Variação hormonal correlacionada à expressão de ... · ligadas ao metabolismo do amido durante...
83
UNIVERSIDADE DE SÃO PAULO FACULDADE DE CIÊNCIAS FARMACÊUTICAS Programa de Pós-Graduação em Ciência dos Alimentos Área de Bromatologia Variação hormonal correlacionada à expressão de enzimas ligadas ao metabolismo do amido durante o desenvolvimento e amadurecimento da manga (Mangifera indica cv Keitt) Claudia Mitsue Koike Dissertação para obtenção do grau de MESTRE Orientador: Prof. Dr. Eduardo Purgatto São Paulo 2007
Transcript of Variação hormonal correlacionada à expressão de ... · ligadas ao metabolismo do amido durante...

UNIVERSIDADE DE SÃO PAULO FACULDADE DE CIÊNCIAS FARMACÊUTICAS
Programa de Pós-Graduação em Ciência dos Alimentos Área de Bromatologia
Variação hormonal correlacionada à expressão de enzimas
ligadas ao metabolismo do amido durante o desenvolvimento e
amadurecimento da manga (Mangifera indica cv Keitt)
Claudia Mitsue Koike
Dissertação para obtenção do grau de
MESTRE
Orientador: Prof. Dr. Eduardo Purgatto
São Paulo
2007

UNIVERSIDADE DE SÃO PAULO FACULDADE DE CIÊNCIAS FARMACÊUTICAS
Programa de Pós-Graduação em Ciência do Alimentos Área de Bromatologia
Variação hormonal correlacionada à expressão de enzimas
ligadas ao metabolismo do amido durante o desenvolvimento e
amadurecimento da manga (Mangifera indica cv Keitt)
Claudia Mitsue Koike
Dissertação para obtenção do grau de
MESTRE
Orientador: Prof. Dr. Eduardo Purgatto
São Paulo
2007

Claudia Mitsue Koike
Variação hormonal correlacionada à expressão de enzimas ligadas ao
metabolismo do amido durante o desenvolvimento e amadurecimento da
manga (Mangifera indica cv Keitt)
Comissão Julgadora
da Dissertação para obtenção do grau de Mestre
Prof. Dr. Eduardo Purgatto
orientador/presidente
1° Examinador
2° Examinador
São Paulo, 20 de julho de 2007

“Cada pessoa que passa em nossa vida, passa sozinha, é porque
cada pessoa é única e nenhuma substitui a outra. Cada pessoa
que passa em nossa vida passa sozinha, e não nos deixa só,
porque deixa um pouco de si e leva um pouquinho de nós. Essa é
a mais bela responsabilidade da vida e a prova de que as pessoas
não se encontram por acaso”.
Charles Chaplin

Agradecimentos Agradeço primeiro a Deus por estar presente em minha vida nos momentos
felizes e nas horas mais difíceis, me dando coragem e força para vencer todos os
obstáculos que encontramos na vida.
Agradeço aos meus pais que puderam com muito esforço me dar a
oportunidade de estudar e permitir que pudesse fazer o meu próprio caminho,
sempre me apoiando, e pela paciência e compreensão. Ao meu irmão que sempre
que pôde, me ajudou e sempre esteve também ao meu lado.
A Cleusa, amiga que sempre pude contar, nos bons momentos e nas horas
mais difíceis; me incentivando a seguir o caminho que escolhi e aconselhando
quando precisei.
As minhas amigas Claudinéia, Cíntia, Patrícia e Roberta que estiveram ao
meu lado me dando força em meu trabalho e nos momentos de tristeza, nos breves
momentos de descontração, e por ter compartilhado parte de nossas vidas neste
período.
Agradeço a todos do laboratório: Aline, Aderuza, Amanda, Ana Cristina, Any,
Eliana, Fernanda, Gabriele, Giseli, Grazielli, Janaína, Juliana, João Paulo, Lúcia,
Lucile, Maria Luiza, Márcia, Marcinha, Milana, Neuza, Renato e Tânia.
Ao Edílson, Mônica, Cléo, Joana, Lurdinha e a todos do departamento que
puderam me ajudar a realizar este trabalho.
A professora Beatriz pelo seu profissionalismo, sua ajuda e compreensão
junto ao meu trabalho.
Ao Eduardo pela orientação durante este período, pelo carinho e pela
dedicação.
A CAPES pela bolsa que me proporcionou os meus estudos.
Quero agradecer a todos de coração por fazerem parte de mais uma etapa da
minha vida.
A todos muito obrigada!

Dedico este trabalho a minha mãe Helena, ao meu pai Katsumi e ao meu irmão Marcelo.

SUMÁRIO
1 INTRODUÇÃO 01 1.1. Amadurecimento de frutos 01 1.2. Metabolismo do amido em frutos 06
1.2.1. α- amilases 07 1.2.2. β- amilases 08 1.2.3. Fosforilases 08 1.2.4. Isoamilases 10 1.2.5. Enzima-D (DPE - Disproportionating Enzymes) 11
1.3. Manga 14 1.4. Degradação do amido em mangas Keitt 16
2 OBJETIVO 22
3 MATERIAL E MÉTODOS 23 3.1. Material 23 3.2. Métodos 24
3.2.1. Extração de Ácido Indol-3-Acético (AIA) e Ácido Abscísico (ABA) 24 3.2.2. Derivatização 25 3.2.3. Determinação e análise dos hormônios vegetais por CG-EM-MSI 25 3.2.4. Extração de RNA Total 26 3.2.5. Síntese do cDNA 27 3.2.6. Northern Blot Reverso 27
3.2.6.1. Síntese de sondas de cDNA 27 3.2.6.2. Transferência para membrana 28 3.2.6.3. Preparo das sondas radioativas e hibridização 28 3.2.6.4. Análise densitométrica 29
3.2.7. Medida de Etileno e CO2 29
3.2.8. Tratamento com 1- Metilciclopropano (1-MCP) 30 3.2.9. Açúcares Solúveis 31 3.2.10. Amido 31
3.2.11. Extração e Atividade de α-amilase e de β-amilase 31
3.2.12. Extração de proteínas e Atividade de fosforilase em gel nativo 32

4 RESULTADOS E DISCUSSÃO 33
4.1. Determinação e análise dos hormônios vegetais por CG-EM-MSI 33 4.2. Northern Blot Reverso 39 4.3. Medida de Etileno e CO2 46 4.4. Amido 48
4.5. Açúcares Solúveis 50
4.6. Atividade de α-amilase e de β-amilase 52
5 CONSIDERAÇÕES FINAIS 56
6 CONCLUSÕES 57
7 REFERÊNCIAS BIBLIOGRÁFICAS 58
8 RESUMO 70
9 ABSTRACT 71

ÍNDICE DE FIGURAS
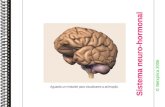
Figura 1. O ciclo de Yang e a biossíntese do etileno (Modificado de Pech et al.,
2003). 02 Figura 2. Esquema da suposta via de degradação de amido em folhas de
Arabidopsis. GWD (glucan-water dikinase), PWD (phosphoglucan-water
dikinase), ISA3 (isoamilase), Pho1 (amido fosforilase plastidial), Pho2
(amido fosforilase citsólica), DPE1 e 2 (enzima-D, disproportionating
enzymes), MEX1 (transportador de maltose), 1- transportador de glicose,
2- transportador de triose fosfato. 13 Figura 3. Perfil da produção de etileno e CO2 (respiração) durante amadurecimento
da manga Keitt. 17 Figura 4. Perfil da síntese da sacarose e da degradação do amido durante o
desenvolvimento e amadurecimento da manga Keitt. 18 Figura 5. Atividade de α-amilase e β-amilase durante o desenvolvimento e
amadurecimento.
19
Figura 6. Diagrama da coleta das amostras da primeira amostragem. À esquerda,
coleta dos frutos verdes e à direita, coleta dos frutos maduros. 24 Figura 7. Perfil dos hormônios AIA ( ) e ABA ( ), quantificados em CG-EM-MSI,
durante o desenvolvimento e amadurecimento da manga Keitt (dpa – dias
pós-antese/ dpc – dias pós-colheita). 34 Figura 8. (A) Análise densitométrica. (B) Northern Blot Reverso da α-amilase.
Normalizado com os valores obtidos da razão entre DO dos transcritos da
α-amilase e DO dos mRNAs do controle actina. (dpa – dias pós-antese/
dpc – dias pós-colheita). 40 Figura 9. (A) Análise densitométrica. (B) Northern Blot Reverso da β-amilase.
Normalizado com os valores obtidos da razão entre DO dos transcritos da
β-amilase e DO dos mRNAs do controle actina. (dpa – dias pós-antese/
dpc – dias pós-colheita). 42 Figura 10. (A) Análise densitométrica. (B) Northern Blot Reverso da isoamilase.
Normalizado com os valores obtidos da razão entre DO dos transcritos da
isoamilase e DO dos mRNAs do controle actina. (dpa – dias pós-antese/
dpc – dias pós-colheita). 44

Figura 11. (A) Análise densitométrica. (B) Northern Blot Reverso da DPE2.
Normalizado com os valores obtidos da razão entre DO dos transcritos da
DPE2 e DO dos mRNAs do controle actina. (dpa – dias pós-antese/ dpc –
dias pós-colheita). 45 Figura 12. Perfil da produção de etileno e respiração (produção de CO2) durante o
amadurecimento de mangas Keitt. (A) Amostras controle ( etileno,
CO2). (B) Tratamento com 1-MCP ( etileno, CO2). 47 Figura 13. Evolução dos teores percentuais de amido em amostras controle (sem
tratamento 1-MCP - ) e tratadas com 1-MCP ( ) durante o
amadurecimento da manga Keitt. 48 Figura 14. Evolução dos teores dos carboidratos em amostras controle (sem
tratamento 1-MCP - frutose, sacarose, glicose) e tratadas com 1-
MCP ( frutose, sacarose, glicose) durante o amadurecimento da
manga Keitt.
50
Figura 15. Atividade de amido fosforilases citosólica e plastidial em gel de
poliacrilamida contendo glicogênio 1% corado com I2/KI em amostras
controle (sem tratamento 1-MCP) e tratadas com 1-MCP de manga Keitt.
53
Figura 16. Evolução dos teores de maltose em amostras controle (sem tratamento 1-
MCP - ) e tratadas com 1-MCP ( maltose) durante o amadurecimento
de mangas Keitt.
56

ÍNDICE DE TABELAS
Tabela 1. Valores da atividade de α-amilase e de β-amilase durante o
amadurecimento de mangas Keitt controle (sem tratamento 1-MCP) e
tratadas com 1-MCP. 52

ABREVIATURAS
ABA Ácido Abscísico
ACC Ácido 1-aminociclopropano-1-carboxílico
ACS Ácido 1-aminociclopropano-1-carboxílico sintase
ACO Ácido 1-aminociclopropano-1-carboxílico oxidase
AIA Ácido Indol-3-acético
(13C6)-AIA Ácido Indol-3-acético marcado com carbono-13
dpa Dias após antese
cDNA DNA complementar
DPA Dias Após Antese
DPC Dias Após Colheita
DPE1 Disproportionating Enzymes 1
DPE2 Disproportionating Enzymes 2 (α-1,4 glicanotransferase)
GA Ácido Giberélico CG-EM-MSI Cromatografia a gás com detecção por espectrometria de
massa – monitoramento seletivo de íons
GWD Glucan-Water Dikinase (glicano-água diquinase)
ISA3 Isoamilase 3
mRNA RNA mensageiro
Pho1 Fosforilase Plastidial
Pho2 Fosforilase Citosólica
PWD Phosphoglucan-Water Dikinase (fosfoglicano-água dikinase)
SAM S-adenosilmetionina
1-MCP 1-metilciclopropeno

1. INTRODUÇÃO
1.1. Amadurecimento de frutos
O amadurecimento do fruto é caracterizado por significativas transformações
no órgão que em última instância levam as alterações na textura, sabor, aroma e cor
do fruto, tornando-o assim, aceitável para o consumo (NASCIMENTO et al., 1998).
Tais alterações são decorrentes de mudanças no padrão de atividade de
enzimas envolvidas nos diversos metabolismos ligados às características acima
citadas. Muitas destas alterações ocorrem, ao menos parcialmente, por mudanças
nos padrões de expressão gênica das enzimas ligadas a diversas vias metabólicas.
A regulação destes processos pode ocorrer em diversos níveis sendo que os
hormônios (particularmente o etileno) desempenham papel chave no processo.
O etileno é um hormônio vegetal que afeta muitos processos do
desenvolvimento, como a germinação de sementes, senescência de flores e folhas,
crescimento e diferenciação de raízes, abscisão de folhas e o amadurecimento dos
frutos (ARORA, 2005; GUO et al., 2003; LIU, et al., 1999; THEOLOGIS, 1992) o que
o levou a ser designado como “hormônio do amadurecimento”.
O etileno é sintetizado a partir de intermediários do ciclo da metionina ou ciclo
de Yang (Yan; Hoffman, 1984 – Figura 1). O passo limitante na sua produção é a
conversão da S-adenosil-metionina no ácido 1-aminociclopropano-1-carboxílico
(ACC), pela ACC sintase (S-adenosil-L-metionina metiltioadenosina liase, EC
4.4.1.14). Em seguida o ACC é convertido pela ACC oxidase (1-aminociclopropano
carboxilato oxidase, E.C. 1.4.3) em etileno, gás carbônico e ácido cianídrico, na
presença de oxigênio.

[O] HPO4-2 HCOO-
CH3 S CH2 CH2 CO COO-
R CH COO-
NH3+
R CO COO-
CH3 S CH2 CH2 CH COO-
NH3+
ATP
ATP
PPi + Pi
CH3 S+
NH3+
COO-CHCH2CH2
CH2
OH OH
AdeO
CH3 S
CH2
OH OH
AdeO
CH3 S
CH2
OH OH
OHO
ADP
CH3 S
CH2
OH OH
OPO3H-
O
Ade
H2CC
H2C
NH3+
COO-
1/2 O2
CO2 + HCN
CH2 CH2
S-Adenosilmetionina(SAM)
5'-metiltioadenosina
5-metiltioribose
5-metiltioribose-1-fosfato
2-oxo-4-metilbutirato
Metionina
ACCSintase
Ácido aminociclopropano carboxílicoACC
Etileno
ACCOxidase
Figura 1: O ciclo de Yang e a biossíntese do etileno (Modificado de Pech et al., 2003).
A influência dos níveis de etileno no amadurecimento dos frutos é muito
variável, mesmo entre cultivares de um mesmo fruto. A sensibilidade a este
hormônio parece aumentar com o avanço do desenvolvimento, culminando com uma
maior sensibilidade durante o amadurecimento. No caso específico da manga,
Tharanathan et al. (2006) observou que níveis abaixo de 0,01µL.h-1 podem
desencadear o amadurecimento em manga.

