
Uma fonte de água jorrando. Onde reina o amor Fraterno amor! Onde reina o amor: Deus aí está!

I
UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CAMPUS DE BOTUCATU
UTILIZAÇÃO DO CARBONO-13 COMO MARCADOR NA PARTIÇÃO DE
FOTOASSIMILADOS EM FIGUEIRA
ANDRÉA CARVALHO DA SILVA
Dissertação apresentada à Faculdade de Ciências Agronômicas da UNESP - Campus de Botucatu, para obtenção do título de Mestre em Agronomia - Área de Concentração em Horticultura.
BOTUCATU - SP
Fevereiro - 2009

II
UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CAMPUS DE BOTUCATU
UTILIZAÇÃO DO CARBONO-13 COMO MARCADOR NA PARTIÇÃO DE
FOTOASSIMILADOS EM FIGUEIRA
ANDRÉA CARVALHO DA SILVA
Orientadora: Profª. Drª. Sarita Leonel
Co-orientador: Prof. Dr. Carlos Ducatti
Dissertação apresentada à Faculdade de Ciências Agronômicas da UNESP - Campus de Botucatu, para obtenção do título de Mestre em Agronomia - Área de Concentração em Horticultura.
BOTUCATU - SP
Fevereiro - 2009

III

IV

I
A Alderico Carvalho da Silva, meu pai, Paulina Maria de Azevedo Silva, minha mãe,
A Wanderson Carvalho da Silva meu irmão e
Adilson Pacheco de Souza, meu amor
DEDICO.

II
AGRADECIMENTOS
À DEUS, por ser um pai tão amoroso e compassivo por me permitir
concluir mais esta etapa da vida profissional, o que me traz muita alegria e realização.
À Alderico Carvalho da Silva, meu pai, que soube incentivar, encorajar e
compreender as inúmeras vezes que nunca estive em casa.
Á Paulina Maria de Azevedo Silva, por suas orações e afeto mesmo a
quilômetros de distância.
Á Wanderson Carvalho da Silva, meu grande irmão pelo carinho e por
nunca deixar que o sentimento de solidão reina-se em mim.
“...Manda dizer-te que foi porque teu exemplo convenceu.
Houve alguém que venceu... e manda dizer-te que foi porque
tuas lições permaneceram...” (autor desconhecido)
À Adilson Pacheco de Souza, meu amigo amante, pelo ombro, pelo afago
e por recolher minhas lágrimas nos momentos mais difíceis.
Ao grande amigo, Aldir Carlos Silva, pelo grande apoio e
companheirismo.
À Profª Drª Sarita Leonel, onde encontrei apoio, zelo, amizade e
orientação nos momentos de tomadas de decisão durante estes dois anos.
À Prof. Dr. Carlos Ducatti, pela dedicação e abertura com que me recebeu
e pela oportunidade de conhecer uma área de pesquisa tão fascinante como a dos isótopos estáveis
ambientais.
À Marco Antonio da Silva Vasconcellos, pela orientação, estímulo e
confiança durante a realização deste trabalho.
À Prof. Dr. João Domingos Rodrigues, pela paciência, ensinamento e
amizade durante a realização deste trabalho.
Ao técnico de laboratório do Centro de Isótopos Estáveis: Evandro Tadeu
da Silva, Silvia e Sibeli, pela amizade eficiência e competência.
Aos funcionários do Departamento de Horticultura e Pomar.

III
Aos amigos Emi Lorenzetti, Roselani Araldi, Adriana Tanaka, Thaise
Ribeiro, Francine Vercese, Edvar de Sousa e Manoel Euzébio de Souza pelos momentos de risadas
e descontrações durante estes dois anos que muito colaborou em todas as etapas desta pesquisa.
Às minhas eternas irmãs de república Danila Monte Conceição e Simone
Fernandes Ciavatta pelo apoio e amizade sempre.
“Se alguma coisa me consome e me envelhece é que a roda furiosa da vida não me permite ter sempre ao meu lado,
morando comigo, andando comigo, falando comigo, vivendo comigo, todos os meus amigos, e, principalmente os que só
desconfiam ou talvez nunca vão saber que são meus amigos!”(Vinicius de Moraes)
À Faculdade de Ciências Agronômicas de Botucatu pelo apoio em todas
as atividades desenvolvidas durante o período de experimentação.
Aos Colegas do Ministério Universidades Renovadas: Flavinha, João,
Capitu, Leonardo, Meire, Maila, Nadja, Nicolas pela força e continuidade da caminhada rumo ao
céu.
Aos amigos da pós-graduação, pela grande e sincera amizade,
companhia, pelos risos, abraços, pelas palavras de apoio! Cada um seguirá o seu caminho,
talvez em direções opostas... Em mim permanece a certeza de que a vida nos proporcionará
muitos reencontros!
A todos aqueles que anonimamente ajudaram para que este trabalho fosse
concluído, meus sinceros agradecimentos e que o Senhor Jesus recompense de maneira graciosa
todo o empenho disponibilizado.
“Aos que se tornaram familiares, aos que nasceram familiares
e aos que conheci antes de ontem; Aos que me deixaram louco e aos que enlouqueci;
Aos que me criticaram em tudo e a um ou outro que aturou minha “chatura”; Aos amigos que passaram e aos que se estagnaram em mim;
Aos que me consideram muito e aos que com razão fizeram pouco; Aos que conhecem o que penso e aos que só conhecem o que faço;
Aos que passam o dia todo comigo e aos que estão o tempo todo em mim. Este trabalho é a soma de todos vocês. E se não é melhor, é por falta de memória,
mas não por falta de amigos”. (Efraim Rodrigues)
Meus sinceros agradecimentos.....

IV
SUMÁRIO
Página RESUMO......................................................................................................................VII
SUMMARY....................................................................................................................IX
1. INTRODUÇÃO.......................................................................................................... ..1
2. REVISÃO DE LITERATURA ....................................................................................4
2.1. A fruticultura como atividade agrícola..........................................................4
2.2. Situação econômica da ficicultura.................................................................6
2.3. A figueira.......................................................................................................6
2.4. Caracterização Botânica ................................................................................8
2.5. Ecofisiologia da Figueira.............................................................................11
2.6. Fotossíntese e Reserva.................................................................................12
2.7. Translocação de fotoassimilados.................................................................19
2.8. Isótopos e sua Utilização .............................................................................23
3. MATERIAL E MÉTODOS........................................................................................27
3.1. Localização e descrição da área experimental.............................................27
3.2. Caracterização das Trocas Gasosas .............................................................29
3.3. Metodologia de enriquecimento em 13C......................................................30
3.4. Metodologia de coleta do material enriquecido ..........................................32
3.5. Secagem e moagem das amostras................................................................33
3.6. Analise do enriquecimento relativo.............................................................33
4. RESULTADOS E DISCUSSÃO ...............................................................................35
4.1. Caracterização fotossintética de figueiras ‘Roxo de Valinhos’...................35
4.2. Enriquecimento relativo natural da razão 13C/ 12C em figueiras..................39
4.3. Partição de Fotoassimilados em função dos drenos ....................................44
4.4. Partição de Fotoassimilados em função do tempo......................................58
5. CONSIDERAÇÕES FINAIS .....................................................................................72
6. CONCLUSÕES..........................................................................................................74
7. REFERÊNCIAS BIBLIOGRÁFICAS........................................................................75

V
LISTA DE TABELAS
Página
Tabela 1. Médias mensais das temperaturas mínima, máxima e média (ºC), precipitação
pluviométrica (mm) e radiação solar global (W m-2) medidas no período de
janeiro de 2007 a agosto de 2008.. ...........................................................................28
Tabela 2. Dados de trocas gasosas de folhas de figueira ‘Roxo de Valinhos’ em
Botucatu, SP. ............................................................................................................36
Tabela 3. Distribuição natural dos valores de δ 13C nas partes vegetativas dos ramos da
figueira ‘Roxo de Valinhos’. ....................................................................................40
Tabela 4. Distribuição natural do valor δ13C nas partes lenhosas da figueira ‘Roxo de
Valinhos’.. ................................................................................................................40
Tabela 5. Distribuição natural de massa seca e carbono total em figueiras ‘Roxo de
Valinhos’. .................................................................................................................43
Tabela 6. Dados morfológicos médios das plantas alimentadas com Carbono-13, no
experimento fonte-dreno...........................................................................................44
Tabela 7. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de
Valinhos’ sem a presença de drenos.........................................................................47
Tabela 8. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de
Valinhos’ com apenas brotações como drenos.........................................................50
Tabela 9. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de
Valinhos’ com frutos como drenos...........................................................................53
Tabela 10. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de
Valinhos’ com frutos e brotações como drenos........................................................56
Tabela 11. Valores de δ‰ 13C nas diferentes partições vegetativas de ramos de figueira
‘Roxo de Valinhos’ em função do tempo após enriquecimento...............................64
Tabela 12. Valores de δ‰ 13C nas diferentes partições lenhosas e no sistema radicular
da figueira ‘Roxo de Valinhos’ em função do tempo após enriquecimento. ...........65
Tabela 13. Valores da equação exponencial experimental (turnover) e da meia-vida do
carbono, em diferentes partições vegetativas de Ficus carica . ...............................70
Tabela 14. Valores estimados e mensurados de δ‰ 13C e a porcentagem de carbono-13
trocado, para as diferentes partições vegetativas de Ficus carica.. ..........................71

VI
LISTA DE FIGURAS
Página
Figura 1. Câmara de enriquecimento de 13CO2 ..........................................................................31
Figura 2. Taxa de assimilação líquida de CO2 de folhas de Ficus carica L. em função da
área foliar (padrão de medida em 6,0 cm²)...............................................................38
Figura 3. Assimilação líquida de CO2 e taxa de transpiração de uma folha de Ficus
carica L. em função da sua posição ao longo do ramo ............................................38
Figura 4. Régua isotópica apresentando os sentidos de enriquecimento de δ 13C em
relação ao padrão Lm ...............................................................................................42
Figura 5. Relação existente entre massa seca de uma dada partição e o seu carbono total
existente....................................................................................................................43
Figura 6. Valores instantâneos de temperatura e radiação solar global coletados nos dias
16 e 17 de março de 2008.........................................................................................46
Figura 7. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de
Valinhos’ sem a presença de drenos, sendo a 4ª folha alimentada...........................48
Figura 8. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de
Valinhos’ com apenas brotações como drenos, sendo a 4ª folha alimentada...........51
Figura 9. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de
Valinhos’ com apenas frutos como drenos, sendo a 4ª folha alimentada.................54
Figura 10. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de
Valinhos’ com frutos e brotações como drenos, sendo a 4ª folha alimentada..........57
Figura 11. Valores instantâneos de temperatura e radiação solar global coletados nos
dias 11, 12 e 13 de março de 2008. ..........................................................................58
Figura 12. Modelo exponencial para os isotopos estáveis do carbono na gema apical e
nos frutos de Ficus carica L., considerando apenas o ramo da folha marcada.. ......69

VII
UTILIZAÇÃO DO CARBONO-13 COMO MARCADOR NA PARTIÇÃO DE
FOTOASSIMILADOS EM FIGUEIRA. Botucatu, 2009. 94p. Dissertação (Mestrado em
Agronomia/Horticultura) – Faculdade de Ciências Agronômicas, Universidade estadual
Paulista.
Autor: ANDRÉA CARVALHO DA SILVA
Orientadora: SARITA LEONEL
Co-orientador: CARLOS DUCATTI
RESUMO
O trabalho teve como objetivo avaliar, a viabilidade da metodologia
para avaliação da translocação e alocação de fotoassimilados, utilizando o isótopo estável do
carbono-13 como marcador, assim como, avaliar a relação fonte-dreno nos diferentes órgãos
bem como, saber em que órgão se encontraria o carbono-13 após determinados intervalos de
tempo, em plantas em estádio reprodutivo da espécie Ficus carica L. Para tanto, uma folha da
figueira considerada adulta através da caracterização fotossintética com um medidor portátil
de fotossíntese IRGA-6400, foi colocada dentro de uma câmara de acrílico construída para
este fim, e submetida a um ambiente com enriquecimento de 13CO2 por 30 minutos. Após 24
horas, os diferentes órgãos presentes nas plantas foram coletados no experimento órgãos
drenos, nas demais plantas seguiu-se a experimentação tempo de alocação, onde as plantas de
Ficus carica L. foram arrancadas ao longo das horas: 6, 24, 48, 72, 120, 168, 360. Após o
tempo especificado as partes (meristema apical, folhas, ramos, caule e sistema radicular) das
plantas em estudo foram coletados e imediatamente imersos em nitrogênio líquido (-196
ºC), para evitar que os tecidos permanecessem vivos e conseqüentemente pudessem consumir
os fotoassimilados no processo da respiração. As amostras, previamente identificadas foram
secas em estufa de circulação forçada a 65º C por 72 horas, em seguida moídas em moinho
criogênico, para que houvesse uma perfeita homogeneização, sendo posteriormente analisadas
no espectrômetro de massas de razão isotópica, para a determinação do enriquecimento

VIII
relativo de 13C. Naturalmente a planta apresenta valores de enriquecimento relativo de δ 13C -
27,92‰, alocando acima de 90% da massa seca e do carbono total nas partes lenhosas. Os
resultados obtidos indicaram que figueira se caracteriza como uma planta do ciclo
fotossintético C3, tanto pelos valores da taxa assimilação líquida de CO2 quanto pelos valores
isotópicos, tendo a quarta folha como a mais fotossinteticamente ativa e principal exportadora
de fotoassimilados. A planta apresenta crescimento vegetativo e fase reprodutiva
concomitantes, existindo preferência na alocação de 13C nas partes meristemáticas, seguidas
pelos órgãos reprodutivos, com uma atividade fisiológica que possibilita a renovação do 13C
dentro de 24 horas, existindo novos ciclos de translocação entre 72 e 168 horas após o
enriquecimento.
___________________________________________
Palavras chaves: Isótopo estável, translocação de 13C, figo, reservas de carboidratos, fonte-
dreno.

IX
UTILIZATION OF THE CARBON-13 AS MARKER IN THE PARTITION OF
PHOTOSYNTHATES IN FIG TREE. Botucatu, 2009. 94p. Dissertation (Master in
Agronomy/Horticulture) – Faculty of Agronomic Sciences, state University native of São
Paulo.
Author: ANDRÉA CARVALHO DA SILVA
Adviser: SARITA LEONEL
Co-adviser: CARLOS DUCATTI
SUMMARY
The work have planned be quizzed, the feasability of the methodology
for evaluation of the fhotosynthates translocation and allocation, using the carbon 13 stable
isotope a marker, evaluate the source-sink relation in the different organs and, know in that
organ would be found the carbon-13 after determined time periods, in Ficus carica L. plants
in reproductive stadium. A fig tree leaf considered adult through the photosynthetic with a
portable meter of photosynthesis IRGA-6400 characterization. Was put inside a acrylic
chamber, and submitted to an environment with 13CO2 enrichment for 30 minutes. After 24
hours, the different organs present in the plants were collected in the drains organs experiment,
in the other plants followed the allocation time experimentation where the plants of Ficus
carica L. were lurches to the long one of the hours: 6, 24, 48, 72, 120, 168, 360. After the
time specified the studied plant parts (meristem, sheets, branches, stem and system roots) were
collected and immediately immersed in liquid nitrogen (-196 ºC), for avoid that the fabrics
remained alive and consequently could consume the fhotosynthates in the breath trial. The
samples, previously identified were droughts in of circulation forced to 65º C for 72 hours,
right away ground in cryogenic mill, for that had a perfect homogenization, being
subsequently analyzed in the mass spectrometry of isotopic reason, for the relative enrichment

X
13C determination. Naturally the plant presents values of 13C relative enrichment of -27,92 ‰,
allocating more than 90% of the dry mass and the total carbon in the woody part. The results
obtained indicated that fig tree is characterized like a plant of the C3 cycle photosynthetic,
because the values of the CO2 rate liquid assimilation and by the isotopes values, having the
four leaf as the most photosynthetic active and main exporter of 13C-photosynthates. The
plant presents vegetative growth and reproductive phase concomitant, existing preference in
the 13C allocation in meristems areas, succession by reproductive organs, with an physiologic
activity that enables the renewal of the 13C in 24 hours, existing new translocation cycles
between 72 and 168 hours after the enrichment.
___________________________________________
Keywords: stable isotope, 13C-translocation, fig tree, carbohydrate reserves, source-sink.

1
1. INTRODUÇÃO
A figueira (Ficus carica L.) é uma das mais antigas frutíferas
cultivadas no mundo, originária da Ásia menor e da Síria, na região mediterrânea,
apresentando excelente adaptação a diferentes climas, sendo cultivada tanto em regiões sub-
tropicais quentes, como em regiões de clima temperado, sendo introduzida no Brasil pela
primeira expedição colonizadora no ano de 1532 (ABRAHÃO et al., 1990).
O figo é cultivado em cerca de 40 países, tendo como principais países
produtores a Turquia, Portugal, Grécia, Itália, Espanha, Argélia e Marrocos. O Brasil é o
maior produtor e o segundo maior exportador de figo in natura no mundo, superado apenas
pela Turquia (VALLI, 2002; FAO, 2003; IBRAF, 2005).
No Brasil, a figueira é cultivada com o emprego de uma única
variedade, a ‘Roxo de Valinhos’, caracterizada por apresentar grande valor econômico,
rusticidade, elevado vigor e produtividade, além de boa adaptação a podas drásticas. Seus
frutos podem ser utilizados tanto para consumo in natura como para a indústria (MAIORANO
et al, 1997; PENTEADO, 1999).
A colheita brasileira do figo ocorre num período de entressafra da
produção da fruta fresca no Hemisfério Norte e nos demais países do Mercosul. Assim, são
amplas as possibilidades de exportação, pois o produto brasileiro entra no mercado

2
internacional a partir de dezembro, logo após a safra dos países mediterrâneos (FRANCISCO
et al., 2005).
Conforme dados do Ministério da Agricultura (2008), o Brasil
produziu 23.627 toneladas de figos em 2005, numa área de 2.911 ha, resultando numa média
de produtividade nacional de 8,14 t/ha. Apesar do cultivo ser bastante antigo, o Brasil não
apresenta boa produtividade e a área colhida diminuiu em mais da metade, passando de 5 mil
para pouco mais de 2 mil hectares, entre as décadas de 70 e 90 (IBGE, 1996).
Atualmente, os maiores estados produtores brasileiros são Rio Grande
do Sul, São Paulo e Minas Gerais. No Estado de São Paulo, são produzidas cerca de 7 mil
toneladas (IBRAF, 2005), respondendo por todas as exportações brasileiras de figo maduro, as
quais, na média dos últimos cinco anos, foram de 837 t/ano (MINISTÉRIO DA
AGRICULTURA, 2008). Na região de Campinas destaca-se, principalmente o município de
Valinhos, onde a cultura se desenvolveu inicialmente no estado, e concentra mais de 80% da
produção paulista de figo.
Apesar da figueira encontrar condições satisfatórias para o seu
desenvolvimento, seu cultivo vem sendo feito de maneira tradicional, sem muitas inovações
ou melhorias técnicas (GIACOBBO et al., 2007). Todavia, as perspectivas e possibilidades de
expansão do cultivo da figueira no Estado de São Paulo são promissoras, com um potencial
expansivo de produção no interior paulista, principalmente em função da boa adaptação da
cultura e da proximidade do mercado consumidor, além das significativas exportações de figo
ao natural.
Os estudos sobre a economia de carboidratos, para a produção de
espécies hortícolas são de grande importância para a agricultura, particularmente de frutas,
devido ao potencial de modificação na alocação de carbono na planta, com reflexos no
aumento ou diminuição da produção de frutos comerciais. Estas alterações são diretamente
influenciadas diretamente pelas práticas culturais de uma dada cultura que causam efeitos
significativos na translocação e alocação de carbono fixado durante o processo fotossintético.
Esses efeitos visam a obtenção de frutos em quantidade e qualidade, sem alternância de
produção.
O estudo de translocação e alocação de fotoassimilados em plantas
iniciaram-se na década de 60, com o uso do carbono radioativo 14 (XAVIER et al., 2007),

3
contudo, problemas relacionados a legislação e risco de contaminação humana limitaram o uso
dessa técnica. Nesse contexto, a utilização dos isótopos estáveis, por serem de ocorrência
natural no ambiente substituíram o uso do radioisótopo 14C e passaram a ser uma boas
ferramentas nos estudos como marcadores naturais para indicação de processos metabólicos.
Como os frutos da figueira são comercializados na forma in natura, a
sua qualidade interna e externa, assume grande importância. Dessa forma, maior
conhecimento sobre os parâmetros fisiológicos direta ou indiretamente relacionados com a
produção e qualidade dos frutos, como por exemplo, a atividade fotossintética, as relações
fonte-dreno e alocações de carbono da planta, são fundamentais.
Desse modo, este trabalho teve como objetivo caracterizar as trocas
gasosas da figueira cv. Roxo de Valinhos em condições de campo e determinar o tempo e o
direcionamento da translocação e a alocação dos fotoassimilados, usando o 13C isótopo estável
como traçador.

4
2. REVISÃO DE LITERATURA
2.1. A fruticultura como atividade agrícola
De acordo com Nachreiner e Santos (2002), Conhecer o mercado
internacional vem sendo um desafio para os produtores brasileiros de frutas frescas que, cada
vez mais, querem conquistar uma fatia desse atraente canal de comercialização. O Brasil, em
função de suas condições climáticas, apresenta um enorme potencial para se tornar um dos
maiores pólos produtivos de frutas frescas para o mercado mundial, aproveitando a “onda
naturalista” que o mundo atravessa. Os Estados Unidos, apesar de ser um importador em
potencial, não são um grande comprador da fruta brasileira, que, na maioria dos casos, não
consegue ultrapassar as barreiras – legislativas e sanitárias – definidas por aquele país
conforme demonstra a participação do Brasil nas importações norte-americanas (FAO, 2003).
Geradora de emprego, renda e de alimentos com elevada qualidade
nutricional, a fruticultura tem se consolidado como atividade lucrativa e de sustentabilidade
alimentar em todo o território nacional. No entanto, à medida que se incentiva a expansão da
fruticultura, principalmente visando o aumento da competitividade de seus produtos (in natura
ou processados), é necessário propiciar aos produtores, (grandes, médios ou pequenos)

5
informações tecnológicas e de mercado que os auxiliem no desenvolvimento e consolidação
desta atividade agrícola.
De acordo com o Ministério da Agricultura (2008) e o Instituto
Brasileiro de Geografia e Estatística (2004), a fruticultura é estratégica para o agronegócio
brasileiro. Com um superávit de US$ 267 milhões em 2003, o setor ocupa uma área de 3,4
milhões de hectares. A produção de frutas permite obter um faturamento bruto entre R$ 1 mil
e R$ 20 mil por hectare. Hoje, o mercado interno absorve 21 milhões de toneladas/ano e o
excedente exportável é de cerca de 17 milhões de toneladas. Com uma fruticultura
diversificada, o Brasil é um dos maiores pólos mundiais de produção de sucos de frutas. No
ano passado, as exportações do setor alcançaram US$ 1,25 bilhão. Do total, 95,5%
corresponde ao suco de laranja, do qual o país é o maior produtor e exportador. O setor gerou
receitas cambiais de US$ 1,2 bilhão em 2003, um resultado 14,6% acima do valor vendido ao
mercado externo em 2002. Os principais destinos foram Bélgica, Países Baixos, Estados
Unidos e Japão. O Brasil é o terceiro pólo mundial de fruticultura, com uma produção anual de
cerca de 38 milhões de toneladas. Em 2003, as vendas externas de frutas frescas alcançaram
US$ 335,3 milhões, com um aumento de 39% em comparação aos US$ 241 milhões obtidos
em 2002. Consciente do enorme potencial do país na área de fruticultura, com plenas
condições de ampliar sua participação do mercado internacional, o Ministério da Agricultura,
Pecuária e Abastecimento e os produtores no setor estão investindo em um sistema de cultivo
de frutas de alto padrão de qualidade e sanidade. É o programa de Produção Integrada de
Frutas (PIF), que prevê o emprego de normas de sustentabilidade ambiental, segurança
alimentar, viabilidade econômica e socialmente justa, mediante o uso de tecnologias não
agressivas ao meio ambiente e ao homem. As frutas cultivadas no sistema de produção
integrada vão para o mercado com um selo de conformidade, atestando a sua qualidade e
sanidade. Desde que foi implantada, a PIF permitiu uma redução de 63% no uso de
agrotóxicos nos pomares de manga; de 50% no mamão; de 32% na uva; e de 30% na maçã.
Para Buainain e Batalha (2007), o Brasil, apesar de ser considerado
grande produtor de frutas, ocupando a terceira posição na produção mundial, possui baixo
consumo per capita de frutas frescas. O consumo nacional encontra-se em torno de 57,0
Kg/ano, valor considerado baixo em relação a outros países como a Espanha (120,0 Kg/ano),
Alemanha (112,0 Kg/ano), Estados Unidos (67,0 Kg/ano) e Japão (61,8 Kg/ano). Cabe

6
ressaltar que a Organização Mundial de Saúde (OMS) recomenda um consumo de 146 kg de
frutas/ano.
2.2. Situação econômica da ficicultura
A produção de figo nos países localizados na Bacia Arábica do
Mediterrâneo representa a maior parte da produção mundial. Em 2004, a Turquia respondeu
por 26% do total; o Egito, por 18%; a Grécia e o Irã, por 7% cada um; Marrocos e Espanha,
por 6% e o Brasil, na 10a colocação, por 2% da produção mundial (FAO, 2004).
O figo está entre as vinte principais frutas exportadas pelo Brasil e vem
mantendo a terceira posição no ranking de volume comercializado, entre as frutas de clima
temperado, com 0,9 mil toneladas. Fica atrás da maçã com 153,0 mil toneladas e da uva com
28,8 mil toneladas, atingindo o patamar de US$ FOB 2,109 milhões (SECEX, 2004). Os
maiores importadores do figo brasileiro são Alemanha, França, Países Baixos, Reino Unido e
Suíça, dentre mais de dez países para onde, costumeiramente, são feitos embarques aéreos. No
Estado de São Paulo, os municípios produtores de maior importância são os de Valinhos,
Campinas, Louveira e Bragança Paulista, com a principal produção voltada para mesa,
destinada tanto ao mercado interno quanto ao externo. No ano de 2004, essa região exportou
40% da sua produção de figo.
Embora haja países com grandes produções, estas se destinam
principalmente ao mercado interno, ficando o Brasil (figo tipo comum: Roxo de Valinhos) e a
Turquia (tipo polinizado, Smirna) como importantes fornecedores de figo ao mercado
internacional.
2.3. A figueira
A figueira é chamada em espanhol de higuera, em francês de figuier;
em árabe, kerma; em inglês, common fig; fig em italiano. É uma das frutas mais antigas de
que se tem relato, foi encontrada em uma pintura egípcia de Beni-Hassan uma coleção de
figos, pintura esta datada de 4500 anos A.C, também é mencionado no Antigo Testamento da
Bíblia Sagrada como sendo um dos quesitos de abundancia da terra prometida

7
(DOMINGUEZ, 1990). Assim como o óleo de oliva, os pães ázimos, o figo constitui um dos
alimentos mais populares que sustentam a humanidade desde o começo de sua história. Os
figos foram provavelmente uma das primeiras frutas a serem secadas e armazenadas pelo
homem. O povo da cidade de Ática conhecida por seus figos considerava tal fruto como sendo
sagrado, fato este que se difundiu para todos os países do Sudoeste da Ásia, no Egito na
Grécia e na Itália.
A figueira cultivada possui o nome botânico de Ficus carica L., e
pertence a família botânica, das Moráceas. O gênero Ficus engloba cerca de mil espécies,
sendo muitas destas espécies largamente usadas como plantas ornamentais. A figueira possui
uma boa adaptabilidade climática por isso é encontrada tanto nos regiões subtropicais quentes
como nas regiões de clima bem temperado, contudo segundo Penteado (1999), as melhores
condições para a cultura são o inverno frio, chuvas bem distribuídas e umidade atmosférica
baixa.
A figueira é uma planta de folhas caducas, nativa da região
Mediterrânea, onde predomina o clima subtropical temperado.
No Brasil e provavelmente no estado de São Paulo, a figueira tenha
sido introduzido com o advento da primeira expedição colonizadora de Martin Afonso de
Souza, no ano de 1532 (RIGITANO, 1955). Somente a partir de 1910 passou a ser cultivada
comercialmente na região compreendida pelo antigo distrito de Valinhos, ainda pertencente na
época a cidade de Campinas. Hoje o município de Valinhos é famoso e conhecido como a
capital nacional do Figo Roxo.
Penteado (1999) traz relatos de que o introdutor do Figo Roxo em
Valinhos tenha sido o Sr. Lino Busatto, imigrante italiano, que chegou por volta de 1898, e
teve a iniciativa de mandar vir de uma região da Itália, próxima ao Mar Adriático, algumas
mudas de figueiras, que ao chegarem encontraram fácil adaptação; tratadas com cuidado
prosperaram, e seus figos, de coloração roxo-escura, tornaram-se desde então conhecidos
como “Roxo de Valinhos”.

