
Varredura de uma biblioteca de cDNA da Lasiodora sp ...
126
Universidade Federal de Ouro Preto Núcleo de Pesquisa em Ciências Biológicas Programa de Pós-Graduação em Ciências Biológicas Varredura de uma biblioteca de cDNA da aranha Lasiodora sp utilizando-se de antisoro policlonal anti-veneno total e clonagem das seqüências codificantes para LTx1 e LTx3 no vetor de expressão bacteriano pET-11a André Luiz Gomes Vieira Orientador: Dr. Ieso de Miranda Castro Co-Orientador: Evanguedes Kalapothakis Ouro Preto Março 2005 Dissertação apresentada ao programa de pós- graduação do Núcleo de Pesquisa em Ciências Biológicas da Universidade Federal de Ouro Preto, como parte integrante dos requisitos para a obtenção do título de mestre em ciências biológicas. Área de concentração: Biologia Molecular.
Transcript of Varredura de uma biblioteca de cDNA da Lasiodora sp ...

Universidade Federal de Ouro Preto Núcleo de Pesquisa em Ciências Biológicas
Programa de Pós-Graduação em Ciências Biológicas
Varredura de uma biblioteca de cDNA da aranha Lasiodora sp utilizando-se de antisoro policlonal anti-veneno total e clonagem das seqüências codificantes para LTx1 e LTx3 no vetor de expressão bacteriano pET-11a
André Luiz Gomes Vieira
Orientador: Dr. Ieso de Miranda Castro Co-Orientador: Evanguedes Kalapothakis
Ouro Preto Março 2005
Dissertação apresentada ao programa de pós-graduação do Núcleo de Pesquisa em Ciências Biológicas da Universidade Federal de Ouro Preto, como parte integrante dos requisitos para a obtenção do título de mestre em ciências biológicas. Área de concentração: Biologia Molecular.

Este trabalho tem sido desenvolvido com o apoio financeiro das seguintes instituições:
• Fundação de amparo à Pesquisa do Estado de Minas Gerais – FAPEMIG • Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq • Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – CAPES • Universidade Federal de Ouro Preto - UFOP

II
“Do trabalho das tuas mãos comerás, feliz serás e tudo te irá bem.”
Salmos 128:2
À
Ana Luíza
Vitória
Daniel

III
Agradecimentos
• A Deus, pela fidelidade inabalável e pela certeza de que, mesmo quando estou dormindo, o melhor tem sido planejado para mim.
• Aos meus pais (José Célio e Elza Gomes), meus irmãos (Helen, Jaqueline, Neemias
e Tiago), meus sobrinhos (Ana Luiza, Vitória e Daniel) e meus cunhados (Ronald, Patrícia e Glauciene), razão de tudo isso. Amo vocês!
• Ao professor Dr. Ieso de Miranda Castro, pelo exemplo de seriedade e compromisso
com a pesquisa. Muito obrigado pela orientação e oportunidade de realização deste trabalho.
• Ao co-orientador deste trabalho, professor Dr. Evanguedes Kalapothakis pela
indispensável participação na construção da biblioteca de cDNA e discussão dos resultados obtidos.
• Ao professor Dr. Rogelio Lopes Brandão pelo empenho em fazer do Programa de
Pós-graduação em Ciência Biológicas da Universidade Federal de Ouro Preto um programa de destaque.
• Ao professor Dr. Helio Hideo Babá pela amizade e pelas dúvidas científicas
esclarecidas.
• Aos professores Dr. Luciano Fietto e Dra. Juliana Fietto, pela amizade e pela boa vontade e prontidão com que compartilham seus conhecimentos científicos. Muito Obrigado por tudo!
• À minha colega de graduação que se tornou amiga e que se tornou irmã, Michelle
Bueno de Moura Pereira. Já estou com uma saudade!
• À Cida, como é difícil expressar a gratidão por toda sua simpatia e competência. Ninguém que passa por aqui te esquece!
• À Maria José Trópia (Zezé), simpatia do LBCM.
• Aos colegas de trabalho do LBCM: Maristela, Jú Boechat, Kátia, Pilar (saudades),
Michele Barbi (saudades), Tiago, Fernandinha, Raquel, Giovana, Matheus, Fred (Saudades), Ana Paula (saudades), Mr. Totola, Val, Lis, Aninha, Xisto, Murilo, Mônica, Lucas, Dani Mel, Ramon, Patrick e Sr. Brás. Não temos rádio, mas damos boas risadas...rs...
• À Milene, pela ajuda fundamental na realização deste trabalho e por me aturar nos
momentos difíceis.

IV
• À amiga Danizinha, pelo empurrão científico. Obrigado.
• Às amigas Inês e Mariinha pelo referencial de família que se tornaram para mim em
Ouro Preto.
• Aos amigos da república Brejo, Paulo Schermack, Vinícius Hermani e Wagner Couto pelo convívio e pelas risadas.
• Aos meus tios e primos que torceram e não se esquecem de mim.
• A todos que, direta ou indiretamente, contribuíram para a realização deste trabalho.

V
Resumo
Venenos de aranhas são sistemas de multicomponentes que podem ser agrupados em 3
classes químicas principais: moléculas orgânicas de baixo peso molecular, peptídeos e
proteínas de alto peso molecular. A grande maioria de toxinas de aranhas identificadas até o
momento é composta de polipeptídeos com uma massa molecular de 3000-8000 Da
altamente reticulados por pontes dissulfeto. A purificação de peptídeos tóxicos de venenos
de aranhas nos propicia um arsenal de ferramentas moleculares para estudos
eletrofisiológicos, farmacológicos e estudo de canais iônicos. Este trabalho descreve o
resultado da varredura de uma biblioteca de cDNA, utilizando-se da técnica de ELISA, com
o objetivo de identificar proteínas imunogênicas. A biblioteca de cDNA foi construída a
partir do mRNA isolado de glândulas de veneno de Lasiodora sp. Anti-soro contra o
veneno total foi usado na varredura. Os clones positivos foram caracterizados por
seqüenciamento de DNA. Nossos resultados mostram que os precursores das toxinas
descritas apresentam um peptídeo sinal caracterizado por um alto conteúdo de aminoácidos
hidrofóbicos, um propeptídeo interposto entre a seqüência sinal e a toxina madura, e a
seqüência de aminoácidos da toxina madura. Os cDNAs que codificam as toxinas LTx1 e
LTx3 foram inseridos no vetor de expressão pET-11a, gerando plasmídeos com a
capacidade de codificar as toxinas maduras. O sistema pET-11a-LTx1 produziu a toxina
LTx1 recombinante na forma solúvel.

VI
Abstract
Spider venoms are multicomponent systems that can be grouped into three major chemical
classes: low molecular mass organic molecules, polypeptides and high molecular mass
protein. The vast majority of spider toxins identified up to date are polypeptides with a
molecular mass of 3000-8000 Da, highly reticulated by disulfide bridges. Purification of
peptides toxins from spider venoms gives an arsenal of molecular tools for
electrophysiological, pharmacological and structural studies of ion channels. The present
work describes the results of a screening carried out using ELISA and a cDNA expression
library for identification of immunogenic proteins. The cDNA library was previously
constructed with mRNA isolated from Lasiodora sp venom glands. Antibodies against the
whole venom were used in this screening. Positive clones were characterized by DNA
sequencing. The sequences revealed that the precursors of proteins contain signal peptides
characterized by a very hydrophobic core, an intervening propeptide region, and the mature
toxin. The cDNAs encoding the LTx1 and LTx3 toxins were inserted into the expression
vector pET-11a, generating plasmids with coding capacity for mature toxins. The
recombinant toxin LTx1 was found primarily in soluble fraction of the host cell.

Índice
Resumo V
Abstract VI
1 Lista de Figuras VII
2 Tabela IX
3 Abreviaturas X
4 Introdução 1
4.1 Aspectos gerais 2
4.2 Composição de venenos de aranhas 4
4.3Utilização de toxinas de venenos no estudo de canais iônicos e em neurobiologia 5
4.4 Atividade bioinseticida de venenos de aranhas 12
4.5 Atividade bactericida de venenos de aranhas 14
4.6 Bioquímica e biologia molecular de toxinas da aranha Lasiodora sp 15
5 Objetivos 18
5.1 Objetivos Geral 19
5.2 Objetivos Específicos 19
6 Materiais e Métodos 20
6.1 Construção da Biblioteca de cDNA 21
6.2 Dosagem do mRNA 23
6.3 Preparação do fago auxiliar M13 23
6.4 Titulação do fago auxiliar M13 24
6.5 Excisão em massa do fagemídeo pBK-CMV 24
6.6 Preparo do estoque de bactérias recombinantes 27
6.7 Preparo do extrato livre de células 27
6.8 ELISA (Enzyme Linked Imunosorbent Assay) 28
6.9 Extração do DNA plasmidial 29
6.10 Preparo do gel de agarose 30
6.11 Eletroforese do DNA em gel de agarose 30
6.12 Seqüenciamento do DNA 31
6.13 Purificação dos produtos de extensão 32
6.14 Análise das seqüências de nucleotídeos obtidas 32
6.15Clonagem das seqüências codificantes para LTx1 e LTx3 no sistema de expressão
bacteriano pET-11a e pET-21b Novagen
® 32
6.15.1Amplificação da região correspondente às toxinas maduras LTx1 e LTx3 para
clonagem em sistema PCR®
2.1 TOPO TA Cloning Invitrogen® 33
6.15.2 Preparo de gel de poliacrilamida 6 % 35
6.15.3 Purificação do produto de PCR em gel de poliacrilamida 35
6.15.4Clonagem das seqüências codificantes para LTx1 e LTx3 em PCR
®2.1-TOPO TA
Cloning Invitrogen®
36
6.15.5 Preparo de células E. coli Top 10 F’competentes (Método CaCl2) 38
6.15.6Transformação de células de E. coli Top 10 F’ com o produto de ligação em PCR
®
2.1-TOPO®
38
6.15.7Reação em cadeia da polimerase de colônias provenientes da transformação de E.
coli Top 10 F’ com o produto de ligação em PCR®
2.1-TOPO® 39
6.15.8Extração do DNA plasmidial de colônias provenientes da transformação de E. coli
Top 10 F’ com o produto de ligação em PCR®
2.1-TOPO® 40
6.15.9
Digestão dos plasmídeos PCR®
2.1-TOPO®-LTx1 (e LTx3) com as enzimas Nde I e
BamH I para clonagem do fragmento liberado no sistema de expressão pET-11a
(Novagen®)
41
6.15.10 Purificação dos produtos de digestão 41
6.15.11Digestão do vetore de expressão pET-11a com as enzimas de restrição Nde I e
Bam H I42

6.15.12 Purificação do vetor de expressão utilizando papel Whatman DE81 42
6.15.13Clonagem dos fragmentos que codificam as toxinas LTx1 e LTx3 no vetor de
expressão pET-11a44
6.15.14Transformação de células de E. coli Top 10 F’ com o produto de ligação no vetor de
expressão pET-11a 46
6.15.15
Reação em cadeia da polimerase de colônias provenientes da transformação de
células de E. coli Top 10 F’ com os produtos de ligação no vetor de expressão pET-
11a
46
6.15.16Transformação de bactérias de expressão (E. coli linhagem BL-21 DE3) com a
construção pET-11a-LTx1 e pET-11a-LTx348
6.16Indução da expressão da toxina recombinante LTx1 clonada no vetor de expressão
pET-11a 48
6.16.1 Produção da LTx1 recombinate e solubilidade da proteína 49
6.16.2 Eletroforese em gel de poliacrilamida SDS-PAGE 50
6.16.3 Coloração pelo método da prata 50
6.16.4 Western Blot 51
6.17 Genótipos das linhagens de bactérias hospedeiras utilizadas neste trabalho 52
6.18 Meios de cultura 52
6.19 Reagentes e material de uso 53
6.20 Soluções 54
6.20.1 Soluções utilizadas na ELISA 55
6.20.2 Soluções utilizadas no gel SDS-PAGE 56
6.20.3 Coloração pelo método da prata 57
6.21 Equipamentos utilizados no laboratório 58
7 Resultados 59
7.1 Varredura dos clones provenientes das excisões em larga-escala 60
7.2Análise das seqüências nucleotídicas dos cDNAs dos clones que apresentaram
similaridade com toxinas de aranhas 62
7.2.1 Tradução dos cDNAs que codificam para as toxinas LTx1, LTx2 e LTx3 64
7.2.2 Análise estrutural das toxinas LTx1, LTx2 e LTx3 66
7.2.3
Alinhamento das seqüências peptídicas das toxinas das aranhas Lasiodora sp
(LTx1, LTx2, LTx3), E. Californicum (ESTx), B. smithii (BsTx), L. parahybana
(LpTx1 e LpTx2)
69
7.2.4Alinhamento das seqüências peptídicas completas das toxinas LTx1, LTx2, LTx3 e
HwTx-II (Ile)70
7.3 Clonagem das toxinas LTx1 e LTx3 em PCR®
2.1-TOPO TA Cloning Invitrogen® 71
7.4 Clonagem das toxinas LTx1 e LTx3 no vetor de expressão pET-11a 73
7.5 Indução da toxina LTx1 recombinante 75
7.6Análise da seqüência nucleotídica dos cDNAs dos clones (16-19) que apresentaram
similaridade com a toxina Magi 7 da aranha Macrothele gigas 78
7.7Análise das seqüências que apresentaram similaridade com outras proteínas
depositadas no GenBank82
7.7.1 Análise da seqüência do DNA complementar do clone 20 82
7.7.2 Análise da seqüência do DNA complementar do clone 21 85
7.8Análise das seqüências dos clones que codificam para peptídeos que não
apresentaram similaridades, quando submetidos à análise pelo BlastP88
7.8.1 Análise da seqüência do DNA complementar do clone 22 89
7.8.2 Análise da seqüência do DNA complementar do clone 23 93
7.8.3 Análise da seqüência do DNA complementar do clone 24 94
8 Discussão 95
9 Conclusões 106
10 Perspectivas 109
11 Referências Bibliográficas 111
Anexo 119

VII
1- Lista de Figuras Figura 1 Foto de um espécime do gênero Lasiodora 3
Figura 2 Estrutura tetramérica da latrotoxina αLTX 7
Figura 3 Modelo de poro da toxina αLTX 7
Figura 4 Mecanismos de ação da αLTX 9
Figura 5 Localização de VDSC em células GH3 e co-localização de manchas AF568-TiTX-γ com AF488-TsTX
11
Figura 6 Construção da molécula de cDNA 22
Figura 7 Ligação do cDNA no vetor ZAP Express e excisão em massa do fagemídeo pBK-CMV 26
Figura 8 Mapa do vetor de clonagem PCR® 2.1-TOPO TA Cloning Invitrogen® 37
Figura 9 Mapa esquemático do vetor de expressão bacteriano pET-11a Novagen® 45
Figura 10 Alinhamento dos cDNAs dos clones que apresentaram similaridade com toxinas de aranhas S. huwena, E. californicum, B. smithi e Lasiodora parahibana 63
Figura 11 Tradução das seqüências nucleotídicas que codificam para as toxinas LTx1, LTx2 e LTx3 65
Figura 12 Predição do ponto de clivagem entre os peptídeos sinal e peptídeo intermediário na seqüência dos precursores das toxinas LTx1, LTx2 e LTx3 67
Figura 13 Gráfico de hidropaticidade das seqüências de aminoácidos traduzidas a partir dos cDNAs dos clones que codificam os precursores das toxinas LTx1, LTx2 e LTx3 68
Figura 14
Alinhamento das seqüências de aminoácidos das toxinas das aranhas Lasiodora sp (LTx1, LTx2 e LTx3), E. Californicum (ESTx), B. smithii (BsTx), e L. parahybana (LpTx1 e LpTx2)
69
Figura 15 Alinhamento das seqüências de aminoácidos das toxinas das aranhas Lasiodora sp (LTx1, LTx2 e LTx3) e Selenocosmia huwena (HwTx-II) 70
Figura 16
PAGE dos produtos de PCR de colônias obtidas na transformação de células de E. coli linhagem TOP 10 F’ com os produtos das ligações das seqüências codificantes para as toxinas LTx1 e LTx3 com o vetor PCR® 2.1-TOPO
72
Figura 17
PAGE dos produtos de PCR de colônias obtidas na transformação de células de E. coli linhagem TOP 10 F’ com produto de clonagem das seqüências codificantes para LTx1 e LTx3 nos vetores de expressão pET-11a e pET-21b
74
Figura 18
Gel SDS-PAGE da indução da expressão de LTx1 recombinante
76
Figura 19
Western Bloting da toxina LTx1 recombinante
77
Figura 20
Tradução do cDNA que codifica a toxina LTx5
79

VIII
Figura 21 Alinhamento das seqüências peptídicas dos pró-peptídeos Magi 7 (Macrothele gigas) e LTx5 (Lasiodora sp) 79
Figura 22
Predição do ponto de clivagem entre os peptídeos sinal e intermediário na seqüência do precursor da toxina LTx5
80
Figura 23 Gráfico de hidropaticidade da seqüência de aminoácidos traduzida a partir do cDNA dos clones que codificam o precursor da toxina LTx5 81
Figura 24
Alinhamento da seqüência peptídica codificada pelo cDNA do clone 20 com a cadeia β-1 da hemoglobina de Mus musculus, cadeia β-2 da globina de Rattus norvegicus e hemoglobina quimérica humano/camundongo (zeta 2/beta 2)
84
Figura 25
Alinhamento da seqüência peptídica codificada pelo cDNA do clone 21 com as proteínas CG7580-PA da mosca Drosophila melanogaster, agCP10691 do mosquito Anopheles gambiae e Qpc-pending do camundongo Mus musculus
87
Figura 26
Predição do ponto de clivagem entre os peptídeos sinal e intermediário na seqüência do peptídeo LsC3a
91
Figura 27
Gráfico de hidropaticidade dos aminoácidos traduzidos a partir do cDNA dos clones que codificam o precursor do componente clonado LsC3a
92
Figura 28
Perfil molecular de peptídeos de venenos de 55 tarântulas utilizando espectrometria de massa MALDI-TOF
99
Figura 29
Alinhamento das seqüências peptídicas das toxinas maduras das aranhas Lasiodora sp e Lasiodora parahybana
100
Figura 30
Alinhamento (ClustalW) de toxinas de tarântulas
102
Figura 31
Alinhamento dos precursores das toxinas da aranha Lasiodora sp LTx1, LTx2, LTx3, LTx4 e LTx5
103
Figura 32
Árvore filogenética das toxinas da aranha Lasiodora sp baseada no programa de alinhamento ClustalW
104

IX
2- Tabela Tabela 1 Resultados do teste de ELISA e similaridade utilizando o programa BlastP 62

X
3- Abreviaturas
ATP: adenosina trifosfato
Blast: “Basic Local Alignment Search Tool”
BsTx: toxina do veneno da aranha Brachypelma smithii
cDNA: ácido desoxirribonucléico complementar
DL50: dose letal mediana
DMSO: Dimetil sulfóxido
DNA: ácido desoxirribonucléico
dNTP: desoxirribonucleotídeo(n)trifosfato, n= adenosina, citosina, guanosina ou timina
DO: densidade ótica
EDTA: ácido etilenodiamino tetracético
ELISA: Enzyme Linked Imunosorbent Assay
ESTx: toxina do veneno da aranha Eurypelma californicumi
GABA: ácido γ-aminobutírico
HPLC: High-Performance Liquid Chromatography
HwTx-(I e II): toxinas I e II do veneno da aranha Selenocosmia huwena
IgG: imunoglobulina G
IPTG: isopropil-tio-β-D-galactosídio
LpTx : toxinas da aranha Lasiodora parahibana
LsC : componente do veneno da aranha Lasiodora sp
LTx: toxinas da aranha Lasiodora sp
MCS: “Multiple Cloning Site” (sítio múltiplo de clonagem)
mRNA: ácido ribonucléico mensageiro
NRX: Neurexina
OPD: O-fenilenodiamino
PBS: tampão de salina fosfato
PBST: tampão de salina fosfato tween 20
PCR: “Polymerase Chain Reaction” (reação da polimerase em cadeia)
PEG: polietilenoglicol
PLC: Fosfolipase C

XI
PMSF: fluoreto fenilmetilsulfonil
PSA: persulfato de amônia
rATP: ATP reativo
RNA: ácido ribonucléico
Rnase: ribonuclease
ssDNA: single strand DNA (DNA fita simples)
SDS: dudecil sufato de sódio
SDS PAGE: Eletroforese em gel de poliacrilamida com SDS
TAE: tampão tris acetato EDTA
TE: tampão tris EDTA
TEMED: N, N, N`, N` tetrametiletilenodiamino
TiTx-γ: Tityus Toxina Gama
Tris HCl: tris(hidroximetil) aminometano acidificado com ácido clorídrico
TTX: tetrodotoxina
VDSC: Voltage Dependent Sodium Channel
X- Gal: 5-bromo-4-cloro3-indolil-β-D-galactosídio

2
4- Introdução
4.1- Aspectos gerais
A aranha Lasiodora sp é conhecida popularmente no Brasil como caranguejeira.
Assim como outras aranhas da família Theraphosidae (subordem Mygalomorphae), a
aranha caranguejeira pertence a um grupo de aranhas também conhecidas mundialmente
como tarântulas. Das 37.972 espécies de aranhas descritas até o momento (totalizando 3526
gêneros), apenas 860 espécies pertencem à família Theraphosidae (composta por 107
gêneros) (Escoubas & Rash, 2004). As tarântulas podem ser encontradas em áreas tropicais
e semitropicais, savanas, desertos, florestas úmidas ou ambientes semitemperado. Essa
diversidade de nichos ecológicos, associada à diversidade de comportamentos de captura de
presas, contribui para a grande diversidade de seus venenos. Assim como outras aranhas, as
tarântulas são predadoras e se alimentam de uma variedade de animais vertebrados e
invertebrados. A grande habilidade das tarântulas para capturar presas grandes e fortes sem
a ajuda de teias (as tarântulas são aranhas errantes) implica não apenas em força física, mas
também em venenos altamente eficientes que agem rapidamente sobre o sistema nervoso
central e periférico de suas presas. Esta característica sugere que estes venenos sejam fontes
promissoras de compostos a serem utilizados no estudo de receptores e canais iônicos de
vertebrados. Dentre as inúmeras espécies de tarântulas conhecidas podem-se citar
Cithariscius crawshayi, Scodra griseipes, Hysterocrates Hercules (encontradas na África),
Selenocosmia Lyra, Haplopelma lividum, Cyriopagopus paganus (encontradas na Ásia),
Brachypelma smithi, Avicularia magdalena, Aphonopelma texense (encontradas na
América Central e do Norte) e Lasiodora parahybana, Grammostola spatulata, Paraphysa
manicata (encontradas na América do Sul) (Escoubas & Rash, 2004). A toxicidade do
veneno das aranhas, na maioria dos casos, não está diretamente relacionada com o tamanho
corporal. Dados recentes sobre picadas de aranhas da família Theraphosidae da América do
Sul (48 casos), Ásia/África (4 casos) e Austrália (9 casos) sugerem que essas aranhas são
inofensivas a humanos (Escoubas & Rash, 2004). Dentre os sintomas decorrentes de
picadas de tarântulas em humanos estão dor local moderada a severa, coceira forte, edema,
eritema, inchaço, ardência e cãibra. Nos casos mais severos são observados cãibras fortes e

3
espasmos musculares, os quais podem persistir por várias horas (Escoubas & Rash, 2004).
A dor decorrente da picada pode estar relacionada a uma combinação de injúria mecânica
(provocada pelas grandes quelíceras), baixo pH do veneno (tipicamente pH 5), efeitos das
aminas biogênicas (serotonina e histamina), adenosina e ATP (Escoubas & Rash, 2004). No
entanto, até mesmo venenos de baixa toxicidade para humanos podem ser fontes muito
importantes de moléculas que podem ser usadas como ferramentas bioquímicas para
estudos de processos farmacológicos, fisiológicos e neurológicos de vertebrados.
A figura 1 ilustra um representante do gênero Lasiodora.
Figura 1: Foto de um espécime do gênero Lasiodora. (Disponível em: www.bioterium.com)

4
4.2- Composição de venenos de aranhas
Tal como venenos de outros animais, como serpentes e escorpiões, venenos de
aranhas são uma rica fonte de toxinas, incluindo peptídeos de baixo peso molecular,
neurotoxinas, toxinas dermonecróticas (que são um grupo de proteínas com intensa
atividade sobre a matriz celular), poliaminas livres, ATP, ADP, AMP, sais inorgânicos,
aminoácidos livres, nucleotídeos, acilpoliaminas (que em venenos de aranhas do gênero
Nephila agem como neurobloqueador com atividade letal), polipeptídeos com mais de 1000
aminoácidos (principais componentes letais do gênero Latrodectus, causadores de uma
liberação maciça de neurotransmissores de uma diversidade de terminais nervosos), fatores
que podem modificar vários componentes da circulação sanguínea (tal como as
esfingomielinases, às quais se tem atribuído ações hemolíticas que podem causar anemia,
insuficiência renal aguda bem como ulcerações cutâneas) e neurotransmissores, dentre os
quais pode-se citar o glutamato, aspartato, ácido γ-aminobutílico (GABA), histamina,
dopamina, serotonina e epinefrina (Tambourgi, e cols., 2004; Horni e cols., 2001; Savel-
Niemann, 1989; Chan e cols., 1975; Kalapothakis e cols., 2002; Turkov e cols., 1997;
Escoubas e cols., 2000; Atkinson & Wright, 1992a; Rash & Hodgson, 2002).
Nem todos os componentes encontrados em glândulas de venenos de aranhas são
produzidos por células desse órgão. A presença de nucleotídeos de adenosina (ATP, ADP e
AMP) em venenos de aranhas foi demonstrada em 1975 por Chan e cols. no veneno de
Dugesiella hentzi e da tarântula Aphonopelma sp. Demonstrou-se que o composto
adenosina 5`-trifosfato (ATP) é relativamente pouco tóxico para ratos comparado com a
necrotoxina purificada do veneno de D. hentzi, com uma DL50 (ATP) de 2.4 mg por grama
de peso corporal. A necrotoxina de D. hentzi apresentou uma DL50 de 8,5µg por grama de
peso corporal. Não obstante, a toxicidade da necrotoxina foi notavelmente aumentada pela
presença de ATP. Os nucleotídeos presentes em venenos podem ser produzidos diretamente
em glândulas de venenos, embora não haja evidência experimental da presença de
nucleotídeos em glândulas de venenos de tarântulas. O ATP, um composto rico em energia,
pode ser necessário para a liberação do veneno, sendo secretado juntamente com outras
substâncias quando a aranha é excitada (Chan e cols., 1975). Considerando-se as
concentrações relativamente altas encontradas nos experimentos de Chan e cols. (1975), o