Embora a classificação dos frutos em climatéricos e não climatéricos tenha
inicialmente levado em consideração o comportamento respiratório (LOHANI et al.,
2004) verificou-se, anos depois, que esse era precedido de um aumento na
produção de etileno durante o amadurecimento, no caso dos frutos climatéricos. Isto
os difere dos não climatéricos que produzem pouco etileno endógeno e apresentam
um declínio na taxa de respiração após a colheita. (THARANATHAN et al., 2006;
PHILLIPS et al., 2004; WHILE, 2002).
Todos os frutos são capazes de produzir etileno, mas apenas nos frutos
climatéricos essa síntese pode ocorrer de maneira autocatalítica, ou seja, o próprio
hormônio induz a expressão das enzimas responsáveis por sua produção. Desta
forma sugere-se a existência de dois sistemas reguladores da biossíntese de etileno
em plantas superiores. O Sistema I, atuando tanto nos frutos climatéricos como nos
não-climatéricos e também nos tecidos vegetais, responsável pela produção de
níveis baixos de etileno. O Sistema II é responsável pelo elevado aumento na
produção de etileno durante o amadurecimento dos frutos climatéricos (LELIÈVRE et
al., 1997).
A maior parte dos estudos sobre os mecanismos de ação do etileno tem sido
feitos na planta modelo Arabidopsis thaliana e em tomate. Os modelos desses
trabalhos têm indicado que a ação do etileno sobre os receptores (identificados
como proteínas quinase) deve ser inibitória, ou seja, os receptores estão ativos na
ausência de etileno e assim inibem as respostas ao hormônio, e são desativados
quando ocorre a ligação do hormônio (ARORA, 2005), liberando a via de sinalização
e promovendo as respostas etileno-dependentes.
Pouco se sabe a respeito dos fatores que coordenam as mudanças para a
sensibilização dos tecidos dos frutos climatéricos para responder ao etileno, apesar

de o etileno desempenhar papel importante no amadurecimento, é provável que os
mecanismos de controle sejam o resultado da interação entre o etileno e outros
hormônios como auxinas, ácido abscísico e giberelinas, ou seja, um balanço
hormonal. Neste quadro, dois hormônios em particular, o ácido indol-3-acético (AIA)
e o ácido abscísico (ABA) vêm recebendo maior destaque.
O ácido indol-3-acético (AIA) atua sobre o desenvolvimento, crescimento e
amadurecimento dos frutos, influenciando nos processos celulares como o
alongamento, divisão e diferenciação celular (THEOLOGIS, 1996; TINGWA;
YOUNG,1975). Evidências mostram que o AIA constitui um fator de resistência ao
amadurecimento, sendo demonstrado seu efeito no atraso deste processo em
bananas e pêras tratadas com auxina na fase pré-climatérica (PURGATTO et al.,
2001; TINGWA; YOUNG, 1975; FRENKEL; DYCK, 1973).
O AIA se apresenta na forma livre ou conjugada com açúcares, aminoácidos
e alguns peptídeos. O AIA conjugado parece ter função no transporte, no acúmulo
do hormônio consistindo a maior parte do AIA total durante o crescimento vegetal
(ÖSTIM et al., 1998), servindo como reserva que pode ser mobilizada conforme a
necessidade do tecido no qual está acumulado.
Estudos recentes sobre o amadurecimento da banana indicaram que além do
etileno, o ácido indol-3-acético também estaria ligado ao amadurecimento desse
fruto (LOHANI et al., 2004). Outras evidências demonstram que vários fatores de
transcrição responsivos a auxinas são ativados durante o amadurecimento do
tomate (WANG et al. 2003), sendo que estes também são modulados pela
sinalização do etileno. Embora os genes-alvo destes fatores de transcrição ainda
não tenham sido identificados, a observação sugere a participação do AIA como
modulador do amadurecimento de frutos.

O ácido abscísico (ABA) influencia o amadurecimento tanto em frutos
climatéricos como nos não-climatéricos, e o seu efeito no amadurecimento parece
estar ligado à evolução do etileno, mas o mecanismo pelo qual o ABA estimula a
síntese de etileno ainda é desconhecido. Um provável mecanismo indica que o ABA
pode afetar a síntese de etileno pelo aumento da síntese de ACC, resultando na
aceleração do amadurecimento (VENDRELL; LARA, 2003).
Segundo Jiang et al. (2000) o ABA parece ser o desencadeador do aumento
da sensibilidade ao etileno, levando em conta que o etileno seja o desencadeador do
amadurecimento. Em bananas foi observado que o tratamento com altas
concentrações de ABA estimulou a síntese de etileno e provocou aumento na taxa
respiratória. Em tratamento com 1-MCP, um potente antagonista do etileno, por
algumas horas e seguida pela adição do ABA, não houve efeito aparente no fruto,
indicando que a influência do ABA sobre o amadurecimento depende da resposta ao
etileno. Ao aplicar o ABA imediatamente antes ou depois do tratamento com 1-MCP
houve o aumento na taxa de respiração e do etileno, indicando que em frutos
climatéricos, o climatério, a respiração e a maciez sejam mais sensíveis ao ABA do
que se supunha. O tratamento com etileno juntamente com o ABA resultou no
aceleramento da produção de etileno, respiração, mudança de cor e na maciez em
relação ao tratamento com etileno apenas. O que se sugere que o ABA possa atuar
também como coordenador do amadurecimento.
Lohani et al. (2004) demonstraram que o tratamento com o hormônio promove
a diminuição da firmeza da banana, em menor escala quando comparado ao
tratamento do ácido abscísico mais etileno, ao contrário do que ocorreu com
tratamento com AIA com e sem etileno, que inibiu a atividade enzimática hidrolítica
sobre os polissacarídeos constituintes da parede celular.

1.2. Metabolismo do amido em frutos
Um dos principais eventos bioquímicos que ocorrem durante o
amadurecimento é a degradação do amido, nos frutos que acumulam esse
carboidrato como reserva. Na manga o amido armazenado é a principal reserva de
carbono, assim como na banana, sendo praticamente todo degradado durante o
amadurecimento do fruto para a síntese de sacarose após a colheita.
O grânulo de amido possui uma estrutura complexa, composta por dois tipos
de polímeros; a amilose, cadeia de glicose formada por ligações α-1,4, tendo poucas
ramificações, e a amilopectina, cadeia de glicose altamente ramificada, formada por
ligações α-1,4 como a amilose e por ligações α-1,6 nas ramificações, constituindo a
maior parte da massa de grânulos de amido de várias espécies vegetais (TETLOW
et al., 2004; ZEEMAN et al., 2004).
A estrutura dos grãos de amido varia de planta para planta e mesmo de tecido
para tecido na mesma planta e, embora muitos estudos tenham se concentrado na
elucidação do processo de formação do grão de amido, a forma como ocorre o
empacotamento das cadeias de amilose e amilopectina para sua formação ainda
não está totalmente esclarecida.
De maneira geral e tomando por base os modelos mais aceitos (GALLANT et
al., 1997; MYERS et al., 2000), as cadeias de amilose e amilopectina se organizam
formando anéis concêntricos, delimitados por diferenças na cristalinidade de duas
regiões distintas: uma região amorfa menos compacta e uma semi-cristalina, na qual
predomina principalmente o arranjo mais compacto das cadeias de amilopectina.
Várias enzimas participam da degradação do grânulo de amido, nas várias
situações onde a mobilização da reserva é necessária.

No amadurecimento de frutos já foram identificadas tanto hidrolases (que
incluem as α e β-amilases e enzimas desramificadoras) como fosforilases,
resultando em reações complexas que produzem metabólitos que podem ser
transportados dos plastídeos para o citosol (ZEEMAN et al., 2004) e servirem tanto
como substratos energéticos como para a síntese da sacarose, promovendo, desta
forma, o adoçamento do fruto.
1.2.1. α- amilases
As α-amilases são endoamilases responsáveis pela hidrólise das ligações α-
1,4 da amilose e amilopectina, mas são capazes, embora com baixa taxa de
eficiência, de hidrolisar as ligações α-1,6, produzindo malto-oligossacarídeos
(VIEIRA JR et al., 2006).
Dentre os exemplos encontrados na literatura, a atividade de α-amilases em
frutos foi descrita em bananas (VIEIRA JR et al., 2006; GARCIA; LAJOLO, 1988;
MAO; KINSELLA, 1981), kiwi (WEGRZYN; MAcRAE, 1995) e mangas
(BERNARDES-SILVA et al., 2007; TANDON et al., 1983; FUCHS, et al., 1980), entre
outros.
Em bananas, Vieira Jr et al. (2006) observaram que a atividade e a expressão
gênica de uma α-amilase aumentavam durante o amadurecimento precedendo a
fase de mobilização do amido.
Wegrzyn et al. (2000) demonstraram para maçãs a presença de um gene que
codifica para uma proteína muito semelhante às α-amilases e que tem a expressão
gênica aumentada em resposta a baixas temperaturas. Essa isoforma apresentou
diferenças significativas na estrutura de dois domínios da proteína quando
comparada a outras formas já conhecidas e descritas, e também pela ausência de

um peptídeo sinal, comumente encontrado em outras α-amilases, conforme pode ser
deduzido de sua seqüência de nucleotídeos.
1.2.2. β- amilases
As β-amilases hidrolisam as ligações α-1,4-glicosídicas, removendo
sucessivamente unidades de maltose das extremidades não-redutoras das cadeias
glicosídicas que compõem os oligossacarídeos provenientes da degradação do
grânulo de amido. As β-amilases têm sido descritas como proteínas abundantes em
plantas, sementes e tubérculos (BERNARDES-SILVA, 2004; PURGATTO et al.,
2001; MITA et al., 1995).
Em bananas, foi observada uma correlação entre o aumento de atividade de
β-amilase e o início da degradação do amido (NASCIMENTO et al. 2006), e essa
enzima correspondeu a um dos mRNAs com abundância aumentada durante o
amadurecimento, conforme análise diferencial de uma biblioteca de cDNA de
banana madura (MEDINA-SUAREZ et al., 1997). De acordo com Purgatto et al.
(2001), o aumento de transcrição durante o amadurecimento, associado com a
quebra do amido, pode ser afetado negativamente pela infiltração de ácido indol-3-
acético em fatias de banana. Uma vez que o início da degradação do amido também
foi atrasado, foi proposto que a β-amilase poderia ter uma importante contribuição
para esse processo.
1.2.3. Fosforilases
As amido fosforilases (α-1,4-glicano fosforilases) são enzimas que catalisam a
quebra reversível das ligações α-1,4 dos glicanos derivados da quebra do amido a
partir da extremidade não redutora das cadeias de amilose e amilopectina

produzindo glicose 1-fosfato. Contudo estas enzimas também são capazes de
alongar cadeias de oligossacarídeos e polissacarídeos quando na presença de
glicose 1-fosfato e de um oligossacarídeo iniciador.
Em plantas superiores as amido fosforilases podem ser encontradas em pelo
menos duas isoformas sendo uma plastidial (amiloplastos e cloroplastos) e a outra
citosólica (SCHUPP et al., 2004).
A amido fosforilase plastidial (Pho1) parece participar nos estágios finais da
degradação, tendo ação sobre os produtos liberados pelas enzimas capazes de
atuar sobre os grânulos de amido, agindo sobre os glicanos lineares, como a
maltopentaose. A glicose 1-fosfato liberada é translocada para o citosol como
glicose-6-fosfato (após ação da fosfoglicomutase) por um transportador específico,
sendo então utilizada como substrato energético e para a síntese de sacarose (LU;
SHARKEY, 2006).
A forma citosólica (Pho2) parece atuar sobre glicanos de alto peso molecular.
Tal observação foi tomada por muitos como uma contradição, visto que o glicano de
alto peso molecular sobre o qual a enzima poderia agir seria o amido que está
presente dentro do plastídeo, ou seja, fora do alcance da enzima. Recentemente,
Fettke et al. (2004) encontraram um heteroglicano de alto peso molecular localizado
no citosol de células de folhas de ervilhas (Pisum sativum), sobre o qual a Pho2
atuava com alta afinidade. Este mesmo tipo de glicano foi identificado em células de
Arabidopsis thaliana (Fettke et al., 2005a) e em folhas de batata (Fettke et al.,
2005b) e acredita-se que este possa ser o substrato natural desta isoforma da amido
fosforilase.
O modelo mais recente proposto para folhas (Fettke et al., 2005b) parece
indicar que a amido fosforilase citosólica atua na liberação de glicose 1-fosfato a

partir de um provável heteroglicano citosólico, formado a partir da maltose produzida
da degradação do amido, por ação da β-amilase e transportado para o citosol via um
transportador específico. Uma das unidades de glicose provenientes da maltose
seriam transferidas para o glicano por meio de uma transglicosidase, conhecida
como disproportionating enzyme 2 (DPE2).
Em frutos de bananeira foram identificadas e caracterizadas por Mota et al.
(2002) as duas isoformas de amido fosforilase e essas se diferenciaram pela
localização e pela afinidade por substratos. A Pho2, 115KDa com subunidade
90,6KDa mostrou alta afinidade por glicanos ramificados e a Pho1, 290KDa com
subunidade 112KDa exibiu baixa afinidade por glicanos ramificados.
A atividade da fosforilase na banana se mostrou constante na sua variação
conforme os obtidos por Mota et al. (2002), diferindo do que Hubardd et al. (1990)
observou; onde a atividade da fosforilase aumentou durante todo o processo de
amadurecimento. Hubardd et al. (1990) tratou os frutos com etileno obtendo tal
aumento, mas resultados obtidos por Mainardi et al. (2006) demonstraram que
bananas tratadas com etileno não mostraram nenhum aumento na atividade e
quando tratadas com 1-metilciclopropeno (1-MCP), um antagonista do etileno,
obteve-se um aumento na atividade, sugerindo, então, que o bloqueio na percepção
ao etileno possui um efeito positivo na atividade da fosforilase.
1.2.4. Isoamilases
As isoamilases compreendem um grupo de enzimas com atividade α-1,6
glicosidásica. Dada a alta quantidade de amilopectina e, por conseguinte de ligações
α-1,6 presentes em boa parte dos grãos de amido, supõe-se que tais enzimas
desempenhem importante papel na degradação do amido.

Bierhals et al. (2006) Identificaram uma isoforma de isoamilase cuja
expressão gênica e atividade aumentavam durante o amadurecimento da banana.
Porém, é possível que outras isoformas da enzima sejam expressas neste fruto,
dada a aparente diferença de perfis encontrados para a atividade de extrato bruto
protéico quando medidas com diferentes substratos.
1.2.5. Enzima-D (DPE - Disproportionating Enzymes)
A enzima-D (4-α-D-glicanotransferase - Disproportionating Enzyme - DPE) é
encontrada tanto nos plastídeos como no citosol, e está envolvida no metabolismo
dos maltooligossacarídeos provenientes da degradação do amido. Seu papel está
bem descrito principalmente em folhas de Arabidopsis thaliana, durante a
degradação do amido transitório. Mutantes defectivos na expressão desta enzima
tiveram um grande acúmulo de maltotriose no período da noite (CRITCHLEY et al.,
2005) no ciclo dioturno em Arabidopsis thaliana.
Os maltooligossacarídeos são os substratos naturais das DPEs que possuem
atividade de transglicosidase, catalisando principalmente a conversão da maltotriose
em maltooligossacarídeos de maior peso molecular (no caso da DPE1), substratos
estes mais adequados para a ação de β-amilases e amido fosforilases que tem
como produto a maltose e a glicose-1-fosfato que são exportadas para o citosol por
transportadores específicos.
A DPE1, aparentemente não pode utiliza a maltose, produto da ação da β-
amilase, mas estudos recentes em Arabidopsis thaliana mostram que há uma
isoforma citosólica desta enzima capaz de atuar sobre este dissacarídeo
demonstrando semelhante atividade transglicosidásica, sendo denominada DPE2.

Em mutantes com perda desta enzima foi observada a inibição da degradação do
amido e acúmulo de maltose.
Em Arabidopsis thaliana, foi atribuída a DPE2 a transferência de unidades de
glicose da maltose para polissacarídeos, heteroglicanos, formados principalmente
por arabinose e galactose. Estes, após sucessivas glicosilações promovidas pela
enzima, atingiriam tamanho tal que se tornariam substratos adequados para ação da
amido fosforilase citosólica que possui afinidade por glicanos de alto peso molecular.
Porém, estudos em folhas de batata, com anticorpos específicos contra a
DPE2, mostraram sua localização no plastídeo, contrastando com aqueles
observados em Arabidopsis thaliana, dando assim margem a uma outra hipótese do
papel desta enzima na degradação do amido, diferente daquela levantada para a
planta modelo. Com a DPE2 situada no cloroplasto esta poderia usar os produtos de
degradação de amido como aceptor de glicose fornecendo maltooligossacarídeos de
tamanho adequado para a ação da β-amilase. A maltose seria então exportada para
o citosol onde seria degradada por outra enzima, como a α-glicosidase (LLOYD et
al., 2005).
A atividade ou expressão gênica de outras enzimas correlacionadas à
degradação do amido em outros modelos vem sendo identificadas em frutos que
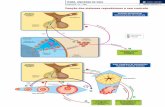
acumulam amido. Dentre estas destaca-se a glucano-água diquinase, detectada em
bananas e maçãs em amadurecimento. Esta parece desempenhar papel
fundamental na mobilização do amido transitório em folhas, conforme descrito na
planta modelo Arabidopsis thaliana (Smith et al. 2005 – Figura 2). Sua presença em
frutos parece indicar uma semelhança entre as vias de degradação de amido neste
órgão e na folha.