8
2.4. Caracterização Botânica
A figueira comum, como citado anteriormente, Ficus carica L.,
pertence à família Moraceae e apresenta um número diplóide (2n) de cromossomas igual a 26.
A família Moraceae inclui 60 gêneros e mais de 2000 espécies de árvores, arbustos,
trepadeiras e pequenas ervas. A espécie Ficus carica L. pertence ao subgênero Eusyce, que é
caracterizado por apresentar flores unissexuais e por ginodioicismo.
Segundo Dominguez, (1990), por ser uma espécie muito difundida e
estar cultivada em distintas situações, pode existir uma ampla gama de variedades, com
morfologia e aspectos de árvores diferentes quanto ao comportamento natural. Podendo em
zonas favoráveis alcançar de 8 a 10 metros de altura, contrariamente não passa de formas
arbustivas em zonas consideradas desfavoráveis com invernos e secas prolongadas que
condicionam o mau crescimento.
Em São Paulo, no entanto, as técnicas culturais utilizadas,
especialmente as podas anuais de inverno, condicionam as plantas a um porte arbustivo, com
longevidade econômica de cerca de 30 anos, longevidade esta, considerando um pomar bem
manejado e sem a incidência de nematóides ou brocas.
Seus ramos, normalmente possuem um grande diâmetro nos países
europeus, contudo no Brasil são considerados finos, estão cobertos por uma casca fina e sem
rugosidades, que em principio são verdes, tornando-se de coloração cinza pálida, todas as
partes verdes da planta contém um látex leitoso de coloração esbranquiçada, que se espessa
quando em contato com o ar. Nesse látex existe uma enzima proteolítica denominada de
ficina, que causa irritação na pele, quando em contato com a mesma, principalmente durante a
desbrota e a colheita e embalagem dos frutos.
O sistema radicular da figueira caracteriza-se como fasciculado. Não
havendo predominância de uma raiz principal, tendo várias em torno do tronco, com
disposição radial e facilmente visível em exemplares adultos. As raízes são bastante
superficiais, fibrosas, abundantes e muito frágeis, por volta de 80% delas se encontram nas
profundidades entre 20 e 45 cm. Utilizando-se cobertura morta para a proteção do solo, podem
apresentar raízes com comprimento superior a 8 metros. O sistema radicular da figueira é

9
considerado um órgão de extrema importância, pois é ele quem armazena as reservas
(carboidratos), durante o inverno, período em que a planta permanece em repouso vegetativo
(MATSUURA et al., 2001).
O tronco possui formação muito variada e madeira pouco densa. A
inserção dos ramos primários, secundários e ramos produtivos são grossos, com tendência a
formar arcos, contudo são pouco visíveis no sistema de produção que exige podas drásticas.
As folhas da figueira são grandes, palmadas, alternas e com grandes
estípulas. Inserem-se em um pecíolo largo e grosso. Seu contorno esta marcado por um
numero de lóbulos interiores entre três e sete, profundamente marcados, que, segundo
Dominguez, (1990), servem de base para definir varietalmente um individuo. Apresentam
coloração verde intenso e brilhante, são ásperas ao tato por sua pilosidade rígida. Existem
variedades praticamente isentas de aspereza, recordando as outras espécies de figueiras, dando
por esse motivo bases para sua classificação varietal. As nervuras principais são também em
forma palmeadas, muito marcadas assim como as nervuras secundárias, a coloração é menos
intensa do que a coloração da folha, porém contém a mesma aspereza.
As gemas frutíferas e vegetativas são grandes e muito pontiagudas,
aparecem nos ramos, junto ás axilas das folhas, durante a fase vegetativa (crescimento), em
geral as figueiras apresentam duas séries de gemas frutíferas em cada nó, o que pode resultar
em duas colheitas distintas (principal e bredas).
As flores da figueira apresentam características particulares, sendo
unissexuadas. As flores femininas se encontram repartidas dentro de um receptáculo carnoso e
lobular, cuja única saída para o exterior é o ostíolo onde se inserem as flores masculinas
pomologicamente chamadas de sicônio, que nada mais é do que o próprio figo, havendo duas
distintas formas de plantas, o caprifigo, que é monóico, e o figo, que é pistilado. As flores do
figo são pequenas, pediceladas, hipóginas e com perianto simples pentapartido. Existem três
tipos de flores: as pistiladas (femininas) com estilo curto, as pistiladas femininas com estilo
longo e as estaminadas (masculinas).
De acordo com Pereira e Nachtigal, (1999), ambas as flores pistiladas
são simples, carpeladas e com um estigma bífido. As flores pistiladas (femininas) de estilo
curto apresentam um ovário, aproximadamente, globoso e um estilo em cerca de 0,7 mm de
comprimento, sendo adaptadas a ovoposição da vespinha-do-figo (Blastophaga psenes,

10
Cavalini). As flores de estilo longo apresentam um ovário mais ou menos ovóide ou elipsóide
e o estilo com 1,75 mm de comprimento, não adaptado a ovoposição da vespinha-do-figo.
O tipo de fruto é o sicônio, ou seja, um fruto carnoso agregado, no qual
os ovários são originados de um aumento na cavidade do receptáculo (a fruta, comumente
chamada de figo, não é, pois um fruto, mas uma infrutescência). Na parte terminal do fruto
existe um orifício, que liga a cavidade do receptáculo com o exterior (SIMÃO, 1971).
Os frutos verdadeiros da figueira são os aquênios, que se formam pelo
desenvolvimento dos ovários. Os aquênios normais apresentam um embrião envolvido pelo
endosperma e pelo tegumento. Os figos não polinizados podem apresentar aquênios com o
ovário esclerificado, porem ocos. A parte suculenta do figo comestível consiste,
principalmente, de tecido parenquimatoso dos órgãos florais, cujas células se tornam maiores e
armazenam substancias de reserva.
O cultivo desta importante fruteira, no Brasil baseia-se unicamente na
plantação de um único material, a variedade Roxo de Valinhos. De acordo com dados oficiais
é uma variedade muito cultivada e conhecida em outros países, com as seguintes
denominações: San Piero, Borwn Turkey, Negro Largo, Douro Blank, Negro d’Espagne,
Aubique Nooire, Portugal, Albicone, Rubicone e Brunswich.
A variedade Roxo de Valinhos é caracterizada como do tipo comum,
formada partenocarpicamente, dispensando o estimulo da caprificação (polinização) e da
formação de sementes. O figo é uma fruta de tamanho grande, quando destinada ao consumo
“in natura”, podendo pesar de 70 a 100 gramas, piriforme alongado, com pedúnculo curto. Na
parte basal do fruto fica o ostíolo, conhecido como “olho” do figo, muito aberto, podendo
apresentar o inconveniente de facilitar a entrada de fungos e insetos. A coloração externa é
roxo-escura e a polpa, rosada-violácea. Apresenta uma cavidade central ampla. Os figos
quando maduros são destinados ao mercado de mesa; o figo meio maduro, para a produção do
doce de figo, os figos verdes pesando por volta de 20 a 30 gramas são empregados para
produção de doces cristalizados ou compotas, para fins de industrialização, outros tipos como:
“inchado”, constituído por figos já bastante desenvolvidos, pesando 40 a 50 gramas e
apresentando inicio de coloração roxo-avermelhada.
Para Penteado (1999), a colheita deve ser realizada no estádio de
maturação designado de verde-arroxeado, porém fisiologicamente maduros, ou seja, quando se

11
apresentam completamente inchados, com coloração verde-escura modificada para roxo-
bronzeado, estando na fase inicial de perda de consistência firme. O roxo de Valinhos quando
maduro, deteriora-se com facilidade, principalmente quando amadurece em épocas chuvosas e
quentes, por isso deve ser colhido logo que atinja o ponto de maturação, também designado
“de vez”.
2.5. Ecofisiologia da Figueira
Quanto à ecofisiologia da figueira pode-se dizer que é uma árvore
pouco exigente em solos, cresce bem em lugares pedregosos, áridos e até em solos pouco
profundos, no entanto prefere solos com boa permeabilidade, férteis e bem drenados Para
produzir frutos de qualidade requer solos ricos em cálcio, e que não sejam excessivamente
úmidos. Podendo ser solos arenosos de pH ligeiramente ácido a neutro (sendo o ideal para a
cultura em torno de 5,6 a 6,8.) A figueira adapta-se bem a diversos tipos de solos, porém de
maneira geral, os solos mais apropriados são os solos areno-argilosos, bem drenados e com
bom teor de matéria orgânica (BOLIANI e CORRÊA, 1999).
Dominguez, (1990), relata que a aeração dos solos é fundamental, pois
a figueira é uma planta muito sensível a podridões radiculares, não suportando solos
encharcados nem a umidade em excesso, porém é uma planta bastante resistente a salinidade,
podendo em situações limites de produção suportar águas que contenham até 2 g de cloreto de
sódio/litro de água.
O sucesso da cultura é limitado mais pela baixa temperatura de inverno
que pelas altas temperaturas de verão, as plantas estão sujeitas aos danos causados pelas
geadas, no estádio vegetativo, resistindo bem quando em dormência (SIMÃO, 1998).
Entre os fatores climáticos, merecem destaque a temperatura,
precipitação, vento, umidade relativa e luz. A figueira tolera temperaturas de 35 a 42 °C.
Temperaturas acima de 40º C antecipam a maturação dos frutos, alterando também a
consistência da casca, que fica dura e coriácea (SIMÃO, 1998).
Nos trópicos, as plantas se desenvolvem rapidamente, porém só
produzem bem em altitudes entre 900 e 1500 m. Figos do tipo comum crescem bem em

12
regiões subtropicais ou considerados semitemperadas. Simão, (1998), reporta que a figueira
adulta resiste bem a temperaturas até -1,5º C, entretanto os brotos são bastante sensíveis. A
exigência em frio hibernal para quebra de dormência das gemas varia de 100 a 300 horas de
frio (abaixo de 7,2º C). Apesar disso, há boa adaptação da figueira em regiões de clima quente,
com a vantagem adicional de poder-se produzir frutas durante o ano todo, visto que a irrigação
e a poda condicionam a frutificação. Nas regiões quentes, as safras são maiores e os figos,
mais doces.
A cultura exige, no período vegetativo, chuvas freqüentes e bem
distribuídas, sendo adequadas precipitações em torno de 1200 mm anuais. Antunes et al.
(1997) relata que o emprego da cobertura morta do solo do pomar permite preservar a
umidade do solo, fundamental para o bom desenvolvimento da figueira. Em locais com
precipitações irregulares, pequenas estiagens são sentidas pelas plantas, causando a queda das
folhas, com prejuízos à produção. Neste caso a cultura deve ser irrigada. Por outro lado, a alta
umidade pode predispor as frutas ao ataque de doenças bem como causar rachaduras nas frutas
quando elas se encontram no estádio de maturação (SIMÃO, 1998).
Com relação aos ventos os mesmos não costumam causar danos à
figueira, a não ser quando excessivamente fortes. Durante o desenvolvimento dos figos, até a
época de maturação, ventos fortes causam o contato das folhas com os frutos, produzindo
nestes grandes escoriações, que os depreciam (PEREIRA, 1981). Nos locais muito sujeitos a
ventos fortes, pode-se recomendar a instalação de quebra-ventos.
A figueira em ambiente altamente iluminado, apresenta um
crescimento vigoroso e produz frutas de excelente qualidade. A coloração e a forma das frutas
são afetadas pelo clima (luz, temperatura, umidade) e outros fatores (SIMÃO, 1998).
2.6. Fotossíntese e Reserva
Dos fatores envolvidos na produtividade agrícola, a fotossíntese é um
dos mais determinantes. A elevação das taxas de fotossíntese depende, dentre outros fatores,
do máximo aproveitamento da luz disponível, o qual pode ser obtido por tratos culturais e
manejos. As formas de manejo influem no número de plantas da população adequadas ao

13
objetivo da exploração, arranjos foliares mais erectófilos, disposição das linhas de plantio na
direção norte-sul e técnicas de manejo da copa, tais como podas, desfolhamento e modificação
da arquitetura da planta (JACKSON, 1980; BERNARDES, 1987).
A produtividade é influenciada por características morfológicas e
fisiológicas da fonte (órgãos fotossintetizantes) e do dreno (órgãos consumidores dos
metabólicos fotossintetizados e carboidratos principalmente). Toda produção de fitomassa
depende da atividade fotossintética da fonte, sendo a assimilação do CO2 um dos muitos
fatores que influenciam o crescimento e desenvolvimento vegetal (FOYER e GALTIER,
1996). Desta forma, buscar mais informações sobre a fisiologia da fonte torna-se de
fundamental importância, e uma forma muito utilizada para estudá-la é por meio de medidas
de trocas gasosas.
Os açúcares provenientes da fotossíntese agem como substrato para o
metabolismo energético e biossíntese de hidratos de carbono, fornecendo condições de
crescimento e desenvolvimento aos tecidos dreno. Além disso, os açúcares podem funcionar
como mensageiros secundários assegurando que a planta continue a se desenvolver, mesmo
após estresses bióticos ou abióticos (HAMMOND e WHITE, 2008).
O amido e a sacarose são fotossintatos de grande importância
acumulados pelas plantas. O amido é o carboidrato de reserva mais abundante nas plantas e, é
encontrado em folhas, diferentes tipos de hastes e raízes, assim como em flores frutos e
sementes, onde é utilizado como fonte de energia durante períodos de dormência, estresse ou
início de crescimento (LAPOINTE, 1998). Alguns autores descrevem o amido como sendo a
principal reserva de carbono utilizado na síntese de sacarose, um açúcar predominante em
frutas após o amadurecimento (BERNARDES-SILVA et al., 2003). As conversões de amido
para açúcar ocorrem em ocasiões de crescimento vegetativo intenso, quando as reservas são
usadas para suportar a atividade meristemática de ápices caulinares e também o crescimento
de frutos (PRIESTLEY, 1963). Em embriões de legumes a sacarose é o açúcar primário que é
capturado por sistemas saturados e insaturados (ZAMSKI, 1995), sendo a sacarose, o principal
fotossintato translocado (HARTT e KORTSHACK, 1964), em plantas superiores a partir dos
tecidos fonte para os tecidos drenos para promoção do desenvolvimento da planta
(ECKARDT, 2003).

14
Os solutos translocados no floema são principalmente carboidratos,
sendo a sacarose o açúcar mais comumente translocado. Nos drenos, os açúcares
transportados, são alocados para os processos de crescimento ou reserva (COLL et al., 2001;
KOZLOWSKI e PALLARDY, 1997). O transporte de fotoassimilados até os vasos
tranportadores é realizado em forma de sacarose, ou de compostos provenientes da rafinose,
dependendo da espécie da planta e do tipo de carga e descarga do floema (via simplasto e
apoplasto) (TOFIÑO et al., 2006).
A luz é importante para a produção de frutos, pois todos os aspectos do
crescimento da planta e dos frutos e o desenvolvimento de gemas floríferas requerem
carboidratos que são produzidos pela fotossíntese nas folhas (RAJAPAKSE et al., 1999;
MARINI, 2002). Na transição para o florescimento, ocorre aumento no suprimento de
carboidratos nas gemas vegetativas, pelo aumento da atividade fotossintética e hidrólise do
amido. A sacarose é acumulada no meristema para o fornecimento de energia para o processo
de ativação mitótica (BODSON e OUTLAW JUNIOR, 1985).
O sombreamento leva à redução da fotossíntese total da planta, pois,
folhas sombreadas apresentam menores taxas fotossintéticas e assim contribuem menos ou
negativamente para a produção da planta do que as folhas expostas ao sol. Folhas de sol
caracterizam-se, entre outras, pelo maior teor de clorofila e rubisco, maior densidade
estomática, menor área foliar e maior peso de folha por superfície (BERNARDES, 1987;
LARCHER, 2000).
Em experimento realizado por Caetano et al. (2005) a penetração da
radiação na copa das figueiras foi avaliada nas leituras do fluxo de fótons fotossintéticos
(µmol.m2.s-1), onde se observou redução linear no fluxo de fótons fotossintéticos (Y= -60.192
X + 2321, R2 = 0,94) no interior da copa das plantas com o aumento do número de ramos
produtivos conduzidos, o que indica a ocorrência de auto-sombreamento. O aumento do
número de ramos conduzidos não elevou a produtividade de figos verdes de forma crescente,
pois o auto-sombreamento proporcionado por uma estrutura de copa com mais ramos diminuiu
o número frutos formados. A maior produtividade observada de figos verdes foi obtida quando
as plantas foram conduzidas com 24 ramos, sendo que, neste tratamento, a área foliar média de
cada planta foi de 6,2 m2.

15
A produção fotossintética não aumenta indefinidamente com o IAF
(Índice de área foliar), sendo limitada pelo sombreamento que as folhas superiores exercem
sobre as inferiores. O auto-sombreamento no dossel provoca decréscimo na taxa fotossintética
média em função do aumento do IAF e reduz a formação de gemas reprodutivas (JACKSON,
1980; BERNARDES, 1987; LUCCHESI, 1987).
A poda insuficiente em fruteiras resulta em aumento da estrutura
vegetativa da planta. Este crescimento causa pesado sombreamento e as gemas frutíferas
param de se desenvolver. Desta forma, em poucos anos, as gemas frutíferas se desenvolverão
somente no topo e lados da planta onde há incidência da luz solar. Podando-se uma larga
porção das folhas e ramos dentro da copa, aumenta a penetração da luz solar, que promoverá o
desenvolvimento de gemas frutíferas no interior da área da planta (KADIR, 2003). A
utilização da poda permite, portanto, adequar a copa da planta para obtenção de máxima
produtividade e qualidade do produto colhido.
Os estômatos atuam como reguladores da perda de água pela
transpiração, respondendo ao déficit hídrico com a alteração da abertura do poro a uma faixa
crítica de valores do potencial hídrico foliar (LARCHER, 2000).
A condutância estomática varia com a espécie/cultivar, a idade da
folha e com o pré-condicionamento das plantas (DAÍ et al., 1992). Essas variações afetam o
uso da água, haja vista que a taxa de assimilação de CO2 e de transpiração respondem
diferentemente à abertura dos estômatos (MACHADO e LAGÔA, 1994).
A transpiração resulta da difusão de vapores de água através dos
estômatos abertos, mas quando estes estão fechados, se estabelece nas plantas certa resistência
à perda de água, com reflexos sobre as atividades metabólicas. Calbo e Moraes (1997)
observaram que a transpiração de folhas de plantas de buriti (Mauritia vinifera Mart.) foi
reduzida ao nível de 10%, após o sexto dia de suspensão da irrigação, e que a condutância
estomática atingiu valor próximo de zero, indicando haver certa correlação entre essas
variáveis biofísicas.
No caso das plantas frutíferas, o fruto é o dreno de importância
econômica, de forma que a relativa partição de matéria seca direcionada para o fruto irá
determinar, em parte, a sua qualidade final. Contudo, o potencial da quantidade de

16
fotoassimilados que podem ser transportados para os drenos está diretamente ligado à
atividade fotossintética de uma fonte produtora de fotoassimilados (ZAMSKI, 1996).
Segundo Larcher (2000), em espécies lenhosas, a formação das flores,
a freqüência do florescimento, a quantidade de frutos e o amadurecimento das sementes são
regulados por uma combinação de fatores nutricionais, alocação de assimilados e mecanismos
endógenos de controle.
Toda produção de biomassa depende da fotossíntese. Plantas
assimilam o CO2 da atmosfera e reduzem ao nível de triose-fosfato, a qual pode então ser
usada para produzir carboidratos, principalmente sacarose e amido. A assimilação
fotossintética do carbono é apenas um dos fatores que influenciam o crescimento e
desenvolvimento da planta. Outras etapas críticas são a síntese e transporte de sacarose, a
partir do mesófilo foliar, o carregamento do floema e a partição na planta (FOYER e
GALTIER, 1996).
Segundo Pimentel (1998), quando o carbono é fixado no cloroplasto,
por plantas do ciclo fotossintético C3 cerca de 70 a 80% da triose-P formada é reciclada para a
regeneração da enzima de carboxilação (ribulose-1,5 difosfato), sendo o restante utilizado para
a síntese de amido (transitório) no cloroplasto, e outros carboidratos solúveis, no citossol ou
vacúolo. Estes apresentam diferentes funções fisiológicas como armazenamento, translocação
e utilização do carbono, além de promover a proteção da planta a vários tipos de estresses,
como por exemplo, à salinidade, à seca e as altas e baixas temperaturas.
As fontes normalmente são órgãos que atingiram um grau de
desenvolvimento que lhes permite absorver quantidades adequadas de água e nutrientes pela
corrente transpiratória e ter uma fotossíntese líquida capaz de torná-los autotróficos, enviam
fotoassimilados para drenos com os quais mantêm relações vasculares diretas (PRESTON,
1998).
Na transição da folha de um órgão dreno para fonte, mudanças
ontogênicas ocorrem durante o desenvolvimento, como reflexo da interação da planta com as
condições ambientais. O inicio da fotossíntese, durante a ontogênese da folha, requer a
coordenação de numerosos eventos, que são modificados por controles endógenos e
ambientais. Dois eventos assumem importância primordial nas interações fonte-dreno: o

17
primeiro está relacionado com desenvolvimento da capacidade fotossintética da folha e o
segundo com os comportamentos como importadores ou exportadores dos tecidos, associados
às mudanças no metabolismo dos carboidratos. O desenvolvimento da capacidade
fotossintética está associado com a emergência da folha. Na ausência de luz não ocorre à
síntese de pigmentos fotossintéticos, após a sua emergência, a folha intercepta a luz e muito
rapidamente os pigmentos são sintetizados. Embora exista considerável variação entre
espécies, o máximo da capacidade fotossintética ocorre antes ou no momento da plena
expansão da folha, freqüentemente antes do conteúdo máximo da clorofila ter sido atingido,
podendo ser citado o exemplo da beterraba onde suas folhas atingem o máximo de sua
capacidade fotossintética quando apresentam apenas 22% de seu comprimento final (FOYER
e GALTIER, 1996).
A maioria dos carboidratos acumulados nos sítios de estocagem é
translocada durante a estação de crescimento e este fenômeno está associado à presença de
drenos fortes. A sacarose é o carboidrato mais comumente translocado (pelo floema) entre as
fontes, órgãos exportadores de C, e os drenos, órgãos consumidores de C.
Em relação à fonte, de acordo com Pimentel (1998), as plantas que
apresentarão uma grande área foliar podem interceptar mais energia luminosa, porém também
apresentará uma grande superfície de transpiração, o que é indesejável principalmente para
plantas C3 que têm baixa eficiência no uso da água. Como a energia luminosa não é um fator
limitante para a agricultura em regiões tropicais, deve ser feita à seleção de plantas que
apresentem menor área especifica de folhas, ou seja, plantas com maior peso de folha por
unidade de área. Como a atividade fotossintética é função do número de cloroplastos, seja
disposta horizontalmente (maior área foliar) ou verticalmente (maior espessura e área
especifica), uma folha mais espessa e menos larga (permitiria o adensamento de plantio)
manterá com isso alta atividade fotossintética por unidade de área, com menor superfície de
transpiração.
Os fotoassimilados seguem pelo floema por fluxo de massa até a
região do dreno. Existe uma pressão entre a fonte e o dreno, gerada pelo carregamento de
sacarose na fonte e descarregamento no dreno (COLL et al., 2001; HOPKINS, 1995). O
carregamento de sacarose na fonte provoca um aumento no potencial osmótico, levando à
entrada de água, conseqüentemente aumento do potencial de pressão. No dreno, o

18
descarregamento de sacarose leva a uma redução no potencial osmótico com a conseqüente
saída de água e redução do potencial de pressão. Deste modo forma-se um gradiente de
pressão entre a fonte e o dreno que determina o transporte da seiva por fluxo de massa (TAIZ
e ZEIGER, 2004). A importância relativa dos drenos muda ao longo do ciclo da planta de
acordo com a distribuição espacial dos tecidos em crescimento, durante a fase vegetativa os
meristemas: apical e radicular são mais importantes, mas durante a fase reprodutiva os frutos
se tornam os drenos preferenciais (HOPKINS, 1995).
Lai et al. (1988), relatam que não foi observada a translocação dos
fotoassimilados produzidos pela folha do nó 19 em plantas de kiwi, que apresentava 49% de
expansão da área foliar total, sugerindo que nesta fase de seu desenvolvimento esta folha não
estaria atuando como uma folha fonte, e sim ainda como um possível dreno. Folhas com 64%
de expansão total de sua área foliar já atuavam como exportadoras de fotoassimilados, ainda o
mesmo autor citando Watson e Bowers (1965), reportam que folhas da maioria das plantas
tornam-se exportadoras de fotoassimilados a partir de 1/3 a 50% de seu pleno
desenvolvimento final (tamanho). Contudo, em cerejeira (Prunus cerasus L.). Flore e Layne
(1999) relataram que as folhas das brotações terminais só começaram a exportar
fotoassimilados 17 dias após sua emergência, quando apresentavam 27% do total de peso de
matéria seca da folha.
Matsuura et al. (2001), trataram folhas de figueira com 13C em
diferentes alturas do ramo e fases de desenvolvimento dos frutos. A maior parte dos
fotoassimilados produzidos pelas folhas da base dos ramos foi armazenada nos frutos da axila
dessas folhas e na base dos ramos, enquanto os produzidos pelas folhas da parte mais apical do
ramo foram armazenados nas folhas e grande parte direcionados para os ápices dos brotos,
indicando forte concorrência entre o crescimento vegetativo e a formação de frutos.
Nos tecidos fotossinteticamente ativos, como folhas maduras, a
produção de carboidrato é maior do que a sua necessidade para manutenção do metabolismo e
crescimento, desta forma, exportam excedentes, na forma de sacarose, para tecidos que são
fotossinteticamente menos ativos ou inativos, como folhas jovens , raízes cachos ou ramos
(DANTAS et al., 2007).
Pigé et al. (2001), avaliaram os parâmetros de troca de carbono em
folha e frutos de ramos de figueira com 1 ano de idade durante a estação primavera-outono,

