

![EFEITO DA ATIVIDADE FSICA NA NEUROQUMICA DE … · Estudo da imunorreatividade das proteínas ligantes de cálcio na neuroquímica da medula ... o aumento da [Ca2+]i nos neurônios](https://static.fdocumentos.com/doc/165x107/5be67bc409d3f22b448db4b1/efeito-da-atividade-fsica-na-neuroqumica-de-estudo-da-imunorreatividade-das.jpg)





Línguas
Páginas
Legal


LEONARDO LUCAS DO NASCIMENTO SIQUEIRA
CARACTERIZAÇÃO CITOARQUITETÔNICA E NEUROQUÍMICA DO
SISTEMA DOPAMINÉRGICO MESENCEFÁLICO DE TARTARUGA
MARINHA (Chelonia mydas)
Dissertação apresentada ao Programa de Pós-
graduação em Biologia Estrutural e Funcional
da Universidade Federal do Rio Grande do
Norte
Natal-RN
2018

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
DEPARTAMENTO DE MORFOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA ESTRUTURAL E
FUNCIONAL
CARACTERIZAÇÃO CITOARQUITETÔNICA E NEUROQUÍMICA DO
SISTEMA DOPAMINÉRGICO MESENCEFÁLICO DE TARTARUGA
MARINHA (Chelonia mydas)
ORIENTADORA
Simone Almeida Gavilan
CO-ORIENTADOR
Expedito Silva do Nascimento Júnior
Natal-RN
2018

Siqueira, Leonardo Lucas do Nascimento. Caracterização citoarquitetônica e neuroquímica do sistemadopaminérgico mesencefálico de tartaruga marinha (Cheloniamydas) / Leonardo Lucas do Nascimento Siqueira. - Natal, 2018. 75 f.: il.
Dissertação (Mestrado) - Universidade Federal do Rio Grandedo Norte. Centro de Biociências. Programa de Pós-Graduação emBiologia Estrutural e Funcional. Orientadora: Profa. Dra. Simone Almeida Gavilan. Coorientador: Prof. Dr. Expedito Silva do Nascimento Júnior.
1. Quelônio - Dissertação. 2. Sistema nervoso - Dissertação.3. Citoarquitetura - Dissertação. 4. Neurotransmissores -Dissertação. 5. Dopamina - Dissertação. I. Gavilan, SimoneAlmeida. II. Nascimento Júnior, Expedito Silva do. III. Título.
RN/UF/BSE-CB CDU 598.13
Universidade Federal do Rio Grande do Norte - UFRNSistema de Bibliotecas - SISBI
Catalogação de Publicação na Fonte. UFRN - Biblioteca Setorial Prof. Leopoldo Nelson - Centro de Biociências - CB
Elaborado por KATIA REJANE DA SILVA - CRB-15/351

LEONARDO LUCAS DO NASCIMENTO SIQUEIRA
CARACTERIZAÇÃO CITOARQUITETÔNICA E
NEUROQUÍMICA DO SISTEMA DOPAMINÉRGICO
MESENCEFÁLICO DE TARTARUGA MARINHA (Chelonia mydas)
Dissertação apresentada ao Programa de Pós-
graduação em Biologia Estrutural e Funcional da
Universidade Federal do Rio Grande do Norte
Aprovado em: 15 de maio de 2018.
BANCA EXAMINADORA
____________________________________
Profa. Dra. Simone Almeida Gavilan
____________________________________
Prof. Dr. Expedito Silva do Nascimento Júnior
____________________________________
Prof. Dr. Flávio José de Lima Silva
Natal-RN
2018

AGRADECIMENTOS
Agradeço antes de tudo ao meu Deus que criou situações que me levaram a participar da
seleção de mestrado PPGBIOEF e por isso estou escrevendo esses agradecimentos agora. A
Ele toda glória e louvor por todas as coisas que faz, pela vida, por sua graça, amor e salvação.
Tudo começou no IFRN com a professora Mônica Almeida, irmã gêmea da professora
Simone. Então serei sempre grato pelo carinho e consideração que teve. Uma pessoa incrível
e sempre pronta a ajudar. Obrigado pela pessoa que és.
Por vários amigos do curso de Comércio Exterior que me incentivaram a seguir em frente em
um novo desafio. Em especial a minha amiga Regirlane e sua mãe, que me deram muito apoio
com conversas e orações.
A professora Simone Almeida que me recebeu muito bem no laboratório e aceitou me
orientar, me confiando desenvolver a pesquisa de um projeto desafiador e de muita
importância. Sou muito grato pela pessoa que és e a tenho como referência.
A minha amiga Alana, que me ajudou durante o processo seletivo para o mestrado. Ajudando-
me a revisar todo o assunto, mesmo não sendo da área. Obrigado pelo tempo que dedicou em
me ajudar.
Agradeço a toda equipe do Projeto Cetáceos da Costa Branca/UERN que acabou contribuindo
de várias maneiras para o desenvolvimento do projeto. Em especial aos que durante a
necropsia, retiraram os encéfalos utilizados neste trabalho. Sei o quanto é trabalhoso, obrigado
pela contribuição de vocês.
Obrigado a todos que fazem parte do laboratório de Morfofisiologia Comparada dos
Vertebrados. Obrigado a Rafael, Rebeca, Raquel e Silmara pelo tempo que vocês ofereceram
para me ajudar durante os procedimentos de laboratório.
Agradeço ao meu co-orientador, professor Expedito Nascimento, pelas orientações e pela
disponibilidade do laboratório de Neurociências (Labneuro). Sou grato a todos que fazem
parte do Labneuro, em especial, gostaria de destacar Regina pelo auxílio com os reagentes, a
Hélder, que deu o pontapé inicial e me ensinou como fazer os procedimentos, a Paulão, que
dedicou um pouco de seu tempo nas férias e me socorreu quando eu estava perdido na
imunoistoquima. Muito obrigados a todos.
Obrigado a minha família, pelo apoio. Em especial aos meus pais que sempre investiram na
educação de seus filhos. Obrigado mãe por criar todo o ambiente favorável para eu conseguir
me concentrar e por te levado café para mim todo o tempo (risos).
Agradeço a todos que de alguma forma contribuiu para que isso fosse possível.

LISTA DE ABREVIATURAS
CA: Catecolamina
DA: Dopamina
FLM: Fascículo longitudinal medial
GC: Griseum centrale
IP: Núcleo interpendicular
IS: Núcleo istmico
ISMC: Núcleo ístmico pars magnocelular
ISPC: Núcleo ístmico pars parvocelular
NIDT: Decussação tegmental do núcleo intersticial
nIII: Núcleo do nervo oculomotor
nIV: Núcleo do nervo troclear
nPM: Núcleo profundo mesencefálico
PV: Pendúnculo ventral do fascículo prosencefálico lateral
RA8: Homólogo reptiliano do grupamento A8 em mamíferos
Rs: Núcleo reticular superior
Ru: Núcleo rubro
SGC: Stratum griseum centrale
SGF: Stratum griseum et fibrosum superficiale
SGP: Stratum griseum profundum
SN: Substância negra
SNc: Subsância negra, pars compacta
SNr: Subsância negra, pars reticulata
SO: Stratum opticum
TSC: Núcleo central do torus semicircular
TT: Tracto tecto-talâmico
VTA: Área tegmental ventral

LISTA DE FIGURA
Figura 1. Síntese de dopamina, as enzimas envolvidas e sua liberação na fenda sináptica.
Fonte: Acervo, pessoal, 2018. .................................................................................................. 09
Figura 2. Representação de receptor de dopamina transmembana. Esquema da estrutura
putativa da isoforma de receptor D2R. A inserção de aminoácido D2L específica é indicada
junto com os locais potenciais. Resíduos aminoácidos potenciais envolvidos em N-
glicosilação, fosforilação de PKA, palmitoilação e ligantes de agonistas também são
mostrados. Fonte: VALLONE; PICETTI; BORRELI, 2000. .................................................. 11
Figura 3. Distribuição de grupos celulares neuronais DA em cérebro de roedor em
desenvolvimento (a) e adulto (b). Os neurônios dopaminérgicos no cérebro de mamíferos
estão localizados em nove grupos celulares distintos, distribuídos a partir do mesencéfalo ao
bulbo olfatório, como ilustrado esquematicamente, em uma visão sagital. As projeções
principais de grupos celulares DA estão ilustrados por flechas. Abreviações: Ige, eminência
ganglionar lateral; mge, eminência ganglionar medial; p1-p3, prosômeros 1-3. Fonte:
BJÖRKLUND; DUNNETT, 2007. .......................................................................................... 13
Figura 4. Secção transversa através da metade rostral do mesencéfalo, evidenciando
numerosas células fluoresecentes verdes (catecolaminérgicas) dentro do tegmento
mesencefálico da tartaruga pintada (Chrysemys picta) através de técnica de histofluorescência.
Aumento em 100 vezes. Fonte: PARENT; POIRIER, 1971. ................................................... 17
Figura 5. Mesencéfalo caudal. Corpos celulares da substância negra (grupo A9) e área
tegmental ventral (grupo A10) imunorreativos a tirosina hidroxilase em um lagarto não
tratado com colchicina. Um amplo feixe de fibras fluorescentes finas em seção transversal é
visível dorsomedial a substância negra. Transversalmente fibras cortadas mais espessas
podem ser vistas ventrolateral e ventromedial ao grupo celular CA da área tegmental ventral
(Barra: 150 μm). Neurônios, provavelmente dopaminérgicos, de tamanho médio na substância
negra. Neurônios, provavelmente dopaminérgicos, de tamanho pequeno e médio contendo TH
na área tegmental ventral. Fonte: WOLTERS et al., 1984 ....................................................... 18
Figura 6. Fotomicrografia de secção coronal através do mesencéfalo do lagarto Gekko gecko
mostrando neurônios e fibras imunorreativas a dopamina na área tegmentar ventral e porção
rostral da substância negra (E). Fonte: SMEETS; HOOGLAND; VOORN, 1986. ................. 19
Figura 7. Fotomicrografia mostrando células imunorreativas a dopamina no complexo
VTA/SN de Python regius. Note a distribuição aleatória dos processos das células DA na
porção lateral do complexo. Barra: 100 pm. Fonte: SMEETS, 1988 ....................................... 20
Figura 8. Localização da área de estudo, Costa Branca, RN, Brasil ....................................... 21
Figura 9. Células e fibras imunorreativas a TH no mesencéfalo do jacaretinga (Caiman
crocodilus) em nível de substância negra (A, C) e área ventral de Tsai (B, D). (B). Barra:
0,01 mm. Fonte: BRAUTH, 1988 ............................................................................................ 22
Figura 10. Fotomicrografia do mesencéfalo de tartaruga mostrando um subdivisão da
substância negra (pars compacta e pars reticulata) a partir da imunoistoquímica para TH.
Fonte: REINER, 2017. ............................................................................................................. 23

Figura 11. Representação de encéfalo de tartaruga marinha baseada em encéfalo de tartaruga
verde (Chelonia mydas) juvenil. O encéfalo pode ser dividido em três regiões: cérebro
anterior (telencéfalo e diencéfalo), cérebro médio (mesencéfalo) e cérebro posterior
(metencéfalo e mielencéfalo). Algumas estruturas do encéfalo podem ser observadas: (1)
Nervo olfatório; (2) bulbo olfatório; (3) cérebro; (4) nervo óptico; (5) nervo motor ocular
comum; (6) lóbulo óptico; (7) cerebelo; (8) medula; e (9) medula espinal. Fonte: Acervo
pessoal. ..................................................................................................................................... 25
Figura 12. Localização da área de estudo, Bacia Potiguar, RN, Brasil. ................................... 28
Figura 13. Exemplar juvenil da espécie Chelonia mydas. Fonte: WWF, 2018 ........................ 29
Figura 14. Distribuição de tartaruga-verde (Chelonia mydas) ao longo de regiões tropicais e
subtropicais de todo o mundo. Fonte: IUCN, 2017. ................................................................. 30
Figura 15. (A) Biólogo atendendo a encalhe de tartaruga marinha registrada durante
monitoramento junto com a equipe de resgate, responsável pela translocação do animal até a
Base de Reabilitação. (B) Recinto de tartarugas marinhas da Base de Reabilitação de Animais
Marinhos- PCCB-UERN, Praia de Upanema, Areia Branca. (C) Tartaruga marinha recebendo
atendimento da equipe médica veterinária do PCCB-UERN. (D) Animal com melhora
significativa em seu quadro clínico recebendo alimentação. (E e F) Soltura de tartarugas
marinhas aptas para a reintrodução ao seu habitat. Fonte: Projeto Cetáceos da Costa Branca –
PCCB/UERN, 2018. ................................................................................................................. 31
Figura 16. Unidade de Necropsia instalada na Base de Reabilitação de Animais Marinhos-
PCCB-UERN, Mossoró- RN. ................................................................................................... 32
Figura 17. Tartaruga-verde (Chelonia mydas) juvenil macho que veio a óbito. (A) Visão
dorsal. (B) Visão ventral. (C) Retirada de plastrão sem danificar os grandes vasos sanguíneos.
(D) Incisão de agulha no ventrículo para passagem de solução fixadora (paraformaldeído 4%)
pelo sistema sanguíneo do animal com auxílio de seringa. (E) Ruptura da região dorsal da
caixa craniana para acesso ao encéfalo; (F) visão lateral do encéfalo...................................... 34
Figura 18. Tartaruga adulta da espécie Chelonia mydas (tartaruga verde). (A) Ruptura da
região dorsal da caixa craniana para acesso ao encéfalo; (B) (C) visão geral do encéfalo em
relação à cabeça; (D) encéfalo retirado e encaminhado para o processo de pós-fixação. ........ 34
Figura 19. (A) Procedimento de corte do encéfalo utilizando criostato em temperatura média
de – 20°C e secções coronais de 30 μm. (B) Encéfalo de tartaruga marinha orientado em
posição longitudinal, em sentido rostrocaudal, e fixado a um suporte e envolvido por meio de
uma cola especial. Após a retirada dos cortes do criostato com o auxílio de um pincel, eles são
colocados em placas de ensaio de uso laboratorial. (C) A placa da esquerda contém solução
anticogelante e foi destinada as secções que passaram (imunoistoquímica para TH) ou
passarão (outros procedimentos) por métodos imunológicos, enquanto a placa da direita foi
destinada as secções que passaram pelo método de Nissl. O compartimento 1 recebeu as
secções na ordem a,b,c,d em diante, os compartimentos 2, 4, 5 e 6 foram transferidos para
recipientes com solução anticongelante e mantidos em temperatura menor que 0°C até serem
usados. ...................................................................................................................................... 35
Figura 20. (A) Secção coronal do mesencéfalo de tartaruga marinha imersa em solução
montagem para ser posicionada em lâmina histológica gelatinizada. (B) Cortes já organizados
em lâminas passando pelo procedimento de secagem em temperatura ambiente. (C) Lâminas
passando pelas baterias do método de Nissl ............................................................................. 38

Figura 21. Alguns dos procedimentos no método da imunoistoquímica. (A) Visão geral da
bancada exibindo ferramentas utilizadas, como ponteiras de pipetas, solução tampão fosfato
0,1 M, pH 7,4 e Triton X-100 contendo cloreto de sódio, placa de ensaio imunológico,
cestinha com fundo em tela para receber as secções e mantê-las em contato com as soluções
oferecidas e rotor. (B) Cortes dentro de cestinhas com solução tampão fosfato 0,1 M, pH 7,4
passando pelo procedimento das lavagens através de rotação. (C) Pipetagem de anticorpo
primário – uma das várias etapas em que se usam pipetas para colocar a quantidade correta,
medidas em μl de reagentes. (D) Secções do mesencéfalo na etapa de imersão em meio
contendo peróxido de hidrogênio como substrato e DAB como cromógeno para visualização
da reação, procedimento que ocorre dentro de capela e com maior atenção devido a toxicidade
do reagente DAB. ..................................................................................................................... 39
Figura 22. Encéfalo de tartaruga verde juvenil. De cima para baixo, são representadas as faces
lateral, dorsal e ventral do encéfalo e da esquerda para direita uma visão anteroposterior. (A)
Encéfalo de tartaruga juvenil mostrando as principais divisões da morfologia externa. ......... 42
Figura 23. Visão para sagital da região cefálica de tartaruga-verde juvenil exibindo a (A)
Morfologia externa do encéfalo e alguns (B) nervos craniais destacados pelas setas. ............. 44
Figura 24. Desenho esquemático de secção coronal através do mesencéfalo de tartaruga-verde
(Chelonia mydas), ilustrando a distribuição de corpos celulares dopaminérgicos
imunorreativos a TH e suas fibras. (A) Mesencéfalo rostral. (B) Mesencéfalo rostromedial.
(C) Mesencéfalo médiocaudal. (D) Mesencéfalo caudal. Observar o surgimento da área
tegmental ventral (VTA) e substância negra (SN) em porções mais rostrais e do grupamento
homólogo reptiliano A8 de mamíferos (RA8) em porções mais caudais. Fonte: Acervo
pessoal, 2018. ........................................................................................................................... 44
Figura 25. Tipos de neurônios (triangular multipolar, fusiforme e ovoide bipolar)
imunorreativos a tirosina hidroxilase encontrados ao longo do tegmento mesencefálico em
tartaruga-verde (Chelonia mydas). Fonte: Acervo pessoal, 2018. ........................................... 45
Figura 26. Fotomicrografias em campo claro de secções coronais em sentido rostrocaudal do
mesencéfalo de tartaruga-verde (Chelonia mydas) exibindo a citoarquitetura através do
método de Nissl (A, B, C, D e E) e evidenciando neurônios dopaminérgicos imunorreativos a
tirosina hidroxilase (A’, B’, C’, D’ e E’). Barra: 1000μm. ..................................................... 46
Figura 27. Fotomicrografias em campo claro de secções coronais rostrais do mesencéfalo de
tartaruga-verde (Chelonia mydas) com corpos de neurônios e fibras imunorreativas a tirosina
hidroxilase (TH+). (A) Substância negra. Barra: 500 μm. (B) Pars compacta da SN,
localizada em posição ventrolateral no tegmento mesencefálico, exibindo corpos de neurônios
e dendritos dispostos sem uma orientação de sentido único. (C) Pars reticulata da SN,
localizada em lateralmente a SNc, com um menor número de corpos de neurônios e fibras
longas dispostas, principalmente, em sentido mediolateral. B e C são caixas homônimas em
A. Barra: 100μm. ...................................................................................................................... 47
Figura 28. Fotomicrografias em campo claro de secções coronais rostromediais do
mesencéfalo de tartaruga-verde (Chelonia mydas) com corpos de neurônios e fibras
imunorreativas a tirosina hidroxilase (TH+). (A) Área tegmental ventral em posição
ventromedial e substância negra ventrolateral. Barra: 500 μm. (B) Pars compacta da SN
(SNc), localizada em posição ventrolateral no tegmento mesencefálico, exibindo corpos de
neurônios e dendritos dispostos sem uma orientação de sentido único e Pars reticulata da SN
(SNr), localizada lateralmente a SNc, com um menor número de corpos de neurônios e fibras

longas dispostas, principalmente, em sentido mediolateral. (C) Corpos de neurônios e suas
fibras na SNr. Seta indica neurônio do tipo triangular multipolar. B é caixa homônima de A.
Barra: 100μm ............................................................................................................................ 48
Figura 29. Fotomicrografias em campo claro de secções coronais mediocaudais do
mesencéfalo de tartaruga-verde (Chelonia mydas) com corpos de neurônios e fibras
imunorreativas a tirosina hidroxilase (TH+). (A e C) Equivalente reptiliano do grupamento
A8 em mamíferos (RA8). (B) RA8 com corpos de neurônios dispersos e prolongamentos
axonais com orientação, predominantemente, dorsoventral. (D) Seta indica neurônio ovoide
bipolar no grupamento RA8. B e D são caixas homônimas em A e C, respectivamente. (A)
Barra: 500 μm. (B, C e D) Barra: 100μm ................................................................................. 49
Figura 30. Fotomicrografias em campo claro de secções coronais caudais do mesencéfalo de
tartaruga-verde (Chelonia mydas) com corpos de neurônios e fibras imunorreativas a tirosina
hidroxilase (TH+). (A) Área tegmental ventral (VTA) localizada ventromedialmente no
tegmento mesencefálico (B) Seta inidica neurônio ovoide bipolar na VTA, observar fibras
curtas com disposição difusa. B é caixa homônima em A. Barra: 100μm ............................... 50