GWDPWD
α-amilaseISA3
ISA3α-amilase
GlicanosRamificados
GlcanosLineares
Glicose-1-P
Amido
Pho1
Maltose
Maltotriose
β-amilase
Glicose
DPE1
Triose-P
MEX1
Maltose
Glicose
DPE2
Heteroglicanos
Hexose-P
Hexoquinase
Pho2
Sacarose
Cloroplasto
Triose-P
1
2
GWDPWD
α-amilaseISA3
ISA3α-amilase
GlicanosRamificados
GlcanosLineares
Glicose-1-P
AmidoAmido
Pho1
Maltose
Maltotriose
β-amilase
Glicose
DPE1
Triose-P
MEX1
Maltose
Glicose
DPE2
Heteroglicanos
Hexose-P
Hexoquinase
Pho2
Sacarose
Cloroplasto
Triose-P
1
2
Smith et al., 2005
GWDPWD
α-amilaseISA3
ISA3α-amilase
GlicanosRamificados
GlcanosLineares
Glicose-1-P
Amido
Pho1
Maltose
Maltotriose
β-amilase
Glicose
DPE1
Triose-P
MEX1
Maltose
Glicose
DPE2
Heteroglicanos
Hexose-P
Hexoquinase
Pho2
Sacarose
Cloroplasto
Triose-P
1
2
GWDPWD
α-amilaseISA3
ISA3α-amilase
GlicanosRamificados
GlcanosLineares
Glicose-1-P
AmidoAmido
Pho1
Maltose
Maltotriose
β-amilase
Glicose
DPE1
Triose-P
MEX1
Maltose
Glicose
DPE2
Heteroglicanos
Hexose-P
Hexoquinase
Pho2
Sacarose
Cloroplasto
Triose-P
1
2
GWDPWD
α-amilaseISA3
ISA3α-amilase
GlicanosRamificados
GlcanosLineares
Glicose-1-P
AmidoAmido
Pho1
Maltose
Maltotriose
β-amilase
Glicose
DPE1
Triose-P
MEX1
Maltose
Glicose
DPE2
Heteroglicanos
Hexose-P
Hexoquinase
Pho2
Sacarose
Cloroplasto
Triose-P
1
2
GWDPWD
α-amilaseISA3
ISA3α-amilase
GlicanosRamificados
GlcanosLineares
Glicose-1-P
AmidoAmido
Pho1
Maltose
Maltotriose
β-amilase
Glicose
DPE1
Triose-P
MEX1
Maltose
Glicose
DPE2
Heteroglicanos
Hexose-P
Hexoquinase
Pho2
Sacarose
Cloroplasto
Triose-P
1
2
DPE1
Triose-P
MEX1
Maltose
Glicose
DPE2
Heteroglicanos
Hexose-P
Hexoquinase
Pho2
Sacarose
Cloroplasto
Triose-P
1
2
Smith et al., 2005
Figura 2. Esquema da suposta via de degradação de amido em folhas de Arabidopsis
thaliana. GWD (glucan-water dikinase), PWD (phosphoglucan-water dikinase), ISA3
(isoamilase), Pho1 (amido fosforilase plastidial), Pho2 (amido fosforilase citsólica), DPE1 e 2
(enzima-D, disproportionating enzymes), MEX1 (transportador de maltose), 1- transportador
de glicose, 2- transportador de triose fosfato.
Embora a mobilização de amido no amadurecimento venha sendo estudada
em frutos como a banana e maçã, outros frutos que acumulam quantidades
significativas deste polissacarídeo e que apresentam grande importância econômica
ainda não foram estudados com o mesmo grau de profundidade.
Dentre estes frutos destaca-se a manga, que acumula teores que variam de 4
a 8% do seu peso fresco. A mobilização do amido é de fundamental importância
para o adoçamento do fruto e, portanto, para a sua aceitação pelo consumidor.

1.3. Manga
A manga (Mangifera indica Linn.), um fruto tropical de importância comercial,
é uma dicotiledônea da família Anacardiaceae, produzida em países tropicais, tendo
como principal produtor a Ásia. (LIZADA, 1993).
A maior parte da produção se concentra em países como México, Filipinas,
Índia, Paquistão, África do Sul e Brasil. A produção de manga no Brasil vem
crescendo, em relação à exportação, que explora a entressafra do mercado,
utilizando técnicas de indução floral e de pós-colheita. A maior parte da produção
provém do Nordeste e também de São Paulo e Minas Gerais, e esta produção é
principalmente destinada ao mercado europeu e norte americano (PEROSA et. al.,
2006). As principais variedades produzidas são Tommy Atkins, Keitt, Kent, Van
Dyke, Palmer e Haden. O Estado maior produtor de manga é a Bahia com 23%,
seguido de São Paulo com 21%, enquanto as produções de Pernambuco e Minas
Gerais totalizam 21% e as do Ceará e Paraíba 14%.
Sua comercialização e transporte à longa distância são dificultados por sua
alta pericibilidade, particularmente a alta susceptibilidade a doenças, extremos de
temperatura e injúrias mecânicas. Muitas das tecnologias pós-colheita são
designadas para o controle de doenças e proteção contra injúrias durante a
embalagem e o transporte (LIZADA, 1993) no sentido de aumentar a vida de
prateleira do fruto. Alguns exemplos são a embalagem em atmosfera modificada,
irradiação e aplicação de hormônios, como ácido giberélico. No entanto, mesmo
essas medidas não têm apresentado resultados satisfatórios (BERNARDES-SILVA,
2004; LIZADA, 1993).

A qualidade da manga é influenciada pela variedade, valor nutricional e pelas
condições ambientais durante o desenvolvimento da planta (THARANATHAN, et al.,
2006) e também depende dos processos fisiológicos que ocorrem durante o
amadurecimento e desenvolvimento do fruto (LIZADA, 1993). A composição química
da manga varia com o local onde foi cultivada, a variedade e estágio de maturidade.
Os maiores componentes da manga são a água, carboidratos, ácidos orgânicos,
minerais, pigmentos, taninos e vitaminas (THARANATHAN et al., 2006).
Como fruto climatérico, apesar de não estar estabelecido quando ocorre o
climatério, se antes ou depois da colheita comercial (BERNARDES-SILVA, 2004), o
amadurecimento da manga é caracterizado pelas mudanças na cor, aroma, textura e
sabor (WHILE, 2002), devido à degradação da clorofila (que em alguns casos pode
mascarar a presença de pigmentos), síntese de antocianinas e acúmulo de
carotenóides como o β-caroteno, xantofilas e licopeno, embora algumas cultivares
possam reter a maior parte da cor verde, mesmo após alcançar o amadurecimento
pleno; aumento do aroma devido à produção de compostos voláteis e degradação
de flavonóides e taninos, além do aumento da síntese de etileno e da respiração
(THARANATHAN et al., 2006; SATYAN et al., 1986), tornando o fruto atrativo para o
consumo.
A mudança na composição dos compostos voláteis, sintetizados pelo fruto na
transição do estágio verde para o maduro é fundamental para o desenvolvimento de
um dos principais atributos de qualidade da manga. Embora o aroma do fruto
maduro seja facilmente reconhecível, este não pode ser atribuído a um único
composto, a exemplo do que ocorre com a maioria dos aromas. Porém, MacLeod e
Pieris (1984) atribuem ao dimetilestireno, encontrado nas principais cultivares de
manga comercializadas, o aroma característico do fruto maduro.

1.4. Degradação do amido em mangas Keitt
Ao estudar as variedades de manga Palmer, Haden, Tommy Atkins e Van
Dyke, Bernardes-Silva et al. (2003) observaram que estas variedades apresentavam
um perfil de acúmulo de amido durante boa parte do desenvolvimento do fruto. No
entanto, a degradação da reserva se iniciava dias antes do ponto considerado pelos
produtores como o ideal para a colheita, indicando uma provável atividade amilolítica
associada a este processo.
No entanto, a manga Keitt, também analisada pelos autores acima citados,
possui um padrão de amadurecimento muito particular em relação às demais
cultivares estudadas por Bernardes-Silva e Cordenunsi (2004). Esta variedade não
amadurece antes da colheita e não sofre abscisão da árvore. O amido, por sua vez
acumula tanto durante o desenvolvimento quanto após a colheita e é todo
degradado após a colheita. Tais características a tornam um interessante modelo de
estudo do padrão de degradação e síntese de açúcares em mangas.
A variedade Keitt possui frutos grandes (até 15 cm de comprimento, 600g a
800g), ovalados, de casca amarelo-esverdeada, geralmente com vestígios leves cor-
de-rosa, polpa de tom amarelo-intenso, sem fibras, firme, suculenta e doce
(LACERDA, et al., 2004). Originária de Homestead, Flórida foi introduzida no Brasil
na década de 60, é a variedade mais tardia, tendo a sua colheita nas entressafras
de outras variedades de manga.
Em um estudo posterior, Bernardes-Silva et al. (2004), analisaram com mais
detalhes a degradação do amido e a atividade de enzimas ligadas ao processo,
durante o desenvolvimento e amadurecimento da manga Keitt.

Esta manga, aparentemente não sintetiza grandes quantidade de etileno (0,1
µL.kg–1.h-1) e CO2 (45 mL.kg–1.h –1 ) na pós-colheita (Figura 3). Contudo os resultado
parecem coerentes com os encontrados em outras variedades de manga, como a
variedade Alphonso, que produz de 0,02-0,18 µL.kg–1.h-1 de etileno durante o
amadurecimento.
0 2 4 6 8 10 12 14 160,00
0,02
0,04
0,06
0,08
152025303540455055
mL
CO
2/K
g/h
uL e
tilen
o/K
g/h
Dias após colheita
Bernardes Silva e Cordenunsi, 2004 Figura 3. Perfil da produção de etileno e CO2 (respiração) durante amadurecimento da
manga Keitt.
Os principais açúcares solúveis acumulados durante o desenvolvimento e
amadurecimento da manga Keitt são a glicose, frutose e sacarose, com
predominância da última. Em um dos experimentos conduzidos por Bernardes-Silva
e Cordenunsi (2004), os teores de glicose detectados foram de aproximadamente de
1,5%; já a frutose aumentou durante o desenvolvimento e amadurecimento
chegando a 3,3% e a sacarose atingiu cerca de 5,3% no 9° dia pós-colheita. O início
do acúmulo da sacarose coincidiu com o início da degradação do amido por volta do

5° dia pós-colheita. Este padrão foi observado em vários ensaios independentes,
segundo os autores.
O amido é acumulado na manga Keitt tanto durante o desenvolvimento
quanto após a colheita, chegando a 8% do peso fresco do fruto, por volta do quinto
dia pós-colheita, onde se dá início ao processo de degradação (Figura 4 -
BERNARDES-SILVA et al., 2004). Porém, no caso desta cultivar, o processo de
amadurecimento começa com o fruto ainda ligado à planta e nem sempre se
observam mudanças próprias do amadurecimento, exceto pela perda de firmeza.
Amido
Sacarose
Dias após colheita
Bernardes Silva e Cordenunsi, 2004
90 120 180
Figura 4. Perfil da síntese da sacarose e da degradação do amido durante o
desenvolvimento e amadurecimento da manga Keitt.
Algumas das enzimas conhecidas por participarem da degradação do amido
foram analisadas quanto às variações de atividade em extratos protéicos obtidos
durante o período de desenvolvimento e amadurecimento. A atividade de α-amilase
aumentou durante o desenvolvimento do fruto. No amadurecimento, contudo, foram

detectadas flutuações na atividade da enzima, o que sugere a expressão de
isoformas da α-amilase.
A atividade de β-amilase foi pouco expressiva durante o desenvolvimento,
aumentando durante o amadurecimento, coincidindo com o início da degradação do
amido e síntese de sacarose (Figura 5), perfil semelhante ao encontrado em banana
(NASCIMENTO et al., 2006).
As atividades das amido fosforilases também foram analisadas indicando
aumentos muito pronunciados no período de síntese do amido, ou seja, foram altas
durante o desenvolvimento até o ponto da colheita, após o qual variaram pouco.
Este perfil também é encontrado na banana (MOTA et al., 2003), e sugere que, dada
a atividade tanto sintética como fosforolítica destas enzimas, na fase de
desenvolvimento as amido fosforilases estariam ligadas ao processo de formação do
grânulo de amido, ao passo que, durante o amadurecimento, talvez exerçam um
papel na mobilização do polissacarídeo, paralelamente ao das enzimas hidrolíticas.
β-amilase
α-amilase
Dias após colheita
90 120 180
Bernardes Silva e Cordenunsi, 2004
Figura 5. Atividade de α-amilase e β-amilase durante o desenvolvimento e amadurecimento.

Os dados produzidos por Bernardes-Silva e colaboradores avançaram de
forma significativa o entendimento dos processos relacionados ao metabolismo
amido-sacarose em mangas, particularmente na variedade Keitt. Do mesmo modo,
abriram várias possibilidades de estudos deste metabolismo em outras frentes. Uma
de particular interesse refere-se ao sinais hormonais correlacionados ao início da
degradação do amido.
O perfil de produção de etileno (Figura 3) não permite estabelecer uma
correlação temporal com o início da degradação do amido, visto que o máximo de
produção do hormônio parece ocorrer em um período posterior à fase de rápida
mobilização do polissacarídeo. Além disso, o fato deste pico de etileno ter sido
detectado após o pico respiratório e pelos níveis em que foram observados,
também colocam em dúvida a possibilidade de tratar-se realmente de um pico de
produção de etileno característico de um fruto dito climatérico. Também não é
possível descartar a hipótese de o perfil obtido ser decorrente de uma dificuldade em
detectar a emissão de etileno na manga, em vista dos reduzidos valores
encontrados, comparativamente a frutos como a banana.
Dado os precedentes citados nesta introdução que indicam um possível papel
de hormônios como ácido indol-3-acético (AIA) e o ácido abscísico (ABA) no
amadurecimento de frutos, pode-se aventar a hipótese na qual tanto o AIA quanto o
ABA possam atuar durante o desenvolvimento e amadurecimento da manga,
particularmente da variedade Keitt, produzindo efeitos no metabolismo do amido e,
conseqüentemente, contribuindo para a modulação do processo de adoçamento do
fruto.
Além da regulação hormonal envolvida no metabolismo do amido na manga
Keitt outro aspecto a ser explorado, derivado do trabalho de Bernardes-Silva e

Cordenunsi, diz respeito à expressão gênica das enzimas ligadas a degradação do
amido, cujos perfis são muito pouco conhecidos, não apenas nesta, mas também em
outras cultivares de manga.
Tais estudos, além de aprofundar o entendimento de um processo
fundamental para a qualidade dos frutos de Mangifera, também fornece um
contraponto interessante vindo de outra espécie, neste caso uma dicotiledônea, ao
modelo mais estudado de degradação de amido em frutos, que é o encontrado na
banana, uma monocotiledônea.

2- OBJETIVOS
O objetivo geral visa estudar a correlação entre os níveis dos fitormônios
ácido indol-3-acético e ácido abscísico sobre o metabolismo do amido, durante o
amadurecimento e desenvolvimento da manga.
Os objetivos específicos visam avaliar as variações nos níveis do ácido indol-
3-acético e ácido abscísico e avaliar as variações de expressão gênica das enzimas
α-amilase, β-amilase, amido fosforilase, durante o desenvolvimento e o
amadurecimento de mangas da cultivar Keitt.