19
observando que a taxa de fotossíntese líquida variou de 15 a 20 mmol CO2 d-1 AGM-1 sendo
que também os frutos em sua fase inicial de desenvolvimento assimilaram uma pequena taxa
de CO2, evidenciando com isso sua contribuição na relação fonte-dreno, os mesmos autores
apontam valores de radiação fotossinteticamente ativa entre 750 e 800 µmol photons. m-2 s-1,
faixa na qual ocorreu a máxima assimilação líquida de CO2.
Em trabalho realizado por Silva, et al. (2008a) valores das trocas
gasosas e assimilação líquida de CO2 em Botucatu/SP, no mês de março de 2007, foram de
12 a 16 µmol m-2 s-1, corroborando com os dados apresentados por Pigé et al. (2001), sendo a
figueira uma planta temperada adaptou-se muito bem ao clima da região não apresentando
diferenças significativas de fotossíntese. A variação sazonal da taxa de fotossíntese e da
condutância dos estômatos em espécies arbóreas, como a figueira nas regiões tropicais, está
relacionada com as condições de déficit de pressão de vapor, temperatura do ar e
principalmente, umidade do solo, bem como a característica de cada estação do ano.
2.7. Translocação de fotoassimilados
As interações fisiológicas existentes entre os órgãos vegetais capazes
de exportar carboidratos (fontes) e os órgãos que demandam estes compostos (drenos) são
conhecidas como relações fonte-dreno. Tais relações são importantes no desenvolvimento das
plantas, pois influenciam na sua produção e no tamanho dos frutos (MINCHIN et al., 1997).
Os principais carboidratos acumulados são amido e açúcares solúveis redutores e não
redutores, sendo a sacarose o principal açúcar não redutor, mobilizado nos processos de
transporte na direção fonte-dreno, também observado por Zhang et al. (2005b), onde o
crescimento do fruto de pêra Japonesa foi dependente do fluxo de fotoassimilados produzidos
pelas fontes (folhas), bem como da área foliar e habilidade fotossintética das mesmas,
controlando ainda o número, tamanho, atividade e duração do crescimento do dreno.
A maioria dos carboidratos acumulados nos sítios de estocagem é
translocada durante a estação de crescimento e este fenômeno está associado à presença de

20
drenos fortes. A sacarose é o carboidrato mais comumente translocado (pelo floema) entre as
fontes, órgãos exportadores de C, e os drenos, órgãos consumidores de C (JACKSON, 2003).
Em outras espécies frutíferas como as Rosáceas, Buckhout e Tubbe
(1996), relatam que os carboidratos solúveis (frutose, glicose, sacarose e sorbitol) têm
importância na regulação osmótica. A dinâmica da água nos tecidos está muito relacionada
com a dinâmica dos carboidratos solúveis, embora a movimentação passiva destes também
ocorra a pequenas distâncias por meio da difusão simples ou difusão facilitada por proteínas
transportadoras da membrana. Eles podem ser estocados como a sacarose em quantidades
importantes dentro dos vacúolos (YAMAKI, 1982).
Nas frutíferas temperadas como a pêra, o sorbitol é o açúcar de
translocação mais importante (OLIVEIRA e PRIESTLEY, 1988). Este exerce o mesmo papel
que a sacarose em outras espécies, como forma de transporte dos produtos da fotossíntese
desde as folhas até outros órgãos através da seiva do floema (MOING et al., 1992) onde
podem estar em quantidades equivalentes ou superiores a sacarose (BIELESKI, 1982). A
temperatura exerce influência sobre o transporte no floema. A primeira resposta da planta em
relação à baixa temperatura é a inibição do transporte, porém, a retomada do fluxo pode
acontecer completamente mesmo sob a continuidade das baixas temperaturas. A ação da baixa
temperatura no transporte, possivelmente, está relacionada com a alteração da viscosidade da
solução de carboidratos. Da mesma forma, não somente a baixa temperatura atingida pode
alterar o fluxo, mas também a magnitude da redução ou mesmo a taxa de resfriamento. Após
vários tratamentos com baixas temperaturas, algumas plantas podem se tornar insensíveis às
variações térmicas impostas, tornando-se adaptadas às novas condições de crescimento. O
resfriamento lento parece induzir mudanças na natureza do fluxo, causando re-direcionamento
para vários drenos na planta (THORPE e MINCHIN, 1996).
De acordo com Rakngan (1995), as gemas de pereiras do cultivar
Nijisseiki, aumentam a concentração de açúcares quando as plantas entram em repouso e
diminuem antes ou durante a brotação. Em experimento realizado por Herter et al,. (2002),
observou-se que os níveis de carboidratos em gemas floríferas de pereira japonesa submetidas
à flutuação térmica durante o período de dormência não apresentaram diferenças, exceto
quanto ao sorbitol, que foi considerado o principal açúcar de translocação em pereiras cv.
Nijisseiki.

21
Assim como a figueira, o caquizeiro é uma espécie lenhosa que para
Mowat e George (1994), apresenta alternância entre períodos de crescimento e dormência, de
acordo com a sazonalidade climática nas diferentes regiões onde é cultivado. Na estação de
crescimento, ocorre o alongamento das brotações e a expansão foliar, cujo desenvolvimento se
completa pouco antes do florescimento. Durante o período de atividade fotossintética, o
eventual excedente em compostos fotoassimilados produzidos pela planta fica também
imobilizado na forma de carboidratos insolúveis em órgãos aéreos e subterrâneos da planta,
sendo, então, mobilizados gradativamente em carboidratos solúveis durante o período de
dormência (MOWAT e GEORGE, 1994).
Com o fim da dormência, essa mobilização é acelerada, sendo os
carboidratos solúveis conduzidos para as gemas em brotação que, por sua vez, formarão novos
ramos e folhas; posteriormente, as flores e os frutos são supridos, seguidos pelo câmbio, por
novas gemas em formação e, finalmente, pelos tecidos que servem como depósito de
carboidratos em órgãos subterrâneos e aéreos da planta (WARDLAW, 1990; LARCHER,
2000). O amido é o principal carboidrato de reserva do caquizeiro, sendo facilmente
mobilizado para formas solúveis durante o seu desenvolvimento (MOWAT e GEORGE,
1994). A época da mobilização dos carboidratos presentes nos órgão lenhosos da planta está
diretamente ligada aos eventos climáticos, sobretudo à temperatura, tendo grande importância
nos estudos de adaptação de frutíferas de clima temperado (HERTER et al., 2001). A
intensidade dessa mobilização influência, por sua vez, no desenvolvimento fenológico da
planta, como no crescimento de ramos, no florescimento e na produção de frutos (LIU et al.,
1999; LARCHER, 2000).
Em trabalho realizado por Matsuura et al. (2001), folhas de figueiras
(Ficus carica L.) 'Masui Daufine' com dois anos de idade foram expostas a 13CO2 no mês de
outubro, e as analises mostraram que 13C-fotossintatos foram armazenadas durante o período
de dormência e remobilizado na primavera seguinte. O excesso de % átomos 13C durante o
período de dormência foi elevado nas raízes que apresentavam o menor calibre decrescendo
em seguida, para raízes finas, raízes de tamanho intermediário (médio), raízes com maior
diâmetro, tronco, ramos de dois anos de idade e ramos de um ano de idade.
Embora, 25 dias depois do surgimento da brotação, novas brotações e
raízes exibissem relativamente nos órgãos velhos (maduros) um elevado excesso de % átomos

22
13C, as novas raízes retiveram altos níveis de 13C, após 45 dias da pausa da brotação, embora o
excesso de átomos (%) 13C, em ramos de 1 e 2 anos de idade e raízes de diâmetros maiores,
drasticamente decresceram durante os primeiros 25 dias depois do rompimento da brotação.
Além disso, o decréscimo significativo nos níveis de átomos 13C no
tronco e nas raízes de tamanho intermediário (médias) ocorreu entre 25º e 45º dia depois do
surgimento das brotações. O excesso de átomos (%) 13C em novas brotações foi menor nas
partes mais altas da planta do que em partes baixas, 45 dias depois da brotação. Neste caso,
observou-se que o crescimento de novas brotações e raízes na primavera, dependeu
principalmente das reservas de carboidratos dos ramos mais velhos e próximos a brotação e
raízes de tamanhos maiores no mínimo 25 dias depois do rompimento da brotação.
Subseqüentemente, para os próximos 20 dias, novos crescimentos contam com as reservas
armazenadas no tronco e nas raízes de médio tamanho (MATSUURA, et al., 2001).
Nota-se ainda que nos dados de Matsuura et al. (2001), o sistema
radicular é um dos principais órgãos de reserva da planta, sendo o mesmo quem fornece o
aporte de reservas necessárias para a brotação dos novos ramos e recuperação de todo o
dossel do próximo ciclo, pois no sistema de produção brasileiro continuamente a planta se
encontra em crescimento devido as podas anuais. Portanto, faz-se necessário dar condições de
aeração adequada de solo, adubação equilibrada e irrigação para que as reservas acumuladas
de sacarose, amido e outros garantam o surgimento de novos ramos, folhas e frutos.
Silva et al. (2008b), estudando a variação natural de carbono 13
constataram que as partes novas dos ramos do cultivar Roxo de Valinhos apresentaram valores
isotópicos médios de -28,41‰; -28,43‰ e -28,51‰, que indicaram maiores valores de δ 13C
nas folhas recém abertas, gema apical e frutos. O ramo 02 apresentou órgãos vegetais com
valores isotópicos maiores quando comparados ao ramo 01, mostrando que este ramo possui
uma idade fenológica maior entre a poda e época de coleta. Verificou-se que os frutos do ramo
2 apresentaram maiores valores de δ 13C que as brotações, evidenciando tendências da relação
fonte-dreno da planta. Considerando que a planta estudada possuía 07 anos de idade, ou seja,
uma planta ainda jovem visto a idade que as figueiras podem alcançar, foi observado um
pequeno gradiente da distribuição natural do valor δ 13C com pequeno aumento nos tecidos
novos (ramo – parte apical e radicelas), entretanto, nos tecidos mais velhos da planta
verificou-se predominância do 12C. Contudo, o valor isotópico médio -28,94 ± 0,361‰

23
observado nos resíduos de poda, diferiu destes valores, pelo fato do agrupamento equivaler a
resíduos de tecidos vegetais de podas sucessivas dos últimos 5 anos.
2.8. Isótopos e sua Utilização
Os isótopos são elementos naturais que apresentam em sua
constituição o mesmo número atômico, porém com números de massa diferentes, ou sejam,
apresentam o mesmo número de prótons e elétrons e diferentes números de nêutrons,
ocupando o mesmo lugar na tabela periódica. Dessa forma, os isótopos apresentam as mesmas
propriedades químicas, sendo classificados em radioativos ou estáveis.
Os elementos naturais que constituem os isótopos estáveis não
possuem as propriedades de emissão de radiações e são caracterizados pela sua abundancia
natural, expressa na unidade átomos %. Exemplos: 12C (98,89%) e 13C (1,11%); 14N (99,62) e 15N (0,38%); e 16O (99,796%) e 18O (0,204%) e o 1H (99,8844%) e 2H (0,1156%). A
terminologia comumente empregada na detecção das concentrações e naturais expressa-se pela
linguagem delta per mil e por átomos por cento para compostos enriquecidos (BARRIER e
PROSSER, 1996), citados por Vasconcellos (2001). De acordo com Schimel (1993) os
isótopos estáveis são usados para seguir movimentos e transformações químicas em sistemas
biológicos e ambientais, podendo ser introduzidos na planta, solo ou sistemas aquáticos e
monitorados com grande sensibilidade e precisão por espectrômetros de massa.
Segundo Xavier et al. (2007), os isótopos radioativos 14C, foram
descobertos em 1940, sendo inicialmente utilizados em estudos sobre datação em arqueologia.
A aplicação do 14C no estudo sobre a fisiologia de plantas ganhou impulso a partir das décadas
de 50 e 60, permitindo o conhecimento mais aprofundado sobre a fotossíntese, translocação e
alocação dos fotoassimilados e das relações fonte-dreno, em diversas espécies de plantas.
Esses isótopos radioativos ou radioisótopos emitem partículas e/ou radiações (α, β, γ e raios
x), os quais se desintegram, transformando-os em átomos de outro elemento ou do mesmo
elemento. Uma importante unidade dos elementos radioativos refere-se à meia-vida do átomo,
a qual se define como o tempo requerido para que a metade da população de um átomo se
desintegre.

24
A determinação dos valores isotópicos, ou sinal isotópico nas
diferentes espécies de plantas é conseguido a partir de amostras do material orgânico da
planta, que são analisadas pelo equipamento conhecido como espectrômetro de massas de
razão isotópica (IRMS). Neste sistema, de acordo com Ducatti (2007), a amostra e o padrão
são admitidos na forma de dióxido de carbono e após a passagem por uma fonte de ionização,
os feixes dos íons gerados são separados por um campo magnético de acordo com as suas
relações massa/carga. Basicamente, compara-se a razão do 13CO2 (massa 45)/ 12CO2 (massa
44) com uma amostra padrão. O resultado em termo de enriquecimento relativo (δ) da amostra
em relação ao padrão é expresso em partes por mil (‰), conforme equação 1.
3
PADRÃO
PADRÃOAMOSTRA13 10R
RRC ×
−=δ (eq. 01)
em que: R é a razão isotópica obtida entre o isótopo pesado sobre o isótopo leve (13C/12C) da
amostra e do padrão, respectivamente. Como os valores numéricos das diferenças são
pequenas, costuma-se multiplicar a expressão por 1000, obtendo-se a terminologia em delta
per mil [δ‰(amostra, padrão)].
Os isótopos estáveis do carbono (12C e 13C) tornaram-se uma
ferramenta muito útil na pesquisa sobre aspectos relacionados à fisiologia de plantas, uma vez
que as razões entre estes dois isótopos podem auxiliar diretamente no estudo da fotossíntese,
determinação dos ciclos fotossintéticos, translocação e alocação de carbono e estresse hídrico,
alem de indiretamente servirem de base no estudo sobre o melhoramento de plantas tolerantes
ao estresse hídrico e mesmo para trabalhos relacionados a desbaste ou poda de plantas,
notadamente de fruteiras (EHLERINGER et al., 1993).
De acordo com Ludlow et al (1976), aproximadamente 99% de todo
carbono na natureza está na forma do isótopo 12C e apenas 1% estaria na forma do isótopo 13C.
Estes dois isótopos estáveis do carbono se comportam de forma diferente nas reações físicas e
químicas, resultando em proporções variáveis destes isótopos nos diferentes materiais. Para
Schimel (1995), os isótopos estáveis são usados para seguir movimentos e transformações
químicas em sistemas biológicos e ambientais, podendo ser introduzidos na planta, solo ou

25
sistemas aquáticos e monitorados com grande sensibilidade e precisão por espectrômetros de
massa. Tornaram-se uma ferramenta muito útil nas pesquisas sobre aspectos relacionados à
fisiologia de plantas, uma vez que as razões entre estes dois isótopos podem auxiliar
diretamente no estudo da fotossíntese, determinação dos ciclos fotossintéticos, translocação e
alocação de carbono e estresse hídrico, além de indiretamente servir de base no estudo sobre o
melhoramento de plantas tolerantes ao estresse hídrico e mesmo para trabalhos relacionados a
desbaste ou poda de plantas, notadamente de fruteiras (EHLERINGER et al., 1993).
As plantas em sua maioria podem ser classificadas quanto ao ciclo
fotossintético em dois grupos principais: plantas do ciclo C3 e C4, existindo também espécies
chamadas de CAM, que não podem ser classificadas pelos critérios padrões nem como C3
nem como C4, por terem características de ambos os grupos. Ambos os grupos apresentam
diferenças na razão entre o 12C e 13C presentes em suas folhas, sendo estas diferenças reflexos
dos processos de fracionamento isotópico, que determinam uma discriminação, contra ou a
favor do 13C, ocorrido durante a fotossíntese. Basicamente, nas plantas C3 estas discriminações
ocorrem na difusão do CO2 pelos estômatos até os cloroplastos, pela ação da enzima de
carboxilação (rubisco) e pela diferença nas concentrações externas e internas de CO2. Nestas, a
discriminação isotópica ocorre em maior valor pela ação da enzima rubisco, sendo que nesta
fase a discriminação é contra o 13C.
Como resultado, espécies de plantas do ciclo fotossintético C3
apresentam valores na razão isotópica que variam de -22 a – 34 ‰, tendo como media, valores
na ordem de -27‰. As plantas do ciclo C4 apresentam além das etapas citadas para as plantas
C3, discriminação isotópica na formação do HCO-3 e sua incorporação pela PEP-carboxilase,
alem do fator φ que representa a taxa de CO2 que escapa das células da bainha e podem ser ou
não ser reincorporadas, ocorrendo nestas fases, discriminações a favor do 13C.
Dessa forma, o resultado do enriquecimento isotópico relativo (δ 13C)
nas espécies do ciclo C4 variam de –9 a –16 ‰, com valores médios de -14‰ e as plantas
CAM utilizam a via C4, porém de uma forma distinta das plantas C4. As plantas C4 fazem
uma separação espacial dos eventos, enquanto as plantas CAM fazem uma separação
temporal, dependendo da condição ambiental em que se encontram assumem comportamento
de plantas com ciclo CAM obrigatório que apresentam valores isotópicos comparáveis ao das
plantas do ciclo C4 (–9 a –16 ‰) ou facultativas que apresentam valores de -20 ‰ a -15‰.

26
(FARQUHAR et al., 1989; O’LEARY, 1993). Diz-se, portanto que plantas do ciclo
fotossintético C3 são mais pobres em 13C em relação às plantas do ciclo fotossintético C4. E
essa variação na quantidade de 13C nas diferentes espécies (C3 e C4), faz com as mesmas
possuam um valor isotópico próprio.
Segundo Vasconcellos (2001), assim como no uso de isótopos
radioativos, pode ser realizado o enriquecimento da fonte em termos de seus isótopos estáveis.
Por exemplo, plantas colocadas em ambiente controlado, enriquecido em 13CO2, produzirão
fotoassimilados enriquecidos em 13C, conseqüentemente alterando seu sinal isotópico e
possibilitando sua determinação (direção de translocação) onde quer que estejam alocados.
Dessa forma, podem-se usar os isótopos estáveis como “traçadores” ou “marcadores” à
semelhança dos isótopos radioativos.
Para uma determinação quantitativa da translocação e alocação dos
assimilados nas diferentes partes vegetais, utilizando isótopos radioativos ou estáveis, o
equilíbrio entre os assimilados marcados e os não marcados deve ser obtido, de forma que o
tempo necessário para que ocorra este equilíbrio varie conforme a metodologia utilizada
(DELÉENS et al., 1994).
O equilíbrio entre compostos enriquecidos naturais segue o fenômeno
de troca, que está baseado na aplicação da lei de diluição isotópica, utilizados nos estudos das
reações de troca isotópica. Uma reação de troca isotópica é uma reação segundo a qual dois
átomos com o mesmo número atômico, mas com número de massa diferente substituem-se
mutuamente. Deste modo, a quantidade de um elemento marcado fixado em um órgão ou
tecido, em um dado período de tempo, fornece uma estimativa de troca, ou do modelo de
partição do referido elemento. Porém, a medida desta estimativa requer condições específicas
para uma determinação precisa. Para tanto, uma analise das medidas do isótopo marcado nos
diferentes órgãos, durante o período de enriquecimento, acompanhado de uma analise
compartimental, permite a elaboração de um modelo a partir do qual o tamanho dos
compartimentos e os parâmetros de transferência possam ser avaliados. O tempo da diluição
isotópica também é importante, pois enriquecimento por longo tempo permite obter um
equilíbrio de troca entre a fonte enriquecida e os vários órgãos ou tecidos, de forma que o
fluxo do elemento marcado será proporcional ao do elemento não marcado (DELÉENS et al.,
1994).

27
3. MATERIAL E MÉTODOS
3.1. Localização e descrição da área experimental
Os experimentos foram conduzidos em condições de campo na área
experimental do pomar do Departamento de Produção Vegetal Horticultura da Faculdade de
Ciências Agronômicas, FCA/UNESP, câmpus de Botucatu-SP. As coordenadas geográficas
locais, de acordo com Tubelis e Salibe (1989) são: 22º 52’ 47” latitude S, 48º 25’ 12”
longitude W e altitude de 810 m.
O tipo climático predominante no local, segundo Tubelis et al. (1972)
e Curi (1972), baseados no sistema Internacional de Köppen, está incluído no tipo Cwa,
caracterizado como temperado quente (mesotérmico) com chuvas no verão e seca no inverno,
com precipitação e temperatura média anual de 1530 mm e 21°C, respectivamente (CUNHA
et al., 1999). Os dados de temperatura média, máxima e mínima mensais, precipitação
pluviométrica e radiação solar global mensal foram fornecidos pelo Posto Meteorológico do
Departamento de Recursos Naturais da Faculdade de Ciências Agronômicas – UNESP,
câmpus de Botucatu-SP e encontram-se na Tabela 01.

28
Tabela 1. Médias mensais das temperaturas mínima, máxima e média (ºC), precipitação
pluviométrica (mm) e radiação solar global (W m-2) medidas no período de janeiro de 2007 a
agosto de 2008. Fonte: Departamento de Recursos Naturais – FCA/ UNESP, Botucatu/SP.
Temperatura (º C) Mês e ano
Mínima Máxima Média
Precipitação pluviométrica
(mm)
Radiação solar global
(W m-2)
Janeiro / 2007 19,2 26,9 23,0 410,8 310,7 Fevereiro / 2007 19,0 29,3 24,1 108,9 416,4 Março / 2007 19,4 30,2 24,8 49,0 400,0 Abril / 2007 18,0 27,6 22,8 40,3 341,7 Maio / 2007 13,6 23,3 18,5 42,1 302,4 Junho / 2007 14,5 25,2 19,8 23,4 311,9 Julho / 2007 13,0 22,7 17,9 172,6 260,4 Agosto / 2007 15,1 25,6 20,4 0,0 360,6 Setembro / 2007 17,3 28,9 23,1 10,0 392,1 Outubro / 2007 17,5 30,3 23,9 77,7 380,1 Novembro / 2007 17,5 26,5 22,0 177,0 358,0 Dezembro / 2007 18,4 28,1 22,8 180,6 401,2 Janeiro / 2008 18,6 26,4 22,5 279,3 293,4 Fevereiro / 2008 19,0 28,0 23,5 94,6 356,1 Março / 2008 18,6 27,5 23,0 60,9 383,7 Abril / 2008 17,4 25,8 21,6 102,8 293,3 Maio / 2008 13,8 22,8 18,3 115,7 272,3 Junho / 2008 13,5 24,5 17,6 30,8 276,9 Julho / 2008 13,9 24,6 19,1 0,0 346,7 Agosto / 2008 15,1 25,5 20,3 104,1 302,2
O solo da área onde as plantas estavam instaladas foi classificado
como Nitossolo Vermelho, segundo os critérios da EMBRAPA (1999).
A área total do experimento continha 190 plantas de figueira do
cultivar Roxo de Valinhos. O pomar foi implantado em setembro de 2001, num espaçamento
de 3 m entre linhas e 2 m entre plantas, com uma área de 6 m²/planta. As plantas receberam
tratos culturais como poda de formação, frutificação e desbrotas com a finalidade de
adquirirem a conformação de 6 a 9 ramos produtivos (PENTEADO e FRANCO, 1997), que é
considerada a mais recomendada para a cultura, bem como adubações e correções de pH, com
base na analise do solo seguindo as recomendações técnicas de Campo Dall’Orto et al. (1996).
O controle fitossanitário seguiu a recomendação de Penteado e Franco (1997).

29
O manejo de plantas invasoras foi realizado através de roçada nas
linhas e entrelinhas de plantio. Ao redor da coroa das plantas foram realizadas capinas.
3.2. Caracterização das Trocas Gasosas
As determinações de trocas gasosas foram realizadas com um medidor:
sistema aberto portátil de fotossíntese, IRGA, modelo LI-6400 (LI-COR), sempre na região
mediana das folhas (folíolo central de folhas novas completamente desenvolvidas),
completamente expandidas, totalmente expostas à radiação solar, no período das 09h00min às
10h30min da manhã, com o intuito de determinar a folha fotossinteticamente ativa. Sendo
medidos os seguintes parâmetros: Assimilação líquida de CO2 (A) em µmol m-2 s-1,
(quantidade líquida de CO2 assimilado por unidade de área foliar por unidade de tempo); taxa
de transpiração (E) nas folhas mmol m-2 s-1 (quantidade de vapor d’água transpirado por
unidade de área foliar por unidade de tempo (µmol de H2O por segundo por metro quadrado));
condutância estomática nas folhas (gs) em mmol m-2 s-1 (fluxo de entrada e saída de CO2 e
vapores de H2O através dos estômatos); concentração de CO2 nos espaços intercelulares (Ci)
em mmol m-2 s-1; (quantidade de CO2 que fica armazenada na câmara subestomática);
eficiência do uso da água (EUA) (foi obtida através da relação entre a taxa de assimilação
líquida de CO2 e a taxa de transpiração das folhas (A/E)).
Em cinco plantas da espécie em estudo, com 4 ramos cada uma, foi
determinada a área foliar (cm2) individual das folhas, de acordo com a localização da folha no
ramo. A avaliação foi realizada através do aparelho de medição de área foliar, integrador
fotoelétrico modelo (LI-3000, LICOR).
O experimento foi instalado em delineamento estatístico inteiramente
casualizado, constando de cinco plantas com quatro ramos, sendo cada folha da planta
considerada um tratamento. Os dados foram submetidos à análise de variância, pelo teste F e
as diferenças entre médias, comparadas pelo teste de Tukey, a 5 % de significância
(PIMENTEL GOMES, 1990).