5
ATP pode ter uma função especial no veneno de D. hentzi e Aphonopelma sp, agindo como
potenciador da toxicidade de fatores necróticos do veneno. É provável também que os
grupos fosfatos liberados a partir da hidrólise do ATP sejam usados para fosforilar um ou
mais resíduos de aminoácidos de peptídeos tóxicos do veneno, tornando-os mais ativos.
Em 1984, Geren & Odell observaram também a presença de fluidos digestivos em
venenos de aranhas. Tais enzimas digestivas não são produzidas nas glândulas de veneno,
mas, como sugerido por Atkinson & Wright (1992a), decorrem de uma contaminação
natural do veneno com fluidos digestivos. As enzimas colagenases são os principais
responsáveis pela ação necrótica de muitos venenos (Atkinson & Wright, 1992a).
4.3. Utilização de toxinas de venenos no estudo de canais iônicos e em neurobiologia
Os canais iônicos são proteínas integrais de membrana que formam poros capazes
de permitir a passagem de íons através da bicamada lipídica e, desta forma, possibilitam a
alteração do potencial de membrana de uma célula. Este fluxo de íons através dos canais
iônicos presentes na membrana celular gera os chamados potencias de ação, que são os
sinais elétricos responsáveis pela condução do impulso nervoso.
Alguns peptídeos tóxicos de baixo peso molecular de venenos animais agem
especificamente sobre canais iônicos incluindo canais de Ca2+ , canais de K+ , canais de
Na + e canais de Cl- , (Atkinson & Wright, 1992b e Rash & Hodgson, 2002). Atualmente,
muito se tem estudado a respeito das neurotoxinas. Os modos de ação dessas substâncias
têm permitido aos pesquisadores estudar os mecanismos envolvidos na neurotransmissão
uma vez que essas toxinas, ao interagir de forma específica com receptores de membranas
neuronais, podem estimular ou inibir a liberação de neurotransmissores, tais como o
glutamato (Barral e cols., 2001), bem como afetar a transmissão colinérgica. Peptídeos
tóxicos de venenos têm contribuído também com estudos de caracterização e
funcionamento de canais iônicos (Araújo e cols., 1993 e Rash & Hodgson, 2002).
Ushkaryov e cols. (2004) determinaram a estrutura tridimensional de uma toxina
presente no veneno de uma aranha pertencente ao gênero Latrodectus e conhecida
popularmente como viúva negra. As toxinas presentes no veneno da aranha viúva negra

6
(latrotoxinas) têm sido extensivamente usadas no estudo de mecanismos moleculares
envolvidos em neurosecreção em vertebrados, insetos e crustáceos. Recentes avanços nas
análises estruturais e funcionais das latrotoxinas, sobretudo da toxina αLTX, têm permitido
entender os mecanismos envolvidos na liberação de neurotransmissores e têm revelado o
grande potencial desses peptídeos tóxicos como ferramentas para compreender fenômenos
neurobiológicos. À exceção da toxina δLIT, todas as demais latrotoxinas agem somente
sobre células neuronais e a αLTX apresenta forte afinidade por receptores de membranas
neuronais de vertebrados (Ushkaryov e cols., 2004). A estrutura tridimensional da αLTX é
formada por um complexo que consiste em quatro monômeros (complexo tetramérico)
arranjados ao redor de um eixo central assemelhando-se a uma hélice de quatro lâminas
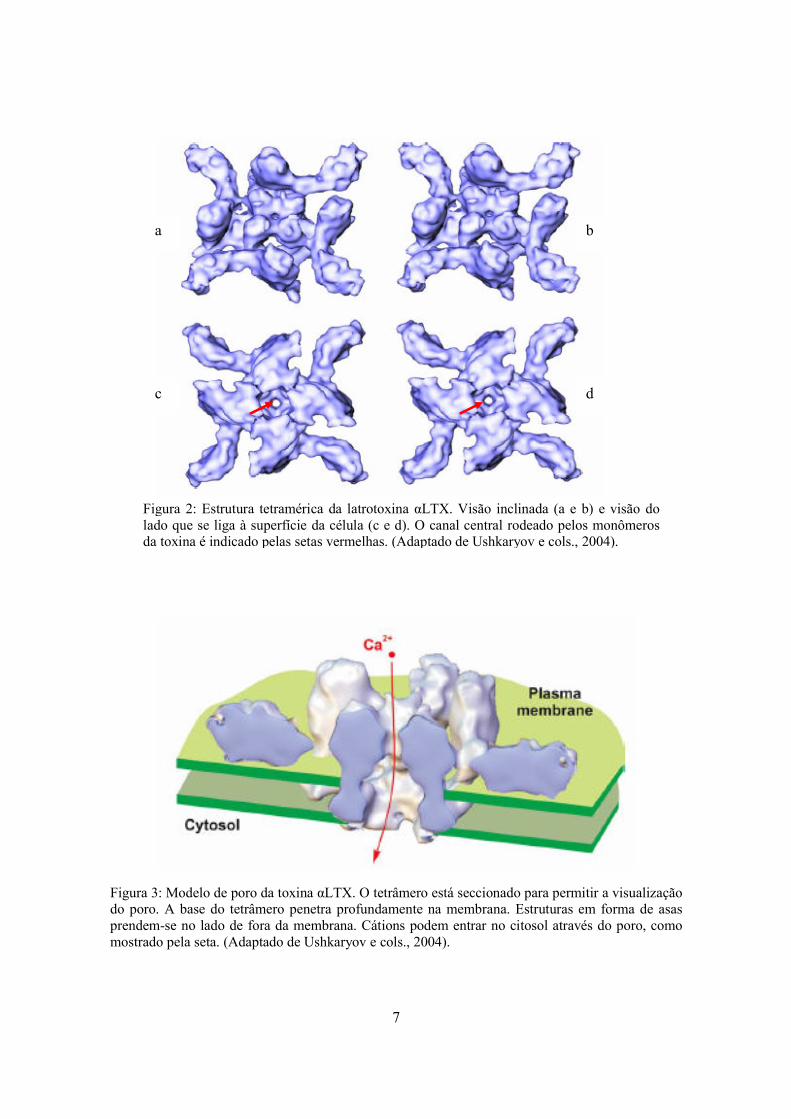
com um diâmetro de 250 Å e uma espessura de 100 Å (Figura 2 a-d) (Ushkaryov e cols.,
2004). Esse tetrâmero possui uma região composta por aminoácidos hidrofóbicos que,
provavelmente, interage com a membrana (Ushkaryov e cols., 2004). Somente através da
determinação da estrutura quaternária da toxina αLTX pôde-se explicar como essa
molécula é capaz de se inserir na membrana tornando-a permeável a íons Ca2+. A
característica mais relevante do tetrâmero αLTX é um canal central rodeado pelos
monômeros da toxina (Figura 2c e 2d). O tetrâmero formado pela αLTX é anfipático sendo,
portanto, capaz de se aderir em superfícies hidrofóbicas. Dessa forma, o tetrâmero αLTX
inserido na membrana tornaria a bicamada lipídica permeável a todas as substâncias que
pudessem passar pelo canal central (pequenas moléculas e cátions hidratados) (Figura 3). O
entendimento desse mecanismo de ação (mecanismo do poro) tem contribuído para a
identificação de seus efeitos específicos sobre a liberação de neurotransmissores e tem
ajudado no estudo das ações da αLTX mediadas por receptor (Ushkaryov e cols., 2004).
7
Figura 3: Modelo de poro da toxina αLTX. O tetrâmero está seccionado para permitir a visualização do poro. A base do tetrâmero penetra profundamente na membrana. Estruturas em forma de asas prendem-se no lado de fora da membrana. Cátions podem entrar no citosol através do poro, como mostrado pela seta. (Adaptado de Ushkaryov e cols., 2004).
Figura 2: Estrutura tetramérica da latrotoxina αLTX. Visão inclinada (a e b) e visão do lado que se liga à superfície da célula (c e d). O canal central rodeado pelos monômeros da toxina é indicado pelas setas vermelhas. (Adaptado de Ushkaryov e cols., 2004).
a b
c d

8
Embora as ações da αLTX sejam complexas, elas podem ser divididas em duas vias
principais. Após a ligação a um receptor neuronal específico, a αLTX pode desencadear
uma sinalização intracelular ou se inserir na membrana levando à formação de um poro.
Ambas as vias podem estimular a liberação de neurotransmissores, embora seus
mecanismos sejam diferentes. Os mecanismos de ação da αLTX são mostrados na figura 4.
Como pode ser visto na figura, a interação de αLTX com receptores específicos da
membrana neuronal, tais como latrofilinas (LPH) ou neurexinas (NRX), desencadeia um
influxo de Ca2+ através do poro formado pelo tetrâmero. Esse Ca2+ age sobre as vesículas
sinápticas desencadeando a liberação de neurotransmissores (via 1). A via 2 da figura 4
mostra um mecanismo de liberação de neurotransmissores independente de Ca2+, onde a
interação do tetrâmero com receptores específicos permite a saída dos neurotransmissores
através do poro formado pelo tetrâmero. A terceira via mostra a interação do dímero αLTX
com seus receptores específicos. Essa interação ativa uma proteína G que, por sua vez,
desencadeia uma cascata de reações envolvendo uma fosfolipase C (PLC) e
fosfotidilinositol (PIP3), culminando na liberação de Ca2+ de reservatórios intracelulares e,
conseqüentemente, na liberação de neurotransmissores das vesículas sinápticas. Um quarto
modo de ação da toxina αLTX na liberação de neurotransmissores independente de Ca2+
tem sido proposto, no entanto, o mecanismo envolvido nessa liberação ainda não foi
completamente esclarecido (Ushkaryov e cols., 2004).
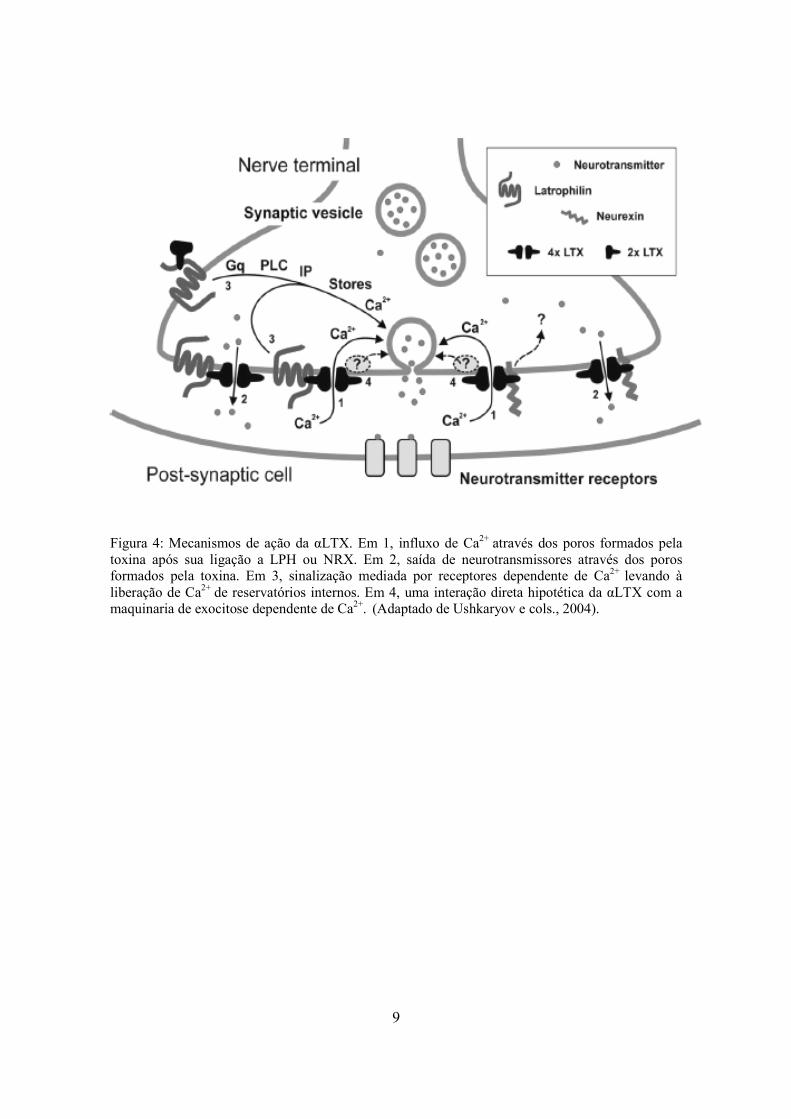
9
Figura 4: Mecanismos de ação da αLTX. Em 1, influxo de Ca2+ através dos poros formados pela toxina após sua ligação a LPH ou NRX. Em 2, saída de neurotransmissores através dos poros formados pela toxina. Em 3, sinalização mediada por receptores dependente de Ca2+ levando à liberação de Ca2+ de reservatórios internos. Em 4, uma interação direta hipotética da αLTX com a maquinaria de exocitose dependente de Ca2+. (Adaptado de Ushkaryov e cols., 2004).

10
Um tipo especial de canal de Na+ (dependente de voltagem) têm sido alvo de muitos
estudos neurobiológicos devido ao fato desses canais apresentarem múltiplas funções na
geração de potenciais de ação em muitos tipos de neurônios do sistema nervoso central.
Além de produzir uma grande corrente transiente responsável pela mudança no potencial de
membrana, os canais de sódio afetam a duração e a freqüência de disparos neuronais
repetitivos (Massensini e cols., 2002). A distribuição diferencial dos canais de Na+
dependentes de voltagem (VDSC) é crítica na determinação de propriedades funcionais
distintas de um neurônio. Embora haja várias técnicas para a localização de VDSCs
(“patch-clamp”, “immunostaining”, marcadores específicos de canais iônicos e etc)
nenhuma delas é capaz de determinar a distribuição de VDSCs funcionais em toda a
extensão da membrana de células vivas (Massensini e cols., 2002). Contudo, toxinas de
venenos animais têm sido usadas como potentes ferramentas para distinguir as várias
isoformas de canais iônicos dependentes de voltagem presentes nas membranas celulares.
Algumas dessas toxinas são conhecidas por interagir especificamente com VDSCs e
portanto, estão sendo utilizadas como ferramentas no estudo de funções de canais iônicos e
de excitabilidade celular em tempo real.
Massensini e cols. (2002) localizaram canais de Na+ voltagem-dependentes (VDSC),
em células vivas, utilizando-se de toxinas do veneno do escorpião Tityus serrulatus
marcadas com fluorescência. A visualização da distribuição espacial dos VDSCs em células
GH3 foi revelada utilizando-se derivados fluorescentes das toxinas TiTX-γ marcada com
Alexa Flúor 568 (AF568-TiTX-γ) e TsTX marcada com Alexa Flúor 488 (AF488-TsTX)
(Molecular Probes Inc., Eugene, OR) do escorpião Tityus serrulatus juntamente com
técnicas de microscopia confocal de varredura a laser (LSCM). Esse método permitiu uma
localização precisa de grupos individuais de cada toxina marcada, AF568-TiTX-γ e AF488-
TsTX. Sobreposições de manchas fluorescentes das toxinas marcadas utilizadas também
foram visualizadas, indicando regiões de co-localização de VDSCs em partes específicas da
célula (Figura 5).
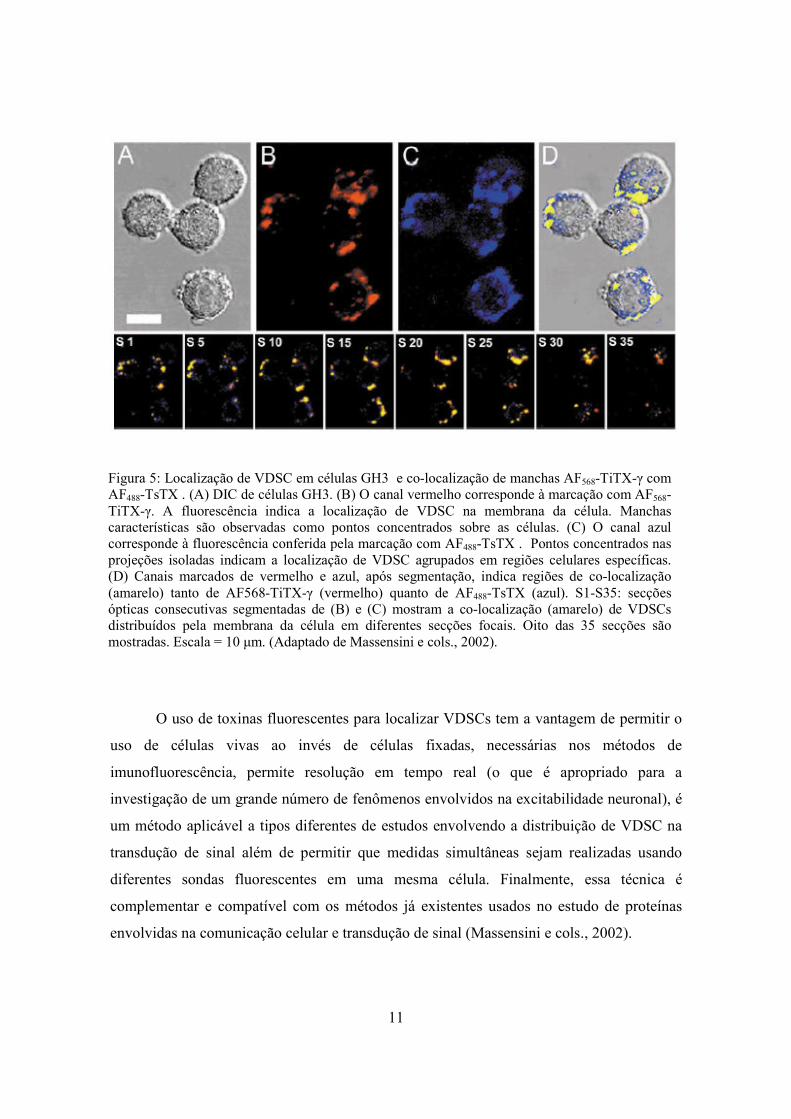
11
O uso de toxinas fluorescentes para localizar VDSCs tem a vantagem de permitir o
uso de células vivas ao invés de células fixadas, necessárias nos métodos de
imunofluorescência, permite resolução em tempo real (o que é apropriado para a
investigação de um grande número de fenômenos envolvidos na excitabilidade neuronal), é
um método aplicável a tipos diferentes de estudos envolvendo a distribuição de VDSC na
transdução de sinal além de permitir que medidas simultâneas sejam realizadas usando
diferentes sondas fluorescentes em uma mesma célula. Finalmente, essa técnica é
complementar e compatível com os métodos já existentes usados no estudo de proteínas
envolvidas na comunicação celular e transdução de sinal (Massensini e cols., 2002).
Figura 5: Localização de VDSC em células GH3 e co-localização de manchas AF568-TiTX-γ com AF488-TsTX . (A) DIC de células GH3. (B) O canal vermelho corresponde à marcação com AF568-TiTX-γ. A fluorescência indica a localização de VDSC na membrana da célula. Manchas características são observadas como pontos concentrados sobre as células. (C) O canal azul corresponde à fluorescência conferida pela marcação com AF488-TsTX . Pontos concentrados nas projeções isoladas indicam a localização de VDSC agrupados em regiões celulares específicas. (D) Canais marcados de vermelho e azul, após segmentação, indica regiões de co-localização (amarelo) tanto de AF568-TiTX-γ (vermelho) quanto de AF488-TsTX (azul). S1-S35: secções ópticas consecutivas segmentadas de (B) e (C) mostram a co-localização (amarelo) de VDSCs distribuídos pela membrana da célula em diferentes secções focais. Oito das 35 secções são mostradas. Escala = 10 µm. (Adaptado de Massensini e cols., 2002).

12
4.4- Atividade bioinseticida de venenos de aranhas
Os mecanismos de ação dos peptídeos tóxicos, em insetos e mamíferos, têm sido
estudados também com o intuito de se desenvolver novas drogas, tendo em vista as
diferentes atividades farmacológicas potenciais das neurotoxinas. Por outro lado, a
perspectiva de se descobrir, dentre os vários componentes de venenos de aranhas, algum
peptídeo que possa ser utilizado na produção de bioinseticidas, tem impulsionado várias
pesquisas com o objetivo de conhecer os mecanismos de ação envolvidos na paralisação e
morte de insetos atacados por aranhas.
O controle de pragas utilizando inseticidas químicos tem submetido diversas
populações de insetos à seleção darwiniana, ocasionando um inevitável desenvolvimento de
resistência. Estima-se que desde 1992, mais de 500 espécies de insetos adquiriram
resistência a uma ou mais classes de inseticidas químicos, incluindo 56 mosquitos do
gênero Anopheles e 39 do gênero Culex e nove espécies de carrapatos (Tedford e cols.,
2004).
Dentre os componentes de venenos de aranhas envolvidos na paralisação de insetos
estão as poliaminas e polipeptídeos. As poliaminas são inibidores pós-sinápticos capazes de
bloquear reversivelmente canais de cálcio, sensíveis ao glutamato, de músculos
locomotores de insetos o que leva à paralisia dos mesmos (Atkinson & Wright 1992b). Já
os polipeptídios tóxicos agem pré-sinapticamente de maneira essencialmente irreversível
induzindo ou inibindo a liberação de neurotransmissores (Atkinson & Wright 1992b).
Atualmente mais de 60 peptídeos tóxicos têm sido descritos. Muitos deles parecem agir
essencialmente sobre canais iônicos podendo bloquear a liberação de neurotransmissores,
por afetar a exocitose de vesículas pré-sinápticas, e induzir modificações anormais da
transmissão sináptica, resultando em paralisia flácida. Outros peptídeos podem provocar
paralisia excitatória induzida pela excessiva despolarização da membrana (Escoubas e cols.,
2000).
Três polipeptídios tóxicos com ação inseticida, isolados do veneno da aranha
Segestria florentina, causaram paralisia completa da larva Heliothis virescens (Ordem:
Lepdoptera/Família: Noctuidae). Uma toxina com atividade inseticida denominada SIT,
isolada do veneno da aranha Segestria florentina induziu também paralisia irreversível em

13
baratas (Lipkin e cols., 2002). A comparação da estrutura primária desses peptídeos com
peptídeos tóxicos de outras aranhas sugeriu que essa família de toxinas compartilha
relações estruturais e funcionais com outras neurotoxinas de aranhas, várias delas com
ações agonistas e antagonistas sobre diferentes canais de cálcio voltagem-dependentes
(Lipkin e cols.,2002).
Figueredo e cols. (1995) isolaram um componente neurotóxico denominado Tx4(6-
1) da fração PhTx4 do veneno da aranha Phoneutria nigriventer, que parece ser específica
para insetos. A seqüência completa de aminoácidos da Tx4(6-1) indica que se trata de um
polipeptídeo de cadeia simples contendo 48 aminoácidos com um peso molecular de
5251Da, e ensaios de atividade mostraram que a fração é capaz de agir sobre mosca
doméstica e barata.
Uma das principais vantagens do uso de inseticidas biológicos é a sua alta
especificidade em relação à praga alvo, não afetando outros insetos, plantas e animais
(Bergmann & Pinedo 2001). Portanto, vários peptídeos neurotóxicos com atividade
inseticida têm sido clonados a fim de se elucidar seus modos de ação sobre canais iônicos,
o que pode revelar a eficácia de tais peptídeos no controle de pragas agrícolas. Uma
estratégia que tem sido testada com sucesso consiste na utilização de baculovírus
geneticamente modificado para expressar neurotoxinas. Os baculovírus são patógenos
usados como inseticidas biológicos que agem especificamente em insetos. No entanto, os
baculovírus agem lentamente, permitindo, em muitos casos, que a praga destrua grande
parte da lavoura. Zlotkin e cols. (2000) clonaram uma neurotoxina do escorpião
Androctonus australis, denominada AaIT, em vetor de baculovírus e mostraram que o vírus
atua como um vetor da toxina recombinante dirigindo-a ao sistema nervoso central do
inseto, o que produz um aumento da toxicidade.

14
4.5- Atividade bactericida de venenos de aranhas
A perspectiva de se encontrar peptídeos de glândulas de veneno de aranhas com
atividade bactericida tem sido impulsionada pelo surgimento, cada vez maior, de bactérias
resistentes a antibióticos. O mecanismo de ação desses peptídeos, mais bem conhecido, é
através de sua inserção na membrana celular causando destruição ou permeabilização da
mesma, levando o microrganismo à morte. Alternativamente, os peptídeos antimicrobianos
podem se ligar a um receptor de membrana levando a uma perda específica de sua função.
Além disso, ao se translocarem através da membrana, essas moléculas com propriedade
antibiótica podem atuar intracelularmente, impedindo a síntese de metabólitos importantes
para o microrganismo (Lohner, 2001).
Em 1989, Xu & Qu encontraram um peptídeo antibiótico no veneno da aranha
Lycosa singoriensis. Recentemente, Yan & Adams (1998) descreveram a existência de
peptídeos com ação inseticida e bactericida no veneno da aranha Lycosa carolinensis,
denominados licotoxinas. Kuhn-Nentwig e cols. (1998) publicaram a estrutura de um
peptídeo com ação inseticida e bactericida, denominado CSTX-4, do veneno da aranha
errante Cupiennius salei. Dois anos depois, Haeberli e cols. (2000) caracterizaram com
maiores detalhes a atividade microbiana dos peptídeos do veneno desta mesma aranha.
Cinco diferentes peptídeos foram isolados do veneno da aranha Cupiennius salei e
suas atividades bactericidas foram testadas contra cinco diferentes espécies de bactérias,
tanto gram-negativas (Escherichia coli, Pseudomonas putida, Paracoccus denitrificans)
como gram-positivas (Staphylococcus epidermidis e Bacillus subtilis). Todas as cinco
bactérias foram susceptíveis a todos os cinco peptídeos em concentrações que variaram de
0,18 a 18µM e o veneno bruto causou 100% de inibição no crescimento em diluições até
1:55.000 (Haeberli e cols., 2000).
Belokoneva e cols. (2004) descreverem a estrutura de dois peptídeos catiônicos
lineares denominados Pandinina 2 (Pin2 – 24 resíduos de aminoácidos) e Oxyopinina 1
(Oxki1 – 48 resíduos de aminoácidos) isolados do veneno bruto do escorpião Pandinus
imperator e da aranha Oxyopes kitabensis, respectivamente. Ambos os peptídeos
mostraram exercer efeitos inibitórios sobre o crescimento de bactérias tanto Gram-
negativas quanto Gram-positivas e atividade hemolítica sobre eritrócitos contendo

15
quantidades abundantes de fosfatidilcolina na membrana. Os peptídeos Pin2 e Oxki1 ligam-
se a fosfocolinas de membrana desencadeando a formação de poros na bicamada lipídica
(Belokoneva e cols., 2004). Os peptídeos antimicrobianos parecem fazer parte de uma
família de peptídeos catiônicos, α-helicoidais, anfipáticos e sem resíduos de cisteínas. Os
membros desse grupo de peptídeos são amplamente distribuídos em uma miríade de
venenos animais e fazem parte de um mecanismo inato de defesa contra diferentes tipos de
patógenos (Moerman e cols., 2003).
4.6. Bioquímica e biologia molecular de toxinas da aranha Lasiodora sp
Embora vários estudos tenham sido realizados com venenos de outras aranhas,
pouco se sabe a respeito das ações das toxinas presentes no veneno da aranha Lasiodora sp.
Todavia, estudos recentes com venenos desta aranha identificaram algumas das ações
farmacológicas deste veneno, tais como, bloqueio do canal de cálcio do tipo-L e modulação
da cinética e voltagem de canais de Na+ (Kushmerick e cols., 2001). Os experimentos
realizados demonstraram que, quando o veneno da Lasiodora sp foi administrado a células
GH3 na presença de TTX (tetrodotoxina), que bloqueia canais de Na+ sensíveis à TTX, as
oscilações de Ca2+ foram rapidamente abolidas, reduzindo assim o nível basal de Ca2+
intracelular. Quando administrado na ausência de TTX, o veneno causou um lento aumento
do Ca2+ intracelular, o que sugere que, substâncias tais como neurotransmissores ou outras
moléculas pequenas presentes no veneno bloqueiam canais de Ca 2+ Tipo-L (Kushmerick e
cols., 2001). Como a única diferença experimental foi a presença ou ausência de TTX, os
autores concluíram que, além de seus efeitos sobre canais de Ca2+, o veneno age também
sobre canais de Na+.
Kushmerick e cols. (2001) também observaram que quando o veneno foi
administrado em altas concentrações (500µg ml- de proteína) ou aplicado na presença de
nifedipina ou Cd2+ , ambos bloqueadores de canais do Tipo-L, não ocorre aumento do Ca2+
intracelular, o que sugere que este aumento depende da disponibilidade de canais de Na+ e
Ca2+ do Tipo-L. Os autores sugeriram então, que o veneno causa o aumento de Ca2+
intracelular via abertura de canais de Na+ causando, portanto, despolarização da membrana

