresentam um complexo padrão de uso do hábitat, em que fatores
físicos e biológicos afetam aspectos ecológicos, incluindo o comportamento e a
distribuição espacial (Azevedo et al. 2007). A escolha da área, o tipo de atividade
executada em cada local e a estrutura da comunidade são influenciados por fatores
ambientais (fotoperíodo, profundidade, temperatura da superfície, abundância e
distribuição de presas, maré e indiretamente o vento) (Simão e Poletto, 2002; Daura-
Jorge et al. 2005; Rossi-Santos, 2006; Araújo et al. 2007) e físicos (geomorfologia do
fundo) (Hui, 1979; Harzen, 1998; Bearzi et al. 1999; Domit, 2006).
Além do uso do hábitat, o ambiente influencia no ritmo dos padrões
comportamentais apresentados pelos golfinhos. Esse ritmo se dá como uma adaptação

13
às variações diurnas e sazonais dos fatores ambientais e são o resultado de um complexo
ajuste entre o forrageio ótimo e o tempo de forrageio, atividades sociais e as restrições
ambientais (Karczmarski e Cockcroft, 1999; Silva, 2001). A ocorrência de ritmos
diurnos, tanto em animais cativos quanto em animais livres na maioria dos animais
terrestres e aquáticos, sugere que as variáveis fotoperíodicas exercem uma influência
significativa sobre o comportamento (Saayman, 1972; Silva, 2001).
Eberhardt propôs em 1977 a utilização do orçamento de atividades para o acesso
ao status de populações de mamíferos marinhos, baseando-se em algumas linhas de
pensamento utilizadas até hoje. A primeira é que o alimento é um fator limitante
importante para praticamente todas as espécies e a segunda é que o percentual de tempo
de forrageio incluindo procura e perseguição aumenta quando a abundância e a
qualidade das presas diminuem (Gelatt, 2002). A importância da disponibilidade de
alimento assume então uma relevada importância, fazendo com que muitos animais
tolerem distúrbios ambientais e lugares extremamente impactados pelo homem em
função da facilidade na obtenção de alimento (Maze e Würsig, 1999; Bearzi et al. 1999;
Azevedo et al. 2007).
Bearzi et al. (1999) observaram em uma população de golfinhos Turciops sp.
que a grande flexibilidade comportamental desses animais contribui para sua
sobrevivência em locais com elevadas variações ambientais. Isso faz com que o
percentual de tempo gasto pelos animais em forrageio forneça um panorama da
disponibilidade de presas e indiretamente uma visão geral do status ecológico da área.
O boto Sotalia guianensis (Van Bénéden, 1864) é uma espécie que apresenta
áreas preferenciais para cada tipo de atividade (Simão e Poletto, 2002; Cremer et al.
2004; Daura-Jorge et al. 2005). Estes animais têm preferência por águas costeiras,
abrigadas e sua distribuição vai do sul do Brasil até Honduras (Borobia et al. 1991; Carr

14
e Bonde, 2000; Valle, 2006; Azevedo et al. 2007). São generalistas, alimentando-se de
peixes, moluscos e crustáceos, utilizando inclusive recifes submersos e áreas de
estuários para forragear (Borobia e Barros, 1989; Valle, 2006). Possuem pequena área
de uso, inferior a 40 km² na Baía Norte em Santa Catarina (Flores e Bazzalo, 2004), 79
km² para a Baía de Babitonga também em Santa Catarina (Cremer, 2007) e 136,9 km²
para baía de Guanabara no Rio de Janeiro (Azevedo et al. 2007). Em geral não
costumam passar muito tempo em mar aberto (Flores e Bazzalo, 2004). São cetáceos
pequenos com comprimento máximo do corpo de 1,98 m para os machos (Rosas, 2000)
e 2,20 m para fêmeas, sendo considerado robusto e com nadadeiras proporcionalmente
grandes (Hetzel e Lodi, 1993; Da Silva e Best, 1996).
Apesar da flexibilidade comportamental e aparente adaptação a ambientes
poluídos esses animais se encontram sobre forte pressão em função da degradação do
ambiente em que vivem, capturas acidentais e intencionais. Tal pressão se reflete com
maior intensidade nas espécies que freqüentam as regiões próximas da costa (Hetzel e
Lodi, 1993; Bearzi et al. 1999; Maze e Würsig, 1999; Pereira et al. 2007). Essas
regiões, em geral baías, enseadas, portos e estuários, estão sujeitas à atracação de
navios, embarcações de pesca e atividades recreativas, que geram intensa poluição
sonora e trânsito marítimo. Além disso, em geral recebem grande aporte de esgotos e
dejetos industriais.
A ocorrência do boto S. guianensis em Alagoas e no Porto de Maceió já foi
relata em estudos anteriores (Barros e Teixeira, 1994; Campos et al. 2007), porém
foram relativos apenas a registros de ocorrência ou acerca de comportamentos
específicos. A região é vulnerável no que diz respeito a impactos antropogênicos:
intensa urbanização, tráfego de embarcações e grande aporte de dejetos orgânicos e
industriais.

15
Com isso, partindo do princípio de que entender as bases do comportamento de
uma espécie é fundamental para o conhecimento da ecologia e para o estabelecimento
de medidas que possam auxiliar na conservação desta espécie (Daura-Jorge et al. 2005)
e que o uso de indicadores para o aceso ao status populacional de predadores de topo de
cadeia é bastante relevante para o manejo e para a conservação da vida selvagem como
um todo (Gelatt, 2002), o estudo proposto visou caracterizar o uso do habitat, a
distribuição das atividades comportamentais realizadas ao longo do dia e a influência do
tipo de substrato do fundo, profundidade, das variações da maré e do tráfego de
embarcações sobre a expressão dos comportamentos de S. guianensis na região do Porto
de Maceió – Alagoas.
Os resultados obtidos podem ampliar o conhecimento sobre a biologia da
espécie, assim como fornecer bases de comparação com outras populações e subsidiar a
adoção de medidas de conservação e manejo de cetáceos, em especial para populações
que ocupam regiões impactadas por ações humanas como regiões de portos.

16
ARTIGO 1 – Uso do Habitat e Influência da Geomorfologia do Fundo e da Maré
no Comportamento do Boto (Sotalia guianensis Van BÉNÉDEN, 1864) no Porto de
Maceió – Alagoas – Brasil.
Karlla de Moraes Souza Lima Collaço 1 e Flávio José de Lima Silva1,2
1 Programa de Pós-Graduação em Psicobiologia. Universidade Federal do Rio Grande
do Norte (UFRN). Campus UFRN. Caixa Postal 1511 - CEP 59078-970. Natal, RN.
Brazil.
e-mail: [email protected]
2 Universidade do Estado do Rio Grande do Norte (UERN). Laboratório de Sistemática
e Ecologia Animal. Dept. Ciências Biológicas – Fac. Ciências Naturais. Av. Prof.
Antônio de Campos, s/n . Campus Central UERN. CEP 59600-970. Mossoró, RN.
Brazil.
e-mail: [email protected]
A ser traduzido para inglês e submetido ao periódico: Marine Mammal Science

17
RESUMO
Sotalia guianensis é um pequeno boto que habita regiões costeiras e que ocorre desde
Honduras até o Sul do Brasil. Apesar da flexibilidade e adaptabilidade da espécie, ela é
bastante vulnerável em função de seu hábito costeiro, estando constantemente em
contato com impactos causados pelo homem. Sabendo que a escolha da área, o tipo de
atividade executada em cada local e a estrutura da comunidade são influenciadas por
fatores ambientais e que a distribuição de uma população está ligada ao que o habitat
escolhido pode fornecer buscou-se, associando os fatores comportamento e tipo de
habitat, caracterizar o uso do habitat e a influência da geomorfologia do fundo,
profundidade e das variações da maré sobre a expressão dos comportamentos do boto S.
guianensis na região do Porto de Maceió – Alagoas. Os animais apresentaram maior
freqüência de atividades no setor com nove metros de profundidade, pouca declividade
e fundo arenoso. O uso da área parece estar ligado principalmente à disponibilidade de
presas e segurança dos animais. Sendo a região é bastante impactada este trabalho
poderá auxiliar no estabelecimento de medidas mitigadoras e de conservação para a
população que ocorre não só na área do Porto de Maceió, mas para outras áreas
semelhantes.
Palavras-chave: Sotalia, Uso do habitat, Comportamento.

18
INTRODUÇÃO
Delphinideos apresentam um complexo padrão de uso do hábitat, em que fatores
físicos e biológicos afetam aspectos ecológicos, incluindo o comportamento e a
distribuição espacial (Azevedo et al. 2007). A escolha da área, o tipo de atividade
executada em cada local e a estrutura da comunidade são influenciadas por fatores
ambientais (fotoperíodo, profundidade, temperatura da superfície, abundância e
distribuição de presas, maré e indiretamente o vento) (Simão and Poletto 2002, Daura-
Jorge et al. 2005, Rossi-Santos 2006, Araújo et al. 2007) e físicos (geomorfologia do
fundo) (Hui 1979, Harzen 1998, Bearzi et al. 1999, Domit, 2006).
A distribuição de uma população está ligada ao que o habitat escolhido pode
fornecer (Ballance 1992, Hastie et al. 2004, Rossi-Santos 2006, Azevedo et al. 2007,
Araújo et al. 2007). No entanto, apenas a informação sobre a distribuição não revela a
função ecológica da área utilizada. Essa função somente é determinada quando os
fatores comportamento e tipo de habitat são analisados em conjunto (Grigg and
Markowitz 1997, Hastie et al. 2004).
Para isso entender as bases do comportamento torna-se essencial, não só para o
conhecimento dos ritmos, mas da ecologia de uma população e sua conservação (Daura-
Jorge et al. 2005).
O boto Sotalia guianensis (Van Bénéden, 1864) é uma espécie que sabidamente
escolhe áreas preferenciais para cada tipo de atividade (Cremer et al. 2004, Daura-Jorge
et al. 2005). Estando presentes em geral em zonas costeiras abrigadas, do sul do Brasil
até Honduras (Borobia et al. 1991, Carr and Bonde 2000, Valle 2006, Azevedo et al.
2007). São generalistas, alimentando-se de peixes, moluscos e crustáceos, utilizando

19
recifes submersos e estuários para forragear (Borobia and Barros 1989, Valle 2006). A
espécie possui pequena área de uso com um máximo de 136,9 km² ocorrendo na baía de
Guanabara no Rio de Janeiro (Azevedo et al. 2007). Segundo Flores and Bazzalo (2004)
não costumam passar muito tempo em mar aberto. São considerados animais pequenos
e robustos com comprimento máximo do corpo de 2,06 m (Da Silva and Best 1996).
Apesar da flexibilidade comportamental e aparente adaptação a ambientes
poluídos esses animais se encontram sobre forte pressão em função da degradação do
ambiente em que vivem, capturas acidentais, intencionais e transito de navios,
embarcações de pesca e atividades recreativas, que geram intensa poluição sonora e
trânsito marítimo. (Hetzel and Lodi 1993, Bearzi et al. 1999, Maze and Würsig 1999,
Pereira et al. 2007).
Este estudo visou caracterizar a influência da profundidade, do substrato do fundo,
das variações da maré e do tráfego de embarcações sobre a expressão dos
comportamentos do boto S. guianensis na região do Porto de Maceió – Alagoas. A
partir dos resultados obtidos pode-se não só ampliar o conhecimento sobre a biologia e
ecologia da espécie, mas fornecer bases para adoção de medidas de conservação e
manejo de cetáceos, especialmente para populações que ocupam regiões urbanas
impactadas e de portos.

20
MATERIAIS E MÉTODOS
Área de Estudo
O porto de Maceió (09º 40' 2” S e 35º 42' 54” W) encontra-se no centro do
litoral do Estado de Alagoas entre duas praias com intensa urbanização: Avenida e
Pajuçara. Região impactada por atividades antrópicas é cercada por estruturas que
merecem ser descritas para um melhor entendimento das condições da região. A três
quilômetros a noroeste está o emissário submarino, estrutura construída para lançar
parte dos dejetos da cidade, em princípio tratados, no mar. Um quilômetro também a
noroeste do local das observações está o “Riacho Salgadinho”, córrego que recebe
esgoto clandestino, carreando lixo para o cais do Porto durante a maré cheia.
A região possui ainda uma colônia de pescadores, a menos de um quilômetro a
norte do ponto de observação, que produz um intenso tráfego de embarcações, além
disso, a maior marina e as piscinas naturais mais freqüentadas do Estado estão a menos
de dois quilômetros a norte e a leste respectivamente do local de observação,
intensificando ainda mais a circulação de embarcações.
O cais do Porto possui três berços, um píer de granéis líquidos, um cais do
terminal açucareiro e um cais comercial que recebe também navios de turismo.
Em função da configuração geográfica da costa, prevalece a corrente de maré,
cuja direção é sul na enchente e norte na vazante, com amplitude máxima 2,60 metros
(DHN 2008, INPH 1993).
Coleta dos dados
Uma plataforma fixa com cerca de 3 metros de altura, localizada na base do
Farol do porto, que possui cerca de 4 metros em relação ao nível do mar, totalizando 7

21
metros de altura, foi utilizada como ponto de observação. Como a plataforma está
fixada na entrada do cais comercial é possível visualizar a maior parte da área do cais e
toda a área referente a mar aberto num ângulo de 240°. A área de estudo foi dividida em
quatro setores para um melhor aproveitamento dos dados de acordo com a profundidade
e as características físicas de cada subárea (Figura 1). A divisão foi feita utilizando
pontos de referência no Porto ou em terra. Estando, a Nordeste o Setor 1, composto
parte pelo canal de entrada e saída dos navios do porto e parte pela rota de saída dos
barcos da colônia de pesca e da marina com profundidade que varia de 6,0m a 8,0m
metros, intenso tráfego de embarcações e fundo arenoso/orgânico. A Sul, mar aberto, se
localiza o Setor 2 com profundidade média de 9,0m, fundo arenoso e uma pequena
porção de recife submerso. Parte deste setor está no canal de entrada e saída dos grandes
navios. A leste está o cais comercial, onde está localizado o Setor 3 com profundidade
média de 10,5 metros. Este setor sofreu uma dragagem em 1999 e possui fundo de
argila siltosa/orgânico (INPH 1993). O Setor 4 a sudeste tem profundidade média de 2m
e fundo com formação de recife (Porto de Maceió 2002).
Os dados analisados neste trabalho foram coletados com auxílio de binóculos 12
X 50 mm, em dias aleatórios de janeiro a abril de 2007 e de janeiro a julho de 2008. As
observações foram alternadas sendo uma no período da manhã de 07:00hs as 12:00hs e
outra no período da tarde de 12:00hs as 17:00 hs de forma que se obtivesse o mesmo
número de dias e horas de observação para os dois períodos.
Para coleta dos dados comportamentais utilizou-se como método amostral a
amostragem comportamental com registro de todas as ocorrências (Altmann 1974,
Martin and Bateson 1993). Esse método foi possível pelo pequeno número de animais,
tamanho da área e pelo tipo de visão proporcionado pela localização da plataforma, que

