Uso de marcadores citogenéticos clássicos e...
72
INSTITUTO NACIONAL DE PESQUISAS DA AMAZONIA Programa de Pós Graduação em Genética, Conservação e Biologia Evolutiva NATÁLIA DAYANE MOURA CARVALHO Manaus - AM 2011 Uso de marcadores citogenéticos clássicos e moleculares para a caracterização de Synbranchus spp. (Synbranchiformes) da Amazônia central
-
Upload
truongdiep -
Category
Documents
-
view
215 -
download
0
Transcript of Uso de marcadores citogenéticos clássicos e...

INSTITUTO NACIONAL DE PESQUISAS DA AMAZONIA
Programa de Pós Graduação em Genética, Conservação e Biologia
Evolutiva
NATÁLIA DAYANE MOURA CARVALHO
Manaus - AM
2011
Uso de marcadores citogenéticos clássicos e moleculares para a caracterização de Synbranchus spp. (Synbranchiformes) da
Amazônia central

ii
NATÁLIA DAYANE MOURA CARVALHO
Orientadora: Eliana Feldberg, Dra.
Co-orientadora: Maria Claudia Gross, Dra.
Manaus – AM
2011
Uso de marcadores citogenéticos clássicos e moleculares para a caracterização de Synbranchus spp. (Synbranchiformes) da
Amazônia central
Dissertação apresentada ao Instituto Nacional de Pesquisas da Amazônia como parte dos requisitos para obtenção do título de Mestre em Genética, Conservação e Biologia Evolutiva.

iii
FICHA CATALOGRÁFICA
Sinopse:
São apresentados dados cromossômicos de quatro espécies de Synbranchus: S. madeirae, S. cf. lampreia, Synbranchus sp.1 e Synbranchus sp.2, coletadas em simpatria e sintopia na região de confluência dos rios Negro e Solimões. As espécies foram estudadas mediante análise de citogenética clássica (coloração convencional, detecção de heterocromatina e regiões organizadoras de nucléolo) e molecular (hibridização fluorescente in situ com sondas de DNAr 18S, DNAr 5S e sequências teloméricas) e foram comparadas com as espécies descritas na literatura. Foram também discutidas as possíveis tendências evolutivas para este grupo de peixes.
Palavras-chave: Mussum, Variabilidade cromossômica, Heterocromatina constitutiva, Região organizadora de nucléolo, FISH, Amazônia
C331 Carvalho, Natalia Dayane Moura
Uso de marcadores citogenéticos clássicos e moleculares para a caracterização de Synbranchus spp. (Synbranchiformes) da Amazônia Central / Natália Dayane Moura Carvalho. --- Manaus : [s.n.], 2011.
xvi, 57 f. : il. color.
Dissertação (mestrado) -- INPA, Manaus, 2011
Orientador : Eliana Feldberg
Co-orientador : Maria Claudia Gross
Área de concentração : Genética, Conservação e Biologia Evolutiva
1. Mussum (peixe) – Amazônia. 2. Variabilidade cromossômica. 3.Região organizadora de nucléolo. 4. FISH (técnica). I. Título.
CDD 19. ed. 597.10415

iv
Dedico esta dissertação aos meus pais
Nilce e Maurício, aos meus irmãos
Maurício, Márcio e Júnior, à minha
cunhada Paula, aos meus pequenos
sobrinhos Maurício Vinícius e Paula Sofia
e ao meu noivo Rogério Neves.

v
A realização deste trabalho foi possível devido:
Ao Programa de Pós-graduação em Genética, Conservação e Biologia Evolutiva,
do INPA.
Ao laboratório de Genética Animal do INPA, Coordenação de Pesquisas em
Biologia Aquática (CPBA), onde este trabalho foi desenvolvido, com financiamento
proporcionado pelos projetos: “Caracterização genética (cromossomos, proteínas e
DNA) de vertebrados amazônicos” (INPA/MCT) e “Citotaxonomia de Arraias de água
doce da Amazônia central: conhecer para conservar” (PIPT/FAPEAM).
À Fundação de Amparo de Pesquisas do Estado do Amazonas (FAPEAM) pela
concessão da bolsa de estudo durante a realização deste trabalho.

Agradecimentos
Agradeço a TODOS que contribuíram de alguma forma para a realização deste trabalho.
Em especial:
A Deus, pois, sem sua ajuda, nada teria sido possível.
À Dra. Eliana Feldberg pela excelente orientação, amizade, conselhos, ensinamentos,
confiança e principalmente por ser essa profissional esplêndida e admirável. Obrigada
por ter me acolhido em seu laboratório e por ser tão especial na minha vida,
simplesmente a minha mãe científica.
À Dra. Maria Claudia Gross pela primorosa co-orientação, apoio, incentivo, sugestões e
principalmente pelo auxílio no laboratório. Sem a sua ajuda este trabalho não teria sido
realizado. Obrigada por tudo e principalmente pela confiança depositada em mim.
Ao Dr. Jorge Porto pela amizade e contribuição a este trabalho.
Ao Dr. Jansen Zuanon pela identificação das espécies analisadas.
Aos colegas do laboratório de Genética Animal: Carlos, Leandra, Leila, Heidi,
Thatianna, Brenda, Carlos Valentin, Denise e Érica por todos os momentos que
passamos juntos, pelos debates, sugestões, críticas científicas e às festinhas no
laboratório. Em especial, as minhas amigas Heidi Luz e Thatianna de Lira pela
companhia em nossos passeios (grandes aventuras), conversas, discussões científicas, e
principalmente por ter me aguentado nesse dois anos de mestrado. Obrigada, vocês já
fazem parte da minha vida.
Aos amigos do curso de mestrado – GCBEv/ 2009: Heidi, Alessandro, Carolina,
Luciana, Diana, Ramon, Jaqueliny Zocca, Jaqueline Fortuna, Tatiana e Gilson pelos
bons momentos extra-laboratório.
Aos meus pais Maurício e Nilce que são a razão da minha vida, para mim motivo de
orgulho. Obrigada por tudo e principalmente por acreditarem em mim. Tudo que puder
fazer por vocês farei, por que simplesmente amo vocês.
Aos meus irmãos Maurício, Márcio e Junior e minha cunha Paula pelo apoio
incondicional, amor e amizade.

vii
Ao meu pequeno príncipe Maurício Vinicius e à minha pequena princesa e afilhada
Paula Sofia por iluminar minha vida.
Ao meu noivo Rogério de Oliveira Neves pelo amor, carinho, paciência, atenção e por
mostrar-me o caminho maravilhoso de um amor e amizade, transmitindo sempre de
forma entusiasmada, clara e objetiva.

viii
Resumo
A família Synbranchidae pertence à ordem Synbranchiformes e é composta por quatro gêneros: Macrotrema, Ophisternon, Monopterus e Synbranchus. De acordo com a classificação taxonômica vigente, Synbranchus compreende três espécies válidas: S. marmoratus Bloch, 1795, S. madeirae Rosen & Rumney, 1972 e S. lampreia Favorito, Zanata & Assumpção, 2005. Contudo, estudos citogenéticos têm evidenciado uma variação no número diploide de exemplares identificados como “S. marmoratus”, coletados nas regiões sul, sudeste e centro-oeste do Brasil, os quais apresentam 42, 44 e 46 cromossomos e diversas fórmulas cariotípicas. Para as outras duas espécies válidas nenhuma característica citogenética havia sido publicada até o momento. Diante deste contexto, foram realizadas análises citogenéticas clássicas (coloração convencional, Banda C e RON) e moleculares (FISH com sondas de DNAr 18S, 5S e sequências teloméricas) em 14 indivíduos de S. madeirae, oito de S. cf. lampreia, um indivíduo de Synbranchus sp.1 e um de Synbranchus sp. 2, coletados em simpatria e sintopia no lago Catalão, situado na confluência dos rios Negro e Solimões. Para S. madeirae foi encontrado 2n=46 cromossomos, sendo 6m+2st+38a, NF=54, S. cf. lampreia 2n=44 cromossomos, sendo 6m+2st+36a, NF=52, Synbranchus sp.1 2n=42 cromossomos, sendo 6m+4sm+32a, NF=52 e Synbranchus sp.2 2n=42 cromossomos, sendo 6m+2sm+4st+30a, NF=54. A heterocromatina constitutiva apresentou-se distribuída nas regiões centroméricas, intersticiais, terminais, proximais e distais da maioria dos cromossomos para as quatro espécies de Synbranchus. A região organizadora de nucléolo (RON), apresentou-se como RON simples para S. cf. lampreia e múltipla para S. madeirae, Synbranchus sp.1 e Synbranchus sp. 2, com variação interespecífica quanto à sua distribuição, evidenciada tanto por impregnação com AgNO3 como pela hibridização de sondas DNAr 18S. A FISH com sonda DNAr 5S evidenciou uma marcação intersticial nos braços longos do par 7 de S. cf. lampreia e S. madeirae e do par 10 em Synbranchus sp.1. A sonda telomérica evidenciou marcações nos telômeros de todos os cromossomos além de marcações em regiões intersticiais em S. madeirae, S. cf. lampreia e Synbranchus sp.1. O somatório destes dados indica a ocorrência de uma variabilidade cariotípica interespecífica no gênero Synbranchus, incluindo heterocromatinização ou adição de heterocromatina, presença de rearranjos robertsonianos e não-robertsonianos na evolução cromossômica do grupo. Assim, acreditamos que a tendência de evolução cromossômica mais provável para este grupo seja a partir de um ancestral com 2n=48 cromossomos com a redução do número diploide.

ix
Abstract
Synbranchidae belongs to the Synbranchiformes order and it is composed by four genus: Macrotema, Ophisternon, Monopterus and Synbranchus. According with the present taxonomic arrangement, Synbranchus comprises three valid species: S. marmoratus Bloch, 1795, S. madeirae Rosen & Rumney, 1972, and S. lampreia Favorito, Zanata & Assumpçao, 2005. However, cytogenetic studies has made evident a variance in the diploid number of specimens of S. marmoratus collected in the south, south-east and center-west of Brasil, presenting 42, 44 and 46 chromosomes and diverse karyotypic formulas. Until nowadays, none cytogenetic characteristics have been registered for the two other valid species. Thereafter this contexture, there have been realized classic cytogenetic analysis (conventional coloration, C Band and NOR) and molecular as well (FISH with rDNA 18S, 5S probers and telomeric sequences) in fourteen individuals of S. madeirae, eight of S. cf. lampreia, one individual of Synbranchus sp.1 and one of Synbranchus sp.2 collected in compatibility at the Catalão lake situated at the confluence of the Negro and Solimões rivers next to Manaus, Amazonas. It was found 2n=46 chromosomes for S. madeirae, being 6m+2st+38a, NF=54, S. cf. lampreia 2n=44 chromosomes, being 6m+2st+36a, NF=52, Synbranchus sp1 2n=42 chromosomes, being 6m+4sm+32a, NF =52 and Synbranchus sp2 2n=42 chromosomes, being 6m+2sm+4st+30a, NF=54. Constitutive heterochromatic presented distributed in the centromeric, interstitial, terminal and proximal and distal of chromosomes majority for the four Synbranchus species. It has been inferred by the nucleolus organizer region (NOR) that simple NOR for S. cf. lampreia and multiple for S. madeirae, Synbranchus sp.1 and Synbranchus sp.2, with interspecific variance as for its distribution, being proved by its impregnation with AgNOR, as much as by r DNA 18S hybridization probe. It has also been proved by FISH with rDNA 5S probe an interstitial marker on pair 7 long arms of S. cf. lampreia and S. madeirae and of pair 10 in Synbranchus sp1. A telomeric probe has evinced marks in all telomeric chromosomes beyond marks in interstitial regions in S. madeirae, S. cf. lampreia and Synbranchus sp.1. It was inferred by the amount of this data that the occurrence of an interspecific karyotypic variability on Synbranchus genus, including heterochromatinization or heterochromatin addition, appearance of Robertsonian and non-Robertsonian rearrangements in chromosomic evolution of this group. Meanwhile, we believe that the evolutive tendency most likely for this group is from an ancestor with 2n = 48 chromosomes with reduction of the diploid number.

x
Sumário
1 Introdução.......................................................................................................................1
1.1 Considerações gerais............................................................................................... 1
1.2 Biologia de Synbranchidae ..................................................................................... 1
1.3 Citogenética de Synbranchiformes ......................................................................... 3
1.5 Objetivos ................................................................................................................. 8
1.5.1 Geral ................................................................................................................ 8
1.5.2 Específicos ........................................................................................................ 8
2 Material e Métodos.........................................................................................................9
2.1 Material ................................................................................................................... 9
2.2 Métodos ................................................................................................................ 12
2.2.1 Indução de mitoses........................................................................................... 12
2.2.2 Obtenção de cromossomos mitóticos ................................................................. 12
2.2.3 Detecção das regiões organizadoras de nucléolo – RONs................................. 13
2.2.4 Detecção da heterocromatina constitutiva (Banda C) ........................................... 13
2.2.5 Extração de DNA total....................................................................................... 14
2.2.6 Isolamento de sequências repetitivas por PCR (Polymerase Chain Reaction) e marcação por nick translation (Bionick labeling system-Invitrogen) ............................. 15
2.2.7 Hibridização in situ por fluorescência - FISH...................................................... 16
2.2.8 Análise cariotípica ............................................................................................. 18
2.2.9 Análise comparativa das medidas cromossômicas entre as espécies de Synbranchus ....................................................................................................................18
3 Resultados.....................................................................................................................20
3.1 Synbranchus madeirae.......................................................................................... 20
3.2 Synbranchus cf. lampreia ..................................................................................... 25
3.3 Synbranchus sp. 1 ................................................................................................. 29
3.4 Synbranchus sp.2 ................................................................................................. 32
3.5 Medidas cromossômicas ....................................................................................... 36

xi
4 Discussão......................................................................................................................37
5 Conclusões....................................................................................................................45
6 Perspectivas ..................................................................................................................46
7 Referências Bibliográficas............................................................................................47

xii
Lista de figuras
Figura 1 Exemplares de Synbranchus madeirae (a, b), S. cf. lampreia (c, d),
Synbranchus sp.1 (e, f) e Synbranchus sp.2 (g, h). Formato da cabeça (a, c, e, g) e o
padrão de manchas ao longo do corpo (b, d, f, h)............................................................11
Figura 2 Cariótipos de Synbranchus madeirae: a) em coloração convencional Giemsa;
b) regiões de heterocromatina constitutiva evidenciada por meio da técnica de banda C;
c) em destaque os pares nucleolares impregnados por AgNO3. m=Metacêntrico,
st=Subtelocêntrico, a=Acrocêntrico. Barra igual a 10µm...............................................22
Figura 3 Cariótipos de Synbranchus madeirae: a-b) localização física cromossômica do
DNAr 18S (amarelo); c) localização física cromossômica do DNAr 5S (amarelo); d)
Localização física cromossômica da sequência telomérica (amarelo). m=Metacêntrico,
st=Subtelocêntrico, a=Acrocêntrico. Barra igual a 20µm...............................................24
Figura 4 Cariótipos de Synbranchus cf. lampreia: a) em coloração convencional
Giemsa; b) regiões de heterocromatina constitutiva evidenciada por meio da técnica de
banda C; c) em destaque o par nucleolar impregnado por AgNO3. m=Metacêntrico,
st=Subtelocêntrico, a=Acrocêntrico. Barra igual a 10 µm..............................................27
Figura 5 Cariótipos de Synbranchus cf. lampreia: a) Localização física cromossômica
do DNAr 18S (amarelo); b) Localização física cromossômica do DNAr 5S (amarelo); c)
Localização física cromossômica da sequência telomérica (amarelo). m=Metacêntrico,
st=Subtelocêntrico, a=Acrocêntrico. Barras iguais a 20µm............................................28
Figura 6 Cariótipos de Synbranchus sp. 1: a) em coloração convencional Giemsa; b)
regiões de heterocromatina constitutiva evidenciada por meio da técnica de banda C; c)
em destaque os pares nucleolares impregnados por AgNO3. m=Metacêntrico,
sm=Submetacêntrico, a=Acrocêntrico. Barra igual a 10µm...........................................30
Figura 7 Cariótipos de Synbranchus sp.1: a) Localização física cromossômica do DNAr
18S (amarelo); b) em destaque os pares portadores dos sítios de DNAr 18S; c)
Localização física cromossômica do DNAr 5S (amarelo); d) Localização física
cromossômica da sequência telomérica (amarelo). m=Metacêntrico,
sm=Submetacêntrico, a=Acrocêntrico. Barra igual a 20µm...........................................31