16
e entrada de Ca2+ via canais de Ca2+ voltagem-dependentes. Assim sendo, os efeitos
observados pela administração do veneno da aranha Lasiodora sp demonstraram que o
mesmo é uma fonte de toxinas ativas em canais iônicos, possuindo, portanto, um valor
inestimável para a caracterização dos mesmos.
Kalapothakis e cols. (2003) obtiveram dados que sugerem que o veneno da aranha
Lasiodora sp ocasiona, em coração de rato, um aumento na liberação vesicular de
acetilcolina na extremidade de nervos parassimpáticos por ativar canais de Na + resistentes
à TTX. Uma dose equivalente a 100µg do veneno da aranha Lasiodora sp causou
bradicardia, distúrbios rítmicos e parada cardíaca transitória no modelo experimental
utilizado. Os resultados sugerem que o veneno dessa aranha parece agir sobre canais de Na+
resistentes à TTX uma vez que 200nM dessa substância não foram suficientes para inibir o
efeito do veneno sobre o coração.
O fracionamento do veneno total parcialmente purificado da aranha Lasiodora sp
através de cromatografia de filtração molecular em coluna Sephadex G50 Fine (Pharmacia)
foi realizado por de Deus e cols (2003). Os autores obtiveram três frações denominadas P1,
P2 e P3. A atividade tóxica de cada pico foi avaliada tanto em insetos quanto em
mamíferos. de Deus e cols. (2003) observaram que as três frações possuíam atividade sobre
mamíferos, enquanto que somente as frações P2 e P3 possuíam atividade sobre o sistema
nervoso de insetos.
Moura e cols. (2004) realizaram a varredura de uma sub-biblioteca de cDNA da
glândula de veneno da aranha Lasiodora sp utilizando-se de antisoro anti-P3. A fração de
IgG de coelho anti-P3 purificada a partir de IgGs totais foi utilizada como primeiro
anticorpo na varredura de sub-bibliotecas para identificar clones que codificam toxinas
presentes na fração P3 do veneno total, utilizando-se a técnica de ELISA como descrito por
Kalapothakis e cols. (2001). Na varredura, os autores encontraram sete clones que
codificam para a toxina LTx1. Essa toxina foi previamente descrita em nosso laboratório
(Vieira e cols., 2004) e é idêntica à toxina LpTx1 descrita por Escoubas e cols. (1997).
Outros quatro clones encontrados codificam uma toxina denominada LTx4 , similar
às toxinas Magi 1 e Magi 4 (Corzo e cols., 2003), isoladas da glândula de veneno da aranha
Macrothele gigas, à Conotoxina Vx-VIb (Wang e cols., GenBank: ANN71750 [gi:
25140450), isolada do molusco marinho Conus vexillum e à neurotoxina Tx3-2 (Cordeiro e

17
cols., 1993), isolada da aranha Phoneutria nigriventer. Ensaios biológicos utilizando as
toxinas Magi, realizados por Corzo e cols. (2003), demonstraram que a Magi 1 não possui
ação tóxica em larvas da lagarta do tabaco ou em camundongos. A toxina Magi 4 induz
paralisia flácida nas larvas podendo levá-las a morte, e é letal para camundongos na DL50
de 0,15 pmol/g. Cordeiro e cols. (1993) demonstraram que a neurotoxina Tx3-2 produz
efeitos motores e paralisia flácida em camundongos seis horas após a injeção intracérebro-
ventricular da toxina, e Kalapothakis e cols. (1998) demonstraram que a toxina Tx3-2
purificada bloqueia canais do tipo L em células GH3, sugerindo que a toxina é um novo
peptídeo que atua em canais iônicos do tipo L. Embora nenhum ensaio de atividade tenha
sido realizado com a toxina LTx4, Moura e cols. (2004) inferiram, baseados em ensaios de
atividade das toxinas com as quais LTx4 apresentou similaridades, que LTx4 possa também
possuir ação sobre o sistema nervoso de vertebrados.
Tendo em vista o vasto espectro de atuação das moléculas presentes em venenos de
aranhas, sobretudo sobre os canais iônicos, bem como a relação destes com distúrbios
neurológicos, faz-se necessário conhecer não só a estrutura e a função desses canais para a
identificação de outros distúrbios ligados aos mesmos, mas também a descoberta de novos
fármacos com eficácia terapêutica. Assim sendo, a utilização de neurotoxinas ativas para
esses estudos é de suma importância. Contudo, o fracionamento do veneno em seus
componentes tóxicos é um processo árduo e oneroso. Frente a isso, a clonagem de toxinas
surge como ferramenta fundamental deste trabalho, pois além do conhecimento molecular,
a obtenção em grandes quantidades das toxinas recombinantes será de enorme valor em
investigações neurobiológicas. Neste trabalho, propomos a caracterização de clones de
cDNAs utilizando-se de uma biblioteca de cDNA produzida a partir do mRNA extraído da
glândula de veneno da aranha Lasiodora sp bem como a clonagem, em vetores de
expressão, de genes que codificam para toxinas presentes no veneno da aranha em questão.

19
5. Objetivos
5.1. Objetivo Geral
Proceder a varredura de uma biblioteca de cDNA, preparada a partir de glândulas de
veneno da aranha Lasiodora sp, com o objetivo de identificar toxinas.
5.2. Objetivos Específicos
• Preparo de uma biblioteca de cDNA a partir de mRNAs obtidos da glândula de
veneno da aranha Lasiodora sp;
• Varredura da biblioteca de cDNA utilizando a técnica de ELISA;
• Seqüenciamento e análise dos fragmentos gênicos que codificam para proteínas
imunogênicas;
• Clonagem, em vetores de expressão bacterianos, de fragmentos gênicos que
apresentarem similaridade com seqüências codificantes para toxinas depositadas no
GenBank;
• Expressão e purificação de toxinas recombinantes.

21
6. Materiais e métodos
6.1- Construção da Biblioteca de cDNA
A biblioteca de cDNA da glândula de veneno da aranha Lasiodora sp foi construída
no Laboratório de Biotecnologia e Marcadores Moleculares do Departamento de
Farmacologia da Universidade Federal de Minas Gerais (UFMG) pelos professores Dr.
Evanguedes Kalapothakis, Dr. Ieso de Miranda Castro e Dr. Elio Hideo Babá.
As glândulas de veneno das aranhas foram extraídas manualmente, pulverizadas em
presença de nitrogênio líquido usando pilão e pistilo e lisadas na presença de tiocianato de
guanidina, segundo o protocolo de Chirgwin e cols. (1979). O mRNA foi purificado através
de uma coluna de oligo (dT)-celulose usando Poly (A) Quick mRNA Isolation Kit
(Pharmacia) e o mRNA foi eluído de acordo com as recomendações do fabricante. A
síntese do DNA complementar foi realizada usando o Kit síntese ZAP-cDNA (Stratagene).
A figura 6 ilustra os passos de obtenção da molécula de cDNA. As moléculas de cDNA
foram inseridas direcionalmente entre os sítios Eco RI e Xho I do vetor Uni-ZapTM e
subseqüentemente empacotadas em partículas de fago Lambda ZAP usando o Kit Gigapack
III Packaging Extracts (Stratagene) (Figura 7).

22
Figura 6: Construção da molécula de cDNA. A transcriptase reversa catalisa a reação de síntese da primeira fita de cDNA usando como molde o mRNA isolado da glândula de veneno da aranha e um iniciador oligo(dT) contendo o sítio Xho I. O 5-metil dCTP é usado para adicionar grupos metil à primeira fita do cDNA. Uma RNase H cliva o mRNA molde permitindo assim a ação da DNA polimerase I e a síntese da segunda fita do cDNA. Com o auxílio da DNA ligase T4, adaptadores EcoR I são ligados às extremidades cegas do cDNA e em seguida são digeridas por suas respectivas enzimas de restrição (Xho I e Eco RI) (Figura adaptada do manual do fabricante Stratagene®).
5`GAGAGAGAGAGAGAGAGAGACTCGAGTTTTTTTTTTTTTTT3` AAAAAAAAAAAA 5`
5`GAGAGAGAGAGAGAGAGAGACTCGAGTTTTTTTTTTTTTTT3` AAAAAAAAAAAA 5`
CH3CH3 CH3
5`GAGAGAGAGAGAGAGAGAGACTCGAGTTTTTTTTTTTTTTT3`CTCTCTCTCTCTCTCTCTCTCT GAGCTC A AAAAAAAAAAA
CH3 CH3CH3
3`
5`
5`GAGAGAGAGAGAGAGAGAGACTCGAGTTTTTTTTTTTTTT3`CTCTCTCTCTCTCTCTCTCTCT GAGCTC A AAAAAAAAAA
3`5`
CH3 CH3CH3
dATP, dGTP, dTTP, 5-metil dCTP e MMLV-RT
RNase H, DNA polimerase IdNTPs e DNA ligase
Adapitadores Eco RIDNA ligase T4
Primer mRNA
5 CTCGAGTTTTTTTTTTTTTT3` C A AAAAAAAAAA
G 3`CTTAAG 5`
CH3 CH3 CH3
cDNA
GAGAGAGAGAGAGAGAGAGACTCGAGTTTTTTTTTTTTTTCTCTCTCTCTCTCTCTCTCTCT GAGCTC A AAAAAAAAAA
CH3 CH3CH3
GAATTC 3`CTTAAG 5`
Xho I Eco RI
Enzimas de restriçãoXho I e Eco RI
5`GAATTC3`CTTAAG

23
6.2- Dosagem do mRNA
A quantificação do mRNA purificado foi realizada através de leitura no
espectrofotômetro a 260nm. O fator de conversão de densidade óptica (DO) = 1
corresponde à concentração de RNA de 40 µg/ml, de acordo com o protocolo descrito por
Sambrook e cols. (1989). As razões entre as leituras A260/A280 nm foram estimadas para
verificar a pureza do mRNA. A amostra foi considerada pura quando as razões acima
apresentaram valores entre 1.8 e 2.0.
6.3- Preparação do fago auxiliar M13
Inoculou-se 15µl de células E. coli, linhagem XL1-BLUE, em 10ml de meio
“superbroth” contendo tetraciclina na concentração de 12,5µg/ml e incubado a 37ºC por
uma hora. Em seguida, adicionou-se às células 2µl de fago e o sistema foi incubado a 37ºC
por 2 horas. Após esta incubação, as células foram transferidas para 500µl de meio
“superbroth” contendo tetraciclina na concentração de 12,5µg/ml e incubou-se sob agitação
à temperatura de 37ºC por 14-16 horas. No dia seguinte, as células foram centrifugadas por
15 minutos a 2500 g. O sobrenadante foi incubado por 20 minutos a 70ºC e então
novamente centrifugado. Após a centrifugação, o sobrenadante foi estocado em volumes de
50 µl a 4ºC, em presença de DMSO 7% (v/v).

24
6.4- Titulação do fago auxiliar M13
Cerca de 1µl de diluições seriadas do fago M13 (10-9, 10-10, 10-12, 10-38, 10-39, 10-40 e
10-42) em meio LB foram adicionados a alíquotas de 100µl de uma cultura de bactérias
XL1-Blue (DO600 = 1) e incubadas durante 30 minutos à temperatura de 37ºC. Após a
infecção, 5ml de meio top ágar foi adicionada à mistura que foi então semeada em meio
“superbroth” ágar. As placas foram incubadas por 16 horas a 37ºC. O título da preparação
em unidades formadoras de colônia por ml (pfu/ml) foi obtido através da fórmula:
Número de placas de lise (pfu) X fator de diluição X 1000µl/ml Volume plaqueado
6.5- Excisão em massa do fagemídeo pBK-CMV
Inicialmente, as células XL1-Blue e XLOLR, provenientes do estoque, foram
crescidas em meio LB-ágar tetraciclina (50µg/ml) com objetivo de se obter células novas.
No dia seguinte, uma colônia de cada bactéria foi novamente inoculada em 3 ml de meio
LB-caldo contendo tetraciclina na mesma concentração anterior, e procedeu-se o
crescimento por aproximadamente 3 horas a 37ºC. Em seguida, as células foram inoculadas
em 25 ml de meio NZY contendo maltose 0.2% e MgSO4 100mM. As células XL1-Blue e
XLOLR foram então centrifugadas a 1.000g durante 10 minutos e suspensas em Mg2SO4
10mM de forma a obter suspensões com DO igual a 5.0 e 0.75, respectivamente. O passo
seguinte consistiu na incubação de 100µl de biblioteca com 200µl de XL1-Blue e 1µl de
fago auxiliar M13 a 37ºC por 30 minutos. Decorridos os 30 minutos, foi acrescentado 1 ml
de meio NZY e o tubo foi incubado por 2 horas a 37ºC. Em seguida, o sistema foi incubado
por 15 minutos a 70ºC. Após a incubação, as células foram centrifugadas a 1000 g por 15
minutos. Após a centrifugação, uma alíquota de 200µl de sobrenadante foi recolhida e
adicionada a 200µl de XLOLR que foram então incubados por 30 minutos a 37ºC.
Transcorrido o tempo de incubação, foram adicionados 300µl de meio NZY e o sistema foi
incubado por 45 minutos a 37ºC sob agitação (shaker). Em seguida, alíquotas de 100µl
foram plaqueados em meio LB-ágar contendo canamicina (50µg/ml) e X-Gal, totalizando 7

25
placas. À placa controle, adicionou-se 100µl de XLOLR, apenas. Como a XLOLR usada na
placa controle não passou pelo processo de infecção, e portanto não possui o DNA
recombinante, ela é sensível à canamicina, e portanto não é capaz de crescer. As placas
foram incubadas a 37ºC durante a noite. As placas contendo colônias de bactérias
recombinantes foram estocadas a 4ºC, por no máximo uma semana.
A figura 7 ilustra a inserção do cDNA dentro do genoma do fago Lambda, bem como
sua posterior excisão e recircularização na forma de fagemideo pBK-CMV contendo o
inserto clonado.

26
Figura 7: Ligação do cDNA no vetor ZAP Express e excisão em massa do fagemídeo pBK-CMV. O cDNA (DNA insert) é inserido unidirecionalmente no sitio múltiplo de clonagem (CMV), flanqueado pelos promotores T3 e T7, do vetor ZAP Express, entre os sítios de restrição Xho I e Eco RI e posteriormente empacotado em partículas viáveis de fago (biblioteca). A excisão do fagemideo pBK-CMV contendo inserto, bem como sua infecção em células E. coli (linhagem XLOLR), foi realizada com o auxilio de um fago helper (fago auxiliar). (Fonte: http//www.stratagene.com).

27
6.6- Preparo do estoque de bactérias recombinantes
As colônias de bactérias provenientes da excisão em massa foram coletadas usando-se
um palito e inoculadas em tubos contendo meio LB-canamicina. Procedeu-se o crescimento
em um agitador rotatório (shaker) a 37ºC por 14-16 horas. No dia seguinte, as culturas
foram transferidas para tubos de microcentrífuga (1,7ml), identificadas, estocadas em
presença de glicerol (30% v/v final) e armazenadas à temperatura de –80ºC.
6.7- Preparo do extrato livre de células
As bactérias recombinantes foram crescidas em LB-caldo contendo canamicina
(50µg/ml) e IPTG (50µg/ml) por 14-16 horas, a 37ºC, sob agitação. No dia seguinte, as
culturas foram centrifugadas em tubos de vidro por 5-8 minutos a 1.000g e o sobrenadante
foi desprezado. As células foram suspensas em 500µl de água Milli-Q e submetidas a uma
nova centrifugação. O sobrenadante foi desprezado e as células suspensas em 400µl de uma
solução de tampão carbonato (Na2CO3 15mM, NaHCO3 35mM pH 9,6) contendo PMSF
0.5mM. As células foram rompidas com pérolas de vidro (aproximadamente 1/3 do volume
de solução de tampão carbonato) com o auxílio de um agitador de tubos. Após a lise,
procedeu-se nova centrifugação por 5 minutos a 1.000g e os sobrenadantes fora transferidos
para tubos de 1,5ml, devidamente identificados com o número do clone, e estocados a
–20ºC para posterior análise (ELISA).

28
6.8- ELISA (Enzyme Linked Imunosorbent Assay)
A varredura da biblioteca de cDNA da glândula de veneno da aranha Lasiodora sp foi
realizada através da técnica de ELISA, como descrito por Kalapothakis e cols (2001).
Placas de ELISA com 96 poços foram sensibilizadas durante 16 horas com 100µl de
extratos celulares a 4ºC. No dia seguinte, as placas foram lavadas por 4 vezes com uma
solução de NaCl 0,15M - Tween 20 - 0,05%. Os poços foram bloqueados com 100µl de
solução de caseína 1% preparada em PBS 1x - Tween 20 - 0,05% e a placa mantida a 37ºC
por 1 hora. Após o bloqueio, as placas foram lavadas 4 vezes com NaCl 0,15 M - Tween 20
- 0,05%. Aos poços foram então adicionados 100µl de IgGs de coelho (anti-veneno total)
diluído 1:5000 (2µl para 10ml de PBS 1x - caseína 0,2% - Tween 20-0,05%). Incubou-se
novamente a placa, a 37ºC por 2 horas. Transcorrido o tempo de incubação, os poços foram
lavados 4 vezes com NaCl 0,15M - Tween 20 - 0,05%. Após a lavagem, adicionou-se a
cada poço 100µl de anti-IgG de coelho-peroxidase diluído 1:4000 (2,5µl em 10ml de PBS
1x -caseína 0,2% - Tween 20-0,05%). Incubou-se por mais 1 hora à temperatura de 37ºC e
em seguida lavou-se 10 vezes com NaCl 0,15M - Tween 20 - 0,05%. A cada poço foram
adicionados 100µl de OPD 0.2mg/ml solubilizado em tampão citrato (Na2HPO4 50mM,
ácido cítrico 24mM pH 5.0) mais H2O2 (30%v/v) 0,2µl/ml. A reação foi incubada na
ausência de luz por 25 minutos e em seguida, interrompida com 20µl de H2SO4 diluído
1:20. A leitura da placa foi feita a 492nm usando-se um leitor de placas de ELISA
(Molecular Devices Emax Spectrophotometer). Os clones que se mostraram positivos
foram confirmados em um segundo ensaio imunológico. Como controle negativo da reação,
nós utilizamos extratos de bactérias XLOLR não infectadas.

29
6.9- Extração do DNA plasmidial
Os clones que se mostraram positivos no teste de ELISA foram crescidos a 37ºC por
16 horas em 3 ml de meio LB-caldo contendo canamicina na concentração final de
50µg/ml. No dia seguinte, duas alíquotas de 1,5 ml de cada clone foram centrifugadas em
tubos de microcentrífuga (1,5 ml) por 3 minutos a 3000 g. O sobrenadante foi desprezado e
as células foram suspensas em 200µl de STET e 5µl de lisozima. Procedeu-se então uma
incubação por 5 minutos à temperatura ambiente e posteriormente a 95ºC por 60 segundos.
A mistura foi centrifugada por 10 minutos a 10.000g. Após a remoção dos restos celulares,
adicionou-se 8µl de CTAB e procedeu-se nova centrifugação, a 10.000g, por 10 minutos.
Após desprezar o sobrenadante, o precipitado foi suspenso em 200µl de uma solução 1,2 M
de NaCl. Os tubos foram levados rapidamente ao vortex e em seguida, adicionou-se 400µl
de fenol/clorofórmio a cada tubo. O conteúdo dos tubos foi agitado vigorosamente por 2
minutos com o auxílio de um agitador de tubos. Em seguida, os tubos foram submetidos à
centrifugação por 10 minutos a 10.000g à temperatura ambiente. Após a centrifugação, a
camada superior foi transferida para outro tubo de microcentrífuga e tratada com 5µl de
RNase por 30 minutos à temperatura ambiente. Decorridos os 30 minutos, o DNA foi
precipitado pela adição de etanol 100% (2,5 vezes o volume da solução de DNA) e
incubação por 1 ou 2 horas a –20ºC. Após a incubação, o DNA foi obtido por centrifugação
por 10 minutos, a 10.000g e o sobrenadante desprezado. O DNA foi lavado (1 ou 2 vezes)
com 400µl de etanol 70% e novamente centrifugado por 10 minutos a 10.000g. O etanol
residual foi eliminado por evaporação usando-se um bloco térmico (40ºC) ou o sistema
speed vac.

30
6.10- Preparo do gel de agarose
Pesou-se 1g de agarose e adicionou-se a 100ml de TAE 1x (40mM de Tris-acetato,
1mM de EDTA, pH 8,0 ajustado com ácido acético). A solução foi aquecida em forno de
microondas até completa solubilização da agarose.
6.11- Eletroforese do DNA em gel de agarose
Com o objetivo de quantificar o DNA plasmidial extraído, procedeu-se a eletroforese
de uma alíquota do DNA em gel de agarose 1% (p/v) durante 1 hora a 100mV. A um
volume de 3µl de DNA acrescentou-se 6µl de água e 1µl de tampão da amostra (0,25 % de
azul de bromofenol, 0,25 % de xilenocianol e 30% p/v de glicerol). Após a aplicação da
amostra e o desenvolvimento da eletroforese, o gel foi corado em solução de brometo de
etídio (0,5 mg/ml) diluído em TAE 1X, por 15 minutos. O gel foi visualizado em um
transluminador a 356 nm e fotografado por um sistema Sony (Modelo SSC-M 370CE)
acoplado a um visor e a uma impressora. No processo de quantificação utilizamos como
referência o padrão de DNA (1Kb - Invitrogen®)

31
6.12- Seqüenciamento do DNA
Os clones que se mostraram positivos após varredura utilizando a técnica de ELISA
foram seqüenciados de acordo com o método descrito por Sanger e cols. (1977) utilizando-
se do kit de seqüenciamento DYEnamic ET Dye Terminator Cycle Sequencing Kit for
Mega BACE DNA Analysis System (Amersham Biosciences). O seqüenciamento foi feito
em ambas as fitas utilizando-se os iniciadores BK reverse e o T7 primer, que se anelam no
vetor pBK-CMV.
A reação de amplificação foi preparada como descrito abaixo:
• 4µl de Mix (kit seqüenciamento)
• 1 µl de iniciador (5 pMol) direto ou reverso
• 4 µl de DNA (200ng)
• 1 µl de água
As reações foram conduzidas em um termociclador (Mastercycler, Eppendorff),
conforme o programa descrito a seguir:
1. Rápida elevação da temperatura a 95ºC
2. 95ºC por 25 segundos
3. 50ºC por 20 segundos
4. 60ºC por 1 minuto
O ciclo foi repetido por 30 vezes.
Abaixamento da temperatura a 4ºC.
As seqüências dos iniciadores utilizados nas reações de sequenciamento encontram-se
descrita abaixo.
Iniciadores Seqüências BK-reverse 5`-ACAGGAAACAGCTATGACCTTG-3`
T7 primer 3`-CGGGATATCACTCAGCATAATG-5`

32
6.13- Purificação dos produtos de extensão
Adicionou-se 80µl de isopropanol (75%) a 10µl do produto da reação de
sequenciamento. Após 15 minutos, à temperatura ambiente, procedeu-se uma centrifugação
por 30 minutos a 1.000 g e desprezou-se o sobrenadante. Em seguida, lavou-se o
precipitado com 250µl de isopropanol a 75% e procedeu-se nova centrifugação por 5
minutos a 1.000 g. Desprezou-se o sobrenadante e secou-se o precipitado à temperatura
ambiente por 1 hora. Após a secagem, suspendeu-se o DNA em 10µl de tampão da amostra
(azul de bromofenol 0.25%, xilenocianol 0.25% e glicerol 30). Uma alíquota do DNA a ser
seqüenciado foi submetida à eletroforese em gel de agarose com o objetivo de estimar a
concentração de DNA obtida. Após a quantificação do DNA, procedeu-se à análise do
DNA em um aparelho de seqüenciamento automático Mega BACE 500.
6.14- Análise das seqüências de nucleotídeos obtidas
Todas as seqüências nucleotídicas dos clones que se mostraram positivos em dois
ensaios imunológicos de ELISA foram submetidas a análises pelos programas Blastn,
Blastx, BlastP (Basic Local Alignment Search Tool – Altschul e cols., 1990 – disponível em
http//:www.ncbi.nlm.nih.gov), SIXFRAME (disponível em http//:workbench.sdsc.edu),
SignalP e ProtScale (disponíveis em http//:www.cbs.dtu.dk) .
6.15- Clonagem das seqüências codificantes para LTx1 e LTx3 em sistema de expressão bacteriano pET-11a Novagen®
O procedimento de clonagem foi desenhado com o intuito de inserir apenas a
seqüência que codifica as toxinas maduras dos clones selecionados na varredura da
biblioteca de cDNA da aranha Lasiodora sp no vetor selecionado. O sistema de expressão
pET é um poderoso sistema desenvolvido para a clonagem e expressão de proteínas
recombinantes em células E. coli. Os vetores de expressão pET apresentam um sistema de

33
indução baseado no promotor da T7 RNA polimerase, desenvolvido por Studier e cols.
(1990). Nesses vetores de expressão, o operador Lac encontra-se inserido entre o promotor
T7 e seqüências de iniciação de tradução, o que permite desrepressão, mediada por IPTG,
do promotor T7 em adição à indução mediada por IPTG da T7 polimerase do promotor lac
UV5 em cepas de bactérias DE3. As etapas da clonagem das toxinas, no sistema de
expressão pEt-11a, estão descritas a seguir.
6.15.1- Amplificação da região correspondente às toxinas maduras LTx1 e LTx3 para clonagem em sistema PCR® 2.1 TOPO TA Cloning Invitrogen®
Com o objetivo de proceder a clonagem, apenas da região que codifica as toxinas
maduras LTx1 e LTx3, no vetor de expressão bacteriano pET-11a (Novagen®), desenhamos
pares de iniciadores contendo os sítios de restrição Nde I e BamH I. As seqüências dos
iniciadores utilizados nas reações de PCR para a amplificação das toxinas maduras LTx1 e
LTx3 encontram-se descritas abaixo:
Iniciadores Seqüências Forward 5’-CCATATGTTTTTCGAATGTAC-3’
Reverse 5’-GGGATCCCTAAAACTTCAAAC-3’

34
Para amplificar a região correspondente as toxinas maduras LTx1 e LTx3,
procedeu-se a reação em cadeia da polimerase utilizando-se o DNA plasmidial extraído
previamente como molde da reação (DNA template). O volume dos reagentes utilizados na
reação de PCR está descrito a seguir.
• 1µl de DNA plasmidial
• 3.0µl de dNTP (solução estoque 2.5mM)
• 3.0µl de tampão Taq 10x
• 1.5µl de MgCl2 (25mM)
• 1.0µl de iniciador F (20 pmoles)
• 1.0µl de iniciador R (20 pmoles)
• 0.5µl de Taq DNA polimerase
• 19µl de água Milli-Q estéril
Após a adição de uma gota de óleo mineral, procedeu-se a reação, em um
termociclador, conforme o programa descrito abaixo.
1. 95ºC por 4 minutos
2. 92ºC por 1 minuto
3. 43ºC por 1 minuto
4. 72ºC por 1 minuto
5. Etapas 2 a 4 (34 vezes)
6. 72ºC por 5 minutos
7. Abaixamento da temperatura a 4ºC, até purificação.