22
permite observar todos os indivíduos que estiverem dentro da área ao mesmo tempo.
Esse formato de observação foi baseado no que Baird and Hanson (2004) utilizaram
para orcas. Eles afirmam para uma área semelhante à desse trabalho que o foco estando
voltado para o comportamento e não para o grupo reduz a probabilidade de extrapolar
um comportamento para todo o grupo, evitando a subestimação de comportamentos
menos freqüentes como o descanso. A metodologia relativa a coleta dos dados
referentes a encontros com embarcações foi baseada em trabalhos semelhantes
executados por Constantine (2004), Pereira et al. (2007) e Araújo et al. (2008), onde os
encontros eram contabilizados e categorizados para posterior análise. Para esse trabalho
as embarcações foram divididas em: Navios e embarcações portuárias, pesca e turismo.
Consideramos apenas duas categorias etárias: adultos, e infantes para animais com
comprimento visualmente estimado inferior a 2/3 do tamanho do adulto, regularmente
acompanhado de animal adulto (Maze and Würsig 1999, Azevedo et al. 2005). Grupo
foi definido com todos os animais que estavam ao mesmo tempo dentro da área de
observação.
Dados referentes à profundidade e composição geomorfológica do fundo foram
extraídos a partir das Cartas Náuticas 901 e 920, e relatórios de pesquisas anteriores
cedidos pela administração do Porto de Maceió (INPH 1993, Oliveira and Campion
1999). As profundidades estabelecidas para os setores são referentes à baixa mar de
sizígia. A tábua de maré utilizada foi calculada pela Marinha do Brasil e para as análises
dos dados os ciclos da maré foram divididos em enchente/cheia e vazante/vazia.
Categorias comportamentais e encontros com embarcações
Quatro categorias comportamentais e três tipos de reações após encontros ou
interações com embarcações foram considerados e analisados neste trabalho, definidos
com base em trabalhos anteriores e descritos a seguir:

23
Forrageio: Todo comportamento individual ou em grupo ligado à alimentação direta ou
indiretamente, culminando ou não na captura e consumação da presa. Sendo
consideradas as atividades de forrageio descritas para a espécie anteriormente (Geise
1991, Nascimento 2002, Monteiro et al. 2006, Souto et al. 2006);
Deslocamento: Deslocamento de um ou mais indivíduos com trajetória definida
(direção constante), velocidade variada, sem evidencia de outra atividade e com os
animais claramente em transito de uma área a outra (Harzen 1998, Edwards and Schnell
2001). A categoria deslocamento não foi contabilizada quando inserida no contexto de
outra categoria analisada neste trabalho. Por exemplo: O deslocamento executado
durante a perseguição a uma presa não foi contabilizado na categoria deslocamento e
sim na categoria forrageio, por entender que tal deslocamento está inserido nesse
contexto;
Descanso: Os animais se movimentam lentamente, rente à superfície, sem função
aparente, em geral realizado em grupo e em sincronia, onde se observa apenas o dorso
dos animais quando emergem para respirar. Esses animais mantêm ainda uma
orientação regular podendo, no entanto mudar de direção (Wursig and Wursig 1980,
Karczmarski and Cockcroft 1999);
Socialização: Interações entre os indivíduos usualmente com contato corporal e fora do
contexto de outro tipo de atividade. Incluindo atividades aéreas, interação com objetos,
interações agressivas entre indivíduos e comportamentos agonísticos na presença de
embarcações (Harzen 1998, Edwards and Schnell 2001, Daura-Jorge et al. 2005);
Reações a encontros com embarcações
Positiva: Deslocamento dos animais em direção a embarcação ou natação nas ondas
formadas pelo deslocamento da embarcação (Pereira et al. 2007, Araújo et al. 2008);

24
Neutra: Os animais não alteram o comportamento quando a embarcação se aproxima,
ou simplesmente distanciam-se da embarcação e continuam executando a mesma
atividade anterior ao encontro (Pereira et al. 2007, Araújo et al. 2008);
Negativa: Expressão de atividade à superfície ou comportamento agonístico (batida de
cabeça, batida de cauda, saltos), alteração ou modificação da atividade realizada
anteriormente, saída da área (Pereira et al. 2007, Araújo et al. 2008).
Análise dos dados
Por não apresentarem distribuição normal os dados foram comparados a partir
de testes não-paramétricos com o auxílio do programa estatístico Graphpad Instat 3.0.
Para verificar a diferença entre dois grupos independentes de amostras (diferenças entre
dois setores ou duas atividades) foi utilizado o Teste de Man-Whitney (U-Test). Para
analisar a diferença entre mais de dois grupos independentes de amostras (diferenças
entre setores e atividades, atividades e marés) foi utilizado o teste de Kruskal-Wallis. A
correlação de Spearman foi utilizada para verificar a existência de relação entre
profundidade e os comportamentos. Em todos os testes foi adotado um nível de
significância de 5%. Para a análise da influência da interação com embarcações não foi
feita distinção quanto ao tipo de embarcação presente na área.

25
RESULTADOS
Para efeito de análise de dados neste trabalho foram realizados 38 dias de
observação, 5 horas por dia, totalizando 190 horas de esforço amostral e 87 horas de
esforço efetivo. Em 31(81,59%) dos dias de observação os animais estavam presentes e
em apenas 7 dias (18,42%) não houve animais na área. Houve presença de infantes em
30 (96,77%) dos 31 dias com animais na área. Dez animais foi o maior número
observado na área ao mesmo tempo e o menor foi 1 animal, sendo a média de 4,15
animais. A maior freqüência foi de 3 animais (n=18) e a menor 8 e 10 animais (n=1).
Dos indivíduos observados 66,08% eram adultos, 27,65% infantes e 6,27% não foi
possível identificar a faixa etária. Houve maior concentração do número de infantes em
relação ao número de adultos no período da manhã (62,67% adultos/28,12% infantes)
do que no período da tarde (68,55% adultos/27,31% infantes), diferença considerada
não significativa (p>0,05).
O comportamento mais freqüente foi o forrageio representando 34,25% das
atividades, seguido pelo deslocamento com 32,98%. A categoria socialização
representou 27,59% e o descanso 5,18%, do total das atividades. Os testes estatísticos
não demonstraram diferença significativa quando analisados os totais das atividades
(p<0, 2301). Quando comparadas as categorias uma a uma verificou-se diferença
significativa entre forrageio e descanso (p<0,01), socialização e descanso (p<0,05) e
entre deslocamento e descanso (p<0, 001). Entre os eventos registrados na categoria
socialização foi possível observar seis episódios de brincadeira com objeto, sendo cinco
eventos referentes a infantes com algas ou sacos plásticos e um de um adulto com um
pedaço de madeira solto na água.

26
A freqüência de atividades foi maior no período da tarde, que no período da
manhã, exceto o comportamento de forrageio, que se apresentou praticamente
equitativo, com leve predomínio no período da manhã (50,06%). A categoria descanso
foi mais freqüente no período da tarde (66,52%), contudo não se verificou diferença
significativa na freqüência desse comportamento entre os períodos manhã e tarde
(Figura 2).
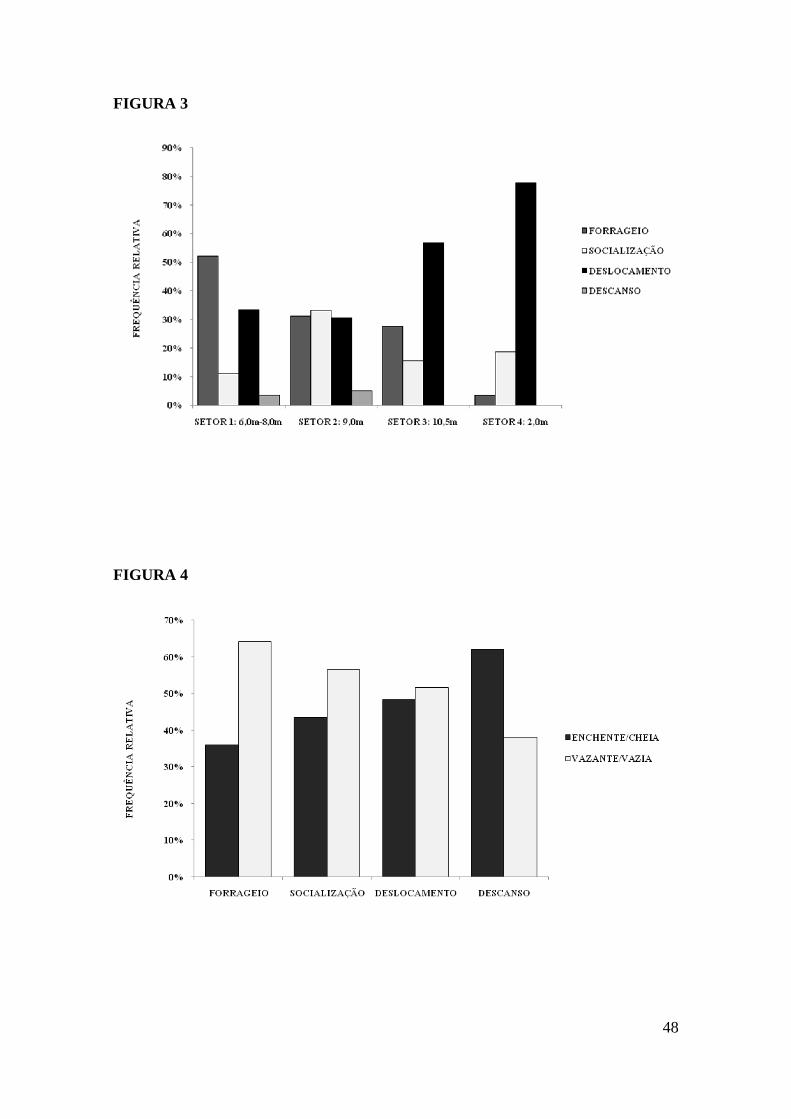
O Setor 2 (profundidade 9,0 m, fundo arenoso e baixa declividade) foi o mais
utilizado pelos animais. Do total das atividades observadas 70,96% ocorreram nesse
setor. No Setor 1 (profundidade 6,0 – 8,0m, fundo arenoso/orgânico) foram executadas
19,18% do total, seguido pelo Setor 4 (profundidade 3,0 m, fundo de recifes de coral)
com 6,68% e pelo Setor 3 (profundidade 10,5 m, fundo argila siltosa/orgânico) com
3,18% do total de atividades observadas. Verificou-se diferença significativa na
freqüência de uso dos setores pelos animais (Kruskal Wallis p=0, 0302). Quando
analisados dois a dois verificou-se que a diferença ocorre apenas entre os setores 2 e 3
(p=0, 0286). Houve diferença significativa em relação as freqüências das atividades
apresentadas em cada um dos setores. No setor 1 a diferença mais marcante se deu entre
forrageio e descanso (p<0,05) e entre deslocamento e descanso (p<0,001), no setor 2 as
diferenças foram mais significativas entre o forrageio e o descanso (p<0,001) e nos
setores 3 e 4 a diferença maior se deu entre as freqüências de deslocamento e de
descanso, ambas com valor de p< 0,001 (Figura 3). O teste de Spearman mostrou que
não existe correlação entre as atividades e a profundidade do setor (p=0, 331).
A área com profundidade de 9,0m, fundo arenoso e com baixa declividade,
destacou-se não só por ser a área mais utilizada, mas todas as categorias
comportamentais observadas foram mais freqüentes neste setor. A área foi responsável

27
por 66,64% de todo o forrageio observado, 85,81% da socialização, 61,80% do
deslocamento e 84,44% de todos os eventos de descanso presenciados.
O Setor 1 foi utilizado pelos golfinhos para o forrageio e o deslocamento. Essas
categorias comportamentais representaram 52,04% e 33,37% respectivamente, das
atividades realizadas neste setor. No Setor 2 a distribuição das atividades foi mais
equitativa, exceto pelo descanso que representou apenas 5,08% das atividades realizadas
neste setor. Já o Setor 3 foi utilizado em grande parte para o deslocamento, 56,72% das
atividades foram para este fim, seguido pelo o forrageio com 27,61% do total. O
deslocamento também foi a atividade mais observada no Setor 4, representando 77,66%
do total. Este setor foi utilizado basicamente como área de entrada e saída dos animais.
Não foi observada a categoria descanso nestes dois últimos setores (Figura 3).
A maré não mostrou influência estatisticamente significativa nas atividades
realizadas pelos golfinhos. Embora não estatisticamente significante houve maior
freqüência de atividades durante a maré vazante/vazia. Do total de atividades de
forrageio 64,08% (p=0, 3429) ocorreram na maré vazante/vazia. Com relação à
socialização 56,50% ocorreram na maré vazante/vazia e do total de observações
referentes ao deslocamento 51,66% foram na maré vazante/vazia, somente o descanso
foi mais freqüente na maré enchente/cheia, 61,98% (Figura 4).
Quando observamos as categorias a partir das marés, vemos que das atividades
realizadas no período de maré enchente/cheia o deslocamento foi o mais freqüente,
representando 35,80% do total. Na maré vazante/vazia o comportamento mais freqüente
foi o forrageio com 38,74%.
Foram observados 124 episódios de interação com embarcações (navios e
embarcações portuárias, turismo e pesca). Destes encontros 113 foram considerados
negativos, 9 neutros e apenas 2 foram tidos como positivos. Em quatro ocasiões os

28
animais foram vistos forrageando na lama deixada pelo rastro dos navios de grande
porte e rebocadores.
DISCUSSÃO
Os resultados gerais de tamanho de grupo, freqüência de ocorrência e categorias
comportamentais observados, foram bastante semelhantes aos obtidos anteriormente em
trabalhos realizados no mesmo tipo de área (Geise et al. 1999, Guilherme-Silveira and
Lima 2007; Azevedo et al. 2007, Araújo et al. 2008). A freqüência de ocorrência dos
animais na área (81,59%) permite classificar o Porto de Maceió como área de
concentração para a espécie, segundo a definição proposta por Cremer et al. (2004).
Araújo et al. (2007) em estudo recente nos Portos de Recife e Suape em Pernambuco
observaram grupos variando entre 1 e 10 indivíduos, assim como observado por nós
para o Porto de Maceió.
Pequenas agregações são comuns para a espécie, que tem preferência por áreas
fechadas como baías e Portos, e também foram observadas em outros trabalhos
realizados em áreas semelhantes (Geise et al. 1999, Araújo et al. 2001, Azevedo et al.
2005, Souto et al. 2006, Cremer 2007, Guilherme-Silveira and Lima 2007). No entanto,
já foram avistadas grandes agregações desses animais na Baía de Sepetiba e Paraty no
Rio de Janeiro, sendo o maior grupo já encontrado composto por 450 indivíduos (Lodi
and Hetzel 1998, Lodi 2003). Aparentemente existe uma tendência à formação de
pequenos grupos no Norte/Nordeste do Brasil e grupos maiores no Sul/Sudeste que
pode estar relacionada às características oceanográficas e fisiográficas de cada região,
além das atividades humanas presentes na área (Parente et al. 2007, Araújo et al. 2008).