xiii
Figura 8 Cariótipos de Synbranchus sp. 2: a) em coloração convencional Giemsa; b)
regiões de heterocromatina constitutiva evidenciada por meio da técnica de banda C; c)
em destaque o par nucleolar impregnado por AgNO3. m=Metacêntrico,
sm=Submetacêntrico, st=Subtelocêntrico, a=Acrocêntrico. Barra igual a 10 µm..........33
Figura 9 Cariótipo de Synbranchus sp. 2: a) Localização física cromossômica do DNAr
18S (amarelo). m=Metacêntrico, sm=Submetacêntrico, st=Subtelocêntrico,
a=Acrocêntrico. Barra igual a 20µm...............................................................................34
Figura 10 Idiograma comparativo de (a) Synbranchus madeirae; (b) Synbranchus cf.
lampreia; (c) Synbranchus sp.1; (d) Synbranchus sp.2; em preto, heterocromatina
constitutiva; em amarelo, genes de DNAr 18S; em rosa, genes de DNAr 5S...............35
Figura 11 Análise comparativa das medidas cromossômicas entre as espécies
estudadas. Eixo X: número de classes (classe I: 2,53 a 4,40; classe II: 4,41 a 6,28; classe
III: 6,29 a 8,16; classe IV: 8,17 a 10,04; classe V: 10,05 a 12,87); eixo Y: número de
cromossomos por classe de tamanho...............................................................................36

xiv
Lista de tabelas
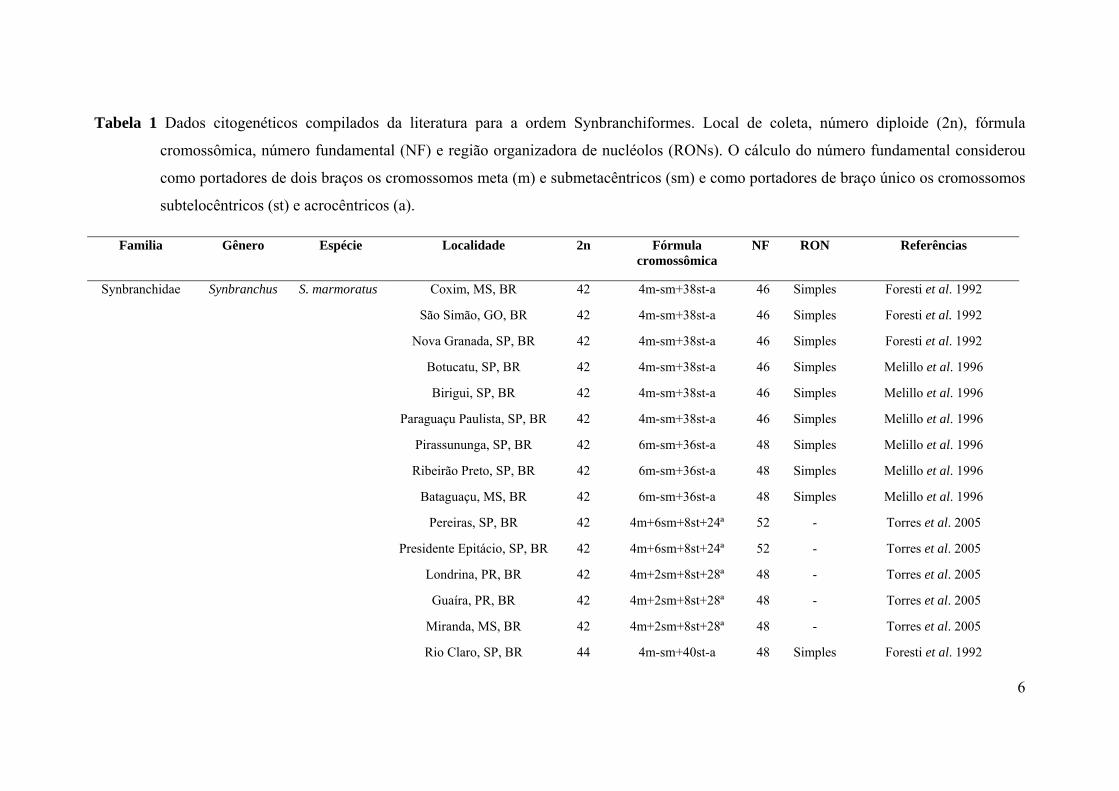
Tabela 1 Dados citogenéticos compilados da literatura para a ordem Synbranchiformes.
Local de coleta, número diploide (2n), fórmula cromossômica, número fundamental
(NF) e região organizadora de nucléolos (RONs). O cálculo do número fundamental
considerou como portadores de dois braços os cromossomos metacêntricos (m) e
submetacêntricos (sm) e como portadores de braço único os cromossomos
subtelocêntricos (st) e acrocêntricos (a)..........................................................................06
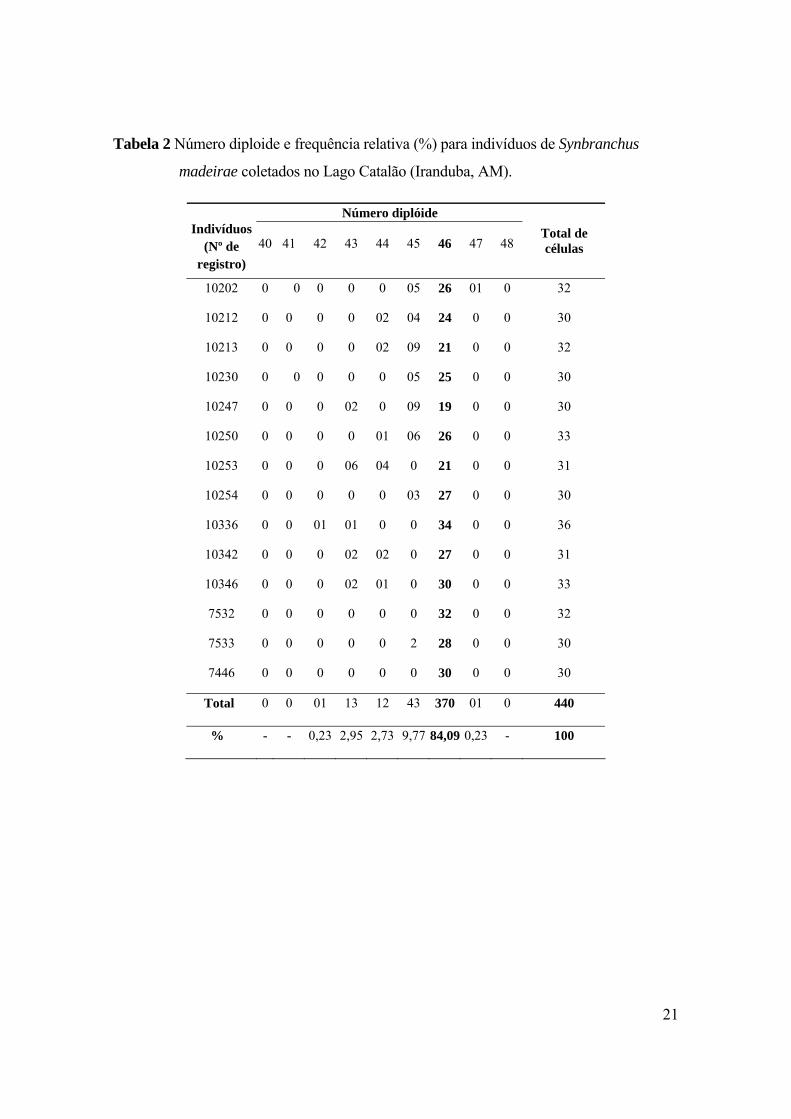
Tabela 2 Número diploide e frequência relativa (%) para indivíduos de Synbranchus
madeirae coletados no Lago Catalão (Iranduba, AM)..........................................................21
Tabela 3 Número diploide e frequência relativa (%), para indivíduos de Synbranchus cf.
lampreia coletados no Lago Catalão (Iranduba, AM)..........................................................26
Tabela 4 Número diploide e frequência relativa (%), para um indivíduo de Synbranchus
sp.1 coletado no Lago Catalão (Iranduba, AM)...................................................................29
Tabela 5 Número diploide, frequência relativa (%) e local de coleta, para um indivíduo de
Synbranchus sp.2 coletado no Lago Catalão (Iranduba, AM)............................................32

xv
Lista de abreviaturas
Banda C Técnica de detecção da heterocromatina constitutiva
DNAr DNA ribossomal
dNTP Desoxirribonucleotídeo
EDTA Ácido etilenodiaminotetracético
FISH Hibridização in situ fluorescente
FITC Fluoresceina isotiocianato
IGS Espaçador intergênico não transcrito
NTS Espaçador não transcrito
PBD Tampão fosfato dextrano
PCR Reação em cadeia da polimerase
Rex Retroelemento isolado primariamente de Xiphophorus
RNAr RNA ribossomal
RON Região organizadora de nucléolo
SDS Dodecil sulfato de sódio
SSC Solução salina de citrato padrão
Tampão C Tampão de acoplamento

1
1 Introdução
1.1 Considerações gerais A região Neotropical, que compreende a parte sul da América do Norte e as
Américas Central e do Sul, abriga a mais rica e diversa fauna de peixes de água doce do
mundo, sendo estimada a presença de 8.000 espécies, com 4.475 já descritas (Reis et al.
2003). Dentro desta região, a Bacia Amazônica se sobressai por sua diversidade
ictiofaunística, reunindo um grande número de espécies que pode chegar a 5.000, onde
são encontrados representantes das ordens Characiformes, Perciformes, Gymnotiformes,
Siluriformes, Myliobatiformes, Synbranchiformes, entre outras (Santos e Ferreira
1999).
1.2 Biologia de Synbranchidae A ordem Synbranchiformes compreende três famílias de peixes teleósteos:
Chaudhuriidae, Mastacembelidae e Synbranchidae (Nelson 2006). Esta ordem é
composta por 99 espécies válidas, as quais encontram-se amplamente distribuídas nas
regiões tropicais e subtropicais do Novo e Velho Mundo (Rosen e Greenwood 1976;
Schofield e Nico 2009).
As espécies de Synbranchidae apresentam corpo cilíndrico na região anterior e
comprimido na região posterior, são desprovidas de escamas e na idade adulta chegam a
medir cerca de 150 cm de comprimento. As nadadeiras dorsal e anal são rudimentares, a
caudal é curta, rudimentar ou ausente, e não possuem as pélvicas e peitorais.
Apresentam abertura branquial única localizada na região ventral da cabeça, cabeça
proeminente com olhos pequenos, narinas anteriores, boca grande provida de dentes
pequenos e não possuem bexiga natatória. A pele produz um muco que recobre todo o
corpo (Britski et al. 1999; Kullander 2003; Nelson 2006).
Com relação às características reprodutivas dos Synbranchidae, o
hermafroditismo protogínico diândrico é a forma de expressão sexual dominante
(Antoneli 2006). Ou seja, ocorre mudança de sexo nos indivíduos adultos, envolvendo a
degeneração do tecido gonadal feminino e o crescimento e maturação do tecido sexual
masculino, sobretudo apresentando dois tipos diferentes de machos. Os machos do
primeiro tipo são aqueles que se desenvolvem diretamente dos ovos e são denominados

2
machos primários, e os do segundo são machos resultantes da mudança de sexo a partir
de uma fêmea, por reversão do sexo e são chamados machos secundários (Lo Nostro e
Guerrero 1996; Antoneli 2002; Lo Nostro et al. 2003; Ravaglia e Maggese 2003). A
reversão do sexo de indivíduos identificados como S. marmoratus ocorre
predominantemente durante a estação seca e está relacionada à idade e ao comprimento
do indivíduo (Antoneli 2002; 2006). Deste modo, os machos secundários exibem
comprimento entre 25 e 54,9 cm, enquanto os machos primários podem ser encontrados
em qualquer faixa de comprimento (Lo Nostro e Guerrero 1996; Antoneli 2006).
Com relação à distribuição de Synbranchidae, são encontrados na África, Ásia,
Austrália, México e Américas Central e do Sul e compreende quatro gêneros:
Synbranchus, Ophisternon, Monopterus e Macrotrema. Na região Neotropical são
conhecidos apenas dois gêneros, Ophisternon e Synbranchus. Ophisternon é composto
por seis espécies, sendo que nesta região ocorrem duas: O. infernale Hubbs 1938 e O.
aenigmaticum Rosen e Greenwood 1976. Synbranchus compreende três espécies
válidas: S. marmoratus Bloch 1795, S. madeirae Rosen e Rumney 1972 e S. lampreia
Favorito, Zanata e Assumpção 2005 (Nelson 2006). Segundo Favorito-Amorim (1992)
Synbranchus marmoratus, considerada por vários autores como um complexo de
espécies, é representada por, pelo menos, sete táxons distintos, incluídas
provisoriamente dentro de Synbranchus. Deste modo, a autora sugere uma revisão
taxonômica mais ampla das espécies de Synbranchidae, para que a caracterização das
mesmas possa ser resolvida satisfatoriamente.
As espécies de Synbranchus alimentam-se de peixes, moluscos e pequenos
crustáceos e estão presentes em uma variedade de habitats, incluindo estuários,
pântanos, riachos, lagoas, charcos e corredeiras, mas têm preferência por águas com
baixa concentração de oxigênio e durante os períodos de seca sobrevivem enterradas na
lama úmida. Esta adaptabilidade a diferentes ambientes está relacionada aos tipos de
respiração, uma vez que além da típica respiração branquial estas espécies apresentam
respiração bucofaríngea e acessória, na qual as paredes da câmara branquial são
altamente vascularizadas e contribuem para as trocas gasosas, o que possibilita a
permanência fora da água por várias horas e a obtenção de oxigênio atmosférico durante
períodos de seca (Ghaham et al. 1995; Britski et al. 1999; Shibatta 2006). Segundo

3
Shibatta (2006) são peixes não migratórios, de hábitos noturnos, fecundação externa e
apresentam cuidado parental.
De acordo com a literatura, S. marmoratus tem sua distribuição desde o México
até o norte da Argentina; S. madeirae é restrita ao rio Madeira (Rosen e Rumney 1972)
e S. lampreia é endêmica da Ilha de Marajó, no estado do Pará (Favorito et al. 2005).
Entretanto, em levantamentos ictiofaunísticos recentes foi observado que S. marmoratus
parece ocorrer somente no Suriname e as outras duas espécies aparentemente possuem
uma ampla distribuição na Bacia Amazônica (T. Roberts comunicação pessoal), estando
a taxonomia deste grupo ainda não elucidada, o que enfatiza a necessidade de estudos
complementares.
1.3 Citogenética de Synbranchiformes A citotaxonomia pode ser definida como a identificação de táxons, utilizando
técnicas de citogenética, a qual permite individualizar espécies por meio de
características cromossômicas. A citotaxonomia está intimamente ligada à citogenética,
que é a ciência que estuda os cromossomos, seja de forma isolada ou direcionada ao
conjunto cromossômico de uma espécie, abrangendo aspectos morfológicos, funcionais
e evolutivos (Guerra, 1988).
Investigações cromossômicas em espécies de peixes têm contribuído para o
entendimento de sua sistemática, conhecimento da diversidade genética e da evolução,
principalmente quando combinadas com análises morfológicas e moleculares. Este tipo
de análise em peixes amazônicos já possibilitou a identificação de espécies crípticas
(Nakayama et al. 2001; Teixeira et al. 2006; Nakayama et al. 2008), detecção de
polimorfismos intra-específicos (Nakayama et al. 2000; Centofante et al. 2002;
Mesquita et al. 2008), detecção de cromossomos supranumerários (Feldberg et al.
2004), identificação de híbridos (Alves-Brinn et al. 2004), determinação de sistemas de
cromossomos sexuais (de Oliveira et al. 2007; 2008; Terencio et al. 2008) e evolução
cromossômica (Benzaquem et al. 2008).
Os estudos citogenéticos realizados com peixes têm como principais ferramentas
para a compreensão da estrutura organizacional dos cromossomos a determinação do
número diploide e a fórmula cariotípica. Outros marcadores eficazes na caracterização
de espécies são a localização das regiões organizadoras de nucléolo (RONs), as quais
estão envolvidas na transcrição de RNA ribossomal e o padrão de distribuição da