30
Dentre as funções analisadas na análise de regressão, o modelo
significativo foi conseguido com a função exponencial polinomial do 2º grau, tanto para a
posição da folha no ramo, quanto para a área foliar.
3.3. Metodologia de enriquecimento em 13C
Para a realização desta prática, foi construída uma câmara em material
acrílico e adquiridos equipamentos como seringa e mangueiras de látex que possibilitaram a
aplicação do 13CO2.
A câmara foi baseada no modelo descrito por Vasconcellos (2001), de
forma a possibilitar a retirada do ar ambiente e a injeção do gás enriquecido, imediatamente
após a inserção da folha no seu interior e seu completo fechamento, havendo para isso uma
saída lateral que foi fechada por um pedaço de mangueira (látex), para evitar que houvesse a
difusão do ar enriquecido da câmara para o ambiente. Contudo neste experimento não foi
retirado o ar ambiente da câmara, somente injetado o gás enriquecido, tendo a câmara, as
seguintes dimensões: 28,0 cm de comprimento x 35 cm de largura x 4,0 cm de altura x 0,5 cm
de espessura, perfazendo um volume de 4000 cm3.
Nos ramos das plantas adultas em estádio reprodutivo, previamente
foram selecionadas folhas adultas pela caracterização fotossintética sendo ela a quarta folha do
ramo a contar do ápice. Esta folha foi cuidadosamente colocada na câmara de acrílico, sendo
esta fechada e vedada com vaselina, assim como, a região do pecíolo que também foi envolta
em pomada vaselina, de forma a ocupar todo o espaço entre o pecíolo da folha e a câmara.
A injeção do gás na câmara foi feita por um dispositivo que regula o
direcionamento do fluxo do cilindro de 13CO2 para a seringa e posteriormente, da seringa para
a câmara, introduzindo-se 8,0 ml de 13CO2, a 99 átomos %, ou seja, de cada 100 átomos de
CO2 deste gás, 99 % dos átomos são formados por 13C e 1% de 12C, que propiciaram um δ13C
de 35.693,27 % de 13C (29,19 átomos % de 13C em excesso). O tempo de enriquecimento foi
de 30 minutos, e após este período foi retirada a câmara.
No campo, as folhas foram colocadas na câmara entre os horários de
09h 00min e 11h 00min, para uma maior eficiência fotossintética, de forma que a câmara não

31
recebesse as maiores taxas instantânea de radiação solar global diária, que provocaria a
inibição da fotossíntese com a elevação da temperatura. A temperatura interna foi medida com
a introdução de um termômetro dentro da câmara. O período de condução do experimento foi
de setembro de 2007 a março de 2008.
Figura 1. Câmara de enriquecimento de 13CO2. FCA/UNESP, Botucatu-SP.

32
3.4. Metodologia de coleta do material enriquecido
No primeiro experimento de alocação dos fotoassimilados, as amostras
foram coletadas 24 horas após o período de enriquecimento, por volta das 10h00min do dia
seguinte, em que foi realizado o enriquecimento. A experimentação constou na escolha de 04
plantas amostradas coletadas ao acaso, na área experimental para evidenciar os tratamentos:
T1- planta com ausência de drenos (frutos e brotação);
T2-planta com presença do dreno fruto;
T3- planta com presença do dreno broto;
T4- planta com a presença dos dois drenos.
No momento da coleta dos diferentes órgãos da planta trabalhada
(folha, ramo, tronco e raízes), estes foram devidamente, identificados quanto a sua localização
na planta, e imediatamente imersos em nitrogênio liquido (-196º C), para provocar a morte dos
tecidos, evitando as alterações do enriquecimento.
No experimento de determinação do tempo de alocação dos
fotoassimilados, os tratamentos foram os tempos de arranquio das plantas no momento da
coleta dos diferentes órgãos amostrados (folhas, ramos, tronco e raízes):
T1-6 horas após o enriquecimento com 13C;
T2-24 horas após o enriquecimento com 13C;
T3-48 horas após o enriquecimento com 13C;
T4-72 horas após o enriquecimento com 13C;
T5-120 horas após o enriquecimento com 13C;
T6-168 horas após o enriquecimento com 13C;
T7-360 horas após o enriquecimento com 13C.

33
3.5. Secagem e moagem das amostras
No laboratório do Centro de Isótopos Estáveis Ambientais, do Instituto
de Biociências da Unesp, Campus de Botucatu-SP, as amostras previamente identificadas
foram colocadas em estufas com circulação de ar forçada à 65º C por 72 horas, para secagem e
moagem. Para isso foi utilizado o moinho criogênico à base de nitrogênio líquido (Spex –
Modelo 6700). No moinho as amostras foram colocadas em tubos individualizados, imersas
em nitrogênio líquido. O uso deste equipamento permitiu a obtenção de um material com
finíssima granulometria (com aspecto de talco), o qual permitiu uma perfeita homogeneização
da amostra. Devido à grande sensibilidade e precisão do espectrômetro de massas (±0,2‰),
isso para minimizar as diferenças entre repetições (em número de três), de cada amostra, nas
leituras dos resultados, devido ao seu tamanho e dureza, foram primeiramente moídas, no
moinho martelo (Culatti TYP MIC CZ13).
Após a moagem, foram retiradas, de cada amostra, uma alíquota que
variou entre 50 e 70 µg (tendo o devido valor de peso anotado) e acondicionadas em cápsulas
de estanho, com 6 mm de altura e 4 mm de diâmetro (Modelo D1106-Elemental Microanalysis
Limetd).
3.6. Analise do enriquecimento relativo
Na etapa seguinte, as amostras foram colocadas no analisador
elemental (Carlo Erba EA 1108 – Fisons, Milão – Itália), o qual está acoplado ao
espectrômetro de massas. Neste aparelho, através da combustão das amostras, são liberados o
CO2 e o N2, separados na coluna de cromatografia, não sendo realizada a analise do N2, o
produto de interesse CO2 em seguida é transferido para o ConFlo (Finnigan Mat – Alemanha),
onde ocorre o ajuste entre altura, em volts da amostra e do padrão. Após este ajuste a amostra
seguiu para o espectrômetro de massas para razões isotópicas (IRMS) (Modelo Delta S -
Finnigan Mat – Alemanha), onde a razão 13C/ 12C foi determinada e expressa em δ‰, notação
relativa ao padrão PDB, equação 1.

34
Das plantas em estádio reprodutivo, uma foi escolhida para a avaliação
do enriquecimento natural de 13C e 12C, em seus diversos órgãos e posição na planta visando à
determinação do valor limite do enriquecimento relativo natural (δ 13C), valor esse a ser usado
como comparação com os resultados obtidos nas amostras coletadas nos ramos, nos quais as
folhas foram submetidas ao ambiente de enriquecimento de 13CO2. Ao final seria possível a
caracterização e mapeamento do enriquecimento relativo natural de 13C de toda a planta. Em
seguida foram seccionadas as seguintes partes: gema apical, folha recém-aberta, folha
fotossinteticamente ativa de cada nó em todo o ramo, brotações laterais, frutos, sendo os
ramos divididos em três partes (apical, mediana e basal) objetivando a igualdade no número de
nós entre as partes. Os dois ramos partiram do mesmo resíduo de poda, que foi seccionado em
função do número de podas realizadas. Seqüencialmente foram seccionados: caule, epiderme
do caule, raiz primária (diâmetro maior que 40 mm até o caule), raízes secundárias (diâmetro
entre 40 e 15 mm) e terciárias (diâmetro compreendido entre 15 e 2 mm), sendo estas últimas
subdivididas em grossa, média e fina, e as radicelas (diâmetro menores que 2mm).
Agrupando todos os constituintes de cada parte da planta, foram
calculadas as médias do valor isotópico natural e o desvio padrão destas.

35
4. RESULTADOS E DISCUSSÃO
4.1. Caracterização fotossintética de figueiras ‘Roxo de Valinhos’
O metabolismo do carbono das plantas está vinculado à atmosfera
através das trocas gasosas. Durante o período em que foram realizadas as avaliações das trocas
gasosas, a temperatura do ar variou de 22,46 a 31,92 ºC, a umidade relativa do ar de 64,3 a
77,2% e a radiação fotossinteticamente ativa de 119 a 1.868 µmol fótons m-2 s-1. Em geral, os
valores da temperatura da folha estiveram 3,7% (1,1 ºC) acima da temperatura do ar para todos
os dias de avaliação, corroborando com Nogueira et al. (1999) que encontraram diferenças de
1,0 a 2,5 ºC em folhas de mangabeira expostas ao sol. Para Machado et al. (2002), a variação
sazonal das condições do ambiente ao longo do ano podem causar variações significativas nas
trocas gasosas em espécies arbóreas. Assim, para estudos com esse enfoque, as folhas
necessitam estar com as mesmas idades, ou então, procurar avaliar folhas em diferentes
estádios dentro das mesmas condições climáticas.
Os parâmetros avaliados nas trocas gasosas (taxa de assimilação
líquida de CO2, condutância estomática, concentração interna de CO2, taxa de transpiração e

36
eficiência do uso da água) em folhas de figueira ‘Roxo de valinhos’ em diferentes posições no
ramo da planta diferiram estaticamente entre si (Tabela 02), todavia, foram semelhantes entre
os ramos de uma mesma planta.
Pimentel (1998) relata que as plantas C3 apresentam atividade
fotossintética variando entre 12 a 25 µmol m-2 s-1, as plantas C4 entre 25 a 40 µmol m-2 s-1 e as
plantas CAM entre 2,5 a 7,6 µmol m-2 s-1, dependendo das condições ambientais. Pelos
valores de taxa de assimilação líquida de CO2 fica evidenciado que a figueira ‘Roxo de
Valinhos’ é uma planta C3. Contudo, os valores médios aqui apurados podem ser considerados
baixos para a espécie Ficus carica L., pois, Pigé et al. (2001), avaliando os parâmetros de
troca de carbono em folha e frutos de ramos da figueira com 1 ano de idade durante a estação
primavera-outono, observaram que a taxa de fotossíntese líquida variou de 15 a 20 mmol CO2
d-1 AGM-1, o mesmo autor aponta valores de radiação fotossinteticamente ativa entre 750 e
800 µmol photons m-2 s-1 faixa na qual ocorreu a máxima assimilação líquida de CO2, valores
estes diferentes dos encontrados para a radiação fotossinteticamente ativa em Botucatu,
ocasionando a saturação por luminosidade.
Tabela 2. Dados de trocas gasosas de folhas de figueira ‘Roxo de Valinhos’ em Botucatu, SP.
FCA / UNESP, 2007.
Folha
Taxa de assimilação líquida de CO2 (µmol m-2 s-1)
Condutância estomática
(mmol m-2 s-1)
Concentração interna de CO2
(mmol m-2 s-1)
Taxa de transpiração
(mmol m-2 s-1)
Eficiência do uso da água
1 -0,370 c 0,159 b 391,58 a 2,195 b -0,202 b 2 8,839 b 0,316 ab 308,42 b 3,787 ab 2,867 a 3 11,104 ab 0,389 ab 301,17 b 3,857 ab 3,344 a 4 14,378 a 0,415 ab 291,92 b 4,226 ab 3,873 a 5 12,832 a 0,383 ab 295,58 b 4,134 ab 3,778 a 6 14,117 a 0,427 a 299,67 b 4,413 a 3,659 a 7 13,147 a 0,407 ab 306,83 b 4,521 a 3,291 a 8 13,839 a 0,416 ab 308,5 b 4,786 a 3,250 a
CV 55,01 % 25,77 % 16,39 % 20,47 % 48,63 % *Médias seguidas da mesma letra na coluna, não diferem pelo Teste de Tukey a 5% de probabilidade. *Médias de 05 plantas, com quatro ramos cada uma.
As folhas jovens apresentaram as menores taxas de assimilação líquida
CO2, transpiração e condutância estomática, refletindo diretamente na momentânea

37
ineficiência do uso da água por esta folha, pois apresentaram elevada concentração de CO2 na
câmara subestomática, dada pela expansão dos mecanismos de absorção da energia radiante e
a baixa conversão fotoquímica. Segundo Larcher (2000) a deficiência de clorofila pode ser
reconhecida pela coloração pálida e ocasiona uma considerável redução na intensidade
fotossintética, sendo que a deficiência de clorofila ocorre principalmente no começo do
desenvolvimento da folha.
A condutância estomática apresentou padrões similares ao da taxa de
fotossíntese, sugerindo que a queda da assimilação líquida de CO2 esteja relacionada com o
fechamento parcial dos estômatos. Campostrine e Yamanishi (2001) citam que a correlação
entre a assimilação de CO2 e a condutância estomática podem ser explicadas, como uma
função da alta concentração de CO2 no sítio de carboxilação quando ocorre o aumento da
condutância estomática, ou que a fixação de CO2 pode estar restringida pelo metabolismo de
carboidrato no citossol da célula e pelos processos supracelulares de transporte de
carboidratos, quando existe baixa atividade nas regiões dos drenos de fotoassimilados (BEL,
1992).
Efetivamente foi observado que folhas com área foliar acima de 60
cm2 começaram a assimilar CO2 e produzir fotoassimilados, pois começam a apresentar taxas
de assimilação de CO2 positivas com absorção dos fótons pelos cloroplastos emitidos pela
radiação fotossinteticamente ativa (Figura 2). Por conseguinte, folhas da figueira ‘Roxo de
Valinhos’ apresentaram máxima taxa de assimilação líquida de CO2, quando atingiram 172,42
cm2, valor este 4,65% superior à área foliar média da 4ª folha (164,39 cm2). Em média, nas
condições ambientais do mês março em Botucatu, as folhas alcançaram esse valor de área
foliar após 26 dias da sua abertura, corroborando com as ponderações de Pimentel (1998) que
plantas C3 têm sua maior produção de fotoassimilados após 21 dias da antese foliar.
A partir da 4ª folha foi verificado praticamente o mesmo
comportamento quanto às trocas gasosas, permitindo caracterizá-las como folhas adultas
(Figura 3). Ao longo do ramo vegetativo, foi observado que as maiores taxas de assimilação
líquida de CO2 e transpiração são encontradas nas folhas 6 e 7. Por apresentar menor taxa de
transpiração e uma atividade fotossintética semelhante as demais folhas adultas, além de
apresentar menor possibilidade da incidência do inoculo de ferrugens, foi escolhida a folha 4,
como aquela com potencial para enriquecimento com carbono marcado.
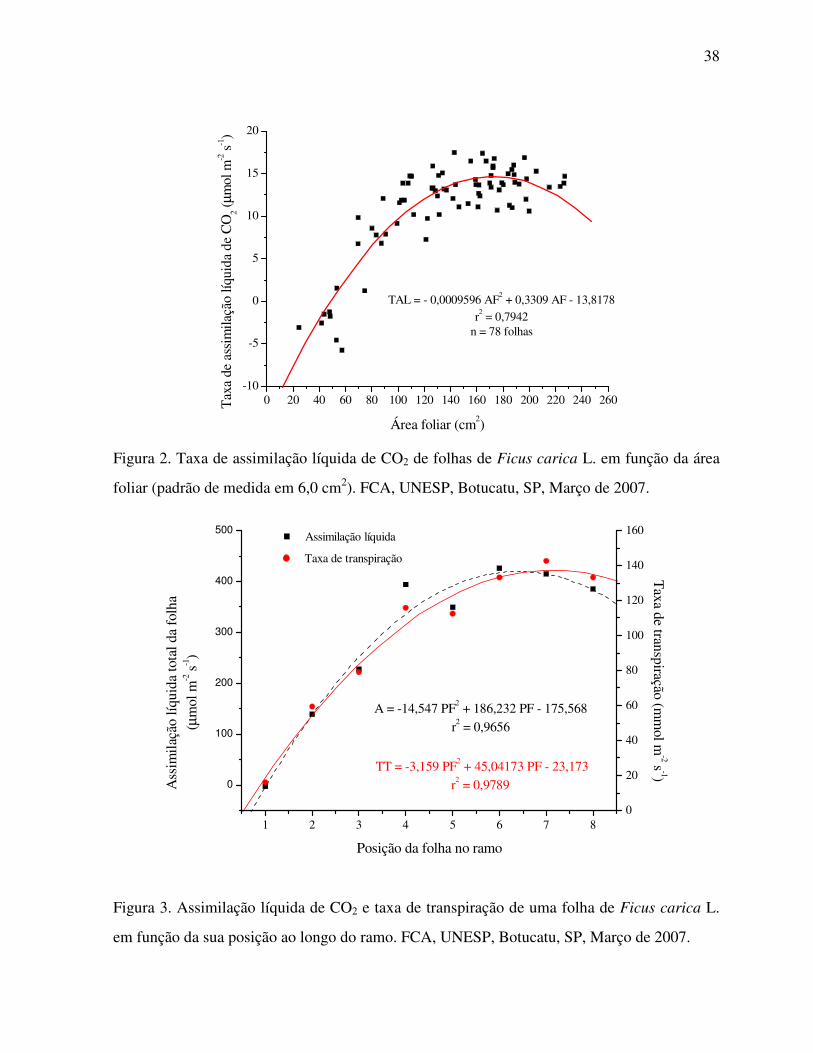
38
0 20 40 60 80 100 120 140 160 180 200 220 240 260-10
-5
0
5
10
15
20
TAL = - 0,0009596 AF2 + 0,3309 AF - 13,8178r2 = 0,7942
n = 78 folhas
Tax
a de
ass
imila
ção
líqui
da d
e C
O2 (
µm
ol m
-2 s
-1)
Área foliar (cm2)
Figura 2. Taxa de assimilação líquida de CO2 de folhas de Ficus carica L. em função da área
foliar (padrão de medida em 6,0 cm2). FCA, UNESP, Botucatu, SP, Março de 2007.
1 2 3 4 5 6 7 8
0
100
200
300
400
500
A = -14,547 PF2 + 186,232 PF - 175,568r2 = 0,9656
Assimilação líquida
Posição da folha no ramo
Ass
imil
ação
líqu
ida
tota
l da
folh
a (µ
mol
m-2 s
-1)
0
20
40
60
80
100
120
140
160
TT = -3,159 PF2 + 45,04173 PF - 23,173r2 = 0,9789
Taxa de transpiração
Taxa de transpiração (m
mol m
-2 s-1)
Figura 3. Assimilação líquida de CO2 e taxa de transpiração de uma folha de Ficus carica L.
em função da sua posição ao longo do ramo. FCA, UNESP, Botucatu, SP, Março de 2007.

39
4.2. Enriquecimento relativo natural da razão 13C/ 12C em figueiras
Um indicador do processo de assimilação de carbono como um todo é
a razão isotópica entre 13C e 12C na matéria seca das plantas, registrada em partes por mil
tendo por referência um padrão. Segundo Larcher (2000), atualmente o 13CO2 está sendo
encontrado em uma concentração de aproximadamente 1,1% em relação ao 12CO2 na
atmosfera. A ocorrência do 13C na massa vegetal é baixa, porque durante a carboxilação o 12C
é favorecido pelas enzimas. A Rubisco favorece mais fortemente o 12C, discriminando mais o 13C, quando comparada com a PEP-carboxilase (–28‰ e –9‰, respectivamente), comparando
enzimas de plantas C3 e C4.
Os ramos considerados neste estudo de variação natural foram
originados da mesma seção podada (com 5 resíduos de poda). O ramo 1 possuía 1,42 m de
comprimento e 35 nós, enquanto o ramo 2 tinha 1,52 m e 38 nós.
As partes novas dos ramos apresentaram valores isotópicos médios de
-28,18‰; -28,23‰ e -28,40‰, indicando valores superiores de δ13C nas folhas recém abertas,
gema apical e frutos (Tabela 3). As folhas dos nós em que haviam frutos formados
apresentaram menores valores isotópicos naturais, mostrando que existe uma discriminação do 13C destas folhas para a formação dos tecidos de reserva destes frutos.
A diferença dos valores isotópicos entre os ramos não foi significativa,
contudo o ramo 02 apresentou constituintes com valores isotópicos (-29,28 ‰), levemente
inferiores quando comparados com o ramo 01 (-28,98‰), sugerindo uma leve diluição
isotópica devido às diferentes idades fenológicas existente entre os ramos. Por conseguinte, no
ramo 02, os frutos apresentaram maiores valores de δ13C (-28,40‰) que as brotações (-29,98),
mostrando as tendências da diluição isotópica entre os drenos da planta.
Considerando que a planta estudada possuía 07 anos de idade, ou seja,
uma planta ainda jovem, visto que as figueiras podem chegar a 30 anos de idade com
produção satisfatória, foi observado um pequeno gradiente da distribuição natural do valor
δ13C, com tendência a aumento nos tecidos lenhosos novos, como a parte apical do ramo e as
radicelas, todavia, nos tecidos mais velhos da planta foi verificado a predominância do 12C.
(Tabela 4). Contudo, o valor isotópico médio -28,94 ± 0,35‰ verificado para os resíduos
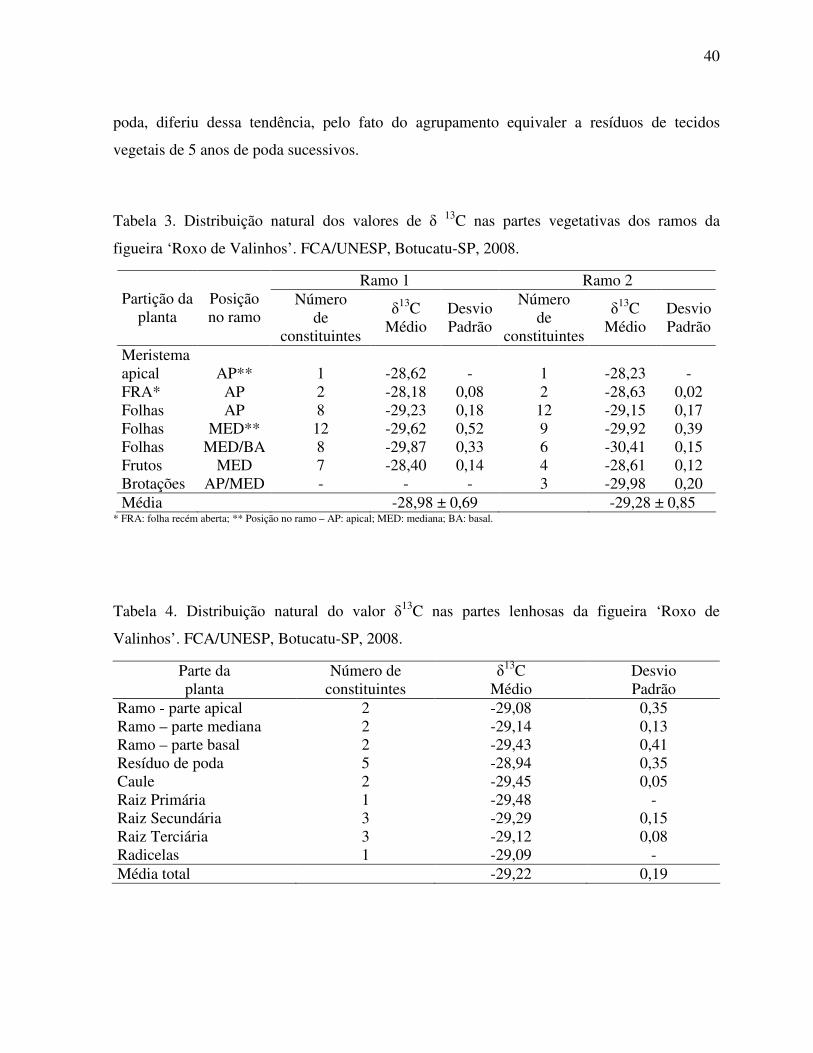
40
poda, diferiu dessa tendência, pelo fato do agrupamento equivaler a resíduos de tecidos
vegetais de 5 anos de poda sucessivos.
Tabela 3. Distribuição natural dos valores de δ 13C nas partes vegetativas dos ramos da
figueira ‘Roxo de Valinhos’. FCA/UNESP, Botucatu-SP, 2008.
Ramo 1 Ramo 2 Partição da
planta
Posição no ramo
Número de
constituintes
δ13C
Médio Desvio Padrão
Número de
constituintes
δ13C
Médio Desvio Padrão
Meristema apical AP** 1 -28,62 - 1 -28,23 - FRA* AP 2 -28,18 0,08 2 -28,63 0,02 Folhas AP 8 -29,23 0,18 12 -29,15 0,17 Folhas MED** 12 -29,62 0,52 9 -29,92 0,39 Folhas MED/BA 8 -29,87 0,33 6 -30,41 0,15 Frutos MED 7 -28,40 0,14 4 -28,61 0,12 Brotações AP/MED - - - 3 -29,98 0,20 Média -28,98 ± 0,69 -29,28 ± 0,85
* FRA: folha recém aberta; ** Posição no ramo – AP: apical; MED: mediana; BA: basal.
Tabela 4. Distribuição natural do valor δ13C nas partes lenhosas da figueira ‘Roxo de
Valinhos’. FCA/UNESP, Botucatu-SP, 2008.
Parte da planta
Número de constituintes
δ13C
Médio Desvio Padrão
Ramo - parte apical 2 -29,08 0,35 Ramo – parte mediana 2 -29,14 0,13 Ramo – parte basal 2 -29,43 0,41 Resíduo de poda 5 -28,94 0,35 Caule 2 -29,45 0,05 Raiz Primária 1 -29,48 - Raiz Secundária 3 -29,29 0,15 Raiz Terciária 3 -29,12 0,08 Radicelas 1 -29,09 - Média total -29,22 0,19

41
Não foram verificadas diferenças entre, os valores de δ13C das partes
vegetativas (Tabela 03) e lenhosas (Tabela 04), apresentando uma média geral de -29,13 ±
0,46‰. Por esse motivo, o intervalo de confiança, calculado para determinar o valor limite
inferior, para o valor do enriquecimento relativo da razão 13C/12C, das distintas partes será o
mesmo.
O limite mínimo do δ13C foi determinado pela equação:
Lm = Média do δ13C – IC
em que: Lm – é o valor limite mínimo do δ13C; IC – é o valor do
intervalo de confiança, ao nível de 1% de probabilidade.
O valor de Lm obtido, foi:
Lm = -29,13‰ - (-0,88‰) = -28,25‰
Como a precisão do espectrômetro de massa 13CO2 utilizado, é de
±0,2‰, o limite mínimo determinado de δ13C seria de -28,25‰ - (0,2‰) = -28,05‰. Porém,
como o menor valor do enriquecimento natural de δ13C obtido, nas determinações das
amostras avaliadas no experimento, foi de -28,12‰ (não apresentado nas tabelas por ser valor
absoluto de uma amostra), portanto 0,13‰ inferior ao limite mínimo calculado, o limite
mínimo de δ‰13C, PDB considerado, para uma maior segurança na identificação do
enriquecimento em 13C, será: -28,05‰ – (- 0,13‰) = -27,92‰. Dessa forma, o valor do limite
mínimo será o novo padrão para a determinação do enriquecimento ou não das amostras em 13C, permitindo inferir que, amostras que apresentam valores maiores que -27,92‰, indicam
estarem mais ricas em 13C que o padrão, sendo este enriquecimento reflexo da translocação e
alocação dos fotoassimilados enriquecidos em 13C produzidos pelas folhas marcadas,
indicando que essas amostras são consideradas drenos para a folha fonte marcada (alimentada
com 13CO2).
Na régua isotópica (Figura 4), estes dados podem ser mais facilmente
visualizados, a partir de que valores de δ13C as amostras podem ser consideradas ricas em 13C.