29
Uma maior freqüência de grupos foi observada no período da tarde, diferente
do que foi observado por Geise et al. (1999) em Cananéia e Lodi (2003) em Paraty.
Porém a diferença não foi significativa estatisticamente.
Infantes foram efetivamente observados ao longo de todo o estudo, estando
presentes em quase todas as avistagens. Essa presença de infantes nos grupos ao longo
de todo o ano já foi reportada por outros autores como Rosas and Monteiro-filho (2002).
Este fator associado a grande freqüência do comportamento de socialização envolvendo
infantes indicam que apesar de impactada por diversos fatores antrópicos a região do
Porto de Maceió oferece boas condições de segurança e alimentação para ser utilizada
como área de cria (Mattila et al. 1994, Santos et al. 2001, Lodi 2003).
A proporção por faixa etária foi de quase um infante para cada dois adultos
avistados. Este padrão é um pouco incomum para Sotalia guianensis, mas já foi
observado por Geise et al. (1999) para a região do estuário de Cananéia. Grupos com
alto índice de infantes em geral possuem um maior número de indivíduos. Alguns
autores associam esse aumento no tamanho do grupo a uma maior proteção aos recém-
nascidos e a um incremento no desenvolvimento dos processos de aprendizagem
(Karczmarki 1999, Lodi 2003, Azevedo et al. 2005). O pequeno tamanho de grupos
com infantes no Porto pode estar associado ao tamanho da área e a proteção conferida
pela estrutura do cais (Araújo et al. 2007).
Os animais estiveram envolvidos em atividade de forrageio durante a maior
parte do tempo seguida pelo deslocamento. Estas duas categorias comportamentais se
revezam entre as mais freqüentes em quase todos os trabalhos relacionados a uso de
habitat já descritos para a espécie em áreas semelhantes (Geise 1999, Edwards and
Schnell 2001, Daura-Jorge et al. 2005, Rossi-Santos 2006, Azevedo et al. 2007) e para
outros delfins (Ballance 1992, Hanson and Defran 1993, Bearzi et al. 1999,

30
Karczmarski and Cockcroft 1999). Para alguns autores as diferenças nas freqüências
comportamentais podem estar relacionadas ao tipo de metodologia utilizada (Mann
1999, Daura-Jorge et al. 2005). No caso do Porto de Maceió acredita-se que a
distribuição observada das atividades está relacionada principalmente a movimentação
das embarcações e a distribuição e abundancia de presas.
A maioria dos episódios de forrageio foi relacionada a estratégias individuais ou
em pequenos grupos de até três indivíduos. Por quatro vezes os animais foram
observados forrageando na pluma levantada pela passagem dos navios em estratégia
semelhante ao que foi descrito por Rossi-Santos and Wedekin (2006).
A proporção de tempo gasto em atividades de socialização e descanso foi
considerada alta. A presença constante de infantes no grupo participando das atividades
dos adultos e eventualmente a formação de grupos compostos apenas por infantes
parece ter contribuído para os longos períodos de intensa socialização. Lodi (2003)
também observou em Paraty grupos formados apenas por infantes e a presença deles nas
atividades dos adultos, o que foi reafirmado por Monteiro et al. (2006). Esta
participação dos infantes em atividades de adultos está relacionada à aprendizagem e
treinamento dos comportamentos que serão utilizados nas próximas fases de suas vidas
(Spinelli 2002, Monteiro et al. 2006, Araújo et al. 2008).
Aparentemente o tempo gasto com comportamentos não alimentares é
proporcional a eficiência do forrageio. Quanto maior a eficiência menos tempo é gasto
procurando por presas e mais tempo é disponibilizado para outras atividades. Essa
eficiência está diretamente ligada à riqueza do habitat, abundância e disponibilidades de
alimento (Hui 1979, Karczmarski and Cockcroft 1999, Bearzi et al. 1999, Neumann
2001, Paisley and Garshelis 2006). No caso do Porto de Maceió a alta freqüência de
socialização e descanso, aparenta estar ligada a esse fator uma vez que a área é bastante

31
conhecida pelos pescadores locais como sendo de concentração de cardumes de peixes
que são sabidamente presas do boto Sotalia guianensis, como tainha (Mugil sp.), espada
(Trichiurus lepturus), linguado (Citharichthys sp.), sardinha (Pellona harrowere) e
manjuba (Anchoa filifera) (Borobia and Barros 1989, Di Beneditto and Ramos 2004,
Cremer 2007).
As categorias comportamentais socialização, deslocamento e descanso
apresentaram maior freqüência no período da tarde e o forrageio se distribuiu
equitativamente nos dois períodos, a diferença, contudo não foi significativa. Uma
maior freqüência do comportamento de forrageio é comumente associada ao período da
manhã ou a um pico pela manhã e outro no fim da tarde, não só para Sotalia guianensis,
mas também para outros delfins. Este fator tem sido imputado a variações sazonais
ambientais como fotoperíodo, estação do ano e maré, mas muito mais a variação
sazonal diária das presas consumidas por esses animais (Saayman, et al.1972, Hanson
and Defran 1993, Harzen 1998, Geise et al. 1999, Karczmarski and Cockcroft 1999,
Azevedo et al. 2007).
Os resultados corroboram com a afirmação de que a freqüência e distribuição
dos atividades comportamentais se dá de acordo com a distribuição e abundância das
presas. Isso se pode concluir porque as águas nesta região apresentam colunas
homogêneas, isotérmicas e isosalinas não apresentando estratificação considerável e as
correntes na camada superior são bastante influenciadas pelos ventos enquanto que na
camada inferior são principalmente governadas pela maré, que tem amplitude média de
2,4m em sizígia (INPH 1993). A princípio esperávamos que influência da maré na
freqüência comportamental fosse significativa, como já foi antes observado em baías
fechadas e marés com grande amplitude (Karczmarski and Cockcroft 1999), porém isto
não se confirmou.

32
Mesmo a diferença não se mostrando estatisticamente significativa, houve maior
concentração das freqüências comportamentais no período da maré vazante/vazia,
exceto pelo descanso, que foi mais freqüente na maré enchente/cheia. O forrageio foi à
atividade mais observada na maré vazante/vazia e o deslocamento a mais freqüente na
maré enchente/cheia. Essa predominância é comum em S. guianensis e já foi observada
em locais com grande variação na amplitude da maré como observado por Reis (2002)
em Ilhéus na Bahia. A maior concentração das atividades de forrageio na maré
vazante/vazia é atribuída à diminuição da massa de água, facilitando a ecolocalização e
a captura das presas que provavelmente estão mais concentradas (Monteiro-Neto et al.
1995, Reis 2002, Flores and Bazzalo 2004, Daura-Jorge et al. 2005). Pode estar também
associada à distribuição dos nutrientes e do zooplancton, regida pelas correntes e pela
maré, o que afeta diretamente a distribuição das presas (Ballance 1992, Hanson and
Defran 1993, Harzen 1998). Em relação ao local de estudo pode-se dizer que a
qualidade da água também influencia na disponibilidade de presas. É comum em região
de portos haver contaminação por metais pesados e outros contaminantes, a depender do
tipo de material comercializado. E essa contaminação interfere diretamente na riqueza
de espécies. Isso, porém não ocorre na região estudada conforme resultados obtidos por
Oliveira and Campion (1999), que mostram que não há contaminação significativa por
metais pesados na água, na pluma nem tão pouco no solo da região do Porto de Maceió.
Apesar da proximidade com possíveis fontes poluidoras a contaminação da região está
em grande parte associada a efluentes domésticos e não industriais.
O setor com profundidade média de 9,0m e fundo arenoso e baixo declive foi o
responsável por 70,96% de todas as ocorrências comportamentais. A presença de S.
guianensis já foi observada em diversas profundidades, tipos de fundo e temperaturas,
sendo considerada uma espécie generalista de hábitos e habitats (Edwards and Schnell

33
2001, Cremer et al. 2004, Daura-Jorge et al. 2005, Rossi-Santos et al. 2006, Valle 2006,
Azevedo et al. 2007), assim como outras espécies de delfins costeiros como Tursiops
truncatus e Souza chinensis (Shane 1980, Karczmarski et al. 2000) reafirmando sua
distribuição em função da disponibilidade de recurso alimentar.
O que chama atenção neste caso é que este setor compreende boa parte do canal
de entrada dos navios de grande porte e onde se concentra o maior tráfego de
embarcações, por esse motivo, a princípio não esperávamos que esse setor apresentasse
as maiores freqüências de atividade. Porém em função da composição do fundo do setor
3 e do interior do cais (argila siltosa/matéria orgânica) além do material orgânico que é
carreado para a água acidentalmente em função dos ventos, é comum verificar a entrada
de grandes cardumes nessa direção (setores 3 e cais) e para isso esses cardumes tem que
passar pelo setor 2.
A concentração dos comportamentos no canal de entrada pode ser interpretada
como uma estratégia de forrageio em que os animais têm acesso a uma maior
quantidade de presas em função do movimento e a abundancia das presas nesse local
(Ballance 1992, Harzen 1998). Neste caso o perigo do tráfego seria compensado pela
grande quantidade de recursos disponíveis. Por outro lado, sendo o setor 2 mais aberto,
permite a dispersão dos animais quando da aproximação de embarcações, o que o cais
fechado não permitiria. Este tipo de resposta comportamental foi também observado por
Pereira et al. (2007) para a baía Norte de Santa Catarina.
Esta condição se confirmou quando no final do período de coleta de dados houve
a construção de um novo berço no cais comercial fazendo com que muitos cardumes
não seguissem mais o regime de entrada e saída em função da maré e se fixassem nos
pilares do novo cais. Neste mesmo período os botos passaram a entrar muito mais
freqüentemente no cais para forragear, executando estratégias em grupo e utilizando a

34
parede do cais como anteparo. Porém, isso somente acontecia quando o cais estava
praticamente vazio e não foram vistos grupos com infantes nesses episódios.
Os resultados da análise dos encontros com embarcações indicam que esse fator
apresenta forte influência no comportamento dos golfinhos. Menos de 10% dos
encontros foi considerado neutro ou positivo. Ficou claro que os barcos de pesca e as
lanchas de turistas interferem mais que os navios de grande porte ou rebocadores. A
aproximação de barcos de pesca em geral culminava na saída do grupo dá área,
enquanto a reação à passagem dos navios normalmente causava apenas o afastamento
temporário dos animais.
Os barcos de pesca e lanchas de passeio apesar de serem menores estão sempre
com os motores ligados ao passar pelo local e se deslocam em maior velocidade que os
navios, além disso, é comum observarmos estes barcos desviando de seu curso e indo na
direção dos botos. Em relação aos barcos de pesca o direcionamento é visivelmente
proposital, chegando algumas vezes a perseguirem os animais. Esses episódios
diminuíram após algum tempo de nossa presença na área. Quanto às lanchas a
aproximação se dá no intuito de observar os animais, que logo se afastam. No caso dos
navios a aproximação é muito mais lenta e constante, em geral com o motor desligado,
curso previsível e sem tentativa de contato com os animais (Constantine 2004, Araújo et
al. 2008). Em relação aos rebocadores, como todas as embarcações de responsabilidade
do Porto de Maceió, são orientados pela direção a evitarem qualquer tipo de
interferência no comportamento dos botos. Nesse caso, a ameaça é mínima, pois eles
desviam dos animais ainda distantes evitando a rota de colisão.
Pereira et al. (2007) sugere que o barulho e a freqüência dos motores seja motivo
de aumento das reações negativas em Santa Catarina, enquanto Lusseau (2003) afirma
que o tipo de aproximação é o fator mais impactante na determinação de uma resposta

35
comportamental dos animais do que o tipo de embarcação, hipótese corroborada por
Santos-Jr et al. (2006) e Tosi and Ferreira (2008) em estudos recentes em Pipa no Rio
Grande do Norte. No caso do Porto de Maceió observamos uma junção destes dois
fatores se compararmos o tipo de aproximação e a intensidade do barulho produzido
pelos navios e pelas embarcações de pesca e de turismo.
Nossos resultados demonstram que a região do Porto de Maceió apresenta as
condições necessárias para a manutenção da presença dos golfinhos na área. Esses
animais vem em busca da proteção e facilitação conferida pela estrutura do cais para
forragear e cuidar das crias. A composição do fundo, a profundidade e a maré não são
fatores condicionantes para a presença dos animais na área. Esses fatores também não
detêm grande influencia em relação ao comportamento dos golfinhos na área. É
necessário estabelecer um programa de educação ambiental em relação a marina e a
colônia de pescadores para evitar o encontro negativo entre as embarcações de turismo e
pesca e os animais.
Futuros trabalhos para identificação dos animais que freqüentam a região são
importantes no intuito de descobrir se a população é residente ou não e o
monitoramento desses animais pode vir a acrescentar novas informações sobre a
ecologia dessa espécie e a ocorrência dessas populações em regiões de portos.

36
AGRADECIMENTOS
Os autores agradecem a administração do Porto de Maceió pela permissão concedida
para realização deste trabalho na pessoa do Sr. Berenaldo Lessa, e por todo o apoio
estrutural. A MAMMALIA - Conservação & Pesquisa pelo auxílio técnico no
transporte para a coleta de dados. Aos pesquisadores Bruno Collaço, Ana Eduarda
Campos e Juliana Glasherster pelo auxilio no trabalho de campo. Agradecem também
ao Programa de Pós-graduação da Universidade Federal do Rio Grande do Norte –
UFRN.

37
LITERATURA CITADA
ALTMAN, J. 1974. Observational study of behaviour: sampling methods. Behaviour,
Leiden, 49: 227-265.
ARAÚJO, J.P., PASSAVANTE, J.Z.O., and A.S. SOUTO. 2001. Behavior of the
estuarine dolphin, Sotalia guianensis, at Dolphin Bay – Pipa – Rio Grande do Norte –
Brazil. Tropical Oceanography 31 (2): 101-112.
ARAÚJO, J.P., ARAÚJO, M.E.,SOUTO, A.S., PARENTE, C.L., and L. GEISE.
2007.The influence of seasonality, tide and time of activities on the behavior of Sotalia
guianensis (Van Bénéden) (Cetacea, Delphinidae) in Pernambuco, Brazil. Revista
Brasileira de Zoologia 24 (4): 1122–1130.
ARAÚJO, J.P., SOUTO, A.S., GEISE, L., and M.E. ARAÚJO. 2008. The behavior of
Sotalia guianensis (Van Bénéden) in Pernambuco coastal waters, Brazil, and a further
analysis of its reaction to boat traffic. Revista Brasileira de Zoologia 25 (1): 1–9.
AZEVEDO, A.F., VIANA, S.C., OLIVEIRA, A.M., and M.V. SLUYS. 2005. Group
characteristics of marine tucuxis (Sotalia fluviatilis) (Cetacean: Delphinidae) in
Guanabara Bay, South Eastern Brazil. Journal Marine Biological.85: 209-212.
AZEVEDO, A.F., VIANA, S.C., OLIVEIRA, A.M., and M.V. SLUYS. 2007. Habitat
use by marine tucuxis (Sotalia guianensis) (Cetacea: Delphinidae) in Guanabara Bay,
south-eastern Brazil. Journal of Marine Biology, 87: 201–205.

38
BAIRD, R. W., and M. B. HANSON. 2004. Diet studies of “Southern Resident” killer
hales: prey sampling and behavioral cues of predation. Cascadia Research Collective,
Olympia, WA. Nacional Oceanic and Atmospheric Administration, Seattle, WA.
BALLANCE, L. T. 1992. Habitat use patterns and ranges of the bottlenose dolphin in
the Gulf of California, Mexico. Marine Mammal Science, 8(3): 262-274.
BEARZI, G., POLITI, E., and G.N. DI SCIARA. 1999. Diurnal behavior of free-
ranging bottlenose dolphins in the Kvarneric (Northern Adriatic sea). Marine Mammal
Science, 15(4): 1065 – 1097.
BOROBIA, M., and N.B. BARROS. 1989. Notes on the diet of marine Sotalia
fluviatilis. Marine Mammal Science 5 (4): 395-399.
BOROBIA, M., SICILIANO, S., LODI, L., and W. HOEK. 1991. Distribution of the
South American dolphin Sotalia fluviatilis. Canadian Journal of Zoology, 69: 1025-
1039.
CABALLERO, S., TRUJILLO, F., VIANNA, J.A., BARRIOS-GARRIDO, H.,
MONTIEL, M.G., BELTRÁN-PEDREROS, S., MARMONTEL, M., SANTOS, M.C.,
ROSSI-SANTOS, M., SANTOS,F.R., and C.S. BAKER. 2007. Taxonomic status of
the genus Sotalia: species level ranking for “tucuxi” (Sotalia fluviatilis) and “costero”
(Sotalia guianensis) dolphins. Marine Mammal Science.
CARR, T., and R.K. BOND. 2000. Tucuxi (Sotalia fluviatilis) Occurs in Nicaragua, 800
Km North Of Its Previously Known Range. Marine Mammal Science, 16(2): 447-452.