35
6.15.2- Preparo de gel de poliacrilamida 6 %
Para o preparo de gel de poliacrilamida 6%, procedeu-se à mistura dos seguintes
reagentes.
• 1.5 ml de acrilamida 40%
• 1.0 ml de TAE 10x
• 7.5 ml de água Milli-Q
• 75µl de persulfato de amônio 10%
• 5.0µl de TEMED
6.15.3- Purificação do produto de PCR em gel de poliacrilamida
Após a reação de amplificação da região correspondente às toxinas maduras LTx1 e
LTx3, o produto de PCR foi submetido à eletroforese em gel de poliacrilamida (tampão
Tris-acetato 1X) por 50 minutos a 100 volts. O gel foi corado com brometo de etídio por 5
minutos sob baixa agitação. As bandas correspondentes às toxinas maduras LTx1 e LTx3
(aproximadamente 170 pares de bases) foram extraídas com o auxilio de uma lâmina de
bisturi e transferidas para um tubo de microcentrifuga (1.5ml) contendo 500µl de solução
TE 1X NaCl 1M. O conteúdo do tubo foi agitado com o auxilio de um agitador de tubos e
imediatamente incubado a 37ºC por 2 horas. Após incubação, procedeu-se uma
centrifugação por 10 minutos a 10.000g. O sobrenadante foi transferido para outro tubo de
microcentrifuga ao qual adicionaram-se 400µl de fenol/clorofórmio alcalino. Após agitação
do conteúdo presente no tubo com o auxílio de um agitador de tubos, procedeu-se nova
centrifugação por 10 minutos a 10.000g. A suspensão aquosa (contendo o DNA) foi
transferida para outro tubo de microcentrifuga ao qual adicionou-se 2 volumes de etanol
100% e 1µl de tRNA (concentração estoque 10mg/ml). O conteúdo do tubo foi mantido a -
80ºC por 40 minutos. Após a precipitação, procedeu-se nova centrifugação por 15 minutos

36
a 10.000g. Descartou-se o sobrenadante e adicionou-se 400µl de etanol 70%. Procedeu-se
nova centrifugação por 10 minutos a 10.000g. O sobrenadante foi desprezado e o DNA foi
mantido a 65ºC por 15 minutos em um bloco térmico. Após a secagem, o DNA foi
suspenso em 20µl de água Milli-Q estéril.
6.15.4- Clonagem das seqüências codificantes para LTx1 e LTx3 em PCR® 2.1-TOPO TA Cloning Invitrogen®
A clonagem dos fragmentos (produtos de PCR) que codificam as toxinas maduras
LTx1 e LTx3 em sistema PCR® 2.1-TOPO TA Cloning Invitrogen® foi realizada como uma
estratégia para facilitar a clonagem das toxinas maduras no vetore pET-11a. A figura 8
ilustra o mapa do vetor PCR® 2.1 TOPO TA Cloning Invitrogen® utilizado.
Após a purificação dos produtos de PCR, correspondentes aos fragmentos que
codificam as toxinas maduras (LTx1 e LTx3), como descrito em 6.15.3, os mesmos foram
inseridos no vetor PCR® 2.1-TOPO® conforme protocolo descrito a seguir:
• 2µl de DNA (produto de PCR)
• 1µl de salt solution
• 1µl de vetor TOPO®
• 2µl de água milli-Q estéril
A reação foi mantida à temperatura ambiente por 10 minutos.

37
Figura 8: Mapa do vetor de clonagem PCR® 2.1-TOPO TA Cloning Invitrogen® (Disponível em: www.invitrogen.com)

38
6.15.5- Preparo de células E. coli Top 10 F’competentes (Método CaCl2)
Células de E. coli, linhagem TOP 10 F’, foram pré-inoculadas em 3ml de meio LB
caldo contendo estreptomicina (10µg/ml) e tetraciclina (10µg/ml). As células foram
crescidas sob agitação constante (200 rpm), por 14-16 horas, à temperatura de 37ºC. Após o
crescimento, as bactérias foram transferidas para um erlenmeyer contendo 100µl de LB
sem antibiótico. Procedeu-se nova incubação a 37ºC, 200 rpm por um período de 2-3 horas
até que as células atingissem uma densidade ótica entre 0.6 e 0.8. Atingida a densidade
ótica ótima, as bactérias foram submetidas à centrifugação por 8 minutos a 1.000g em
centrifuga refrigerada (4ºC). O sobrenadante foi desprezado e as células suspensas em 40
ml de CaCl2 0.1M gelado. Incubou-se em gelo por pelo menos 1 hora. Após incubação,
procedeu-se nova centrifugação por 5 minutos a 1.000g em centrifuga refrigerada.
Desprezou-se o sobrenadante, e as células foram suspensas em 1ml de CaCl2 0.1M.
Alíquotas de 100µl foram estocadas em tubos de microcentrífuga, contendo 30% de
glicerol, e mantidas a temperatura de -80ºC.
6.15.6. Transformação de células de E. coli Top 10 F’ com o produto de ligação em PCR® 2.1-TOPO®
Células de E. coli (linhagem Top 10 F’) competentes foram descongeladas e
mantidas em gelo. Adicionou-se 6µl do produto de ligação (PCR® 2.1-TOPO® mais
fragmentos) ao tubo contendo 100µl de células competentes. Incubou-se em gelo por 15-30
minutos. Após a incubação, a mistura foi submetida a um choque térmico a temperatura de
42ºC por 2 minutos. Imediatamente após o choque térmico, um volume de 1 ml de meio LB
foi adicionado à mistura. Procedeu-se nova incubação por 50 minutos a temperatura de
37ºC. Após incubação, procedeu-se uma centrifugação por 4 minutos a 3.000g. O excesso
de sobrenadante foi desprezado e as células foram suspensas em aproximadamente 100µl

39
de meio LB restante no tubo. As células foram semeadas em placas contendo meio LB
ampicilina com o auxilio de pérolas de vidro e incubadas a 37ºC por 14-16 horas.
6.15.7- Reação em cadeia da polimerase de colônias provenientes da transformação de E. coli Top 10 F’ com o produto de ligação em PCR® 2.1-TOPO®
Colônias de bactérias provenientes da transformação foram submetidas à reação em
cadeia da polimerase para confirmar a presença dos fragmentos que codificam as toxinas
maduras LTx1 e LTx3 no vetor PCR® 2.1-TOPO®. Os iniciadores utilizados na reação
foram:
Iniciadores Seqüências M13 Reverse Primer 5’-GTCCTTTGTCGATACTG-3’
M13 Forward Primer 5’-GACCGGCAGCAAAATG-3’
Para amplificar a região correspondente às toxinas maduras LTx1 e LTx3,
procedeu-se a reação em cadeia da polimerase utilizando-se 0.5µl de cultura dos clones
provenientes da transformação como molde da reação (DNA template). O sistema da
reação de PCR encontra-se descrito abaixo:
• 0.5µl da cultura (clone)
• 3.0µl de dNTP (solução estoque 2.5mM)
• 3.0µl de tampão Taq 10x
• 1.5µl de MgCl2 (25mM)
• 1.0µl de iniciador F (20 pmoles)
• 1.0µl de iniciador R (20pmoles)
• 0.5µl de Taq DNA polimerase
• 19.5µl de água Milli-Q estéril

40
Após a adição de uma gota de óleo mineral, procedeu-se a reação em um
termociclador, conforme o programa descrito abaixo.
1. 95ºC por 4 minutos
2. 92ºC por 1 minuto
3. 43ºC por 1 minuto
4. 72ºC por 1 minuto
5. Etapas 2 a 4 (34 vezes)
6. 72ºC por 5 minutos
7. Abaixamento da temperatura a 4ºC, até purificação.
6.15.8- Extração do DNA plasmidial de colônias provenientes da transformação de E. coli Top 10 F’ com o produto de ligação em PCR® 2.1-TOPO®
As colônias que se mostram positivas na reação de PCR (apresentaram uma
amplificação de aproximadamente 374 pb) foram inoculadas em 5 ml de meio LB caldo
contendo ampicilina na concentração de 10µg/ml e incubadas por 14-16 horas a
temperatura de 37ºC sob agitação de 200 rpm. A extração do plasmídeo PCR® 2.1-TOPO®
contendo inserto foi realizada utilizando-se o kit de extração de DNA plasmidial Wizard
Plus SV DNA Minipreps Promega®.

41
6.15.9- Digestão dos plasmídeos PCR® 2.1-TOPO®-LTx1 (e LTx3) com as enzimas Nde I e BamH I para clonagem do fragmento liberado no sistema de expressão pET-11a (Novagen®)
Vetores PCR® 2.1-TOPO® contendo insertos que codificam para as toxinas LTx1 e
LTx3 foram digeridos com as enzimas de restrição Nde I e Bam H I para a posterior
clonagem do produto de digestão no sistema de expressão pET-11a, conforme protocolo a
seguir.
• 7.0µl de DNA plasmidial
• 3.0µl de tampão K (Amersham)
• 0.3µl BSA
• 0.7µl de Nde I
• 1.0µl de BamH I
• 18.0µl de água Milli-Q estéril
Incubação por 2 horas a 37ºC.
6.15.10- Purificação dos produtos de digestão
Os produtos de digestão foram submetidos à eletroforese em gel de poliacrilamida
por 50 minutos a 100 volts com o objetivo de purificar as bandas correspondentes aos
insertos que codificam as toxinas maduras LTx1 e LTx3 (aproximadamente 170 pb). A
purificação da banda de interesse foi conduzida conforme protocolo descrito em 6.15.3.

42
6.15.11- Digestão do vetores de expressão pET-11a e pET-21b com as enzimas de restrição Nde I e Bam H I
Vetor pET-11a (Novagen®) (Figura 9) obtido a partir de mini-preparação de DNA
plasmidial como descrito na secção 6.9, foi digerido com as enzimas de restrição Nde I e
Bam H I para a posterior clonagem dos fragmentos que codificam as toxinas maduras LTx1
e LTx3 seguindo o protocolo descrito abaixo.
• 7.0µl de pET-11a
• 3.0µl de tampão K (Amersham)
• 0.3µl BSA
• 0.7µl de Nde I
• 1.0µl de BamH I
• 18.0µl de água Milli-Q estéril
A mistura de reação foi incubada por 2 horas a 37ºC. Após a digestão, procedeu-se a
defosforilação do vetor com o objetivo de evitar a sua recircularização. Ao produto de
digestão, adicionou-se 3µl de tampão e 0.5µl de CIP (calf intestinal phosphatase). A
mistura foi incubada a 37ºC por 30 minutos. A inativação da enzima procedeu-se a 75ºC
por 10 minutos.
6.15.12- Purificação do vetor de expressão utilizando papel
Whatman DE81
Após a digestão do vetor de expressão pelas enzimas de restrição, todo o volume da
reação foi aplicado em gel de agarose 1% e submetido à eletroforese por 1 hora e 120 volts.
O gel foi corado com brometo de etídio e as bandas visualizadas em luz ultravioleta com o
auxílio de um transiluminador. Com o auxílio de uma lâmina de bisturi, procedeu-se um
corte no gel à frente da banda correspondente ao DNA digerido. Introduziu-se no corte
meio disco de papel Whatman DE81. O gel foi novamente submetido à eletroforese por 10
minutos e 120 volts para propiciar a ligação do DNA ao papel Whatman DE81. O gel foi

43
novamente levado ao transluminador para certificar que todo o DNA havia se ligado ao
papel. Com o auxílio de uma pinça, o papel foi transferido para um tubo de microcentrífuga
(1,5 ml) ao qual adicionou-se 500µl de TE 1X NaCl 1M. Agitou-se por 30 segundos com o
auxílio de um agitador de tubos e incubou-se por 2 horas em estufa a 37ºC. Após
incubação, centrifugou-se por 5 minutos a 10.000g e o sobrenadante foi transferido para um
novo tubo de 1,5ml. Adicionou-se 400µl de fenol/clorofórmio alcalino e o conteúdo do
tubo foi agitado vigorosamente com o auxílio de um agitador de tubos. Procedeu-se nova
centrifugação por 10 minutos a 10.000g. Após a centrifugação, o sobrenadante (fase
aquosa) foi transferido para outro tubo de microcentrífuga ao qual adicionou-se 2 volumes
de etanol 100%. O tubo foi mantido por 40 minutos à temperatura de -80ºC para permitir a
precipitação do DNA. Após a precipitação do DNA, procedeu-se nova centrifugação por
15 minutos a 10.000g. Desprezou-se o sobrenadante e adicionou-se 400µl de etanol 70%. O
tubo contendo o DNA foi submetido a uma nova centrifugação por 10 minutos a 10.000g.
Após desprezar o sobrenadante, procedeu-se a secagem do DNA em um speed-vac. O DNA
foi suspenso em 20µl de água Milli-Q estéril e estocado a -80ºC.

44
6.15.13- Clonagem dos fragmentos que codificam as toxinas LTx1 e LTx3 no vetor de expressão pET-11a
Após a digestão dos plasmídeos PCR® 2.1-TOPO®, contendo as seqüências
codificantes para LTx1 e LTx3, e dos vetores de expressão pET-11a (Figura 9) com as
enzimas de restrição Nde I e BamH I, ambos os fragmentos foram submetidos a reação de
ligação no vetor de expressão utilizando-se a enzima T4 DNA ligase, conforme o protocolo
descrito abaixo.
• 3µl de DNA vetor pET-11a
• 5µl DNA inserto
• 0.7µl de T4 DNA ligase Promega®
• 10µl de Tampão da ligase 2x
• 1.3µl de água Milli-Q estéril
• Incubação por 14-16 horas a temperatura de 16ºC em um bloco térmico
(Termomixer Eppendorff®) mantido a 4ºC em câmara fria.
45
Figura 9: Mapa esquemático do vetor de expressão bacteriano pET-11a Novagen®. O sistema pET-11a possui um sistema de detecção e purificação da proteína recombinante baseado em uma pequena seqüência T7-Tag.

46
6.15.14- Transformação de células de E. coli Top 10 F’ com o produto de ligação no vetor de expressão pET-11a
Após a ligação dos fragmentos que codificam para as toxinas maduras LTx1 e LTx3
no vetor de expressão pET-11a, como descrito em 6.15.13, procedeu-se a transformação de
células de E. coli Top 10 F’ utilizando-se todo o produto das ligações. Os demais passos da
transformação foram conduzidos conforme descrito na secção 6.15.6.
6.15.15- Reação em cadeia da polimerase de colônias provenientes da transformação de células de E. coli Top 10 F’ com os produtos de ligação no vetor de expressão pET-11a
Colônias de bactérias provenientes da transformação descrita em 6.15.14 foram
submetidas à reação em cadeia da polimerase para confirmar a presença dos fragmentos
que codificam as toxinas maduras LTx1 e LTx3 no vetor de expressão pET-11a. Os
iniciadores utilizados na reação foram.
Iniciadores Seqüências PET-F PET-R
3’-TATAGGGGAATTGTGAGCGG-5’ 3’-CTTTCGGGCTTTGTTAGCAG-5’

47
Para amplificar a região correspondente às toxinas maduras LTx1 e LTx3,
procedeu-se a reação em cadeia da polimerase utilizando-se 0.5µl de cultura dos clones
provenientes da transformação como molde da reação. O sistema da reação de PCR
encontra-se descrito abaixo:
• 0.5µl de cultura
• 3.0µl de dNTP (solução estoque 2.5mM)
• 3.0µl de tampão Taq 10x
• 1.5µl de MgCl2 (25mM)
• 1.0µl de iniciador F (20 pmoles)
• 1.0µl de iniciador R (20 pmoles)
• 0.5µl de Taq DNA polimerase
• 19.5µl de água Milli-Q estéril
Após a adição de uma gota de óleo mineral, procedeu-se a reação, em um
termociclador, conforme o programa descrito abaixo.
1. 95ºC por 4 minutos
2. 92ºC por 1 minuto
3. 50ºC por 1 minuto
4. 72ºC por 1 minuto
5. Etapas 2 a 4 (34 vezes)
6. 72ºC por 5 minutos
Abaixamento da temperatura a 4ºC, até purificação.

48
6.15.16- Transformação de bactérias de expressão (E. coli linhagem BL-21 DE3) com a construção pET-11a-LTx1 e pET-11a-LTx3
Células competentes E. coli linhagem BL-21 (DE3) foram descongeladas em gelo e
transformadas com as construções pET-11a-LTx1 e pET11a-LTx3, como descrito em
6.15.6.
6.16- Indução da expressão da toxina recombinante LTx1 clonada no vetor de expressão pET-11a
Um volume de 20µl de células E. coli linhagem BL-21 transformada com a
construção pET-11a-LTx1 foi inoculado em 4ml de meio LB-caldo contendo ampicilina
(100µg/ml) e incubado a 37ºC, 200 rpm por 14-16 horas. No dia seguinte, o pré-inóculo foi
vertido em 100ml de meio LB-caldo contendo ampicilina (100µg/ml) e incubado a 30ºC,
200 rpm, até que a DO600 atingisse um valor entre 0,6-0,8. Ao atingir a DO600 desejada,
uma alíquota de 100µl de células (correspondente ao tempo zero de indução) foi
centrifugada em uma microcentrífuga por 2 minutos a 10.000g. Descartou-se o
sobrenadante e as células foram suspensas em 100µl de tampão da amostra (SDS-PAGE
2X) contendo agente redutor para posterior análise em gel SDS-PAGE. A expressão da
LTx1 recombinante foi induzida pela adição de IPTG (0,6mM concentração final) à cultura
e incubação à temperatura de 30ºC sob agitação (200 rpm) por 5 horas. Após diferentes
tempos de indução (1h, 2h, 3h, 4h e 5h), alíquotas de 100µl de células foram centrifugadas
e tratadas com o tampão da amostra (SDS-PAGE 2X) como descrito acima. A
quantificação da proteína de cada alíquota foi realizada pelo método de Bradford. As
alíquotas suspensas em tampão da amostra (SDS-PAGE 2X) foram submetidas à
temperatura de 95ºC por 5 minutos e um volume contendo 20µg de proteína foi analisado
em gel SDS-PAGE.

49
6.16.1- Produção da toxina LTx1 recombinante e solubilidade da proteína
Após 5 horas de indução (como descrito em 6.16), a cultura (100ml) foi
centrifugada por 15 minutos, 4ºC, 1.000g. O sedimento celular foi suspenso em 10ml de
solução de lise (50mM Tris pH 8.0, 300mM NaCl e 10mM imidazol) contendo 80µl de
lisozima (solução estoque 50mg/ml), 1µl de aprotinina (solução estoque 10mg/ml), 1µl de
pepstatina (solução estoque 10mg/ml) e 200µl de PMSF (solução estoque 50mM) e
mantido em gelo por 30 minutos. Após incubação, as células foram submetidas à sonicação
(200 a 300 w) por 6 vezes com pulsos de 10 segundos e intervalos de 10 segundos em gelo.
Após a sonicação, adicionou-se à solução 0,1% de Triton X100. As células sonicadas foram
submetidas à centrifugação por 30 minutos, 1.000g, a 4ºC. O sobrenadante foi transferido
para um tubo falcon (15ml) e concentrado para análise em SDS-PAGE.
Para testar a produção da proteína sob a forma de corpos de inclusão, o sedimento
proveniente da sonicação da cultura de células induzidas por 5 horas foi submetido a 2
lavagens (20ml cada) com tampão de ligação 1 (50mM Tris, 500mM NaCl, 10mM
mercaptoetanol e 2M uréia). Após as lavagens e centrifugação por 5 minutos, a 4ºC,
1.000g, o precipitado foi suspenso, com o auxílio de uma pipeta, em 375 µl de tampão X
(50mM Tris, 500mM NaCl, 10mM β-mercaptoetanol e 8M uréia). Um volume de 35ml de
tampão de ligação 2 pH 8.0 (50mM Tris, 500mM NaCl e 5mM imidazol) foi
vagarosamente adicionado ao tubo contendo o precipitado suspenso em tampão X e
incubado por 24 horas à temperatura de 4ºC para permitir a renaturação das proteínas
eventualmente presentes nos corpos de inclusão. No dia seguinte, procedeu-se nova
centrifugação por 15 minutos, 1.000g, a 4ºC. O sobrenadante (35ml) foi concentrado para
análise posterior.

50
6.16.2- Eletroforese em gel de poliacrilamida SDS-PAGE
Para a eletroforese das frações contendo, eventualmente, a toxina LTx1
recombinante, adotou-se o sistema de gel desnaturante (SDS-PAGE) descrito por Laemmli
e cols. (1970). As concentrações dos géis foram de 4% para o gel de concentração e 18%
para o gel de separação. As amostras (30µg) foram tratadas com o tampão da amostra SDS-
PAGE 2X contendo o agente redutor β-mercaptoetanol, fervidas durante 5 minutos, e
aplicadas no gel. A eletroforese procedeu-se verticalmente a 100V, em tampão Tris 0,025M
pH 8,3 contendo 0,96M de glicina e 0,1% de SDS. Após a eletroforese o gel foi revelado
em solução de nitrato de prata. As soluções usadas encontram-se descritas no final deste
capítulo.
6.16.3- Coloração pelo método da prata
Após a eletroforese, o gel foi transferido para uma cuba contendo solução fixadora e
procedeu-se a incubação por 1 hora. Em seguida o gel foi lavado com etanol 50% (solução
de lavagem), por 3 vezes, por 20 minutos. Após as lavagens, o gel foi tratado com uma
solução de tiossulfato de sódio (0,4g/ml) por 2 minutos. Em seguida, procedeu-se nova
lavagem com água destilada, por 3 vezes, 20 minutos. Após as lavagens, procedeu-se a
incubação do gel por 20 minutos em solução de impregnação. Após a impregnação, o gel
foi incubado em solução de desenvolvimento até o aparecimento das bandas. A reação foi
interrompida pela adição da solução de parada. A composição das soluções utilizadas nesse
método de coloração encontra-se no final deste capítulo.

51
6.16.4- Western Blot
As proteínas separadas eletroforeticamente em gel de poliacrilamida-SDS 18%
foram transferidas para um filtro de nitrocelulose (Amersham, poros de 0,45 µm), com o
auxílio de um sistema de transferência Pharmacia LKB-PhastSystem, por 40 minutos, 5 v,
50mA. O sistema foi montado fazendo-se um “sanduíche” com 3 folhas de papel de filtro
3mm, 1 folha de membrana de nitrocelulose, o gel contendo amostras a serem transferidas e
mais 3 folhas de papel de filtro 3mm, todos embebidos no tampão de transferência contínua
descrito no manual. Após a transferência, a membrana foi bloqueada, durante 1 hora, com
leite em pó 5% em 0,05% Tween/PBS (PBS-T). Após o bloqueio a membrana foi lavada 3
vezes com PBS-T por 10 minutos e incubada com antisoro policlonal anti-veneno total
diluído 1:2000 em leite em pó 5% PBS-T por 1 hora. Procedeu-se nova lavagem como
descrito acima. A membrana foi incubada com anti-IgG de coelho-peroxidase diluído
1:20000 em leite em pó 5% PBS-T, lavada conforme descrito acima e revelada com ECL
reagente (Amersham).

52
6.17- Genótipos das linhagens de bactérias hospedeiras utilizadas neste trabalho
Linhagem XL1-Blue MRF`: ∆(mcrA)183 ∆(mcrCB-hsdSMR-mrr) 173 endA1 supE44
thi-1 recA1 gyrA96 relA1 lac[F`proAB laclqZ∆M15 Tn10(Tetr)]
Linhagem XLOLR: ∆(mcrA)183 ∆(mcrCB-hsdSMR-mrr) 173 endA1 thi-1 recA1 gyrA96
relA1 lac[F`proAB laclqZ∆M15 Tn10(Tetr)] Su- (nonsuppressing) λr (lambida resistent).
Linhagem TOP 10F’: F’, mcrA, (mrr-hsdRMS-mcrBC), Φ80∆lac∆-M15, ∆lacX74, deo R,
recA1, ara∆139, ∆ (ara, leu), 7697, galU, galK, λ-, rslp,endA1, nupG.
Linhagem BL-21 (DE3): F-ompT hsdSb(r-B m-B) gal dcm.
6.18- Meios de cultura
LB Broth: 10g de NaCl, 10g de bactotriptona e 5g de extrato de levedura. O pH foi
ajustado para 7,5 com NaOH e o volume acertado para 1 litro. Autoclavar a solução.
LB-ágar: Possui a mesma composição do LB Broth, acrescido de 20g de agar por litro de
solução. Autoclavar a solução.
LB-ágar canamicina: Possui a mesma composição do LB-ágar acrescido de kanamicina na
concentração de 50µg/ml.
LB-ágar tetraciclina: Possui a mesma composição do LB-ágar acrescido de tetraciclina na
concentração final de 12,5µg/ml.
LB top ágar: Possui a mesma composição do LB-broth acrescido de agarose 0,7% (w/v).
LB XIA: LB agar contendo 20µg/ml de ampicilina e 1mM de IPTG

53
2xYP: 16g de bactopeptona, 10g de extrato de levedura, ajustar o pH para 7.5 e o volume
para 1 litro. Autoclavar a solução.
6.19- Reagentes e material de uso
Tetraciclina: A tetraciclina foi dissolvida em H2O estéril na concentração de 15mg/mL e
estocada a –20ºC. A solução foi utilizada na concentração final de 12,5 µg/mL.
Canamicina: A canamicina foi dissolvida em H2O estéril na concentração de 50mg/mL e
estocada a –20ºC. A solução foi utilizada na concentração final de 100µg/mL.
Ampicilina: A ampicilina foi dissolvida em água deionizada estéril na concentração final
de 100mg/ml (solução estoque 1.000X concentrada) e estocada a -20ºC.
Estreptomicina: A estreptomicina foi dissolvida em água deionizada estéril na
concentração de 20mg/ml (solução estoque 1.000X concentrada) e estocada a -20ºC.
IPTG: O IPTG foi dissolvido em água Milli-Q estéril na concentração de 500mM e
estocado a -20ºC.
Maltose: Dissolveu-se 20g de maltose em 100 ml de água (solução 20%). Autoclavar a
solução.
PMSF: O PMSF foi dissolvido em isopropanol na concentração final de 100mM e estocado
a 4ºC
X-Gal: O X-Gal foi dissolvido em dimetilformamida na concentração de 20mg/ml e
estocado a –20ºC.
Β-Mercaptoetanol 700mM: 5 µl de β-mercaptoetanol 14 M e 95 µl de água deionizada.