42
Figura 4. Régua isotópica, apresentando os sentidos de enriquecimento em 13C, em relação ao
padrão numérico Lm, onde Xi representa as amostras comparadas ao padrão Lm.
Em uma caracterização geral das figueiras, verificou-se que as raízes
primárias apresentaram os maiores valores médios de massa seca 270,5 g, representando 38%
de toda a massa seca da planta e, por conseguinte, 36% do carbono total encontrado em uma
figueira de 7 anos de idade. Os órgãos em crescimento foram os que arraigaram os menores
valores médios de massa seca e carbono total, onde as partições seguem ordem crescente de
armazenamento de carbono total de: gema apical < frutos < radicelas < brotações < folhas
novas, quando verificadas suas quantidades na planta como um todo (Tabela 5).
O sistema radicular da figueira representa 55,68% da distribuição total
de carbono da planta, ou seja, praticamente, acima de 50% do CO2 assimilado pela planta
através das trocas gasosas, foi alocado para o crescimento e manutenção do sistema radicular,
enquanto que os órgãos de sustentação da planta (resíduos de podas, ramos e caule),
representam juntos 42,5 %.
Em contrapartida, a massa vegetativa representa apenas 3,21% e
2,67% de toda massa seca e carbono assimilado pela planta, sendo as folhas os órgãos
vegetativos onde ocorre a maior demanda de carbono. As folhas da figueira pelos dados de
massa seca apresentados convertem uma elevada quantidade de CO2 atmosférico em lenho
(esqueleto de carbono), visto que são pelas folhas que ocorre a entrada do CO2 substrato das
enzimas que atuam na fotossíntese.

43
Tabela 5. Distribuição natural de massa seca e carbono total em figueiras ‘Roxo de Valinhos’.
FCA/UNESP, Botucatu-SP, 2008.
Massa seca (g) Carbono total (g) Distribuição (%) Partição
Média* Desvio Média* Desvio Massa seca** Carbono total Gema apical 0,14 e 0,05 0,056 d 0,02 0,02 0,02 Folhas novas 2,08 e 0,15 0,83 d 0,09 0,29 0,23 Folhas adultas 17,63 d 0,28 7,39 c 0,12 2,49 2,08 Frutos 1,30 e 0,20 0,55 d 0,09 0,18 0,16 Brotações 1,65 e 1,07 0,65 d 0,40 0,23 0,18 Ramo 50,94 cd 5,18 25,44 bc 2,39 7,19 7,16 Resíduos de poda 41,84 cd 20,85 24,57 bc 2,09 5,91 6,91 Caule 208,45 b 100,59 98,05 ab 46,81 29,44 27,58 Raízes primárias 270,51 a 93,00 128,11 a 58,74 38,21 36,04 Raízes secundárias 94,73 c 40,71 42,83 b 18,11 13,38 12,05 Raízes terciárias 17,23 d 6,78 26,27 bc 2,04 2,43 7,39 Radicelas 1,48 e 0,82 0,73 d 0,40 0,21 0,20
Total 707,97 - 355,47 - 100 100 *Médias seguidas da mesma letra na coluna não diferem pelo Teste de Tukey a 5% de probabilidade. **Média de 4 plantas, com 4 ramos cada.
A relação existente entre a massa seca de qualquer partição da figueira
com o carbono total foi altamente significativa, visto que em média a cada 100g de massa
seca, são esperadas 46,8 g de carbono total da planta como um todo. Analisando cada partição
em particular, não foram encontradas relações diferentes da apresentada na Figura 5.
0 50 100 150 200 250 300 350 400 450 5000
50
100
150
200
250
CT = 0,4682 MS
r2 = 0,9989n = 225
Car
bono
tota
l (g)
Massa seca (g) Figura 5. Relação existente entre massa seca de uma dada partição e o carbono total existente.
FCA/UNESP, Botucatu-SP, 2008.

44
4.3. Partição de Fotoassimilados em função dos drenos
Os dados morfológicos das plantas, como comprimento dos ramos,
número de folhas, número de entrenós, número de brotações por ramos e número de frutos do
experimento fonte-dreno revelam a proximidade morfológica das plantas escolhidas para o
enriquecimento, não havendo diferenças estatísticas para os parâmetros de número de folhas
por ramo, comprimento do ramo e número de frutos por planta (Tabela 6), permitindo
evidenciar a resposta fisiológica das plantas, em função do dreno presente, ou seja, é a
proximidade destes parâmetros morfológicos nas diferentes plantas que nos garantem que a
resposta da translocação com o uso do 13C é devido ao dreno em questão e não a fatores
genéticos ou ambientais. Contudo, a diferença observada no número de brotações e ausência
de frutos em duas plantas caracteriza os tratamentos adotados.
Tabela 6. Dados morfológicos médios das plantas alimentadas com Carbono-13, no
experimento fonte-dreno. FCA/UNESP, Botucatu-SP, 2008.
Tratamento Nº
Folhas Comprimento do ramo (cm)
Número de entrenós
Número de brotações
Número de frutos
Planta com brotações e frutos 6,8 a 124,5 a 46,8 a 3,0 a 0,3 a Planta com frutos 6,3 a 99,4 a 45,0 ab 0,0 b 1,3 a Planta com brotações 7,0 a 99,0 a 37,3 b 1,8 ab 0,0 a Planta sem drenos 6,8 a 104,0 a 43,8 ab 0,0 b 0,0 a
CV (%) 8,9 14,7 9,8 33,6 10,8 *Médias seguidas da mesma letra na coluna não diferem pelo Teste de Tukey a 5% de probabilidade.
A temperatura é um dos fatores externos de maior importância
relacionada à fotossíntese, pois as enzimas participantes desse processo necessitam de faixas
de temperaturas específicas para o seu funcionamento, para Larcher (2000), essa faixa para as
plantas C3, onde ocorre a aumento da abertura estomática situa-se entre 15 e 35 ºC.
As plantas foram enriquecidas na transição entre o verão e o outono de
2008, sendo a planta com presença de frutos e brotos e a planta apenas com frutos
enriquecidas no dia 17/03, enquanto que a planta apenas com brotações e a planta sem drenos

45
foram enriquecidas no dia 16/03. Ambos os enriquecimentos ocorreram entre as 09:00 e 11:00
horas. Os dados de temperatura do ar e radiação solar global, medidas a cada 15 minutos, no
ambiente externo à câmara de enriquecimento são apresentados na Figura 6.
Foram observadas variações de temperatura dentro da câmara de
enriquecimento em relação ao ambiente externo, todavia, estas variações não impediram a
assimilação de CO2 e do carbono marcado. Para a planta apenas com brotações, houve um
incremento de 4,5 ºC, do momento inicial ao final do enriquecimento dentro da câmara, e em
relação à temperatura externa em média a temperatura dentro da câmara foi 3,5 ºC superior.
Para a planta sem drenos foi verificado um aumento de 6,5º C entre o inicio e fim do
enriquecimento dentro da câmara, sendo em média a temperatura dentro da câmara 6,9 ºC
superior a temperatura ambiente. Neste último caso, a média de temperatura dentro da câmara
foi de 26,8 ºC, corroborando com dados supracitados por Larcher (2000). Essa diferença no
incremento de temperatura dentro da câmara de 2,4 ºC entre os dois enriquecimentos são
justificados pelo aumento da radiação solar global na parte final do enriquecimento da planta
sem drenos, observado na Figura 6b.
No tratamento onde os drenos foram os frutos, ocorreu um aumento de
2,5 ºC entre o inicio e o final do enriquecimento dentro da câmara, sendo em média a
temperatura dentro da câmara 2,3 ºC superior a temperatura externa. Já para a planta com
brotações e frutos, houve um aumento de 3,0 ºC do inicio ao fim do enriquecimento dentro da
câmara. Em média a temperatura dentro da câmara foi 6,8 ºC superior a temperatura ambiente
(26,5 ºC), causada pelo aumento instantâneo da radiação solar global ao final do
enriquecimento (aproximadamente 0,4 W m-2).
A exposição de plantas jovens à luz solar plena pode levar ao aumento
significativo na temperatura foliar que, eventualmente, causa quedas drásticas no potencial
hídrico da folha e intensifica os efeitos da fotoinibição (VALLADARES e PEARCY, 1997).
As temperaturas consideradas moderadamente altas (35 a 42°C) podem causar danos diretos
ao aparato fotossintético (WISE et al., 2004), por provocar mudanças na membrana do
tilacóide e alterar as propriedades físico-químicas, e também a organização funcional dessas
estruturas celulares (BERRY e BJÖRKMAN, 1980). Em elevadas temperaturas, a taxa
máxima de assimilação de carbono pode ser inibida por diminuir a condutância estomática
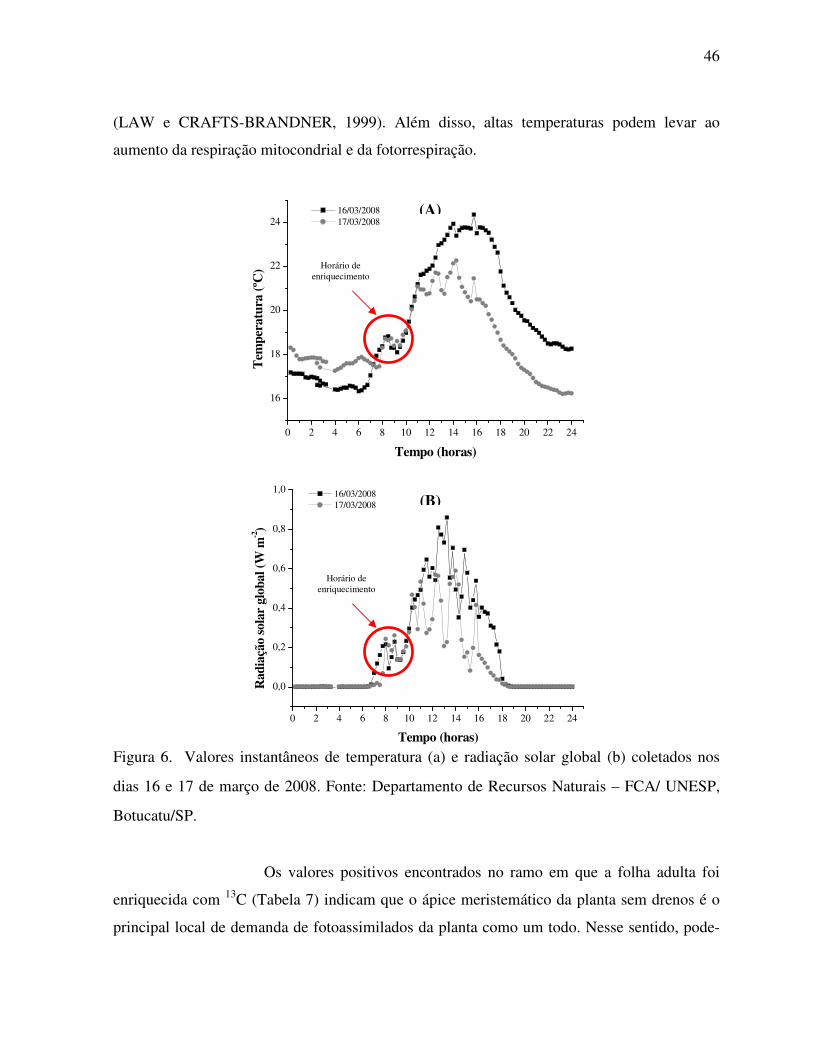
46
(LAW e CRAFTS-BRANDNER, 1999). Além disso, altas temperaturas podem levar ao
aumento da respiração mitocondrial e da fotorrespiração.
0 2 4 6 8 10 12 14 16 18 20 22 24
16
18
20
22
24 16/03/2008 17/03/2008
Tem
pera
tura
(ºC
)
Tempo (horas)
0 2 4 6 8 10 12 14 16 18 20 22 24
0,0
0,2
0,4
0,6
0,8
1,0 16/03/2008 17/03/2008
Rad
iaçã
o so
lar
glob
al (W
m-2)
Tempo (horas) Figura 6. Valores instantâneos de temperatura (a) e radiação solar global (b) coletados nos
dias 16 e 17 de março de 2008. Fonte: Departamento de Recursos Naturais – FCA/ UNESP,
Botucatu/SP.
Os valores positivos encontrados no ramo em que a folha adulta foi
enriquecida com 13C (Tabela 7) indicam que o ápice meristemático da planta sem drenos é o
principal local de demanda de fotoassimilados da planta como um todo. Nesse sentido, pode-
Horário de enriquecimento
Horário de enriquecimento
(A)
(B)

47
se dizer que mesmo durante a fase reprodutiva a planta não cessa o seu crescimento, pois os
carboidratos tornaram-se substratos universais para a respiração e o ponto de partida para
diferentes biossínteses.
Tabela 7. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de Valinhos’ sem
a presença de drenos. FCA/UNESP, Botucatu-SP, 2008.
Partição Ramo 1 Ramo 2 Ramo 3 Ramo 4 Parte vegetativa
Gema Apical 118,3 -28,84 -28,4 -28,31 FNP* 68,21 -28,95 -29,61 -28,89 FNM** -25,88 -30,11 -29,49 -30,43 F3 -28,76 -29,7 -29,24 -29,54 F4 132,16 -29,64 -28,61 -28,61 F5 -29,01 -29,57 -29,52 -28,55 F6 -28,56 -29,00 -28,88 -28,56 F7 -28,37 - -29,02 -28,09
Parte lenhosa Ramo - parte apical -21,83 -28,47 -28,50 -28,36 Ramo - parte mediana -24,63 -28,52 -28,66 -28,56 Ramo - parte basal -26,45 -28,75 -28,61 -28,47 Resíduo de poda -28,26 - -28,57 - Caule -28,42 - - - Raiz Primária -28,03 - - - Raiz Secundária -28,26 - - - Raiz Terciária -28,26 - - - Radicelas -16,5 - - - *FNP-Folha nova pequena; ** FNM- Folha nova média.
Outra consideração importante, é que houve variação do
enriquecimento relativo de δ13C apenas no ramo que continha a folha enriquecida e nas
radicelas, mostrando a tendência de translocação realizada pela planta dos fotoassimilados
produzidos por um ramo não serem alocados em outro ramo, independente de pertencerem ao
mesmo resíduo de poda ou não. Nos demais ramos (2, 3 e 4), em todas as partições estudadas,
de acordo com a régua isotópica (Figura 4) não houve enriquecimento relativo.
O maior enriquecimento da parte lenhosa foi encontrado no sistema
radicular, mais especificamente nas radicelas (Figura 7), que são órgãos de reserva da planta.
Conforme constatato por Matsuura et al. (2001) através da técnica isotópica, com a exposição

48
de folhas de Ficus carica L. 'Masui Daufine', expostas a 13CO2 em outubro, que mostraram 13C-fotossintatos armazenados durante o período de dormência e remobilizados na primavera
seguinte. O excesso de átomos % 13C durante o período de dormência foi elevado nas raízes
que apresentavam o menor diâmetro, decrescendo seguidamente por raízes finas, raízes de
tamanho intermediário (médio), raízes com maior diâmetro, tronco, ramos de dois anos de
idade e ramos de um ano de idade.
Figura 7. Valores de enriquecimento relativo de δ13C em figueiras ‘Roxo de Valinhos’ sem a
presença de drenos, sendo a 4ª folha alimentada. FCA/UNESP, Botucatu-SP, 2008.
AP: parte apical do ramo MD: parte mediana do ramo BA: parte basal do ramo RP1 e RP2: resíduos de poda RP: raiz primária RS: raiz secundária RT: raiz terciária Rd: radicelas
118,30 -28,84 -28,40 -28,31
68,21* -28,95* -29,61* -28,89*
-25,88** -30,11** -29,49** -30,43**
-28,76 -29,64 -28,61 -28,61
132,16 -29,70 -29,24 -29,54
-29,01 -29,57 -29,52 -28,55
-28,56 -29,00 -28,88 -28,56
-28,37 -29,02 -28,09
AP: -21,83 MD: -24,63 BA: -26,45
AP: -28,50 MD: -28,66 BA: -28,61
AP: -28,36 MD: -28,56 BA: -28,47
AP: -28,47 MD: -28,52 BA: -28,75
RP1: -28,26 RP2: -28,57 Caule -28,42
RP: -28,03 RS: -28,26 RT: -28,26 Rd: -16,50

49
Fica evidente que a alocação de fotoassimilados em plantas onde não
existem órgãos drenos acontece para todas as partes da planta, possibilitando a esta, o estoque
de reservas em órgãos lenhosos como ramos e raízes. Tendo o amido e a sacarose como
fotossintatos de grande importância acumulados pelas plantas, sendo o primeiro o carboidrato
de reserva mais abundante e, encontrado em folhas, diferentes tipos de hastes e raízes, flores,
frutos e sementes, utilizado como fonte de energia durante períodos de dormência, estresse ou
início de crescimento (LAPOINTE, 1998).
Observa-se que na planta onde os drenos são as brotações, que as
partições mantiveram o mesmo padrão de distribuição ao longo da planta, ou seja, o marcador
que entra na forma de CO2 é visto apenas no ramo que possuía a 4ª folha enriquecida (Tabela
8). A brotação presente neste ramo apresentou valores isotópicos que a caracterizou como
enriquecida, contudo nota-se que houve uma grande diluição isotópica na presença das demais
brotações da planta, pois a mesma quantia de carbono-13 foi aplicada nas quatro plantas
estudadas, e nesta planta em particular, menores valores isotópicos foram constatados.
Neste tratamento ocorreu uma mudança da relação fonte e dreno, visto
que na presença de um maior número de brotações, a alocação de fotoassimilados para o
sistema radicular ficou prejudicada, não sendo evidenciado em nenhuma parte lenhosa da
planta sinais de enriquecimento relativo com carbono-13. Segundo Foyer e Galtier (1996), este
fenômeno pode ser influenciado por características morfológicas e fisiológicas da fonte
(órgãos fotossintetizantes) e do dreno (órgãos consumidores dos metabólicos fotossintetizados,
principalmente carboidratos). Toda a produção de fitomassa depende da atividade
fotossintética da fonte, porém a assimilação do CO2 é apenas um dos muitos fatores que
influenciam o crescimento e desenvolvimento vegetal.
Apesar da grande demanda de fotoassimilados gerada pelas brotações
presentes na planta, o padrão de alocação de fotoassimilados para o ápice meristemático foi
mantido, contudo foram verificadas menores quantidades de 13C quando comparadas com a
planta sem drenos. Nos tecidos fotossinteticamente ativos, como folhas maduras, a produção
de carboidrato é maior do que a necessidade para manutenção do seu metabolismo e
crescimento, desta forma, exportam excedentes, na forma de sacarose, para tecidos que são
fotossinteticamente menos ativos ou inativos, como folhas jovens , raízes cachos ou ramos
(DANTAS et al., 2007).
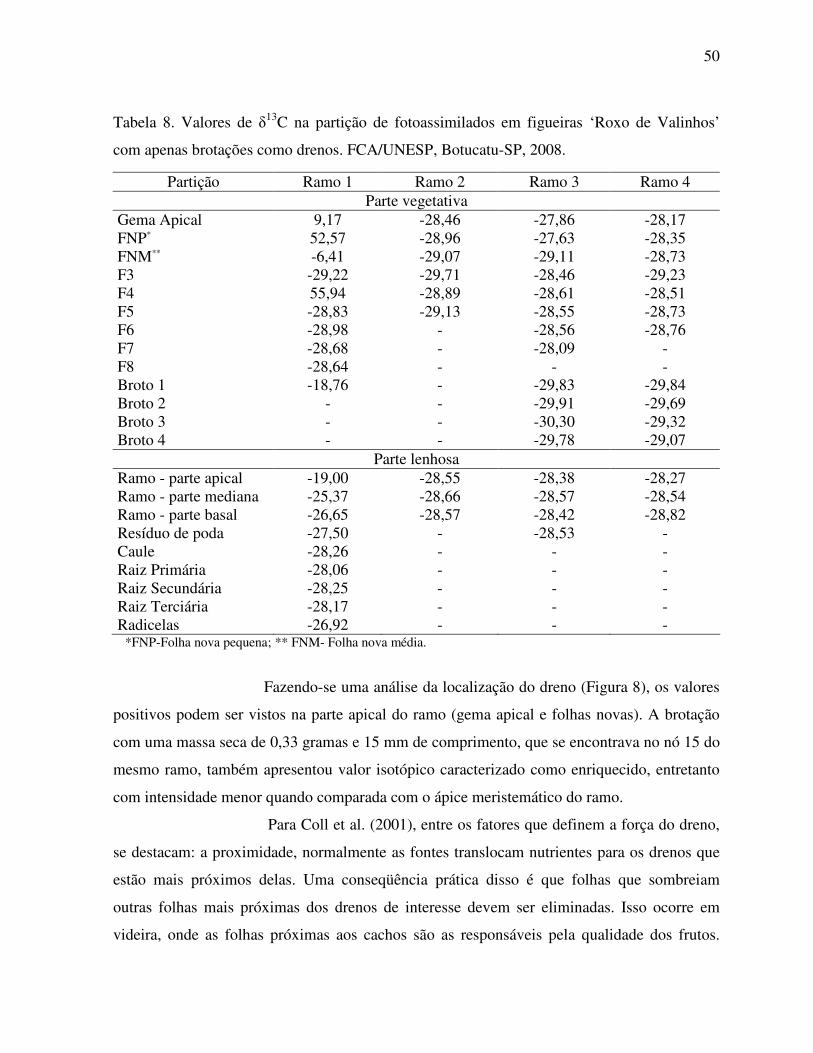
50
Tabela 8. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de Valinhos’
com apenas brotações como drenos. FCA/UNESP, Botucatu-SP, 2008.
Partição Ramo 1 Ramo 2 Ramo 3 Ramo 4 Parte vegetativa
Gema Apical 9,17 -28,46 -27,86 -28,17 FNP* 52,57 -28,96 -27,63 -28,35 FNM** -6,41 -29,07 -29,11 -28,73 F3 -29,22 -29,71 -28,46 -29,23 F4 55,94 -28,89 -28,61 -28,51 F5 -28,83 -29,13 -28,55 -28,73 F6 -28,98 - -28,56 -28,76 F7 -28,68 - -28,09 - F8 -28,64 - - - Broto 1 -18,76 - -29,83 -29,84 Broto 2 - - -29,91 -29,69 Broto 3 - - -30,30 -29,32 Broto 4 - - -29,78 -29,07
Parte lenhosa Ramo - parte apical -19,00 -28,55 -28,38 -28,27 Ramo - parte mediana -25,37 -28,66 -28,57 -28,54 Ramo - parte basal -26,65 -28,57 -28,42 -28,82 Resíduo de poda -27,50 - -28,53 - Caule -28,26 - - - Raiz Primária -28,06 - - - Raiz Secundária -28,25 - - - Raiz Terciária -28,17 - - - Radicelas -26,92 - - -
*FNP-Folha nova pequena; ** FNM- Folha nova média.
Fazendo-se uma análise da localização do dreno (Figura 8), os valores
positivos podem ser vistos na parte apical do ramo (gema apical e folhas novas). A brotação
com uma massa seca de 0,33 gramas e 15 mm de comprimento, que se encontrava no nó 15 do
mesmo ramo, também apresentou valor isotópico caracterizado como enriquecido, entretanto
com intensidade menor quando comparada com o ápice meristemático do ramo.
Para Coll et al. (2001), entre os fatores que definem a força do dreno,
se destacam: a proximidade, normalmente as fontes translocam nutrientes para os drenos que
estão mais próximos delas. Uma conseqüência prática disso é que folhas que sombreiam
outras folhas mais próximas dos drenos de interesse devem ser eliminadas. Isso ocorre em
videira, onde as folhas próximas aos cachos são as responsáveis pela qualidade dos frutos.

51
Como critério geral, as folhas da porção superior da planta costumam translocar nutrientes
para as folhas novas e caules em crescimento e as folhas da porção basal tendem a exportar
para o sistema radicular; o desenvolvimento, durante a fase vegetativa os maiores drenos são
raízes e ápices caulinares e na fase reprodutiva os frutos se tornam dominantes; e a conexão
vascular, fontes translocam assimilados preferencialmente para drenos com os quais elas têm
conexão vascular direta. O carregamento do floema na região da fonte envolve o movimento
dos produtos dos cloroplastos nas células do mesofilo para as células do tubo crivado, esse
processo ocorre nas nervuras terminais das folhas (TAIZ e ZEIGER, 2004; ZHANG et al.
2005a).
Figura 8. Valores de enriquecimento relativo de δ13C em figueiras ‘Roxo de Valinhos’ com
apenas brotações como drenos, sendo a 4ª folha alimentada. FCA/UNESP, Botucatu-SP, 2008.
9,17 -28,46 -27,86 -28,17
52,57* -28,96* -29,63* -28,35*
-6,41** -29,07** -29,11** -28,73**
-29,22 -29,07 -28,46 -29,23
55,94 -29,71 -28,61 -28,51
-28,83 -28,89 -28,55 -28,73
-28,98 -29,13 -28,56 -28,76
-28,68 -28,09
Br1: -29,83
AP: -19,00 MD: -25,37 BA: -26,65
AP: -28,38 MD: -28,57 BA: -28,42
AP: -28,27 MD: -28,54 BA: -28,82
AP: -28,55 MD: -28,66 BA: -28,57
RP1: -27,50 RP2: -28,53 Caule -28,26
RP: -28,06 RS: -28,25 RT: -28,17 Rd: -26,92
-28,64
Br1: -18,76
Br2: -29,91 Br3: -30,30
Br4: -29,78
Br1: -29,84
Br2: -29,69
Br3: -29,32
Br4: -29,07
AP: parte apical do ramo MD: parte mediana do ramo BA: parte basal do ramo RP1 e RP2: resíduos de poda RP: raiz primária RS: raiz secundária RT: raiz terciária Rd: radicelas Br: brotações

52
Nota-se, ainda que apesar da grande diluição isotópica existente no
tratamento onde os drenos foram às brotações, as partes lenhosas do ramo (apical, mediana e
basal) ficaram levemente enriquecidos em relação ao padrão, bem como o resíduo de poda e
também as radicelas. Esse fato evidencia que apesar das brotações dos demais ramos (onde
não houve enriquecimento) contribuírem para a diluição, existe uma grande força dos órgãos
lenhosos, caracterizando claramente as partições de movimentação dos fotoassimilados e os
órgãos de reserva (radicelas). Em relação a essas brotações, provavelmente sejam suas
próprias folhas fontes que as sustentem, havendo um equilíbrio na rota de translocação da
planta, restringindo a mesma a curtas distâncias.
Os resultados do enriquecimento relativo de δ13C da planta que
apresentava frutos como dreno são apresentados na Tabela 9. Dadas as condições de estudo,
onde a mesma quantidade de 13C foi injetada na câmara de enriquecimento para as quatro
plantas e todas as plantas apresentavam o mesmo estádio fenológico, podemos considerar que,
na comparação entre os drenos frutos com os drenos brotações, os frutos se apresentaram
como drenos mais fortes do que as brotações, visto que foi observado o enriquecimento
relativo de todos os frutos presentes na planta (em dois ramos distintos, porém pertencentes ao
mesmo resíduo de poda), enquanto que apenas a brotação presente no ramo que continha a
folha enriquecida apresentou valor isotópico superior ao limite mínimo para enriquecimento
(Figura 8).
Valores isotópicos maiores podem ser vistos na parte apical, local este
que apresentava folhas jovens em desenvolvimento, portanto dreno preferencial em relação ao
fruto. Nas fontes, os mecanismos reguladores da alocação determinam as quantidades de
carbono fixado, que serão armazenadas, normalmente como amido, metabolizadas nas células
da fonte, ou imediatamente transportada para os tecidos-dreno (HOPKINS, 1995).
Houve um fracionamento isotópico do 13C nesta planta, pois foi
constatado o enriquecimento relativo dos demais ramos, principalmente nos tecidos novos
quando comparados ao valor padrão delimitado pela régua isotópica (Figura 4).
Provavelmente esse fenômeno foi causado pela renovação constante da relação 12C/13C através
da assimilação e incorporação do CO2 pelo ciclo de Calvin Benson.
Existe uma relativa competição entre os frutos drenos do ramo
marcado, pois os mesmos apresentavam o mesmo padrão de desenvolvimento, tendo o fruto 1

53
dimensões de 2,75 x 4,84 mm (diâmetros equatorial e longitudinal, respectivamente) e massa
seca de 1,22 g; o fruto 2, dimensões de 2,76 x 4,90 mm e massa seca de 1,36 g e o fruto 3,
dimensões de 2,86 x 5,37 mm e massa seca de 1,49 g). Permitindo evidenciar que a
localização do dreno interferiu na alocação do fotoassimilado, visto que o fruto 2 estava na
mesma filotaxia da 4ª folha alimentada (Figura 9).
A força de um dreno pode ser determinada através de seu tamanho
(massa total do tecido) e atividade (taxa de importação e assimilados), mas também depende
de outros fatores como, suprimento de carboidratos, presença de outros drenos e resistência ao
transporte ao longo do floema (TORPE e MINCHIM, 1996).
Tabela 9. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de Valinhos’
com frutos como drenos. FCA/UNESP, Botucatu-SP, 2008.
Partição Ramo 1 Ramo 2 Ramo 3 Ramo 4 Parte vegetativa
Gema Apical 119,51 -27,6 -28,23 -27,78 FNP* 371,19 -28,53 -28,41 -27,64 FNM** -28,85 -28,99 -29,7 -29,17 F3 -28,85 -29,78 -29,3 -28,69 F4 218,68 -29,28 -29,21 -28,97 F5 -27,79 -29,59 -28,14 -27,66 F6 -28,48 -28,38 -28,55 -28,59 F7 - -28,58 - - F8 - - - - Fruto 1 -27,00 -27,56 - - Fruto 2 -25,33 -27,74 - - Fruto 3 -27,87 - - -
Parte lenhosa Ramo - parte apical -14,85 -28,10 -28,28 -28,37 Ramo - parte mediana -30,26 -28,18 -28,3 -28,31 Ramo - parte basal -17,92 -28,53 -28,31 -28,82 Resíduo de poda -27,57 - -28,88 - Caule -28,63 - - - Raiz Primária -27,58 - - - Raiz Secundária -28,15 - - - Raiz Terciária -28,84 - - - Radicelas -17,05 - - -
*FNP-Folha nova pequena; ** FNM- Folha nova média.