RESUMO
São conhecidas no mundo sete espécies de tartarugas marinhas, dentre essas, há registros de
ocorrência de cinco espécies no litoral do Brasil: Dermochelys coriacea, Chelonia mydas,
Caretta caretta, Eretmochelys imbricata e Lepidochelys olivacea. De acordo com a Lista
Vermelha da União Internacional para a Conservação da Natureza, todas as espécies de
tartarugas marinhas existentes no Brasil, se encontram ameaçadas de extinção, estando C.
mydas classificada como “Em perigo” e E. imbricata “Criticamente em perigo”. As espécies
C. caretta, L.olivacea e D. coriacea estão classificadas como “Vulneráveis”. Embora se saiba
sobre a morfologia externa do sistema nervoso de tartarugas marinhas, pouco se conhece
sobre a neurobiologia desses animais. O sistema dopaminérgico tem papel crítico em uma
grande variedade de funções e está presente no sistema nervoso central de todos os
vertebrados, sendo responsável pela neurotransmissão de dopamina. A dopamina é
reconhecida pela atividade que exerce no controle de processos complexos, como
programação da atividade motora e comportamentos motivados. Entendendo a importância
deste neurotransmissor e a carência de trabalhos que descrevam a citoarquitetura do encéfalo
de tartaruga marinha, esta pesquisa se propôs a caracterizar pela primeira vez os grupamentos
neuronais dopaminérgicos do mesencéfalo em tartaruga verde (C. mydas). Para isto, foram
utilizados dois métodos: o método de Nissl para descrever a citoarquitetura e a
imunoistoquímica para TH como forma de marcar os neurônios DA. Foram utilizados quatro
exemplares de C. mydas encalhadas que vieram a óbito na base de Reabilitação do Projeto
Cetáceos da Costa Branca ou que foi encontrado morto durante o monitoramento de praias
entre os anos de 2016 a 2017. Os animais foram necropsiados e retirados o encéfalo, tendo
passado pelo procedimento de perfusão (formol 4%) ou não (fixado em formol 10% por
imersão). Após fixados, os encéfalos foram mantidos em sacarose 30% até ser realizada a
microtomia através de criostato. As lâminas confeccionadas foram descritas utilizando
microscópio óptico, sendo realizadas capturas de imagens através de câmera acoplada ao
microscópio. O mesencéfalo de tartaruga verde apresenta um padrão de organização neuronal
semelhante ao encontrado em outros répteis. Os neurônios DA imunorreativos podem ser
divididos em um grupamento ventromedial (área tegmental ventral/A10), um grupamento
dorsolateral (substância negra/A9) e uma extensão caudal de A9 (homólogo reptiliano de A8
em mamíferos/RA8). Os neurônios destes grupamentos são bipolares ou multipolares e
ovoides, fusiformes ou triangulares em forma. Desta forma, viabilizamos as primeiras
descrições do sistema dopaminérgico que podem auxiliar no entendimento de questões
ecológicas e fornecer bases neuroanatômicas para futuros estudos funcionais da motricidade.
Palavras-chave: quelônio; sistema nervoso; citoarquitetura; neurotransmissores; dopamina.

ABSTRACT
Seven species of sea turtles are known in the world, among them, five species occur on the
coast of Brazil: Dermochelys coriacea, Chelonia mydas, Caretta caretta, Eretmochelys
imbricata and Lepidochelys olivacea. According to the Red List of the International Union for
the Conservation of Nature all species of sea turtles in Brazil are threatened with extinction,
with C. mydas classified as "Endangered" and E. imbricata "Critically endangered". The
species C. caretta, L.olivacea and D. coriacea are classified as "Vulnerable". Little is known
about the neurobiology of these animals. The dopaminergic system plays a critical role in a
wide variety of functions and is present in the central nervous system of all vertebrates and is
responsible for dopamine neurotransmission. Dopamine is recognized for its activity in the
control of complex processes, such as motor activity programming and motivated behaviors.
Understanding the importance of this neurotransmitter and the lack of studies describing the
cytoarchitecture of the marine turtle brain, this research aimed to characterize for the first time
the dopaminergic neuronal dopaminergic clusters of the midbrain in green turtle (C. mydas).
For this, two methods were used: the Nissl method to describe the cytoarchitecture and the
immunohistochemistry for TH as a way of marking DA neurons. Four C. mydas stranded
specimens were used. They died at the Rehabilitation base of the Projeto Cetáceos da Costa
Branca or were found dead during the monitoring of beaches between 2016 and 2017. The
animals were necropsied and the brain was removed, having undergone the perfusion
procedure (formalin 4%) or not (fixed in 10% formalin by immersion). After fixation, the
brains were maintained in 30% sucrose until the microtomy was performed by cryostat. The
prepared slides were described using an optical microscope, and images were taken through a
camera coupled to the microscope. The green turtle midbrain presents a pattern of neural
organization similar to that found in other reptiles. Immunoreactive DA neurons can be
divided into a ventromedial group (ventral tegmental area/A10), a dorsolateral group
(substantia nigra/A9) and a caudal extension of A9 (reptilian equivalent
of mammalian A8/RA8). The neurons of these groups are bipolar or multipolar and ovoid,
fusiform and triangular in shape. In this way, we enabled the first descriptions of the
dopaminergic system that can aid in the understanding of ecological issues and provide
neuroanatomic bases for future functional motor studies.
Keywords: chelonian; nervous system; cytoarchitecture; neurotransmitters; dopamine.

7
SUMÁRIO
Resumo ........................................................................................................................ 05
Abstract ........................................................................................................................ 06
1. Introdução .................................................................................................................... 08
1.1. Dopamina ..................................................................................................................... 08
1.2. Receptores Dopaminérgicos ........................................................................................ 10
1.3. Grupamentos Neuronais Dopaminérgicos ................................................................... 12
1.4. Grupamentos Neuronais Dopaminérgicos Mesencefálicos ......................................... 13
1.5. Grupamentos Neuronais Dopaminérgicos Mesencefálicos em Répteis ...................... 16
1.6. Tartaruga-marinha ....................................................................................................... 24
2. Objetivos ...................................................................................................................... 27
2.1. Objetivo Geral ............................................................................................................. 27
2.2. Objetivos Específicos .................................................................................................. 27
3. Materiais e Métodos .................................................................................................... 28
3.1. Área de Coleta ............................................................................................................. 28
3.2. Modelo Experimental .................................................................................................. 28
3.3. Coleta de Dados ........................................................................................................... 31
3.3.1. Remoção de encéfalo, fixação e armazenamento ........................................................ 32
3.3.2. Microtomia .................................................................................................................. 34
3.3.3. Método de Nissl ........................................................................................................... 36
3.3.4. Imunoistoquímica para Tirosina Hidroxilase .............................................................. 38
3.3.5. Obtenção das Imagens e Análise dos Resultados ........................................................ 39
3.3.6. Análise Estatística ........................................................................................................ 40
4. Resultados .................................................................................................................... 41
4.1. Morfologia Externa ...................................................................................................... 41
4.2. Microscopia ................................................................................................................. 43
5. Discussão ..................................................................................................................... 51
6. Conclusão .................................................................................................................... 55
7. Referências Bibliográficas .......................................................................................... 56

8
1. INTRODUÇÃO
1.1. Dopamina
A 3-hidroxitiramina/dopamina (DA) é uma monoamina pertencente ao grupo das
catecolaminas (CA) e atua como um dos neurotransmissores no sistema nervoso de
vertebrados. Catecolaminas é o termo que agrupa compostos orgânicos com uma estrutura de
anel benzeno com dois grupos hidroxilados adjacentes e uma cadeia amina oposta
lateralmente (SMEETS; GONZÁLEZ, 2000). A biossíntese de DA ocorre em via comum com
outras duas CAs: noradrenalina e adrenalina (PRAKASH; WURST, 2006). Dopamina,
noradrenalina e adrenalina constituem as CAs mais utilizadas pelo sistema nervoso. DA pode
ser tanto precursora para a síntese de outras CAs, como também pode desempenhar o papel de
um dos principais neurotransmissores de modulação no cérebro (JONES; PILOWSKI, 2002;
MARÍN et al., 2005).
Os neurônios em que se pode encontrar dopamina são rotulados como neurônios
dopaminérgicos. Em suas funções básicas, neurônios dopaminérgicos são capazes de
sintetizar, transmitir e recuperar a DA (RIDDLE, R.; POLLOCK, J. D, 2003). Para a síntese
deste neurotransmissor é utilizado o aminoácido tirosina como precursor. A presença de
tirosina no organismo é resultado, principalmente, de sua absorção através de dieta, mas
também, porque a enzima hepática fenilalanina-hidroxilase é capaz de hidroxilar fenilalanina
e a converter em tirosina. Quando a tirosina chega ao interior dos neurônios a biossíntese de
DA é iniciada. Através da ação de tirosina hidroxilase (TH), a tirosina é convertida em 3,4-
dihidroxifenilalanina (DOPA) (MARÍN et al., 2005; CHEN et al., 2008). Para o
funcionamento eficaz de TH intracelular, é necessário a participação do co-fator 6-
tetrahidrobiopterina (BH4), que tem sua síntese modulada pela enzima GTP-ciclohidrolase I
(GTPCHI) (NAGATSU; STJARNE, 1998; NAGATSU; ICHINOSE, 1999). Outra enzima, a
DOPA-descarboxilase, age sobre DOPA a convertendo em 3-hidroxitiramina/dopamina. A
dopamina ainda pode ser convertida em noradrenalina através da ação enzimática da
dopamina-beta-hidroxilase (NAGATSU et al., 1964; PRAKASH; WURST, 2006).
Finalizada sua síntese, a DA é armazenada em vesículas sinápticas por meio do
transportador vesicular de monoamina 2 (VMAT2), que se liga aos terminais pré-sinápticos e
torna as vesículas aptas para serem liberadas por exocitose após estímulo de íons Ca+²
(LAWLOR; DURING, 2004; BERNSTEIN et al., 2014). Após terminar sua atuação nos

9
receptores de neurônios pós-sinápticos, a DA é metabolizada na fenda sináptica pela ação da
enzima catecol-O-metiltransferase (COMT), podendo também, ser recapturada na fenda por
meio da ação do transportador dopaminérgico (DAT) para, posteriormente, ser metabolizada
através da mono-aminooxidase (MAO) ou rearmazenada em vesículas pela VMAT2 (SHIH et
al., 2006; GOOLE; AMIGHI, 2009) (Figura 1).
Figura 1. Síntese de dopamina, as enzimas envolvidas e sua liberação na fenda sináptica. Fonte: Acervo pessoal,
2018.
A DA é reconhecida pela atividade que exerce no controle de processos complexos.
Dentre suas atuações, pode-se citar a modulação sobre os passos iniciais de percepção
sensorial no bulbo olfatório e retina, participação na programação motora, aprendizado,
memória, processos afetivos e motivacionais, controle da temperatura corporal, ingestão de
alimentos, função renal e motilidade intestinal (BONELLI, 2007; YAMAMOTO; VERNIER,
2011; DE LA MORA et al., 2010; URBAN et al., 2012; SCHULTZ, 2013; GONZALEZ et
al., 2013; PACHECO et al., 2014; BARROT, 2014; IKEMOTO et al., 2015). Novas
evidências ainda apontam a DA como um regulador chave no processo inflamatório (YAN et
al., 2015).
O estudo de neurônios catecolaminérgicos (que engloba os neurônios dopaminérgicos,
adrenérgicos e noradrenérgicos) começou por volta de 1960 com as pesquisas de Carlsson,
Falck e Hillarp utilizando o método de histofluorêscencia de formaldeído (CARLSSON,
1962). Estes pesquisadores foram os pioneiros na identificação das duas catecolaminas
primárias: noradrenalina e dopamina. Poucos anos depois, surge uma explicação detalhada da
distribuição de neurônios contendo catecolamina e serotonina no cérebro de rato por
Dahlström e Fluxe (UNGERSTEDT, 1971).
NEURÔNIO PÓS-SINÁPTICO
TERMINAIS PRÉ-SINÁPTICOS
FENILALANINA TIROSINA DOPA
DOPAMINA
NORADRENALINA
FENILALANINA-HIDROXILASE TIROSINA-HIDROXILASE
DOPA-DESCARBOXILASE
DOPAMINA-BETA-HIDROXILASE
VMAT2 Ca+²
DAT
COMT

10
Embora a técnica de histofluorescência tenha trazido uma boa base para o
entendimento do sistema CA em vertebrados, é o método de imunoistoquímica que vem
sendo utilizado por ser mais sensível e específico (SMEETS; GONZÁLEZ, 2000). Para
detectar CA, podem ser utilizados os métodos que detectem as próprias substâncias
catecolaminérgicas ou métodos que detectem suas enzimas sintetizantes, as enzimas
degradativas ou os complexos de captação (SMEETS; GONZALÉZ, 2000).
O método de imunoistoquímica para TH detecta a enzima sintetizante participante na
conversão de tirosina para DOPA. Este método pode ser aplicado para determinar se
neurônios utilizam DA como neurotransmissor. Porém, TH é enzima que participa na síntese
não só de neurônios dopaminérgicos, mas também de neurônios noradrenérgicos e
adrenérgicos. Para determinar a especificidade para síntese de DA seria mais adequado testar
a presença da dopamina-beta-hidroxilase, que só estará presente se os neurônios forem
noradrenérgicos. Mesmo tendo essas especificidades em vista, o uso da imunoistoquímica
para TH para determinar se os neurônios são dopaminérgicos não é incorreto, desde que
existam trabalhos que comprovem a existência de neurônios DA na mesma área em animais
da mesma ordem (PRAKASH; WURST, 2006). Isso foi proposto, por exemplo, por Manger
(2005) quando ele falou que independente do tamanho, fenótipo ou hábito de vida, o mesmo
complemento de núcleos moduladores está presente nos diferentes representantes de
mamíferos. Os neurônios imunorreativos a TH no tegmento mesencefálico de vertebrados
quase sempre possuem DA como neurotransmissor monoaminérgico (SMEETS;
GONZÁLEZ, 2000). Inclusive estudos em répteis, com imunoistoquímica para DA e TH,
revelam grupamentos neuronais sintetizantes de DA no mesencéfalo de diferentes
representantes desse grupo taxonômico (SMEETS; JONKER; HOOGLAND, 1987; SMEETS,
1988).
1.2. Receptores dopaminérgicos
A dopamina opera mediante a ligação com seus diferentes receptores. Todos eles são
proteínas de membrana que pertencem a superfamília de receptores acoplados a proteína G
(Figura 2). São cinco os receptores específicos para DA que estão agrupados em subgrupos
(D1 e D2) (STRANGE, 1993; CALLIER et al., 2003). Fazendo parte de D1 estão D1 e D5 e
de D2 os subtipos D2, D3 e D4 (SPÜHLER; HAURI, 2013). A classificação dos receptores
dopaminérgicos foi feita tendo como base a homologia de sequências, maquinaria de
transdução de sinal e propriedades farmacológicas (SIBLEY et al., 1993). Os subtipos de

11
receptores D1 e D5, frequentemente, são acoplados com subunidades G estimulatórias,
enquanto os subtipos D2, D3 e D4, geralmente, são acoplados a subunidades G inibitórias
(SIIBLEY et al., 1993). Os receptores de dopamina são sensíveis de forma diferente à
presença de seu neurotransmissor. D3 mostra maior afinidade para a dopamina, em seguida e
em ordem decrescente vêm D5, D4, D2 e D1 (WU et al., 2005).
A expressão de receptores D1 ocorre principalmente no caudado e putâmen, núcleo
accumbens, tubérculo olfatório, córtex cerebral e amígdala. Os receptores D1 na substância
negra parecem ser expressos no estriado e transportados por axônios na via direta estriato-
nigral. Já receptores D5 são expressos no hipocampo, núcleo mamilar lateral e no núcleo
parafascicular do tálamo. O receptor D2 é expresso em grande parte no caudado e putâmen,
tubérculo olfatório e núcleo accumbens, mas também é detectado na substância negra pars
compacta e na área tegmental ventral. Eles ainda podem ser localizados em regiões não
cerebrais (retina, rins, sistema vascular e glândula pituitária). Receptores D3 estão menos
distribuídos e encontram-se nas ilhas de Calleja, alguns núcleos septais, hipotálamo e
diferentes regiões talâmicas e cerebelares, assim como, na substância negra pars compacta.
Por último, receptores D4 são expressos no córtex frontal, amígdala, bulbo olfatório,
hipocampo, hipotálamo e mesencéfalo (JACKSON; WESTLIND-DANIELSSON, 1994). Em
síntese, D1, D2 e D5 localizam-se no estriado, sistema límbico, bulbo olfatório e nos córtices
pré-frontal, pré-motor, cingulado e entorrinal, enquanto D3 e D4 estão restritos à área límbica
(CAVE; BAKER, 2009).
Figura 2. Representação de receptor de dopamina transmembana. Esquema da estrutura putativa da isoforma de
receptor D2R. A inserção de aminoácido D2L específica é indicada junto com os locais potenciais. Resíduos
aminoácidos potenciais envolvidos em N-glicosilação, fosforilação de PKA, palmitoilação e ligantes de
agonistas também são mostrados. Fonte: VALLONE; PICETTI; BORRELI, 2000.