39
CONSTANTINE R., BRUNTON D.H., and T. DENNIS. 2004. Dolphin-watching tour
boats change bottlenose dolphin (Tursiops truncatus) behaviour. Biological
Conservation 117:299–307.
CREMER, M. J. 2007. Ecologia e conservação de populações simpátricas de pequenos
cetáceos em ambiente estuarino no Sul do Brasil. Tese de Doutorado. Universidade
Federal do Paraná. Curitiba.
CREMER, M. J., HARDT, F. A. S., TONELLO JÚNIOR, A. J., SIMÕES-LOPES, P.
C., and J. S. R. PIRES. 2004. Core areas changes in Sotalia guianensis (Cetacea,
Delphinidae) population in Babitonga Bay, Santa Catarina. Revista Univille 9 (Edição
Especial): 12-16.
DAURA-JORGE, F. G., WEDEKIN, PIACENTINI, V. DE Q., and P. C. SIMÕES-
LOPES. 2005. Seasonal and daily patterns of group size, cohesion and activity of the
estuarine dolphin, Sotalia guianensis (P. J. van Benédén) (Cetacea, Delphinidae), in
southern Brazil. Revista Brasileira de Zoologia, 22 (4): 1014 – 1021.
DI BENEDITTO, A.P.M., and R.M.A RAMOS. 2004. Biology of the marine tucuxi
dolphin (Sotalia fluviatilis) in South-Eastern Brazil.Journal of the Marine Biological
Association of the United Kingdom 84: 1245-1250.
DNH. 2008. Cartas Náuticas 901, 920 - Brasil Costa Leste - Porto de Maceió.
Departamento de Navegação e Hidrologia. Marinha do Brasil. Pode ser acessado em:
www.mar.mil.gov.br

40
DOMIT, C. 2006. Comportamento de pesca do boto-cinza na região do Complexo
Estuarino-Lagunar de Cananéia/Paranaguá, Brasil. Dissertação de Mestrado (Zoologia).
Universidade Federal do Paraná. Curitiba, Paraná.
EDWARDS, H. H., and G. D. SCHNELL. 2001. Status and ecology of Sotalia
fluviatilis in the Cayos Miskito Reserve, Nicaragua. Marine Mammal Science 17(3):
445-472.
FLORES P. A. C., and M.BAZZALO. 2004. Home ranges and movement patterns of
the marine tucuxi dolphin, Sotalia fluviatilis, in Baía Norte, southern Brazil. Lajam -
Latin American Journal of Aquatic Mammals, 3(1): 37-52.
GEISE, L. 1991. Sotalia guianensis (Cetacea, Delphinidae) population in the Guanabara
Bay, Rio de Janeiro, Brazil. Mammalia 55(3) 371-379.
GEISE, L. 2006. The correct use of the vernacular names of South American dolphins
genus Sotalia Gray (Mammalia, Cetacea, Dephinedae). Revista Brasileira de Zoologia
23(4): 1275-1276.
GEISE, L.,GOMES, N., and R. CERQUEIRA. 1999. Behaviour, habitat use and
population size of Sotalia fluviatilis (Gervais, 1853) in the Cananéia estuary region, São
Paulo, Brasil. Revista Brasileira de Biologia 59(2): 183-194.
GRIGG, E., and H.MARKOWITZ. 1997. Habitat use by bottlenose dolphins (Tursiops
truncatus) at Turneffe Atoll, Belize. Aquatic Mammals., v. 23 (3):163-170.
GUILHERME-SILVEIRA, F.R., AND F.J.L. SILVA. 2007. Behavioural seasonality of
the estuarine dolphin, Sotalia guianensis, on the North-Eastern Brazilian coast. JMBA2-
Biodiversity Records Published on-line, 1–5.

41
HANSON, M.T., and R.H. DEFRAN. 1993. The behaviour and feeding ecology of the
Pacific coast bottlenose dolphin, Tursiops truncatus. Aquatic Mammals 19:127–142
HARZEN, S. 1998. Habitat use by the bottlenose (Tursiops truncatus) in the Sado
estuary, Portugal. Aquatic Mammals, 24(3) 117-128.
HASTIE, G. D., WILSON, B., WILSON L. J., PARSONS K. M., and P. M.
THOMPSON. 2004. Functional mechanisms underlying cetacean distribution patterns:
hotspots for bottlenose dolphins are linked to foraging. Marine Biology, 144: 397-403.
HETZEL, B., and L. LODI. 1993. Baleias, botos e golfinhos. Guia de identificação para
o Brasil. Ed. Nova Fronteira. Rio de Janeiro.
HUI, C. 1979. Undersea topography and distribution of dolphins of the genus Delphinus
in the Southern California Bight. Journal of Mammalogy, 60(3): 521-527.
INMET. 2008. Instituto Nacional de Meteorologia - Estação de Referência 82994 –
Maceió, 1961-1977. Pode ser acessado em: www.inmet.gov.br.
INPH. 1993. Parecer técnico sobre dragagens e despejos dos dragados na área portuária
de Maceió. Rio de janeiro.
KARCZMARSKI, L., and V. G. COCKCROFT. 1999. Daylight behaviour of
Humpback dolphins Sousa chinensis in Alagoa Bay, South Africa. International Journal
of Mammalian Biology. 64: 19-29.
LODI, L.2003. Tamanho e composição de grupo dos botos-cinza, Sotalia guianensis
(Van Bénéden,1864) (Cetacea, Delphinidae), na Baía de Paraty, Rio de Janeiro, Brasil.
Atlântica, Rio Grande, 25(2): 135-146.

42
LODI, L., and B. HETZEL. 1998. Grandes agregações do boto-cinza (Sotalia fluviatilis)
na Baía da Ilha Grande, Rio de Janeiro. Bioikos, 12 (2): 26-30.
LUSSEAU, D. 2003. Efects of tour boats on the behavior of bottlenose dolphins: using
Markov chains to model anthropogenic impacts. Conservation Biology 17(6):1785–
1793.
MARTIN, P., and P. BATESON. 1993. Measuring Behaviour: an introductory guide.
Cambridge, Cambridge University Press, 200pp.
MATTILA, D.K., CLAPHAM, P.J., VÁSQUEZ, O., and R.S.BOWMAN. 1994.
Ocurrence, population composition and habitat use of humpback whales in Samana
Bay, Dominican Republic. Canadian Journal of Zoology 72: 1898-1907.
MAZE, K. S., and B. WÜRSIG. 1999. Bottle nose dolphin of San Luis Pass, Texas:
Occurrence patterns, site-fidelity, and habitat use. Aquatic Mammals, 25.2: 91 – 103.
MONTEIRO, M.S., SOUTO, A., and L. F. NASCIMENTO. 2006. Comparações entre
os comportamentos de forrageio nas diferentes faixas etárias do boto-cinza (Sotalia
guianensis) (Cetacea; Delphinidae) na Baía dos Golfinhos, Praia de Pipa, RN, Brasil.
Revista de Etologia 8 (1): 13-25.
MONTEIRO-NETO, C., OLIVEIRA, J.A. DE, ÁVILA, F.J.C., JÚNIOR, T.T.A., and
M.A.A FURTADO-NETO. 1995. Monitoramento do boto cinza, Sotalia fluviatilis
Cetacea:Delphinidae) em Fortaleza, Estado do Ceará, Brasil. Arquivo Ciências do Mar,
Fortaleza, 29(1-2): 28-35.

43
NASCIMENTO, L. F. 2002. Descrição comportamental do boto cinza (Sotalia
fluviatilis, GERVAIS, 1853) no litoral sul do estado do Rio Grande do Norte.
Dissertação de Mestrado, Universidade Federal do Rio Grande do Norte, Natal, Brasil.
NEUMANN, D.R. 2001. The activity budget of free-ranging commom dolphins
(Delphinus delphis) in the Northwestern Bay of Plenty, New Zeland. Aquatic mammals.
27(2): 121-136.
OLIVEIRA, A.M., and G.L. CAMPION 1999. Monitoramento da pluma de sedimentos
e matais pesados do material dragado do Porto de Maceió. Universidade Federal de
Alagoas-FUNDEPS.
PAISLEY, S., and D.L. GARSHELIS. 2006. Activity patterns and time budget of
Andean bears (Tremarctos ornatus) in the Apolobamba Range of Bolívia. Journal of
Zoology, 268:25-34.
PARENTE, C.L., ARAÚJO, J.P., and M.E. ARAÚJO. 2007. Diversity of cetaceans as
tool in monitoring environmental impacts of seismic surveys. Biota Neotropica 7 (1): 1-
7.
PEREIRA, M.G., BAZZALO, M., and P.A.C. FLORES. 2007. Reações
comportamentais de Sotalia guianensis (Cetacea, Delphinidae) durante encontros com
embarcações na Baía Norte de Santa Catarina. Revista Brasileira de Zoociências 9 (2):
123-135.
Porto de Maceió. 2002. Plano de Emergência da APMC. CD – ROM.
REIS, M.S.S. O boto Sotalia guianensis (Gervais, 1853) (Cetacea, Delphinidae) no
litoral de Ilhéus, Bahia: Comportamento e interações com atividades pesqueiras.
Dissertação de Mestrado, Universidade Estadual de Santa Cruz, Ilhéus.

44
ROSAS, F.C.W., and E.L.A. MONTEIRO-FILHO. 2002. Reproduction of the estuarine
dolphin (Sotalia guianensis) on the coast of Paraná, Southern Brazil. Journal of
Mammalogy. 83(2):507-515.
ROSSI-SANTOS, M.R. 2006 Ecologia comportamental do boto cinza, Sotalia
guianensis (Van Bénedén, 1874) (Cetacea: Delphinidae) na região extremo sul do
Estado da Bahia, Nordeste do Brasil. Dissertação de Mestrado. UFPR.
ROSSI-SANTOS, M.R., and L.L. WEDEKIN. 2006. Evidence of bottom contact
behavior by estuarine dolphins (Sotalia guianensis) on the eastern coast of Brazil.
Aquatic Mammals, 32(2), 140-144.
ROSSI-SANTOS, M.R., WEDEKIN, L.L., and R.S. SOUSA-LIMA. 2006. Distribution
and habitat use of small Cetaceans off Abrolhos bank, Eastern Brazil. LAJAM - Latin
American Journal of Aquatic Mammals, 5(1): 23-28.
SAAYMAN, G.S., TAYLER, C.K., and D. BOWER. 1972. Diurnal activity cycles in
captive and free-ranging Indian Ocean bottlenose dolphins (Tursiops aduncus
ehrenburg). Behaviour, 44(3/4): 213-233.
SANTOS, M.C.O., ACUÑA, L.B., and S. ROSSO. 2001. Insights on site fidelity and
calving intervals of the marine tucuxi dolphin (Sotalia fluviatilis) in southern Brazil.
Journal of Marine Biology. Ass. UK 81: 1049-1052.
SANTOS-JR, E., PANSARD, K.C., YAMAMOTO, M.E., and S. CHELLAPPA. 2006.
Comportamento do boto-cinza, Sotalia guianensis (Van Bénédén) (Cetacea,
Delphinidae) na presença de barcos de turismo na praia de Pipa, Rio Grande do Norte,
Brasil. Revista Brasileira de Zoologia. 23(3):661–666.

45
SHANE, S.H.1980. Ocurrence, moviments and distribution of bottlenose dolphin
(Tursiops truncatus) in southern Texas. Fishery Bulletin, 78(3):593-601.
da SILVA, V.M.F., and R.C. BEST. 1996. Sotalia fluviatilis, Mammalian Species.
American Society of Mammalogists.
SIMÃO, S.M., and F.R. POLETTO. 2002. Áreas preferenciais de pesca e dieta do
ecótipo marinho do boto cinza (Sotalia fluviatilis) na Baía de Sepetiba, Rio de Janeiro.
Floresta e ambiente. 9 (1): 18-25.
SOUTO, A., ARAÚJO, J.P., GEISE, L., and M.E. ARAÚJO. 2006. The surface
behavior of the estuarine dolphin in Baía dos Golfinhos, RN, Brazil: a field and
comparative study. Revista Brasileira de Zoociências 8 (2): 183-192.
SPINELLI, L.H.P., NASCIMENTO, L.F., and M.E. YAMAMOTO. 2002. Identificação
e descrição da brincadeira em uma espécie pouco estudada, o boto cinza (Sotalia
fluviatilis),em seu ambiente natural. Estudos de Psicologia 7(1): 165-171.
TOSI, C. H., and R.G. FERREIRA. 2008. Behavior of estuarine dolphin, Sotalia
guianensis (Cetacea, Delphinidae), in controlled boat trafic situation at Southern coast
of Rio Grande do Norte, Brazil. Biodiversity and Conservation.
VALLE, A. L. 2006. Eficiência e uso de hábitat para a captura de presas por golfinhos
Sotalia Fluviatilis: Comparação entre adultos e juvenis. Acta Biológica Leopondensia,
28 (1):55-60.
WÜRSIG, B., and M. WÜRSIG. 1980. Behavior and ecology of dusky dolphins,
Lagenorhynchus obscurus, in the south Atlantic. Fishery Bulletin, Seattle, 77: 871- 891.

46
LISTA DE FIGURAS
FIGURA 1: Foto aérea do Porto de Maceió (9°40’2’’S e 35°42’54’’W), destacando o
ponto de observação e os setores de estudo. Fonte: Porto de Maceió, 2002.
FIGURA 2: Freqüência relativa de ocorrência das categorias comportamentais de S.
guianensis no Porto de Maceió – AL por período do dia.
FIGURA 3: Freqüência relativa de ocorrência das categorias comportamentais de S.
guianensis em relação a profundidade no Porto de Maceió – AL.
FIGURA 4: Freqüência relativa total das atividades observadas de acordo com as
marés.

47
FIGURAS
FIGURA 1
FIGURA 2
48
FIGURA 3
FIGURA 4

49
ANEXO
Normas para publicação - Marine Mammal Science
MANUSCRIPT PREPARATION
The manuscript should be concise, logical, and unequivocal. Publication is facilitated if
authors double-check for typographical and other errors and ensure that manuscripts and
figures meet the requirements outlined below. Manuscripts that do not conform will be
returned for correction before consideration. If in doubt, consult the Editorial Office.
Authors are charged for excessive changes in proof.
A full-length Article includes a title page, abstract, keywords, text, acknowledgments,
literature citations, tables, figure captions, and figures. Notes and Letters do not have an
abstract and are prepared in narrative form without headings, except for "Literature
Cited."
All parts of the manuscript, including footnotes, tables, and figure captions, should be
typewritten, double-spaced with margins at least 2.5 cm wide. Number all pages of the
manuscript beginning with the title page and include line numbers on each page.
Underline only when the material is to be set in italics. Use capital letters only when the
letters or words are to be capitalized. Do not end a line of text with a hyphen.
Title Page
The first page should contain only the title and the name, affiliation, and complete
address (plus current address, if different) of the author(s). The title should be brief and
contain words useful for indexing and information retrieval.
Abstract and Key Words
The abstract, of not more than 200 words typed on a separate page, should precisely
reflect the contents of the paper, and focus attention on the purpose of the study,

50
principal results, conclusions and their significance. Below the abstract, provide and
identify as such three to ten key words or short phrases that will assist in cross-indexing
your article.
Text
As a general guide, The Chicago Manual of Style is recommended. Spelling should be
standard U.S. (not British) to conform to Webster's Third New International Dictionary.
Units should conform to the Système International d'Unités (SI). Non-standard
abbreviations must be defined the first time they appear. Mathematical symbols, Greek
letters, and unusual symbols should be identified clearly; superscripts and subscripts
should be legible and carefully placed.
Standard Abbreviations
General: s, min, h (hour), d (day), wk, mo, yr, g (gram), mg, kg, Hz, kHz, MHz, km, m
(meter), mm, cm, cc, mi (mile), ft, in. (note period), kn (knot), ha, gal, ml, l (liter, spell
out when used alone).
Statistics: P (probability), &Xmacr; (mean), SD, SE, CV, SEM, n (sample size), df, r
(correlation coefficient), t, F, U, Z (statistical tests); letters in equations are italicized.
Latin words and phrases (always italicized): i.e., (note comma); e.g., (note comma),
ca.; cf; in vivo; in situ; vs.; etc.; per se; et al.; via; sensu; sensu faro; sensu stricto; a
priori.
Acknowledgements
List all acknowledgments briefly under a single heading at the end of the text on a
separate page. If applicable, give the permit number under which the work was
conducted.