54
6.20- Soluções
TAE (Tampão Tris-acetato): Solução estoque 50X: 242g de Tris, 57,1mL de ácido
acético glacial e 100mL de EDTA, pH 8,0. Completar o volume para 1 litro.
TE (Tampão Tris-EDTA): Solução estoque Tris-HCl 1M, pH 8,0 e EDTA 0,5M (pH 8,0).
Juntar as soluções estoque nas concentrações Tris-HCl 10mM pH 8,0 e EDTA 1mM pH 8,0
de acordo com o volume a ser preparado. Autoclavar e guardar à temperatura ambiente.
Solução de MgCl2: Solução preparada em duas concentrações, 100mM e 10mM.
10x sample buffer: Tris-HCl 10mM (pH 7.5), EDTA 1mM, NaCl 5M.
Corante: 0.25% de azul de bromofenol, 0.25% de xilenocianol FF e 30% de glicerol.
STET: 8% p/v de sacarose, 0.1% v/v de Triton X-100, 50mM de EDTA, 50mM de Tris-
HCl. Ajustar o pH para 8,0.
Lisozima: A lisozima foi diluída em água deionizada para uma concentração de 50mg/ml.
CTAB: Cetil trimetil brometo de amônio diluído em água deionizada para uma
concentração de 50mg/ml.
Solução saturada de fenol/clorofórmio: 500g de fenol (Sigma-P1037) são aquecidas
(68ºC), até a completa dissolução. Hidroxiquinolina é adicionada na concentração de 0,1%.
Ao fenol dissolvido adiciona-se 1 volume de tampão Tris-HCl 0,5M, pH 8,0. A mistura é
mantida sob agitação por 15 minutos e depois em repouso até ocorrer a separação completa
das fases. A fase aquosa é então removida e adiciona-se 1 volume de solução Tris-HCL
0,1M, pH 8,0 repetindo o procedimento acima, até obtenção da solução final com pH> 7,8.
Após o equilíbrio da solução e remoção da fase aquosa, adiciona-se 0,1 volume de Tris-HCl
0,1M, pH 8,0 contendo 0,2% de β-mercaptoetanol. A está mistura adicionou-se igual
volume de clorofórmio (MERCK). Armazenar a 4ºC, protegido da luz.

55
6.20.1- Soluções utilizadas na ELISA
PBS pH 7,4: 0,05M Na2HPO4 e 0,15M de NaCl.
Tampão de ligação: Na2CO3 1,59g, NaHCO3 2,93g pH 9,6. Diluir em água q.s.p 1L.
Estocar a 4ºC.
Solução de bloqueio: Caseína 2%, diluir em PBS 1X. Estocar a –20ºC.
Tampão de incubação: Caseína 0,25%, Tween20 0,05%, diluir em PBS 1X. Estocar a –
20ºC.
Solução de lavagem: NaCl 9,0g, Tween 20 0,5ml, diluir em água q.s.p. 1L. Estocar a 4ºC.
Solução reveladora: 10ml de tampão citrato, 4mg de OPD, 2µl de água oxigenada 30%.

56
6.20.2- Soluções utilizadas no gel SDS-PAGE
Solução A: 39g de acrilamida, 1g de bisacrilamida, água q.s.p. 100ml. Filtrar a solução e
armazenar em frasco escuro a 4ºC.
Solução B: 18,15g de Tris, ajustar o pH para 8,8 com HCl concentrado, água deionizada
q.s.p. 100ml. Armazenar a 4ºC.
Solução C: 6g de Tris, ajustar o pH para 6,8 com HCl concentrado, água deionizada q.s.p.
para 100ml. Armazenar a 4ºC.
SDS 10%: 10g de SDS diluídos em água deionizada q.s.p. 100ml. Armazenar à
temperatura ambiente.
PSA 10%: Persulfato de amônio dissolvido em água deionizada na concentração de
0,1g/ml.
Tampão da amostra (sem agente redutor – 2X concentrado): 1ml de solução C, 0.8 ml de
glicerol, 1.6ml de SDS 10%, 0.2ml de azul de bromofenol 0.2%, 0.4ml de água deionizada.
Armazenar à temperatura ambiente.
Tampão da amostra (com agente redutor – 2X concentrado): Substituir a água do tampão
sem agente redutor por 0,4ml de β-mercaptoetanol.
Tampão de corrida (5X concentrado): 4,5g de Tris (0,125M), 21,6g de glicina (0,96M),
1,5g de SDS (0,5%), água deionizada q.s.p. 300ml. Acertar o pH para 8,3. Armazenar a
4ºC.

57
6.20.3- Coloração pelo método da prata
Solução fixadora: 50ml de metanol, 12ml de ácido acético, água deionizada q.s.p. 100ml.
Solução de lavagem: 50ml de etanol e água deionizada q.s.p. 100ml.
Solução de tratamento: tiossulfato de sódio diluído em água na concentração de 0,4g/l.
Solução de impregnação: 0,1g de prata, 75µl de formaldeído e água deionizada q.s.p.
100ml.
Solução de desenvolvimento: 6g de carbonato de cálcio, 50µl de formaldeído, 2ml da
solução de pré-tratamento e água deionizada q.s.p. 100ml. Preparar a solução no momento
do uso.
Solução de parada: 50ml de etanol, 12ml de ácido acético e água deionizada q.s.p. 100ml.

60
7- Resultados
7.1- Varredura dos clones provenientes das excisões em larga-escala
Neste trabalho, realizou-se a varredura de aproximadamente 500 clones
provenientes de excisões em massa de três diferentes sub-bibliotecas. Para a realização da
varredura utilizou-se a técnica de ELISA (Enzyme Linked Imunosorbent Assay) conforme
descrito em materiais e métodos. O anticorpo primário foi obtido a partir da imunização de
coelhos com o veneno total da aranha Lasiodora sp. A reação foi revelada utilizando-se de
o-fenilenodiamino (OPD).
Um total de 43 clones mostrou-se positivo em dois ensaios imunológicos. Dezenove
clones apresentaram similaridade com toxinas, 2 clones apresentaram similaridade com
outras proteínas e 22 clones não apresentaram similaridade significativa com proteínas
depositadas no banco de dados quando suas seqüências nucleotídicas traduzidas foram
submetidas a análise pelo programa BlastP (tabela 1).
A reação de seqüenciamento dos insertos clonados nos plasmídeos e a posterior
análise de suas seqüências nucleotídicas revelou que alguns cDNAs apresentam
similaridade com seqüências de proteínas depositadas no GenBank, que vão desde
peptídeos tóxicos como BsTx, ESTx, HwTx-II, Magi 7 e LpTx (1 e 2) das aranhas
Brachypelma smithii, Eurypelma californicum, Selenocosmia huwena (atualmente
conhecida como Ornithoctonus huwena), Macrothele gigas e Lasiodora parahibana,
respectivamente, assim como similaridade com algumas proteínas como a cadeia beta da
hemoglobina de camundongo (Mus musculus). Todas as seqüências nucleotídicas dos
clones que se mostraram positivos em dois ensaios imunológicos de ELISA foram
submetidas a análises pelos programas Blastn, Blastx e BlastP (Basic Local Alignment
Search Tool – Altschul e cols., 1990 – disponível em http://www.ncbi.nlm.nih.gov). A
análise pelo Blastn foi realizada com o objetivo de se detectar a presença de seqüências de
vetor de clonagem (vetor pBK-CMV). Após a eliminação dessas seqüências, apenas a
seqüência correspondente ao inserto clonado foi submetida à análise pelo Blastx, que
compara uma determinada seqüência nucleotídica, traduzida em suas seis possíveis janelas
de leitura (frames), com seqüências depositadas no GenBank. A análise inicial das

61
seqüências utilizando o programa BlastX faz-se necessária porque essa análise permite a
detecção de similaridades mesmo quando as seqüências nucleotídicas são de baixa
qualidade. Após a análise pelo programa BlastX, as seqüências dos cDNAs foram
traduzidas em suas seis possíveis janelas de leitura utilizando o programa SIXFRAME
(disponível em http://workbench.sdsc.edu). As seqüências peptídicas obtidas a partir da
tradução das seqüências de cDNAs dos clones positivos foram submetidas à análise pelo
BlastP. A análise pelo programa BlastP compara uma seqüência peptídica com outras
seqüências depositadas no banco de dados de proteínas.
As seqüências peptídicas que apresentaram similaridades com toxinas de outras
aranhas depositadas no banco de dados foram designadas pela sigla LTx (toxina de
Lasiodora sp). Os clones cuja seqüência nucleotídica apresentou similaridades com outras
proteínas ou que não apresentou similaridades com proteínas depositadas no GenBank mas
apresentou pelo menos uma matriz aberta de leitura (ORF), foram designados pela sigla
LsC (componente clonado da glândula de veneno de Lasiodora sp). A tabela 1 apresenta os
resultados obtidos na varredura utilizando o teste de ELISA bem como o resultado da
análise de similaridade (BlastP).

62
Tabela 1: Resultados do teste de ELISA e similaridade utilizando o programa BlastP.
Clone Similar a Identidade Organismo Referências
1 a 15* BsTx 80% Brachypelma smithii Kaiser e cols., 1994
ESTx 80% Eurypelma californicum Savel-Niemann e cols.,
1989
LpTx1/LpTx2 95-100% Lasiodora parahibana Escoubas e cols., 1997
HwTxI/HwTxII 69% Selenocosmia huwena Shu & Liang, 1999
16 a 19* Magi 7 32% Macrothele gigas Satake e cols., 2004
20 Cadeia β-1 de hemoglobina 98% Mus musculus
Cadeia beta-2 de globina 92% Rattus norvegicus
Hemoglobina quimérica
humano/camundongo(zeta
2/beta 2) 100%
21 CG7580-PA 62% Drosophila
melanogaster
Proteína agCP10691 58% Anopheles gambiae
Proteína Qpc-pending 46% Mus musculus
22 a 43 Sem similaridade
significativa
* Clones 1-12, 13, 14-15 e 16-19 correspondem, respectivamente, às toxinas LTx1, LTx2, LTx3 e LTx5.
7.2- Análise das seqüências nucleotídicas dos cDNAs dos clones que apresentaram similaridade com toxinas de aranhas
Dos 19 clones que apresentaram similaridade com peptídeos tóxicos de outras
aranhas, 15 codificam toxinas pertencentes a uma mesma família (LTx1, LTx2 e LTx3),
sendo que 12 clones codificam a mesma toxina, denominada nesse trabalho LTx1. O clone
que codifica a toxina LTx2 foi encontrado apenas uma vez e dois clones encontrados
codificam a toxina LTx3.
A análise das seqüências dos cDNAs desses 15 clones (utilizando o programa
SignalP – disponível em http://www.cbs.dtu.dk/services/signalp/) revelou a presença de um
peptídeo sinal, um peptídeo intermediário e um peptídeo maduro compostos
respectivamente por 23, 27 e 49 resíduos de aminoácidos.
A figura 10 apresenta apenas as seqüências nucleotídicas alinhadas dos cDNAs que
codificam para as toxinas LTx1, LTx2 e LTx3.

63
1 60
LTx1 ggcaccag------c------------------ttcgtagtatctgaactt-caggagaa
LTx2 ggcaccag------cgcaaatagcagatttcattccgtagtatctgaactt-caggagaa
LTx3 ggcacgagtattttcgcaaataccgagatttcattcgtagtatctgaactttcaggagaa
***** ** * * **************** ********
61 120
LTx1 tttcaagcaaaagATGAGATCTTTGACGTTGGCTGCTCTTCTGCTATGCAGCTTGCTGTT LTx2 tttcaagcaaaagATGAGATCTTTGACGTTGGCTGCTCTTCTGCTATGCAGCTTGCTGTT LTx3 tttcaagcaaaagATGAGATCTTTGACGTTGGCTGCTCTTCTGCTATGCAGCTTGCTGTT *************...********************************************
121 180
LTx1 AGTTTTTCACACTTCAGCAGCTGCAGAGCTTGAAGCTCAGGAAGGCCATCTGATGATCCC LTx2 AGTTTTTCACACTTCAGCAGCTGAAGAGCTTCAAGCTCAGGAAGGCCATCTGATGATCCC LTx3 AGTTTTTCACACTTCAGCAGCTGCAGAGCTTGAAGCTCAGGAAGGCCATCTGATGATCCC *********************** ******* ****************************
181 240
LTx1 CGGTGATACTGACACCGCCCTAGAAACTGTCGATGATGAAAGATTTTTCGAATGTACTTT LTx2 CGGTGATACTGACACCGCACTAGAAACTGTCGATGATGAAAGACTTTTCGAATGTACTTT LTx3 CGGTGATACTGACACCGCCCTAGAAACTGTCGATGATGAAAGATTTTTCGAATGTACTTT ****************** ************************ ****************
241 300
LTx1 TGAATGCGACATTAAAAAAGAAGGCAAACCATGCAAACCCAAGGGATGCAAGTGTAAGGA LTx2 TGAATGCGACATTAAAAAAGAAGGCAAACCATGCAAACCCAAGGGATGCAAGTGTGACGA LTx3 TGAATGCGACATTAAAAAAGAAGGCAAACCATGCAAACCCAAGGGATGCAAGTGTGACGA ******************************************************* * **
301 360
LTx1 CAAGGATAACAAGGATCACAAGAAATGTTCTGGTGGATGGAGATGTAAGCTCAAGCTCTG LTx2 CAAGGATAACAAGGATCACAAGAAATGTTCTGGTGGATGGAGATGTAAGCTCAAGCTCTG LTx3 CAAGGATAACAAGGATCACAAGAAATGTTCTGGTGGATGGAGATGTAAGCTCAAGCTCTG ************************************************************
361 420
LTx1 TTTGAAGTTTtagtatgaaaactggccaggtgctgccatcactacgagaaaactcaccat LTx2 TTTGAAGATTtagtatgaaaactggccaggtgctgccatcactacgagaaaactcaccat LTx3 TTTGAAGTTTtagtatgaaaactggccaggtgctgccatcactacgagaaaactcaccat ******* ****************************************************
421 480
LTx1 aacatgcacacgaagagctattcttggtgatgctgtagcaaatgtattaacatgttagcc
LTx2 aacatgcacacgaagagctattcttggtgatggtgtagcaaatgtattaacatgttagcc
LTx3 aacatgcacacgaagagctattcttggtgatgctgtagcaaatgtattaacatgttagcc
******************************** ***************************
481 540
LTx1 agaaatgttgtttgaattatattatgtatctgaaaaatcgttaataaaatagtttcaac-
LTx2 agaaatgttgtttgaattatattatgtatctgaaaaatcgttaataaaatagtttcaac-
LTx3 agaaatgttgtttgaattatattatgtttctgaaaaatcgttaataaaatagtttcaaca
*************************** **************.......**********
541 570
LTx1 ctttcaaaaaaaaaaaa
LTx2 ctttcaaaaaaaaaaaa
LTx3 ctttcaaaaaaaaaaaa
*****************
Figura 10: Alinhamento dos cDNAs dos clones que apresentaram similaridade com toxinas de
aranhas S. huwena, E. californicum, B. smithi e Lasiodora parahibana. As regiões 5’ e 3’ não
codificantes são mostradas em letras minúsculas. Em vermelho aparece a seqüência que codifica
para o peptídeo sinal seguida da seqüência codificante do peptídeo intermediário (verde). A
seqüência nucleotídica correspondente à região da toxina madura é mostrada em azul, enquanto o
códon de parada é mostrado em rosa. O códon de iniciação e o sinal de poliadenilação estão
indicados por pontos (.). Asteriscos (*) indicam regiões consenso a todas as seqüências.

64
7.2.1- Tradução dos cDNAs que codificam para as toxinas LTx1, LTx2 e LTx3
A figura 11 mostra a tradução das seqüências nucleotídicas dos cDNAs que codificam
para as toxinas LTx1, LTx2 e LTx3. As regiões das seqüências nucleotídicas destacadas em
vermelho, verde e azul, codificam para o peptídeo sinal, peptídeo intermediário e toxina
madura, respectivamente. A tradução das seqüências nucleotídicas foi obtida pelo programa
SIXFRAME (http://workbench.sdsc.edu).

65
Toxina LTx1
ATG AGA TCT TTG ACG TTG GCT GCT CTT CTG CTA TGC AGC TTG CTG TTA GTT M R S L T L A A L L L C S L L L V TTT CAC ACT TCA GCA GCT GCA GAG CTT GAA GCT CAG GAA GGC CAT CTG ATG F H T S A A A E L E A Q E G H L M ATC CCC GGT GAT ACT GAC ACC GCA CTA GAA ACT GTC GAT GAT GAA AGA TTT I P G D T D T A L E T V D D E R F TTC GAA TGT ACT TTT GAA TGC GAC ATT AAA AAA GAA GGC AAA CCA TGC AAA F E C T F E C D I K K E G K P C K CCC AAG GGA TGC AAG TGT AAG GAC AAG GAT AAC AAG GAT CAC AAG AAA TGT P K G C K C K D K D N K D H K K C TCT GGT GGA TGG AGA TGT AAG CTC AAG CTC TGT TTG AAG TTT S G G W R C K L K L C L K F
Toxina LTx2
ATG AGA TCT TTG ACG TTG GCT GCT CTT CTG CTA TGC AGC TTG CTG TTA GTT M R S L T L A A L L L C S L L L V TTT CAC ACT TCA GCA GCT GAA GAG CTT CAA GCT CAG GAA GGC CAT CTG ATG F H T S A A E E L Q A Q E G H L M ATC CCC GGT GAT ACT GAC ACC GCC CTA GAA ACT GTC GAT GAT GAA AGA CTT I P G D T D T A L E T V D D E T L TTC GAA TGT ACT TTT GAA TGC GAC ATT AAA AAA GAA GGC AAA CCA TGC AAA F E C T F E C D I K K E G K P C K CCC AAG GGA TGC AAG TGT GAC GAC AAG GAT AAC AAG GAT CAC AAG AAA TGT P K G C K C D D K D N K D H K K C TCT GGT GGA TGG AGA TGT AAG CTC AAG CTC TGT TTG AAG ATT S G G W R C K L K L C L K I
Toxina LTx3
ATG AGA TCT TTG ACG TTG GCT GCT CTT CTG CTA TGC AGC TTG CTG TTA GTT M R S L T L A A L L L C S L L L V TTT CAC ACT TCA GCA GCT GCA GAG CTT CAA GCT CAG GAA GGC CAT CTG ATG F H T S A A A E L Q A Q E G H L M ATC CCC GGT GAT ACT GAC ACC GCC CTA GAA ACT GTC GAT GAT GAA AGA TTT I P G D T D T A L E T V D D E R F TTC GAA TGT ACT TTT GAA TGC GAC ATT AAA AAA GAA GGC AAA CCA TGC AAA F E C T F E C D I K K E G K P C K CCC AAG GGA TGC AAG TGT GAC GAC AAG GAT AAC AAG GAT CAC AAG AAA TGT P K G C K C D D K D N K D H K K C TCT GGT GGA TGG AGA TGT AAG CTC AAG CTC TGT TTG AAG TTT
S G G W R C K L K L C L K F
Figura 11: Tradução das seqüências nucleotídicas que codificam para as toxinas LTx1, LTx2 e
LTx3 utilizando-se o programa SIXFRAME. A seqüência correspondente ao peptídeo sinal,
peptídeo intermediário e à toxina madura são mostrados em vermelho, verde e azul,
respectivamente.

66
7.2.2- Análise estrutural das toxinas LTx1, LTx2 e LTx3
A figura 12 mostra o gráfico gerado pela análise das seqüências peptídicas das
toxinas maduras de LTx1, LTx2 e LTx3 no programa SignalP, que prediz o sítio de
clivagem de peptídeos sinal. A figura 13 mostra o gráfico de hidropaticidade bem como o
alinhamento das seqüências peptídicas das regiões correspondentes ao peptídeo sinal
(vermelho), peptídeo intermediário (verde) e proteína madura (azul) das toxinas LTx1,
LTx2 e LTx3 encontradas neste trabalho. O gráfico de hidropaticidade gerado pela análise
das seqüências peptídicas das três toxinas, quando submetidas ao programa ProtScale
(disponível em http://www.cbs.dtu.dk/services/signalp/), revelou uma região inicial da
seqüência composta predominantemente por aminoácidos hidrofóbicos, característica de
peptídeos sinal, comum em toxinas. A região do peptídeo intermediário é rica em
aminoácidos hidrofílicos e concentra a maior parte dos resíduos de glutamato (E) do pró-
peptídeo. A toxina madura apresenta um perfil predominantemente hidrofílico.

67
Figura 12: Predição do ponto de clivagem entre os peptídeos sinal e peptídeo intermediário na
seqüência dos precursores das toxinas LTx1, LTx2 e LTx3. O pico em vermelho indica o ponto de
clivagem entre os peptídeos sinal e intermediário (SAA-AE). O pico em azul escuro indica o caráter
hidrofóbico comum aos peptídeos sinal. Os picos verde e azul claro indicam as regiões N- e C-
terminais do peptídeo sinal, respectivamente.

68
Figura 13: Gráfico de hidropaticidade das seqüências de aminoácidos traduzidas a partir dos cDNAs
dos clones que codificam os precursores das toxinas LTx1, LTx2 e LTx3. Neste gráfico estão
representados o peptídeo sinal hidrofóbico (23 aminoácidos), o peptídeo intermediário (27
aminoácidos) e a toxina madura (49 aminoácidos). As regiões acima e abaixo do ponto 0 (seta
vermelha) indicam a presença de aminoácidos com características hidrofóbicas e hidrofílicas,
respectivamente.
Peptídeo sinal Peptídeo intermediário Toxina madura
MRSLTLAALLLCSLLLVFHTSAA MRSLTLAALLLCSLLLVFHTSAA MRSLTLAALLLCSLLLVFHTSAA ---------23-----------
AELEAQEGHLMIPGDTDTALETVDDER EELQAQEGHLMIPGDTDTALETVDDER AELQAQEGHLMIPGDTDTALETVDDER ------------27-------------
FFECTFECDIKKEGKPCKPKGCKCKDKDNKDHKKCSGGWRCKLKLCLKF LFECTFECDIKKEGKPCKPKGCKCDDKDNKDHKKCSGGWRCKLKLCLKI FFECTFECDIKKEGKPCKPKGCKCDDKDNKDHKKCSGGWRCKLKLCLKF -----------------------49------------------------

69
7.2.3- Alinhamento das seqüências peptídicas das toxinas das aranhas Lasiodora sp (LTx1, LTx2, LTx3), E. Californicum (ESTx), B. smithii (BsTx), L. parahybana (LpTx1 e LpTx2)
A figura 14 mostra o alinhamento das seqüências peptídicas das toxinas da aranha
Lasiodora sp (LTx1, LTx2 e LTx3) com toxinas das aranhas E. Californicum (ESTx), B.
smithii (BsTx), e L. parahybana (LpTx1 e LpTx2).
LTx1 FFECTFECDIKKEGKPCKPKGCKCKDKDNKDHKKCSGGWRCKLKLCLKF LTx2 LFECTFECDIKKEGKPCKPKGCKCDDKDNKDHKKCSGGWRCKLKLCLKI LTx3 FFECTFECDIKKEGKPCKPKGCKCDDKDNKDHKKCSGGWRCKLKLCLKF LpTx1 FFECTFECDIKKEGKPCKPKGCKCKDKDNKDHKKCSGGWRCKLKLCLKF LpTx2 FFECTLECDIKKEGKPCKPKGCKCNDKDNKDHKKCSGGWRCKLKLCLKF ESTX IFECVFSCDIEKEGKPCKPKG----------EKKCSGGWKCKIKLCLKI BsTx IFECVFSCDIEKEGKPCKPKG----------EKKCSGGWKCKIKLCLKI
Figura 14: Alinhamento das seqüências de aminoácidos das toxinas das aranhas Lasiodora sp
(LTx1, LTx2 e LTx3), E. Californicum (ESTx), B. smithii (BsTx), e L. parahybana (LpTx1 e
LpTx2). Os resíduos completamente conservados entre as seqüências são mostrados em azul. Em
verde são mostrados os resíduos altamente conservados enquanto aqueles pouco conservados são
apresentados em vermelho. Em preto é mostrada uma região onde não há consenso com as
seqüências dos peptídeos ESTx e BsTx.

70
7.2.4- Alinhamento das seqüências peptídicas completas das toxinas LTx1, LTx2, LTx3 e HwTx-II (Ile)
A figura 15 mostra o alinhamento das seqüências peptídicas das toxinas da aranha
Lasiodora sp (LTx1, LTx2 e LTx3) com uma toxina composta por 37 resíduos de
aminoácidos, com atividade inseticida, encontrada na glândula de veneno da aranha
Selenocosmia huwena.
LTx1 FFECTFECDIKKEG-KPCKPKGCKCKDKDNKDHKKCSGGWRCKLKLCLKF LTx2 LFECTFECDIKKEG-KPCKPKGCKCDDKDNKDHKKCSGGWRCKLKLCLKI LTx3 FFECTFECDIKKEG-KPCKPKGCKCDDKDNKDHKKCSGGWRCKLKLCLKF HwTx-II LFECSFSCEIEKEGDKPCK-------------KKKCKGGWKCKFNMCVKV
Figura 15: Alinhamento das seqüências de aminoácidos das toxinas das aranhas Lasiodora sp
(LTx1, LTx2 e LTx3) e Selenocosmia huwena (HwTx-II). Os resíduos completamente conservados
entre as seqüências são mostrados em azul. Em verde são mostrados os resíduos altamente
conservados enquanto aqueles pouco conservados são apresentados em vermelho. Em preto são
mostradas as regiões onde não há consenso com a seqüência do peptídeo HwTx-II.