54
A parte lenhosa, da planta que apresentava apenas frutos como drenos,
concentraram a maior quantidade de 13C, quando comparado com os demais tratamentos,
mantendo a tendência de armazenamento de fotoassimilados marcados nos ramos (apical e
basal), bem como no sistema radicular, tendo as radicelas como o principal órgão armazenador
de fotoassimilados da planta. Esses dados corroboram com os encontrados por Simkhada et al.
(2007) em raízes de Diopyros kaki, usando o 13C como marcador.
Figura 9. Valores de enriquecimento relativo de δ13C em figueiras ‘Roxo de Valinhos’ com
apenas frutos como drenos, sendo a 4ª folha alimentada. FCA/UNESP, Botucatu-SP, 2008.
119,51 -27,60 -28,23 -27,78
371,19* -28,53* -28,41* -27,64*
-28,85** -28,99** -29,70** -29,17**
-28,85 -29,78 -29,30 -28,69
218,68 -29,28 -29,21 -28,97
-27,79 -29,59 -28,14 -27,66
-28,48 -28,38 -28,55 -28,59
Fr1: -27,56
AP: -14,85 MD: -30,26 BA: -17,92
AP: -28,28 MD: -28,30 BA: -28,31
AP: -28,37 MD: -28,31 BA: -28,82
AP: -28,10 MD: -28,18 BA: -28,53
RP1: -27,57 RP2: -28,88 Caule
-28,63
RP: -27,58 RS: -28,15 RT: -28,84 Rd: -17,05
Fr1: -27,00
Fr2: -27,74 Fr2: -25,33 Fr3: -27,87
-28,58
AP: parte apical do ramo MD: parte mediana do ramo BA: parte basal do ramo RP1 e RP2: resíduos de poda RP: raiz primária RS: raiz secundária RT: raiz terciária Rd: radicelas Fr: frutos

55
Quando comparado as partes lenhosas dos tratamentos onde os drenos
foram separadamente o fruto e a brotação, constatou-se uma elevação na diluição isotópica das
plantas, pois nas partes lenhosas do tratamento brotação houve um menor acúmulo de 13C-
fotossintatos (Tabela 8), enquanto na presença de frutos houve um maior acumulo de
fotoassimilados nas partes lenhosas (Tabela 9), evidenciando que ocorreu uma maior taxa de
renovação de CO2, na massa vegetativa da planta onde existiam as brotações.
Durante o período de atividade fotossintética, o eventual excedente em
compostos fotoassimilados produzidos pela planta fica imobilizado na forma de carboidratos
insolúveis em órgãos aéreos e subterrâneos da planta, sendo, então, mobilizados
gradativamente em carboidratos solúveis durante o período de dormência. Com o fim da
dormência, essa mobilização é acelerada, sendo os carboidratos solúveis conduzidos para as
gemas em brotação que, por sua vez, formarão novos ramos e folhas; posteriormente, as flores
e os frutos são supridos, seguidos pelo câmbio, por novas gemas em formação e, finalmente,
pelos tecidos que servem como depósito de carboidratos em órgãos subterrâneos e aéreos da
planta (WARDLAW, 1990; LARCHER, 2000).
Para Herter et al. (2001), a época da mobilização dos carboidratos
presentes nos órgão lenhosos da planta está diretamente ligada aos eventos climáticos,
sobretudo à temperatura, tendo grande importância nos estudos de adaptação de frutíferas de
clima temperado. A intensidade dessa mobilização influencia, por sua vez, no
desenvolvimento fenológico da planta, como no crescimento de ramos, no florescimento e na
produção de frutos (LIU et al., 1999; LARCHER, 2000).
Na planta com a presença de brotações e frutos, o dreno preferencial
foi definido como o fruto, pelo maior valor isotópico (-23,75‰) quando comparado com os
demais valores isotópicos das brotações (Tabela 10e Figura 10). Essas informações
corroboram com os relatos de Rena e Carvalho (2003) em cafeeiro, que os frutos são drenos
preferenciais de fotoassimilados no período reprodutivo, existindo um elevado grau de
dependência do estado nutricional da planta e da relação folha e fruto.
Nos processos físicos, segundo Ducatti (2007) as pequenas diferenças
de comportamento podem conduzir ao fracionamento isotópico, uma variação nas
concentrações das várias espécies isotópicas 13C ou 12C em diferentes regiões de um sistema
físico sujeito a um determinado processo. Semelhantemente a esse fato, na planta onde os
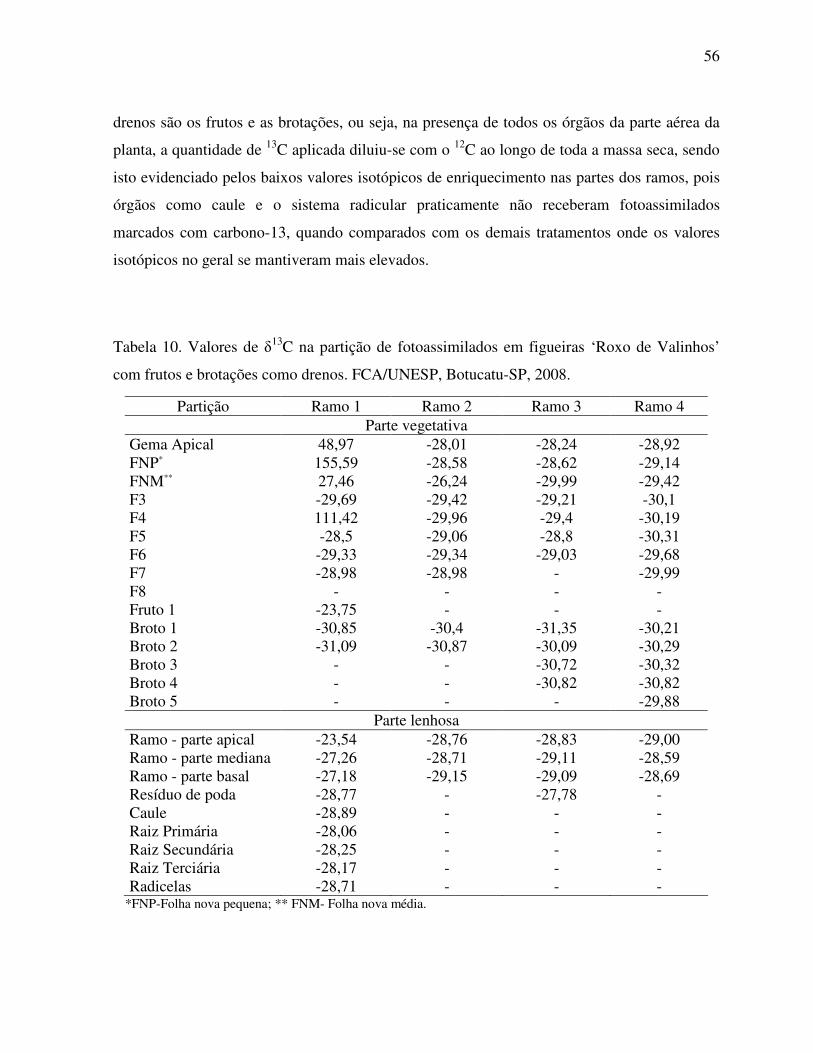
56
drenos são os frutos e as brotações, ou seja, na presença de todos os órgãos da parte aérea da
planta, a quantidade de 13C aplicada diluiu-se com o 12C ao longo de toda a massa seca, sendo
isto evidenciado pelos baixos valores isotópicos de enriquecimento nas partes dos ramos, pois
órgãos como caule e o sistema radicular praticamente não receberam fotoassimilados
marcados com carbono-13, quando comparados com os demais tratamentos onde os valores
isotópicos no geral se mantiveram mais elevados.
Tabela 10. Valores de δ13C na partição de fotoassimilados em figueiras ‘Roxo de Valinhos’
com frutos e brotações como drenos. FCA/UNESP, Botucatu-SP, 2008.
Partição Ramo 1 Ramo 2 Ramo 3 Ramo 4 Parte vegetativa
Gema Apical 48,97 -28,01 -28,24 -28,92 FNP* 155,59 -28,58 -28,62 -29,14 FNM** 27,46 -26,24 -29,99 -29,42 F3 -29,69 -29,42 -29,21 -30,1 F4 111,42 -29,96 -29,4 -30,19 F5 -28,5 -29,06 -28,8 -30,31 F6 -29,33 -29,34 -29,03 -29,68 F7 -28,98 -28,98 - -29,99 F8 - - - - Fruto 1 -23,75 - - - Broto 1 -30,85 -30,4 -31,35 -30,21 Broto 2 -31,09 -30,87 -30,09 -30,29 Broto 3 - - -30,72 -30,32 Broto 4 - - -30,82 -30,82 Broto 5 - - - -29,88
Parte lenhosa Ramo - parte apical -23,54 -28,76 -28,83 -29,00 Ramo - parte mediana -27,26 -28,71 -29,11 -28,59 Ramo - parte basal -27,18 -29,15 -29,09 -28,69 Resíduo de poda -28,77 - -27,78 - Caule -28,89 - - - Raiz Primária -28,06 - - - Raiz Secundária -28,25 - - - Raiz Terciária -28,17 - - - Radicelas -28,71 - - -
*FNP-Folha nova pequena; ** FNM- Folha nova média.

57
Os diferentes ramos da planta são, provavelmente, supridos de sua
demanda de fotoassimilados pelas folhas existentes em cada ramo, ou seja, folhas de um
determinado ramo não são fontes de fotoassimilado para outro ramo onde a identificação de
um tecido como fonte ou dreno depende da direção do transporte dos assimilados. Do ponto de
vista bioquímico as plantas exibem diferentes características de fonte (ZAMSKI, 1995).
Figura 10. Valores de enriquecimento relativo de δ13C em figueiras ‘Roxo de Valinhos’ com
frutos e brotações como drenos, sendo a 4ª folha alimentada. FCA/UNESP, Botucatu-SP,
2008.
48.97 -28,01 -28,24 -28,92
155,59* -28,58* -28,62* -29,14*
27,46** -26,24** -29,99** -29,42**
-29,69 -29,42 -29,21 -30,10
111,42 -29,96 -29,40 -30,19
-28,50 -29,06 -28,80 -30,31
-29,33 -29,34 -29,03 -29,68
Br1: -30,40
AP: -23,54 MD: -27,26 BA: -27,18
AP: -28,83 MD: -29,11 BA: -29,09
AP: -29,00 MD: -28,59 BA: -28,69
AP: -28,76 MD: -28,71 BA: -29,15
RP1: -28,77 RP2: -27,78 Caule
-28,89
RP: -28,06 RS: -28,25 RT: -28,17 Rd: -28,71
Fr1: -23,75
Br2: -30,87 Br1: -30,85
Br2: -31,09
-28,98 -28,98
Br2: -30,09
Br1: -31,35
Br3: -30,72
Br4: -30,82
Br1: -30,21
-29,99
Br2: -30,29
Br3: -30,32
Br4: -30,82
Br5: -29,88
AP: parte apical do ramo MD: parte mediana do ramo BA: parte basal do ramo RP1 e RP2: resíduos de poda RP: raiz primária RS: raiz secundária RT: raiz terciária Rd: radicelas Fr: frutos Br: brotações
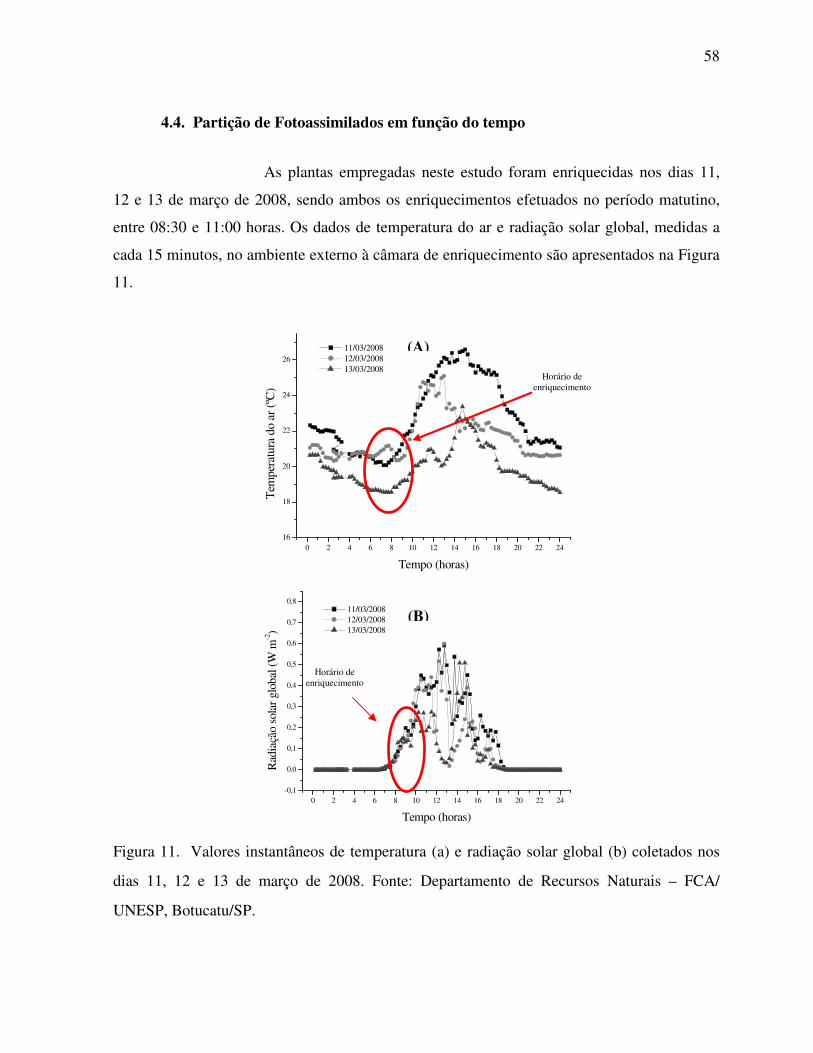
58
4.4. Partição de Fotoassimilados em função do tempo
As plantas empregadas neste estudo foram enriquecidas nos dias 11,
12 e 13 de março de 2008, sendo ambos os enriquecimentos efetuados no período matutino,
entre 08:30 e 11:00 horas. Os dados de temperatura do ar e radiação solar global, medidas a
cada 15 minutos, no ambiente externo à câmara de enriquecimento são apresentados na Figura
11.
0 2 4 6 8 10 12 14 16 18 20 22 2416
18
20
22
24
26
11/03/2008 12/03/2008 13/03/2008
Tem
pera
tura
do
ar (
ºC)
Tempo (horas)
0 2 4 6 8 10 12 14 16 18 20 22 24-0,1
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8 11/03/2008 12/03/2008 13/03/2008
Rad
iaçã
o so
lar
glob
al (
W m
-2)
Tempo (horas)
Figura 11. Valores instantâneos de temperatura (a) e radiação solar global (b) coletados nos
dias 11, 12 e 13 de março de 2008. Fonte: Departamento de Recursos Naturais – FCA/
UNESP, Botucatu/SP.
Horário de enriquecimento
Horário de enriquecimento
(B)
(A)

59
Dentro da câmara de enriquecimento foram verificadas temperaturas
médias de 23,4; 23,4; 27,00; 27,0; 27,1; 25,1 e 22,1 ºC, durante o período em que as plantas
foram alimentadas com 13C, com uma variação de 5,0; 4,1; 3,5; 3,0; 6,0; 5,5 e 4,0 ºC entre o
início e o fim do enriquecimento, para o estudo dos tempos de translocação de 6, 24, 48, 72,
120, 168 e 360 horas após o enriquecimento, respectivamente. Em média a temperatura
dentro da câmara foi 2,9; 4,2; 4,8; 3,9; 3,7; 3,2 e 1,8 ºC superior à temperatura ambiente
externa, para os mesmos tempos de translocação acima. Já a radiação solar global variou entre
0,1 e 0,4 W m-2 durante os enriquecimentos, favorecendo essas variações de temperatura entre
as diferentes alimentações.
Plantas de clima temperado seguem padrões específicos de deposição e
uso de carboidratos por toda planta de acordo com a estação (KRAMER e KOZLOWSKI,
1979). Esses padrões são modificados segundo as espécies (LOESCHER et al.,1990).
Ás 6 horas após o enriquecimento, a partição gema apical do ramo 1,
continha o maior valor isotópico dentre todas as gemas vegetativas (Tabela 11). Como todas
as partições do mesmo ramo estavam enriquecidas, o sentido de translocação dos
fotossintatos-13 foi direcionado para as demais gemas (meristemas apicais) da planta. O
tempo de seis horas após o enriquecimento pode ser considerado como um pequeno intervalo
de tempo, porém foi suficiente para a observação da translocação na figueira cultivada com
quatro ramos nas condições brasileiras, corroborando com o encontrado por Simkhada et al.
(2007) para o caqui.
Entre 6 e 24 horas após a alimentação com o gás 13C, o sinal isotópico
decresceu 92,4% nas gemas vegetativas, evidenciando o compartilhamento dos
fotoassimilados com os demais tecidos e órgãos da figueira.
Às 48 horas depois do enriquecimento, apesar do brusco decréscimo
do sinal isotópico, ainda foi constatado que a gema apical do ramo 1 apresentava sinais de
enriquecimento.
No entanto, em 72 horas pode ser observado um novo fluxo de
demanda de fotoassimilados pela partição, havendo uma elevação do sinal em todos os ramos.
Em 120 horas o sinal ficou positivo novamente apenas no ramo 1 e caindo para -14,45 ‰ às
168 horas, com tendência a variação natural com 360 horas (-25,07 ‰). Por conseguinte, o
órgão ainda estava com um valor 10,2% mais rico em fotossintatos-13 que o limite de

60
confiança (Figura 4), exigindo para que houvesse a renovação de todos os fotoassimilados-13
uma exposição de mais 10,8 horas de assimilação do CO2 atmosférico.
As gemas apicais dos demais ramos estavam enriquecidas 6 horas após
a alimentação, no entanto, foram verificados valores próximos ao valor limite da variação
natural do restante dos intervalos de tempo estudados. Apenas a gema apical do ramo 4
recebeu novo fluxo de fotossintatos ricos em carbono-13 após 360 horas (-26,29 ‰).
Simkhada et al,. (2007), também encontraram diferenças nos padrões da partição do 13C em
direção aos órgãos de caqui com respeito ao tempo depois de alimentação.
As folhas jovens da planta foram as partições (tecidos vegetais) onde
se evidenciaram os maiores sinais isotópicos dentre todas as partições estudadas (excluindo-se
a folha alimentada), porém, após 24 horas restava apenas 28,14% do valor isotópico do
primeiro tempo estudado (6 horas). Não obstante, com 48 horas o sinal continuava em queda,
evidenciando a entrada de novas taxas de CO2 atmosférico.
Um novo fluxo de seiva foi observado após as 72 horas para as folhas
jovens, visto que estes órgãos apresentavam contínuo crescimento em detrimento a fase
reprodutiva da planta, alcançando a sinalização positiva dentro da régua isotópica com valor
de 24,25 ‰. Este sinal ainda decresceu após as 120 horas (Tabela 11), elevou-se novamente
para 7,55 ‰, às 168 horas e decaiu no tempo de 360 horas para valores negativos, contudo
ficando bem acima dos valores da variação natural da planta, permitindo identificar as folhas
jovens como órgãos ricos em fotossintatos-13 após 15 dias do enriquecimento.
As demais folhas jovens dos ramos 2, 3 e 4 se apresentaram
enriquecidas somente até 24 horas, não sendo mais evidenciado sinal de enriquecimento após
48 horas da alimentação. Com 72 horas a média do sinal isotópico destas partições era de
-27,70 ‰, valor este muito próximo do limite de confiança da variação natural do δ 13C.
Entretanto, com120 horas essas partições não estavam enriquecidas, enquanto que após 168
horas foram verificados sinas de enriquecimento próximos ao limite estabelecido pela régua
isotópica (Figura 4). Ou seja, no intervalo de 360 horas após a alimentação, as folhas jovens
em franco desenvolvimento nos ramos 2, 3 e 4 renovaram a razão 12C/13C de suas células,
voltando ao valor padrão natural.
Os valores isotópicos de δ13C em ‰ da folha fonte adulta, que estava
acima da folha fonte alimentada no ramo 1, encontraram-se levemente enriquecidas entre o

61
tempo de 6 às 48 horas após a alimentação (Tabela 11). Um novo fluxo de translocação de
fotoassimilados-13, no período entre as 72 e 360 horas após a alimentação também foi
verificado para essa partição. Contudo, o maior valor isotópico dentre os tempos após a
alimentação foi evidenciado ás 120 horas (-20,28‰), sendo que essas folhas adultas
permaneceram com presença de 13C em seus tecidos até 360 horas após a alimentação. As
demais folhas adultas dos ramos 2, 3 e 4 na mesma posição não mostraram sinais de
enriquecimento, estando seus valores sempre próximos do valor limite da variação natural em
média -27,83,-28,22 ,-29,41, -29, -29,44, -28,84, -28,67‰, respectivamente às 6, 24, 48, 72,
120, 168 e 360 horas após a alimentação da folha adulta fonte presente no nó 4.
Assim como nas demais partições o maior valor isotópico foi
observado no tempo de 6 horas na folha alimentada, sendo este o valor mais elevado
encontrado em todo o experimento e esta partição manteve-se com valores positivos ao longo
das 360 horas. Observou-se uma queda do sinal isotópico na ordem de 92,3%, no período de
24 horas após a alimentação, e o menor valor foi verificado as 72 horas (5,01 ‰) após o
enriquecimento.
Nas demais folhas fontes dos outros ramos (2, 3 e 4), não foram
verificados sinais de enriquecimento, exceto a quarta folha do ramo 2 que possivelmente pela
proximidade dos ramos, ficou enriquecida em relação ao padrão (-27,92 ‰) no tempo de 6
horas. Como são folhas fontes, houve uma elevada taxa de renovação do CO2 atmosférico.
Constatou-se que a quantidade injetada de 8,0 ml do gás contendo 13C
dentro da câmara foi suficiente para que o 13C fosse rastreado nas demais partes da planta, pois
as folhas fontes escolhidas apresentavam as maiores áreas foliares e aparatos fotossintéticos
em perfeitas condições fisiológicas. Apesar da alta especificidade da enzima Rubisco pela
forma isotópica do 12C, carbono este com menor de massa (44), a concentração de 2273, 97 ‰
de 13C não foi o suficiente para saturar a enzima.
A partição das folhas fontes posicionadas abaixo da folha alimentada
apresentaram enriquecimento relativo de fotoassimilados-13 no tempo de 6 horas, em média
-1‰ mais pobre em 13C que as folhas adultas e fontes posicionadas acima da folha alimentada,
exportando mais carboidratos que a folha do nó 3, por exemplo. Nas folhas que se
encontravam na mesma posição (quinto nó), nos demais ramos não foram observados sinais de

62
enriquecimento e as taxas de renovação de CO2 possivelmente foram altas, havendo a troca do 13C pelo 12C.
Mantendo a tendência de enriquecimento no tempo de 6 horas após a
alimentação, assim como as demais partições, a brotação apresentou valor isotópico de
-2,34‰, não sendo observado sinal de enriquecimento nos tempos compreendidos entre 24 e
72 horas. Contudo, essa partição apresentou-se rica em fotossintatos-13 ás 120 horas após
alimentação, com tendência da partição em igualar o seu valor isotópico ao da variação natural
no período das 168 às 360 horas.
O valor de enriquecimento do fruto às 6 horas foi menor do que o valor
isotópico da brotação para o mesmo tempo, entretanto os frutos ainda estavam ricos em
fotossintatos-13, nos tempos de 24, 48 e 72 horas após a alimentação, se mantendo estáveis
nos tempos de 120 e 168 horas e enriquecidos novamente após as 360 horas da alimentação.
Nas primeiras 6 horas o dreno foi considerado a brotação, pois a
mesma se encontrou 6,5 vezes mais rica em 13C que o fruto, todavia, na competição entre
drenos, os frutos foram caracterizados como drenos preferenciais, visto que ocorreram
demandas de fotoassimilados por estes órgãos ao longo das 360 horas. Teng et al. (1999),
alimentando pêra ‘Nijisseiki’ na primavera encontrou 13C em frutos, folhas e brotos novos e a
quantidade de 13C nos órgãos vegetativos das partes aéreas aumentou, enquanto a quantidade
de 13C no fruto decaiu drasticamente.
A parte superior do ramo, considerada como apical (onde se
localizava a folha alimentada – 4ª folha), mostrou elevado valor isotópico no tempo de 6
horas, valor este acima dos evidenciados para as folhas acima e abaixo da folha alimentada,
brotações e frutos. Isto se deve ao fato de que essa é uma partição de transporte dos
fotoassimilados marcados e também pela grande quantidade de partições em franco
desenvolvimento celular (gemas e folhas novas). Após 24 horas da alimentação com 13C o
valor isotópico alcançou a sinalização negativa, decrescendo até as 72 horas, com uma nova
demanda de 13C a partir das 120 horas em torno de -3,2 ‰. Todavia, permaneceu rico em 13C
em relação ao valor isotópico da variação natural após 360 horas. Não foram verificados sinais
de enriquecimento nas partes apicais dos ramos 2, 3 e 4, pois os valores de δ13C se mantiveram
próximos ou abaixo do limite de confiança caracterizado na régua isotópica.