51
Literature Cited
References should be cited in the text in the following form: Smith (1982); Smith
(1982a, b); Smith (1983, 1984); Smith and Jones (1984); (Smith 1986); (Smith 1986,
Jones 1987); (Smith 1986; Jones 1986, 1987); more than two authors, Smith et al.
1987.
References should be double-spaced and listed alphabetically as "Literature Cited" in
the following standard form, giving the journal titles in full and author's last names
in small capital except for the first letter in full capital:
ARMSTRONG, W.A., and C.W. OLIVER. 1995. Recent use of fish aggregating device
in the eastern tropical Pacific tuna purse-seine fishery: 1990-1994. National Marine
Fisheries Service Center Administrative Report LJ-95-14 (unpublished). 47 pp.
Available from SWFC, P.O. Box 271, La Jolla, CA 92038.
GENTRY, R.L., and J.R. HOLT. 1982. Equipment and techniques for handling northern
fur seals. U.S. Department of Commerce, NOAA Technical Report NMFS SSRF-758.
15 pp.
HUBBS, C.L., W.F. PERRIN and K.C. BALCOMB. 1973. Stenella coeruleoalba in the
eastern and central tropical Pacific. Journal of Mammalogy 54:549-552.
LEATHERWOOD, S., and R.R. REEVES. 1983. The Sierra Club handbook of whales
and dolphins. Sierra Club Books, San Francisco, CA.
MURCHISON, A.E. 1980. Detection range and range resolution of echolocating
bottlenose porpoise (Tursiops truncatus). Pages 43-70 in R.-G. Busnel and J.F. Fish,
eds. Animal sonar systems. Plenum Press, New York, NY.
Multiple citations for an author and single co-author are arranged alphabetically

52
according to co-author. If there is more than one co-author, citations are arranged
chronologically.
Issue numbers are not used unless page numbering begins at 1 with each issue. The
number of pages is not given for books, but should be included for unpublished
documents, theses, and "gray literature" (government reports, technical bulletins, etc.)
Personal communications and unpublished data are not to be included under "Literature
Cited" but may be cited as footnotes, which shall include the complete name and
address of the source and the month and year of the communication or notification of
the unpublished data. A paper may be cited "in press" only if it has been accepted in
final form by a journal. Papers "submitted" or "in preparation" may not be cited as such,
but information in them may be cited as "personal communication." Any citation of a
personal communication, unpublished data, manuscript submitted or in preparation, or
unpublished report must be with the explicit permission of the lead author or person
who provided the information. Reference to non-refereed documents (e.g., contract
reports, environmental impact statements, meeting working papers) is discouraged.
Citations of these documents must be accompanied by the address where they can be
obtained. Meeting abstracts should not be cited. Any document bearing a "Do not cite
without permission" statement may be cited only with the explicit permission of the lead
author. A statement that all necessary permissions have been obtained must be included
in the cover letter accompanying the submitted manuscript. The use of gray literature is
discouraged and should only be cited when there is no primary literature to support
important findings or the interpretation of those findings presented in the manuscript.
Authors must double-check all literature cited; they are solely responsible for its
accuracy.
Taxonomic usage in Marine Mammal Science in general follows D. W. Rice (1998)

53
"Marine mammals of the world: Systematics and distribution," Society for Marine
Mammalogy Special Publication Number 4. Authors wishing to use a different
nomenclature should explain the departure in a footnote.
Tables
Excessive tabular data are discouraged. Tables should be typed separately and double-
spaced. Tables should be numbered with Arabic numerals in the sequence first
referenced in the text and have a brief title. Column headings and descriptive footnotes
should be brief. Do not use vertical rules.
Figures
Figures are costly and should be used with discretion. An illustration is justified only if
it clarifies or reduces the text.
Please note that if accepted, figures will be requested in TIFF or EPS format. Please
save line artwork (vector graphics) as Encapsulated PostScript (EPS) and bitmap files
(halftones or photographic images) as Tagged Image Format (TIFF), with a resolution
of at least 300 dpi at final size. More detailed information on the submission of
electronic artwork can be found at:
http://www.blackwellpublishing.com/bauthor/illustration.asp.

54
ARTIGO 2 – Orçamento temporal do boto (Sotalia guianensis) no Porto de Maceió
– Alagoas, Brasil
Karlla de Moraes Souza Lima Collaço 1 e Flávio José de Lima Silva1,2
1 Programa de Pós-Graduação em Psicobiologia. Universidade Federal do Rio Grande
do Norte (UFRN). Campus UFRN. Caixa Postal 1511 - CEP 59078-970. Natal, RN.
Brazil.
e-mail: [email protected]
2 Universidade do Estado do Rio Grande do Norte (UERN). Laboratório de Sistemática
e Ecologia Animal. Dept. Ciências Biológicas – Fac. Ciências Naturais. Av. Prof.
Antônio de Campos, s/n . Campus Central UERN. CEP 59600-970. Mossoró-RN.
Brazil.
e-mail: [email protected]
A ser traduzido para inglês e submetido ao periódico:
JMBA - Journal of the Marine Biological Association of the United Kingdom

55
RESUMO
O ritmo apresentado pelos animais em suas atividade representa uma
adaptação às variações ambientais e são o resultado de um complexo sistema que
envolve o forrageio ótimo, tempo de alimentação, atividades sociais e fatores físicos e
ambientais. Comparações entre populações sabidamente abaixo do equilíbrio e
populações equilibradas podem ser utilizadas para identificar indicadores de status
populacional e futuras medidas de conservação. O orçamento temporal e de atividades
vem sendo utilizado para fornecer bases para essas comparações, informações acerca
de como a população é afetada por mudanças em seu ambiente. Sotalia guianesis (Van
Bénéden, 1864) é um pequeno boto costeiro que por seus hábitos está sujeito a diversos
impactos antropogênicos e é considerado um bom bioindicador. É avistado com grande
freqüência na região do Porto de Maceió, que está sujeita a impactos com grande
tráfego de embarcações de carga, pesca e turismo, além de estar localizada próxima a
um ponto de despejo de efluentes domésticos. A partir disso o objetivo deste trabalho
foi delinear a distribuição das atividades comportamentais realizadas ao longo do dia e
a influência da geomorfologia do fundo e da profundidade sobre o orçamento temporal
e de atividades boto S. guianensis na região do Porto de Maceió – Alagoas. Foram
observados dois picos de atividade ao longo do dia distribuídos em função da atividade
de forrageio, o tipo de fundo e a profundidade pareceu não influenciar diretamente na
distribuição dos animais que se distribuíram em função das presas.
Palavras-chave: Sotalia, Orçamento temporal, Comportamento.

56
INTRODUÇÃO
O ritmo e o tempo gasto com cada tipo de atividade nos animais representa uma
adaptação às variações sazonais e diurnas dos fatores ambientais e são o resultado de
um complexo entendimento entre o forrageio ótimo, disponibilidade de alimento, tempo
de alimentação, atividades sociais, competições, restrições ambientais e demais fatores
ecológicos (Krebs & Davies, 1996; Karczmarski & Cockcroft, 1999; Silva, 2001). A
qualidade do habitat pode ser um fator determinante de diferenças apresentadas no
orçamento de atividades entre populações de uma mesma espécie (Li & Rogers, 2004).
Essas medidas de qualidade são hoje comumente empregadas na ecologia e na biologia
da conservação como base para o manejo de espécies.
O orçamento temporal e de atividades vem sendo utilizado porque pode
proporcionar valiosas informações de como os animais interagem uns com os outros e
com seu meio ambiente, como a população é afetada por mudanças no hábitat,
suprimentos alimentares, esforços reprodutivos e outros parâmetros fisiológicos, sociais
e ambientais (Neumann, 2001; Queiroz & Ferreira, 2006). Partindo dos resultados
gerados por esse tipo de trabalho pode-se utilizar o contraste entre populações
sabidamente abaixo do equilíbrio e populações equilibradas para identificar e
categorizar indicadores de status populacional e para determinar futuras ações
conservacionistas (Gellat, 2002).
O boto Sotalia guianensis (Van Bénéden, 1864) comumente utiliza áreas
específicas para cada tipo de atividade (Cremer et al., 2004; Daura-Jorge et al., 2005).
Distribui-se em regiões costeiras e abrigadas do Sul do Brasil a Honduras (Borobia et
al., 1991; Carr & Bonde, 2000; Valle, 2006; Azevedo et al., 2007), não passando muito
tempo em mar aberto (Flores & Bazzalo, 2004). Animais de hábitos generalistas,

57
utilizam desde estuários a recifes submersos para o forrageio, alimentando-se de peixes,
moluscos e crustáceos (Borobia & Barros, 1989; Valle, 2006). Sua área de uso é
considerada pequena, com tamanho máximo registrado de 136,9 km² para baía de
Guanabara no Rio de Janeiro (Azevedo et al., 2007). O tamanho máximo registrado
para a espécie foi de 2,06 m para as fêmeas (da Silva & Best, 1996) e 1,98 para os
machos (Rosas, 2000).
Em função da sua preferência por regiões costeiras esses animais estão sujeitos à
intensa interferência humana, seja diretamente por capturas acidentais ou intencionais,
seja pelo transito de embarcações nos locais de ocorrência dos animais ou ainda pelo
despejo de dejetos industriais e orgânicos (Hetzel & Lodi, 1993; Bearzi et al., 1999;
Pereira et al, 2007; Tosi & Ferreira, 2008). A região do Porto de Maceió está sujeita a
todos esses impactos, uma vez que possui grande tráfego de embarcações de carga,
pesca e turismo, além de estar localizada próxima a um ponto de despejo de efluentes
domésticos.
O estabelecimento do orçamento temporal e de atividades para esta população
pode, através da comparação com outras populações da mesma espécie, auxiliar na
determinação de seu status e no delineamento de futuras medidas de conservação não só
para a população que ocorre nesta região, mas para a outras populações ocorrentes em
regiões semelhantes.
O objetivo deste trabalho foi delinear a distribuição das atividades
comportamentais realizadas ao longo do dia e analisar se há influência da
geomorfologia do fundo e da profundidade sobre o orçamento temporal do boto S.
guianensis na região do Porto de Maceió – Alagoas.

58
MATERIAIS E MÉTODOS
Área de Estudo
O porto de Maceió (09º 40' 2” S e 35º 42' 54” W) encontra-se no centro do
litoral do Estado de Alagoas entre duas praias com intensa urbanização: Avenida e
Pajuçara. Região impactada por atividades antrópicas é cercada por estruturas que
merecem ser descritas para um melhor entendimento das condições da região. A três
quilômetros a noroeste está o emissário submarino, estrutura construída para lançar
parte dos dejetos da cidade, em princípio tratados, no mar. Um quilômetro também a
noroeste do local das observações está o “Riacho Salgadinho”, córrego que recebe
esgoto clandestino, carreando lixo para o cais do Porto durante a maré cheia.
A região possui ainda uma colônia de pescadores, a menos de um quilômetro a
norte do ponto de observação, que produz um intenso tráfego de embarcações, além
disso, a maior marina e as piscinas naturais mais freqüentadas do Estado estão a menos
de dois quilômetros a norte e a leste respectivamente do local de observação,
intensificando ainda mais a circulação de embarcações.
O cais do Porto possui três berços, um píer de granéis líquidos, um cais do
terminal açucareiro e um cais comercial que recebe também navios de turismo.
Em função da configuração geográfica da costa, prevalece a corrente de maré,
cuja direção é sul na enchente e norte na vazante, com amplitude máxima 2,60 metros
(DHN 2008, INPH 1993).
Coleta dos dados
Uma plataforma fixa com cerca de 3 metros de altura, localizada na base do
Farol do porto, que possui cerca de 4 metros em relação ao nível do mar, totalizando 7

59
metros de altura, foi utilizada como ponto de observação. Como a plataforma está
fixada na entrada do cais comercial é possível visualizar a maior parte da área do cais e
toda a área referente a mar aberto num ângulo de 240°. A área de estudo foi dividida em
quatro setores para um melhor aproveitamento dos dados de acordo com a profundidade
e as características físicas de cada subárea. A divisão foi feita utilizando pontos de
referência no Porto ou em terra. Estando, a Nordeste o Setor 1, composto parte pelo
canal de entrada e saída dos navios do porto e parte pela rota de saída dos barcos da
colônia de pesca e da marina com profundidade que varia de 6,0m a 8,0m metros,
intenso tráfego de embarcações e fundo arenoso/orgânico. A Sul, mar aberto, se localiza
o Setor 2 com profundidade média de 9,0m, fundo arenoso e uma pequena porção de
recife submerso. Parte deste setor está no canal de entrada e saída dos grandes navios. A
leste está o cais comercial, onde está localizado o Setor 3 com profundidade média de
10,5 metros. Este setor sofreu uma dragagem em 1999 e possui fundo de argila
siltosa/orgânico (INPH 1993). O Setor 4 a sudeste tem profundidade média de 2m e
fundo com formação de recife (Porto de Maceió 2002) (Figura 1).

60
Fig 1: Foto aérea do Porto de Maceió (9°40’2’’S e 35°42’54’’W), destacando o ponto de observação
e os setores de estudo. Fonte: Porto de Maceió, 2002.
Os dados analisados neste trabalho foram coletados com auxílio de binóculos 12
X 50 mm, em dias aleatórios de janeiro a abril de 2007 e de janeiro a julho de 2008. As
observações foram alternadas sendo uma no período da manhã de 07:00hs as 12:00hs e
outra no período da tarde de 12:00hs as 17:00 hs de forma que se obtivesse o mesmo
número de dias e horas de observação para os dois períodos.
Para coleta dos dados comportamentais utilizou-se como método amostral a
amostragem comportamental com registro de todas as ocorrências (Altmann 1974,
Martin and Bateson 1993). Esse método foi possível pelo pequeno número de animais,
tamanho da área e pelo tipo de visão proporcionado pela localização da plataforma, que
permite observar todos os indivíduos que estiverem dentro da área ao mesmo tempo.
Esse formato de observação foi baseado no que Baird and Hanson (2004) utilizaram
para orcas. Eles afirmam para uma área semelhante à desse trabalho que o foco estando
voltado para o comportamento e não para o grupo reduz a probabilidade de extrapolar

61
um comportamento para todo o grupo, evitando a subestimação de comportamentos
menos freqüentes como o descanso.
Consideramos apenas duas categorias etárias: adultos, e infantes para animais com
comprimento visualmente estimado inferior a 2/3 do tamanho do adulto, regularmente
acompanhado de animal adulto (Maze and Würsig 1999, Azevedo et al. 2005). Grupo
foi definido com todos os animais que estavam ao mesmo tempo dentro da área de
observação.
Dados referentes à profundidade e composição geomorfológica do fundo foram
extraídos a partir das Cartas Náuticas 901 e 920, e relatórios de pesquisas anteriores
cedidos pela administração do Porto de Maceió (INPH 1993, Oliveira and Campion
1999). As profundidades estabelecidas para os setores são referentes à baixa mar de
sizígia.
Categorias comportamentais
Quatro categorias comportamentais foram consideradas e analisados neste
trabalho, definidos com base em trabalhos anteriores e descritas a seguir:
Forrageio: Todo comportamento individual ou em grupo ligado à alimentação direta ou
indiretamente, culminando ou não na captura e consumação da presa. Sendo
consideradas as atividades de forrageio descritas para a espécie anteriormente (Geise
1991, Nascimento 2002, Monteiro et al. 2006, Souto et al. 2006);
Deslocamento: Deslocamento de um ou mais indivíduos com trajetória definida
(direção constante), velocidade variada, sem evidencia de outra atividade e com os
animais claramente em transito de uma área a outra (Harzen 1998, Edwards and Schnell
2001). A categoria deslocamento não foi contabilizada quando inserida no contexto de
outra categoria analisada neste trabalho. Por exemplo: O deslocamento executado
durante a perseguição a uma presa não foi contabilizado na categoria deslocamento e