3- MATERIAL E MÉTODOS
3.1. Material
Para as análises efetuadas no presente trabalho foram utilizados duas
amostragens de mangas da cultivar Keitt: a primeira foi realizada por Bernardes-
Silva e Cordenunsi, e resultaram nos dados apresentados na Introdução. A partir
deste material foram realizados os ensaios relativos à determinação dos hormônios
ácido 3-indol-acético (AIA) e ácido abscísico (ABA), e Northern Blot Reverso para a
avaliação da transcrição de genes de enzimas correlacionadas à degradação do
amido.
Deste modo, nesta primeira amostragem foram utilizadas mangas da
variedade Keitt obtidas da região de Joanópolis (SP), em duas fases: durante a
maturação fisiológica, com 90, 120 e 180 dias pós-antese (dpa) e durante o
amadurecimento. Para os testes relativos ao amadurecimento, as amostras foram
coletadas com cerca de 207dpa e armazenadas em câmara tipo BOD, com
temperatura e umidade controladas, sendo amostradas durante o amadurecimento.
Os frutos (no mínimo dois) foram descascados, picados e congelados em nitrogênio
líquido e armazenados a -80ºC. Todas as amostras foram pulverizadas com o auxílio
de nitrogênio líquido antes da sua utilização. O amadurecimento foi monitorado
através de medidas diárias da produção de etileno e da respiração (CO2 liberado).

90 120 180 207 208 210 212 216
0 1 3 5 9
Dias após antese (dpa)
Dias após colheita (dpc)
90 120 180 207 208 210 212 216
0 1 3 5 9
Dias após antese (dpa)
Dias após colheita (dpc)Figura 6. Diagrama da coleta das amostras da primeira amostragem. À esquerda, coleta
dos frutos verdes e à direita, coleta dos frutos maduros.
Uma segunda amostragem, obtida na safra 2006-2007, foi utilizada para
avaliar os efeitos de tratamentos hormonais e do 1-MCP sobre as atividades das
enzimas de degradação de amido. Neste ensaio foram feitas as medições de CO2 e
etileno, açúcares solúveis, determinação de amido, atividade de α-amilase, β-
amilase e fosforilase, e foram utilizadas mangas da variedade Keitt coletadas no
ponto de colheita na mesma região de Joanópolis (SP). Os frutos foram
armazenados em câmara tipo BOD, com temperatura e umidade controladas, sendo
amostrados durante o amadurecimento. Os frutos foram descascados, picados e
congelados em nitrogênio líquido e armazenados a -80ºC. Todas as amostras foram
pulverizadas com o auxílio de nitrogênio líquido antes da sua utilização.
3.2. Métodos
3.2.1. Extração de Ácido Indol-3-Acético (AIA) e Ácido Abscísico (ABA)
O método para a extração e purificação parcial do AIA e do ABA da polpa de
mangas Keitt foi o descrito por Chen et al. (1988) com modificações.
Para tal, 0,5 g de polpa de manga congelada e 1 µg de padrão interno (13C6)-
AIA (Cambridge Isotopes, Inc) foram homogeneizados por 1 minuto em solução de 2
mL de isopropanol 65% em tampão imidazol 0,2 M, pH 7,0 e centrifugados a 12.000
x g por 10 minutos. O sobrenadante, diluído 6 vezes em água deionizada, foi
aplicado em coluna de extração de fase sólida (aminopropil), pré-lavada com 2 mL

de hexano, 2 mL de acetonitrila, 2 mL de água e 2 mL de tampão imidazol e lavada
após a aplicação da amostra na coluna, com 2 mL de hexano, 2 mL de acetato de
etila, 2 mL de acetonitrila e 2 mL de metanol. A fração contendo os hormônios foi
eluída em 3 mL de metanol com 2% de ácido acético, seca em nitrogênio e
ressuspendida em 20 µL de metanol.
3.2.2. Derivatização
Para a derivatização do AIA e do ABA, foi adicionado 1 mL de diazometano
em éter, obtido a partir do reagente N-Methyl-N-Nitroso-P-Toluenesulfonamida
(Diazald - Sigma) conforme Chen (1988) em 20 µL de amostras por 15 minutos a
temperatura ambiente. A mistura de reação foi seca em nitrogênio e ressuspendida
em 20 µL de acetato de etila.
3.2.3. Determinação e análise dos hormônios vegetais por CG-EM-MSI
As análises foram realizadas em cromatógrafo a gás Hewlett-Packard modelo
6890 acoplado a detector por espectrometria de massa modelo 5973. A coluna
utilizada nas separações foi a HP-1701 (30 m, D.I. 0,25 mm, 0,50 µm de espessura
do filme interno) tendo hélio como gás carreador, com fluxo de 1 mL/min. As injeções
foram feitas automaticamente por meio do injetor Hewlett-Packard modelo LS-1100 e
o volume injetado foi de 1 µL, sem a utilização do divisor de amostras (splitless),
com fluxo de ventilação de 20 mL/min após meio minuto da injeção. A coluna foi
mantida a 150oC por 3 minutos, seguida de rampa de temperatura de 2oC min-1 até
200oC. Os íons monitorados foram aqueles com relação massa/carga (m/z) em 130
e 189 correspondentes ao AIA endógeno e 136 e 195 correspondentes ao padrão
interno de [13C6]-AIA. Para o ABA foram monitorados os íons com m/z 134, 162 e

190. A concentração de AIA endógeno foi obtida pela relação entre as áreas dos
picos de AIA nos cromatogramas extraídos em m/z 130 e 136 e a concentração de
ABA por meio de curva padrão (0,1 µg a 10,0 µg) de ABA.
3.2.4.Extração de RNA Total
A extração de RNA total foi realizada segundo a metodologia descrita por
López-Gomes e Gomes-Lim (1992) com modificações, utilizando pedaços de polpas
congeladas de manga, 3 amostras antes da colheita (90 dias pós-antese - dpa;
120dpa; 180dpa), 1 em ponto de colheita (207dpa) e 4 após a colheita (1, 3, 5, 9
dpc). Amostras com 2 g de fruto foram trituradas em gral de porcelana e mantidas
congeladas pela adição de nitrogênio líquido. Lentamente, ao material ainda
congelado, foram adicionados 10 mL de solução extratora (3 mL de Tris-borato
0,5M, pH 7,5; 1 mL de EDTA 0,5M; 2 mL de SDS 0,5M; 100 µL de Mercaptoetanol;
3,9 mL de água), sob constante agitação com pistilo até o completo
descongelamento. A mistura foi transferida para um tubo de 50 mL e centrifugada a
12000 x g, a 4°C por 15 minutos. Ao sobrenadante, sem o precipitado, mantido em
gelo, foi adicionado 2,5 mL de etanol absoluto e 1,1 mL de acetato de potássio 5 M
(KAc) e agitado por inversão por 1 minuto. Seguiu-se a adição de 10 mL de
clorofórmio e centrifugação por 10 minutos a 12000 x g a 4°C. Foi então adicionado
ao sobrenadante (fase aquosa, que contém RNA) 10 mL de fenol:clorofórmio (1:1),
centrifugada por 10 minutos a 12000 x g a 4°C e novamente separada e extraída
com 10 mL de clorofórmio. Após centrifugação a 12000 x g por 10 minutos a 4°C, a
fase aquosa foi transferida para um tubo de vidro COREX e o RNA foi precipitado
por adição de LiCl 10M (5 mL) e mantido a –20°C por 16 horas.

O tubo contendo o RNA precipitado foi centrifugado por 1 hora a 10800x g a
4°C, o sobrenadante foi descartado e o precipitado foi retomado em 2 mL de tampão
TE pH 8,0 (10mM Tris-HCl pH 8,0, 1mM EDTA). Ao RNA solubilizado foram
adicionados 6 mL de etanol absoluto e 200 µL de acetato de sódio (NaAc) 3M, pH
5,2. O tubo foi mantido a –20°C por 2 horas, centrifugado por 1 hora a 10800 x g a
4°C e o precipitado foi retomado em 200 µL de tampão TE pH 8,0.
Para avaliar a integridade do material, foi realizada uma eletroforese em gel
de agarose (1%, 80V, 15 minutos). Ao material solubilizado em água foi adicionado
0,1 volume de acetato de sódio 3M e 3 volumes de etanol absoluto gelado, para
manter o RNA precipitado e armazenado em freezer -80°C.
3.2.5. Síntese do cDNA
A síntese de cDNA de manga dos 9 pontos da extração de RNA total foi feita
a partir de 1 µg do RNA total por reação de transcrição reversa utilizando o kit Im
Prom II – Reverse Transcription System seguindo as instruções do fabricante
(Promega, Inc.). Como iniciador de síntese da cadeia foi utilizado Oligo dT15.
3.2.6. Northern Blot Reverso
3.2.6.1. Síntese de sondas de cDNA
As sondas foram sintetizadas a partir de fragmentos dos genes de α-amilase,
β-amilase, fosforilase e actina obtidos por reação em cadeia da polimerase (PCR)
utilizando “primers” específicos para cada fragmento, tendo o cDNA de banana como
molde. A amplificação dos fragmentos seguiu o seguinte protocolo: desnaturação a
94°C por 5 minutos; 30 ciclos com desnaturação a 94°C por 30 segundos,
temperatura de pareamento variável (de 50°C a 60°C) por 30 segundos, extensão a

72°C por 2 minutos, extensão final a 72°C por mais 7 minutos e incubação a 4°C.
Para α-amilase a temperatura foi de 58°C, para β-amilase foi de 57°C.
3.2.6.2. Transferência para membrana
Para a transferência dos fragmentos amplificados de α-amilase, β-amilase,
fosforilase e actina, aos produtos de PCR obtidos nas respectivas reações foi
adicionado SSC (NaCl 0,15 M; citrato de sódio 0,015 M) de forma a obter solução
final em 10 X SSC. As reações foram aquecidas a 95°C por 30 min para
desnaturação do DNA e colocadas imediatamente no gelo. As reações foram
transferidas, sob vácuo, para membrana de nylon (HybondN+, Amersham) utilizando
o aparelho Bio-Dot (Bio-Rad). Após a transferência a membrana foi colocada sobre
papel toalha umedecido com solução de desnaturação (NaOH 50mM, NaCl 0,5 M)
por 5 minutos e transferida para outro papel toalha umedecido com solução de
neutralização (Tris-HCl 100mM, pH 7,0, NaCl 0,5 M). Após secagem em temperatura
ambiente, os cDNAs foram fixados à membrana por tratamento com ultravioleta no
forno ( UV-crosslinker da Hoefer) sob condições de energia e tempo padrão do
aparelho. Para avaliação da uniformidade da transferência, as membranas foram
coradas com solução de azul de metileno.
3.2.6.3. Preparo das sondas radioativas e hibridização
Para a reação de marcação das sondas radioativas foram utilizados como
moldes os RNAs totais extraídos das polpas de manga Keitt amostrados aos 90,
120, 180, 207 dias pós antese; 1, 3, 5, 9 dias pós-colheita.
As reações de marcação das sondas de cDNA foram compostas de 10 µg de
RNA total (previamente desnaturado a 65oC por 5min), 2 µL de oligo (dT)18 (1

µg/µL), 10 µL de tampão Superscript 5 X, 0,5 µL RNAsin, 5 µL de DTT 0,1M, 5 µL
d(AGT) 10 mM, 5 µL de α-[32P]-dCTP (50µCi), 1 µL de transcriptase reversa
Superscript II. A mistura foi incubada por 1 hora a 42oC, adicionada de 2 µL de dCTP
25 mM não marcado e incubada por mais 45 minutos a 42oC. A sonda foi purificada
em coluna Sephadex G-50 (Microspin Columns, Amersham Biosciences) conforme
instruções do fabricante.
As membranas contendo os fragmentos dos genes de estudo foram pré-
hibridizadas com 10 mL de solução de bloqueio PerfectHyb da Sigma a 65oC por 2
horas em garrafas de vidro. As sondas purificadas, após desnaturação a 95oC por 5
min, foram adicionadas diretamente à solução de pré-hibridização, e mantidas nesta
solução a 65oC por 16 horas para hibridização. Ao fim do período as membranas
foram lavadas 2 vezes com solução 2 X SSC/0,1% SDS a temperatura ambiente,
embaladas em filme de PVC e expostas a filmes de raios-X a –80oC (Sambrook et.
al.,1989). As membranas foram lavadas conforme as instruções do fabricante da
membrana de nylon Hybond N+ (Amersham Biosciences).
3.2.6.4. Análise densitométrica
Após a revelação, as bandas visíveis no filme de raio-X foram digitalizadas no
densitômetro GS-700 da Bio-Rad, e analisadas com o software Quantity One v5.2 da
mesma empresa. As bandas referentes ao gene da actina foram utilizadas para a
normalização das bandas dos genes em estudo.
3.2.7. Medida de Etileno e CO2
Para as análises de etileno e respiração, os frutos em ponto de colheita foram
mantidos em câmara BOD (demanda bioquímica de oxigênio) com temperatura de

20°C. Os frutos foram encerrados individualmente em frascos hermeticamente
fechados. Depois de 1h 30 min, 10 mL (para análise de etileno) e 1 mL (para análise
de CO2) foram coletados usando seringas “gas-tight”, e foram medidos através de
cromatografia a gás (HP-6890, Agilent Tecnologies).
Para determinação do etileno foi utilizado o detector de ionização de chamas
(FID) e para o CO2 foi utilizado o detector de condutividade térmica (TCD). Em
ambas as análises foram utilizadas a coluna HP-Plot Q (30 m de comprimento, 0,53
mm de diâmetro, Agilent Technologies), a temperatura do injetor e do detector foi de
250°C e a temperatura isotérmica foi de 30°C. O hélio foi usado como gás de arraste
em um volume de 1 mL min-1 para o etileno e 4 mL min-1 para o CO2. O modo de
injeção para o etileno foi o “pulsed splitless” e para o CO2 foi o “split 50:1”. Após as
análises, amostras de diferentes dias foram descascadas, picadas e congeladas em
nitrogênio líquido e guardados a –80°C.
3.2.8. Tratamento com 1- Metilciclopropeno (1-MCP)
Os frutos em ponto de colheita (aproximadamente com 205 dias pós antese)
foram mantidos em câmara BOD com temperatura de 20°C foram encerradas em
frascos e tratados com 1-MCP (100 nL/L - Ethylbloc, Rohm e Haas, Co) por 16 horas
e frutos que não foram tratados com o 1-MCP foram considerados como controle.
Após o tratamento com 1-MCP foram realizadas as análises em cromatografia
a gás e as amostras de diferentes dias foram descascadas, picadas e congeladas
em nitrogênio líquido e guardada a –80°C.