4
heterocromatina constitutiva no cariótipo, que indica como o genoma eucariótico está
organizado (Feldberg et al. 1993; Almeida-Toledo 1998; Margarido e Galetti Jr. 1999).
Ainda, a utilização de técnicas de manipulação do material genético associada à
detecção de sequências específicas de DNA fez surgir o que se conhece hoje por
citogenética molecular, sendo o fator fundamental para a expansão desses estudos a
utilização de marcações não-radioativas do DNA, com o emprego de corantes
fluorescentes. Esta técnica permite a detecção de sequências específicas em
cromossomos metafásicos e núcleos interfásicos através da hibridização de sequências
complementares, sendo utilizadas principalmente sequências repetitivas em tandem, tais
como genes de RNA ribossomais e sequências teloméricas (Artoni et al. 2000).
Citogeneticamente, a família Synbranchidae é a melhor conhecida da ordem
Synbranchiformes, com dados para três espécies dos gêneros Monopterus, Ophisternon
e Synbranchus. Monopterus albus apresenta 2n=24 cromossomos telocêntricos (Yu et
al. 1987) e Ophisternon bengalense 2n=46 cromossomos, sendo 6m+40st (Arkhipchuk
1999). Já os indivíduos identificados como S. marmoratus provenientes das regiões sul,
sudeste e centro-oeste do Brasil e da Argentina apresentam uma grande variabilidade,
com relação ao número diploide, que varia de 42 a 46 cromossomos e diversas fórmulas
cariotípicas (Tabela 1). Para a família Mastacembelidae, também pertencente à ordem
Synbranchiformes, duas espécies têm análises cromossômicas publicadas,
Mastacembelus armatus e M. aculeatus, ambas com 2n=48 cromossomos (Tabela 1).
Quanto ao padrão de distribuição da heterocromatina neste grupo de peixes, em
indivíduos identificados como S. marmoratus foram evidenciados blocos
heterocromáticos na região pericentromérica de quase todos os cromossomos, sendo que
em alguns foram também evidenciados na região intersticial (Foresti et al. 1992). Em
M. armatus foram localizados na região centromérica de quase todos os cromossomos
(Oliveira et al. 1997).
Com relação ao número de regiões organizadoras de nucléolo (RONs), todas as
populações identificadas como S. marmoratus e amostradas no Brasil, apresentaram
RONs simples, enquanto que três populações da Argentina apresentaram RONs
múltiplas (Tabela 1). Mastacembelus armatus também apresentou RON simples, que

5
segundo Oliveira et al. (1997) é uma característica considerada ancestral para a ordem
Synbranchiformes.
Análises citogenéticas moleculares até o momento não foram realizadas em
espécies da família Synbranchidae, bem como as espécies amazônicas ainda não
tiveram a sua composição cariotípica revelada. Deste modo, dada a ampla distribuição
de Synbranchus spp, a taxonomia incerta do grupo e a grande variabilidade cariotípica
já descrita para S. marmoratus de outras regiões brasileiras e argentina, a caracterização
cromossômica das espécies encontradas no lago Catalão, um ecótono de águas mistas,
situado na confluência dos rios Solimões (água branca) com o Negro (água preta), é de
fundamental importância para o conhecimento do grupo. Ainda, a utilização de
elementos repetitivos no mapeamento físico cromossômico em Synbranchus spp.
poderá proporcionar uma melhor visão do genoma das espécies, de como este está
organizado e como ocorreu a sua evolução.
6
Tabela 1 Dados citogenéticos compilados da literatura para a ordem Synbranchiformes. Local de coleta, número diploide (2n), fórmula
cromossômica, número fundamental (NF) e região organizadora de nucléolos (RONs). O cálculo do número fundamental considerou
como portadores de dois braços os cromossomos meta (m) e submetacêntricos (sm) e como portadores de braço único os cromossomos
subtelocêntricos (st) e acrocêntricos (a).
Familia Gênero Espécie Localidade 2n Fórmula cromossômica
NF RON Referências
Synbranchidae Synbranchus S. marmoratus Coxim, MS, BR 42 4m-sm+38st-a 46 Simples Foresti et al. 1992
São Simão, GO, BR 42 4m-sm+38st-a 46 Simples Foresti et al. 1992
Nova Granada, SP, BR 42 4m-sm+38st-a 46 Simples Foresti et al. 1992
Botucatu, SP, BR 42 4m-sm+38st-a 46 Simples Melillo et al. 1996
Birigui, SP, BR 42 4m-sm+38st-a 46 Simples Melillo et al. 1996
Paraguaçu Paulista, SP, BR 42 4m-sm+38st-a 46 Simples Melillo et al. 1996
Pirassununga, SP, BR 42 6m-sm+36st-a 48 Simples Melillo et al. 1996
Ribeirão Preto, SP, BR 42 6m-sm+36st-a 48 Simples Melillo et al. 1996
Bataguaçu, MS, BR 42 6m-sm+36st-a 48 Simples Melillo et al. 1996
Pereiras, SP, BR 42 4m+6sm+8st+24ª 52 - Torres et al. 2005
Presidente Epitácio, SP, BR 42 4m+6sm+8st+24ª 52 - Torres et al. 2005
Londrina, PR, BR 42 4m+2sm+8st+28ª 48 - Torres et al. 2005
Guaíra, PR, BR 42 4m+2sm+8st+28ª 48 - Torres et al. 2005
Miranda, MS, BR 42 4m+2sm+8st+28ª 48 - Torres et al. 2005
Rio Claro, SP, BR 44 4m-sm+40st-a 48 Simples Foresti et al. 1992

7
Pentecostes, CE, BR 44 4m-sm+40st-a 48 Simples Foresti et al. 1992
Botucatu, SP, BR 44 4m-sm+40st-a 48 Simples Melillo et al. 1996
Birigui, SP, BR 44 4m-sm+40st-a 48 Simples Melillo et al. 1996
Bataguaçu, MS, BR 44 4m-sm+40st-a 48 Simples Melillo et al. 1996
Ituzaingo, Corrientes, AR 44 4m-sm+40st-a 48 Múltiplas Sanchez e Fenocchio 1996
Reconquista, Santa Fé, AR 44 4m-sm+40st-a 48 Múltiplas Sanchez e Fenocchio 1996
Garabato, Santa Fé, AR 44 4m-sm+40st-a 48 Múltiplas Sanchez e Fenocchio 1996
Pirassununga, SP, BR 44 4m+2sm+8st+30ª 50 - Torres et al. 2005
Pirassununga, SP, BR 46 4m-sm+42st-a 50 Simples Melillo et al. 1996
Ribeirão Preto, SP, BR 46 4m-sm+42st-a 50 Simples Melillo et al. 1996
Bandeirantes, PR, BR 46 4m+2sm+8st+32ª 52 - Torres et al. 2005
Miranda, MS, BR 46 6m+2sm+6st+32ª 54 - Torres et al. 2005
Ophisternon O. bengalense - 46 6m+40st 52 - Arkhipchuk 1999
Monopterus M. albus Wuhan (China) 24 24t 24 Simples,
múltipla-
Ji et al. 2003
Mastacembelidae Mastacembelus M. armatus - 48 10m+6sm+4st+28a 68 Simples Oliveira et al. 1997
M. aculeatus China 48 - - Liu et al. 2002
Sinobdella S. sinensis - 48 15m+4sm+3st+26a 70 - Yajuan e Qixing 1991
Macrognathus M. aculeatum - 48 8m+2sm+38st,a 58 - Khuda-Bukhsh e Barat 1987
M. pancalus - 48 14m+12sm+14st+8a 88 - Khuda-Bukhsh e Barat 1987
48 16m+6sm+8st+18a 78 - Manna e Prasad 1977

8
1.5 Objetivos
1.5.1 Geral • Analisar as espécies de Synbranchus da região de confluência dos rios Negro e
Solimões, utilizando marcadores citogenéticos clássicos e moleculares.
1.5.2 Específicos • Determinar o cariótipo de indivíduos de Synbranchus spp., coletados na região
de confluência dos rios Negro e Solimões.
• Estabelecer os padrões de distribuição da heterocromatina constitutiva (Banda
C) e da região organizadora de nucléolo (RONs) em cromossomos mitóticos.
• Localizar as sequências de DNA ribossômico 5S e 18S e sequências teloméricas
em cromossomos mitóticos pela técnica de hibridização in situ fluorescente
(FISH).
• Verificar se os cariótipos observados correspondem às espécies válidas e se
existe variabilidade cromossômica inter e intraespecífica.
• Inferir possíveis tendências evolutivas para o gênero Synbranchus.

9
2 Material e Métodos
2.1 Material Foram efetuadas preparações cromossômicas de 96 indivíduos de Synbranchus,
coletados no lago Catalão (03º10’45” S; 59º54’25.4” W) situado na confluência dos rios
Solimões e Negro, Amazonas, Brasil, porém, apenas 21 apresentaram metáfases em
boas condições de análise. Destes, 14 indivíduos foram classificados como Synbranchus
madeirae, oito como Synbranchus cf. lampreia, um como Synbranchus sp.1 e outro
como Synbranchus sp.2 (Figura 1). Entretanto, somente foi possível identificar o sexo
de três indivíduos de Synbranchus madeirae, sendo 2 machos e uma fêmea. Todos os
exemplares foram identificados pelo pesquisador Dr. Jansen Zuanon (CPBA/INPA).
Para uma melhor caracterização das espécies estudadas foi feita uma breve
descrição morfológica das mesmas com base no formato da cabeça e padrão de
coloração.
Synbranchus madeirae
Esta espécie apresenta a região dorsal do corpo com coloração de fundo escuro
com manchas claras circulares bem definidas; a região ventral com coloração de fundo
marrom claro com manchas escuras. A cabeça bastante afilada, tanto lateralmente
quanto dorsalmente e apresenta uma faixa negra alongada abaixo dos olhos, iniciando-
se na extremidade anterior do focinho e estendendo-se até aproximadamente a
extremidade posterior do osso maxilar.
Synbranchus cf. lampreia
Esta espécie apresenta a região dorsal do corpo com coloração de fundo marrom
com manchas escuras, inclusive na cabeça, onde são menores; a região ventral com
coloração de fundo claro, com manchas escuras. A cabeça com saliência muscular
dorsal.
Synbranchus sp.1

10
Esta espécie apresenta a região dorsal do corpo com coloração de fundo cinza
escuro com ausência de manchas; na região ventral, coloração de fundo cinza claro. A
cabeça bastante afilada, tanto lateralmente quanto dorsalmente.
Synbranchus sp. 2
Esta espécie apresenta a região dorsal do corpo com coloração de fundo escuro
quase preto com ausência de manchas; a região ventral com coloração de fundo claro. A
cabeça com saliência muscular dorsal.
Os peixes foram capturados com o uso de redes e rapichés e transportados vivos
em caixas térmicas tipo isopor, para o Laboratório de Genética Animal do Instituto
Nacional de Pesquisas da Amazônia – INPA, Coordenação de Pesquisas em Biologia
Aquática ou para o laboratório flutuante do Lago Catalão, onde foram preparadas as
suspensões celulares. Para tanto, os peixes foram submetidos a uma dose letal do
anestésico eugenol (2%) e após terem sido retirados os tecidos de interesse, os mesmos
foram numerados, registrados, fixados em formol 10% por 24h, lavados em água
corrente e acondicionados em recipientes contendo álcool 70%, visando seu depósito na
Coleção de Peixes do INPA (Licença IBAMA: Número 10609-1 de 21/11/2007).

11
Figura 1 Exemplares de Synbranchus madeirae (a, b), S. cf. lampreia (c, d),
Synbranchus sp.1 (e, f) e Synbranchus sp.2 (g, h). Formato da cabeça (a, c, e, g) e o
padrão de manchas ao longo do corpo (b, d, f, h).

12
2.2 Métodos
2.2.1 Indução de mitoses Para se obter um maior número de células em metáfase foi utilizada a técnica de
indução de mitoses descrita por Oliveira et al. (1988), que consiste em preparar uma
solução de fermento biológico na seguinte proporção: 0,5 g de fermento, 0,5 g de açúcar
e 20 mL de água destilada. Em seguida, esta solução foi incubada em banho-maria ou
estufa a 40 °C por cerca de 20 minutos, e posteriormente injetada na região
intraperitoneal do animal, na proporção de 1 mL para cada 100 g de peso vivo. Os
peixes foram mantidos em aquários aerados por um período de 24 horas antes da coleta
das amostras para análise citogenética.
2.2.2 Obtenção de cromossomos mitóticos Após 24 horas da aplicação da solução de fermento, foi injetada,
intraperitonealmente uma solução aquosa de colchicina 0,0125% na proporção de 1 mL
para cada 100 g de peso vivo, por 45 a 60 minutos (Bertollo et al. 1978). Após esse
tempo, os peixes foram submetidos a uma dose letal do anestésico eugenol 2% e
procedeu-se a retirada da porção anterior do rim, que é o órgão hematopoiético. Este
tecido foi lavado em solução hipotônica de KCl a 0,075M e transferido para outro
recipiente de vidro contendo cerca de 10 mL da solução hipotônica de KCl, onde foi
dissociado com pinças de dissecação. A suspensão celular obtida foi colocada em estufa
a 37 °C por 20 minutos. Em seguida o material foi ressuspendido cuidadosamente com
o auxílio de uma seringa desprovida de agulha e transferido para um tubo de centrífuga,
utilizando-se uma pipeta Pasteur. Foram adicionadas 4 gotas de fixador Carnoy 3:1
(metanol: ácido acético), recém preparado e gelado e a amostra foi centrifugada por 10
minutos a 900 rpm, descartando-se o sobrenadante. Adicionou-se com cuidado 8 mL de
fixador Carnoy 3:1, ressuspendeu-se o material cuidadosamente com o auxílio de uma
pipeta Pasteur e esta lavagem foi repetida por mais duas vezes. Após a última
centrifugação e eliminação do sobrenadante foram adicionados 1,5 mL de fixador e o
material foi ressuspendido com cuidado. Esta suspensão celular foi guardada em tubos
eppendorf e mantida em freezer para posterior utilização.
Para a preparação das lâminas, as mesmas foram colocadas em solução
sulfocrômica por 24 horas. Após este período, foram retiradas, lavadas em água corrente

13
e destilada e armazenadas em álcool 100%. As lâminas foram imersas em água destilada
a 45 °C, em banho-maria. Após 5 minutos, foram retiradas da água e a suspensão
celular foi gotejada sobre três pontos diferentes da lâmina, que foi seca diretamente ao
ar. Em seguida, as amostras foram coradas com Giemsa 5% diluído em tampão fosfato
0,06M e pH 6,8, por 15 minutos, lavadas em água destilada e secas ao ar.
2.2.3 Detecção das regiões organizadoras de nucléolo – RONs Para a detecção das regiões organizadoras de nucléolo (RONs) foi utilizada a
técnica descrita por Howell e Black (1980), com modificações. Sobre as lâminas com a
suspensão foram adicionadas 2 a 3 gotas de uma solução coloidal de gelatina (2 g de
gelatina comercial sem sabor, dissolvida em 100 mL de água destilada, acrescida de
1mL de acido fórmico) e, sobre cada gota de gelatina, duas gotas de solução aquosa de
nitrato de Prata (AgNO3) a 50%, agitando-se levemente a lâmina, que foi coberta com
lamínula. A lâmina foi colocada em câmara úmida, em banho-maria a 60 °C, durante 3
a 8 minutos. Após esse tempo, quando a lâmina adquiriu uma coloração marrom
dourada, a mesma foi lavada em água destilada, permitindo que a lamínula fosse
retirada naturalmente pela própria água e seca ao ar.
2.2.4 Detecção da heterocromatina constitutiva (Banda C) Para a detecção da heterocromatina constitutiva foi utilizada a técnica descrita
por Sumner (1972), com algumas modificações. As lâminas com a suspensão celular
foram tratadas com HCl 0,2N a 45 °C, por 2 minutos, lavadas rapidamente em água
destilada à temperatura ambiente e secas ao ar. Após isso as lâminas foram incubadas
por cerca de 40 segundos em solução de hidróxido de bário a 5%, recém preparada e
filtrada a 42 °C. Posteriormente, a ação do hidróxido de bário foi interrompida
imergindo-se rapidamente a lâmina em solução de HCl 0,2N (temperatura ambiente),
lavada em água destilada e deixada secar ao ar. Após secas, as lâminas foram incubadas
em solução 2xSSC (cloreto de sódio 0,3M e citrato trisódico 0,03M, pH 6,8) em banho-
maria a 60 °C, por 15 minutos. Em seguida, foram lavadas várias vezes em água
destilada e secas ao ar. As lâminas foram coradas com solução de Giemsa (diluída a 5%
em tampão fosfato 0,06M e pH 6,8) durante 10 minutos, lavadas em água destilada e
secas ao ar.