62
sim na categoria forrageio, por entender que tal deslocamento está inserido nesse
contexto;
Descanso: Os animais se movimentam lentamente, rente à superfície, sem função
aparente, em geral realizado em grupo e em sincronia, onde se observa apenas o dorso
dos animais quando emergem para respirar. Esses animais mantêm ainda uma
orientação regular podendo, no entanto mudar de direção (Wursig and Wursig 1980,
Karczmarski and Cockcroft 1999);
Socialização: Interações entre os indivíduos usualmente com contato corporal e fora do
contexto de outro tipo de atividade. Incluindo atividades aéreas, interação com objetos,
interações agressivas entre indivíduos e comportamentos agonísticos na presença de
embarcações (Harzen 1998, Edwards and Schnell 2001, Daura-Jorge et al. 2005);
Análise dos dados
Por não apresentarem distribuição normal os dados foram comparados a partir de
testes não-paramétricos com o auxílio do programa estatístico Graphpad InStat 3.0. Para
verificar a diferença entre dois grupos independentes de amostras (diferenças entre dois
setores ou duas atividades) foi utilizado o Teste de Man-Whitney (U-Test). Para analisar
a diferença entre mais grupos independentes de amostras (diferenças entre setores e
atividades, atividades e marés) foi utilizado o teste de Kruskal-Wallis. Em todos os
testes foi adotado um nível de significância de 5%.
RESULTADOS
Presença dos animais e Orçamento temporal
Foram realizados 38 dias de observação, 5 horas por dia, totalizando 190 horas
de esforço amostral e 87 horas de esforço efetivo. Em 31(81,59%) dos dias de
observação os animais estavam presentes e em apenas em 7 dias (18,42%) não

63
houveram animais na área. A presença de infantes foi significativa, em 30 (96,77%) dos
dias observados. O maior número de animais na área ao mesmo tempo foi igual a 10
animais e o menor 1 animal, sendo a média igual a 4,15 animais. A maior freqüência foi
de 3 animais ao mesmo tempo (n=18) e os números menos freqüentes 8 e 10 animais
(n=1).
O número de avistagens em relação à presença de indivíduos teve maior
efetividade no período da tarde. Do número total de avistagens 57,85% ocorreram no
período da tarde. Em relação à faixa etária, 60,02% do total de indivíduos adultos
observados e 57,14% dos infantes foram observados no período da tarde e 39,97% do
total de adultos e 42,85% dos infantes foram observados no período da manhã. As
diferenças, contudo não foram consideradas significativas pelos testes estatísticos.
Dois picos de ocorrência dos animais na área foram observados, um das 8:00 as
11:00 e o segundo das 13:00 as 16:00. Sendo o horário entre 13:00 e 13:59 o de maior
número de avistagens. Por outro lado das 11:00 as 11:59 houve um grande decréscimo
no número de animais presentes nas área, voltando a ocorrer uma diminuição entre
16:00 e 17:00. Também no intervalo entre 16:00 e 17:00 ocorreu um incremento na
proporção adultos/infantes presentes na área.
Forrageio foi a categoria comportamental mais freqüente representando 34,25%
das atividades, seguido pelo deslocamento com 32,98% e pela socialização com
27,59%. O descanso foi responsável por apenas 5,18%, do total das atividades.
Estatisticamente não houve diferença significativa entre as atividades quando analisados
os totais (p<0,2301), porém quando analisadas as categorias uma a uma o teste de
Kruskal-Wallis mostrou que existe diferença entre forrageio e descanso (p<0,01),
socialização e descanso (p<0,05) e entre deslocamento e descanso (p<0,001).

64
Quase todas as atividades obtiveram maior freqüência no período da tarde,
apenas forrageio obteve um percentual ligeiramente maior no período da manha
(50,06%). A categoria descanso foi a que obteve maior diferença entre as freqüências
apresentadas nos períodos manhã e tarde. Do total, 66,52% desse tipo de
comportamento ocorram no período da tarde e 33,48% no período da manhã. O teste de
Wilcoxon no entanto, não mostrou significância entre os dois períodos (p=0,5) para o
descanso.
A Socialização apresentou 57,85% de freqüência a tarde e 42,15% no período
da manhã e o deslocamento 57,81% para a tarde e 42,18% para a manhã. O teste de
Kruskal Wallis não mostrou diferença significativa em relação às atividades e os
períodos do dia (p<0,4408).
As atividades apresentaram maior número de ocorrências em determinados
horários do dia, sendo forrageio e socialização as mais observadas ao longo da coleta de
dados. O forrageio obteve um pico no número de ocorrências pela manhã (08:00 as
09:59) e outro pico, de maior freqüência, à tarde (13:00 as 13:59). A socialização obteve
também um pico pela manhã no mesmo horário do forrageio e outro à tarde, sendo o
horário entre 15:00 e 17:00 o que obteve maior número de ocorrência deste
comportamento. Socialização foi, além disso, a atividade mais observada no período
entre 16:00 e 17:00 quando analisados os totais de números de ocorrência. O
deslocamento teve seu ápice no intervalo entre 15:00 e 16:00, o que coincidiu com o
início do horário de maior freqüência de saída dos animais da área de observação. O
descanso também foi observado com maior freqüência no fim da tarde entre 15:00 e
17:00 (Figura 2).
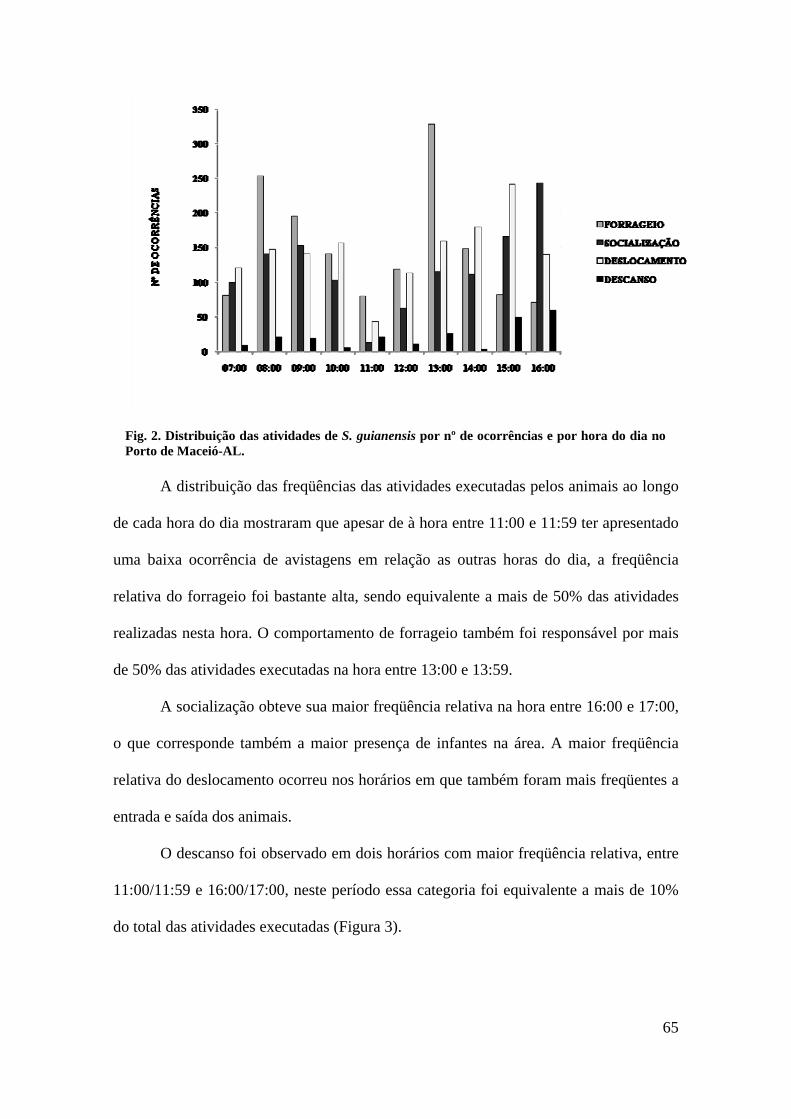
65
Fig. 2. Distribuição das atividades de S. guianensis por nº de ocorrências e por hora do dia no Porto de Maceió-AL.
A distribuição das freqüências das atividades executadas pelos animais ao longo
de cada hora do dia mostraram que apesar de à hora entre 11:00 e 11:59 ter apresentado
uma baixa ocorrência de avistagens em relação as outras horas do dia, a freqüência
relativa do forrageio foi bastante alta, sendo equivalente a mais de 50% das atividades
realizadas nesta hora. O comportamento de forrageio também foi responsável por mais
de 50% das atividades executadas na hora entre 13:00 e 13:59.
A socialização obteve sua maior freqüência relativa na hora entre 16:00 e 17:00,
o que corresponde também a maior presença de infantes na área. A maior freqüência
relativa do deslocamento ocorreu nos horários em que também foram mais freqüentes a
entrada e saída dos animais.
O descanso foi observado em dois horários com maior freqüência relativa, entre
11:00/11:59 e 16:00/17:00, neste período essa categoria foi equivalente a mais de 10%
do total das atividades executadas (Figura 3).

66
Fig. 3. Distribuição hora a hora das categorias comportamentais executadas por S. guianensis no Porto de Maceió, AL.
Influência da Geomorfologia e da Profundidade
Não houve preferência por setor em relação ao período do dia. O Setor 2 foi
preponderante sobre os outros, 70,96% do total das atividades foram executadas neste
setor.
A distribuição das atividades mostrou que o Setor 2 (profundidade 9,0m, fundo
arenoso) obteve uma distribuição equitativa das atividades ao longo do dia, exceto no
horário entre 11:00 e 11:59, horário que obteve a menor freqüência de ocorrência de
atividades em todos os setores. No Setor 1 (profundidade 6-8m, fundo
arenoso/orgânico) houve um pico na atividade de forrageio na hora entre 13:00 e 13:59
Esse setor foi utilizado basicamente para forrageio e deslocamento. No Setor 3
(profundidade 10,5m, fundo argila siltosa/orgânico) o deslocamento foi o
comportamento preponderante. Apenas no horário entre 10:00 e 10:59 houve ocorrência
significativa dos comportamentos de forrageio e socialização. Entre 11:00 e 11:59 não
foram registradas atividades nesse setor. O deslocamento foi a atividade mais observada

67
no Setor 4 (profundidade 2m, fundo recife), estando esse comportamento associado ao
horário de maior freqüência de entrada e saída dos animais. A socialização esteve
presente neste setor com incidência significativa também associada aos horários de
entrada e saída dos animais. Semelhante ao Setor 3 não foram registradas atividades
neste setor no horário entre 11:00 e 11:59 (Figura 4).
N
º d
e o
corr
ênci
as
Fig. 4. Distribuição horária das categorias comportamentais executadas por S. guianensis em cada um dos setores observados.

68
DISCUSSÃO
Presença dos animais e Orçamento temporal
O tamanho dos grupos não diferiu dos observados na maioria das áreas de
ocorrência para a espécie e não foram observadas variações significativas nos tamanhos
de grupo relativas ao tipo de atividade apresentada (Geise et al., 1999; Daura-Jorge et
al., 2005; Souto et al., 2006). Apesar disso pode-se verificar que a maioria dos eventos
de forrageio foram individuais e os referentes à socialização e descanso em grupos um
pouco maiores ou em pares adulto/infante.
O período da tarde foi o de maior freqüência da presença dos animais na área,
também o período em que ocorreram o maior número de registros dos comportamentos,
com um grande decréscimo no horário entre 11:00 e 12:00 horas, o que difere da
maioria dos trabalhos com a espécie, em que é comum uma maior freqüência no
começo da manhã ou durante as horas mais quentes do dia (Geise et al., 1999; Lodi,
2003; Queiroz & Ferreira, 2006), contudo Edwards & Schnell (2001) encontraram
horários de freqüência diferentes para duas áreas da Reserva de Miskito na Nicarágua,
sendo uma pela manhã e outra à tarde. Eles atribuíram as diferenças a fatores físicos e
ambientais como topografia, profundidade, temperatura da água e salinidade, além da
distribuição e abundância de presas. Para o Porto de Maceió acreditamos que a
freqüência da presença dos animais esteja mais relacionada com o movimento diário das
presas, e este diretamente ligado a fatores ambientais como observado por Würsig &
Würsig (1980), Hanson & Defran (1993) e Karczmarski & Cockcroft (1999) para outros
delfins. Além disso, o padrão de horários da presença dos animais na área e do
comportamento de forrageio foi bastante semelhante ao encontrado por Sayman (1972)

69
e Hanson & Defran (1993) para Tursiops truncatus e também por Azevedo et al. (2007)
para Sotalia guianensis, com dois picos para a atividade, no início da manhã com
declínio no meio do dia e novo pico no final da tarde.
O orçamento temporal apresentado pelos animais foi em parte semelhante aos
obtidos para outras populações em condições semelhantes (Geise et al., 1999; Edwards
& Schnell, 2001; Daura-Jorge et al., 2005; Rossi-Santos, 2006; Azevedo et al., 2007) e
para outros delfins (Ballance, 1992; Hanson & Defran, 1993; Bearzi et al., 1999;
Karczmarski & Cockcroft, 1999). Exceto pela alta porcentagem de tempo gasto em
socialização que tem sido pouco comum para a espécie, e que se explica por fatores
como a alta freqüência de infantes na área por todo o período de coleta de dados e pela
grande disponibilidade de presas, que faz com que os animais necessitem gastar menos
tempo forrageando e disponham de mais tempo para outras atividades.
As freqüências das demais categorias comportamentais aparentemente estiveram
bastante ligadas ao forrageio. Os picos de socialização se distribuíram logo após os
picos de forrageio, e o horário de maior número de ocorrências desse comportamento
esteve ligado ao momento da saída do grupo da área de observação. A maior ocorrência
do deslocamento foi referente aos horários de entrada e saída dos animais na área. Este
mesmo padrão de distribuição dos comportamentos de forrageio e socialização é
apresentado por Shane et al. (1986) e por Saayman (1972) para os golfinhos Tursiops
truncatus e Tursiops aduncus no Texas e na Africa do Sul respectivamente.
O descanso foi pouco significativo, mas teve sua maior freqüência de ocorrência
anterior aos momentos de saída dos animais da área e posterior aos picos de
alimentação. Azevedo et al. (2007) sugerem que a baixa freqüência do descanso nas
observações diurnas pode estar relacionada a uma maior freqüência desse

70
comportamento no período noturno. Atem & Monteiro-filho (2006) em observações
noturnas afirma não ter detectado esse comportamento em função deste ser o mais
difícil de ser identificado.
Este padrão comportamental corrobora com o que já foi descrito para outras
espécies de delfins, no que diz respeito à distribuição das atividades desses animais. O
sucesso de forrageio é o que determina o tempo a ser gasto com as outras atividades
(Karczmarski & Cockcroft, 1999). Estas serão mais freqüentes apenas quando as
necessidades nutricionais forem satisfeitas (Neumann, 2001).
O horário entre 11:00 e 11:59 apresentou um decréscimo considerável na
presença dos animais na área e conseqüentemente na freqüência das atividades.
Saayman et al. (1972), encontraram o mesmo padrão para o delfim Tursiops aduncus.
Este mesmo autor sugere que a variação horária em relação ao forrageio esteja ligada à
influência do fotoperíodo na disponibilidade das presas. Quanto à diminuição das outras
categorias comportamentais o autor afirma que podem estar relacionados a alguma outra
variável ecológica não especificada. Em nosso caso apesar da diminuição na freqüência
de ocorrência dos animais neste horário, proporcionalmente o comportamento de
forrageio foi bastante alto, não justificando, portanto, a diminuição na disponibilidade
de presas. Uma observação mais profunda deve ser feita sobre este aspecto para
esclarecê-lo melhor.
Influência da Geomorfologia do Fundo e da Profundidade
Em relação aos setores o orçamento temporal e de atividades pôde mostrar o tipo
de uso de cada setor pelos os animais. A utilização de habitats diferentes para cada tipo
de atividade é comum para S. guianensis e a determinação do uso parece estar ligada à

71
estrutura de cada região e a adaptação das populações as diferenças físicas e ambientais
encontradas nessas áreas (Edwards & Schnell, 2001; Cremer et al., 2004; Daura-Jorge
et al., 2005; Rossi-Santos et al., 2006; Valle, 2006; Azevedo et al., 2007).
O setor 2 (profundidade 9m e fundo arenoso) foi o responsável pela maioria das
atividades e estas se distribuíram como já foi descrito acima. O setor 1 (6-8m, fundo
arenoso/orgânico) apresentou um número de ocorrências muito baixas de todos as
categorias comportamentais ao longo do dia exceto no horário correspondente ao
segundo pico do forrageio, aumento este relacionado a localização do setor, vizinho ao
setor 2.
O setor 3 (10,5m, argila siltosa/orgânico) apresentou poucas ocorrências de
atividades, com um pequeno pico na atividade de forrageio equivalente ao horário de
maior freqüência desse comportamento pela manhã e um pico na freqüência do
deslocamento equivalente ao horário de saída dos animais da área. O setor 4 (2m, recife)
foi utilizado basicamente para entrada e saída dos animais. O gráfico mostra claramente
os horários utilizados para esse fim, entrada no inicio da manhã com saída no meio do
dia, retorno logo a seguir e nova saída no fim da tarde. É ainda possível observar os
picos de socialização ligados à entrada e saída dos animais.
A maior freqüência de atividades na profundidade 9m, com fundo arenoso e
menor declive, pode estar ligada tanto à distribuição das presas, quanto a segurança dos
animais em relação ao tráfego de embarcações na área. O uso mais freqüente de áreas
mais profundas e menor declive por S. guianensis já foi reportada anteriormente por
Geise et al. (1999), Simão & Poletto (2002) e Rossi-Santos (2006).