12
1.3. Grupamentos neuronais dopaminérgicos
Grupos celulares catecolaminérgicos, normalmente, não estão arranjados de forma
isolada a estruturas anatômicas bem definidas. O padrão de distribuição dos núcleos contendo
CA pode variar entre espécies dentro de um mesmo grupo de vertebrados e mais ainda quando
se fala de diferentes representantes dentro de vertebrados. Dessa forma, definir tais grupos
com a nomenclatura A1-17 torna mais fácil a comparação entre diferentes espécies animais
(BJÖRKLUND; DUNNETT, 2007).
Dahlström e Fluxe descobriram em 1964 doze grupos de neurônios contendo
catecolamina (A1-A12) e nove de neurônios contendo serotonina (B1-B9) em cérebro de rato
(DAHLSTRÖM; FLUXE, 1964). Os grupos celulares A1-A12 foram definidos a partir do
eixo rostrocaudal, estando distribuídos a partir da medula oblonga até o hipocampo. Na
década de 70, novas técnicas facilitaram o estudo de neurônios catecolaminérgicos. Passou-se
a utilizar a técnica de imunoistoquímica para enzimas sintetizantes de CA: tirosina
hidroxilase, aminoácido aromático descarboxilase e dopamina-B-hidroxilase. Esta nova
ferramenta acabou por permitir o mapeamento do sistema catecolaminérgico em maior
detalhe e tornou mais precisa a distinção entre diferentes catecolaminas (BJÖRKLUND;
DUNNETT, 2007). Cinco grupos, compreendendo de A13-17, foram acrescentados
posteriormente. Estes núcleos descobertos por último encontram-se no diencéfalo, bulbo
olfatório e retina (HÖKFELT et al., 1984).
Os neurônios dopaminérgicos presentes no cérebro de mamíferos estão situados em
grupos celulares distintos entre o mesencéfalo e bulbo olfatório. Existem nove desses núcleos
descritos, codificados em sentido rostral a caudal da seguinte maneira: (I) No mesencéfalo os
neurônios DA estão arranjados em substância negra pars compacta (SNc/A9), área tegmental
ventral (VTA/A10) e zona retrorubral (RrF/A8); (II) em seguida é encontrado grupamentos
celulares no diencéfalo referentes aos grupos A11-15, sendo os maiores A11 (grupo de
neurônios do hipotálamo posterior) e A13 (grupos de neurônios da zona incerta no tálamo
ventral); Por último, vem uma pequena população de neurônios dopaminérgicos localizados
no telencéfalo que compreendem os interneurônios periglomerulares do bulbo olfatório (A16)
e os interneurônios amácrinos da retina (MARÍN et al., 2005; BJÖRKLUND; DUNNETT,
2007; PRAKASH; WURST, 2006) (Figura 3).

13
Figura 3. Distribuição de grupos celulares neuronais DA em cérebro de roedor em desenvolvimento (a) e adulto
(b). Os neurônios dopaminérgicos no cérebro de mamíferos estão localizados em nove grupos celulares distintos,
distribuídos a partir do mesencéfalo ao bulbo olfatório, como ilustrado esquematicamente, em uma visão sagital.
As projeções principais de grupos celulares DA estão ilustrados por flechas. Abreviações: Ige, eminência
ganglionar lateral; mge, eminência ganglionar medial; p1-p3, prosômeros 1-3. Fonte: BJÖRKLUND;
DUNNETT, 2007.
1.4. Grupamentos neuronais dopaminérgicos mesencefálicos
A variedade do sistema dopaminérgico mesencefálico surge a partir do processo
ontogenético em que um conjunto comum de precursores passa pela especificação, migração e
diferenciação e gera os padrões de conectividade conhecidos como resultado dos caminhos
axonais na fase de desenvolvimento (VAN DEN HEUVEL; PASTERKAMP, 2008). Cascatas
de eventos levam ao desenvolvimento dos neurônios dopaminérgicos. Eventos
desenvolvimentais iniciais formam um organizador no limite mesencefálico/metencefálico.
Este organizador, também chamado Iso, é responsável pela proliferação de precursores de
neurônios dopaminérgicos em desenvolvimento e fornecedor de pistas iniciais. Ele atua junto
com a linha média, que fornece informação adicional. Dentre os vários fatores, pode-se citar
FGF8 (fator de crescimento de fibroblastos 8) liberado a partir do Iso e sonic hedgehog (shh)
liberado a partir da linha média. Estes fatores são necessários para especificação inicial de
neurônios dopaminérgicos em desenvolvimento. Ao amadurecerem, eles dão origem a
neurônios que estarão envolvidos em uma grande escala de funções cerebrais (WURST;
BALLY-CUIF, 2001; LIU; JOYNER, 2001; HIDALGO-SANCHEZ; ALVARADO-
MALLART, 2002; KIM, et al., 2003).
A conectividade estrutural entre diferentes áreas cerebrais permite que os sinais por
estas desencadeados sejam integrados, o que permite o desenvolvimento e execução de planos
de ação apropriados a diferentes situações. Através desta integração, neurônios
dopaminérgicos mesencefálicos estão envolvidos em uma série de questões comportamentais,

14
cognitivas, motivacionais, de recompensa, de controle motor e também de algumas desordens
neurológicas e/ou psiquiátricas (BJÖRKUND; DUNNETT, 2007; YETNIKOFF et al., 2014;
HABER, 2014). O estriado se comunica com o córtex pré-frontal, ínsula e mesencéfalo e,
acredita-se, que a integração dos sinais destas áreas seja crítico para respostas adaptativas do
organismo (SALAMONE; CORREA, 2002; KOHNO et al., 2017). Projeções que saem do
mesencéfalo ao estriado sinalizam a existência de eventos motivantes e respostas
antecipatórias moduladas por dopamina estão envolvidas nas preferências que envolvem risco
(SUGAM et al., 2012). Componente importante na tomada de decisão dirigida a objetivos é a
capacidade de antes avaliar bem os aspectos diferentes de recompensa, tais como, valor versus
risco e previsibilidade e ainda inibir comportamentos não adaptativos com base em
experiências não bem-sucedidas. Vias neuronais dopaminérgicas que integra regiões do
mesencéfalo com outras áreas cerebrais estão relacionadas a estas funções. São elas as vias
mesolímbica, mesocortical e mesoestriatal (WISE, 2004; HABER, 2014).
Existem três circuitarias formadas a partir de núcleos que estão no mesencéfalo:
mesoestriatal, mesocortical e mesolímbica. A via mesoestriatal se origina na SNc (A9) e
projeta principalmente para os domínios dorsolaterais do estriado e está envolvida no
movimento voluntário e postura corporal (PRAKASH; WURST, 2006; DELONG;
WICHMANN, 2007; MONEY; STANWOOD, 2013). A via mesocortical, que inerva
diferentes regiões do córtex frontal, refere-se às projeções dopaminérgicas a partir do
mesencéfalo ventral, inclusive a VTA e SN, para o córtex pré-frontal e está relacionada ao
aprendizado e memória (YETNIKOFF et al., 2014). Por último, o sistema mesolímbico é
formado por neurônios dopaminérgicos localizados na VTA e suas projeções para o Núcleo
Accubens (NAc), córtex, amígdala e o hipocampo, que participam da configuração de
comportamentos motivacionais (BJÖRKUND; DUNNETT, 2007; NIEH et al., 2013). A RRF
também se comunica com esses três circuitos através de suas projeções eferentes
(BENTIVOGLIO; MORELLI, 2005). Embora estes circuitos se caracterizem por
apresentarem localizações anatômicas e padrões de atividade distintos, suas células neuronais
estão interconectadas (BENTIVOGLIO; MORELLI, 2005). Dessa forma, as projeções a partir
da SN não estão exclusivamente destinadas ao estriado, assim como, projeções a partir da
VTA não se restringem apenas as vias mesoestriatal e mesolímbica (BJÖRKUND;
DUNNETT, 2007).

15
Degeneração destas vias leva a desordens neurológicas, como por exemplo, a doença
de Parkinson que dificulta a iniciação de movimentos e a postura corporal. Outros exemplos
são esquizofrenia, doença de Huntington, vícios a drogas (desencadeados por drogas de
abuso, como por exemplo, os psicoestimulantes e opióides), distúrbios de humor e transtorno
do déficit de atenção com hiperatividade (ADHD) (LAU et al., 2013; BARROT, 2014; WISE;
KOOB, 2014; BORGONOVO et al., 2016).
Existem estudos disponíveis que investigaram sobre a presença de células
dopaminérgicas mesencefálicas em diferentes classes de vertebrados. Análise comparativa
dos grupos celulares CA no mesencéfalo de vertebrados revela que em peixes ciclóstomos não
há a presença de nenhum desses grupamentos, enquanto osteíctes pode apresentar o
grupamento A10 e condrictes pode exibir tanto o grupamento A9 quanto o A10. Em Anfíbios,
o grupamento A9 e A10 também podem ser observados. Por sua vez, répteis, aves e
mamíferos podem apresentar uma divisão em três grupamentos, podendo ainda ser
encontrados grupos adicionais (SMEETS; GONZÁLEZ, 2000).
Em peixes ciclóstomos e actinopterígeos, estudos imunoistoquímicos não revelaram
grupamentos de células dopaminérgicas no mesencéfalo (RINK; WULLIMANN, 2001;
BARREIRO-IGLESIAS et al., 2008). Embora os actinopterígeos não tenham apresentado
células imunorreativas a DA no mesencéfalo através dos estudos realizados até agora, outra
classe representante de osteíctes, os sarcopterígeos, apresenta espécies em que se pode
identificar grupo de corpos celulares imunorreativos a TH (LÓPEZ; GONZÁLEZ, 2017). Por
exemplo, o mesencéfalo de peixes dipnoicos, uma subclasse de sarcopterígeos, dispõe de
células dopaminérgicas comparáveis a área tegmental ventral/substância negra pars compacta
encontradas em amniotas (SMEETS; GONZÁLEZ, 2000). Tais células em peixes dipnoicos,
não estão restritas apenas na região mesencefálica, ao invés disso, podem se estender até
regiões diencefálicas ventrais (VERNEY et al., 2001; LOPEZ; GONZÁLEZ, 2017). Em
peixes cartilaginosos (condrictes) já foi descrito em todos os representantes de
elasmobrânquios (tubarão, mantas e raias) estudados um grupamento DA mesencéfalico em
torno da sua extensão rostrocaudal, que também podem se estender para o tegmento
diencefálico (MEREDITH; SMEETS, 1987; STUESSE et al., 1994).
Anfíbios, assim como peixes elasmobrânquios, apresentam uma extensão de corpos
celulares mesencefálicos imunorreativos a TH/DA ao longo de sua extensão rostrocaudal.
Estas células estão presentes a partir do limite diencéfalo-mesencéfalo até a saída do nervo

16
oculomotor e estão restritas a linha média, não sendo possível fazer uma subdivisão desse
grupamento. Na rã Rana ridibunda, um único grupo de células imunorreativas a TH/DA são
observadas ao longo da linha média se estendendo até a saída do nervo oculomotor. São
células que parecem pertencer a um grupo dorsomedial proveniente do tubérculo posterior, e
se assemelha a área tegmental ventral (GONZÁLEZ; SMEETS, 1991;). Esse mesmo padrão é
observado no sapo Physalaemus pustulosus (O’CONNELL et al., 2010)
Em aves, é possível fazer uma divisão das células CA em três grupamentos tendo
como base a sua posição (A8, A9 e A10), além de ser possível encontrar grupos adicionais.
Por exemplo, foi observado células CA no cinza mesencefálico periaquedutal. Essas células
foram consideradas a parte mais caudal do grupo celular A11, que se estende à níveis mais
caudais do mesencéfalo. Os grupos celulares A8-A10 representa um campo grande e contínuo
de neurônios dopaminérgicos no mesencéfalo de aves. Normalmente, A10 é o grupamento
mais rostral e medial, enquanto A9 é mais intermediário e lateral e A8 é o mais caudal
(REINER et al., 1994; MOONS et al., 1994).
Células dopaminérgicas no mesencéfalo de mamíferos inicialmente foram
classificadas em três grupos (A8, A9 e A10). Atualmente, é possível subdividir A9 e A10. O
grupo A10 pode ser subdividido em um grupo caudal A10c, que consiste de células no núcleo
de Edinger Westphal, núcleo linear central e rostral e o núcleo interfascicular. A área
tegmental ventral é subdividida em núcleo pigmentado paranigral e núcleo pigmentado
parabraquial. O grupo supramamilar constitui o subgrupo A10dr (dorsal, ventral). O grupo
A10dc (dorsal, caudal) pode ser encontrado na rafe, enquanto o grupo A10dr (dorsal, rostral)
é encontrado dentro ou na proximidade do complexo habenular de ratos (SMEETS;
GONZÁLEZ, 2000). A SN pode ser dividida em camada dorsal e camada ventral. A diferença
entre as camadas da SN é que as células da camada dorsal têm dendritos orientados,
principalmente, em direção mediolateral, são imunorreativas a TH e proteína ligante de cálcio,
Calbindina-D28k e projetam, em grande parte, para a matriz do estriado. As células da camada
ventral, por sua vez, têm uma organização dentrítica de sentido, predominantemente,
dorsoventral, não são imunorreativas a calbindina-D28k e inerva, principalmente, trechos
estriatais (SMEETS; GONZÁLEZ, 2000). Esta nomenclatura e classificação podem ser
adaptadas para outros grupos dentro de mamíferos (MASEKO et al., 2007; DELL et al., 2010;
MEDEIROS et al., 2016; CAVALCANTI et al., 2016).
1.5. Grupamentos neuronais dopaminérgicos mesencefálicos em répteis

17
Os primeiros estudos dos grupamentos neuronais em répteis utilizaram a técnica de
fluorescência induzida por formaldeído (técnica FIF – do inglês formaldehyde-induced
fluorescence) com a finalidade de identificar o sistema monoaminérgico (PARENT;
POIRIER, 1971 – Figura 4). A técnica FIF permite discriminar entre serotonina e
catecolamina, mas não entre os diferentes neurotransmissores catecolaminérgicos (dopamina,
noradrenalina e adrenalina), além de ser pouco sensível, prejudicando assim, a interpretação
dos resultados em áreas em que o sistema dopaminérgico se sobrepõe com o sistema
serotoninérgico (SMEETS; JONKER; HOOGLAND, 1987). Outras formas de visualizar o
sistema monoaminérgico, descobertas sem seguida, foi através da técnica de
imunoistoquímica usando anticorpos contra as enzimas sintetizantes de neurotransmissores e
contra os próprios neurotransmissores (HALÁSZ et al., 1982; GEFFARD, 1984).
Figura 4. Secção transversa através da metade rostral do mesencéfalo, evidenciando numerosas células
fluoresecentes verdes (catecolaminérgicas) dentro do tegmento mesencefálico da tartaruga pintada (Chrysemys
picta) através de técnica de histofluorescência. Aumento em 100 vezes. Fonte: PARENT; POIRIER, 1971.
Assim como em aves e mamíferos, uma divisão de núcleos dopaminérgicos
mesencefálicos, tendo como base a posição, pôde ser feita em três subdivisões. Sendo ainda
possível identificar células dopaminérgicas fora do complexo A8-A10 (SMEETS;
GONZÁLEZ, 2000). Neurônios DA/CA mesencefálicos foram caracterizados em lagartos,
jacaré, serpente e tartaruga.
Através da técnica de imunofluorescência indireta foi possível observar grupos
celulares contendo CA no tronco encefálico do lagarto Varanus exanthematicus. No
mesencéfalo deste lagarto os grupos celulares dopaminérgicos foram divididos em (I)
substância negra, (II) um grupo celular medial a substância negra que se estende rostralmente
e se compara a área tegmental ventral e (III) uma extensão caudodorsal a substância negra.