14
2.2.5 Extração de DNA total A extração de DNA foi realizada a partir do tecido muscular, utilizando o
protocolo básico de Sambrook e Russel (2001) com algumas modificações. Foram
macerados aproximadamente 20 mg de tecido muscular e transferidos para um tubo de
volume de 1,5 mL. Em seguida foram adicionados 500 µL de tampão de lise (Tris-HCl
10 mM em pH 8,0, NaCl 0,3 M, EDTA 10 mM, SDS 1%) conforme Estoup et al.
(1993) e com o acréscimo de Uréia 4 M conforme Asahida et al. (1996). Posteriormente
foram acrescentados: 15 μL de proteinase K (10 mg/mL) e 6 μL de RNAse (10 mg/mL).
As amostras foram incubadas a 60 ºC por aproximadamente 2 horas para que o tecido
fosse totalmente digerido. Foram adicionados 100 µL de acetato de amônio 3M, 1
volume (600 µL) de fenol-clorofórmio (1:1) e agitado por inversão, por alguns minutos.
Após isso as amostras foram centrifugadas por 30 minutos a 14.000 rpm em
temperatura ambiente. O sobrenadante foi transferido para um novo tubo, adicionados
600 µL de clorofórmio e misturado, cuidadosamente, por alguns minutos, por inversão.
Em seguida, a mistura foi centrifugada por 20 minutos a 14.000 rpm. O sobrenadante
foi transferido para um novo tubo e foram acrescentados 100 µL de acetato de amônio
3M, 1 volume (600 µL) de isopropanol gelado e misturado gentilmente por inversão. O
precipitado foi deixado a -20 °C por cerca de 14 horas ou a -80 °C por 1 hora. O
material foi retirado do freezer e centrifugado por 30 minutos a 14.000 rpm,
descartando-se o sobrenadante. O pellet foi lavado com 1 mL de etanol 70% e
centrifugado por 20 minutos a 14.000 rpm. Novamente, o sobrenadante foi descartado,
o pellet foi seco em estufa a 55 °C, ressuspendido em 100 μl de TE 0,2X ou água milli-
Q e deixado eluindo por 14 horas. Para possibilitar a análise da quantidade e integridade
do material, o DNA extraído foi quantificado por comparação com marcador de DNA
bacteriófago Lambda, em eletroforese padrão (com tampão Tris-Borato-EDTA 0,5X e
corrida a 70 V por 40 minutos) em gel de agarose 0,8% e corado com GelRed Acid Gel
Stain (Biotium 1:500). A visualização e análise do DNA no gel foram feitas no
fotodocumentador Easy Doc 100 (BioAgency), o qual possui acoplado um
transluminador de luz ultravioleta (260 nM).

15
2.2.6 Isolamento de sequências repetitivas por PCR (Polymerase Chain Reaction) e
marcação por nick translation (Bionick labeling system-Invitrogen)
Sequências repetitivas já identificadas e caracterizadas como conservadas em
outras espécies de peixes, tais como 18S, 5S e teloméricas foram estudadas. Para a
obtenção das sondas de DNAr 5S, DNAr 18S e sequências teloméricas foi utilizado o
DNA genômico extraído do músculo de Synbranchus madeirae e Synbranchus cf.
lampreia. As sondas foram obtidas por amplificação por Reação em Cadeia da
Polimerase (PCR), conforme Saiki et al. (1988) utilizando os primers:
-DNAr 5S (A 5’-TACGCCCGATCTCGTCCGATC-3’ e B 5’-
CAGGCTGGTATGGCCGTAAGC-3’) (Komiya e Takemura 1979);
-DNAr 18S (IpF 5’CCGCTTTGGTGACTCTTGAT e IpR
5’CCGAGGACCTCACTAAACCA) (Gross et al. 2010b).
- Sequências teloméricas (TTAGGG)5 e (CCCTAA)5 (Ijdo et al. 1991).
Os ciclos de amplificação seguiram as seguintes etapas:
a) 5S: 5 minutos a 94 ºC (desnaturação); 2 ciclos de 1 minuto a 95 ºC, 30 segundos
a 61 ºC e 45 segundos a 72 ºC; 2 ciclos de 1 minuto a 95 ºC, 30 segundos a 59 ºC e 45
segundos a 72 ºC; 2 ciclos de 1 minuto a 95 ºC, 30 segundos a 57 ºC e 45 segundos a 72
ºC; 25 ciclos de 1 minuto a 95 ºC, 30 segundos a 61 ºC e 45 segundos a 72 ºC
(amplificação); 7 minutos a 72 ºC (extensão);
b) 18S: 2 minutos a 94 ºC (desnaturação); 35 ciclos de 1 minuto a 95 ºC, 30
segundos a 55 ºC, 1 minuto e 40 segundos a 72 ºC (amplificação); 7 minutos a 72 ºC
(extensão).
c) Telomérica: 10 minutos a 94 ºC (desnaturação); 30 segundos a 55 ºC; 1 minuto
a 72 ºC (amplificação); 30 ciclos de 1 minuto a 94 ºC, 30 segundos a 60 ºC e 1 minuto e
30 segundos a 72 ºC (amplificação); 5 minutos a 72 ºC (extensão).
As reações de PCR foram feitas em um termociclador Eppendorf -
Mastercycler Gradient, para volume final de 15 μL (1 μL de DNA genômico (100 ng);
1,5 μL de Tampão 10X com cloreto de magnésio (1,5 mM); 0,15 μL de Taq DNA

16
polimerase (5U/µl); 3,0 μL de dNTP (1 mM); 0,6 μL de cada primer (5 mM); água
milli-Q para completar o volume).
As sondas foram marcadas por biotina 14-dATP por nick translation, seguindo
protocolo do fabricante (Bionick labeling system-Invitrogen). Para tanto, em um tubo de
eppendorf de 1,5 mL, mantido no gelo, foi preparada uma solução contendo 1 μL de
Mix dNTP 10x; 1 μL de sonda de DNA (200 ng/ μL); 1 μL de Mix de enzima 10x; 6 μL
de água milli-Q, totalizando 9 μL, para cada lâmina que foi hibridizada. Esta solução foi
homogeneizada, centrifugada brevemente e incubada a 16 ºC por 45 minutos no
termociclador. Em seguida, para interromper a reação, foi adicionado 1 μL de stop
buffer contendo 1 μL de acetato de sódio 3M e após isso foi mantida em freezer para
posterior utilização.
2.2.7 Hibridização in situ por fluorescência - FISH
Foi utilizada a técnica de hibridização in situ por fluorescência (FISH) descrita
por Pinkel et al. (1986) com algumas modificações.
Tratamento das lâminas As lâminas foram lavadas em tampão PBS 1x por 5 minutos em temperatura
ambiente. As lâminas foram desidratadas em série alcoólica gelada (70%, 85% e 100%)
durante 5 minutos cada. Em seguida foram tratadas com 90 µl de RNase 10 µg/mL (5
µL de RNase 10 mg/mL e 975 µL de 2XSSC) por 1 hora em câmara úmida a 37 ºC. As
lâminas foram lavadas três vezes em 2xSSC durante 5 minutos cada. Após isso, as
lâminas foram lavadas em PBS 1x durante 5 minutos.
Fixação
As lâminas foram fixadas em formaldeído 1% em PBS 1X/50mM MgCl2 durante
10 minutos à temperatura ambiente. Posteriormente lavadas em PBS 1x por 5 minutos.
Após, as lâminas foram desidratadas em série alcoólica gelada (70%, 85% e 100%)
durante 5 minutos cada. Deixou-se secar ao ar. Em seguida foram desnaturadas em
formamida 70% em 2xSSC a 70 ºC por 5 minutos e novamente desidratadas em etanol
gelado 70%, 85% e 100% por 5 minutos cada. Deixou-se secar ao ar.

17
Solução de hibridização
Em um tubo eppendorf, foram adicionados 6 µl de sonda, 15 µl de formamida, 6
µl de sulfato de dextrano 50% e 3 µl de 20xSSC. A sonda foi desnaturada a 99 °C por
10 minutos e passada imediatamente ao gelo.
Hibridização
Foram colocados 30 µl de solução de hibridização sobre uma lamínula e a
lâmina foi invertida sobre a lamínula. As lâminas foram mantidas com o material
voltado para baixo em câmara úmida (H20 destilada) a 37 °C por cerca de 14 horas.
Lavagens
As lamínulas foram removidas das lâminas. Em seguida foram lavadas em
2xSSC a 72 °C por 5 minutos. Após, foram transferidas para solução de PBD 1x (leite
em pó desnatado; Triton X-100; 20xSSC; diluído em água destilada ajustando para pH
7,0) a temperatura ambiente.
Detecção Foram colocadas sobre cada lâmina 30 µL de avidina-FITC 0,07% em tampão C
(0,1 M de bicarbonato de sódio, pH 8,5 e 0,15 M de NaCl). Em seguida foram cobertas
com lamínula e deixadas por 30 minutos em câmara úmida com água destilada.
Posteriormente as lamínulas foram removidas e as lâminas foram lavadas três vezes em
solução de PBD 1x recém preparado a 45 ºC por 2 minutos cada. Após a lavagem, foi
colocada sobre cada lâmina 20 µL de anti-avidina biotina-conjugada 2,5% (1 µL de
anti-avidina estoque em 19 µL de PBD 1x), coberta com lamínula e deixada em câmara
úmida com água destilada a 37 °C por 10 minutos. Após, as lâminas foram lavadas três
vezes em solução de PBD 1x a 45 ºC por 2 minutos cada. Posteriormente, foi colocada
sobre cada lâmina 30 µL de avidina-FITC 0,07% em tampão C, coberta com lamínula e
deixada em câmara úmida com água destilada a 37 °C por 10 minutos.
Montagem das lâminas Adicionou-se a cada lâmina solução de iodeto de propídio diluído em antifade
Vector (20 µL de antifade e 0,4 µL de iodeto de propídio).

18
2.2.8 Análise cariotípica Após a análise e contagem dos cromossomos ao microscópio óptico foi
estabelecido o número diploide modal e suas frequências relativas para cada indivíduo.
As melhores metáfases obtidas com as técnicas clássicas (convencional, RON e banda
C) foram fotografadas, com objetiva de imersão em câmera digital Canon Power shot A
650 IS. As metáfases submetidas a técnicas moleculares (FISH) foram analisadas e
capturadas em um fotomicroscópio de epifluorescência Olympus Bx-51, em objetiva de
imersão.
Para a montagem dos cariótipos foi utilizado o programa Adobe Photoshop 7.0,
versão CS3, a partir de cromossomos metafásicos mitóticos, os quais foram recortados e
tentativamente emparelhados. Os cromossomos foram medidos, utilizando o programa
livre ImageJ, agrupados de acordo com a sua morfologia e colocados em ordem
decrescente de tamanho. A morfologia cromossômica foi determinada de acordo com os
critérios de relação de braços (RB=BM/Bm, onde BM = braço maior e Bm = braço
menor) segundo Levan et al. (1964). Os cromossomos foram separados em
metacêntricos (m), que apresentam RB=1,00 a 1,70, submetacêntricos (sm) RB=1,71 a
3,00, subtelocêntricos (st) RB=3,01 a 7,00 e acrocêntricos (a) RB > 7,00. O número
fundamental (NF) foi determinado de acordo com o número de braços cromossômicos,
considerando-se os metacêntricos, submetacêntricos e subtelocêntricos como tendo dois
braços e acrocêntricos como tendo apenas um braço.
2.2.9 Análise comparativa das medidas cromossômicas entre as espécies de Synbranchus
A partir dos dados obtidos com as medidas cromossômicas foi feita uma análise
comparativa entre as espécies estudadas. De cada espécie obteve-se a frequência do
comprimento relativo dos pares cromossômicos por classes de tamanho. Para
determinar o número ideal de classes, para a distribuição dos pares por classe de
tamanho relativo, foi utilizada a formula de Sturges (Fonseca e Martins 1982):
n= 1+ 3,32 * LogN, onde n= número de classes e N= lote haplóide (em número).
Depois de determinar o número ideal de classes, foi feita a subtração entre o
maior e o menor valor do comprimento relativo dos cromossomos, em µm, de cada
espécie. O valor encontrado foi dividido pelo número ideal de classes, e o resultado

19
encontrado corresponde ao intervalo de cada classe, onde: classe 1=2,53-4,40; classe
2=4,41-6,28; classe 3=6,29-8,16; classe 4=8,17-10,04; classe 5=10,05-12,87.