71
7.3- Clonagem das toxinas LTx1 e LTx3 em PCR® 2.1-TOPO TA Cloning Invitrogen®
Com o objetivo de facilitar a clonagem dos fragmentos que codificam as toxinas
LTx1 e LTx3 no vetor de expressão pET-11a Novagen®, os fragmentos obtidos por PCR
utilizando iniciadores contendo os sítios de restrição Nde I e Bam H I foram primeiramente
clonados em vetor PCR®
2.1-TOPO TA Cloning Invitrogen®. Essa estratégia faz-se
necessária devido à dificuldade apresentada por varias enzimas de restrição, por exemplo
Nde I, em digerir sítios localizados nas extremidades de fragmentos de PCR. A figura 16
mostra o produto da amplificação dos fragmentos correspondentes às toxinas maduras
LTx1 e LTx3 clonadas em vetor PCR® 2.1-TOPO TA Cloning Invitrogen
®. As amostras
foram submetidas à eletroforese em gel de poliacrilamida 6%. Na canaleta 1, aplicou-se 6µl
de padrão de peso molecular 1kb da Invitrogen®. Nas canaletas (2 e 3) e (4 e 5), são
apresentados os produtos de PCR (aproximadamente 350pb) dos fragmentos que codificam
para a toxina LTx1 e LTx3, respectivamente.
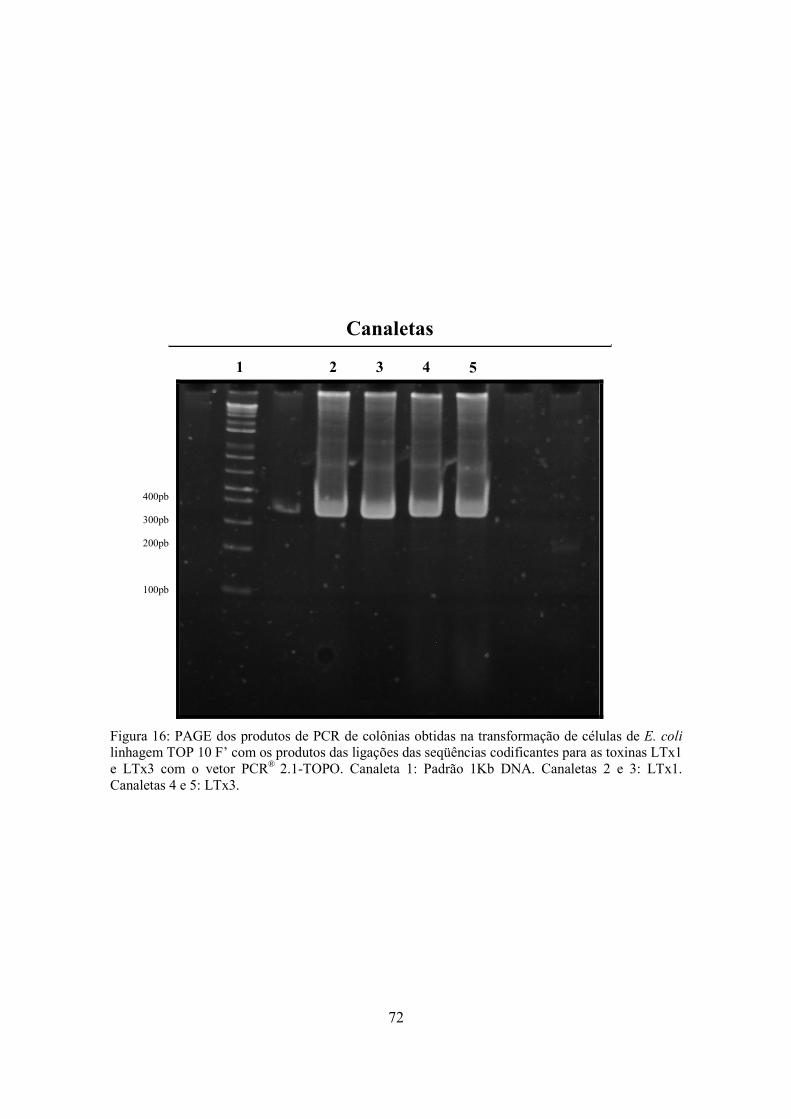
72
Figura 16: PAGE dos produtos de PCR de colônias obtidas na transformação de células de E. coli
linhagem TOP 10 F’ com os produtos das ligações das seqüências codificantes para as toxinas LTx1
e LTx3 com o vetor PCR® 2.1-TOPO. Canaleta 1: Padrão 1Kb DNA. Canaletas 2 e 3: LTx1.
Canaletas 4 e 5: LTx3.
1 2 3 4 5
Canaletas
100pb
200pb
300pb
400pb

73
7.4- Clonagem das toxinas LTx1 e LTx3 no vetor de expressão pET-11a
Após a digestão de DNAs extraídos dos clones obtidos com o sistema PCR® 2.1-
TOPO®, com as enzimas de restrição Nde I e BamH I, os fragmentos que codificam as
toxinas LTx1 e LTx3 foram purificadas como descrito em Materiais e Métodos. Procedeu-
se então a clonagem no vetor de expressão pET-11a. As colônias selecionadas na
transformação foram submetidas a uma PCR para verificar a presença do fragmento
clonado (Figura 17). O produto de amplificação correspondente à clonagem das toxinas
LTx1 e LTx3 no vetor pET-11a é de aproximadamente 240 pb (canaletas 2 e 3).

74
Figura 17: PAGE dos produtos de PCR de colônias obtidas na transformação de células de E. coli
linhagem TOP 10 F’ com produto de clonagem das seqüências codificantes para LTx1 e LTx3 no
vetor de expressão pET-11a (canaletas 2 e 3). Na canaleta 4 foi aplicado o produto de amplificação
do vetor pET-11a vazio.
200pb
300pb
400pb
100pb
1 2 3 4

75
7.5- Indução da toxina LTx1 recombinante Após a expressão da toxina LTx1 recombinante em diferentes tempos de indução (1,
2, 3, 4 e 5 horas) como descrito em materiais e métodos (6.16), os extratos celulares foram
submetidos à eletroforese em gel SDS-PAGE. A análise da expressão de LTx1 revelou
uma indução basal (tempo 0) e um tempo ótimo de indução em torno de 5 horas após a
adição de IPTG 0.6mM (concentração final) e incubação a 30ºC (200 rpm) (Figura 18).
Alíquotas de aproximadamente 20µg de proteínas provenientes do veneno bruto, da fração
solúvel e da fração insolúvel (corpos de inclusão) foram submetidas à eletroforese em gel
SDS-PAGE. As proteínas presentes no gel foram transferidas para uma membrana de nylon
e analisadas por western bloting usando o antisoro contra veneno total como anticorpo
primário. A figura 19 mostra o reconhecimento de proteínas do veneno bruto de Lasiodora
sp (canaleta 1) e da toxina LTx1 recombinante (canaletas 2). Como pode ser observado, a
toxina LTx1 recombinante foi encontrada, sobretudo na fração solúvel, sugerindo que não
há formação de corpos de inclusão uma vez que não foi observada a presença mensurável
de LTx1 no precipitado da sonicação (canaleta 3).

76
Figura 18: Gel SDS-PAGE da indução da expressão de LTx1 recombinante. Os tempos de indução
são indicados acima de cada canaleta. A toxina LTx1 recombinante está indicada pela seta. Uma
indução basal pode ser observada (0 hora de indução).
0 1 2 3 4 5
Tempos de Indução (h)
LTx1

77
Figura 19: Western Bloting da toxina LTx1 recombinante. Uma fração do veneno bruto e as
eluições de LTx1 (sobrenadante e corpos de inclusão) foram aplicados nas canaletas 1, 2 e 3,
respectivamente. A toxina LTx1 recombinante, presente no sobrenadante está indicada pela seta.
1 2 3
Canaletas

78
7.6- Análise da seqüência nucleotídica dos cDNAs dos clones (16-19) que apresentaram similaridade com a toxina Magi 7 da aranha Macrothele gigas
Dos 19 clones que apresentaram similaridades com toxinas de outras aranhas, 4
apresentaram similaridade com o peptídeo precursor da toxina Magi 7, descrito por Satake
e cols. (2004), composto por 124 resíduos de aminoácidos, presente na glândula de veneno
da aranha Macrothele gigas. O cDNA dos 4 quatro clones que apresentaram similaridade
com o precursor da toxina Magi 7 foi traduzido no frame +2 (utilizando o programa
SIXFRAME) e originou uma janela aberta de leitura que codifica 116 resíduos de
aminoácidos (Figura 20). A figura 21 mostra o alinhamento do peptídeo precursor de LTx5
com o peptídeo precursor de Magi 7. O novo peptídeo encontrado foi designado pela sigla
LTx5. A análise do peptídeo LTx5 pelo programa SignalP (disponível em
http://www.cbs.dtu.dk/services/signalp/) revelou a existência de um peptídeo sinal
composto por 17 resíduos de aminoácidos, com o sítio de clivagem entre o peptídeo sinal e
o peptídeo intermediário localizado entre os aminoácidos das posições 17 e 18 (AVA-TW)
(Figura 22). O peptídeo intermediário apresenta 28 resíduos de aminoácidos, com o ponto
de clivagem localizado no resíduo de arginina (R) da posição 45 (Arg45). A análise do
peptídeo LTx5 pelo programa BlastP revelou 32 % de similaridade com o peptídeo
precursor da toxina Magi 7. A figura 21 mostra o alinhamento das seqüências peptídicas de
Magi 7 e LTx5 obtido na análise pelo programa ClustalW.
A figura 23 mostra o perfil de hidropaticidade bem como as regiões correspondentes
ao peptídeo sinal (vermelho), peptídeo intermediário (verde) e proteína madura (azul) da
toxina LTx5. O gráfico de hidropaticidade gerado pela análise da seqüência peptídica da
toxinas LTx5 quando submetida ao programa ProtScale (disponível em
http://www.cbs.dtu.dk/services/protscale/) revelou características similares às seqüências
LTx1, LTx2 e LTx3, isto é, uma região inicial composta predominantemente por
aminoácidos hidrofóbicos, uma região rica em aminoácidos hidrofílicos e concentrando
grande parte dos resíduos de glutamato. A toxina madura apresenta um perfil
predominantemente hidrofílico.

79
gaattcggcacgagcagaaacaaatattgctggaatttcactcaagATG AAG CTG AGC ACT TTT ATC M K L S T F I ATA ATG ATA AGT CTG GCT GTG GCC GTT GCT ACC TGG CCT TCA GAG CAC ATT GAA I M I S L A V A V A T W P S E H I E GGA AGT GAT TCT GAA ACC AAA CTG AAT GTG GAA CTC GGG CCA TAT GCA TTA GCT G S D S E T K L N V E L G P Y A L A GAT CGT GCA GAA AAG GGA AAA AAG GAC TCC CTT AAC AAA GGA GAG CCA TGC CAA D R A E K G K K D S L N K G E P C Q TTT CAT TGC GAG TGT CGT GGT GCT AGT GTT CTC TGC GAA GCT GTT TAT GGA ACA F H C E C R G A S V L C E A V Y G T AGA AGT CCG ATG TAT AAA TGC ATG ATC AAA AGA TTA CCT ATT TCG GTA TTG GAC R S P M Y K C M I K P L P I S V L D ATA ATG TAT CAG GCA GAG CGT GCT TTA GAA AAA CTT GCT TCT TCA TTT CGG TGT I M Y Q A E R A L E K L A S S F R C GAGTAAataaggttaactcgttcagggcaacgaatgactgaatgttacataacaatgcataatacttctgtaa E * aatctgtattggcaaaagaattcgtttagaaaaaaacaataaagcgagatatatgcagaaaaaaaaactcgag
Figura 20: Tradução do cDNA que codifica a toxina LTx5. As regiões 5’ e 3’ não traduzidas são
mostradas em letras minúsculas enquanto a região codificante aparece em maiúsculas. As regiões
que codificam para os peptídeos sinal, intermediário e maduro são mostrados em vermelho, verde e
azul, respectivamente.
Magi_7 MKLSALVFVASVMLVAASPVKDVEEPVETHLAADLKTIEELAKYEEAAVQKRSCIVGSKN LTx5 MKLSTFIIMISLAVAVATWPSEHIEGSD----SETKLNVELGPYALADRAEK-GKKDSLN Magi_7 IGETCVASCQCCGATVRCIGEGTKGICNNYQTNNILGQILLYAKDTVVNTAGLLVCAQDL LTx5 KGEPCQFHCECRGASVLCEAVYGT---RSPMYKCMIKRLPISVLDIMYQAERALEKLASS Magi_7 SEYE LTx5 FRCE
Figura 21: Alinhamento das seqüências peptídicas dos pró-peptídeos Magi 7 (Macrothele gigas) e
LTx5 (Lasiodora sp). Os resíduos completamente conservados entre as seqüências são mostrados
em azul. Em verde são mostrados os resíduos altamente conservados enquanto aqueles pouco
conservados são apresentados em vermelho. Em preto são mostradas as regiões onde não há
consenso com a seqüência do peptídeo.

80
Figura 22: Predição do ponto de clivagem entre os peptídeos sinal e intermediário na seqüência do
precursor da toxina LTx5. O pico em vermelho indica o ponto de clivagem entre os peptídeos sinal
e intermediário (AVA-TW). O pico em azul escuro indica o caráter hidrofóbico comum aos
peptídeos sinais. Os picos verde e azul claro indicam as regiões N- e C-terminais do peptídeo sinal,
respectivamente.
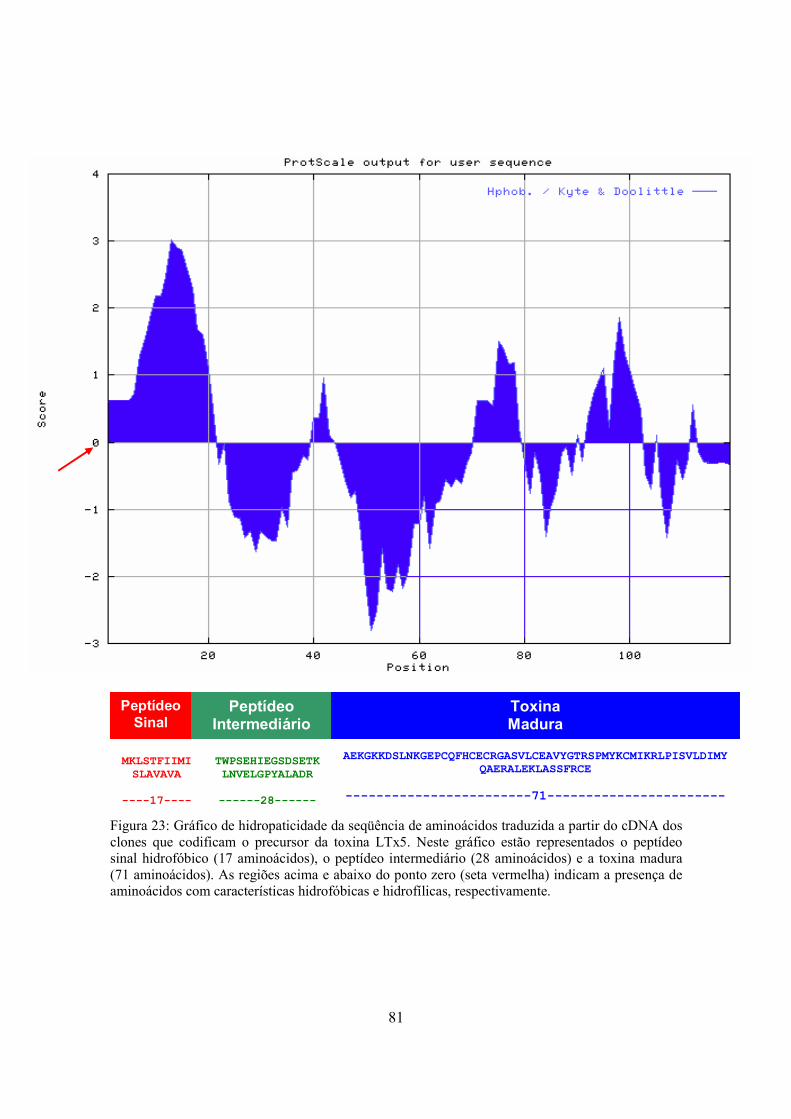
81
Figura 23: Gráfico de hidropaticidade da seqüência de aminoácidos traduzida a partir do cDNA dos
clones que codificam o precursor da toxina LTx5. Neste gráfico estão representados o peptídeo
sinal hidrofóbico (17 aminoácidos), o peptídeo intermediário (28 aminoácidos) e a toxina madura
(71 aminoácidos). As regiões acima e abaixo do ponto zero (seta vermelha) indicam a presença de
aminoácidos com características hidrofóbicas e hidrofílicas, respectivamente.
Peptídeo
Sinal Peptídeo
Intermediário Toxina Madura
MKLSTFIIMISLAVAVA
----17----
TWPSEHIEGSDSETKLNVELGPYALADR
------28------
AEKGKKDSLNKGEPCQFHCECRGASVLCEAVYGTRSPMYKCMIKRLPISVLDIMYQAERALEKLASSFRCE
------------------------71-----------------------

82
7.7- Análise das seqüências que apresentaram similaridade com outras proteínas depositadas no GenBank
Em uma primeira análise (BlastN e BlastX apenas), dos 43 clones que apresentaram
reação positiva no teste de ELISA 22 apresentaram similaridades com outras proteínas
depositadas no GenBank. Esses clones foram posteriormente analisados utilizando os
programas SIXFRAME (workbench.sdsc.edu) e BlastP. A seguir são apresentadas as
análises apenas dos clones que apresentaram uma ou mais janelas de leitura abertas e cujo
peptídeo codificado apresentou alguma similaridade com proteínas depositadas no
GenBank quando submetidos à análise pelo BlastP.
7.7.1- Análise da seqüência do DNA complementar do clone 20
A seqüência do cDNA do clone 20 apresenta 629 pares de bases. Os sítios de
restrição Xho I e Eco RI são mostrados em vermelho. O códon de iniciação e o de parada
são mostrados em rosa e verde, respectivamente. A tradução do cDNA do clone 20
utilizando o programa SIXFRAME procedeu-se no frame +2 e originou uma proteína
composta por 109 aminoácidos denominada LsC1, cuja seqüência é mostrada a seguir.
cDNA
GAATTCGGCACGAGCAGAAACAGACATCATGGTGCACCTGACTGATGCTGAGAAGGCTGCTGTCTCTGGC CTGTGGGGAAAGGTGAACGCCGATGAAGTTGGTGGTGAGGCCCTGGGCAGGCTGCTGGTTGTCTACCCTT GGACCCAGCGGTACTTTGATAGCTTTGGAGACCTATCCTCTGCCTCTGCTATCATGGGTAATGCCAAAGT GAAGGCCCATGGCAAGAAAGTGATAACTGCCTTTAACGATGGCCTGAATCACTTGGACAGCCTCAAGGGC ACCTTTGCCAGCCTCAGTGAGCTCCACTGTGACAAGCTGCATGTGGATCCTGAGAACTTCAGGCTCCTGG GCAAATGATCGTGATTGTGCTGGGCCACCACCTGGGCAAGGATTTCACCCCCGCTGCACAGGCTGCCTTC CAGAAGGTGGTGGCTGGAGTGGCCGCTGCCCTGGCTCACAAGTACCACTAAACCCCCTTTCCTGCTCTTG CCTGTGAACAATGGTTAATTGTTCCCAAGAGAGCATCTGTCAGTTGTTGGCAAAATGATAAAGACATTTG AAAATCTGTCTTCTGACAAATAAAAAGCATTTATTTCACTGCAAAAAAAAAAAAAAAAAAAAACTCGAG

83
LsC1
MVHLTDAEKA AVSGLWGKVN ADEVGGEALG RLLVVYPWTQ RYFDSFGDLS SASAIMGNAK VKAHGKKVIT AFNDGLNHLD SLKGTFASLS ELHCDKLHVD PENFRLLGK
A análise da seqüência do cDNA do clone 20 revelou alta similaridade (98%) com a
cadeia β-1 da hemoglobina do camundongo pertencente à espécie Mus musculus. A cadeia
β-1 possui 146 aminoácidos. Uma elevada similaridade (92%) com a cadeia β-2 da globina
do camundongo Rattus norvegicus também foi detectada. Esta cadeia apresenta 147
resíduos de aminoácidos. A seqüência do cDNA do clone 20 apresentou uma terceira
similaridade (100%) com uma hemoglobina quimérica humano/camundongo (cadeias zeta
2/beta 2) de 146 aminoácidos.
A figura 24 mostra o alinhamento da seqüência peptídica codificada pelo cDNA do
clone 20 com as proteínas com as quais apresentou similaridade.
Cadeia ββββ-1 da hemoglobina do camundongo Mus musculus
Score = 210 bits (535), Expect = 3e-54 Identities = 106/108 (98%), Positives = 107/108 (99%)
Query:1 MVHLTDAEKAAVSGLWGKVNADEVGGEALGRLLVVYPWTQRYFDSFGDLSSASAIMGNAK 60 MVHLTDAEKAAVS LWGKVN+DEVGGEALGRLLVVYPWTQRYFDSFGDLSSASAIMGNAK Sbjct:1 MVHLTDAEKAAVSCLWGKVNSDEVGGEALGRLLVVYPWTQRYFDSFGDLSSASAIMGNAK 60
Query:61 VKAHGKKVITAFNDGLNHLDSLKGTFASLSELHCDKLHVDPENFRLLG 108 VKAHGKKVITAFNDGLNHLDSLKGTFASLSELHCDKLHVDPENFRLLG Sbjct:61 VKAHGKKVITAFNDGLNHLDSLKGTFASLSELHCDKLHVDPENFRLLG 108
• Query: clone 20
• Sbjct: β-1 hemoglobina
(a)

84
Cadeia beta-2 da globina do camundongo Rattus norvegicus
Score = 202 bits (515), Expect = 7e-52
Identities = 100/108 (92%), Positives = 103/108 (95%)
Query: 1 MVHLTDAEKAAVSGLWGKVNADEVGGEALGRLLVVYPWTQRYFDSFGDLSSASAIMGNAK 60 MVHLTDAEKAAV+GLWGKVN D+VGGEALGRLLVVYPWTQRYFDSFGDLSSASAIMGN K Sbjct: 1 MVHLTDAEKAAVNGLWGKVNPDDVGGEALGRLLVVYPWTQRYFDSFGDLSSASAIMGNPK 60
Query: 61 VKAHGKKVITAFNDGLNHLDSLKGTFASLSELHCDKLHVDPENFRLLG 108 VKAHGKKVI AFNDGL HLD+LKGTFA LSELHCDKLHVDPENFRLLG Sbjct: 61 VKAHGKKVINAFNDGLKHLDNLKGTFAHLSELHCDKLHVDPENFRLLG 108
• Query: clone 20
• Sbjct: β-2 globina
Hemoglobina quimérica humano/camundongo(zeta 2/beta 2)
Score = 211 bits (537), Expect = 2e-54
Identities = 107/107 (100%), Positives = 107/107 (100%)
Query: 2 VHLTDAEKAAVSGLWGKVNADEVGGEALGRLLVVYPWTQRYFDSFGDLSSASAIMGNAKV 61 VHLTDAEKAAVSGLWGKVNADEVGGEALGRLLVVYPWTQRYFDSFGDLSSASAIMGNAKV Sbjct: 1 VHLTDAEKAAVSGLWGKVNADEVGGEALGRLLVVYPWTQRYFDSFGDLSSASAIMGNAKV 60
Query:62 KAHGKKVITAFNDGLNHLDSLKGTFASLSELHCDKLHVDPENFRLLG 108 KAHGKKVITAFNDGLNHLDSLKGTFASLSELHCDKLHVDPENFRLLG Sbjct:61 KAHGKKVITAFNDGLNHLDSLKGTFASLSELHCDKLHVDPENFRLLG 107
• Query: clone 20
• Sbjct: Hemoglobina quimerica
Figura 24: Alinhamento da seqüência peptídica codificada pelo cDNA do clone 20 com a cadeia β-1
da hemoglobina de Mus musculus (a), cadeia β-2 da globina de Rattus norvegicus (b) e hemoglobina
quimérica humano/camundongo (zeta 2/beta 2) (c). Os resíduos altamente conservados são mostrados
em azul enquanto os grupos fortemente conservados aparecem em verde. Em preto são mostrados
resíduos que não apresentam consenso.
7.7.2. Análise da seqüência do DNA complementar do clone 21
(b)
(c)

85
A seqüência do cDNA do clone 21 apresenta 434 pares de bases. Os sítios de
restrição Xho I e Eco RI são mostrados em vermelho. Os códons de iniciação e parada são
mostrados em rosa e verde, respectivamente. A tradução do cDNA do clone 21 realizada
pelo programa SIXFRAME procedeu-se no frame +2 e originou uma proteína de 82
aminoácidos denominada LsC2, cuja seqüência é mostrada a seguir.
cDNA
GAATTCGGCACGAGTGATTTAGTTGTTCCGATCGGGAAGTACTGGAATAAAGGAAGCAATGGGTAAACAG TTTGGACAGCTAGCAAAAATGAGGGGCATTGTACAATTCAGAATATCACCGTATGAGCAAAATCCACTTG CTGGAATAATTTCTAAAGGGTTTCCAAATATGATTCGACGTATTCGTTCCAATATATTTTCCATGGCTCC ACCATTTGTCCTTGGATATCTCGTTTATGATTGGACTGAAAAAGAACATGCCCGTCTGCAGAGGAAGAAC CCAGCTGATTATGCAAATGATGAATGAATGATATTTGTTTACATAGAAAATTCTGTAGTTCAGTTATTCT GTACATTTTTGTAGTTCATTATTGAAATAAATATAAGAAAAATAAAGAAGTGATTCTGGTTTCTAAAAAA AAAAAAAACTCGAG
LsC2
MGKQFGQLAK MRGIVQFRIS PYEQNPLAGI ISKGFPNMIR RIRSNIFSMA PPFVLGYLVY DWTEKEHARL QRKNPADYANDE
A análise da seqüência do cDNA do clone 21 apresentou uma similaridade de 62%
com uma proteína de 89 aminoácidos da mosca Drosophila melanogaster, denominada
CG7580-PA. Uma similaridade menor (58%) foi verificada entre o peptídeo codificado
pelo cDNA do clone 21 e uma proteína de 88 aminoácidos do mosquito Anopheles
gambiae, denominada agCP10691. A seqüência do cDNA do clone 21 apresentou uma
terceira similaridade (46%) com uma proteína de 88 aminoácidos denominada Qpc-pending
do camundongo Mus musculus.
A figura 25 mostra o alinhamento da seqüência peptídica codificada pelo cDNA do
clone 21 com as proteínas com as quais apresentou similaridade.