18
Tais grupos celulares são comparáveis aos grupos A9, A10 e A8 de mamíferos,
orespectivamente (WOLTERS et al., 1984 – Figura 5). Além disso, outras células podem
aparecer fora da divisão tripla em A8, A9 e A10. No lagarto V. exanthematicus e A.
carolinensis, células imunorreativas a TH/DA ocorrem na zona periventricular do
mesencéfalo e na tartaruga P. scripta elegans, algumas células dispersas foram encontradas
no núcleo laminar do torus semicircular (WOLTERS et al., 1984; SMEETS, 1994).
Figura 5. Mesencéfalo caudal. Corpos celulares da substância negra (grupo A9) e área tegmental ventral (grupo
A10) imunorreativos a tirosina hidroxilase em um lagarto não tratado com colchicina. Um amplo feixe de fibras
fluorescentes finas em seção transversal é visível dorsomedial a substância negra. Transversalmente fibras
cortadas mais espessas podem ser vistas ventrolateral e ventromedial ao grupo celular CA da área tegmental
ventral (Barra: 150 μm). Neurônios, provavelmente dopaminérgicos, de tamanho médio na substância negra.
Neurônios, provavelmente dopaminérgicos, de tamanho pequeno e médio contendo TH na área tegmental
ventral. Fonte: WOLTERS et al., 1984.
Algumas células imunofluorescentes catecolaminérgicas de pequeno e médio porte
foram encontradas dispersas no mesencéfalo de V. exanthematicus. Na área ventral e medial
aos núcleos vermelhos (denominada VTA), uma quantidade considerável de células positivas
a CA é encontrada entre fibras do nervo oculomotor. A SN apresenta numerosas células
catecolaminérgicas arredondadas ou ovais de tamanho médio. Células imunorreativas a TH e
DA também estavam presentes na zona periventricular do mesencéfalo (WOLTERS et al.,
1984; SMEETS, 1994).
A distribuição de dopamina imunorreativa no mesencéfalo do lagarto Gekko gecko foi
estudada usando anticorpos contra dopamina. As células dopaminérgicas foram encontradas
na VTA, SN e presumido equivalente reptiliano do grupo A8 em mamíferos (RA8)
(SMEETS; HOOGLAND; VOORN, 1986 – Figura 6).
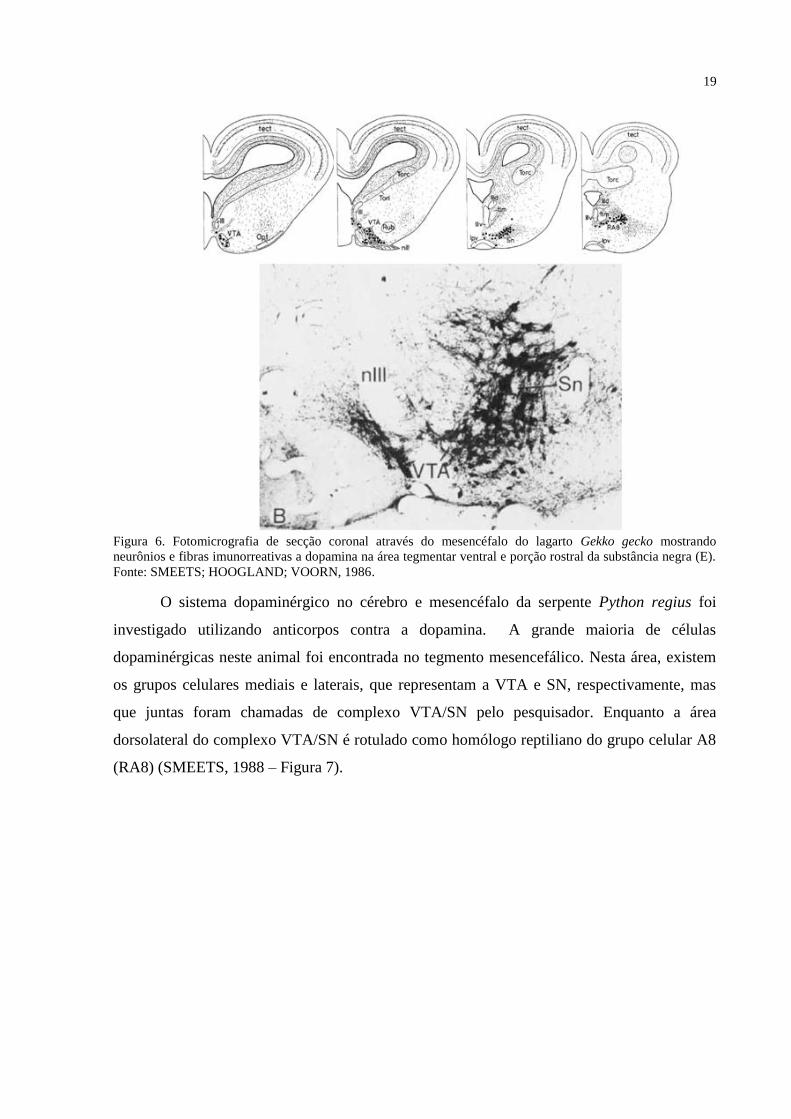
19
Figura 6. Fotomicrografia de secção coronal através do mesencéfalo do lagarto Gekko gecko mostrando
neurônios e fibras imunorreativas a dopamina na área tegmentar ventral e porção rostral da substância negra (E).
Fonte: SMEETS; HOOGLAND; VOORN, 1986.
O sistema dopaminérgico no cérebro e mesencéfalo da serpente Python regius foi
investigado utilizando anticorpos contra a dopamina. A grande maioria de células
dopaminérgicas neste animal foi encontrada no tegmento mesencefálico. Nesta área, existem
os grupos celulares mediais e laterais, que representam a VTA e SN, respectivamente, mas
que juntas foram chamadas de complexo VTA/SN pelo pesquisador. Enquanto a área
dorsolateral do complexo VTA/SN é rotulado como homólogo reptiliano do grupo celular A8
(RA8) (SMEETS, 1988 – Figura 7).
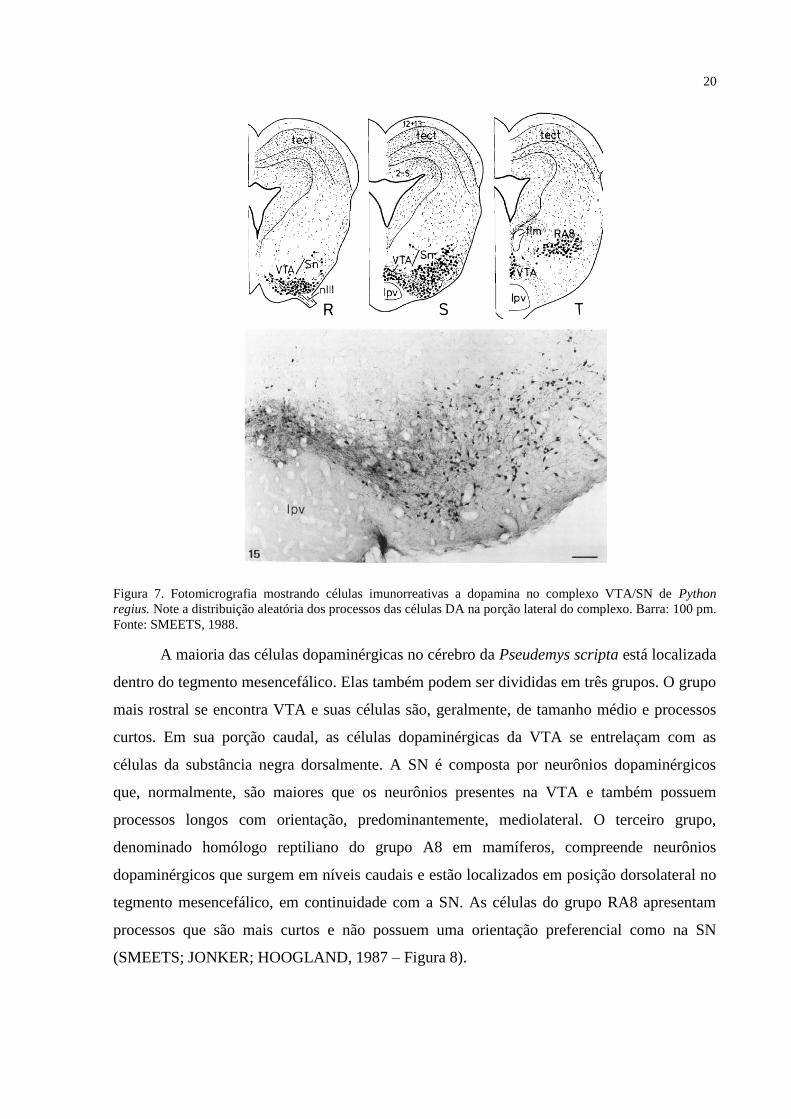
20
Figura 7. Fotomicrografia mostrando células imunorreativas a dopamina no complexo VTA/SN de Python
regius. Note a distribuição aleatória dos processos das células DA na porção lateral do complexo. Barra: 100 pm.
Fonte: SMEETS, 1988.
A maioria das células dopaminérgicas no cérebro da Pseudemys scripta está localizada
dentro do tegmento mesencefálico. Elas também podem ser divididas em três grupos. O grupo
mais rostral se encontra VTA e suas células são, geralmente, de tamanho médio e processos
curtos. Em sua porção caudal, as células dopaminérgicas da VTA se entrelaçam com as
células da substância negra dorsalmente. A SN é composta por neurônios dopaminérgicos
que, normalmente, são maiores que os neurônios presentes na VTA e também possuem
processos longos com orientação, predominantemente, mediolateral. O terceiro grupo,
denominado homólogo reptiliano do grupo A8 em mamíferos, compreende neurônios
dopaminérgicos que surgem em níveis caudais e estão localizados em posição dorsolateral no
tegmento mesencefálico, em continuidade com a SN. As células do grupo RA8 apresentam
processos que são mais curtos e não possuem uma orientação preferencial como na SN
(SMEETS; JONKER; HOOGLAND, 1987 – Figura 8).

21
Figura 8. Desenhos de uma série de secções coronais, em sentido rostrocaudal, através do mesencéfalo da
tartaruga Pseudemys scripta mostrando a distribuição de células imunorreativas à dopamina (pontos grandes) e
fibras (pontos curtos e linhas contínuas) (A) e fotomicrografia mostrando células imunorreativas a dopamina na
substância negra, evidenciando os processos longos e com orientação mediolateral (B). Barra: 0,01 mm. Fonte:
SMEETS; JONKER; HOOGLAND, 1987b.
Em Caiman crocodilus, um grande grupo de células positivas a CA foi observado no
tegmento pedunculopontino (TP) e em algumas células mediais a ele que podem ser
comparadas a VTA (BRAUTH; KITT, 1980; REINER; KARTEN, 1980 – Figura 9).
Em duas espécies de quelônio do gênero Chrysemys foi revelado que células
catecolaminérgicas estavam presentes no tegmento mesencefálico medial e lateral, similar a
VTA e SN, respectivamente (PARENT; POIRIER, 1971; BRAUTH et al., 1983). Em outra
espécie tartaruga (Pseudemys scripta elegans) algumas células esparsamente distribuídas
foram observadas no núcleo laminar do torus semicircular (TSC) (SMEETS, 1994).
B
A

22
Figura 9. Células e fibras imunorreativas a TH no mesencéfalo do jacaretinga (Caiman crocodilus) em nível de
substância negra (B, C) e área ventral de Tsai (A, D). (B). Barra: 0,01 mm. Fonte: BRAUTH, 1988.
A maioria das células catecolaminérgicas no tronco encefálico de tartaruga está
concentrada na porção rostral do tegmento mesencefálico. Observando no nível do núcleo
oculomotor, estas células formam uma grande coleção que se estendem da região
ventromedial a dorsolateral do tegmento. A distribuição destas células nervosas no cérebro de
tartaruga pintada (Chrysemys picta) foi investigada utilizando a técnica de histofluorescência
de Falck e Hillarp. Foi observado que a organização destas células é muito similar ao
observado no cérebro de rato (PARENT; POIRIER, 1971). É concebível que estas células
correspondam a SN observada em mamíferos e que formem a via “nigroestriatal” a partir de
achados que mostram numerosas terminações catecolaminérgicas encontradas no nível do
estriado (PARENT; OLIVIER, 1970; PARENT; POIRIER, 1971; REINER; KARTEN,
1980).
Nos estudos feitos com tetrápodes, a inervação dopaminérgica dos gânglios da base é
dividido em sistema mesoestriatal dorsal e ventral, sendo a SNc e a VTA as principais fontes
de entrada DA em regiões estriatais (SMEETS; GONZÁLEZ, 2000). Os répteis apresentam
uma forte segregação a partir destes dois grupamentos neuronais, a SNc inerva
substancialmente o estriado e VTA envia projeções para o núcleo accumbens (SMEETS,
1994; PEREZ-SANTANA; SMEETS, 1997). A dopamina age no estriado reptiliano a partir
dos receptores D1 e D2, assim como ocorre em mamíferos (BARRA et al., 2010).

23
Em répteis, os neurônios da SNr estão localizados lateral ou dorsolateral aos neurônios
dopaminérgicos da SNc. Assim como em mamíferos, os neurônios da SNr projetam para o
tectum mesencefálico e estes para os centros pré-motores e motores no tronco encefálico e
medula espinal (MEDINA; REINER, 1994). A rota do gânglio basal para o tectum
mesencéfalico através da SNr provavelmente está envolvida em promover movimento em
répteis (REINER, 2017 – Figura 10).
Figura 10. Fotomicrografia do mesencéfalo de tartaruga mostrando um subdivisão da substância negra (pars
compacta e pars reticulata) a partir da imunoistoquímica para TH. Fonte: REINER, 2017.
Desordens em vias dopaminérgicas, assim como mencionado anteriormente, trazem
alterações neurológicas. Em vertebrados, os efeitos dessas desordens são semelhante à doença
de Parkinson, com anormalidades na postura, rigidez dos músculos, tremor e bradicinesia.
(TEMPLE et al., 2002). Estas alterações podem ser induzidas por substâncias neurotóxicas. A
fim de se conhecer as mudanças neurológicas em répteis, usou-se a substância sintética
neurotóxica, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) que após ser convertida
em seu metabólito 1-methyl-4-phenylpyridinium (MPP+) é capaz de induzir uma série de
desordens de movimento através da destruição de neurônios dopaminérgicos no trato
nigroestriatal e consequentemente gerar uma redução de DA no estriado. Foi observado no
lagarto Anolis, que a MPTP causou danos aos terminais do estriado no cérebro do animal e
induziu uma variedade de sintomas parkinsonianos (hipocinesia, rigidez e movimentos da
cabeça e pescoço estereotipados) (GREENBERG et al., 1989). Da mesma forma, depleção de
DA causada por MPTP demonstrou os mesmo efeitos comportamentais relatados a outros
vertebrados em duas espécies de serpentes do gênero Elaphe (TEMPLE et al., 2002).

24
1.6. Tartarugas marinhas
Os répteis existentes compreendem os sphenodons (tuatara), lagartos, serpentes,
tartarugas e crocodilianos. Ao todo, são cerca de mais de 10.000 espécies. Assim como os
mamíferos, os répteis também expressam um número relacionado de comportamentos
complexos. Eles são capazes, por exemplo, de aprender a navegar labirintos (NAUMANN et
al., 2015).
Tartarugas marinhas possuem distribuição cosmopolita, realizando grandes migrações
entre áreas de alimentação e reprodução (MEYLAN; MEYLAN, 1999; LOHMANN et al.,
2008). Tartaruga marinha encontra-se dentro da classe Reptilia, ordem Testudines e família
Cheloniidae e estão agrupadas em duas famílias: Dermochelyidae e Cheloniidae. No mundo
são conhecidas apenas sete espécies, dentre essas, há registros de ocorrência de cinco no
litoral do Brasil: Dermochelys coriacea (tartaruga de couro), Chelonia mydas (tartaruga-
verde), Caretta caretta (tartaruga-cabeçuda), Eretmochelys imbricata (tartaruga-de-pente) e
Lepidochelys olivacea (tartaruga Oliva) (MARCOVALDI; MARCOVALDI, 1999).
De acordo com a Lista Vermelha da União Internacional para a Conservação da
Natureza (IUCN, 2017), todas as espécies de tartarugas marinhas existentes no Brasil, se
encontram ameaçadas de extinção, sendo três (D. coriacea, L. olivacea e C. caretta)
classificadas como vulneráveis, uma como ameaçada (C. mydas) e uma (E. imbricata) como
criticamente ameaçada.
São animais capazes de voltar ao mesmo local onde nasceram para nidificar, mesmo
percorrendo por longas distâncias. Para fazer isso, estudos sugerem que elas utilizem um
mecanismo de navegação baseado no campo magnético da crosta terrestre e impriting
geomagnético, mas que também usam pistas químicas e visuais (SILVA, 2013; BROTHERS;
LOHMANN, 2015). Ou que ainda utilizem pistas magnéticas e químicas em conjunto e de
forma sequenciada como estratégia para localizar pontos específicos (ENDRES et al., 2016).
Em tartarugas marinhas, o encéfalo está organizado de forma longitudinal ao longo da
linha média do crânio e se encontra alojado dentro da caixa craniana. O encéfalo desses
animais pode ser descrito em três regiões de acordo com o seu desenvolvimento: cérebro
anterior, cérebro médio e cérebro posterior. O cérebro anterior se estende desde o nariz até o
cérebro médio e se refere ao telencéfalo e diencéfalo; o cérebro médio se estende desde o olho
até a porção posterior dos lóbulos ópticos e se refere ao mesencéfalo; e o cérebro posterior se
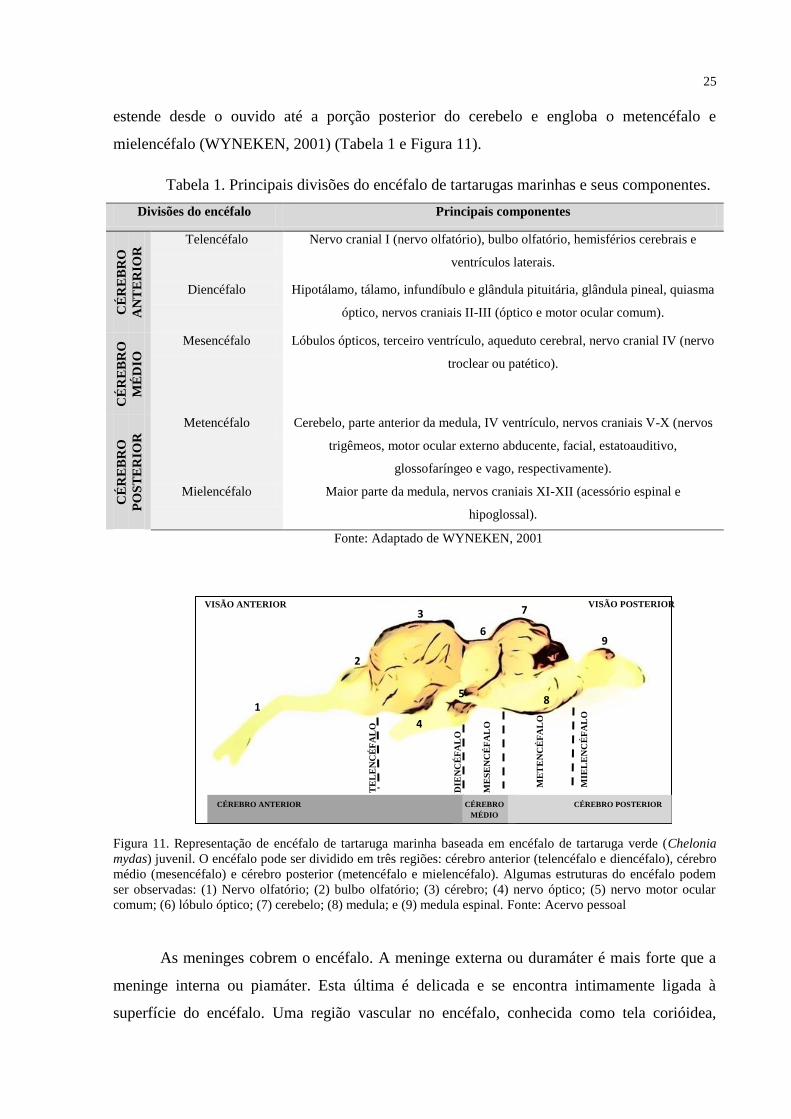
25
estende desde o ouvido até a porção posterior do cerebelo e engloba o metencéfalo e
mielencéfalo (WYNEKEN, 2001) (Tabela 1 e Figura 11).
Tabela 1. Principais divisões do encéfalo de tartarugas marinhas e seus componentes.
Divisões do encéfalo Principais componentes
CÉ
RE
BR
O
AN
TE
RIO
R Telencéfalo Nervo cranial I (nervo olfatório), bulbo olfatório, hemisférios cerebrais e
ventrículos laterais.
Diencéfalo Hipotálamo, tálamo, infundíbulo e glândula pituitária, glândula pineal, quiasma
óptico, nervos craniais II-III (óptico e motor ocular comum).
CÉ
RE
BR
O
MÉ
DIO
Mesencéfalo Lóbulos ópticos, terceiro ventrículo, aqueduto cerebral, nervo cranial IV (nervo
troclear ou patético).
CÉ
RE
BR
O
PO
ST
ER
IOR
Metencéfalo Cerebelo, parte anterior da medula, IV ventrículo, nervos craniais V-X (nervos
trigêmeos, motor ocular externo abducente, facial, estatoauditivo,
glossofaríngeo e vago, respectivamente).
Mielencéfalo Maior parte da medula, nervos craniais XI-XII (acessório espinal e
hipoglossal).
Fonte: Adaptado de WYNEKEN, 2001
Figura 11. Representação de encéfalo de tartaruga marinha baseada em encéfalo de tartaruga verde (Chelonia
mydas) juvenil. O encéfalo pode ser dividido em três regiões: cérebro anterior (telencéfalo e diencéfalo), cérebro
médio (mesencéfalo) e cérebro posterior (metencéfalo e mielencéfalo). Algumas estruturas do encéfalo podem
ser observadas: (1) Nervo olfatório; (2) bulbo olfatório; (3) cérebro; (4) nervo óptico; (5) nervo motor ocular
comum; (6) lóbulo óptico; (7) cerebelo; (8) medula; e (9) medula espinal. Fonte: Acervo pessoal
As meninges cobrem o encéfalo. A meninge externa ou duramáter é mais forte que a
meninge interna ou piamáter. Esta última é delicada e se encontra intimamente ligada à
superfície do encéfalo. Uma região vascular no encéfalo, conhecida como tela corióidea,
TE
LE
NC
ÉF
AL
O
DIE
NC
ÉF
AL
O
CÉREBRO
MÉDIO
1
2
4
3
4
5
6
7
2
ME
SE
NC
ÉF
AL
O
ME
TE
NC
ÉF
AL
O
MIE
LE
NC
ÉF
AL
O
VISÃO POSTERIOR VISÃO ANTERIOR
9
CÉREBRO ANTERIOR CÉREBRO POSTERIOR
8