3.2.9. Açúcares Solúveis
Para a extração dos açúcares foi utilizado 500 mg de polpa de manga
congelada homogeneizado em 3 mL de etanol 80% a 80°C sob agitação mecânica
por 15 min e centrifugados a 12.000 x g por 30 min. A extração foi repetida 3 vezes e
ao sobrenadante (3 repetições) foi adicionado etanol 80% para a correção do
volume final de 10 mL.
A partir de uma alíquota de 1 mL, o etanol foi evaporado a vácuo e
ressuspendido em 1 mL de água. Os açúcares foram identificados e quantificados,
através de HPLC (Dionex, modelo DX 500).
3.2.10. Amido
As análises de amido foram realizadas pelo método otimizado por Áreas e
Lajolo (1980), onde o amido foi solubilizado com hidróxido de sódio e neutralizado
com ácido acético, precipitado com etanol 80% e hidrolisado com amiloglicosidase.
A glicose resultante foi quantificada pelo sistema glicose-oxidase-peroxidase/ABTS
(Bergmeyer, 1974).
3.2.11. Extração e Atividade de α-amilase e de β-amilase
Para a extração, 500 mg de amostra foi homogeneizado em 2,5 mL de
solução extratora (Tampão Hepes-KOH 50 mM pH 7, benzamidina 1 mM, 1% PVP,
20 mM de cisteína) em Turrax por 30 min. Para a medida de proteína total foi
utilizada metodologia segundo Bradford (1976).
A atividade de α-amilase foi medida utilizando o reagente Ceralpha
(Megazyme Internacional Ireland, Ltd). Após incubação a 37°C por 1h 30 min de 20
µL de extrato enzimático e 20 µL do reagente, a reação foi interrompida com adição

de 160 µL de Tris 1,1%. Para a atividade de β-amilase foi utilizado o reagente
Betamyl (Megazyme Internacional Ireland, Ltd). Após incubação a 37°C por 1h 30
min de 40 µL de extrato enzimático e 20 µL do reagente, as reações foram
interrompidas com adição de 160 µL e 140 µL de Tris 1,1%. A absorbância do ρ-
nitofenol liberado foi lida a 410 nm e sua concentração determinada com base na
curva padrão de ρ-nitofenol.
3.2.12. Extração de proteínas e Atividade de fosforilase em gel nativo
Para a extração de proteínas, 500 mg de cada amostra foram
homogeneizados em 2,5 mL de solução extratora (Tampão Hepes-KOH 50 mM pH
7,5; benzamidina 1 mM, 1% PVP, 20 mM de cisteína) em homogeneizador do tipo
Turrax por 30 min. Para a medida de proteína total foi utilizada metodologia segundo
Bradford (1976).
Os géis de separação, para este ensaio, continham 6% de acrilamida/ bis-
acrilamida (30:0,8), 375 mM de Tris-HCl pH 8,8 e 0,4% (p/v) de glicogênio de ostra;
os géis de empilhamento foram compostos por 3,75% de acrilamida/ bis-acrilamida e
62,5 mM de Tris-HCl pH 6,8. Após a eletroforese (15 mA/gel – 4 °C), os géis foram
lavados, durante 15 minutos, com 40 mL de uma solução de citrato-NaOH 100 mM
pH 6,0 e então foram incubados por 1 hora à 37 °C em 40 mL deste mesmo meio
adicionado de 0,05% de amido solúvel de batata e glicose-1-fosfato 20 mM. Após a
incubação os géis foram novamente lavados com a solução de citrato-NaOH e por
fim foram corados com I2 0,67% (p/v) + KI 3,33% (p/v).

4. RESULTADOS E DISCUSSÃO
4.1. Determinação e análise dos hormônios vegetais por CG-EM-MSI
De modo a avaliar as possíveis interações hormonais correlacionadas à
mobilização do amido bem como às atividades enzimáticas estudadas por
Bernardes-Silva e Cordenunsi em mangas Keitt, foram realizadas análises dos níveis
de AIA e ABA no mesmo material no qual as autoras trabalharam até 2004
(conforme descrito em Material e Métodos). A escolha deste material partiu da
conveniência do mesmo estar ainda disponível e viável para análises, o que
acrescentaria maiores detalhes ao conjunto de resultados obtidos pelas
pesquisadoras.
As análises realizadas em CG-MS indicaram altos níveis de AIA no início do
desenvolvimento da manga que decresceram até o ponto de colheita e apresentou
novo aumento no intermédio do amadurecimento, voltando aos níveis da colheita ao
fim do período analisado. O ABA, por outro lado, apresentou um perfil oposto ao do
AIA, baixos níveis durante o desenvolvimento aumentando na fase amadurecimento
atingindo seus valores máximos já próximo à senescência (Figura 7).
Os resultados indicam que a degradação do amido em manga parece estar
relacionado com a diminuição de AIA e aumento de ABA, tendo o início do processo
por volta do quinto dia pós-colheita, onde se tem a interseção da queda nos níveis
de AIA e de aumento do ABA, juntamente com o ponto de máximo de acúmulo de
amido.

90 120 180207 (0)208 (1)210 (3)212 (5)213(6)214(7)215(8)216 (9)217(10)0
20
40
60
80
100
120
140
160
0
10
20
30
40
50
60
70
80
90
100
AIA
(pm
ol/g
)
ABA
(ug/
g)
90 120 180 0 1 3 5 6 7 9 108
dpa dpc
90 120 180207 (0)208 (1)210 (3)212 (5)213(6)214(7)215(8)216 (9)217(10)0
20
40
60
80
100
120
140
160
0
10
20
30
40
50
60
70
80
90
100
AIA
(pm
ol/g
)
ABA
(ug/
g)
90 120 180 0 1 3 5 6 7 9 108
dpa dpc
Figura 7. Perfil dos hormônios AIA ( ) e ABA ( ), quantificados em CG-EM-MSI, durante o
desenvolvimento e amadurecimento da manga Keitt (dpa – dias pós-antese/ dpc – dias pós-
colheita).
O alto nível de AIA encontrado durante o desenvolvimento e no 1° dia pós-
colheita condiz com estudos que demonstram que este hormônio possa atuar como
um fator de resistência ao amadurecimento, afetando alguns aspectos do próprio
amadurecimento. Frenkel e Dyck (1973) observaram um atraso na degradação da
clorofila, no amaciamento da polpa e no aumento da taxa respiratória, e um aumento
na produção de etileno em pêras tratadas com AIA. Porém, na manga Keitt o
pequeno aumento nos níveis de etileno, observado no 9º dia pós-colheita não se
sobrepõe aos eventos já mencionados, o que pode indicar que a ação do AIA não
seja dependente do etileno neste fruto.
Outro trabalho que coloca o AIA como um inibidor do amadurecimento foi
reportado por Lohani et al. (2004), que observaram que esta auxina foi capaz de
inibir enzimas de degradação da parede celular em bananas, independente do
tratamento com etileno, indicando a independência da ação do primeiro hormônio
em relação ao segundo.

Em sorgo, o tratamento com AIA em cultura contendo sacarose levou ao
aumento da quantidade de amido, sendo este associado ao processo da divisão e
extensão celular. Além disso, o hormônio foi capaz de aumentar a atividade da
enzima sacarose sintase, sendo que a sacarose fosfato sintase teve sua atividade
diminuída. (BHATIA; SINGH, 2002). Em manga este efeito parece acontecer de
forma oposta onde o acúmulo de amido ocorre concomitantemente à diminuição da
concentração de AIA (BERNARDES-SILVA; CORDENUNSI, 2004).
Outros resultados que sugerem a ação do AIA como um inibidor do processo
de degradação do amido foram obtidos por Purgatto et al. (2001) com a infiltração da
auxina em amostras de bananas. Neste modelo, os autores observaram a acentuada
inibição na degradação do amido nas amostras infiltradas com o hormônio.
Conseqüentemente, o aumento respiratório e o aumento na síntese de sacarose
foram atrasados em relação às amostras de bananas infiltradas com manitol (fatias
controle).
Em um trabalho posterior (PURGATTO et al., 2002), no qual foram medidos
os níveis da auxina livre tanto em amostras infiltratadas com o AIA como nas
amostras controle (infiltradas com manitol), constatou-se que o decréscimo nos
teores da auxina estavam temporalmente correlacionados ao início da degradação
do amido. Nas fatias tratadas com AIA, a intensa mobilização da auxina na forma
livre para suas formas conjugadas com açúcares e aminoácidos, foi o fator
preponderante para que o processo de degradação do amido fosse iniciado.
Somente após o nível de auxina nas amostras infiltradas decair ao mesmo nível das
controle é que foi registrado o início da degradação do amido. Esta, porém, ocorreu
com atraso de 5 dias.

Neste mesmo estudo, foi observado comportamento semelhante em bananas
inteiras amadurecidas naturalmente. Foi observada a independência destes eventos
em relação ao etileno, uma vez que o pico de produção deste hormônio ocorreu ao
mesmo tempo tanto nas amostras de bananas tratadas com AIA como nas amostras
controle.
Tomando estas observações como base de comparação, é possível observar
um comportamento diferenciado em relação à manga. Nesta, os níveis de AIA, altos
durante o desenvolvimento, decresceram próximo ao ponto de colheita, atingindo
valores praticamente 100 vezes menores. Porém, no início do período de
degradação do amido, no 5º dia pós-colheita (Figura 4), os teores de AIA voltaram a
subir, para novamente decrescer. Este padrão contrasta com aquele encontrado na
banana por Purgatto et al. (2002) indicando que, na manga o AIA não parece
representar um fator de inibição para a degradação do amido. Fica em aberto a
questão de qual seria a fonte da auxina para o aumento observado. Uma
possibilidade seria a mobilização do AIA para formas conjugadas que seriam, então,
hidrolisadas liberando a auxina no período anterior ao início da degradação do
amido e formação de sacarose.
As mudanças no conteúdo de ABA na polpa da manga Keitt (Figura 7)
parecem indicar uma correlação entre o aumento na síntese do hormônio e a
mobilização do amido (Figura 4), visto que estes parecem ocorrer de maneira
concomitante. Do mesmo modo, a atividade de β-amilase (Figura 5) aumentou
seguindo o aumento na síntese do ABA, o que reforçou uma hipótese, a princípio, de
um envolvimento deste hormônio na sinalização para a degradação do amido na
manga.

Em relação ao ABA, estudos em bananas mostram que ocorre uma interação
entre este hormônio e a síntese de etileno e desta forma há estimulação do
amadurecimento em frutos climatéricos, como mostrado em maçãs (LARA;
VENDRELL, 1998), onde houve o acúmulo de ABA e os frutos rapidamente
senesceram.
Em abacate, concentrações de ABA não influenciaram o amadurecimento,
avaliado pela taxa de respiração e produção de etileno, sendo um perfil que
diferente do padrão encontrado para a maçã e o tomate. Neste último, além do
aceleramento do amadurecimento houve o estimulo da síntese de licopeno
(TINGWA e YOUNG, 1975).
Gertman e Fuchs (1972) observaram em mudas de ervilhas a inibição de
etileno quando tratadas com ABA em concentrações ente 0,1µM a 1mM, mas
estimulou a síntese na casca de laranja. Também foi observado que ao tratar com
metionina houve a inibição da conversão do aminoácido em etileno em ervilhas e
nada ocorreu em laranjas, sugerindo que o mecanismo de produção de etileno
possa ser o mesmo nos dois sistemas, porém o mecanismo de controle seja
diferente.
Em bananas tratadas apenas com ABA obteve-se a aceleração da produção
de etileno e da respiração juntamente com mudanças de cor e maciez do fruto
comparado com os frutos tratados só com etileno. E tratamentos com a combinação
dos dois hormônios foram melhores também do que só com etileno, sugerindo que o
ABA seja responsável pela sensibilidade ao etileno, e atuaria como um coordenador
do amadurecimento (JIANG, et al., 1999). Lohani et al. (2004) observaram que
tratamentos com ABA juntamente com etileno em bananas resultou na maior perda

de firmeza em relação à amostra tratada apenas com etileno, indicando novamente
a ação somada do ABA.
Em plantas superiores a síntese de ABA envolve a degradação oxidativa dos
carotenóides (SETHA, et al., 2004), e a expressão de genes responsáveis pela
síntese de carotenóides são coordenados pelo etileno entre outros como a
degradação de clorofila, produção autocatalítica de etileno e conversão do amido em
açúcares (THEOLOGIS, 1992). O papel exercido pelo ABA no amadurecimento de
frutos ainda não é claro, apesar da síntese de ABA serem altos no amadurecimento.
Todos estes são exemplos do envolvimento do ABA em processos ligados ao
amadurecimento de frutos. Porém, um possível papel do ABA como sinalizador para
a síntese de enzimas ligadas ao metabolismo do amido não foi registrado até o
momento para nenhum fruto. O papel do ABA neste metabolismo tem sido relatado
em cereais, nos quais foram registrados diversos efeitos, por vezes contraditórios,
como indução (YANG et al. 2006) e inibição da síntese do amido (BHATIA e SINGH
2002), inibição (NODA et al 1994 ) da degradação.

4.2. Northern Blot Reverso
Para o melhor entendimento das relações entre as variações hormonais
registradas e a degradação do amido foram verificados os padrões de expressão
gênica das enzimas relacionadas ao processo de mobilização do amido, algumas
das quais previamente analisadas quanto à atividade no trabalho de Bernardes-Silva
e Cordenunsi (2004).
Para as extrações de RNAs totais foi considerada a determinação dos
hormônios AIA e ABA, e também a degradação de amido durante o desenvolvimento
e amadurecimento da manga. As amostras selecionadas foram avaliadas quanto a
expressão das enzimas que se acredita participar da degradação do amido: α-
amilase, β-amilase, DPE2 e isoamilase.
Para verificar a expressão destas enzimas foram realizadas várias tentativas
de obter fragmentos a partir de cDNAs de manga, utilizando primers degenerados
construídos a partir de seqüências já conhecidas das enzimas. As amplificações por
PCR, no entanto, não foram efetivas e, desta forma, optou-se por utilizar fragmentos
de enzimas previamente isolados a partir de cDNAs de banana e utilizando cDNAs
de manga marcados radioativamente.
Utilizando a técnica de Northern Blot Reverso foi possível observar a
expressão das enzimas relacionadas ao metabolismo do amido acima citadas.
Altos níveis de expressão da α-amilase foram detectados durante o
desenvolvimento do fruto e amadurecimento (Figura 8). Foram observados dois
aumentos pronunciados no mRNA: um no 207° dia pós-antese e outro no 5° dia pós-
colheita, seguindo as variações observadas na atividade enzimática que foi
determinada por Bernardes-Silva e Cordenunsi (2004).

0,00
0,50
1,00
1,50
2,00
2,50
3,00D
O A
lfa-a
mila
se /D
O A
ctin
a
90 120 180 0 1 3 5 90,00
0,50
1,00
1,50
2,00
2,50
3,00D
O A
lfa-a
mila
se /D
O A
ctin
a
0,00
0,50
1,00
1,50
2,00
2,50
3,00D
O A
lfa-a
mila
se /D
O A
ctin
a
90 120 180 0 1 3 5 990 120 180 0 1 3 5 9
Figura 8. (A) Análise densitométrica. (B) Northern Blot Reverso da α-amilase. Normalizado
com os valores obtidos da razão entre DO dos transcritos da α-amilase e DO dos mRNAs do
controle actina. (dpa – dias pós-antese/ dpc – dias pós-colheita).
Adpa dpc Adpa dpc Adpa dpc Bdpa dpc Bdpa dpc Bdpa dpc090 120 180 1 53 9
α-amilase
Actina090 120 180 1 53 9090 120 180 1 53 9
α-amilase
Actina
α-amilase
Actina
Em cereais a degradação se inicia com a ação da α-amilase, porém em
sistemas como a de Arabidopsis thaliana, na qual foram detectadas três isoformas
da enzima, duas delas parecem não atuar na degradação, pois em folhas de
mutantes que não expressavam duas destas isoformas não foi observada nenhuma
mudança na taxa de degradação do amido. Sugere-se que só uma das isoformas
possa atuar sobre o grânulo (SMITH et al., 2005).
Como a enzima não é capaz de quebrar as ligações α-1,6 da amilopectina
alguns autores sugerem a existência de outras enzimas em Arabidopsis que
possuam tal atividade, dentre as quais estão as enzimas desramificadoras, como as
isoamilases (SMITH et al., 2004; SMITH et al., 2005; LLOYD et al., 2005).
Estudos têm relacionado os níveis hormonais com a atividade expressão da
α-amilase em cereais. Skadsen (1998) observou em cevada que o ácido giberélico
(GA) induziu a transcrição e de mRNAs de α-amilase, mas houve a repressão dos
genes pelo aumento do ácido abscísico, embora este não seja o único a atuar na
repressão destes genes. O mesmo foi observado por Bethke et al. (1999), que tratou
grãos de cevada com ABA e observou baixa atividade da enzima e inibição do
acúmulo de proteína α-amilase. Bhatia e Singh (2002) observaram que a auxina foi

capaz de induzir atividade da enzima em sorgo e o ABA de reduzir sua atividade. Em
cotilédones de feijão foi observada a influência do ABA, a atividade da enzima
diminuiu em relação ao controle e não houve nenhum efeito quando estes foram
tratados com ácido indol-3-acético (AIA) e etileno (TANEYAMA et al., 1995).
Tais precedentes mostram uma gama variada de resultados associados à
regulação hormonal da α-amilase. Em fatias de bananas infiltradas com AIA,
Purgatto et al. (2001) não observaram variações expressivas na atividade de α-
amilase em comparação com fatias controle (tratadas apenas com manitol). No
entanto, o tratamento com etileno foi capaz de induzir a atividade da enzima. Porém
o dado contrastante entre a expressão de α-amilase em banana e manga refere-se
justamente ao nível de expressão. Enquanto na banana os níveis de mRNA
parecem ser baixos ao longo de todo o amadurecimento, na manga os níveis
detectados indicaram alta expressão do mRNA que codifica para a enzima. Este
resultado sugere correlação com o alto nível de marcação para α-amilase obtido em
grânulos de amido de manga isolados e incubados com antisoro anti-amilase de
banana (BERNARDES-SILVA et al. 2004).
Tais resultados sugerem um papel relevante para a α-amilase na degradação
do amido de manga e esta não parece ser regulada por etileno, AIA ou ABA.
A β-amilase também foi expressa durante todo o período analisado, porém os
níveis de expressão foram mais baixos no desenvolvimento e aumentando conforme
a evolução do amadurecimento, tendo valores máximos ao final (Figura 9). O que foi
demonstrado também com a técnica de Northern Blot convencional (dados não
apresentados), na qual foi utilizada sonda marcada radioativamente obtida a partir
de um fragmento de cDNA de β-amilase isolado de banana. Estes padrões foram
também observados por Bernardes-Silva et al. (2004) para a atividade da enzima.