63
A parte mediana do ramo 1 esteve 62% mais pobre em 13C, que a parte
apical do ramo 1, para o tempo de 6 horas. Foi observado um decréscimo do sinal no tempo de
24 horas, ficando estável em -26 ‰ das 48 ás 168 horas após a alimentação. Neste caso, o
fluxo de translocação foi retardado em relação a parte apical do ramo. A parte mediana dos
ramos 2, 3 e 4 não apresentaram sinal de enriquecimento em nenhum dos tempos estudados,
tendo seus valores oscilando em torno de -27,09 ‰ a -28,06 ‰, próximos do valor limite de
confiança da variação natural (-27,09 ‰).
O valor isotópico encontrado na parte basal do ramo 1 ás 6 horas
depois da alimentação com 13C, apesar de positivo (20,92 ‰), ficou 75,3% e 60,2% abaixo
dos valores encontrados na parte apical e mediana, respectivamente. Por conseguinte, a parte
basal do ramo estava mais rica em fotossintatos-13 do que a parte mediana no tempo 24 horas,
tendo sua taxa de CO2 renovada com praticamente 48 horas, pois os valores isotópicos foram
semelhantes ao da variação natural. As 72 horas, nenhum sinal de enriquecimento relativo foi
evidenciado, enquanto que às 360 horas após o enriquecimento valores que permitiam
caracterizá-lo como enriquecido, mostrando o mesmo comportamento de estabilidade
isotópica do 13C observado na parte mediana no período das 48 às 168 horas. Nos demais
ramos, essa partição apresentou a mesma tendência de não enriquecimento ao longo das 360
horas.
A quantidade de exportação de fotoassimilados depende da
composição do ramo e tipo do ápice. À distância de translocação entre ramos e conexão dos
feixes vasculares são mais simples em ramos mais jovens que em galhos mais velhos (TENG
et al., 2002), e os resultados encontrados confirmam estas declarações, como mostrado pela
alta quantidade de 13C na parte mediana e apical que são próximas ao local de exposição do 13C no ramo.
O relacionamento fonte – dreno depende do grau de exposição leve,
localização das folhas no dossel e da carga de colheita. Os novos órgãos com altas taxas de
crescimento são referidos como centros de crescimento que, governam a atividade relativa, ou
capacidade da fonte. Estes diferentes centros de crescimento podem influenciar o padrão de
assimilação e distribuição de fotoassimilados, corroborando com os maiores valores de
enriquecimento encontrados nas partições apicais no menor tempo estudado (QUINLAN e
PRESTON, 1971; QUINLAN e WEAVER, 1970).
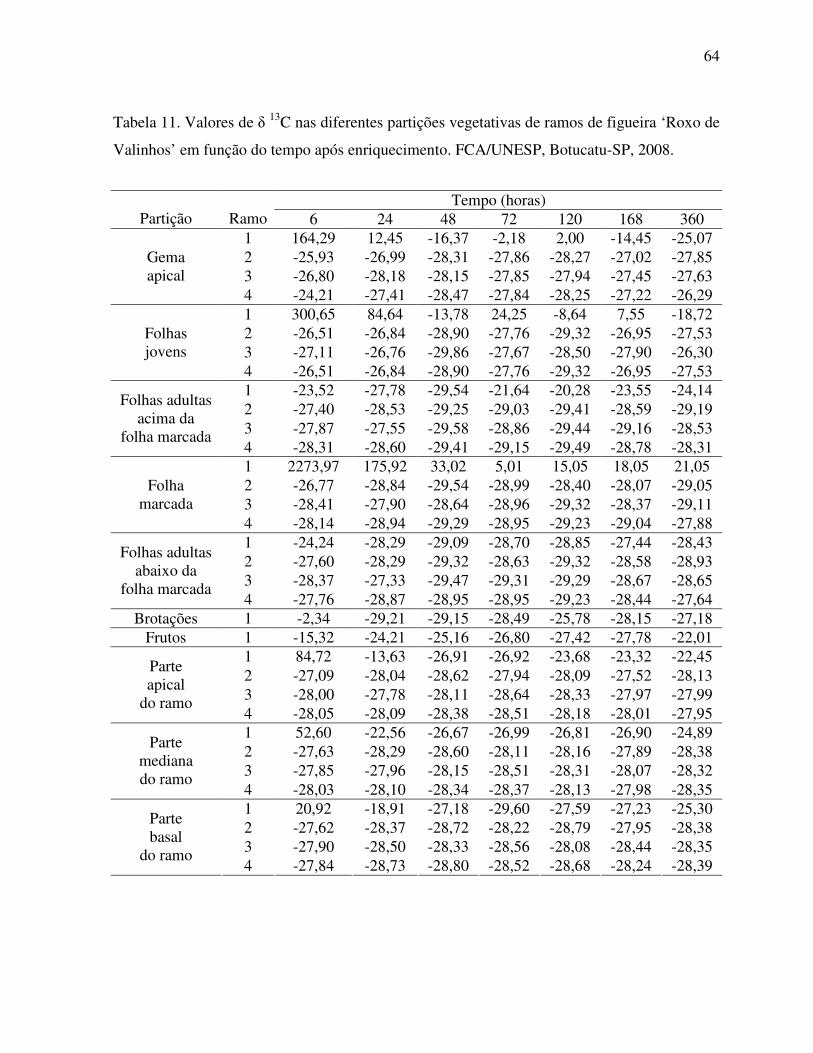
64
Tabela 11. Valores de δ 13C nas diferentes partições vegetativas de ramos de figueira ‘Roxo de
Valinhos’ em função do tempo após enriquecimento. FCA/UNESP, Botucatu-SP, 2008.
Tempo (horas)
Partição Ramo 6 24 48 72 120 168 360 1 164,29 12,45 -16,37 -2,18 2,00 -14,45 -25,07 2 -25,93 -26,99 -28,31 -27,86 -28,27 -27,02 -27,85 3 -26,80 -28,18 -28,15 -27,85 -27,94 -27,45 -27,63
Gema apical
4 -24,21 -27,41 -28,47 -27,84 -28,25 -27,22 -26,29 1 300,65 84,64 -13,78 24,25 -8,64 7,55 -18,72 2 -26,51 -26,84 -28,90 -27,76 -29,32 -26,95 -27,53 3 -27,11 -26,76 -29,86 -27,67 -28,50 -27,90 -26,30
Folhas jovens
4 -26,51 -26,84 -28,90 -27,76 -29,32 -26,95 -27,53 1 -23,52 -27,78 -29,54 -21,64 -20,28 -23,55 -24,14 2 -27,40 -28,53 -29,25 -29,03 -29,41 -28,59 -29,19 3 -27,87 -27,55 -29,58 -28,86 -29,44 -29,16 -28,53
Folhas adultas acima da
folha marcada 4 -28,31 -28,60 -29,41 -29,15 -29,49 -28,78 -28,31 1 2273,97 175,92 33,02 5,01 15,05 18,05 21,05 2 -26,77 -28,84 -29,54 -28,99 -28,40 -28,07 -29,05 3 -28,41 -27,90 -28,64 -28,96 -29,32 -28,37 -29,11
Folha marcada
4 -28,14 -28,94 -29,29 -28,95 -29,23 -29,04 -27,88 1 -24,24 -28,29 -29,09 -28,70 -28,85 -27,44 -28,43 2 -27,60 -28,29 -29,32 -28,63 -29,32 -28,58 -28,93 3 -28,37 -27,33 -29,47 -29,31 -29,29 -28,67 -28,65
Folhas adultas abaixo da
folha marcada 4 -27,76 -28,87 -28,95 -28,95 -29,23 -28,44 -27,64
Brotações 1 -2,34 -29,21 -29,15 -28,49 -25,78 -28,15 -27,18 Frutos 1 -15,32 -24,21 -25,16 -26,80 -27,42 -27,78 -22,01
1 84,72 -13,63 -26,91 -26,92 -23,68 -23,32 -22,45 2 -27,09 -28,04 -28,62 -27,94 -28,09 -27,52 -28,13 3 -28,00 -27,78 -28,11 -28,64 -28,33 -27,97 -27,99
Parte apical
do ramo 4 -28,05 -28,09 -28,38 -28,51 -28,18 -28,01 -27,95 1 52,60 -22,56 -26,67 -26,99 -26,81 -26,90 -24,89 2 -27,63 -28,29 -28,60 -28,11 -28,16 -27,89 -28,38 3 -27,85 -27,96 -28,15 -28,51 -28,31 -28,07 -28,32
Parte mediana do ramo
4 -28,03 -28,10 -28,34 -28,37 -28,13 -27,98 -28,35 1 20,92 -18,91 -27,18 -29,60 -27,59 -27,23 -25,30 2 -27,62 -28,37 -28,72 -28,22 -28,79 -27,95 -28,38 3 -27,90 -28,50 -28,33 -28,56 -28,08 -28,44 -28,35
Parte basal
do ramo 4 -27,84 -28,73 -28,80 -28,52 -28,68 -28,24 -28,39

65
Os valores isotópicos encontrados nas partições, resíduos de poda,
caule e sistema radicular da figueira ‘Roxo de valinhos’, foram os menores evidenciados em
toda a planta para os tempos testados (Tabela 12).
Nenhuma partição apresentou valores positivos, entretanto houve sinal
de enriquecimento no resíduo de poda às 6 horas, dado pela translocação de fotoassimilados
entre os ramos, tendendo à variação natural em 24 horas, não sendo observado novo fluxo de
translocação de fotoassimilados nesta partição.
Os órgãos caule, raiz primária e secundária mantiveram seus valores
muito próximos ao da variação natural, nas primeiras 6 horas, neste mesmo tempo não sendo
observado sinal de enriquecimento nas raízes terciárias, contudo, havendo concentração de 13C
após 48 horas, estando esta partição -4 ‰ acima do valor limite da variação natural. As
radicelas foram consideradas drenos constantes ao longo das 360 horas.
Observou-se uma forte demanda de fotoassimilados pelas partes
vegetativas, pois o sinal isotópico das radicelas tornou-se mais negativo nos tempos das 72 às
168 horas (Tabela 12), voltando a concentrar 13C ás 360 horas após a alimentação. Informação
semelhante foi encontrada por Teng et al. (1999), onde de dezembro para a primavera ocorreu
uma diminuição de 13C foi nas raízes, especialmente as menores que 5 mm em comparação
com as partes aéreas. A redução da quantidade de 13C em raízes são provavelmente para os
processos de respiração e/ou para remobilização ascendente como substrato para novo
crescimento das partes aéreas.
Tabela 12. Valores de δ 13C nas diferentes partições lenhosas e no sistema radicular da figueira
‘Roxo de Valinhos’ em função do tempo após enriquecimento. FCA/UNESP, Botucatu-SP,
2008.
Tempo (horas) Partição 6 24 48 72 120 168 360
Resíduo de Poda -23,30 -27,67 -28,67 -28,75 -28,45 -28,02 -27,88 Caule -27,12 -25,27 - -29,03 -28,72 -28,70 -27,33 Raíz Primária -27,39 -26,29 - -28,05 -27,88 -28,02 -27,66 Raíz Secundária -27,95 -27,83 - -28,20 -28,02 -28,23 -27,31 Raíz Terciária -28,20 -27,66 -23,69 -28,27 -28,58 -28,22 -28,01 Radicelas -26,23 -27,80 -24,83 -27,82 -27,33 -27,07 -26,54

66
Como demonstrado acima, as partições vegetativas da figueira
enriquecida com carbono marcado apresentaram valores isotópicos distintos com o decorrer
do tempo (0 a 360 horas após o enriquecimento). A alternânica dos sítios de alocação entre
elas, após determinado intervalo de tempo, gerou resultados de enriquecimento isotópico
relativo, ou delta per mil (δ ‰) versus tempo, com perfil aproximado de uma onde quadrática,
específica para cada tecido (Jones et al., 1979). Esperou-se que a continuidade dos valores
isotópicos obtidos nas partições fossem descritos por uma função exponencial do tempo. Para
Ducatti et al. (2002), a exploração desta função exponencial abre perspectivas de novas
interpretações nestes experimentos, visando o estudo da reciclagem de carbono-13 em tecidos
vegetais.
Todos os fundamentos teóricos do desenvolvimento e aplicabilidade
do modelo exponencial delta per mil (δ ‰) versus tempo, foram apresentados por Ducatti et
al. (2002), permitindo extrair apenas as equações utilizadas nesse estudo, que relacionou a
concentração inicial e final de carbono-13 em uma partição com o tempo, conforme
apresentado na equação 02.
( ) ( )Kt
FINALINICIALFINALe ][t −δ−δ+δ=δ (eq. 02)
em que: δfinal, representa o valor de delta per mil esperado após um tempo t (em horas); δinicial,
indica o valor de delta per mil proveniente da caracterização natural da partição. A constante k
pode ser interpretada como a constante de troca isotópica com unidades de tempo-1.
Assim ocorrendo, à medida que o tempo aumenta, a concentração dos
isótopos no tecido começa a ser substituída exponencialmente, ocorrendo a troca do carbono-
13 (marcado) pelo carbono-12, dado pela discriminação natural das enzimas da planta pelo
carbono-12.

67
Na condição de metade do enriquecimento final (δf) e metade do
enriquecimento inicial (δi), pode-se obter, a partir da equação (2), o parâmetro de meia vida
(T), expressada em unidade de tempo (horas) dado pela equação (3).
( )K
2lnT = (eq. 03)
A qual será utilizada nas interpretações práticas, da diluição isotópica
nos diversos órgãos amostrados. Ducatti et al. (2002), expressa que quanto menor a meia vida
(T), maior será a taxa de renovação, ou o turnover do tecido.
Por outro lado, admitindo que o valor de δf seja maior do que o valor
de δi na equação 2, pode-se expressar o tempo necessário para a troca de 95% dos átomos (F),
como exemplo, expresso pela equação 4.
( )F1lnK
1t −−= (eq.04)
Parâmetro este, empregado nas interpretações do percentual de troca
(F) do carbono nos diversos órgãos estudados da planta em questão (Tabela 14).
De uma forma geral, a constante K fornece uma idéia da “velocidade”
do processo de eliminação ou troca do carbono-13 no tecido da partição. Como cada órgão da
planta apresenta uma demanda e uma saída de fotoassimilados peculiares, essa constante foi
diferente para cada partição estudada, assim como o valor de meia-vida também distinto para
partições diferentes na mesma planta (Figura 12 e Tabela 13).
Os autores Manetta e Benedito-Cecilio (2003), relatam que o método
de marcação isotópica, quando usado para a determinação da velocidade de processos
metabólitos em animais, fornece resultados com respeito à taxa de turnover. Um dos avanços
mais significativos realizados com este método foi a descoberta de que os componentes
macromoleculares das células e dos tecidos sofrem um processo constante de renovação
metabólica. Existe um estado de equilíbrio dinâmico no interior celular, no qual a biossíntese é

68
constante e exatamente contrabalançada por igual velocidade de degradação. Este processo de
troca metabólica, “renovação celular”, é também chamado de “lavagem celular”, ou turnover.
A função exponencial descrita pela equação 02 apresentou um ajuste
excelente para predizer a relação delta per mil de carbono-13 δ13C versus tempo nas diversas
partições vegetativas estudadas. A Figura 12 apresenta os ajustes para a gema apical e frutos
apenas do ramo que continha a folha enriquecida, permitindo inferir que a taxa de renovação
de fotoassimilados com o tempo é maior para a gema apical, comparativamente ao órgão fruto,
ou seja, a velocidade de troca dos fotoassimilados é maior na gema do que no fruto.
A expressão para a meia-vida sugere que partições que apresentam
meia-vida maior, apresentam uma “baixa velocidade” de metabolização ou incorporação de
carbono no tecido em questão, como o caso das folhas novas que apresentaram os maiores
valores de δ 13C em todo o período estudado (0 a 360 horas). As folhas enriquecidas (folha 4),
não atingiram valores negativos dentro das 360 horas estudadas, contudo pela grande
quantidade de carbono-13 assimilado pela folha e por ser a folha com maior atividade
fotossintética da figueira, apresentaram os menores tempo de meia-vida. Os drenos não se
diferenciaram quanto à taxa de turnover do carbono-13 (Tabela 13).
Por conseguinte, pode ser estabelecida a seguinte sequência de
metabolização do carbono-13 nas partições estudadas: Folhas novas > Frutos > Brotações >
Folhas Adultas > Gema Apical > Partes lenhosas do ramo > Folha marcada.

69
0 50 100 150 200 250 300 350 400
-25
0
25
50
75
100
125
150
175
200
δ 13C = -22,84 + 323,0848 e-0,09135 t
r2 = 0,99944T = 7,6h
(a)
δ 13
Car
bono
Tempo (horas)
0 50 100 150 200 250 300 350 400
-28
-26
-24
-22
-20
-18
-16
-14
δ 13C = -27,10 + 18,76447 e-0,07765 t
r2 = 0,99926T = 8,9h
(b)
δ 13
Car
bono
Tempo (horas)
Figura 12. Modelo exponencial para os isótopos estáveis do carbono em gema apical (a) e
frutos (b) de Ficus carica L., considerando apenas o ramo da folha marcada. FCA / UNESP,
Botucatu – SP, 2008.
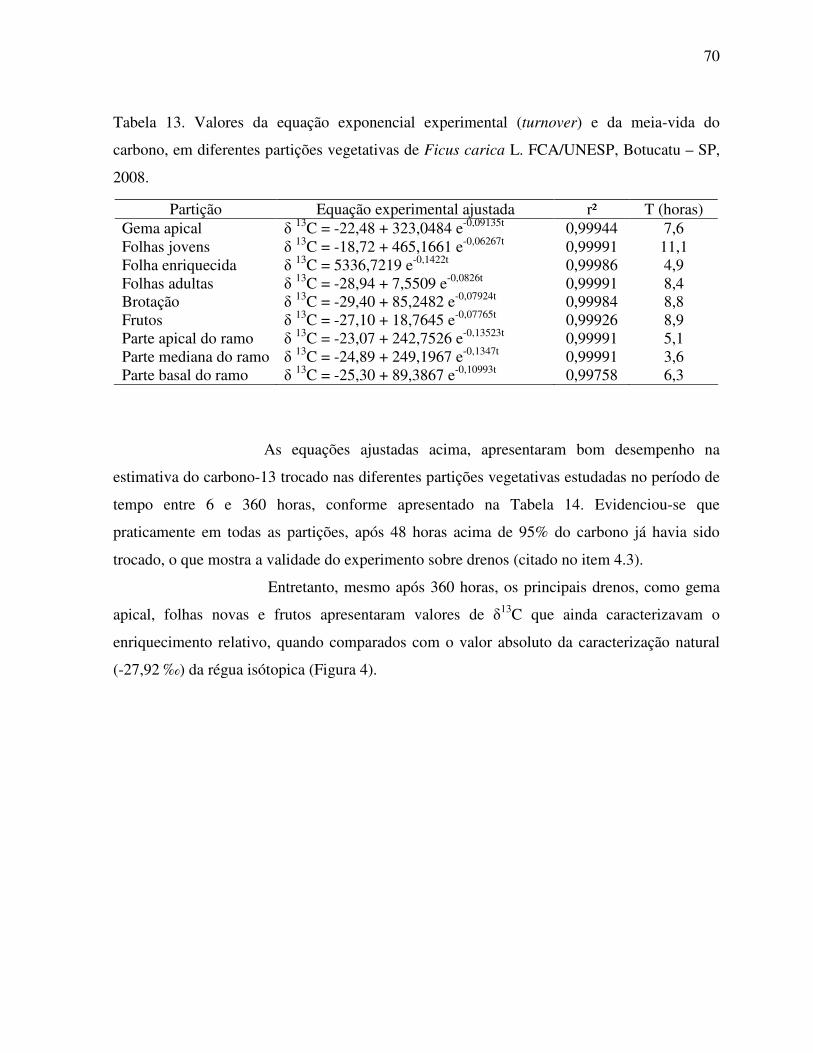
70
Tabela 13. Valores da equação exponencial experimental (turnover) e da meia-vida do
carbono, em diferentes partições vegetativas de Ficus carica L. FCA/UNESP, Botucatu – SP,
2008.
Partição Equação experimental ajustada r² T (horas) Gema apical δ 13C = -22,48 + 323,0484 e-0,09135t 0,99944 7,6 Folhas jovens δ 13C = -18,72 + 465,1661 e-0,06267t 0,99991 11,1 Folha enriquecida δ 13C = 5336,7219 e-0,1422t 0,99986 4,9 Folhas adultas δ 13C = -28,94 + 7,5509 e-0,0826t 0,99991 8,4 Brotação δ 13C = -29,40 + 85,2482 e-0,07924t 0,99984 8,8 Frutos δ 13C = -27,10 + 18,7645 e-0,07765t 0,99926 8,9 Parte apical do ramo δ 13C = -23,07 + 242,7526 e-0,13523t 0,99991 5,1 Parte mediana do ramo δ 13C = -24,89 + 249,1967 e-0,1347t 0,99991 3,6 Parte basal do ramo δ 13C = -25,30 + 89,3867 e-0,10993t 0,99758 6,3
As equações ajustadas acima, apresentaram bom desempenho na
estimativa do carbono-13 trocado nas diferentes partições vegetativas estudadas no período de
tempo entre 6 e 360 horas, conforme apresentado na Tabela 14. Evidenciou-se que
praticamente em todas as partições, após 48 horas acima de 95% do carbono já havia sido
trocado, o que mostra a validade do experimento sobre drenos (citado no item 4.3).
Entretanto, mesmo após 360 horas, os principais drenos, como gema
apical, folhas novas e frutos apresentaram valores de δ13C que ainda caracterizavam o
enriquecimento relativo, quando comparados com o valor absoluto da caracterização natural
(-27,92 ‰) da régua isótopica (Figura 4).

71
Tabela 14. Valores estimados e mensurados de δ‰ 13C e a porcentagem de carbono-13
trocado, para as diferentes partições vegetativas de Ficus carica L. FCA/UNESP, Botucatu –
SP, 2008.
Gema apical Folhas jovens
Tempo δ
13C estimado
δ 13C
mensurado % de C trocado
δ 13C
estimado δ
13C mensurado
% de C trocado
6 163,92 164,29 42,2 300,60 300,65 31,4 T 138,52 - 50,1 213,21 - 50,1 24 13,23 12,45 88,8 84,58 84,64 77,8 48 -18,81 -16,37 98,8 4,22 -13,78 95,1 72 -22,39 -2,18 99,9 -13,63 24,25 98,9
120 -22,83 2,00 100 -18,47 -8,64 99,9 168 -22,84 -14,45 100 -18,71 7,55 100,00 360 -22,84 -25,07 100 -18,72 -18,72 100,00
Folha enriquecida Folhas adultas
Tempo δ
13C estimado
δ 13C
mensurado % de C trocado
δ 13C
estimado δ
13C mensurado
% de C trocado
6 2273,71 2273,97 57,4 -24,34 -24,53 39,1 T 2658,68 - 50,2 -25,17 - 50,0 24 175,84 175,92 96,7 -27,90 -27,13 86,2 48 5,79 33,02 99,9 -28,80 -29,36 98,1 72 0,19 5,01 100,00 -28,92 -29,17 99,7
120 0,00 15,05 100,00 -28,94 -29,17 100,00 168 0,00 18,05 100,00 -28,94 -29,32 100,00 360 0,00 21,05 100,00 -28,94 -28,96 100,00
Frutos Brotações
Tempo δ
13C estimado
δ 13C
mensurado % de C trocado
δ 13C
estimado δ
13C mensurado
% de C trocado
6 -15,32 -15,32 37,2 23,59 23,59 37,8 T -17,70 - 49,9 13,05 - 50,2 24 -24,19 -24,21 84,5 -16,67 -29,5 85,1 48 -26,65 -27,54 97,6 -27,50 -27,5 97,8 72 -27,03 -26,8 99,6 -29,12 -29,59 99,7
120 -27,10 -26,95 99,9 -29,39 -18,76 99,9 168 -27,10 -27,78 100,00 -29,40 -29,4 100,00 360 -27,10 -27,1 100,00 -29,40 -28,83 100,00
(T) tempo de meia-vida da respectiva partição vegetativa.

72
5. CONSIDERAÇÕES FINAIS
O uso da metodologia do carbono-13 como traçador nos estudos da
partição de fotoassimilados em plantas lenhosas apresentou resultados satisfatórios. Todavia,
como essa metodologia visa saber o comportamento ecofisiológico da planta no campo,
apresenta problemas de transpiração da folha dentro da câmara de enriquecimento, sendo
necessário, portanto, estudos posteriores que indiquem quanto tempo de exposição uma folha-
fonte resiste dentro da câmara sem prejudicar suas atividades fisiológicas e possa ser
considerada enriquecida, translocando fotoassimilados às demais partições da planta.
Quanto aos parâmetros avaliados, a continuidade de trabalhos
relacionadas ao tema, poderá mostrar a influência de práticas culturais específicas da cultura,
como as podas na figueira, no desenvolvimento dos ramos e dos órgãos reprodutivos,
indicando o direcionamento e a partição dos fotoassimilados produzidos remanescentes em um
determinado ramo podado ou que tenha sofrido desfolha. Além de evidenciar, quanto tempo
antes dessa prática, a planta começa a remobilizar carbono para os novos processos de
brotação e reconstrução do dossel, associados diretamente com a contribuição climática
durante esse processo.
Trabalhos envolvendo 13C devem ser bem objetivos, pois o uso de
apenas uma câmara de enriquecimento impede a repetibilidade instantânea na aplicação do 13C, que permitiria marcações nas mesmas condições climáticas. Uma alternativa seria a

73
utilização do carbonato de bário (Ba13CO3) como marcador aplicados em sacos plásticos,
permitindo a utilização de vários ao mesmo tempo, porém, ainda existe necessidade de estudos
quanto à densidade do plástico e o tempo de exposição da folha.
De forma geral, os elevados custos das análises podem tornar uma
dificuldade na viabilização do trabalho, principalmente quanto a repetições e delineamentos
estatísticos.

74
6. CONCLUSÕES
A figueira Roxo de Valinhos, em Botucatu/SP, foi caracterizada como
uma planta do ciclo fotossintético C3, tanto pelos valores de assimilação líquida de CO2
(14, 37 µmol m-2 s-1), quanto pelos valores isotópicos (-27,92 ‰), tendo a quarta folha como a
mais fotossinteticamente ativa e principal exportadora de fotoassimilados.
A planta por ter o seu hábito de crescimento modificado pelas podas
drásticas anuais apresenta crescimento vegetativo e fase reprodutiva concomitantes. Neste
contexto, existe uma preferência na alocação de 13C nas partes meristemáticas, seguidas pelos
órgãos reprodutivos. As folhas recém abertas, com aproximadamente 60 cm² de área foliar,
mostraram ser órgão dreno, não exportando fotoassimilados enriquecidos por elas produzidos,
independentemente da presença ou não de órgãos reprodutivos nos ramos.
A planta apresenta uma renovação do 13C dentro de 24 horas, existindo
novos ciclos de retranslocação entre 72 e 168 horas após o enriquecimento para a maioria dos
órgãos, onde estes apresentaram um tempo de meia-vida de duração do carbono-13 inferior a
11 horas.