26
produz o líquido incolor cefalorraquídeo. Este líquido circula através dos ventrículos e
alcança os espaços epidurais (acima da dura-máter) e subdurais (abaixo da dura-máter) para
banhar o encéfalo (WYNEKEN, 2001).
Inicialmente, o encéfalo tem forma de tubo, mas logo sofre várias modificações ao
longo de sua extensão para dar origem as estruturas observadas em tartarugas adultas. Os
ventrículos laterais, terceiro e quarto ventrículo e aqueduto cerebral, por sua vez, são todos
vestígios da cavidade neural que permanecem nos adultos. Durante a ontogenia, o encéfalo é
proporcionalmente maior em recém-nascidos e juvenis que em subadultos e adultos
(WYNEKEN, 2001).
Embora existam trabalhos que descrevam a anatomia do SNC de tartarugas marinhas,
os que descrevem a citoarquitetura do seu encéfalo são inexistentes. Nesse sentido, o presente
trabalho utiliza animais provenientes de encalhes e que vieram à óbito para vencer essa
dificuldade. As primeiras descrições da citoarquitetura do encéfalo de tartarugas marinhas são
feitas na região mesencefálica e investigando os grupamentos neuronais dopaminérgicos,
assim como, da orientação de suas fibras. A investigação dos grupamentos neuronais
dopaminérgicos do mesencéfalo em tartaruga marinha é importante por fornecer informação
do sistema dopaminérgico em um grupo reptiliano, aumentando, assim, o entendimento
acerca do sistema nervoso em répteis. Além de providenciar informação útil no entendimento
das similaridades das estruturas nervosas entre os diferentes grupos de vertebrados.

27
2. OBJETIVOS
2.1. Objetivo geral
Caracterizar morfologicamente os núcleos mesencefálicos imunorreativos a
Tirosina Hidroxilase de tartaruga marinha.
2.2. Objetivos específicos
Identificar os núcleos neuronais imunorreativos a Tirosina Hidroxilase no
mesencéfalo de tartarugas marinhas;
Descrever a morfologia dos neurônios imunorreativos a Tirosina Hidroxilase
presentes no mesencéfalo de tartarugas marinhas;
Delimitar a citoarquitetura dos núcleos mesencefálicos imunorreativos a
Tirosina Hidroxilase a partir do método de Nissl.

28
3. MATERIAIS E MÉTODOS
3.1. Área de coleta
O trabalho desenvolvido integrou um projeto maior intitulado “Monitoramento de
Praias do Litoral Potiguar/Cearense e Embarcado da Biota Marinha, em especial sobre
Mamíferos Marinhos e Quelônios”, resultante de uma condicionante ambiental exigida pelo
IBAMA à PETROBRÁS decorrente de indicação do Parecer Técnico
CGPEG/DILIC/IBAMA através do contrato de número 2500.005657510.2.
A pesquisa foi realizada em praias do litoral Noroeste do Rio Grande do Norte e
Ceará, entre os municípios de Caiçara do Norte - RN (5° 4'1.15"S; 36° 4'36.41"O) e
Icapuí/CE, (4°38'48.28"S; 37 °32'52.08"O), perfazendo uma extensão aproximada de 344 km
(Figura 12).
Figura 12. Localização da área de estudo, Bacia Potiguar, RN, Brasil.
3.2. Modelo experimental
Foram utilizados neste trabalho quatro encéfalos de tartarugas marinhas pertencentes a
espécie Chelonia mydas (Linnaeus, 1758) (Figura 13). Cada indivíduo recebeu um código de
identificação (TA1, TA2, TA3 e TA4). Dois eram machos e duas eram fêmeas. De todos,
apenas TA1 era adulto e os demais eram animais juvenis.

29
C. mydas é uma espécie ameaçada de extinção de acordo com a lista da IUCN (2018).
É importante salientar que nenhum animal sofreu eutanásia neste estudo, foram utilizados
apenas os animais que vieram a óbito.
Figura 13. Exemplar juvenil da espécie Chelonia mydas. Fonte: WWF, 2018
De sete espécies de tartarugas marinhas existentes em todo o mundo, C. mydas é
considerada a segunda maior e a única que vai frequentemente à costa para descansar em
algumas regiões. Os adultos são herbívoros, alimentando-se de ervas marinhas, algas e outros
vegetais marinhos com a ajuda de seu bico afiado e serrilhado. Tartaruga-verde é encontrada
em regiões tropicais e subtropicais de todo o mundo, como observado na imagem abaixo
(Figura 14), onde o destaque laranja representa a distribuição desses animais (SEATURTLES,
2017).
Em relação a sua biologia, indivíduos da espécie C. mydas atingem a maturidade
sexual entre 20 a 40 anos. Quando adultos, medem entre 80 a 120 cm de comprimento,
atingindo até 300 kg. Quando filhotes medem entre 3 a 4 cm e pesam de 25 a 30 g. A
reprodução ocorre a cada dois ou quatro anos, com duas a cinco desovas por temporada, com
uma média de 80-120 ovos que duram, aproximadamente 60 dias para eclodirem (MTSG,
2017).

30
Figura 14. Distribuição de tartaruga-verde (Chelonia mydas) ao longo de regiões tropicais e subtropicais de todo
o mundo. Fonte: IUCN, 2017.
3.3. Coleta de dados
Os animais utilizados no estudo foram provenientes de encalhes de tartarugas
marinhas nas praias da região da Bacia Potiguar. Ao registrar o animal vivo, o biólogo
acionava a equipe de resgate que realizava a translocação para a Base de Reabilitação de
Animais Marinhos localizada na Praia de Upanema, Areia Branca (Figura16). Os animais
recebem tratamento adequado ao seu quadro clínico pela equipe médica veterinária do projeto
Cetáceos da Costa Branca com vistas a sua recuperação e posterior soltura (Figura 15).

31
Figura 15. (A) Biólogo atendendo a encalhe de tartaruga marinha registrada durante monitoramento junto com a
equipe de resgate, responsável pela translocação do animal até a Base de Reabilitação. (B) Recinto de tartarugas
marinhas da Base de Reabilitação de Animais Marinhos- PCCB-UERN, Praia de Upanema, Areia Branca. (C)
Tartaruga marinha recebendo atendimento da equipe médica veterinária do PCCB-UERN. (D) Animal com
melhora significativa em seu quadro clínico recebendo alimentação. (E e F) Soltura de tartarugas marinhas aptas
para a reintrodução ao seu habitat. Fonte: Projeto Cetáceos da Costa Branca – PCCB/UERN, 2018.
Quando porventura, alguns desses animais não resistiam e vinham a óbito, eram
imediatamente encaminhados para necropsia no Laboratório de Monitoramento de Biota
Marinha – UERN – Mossoró, dando prioridade a retirada do encéfalo no caso de animais
utilizados neste estudo (Figura 16).
Os técnicos que atuam no projeto possuem a licença ABIO (AUTORIZAÇÃO DE
CAPTURA, COLETA E TRANSPORTE DE MATERIAL BIOLÓGICO) e Cadastro no
SISBIO (Anexo 1).
A
F E
B
C D

32
Figura 16. Unidade de Necropsia instalada na Base de Reabilitação de Animais Marinhos- PCCB-UERN,
Mossoró- RN.
3.3.1. Remoção de Encéfalo, Fixação e Armazenamento
Para fixação dos encéfalos foram utilizados dois métodos. O primeiro foi por meio da
perfusão através do ventrículo, com a passagem de formol 10% precedido pela passagem de
solução salina. Esse método foi utilizado para um animal que havia morrido pouco antes e
estava em perfeita condições para passar pelo procedimento (TA1). Para os demais animais, o
encéfalo foi retirado com um tempo maior após o animal ter morrido (TA2 e TA3). Para estes,
os encéfalos foram retirados sem passar pela perfusão e foram fixados por imersão em formol
4%. Após a fixação inicial, os encéfalos passaram pela pós-fixação e, em seguida, foram
mantidos em sacarose 30%. O animal TA4 não passou pelo procedimento de fixação, pois foi
utilizado com o objetivo único de descrever a morfologia externa do encéfalo.
3.3.1.1. Perfusão Transcardíaca
Em tartarugas marinhas, o coração está localizado dentro do pericárdio. Seu limite
dorsal é com os pulmões enquanto o limite lateral é com os lobos do fígado. Existem quatro
compartimentos ou câmaras que subdividem o coração em tartarugas: seio venoso, dois
grandes átrios e um ventrículo. Da parte anterior e ventral do coração emergem os grandes
vasos. Um desses vasos é a aorta direita, que supre sangue para a cabeça, membros e parte
inferior do corpo, através do tronco braquiocefálico e posteriormente das artérias carótidas. O
acesso ao coração do animal foi feita com a retirada do plastrão. Os processos acrômios

33
serviram como ponto de referência para a localização exata do coração. Foi feito um corte
longitudinal entre os processos acrômios e rebatida a musculatura peitoral lateralmente. A
partir do ventrículo e direcionado a aorta direita foi introduzida uma agulha (20mm x
0,55mm). O passo seguinte foi a passagem no sistema circulatório de solução salina a 0,9%
em tampão fosfato 0,1M, pH 7,4 (2 litros) como forma de limpar o sistema e potencializar o
efeito do fixador. Em seguida foram passados mais dois litros de solução fixadora composta
por formol 4%. A saída do fluxo vascular foi feito por corte no átrio direito.
Posteriormente, foi realizada a ruptura do crânio do animal com o auxílio de uma mini
serra elétrica, sendo retirado o encéfalo. O encéfalo foi armazenado em uma solução pós-
fixadora contendo formol 10% e sacarose a 30% durante um período de 24 horas (Figuras 18).
E por último, foi mantido em solução sacarose a 30% em tampão fosfato 0,1 M e PH 7,4 a 4º
C até ser submetido a microtomia.
3.3.1.2. Fixação por imersão
Dois animais (TA2 e TA3) não passaram pelo procedimento de perfusão. Ao invés
disso, depois de detectado o óbito do animal, eles tiveram seus encéfalos retirados. O passo
seguinte foi a fixação por imersão em formol 10% durante um média de 20 dias, até ser
detectado que o encéfalo estava consistente. Finalizada a fixação, os encéfalos passaram por
uma graduação crescente de sacarose (10%, 20% e 30%). E foram armazenados em sacarose
30% em tampão fosfato 0,1 M e PH 7,4 a 4º C até serem submetidos a cortes coronais pelo
procedimento de microtomia em criostato.
A B
C D

34
Figura 17. Tartaruga-verde (Chelonia mydas) juvenil macho que veio a óbito. (A) Visão dorsal. (B) Visão ventral. (C) Retirada de plastrão sem danificar os grandes vasos sanguíneos. (D) Incisão de agulha no ventrículo para passagem de solução fixadora (paraformaldeído 4%) pelo sistema sanguíneo do animal com auxílio de seringa. (E) Ruptura da região dorsal da caixa craniana para acesso ao encéfalo; (F) visão lateral do encéfalo.
Figura 18. Tartaruga adulta da espécie Chelonia mydas (tartaruga verde). (A) Ruptura da região dorsal da caixa craniana para acesso ao encéfalo; (B) (C) visão geral do encéfalo em relação à cabeça; (D) encéfalo retirado e encaminhado para o processo de pós-fixação.
3.3.2. Microtomia
Inicialmente, esperou o criostato (Luptec CM 2850) atingir uma temperatura média de
-20°C. Após isso, com o auxílio de uma cola especial (Tissue-T, O.C.T. Compound) o
encéfalo foi fixado longitudinalmente e em sentido rostrocaudal em um suporte para ser
cortado. No procedimento de microtomia, a espessura das secções coronais foi padronizada
E F
A B
C D

35
em 30μm e foram distribuídas em seis compartimentos. O primeiro compartimento, continha
tampão fosfato 0,1 M, pH 7,4 e foi utilizado para o método de Nissl. As secções do primeiro
compartimento obedeceram a ordem rostrocaudal, sendo mantidas uma isoladas das outras na
seguinte ordem: a, b, c, d, em diante. Os demais compartimentos (2, 3, 4, 5 e 6) continham
solução anticogelante e os cortes não foram isolados um dos outros, pois havia como colocá-
los em ordem tendo as lâminas montadas do Nissl. Após a microtomia, os cortes destes
últimos compartimentos foram mantidos em temperatura abaixo de 0°C até serem utilizados
em outro procedimento, como a imunoistoquímica para TH. Os cortes foram colocados nos
compartimentos de maneira cíclica. Primeiro, era colocado no compartimento 1a, depois nos
compartimentos 2, 3, 4, 5 e 6, em ordem crescente, até voltar para o compartimento 1b e
repetir todo o ciclo. A distância entre as secções de um mesmo compartimento foi de
aproximadamente 180 μm (Figura 19).
Figura 19. (A) Procedimento de corte do encéfalo utilizando criostato em temperatura média de – 20°C e
secções coronais de 30 μm. (B) Encéfalo de tartaruga marinha orientado em posição longitudinal, em sentido
rostrocaudal, e fixado a um suporte e envolvido por meio de uma cola especial. Após a retirada dos cortes do criostato com o auxílio de um pincel, eles são colocados em placas de ensaio de uso laboratorial. (C) A placa da esquerda contém solução anticogelante e foi destinada as secções que passaram (imunoistoquímica para TH)
C B
A
Compartimento 1 Compartimentos
a
b
c
g 2
3
4
5
6

36 ou passarão (outros procedimentos) por métodos imunológicos, enquanto a placa da direita foi destinada as secções que passaram pelo método de Nissl. O compartimento 1 recebeu as secções na ordem a,b,c,d em diante, os compartimentos 2, 4, 5 e 6 foram transferidos para recipientes com solução anticongelante e mantidos em temperatura menor que 0°C até serem usados.
3.3.3. Método de Nissl
Foi utilizado o método de Nissl como forma de delimitar a citoarquitetura do
mesencéfalo em tartaruga marinha. Um dos corantes usados neste método é a thionina, que
apresenta afinidade por corpos celulares, especialmente por uma das organelas, o retículo
endoplasmático rugoso. Os cortes do primeiro compartimento foram montados em lâminas
de vidro gelatinizadas, para isso, cada corte era retirado da placa de ensaio, colocado em uma
placa de petri ou prato de vidro contendo solução montagem e com o auxílio de um pincel era
direcionado para a lâmina até ser posicionado da forma preferida. Após esta etapa, as lâminas
com os cortes eram mantidos em temperatura ambiente por pelo menos 24 horas até secarem.
Após isso, foram submetidos ao método de Nissl,
O passo seguinte foi a submissão das lâminas com os cortes já secos no método de
coloração de Nissl, utilizando-se a thionina como corante para permitir uma melhor
demarcação das estruturas. O procedimento foi realizado em duas baterias. A primeira foi a
desidratação/diafanização do tecido e sua posterior reidratação (em sentido inverso) até ser
imerso na thionina. A segunda foi a passagem das lâminas por mais uma bateria de
desidratação. Antes de iniciar o procedimento, a thionina foi tirada da geladeira e quando
alcançou a temperatura ambiente foi aquecida em estufa a 50° C por pelo menos 15 minutos
antes do uso.
1. Primeira bateria:
(desidratação e diafanização)
a. Álcool 70 – de 2 horas à overnigth até no máximo 18 horas;
b. Álcool 95 I e II – 3 minutos cada;
c. Álcool 100 I e II – 3 minutos cada;
d. Xilol I e II – 3 minutos o primeiro e 30 minutos o segundo;
(Rehidratação – mesma bateria em sentido inverso)
e. Xilol I – 2 minutos;
f. Álcool 100 II e I – 2 minutos cada;
g. Álcool 95 II e I – 2 minutos cada;
h. Álcool 70 – 2 minutos;