0,000,200,400,600,801,001,201,401,60
DO
Bet
a-am
ilase
/ DO
Act
ina
90 120 180 0 1 3 5 90,000,200,400,600,801,001,201,401,60
DO
Bet
a-am
ilase
/ DO
Act
ina
0,000,200,400,600,801,001,201,401,60
DO
Bet
a-am
ilase
/ DO
Act
ina
90 120 180 0 1 3 5 990 120 180 0 1 3 5 9 090 120 180 1 53 9
β-amilase
Actina090 120 180 1 53 9090 120 180 1 53 9
β-amilase
Actina
β-amilase
Actina
Figura 9. (A) Análise densitométrica. (B) Northern Blot Reverso da β-amilase. Normalizado
com os valores obtidos da razão entre DO dos transcritos da β-amilase e DO dos mRNAs do
controle actina. (dpa – dias pós-antese/ dpc – dias pós-colheita).
Adpa dpc Adpa dpc Adpa dpc Bdpa dpc Bdpa dpc Bdpa dpc
Segundo a literatura, a β-amilase parece possuir uma complexa regulação,
podendo ser induzida por açúcares como sacarose, glicose ou frutose, e também por
hormônios vegetais. Mita et al. (1995) observaram que a atividade da enzima em
Arabidopsis aumentou quando folhas foram tratadas com sacarose, sendo que o
aumento foi concomitante com o aumento do transcrito. O mesmo efeito foi obtido
com o tratamento com glicose e frutose.
Alterações circadianas da atividade de β-amilase em Arabidopsis seriam
indicações de que a enzima desempenharia um papel relevante na degradação do
amido acumulado nas folhas (CHANDLER et al., 2001). De acordo com estudos
usando mutantes, a isoforma BAM3 parece ser importante para a degradação do
amido transitório em cloroplastos (LU; SHARKEY, 2006; LLOYD, et al., 2005).
A β-amilase parece ser importante na degradação do amido em banana, já
que sua atividade aumenta simultaneamente com a degradação (PURGATTO et al.,
2001), acompanhada do aumento no nível de transcritos da enzima. Ao realizar a
infiltração de ácido indol-3-acético (AIA) em fatias do fruto, foi observado atraso no
aumento da expressão e atividade da enzima, em relação a fatias controle infiltradas
com manitol.

Tal efeito parece contrastar com o resultado observado na análise dos níveis
dos hormônios obtidos no presente trabalho e da atividade de β-amilase realizada
por Bernardes-Silva (2004) (Figura 5). Segundo Bathia e Singh (2002) o AIA foi
capaz de aumentar a atividade de β-amilase em sorgo efeito similar ao observado
para a manga uma vez que o aumento da transcrição e atividade da enzima ocorreu
logo após o aumento na síntese de AIA. Além disso, é possível notar que os teores
de ABA também aumentaram pouco antes do aumento de transcrição e atividade de
β-amilase. Os resultados parecem indicar uma regulação positiva de um ou outro
hormônio, ou mesmo de ambos, sobre a β-amilase de maneira diversa da
encontrada em banana. Neste fruto, Nascimento et al. (2006) observaram que tanto
a expressão quanto a atividade parecem depender em grande parte da sinalização
do etileno.
O perfil de expressão gênica de uma isoamilase (Figura 10) apresentou
correlação muito próxima com o perfil de atividade da enzima obtido por Bernardes-
Silva et al. (2004 – dados não apresentados) indicando dois picos, um durante a
fase de desenvolvimento e outro ao final do amadurecimento. Embora as análises
conduzidas não permitam afirmar de modo categórico, é possível que tal padrão de
expressão e atividade corresponda a possíveis isoformas da isoamilase com
atividades e regulações distintas.
Devido à sua ação sobre as ramificações da amilopectina, tem-se atribuído
um relevante papel das isoamilases tanto na síntese como na degradação do amido
(KUBO et al., 1999). Mutantes de Arabidopsis com perda das enzimas mostraram
deficiência na produção de amido, produzindo no lugar deste um análogo do
glicogênio, denominado fitoglicogênio (DINGES et al., 2003).

0,000,200,400,600,801,001,201,401,60
DO
Isoa
mila
se0/
DO
Act
ina
120 18090 0 1 3 5 90,000,200,400,600,801,001,201,401,60
DO
Isoa
mila
se0/
DO
Act
ina
0,000,200,400,600,801,001,201,401,60
DO
Isoa
mila
se0/
DO
Act
ina
120 18090 0 1 3 5 9120 18090 0 1 3 5 9
Figura 10. (A) Análise densitométrica. (B) Northern Blot Reverso da isoamilase.
Normalizado com os valores obtidos da razão entre DO dos transcritos da isoamilase e DO
dos mRNAs do controle actina. (dpa – dias pós-antese/ dpc – dias pós-colheita).
Adpa dpc Adpa dpc Adpa dpc Bdpa dpc Bdpa dpc Bdpa dpc090 120 180 1 53 9
Isoamilase
Actina090 120 180 1 53 9090 120 180 1 53 9
Isoamilase
Actina
Isoamilase
Actina
Mutantes com perda da isoformas ISA1 e ISA2 em Arabidopsis e em batata
mostraram um acúmulo de maltodextrina e diminuição dos grânulos de amido
comparativamente aos grânulos da variedade selvagem (LU; SHARKEY, 2006;
HUSSAIN et al., 2002).
Bierhals et al. 2004 demonstraram a atividade desramificadora em extratos
protéicos de bananas tendo o glicogênio e a β-limite dextrina como substratos
resultando na produção de maltose, sendo que os melhores resultados, em termos
de quantidade de produto formado, foram obtidos com o segundo substrato. As
variações de atividade da enzima, medida em extratos de vários pontos do
amadurecimento foram concomitantes a mobilização do amido, mesmo com frutos
tratados com etileno. Tal resultado é similar ao encontrado por encontrado por
Bernardes-Silva et al. (2004).
Conforme Lu e Sharkey (2006) a isoforma ISA3 tem alta atividade sobre a β-
limite dextrina, sendo a forma apontada como participante na degradação do amido
transitório em folhas de A. thaliana. Tais observações sugerem uma ação conjunta
da ISA3 e da β-amilase para a produção de maltose (SMITH et al., 2005).
Os padrões de transcrição observados para a DPE2 (Figura 11) foram
similares aos encontrados para a β-amilase (Figura 9), particularmente na fase pós-

colheita. O resultado encontra correlação com dados da literatura que indicam uma
ligação entre as atividades das duas enzimas, visto que a DPE2 atua
preferencialmente sobre a maltose, produto da β-amilase. Embora esta enzima já
tenha sido descrita em folhas, e tubérculos, cabe salientar que além da banana, da
qual foi isolado o fragmento de cDNA que permitiu esta análise, este é um dos
poucos relatos da detecção desta enzima em frutos.
0,000,200,400,600,801,001,201,401,60
DO
DPE
2/ D
O A
ctin
a
90 120 180 0 1 3 5 90,000,200,400,600,801,001,201,401,60
DO
DPE
2/ D
O A
ctin
a
0,000,200,400,600,801,001,201,401,60
DO
DPE
2/ D
O A
ctin
a
90 120 180 0 1 3 5 990 120 180 0 1 3 5 9
Figura 11. (A) Análise densitométrica. (B) Northern Blot Reverso da DPE2. Normalizado
com os valores obtidos da razão entre DO dos transcritos da DPE2 e DO dos mRNAs do
controle actina. (dpa – dias pós-antese/ dpc – dias pós-colheita).
Adpa dpc Adpa dpc Adpa dpc Bdpa dpc Bdpa dpc Bdpa dpc090 120 180 1 53 9
DPE 2
Actina
090 120 180 1 53 9090 120 180 1 53 9
DPE 2
Actina
DPE 2
Actina
Lu e Sharkey (2006) sugerem que a DPE2 esteja envolvida na transferência
de unidades de glicose da maltose para hetroglicanos citosólicos, uma vez que em
mutantes de Arabidopsis com perda desta enzima ocorreu o acúmulo de maltose no
citosol (LLOYD et al., 2005).
Como houve uma expressão também praticamente constante da isoamilase é
possível especular que a expressão e talvez a atividade da DPE2 aumente mesmo
quando a atividade de β-amilase é baixa, como observado no inicio do
desenvolvimento.

4.3. Medida de Etileno e CO2
Embora as análises realizadas indiquem a possibilidade da regulação do
metabolismo do amido na manga Keitt ser modulado por outros hormônios, o papel
do etileno não poderia ser descartado, apesar dos baixos níveis encontrados no
trabalho de Bernardes-Silva e Cordenunsi (Figura 3). Desta forma, uma nova
amostragem foi realizada e parte dos frutos foi tratada com o antagonista de etileno
o 1-metilciclopropeno (1-MCP), visando avaliar a extensão dos efeitos do hormônio
sobre a mobilização do amido durante o amadurecimento.
O nível de 1-MCP usado nos tratamentos teve por base àqueles utilizados por
Wang et al. (2006), embora a literatura apresente muita variabilidade de tempos e
concentrações nos tratamentos de manga com este antagonista de etileno. Dado
que as concentrações encontradas vão de 100 nL.L-1 a 100 µL.L-1 de 1-MCP em
tempos que variam de 1 a 12 horas, optou-se por utilizar a concentração mínima
com o maior tempo. Esta decisão baseou-se na limitação da quantidade de 1-MCP
disponível para o tratamento das mangas.
As análises, iniciadas logo após o tratamento com o 1-MCP, indicam a
existência de um aumento de respiração por volta do 11° dia pós-colheita e de um
aumento da produção de etileno, com os maiores valores sendo registrados por volta
do 12° dia pós-colheita, logo após o da respiração. Em mangas tratadas com 1-
MCP, a produção de etileno foi menor que no controle, onde o aumento ocorreu por
volta do 14° dia pós-colheita. Já a respiração foi maior nas tratadas com 1-MCP
tendo um aumento por volta do 16° dia pós-colheita (Figura 12), o que condiz com a
literatura, onde tratamento com 1-MCP é capaz de atrasar a evolução do etileno em
frutos climatéricos (MAO et al., 2007; BLANKENSHIP; DOLE, 2003).

Os valores encontrados nos tratamentos controle foram muito superiores aos
encontrados por Bernardes-Silva (2004), sendo maiores já com o início da
senescência do fruto. O mesmo ocorrendo com o tratamento com 1-MCP que
produziu níveis de etileno muito maiores do que os registrados por Bernardes-Silva
(2004).
A 0 1 2
B dias
3 4 5 6 7 8 9 10 11 12 130,0
0,2
0,4
0,6
0,8
1,0
1,2
0
20
40
60
80
100
120
dias pós-colheita
Pro
duçã
o Et
ileno
ug.
kg -1
frut
o
Pro
duçã
o C
O2 m
g.kg
-1 fr
uto
0 2 4 6 8 10 12 14 16 18 200,0
0,2
0,4
0,6
0,8
1,0
1,2
0
20
40
60
80
100
120
Prod
ução
Etil
eno
ug.k
g -1 fr
uto
pós-colheita
Prod
ução
CO
2 m
g. k
g-1 fr
uto
Figura 12. Perfil da produção de etileno e respiração (produção de CO2) durante o
amadurecimento de mangas Keitt. (A) Amostras controle ( etileno, CO2). (B) Tratamento
com 1-MCP ( etileno, CO2).

4.4. Amido
Em uma nova amostragem da manga Keitt, verificou-se que o máximo
de acúmulo de amido (Figura 13) ocorreu por volta do 5° dia pós-colheita
(aproximadamente 7%), e este foi praticamente todo degradado na manga madura.
O resultado é próximo ao encontrado por Bernardes-Silva (2004, 2000) para outras
cultivares e para a própria cultivar Keitt.
Os teores encontrados no tratamento com 1-MCP indicam que o máximo de
acúmulo de amido ocorreu por volta do 3° dia pós-colheita (aproximadamente 6%),
sendo também praticamente todo degradado na manga madura. O seu perfil é
semelhante ao controle, porém ao contrário do que ocorreu o amido foi acumulado e
se iniciou a degradação praticamente 2 dias antes do controle.
0 2 4 6 8 10 120
1
2
3
4
5
6
7
8
Teor
de
amid
o (%
)
dias pós-colheita
Figura 13. Evolução dos teores percentuais de amido em amostras controle (sem
tratamento 1-MCP - ) e tratadas com 1-MCP ( ) durante o amadurecimento da manga
Keitt.
O fato observado pode ser devido ao período da colheita, ou seja, os frutos
amostrados poderiam estar em diferentes estágios do amadurecimento, não

perceptíveis no momento da colheita, visto que esta é determinada basicamente em
função de parâmetros visuais. Outra possibilidade diz respeito à quantidade de 1-
MCP aplicada no tratamento, que não seria suficiente para atrasar a degradação do
amido, embora tenha sido para outros metabolismos, como a síntese de etileno
citada acima. Isto indicaria uma suposta menor sensibilidade do metabolismo do
amido ao bloqueio pelo 1-MCP, o que encontra suporte na literatura, na qual
algumas respostas ao etileno são mais ou menos sensíveis a inibição determinada
por este antagonista (BLANKENSHIP; DOLE, 2005). Também não é possível
descartar a hipótese de que o bloqueio da sinalização do etileno pelo 1-MCP tenha
induzido a aceleração na degradação do amido, ao contrário das observações
encontradas na literatura referente a frutos tratados com 1-MCP (BLANKENSHIP;
DOLE, 2005).
Analisando estes resultados em paralelo às curvas de produção de etileno e
respiração (Figura 12), observa-se que o aumento na produção do hormônio e o
climatério iniciam após o início da degradação do amido, indicando uma possível
independência da degradação do amido em manga Keitt em relação à sinalização
do etileno. Essa hipótese poderia ser refutada se forem considerados os ligeiros
aumentos na produção do etileno por volta do 4º e 5º dias pós-colheita, em ambos
os grupos de frutos, como suficientes para disparar a mobilização do amido. Esta
hipótese pressupõe uma grande sensibilidade deste metabolismo em relação à
sinalização mediada pelo etileno, o que pode ser avaliado por alterações na
atividade de enzimas correlacionadas à degradação do amido.