20
3 Resultados De um total de 24 indivíduos coletados, 14 foram identificados como
Synbranchus madeirae, oito como Synbranchus cf. lampreia, um foi nomeado de
Synbranchus sp.1 e um de Synbranchus sp. 2.
3.1 Synbranchus madeirae A Tabela 2 mostra o número dos exemplares, número diploide e total de células
analisadas. O número diploide modal determinado para esta espécie foi igual a 46
cromossomos, sendo 6m+2st+38a e o número fundamental (NF) foi igual a 54 (Figura
2a). Nenhum heteromorfismo relacionado à presença de cromossomos sexuais foi
observado.
A heterocromatina constitutiva foi observada nas regiões centromérica e
terminal da maioria dos cromossomos, mas também foram observados blocos
intersticiais e distais em alguns pares cromossômicos (Figura 2b; 4).
A região organizadora de nucléolo (RON), submetida à impregnação com nitrato
de prata, foi evidenciada com até cinco marcações, sendo considerada múltipla,
localizadas nas regiões terminais dos braços longos do par 2, dos braços curtos do par 6
e na região intersticial do braço longo de um dos homólogos do par 7. Porém, nem
sempre todas estiveram ativas (Quadro 1; Figura 2c).
A hibridização in situ fluorescente (FISH) com sonda de DNAr 18S também
evidenciou até cinco marcações, porém dois padrões de distribuição foram encontrados.
No primeiro padrão (10 indivíduos), um dos homólogos dos pares 1, 2, e 7 apresentou
marcações, assim como ambos os homólogos do par 6 e para um indivíduo foi
observado em ambos os homólogos dos pares 2 e 21 apresentaram sítios ribossomais
18S, bem como um dos homólogos dos pares 6 e 7. Entretanto, para o primeiro padrão e
para um individuo, apenas as marcações dos pares 6 e 7 foram coincidentes com as
marcações com nitrato de Prata. Do par 2 apenas um dos homólogos coincidiu com
AgRON e do par 1, um dos homólogos também apresentou uma marcação na região
terminal dos braços longos (Figura 3 a, b; 4; Quadro 1). A sonda DNAr 5S evidenciou
um sítio localizado na região intersticial dos braços longos do par 7, sendo sintênico e
adjacente a um dos sítios de DNAr 18S (Figura 3c, 4). A sonda telomérica evidenciou
sequências apenas nas regiões teloméricas de todos os cromossomos (Figura 3d).
21
Tabela 2 Número diploide e frequência relativa (%) para indivíduos de Synbranchus
madeirae coletados no Lago Catalão (Iranduba, AM).
Número diplóide Indivíduos
(Nº de registro)
40 41 42 43 44 45 46 47 48 Total de células
10202 0 0 0 0 0 05 26 01 0 32
10212 0 0 0 0 02 04 24 0 0 30
10213 0 0 0 0 02 09 21 0 0 32
10230 0 0 0 0 0 05 25 0 0 30
10247 0 0 0 02 0 09 19 0 0 30
10250 0 0 0 0 01 06 26 0 0 33
10253 0 0 0 06 04 0 21 0 0 31
10254 0 0 0 0 0 03 27 0 0 30
10336 0 0 01 01 0 0 34 0 0 36
10342 0 0 0 02 02 0 27 0 0 31
10346 0 0 0 02 01 0 30 0 0 33
7532 0 0 0 0 0 0 32 0 0 32
7533 0 0 0 0 0 2 28 0 0 30
7446 0 0 0 0 0 0 30 0 0 30
Total 0 0 01 13 12 43 370 01 0 440
% - - 0,23 2,95 2,73 9,77 84,09 0,23 - 100

22
Figura 2 Cariótipos de Synbranchus madeirae: a) em coloração convencional Giemsa;
b) regiões de heterocromatina constitutiva evidenciada por meio da técnica de
banda C; c) em destaque os pares nucleolares impregnados por AgNO3.
m=Metacêntrico, st=Subtelocêntrico, a=Acrocêntrico. Barra igual a 10µm.

23
Quadro 1 Padrões de RONs e DNAr 18S encontrados em indivíduos de S. madeirae,
coletados mo lago Catalão (Iranduba, AM).

24
Figura 3 Cariótipos de Synbranchus madeirae: a-b) localização física cromossômica do
DNAr 18S (amarelo); c) localização física cromossômica do DNAr 5S
(amarelo); d) Localização física cromossômica da sequência telomérica
(amarelo). m=Metacêntrico, st=Subtelocêntrico, a=Acrocêntrico. Barra igual
a 20µm.

25
3.2 Synbranchus cf. lampreia A Tabela 3 mostra o número dos exemplares, número diploide e total de células
analisadas. O número diploide modal determinado para esta espécie foi igual a 44
cromossomos, sendo 6m+2st+36a e o número fundamental (NF) foi igual a 52 (Figura
4a).
A heterocromatina constitutiva foi observada nas regiões proximal, terminal,
distal, centromérica e intersticial da maioria dos cromossomos (Figura 4b).
A região organizadora de nucléolo (RON) submetida à impregnação com nitrato
de prata foi evidenciada como RON simples, localizada na região terminal dos braços
longos do par 19, com um bloco de heterocromatina distal, adjacente à RON (Figura
4c).
A hibridização in situ fluorescente (FISH), com sonda de DNAr 18S, evidenciou
um par cromossômico portador deste sítio, coincidente com a marcação por nitrato de
Prata (Figura 5a). A sonda de DNAr 5S evidenciou um sítio localizado na região
intersticial dos braços longos do par 7 (Figura 5b). A sonda telomérica evidenciou
sequências em todas as regiões teloméricas, além de uma marcação centromérica em um
dos homólogos do par 3 (Figura 5c).

26
Tabela 3 Número diploide e frequência relativa (%) para indivíduos de Synbranchus cf.
lampreia coletados no Lago Catalão (Iranduba, AM).
Número diploide Indivíduos
(Nº de registro)
40 41 42 43 44 45 46 47 48 Total de células
9885 0 0 02 04 37 0 0 0 0 43
9911 0 0 0 08 22 0 0 0 0 30
9912 0 0 0 0 31 0 0 0 0 31
10186 01 0 0 07 24 0 0 0 0 32
10210 0 0 02 08 20 0 0 0 0 30
10340 0 0 0 04 27 0 0 0 0 31
10345 0 0 0 08 22 0 0 0 0 30
10349 0 03 0 09 19 0 0 0 0 31
Total 01 03 04 48 202 0 0 0 0 258
% 0,39 1,16 1,55 18,6 78,30 - - - - 100

27
Figura 4 Cariótipos de Synbranchus cf. lampreia: a) em coloração convencional
Giemsa; b) regiões de heterocromatina constitutiva evidenciada por meio da
técnica de banda C; c) em destaque o par nucleolar impregnado por AgNO3.
m=Metacêntrico, st=Subtelocêntrico, a=Acrocêntrico. Barra igual a 10 µm.

28
Figura 5 Cariótipos de Synbranchus cf. lampreia: a) Localização física cromossômica
do DNAr 18S (amarelo); b) Localização física cromossômica do DNAr 5S
(amarelo); c) Localização física cromossômica da sequência telomérica
(amarelo). m=Metacêntrico, st=Subtelocêntrico, a=Acrocêntrico. Barras
iguais a 20µm.
29
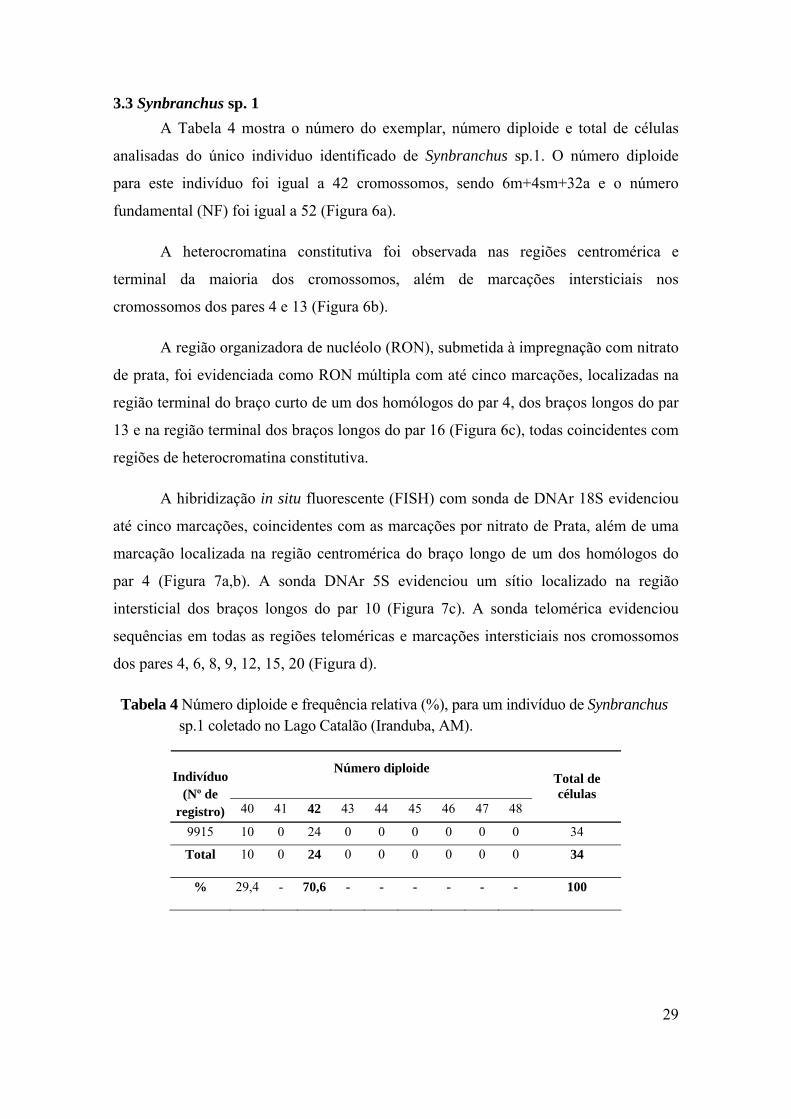
3.3 Synbranchus sp. 1 A Tabela 4 mostra o número do exemplar, número diploide e total de células
analisadas do único individuo identificado de Synbranchus sp.1. O número diploide
para este indivíduo foi igual a 42 cromossomos, sendo 6m+4sm+32a e o número
fundamental (NF) foi igual a 52 (Figura 6a).
A heterocromatina constitutiva foi observada nas regiões centromérica e
terminal da maioria dos cromossomos, além de marcações intersticiais nos
cromossomos dos pares 4 e 13 (Figura 6b).
A região organizadora de nucléolo (RON), submetida à impregnação com nitrato
de prata, foi evidenciada como RON múltipla com até cinco marcações, localizadas na
região terminal do braço curto de um dos homólogos do par 4, dos braços longos do par
13 e na região terminal dos braços longos do par 16 (Figura 6c), todas coincidentes com
regiões de heterocromatina constitutiva.
A hibridização in situ fluorescente (FISH) com sonda de DNAr 18S evidenciou
até cinco marcações, coincidentes com as marcações por nitrato de Prata, além de uma
marcação localizada na região centromérica do braço longo de um dos homólogos do
par 4 (Figura 7a,b). A sonda DNAr 5S evidenciou um sítio localizado na região
intersticial dos braços longos do par 10 (Figura 7c). A sonda telomérica evidenciou
sequências em todas as regiões teloméricas e marcações intersticiais nos cromossomos
dos pares 4, 6, 8, 9, 12, 15, 20 (Figura d).
Tabela 4 Número diploide e frequência relativa (%), para um indivíduo de Synbranchus sp.1 coletado no Lago Catalão (Iranduba, AM).
Número diploide Indivíduo (Nº de
registro) 40 41 42 43 44 45 46 47 48
Total de células
9915 10 0 24 0 0 0 0 0 0 34
Total 10 0 24 0 0 0 0 0 0 34
% 29,4 - 70,6 - - - - - - 100

30
Figura 6 Cariótipos de Synbranchus sp. 1: a) em coloração convencional Giemsa; b)
regiões de heterocromatina constitutiva evidenciada por meio da técnica de
banda C; c) em destaque os pares nucleolares impregnados por AgNO3.
m=Metacêntrico, sm=Submetacêntrico, a=Acrocêntrico. Barra igual a 10µm.

31
Figura 7 Cariótipos de Synbranchus sp.1: a) Localização física cromossômica do DNAr
18S (amarelo); b) em destaque os pares portadores dos sítios de DNAr 18S; c)
Localização física cromossômica do DNAr 5S (amarelo); d) Localização
física cromossômica da sequência telomérica (amarelo). m=Metacêntrico,
sm=Submetacêntrico, a=Acrocêntrico. Barra igual a 20µm.

32
3.4 Synbranchus sp.2 A Tabela 5 mostra o número do exemplar, número diploide e total de células
analisadas do único individuo identificado de Synbranchus sp.2. O número diploide
para este indivíduo foi igual a 42 cromossomos, sendo 6m+2sm+4st+30a e o número
fundamental (NF) foi igual a 54 (Figura 8a).
A heterocromatina constitutiva foi observada nas regiões centromérica e
terminal da maioria dos cromossomos, um dos homólogos do par 1 apresentou dois
blocos intersticiais, sendo um no braço curto e um no longo e alguns cromossomos
apresentaram blocos intersticiais (Figura 8b).
A região organizadora de nucléolo (RON), submetida à impregnação com nitrato
de prata, foi evidenciada como RON simples, localizada na região terminal do braço
curto de um dos homólogos do par 4 (Figura 8c).
A hibridização in situ fluorescente (FISH) com sonda de DNAr 18S evidenciou
até 14 marcações, sendo oito conspícuas. Estas foram intersticiais no braço longo e
curto de um dos homólogos do par 1, terminal nos braços curtos e intersticial nos braços
longos do par 4, intersticial nos pares 12 e 15, terminal nos braços longos do par 13 e
distal nos braços longos do par 14 (Figura 9a). Porém, somente um dos homólogos do
par 4 foi coincidente com o nitrato de Prata.
Tabela 5 Número diploide e frequência relativa (%), para o indivíduo identificado como Synbranchus sp.2 coletado no Lago Catalão (Iranduba, AM).
Número diploide Indivíduo
(Nº de registro) 40 41 42 43 44 45 46 47 48
Total de células
10209 5 0 15 0 0 0 0 0 0 20
Total 5 0 15 0 0 0 0 0 0 20
% 25 - 75 - - - - - - 100

33
Figura 8 Cariótipos de Synbranchus sp. 2: a) em coloração convencional Giemsa; b)
regiões de heterocromatina constitutiva evidenciada por meio da técnica de
banda C; c) em destaque o par nucleolar impregnado por AgNO3.
m=Metacêntrico, sm=Submetacêntrico, st=Subtelocêntrico, a=Acrocêntrico.
Barra igual a 10 µm.

34
Figura 9 Cariótipo de Synbranchus sp. 2: a) Localização física cromossômica do DNAr
18S (amarelo). m=Metacêntrico, sm=Submetacêntrico, st=Subtelocêntrico,
a=Acrocêntrico. Barra igual a 20µm.

35
Figura 10 Idiograma comparativo de (a) Synbranchus madeirae;(b) Synbranchus cf. lampreia; (c) Synbranchus sp.1; (d) Synbranchus sp.2; em preto, heterocromatina constitutiva; em amarelo, genes de DNAr 18S; em rosa, genes de DNAr 5S.

36
3.5 Medidas cromossômicas O comprimento total médio dos cromossomos de Synbranchus cf. lampreia
variou entre 19,255 a 44,049 μm, de S.madeirae variou entre 12,364 a 62,936 μm, de
Synbranchus sp1 variou entre 43,281 a 186,274μm e de Synbranchus sp2 variou entre
41,381 a 183,215μm. O agrupamento dos cromossomos em cinco classes de tamanho
também evidencia a distinta composição cromossômica das quatro espécies (Figura 11).
As classes 1 e 2 fornecem resultados mais expressivos na diferenciação das espécies,
uma vez que na classe 3 ocorre sobreposição entre S. cf. lampreia e Synbranchus sp1, e
na classe 5 entre as espécies S. madeirae, Synbranchus sp1 e Synbranchus sp2.
Figura 11 Análise comparativa das medidas cromossômicas entre as espécies
estudadas. Eixo X: número de classes (classe I: 2,53 a 4,40; classe II: 4,41 a 6,28; classe
III: 6,29 a 8,16; classe IV: 8,17 a 10,04; classe V: 10,05 a 12,87); eixo Y: número de
cromossomos por classe de tamanho.