37
i. Álcool 50 – 2 minutos;
j. Água destilada – 2 minutos;
(Coloração)
k. Thionina – 40 a 60 segundos
2. Segunda bateria:
(desidratação e diafanização)
a. Água destilada I e II – 5 batidinhas na primeira e 10 batidinhas na segunda;
b. Álcool 50 – 2 minutos;
c. Álcool 70 -2 minutos;
d. Álcool 95 I e II – 2 minutos cada;
e. Álcool 100 I, II e III – 2 minutos cada;
f. Xilol I e II – 2 minutos o primeiro e 4 minutos o segundo.
Por último, foram colocadas as lamínulas, fixadas com cola a lâmina, tendo o cuidado
para não ficarem bolhas que viessem atrapalhar na análise em microscópio.
A B
C

38 Figura 20. (A) Secção coronal do mesencéfalo de tartaruga marinha imersa em solução montagem para ser
posicionada em lâmina histológica gelatinizada. (B) Cortes já organizados em lâminas passando pelo procedimento de secagem em temperatura ambiente. (C) Lâminas passando pelas baterias do método de Nissl.
3.3.4. Imunoistoquímica para Tirosina Hidroxilase
Para aplicação do método de imunoistoquímica para tirosina hidroxilase, foi escolhido
um dos compartimentos mantidos em solução anticogelante em temperatura menor que 0°C.
Precedendo a técnica de imunoistoquímica, foi necessário realizar um pré-tratamento dos
cortes, na perspectiva de atenuar ou abolir a presença de artefatos. Para tal, as secções foram
lavadas durante cinco minutos em tampão fosfato 0,1 M, ph 7,4 e, posteriormente,
permaneceu por 20 minutos em solução de peróxido de hidrogênio H2O2 a uma concentração
de 0,03%. Durante todo o procedimento, os cortes ficaram em movimento em rotor
(velocidade de 40 rpm).
O próximo passo consistiu na incubação com anticorpo primário. As secções foram
colocadas em contato com uma solução formada pelo anticorpo anti-TH obtido em
camundongo (Sigma) em diluição de 1:5.000, soro normal de asno a 2% e Triton X-100 a
0,4% onde permaneceu em incubação por 20 horas em rotor com rotação lenta (40 rpm). Ao
fim deste período os cortes foram lavados cinco vezes durante cinco minutos cada lavagem
em tampão fosfato 0,1M, PH 7,4 para depois serem colocados em contato com o anticorpo
secundário anti-camundongo obtido em cavalo (Sigma) diluído a 1:1.000, por 90 minutos a
temperatura ambiente, sob agitação lenta, em rotor.
Após a incubação com o anticorpo secundário, os cortes foram submetidos mais uma vez
a cinco lavagens de cinco minutos cada em tampão fosfato 0,1M, ph 7,4. Em seguida, os
cortes foram colocados na solução do complexo avidina-biotina-HRP (protocolo ABC, Kit
elite da Vector), numa diluição de 1:100 em Triton X-100 a 0,4% contendo NaCl durante 90
minutos em rotação lenta. Para que fosse possível visualizar a reação, as secções foram
colocadas em meio contendo peróxido de hidrogênio (H2O2), como substrato, e a 3,3,4,4
tetrahidrocloretodiaminobenzidina (DAB), utilizada como cromógeno, durante 10 minutos.
Esta etapa foi realizada dentro de capela e com maior atenção, devido a toxicidade do
reagente DAB. Ao final, os cortes foram lavados por cinco vezes, cinco minutos cada, em
tampão fosfato 0,1 M, pH 7,4 em rotação lenta.

39
Os cortes foram montados em lâminas gelatinizadas e, após secas, foram imersas em
solução de tetróxido de ósmio a 0,05% com o intuito de intensificar a reação. A última etapa
foi a desidratação em baterias de álcool de graduação crescente até o álcool absoluto e
diafanização em xilol, sendo montadas as lamínulas ao final do processo.
Figura 21. Alguns dos procedimentos no método da imunoistoquímica. (A) Visão geral da bancada exibindo ferramentas utilizadas, como ponteiras de pipetas, solução tampão fosfato 0,1 M, pH 7,4 e Triton X-100 contendo cloreto de sódio, placa de ensaio imunológico, cestinha com fundo em tela para receber as secções e mantê-las em contato com as soluções oferecidas e rotor. (B) Cortes dentro de cestinhas com solução tampão fosfato 0,1 M, pH 7,4 passando pelo procedimento das lavagens através de rotação. (C) Pipetagem de anticorpo primário – uma das várias etapas em que se usam pipetas para colocar a quantidade correta, medidas em μl de reagentes. (D) Secções do mesencéfalo na etapa de imersão em meio contendo peróxido de hidrogênio como substrato e DAB como cromógeno para visualização da reação, procedimento que ocorre dentro de capela e com maior atenção devido a toxicidade do reagente DAB.
3.3.5. Obtenção das imagens e análise dos resultados
As secções do tronco encefálico, coradas pelo método de Nissl e/ou submetidas a
imunoistoquímica para TH foram examinadas ao microscópio óptico (Olympus BX41) em
campo claro. Imagens digitais foram obtidas de secções representativas usando uma câmera
digital (Nikon DXM1200) acoplada ao microscópio, sendo usadas as objetivas de 4X, 10X e
20X. Com o auxílio do software Canvas X 2018, as imagens foram analisadas, corrigidas
minimamente para brilho e contraste e os resultados foram documentados em
A B
C D

40
fotomicrografias. A partir das fotomicrografias foram construídos esquemas e ilustrações
usando como base o “A Stereotaxic Atlas of Forebrain and Midbrain of the Eastern Painted
Turtle (Chrysemis picta)” (POWERS; REINER, 1980) e a nomenclatura dos trabalhos Smeets
et al., (1987), Smeets (1988) e Power e Reiner (1993).
Para análise da área e perímetro dos corpos dos neurônios foi utilizado o software ImageJ.
Foram escolhidas duas secções representativas de cada grupamento (RA8, A9 e A10) ao
longo da extensão mesencefálica.
3.3.6. Análise Estatística
Para análise estatística foi utilizado Teste Anova de um fator para comparação do
perímetro e área dos corpos de neurônios imunorreativos a TH nas áreas de interesse. O teste
de tukey foi utilizado como teste de post hoc. Para todos os testes realizados, foi considerado
p<0,05 como sendo significativo.
41
4. RESULTADOS
Durante o período de dezembro de 2016 a janeiro de 2018 foram coletados quatro
encéfalos de tartarugas marinhas. Dessas, uma foi classificada como adulta e três como
juvenis/subadultas. Três vieram a óbito no Centro de Reabilitação do Projeto Cetáceos da
Costa Branca e passaram imediatamente pelo procedimento de retirada do encéfalo. Uma
tartaruga marinha foi registrada já morta durante o monitoramento de praia por um dos
biólogos do PCCB/UERN e devido ao seu estado bem conservado, foi possível caracterizar a
morfologia externa do encéfalo ainda dentro da caixa craniana através de uma secção sagital
do crânio (Quadro 01).
Quadro 01. Exemplares de tartaruga-verde (Chelonia mydas) utilizados no estudo, com a descrição do seu sexo,
comprimento x largura (cm) e procedimento em que foi utilizado.
COD Espécie Sexo Comprimento x
Largura do
Casco (cm)
Procedimento aplicado
TA1 Chelonia mydas Fêmea 57,0 x 57,0
(Juvenil)
Descrição da morfologia externa
(foto), Método de Nissl e
Imunoistoquímica para TH.
TA2 Chelonia mydas Macho 91,0 x 82,0
(Adulto)
Método de Nissl e
Imunoistoquímica para TH.
TA3 Chelonia mydas Macho 41,0 x 38,0
(Juvenil)
Método de Nissl e
Imunoistoquímica para TH.
TA4 Chelonia mydas Fêmea 37,0 x 33,0
(Juvenil)
Descrição da morfologia externa
(foto).
Por tratar-se de animais em risco de extinção, a perfusão só pôde ser realizada após a
detecção do óbito do animal no Centro de Reabilitação do PCCB-UERN.
4.1. Morfologia Externa
O encéfalo de tartaruga-verde juvenil e adulta apresenta-se divido com delimitações
bem definidas. Em sentido rostral a caudal e em vista dorsal, observa-se nervos e bulbo
olfatório, cérebro, lóbulo óptico, cerebelo e medula. Em visão lateral e ventral é possível notar
os nervos ópticos e oculomotor, assim como outros nervos craniais (Figuras 22 e 23).
Em visão dorsal e sentido rostrocaudal, nota-se o bulbo olfatório fazendo limite com
os hemisférios cerebrais (telencéfalo) e estes, por sua vez, fazendo limite posteriormente com
a borda anterior dos lóbulos ópticos. O cerebelo está em posição mais caudal, fazendo
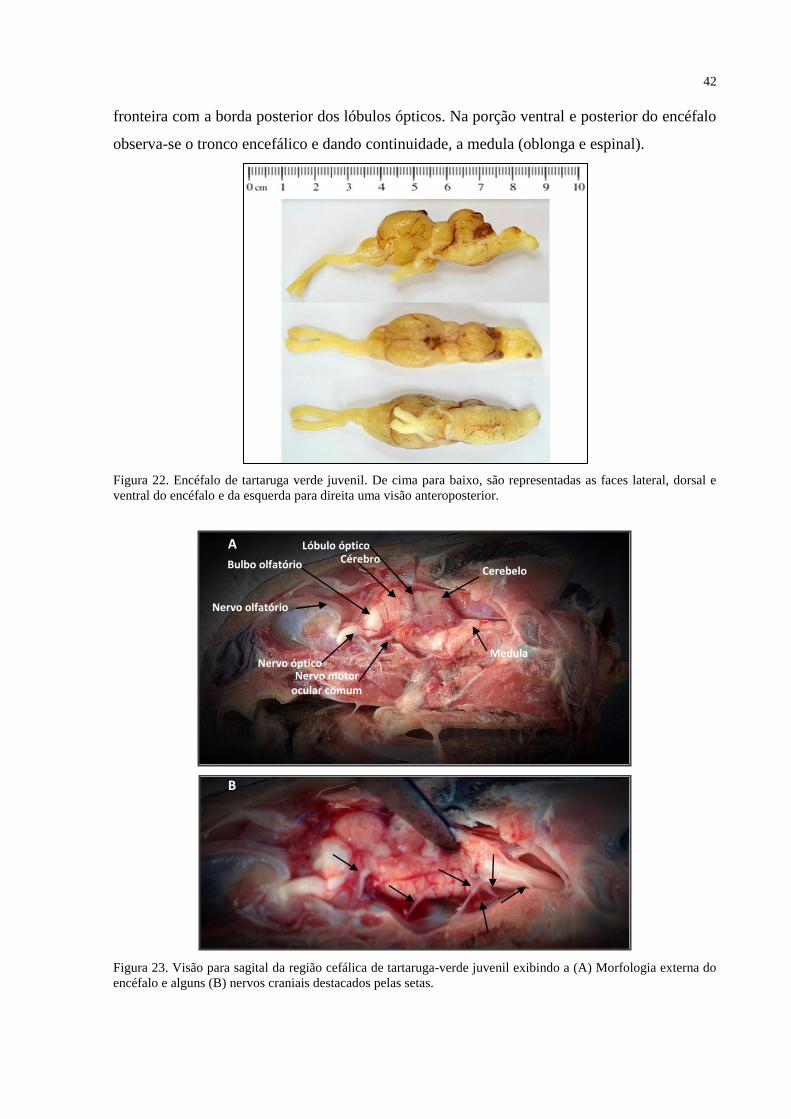
42
fronteira com a borda posterior dos lóbulos ópticos. Na porção ventral e posterior do encéfalo
observa-se o tronco encefálico e dando continuidade, a medula (oblonga e espinal).
Figura 22. Encéfalo de tartaruga verde juvenil. De cima para baixo, são representadas as faces lateral, dorsal e
ventral do encéfalo e da esquerda para direita uma visão anteroposterior.
Figura 23. Visão para sagital da região cefálica de tartaruga-verde juvenil exibindo a (A) Morfologia externa do
encéfalo e alguns (B) nervos craniais destacados pelas setas.
A
B
Medula
Cerebelo
Nervo motor ocular comum
Lóbulo óptico Cérebro Bulbo olfatório
Nervo olfatório
Nervo óptico