4.5. Açúcares Solúveis
A análise dos açúcares solúveis (Figura 14) mostrou que durante o
amadurecimento da manga, após o tratamento com 1-MCP, houve acúmulo de
glicose, frutose e sacarose tanto nas amostras controle como nos frutos tratados,
porém os perfis observados foram diferentes. No controle os teores de glicose e
frutose flutuaram ligeiramente durante todo o amadurecimento e o teor de sacarose
aumentou ao fim do experimento, tendo seu valor máximo por volta do 13° dia pós-
colheita, sendo o principal açúcar solúvel no fruto maduro. Resultados que
confirmam os resultados obtidos por Bernardes-Silva (2004).
Este perfil também foi encontrado no tratamento com 1-MCP. No entanto o
resultado mais expressivo pode ser observado quanto ao aumento de sacarose, que
atingiu valores duas vezes maiores do que os encontrados nos frutos controle, e
também iniciaram o acúmulo com 3 dias de atraso.
0 2 4 6 8 10 120
15
30
45
60
75
90
105
120
mg.
g -1
fruto
dias pós-colheita0 2 4 6 8 10 12 14 16 18 20
0
15
30
45
60
75
90
105
120
mg.
g -1
frut
o
dias pós-colheita
Figura 14. Evolução dos teores dos carboidratos em amostras controle (sem tratamento 1-
MCP - frutose, sacarose, glicose) e tratadas com 1-MCP ( frutose, sacarose,
glicose) durante o amadurecimento da manga Keitt.

Esta observação contrasta com os dados obtidos na análise dos teores de
amido visto que tanto nas amostras de mangas Keitt obtidas por Bernardes-Silva e
Cordenunsi (2004) quanto em seu trabalho anterior com outras cultivares de mangas
(BERNARDES-SILVA; CORDENUNSI, 2000), ocorre o aumento concomitante da
sacarose a partir da mobilização do amido.
Visto que os dados apresentados na Figura 13 indicam que, no mínimo, o
amido tanto das amostras controle quanto das amostras tratadas com 1-MCP havia
praticamente sido todo degradado ao 13º dia pós-colheita, o resultado na análise
dos açúcares, particularmente da sacarose, indica alguma alteração no metabolismo
sintético desta última, ou alguma alteração no fluxo de carbono para a sua via de
biossíntese.
Os aumentos na síntese de sacarose, contudo, parecem estar
correlacionados temporalmente ao aumento na produção de etileno e ao aumento
respiratório (Figura 12).

4.6. Atividade de α-amilase e de β-amilase
Para melhor caracterizar a hipótese de uma alteração na via de degradação
do amido mediada pelo 1-MCP foram medidas as atividades de α-amilase e de β-
amilase na polpa dos frutos. Os resultados (Tabela 1), embora com diferenças
estatisticamente significativas na atividade de ambas as enzimas dos dois grupos,
no 2º e 4º dias pós-colheita, não parecem indicar efeitos muito significativos do 1-
MCP sobre o metabolismo do amido. De certo modo, o resultado indica até uma
menor atividade da β-amilase no 2º dia pós-colheita nas amostras tratadas com 1-
MCP, apesar da degradação do amido ter início dois dias antes em relação às
amostras controle (Figura 13).
A observação sugere que as atividades de α e β-amilase sofreram pouca
influência do 1-MCP, seja pelo nível de antagonista empregado no tratamento ou por
uma possível independência de sua regulação em relação ao etileno, não sofrendo,
portanto, nenhum efeito com o bloqueio da sinalização do hormônio.
Assim sendo, a hipótese da mobilização do amido responder a pequenas
variações na produção de etileno, não parece ser sustentada, pelo menos em parte,
dada a aparente.
Tabela 1. Valores da atividade de α-amilase e de β-amilase durante o amadurecimento de
mangas Keitt controle (sem tratamento 1-MCP) e tratadas com 1-MCP.
Amostras Controle 1-MCP Dias Pós-Colheita Alfa amilase* Beta amilase* Alfa amilase* Beta amilase*
0 5,37 ± 0,76 1,61 ± 0,47 5,37 ± 0,76 1,61 ± 0,47
2 8,25 ± 0,08 3,21 ± 0,05 (**) 7,23 ± 0,51 2,04 ± 0,14 (**)
4 7,93 ± 0,45 2,61 ± 0,15 (**) 7,84 ± 0,38 1,87 ± 0,10 (**) * µmol p-nitrofenol.g-1.h-1.mg-1.proteína.
** indica diferença significativa (p<0,05)

Outras enzimas que foram analisadas quanto aos possíveis efeitos do 1-MCP
foram as amido fosforilases e a DPE2, por meio de ensaio de atividade em gel de
acrilamida após eletroforese em condições não dissociantes.
O resultado (Figura 15) indica que houve efeito do 1-MCP sobre a amido
fosforilase citosólica, em cujo grupo foi observada a maior atividade desta forma da
enzima nas amostras coletadas no 2º dia pós-colheita. O mesmo nível de atividade
nas amostras controle foi atingido apenas no 4º dia pós-colheita.
Figura 15. Atividade de amido fosforilases citosólica e plastidial em gel de poliacrilamida
contendo glicogênio 1% corado com I2/KI em amostras controle (sem tratamento 1-MCP) e
tratadas com 1-MCP de manga Keitt.
Esta observação parece ir de encontro aos resultados relacionados à
degradação do amido nas amostras tratadas com 1-MCP (Figura 13). O estímulo na
atividade da forma citosólica da amido fosforilase também foi reportado por Mainardi
et al. (2006) no tratamento pós-colheita de bananas com 1-MCP, na concentração
de 100 nL.L-1 por 12 horas. Os dados sugerem o efeito semelhante do antagonista
em mangas, porém com intensidade relativamente menor, visto que em bananas o

1-MCP induz efeitos muito mais pronunciados na atividade hidrolítica (Nascimento et
al. 2006) e fosforolítica (MAINARDI et al., 2006) na mobilização do amido.
As análises de atividade da DPE2 não foram efetuadas com sucesso, apesar
de várias tentativas, apontando a necessidade de uma melhor otimização do ensaio
com proteínas extraídas de polpa de mangas. Este método foi utilizado com sucesso
por Mainardi (2007) para avaliação da atividade da DPE2 em polpa de bananas.
No entanto, uma maneira de avaliar as possíveis alterações ocorridas no
metabolismo amido-sacarose em função do tratamento com 1-MCP, foi medir os
níveis de maltose nos frutos controle e nos frutos tratados com o antagonista de
etileno, uma vez que a atividade da β-amilase parece ter sido pouco afetada pelo 1-
MCP. Um possível fator a contribuir para o atraso na síntese de sacarose decorrente
ao efeito sobre a metabolização da maltose.
Os resultados (Figura 16) indicam um perfil nas amostras controle de
decréscimo no nível do açúcar até o 7º dia pós-colheita seguido de uma retomada
na síntese e crescente aumento até o fim do experimento (13º dia pós-colheita)
atingindo valores pouco superiores aos da amostra inicial (1º dia pós-colheita). Nas
amostras tratadas com 1-MCP, contudo, o perfil indica a manutenção dos níveis de
maltose entre 8-10 mg de maltose por grama de polpa fresca (0,8-1,0% em base
fresca) até o 16º dia pós-colheita, após o qual declinou, até o 20º dia pós-colheita,
ao nível similar ao encontrado nos frutos controle no 7º dia pós-colheita.

0 2 4 6 8 10 120
2
4
6
8
10
12
14
16
18m
g. g
-1 fr
uto
dias pós-colheita
0 2 4 6 8 10 12 14 16 18 200
2
4
6
8
10
12
14
16
18
mg.
g-1 fr
uto
dias pós-colheita
Figura 16. Evolução dos teores de maltose em amostras controle (sem tratamento 1-MCP -
) e tratadas com 1-MCP ( maltose) durante o amadurecimento de mangas Keitt.
O resultado demonstra claramente uma alteração no metabolismo da maltose
que pode estar correlacionado ao atraso no aumento de sacarose (Figura 14), visto
que a maltose, como produto do metabolismo do amido, em função da atividade da
β-amilase, serviria de substrato para a biossíntese da sacarose. No entanto para que
isso ocorra, é necessária a clivagem da maltose, seja pela ação de uma DPE2 ou
pela ação de uma α-1,4-glicosidase com atividade maltásica. Neste ponto, é
possível inferir que o fluxo de carbono vindo do amido para a sacarose pode ter
como ponto sensível, etileno-dependente, a metabolização da maltose proveniente
do plastídeo. A medida de atividade ou expressão gênica de enzimas relacionadas à
maltose pode trazer novas informações acerca destas considerações.

5. Considerações Finais
A degradação do amido em mangas Keitt ocorre, em grande parte, devido a
ação de várias enzimas também descritas em bananas sugerindo uma similaridade
nos modelos e, por extensão, um mecanismo conservado em frutos que acumulam
amido. As diferenças do processo entre os frutos residem, aparentemente, nos
níveis de atividade e transcrição das enzimas.
As variações nos níveis de AIA e ABA sugerem um possível papel destes
hormônios na sinalização para a mobilização do amido dada a correlação temporal.
A alta atividade de α-amilase em polpa de mangas Keitt parece ser
conseqüência, ao menos em parte, dos altos níveis de transcrição do mRNA da
enzima, sugerindo participação bastante relevante da α-amilase na degradação pós-
colheita do amido de mangas Keitt.
O tratamento com 1-MCP é capaz de estimular a atividade da forma citosólica
da amido fosforilase, sugerindo uma possível regulação negativa por níveis basais
do etileno, visto que a atividade da enzima aumentou logo após o tratamento com 1-
MCP, este aumento não foi detectado nos frutos controle.
A biossíntese de sacarose foi atrasada por conta do tratamento das mangas
Keitt com 1-MCP. Apesar das amostras controle neste experimento não
apresentarem aumento concomitante, os resultados sugerem uma redução na
velocidade do fluxo de carbono do amido para a síntese da sacarose e indica a
influência do etileno no processo.

6. CONCLUSÕES
A mobilização do amido em mangas Keitt parece, ao menos em parte, estar
dissociada temporalmente da evolução nos níveis de etileno, indicando ser
parcialmente independente da sinalização deste hormônio. As atividades de α e β-
amilase reforçam esta hipótese, dado sofrerem pouca influência do 1-MCP, e serem
pouco inibidas por este antagonista.
O atraso na síntese de sacarose pode ser, ao menos parcialmente, resultado
de alterações no metabolismo da maltose proveniente da degradação do amido,
dada a sua inibição nas amostras de mangas Keitt tratadas com 1-MCP ou devido às
alterações no metabolismo da sacarose, processos que parecem ser etileno-
dependente.

7. REFERÊNCIAS BIBLIOGRÁFICAS
ARORA, A. Ethylene receptors and molecular mechanism of ethylene sensitivity in
plant. Curr. Science. v. 89, n.8, pp. 1348-1361, 2005.
ÂREAS, J.A.G.; LAJOLO, F.M. Starch transformation during banana ripening I: the
phosphorylase and phosphatase behavior in Musa acuminata. J. Food Bioch. v.5,
pp. 19 –37, 1981.
BHATIA, S.; SINGH, R. Phytohormone-mediated transformation of sugars to starch
in relation to the activities of amylases, sucrose-metabolising enzymes in sorghum
grain. Plant Growth Regulation. v. 36, pp. 97-104, 2002.
BERGMEYER, H.U. Methods of enzymatic analysis. 2ed. New York: Academic
Press, 1974, v.3, pp. 1212 – 1215.
BERNARDES-SILVA, A.P.F.; LAJOLO, F.M.; CORDENUNSI, B.R. Evolução dos
teores de amido e açúcares durante o desenvolvimento e amadurecimento de
diferentes cultivares de manga. Ciênc. Tecnol. Aliment. v. 23, pp. 116-120, 2003.
BERNARDES-SILVA A.P.F. Caracterização Química e Enzimática do Processo
de Adoçamento da Manga ‘keitt’, 2004. Tese de Doutorado - Faculdade de
Ciências Farmacêuticas, Universidade de São Paulo, São Paulo, 2004.

BETHKE, P. C.; LONSDALE, J.E.; FATH, A.; JONES; R.L. Hormonally Regulated
Programmed Cell Death in Barley Aleurone Cells. The Plant Cell. v. 11, pp. 1033-
1045, 1999.
BIERHALS, J.; LAJOLO, F.M.; CORDENUNSI, B.R.; NASCIMENTO, R.O. Activity,
Cloning, and Expression of an Isoamylase-Type Starch-debranching Enzyme from
Banana fruit. J. Agric. Food Chem. v. 52, pp. 7412-7416, 2004.
BLANKENSHIP, S.M.; DOLE, J.M. 1-methylcyclopropene: a review. Postharvest
Biology and Techology. v. 28, pp. 1 - 25, 2003.
BRADFORD, M.M.; Rapid and sensitive method for quantification of microgram
quantities of protein utilizing of protein-dye binding. Analy. Bioch. v.72, n.1-2, pp.
248 - 254, 1976.
CHANDLER, J.W.; APEL, K.; MELZER, S. A novel putative β-amylase gene and
ATβ-Amy from Arabidopsis thaliana are circadian regulated. Plant Science. v. 161,
pp. 1019-1024, 2001.
CHEN, K.H.; MILLER, A.N.; PATTERSON, G.W.; COHEN, J.D. A rapid and simple
procedure for purification of indole-3-acetic acid to GC-MS-SIM analysis. Plant
Physiol., Rockville, v.86, n.3, pp. 822 – 825, 1988.
DINGES, J.; COLLEONI, C.; JAMES, M.G.; MYERS, A.M. Mutational Analysis of the
Pullulanase-Type Debranching Enzyme of Maize Indicates Multiple Fuctions in
Starch Metabolism. The Plant Cell. v. 15, pp. 666-680, 2003.

FRENKEL, C.; DYCK, R. Auxin Inhibition of Ripening in Bartlett Pears. Plant
Physiol. v. 51, pp. 6-9, 1973.
FETTKE, J.; ECKERMANN, N.; POESTE, S.; PAULY, M.; STEUP, M. The glycan
substrate of the cytosolic (Pho 2) phosphorylase isozyme from Pisum sativum L.:
identification, linkage analysis and subcellular localization. The Plant Journal. v. 39.
pp. 933 - 946, 2004.
FETTKE, J.; ECKERMANN, N.; TIESSEN, A.; GEIGENBERGER, P.; STEUP, M.
Identification, subcellular localization and biochemical characterization of water-
soluble heteroglycans (SHG) in leaves of Arabidopsis thaliana L.: distinct SHG reside
in the cytosol and in the apoplast. Plant J. v.43, pp. 568 – 586, 2005a.
FETTKE, J.; POESTE, S.; ECKERMANN, N.; TIESSEN, A.; PAULY, M.;
GEIGENBERGER, P.; STEUP, M. Analysis of cytosolic heteroglicans from leaves of
transgenic potato (Solanum tuberosum L.) plants that under-or overexpressed the
Pho2 phosphorylase isozyme. Plant Cell Physiol. v.46, pp. 1984 - 2004, 2005b.
FUCHS, Y.; PESIS, EDNA; ZAUBERMAN, GIORA. Changes in amylase activity,
starch and sugars contents in mango fruit pulp. Scientia horticulturae. v. 13, pp.
155-160, 1980.
GALLANT, D.J.; BOUCHET, B.; BALDWIN, P.M. Microscopy of starch: evidence of a
new level of granule organization. Carbohydrate Polymers. v.32, pp. 177 – 191,
1997.