72
Esse tipo de delineamento do comportamento não havia sido feito ainda para a
espécie e pode ser bastante útil por mostrar o padrão geral da população para
determinada área. O monitoramento constante desses padrões relativos à duração,
distribuição e horários de ocorrência pode mostrar possíveis alterações no hábitat,
suprimentos alimentares e outros padrões fisiológicos, sociais e ambientais (Neumann,
2001), auxiliando na determinação de possíveis medidas de conservação.

73
AGRADECIMENTOS
Os autores agradecem a administração do Porto de Maceió pela permissão concedida
para realização deste trabalho na pessoa do Sr. Berenaldo Lessa, e por todo o apoio
estrutural. A MAMMALIA - Conservação & Pesquisa pelo auxílio técnico no
transporte para a coleta de dados. Aos pesquisadores Bruno Collaço, Ana Eduarda
Campos e Juliana Glasherster pelo auxilio no trabalho e campo. Agradecem também ao
Programa de Pós-graduação da Universidade Federal do Rio Grande do Norte – UFRN.

74
REFERÊNCIAS
Altman, J. 1974. Observational study of behaviour: sampling methods. Behaviour. 49,
227- 265.
Atem, A.C.G., Moteiro-Filho, E.L.A. 2006. Nocturnal activity of the estuarine dolphin (Sotalia guianensis) (Cetacea, Delphinidae) in the Region of Cananéia, São Paulo State, Brazil. Aquatic Mammals 32(2), 236-241.
Azevedo A. F., Oliveira A. M., Viana S. C., Van Sluys. V.2005. Group characteristics of marine tucuxis (Sotalia fluviatilis) (Cetacean: Delphinidae) in Guanabara Bay, South Eastern Brazil. Journal of Marine Biology 85, 209-212.
Azevedo A. F., Oliveira A. M., Viana S. C., Van Sluys, M. 2007. Habitat use by marine tucuxis (Sotalia guianensis) (Cetacea: Delphinidae) in Guanabara Bay, south-eastern Brazil. Journal of Marine Biology 87, 201–205.
Baird, R. W., Hanson, M. B. 2004. Diet studies of “Southern Resident” killer whales: prey sampling and behavioral cues of predation. Cascadia Research Collective, Olympia, WA. Publicação Ocasional. 11pp.
Ballance, L. T. 1992. Habitat use patterns and ranges of the bottlenose dolphin in the Gulf of California, Mexico. Marine Mammal Science, 8(3), 262-274.
Bearzi, G., Politi, E., Di Sciara, G.N. 1999. Diurnal behavior of free-ranging bottlenose dolphins in the Kvarneric (Northern Adriatic sea). Marine Mammal Science 15(4), 1065 – 1097.
Borobia, M., Barros, N.B. 1989. Notes on the diet of marine Sotalia fluviatilis. Marine Mammal Science 5 (4), 395-399. CEPENE.
Borobia, M., Siciliano, S., Lodi, L. and Hoek, W. 1991. Distribution of the South American dolphin Sotalia fluviatilis. Canadian Journal of Zoology 69, 1025-1039.
Caballero, S., Trujillo, F., Vianna, J.A., Barrios-Garrido, H., Montiel, M.G., Beltrán-Pedreros, S., Marmontel, M., Santos, M.C., Rossi-Santos, M., Santos,F.R., Baker, C.S. 2007. Taxonomic status of the genus Sotalia: species level ranking for “tucuxi” (Sotalia fluviatilis) and “costero” (Sotalia guianensis) dolphins. Marine mammal science 23 (2), 358-386.
Carr, T., Bond, R. K. 2000. Tucuxi (Sotalia fluviatilis) Occurs in Nicaragua, 800 Km North Of Its Previously Known Range. Marine Mammal Science 16(2), 447-452.
Cremer, M. J., Hardt, F. A. S., Tonello Júnior, A. J., Simões-Lopes, P. C., Pires, J. S. R.2004. Core areas changes in Sotalia guianensis (Cetacea, Delphinidae) population in Babitonga Bay, Santa Catarina. Revista Univille 9, 12-16.

75
Daura-Jorge, F. G., Wedekin, Piacentini, V. de Q., Simões-Lopes, P. C. 2005. Seasonal and daily patterns of group size, cohesion and activity of the estuarine dolphin, Sotalia guianensis (P. J. van Benédén) (Cetacea, Delphinidae), in southern Brazil. Revista Brasileira de Zoologia 22 (4), 1014 – 1021.
DNH. Cartas Náuticas 901;920 - Brasil Costa Leste - Porto de Maceió. Departamento de Navegação e Hidrologia. Marinha do Brasil. Pode ser acessado em: www.mar.mil.gov.br
Edwards, H. H., Schnell, G. D. 2001. Status and ecology of Sotalia fluviatilis in the Cayos Miskito Reserve, Nicaragua. Marine Mammal Science 17(3), 445-472.
Flores P. A. C., Bazzalo M. 2004. Home ranges and movement patterns of the marine tucuxi dolphin, Sotalia fluviatilis, in Baía Norte, southern Brazil. Lajam - Latin American Journal of Aquatic Mammals 3(1), 37-52.
Geise, L. 1991. Sotalia guianensis (Cetacea, Delphinidae) population in the Guanabara Bay, Rio de Janeiro, Brazil. Mammalia 55(3), 371-379.
Geise, L.,Gomes, N. and Cerqueira, R. 1999. Behaviour, habitat use and population size of Sotalia fluviatilis (Gervais, 1853) in the Cananéia estuary region, São Paulo, Brasil. Revista Brasileira de Biologia 59(2), 183-194.
Gelatt, T. S., Sniiff, D. B., Estes, J. A. 2002. Activity patterns and time-budget of the declining sea otter population at Amchitka island, Alaska. Journal of Wild Life Management 66(1), 29 – 39.
Hanson, M.T., Defran, R.H. 1993. The behaviour and feeding ecology of the Pacific coast bottlenose dolphin, Tursiops truncatus. Aquatic Mammals 19, 127–142
Harzen, S. 1998. Habitat use by the bottlenose (Tursiops truncatus) in the Sado estuary, Portugal. Aquatic Mammals 24(3), 117-128.
INMET – Instituto Nacional de Meteorologia - Estação de Referência 82994. Pode ser acessado em: www.inmet.gov.br.
INPH (1993) Parecer técnico sobre dragagens e despejos dos dragados na área portuária de Maceió. Instituto Nacional de Pesquisas Hidroviárias. Publicação Ocasional. Rio de Janeiro. Brasil. 21pp.
Karczmarski, L. and Cockcroft, V. G. (1999) Daylight behaviour of Humpback dolphins Sousa chinensis in Alagoa Bay, South Africa. International Journal of Mammalian Biology 64, 19-29.
Krebs, J. R. and Davies, N. B. (1996) Introdução à ecologia comportamental. Ed. Atheneu – SP. 420p.
Li, Z. and Rogers, E. (2004) Habitat Quality and Activity Budgets of White-Headed Langurs in Fusui, China. International Journal of Primatology,Vol. 25, No. 1.

76
Lodi, L. (2003) Tamanho e composição de grupo dos botos-cinza, Sotalia guianensis (Van Bénéden,1864) (Cetacea, Delphinidae), na Baía de Paraty, Rio de Janeiro, Brasil. Atlântica, Rio Grande 25(2), 135-146.
Martin, P. and Bateson, P. (1993) Measuring Behaviour: an introductory guide. Cambridge, Cambridge University Press. 200pp.
Mattila, D.K., Clapham, P.J., Vásquez, O. and Bowman, R.S. (1994) Ocurrence, population composition and habitat use of humpback whales in Samana Bay, Dominican Republic. Canadian Journal of Zoology 72, 1898-1907.
Maze, K. S. and Würsig, B. (1999) Bottle nose dolphin of San Luis Pass, Texas: Occurrence patterns, site-fidelity, and habitat use. Aquatic Mammals 25(2), 91 – 103.
Monteiro, M.S., Souto, A. and Nascimento, L. F. (2006) Comparações entre os comportamentos de forrageio nas diferentes faixas etárias do boto-cinza (Sotalia guianensis) (Cetacea; Delphinidae) na Baía dos Golfinhos, Praia de Pipa, RN, Brasil. Revista de Etologia 8 (1), 13-25.
Neumann, D.R. (2001) The activity budget of free-ranging commom dolphins (Delphinus delphis) in the Northwestern Bay of Plenty, New Zeland. Aquatic Mammals 27(2), 121-136.
Oliveira, A.M. and Campion, G.L. (1999) Monitoramento da pluma de sedimentos e metais pesados do material dragado do Porto de Maceió. Universidade Federal de Alagoas-FUNDEPS. Publicação Ocasional. 40pp
Pereira, M.G., Bazzalo, M. and Flores, P.A.C. (2007) Reações comportamentais de Sotalia guianensis (Cetacea, Delphinidae) durante encontros com embarcações na Baía Norte de Santa Catarina. Revista Brasileira de Zoociências 9 (2), 123-135.
Porto de Maceió. (2002) Plano de Emergência da APMC. CD – ROM.
Queiroz, R. E. M. (2006) Estudos sobre orçamento de atividade do boto cinza (Sotalia guianensis) no litoral sul do Rio Grande do Norte. Dissertação de Mestrado. Universidade Federal do Rio Grande do Norte, Brasil.
Rosas, F.C.W. (2000) Interações com a pesca, mortalidade, idade, reprodução e crescimento de Sotalia guianensis e Pontoporia blainvillei (Cetacea, Delphinidae e Pontoporiidae) no litoral sul do estado de São Paulo e litoral do estado do Paraná, Brasil. Tese de doutorado não-publicada, Universidade Federal do Paraná, Brasil.
Rosas, F.C.W. and Monteiro-Filho, E.L.A. (2002) Reproduction of the estuarine dolphin (Sotalia guianensis) on the coast of Paraná, Southern Brazil. Journal of Mammalogy 83(2), 507-515.
Rossi-Santos, M. R. (2006) Ecologia comportamental do boto cinza, Sotalia guianensis (Van Bénedén, 1874) (Cetacea: Delphinidae) na região extremo sul do Estado da Bahia, Nordeste do Brasil. Dissertação de Mestrado. Universidade Federal do Paraná, Brasil.

77
Saayman, G.S., Tayler, C.K. and Bower, D. (1972) Diurnal activity cycles in captive and free-ranging Indian Ocean bottlenose dolphins (Tursiops aduncus ehrenburg). Behaviour 44(3/4), 213-233.
Santos, M.C.O., Acuña, L.B. and Rosso, S. (2001) Insights on site fidelity and calving intervals of the marine tucuxi dolphin (Sotalia fluviatilis) in southern Brazil. Journal of the Marine Biological Association - UK 81, 1049-1052.
Shane, S. H., Wells, R. S. and Würsig, B. (1986) Ecology, behavior and social organization of the bottlenose dolphin: a review. Marine Mammal Science 2(1), 34-63.
Silva, F.J.L. (2001) Descrição de comportamento, sazonalidade de número de indivíduos e reprodução de golfinho rotador Stenella longirostris em Fernando de Noronha. Tese de Doutorado. Universidade Federal do Rio Grande do Norte, Brasil.
da Silva, V.M.F. and Best, R.C. (1996) Sotalia fluviatilis, Mammalian Species. American Society of Mammalogists 527:L7.
Souto, A., Araújo, J.P., Geise, L. and Araújo, M.E. (2006) The surface behavior of the estuarine dolphin in Baía dos Golfinhos, RN, Brazil: a field and comparative study. Revista Brasileira de Zoociências 8 (2), 183-192.
Tosi, C. H. and Ferreira, R.G. (2008) Behavior of estuarine dolphin, Sotalia guianensis (Cetacea, Delphinidae), in controlled boat trafic situation at Southern coast of Rio Grande do Norte, Brazil Biodiversity and Conservation.
Valle, A. L. (2006) Eficiência e uso de hábitat para a captura de presas por golfinhos Sotalia Fluviatilis: Comparação entre adultos e juvenis. Acta Biológica Leopondensia 28 (1), 55-60.
Würsig, B. and Würsig, M. (1980) Behavior and ecology of dusky dolphins, Lagenorhynchus obscurus, in the south Atlantic. Fishery Bulletin 77, 871- 891.

78
LEGENDAS DAS FIGURAS
Figura 1. Foto aérea do Porto de Maceió (9°40’2’’S e 35°42’54’’W), destacando o
ponto de observação e os setores de estudo. Fonte: Porto de Maceió, 2002.
Figura 2. Distribuição das atividades de S. guianensis por nº de ocorrências e por hora
do dia no Porto de Maceió-AL.
Figura 3. Distribuição hora a hora das categorias comportamentais executadas por S.
guianensis no Porto de Maceió, AL.
Figura 4. Distribuição horária das categorias comportamentais executadas por S.
guianensis em cada um dos setores observados.