43
4.2. Microscopia
Através da técnica de imunoistoquímica para tirosina hidroxilase foi possível
identificar grupamentos neuronais no mesencéfalo de tartaruga-verde. O tegmento
mesencefálico de C. mydas contém células dopaminérgicas imunorreativas a TH que podem
ser agrupadas de três formas de acordo com a posição dos neurônios e a orientação de seus
dentritos: área tegmental ventral (VTA/A10), substância negra (SN/A9) e o equivalente
reptiliano do grupo A8 de mamíferos (RA8) (Figura 24).
A imunoistoquímica para TH junto com o método de Nissl permitiu descrever a
citoarquitetura dos grupamentos A10, A9 e RA8 em tartaruga marinha pela primeira vez
(Figuras 26, 27, 28, 29 e 30). Em secções coronais e com sentido rostrocaudal, é possível
identificar os grupamentos A9 e A10 em níveis mais rostrais. O primeiro possui localização
ventrolateral no tegmento mesencefálico e apresenta divisa citoarquitetônica ventral ao núcleo
rubro, e lateralmente faz limite com o pedúnculo ventral do fascículo prosencefálico lateral
(PV) e fascículo longitudinal medial (FLM). Em sua continuidade, faz limite dorsalmente
com o núcleo profundo mesencefálico (NPrM) e em níveis caudais dá lugar ao grupamento
RA8. A SN pode ser classificada em substância negra pars compacta (SNc), rica em corpos
de neurônios e seus processos, e pars reticulata (SNr), com menor quantidade de corpos de
neurônios e processos longos orientados, predominantemente, mediolateralmente.
A VTA encontra-se em posição ventromedial em níveis mesencefálicos rostrais. Em
seu limite mais dorsal, faz fronteira com as células do núcleo do nervo oculomotor (nIII) e
segue direção ventrolateral, se juntando com as células dopaminérgicas da SN lateralmente.
Em direção caudal, a VTA faz limite lateral com o núcleo interpendicular (IP), ventromedial
com o grupamento RA8 e, por último, em extremo caudal, faz limite ventralmente com o
núcleo reticular superior (Rs). Em todo a extensão mesencefálica é possível observar células
dopaminérgicas pertencentes a VTA. Porém, é em níveis mais caudais que há uma forte
marcação dos neurônios dopaminérgicos imunorreativos a TH. Os processos dos neurônios
dopaminérgicos na VTA são caracterizados por serem curtos.
Em níveis caudais, surge o grupamento RA8 como uma extensão da SN que está
localizada dorsolateralmente a VTA. Possui um menor número de corpos de neurônios e seus
processos, assim como os da SNr, são longos, porém, com orientação predominantemente
ventrolaterais.
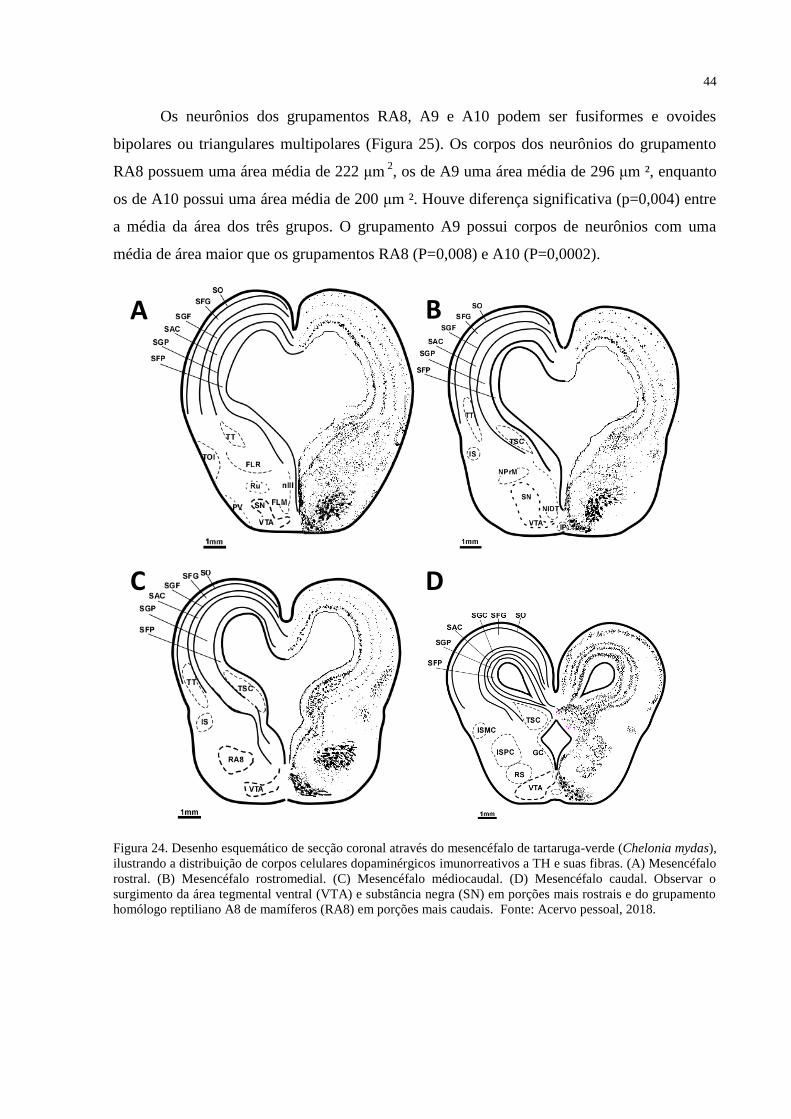
44
Os neurônios dos grupamentos RA8, A9 e A10 podem ser fusiformes e ovoides
bipolares ou triangulares multipolares (Figura 25). Os corpos dos neurônios do grupamento
RA8 possuem uma área média de 222 μm 2
, os de A9 uma área média de 296 μm ², enquanto
os de A10 possui uma área média de 200 μm ². Houve diferença significativa (p=0,004) entre
a média da área dos três grupos. O grupamento A9 possui corpos de neurônios com uma
média de área maior que os grupamentos RA8 (P=0,008) e A10 (P=0,0002).
Figura 24. Desenho esquemático de secção coronal através do mesencéfalo de tartaruga-verde (Chelonia mydas),
ilustrando a distribuição de corpos celulares dopaminérgicos imunorreativos a TH e suas fibras. (A) Mesencéfalo
rostral. (B) Mesencéfalo rostromedial. (C) Mesencéfalo médiocaudal. (D) Mesencéfalo caudal. Observar o
surgimento da área tegmental ventral (VTA) e substância negra (SN) em porções mais rostrais e do grupamento
homólogo reptiliano A8 de mamíferos (RA8) em porções mais caudais. Fonte: Acervo pessoal, 2018.
A B
C D
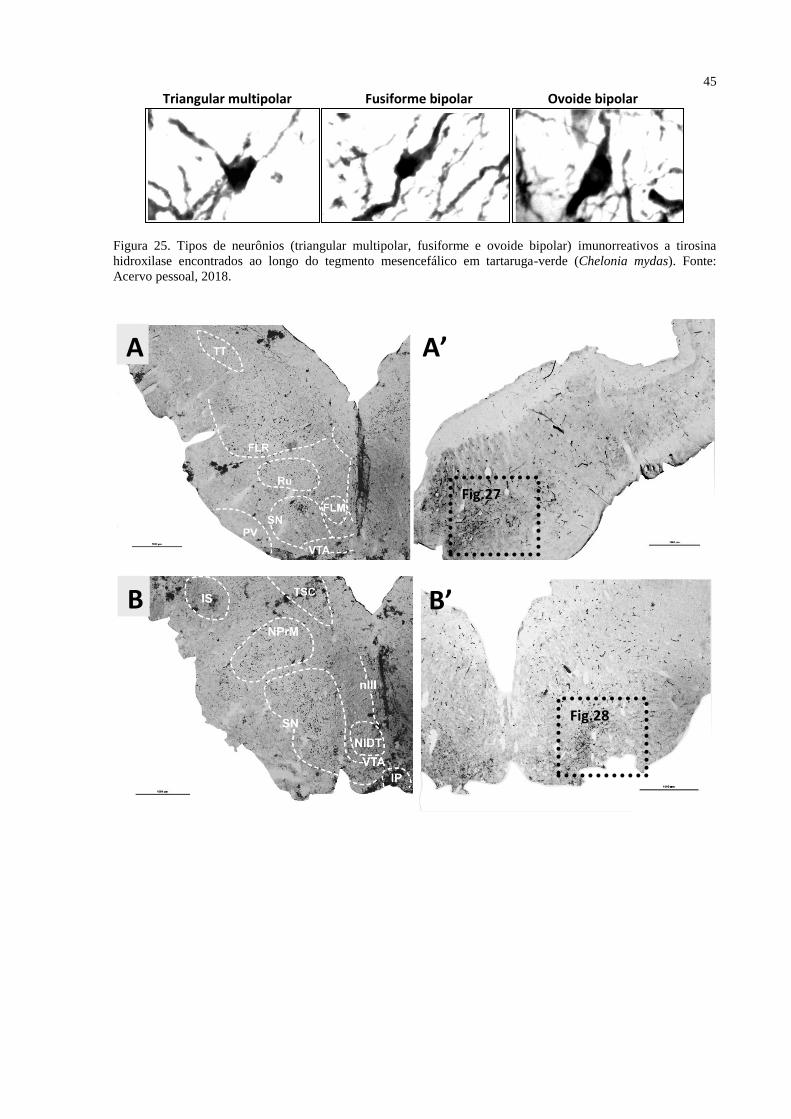
45
Figura 25. Tipos de neurônios (triangular multipolar, fusiforme e ovoide bipolar) imunorreativos a tirosina
hidroxilase encontrados ao longo do tegmento mesencefálico em tartaruga-verde (Chelonia mydas). Fonte:
Acervo pessoal, 2018.
Triangular multipolar Fusiforme bipolar Ovoide bipolar
A A’
B’ B
Fig.28
Fig.27
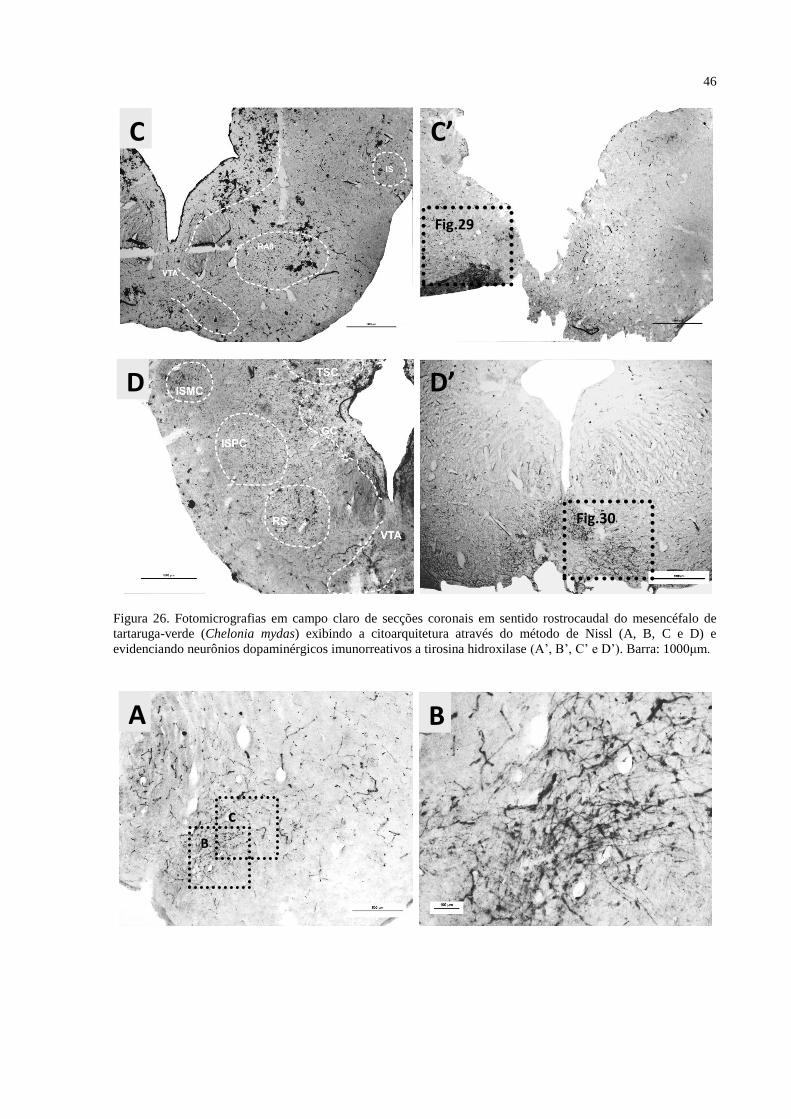
46
Figura 26. Fotomicrografias em campo claro de secções coronais em sentido rostrocaudal do mesencéfalo de
tartaruga-verde (Chelonia mydas) exibindo a citoarquitetura através do método de Nissl (A, B, C e D) e
evidenciando neurônios dopaminérgicos imunorreativos a tirosina hidroxilase (A’, B’, C’ e D’). Barra: 1000μm.
C
D
C’
D’
A B
B
c
Fig.30
Fig.29
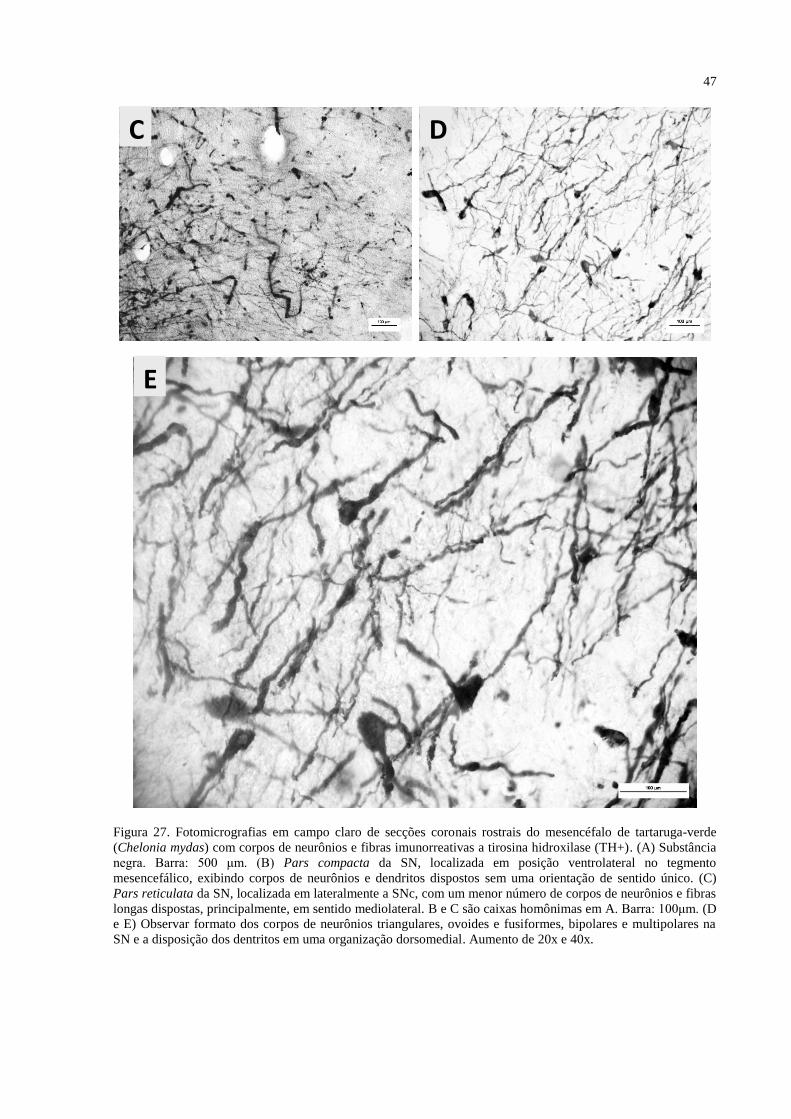
47
Figura 27. Fotomicrografias em campo claro de secções coronais rostrais do mesencéfalo de tartaruga-verde
(Chelonia mydas) com corpos de neurônios e fibras imunorreativas a tirosina hidroxilase (TH+). (A) Substância
negra. Barra: 500 μm. (B) Pars compacta da SN, localizada em posição ventrolateral no tegmento
mesencefálico, exibindo corpos de neurônios e dendritos dispostos sem uma orientação de sentido único. (C)
Pars reticulata da SN, localizada em lateralmente a SNc, com um menor número de corpos de neurônios e fibras
longas dispostas, principalmente, em sentido mediolateral. B e C são caixas homônimas em A. Barra: 100μm. (D
e E) Observar formato dos corpos de neurônios triangulares, ovoides e fusiformes, bipolares e multipolares na
SN e a disposição dos dentritos em uma organização dorsomedial. Aumento de 20x e 40x.
C D
E
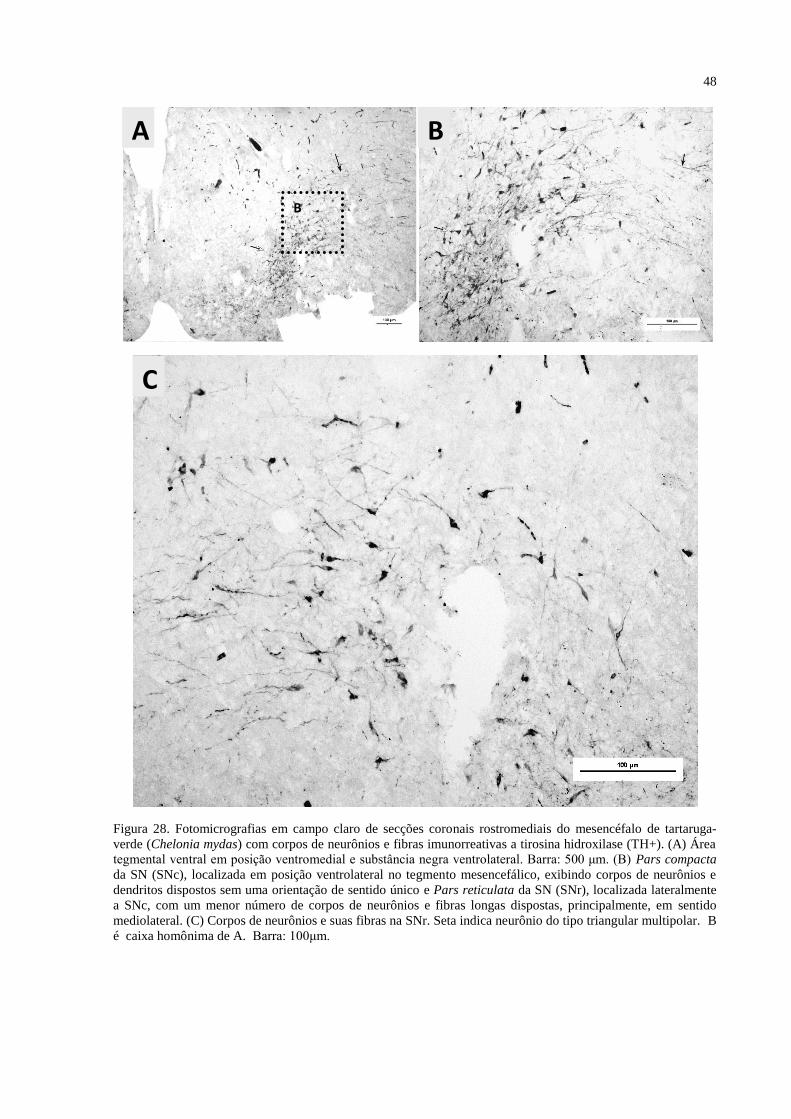
48
Figura 28. Fotomicrografias em campo claro de secções coronais rostromediais do mesencéfalo de tartaruga-
verde (Chelonia mydas) com corpos de neurônios e fibras imunorreativas a tirosina hidroxilase (TH+). (A) Área
tegmental ventral em posição ventromedial e substância negra ventrolateral. Barra: 500 μm. (B) Pars compacta
da SN (SNc), localizada em posição ventrolateral no tegmento mesencefálico, exibindo corpos de neurônios e
dendritos dispostos sem uma orientação de sentido único e Pars reticulata da SN (SNr), localizada lateralmente
a SNc, com um menor número de corpos de neurônios e fibras longas dispostas, principalmente, em sentido
mediolateral. (C) Corpos de neurônios e suas fibras na SNr. Seta indica neurônio do tipo triangular multipolar. B
é caixa homônima de A. Barra: 100μm.
A B
C
B
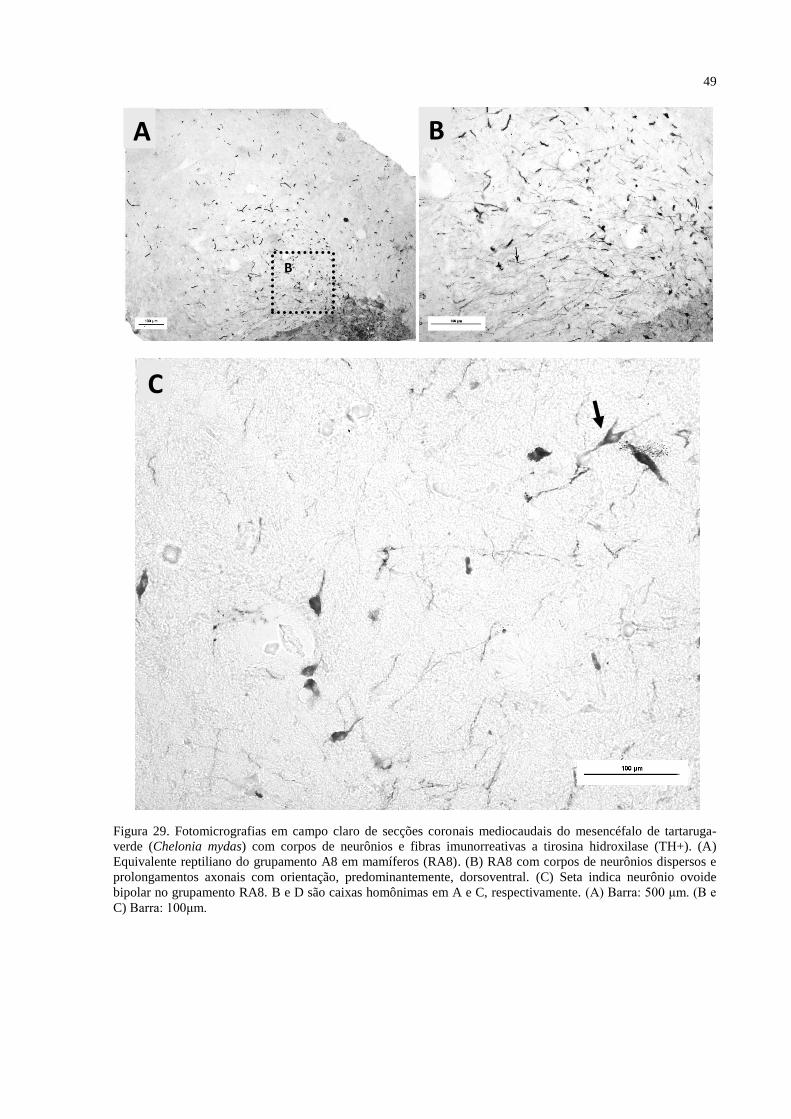
49
Figura 29. Fotomicrografias em campo claro de secções coronais mediocaudais do mesencéfalo de tartaruga-
verde (Chelonia mydas) com corpos de neurônios e fibras imunorreativas a tirosina hidroxilase (TH+). (A)
Equivalente reptiliano do grupamento A8 em mamíferos (RA8). (B) RA8 com corpos de neurônios dispersos e
prolongamentos axonais com orientação, predominantemente, dorsoventral. (C) Seta indica neurônio ovoide
bipolar no grupamento RA8. B e D são caixas homônimas em A e C, respectivamente. (A) Barra: 500 μm. (B e
C) Barra: 100μm.
B
A B
C
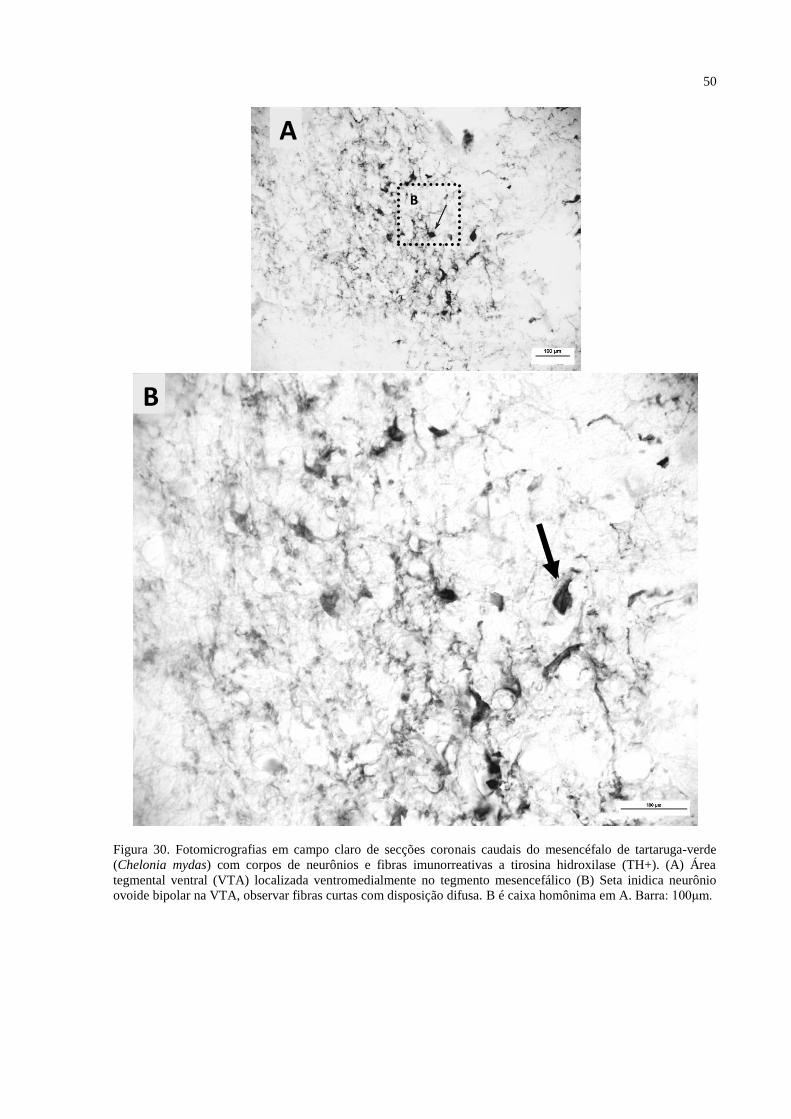
50
Figura 30. Fotomicrografias em campo claro de secções coronais caudais do mesencéfalo de tartaruga-verde
(Chelonia mydas) com corpos de neurônios e fibras imunorreativas a tirosina hidroxilase (TH+). (A) Área
tegmental ventral (VTA) localizada ventromedialmente no tegmento mesencefálico (B) Seta inidica neurônio
ovoide bipolar na VTA, observar fibras curtas com disposição difusa. B é caixa homônima em A. Barra: 100μm.
B
B
A