37
4 Discussão Synbranchiformes é uma ordem de peixes com representantes tanto no Velho
como no Novo Mundo e com ampla distribuição (Nelson 2006). Alguns estudos
envolvendo o conteúdo de DNA nuclear já foram efetuados em algumas espécies desta
ordem, sendo possível evidenciar que Mastacembelus armatus apresenta menor
conteúdo de DNA (1.39 ± 0.08 pg) quando comparado a Macrognathus aculeatus (1,61
pg), ambas espécies com o mesmo número diploide (2n= 48). Em Monopterus albus,
que apresenta somente 2n=24 cromossomos, o conteúdo de DNA (1,24 pg) é muito
similar ao observado para outras espécies de Synbranchiformes, o que indica que
ocorreram poucas alterações na quantidade de DNA durante o processo evolutivo deste
grupo (Jianxun et al. 1991; Oliveira et al. 1997). Em contrapartida, apesar dos estudos
cariotípicos ainda serem escassos comparado com outros grupos de peixes, verifica-se
uma grande variabilidade cromossômica entre as espécies (Tabela 1). O número
diploide varia de 42 a 48 e o no número fundamental (NF) de 24 a 88, sendo que as
espécies do Velho Mundo têm 2n=48 cromossomos.
O número diploide igual a 48 cromossomos acrocêntricos foi considerado por
Ohno (1974) como o cariótipo básico dos teleósteos e segundo Foresti et al. (1992), este
número também seria o ancestral para Synbranchiformes. Visto que a família
Synbranchidae apresenta origem Gondwânica (Rosen 1976), e considerando que as
espécies irmãs do Velho Mundo têm 2n=48 cromossomos (Tabela 1), o cariótipo
ancestral desta família também pode ter sido 2n=48. Porém, não existe consenso com
relação ao número diploide considerado basal para o gênero Synbranchus. Melillo et al.
(1996) sugerem que este número seria 2n=42. Em contrapartida, Sanchez e Fenocchio
(1996) propuseram que as populações de Synbranchus marmoratus da Argentina teriam
se originado a partir de um ancestral com 2n=44, o que teria favorecido a alteração da
fórmula cariotípica e a presença de RONs múltiplas. Ainda, os dados citogenéticos
recentes referentes à S. marmoratus evidenciaram a ocorrência de cinco citótipos,
sugerindo que os números diploides 42 e 44 teriam se originado de um ancestral com
2n=46 cromossomos, com a redução do número cromossômico por meio de rearranjos
robertsonianos do tipo fusão (Torres et al. 2005). Parte desta confusão pode estar
relacionada ao fato de que os indivíduos e populações identificados como S.
marmoratus não formam um grupo monofilético, evidenciado por dados morfológicos

38
(Favorito-Amorim 1992) e moleculares (dois genes mitocondriais) (Perdices et al.
2005).
A análise de mais quatro espécies de Synbranchus identificadas como S.
madeirae (2n=46), S. cf. lampreia (2n=44), Synbranchus sp.1 e Synbranchus sp.2
(2n=42), coletadas em simpatria e sintopia no lago Catalão, Iranduba, no estado do
Amazonas, revelou variabilidade cariotípica interespecífica, sendo verificados três
números diploides e quatro fórmulas cromossômicas. Números diploides semelhantes
aos encontrados no Amazonas foram visualizados em diferentes populações
identificadas de S. marmoratus (Tabela 1). Porém, quando comparamos os dados do
presente trabalho com os disponíveis na literatura, é possível verificar que nenhuma
fórmula cromossômica foi idêntica, ressaltando a presença de rearranjos cromossômicos
não-robertsonianos na evolução cromossômica do grupo. Ainda, apesar de não haver
consenso com relação ao número diploide considerado basal para o gênero
Synbranchus, a evolução cromossômica das espécies deste grupo também englobou
rearranjos robertsonianos que possibilitaram o aparecimento das composições
cariotípicas atuais, os quais favoreceram mudanças no número diploide. Segundo
Sumner (2003), a alteração mais comumente envolvida na evolução cromossômica é o
rearranjo robertsoniano do tipo fusão, mecanismo responsável pela redução do número
diploide e aumento no número de cromossomos com dois braços. No presente trabalho
as duas espécies que apresentaram menor número cromossômico (Synbranchus sp.1 e
Synbranchus sp.2, ambas com 2n=42 cromossomos) também evidenciaram mais
cromossomos do tipo meta-submetacêntricos quando comparado à S. cf. lampreia e S.
madeirae, corroborando a possível ocorrência de fusões cromossômicas.
A presença destes rearranjos cromossômicos também pode ser visualizada pelo
mapeamento físico cromossômico de sequências teloméricas, que consistem de
repetições curtas, ricas em guanina (TTAGGG)n e amplamente conservadas nos
genomas de vertebrados (Guerra 2004), sendo regiões de extrema importância para
manutenção da estabilidade e integridade cromossômica (Nanda et al. 2002; Multani et
al. 2006; Monagham 2010). A hibridização com sonda telomérica evidenciou
marcações nos telômeros de todos os cromossomos das três espécies (S. madeirae, S. cf.
lampreia e Synbranchus sp.1) (não foi obtida para Synbranchus sp. 2), além disso,
foram encontradas marcações intersticiais em duas espécies, o que comprova que na

39
evolução cariotípica do grupo houve a ocorrência de evento de fusão, com a redução do
número diploide. Ainda, essas sequências teloméricas intersticiais podem ser
consideradas resquícios ancestrais de rearranjos cromossômicos, produzidos durante a
evolução cariotípica (Metcalfe et al. 2007). Deste modo, isto é indicativo de um
processo de rearranjo recente ocorrido durante a evolução do grupo (Nanda et al. 2002).
Além disso, rearranjos cromossômicos do tipo inversões e translocações não podem ser
descartados. Estes rearranjos podem contribuir para a reorganização do genoma,
oferecendo novas fórmulas cromossômicas e novos números de braços nos cariótipos.
Assim, os dados do presente trabalho corroboram com a hipótese de Foresti et al.
(1992) e sugerem que o ancestral de Synbranchus possuía 2n=48 cromossomos.
Além da variação interespecífica na fórmula cariotípica e número de
cromossomos, a análise comparativa do tamanho dos cromossomos de S. marmoratus,
S. cf. lampreia, Synbranchus sp.1 e Synbranchus sp.2 também evidenciou diferenças na
organização do genoma das mesmas. Apesar das quatro espécies apresentarem a maioria
dos cromossomos distribuídos na classe de menor tamanho, a frequência dos
cromossomos que estão agrupados nesta classe variou, sendo que 70% dos
cromossomos de S. madeirae apresentam de 2,53 a 4,4µm, 50% dos de S. cf. lampreia,
42% dos de Synbranchus sp.1 e 52% dos de Synbranchus sp.2. Atualmente é
amplamente aceito que a diversidade de tamanho e de organização dos genomas é
influenciada por regiões de DNA repetitivo não codificante, como pseudogenes,
retrotransposons, transposons, DNA satélite e outros, estando a maioria destes
elementos alocados em regiões heterocromáticas (Leitch 2007).
O padrão de distribuição da heterocromatina de algumas espécies de
Synbranchiformes foi estudado e blocos heterocromáticos foram visualizados nas
regiões centromérica e pericentromérica de quase todos os cromossomos, sendo que em
alguns foram também evidenciados em regiões intersticiais (Foresti et al. 1992; Sanchez
e Fenocchio 1996; Oliveira et al. 1997; Liu et al. 2002). As quatro espécies de
Synbranchus analisadas no presente trabalho apresentaram padrões de heterocromatina
espécie-específicos, sendo possível observar além das marcações já encontradas para as
outras espécies, blocos heterocromáticos nas regiões terminal, proximal e distal da
maioria dos cromossomos, caracterizando um padrão muito diferente daqueles já
descritos. Esse padrão distinto de distribuição de heterocromatina, com um possível

40
aumento da mesma, pode ser devido a um processo de heterocromatinização ou adição
de heterocromatina nos indivíduos de Synbranchus spp. coletados no lago Catalão. Esta
diferenciação pode ter ocorrido por trocas desiguais durante processos de crossing over,
transposições ou duplicações dessas sequências, ou mesmo por mecanismos
epigenéticos, como o remodelamento da cromatina e a metilação do DNA, os quais
auxiliam na compactação e organização do genoma em domínios cromáticos (Grewal e
Jia 2007).
Variação na quantidade e localização da heterocromatina constitutiva tem sido
considerada como importante marcador de alguns gêneros de peixes amazônicos, como
observado em Serrasalmus (Characidae, Serrasalminae), onde sete espécies apresentam
um grande bloco proximal no braço longo de um par de cromossomos, coincidente com
o DNAr 5S (Nakayama et al. 2008; Nakayama et al. no prelo). Ainda, em alguns casos,
este marcador pode ser populacional, como evidenciado em diferentes populações de
Geophagus brasiliensis (Vicari et al. 2006). A heterocromatina é atualmente
reconhecida como uma parte importante do genoma dos eucariotos, cujas funções
incluem segregação cromossômica, organização nuclear e regulação da expressão
gênica, e também pode afetar o processo de recombinação gênica (Grewal e Jia 2007;
Skipper 2007; Bühler 2009).
Ainda, evidências mostram que as regiões heterocromáticas podem ter funções
adicionais, incluindo a regulação da mitose, progressão do ciclo celular e proliferação
das células assim como, podem estar associadas a respostas a mudanças no ambiente
como muitas formas de estresses exógenos, como trocas de temperatura, choques
térmicos e hipóxia (Burt e Trivers 2006; Varriale et al. 2008). Para espécies de peixes
amazônicos já foi evidenciado que o padrão de distribuição da heterocromatina
constitutiva nas três espécies de acarás-disco (Symphysodon spp., Cichlidae) pode estar
relacionado ao ambiente em que vivem, visto que cada espécie ocorre preferencialmente
em águas que apresentam padrões físico-químicos distintos (Gross et al. 2010a).
De maneira geral, em peixes, a heterocromatina é rica em sequências repetitivas
e estas têm merecido atenção especial em estudos relativos à estrutura centromérica e
telomérica (Lanfredi et al. 2001; Vicente et al. 2001), origem e evolução de
cromossomos sexuais (Devlin et al. 1998; Stein et al. 2001; Venere et al. 2004),

41
cromossomos supranumerários (Mestriner et al. 2000; Jesus et al. 2003) e evolução dos
genomas (da Silva et al. 2002). Dentre estas sequências repetitivas destacam-se os genes
ribossomais e sequências teloméricas.
Os DNAs codificadores de RNAs ribossomais podem ser divididos em duas
famílias multigênicas, repetidas em tandem, a primeira classe, representada pelo DNAr
45S, que compreende as regiões que transcrevem os genes RNAs 28S, 18S e 5,8S, além
de espaçadores intergênicos, estando estes genes relacionados com as regiões
organizadoras de nucléolo; e a segunda classe, DNAr 5S, consiste de uma sequência
altamente conservada de 120 pares de bases e espaçadores não transcritos (NTS) de
tamanho variável (Martins 2007).
Segundo revisão de Galetti Jr. e Martins (2004), estudos realizados em peixes
neotropicais, como Astyanax spp, Brycon spp., Hoplias malabaricus, Leporinus spp.,
entre outros, têm utilizado a técnica da hibridização fluorescente in situ (FISH) com
sondas de DNA ribossômico, para compreender a organização das regiões
organizadoras de nucléolo e inferir sobre a evolução cromossômica das espécies. Este
método é considerado mais sensível que a impregnação por nitrato de prata porque
localiza as sequências de DNA ribossômico da família 45S em metáfases mitóticas e
células interfásicas, ao invés de marcar as proteínas nucleolares envolvidas com a
atividade transcricional dos genes ribossomais como ocorre com a prata (Miller et al.
1976; Pendás et al. 1993; Phillips e Reed 1996).
Região organizadora de nucléolo simples pode ser considerada uma
característica plesiomórfica em estudos de evolução cromossômica de qualquer grupo
de organismos, e para a ordem Synbranchiformes, a RON simples é considerada uma
condição ancestral (Oliveira et al. 1997). Dentro desta ordem, foram analisadas espécies
das famílias Mastacembelidae e Synbranchidae. Na espécie Mastacembelus armatus,
pertencente à primeira família, RON simples foi verificada e em Synbranchus
marmoratus foi identificada RON simples em pelo menos 15 populações coletadas no
Sul, Sudeste e Centro-oeste do Brasil, porém em posições distintas, enquanto que três
populações da Argentina apresentaram RON múltipla (Tabela 1). Porém, é necessário
enfatizar que estes resultados para M. armatus e S. marmoratus referem-se apenas à
impregnação com nitrato de prata, que tem afinidade com proteínas nucleolares
envolvidas com a atividade transcricional dos genes ribossomais da família 45S,

42
identificando apenas as RONs que estiveram ativas durante a intérfase precedente
(Miller et al. 1976; Howell e Black 1980; Boisvert et al. 2007). Ou seja, não é possível
determinar com exatidão quantos são os sítios ribossomais encontrados nos indivíduos
das diferentes populações de S. marmoratus já analisadas. Entretanto, em M. albus com
o menor número diploide do grupo (2n=24 telocêntricos), o padrão da RONs
evidenciada tanto pela impregnação com AgNO3 como pela FISH com sonda DNAr
18S foi simples e múltipla (Ji et al. 2003). Esses autores acreditam que essa
variabilidade intraespecífica encontrada em M. albus, pode estar relacionada com a
atividade transcricional dos genes ribossomais. No presente estudo, a localização das
regiões organizadoras de nucléolo (RONs), detectada tanto pela impregnação com
AgNO3 como pela FISH com sonda DNAr 18S constituiu num excelente marcador
espécie-específica para as quatro espécies de Synbranchus, ou seja, S. cf. lampreia
apresentou RON simples, evidenciada tanto pela impregnação com AgNO3 como pela
hibridização da sonda homóloga DNAr 18S; Synbranchus sp.1 apresentou RON
múltipla, em ambas as técnicas e Synbranchus sp.2 mostrou apenas um cromossomo
marcado pelo AgNO3 e até 14 marcações pela FISH com a sonda DNAr 18S. Já, S.
madeirae apresentou RON múltipla e com variação intraespecífica para ambas as
técnicas (Quadro1).
A discordância entre os números de RONs e sítios ribossomais 18S encontrada
em S. madeirae provavelmente está relacionada com a atividade transcricional dos
cístrons ribossomais, porém a detecção de dois padrões de distribuição dos genes RNAr
18S demonstra a presença de rearranjos cromossômicos do tipo não-robertsonianos nos
indivíduos desta espécie, os quais podem ser facilitados pela presença de regiões
heterocromáticas adjacentes ou coincidentes a estes cístrons. Ainda, estas regiões
heterocromáticas podem estar abrigando elementos transponíveis, os quais parecem
afetar a mobilidade dos cístrons ribossomais (Guerra 2004). Este fato corroboraria a
hipótese de Sanchez e Fenocchio (1996), que analisando populações de S. marmoratus
da Argentina, sugeriram que a localização da RON em regiões terminal e intersticial
facilitaria eventos de translocação destes sítios no genoma, o que poderia explicar a
origem de RON múltipla nestas populações.Variabilidade inter e intraespecífica quanto
ao número e tamanho dos sítios ribossomais 18S relacionada à mobilização de

43
elementos transponíveis durante a evolução já foram descritas para outras espécies de
peixes amazônicos, tais como os acarás-disco (Gross et al. 2010a).
Em relação ao gene DNAr 5S, para a maioria dos organismos ele está presente
em apenas um par cromossômico, embora sua localização possa ser em região
intersticial ou terminal, e só podem ser observados com a técnica de hibridização in situ
(Martins e Galetti Jr 1999; Guerra 2004; Martins e Wasko 2004; Galetti Jr e Martins
2004). Entretanto, Martins e Galetti Jr. (1999) consideram que a localização desse sítio
em posição intersticial nos cromossomos estaria mais protegida de eventos de
transposição, os quais poderiam agir na dispersão dessas sequências. Esses autores
hipotetizaram que a localização deste gene em apenas um par cromossômico seja a
condição ancestral entre os organismos (Guerra 2004; Martins e Galetti Jr. 1999). A
FISH com a sonda DNAr 5S, nas espécies de Synbranchus aqui analisadas, evidenciou
uma marcação intersticial nos braços longos de um par cromossômico para três
espécies, sendo que em S. cf. lampreia e em S. madeirae foi o par 7 e em Synbranchus
sp.1 o par 10 (dado não obtido para Synbranchus sp 2). Porém, como as fórmulas
cariotípicas são diferentes, pode-se considerar o sitio de DNAr 5S homeólogo para as
três espécies de Synbranchus, sugerindo uma conservação para este gene, e que os
diversos rearranjos ocorridos na evolução cariotípica deste gênero não afetaram o sítio
DNAr 5S. Ainda, em peixes o DNAr 18S e 5S podem estar localizados em sintenia
(Morán et al. 1996; Almeida-Toledo et al. 2002; Cross et al. 2006) ou localizados em
diferentes pares cromossômicos (Martins e Galleti Jr 2001). Em S. madeirae a marcação
5S foi sintênica com uma das marcações do DNAr 18S.
A variabilidade cariotípica encontrada nas espécies de Synbranchus pode estar
associada às características da biologia, fisiologia e reprodução desses peixes, e tais
características podem ter favorecido o surgimento de mecanismos de isolamento
reprodutivo ou interrupção do fluxo gênico (Futuyma, 2002; Ridley 2006),
possibilitando a fixação de rearranjos cromossômicos. De acordo com Galleti Jr. et al.
(2000), grupos de peixes com variação cariotípica apresentam possibilidade de formar
populações isoladas, o que favorece a especiação e a formação de novos táxons, que
nem sempre são reconhecíveis morfologicamente.