GERTMAN, E.; FUCHS, Y. Effect of Acid and Its Interections with Other Plant
Hormones on Ethylene Production in Two Plant Systems. Plant Physiol. v.50,
pp.194-195, 1972.
GUO, H.; ECKER, J.R. The ethylene signaling pathway: new insights. Curr. Op. in
Plant Biol.. V. 7, pp. 40-49, 2003.
HUBARDD, N.L.; PHARR, D.M.; HUBER, S.C. Role of Sucrose Phosphate Synthase
in Sucrose Biosynthesis in Ripening Bananas and Its Relationship to the Respiratory
Climacteric. Plant Physiol. v. 94, n. 201-208, 1990.
HUSSAIN, H.; MANT, A.; SEALE, R.; ZEEMAN, S.; HINCHLIFE, E.; EDWARS, A.;
HYLTON, C.; BORNEMANN, S.; SMITH, A.M.; MARTIN, C.; BUSTOS, R. Three
Isoforms of Isoamylase Contribute Different Catalytic Properties for the Debranching
of Potato Glucans. The Plant Cell. v.15, pp.133 - 149, 2002.
JIANG, Y.; JOYCE, D.C.; MARCNISH, A.J. Effect of Abscisic Acid on Banana Fruit
Ripening in Relation to the Role of Ethylene. J. Plant Growth Regulation. v. 19, pp.
106-111, 2000.
KUBO, A.; FUJITA, N.; HARADA, K.; MATSUDA, T.; SATOH, H.; NAKAMURA, Y.
The Starch-Debranching Enzimes Isoamylase and Pullulanase Are Both Involved in
Amylopectin Biosynthesis in Rice Endosperm. Plant Physiol. v. 121, pp. 399-409,
1999.

LACERDA, M.A.D.; LACERDA, R.D. O Cluster da fruticultura no Pólo Petrolina/
Juazeiro. Revista de Biologia e Ciências da Terra. V.4, n.1, 2004.
LARA, I.; VENDRELL, M. Relationships between ethylene, abscisic acid and quality
during postharvest storage of ‘Granny Smith’ apple. Posthrvest Biology and
Technology. v. 13, pp. 11-18, 1998.
LARA, I.; VENDRELL, M. Cold-induced ethylene biosynthesis is differentially
regulated in peel and pulp tissues of ‘Granny Smith’ apple fruit. Postharvest Biology
and Technology. v.29, pp. 109 -119, 2003.
LELIÈVRE, J.M.; LATCHÈ, A.; JONES, B.; BOUZAYEN, M.; PECH, J.C. Ethylene
and fruit ripening. Physiologia Plantarum, v. 101, pp. 727-739, 1997.
LIZADA, C. Mango. In: SEYMOUR, G.B.; TAYLOR, J.E; TUCKER, G.A.
Biochemistry of Fruit Ripening. London, Chapman & Hall, 1993. Cap. 8, pp. 255-
266.
LIU, X.; SHIOMI, S.; NAKATSUKA, A.; KUBO, Y.; NAKAMURA, R.; INABA, A.
Characterization of Ethylene Biosynthesis Associeted with Ripening in Banana fruit.
Plant Physiol. v. 121, pp. 1257-1265, 1999.
LLOYD, J.; KOSSMANN, J.; RITTE, G. Leaf starch degradation comes out of the
shadows. Trends in Plant Science. v. 10, n. 3, pp.130-138, 2005.

LOHANI, S.; TRIVEDI, P.K.; NATH, P. Changes in activities of cell wall hydrolases
during ethylene-induced ripening in banana: effect of 1-MPC, ABA and IAA.
Posthervest Biology and Technology. v. 31, pp. 119-126, 2004.
LÓPEZ-GÓMES, R.; GÓMES-LIM, M.A. A Method for Extracting Intact RNA from
Fruits Rich in Polysaccharides using Ripe Mango Mesocarp. HortScience. v. 27, n.
5, pp. 440-442, 1992.
LU, Y.; SHARKEY, T.D. The importance of maltose in transitory starch breakdown.
Plant and Cell Envionment. v. 29, pp. 353-366, 2006.
MAcLEOD, A.J.; PIERIS, N.M. Comparison of the volatile components of some
mango cultivars. Phytoch. v. 23, n. 2, pp. 361 - 366, 1984.
MAINARDI, J. A.; PURGATTO, E.; VIEIRA JR, A.; BASTOS, W.A.; CORDENUNSI,
B.R.; NASCIMENTO, R.O.; LAJOLO, F.M. Effects of Ethylene and 1-
Methylcyclopropene (1-MCP) on Gene Expression and Activity Profile of a α-1,4-
glucan-phosphorylase during Banana Ripening. J. Agric. Food Chem. v. 54, pp.
7294-7299, 2006.
MAO, W.W.; KINSELLA, J.E. Amylase Activity in Banna Fruit: Properties and
Changes in Activity with Ripening. Journal of Food Science. v. 36, pp. 1400-1409,
1981.

MAO, L.; WANG, G.; QUE, F. Application of 1-methylcyclopropene prior to cutting
reduces wound responses and maintains quality in cut kiwifruit. J. of Food
Engineering. V.78, pp.361 – 365, 2007.
MEDINA-SUAREZ, R.; MANNING, K.; FLETCHER, J.; AKED, J.; BIRD, C.R.;
SEYMOUR, G. Gene Expression in the Pulp of Ripening Bananas. Plant Physiol. v.
115, pp. 453-461, 1997.
MITA, S.; SUZUKI-FUJII, S.; NAKAMURA, K. Sugar-Inducible Expression pf a Gene
for β-Amylase in Arabidopsis thaliana. Plant Physiol. v. 107, pp. 895-904, 1995.
MOTA, R. V.; CORDENUNSI, B.R.; NASCIMENTO, J.O.; PURGATTO, E.;
ROSSETO, M.R.; LAJOLO, F.M. Activity and expression of banana starch
phosphorylases during fruit development and ripening. Planta. v. 216, pp. 325-333,
2002.
MYERS, A.M.; MORELL, M.K.; JAMES, M.G.; BALL, S.G. Recent progress toward
understanding biosynthesis of the amylopectin crystal. Plant Physiol. v.122, pp. 989 –
997, 2000.
NASCIMENTO, J.R.O.; CORDENUNSI, B.R.; LAJOLO, F.M.; ALCOCER, M.J.C.
Banana sucrose-phosphate synthase gene expression during fruit ripening. Planta.
v.203, pp. 283 – 288, 1998.

NASCIMENTO, J.R.O.; VIEIRA JR, A.; BASSINELLO, P.Z.; CORDENUNSI, B.R.;
MAINARDI, J.A.; PURGATTO, E.; LAJOLO, F.M. Beta-amylase expression and
starch degradation during banana ripening. Postharvest Biology and Technology.
v.40, pp. 41 - 47, 2006.
ÖSTIM, A.; KOWALYCZK, M.; BHALERAO, R.P.; SANDBERG, G. Metanolism of
Índole-3-Acetic Acid in Arabidopsis. Plant Physiol. v. 118, pp. 285-296, 1998.
PEROSA, J.M.; PIERRE, F.C. Técnicas de Pós-colheita e expansão da cultura da
manga no Estado de São Paulo. Rev. Bras. Frutic. Jaboticabal. v. 24, n. 2, pp.381-
384, 2006.
PHILLIPS, L.A.; BARRY, C.; GIOVANNONI, J. Signal transduction systems
regulating fruit ripening. Trend in Plant Science. v. 9, n. 7, pp. 221-338, 2004.
PURGATTO, E.; LAJOLO, F.M.; NASCIMENTO, R.O.; CORDENUNSI, B.R.
Inhibition of β-amylase activity, starch degradation and sucrose formation by indole-
3-acetic acid during banana rinpenig. Planta. v. 212, pp. 823-828, 2001.
PURGATTO, E.; NASCIMENTO, J.R.O.; LAJOLO, F.M.; CORDENUNSI, B.R. The
onset of starch degradation during banana ripening is concomitant to changes in the
content of free and conjugated forms of indole-3-acetic acid. Plant Physiol. v. 159,
pp. 1105-1111, 2002.

SAMBROOK J, FRITSCH EF, MANIATIS T. Molecular Cloning: A Laboratory
Manual, 2nd edn. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York
USA, 1989.
SANGER F, NICKLEN S, COULSON R. DNA sequencing with chain terminating
inhibitors. Proc. Natl. Acad. Sci. USA. v.74, p.5463-5467, 1977.
SATYAN, S.H.; CHAPLIN, G.R.; WILLCOX, M.E. An assessment of fruit quality of
various mango cultivars. Proc. Of the First Aus. Mango Research Workshop.
Cairn, Queensland, pp. 324 - 333, 1986.
SCHUPP, N.; ZIEGLER, P. The Relation of Starch Phosphorylases to Starch
metabolism in Wheat. Plant Cell Physiol. v. 45, n.10, pp. 1471-1484, 2004.
SETHA, S.; KONDO, S.; HIRAI, N.; OHIGASHI, H. Xanthoxin, abscisic acid and its
metabolite levels associated with apple fruit development. Plant Science. v. 166, pp.
493-499, 2004.
SKADSEN, R. Physiological and molecular genetic mechanisms regulating hydrolytic
enzyme gene expression in cereal grains. Physiologia Plantarum. v. 104, pp. 486-
502, 1998.

SMITH, S.M.; FULTON, D.C.; CHIA, T.; THORNEYCROFT, D.; CHAPPLE, A.;
DUSTANN, H.; HYLTON, C.; ZEEMAN, S.C.; SMITH, A.M. Diurnal Changes in the
Transcriptome Encoding Enzymes of Starch metabolism Provide Evidence for Both
Transcriptional and Posttranscriptional Regulation of Starch metabolism in
Arabidopsis leaves. Plant Physiol. v. 136, pp. 2687-2699, 2004.
SMITH, A.; ZEEMAN, S.C.; SMITH, S.M. Starch Degradation. Annu. Rev. plant.
Biol. v. 56, pp. 73-98, 2005.
TANDON, D.K.; KALRA, S.K. Changes in sugars, starch, and amylases activity
during development of mango fruit cv Dashehari. J. of Horticultural Science. v. 58,
n.3, pp. 449-453, 1983.
TANEYAMA, M.; YAMAUCHI, D.; MINAMIKAWA, T. Synthesis and Turnover of α-
amylase in Cotyledons of Germinating Vigna mungo Seeds: Effects of Exogenously
Applied End-Products and Plant Hormones. Plant Cell Physiol. v. 36, n. 1, pp. 139-
146, 1995.
TETLOW, I.J.; MORELL, M. K.; EMES, M.J. Recent developments in understanding
the regulation of starch metabolism in higher plants. J. of Exp. Botany. v. 55, n. 406,
pp. 2131-2145, 2004.
THARANATHAN, R.N.; YASHODA, H.M.; PRABHA, T.N. Mango (Mangifera indica
L.), “The King of Fruits” – An Overview. Food Rev. International. V. 22, pp. 95-123,
2006.

THEOLOGIS, A. One Rotten Apple Spoils the Whole Bushel: The Role of Ethylene in
Fruit Ripening. Cell. v. 70, pp. 181-184, 1992.
THEOLOGIS, A. Plant hormones: More than one way to detect ethylene. Cur.
Biology. v.6. n. 2, pp. 144 – 145, 1996.
TINGWA, P. O.; YOUNG, R.E. The Effect of Indole-3-acetic Acid and Other Growth
Regulatiors on the Ripening of Avacado Fruits. Plant Physiol. v. 55, pp. 937-940,
1975.
VIEIRA JR.; NASCIMENTO, J.R.O.; LAJOLO, F.M. Molecular Cloning and
Characterization of an r-Amylase Occuring in the Pulp of Ripening Bananas and Its
Expressionn Pichia pastoris. J. Agric. Food Chem. v. 54, pp.8222-8228, 2006.
WANG, Y.Z.; ZHANG, D.P. Activities, quantitative changes and subcellular
localization of α-amylase during development of apple fruit. Acta Bot. Sinica. V.44,
n.1, pp. 34 – 41, 2003.
WEGRZYN, T.; MAcRAE, E. Alpha-amylase and starch degradation in kiwifruit. J. of
Plant Physiology. v. 147, pp. 19 - 28 , 1995.
WEGRZYN, T.; REILLY K.; CIPRIANI, G.; MURPHY, P.; NEWCOMB, R.;
GARDNER, R.; MAcRAE, E. A novel α-amylase genes is transiently upregulated
during low temperature exposure in apple fruit. Eur. J. Biochem. v. 267, pp. 1313-
1322, 2000.

WHILE, P. Recent advances in fruit development and ripening: an overview. J. of
Exp. Botany. v. 53, n. 377, pp. 1995-2000, 2002.
YANG, S.F.; HOFFMANN, N.E. Ethylene biosynthesis and its regulation in higher
plants. An. Review Plant Physiology. v. 35, p. 155-189, 1984.
ZEEMAN, S. C.; SMITH, S.M.; SMITH, A.M. The breakdown of starch in leaves. New
Physiologist. v. 163, pp. 247-261, 2004.
BERNARDES-SILVA, A.P.F.; NASCIMENTO, J.R.O.; LAJOLO,F.M.; CORDENUNSI,
B.R. Starch mobilization and sucrose accumulation in the pulp of ‘Keitt’ mangoes
during postharvest ripening. J. of Food Bioch., 2007 (aceito para publicação).

Variação hormonal correlacionada à expressão de enzimas ligadas ao metabolismo do amido durante o desenvolvimento e amadurecimento da manga (Mangifera indica cv Keitt) Claudia Mitsue Koike (Mestrado); Eduardo Purgatto (Orientador). [email protected].
Departamento de Alimentos e Nutrição Experimental, FCF/USP.
O desenvolvimento e o amadurecimento de frutos são processos complexos e
organizados, onde fitormônios, como o ácido indol-3-acético (AIA), o ácido abscísico
(ABA) e o etileno desempenham papel relevante. Dentre os processos
característicos do amadurecimento, a degradação do amido apresenta impacto
direto sobre a qualidade da manga, sendo praticamente todo degradado a síntese
de sacarose. Várias enzimas participam da degradação do grânulo de amido, como
as hidrolases (α e β-amilases, enzimas desramificadoras) e as fosforilases, descritas
também em bananas. A mobilização do amido em mangas Keitt parece estar
dissociada temporalmente da evolução do etileno; e as variações nos níveis de AIA
e ABA sugerem um possível papel na sinalização para mobilização do amido. A
biossíntese da sacarose foi atrasada pelo 1-MCP, um antagonista do etileno,
sugerindo redução na velocidade do fluxo de carbono do amido para síntese da
sacarose que pode ser, resultante das alterações no metabolismo da maltose
proveniente da degradação do amido.
Palavras Chaves: Manga. Mangifera indica. Fitormônios. Amido. β-amilase.

Correlation with hormonal variation and genes expression of enzimes involved in starch metabolic during during development and ripening of mango
(Mangifera indica cv Keitt) Claudia Mitsue Koike (Mestrado); Eduardo Purgatto (Orientador). [email protected].
Lab. de Química, Bioquímica e Biol. Molecular de Alimentos, Depto de Alimentos e
Nutrição Experimental - FCF/USP – SP – Brasil.
The fruit development and ripening are organized and complex processes
where phytohormones, like indol-3-acetic acid (IAA), abscisic acid (ABA) and
ethylene have a pivotal role. In mango fruit one of the main metabolisms that have
direct impact in the mango quality is the starch degradation, practically being all
degraded the synthesis of sacarose. Some enzymes participate of the degradation,
as hidrolases (α and β-amylases, disproportionating enzymes) and phosphorylases,
described also in bananas. The mobilization of the starch in Keitt mangoes it does
not seem to be temporally associated with ethylene; and variations in the IAA and
ABA levels suggest their possible role in the signalling for the starch. Sucrose
biosynthesis was delayed by 1-MCP indicating a reduction in the speed of the carbon
flux from the starch that it can be, resultant of the alterations in the metabolism of
maltose provenient from the starch degradation.
Key Words: Mango. Mangifera indica. Phytohormones. Starch. β-amylase.