86
Proteína CG7580-PA da mosca Drosophila melanogaster
Score = 115 bits (289), Expect = 1e-25
Identities = 51/81 (62%), Positives = 65/81 (80%)
Query: 2 GKQFGQLAKMRGIVQFRISPYEQNPLAGIISKGFPNMIRRIRSNIFSMAPPFVLGYLVYD 61 G+ FG LAK+ GIV +++SP+EQ AG ISKG PNM+RR RSN+F + PPF++GYL+YD Sbjct: 9 GQHFGNLAKVHGIVTYKLSPFEQRAFAGAISKGLPNMVRRFRSNVFIVTPPFIVGYLIYD 68
Query: 62 WTEKEHARLQRKNPADYANDE 82 TE++H L RKNPADYANDE Sbjct: 69 LTERKHTALLRKNPADYANDE 89
• Query: clone 21
• Sbjct: CG7580-PA
Proteína agCP10691 do mosquito Anopheles gambiae
Score = 103 bits (257), Expect = 6e-22
Identities = 47/81 (58%), Positives = 63/81 (77%)
Query: 1 MGKQFGQLAKMRGIVQFRISPYEQNPLAGIISKGFPNMIRRIRSNIFSMAPPFVLGYLVY 60 MG FG+LAK+RGIV +ISP+EQ A +ISKG PN +RRIRS +F +APPFV+GY+VY Sbjct: 7 MGHGFGELAKVRGIVTHKISPFEQRAFANVISKGVPNTLRRIRSQVFIVAPPFVMGYMVY 66
Query: 61 DWTEKEHARLQRKNPADYAND 81 ++ E H ++ RKNPA++ ND Sbjct: 67 NYIENLHTQMNRKNPAEFEND 87
• Query: clone 21
• Sbjct: agCP10691
Proteína Qpc-pending do camundongo Mus musculus
(a)
(b)
(c)

87
Score = 84.0 bits (206), Expect = 4e-16
Identities = 38/82 (46%), Positives = 56/82 (68%)
Query: 1 MGKQFGQLAKMRGIVQFRISPYEQNPLAGIISKGFPNMIRRIRSNIFSMAPPFVLGYLVY 60 MG++FG LA++R ++ + +SP+EQ SKG PN++RR R I +APPFV+ YL+Y Sbjct: 1 MGREFGNLARIRHVISYSLSPFEQRAFPSYFSKGIPNVLRRTRERILRVAPPFVVVYLIY 60
Query: 61 DWTEKEHARLQRKNPADYANDE 82 W +E + +RKNPA Y ND+ Sbjct: 61 TWGNQEFEQSKRKNPAMYENDK 82
• Query: clone 21
• Sbjct: Qpc-pending
Figura 25: Alinhamento da seqüência peptídica codificada pelo cDNA do clone 21 com as proteínas
CG7580-PA da mosca Drosophila melanogaster (a), agCP10691 do mosquito Anopheles gambiae (b)
e Qpc-pending do camundongo Mus musculus (c). Os resíduos altamente conservados são mostrados
em azul enquanto os grupos fortemente conservados aparecem em verde. Em preto são mostrados
resíduos onde não há consenso.
7.8- Análise das seqüências dos clones que codificam para peptídeos que não apresentaram similaridades, quando submetidos à análise pelo BlastP

88
Dentre os clones que apresentaram alguma similaridade com proteínas depositadas
no GenBank, quando submetidos a uma análise geral utilizando os programas BlastN e
BlastX, alguns não apresentaram qualquer similaridade quando os peptídeos codificados
por eles foram submetidos a uma análise mais detalhada pelo BlastP. A seguir são
apresentadas as análises apenas dos clones que apresentaram uma ou mais janelas de leitura
abertas (ORFs) e cujo peptídeo codificado não apresentou similaridade com proteínas
depositadas no GenBank quando submetidos à análise pelo BlastP.
7.8.1- Análise da seqüência do DNA complementar do clone 22

89
A seqüência nucleotídica do clone 22 apresenta 1403 pares de bases. A tradução
dessa seqüência utilizando o programa SIXFRAME revelou a existência de uma janela de
leitura no frame +2 (vermelho) e +3 (azul). A tradução nos frames +2 e +3 originou
peptídeos compostos por 72 e 67 resíduos de aminoácidos, respectivamente.
cDNA
CCCCCTCTGTCTGGGGNTTGGCCCCACNNGGTTGTATGTGTGTGTCTNCTCGCCTCTGCCAATATGGCTG GTTGTCTNGCCGNTGGGGGNTTTTTTATCCGGCCGCGTGTTTGTGTTCCCTTNTGNTTCCGTTGTCCCGC GNTTTCCGTGTTGGGTTTGGTCCTTCATTGGGCCCCCCNGGCTTGTCCAAGTGTGTATACTTTCCGGCTN CGTTGATTNATTGTGGGGGGATTCTGTGATCCTGGGTTAACCCTATTGTTCNAGTCANCCAGGGGGCTCC GACTGTTTGACCCTTTGGNTTTACCGCCCCATANGCCTCGGGTAGTTNTAGCCCCGTNCACTTTATAGAG GGGANCCAANATAGGCCTGTGGGGATGGCTTNACAGGCGCCGCCCCATTGTCCAAGGGTTNGNACAACTT TAACGGTTGGGATTCCCNCAAAGTAATGTTCGGTGGGCAAACGGTGAGNCCAATGCTAACTCCCAAAATG TAATGTTGCGCCGTGGGGAAATTTTTCAACCGTCCAGAAGTATGTGATAGCTGGAGCCCAACTTTGTTAA TCAATTNAATTGGGATTACAGGNTCCATGGGTCTATGGTTGCCCNCAGTGTGTTGCGCTGTTACCCACTT GGGCCCATTCCTAANGGAAGGCCAACCAAGTTTTGGAAACGGGAAAAGTGCTGGCATNTTTCCTGGTAAA TNNACCCCATTGCCTGTGGGTAAATGGTTGTGGAAACCCTTCCGGGCGCGCCAATTAAATCGNGCCAATG TTAAGGCCCTTGGGTATTTCGTTGGCAGGNCATAAAAGGGGGGCACAAATAATAGGGTAACTTCCCNNNG TGNTGAACCACAAACAGGGTAGNGACCCAATGTCCCAAAAGTGTTCAATGTGGCGGGAANGTTGNTTCGT TGGGGTTGTCACTTAGGTGTTTCCTCCTTGCGGAAAGGCCGTGTTTNTATTGGGAAACAATGGAAAGGTT CCCGAATGGTTTANTAAAGTGCCAATTGGAATGCCACTAAAAGGAATGTTTACCCTATTTCGGGTATTTG GACATAAATGGTTAATCAGGCCAGAGCGTGGCCTTTAGTTAAACGTTGCCTTCGTTCAGTTTCGGCTGTG AGTANATTAAGGTTAACTCGTTCAGGGCATACGATATGACTGAATGTTACATAACNAAGTGCATAATACT TCTGTAAAATCTGTATTTGGCAAAAAGAATTCGCTTTAGATATTGGTACATATTATAGCCGAGATATATG CAATGATTTGGAAGCTGTGAGTAGTGATACTCGAGAGTACTTCTAGAGCGCCCGCGGGCCCATCGATTTT CCACCCGGGTGGGTTTACAGCTAAGTACCCTATTCGCCTATAGTGAACTCATGGATTCATGCACACGTTC NAA
LsC3a (Tradução no frame +2)
MGLWLPXVCCAVTHLGPFLXEGQPSFGNGKSAGXFPGKXTPLPVGKWLWKPFRARQLNRANVKALGYFVGRX
LsC3b (Tradução no frame +3)
MCVSXRLCQYGWLSXRWGXFYPAACLCSLXXPLSRXFRVGFGPSLGPPGLSKCVYFPAXLIXCGGIL
A análise da seqüência peptídica de LsC3a pelo programa SignalP revelou a
provável existência de um sítio de clivagem para peptídeo sinal (Figura 26). O caráter
hidrofóbico do peptídeo sinal pode ser visto com maior clareza no gráfico de

90
hidropaticidade gerado pelo programa ProtScale para a seqüência peptídica de LsC3a
(Figura 27). A análise da seqüência peptídica LsC3b pelo programa SignalP não revelou
existência de um provável sítio de clivagem para peptídeo sinal.
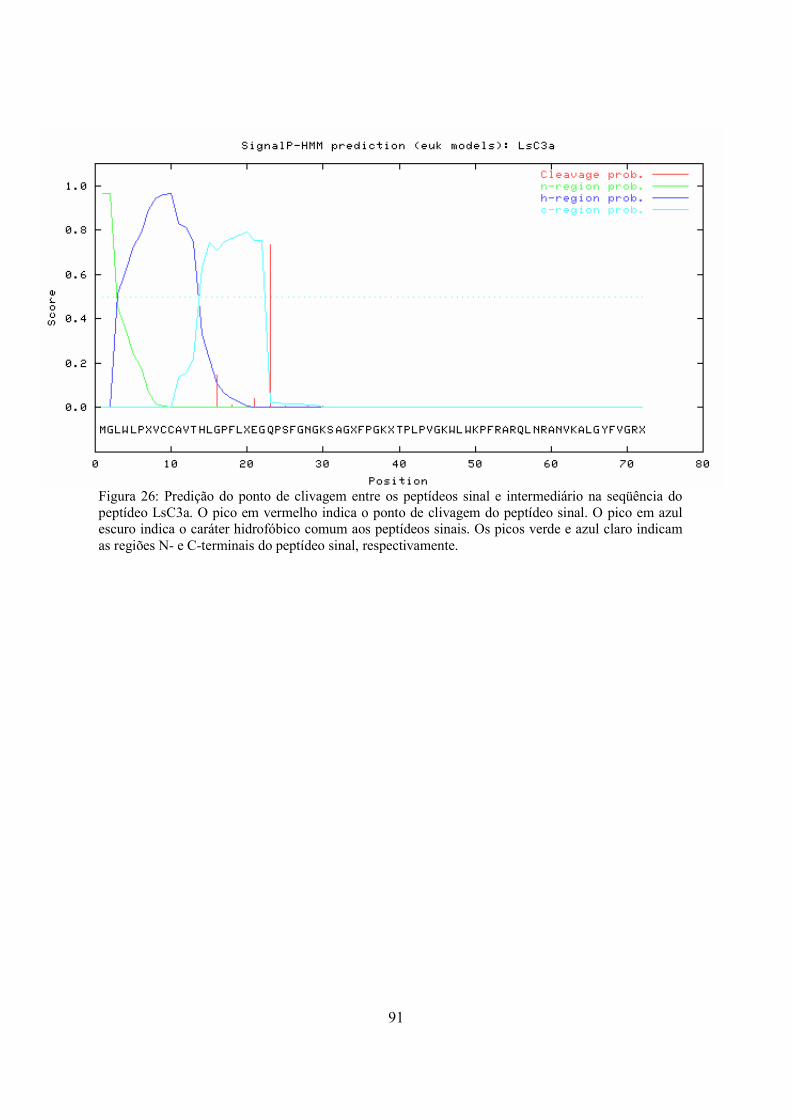
91
Figura 26: Predição do ponto de clivagem entre os peptídeos sinal e intermediário na seqüência do
peptídeo LsC3a. O pico em vermelho indica o ponto de clivagem do peptídeo sinal. O pico em azul
escuro indica o caráter hidrofóbico comum aos peptídeos sinais. Os picos verde e azul claro indicam
as regiões N- e C-terminais do peptídeo sinal, respectivamente.

92
Figura 27: Gráfico de hidropaticidade dos aminoácidos traduzidos a partir do cDNA dos clones que
codificam o precursor do componente clonado LsC3a. As regiões acima e abaixo do ponto 0 (seta
vermelha) indicam a presença de aminoácidos com características hidrofóbicas e hidrofílicas,
respectivamente. A região inicial hidrofóbica (~20 a.a.) sugere a existência de um peptídeo sinal na
seqüência peptídica de LsC3a.

93
7.8.2- Análise da seqüência do DNA complementar do clone 23
A seqüência nucleotídica do clone 23 apresenta 722 pares de bases. A tradução da
seqüência do cDNA do clone 23 utilizando o programa SIXFRAME revelou a existência de
uma janela de leitura no frame +1 (azul) e +2 (vermelho). A tradução nos frames +1 e +2
originou peptídeos compostos por 46 e 42 resíduos de aminoácidos, respectivamente. A
análise desses peptídeos pelo programa BlastP não revelou identidade com seqüências
protéicas depositadas no GenBank.
GAATTCGGCACCAGAGAAAGCTCAACTTGCAGTGCCCGGGAGGGGACAGCTGTTTTTGGAAAAATCCTCC GATGGTGAAATACCATGTCAAGTATATCTATGGAGAGTATGATCAATGTGTGGCCTTTACATGATGTAAA CCCTTCAGTCTCCATACTCATTTCTCTAGGTGATCCCACNAAAGCCCATTTCTCANTTAAGATACCTCTG AACGAGGAGGAGTGACCAAGATCTTTAGGTCACTCTGACTGTTCTGTGGCAGCAGAAGGTTGCCAATGGC AGCAAGGCACTGAGTAAGGGATTTCGTCTTGTGGATGAACTGATGGACCCCATGGTTGTCAGTCTGTTTA TCACTTGCACATCCATGTTTTGGTGGAAGGCAGATGGGCTGGCCACCTGGCTAAGGATAAAAAAACAGGG CCTGTTAGTTTCACTGGATCACTGCTTGTGAATCCTGCTTGACCAGAACTTATAGGGGTTGTAACTAATG ACACACTTATAATACAAAATCTCTTGTTATACTGGCAACTTTTTCTGGGAAGCATTTGTTTTGTTTGATA TTCAAATCAAATTTAGATTTGGCCCGTAACCAATGCCAATTACCAAAACTTATTGCTTTGTTCATGAAAA TACATACATGTATGTGTACTTACCTTTTCTTTTTGTTTGCCTGTTTTGAATAAACCAAACTTTAAGCAAA AAAAAAAAAAAAAAAACTCGAG
LsC4-a (Tradução no frame +1)
MSSISMESMI NVWPLHDVNP SVSILISLGD PTKAHFSXKI PLNEEE
LsC4-b (Tradução no frame +2)
MDPMVVSLFI TCTSMFWWKA DGLATWLRIK KQGLLVSLDH CL

94
7.8.3- Análise da seqüência do DNA complementar do clone 24
A seqüência nucleotídica do clone 24 apresenta 893 pares de bases. A tradução do
cDNA do clone 24 realizada pelo programa SIXFRAME procedeu-se no frame +1 e
originou um peptídeo de 62 aminoácido denominado LsC5. A análise desse peptídeo pelo
programa BlastP não revelou identidade com outras proteínas depositadas no GenBank.
AAAATGTCGCGNCAATCCCTTATCTGATGCCCCAGGGTACCTGNCTCCGAATATTTCCAAGNCCCCAGGT GTGGGCCATAAAGANGTCAAAGTTGTCTTGATCCNAAACAAGAGGGCTTAATGTCAATAGGAAAGGAATG TGAGNNCGTCTTCAATCCCCCGGGGACCGAGAAGGTTTTTTCGTCACCCCGGATATTTAATTTTGGCCCC TTCAGAATATGGAAAGATATTTTGTGGGATAGTCCTTGTGTGTTGCGCCCAGAAATTAATGACGGGAGGG GTTATCCAACAGGCCGTGCATTTCAATAAAGATGGCTCCCTGGGGTTTTCCGTTTGACCAAAAATGATCG TTAAGAAATTAAAATGAAAATTGAAGACGAGTTGAATGAGGACTTATTCCACAAGTCATTCTAGAAACTA AGGGAATACCATATAACCTCACAGGCAAGAAACTGGAAAAGTTGTTTCCAAAGATCATTGATTACAATCA GATTCCAGAAGTGAACAATGTGAAAAATCCGTATTGCCNGAAATCTTTCCTGAATATACCTGAACTTCAG AACTTCTAGGTGAAATGAAATGAGAAACGGGACTTGTTTATTTCTGAATTTTCTTGTTTTCTAATTGTAA CACACGTGATATCCATCTCTGTCGATAGCGAGCTTCAATGAACAGGAATTTTCAAAAATTAGGCCATTGG AGTAACTCAGTGGCATAAAAAAAACAATAATATGCTAAACCCCAAGATAAATAAAAGAATAATAATTTAA GNTGAGATGGAAAACTCGAGAGTAGCTTCTTAGAGCGGCCCGCGGGCCCATCGATTTTCCACCCGGTGGG TTACACAGCTAAGTTACCCATTCGCCTATATCANCGTATGCATCATGGCANTG
LsC5
MSIGKECEXVFNPPGTEKVFSSPRIFNFGPFRIWKDILWDSPCVLRPEINDGRGYPTGRAFQ

96
8- Discussão
Os estudos realizados com venenos de aranhas mostram que a maioria das toxinas
de aranhas descritas até o momento tem ação sobre receptores neurais, canais iônicos ou
proteínas de membranas pré-sinápticas envolvidas na liberação de neurotransmissores.
Embora haja poucos estudos envolvendo componentes tóxicos do veneno da aranha
Lasiodora sp, sabe-se que o veneno é capaz de inibir canais de Ca2+ do tipo L e modular a
cinética de canais de Na+ bem como a entrada de íons para dentro da célula (Kushmerick e
cols., 2001). Os estudos realizados por Kalapothakis e cols. (2003) sobre os efeitos do
veneno da Lasiodora sp em coração isolado de rato sugeriram que o veneno dessa aranha
possa estar envolvido no aumento da liberação vesicular de acetilcolina da extremidade de
nervos parasimpáticos.
Os peptídeos compreendem a principal classe de componentes tóxicos dos venenos
de aranhas, e apresentam uma massa molecular geralmente entre 4 e 10kDa (Savel-
Niemann, 1989). O isolamento e a caracterização funcional desses peptídeos têm sido
descritos em vários trabalhos, contudo, informações acerca da estrutura primária desses
peptídeos ainda são escassas e limitadas a poucas espécies.
Este trabalho foi desenvolvido com o objetivo de se poder inferir a seqüência de
aminoácidos de componentes do veneno bem como expressar as proteínas utilizando-se de
diferentes sistemas, dado a dificuldade de se obter grandes quantidades de veneno bruto.

97
Para a realização da varredura dos clones provenientes da excisão em massa do
fagemídeo pBK-CMV adotou-se a técnica de ELISA. Esta técnica foi escolhida devido a
sua sensibilidade e especificidade na detecção da presença do antígeno. Esse sistema de
varredura foi utilizado na identificação de proteínas imunogênicas de venenos de outros
animais, como por exemplo, os escorpiões Tityus bahiensis e Tityus serrulatus, podendo
gerar uma grande quantidade de informação em um curto período de tempo bem como
contribuir não só para projetos genomas como também para projetos proteomas, por meio
da identificação de proteínas e toxinas importantes, e para o desenvolvimento de
antivenenos para uso clinico (Kalapothakis e cols. 2001).
O seqüenciamento dos clones que apresentaram similaridade com toxinas de veneno
de outras aranhas forneceu a seqüência completa de nucleotídeos que codificam para as
toxinas denominadas LTx1, LTx2, LTx3 e LTx5. Todos os peptídeos precursores
apresentaram a mesma organização estrutural: peptídeo sinal, um peptídeo intermediário e
uma cadeia polipeptídica correspondente à toxina madura. Essa organização tem sido
encontrada em polipeptídeos tóxicos de venenos de outras aranhas e parece ser uma
característica estrutural comum a toxinas de aranhas (Cardoso e cols., 2003; Kalapothakis e
cols., 2002; Penaforte e cols., 2000).
Doze clones codificam seqüências de aminoácidos idênticas à seqüência da toxina
LTx1, um clone apresentou a seqüência codificante para LTx2, dois clones mostraram
codificar para a toxina LTx3 (Vieira e cols., 2004) e quatro clones codificam a toxina
LTx5. O fato do cDNA que codifica a toxina LTx1 ter sido encontrado 12 vezes sugere
uma maior imunogenicidade para LTx1 ou mesmo que o nível de expressão dessa toxina
seja maior, e que a mesma possa desempenhar um papel importante na paralisação e/ou
morte das presas. Essas hipóteses têm sido reforçadas ainda mais pelos achados de Moura e
cols. (2004). Utilizando anticorpos anti-P3, Moura e cols. realizaram a varredura de outras
sub-bibliotecas de cDNA da aranha Lasiodora sp e encontrou 11 clones com similaridade
com toxinas de aranhas, sendo que sete deles codificam para a toxina LTx1.
Os precursores das toxinas LTx1, LTx2 e LTx3, deduzidos a partir das seqüências
de cDNAs apresentaram um peptídeo sinal hidrofóbico, um peptídeo intermediário rico em
resíduos de glutamato e um peptídeo maduro, composto por 49 resíduos de aminoácidos
muito similares entre si diferindo-se em apenas 1-3 resíduos de aminoácidos. A

98
comparação das seqüências de aminoácidos das toxinas maduras LTx1 e LTx2 mostra que
elas se diferem em três resíduos de aminoácidos (posições 1, 25 e 49 – F/L, K/D e F/I). As
toxinas LTx1 e LTx3 diferem-se apenas no resíduo 25 (K/D). Entre as toxinas LTx2 e
LTx3 foram observadas discrepâncias nos resíduos 1 e 49 (L/F e I/F). As toxinas LTx2 e
LTx3 provavelmente exercem funções similares uma vez que os aminoácidos discrepantes
compartilham as mesmas características químicas (não polares). A discrepância observada
no resíduo da posição 25 (K�D) pode conferir às toxinas LTx1 e LTx3 diferentes modos
de ação, sobretudo no que diz respeito a alvos (canais iônicos e/ou receptores) ou
mecanismos de ação dessas toxinas. Estudos realizados com peptídeos de venenos de
tarântulas utilizando espectrometria de massa MALDI-TOF têm revelado que a diversidade
farmacológica dos peptídeos tóxicos está baseada em peptídeos de tamanhos pequenos e
com um número limitado de modelos estruturais (Escoubas & Rash, 2004), de tal maneira
que mesmo diferenças sutis (como a troca de um único aminoácido) na estrutura primária
dessas toxinas podem resultar em diferentes estruturas tridimensionais e conseqüentemente,
em diferentes funções. Isoformas que se diferem em 1 ou 2 resíduos de aminoácidos
encontradas em venenos de aranhas têm sido descritas por outros autores. HaTx1/HaTx2
(Swarts & MacKinnon, 1995), HnTxIV/HnTxV (Liu e cols., 2003; Xiao & Liang, 2003) e
ESTx1/ESTx2 (Savel-Niemann, 1989) são exemplos de isoformas que se diferem em
apenas 1 resíduo de aminoácido; LpTx1/LpTx2 (Escoubas e cols., 1997) diferem em 2
resíduos de aminoácidos.
O tamanho das toxinas descritas neste trabalho, 49 (LTx1, LTx2 e LTx3) e 71
(LTx5) aminoácidos, está de acordo com dados obtidos a partir de estudos de evolução
molecular utilizando espectrometria de massa MALDI-TOF realizado com venenos de 55
tarântulas (Escoubas & Rash, 2004). Esse estudo mostrou uma distribuição bi-modal de
pesos moleculares, onde 57,8% dos peptídeos apresentaram pesos moleculares entre 3500-
4500 Da e 6,9% dos peptídeos possuem massa entre 6500-7000 Da (Figura 28). Isto
significa que a maioria dos peptídeos de venenos de tarântulas possui um tamanho que
varia em média de 31-40 resíduos de aminoácidos (Escoubas & Rash, 2004).

99
Figura 28: Perfil molecular de peptídeos de venenos de 55 tarântulas utilizando espectrometria de
massa MALDI-TOF. A curva representa a freqüência de distribuição de pesos moleculares em
classes de 500 Da para as 1516 massas de peptídeos analisados, mostrando a distribuição bi-modal
(setas). As linhas representam números teóricos de aminoácidos para as classes de pesos
moleculares correspondentes (Adaptado de Escoubas & Rash, 2004).
A figura 29 mostra o alinhamento das seqüências de aminoácidos das toxinas
maduras da aranha Lasiodora sp com a seqüência de duas outras toxinas (LpTx1 e LpTx2)
da aranha Lasiodora parahybana, também de 49 resíduos de aminoácidos, previamente
descritas por Escoubas e cols. (1997). Uma elevada similaridade foi obtida entre as toxinas
isoladas do veneno da aranha Lasiodora parahybana (LpTx1 e LpTx2) e as toxinas
descritas neste trabalho (LTx1, LTx2 e LTx3). A toxina LTx1, descrita neste trabalho, é
idêntica à LpTx1 descrita por Escoubas e cols. (1997). Contudo, LTx2 apresenta diferença
em três resíduos de aminoácidos em relação à LpTx1 (posições 1, 25 e 49) e em 4 resíduos
quando alinhada com LpTx2 (posições 1, 6, 25 e 49). A toxina LTx3 difere de LpTx1
apenas no resíduo da posição 25 e difere da LpTx2 em dois resíduos de aminoácidos
(posições 6 e 25). Escoubas e cols. (1997) mostraram que as frações do veneno da aranha
Lasiodora parahybana tóxicas para insetos contêm peso molecular que varia entre 5000-
Classe Principal
Classe Secundária

100
6000 Da. A alta similaridade entre as toxinas descritas neste trabalho e as toxinas da
Lasiodora parahybana descritas por Escoubas e cols. (1997) poderia levar à especulação de
que se trata da mesma espécie de aranha. Contudo, a espécie Lasiodora parahybana não
tem sido descrita no estado de Minas Gerais, sendo nativa da região Nordeste do Brasil em
áreas como Campina Grande no Estado da Paraíba. Além do mais, pequenas diferenças ou
até 100% de identidade tem sido descrita na literatura entre toxinas de aranhas de espécies
distintas. Esse é o caso da toxina µ-Aga-III isolada do veneno da aranha Agenolepsis aperta
(Skinner e cols., 1989) que é idêntica à toxina CT-II, purificada da aranha Hololena curta
(Stapleton e cols., 1990). As toxinas GsMTx2/PaTx2 (Grammostola
spatulata/Phrixotrichus auratus) (Diochot e cols., 1999; Oswald e cols., 2002) e
TxP1/ESTx2 (Brachypelma smithi/Erypelma californicum) (Savel-Niemann, 1989; Kaiser
e cols., 1994) têm seqüências idênticas, mas foram descritas em diferentes, embora
relacionadas, espécies de aranhas.
1 6 25 49
LTx1 FFECTFECDIKKEGKPCKPKGCKCKDKDNKDHKKCSGGWRCKLKLCLKF
LTx2 LFECTFECDIKKEGKPCKPKGCKCDDKDNKDHKKCSGGWRCKLKLCLKI
LTx3 FFECTFECDIKKEGKPCKPKGCKCDDKDNKDHKKCSGGWRCKLKLCLKF
LpTx1 FFECTFECDIKKEGKPCKPKGCKCKDKDNKDHKKCSGGWRCKLKLCLKF
LpTx2 FFECTLECDIKKEGKPCKPKGCKCNDKDNKDHKKCSGGWRCKLKLCLKF
Figura 29: Alinhamento das seqüências peptídicas das toxinas maduras das aranhas Lasiodora sp e
Lasiodora parahybana. Em azul são mostrados domínios conservados em todas as cinco toxinas.
As posições conservadas dos resíduos de cisteína (em vermelho) nas diferentes seqüências sugerem
que todas podem compartilhar semelhanças na estrutura tridimensional. Os aminoácidos que se
diferem em pelo menos uma delas são mostrados em preto.

101
As toxinas LTx1, LTx2 e LTx3 apresentaram aproximadamente 65% de identidade
com a seqüência de aminoácidos da toxina TXP1, uma neurotoxina descrita por Kaiser e
cols. (1994) purificada do veneno da tarântula mexicana “red knee” (Brachypelma smithii),
e ESTx, uma toxina presente no veneno da aranha Eurypelma californicum descrita por
(Savel-Niemann, 1989). A elevada similaridade compartilhada pelas toxinas LTx1, LTx2,
LTx3, TXP1 e ESTx está relacionada, provavelmente, com o fato dessas aranhas
pertencerem à mesma família, Theraphosidae.
A maioria das seqüências primárias de toxinas de venenos de tarântulas
identificadas até o momento possui em média 31-41 aminoácidos, reticulados por três
pontes dissulfeto (Escoubas & Rash, 2004). O alinhamento das lasiotoxinas (LpTx1 e
LpTx2) descritas para L. parahybana com as toxinas ESTx e HwTx-II (Figura 30) indica
que elas podem adotar uma conformação tridimensional parecida, modificada por um
trecho de 10-13 aminoácidos extras e uma ponte dissulfeto adicional. A conservação dos
resíduos de cisteína sugere que ao longo da evolução muitos domínios protéicos tendem a
ser conservados, e que pequenas modificações de resíduos de aminoácidos observadas nas
seqüências dessas proteínas são suficientes para conferir atividades diferentes. A estrutura
de proteínas tóxicas de escorpiões e serpentes tende a conservar o arranjo das pontes de
dissulfeto e modificar as seqüências peptídicas que estão expostas na superfície da
molécula (Menez e cols., 1992). Ao que parece, os peptídeos tóxicos de aranhas apresentam
a mesma tendência evolutiva. O mecanismo evolucionário envolvido no aumento da
diversidade de toxinas animais parece estar relacionado com mutações pontuais seguidas de
rápidas duplicações gênicas e recombinações. Modificações pós-traducionais do produto
gênico também levam ao aumento de peptídeos e proteínas tóxicos (Mebs, 2001). Um
fenômeno paradoxal que venenos animais podem ter em comum é o fato das regiões não
codificantes de genes de toxinas (regiões 3’ e 5’-não traduzidas e íntrons) apresentarem
elementos estruturais altamente conservados enquanto que partes codificantes (éxons)
freqüentemente exibem grande variabilidade, contrário ao que ocorre normalmente em
outros genes (Mebs, 2001; Afifiyan e cols., 1999; Chuman e cols., 2000; Kuch e cols.,
2003; Chang e cols., 2003; Tani e cols., 2002).