44
Nakamoto et al (1986a, b) analisando populações identificadas como S.
marmoratus coletadas em diferentes regiões de São Paulo, observaram dois padrões de
fenótipos eletroforéticos para a hemoglobina. O primeiro padrão, fenótipo 1, apresentou
três bandas distintas de hemoglobinas: Hb1 (hemoglobina catódica), Hb 2 (hemoglobina
intermediária) e Hb 3 (hemoglobina anódica) nos indivíduos das regiões de Botucatu,
Rio Claro e Ribeirão Preto. O segundo fenótipo também apresentou os três tipos de
hemoglobinas, mas com 12 bandas. Foresti et al (1992), estudando citogeneticamente os
mesmos indivíduos, verificaram uma variação no número diploide (2n=42 a 46). Esses
autores sugerem uma correlação entre o número diploide e o tipo de hemoglobina, ou
seja, os indivíduos identificados pelo fenótipo 1 das hemoglobinas estão associados com
números diploides iguais a 44 e 46 cromossomos, enquanto o fenótipo 2 relaciona-se
apenas com 2n = 42 cromossomos.
Até o momento, apesar da presença de grande variabilidade cariotípica,
cromossomos sexuais diferenciados não foram observados em espécies do gênero
Synbranchus, e esta característica pode facilitar a reversão sexual descrita para essas
espécies. Fato semelhante também ocorre em duas espécies de peixes hermafroditas
sequenciais protogínicos Epinephelus guttatus (Perciformes, Serranidae) e Thalassoma
bifasciatum (Perciformes. Labridae), cujos machos e fêmeas possuem constituição
cromossômica idêntica e a determinação do sexo ocorre por mecanismos gênicos (Ruiz-
Carus 2002).
Diante da variabilidade cromossômica encontrada no presente trabalho e com
base nos dados disponíveis na literatura para o gênero Synbranchus, acreditamos que a
tendência mais provável de evolução cromossômica para este grupo seja a partir de um
ancestral com 2n=48 cromossomos (extinto ou ainda não encontrado) com a redução do
número diploide. Portanto, para que se possa realmente compreender os mecanismos
envolvidos no processo da evolução cariotípica deste grupo de peixes, faz-se necessário
elucidar a sistemática do mesmo, bem como, ampliar os locais de coleta e associar
estudos que envolvam ecologia, biologia e genética de cada táxon.

45
5 Conclusões Através dos estudos citogenéticos realizados no presente trabalho, pode-se
concluir que:
• Foi possível diferenciar quatro espécies de Synbranchus, tanto pela morfologia
como pelos dados cromossômicos, sendo identificadas como: Synbranchus
madeirae, Synbranchus cf. lampreia, Synbranchus sp.1 e Synbranchus sp.2.
• A variabilidade cariotípica interespecífica do gênero Synbranchus pode ser
atribuída a rearranjos robertsonianos e não-robertsonianos.
• O padrão da heterocromatina constitutiva das quatro espécies de Synbranchus
do lago Catalão caracterizou-se como um padrão muito diferente daqueles já
descritos para Synbranchus marmoratus, sugerindo um processo de
heterocromatinização ou adição de heterocromatina, ou ainda por fatores
epigenéticos.
• A variabilidade da RON, simples para uma espécie (S. cf. lampreia) e múltipla
para as outras três (S. madeirae, Synbranchus sp.1 e Synbranchus sp.2), tanto
por impregnação com AgNO3 como por FISH com DNAr 18S, pode estar
relacionada com a atividade transcricional dos cístrons ribossomais ou com uma
possível mobilização de elementos transponíveis.
• O sítio de DNAr 5S pode ser considerado homeólogo para S. madeirae, S. cf.
lampreia e Synbranchus sp.1, sugerindo uma conservação para este gene
ribossomal.
• A hibridização com sondas teloméricas evidenciou a presença destas sequências
em posição intersticial, o que pode sugerir que rearranjos cromossômicos do
tipo fusão e/ou inversão ocorreram em S. madeirae, S. cf. lampreia e
Synbranchus sp.1.

46
6 Perspectivas No decorrer deste trabalho surgiram novas perspectivas de pesquisas.
• A ampliação dos locais de coleta, para verificar se a variabilidade cariotípica
encontrada em Synbranchus no lago Catalão é encontrada em outros locais.
• Mapeamento cromossômico de outras sequências repetitivas como histonas,
elementos transponíveis (Rex1, Rex3, Rex6, entre outros), para que seja possível
compreender os mecanismos evolutivos que provocaram a variação do número
de sítios DNAr 18S em S. madeirae.
• Comparação da organização genômica dos diferentes táxons de Synbranchus por
meio da GISH (hibridização genômica in situ).
• Realização de estudos histológicos para que seja possível determinar o sexo dos
indivíduos de Synbranchus.
• Realização de estudos citogenéticos clássicos e moleculares de outras espécies
da ordem Synbranchiformes, pois existem poucos dados citogenéticos para esta
ordem.

47
7 Referências Bibliográficas
Almeida-Toledo, L.F. 1998. Cytogenetic markers in Neotropical freshwater fishes, p.
583-588. In: Malabarba, L.R.; Reis, R.E.; Vari, R.P.; Lucena, R.P.; Lucena,
Z.M.S.; Lucena, C.A.S (Eds). Phylogeny and classification of Neotropical fishes.
Editora da Pontifícia Universidade Católica, Porto Alegre, Brasil.
Almeida-Toledo, L.F.; Ozouf-Costaz, C.; Foresti, F.; Bonillo, C.; Porto-Foresti, F.;
Daniel-Silva, M. 2002. Conservation of the 5S-bearing chromosome pair and co-
localization with major rDNA clusters in five species of Astyanax (Pisces,
Characidae). Cytogenetic Genome Research, 97: 229–233.
Alves-Brinn, M.N.; Porto, J.I.R.; Feldberg, E. 2004. Karyological evidence for
interspecific hybridization between Cichla monoculus and C. temensis
(Perciformes, Cichlidae) in the Amazon. Hereditas, 141: 252-257.
Antoneli, F.N. 2002. Cinética da inversão de sexo em Synbranchus marmoratus
(Teleostei, Synbranchiformes, Synbranchidae). Dissertação de Mestrado.
Universidade Estadual de Campinas, Campinas, São Paulo. 162 pp.
Antoneli, F.N. 2006. Perfil morfo-funcional da inversão do sexo em Synbranchidae
(Teleostei: Synbranchiformes). Tese de doutorado. Universidade Estadual de
Campinas, Campinas, São Paulo. 167 pp.
Arkhipchuk, V.V. 1999. Database of Fish chromosomes. http://www.fishbase.org.
Artoni, R.F.; Vicari, M.R.; Bertollo, L.A.C. 2000. Citogenética de peixes neotropicais:
métodos, resultados e perspectivas. Publication UEPG Biological and Health
Sciences 6(1): 43-60.
Asahida, T.; Kobayashi, T.; Saitoh, K.; Nakayama, I. 1996. Tissue preservation and
total DNA extraction from fish stored at ambient temperature using buffers
containing high concentration of urea. Fisheries Science, 62(5): 727-730.
Benzaquem, D.C.; Feldberg, E.; Porto, J.I.R.; Gross, M.C.; Zuanon, J.A.S. 2008.
Cytotaxonomy and karyoevolution of the genus Crenicichla (Perciformes,
Cichlidae). Genetics and Molecular Biology, 1: 250-255.

48
Bertollo, L.A.C.; Takahashi, C.S.; Moreira-Filho, O. 1978. Cytotaxonomic
considerations on Hoplias lacerdae (Pisces, Erytrinidae). Brazilian Journal of
Genetic, 7: 103-120.
Boisvert, F.M.; Koningsbruggen, S.V.; Navascués, J.; Lamond, A.I. 2007. The
multifunctional nucleolus. Molecular Cell Biology, 8: 574-585.
Britski, H.A.; Silimon, K.Z.S.; Lopes, B.S. 1999. Peixes do Pantanal: Manual de
Identificação. EMBRAPA-SPI, Brasília, Brasil. 184 pp.
Burt, A.; Trivers, R. 2006. Genes in conflict: the biology of selfish genetic elements.
Harvard University Press, Cambridge, p. 228-300.
Bühler, M. 2009. RNA turnover and chromatin-dependent gene silencing.
Chromosoma, 118: 141–151.
Centofante, L.; Porto, J.I.R.; Feldberg, E. 2002. Choromosomal polymorphism in
Serrasaln spilopleura Kner, 1858 (Characidae, Serrasalminae) from Central
Amazon Basin. Caryologia, 55: 37-45.
Cross, I.; Merlo, A.; Manchado, M.; Infante, C.; Canavate, J.P.; Rebordinos, L. 2006.
Cytogenetic characterization of the sole Solea senegalensis (Teleostei:
Pleuronectiformes: Soleidae): Ag-NOR, (GATA)n, (TTAGGG)n and ribosomal
genes by one-color and two-color FISH. Genetica, 128: 253–259.
Dasilva, C.; Hadji, H.; Ozouf-Costaz, C.; Nicaud, S.; Jaillon, O.; Weissenbach, J.;
Crollius, H.R. 2002. Remarkable compartmentalization of transposable elements
and pseudogenes in the heterochromatin of the Tetraodon nigroviridis genome.
Proceedings of the National Academy of Science USA, 99(21): 1636-1641.
de Oliveira, R.R.; Feldberg, E.; Anjos, M. B.; Zuanon, J. 2007. Karyotype
characterization and ZZ/ZW sex chromosome heteromorphism in two species of
the catfish genus Ancistrus Kner, 1854 (Siluriformes: Loricariidae) from the
Amazon basin. Neotropical Ichthyology, 5: 301–306.
de Oliveira, R.R.; Feldberg, E.; Anjos, M.B.; Zuanon, J. 2008. Occurrence of multiple
sexual chromosomes (XX/XY1Y2 and Z1Z1Z2Z2/Z1Z2W1W2) in catfishes of the

49
genus Ancistrus (Siluriformes: Loricariidae) from the Amazon basin. Genetica,
134: 243–249.
Devlin, R.H.; Stone, G.W.; Smailus, D.E. 1998. Extensive direct-tandem organization
of a long repeat DNA sequence on the Y chromosome of chinoock salmon
(Oncorhynchus tshawytscha). Journal of Molecular Evolution, 46: 277-287.
Estoup, A.; Presa, P.; Krieg, F.; Vaiman, D.; Guyomard, R. 1993. (CT)n and (GT)n
microsatellites: a new class of genetic markers for Salmo trutta L. (brown trout).
Heredity, 71: 488-496.
Favorito-Amorin, S.E. 1992. Revisão sistemática das espécies brasileiras do gênero
Synbranchus (Teleostei, Acanthopterygii). Dissertação de mestrado. Universidade
de São Paulo. 80pp.
Favorito, S.E.; Zanata, A.M.; Assumpção, M.I. 2005. A new Synbranchus (Teleostei:
Synbranchiformes: Synbranchidae) from ilha Marajó, Pará, Brazil, with notes on
its reproductive biology and larval development. Neotropical Ichthyology, 3: 319-
328.
Feldberg, E.; Porto, J.I.R.; Nakayama, C.M. 1993. Karyotype evolution in Curimatidae
(Teleostei, Characiformes) from the Amazon region II Centric fissions in the
genus Potamorhina. Genome, 36:372-376.
Feldberg, E.; Porto, J.I.R.; Alves-Brinn, M.N.; Mendonça, M.N.C.; Benzaquem, D.C.
2004. B chromosomes in Amazonian cichlid species. Citogenetics and Genome
Research, 106: 195–198.
Fonseca, J.S.; Martins, G.A. 1982. Curso de Estatística. Terceira edição, São Paulo.
Atlas. 286pp.
Foresti, F.; Oliveira, C.; Tien, O.S. 1992. Cytogenetic studies in fishes of the genus
Synbranchus (Pisces, Synbranchiformes, Synbranchidae). Naturalia, 17: 129-138.
Futuyma, D.J. 2002. Biologia evolutiva. Editora Ribeirão Preto, São Paulo, Brasil.
567pp.

50
Galetti Jr., P.M.; Aguilar, C.T.; Molina, W.F. 2000. An overiew of marine fish
cytogenetics. Hydrobiologia, 420: 55-62
Galetti Jr, P.M.; Martins, C. 2004. Contribuição da hibridização in situ para o
conhecimento dos cromossomos dos peixes. In: Guerra, M. (Ed). FISH: Conceitos
e Aplicações na Citogenética. Editora da Sociedade Brasileira de Genética,
Ribeirão Preto, Brasil. p. 61-88.
Graham, J.B.; Lai, N.C.; Chiller, D.; Roberts, J.L. 1995. The transition to air breathing
in fishes V. Comparative aspects of cardiorespiratory regulation in Synbranchus
marmoratus and Monopterus albus (Synbranchidae). The Journal of Experimental
Biology, 198: 1455-1467.
Grewal, S.I.S.; Jia, S. 2007. Heterochromatin revised. Nature reviews, 8: 35-46.
Gross, M.C.; Schneider, C.H.; Valente, G.T.; Martins, C.; Feldberg, E. 2010a.
Comparative cytogenetic analysis of the genus Symphysodon (Discus fishes,
Cichlidae): chromosomal characteristics of retrotransposons and minor ribosomal
DNA. Cytogenetic and Genome Research, 127: 43-53.
Gross, M.C.; Schneider, C.H.; Valente, G.T.; Martins, C.; Feldberg, E. 2010b.
Variability of 18S rDNA locus among Symphysodon fishes: chromosomal
rearrangements. Journal of Fish Biology, 76: 1117-1127.
Guerra, M. 1988. Introdução à citogenética geral. Editora Guanabara, Rio de Janeiro,
Brasil. 142 pp.
Guerra, M. 2004. Hibridização in situ: Princípios básicos. In: Guerra, M. (Ed). FISH:
Conceitos e Aplicações na Citogenética. Editora da Sociedade Brasileira de
Genética, Ribeirão Preto, Brasil. p. 1-32.
Howell, W.M.; Black, D.A. 1980. Controlled silver staining of nucleolus organizer
regions with a protective colloidal developer: a 1-step method. Experientia, 3:
1014-1015.
IJdo, J.W.;.Wells, R.A.; Baldini, A.; Reeders, S.T. 1991. Improved telomere detection
using a telomere repeat probe (TTAGGG)n generated by PCR. Nucleic Acids
Research, 19(17): 4780.
Jesus, C.M.; Galetti Jr., P.M.; Valentini, S.R.; Moreira-Filho, O. 2003. Molecular
characterization and chromosomal location of two families of satellite DNA in