79
ANEXO
Normas para a publicação: JMBA - Journal of the Marine Biological Association
of the United Kingdom.

80

81

82
DISCUSSÃO GERAL
A população de Sotalia guianenis que ocorre na região do Porto de Maceió
apresentou padrões comportamentais próximos aos apresentados em outras áreas com
características semelhantes (Geise et al., 1999; Guilherme-Silveira e Lima, 2007;
Azevedo et al., 2007; Araújo et al., 2008). Essa semelhança está possivelmente ligada à
adaptação que essas populações desenvolveram ao longo do tempo em relação às
modificações, antrópicas ou não, sofridas pelo hábitat.
O fato de os animais utilizarem os diferentes setores dentro da região do porto de
maneira distinta confirma as observações feitas por outros pesquisadores de que os
golfinhos podem discriminar entre áreas com diferentes estruturas ecológicas e utiliza-
las apropriadamente, de acordo com suas necessidades (Harzen, 1998). Shane et al.
(1986) afirmam que essa discriminação se deve a capacidade que alguns mamíferos
sociais, como os golfinhos, têm de aprender sobre o seu meio ambiente e guardar esse
conhecimento por toda a vida. Os autores afirmam ainda que os animais podem lembrar
onde e quando podem estar as melhores chances de encontrar presas, por exemplo. De
acordo com Bearzi et al. (1999) a flexibilidade comportamental pode contribuir para a
sobrevivência em ambientes mutantes. Segundo os mesmos autores a variabilidade dos
ambientes costeiros e a distribuição das presas produziram pressões seletivas que podem
tornaram os animais mais generalistas e conseqüentemente melhor adaptados.
Em ambientes impactados como é o caso das regiões de porto em geral, existe
um fator que aparentemente poderia ser decisivo em relação à presença dos animais que
é a poluição. Mais especificamente no caso do Porto de Maceió além da poluição vinda
dos navios, existe também a poluição em decorrência do lançamento de dejetos
orgânicos e industriais na região. No entanto, apesar de o aporte de esgoto ser lançado

83
em grande volume, à direção das correntes faz com que esse dejeto seja levado para
longe do porto na maior parte do tempo e a amplitude de maré impede o lançamento de
esgotos na maré baixa (INPH, 1993). Além disso, apesar de intensamente poluído o
riacho que despeja os dejetos também constitui um ambiente estuarino que por si só é
área de grande produtividade. Unido a esse fator, os produtos orgânicos comercializados
pelo porto, como trigo e soja, apesar de poluírem aumentam a produtividade local,
aumentando a disponibilidade de presas. Harzen (1998) afirma que a alta produtividade
pode prover nutrientes suficientes para manter uma comunidade de golfinhos em
determinada área.
No caso do Porto de Maceió, a segurança também está em questão. O litoral de
Alagoas como um todo é muito aberto, e embora seja cercado por recifes, não existem
muitas enseadas fechadas e abrigadas, fazendo da região do porto uma área abrigada e
segura apesar do intenso movimento de embarcações. Esta idéia é corroborada por
Araújo et al. (2007) em trabalho realizado no Porto de Suape em Pernambuco, região
com características geomorfológicas semelhantes as do Porto de Maceió.
Quanto à distribuição das atividades e dos animais na área, verificamos que
todas as influências estão associadas de alguma forma a distribuição e abundância das
presas ou a segurança dos animais em relação às embarcações (Würsig e Würsig, 1980;
Hanson e Defran, 1993; Karczmarski e Cockcroft, 1999; Constantine, 2004; Azevedo et
al., 2007). A utilização mais freqüente do setor 2, após as análises e comparações,
mostrou que esse setor foi mais utilizado por ser a área mais aberta com maior
facilidade de dispersão dos animais, além de concentrar a maior abundância de presas.
Embora a região esteja bastante impactada o orçamento temporal e de atividades
não diferiu de outros estudos realizados para a espécie independente das condições
ecológicas das regiões estudadas (Geise et al, 1999; Edwards e Schnell, 2001; Daura-

84
Jorge et al., 2005; Rossi-Santos, 2006; Azevedo et al., 2007). Isso ocorre
provavelmente porque os maiores impactos ocorrentes na região são decorrentes de
despejo de dejetos orgânicos, além de alterações físicas da costa em função de
construções e do estabelecimento do porto, não havendo contaminações por metais
pesados e outros contaminantes passíveis de alterar em maior grau a qualidade da água
local.
Com isso é possível afirmar que, embora haja impactos negativos sobre os botos,
os animais estão adaptados às condições ali existentes até o momento, dando suporte
para a sugestão de medidas de conservação que não permitam um maior nível de
degradação da área. Seria possível que as alterações ambientais existentes na região
ainda não houvessem alterado o comportamento dos animais por serem recentes. Porém,
a atual estrutura da região do Porto tem mais de 30 anos, o que reforça a afirmação de
que os animais estão adaptados às condições locais.

85
CONSIDERAÇÕES FINAIS
A alta freqüência da presença dos animais na área e a recorrência de alguns
indivíduos reconhecidos por fortes marcas naturais mostram que a região do Porto de
Maceió é importante para a manutenção da população que ali ocorre e dá subsídios para
futuras pesquisas acerca da identificação dos animais que freqüentam a região.
Os animais presentes na região do Porto de Maceió apresentam um padrão
comportamental típico da espécie e uma distribuição bem determinada comportamentos
com destaque para o forrageio, o que por relatos dos pescadores locais é justificado por
uma alta disponibilidade de presas no local. Fazendo da área importante local de
alimentação para essa população.
A presença constante de infantes na área e nas atividades dos adultos leva a crer
que o local é importante para a população também como área de cuidado e cria.
A distribuição das atividades nos setores foi bastante clara, o setor 1 foi utilizado
basicamente para deslocamento de entrada e saída no cais e para forrageio no segundo
pico da atividade no fim da tarde, o mesmo para o setor 3, que no entanto, teve seu pico
de forrageio pela manhã. O setor 4 foi utilizado basicamente como base para entrada e
saída dos animais na região do porto.
O setor 2 foi o mais utilizado, provavelmente pela localização que propicia
facilidade de acesso dos golfinhos aos cardumes que entram no cais do porto, além de
oferecer facilidade de deslocamento em caso de necessidade de afastamento dos animais
em função do constante tráfego de embarcações.

86
A distribuição das atividades dos animais esteve ligada muito mais a
disponibilidade e abundância das presas que ao tipo de fundo, composição e
profundidades existentes na área do porto.
O orçamento temporal mostrou que as outras atividades se distribuem em função
do forrageio. A socialização esteve diretamente ligada ao início e fim da atividade de
forrageio ou a entrada e saída dos animais da área. As freqüências mais altas de
deslocamento estiveram ligadas à entrada e saída dos animais. Essa visualização foi
possível pelo tipo de definição adotada, uma vez que esse comportamento foi
contabilizado isoladamente e não quando inserido nas outras categorias. O descanso foi
observado em sua maioria no final do dia antes da saída dos animais da área.
O encontro com embarcações foi considerado negativo. Contudo foi verificado
que as interações negativas foram mais expressivas quando ocorriam com barcos de
pesca. O tipo de aproximação e o barulho do motor foram os fatores de maior
importância para a ocorrência das respostas apresentadas pelos animais.
Não foi verificada interferência significativa da maré no comportamento dos
golfinhos, mas houve uma maior freqüência comportamental na maré vazante/vazia.
Este estudo demonstra que a região do Porto de Maceió pode ser considerada
área de concentração para a espécie, servindo como base para futuras pesquisas. O
trabalho dá ainda suporte para o estabelecimento de medidas mitigadores do impacto
causado aos animais e futuras medidas de conservação para a população ali ocorrente e
para outras populações em regiões semelhantes.

87
LITERATURA CITADA (INTRODUÇÃO E DA DISCUSSÃO GERAL)
Araújo, J.P., Araújo, M.E.,Souto, A.S., Parente, C.L. , Geise, L. (2007).The influence of seasonality, tide and time of activities on the behavior of Sotalia guianensis (Van Bénéden) (Cetacea, Delphinidae) in Pernambuco, Brazil. Revista Brasileira de Zoologia, 24 (4): 1122–1130.
Azevedo A. F., Oliveira A. M., Viana S. C. , Van Sluys, M. (2007). Habitat use by marine tucuxis (Sotalia guianensis) (Cetacea: Delphinidae) in Guanabara Bay, south-eastern Brazil. Journal of Marine Biology, 87: 201–205.
Ballance, L. T. (1992). Habitat use patterns and ranges of the bottlenose dolphin in the Gulf of California, Mexico. Marine Mammal Science, 8(3): 262-274.
Barros, N. B. & Teixeira, R. L. (1994). Incidental catch of marine tucuxi, Sotalia fluviatilis, in Alagoas, Northeastern, Brazil. Report of International Whaling Commission, (Special Issue 15): 265-268.
Bearzi, G., Politi, E., Di Sciara, G.N. (1999). Diurnal behavior of free-ranging bottlenose dolphins in the Kvarneric (Northern Adriatic sea). Marine Mammal Science, 15(4): 1065 – 1097.
Borobia, M. & Barros, N.B. (1989). Notes on the diet of marine Sotalia fluviatilis. Marine Mammal Science, 5 (4): 395-399. CEPENE, 197p.
Borobia, M., Siciliano, S., Lodi, L., Hoek, W. (1991). Distribution of the South American dolphin Sotalia fluviatilis. Canadian Journal of Zoology, 69: 1025-1039.
Caballero, S., Trujillo, F., Vianna, J.A., Barrios-Garrido, H., Montiel, M.G., Beltrán-Pedreros, S., Marmontel, M., Santos, M.C., Rossi-Santos, M., Santos, F.R., Baker, C.S. (2007). Taxonomic status of the genus Sotalia: species level ranking for “tucuxi” (Sotalia fluviatilis) and “costero” (Sotalia guianensis) dolphins. Marine Mammal Science, 23(2): 358-386.
Campos, A. E. S. P., da Rocha, J. C. Á. G., Collaço, B. J. R. , Collaço, K. de M. S. L. (2007). Aspectos do Comportamento de Forrageio do Boto – cinza Sotalia Guianensis Van Bénéden, 1864 (Cetácea, Delphinidae), no Porto de Maceió-AL. Anais do VIII Congresso de Ecologia do Brasil. Resumo Estendido. Caxambu – Minas Gerais.
Carr, T. & Bond, R. K. (2000). Tucuxi (Sotalia fluviatilis) Occurs in Nicaragua, 800 Km North Of Its Previously Known Range. Marine Mammal Science, 16(2): 447-452.
Constantine R., Brunton D.H., Dennis T., 2004. Dolphin-watching tour boats change bottlenose dolphin (Tursiops truncatus) behaviour. Biological Conservation. 117:299–307.

88
Cremer, M. J. (2007). Ecologia e conservação de populações simpátricas de pequenos cetáceos em ambiente estuarino no Sul do Brasil. Tese de Doutorado. Universidade Federal do Paraná. Curitiba.
Cremer, M. J., Hardt, F. A. S., Tonello Júnior, A. J., Simões-Lopes, P. C., Pires, J. S. R. (2004). Core areas changes in Sotalia guianensis (Cetacea, Delphinidae) population in Babitonga Bay, Santa Catarina. Revista Univille, 9: 12-16.
Cunha, H.A., Silva, V.M.F., Lailson-Brito Jr., J., Santos, M.C.O., Flores, P.A.C., Martin, A.R., Azevedo, A.F., Fragoso, A.B.L., Zanelatto, R.C., Solé-Cava, A.M. (2005). Riverine and marine ecotypes of Sotalia dolphins are different species. Marine Biology, 148: 449-457.
Daura-Jorge, F. G., Rossi-Santos, M. R., Wedekin, L. L., Simões-Lopes, P. C. (2007). Behavioral patterns and movement intensity of Sotalia guianensis (P.J.Van Bénéden) (Cetecea, Delphinidae) in two different areas on the Brazilian coast. Revista Brasileira de Zoologia, 24 (2): 265-270.
Daura-Jorge, F. G., Wedekin, Piacentini, V. de Q., Simões-Lopes, P. C. (2005). Seasonal and daily patterns of group size, cohesion and activity of the estuarine dolphin, Sotalia guianensis (P. J. van Benédén) (Cetacea, Delphinidae), in southern Brazil. Revista Brasileira de Zoologia, 22 (4): 1014 – 1021.
Domit, C. (2006). Comportamento de pesca do boto-cinza na região do Complexo Estuarino-Lagunar de Cananéia/Paranaguá, Brasil. Dissertação de Mestrado (Zoologia). Universidade Federal do Paraná. Curitiba, Paraná.
Flores P. A. C. & Bazzalo M. (2004). Home ranges and movement patterns of the marine tucuxi dolphin, Sotalia fluviatilis, in Baía Norte, southern Brazil. Lajam - Latin American Journal of Aquatic Mammals, 3(1): 37-52.
Geise, L. (2006). The correct use of the vernacular names of South American dolphins genus Sotalia Gray (Mammalia, Cetacea, Dephinedae). Revista Brasileira de Zoologia, 23(4): 1275-1276.
Gelatt, T. S., Sniiff, D. B., Estes J. A. (2002). Activity patterns and time-budget of the declining sea otter population at Amchitka island, Alaska. Journal of Wild Life Management, 66(1): 29 – 39.
Grigg, E. & Markowitz, H. (1997). Habitat use by bottlenose dolphins (Tursiops truncatus) at Turneffe Atoll, Belize. Aquatic Mammals, v. 23 (3):163-170.
Hanson, M.T. & Defran, R.H. (1993). The behaviour and feeding ecology of the Pacific coast bottlenose dolphin, Tursiops truncatus. Aquatic Mammals, 19:127–142
Harzen, S. (1998). Habitat use by the bottlenose (Tursiops truncatus) in the Sado estuary, Portugal. Aquatic Mammals, 24(3) 117-128.

89
Hastie, G. D., Wilson, B., Wilson L. J., Parsons K. M., Thompson P. M. (2004). Functional mechanisms underlying cetacean distribution patterns: hotspots for bottlenose dolphins are linked to foraging. Marine Biology, 144: 397-403.
Hetzel, B. & Lodi, L. (1993). Baleias, botos e golfinhos. Guia de identificação para o Brasil. Ed. Nova Fronteira. Rio de Janeiro. 280 p.
Hui, C. (1979). Undersea topography and distribution of dolphins of the genus Delphinus in the Southern California Bight. Journal of Mammalogy, 60(3): 521-527.
INPH (1993). Parecer técnico sobre dragagens e despejos dos dragados na área portuária de Maceió. Rio de janeiro.
Karczmarski, L., Cockcroft, V. G. (1999). Daylight behaviour of Humpback dolphins Sousa chinensis in Alagoa Bay, South Africa. International Journal of Mammalian Biology, 64: 19-29.
Maze, K. S. & Würsig, B. (1999). Bottle nose dolphin of San Luis Pass, Texas: Occurrence patterns, site-fidelity, and habitat use. Aquatic Mammals, 25(2): 91 – 103.
Monteiro-Filho, E.L.A., Monteiro, L.R., e Reis, S.F. (2002). Skull shape and divergence in dolphins of the genus Sotalia: a tridimensional morphometric analysis. Journal of Mammalogy, 83(1): 125-134.
Pereira, M.G., Bazzalo, M., Flores, P.A.C. (2007). Reações comportamentais de Sotalia guianensis (Cetacea, Delphinidae) durante encontros com embarcações na Baía Norte de Santa Catarina. Revista Brasileira de Zoociências, 9 (2): 123-135.
Rosas, F.C.W. (2000). Interações com a pesca, mortalidade, idade, reprodução e crescimento de Sotalia guianensis e Pontoporia blainvillei (Cetacea, Delphinidae e Pontoporiidae) no litoral sul do estado de São Paulo e litoral do estado do Paraná, Brasil. Tese de doutorado não-publicada, Universidade Federal do Paraná, Curitiba.
Rossi-Santos, M. R. (2006). Ecologia comportamental do boto cinza, Sotalia guianensis (Van Bénedén, 1874) (Cetacea: Delphinidae) na região extremo sul do Estado da Bahia, Nordeste do Brasil. Dissertação de Mestrado. UFPR.
Saayman, G.S., Tayler, C.K., Bower, D. (1972). Diurnal activity cycles in captive and free-ranging Indian Ocean bottlenose dolphins (Tursiops aduncus ehrenburg). Behaviour, 44(3/4): 213-233.
Shane, S. H., Wells, R. S., Würsig, B. (1986). Ecology, behavior and social organization of the bottlenose dolphin: a review. Marine Mammal Science, 2(1): 34-63.
Silva, F.J.L. (2001). Descrição de comportamento, sazonalidade de número de indivíduos e reprodução de golfinho rotador Stenella longirostris em Fernando de Noronha. Tese de Doutorado. Universidade Federal do Rio Grande do Norte. Natal.

90
da Silva, V.M.F. & Best, R.C. (1996). Sotalia fluviatilis, Mammalian Species. American Society of Mammalogists, 527:L7.
Simão, S. M. & Poletto, F. R. (2002). Áreas preferenciais de pesca e dieta do ecótipo marinho do boto cinza (Sotalia fluviatilis) na Baía de Sepetiba, Rio de Janeiro. Floresta e ambiente, 9 (1): 18-25.
Valle, A. L. (2006). Eficiência e uso de hábitat para a captura de presas por golfinhos Sotalia Fluviatilis: Comparação entre adultos e juvenis. Acta Biológica Leopondensia, 28 (1): 55-60.


