102
Figura 30: Alinhamento (ClustalW) de toxinas de tarântulas. As prováveis interações entre os
resíduos de cisteínas das toxinas HwTx-II (Ile/Gln), ESTx1, ESTx2/TxP1, LpTx1 e LpTx2 são
mostradas pelos algarismos romanos (Adaptado de Escoubas & Rash, 2004).
A análise da expressão de LTx1 revelou uma indução basal (tempo 0) e um tempo
ótimo de indução em torno de 5 horas após a adição de IPTG 0.6mM e incubação a 30ºC
(200 rpm). A toxina recombinante foi também analisada por western bloting juntamente
com uma fração do veneno bruto de Lasiodora sp. A toxina LTx1 foi encontrada sobretudo
na fração solúvel sugerindo que não há formação de corpos de inclusões uma vez que não
foi observada a presença mensurável da mesma no precipitado da sonicação de células
induzidas.
Quatro clones positivos em ensaios imunológicos de ELISA codificam o precursor
da toxina LTx5. A toxina LTx5 é sintetizada na glândula de veneno da aranha Lasiodora sp
inicialmente como um pré-peptídeo contendo um peptídeo sinal hidrofóbico composto por
17 resíduos de aminoácido, um peptídeo intermediário contendo 28 resíduos de
aminoácidos e o peptídeo maduro composto por 71 resíduos de aminoácidos. De acordo
com o perfil molecular do veneno de 55 tarântulas obtido por Escoubas & Rash (2004),
LTx5 pertence à segunda classe de peptídeos encontrados em glândula de veneno de
tarântulas, com peso molecular variando em torno de 7000-8000 Da (Figura 28).
O alinhamento das toxinas da aranha Lasiodora sp LTx1, LTx2, LTx3, LTx5
(descritas neste trabalho) e LTx4 (descrita por Moura e cols., 2004) (Figura 31) sugere
(baseado na similaridade entre as seqüências), a existência de pelo menos três famílias de

103
proteínas de baixo pelo molecular codificadas na glândula de veneno da aranha em questão
(Figura 32).
Figura 31: Alinhamento dos precursores das toxinas da aranha Lasiodora sp LTx1, LTx2, LTx3,
LTx4 e LTx5. Os resíduos destacados em roxo são comuns a todas as seqüências. Os resíduos
conservados nas seqüências de LTx1, LTx2 e LTx3 (azul claro) sugere que essas toxinas
compartilham a mesma família de proteínas. A ausência de domínios conservados entre LTx4
(Moura e cols., 2004 dados não publicados), LTx5 e as três primeiras toxinas sugere a existência de
pelo menos três famílias de proteínas de baixo pelo molecular codificadas na glândula de veneno da
aranha Lasiodora sp.

104
Figura 32: Dendograma das toxinas da aranha Lasiodora sp baseada no programa de alinhamento
ClustalW. Pelo menos três famílias de proteínas parecem ser codificadas na glândula de veneno da
aranha em questão.
A análise do peptídeo codificado pelo cDNA do clone 20 apresentou elevada
similaridade com cadeias β de globinas de diferentes organismos. Embora a probabilidade
de contaminação, no momento da construção da biblioteca de cDNA, com RNAs de
animais utilizados na alimentação das aranhas não seja descartada, fragmentos de
hemoglobina com atividade antimicrobial têm sido descritos. Fogaça e cols. (2000)
identificaram um fragmento da cadeia α da hemoglobina bovina referente à região
compreendida entre os resíduos 33 ao 61, com propriedades antimicrobianas. Esse
peptídeo, com massa molecular de 3.205,6 Da, foi inicialmente isolado do intestino do
carrapato Boophilus microplus. Observou-se que o fragmento da cadeia α da hemoglobina
bovina age em concentrações micromolares apenas contra bactérias Gram-positivas e
contra fungos, não tendo sido ativo contra bactérias Gram-negativas (Fogaça e cols. 2000).
Cadeias de hemoglobina têm sido descritas como uma fonte de peptídeos bioativos
endógenos. Froidevaux e cols. (2001) têm obtido diferentes peptídeos a partir da hidrólise

105
peptídica de hemoglobina bovina. Esses autores obtiveram um fragmento correspondente
aos primeiros 23 resíduos de aminoácidos da extremidade N-terminal da cadeia α da
hemoglobina bovina e testaram sua atividade contra Micrococcus luteus. A inibição do
crescimento das células microbianas foi obtida a uma concentração de 1,5mg/ml (671µM)
de peptídeo α 1-23. A clivagem das subunidades α e β de hemoglobina (inclusive a
humana) utilizando-se brometo de cianogênio tem fornecido fragmentos com atividade
antimicrobial mensuráveis (Parish e cols., 2001). Regiões específicas da molécula de
hemoglobina humana, responsáveis por atividade antimicrobiana, têm sido mapeadas. Os
trinta aminoácidos da extremidade carboxila da subunidade β, responsáveis pela formação
de uma α-hélice catiônica baseada na estrutura cristalina do tetrâmero intacto, apresenta
atividade contra Escherichia coli, Staphilococcus aureus e Candida albicans (Parish e
cols., 2001). Os autores postulam que, em adição à sua função no transporte de oxigênio, a
hemoglobina age como um importante agente antibiótico contra um amplo espectro de
microrganismos.
A análise da seqüência do cDNA do clone 21 apresentou 62% de similaridade com
a proteína CG7580-PA da mosca Drosophila melanogaster, 58% de similaridade com a
proteína agCP10691 do mosquito Anopheles gambiae e 46% de similaridade com a
proteína Qpc-pending do camundongo Mus musculus. Nenhuma atividade tóxica tem sido
descrita para os peptídeos com os quais o cDNA do clone 21 apresentou similaridade.

107
9- Conclusões
� A varredura da biblioteca de cDNA, utilizando-se da técnica de ELISA,
permitiu o isolamento de clones que apresentaram insertos que codificam
seqüências com similaridade com toxinas das aranhas Brachypelma smithii
(TxP1), Eurypelma californicum (ESTx), Selenocosmia huwena (HwTx-II),
Lasiodora parahibana (LpTx1 e LpTx2) e Macrothele gigas (Magi 7).
� As seqüências primárias dos peptídeos precursores para LTx1, LTx2, LTx3 e
LTx5 apresentaram a mesma organização: um peptídeo sinal hidrofóbico
localizado na extremidade N-terminal, um peptídeo intermediário e uma região
correspondente à seqüência da toxina madura.
� O tamanho das toxinas descritas neste trabalho, 49 (LTx1, LTx2 e LTx3) e 71
(LTx5) resíduos de aminoácidos, está em acordo com dados obtidos a partir de
estudos de evolução molecular realizados com venenos de aranhas.
� A análise das seqüências peptídicas de LTx1, LTx2 e LTx3 sugere uma
estrutura organizacional que compreende a presença de quatro ligações
dissulfeto, enquanto a toxina LTx5 deve apresentar apenas três pontes
dissulfeto.
� O cDNA que codifica a toxina LTx1 foi encontrado em 12 clones, em um total
de 19 clones positivos com similaridades com toxinas de outras aranhas. Esse
resultado sugere que LTx1 seja mais imunogênica ou mesmo que LTx1 tenha
um nível de expressão maior.

108
� As toxinas LTx2 e LTx3 provavelmente possuem atividades similares uma vez
que os aminoácidos discrepantes entre as toxinas (posições: 1 e 49)
compartilham as mesmas características químicas, enquanto que a discrepância
observada entre as toxinas LTx1 e LTx2/LTx3 (K�D) pode conferir à LTx1
uma atividade distinta.
� Uma elevada similaridade foi obtida entre as toxinas isoladas do veneno da
aranha Lasiodora sp (LTx1, LTx2 e LTx3) e as toxinas LpTx1, LpTx2 descritas
para L. parahybana, sendo a toxina LTx1 idêntica à LpTx1.
� O alinhamento das toxinas da aranha Lasiodora sp LTx1, LTx2, LTx3, LTx5 e
LTx4 sugere a existência de pelo menos três famílias de toxinas no veneno da
aranha.
� Os experimentos de indução da toxina LTx1 recombinante revelaram um tempo
ótimo de indução em torno de 5 horas e produziu a toxina na sua forma solúvel.

110
10- Perspectivas
Pouco se conhece sobre a composição qualitativa e/ou quantitativa do veneno da
Lasiodora sp. Neste sentido, pretendemos:
� Expressar em outros sistemas as diferentes toxinas identificadas.
� Purificar as toxinas recombinantes.
� Produzir anticorpos específicos contra as toxinas que se mostrarem
ativas. Estes anticorpos poderão ser usados para monitorar processos de
purificação, desenvolvimento de vacinas ou na varredura de outras sub-
bibliotecas.
� Proceder estudos funcionais: ensaios farmacológicos e eletrofisiológicos.
� Estudar a ação das toxinas recombinantes em insetos
� Realizar mutações sítio-dirigidas em aminoácidos que possam ser
críticos do ponto de vista toxicológico ou imunológico.

112
11. Referências Bibliográficas
AFIFIYAN, F.; ARMUGAN, A.; TAN, C. H.; GOPALAKRISHNAKONE, P.;
JEYASEELAN, K. (1999). Postsynaptic α-Neurotoxin gene of the spitting cobra, Naj
naja sputatrix: Structure, Organization, and Phylogenetic Analysis. Genome Research
9: 259-266.
ALTSCHUL, S. F.; GISH, W.; MILLER, W.; MYRES, E. W.; LIPMAN, D. J. (1990)
Basic local alignment search tool. J. Mol. Bio. 215:403-410.
ARAÚJO D. A. M., CORDEIRO M. N., DINIZ, C. R.; BEIRÃO S. L. (1993). Effects
of a toxic fraction, PhTx2, from de spider Phoneutria nigriventer on de sodium current.
Nauyn-Schmeidebergs Arch. Pharmacol. 347: 205-208.
ATKINSON R. K. & WRIGHT L. G. (1992a). The involvement of collagenase in the
necroses induced by the bites of same spiders. Comp. Biochem. Physiol. 102(1): 125-
128
ATKINSON R. K. & WRIGHT L. G. (1992b) The modes of action of spider toxins on
insects and mammals. Biochem. Physiol. 102c, No. 3, pp., 339-342.
BARRAL, J., POBLETTE, F., MENDONZA, E., PINEDA, J. C., GALARRAGA, E.;
BARGAS, J. (2001). High-affinity inhibition of glutamate release from corticostriatal
synapses by ω-agatoxin TK. European Journal of pharmacology. 430: 167-173.
BELOKONEVA, O. S., SATAKE, H., MAL’TSEVA, E. L., PAL’MINA, N. P.,
VILLEGAS, E., NAKAJIMA, T.; CORZO, G. (2004). Pore formation of
phospholipd membranes by the action of two hemolytic arachnid peptides of different
size. BBA 1664(2): 182-188.

113
BERGMANN, M. R.; PINEDO, F. J. R. (2001). Baculovírus recombinante para controle
de pragas. Biotecnologia 22: 50-58.
CARDOSO, F. C.; PACÍFICO, L. G.; CARVALHO, D. C.; VICTÓRIA, J. M. N.;
NEVES, A. L. G.; CHÁVEZ-OLÓRTEGUI, C.; GOMEZ, M. V.;
KALAPOTHAKIS, E. (2003). Molecular cloning and characterization of Phoneutria
nigriventer toxins active on calcium channels. Toxicon 41: 755-763.
CHAN, T. K., GEREN, C. R., HOWELL, D. E.; ODELL, G. V. (1975). Adenosine
Triphosphate in tarantula spider venoms and its synergistic effect with the venom
toxin.. Toxicon 13: 61-66.
CHANG, L. S.; CHUNG, C.; LIOU, J. C.; CHANG, C. W.; YANG, C. C. (2003).
Novel neurotoxins from Taiwan krait (Bungarus multicinctus) venom: purification,
characterization and gene organization. Toxicon 42: 323-330.
CHOMCZYNSKI, P. & SACCHI, N. (1987). Single-step method of RNA isolation by
acid guanidinum thiocyanate-phenol-Cloroform extraction. Anl. Biochem. 162: 156-
159.
CHUMAN, Y.; NOBUHISA, I.; OGAWA, T.; DESHIMARU, M.; CHIJIWA, T.;
TAN, N. H.; FUKUMAKI, Y.; SHIMOHIGASHI, Y.; DUCANCEL, F.;
BOULAIN, J. C.; MÉNEZ, A.; OHNO, M. (2000). Regional and accelerated
molecular evolution in group I snake venom gland phospholipase A2 isozymes. Toxicon
38: 449-462.
CORDEIRO, M. N., FIGUEIREDO, S. G., VALENTIN, A. C., DINIZ, C> R., VON
EICKSTEDT, R. D., GILROY, J.; RICHARDSON, M. (1993). Purificationand
amino acid sequences of six Tx3 type neurotoxins from the venom of the brazilian
“armed” spider Phoneutria nigriventer (Keys.) Toxicon 31: 35-42.

114
CORZO, G., GILLES, N., SATAKE, H., VILLEGAS, E, DAI, L., NAKAJIMA, T.;
HAUPT, J. (2003). Distinct primary structuresof the major peptides toxins from the
venom of the spider Macrothele gigas that bind to sites 3 and 4 in the sodium channel.
FEBS Letters 547: 43-50.
DE DEUS, A. D. T. (2003). Caracterização do veneno bruto da aranha Lasiodora sp.
Dissertação de mestrado – Instituto de Ciências Biológicas da Universidade Federal de
Minas Gerais.
DIOCHOT, S., DRICI, M. D., MOINIER, D., FINK, M. AND LAZDUNSKI, M.
(1999). Effects of phrixotoxins on the Kv4 family of potassium channels and
implications for the role of Ito 1 in cardiac electrogenesis. Br. J. Pharmacol. 126: 251-
263.
ESCOUBAS, P. & RASH, L. (2004). Tarântulas: eight-legged pharmacists and
combinatorial chemists. Toxicon 43: 555-574.
ESCOUBAS, P., CELERIER, M. L., ROMI-LEBRUN, R.; NAKAJIMA, T. (1997).
Two novel peptide neurotoxins from the venom on the tarantula Lasiodora parahibana.
Toxicon 35: 805-806.
ESCOUBAS, P., DIOCHOT, S.; CORZO, G. (2000). Structure and pharmacology of
spider venom neurotoxins. Biochimie 82: 893-907.
FIGUEREDO, S. G., GARCIA, M. E., VALENTIM, A C., CORDEIRO, M. N.,
DINIZ, C. R.; RICHARDSON, M. (1995). Purification and amino acid sequence of
the insecticidual neurotoxin Tx4(6-1) from the venom of the “armed”spider Phoneutria
nigriventer. Toxicon 33: 83-93.

115
FOGAÇA, A. C.; SILVA, Jr. P. I.; MIRANDA, M. T. M.; BIANCHI, A. G.;
MIRANDA, A. P.; RIBOLLA, P. E.; DAFFRE, S. (2000). Antimicrobial activityof a
bovine hemoglobin fragment in the tick Boophilus microplus. J. Biol. Chem. 274:
25330-25334.
FROIDEVAUX, R.; KRIER, F.; NEDJAR-ARROUME, N.; VERCAIGNE-MARKO,
D; KOSCIARZ, E.; RUCKEBUSCH, C.; DHULSTER, P.; GUILLOCHON, D.
(2001).Antibacterial activity of a pepsin-derived bovine hemoglobin fragment. FEBES
Letters. 491: 159-163.
GEREN C. R. & ODELL G. V. (1984). The biochemistry of spider venoms. In insects
poisons, Allergens, and other vertebrate venoms. Handbook of natural toxins (Edited
by Tu A. T.), Vol. 2, pp. 441-481. Marcel Dekker, New York.
HAEBERLI, S., KUHN-NENTWIG, L., SCHALLER, J.; NENTWIG, W. (2000).
Characterisation of antibacterial activity of peptides isolated from the venom of the
spider Cupiennius salei (Araneae: Ctenidae). Toxicon 38: 373-380.
HORNI, A., WEICKMANN, D.; HESSE, M. (2001). The main products of the low
molecular mass fraction in the venom of the spider Latrodectus menavodi. Toxicon 39:
425-428.
KAISER, I. I., GRIFFIN, P. R., AIRD, S. D., HUDIBURG, S., SHABANOWITZ, J.,
FRANCIS, B., JOHN, T. R., HUNT, D. F.; ODELL, G. V. (1994). Primary
structures of two proteins from the venom of the mexican red knee tarantula
(Brachypelma smithii). Toxicon 32 1083-1093.

116
KALAPOTHAKIS, E.; PENAFORTE, C. L.; LEÃO, R. M.; CRUZ, J. S.; PRADO, V.
F.; CORDEIRO, M. N., DINIZ, C. R,; ROMANO-SILVA, M. A.; PRADO, M. A.
M.; GOMEZ, M. V.; BEIRÃO, P. S. L. (1998). Cloning, cDNA sequence anlysis and
pach clamp studies of a toxin from the venom of the armed spider (Phoneutria
nigriventer). Toxicon. 36 (12):1971-1980.
KALAPOTHAKIS, E., JARDIM, S., MAGALHÃES, A. C., MENDES, T. M., DE
MARCO, L., AFONSO, L. C. C.; CHÁVEZ-OLÓRTEGUI, C. (2001). Screening of
expression libraries using ELISA: identification of immunogenic proteins from Tityus
bahiensis and Tityus serrulatus venom. Toxicon 39 679-685.
KALAPOTHAKIS, E., KUSHMERICK, C., GUSMÃO, D. R., FAVARON, G. O. C.,
FERREIRA, A. J., GÓMEZ, M. V.; ALMEIDA. A. P. (2003). Effects of the venom
of a Mygalomorph spider (Lasiodora sp) on the isolated rat heart. Toxicon 41 23-28.
KALAPOTHAKIS, E., SIMONE, C. A.,CIBELE, S. C., TAHIS, M. M., MARCUS, V.
G., OLDEMIR, C. M., IDA, C. G.; CARLOS CHÁVEZ-OLÓRTEGUI. (2002).
Molecular cloning, expression and imunológical properties of LiD1, a protein fron
dermonecrotic family of Loxosceles intermedia spider venoms. Toxicon 40: 1691-1699.
KUCH, U.; MOLLES, B. E.; OMORI-SATOH, T.; CHANHOME, L.; SAMEJIMA,
Y.; MEBS, D. (2003). Identification of alpha-bungarotoxin (A31) as the major
postsynaptic neurotoxin, and complete nucleotide identity of a genomic DNA of
Bungarus candidus from Java with exons of the Bungarus multicinctus alpha-
bungarotoxin (A31) gene. Toxicon 42: 381-390.
KUHN-NENTWIG, L., SCHALLER, J., STRB, B., KAMPFER, U.; NENTWIG, W.
(1998). CSTX-4, a novel bactericidal and insecticidal peptide in de venom of
Cupiennius salei. Toxicon 36: 1276-1277.

117
KUSHMERICK, C., CARVALHO, F. M., MARIA, M., MASSENSINI, A. R.,
ROMANO-SILVA, M. A., GÓMEZ, M. V., KALAPOTHAKIS, E.; PRADO, M.
A. M. (2001). Effects of a Lasiodora spider venom on Ca2+
and Na+
channels. Toxicon
39: 991-1002.
LAEMMLI, U. K. (1970). Cleavage of structural proteins during the assembly of the head
of bacteriophage T4. Nature, 227 (259): 680-685.
LIPKIN, A., KOZLOV, S., SOSYREVA, E., BLAKE, A., WINDASS, J. D.;
GRISHIN, E. (2002). Novel insecticidual toxins from the venom of the spider
Segestria florentina. Toxicon 40: 125-130.
LIU, Z., DAI, J., CHEN, Z., HU, W., XIAO, Y.; LIANG, S. (2003). Isolation and
characterization of hainantoxin-IV, a novel antagonist of tetrodotoxin-sensitive sodium
channels from the Chinese bird spider Selenocosmia hainana. Cell. Mol. Life Sci.
60:972-978.
LOHNER, K. (2001). The role of membrane lipid composition in cell targeting of
antimicrobial peptides. In “Development of novel antimicrobial agents: Emerging
strategies. Lohner, K. (ed), Horizon Scientifc Press, Englend. Pp 149-165.
MASSENSINI, A. R., SUCKLINK, J., BRAMMER, M. J., MORAES-SANTOS, T.,
GÓMEZ, M. V.; ROMANO-SILVA, M. A. (2002). Tracking sodium channels in live
cells: confocal imaging using fluorescetly labeled toxins. Journal of Neuroscience
Methods. 116: 189-196.
MEBS, D. (2001). Toxicity in animals. Trends in evolution? Toxicon 39: 87-96.
MENEZ, A.; BONTEMS, F.; ROUMESTAND, C.; GILQUIN, B.; TOMA, F. (1992).
Structural basis for functional diversity of animal toxins. Proc. Royal Soc. Edinburgh
99b: 83:103.

118
MOERMAN, L., VERDONCK, F., WILLEMS, J., TYTGAT, J.; BOSTEELS, S.
(2003). Antimicrobial peptides from scorpion venom induce Ca2+
signaling in HL-60
cells. BBRC 311: 90-97.
MOURA, M. B. (2004). Varredura de uma biblioteca de cDNA de Lasiodora sp
utilizando-se de anticorpos contra a fração P3 do veneno e caracterização de toxinas.
Dissertação de Mestrado. Núcleo de Pesquisa em Ciências Biológicas – Universidade
Federal de Ouro Preto – MG- Brasil.
OSWALD, R. E., SUCHYNA, T. M., MCFEETERS, R., GOTTLIEB, P.; SACHS, F.
(2002). Solution structure of peptide toxins that block mechanosensitive ion channels.
J.Biol. Chem. 277: 34443-34450.
PARISH, C. A.; JIANG, H.; TOKIWA, Y.; BEROVA, N.; NAKANISHI, K.;
McCABE, D.; ZUCKERMAN, W.; XIA, M. M.; GABAY, J. E. (2001). Broad-
Spectrum Antimicrobial Activity of Hemoglobin. Bioorganic & Medical Chemistry 9:
377-382.
PENAFORTE, C. L., PRADO, V. F., PRADO, M. ª M., ROMANO-SILVA, M. A.,
GUIMARÃES, P. E. M., MARCO, L., GÓMEZ, M. V.; KALAPOTAKIS, E.
(2000). Molecular cloning of cDNAs encoding insecticidual neurotoxic peptides from
the spider Phoneutria nigriventer. Toxicon 38: 1443-1449.
RASH, L. D & HODGSON, W. C. (2002). Pharmacology and biochemistry of spider
venoms. Toxicon 40: 225-254.
SAMBROOK, J.; FRITSH, E. F.; MANIATS, T. (1989). Molecular cloning: A
laboratory manual, 3 eds. Vol 1, 2, 3. Press Cold SpringHarbor, N.Y.

119
SANGER, F.; NICKLEN, S.; MANIATS, T. (1977). DNA sequencing with chain
termiation inhibitors. Proc. Natl. Acad. Sci. USA. 74 5463-5467.
SATAKE, H.; VILLEGAS, E.; OSHIRO, N.; TERADA, K.; SHINADA, T. ; CORZO,
G. (2004). Rapid and efficient identification of cysteine-rich peptides by random
screening of a venom gland cDNA library from the hexathelid spider Macrothele gigas.
Toxicon, 44: 149-156.
SAVEL-NIEMANN, A. (1989). Tarântula (Eurypelma californicum) Venom, a
Multicomponent System. Biol. Chem. Hoppe-Seyler 370: 485-498.
STUDIER, W.; ROSEMBERG, A. H.;DUNN, J. J.; DUBENDORFF, J. W. (1990). Use
of T7 RNA polimerase to direct expression of cloned genes. Methods of enzimology,
185: 60-89.
SHU, Q. & LIANG, S. P. (1999). Purification and characterization of huwentoxin-II, a
neurotoxic peptide from the venom of the Chinese bird spider Selenocosmia huwena. J.
Peptide Res. 53: 486-491.
SKINNER, W. S., ADAMS, M. E., QUISTAD, G. B., KATAOKA, H., CESARIN, B.
J., ENDERLIN, F. E.; SCHOOLEY, D. A. (1989). Purification and characterization
of two classes of neurotoxins from the funnel web, Agenolepis aperta. J. Biol. Chem.
264: 2150-2155.
STAPLETON, A., BLANKENSHIP, D. T., ACKERMAN, B. L., CHEN, T. M.,
GORDER, G. W., MANLEY, G. D., PALFREYMAN, M. G., COUTANT, J. E.;
CARDIN, A. D. (1990). Curtatoxins: neurotoxic insecticidal polypeptides isolated
from the funnel-web spider Hololena curta. J. Biol. Chem. 265: 2054-2059.
SWARTS, K. J., & MACKINNON, R. (1995). An inhibitor of the Kv2.1 potassium
channel isolated from the venom of a Chilean tarantula. Neuron 15: 941-949.

120
TAMBOURGI, D. V., PEDROSA, M. F. F., VAN DEN BERG, C. W., GONÇALVES-
DE-ANDRADE, R. M., FERRACINI, M., PAIXÃO-CAVALCANTE, D.,
MORGAN, B. P.; RUSHMERE, N. K. (2004). Molecular cloning, expression,
function and immunoreactivities of members of a gene family fosphingomyelinases
from Loxoceles venom glands. Molecular Immunology 41: 831-840.
TANI, A.; OGAWA, T.; NOSE, T.; NIKANDROV, N. N.; DESHIMARU, M.;
CHIJIWA, T.; CHANG, C. C.; FUKUMAKI, F.; OHNO, M. (2002).
Characterization, primary structure and molecular evolution of anticoagulant protein
from Agkistrodon actus venom. Toxicon 40: 803-813.
TEDFORD, H. W., SOLLOD, B. L., MAGGIO, F.; KING, G. F. (2004). Australian
funnel-web spiders: master insecticide chemists. Toxicon 43: 601-618.
TURKOV, M., RASHI, S., NOAM, Z.,GORDON, D., KHALIFA, R.
B.,STANKIEWICZ, M.,PELHATE, M.; GUREVITZ, M. (1997). In vitro folding
and functional analysis of an anti-insect selective scorpion depressant neurotoxin
produced in Escherichia coli. Protein Expression and Purification 10: 123-131.
USHKARYOV, Y. A., VOLYNSKI, K. E.; ASHTON, A. C. (2004). The multiple
actions of black widow spider toxins and their selective use in neurosecretion studies.
Toxicon 43: 527-542.
VIEIRA, A. L. G., MOURA, M. B., BABÁ, E. H., CHÁVEZ-OLÓRTEGUI, C.,
KALAPOTHAKIS, E.; CASTRO, I. M. (2004). Molecular cloning of toxins
expressed by the venom gland of Lasiodora sp. Toxicon 44: 949-952.
XIAO, Y. C. & LIANG, S. P. (2003). Purification and characterization of hainantoxin-V,
a tetrodotoxin-sensitive sodium channel inhibitor from the venom of the spider
Selenocosmia hainana. Toxicon 41: 643-650.

121
XU, K., JI, Y. & QU, X. (1989). Purification and characterization of an antibacterial
peptide from venom of Lycosa singoriensis. Acta Zoologica Sinica 35: 300-305.
YAN, L. & ADAMS, M. E. (1998). Lycotoxins, antimicrobial peptides from venom of the
wolf spider Lycosa carolinensis. Jornal of Biological Chemistry 273: 2059-2066.
ZLOTKIN, E., FISHMAN, Y.; ELAZAR, M. (2000). AaIT: From neurotoxin to
insecticide. Biochimie 82: 869-881.


