51
Prochilodus lineatus (Pisces, Prochilodontidae), a species with B chromosomes.
Genetica, 118: 25-32.
Ji , F.Y.; Yu, Q.X.; Li, K.; Ren, X.H.2003. Ag-staining pattern, FISH and ISH with
rDNA probes in the rice field eel (Monopterus albus Zuiew) chromosomes.
Hereditas, 138: 207-212.
Jianxun, C.; Xiuhai, R.; Qxing, Y. 1991. Nuclear DNA content variation in fishes.
Cytologia, 56: 425-429.
Komiya, H.; Takemura, S. 1979. Nucleotide sequence of 5S ribosomal RNA from
rainbow trout (Salmo gairdnerii) liver. The Journal of Biochemistry, 86: 1067-
1080.
Khuda-Bukhsh, A.R.; Barat, A. 1987. Chromosomes in fifteen species of Indian teleosts
(Pisces). Caryologia, 40: 131-144.
Kullander, S.O. 2003. Family Synbranchidae, p. 594-595. In: Reis, R.R.; Kullander,
S.O.; Ferraris. C.J. (Eds). Check list of the freshwater of South and Central
America. Editora da Universidade Católica, Porto Alegre, Brasil.
Lanfredi, M.; Congiu, L.; Garrido-Ramos, M.A.; Herrán, L.D.; Leis, M.; Chicca, M.;
Rossi, R.; Tagliavini, J.; Rejón, C.R.; Rejón, M.R.; Fontana, F. 2001.
Chromosomal location and evolution of a satellite DNA family in seven sturgeon
species. Chromosome Research, 9: 47-52.
Leitch, I.J. 2007. Genome sizes through the ages. Heredity, 99: 121–122.
Levan, A.; Fredga, K.; Sandberg, A.A. 1964. Nomenclature for centromeric position on
chromosomes. Hereditas, 52: 201-220.
Liu, J.D.; Yi, M.S.; Zhao, G.; Zhou, F.; Wang, D.Q.; Yu, Q.X. 2002. Sex chromosomes
in the spiny eel (Mastacembelus aculeatus) revealed by mitotic and meiotic
analysis. Cytogenetic Genome Research, 98: 291-297.
Lo Nostro, F.; Guerrero, G. 1996. Presence of primary and secondary males in a
population of Synbranchus marmoratus, Bloch 1795, a protogynous fish
(Teleostei, Synbranchiformes). Journal of Fish Biology, 49: 788-800.
Lo Nostro, F.; Grier, H.; Andreone, L.; Guerrero, G.A. 2003. Involvement of the
gonadal germinal epithelium during sex reversal and seasonal testicular cycling in

52
the protogynous swamp eel, Synbranchus marmoratus Bloch 1795 (Teleostei,
Synbranchidae). Journal of Morphology, 257: 107-126.
Manna, G.K.; Prasad, R. 1977. Chromosome analysis of five species of freshwater
fishes. Nucleus, 20: 264-271.
Margarido, V.P.; Galetti Jr, P.M. 1999. Heterochromatin patterns and karyotype
relationships within and between the genera Brycon and Salminus (Pisces,
Characidae). /Genetics and Molecular Biology, 22(3): 357-361.
Martins, C.; Galleti Jr., P.M. 1999. Chromosomal localization of 5S rDNA genes
Schizodon (Pisces, Anostomidae) chromosomes. Chromosome Research, 8(4):
353-355.
Martins, C.; Galleti Jr., P.M. 2001. Two 5S rDNA arrays in Neotropical fish species: is
it a general rule for fishes? Genetica, 111: 439-446.
Martins, C.; Wasko, A.P. 2004. Organization and evolution of 5S ribosomal DNA in the
fish genome. In: Williams, C.R. (ed) Focus on Genome Research. Nova Science
Publishers, Hauppauge, NY, USA. p. 335-363.
Martins, C. 2007. Chromosomes and repetitive DNAs: a contribution to the knowledge
of fish genome. In: Pisano, E.; Ozouf-Costaz, C.; Foresti, F.; Kapoor, B.G. (Eds).
Fish Cytogenetics. Science Publisher, Inc., Enfield, New Hampshire, USA. p.
421-453.
Melillo, M.I.F.M.; Foresti, F.; Oliveira, C. 1996. Additional cytogenetic studies on local
populations of Synbranchus marmoratus (Pisces, Synbranchiformes,
Synbranchidae). Naturalia, 21: 201-208.
Mesquita, D.R.; Porto, J.I.R.; Feldberg, E. 2008. Chromosomal variability in the wild
ornamental species of Symphysodon (Perciformes: Cichlidae) from Amazon.
Neotropical Ichthyology, 6: 181-190.
Mestriner, C.A.; Galetti Jr., P.M.; Valentini, S.R.; Ruiz, I.R.G.; Abel, L.D.S.; Moreira-
Filho, O.; Camacho, J.P.M. 2000. Structural and functional evidence that a B
chromosome in the characid fish Astyanax scabripinnis is an isochromosome.
Heredity, 85: 1-9.
Metcalfe, C.J.; Eldridge, M.D.; Johnston, P.G. 2007. Mapping the distribution of the
telomeric sequence (T2AG3)n in the Macropoidea (Marsupialia) by fluorescence

53
in situ hybridization. II. The ancestral 2n=22 macropodid karyotype. Cytogenetics
Genome Research, 116: 212-217.
Miller, E.A.; Dev, V.G.; Tantravahi, R.; Miller, O.J. 1976. Supression of human
nucleolus organizer activity in mouse human somatic hybrid cells. Experimental
Cell Research, 101: 235-243.
Monagham, P. 2010. Telomeres and life histories: the long and the short of it. Annals of
the New York Academy of Sciences, 1206: 130-142.
Morán, P.; Martínez, J.L.; Garcia-Vazquez, E.; Pendás, A.M. 1996. Sex chromosome
linkage of 5S rDNA in rainbow trout. Cytogenetics and Cell Genetics, 75: 145-
150.
Multani, A.S.; Ozen, M.; Furlong, C.L.; Zhao, Y.J.; Hsu, T.C.; Pathak, S. 2006.
Heterochromatin and interstitial telomeric DNA homology. Chromosoma, 110:
214–220.
Nakayama, C.M.; Porto, J.I.R.; Feldberg, E. 2000. Ocorrência de dois citótipos em
Serrasalmus spilopleura Kner, 1858 (Characiformes, Serrasalmidae) da região de
confluência dos rios Negro e Solimões Amazonas, Brasil. Acta Amazonica, 1:
149-154.
Nakayama, C.M.; JéGu, M.; Porto, J.I.R.; Feldberg, E. 2001. Karyological evidence for
a cryptic species of piranha within Serrasalmus rhombeus (Characidae,
Serrasalminae) in the Amazon. Copeia, 3: 866–869.
Nakayama, C.M.; Feldberg, E.; Bertollo, L.A.C. 2008. Mapping of ribosomal genes and
chromosomal markers in three piranha species of the genus Serrasalmus
(Characidae, Serrasalminae) from the Amazon basin. Genetics and Molecular
Biology, 31(4): 868-873.
Nakayama, C.M.; Feldberg, E.; Bertollo, L.A.C. In press. Karyotype differentiation and
cytotaxonomic considerations in species of Serrasalminae (Characiformes,
Characidae) from the Amazon basin. Neotropical Ichthyology

54
Nakamoto, W.; Machado, P.E.A.; Foresti, F. 1986a. Hemoglobins patterns in different
populations of Synbranchus marmoratus (Pisces, Synbranchidae). Comparative
Biochemical and Physiology, 84: 377-382.
Nakamoto, W.; Machado, P.E.A.. 1986b. Characterization of the hemoglobins and globins
of Synbranchus marmoratus (Pisces, Synbranchidae). Comparative Biochemical and
Physiology, 84: 383-386.
Nanda, I.; Schrama, D.; Feichtinger, W.; Haaf, T.; Schartl, M.; Schmid, M. 2002.
Distribuition of telomeric (TTAGGG) sequences in avian chromosomes.
Chromosoma, 111: 215-227.
Nelson, J.S. 2006. Fishes of the world. 4 ed. John Wiley and Sons, New York. 600 pp.
Ohno, S. 1974. Protochordata, Cyclostomata and Pisces. Animal Cytogenetics, 4(1): 1-
92.
Oliveira, C.; Almeida-Toledo, L.F.; Foresti, F.; Toledo-Filho, S.A. 1988.
Supernumerary chromosomes, Robertsonian rearrangement and multiple NORs in
Corydoras aeneus (Pisces, Siluriformes, Callichthyidae). Caryologia, 41: 227-
236.
Oliveira, C.; Almeida Toledo, L.F.; Moli, L; Toledo-Filho, S.A. 1992. Extensive
chromosomal rearrangements and nuclear DNA content changes in the evolution
of the armoured catfishes genus Corydoras (Pisces, Siluriformes, Callichthyidae).
Journal Fish Biology, 40: 419-431.
Oliveira, C.; Torres, R.A.; Favorito, S.; Foresti, F. 1997. Cytogenetic studies of
Mastacembelus armatus (Pisces, Mastacembelidae). Cytobios, 92: 83-89.
Pendás, A.M.; Moran, P.; Garcia-Vasquez, E. 1993. Ribosomal RNA genes are
interspersed throughout a heterochromatic chromosome arm in Atlantic salmon.
Cytogenetic and Cell Genetics, 63: 128-130.
Perdices, A.; Doadrio, I.; Bermingham, E. 2005. Evolutionary history of the
synbranchid eels (Teleostei: Synbranchidae) in Central America and the
Caribbean islands inferred from their molecular phylogeny. Molecular
Phylogenetics and Evolution, 37: 460–473.

55
Pinkel, D.; Straume, T.; Gray, J.W. 1986. Cytogenetic analysis using quantitative, high-
sensitivity, fluorescence hybridization. Proceedings of the Natural Academy of
Science, 83: 2934-2938.
Phillips, R.B.; Reed, K.M. 1996. Application of fluorescence in situ hybridization
(FISH) techniques to fish genetics: a review. Aquaculture, 40: 197-216.
Ravaglia, M.A.; Maggese, M.C. 2003. Ovarian follicle ultrastructure in the teleost
Synbranchus marmoratus Bloch, 1795, with special reference to the vitelline
envelope development. Tissue Cell, 35: 9-17.
Reis, R. E., Kullander S. O., Ferraris, C.J. 2003. Check List of The Freshwater Fishes of
South and Central America. Porto Alegre, EDIPUCRS. 742 pp.
Ridley, M. 2006. Evolução.Editora Artmed, Porto Alegre, Brasil, 752pp.
Rosen, D.E.; Rumney, A. 1972. Evidence of a second species of Synbranchus (Pisces,
Teleostei) in South America. American Museum, 2497: 1-45.
Rosen, D.E.; Greenwood, P.H. 1976. A fourth Neotropical species of synbranchid eel
and the phylogeny and systematics of synbranchiform. Ann Wien von Mus.
Naturg, 157: 1-70.
Rosen, D.E., 1976. A vicariance model of Caribbean biogeography. Systematic Zoology,
2: 431–464.
Ruiz-Carus, R. 2002. Chromosome analysis of the sexual phases of the protogynous
hermaphrodites Epinephelus guttatus and Thalassoma bifasciatum (Serranidae
and Labridae; Teleostei). Caribbean Journal of Science, 38: 44-51.
Saiki, R.K.; Gelfand, D.H.; Stoffel, S.; Scharf, S.J.; Higuchi, R.; Horn, G.T.; Mullis, K.
B.; Erlich, H.A. 1988. Primer-directed enzymatic amplification of DNA with a
thermostable DNA polymerase. Science, 239: 487-491.
Sambrook, J.; Russell, D.W. 2001. Molecular cloning: a laboratory manual. Vol. I.
Cold Spring Harbor Press, Cold Spring Harbor, NY. p. 633–664.
Sanchez, S.; Fenocchio, A.S. 1996. Karyotypic analysis in three populations of the
South-American eel like fish Synbranchus marmoratus. Caryologia, 49: 65-71.

56
Santos, G.M.; Ferreira, E.J.G. 1999. Peixes da Bacia Amazônica. In: Lowe-McConnell,
R.H. (Eds). Estudos ecológicos de comunidades de peixes tropicais. Editora da
Universidade de São Paulo, São Paulo, Brasil. p.345-373.
Schofield, P.; Nico, L.G. 2009. Salinity of non-native Asian swamp eels (Teleostei;
Synbranchidae) in Florida, USA: comparison of three populations and
implications for dispersal. Environmental Biology of Fishes, 85: 51-59.
Scudeler, P.E.S. 2010. Estudo da Estrutura Molecular dos Cromossomos
Supranumerários em Moenkhausia sanctaefilomenae (Teleostei, Characiformes,
Characidae). Tese de doutorado. Universidade Estadual Paulista Júlio de Mesquita
Filho, Botucatu, São Paulo. 136 pp.
Shibata, O.A. 2006. 40 peixes do Brasil: CESP 40 ANOS. Editora Doiis, Rio de Janeiro,
Brasil, 207pp.
Skipper, M. 2007. Mysteries of heterochromatic sequences unraveled. Nature Reviews,
8: 567.
Stein, J.; Phillips, R.B.; Devlin, R.H. 2001. Identification of the Y chromosome in
chinoock salmon (Oncorhynchus tshawytscha). Cytogenetics and Cell Genetics,
92: 108-110.
Sumner, A.T. 1972. A simple technique for demonstrating centromeric
heterochromatin. Experimental Cell Research, 75: 304-306.
Sumner, A.T. 2003. Chromosomes: organization and function. Blackwell Publishing,
New York, USA. 304 pp.
Teixeira, A.S.; Nakayama, C.M.; Porto, J.I.R.; Feldberg, E. 2006. Esterase-D and
chromosome patterns in Central Amazon piranha (Serrasalmus rhombeus
Linnaeus, 1766) from Lake Catalão. Genetics and Molecular Biology, 3: 498-502.
Terencio, M.L.; Schneider, C.H.; Gross, M.C.; Silva, A.M.; Feldberg, E.; Porto, J.I.R.
2008. Comparative cytogenetics of Carnegiella marthae and Carnegiella strigata
(Characiformes, Gasteropelecidae) and description of a ZZ/ZW sex chromosome
system. Genetics and Molecular Biology, 1: 231-234.

57
Torres, R.A. 2000. O gênero Synbranchus (Pisces, Synbranchinae, Synbranchidae):
interrelações citotaxonômicas, evolutivas e natureza da variabilidade cariotipica
Tese de doutorado. Universidade Estadual Paulista Júlio de Mesquita Filho,
Botucatu, São Paulo. 141 pp.
Torres, R.A.; Roper, J.J.; Foresti, F.; Oliveira, C. 2005. Surprising genomic diversity in
the Neotropical fish Synbranchus marmoratus (Teleostei, Synbranchidae): how
many species? Neotropical Ichthyology, 3: 277-284.
Vari, R.P; Malabarba, L.R. 1998. Neotropical ichthyology: an overview, p. 1-11. In:
Malabarba, L.R.; Reis, R.E.; Vari, R.P.; Lucena, R.P.; Lucena, Z.M.S.; Lucena,
C.A.S. (Eds). Phylogeny and classification of Neotropical fishes. Editora da
Pontifícia Universidade Católica, Porto Alegre, Brasil.
Varriale, A.; Torelli, G.; Bernardi, G. 2008. Compositional properties and thermal
adaptation of 18S rRNA in vertebrates. RNA, 14: 1492-1500.
Venere, P.C.; Ferreira, I.A.; Martins, C.; Galetti Jr., P.M. 2004. A novel ZZ/ZW sex
chromosome system for the genus Leporinus (Pisces, Anostomidae,
Characiformes). Genetica, 121: 75-80.
Vicente, V.E.; Jesus, C.M.; Moreira-Filho, O. 2001. Chromosomal localization of 5S
and 18S rRNA genes in three Parodon species (Pisces, Parodontidae).
Caryologia, 54: 365-369.
Vicari, M.R.; Moreira-Filho, O.; Artoni, R.F.; Bertollo, L.A.C. 2006. Basic and
molecular cytogenetics in freshwater Cichlidae (Osteichthyes, Perciformes).
Karyotype and conservationism and divergence. Caryologia, 59: 260-266.
Yajuan, L.; Qixing, Y. 1991. Electron microscopic observation of synaptonemal
complexes in spermatocytes of six species of fishes. Chinese Journal Genetic, 18:
273-282.
Yu, X.; Zhou, T.; Li, Y.; Zhou, M. 1987. On the karyosystematics of cyprinid fishes and
a summary of fish chromosome studies in China. Genetica, 72: 225-236.