75
7. REFERÊNCIAS ABRAHÃO, E.; CHALFUN, N. N. J.; REGINA, M. A.; ALVARENGA, A. A. Influência de diferentes tipos de poda no desenvolvimento e produção da figueira (Fícus carica L.) Roxo de Valinhos. Revista Brasileira de Fruticultura, Cruz das Almas, v. 12, n. 2, p. 27-31, 1990. ANTUNES, L. E. C.; ABRAHÃO, E.; SILVA, V. J. Caracterização da cultura da figueira no Estado de Minas Gerais. Informe Agropecuário, Belo Horizonte, v. 18, n. 188, p. 43-44, 1997. BARRIE, A.; PROSSER, S. J. Automated analysis of light-element stable isotopes by isotope ratio mass spectrometry. In: BOUTTON, T.; YAMASAKI, S. Mass spectrometry of soils. New York: Marcel Dekker, 1996. p. 01-46. BEL, V. A. J. E. Mechanism of sugar transfer. In: BAKER, N. R.; THOMAS, H. (Ed.) Crop Photosynthesis. Amsterdan: Elsevier Sciense, 1992, p. 177-211. BERNARDES, M. S. Fotossíntese no dossel das plantas cultivadas. In: CASTRO, P. R. C., FERREIRA, S. O.; YAMADA, T. (Eds). Ecofisiologia da produção agrícola. Piracicaba: Associação brasileira para pesquisa da potassa e do fosfato, 1987. p. 13-45. BERNARDES-SILVA, A. P. F.; LAJOLO, F. M.; CORDENUNZI, B. R. Evolução dos teores de amido e açúcares solúveis durante o desenvolvimento e amadurecimento de diferentes cultivares de manga. Cienc. Tecnol. Aliment., Campinas, v. 23 (supl), p. 16-20, 2003. BERRY, J.; BJÖRKMAN, O. Photosynthetic response and adaptation to temperature in higher plants. Annual Review of Plant Physiology, v. 31, p. 491-543, 1980. BIELESKI R. I. Sugars alcohols. In: LOEWUS F. A.; TANNER W. (Eds). Plant Carbohydrates, Intracellular carbohydrates. Encyclopedia of Plant Physiology, Berlim:

76
Springer-Verlag, New series, 13 A, 1982, p. 158-192. BODSON, M.; OUTLAW JUNIOR, W. Elevation in sucrose content of the shoot apical meristema of sinapis albaat floral evocation. Plant Physiology, Maryland, v. 79, n. 2, p. 20-24, 1985. BOLIANI, A. C.; CORRÊA, L. S. O cultivo da figueira no Brasil e no Mundo. In: CORRÊA, L. S.; BOLIANI, A. C. (ed.) Cultura da figueira - do plantio à comercialização. Ilha Solteira: FAPESP, 1999. p. 37- 40. BUCKHOUT, T. J.; TUBBE, A. Structure, mechanisms of catalysis, and regulation of sugar transporters in plants. In: SAMSKI, E.; SCHAFFER, A. A. Photoassimilate distribution in plants and crops: source-sink relationships. New York: Marcel Dekker, Inc, 1996. p. 229-260. BUAINAIN, A. M.; BATALHA, M. O. Cadeia Produtiva de Frutas. Série Agronegócios, Brasília: IICA/MAPA/SPA, v.7, 2007. 102 p. CAETANO, L. C. S.; CARVALHO, A. J. C.; CAMPOSTRINE, E.; SOUSA, E. F.; MURAKAMI, K. R. N.; CEREJA, B. S. Efeito do número de ramos produtivos sobre o desenvolvimento da área foliar e produtividade da figueira. Revista Brasileira de Fruticultura, Jaboticabal, v. 27, n. 3, p. 426-429, 2005.
CALBO, M.E.R.; MORAES, J.A.P.V. de. Fotossíntese, condutância estomática, respiração e ajustamento osmótico de plantas de buriti submetidas a estresse hídrico. Revista Brasileira de Fisiologia Vegetal, v. 9, n. 2, p. 117-123, 1997. CAMPO DALL’ORTO, F. A.; CANTARELA, H.; RAIJ, B. V.; PIZA JÚNIOR, C. T. Frutas de clima temperado: II. Figo, maça, marmelo, pêra e pêssego em pomar compacto. In: RAIJ, B. V. et al. (Eds). Recomendações de adubação e calagem para o Estado de São Paulo. 2 ed. Campinas: IAC, 1996. p. 139-140. CAMPOSTRINE, E.; YAMANISHI, O. K.; Influence of mechanical root restriction gas-exchange of four papaya genotypes. Revista Brasileira de Fisiologia Vegetal, v. 13, n. 2, p. 129-138, 2001. COLL, J. B.; RODRIGO, G. N.; GARCÍA, B. S.; TAMÉS, R. S. Transporte por el floema. In: Fisiología vegetal. Madrid, Ediciones Pirámide, 2001, p. 103-120. CUNHA, A. R.; KLOSOWSKI, E. S.; GALVANI, E.; ESCOBEDO, J. F.; MARTINS, D. Classificação climática para o município de Botucatu, SP, segundo Köppen. In: SIMPÓSIO EM ENERGIA NA AGRICULTURA, 1. 1999. Botucatu – SP: Anais... Botucatu: FCA/UNESP, 1999, v.1, p. 487-490.

77
CURI, P. R. Relações entre evaporação média pelo tanque IA-58 e evapotranspiração calculada pelas equações de Thornthwaite e Camargo, para o município de Botucatu. 1972. 88 f. Tese (Doutorado) – Faculdade de Ciências Médicas e Biológicas de Botucatu, Universidade Estadual Paulista, Botucatu, 1972. DAÍ, Z.; EDWARD, G. E.; KU, M. S. B. Control of photosyntesis and stomatal conductance in Ricinus communis L. (Castor bean) by leaf to air vapor pressure deficit. Plant Physiology, v. 99, p. 1426-1434, 1992. DANTAS, B. F.; RIBEIRO, L.S .; PEREIRA, M. S. Teor de açúcares solúveis e insolúveis em folha de videiras, cv. Syrah, em diferentes posições no ramo e épocas do ano. Revista Brasileira de Fruticultura, Jaboticabal, v. 29, n. 1, p. 42-47, 2007. DELÉENS, E., CLIQUET, J. B. & PRIOL, J. L. Use of 13C and 15N plant label near natural abundance for monitoring carbon and nitrogen partitioning. Australian, Journal Plant physiology, v. 21, p. 133-146, 1994. DOMINGUEZ, A. F.; La Higuera: frutal mediterrâneo para climas cálidos. Madri: Editora Multi-Prensa, 1990. 190p. DUCATTI, C.; CARRIJO, A. S.; PEZZATO, A. C.; MANCERA, P. F. A. Modelo teórico e experimental da reciclagem do carbono-13 em tecidos de mamíferos e aves. Scientia Agricola, Piracicaba, v. 59, n. 1, p. 29-33, jan./mar. 2002. DUCATTI, C. Isótopos Estáveis ambientais. Botucatu, Instituto de Biociências, 2007, 201p., apostila. ECKARDT, N. A. The Function of SUT2/SUC3 Sucrose Transporters: The Debate Continues. The Plant Cell, vol. 15, n. 6, p. 1259-1262, 2003. EHLERINGER, J. R.; HALL, A. E.; FARQUHAR, G. D. Stable isotopes and plant carbon-water relations. New York: Academic Press, 1993. 555 p. EMBRAPA. Centro Nacional de Pesquisas de Solos. Sistema Brasileiro de Classificação de Solos. Rio de Janeiro, 1999. 412 p. FAO. Food Agricultural Organization. 2003. Disponível em: <http://apps.fao.org>. Acesso em: 2003. FARQUHAR, G. D.; HUBICK, K. T.; CONDON, A. G.; RICHARDS, R. A. Carbon isotope fractionation and plant water-use efficiency. In: RUNDEL, P. W.; EHLERINGER, J. R.; NAGY, K. A. (ed.) Stable isotopes in ecological research. New York: Springer-Verlag New York Inc., 1989. p. 21-40. FLORE, J. A.; LAYNE, D. R. Photoassimilate production and distribution in cherry. Hort Science, v. 34, n. 6, p. 1015-1019, 1999.

78
FOOD AGRICULTURAL ORGANIZATION – FAO (2004). Statistical – database. Disponível em: <http://www.apps.fao.org>. Acesso em: abr. 2007. FOYER, C. H.; GALTIER, N. Source-sink interaction and communication in leaves. In: ZAMSKI, E.; SCHAFFER, A.A. (ed.) Source-sink relations. New York: Marcel Dekker Inc., 1996. p 311-340. FRANCISCO, V. L. F. S.; BAPTISTELLA, C. S. L.; SILVA, P. R. A cultura do figo em São Paulo. 2005. Disponível em: <http://www.todafruta.com.br>. Acesso em: 10 mai. 2007. GIACOBBO, C. L.; PICOLOTTO, L.; KRÜGER, L. R.; PARISOTTO, E.; TIBOLA, C.; FACHINELLO, J. C. Cultivo da figueira conduzida em quatro diferentes densidades de plantio. Revista Brasileira Agrociência, Pelotas, v. 13, n. 1, p. 43-46, jan-mar, 2007. HAMMOND, J. B.; WHITE, P. J. Sucrose transport in the phloem: integrating root responses to phosphorus starvation. Journal of Experimental Botany, vol. 59, n. 1, p. 93–109, 2008. HARTT, C. E; KORTSCHAK, H. P. Sugar Gradients and Translocation of Sucrose in Detached Blades of Sugarcane. Plant Physiology, vol. 39, n.3, p.470-474, 1964. HERTER, F. G.; WAGNER JÚNIOR, A.; COUTO, M.; CHAVES, A.; MALGARIN, M. B.; VITTI, M. R. Concentrações de carboidratos em gemas florais de pereira cv. Nijisseiki submetidas à flutuação térmica durante o período de dormência. In: CONGRESSO BRASILEIRO DE FRUTICULTURA, 17., 2002, Belém. Anais... Belém: Sociedade Brasileira de Fruticultura, 2002. CD. HERTER, F. G.; VERÍSSIMO, V.; CAMELATTO, D., GARDIN, J. P.; TREVISAN, R. Abortamento de gemas florais de pereira no Brasil. In: SEMINÁRIO DE FRUTICULTURA DE CLIMA TEMPERADO NO BRASIL, 1., 2001, Florianópolis. Anais... p.106-114. HOPKINS, W.G. Introduction to Plant Physiology. Jonh Wiley & Sons, New York, 1995, 464p. INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Produção Agrícola Municipal. Rio de Janeiro, 2004. Disponível em <http://www.sidra.ibge.gov.br>. Acesso em: abr. 2007. IBGE. Anuário Estatístico do Brasil/Fundação Instituto Brasileiro de Geografia e Estatística. Rio de Janeiro: IBGE, 1996. seção 3. p.33. IBRAF – Instituto Brasileiro de Frutas. Comparativos das exportações de Frutas Frescas, 2004/05. Arquivo estatístico IBRAF. Disponível em <http://www.ibraf.org.br/ex-es>. Acesso em: abr. 2007.

79
JACKSON, J. E. Light interception and utilization by orchard systems. Horticultural Reviews, New York, v.2, p.208-267, 1980. JACKSON, J. E. Biology of apples and pears. Cambridge, UK: University Press, 2003. JONES, R. J.; LUDLOW, M. M.; TROUGHTON, J. H.; BLUNT, C. G. Estimation of the proportion of C3 and C4 plant species in the diet of animals from the ratio of natural 12C and 13C isotopes in the faeces. Journal of Agricultural Science, v.92, p.91-100, 1979. KADIR, S. Why fruit trees fail to bear. Kansas: Kansas State University, 2003. 4p. (Horticultural Report). KOLOWSKI, T. T.; PALLARDY, S. G. Photosyntesis. In: Physiology in wood plants. Academic Press, New York, 1997, 411p. KRAMER, P. J.; KOZLOWSKI, T. T. Physiology of woody plants. Academic Press. New York, San Francisco, London. 1979, p. 258-281. LAI, R.; WOOLLEY, D. J.; LAWES, G. S. Patterns of assimilate transport from leaves to fruit within a kiwifruit (Actinidia deliciosa) lateral. Journal of Horticultural Science, v. 63, n. 4, p. 725-730, 1988. LAPOINTE, L. Fruit development in trillium: dependence on stem carbohydrate reserves. Plant Physiology, v. 117, p. 183-188, 1998. LARCHER, W. Ecofisiologia Vegetal. São Carlos: Rima, 2000, p.531. LAW, R. D.; CRAFTS-BRANDNER, S. J. Inhibition and acclimation of photosynthesis to heat stress is closely correlated with activation of Ribulose-1,5-bisphosphate carboxylase/oxygenase. Plant Physiology, v. 120, p. 173-182, 1999. LIU, X.; ROBINSON, P. W.; MADORE, M. A.; WITNEY, G. W.; ARPAIA, M. L. Hass avocado carbohydrate flutuations. I. Growth and phenology. Journal of the American Society for Horticultural Science, Alexandria, v. 124, n. 6, p. 671-675, 1999. LOESCHER, W. H.; MCCAMANT, T.; KELLER, J. D. Carbohydrate reserves, translocation, and storage in woody plant roots. HortScience, v. 25, p. 274-281, 1990. LUCCHESI, A. A. Fatores da produção vegetal. In: CASTRO, P.R.C., FERREIRA, S.O., YAMADA, T. (Eds). Ecofisiologia da produção agrícola. Piracicaba: Associação Brasileira para Pesquisa da Potassa e do Fosfato, 1987, p. 1-11. LUDLOW, M. M.; TROUGHTON, J. H.; JONES, R. J. Atechnique for determining the proportion of C3 and C4 species in plant samples using stable natural isotopes of carbon. Journal Agriculture Science, Cambridge, v. 87, p. 625-632, 1976.

80
MACHADO, E. C.; MEDINA, C. L.; GOMES, M. M. A.; HABERMANN, G. Variação sazonal da fotossíntese, condutância estomática e potencial da água na folha de laranjeira ‘Valência’. Scientia Agrícola, Piracicaba, v. 59, n. 1, p. 53-58, 2002. MACHADO, E. C.; LAGÔA, A. M. M. A. Trocas gasosas e condutância estomática em três espécies de gramíneas. Bragantia, Campinas, v. 53, p. 141-149, 1994. MAIORANO, J. A.; ANTUNES, L. E. C.; REGINA, M. de A. et al. Botânica e Caracterização de cultivares da figueira. Informe Agropecuário, Belo Horizonte, v.18, n. 188, p. 22-24, 1997. MANETTA, G. I.; BENEDITO-CECILIO, E. Aplicação da técnica de isótopos estáveis na estimativa da taxa de turnover em estudos ecológicos: uma síntese. Acta Scientiarum: Biological Sciences, Maringá, v. 25, n. 1, p. 121-129, 2003. MARINI, R.Tree management for improving peach fruit quality. In: Mid atlantic fruit vegetable convention, 2002. Disponível em: <http://www.rce.rutgers.edu/peach/orchard/treemanegement>. Acesso em: 15 abr. 2007. MATSUURA, K, TANABE, K., TAMURA, F., ITAI, A. Storage and translocation of 13C-photosynthates from ‘Masui Daufine’ fig (Ficus carica L.) leaves administrated 13CO2 in autumn. Journal of Japanese Society for Horticultural Science, Tokyo, v. 70, n. 1, p. 66-71, 2001. MINCHIN, P. E. H.; THORPE, M. R.; WUNSCHE, J. N.; PALMER;J. W.; PICTON R. F. Carbon partitioning between apple fruits: short- and long-term response to availability of photosynthate. Journal of Experimental Botany, London, v. 48, n. 7, p. 1.401-1.406, 1997. MINISTÉRIO DA AGRICULTURA, PECUÁIA E ABASTECIMENTO. Estatísticas. Disponível em: < http://www.agricultura.gov.br>. Acesso em: 10 mai. 2008. MOING, A.; CARBONNE, F.; RASHAD, M. H.; GAUDILLÈRE, J. P. Carbon fluxes in mature peach leaves. Plant Physiology, Lancaster, v. 100, p. 1878-1884, 1992. MOWAT, A.D.; GEORGE, A.P. Persimmon. In: Schaffer, B.; Andersen, P.C. Handbook of environmental physiology of fruit crops: temperate crops. Boca Raton: CRC Press, 1994. v. 1, p.209-232. NACHREINER, M. L.; SANTOS, R. R. P. Janelas de Mercado – Oportunidades e entraves no comércio internacional de frutas. Revista Hortifruti Brasil, CEPEA – USP/ESALQ, v. 1, n 4, p. 10-13, agosto 2002. NOGUEIRA, R. J. M. C.; MELO FILHO, P. A.; ARAÚJO, E. L. Expressões ecofisiológicas de germoplasma de Hancornia speciosa Gomes cultivado no litoral de Pernambuco. Ciência Rural, v. 29, n. 4, p. 731-732, 1999.

81
O’LEARY, M. H. Biochemical basis of carbon isotope fractionation. In: EHLERINGER, J. R.; HALL, A. E.; FARQUHAR, G. D. (ed.) Stable isotopes and plant carbon-water relations. Sand Diego: Academic Press Inc., 1993. p. 19-28. OLIVEIRA, C. M.; PRIESTLEY, C. A. Carboydrates reserves in deciduous fruit trees. Horticultural Review, v. 10, p. 403-430, 1988. PENTEADO, S. R. O cultivo da figueira no Brasil e no Mundo. In: CORRÊA, L. S. de; BOLIANI, A. C. (Eds.). Cultura da figueira - do plantio à comercialização. Ilha Solteira: FAPESP, 1999. p.1-16. PENTEADO, S. R.; FRANCO, J. A. M. Figo (Fícus carica L.). Manual técnico das culturas. Campinas: SAA/CATI/DCT, 1997. p. 127-139. PEREIRA, F. M.; NACHTIGAL, J. C. O cultivo da figueira no Brasil e no Mundo. In: CORRÊA, L. S. de.; BOLIANI, A. C. (Ed.) Cultura da figueira - do plantio à comercialização. Ilha Solteira: FAPESP, 1999. p. 25-35. PEREIRA, F. M. Cultura da Figueira. Piracicaba: Livroceres, 1981. 73p. PIGÉ, L. G.; SALAGER, J.L.; MCKEY, M. H.; ROY, J. Carbon allocation to volatiles and other reproductive componentsin male Ficus carica (Moraceae). American Journal of Botany, v. 88, n. 12, p. 2214-2220, 2001. PIMENTEL, C. Metabolismo de carbono na agricultura tropical. Seropédica: Edur, 1998. 150p. PIMENTEL GOMES, F. Curso de estatística experimental. 13 ed. Piracicaba: Nobel, 1990. 468 p. PRESTON, K. A. The effects of developmental stage and source leaf position on integration and sectorial patterns en carbohydrate movement in annual plant, Perilla frutescens (Lamiaceae). American Journal of Botany, v. 85, p. 1695-1703, 1998. PRIESTLEY, C. A. Carbohydrate resources within the perennial plant. Commonwealth Agricultural Bureaux, Farnham Royal, v. 62, n. 1, p. 117-118, 1963. QUINLAN, J. D.; PRESTON, A. P. The influence of shoot competition on fruit retention and cropping of apple trees. Journal Horticulture Science, v. 46, p. 525-534, 1971. QUINLAN, J. D.; WEAVER, R. J. Modification of pattern of the photosynthate movement within and between shoots of Vitis vinifera. Plant Physiology, v. 46, p. 527-530, 1970. RAJAPAKSE, N. C.; YOUNG, R. E.; MCMAHON, M. J.; OI, R. Plant height control by photoselective filters: current status and future prospects. HortTechnology, Alexandria, v. 9, n. 4, p. 618-624, 1999.

82
RAKNGAN, J. Carbohidrate analysis of Japanese pear trees grown inder adverse conditions. In: RAKNGAN, J. (ed.) Phenological and physiological study of Japanese pear grown under adverse condition. Tsukuba, 1995, p. 61-63. RENA, A. B.; CARVALHO, C. H. S. Causas abióticas da seca de ramos e morte de raízes em café. In: ZAMBOLIN, L. (Org.). Produção integrada de café. Viçosa: UFV, 2003. p. 197-222. RIGITANO, O. A figueira cultivada no Estado de São Paulo. Piracicaba, 1955. 59 p. Tese (Doutorado na Escola Superior de agricultura “Luiz de Queiroz”, Universidade de São Paulo. Piracicaba.1955. 59p. SCHIMEL, D. S. Terrestrial ecosystems and the carbon cycle. Global Change Biology, v. 1, p. 77-91, 1995. SCHIMEL, D. S. Theory and application of tracers. San Diego: Academic Press Inc., 1993. 113p. SECRETARIA DE COMÉRCIO EXTERIOR – SECEX. Comércio exterior, 2004. Disponível em <http://www.mdic.gov.br>. Acesso em: abr. 2007. SILVA, A. C.; LEONEL, S.; KLEIN, J.; VASCONCELLOS, M. A. S.; DOMINGOS, J. R. Caracterização das trocas gasosas da figueira (Ficus carica l.) em Botucatu–SP. In: LIV Annual Meeting of the Interamerican Society for Tropical Horticulture, XX,. 2008, Vitória. Anais…Vitória: INCAPER, 2008a. 1 CD ROM. SILVA, A. C.; SOUZA, A. P.; LEONEL, S.; DUCATTI, C.; VASCONCELLOS, M. A. S. Caracterização da Variação Isotópica Natural do Carbono-13 em Ficus carica (L.). In: Simpósio Científico dos Pós-graduandos no CENA/USP, 2008, Piracicaba. Ciência: A Questão Água: Resumos… Piracibaca: CENA/USP, 2008b. 1 CD ROM. SIMÃO, S. Tratado de Fruticultura. Piracicaba: FEALQ, 1998. 760p. SIMÃO, S. Manual de Fruticultura. Piracicaba: Ceres, 1971. 534p. SIMKHADA, E. P.; SEKOZAWA, Y.; SUGAYA, S.; GEMMA, H. Translocation and
distribution of 13
C-photosynthates in ‘Fuyu’ persimmon (Diospyros kaki) grafted onto different rootstocks. Journal of Food, Agriculture & Environment. v. 5, n. 1, p.184-189, 2007. TAIZ, L.; ZEIGER, E. Translocação pelo floema. In: Fisiologia vegetal. 3 ed. Porto Alegre: Artmed, 2004, p.221-249.

83
TENG, Y.; TAMURA, F.; TANABE, K. ; NAKAI, T. Partitioning patterns of photosynthates from different shoot types in ‘Nijisseiki’ pear (Pyrus pyrifolia Nakai). Journal of Horticultural Science and Biotechnology, v. 77, p. 758-765, 2002. TENG, Y; TANABE, K.; TAMURA, F.; ITAI, A. Translocation of 13C- assimilates in the spring following fall assimilation of 13CO2 by ‘Nijisseiki’ pear ( Pyrus pyrifolia Nakai). Journal of Japanese Society for Horticultural Science, v. 68, n.2, p.248-255, 1999. THORPE, M. R.; MINCHIN, P. E. H. Mechanisms of long and short-distance transport from sources to sinks. In: ZAMSKI, E.; SCHAFFER, A. A. Photoassimilate distribution in plants and crops: source-sink relationships. New York: Marcel Dekker, Inc, 1996. p.261-282. TOFIÑO, A., FREGENE, M., CEBALLOS. H., CABAL, D. Regulación de la biosíntesis del almidón en plantas terrestres: perspectivas de modificación. Acta Agronómica, Palmira, Colombia, v. 55, n.1, p. 1-16, 2006. TUBELIS, A.; NASCIMENTO, E. J. L.; FOLONI, L.L. Meteorologia e climatologia. Botucatu. Faculdade de Ciências Médicas e Biológicas, 1972. v.3, p.344-362. TUBELIS, A.; SALIBE, A. A. Relações entre produção de laranjeira ‘Hamlin’ e as precipitações mensais no altiplano de Botucatu. Pesquisa agropecuária brasileira, v. 24, p. 801-806, 1989. VALLADARES, F.; PEARCY, R. W. Interactions between water stress, sun-shade acclimation, heat tolerance and photoinhibition in the sclerophyll Heteromeles arbutifolia. Plant, Cell and Environment, v.20, p.25-36, 1997. VALLI, R. Arboricoltura, Generale e Speciale. Bologna: Calderini Edagricole, 2002. 658p. VASCONCELLOS, M. A. S., Uso do Carbono-13 como marcador na partição de fotoassimilados em maracujazeiro doce (Passiflora alata Dryander.). 2001. 106 f. Doutorado (Doutorado em Agronomia/Horticultura)-Faculdade de Ciências Agronômicas, Universidade estadual, Paulista, Botucatu, 2001. XAVIER, A. M.; LIMA, A. G.; VIGNA, C. R. M.; VERBI, F. M.; BORTOLETO, G. G.; GORAIEB, K.; COLLINS, C. H.; BUENO, M. I. M. S. Marcos da história da radioatividade e tendências atuais. Química Nova, v.30, n.1, p.83-91, 2007. WARDLAW, I. F. The control of carbon partitioning in plants. New Phytologist, Lancaster, v. 116, p. 341-381, 1990. WATSON, D. P.; BOWERS, F. A. Long days produce flowers on passion frui. Hawaii Farm Science, v. 14, n. 2, p. 03-05, 1965.

84
WISE, R. R.; OLSON, A. J.; SCHRADER, S. M.; SHARKEY, T. D. Electron transport is the functional limitation of photosynthesis in field-grown Pima cotton plants at high temperature. Plant, Cell and Environment, v. 27, p. 717-724, 2004. YAMAKI, S. Distribution of sorbitol, neutral sugars, free amino acids, malic acid and some hydrolytic enzymes in vacuoles of apple cotyledons. Plant Cell Physiology, v. 23, p. 881-889, 1982. ZAMSKI, E. Anatomical and physiological characteristics of sink cells. In: Zamski, E. & Schaffer, A.A. (ed) Source-sink relations, Marcel Dekker Inc. New York. 1996. p 283-310. ZAMSKI, E. Transport and accumulation of carbohydrates in developing seeds: the seed as a sink. In: Seed developing and germination. KIGEL, J.; GALILI, G. (Eds.). Marcel Dekker, New York, 1995, p.26-44. ZHANG, C.; TANABE, K.; TAMURA, F.; ITAI, A.; WANG, S. Spur characteristics, fruit growth and carbon partitioning in two late-maturing Japanese pear (Pyrus pyrifolia) cultivars with contrasting fruit size. Journal of the American Society for Horticultural Science, v. 130, p. 252-260, 2005a. ZHANG, C.; TANABE, K.; TAMURA, F.; ITAI, A.; WANG, S. Partitioning of 13C-photosyntate from spur leaves during fruit growth of three pear (Pyrus pyrifolia) cultivars differing in maturation date. Annals of Botany, v.95, p. 685-693, 2005b.


