51
5. DISCUSSÃO
Dentro do subfilo Vertebrata, células dopaminérgicas no mesencéfalo podem estar
ausentes (peixes ciclóstomos), podem apresentar um único grupamento (osteíctes), dois
grupamentos (peixes condrictes e anfíbios) e três grupamentos ou mais (répteis, aves e
mamíferos) (SMEETS; GONZÁLEZ, 2000).
Em mamíferos, há uma maior variação na organização dentrítica, assim como, no tipo
de neurônios dopaminérgicos ao longo do mesencéfalo dependendo da espécie e da divisão e
subdivisão dos grupamentos celulares. Em estudos realizados em primata (CAVALCANTI et
al., 2016), roedores (MOON et al., 2007; LIMACHER et al., 2008; PIETERS et al., 2010;
CAVALCANTI et al., 2014) e quirópteros (MASEKO et al., 2007; DELL et al., 2010;
MEDEIROS et al., 2016), nas subdivisões do complexo VTA/A10, esses neurônios podem se
apresentar na forma ovoide bipolar e piriforme multipolar, com organização dentrítica
variando entre dorsoventral, dorsolateral ou sem um padrão de organização específico. O
complexo SNC/A9 pode apresentar diferentes formatos de corpos de neurônios e
organizações dentríticas também, dependendo da subdivisão e do animal, sendo relatada a
existência de neurônios ovoides, triangulares, piriformes, fusiformes, sendo bipolares ou
multipolares, sem organização dentrítica, com orienação dendrítica, predominantemente,
horizontal e orientação dorsoventral. Os neurônios da RFF/A8 podem ser do tipo ovoide e
fusiformes, do tipo bipolar ou multipolar, sem organização dentrítica especifica ou com
organização no sentido ventrolateral e dorsolateral.
Ao contrário de mamíferos, poucos estudos têm sido feitos descrevendo neurônios
imunorreativos a DA no mesencéfalo de répteis. A maioria dos estudos com esse intuito
foram realizados na década de 80 usando imunoistoquímica para TH e DA.
O mesencéfalo em répteis faz limite anteriormente com o diencéfalo e posteriormente
com o cerebelo. Em sua citoarquitetura, podem-se destacar as áreas do tectum, núcleo
posterodorsal, comissura posterior, núcleo central do torus semicircular, núcleo laminar do
torus semicircular em posição dorsal. Em posição ventral são identificados a VTA, SN, RA8,
fascículo longitudinal medial, núcleo interpendicular pars ventralis, núcleo do nervo
oculomotor (SMEETS; HOOGLAND; VOORN, 1986; SMEETS; JONKER; HOOGLAND,
1987).

52
A grande maioria das células CA/DA em répteis são encontradas no tegmento
mesencefálico. Embora os autores não especifiquem, a partir das fotomicrografias
provenientes das pesquisas é possível notar células ovoides e fusiformes bipolares no
mesencéfalo da tartaruga P. scripta (SMEETS; JONKER; HOOGLAND, 1987), no lagarto G.
gecko (SMEETS; HOOGLAND; VOORN, 1986), na cobra P. regius (SMEETS, 1988) e no
jacaré C. crocodilus (BRAUTH, 1988). Já no lagarto V. exanthematicus são descritos
neurônios com corpos arredondados ou ovoides de tamanho médio na SN (WOLTERS;
DONKELAAR; VERHOFSTAD, 1984). Em C. mydas é possível observar células bipolares
e multipolares com formatos ovais, fusiformes ou estrelados em toda a extensão do tegmento
mesencefálico, não sendo possível identificar a existência significativa de diferença entre os
tipos de neurônios entre os grupamentos neuronais mesencefálicos. No entanto, as células DA
da SN são em média maiores que dos demais grupamentos DA mesencefálicos (VTA e RA8).
Além da tripartição dos grupamentos neuronais imunorreativos a DA no mesencéfalo
de répteis, podem ser detectadas outras áreas com neurônios imunorreativos. Como por
exemplo, no lagarto V. exanthematicus são detectadas poucas células catecolaminérgicas de
tamanho pequeno a médio no núcleo laminar do torus semicircular e núcleo interpendicular
dorsal não observadas em tartaruga marinha C. mydas.
No presente estudo, a distribuição de neurônios dopaminérgicos imunorreativos a TH
no mesencéfalo da tartaruga-verde foi descrita pela primeira vez. Embora seja difícil a
diferenciação entre os diferentes tipos de catecolamina (CA) como dopamina, noradrenalina e
adrenalina usando apenas a técnica de imunoistoquímica para TH, é bem aceito que anticorpo
contra TH demonstre, preferencialmente, o sistema dopaminérgico (SMEETS; HOOGLAND;
VOORN, 1986 cita HÖKFELT et al., 1977). Trabalhos realizados com outros representantes
reptilianos, usando tanto anticorpo contra TH (WOLTERS; DONKELAAR; VERHOFSTAD,
1984; BRAUTH, 1988), como contra DA (SMEETS; HOOGLAND; VOORN, 1986),
mostraram que os grupamentos neuronais dopaminérgicos manteram um padrão básico no
mesencéfalo. Dessa forma, é justificável utilizar imunoistoquímica para TH, ao invés de para
DA, na descrição de tais grupamentos. O padrão organizacional dos grupamentos neuronais
DA mesencefálicos em tartaruga-verde é claramente comparável com o de outros
representantes reptilianos, o que corrobora com a eficácia de anticorpos contra TH em marcar
estruturas neuronais DA no mesencéfalo de répteis.
Comparando os estudos feitos em lagarto (Gekko gecko), jacaré (Caiman crocodilus),
cobra (Python regius) e tartaruga de água doce (Pseudemys scripta elegans) pode-se observar

53
um padrão básico da distribuição de neurônios dopaminérgicos no mesencéfalo de répteis,
definido, mais ou menos, da seguinte maneira: um grupamento medial (VTA); um
grupamento lateral (SN); e, em níveis mesencefálicos caudais, pode ser encontrado um
grupamento dorsolateral, denominado grupamento RA8 (SMEETS; HOOGLAND; VOORN,
1986; SMEETS et al., 1987).
Tartaruga-verde apresenta o padrão de células dopaminérgicas semelhante ao
observado na tartaruga P. scripta e lagarto G. gecko, onde uma tripartição dos grupos
celulares pode ser feita tendo como base, principalmente, a posição e direção dos processos
dos neurônios sintetizantes de dopamina (REINER et al., 1984; SMEETS; HOOGLAND;
VOORN, 1986). A SNr localizada, principalmente, lateral a SNc e possui processos longos e
direcionados lateralmente. Por outro lado, na cobra P. regius existe uma parte medial e lateral
de grupamentos celulares dopaminérgicos que poderia ser considerado a VTA e a SN,
respectivamente, mas que o autor considera um único grupamento e o denomina de
grupamento do complexo VTA/SN. A região presumida como SN não mostra processos que
são direcionados lateralmente. O motivo pelo qual os processos na P. regius não sejam
longos, nem direcionados lateralmente, pode ser explicado pela posição da SNr em relação ao
seu plexus da substância P, onde a localização das células dopaminérgicas coincide quase em
sua totalidade com a área que mostra maior imunorreatividade para a substância P (SMEETS,
1988).
A distribuição de dopamina no mesencéfalo do lagarto G. gecko, assim como foi
observado em tartaruga marinha C. mydas apresentou a tripartição dos grupamentos DA de
forma semelhante. O grupamento mais rostral está localizado na VTA e SN e na parte mais
caudal do mesencéfalo é identificado o RA8. VTA assume posição ventromedial no tegmento,
enquanto SN e RA8 se posicionam dorsolateralmente a VTA. Outro fato que coincide é que
as células dopaminérgicas da SN são, em geral, um pouco maiores que as células da VTA e
seus processos são longos e de orientação, predominantemente, mediolateral. O RA8
apresente neurônios DA com processos orientados, principalmente, em sentido dorsoventral
(SMEETS; HOOGLAND; VOORN, 1986).
Além dos grupamentos RA8, A9 e A10, são identificadas células imunorreativas a TH
e DA em outras áreas mesencefálicas reptilianas. Em duas espécies de lagartos (V.
exanthematicus e A. carolinensis) foram encontradas células imunorreativas a TH/DA na zona
periventricular. Na tartaruga P. scripta células adicionais imunorreativas a DA foram
observadas no núcleo laminar do torus semicircular (TSC) (SMEETS; GONZÁLEZ, 2000).

54
No presente estudo, não foi observado nenhum grupo adicional imunorreativo a TH no
mesencéfalo de tartaruga marinha.

55
6. CONCLUSÃO
O presente estudo, por meio do método de Nissl e da imunoistoquímica para TH,
revela os grupamentos dopaminérgicos do mesencéfalo da tartaruga-verde (Chelonia mydas),
sendo descritos pela primeira vez. Estes caracterizam-se por apresentarem citoarquitetura e
distribuição de neurônios semelhante ao observado em outro répteis. Os grupamentos
neuronais dopaminérgicos imunorreativos a TH no mesencéfalo apresentam uma tripartição,
apresentando-se como A10, A9 e RA8.
Sugere-se o desenvolvimento de estudos complementares que descrevam a
citoarquitetura do sistema nervoso em répteis, com o intuito da realização de uma descrição
geral do sistema dopaminérgico nesses animais, aumentando assim, o entendimento das
variações entre os diferentes representantes reptilianos e entre esses e outros representantes
em vertebrados.

56
7. REFERÊNCIAS BIBLIOGRÁFICAS
BARROT, Michel. The ventral tegmentum and dopamine: a new wave of diversity.
Neuroscience, v. 282, p. 243-247, 2014.
BARREIRO-IGLESIAS, Antón; VILLAR-CERVINO, Verona; ANADÓN, Ramón;
RODICIO, Maria Celina. Descending brain-spinal cord projections in a primitive vertebrate,
the lamprey: cerebrospinal fluid-contacting and dopaminergic neurons. Journal of
Comparative Neurology,v.511, pp. 711–723, 2008.
BEAR, Mark F.; CONNORS, Barry W.; PARADISO, Michael A. Neurociências:
desvendando o sistema nervoso. 2 ed. Porto Alegre: Artmed. 2002. 857p.
BARRAL, Jaime; GALARRAGA, Elvira; TAPIA, Dagoberto; FLORES-BARRERA, Edén;
REYES, Arturo; BARGAS, José. Dopaminergic modulation of spiny neurons in the turtle
striatum. Cellular and Molecular Neurobiology, v. 30, pp. 743-750, 2010.
BENTIVOGLIO, Marina; MORELLI, Micaela. The organization and circuits of
mesencephalic dopaminergic neurons and the distribution of dopamine receptors in the brain.
In Dunnett, S.B. et al., Handbook of chemical Neuroanatomy, v. 21, ed Elsevier, pp. 1-107,
2005.
BERNSTEIN, Alison I.; STOUT, Kristen A.; MILLER, Gary W. The vesicular monoamine
transporter 2: An underexplored pharmacological target. Neurochemistry International, v. 73,
pp. 89-97, 2014.
BJÖRKLUND, Anders; DUNNETT, Stephen B. Dopamine neuron systems in the brain: an
update. Trends in Neurosciences, v. 30, p. 194-202, 2007.
BONELLI, Raphael M; CUMMINGS, Jeffrey L. Frontal-subcortical circuitry and behavior.
Dialogues in Clinical Neuroscience, v. 9, pp. 141-151, 2007.
BRAUTH, Steven E.; KITT, Cheryl A. The paleostriatal system of Caiman crocodile. The
Journal of Comparative Neurology, v. 189. pp. 437-465, 1980.
BRAUTH, Stevens E.; REINER, Anton.; KITT, Cheryl A.; KARTEN, Harvey J. The
substance P-containing striatotegmental path in reptiles: an immunohistochemical study. The
Journal of Comparative Neurology, v. 219, pp. 305-327, 1983.
CARLSSON, Arvid; BENGT, Falck; HILLARP, Nils-Ake. Cellular localization of brain
monoamines. Acta Physiologica Scandinavica, Suppl, v. 56, suppl, 196, pp. 1–28, 1962.
CAVE, John W.; BAKER, Harriet. Dopamine systems in the forebrain. Advances in
Experimental Medicine and Biology. v. 651, p. 15-35, 2009.
CAVALCANTI, José Rodolfo; SOARES, Joacil Germano; OLIVEIRA, Francisco Gilberto;
GUZEN, Fausto Pierdoná; PONTES, André Luiz Bezerra; SOUSA, Twyla Barros.de;
CAVALCANTE, Jeferson Souza; NASCIMENTO-JR, Expedito Silva; CAVALCANTE,
Judney Cley; COSTA, Miriam Stela. A cytoarchitectonic and TH-immunohistochemistry
characterization of the dopamine cell groups in the substantia nigra, ventral tegmental area

57
and retrorubral field in the rock cavy (Kerodon rupestris). Journal of Chemical
Neuroanatomy, v.55, pp. 55-68, 2014.
CAVALCANTI, José Rodolfo Lopes de Paiva; PONTES, André Luiz Bezerra; FIUZA, Felipe
Porto; SILVA, Kayo Diógenes de Azevedo; GUZEN, Fausto Pierdoná; LUCENA, Eudes
Euler de Souza; NASCIMENTO-JÚNIOR, Expedito Silva; CAVALCANTE, Judney
Cley; COSTA, Miriam Stela Maris de Oliveira; ENGELBERTH, Rovena Clara Galvão
Januário; CAVALCANTE, Jeferson Souza. Nuclear organization of the substantia nigra,
ventral tegmental area and retrorubral field of the common marmoset (Callithrix jacchus): A
cytoarchitectonic and TH-immunohistochemistry study. ). Journal of Chemical
Neuroanatomy, v. 77, pp. 100-109, 2016.
CHEN, Xiqun; XU, Lu; RADCLIFFE, Pheona; SUN, Baoyong; TANK, William A.
Activation of Tyrosine Hydroxylase mRNA Translation by cAMP in Midbrain Dopaminergic
Neurons. Molecular Pharmacology, v. 73 (6), pp. 1816-1828, 2008.
DAHLSTRÖM, Annica. and FUXE, Kjell. Evidence for the existence of monoamine-
containing neurons in the central nervous system. I. Demonstration of monoamines in the cell
bodies of brain stem neurons. Acta Physiologica Scandinavica. v. 62, Suppl. 232, pp. 1–55,
1964.
DE LA MORA, Miguel Pérez; GALLEGOS-CARI, Andrea; ARIZMENDI-GARCÍA, Yexel;
MARCELLINO, Daniel; FUXE, Kjell. Role of dopamine receptor mechanisms in the
amygdaloid modulation of fear and anxiety: Structural and functional analysis. Progress in
Neurobiology, v. 90, p. 198-216, 2010.
DELL, Leigh-Anne; KRUGER, Jean-Leigh; BHAGWANDIN, Adhil; JILLANI, Ngalla E.;
PETTIGREW, John D.; MANGER, Paul R. Nuclear organization of cholinergic, putative
catecholaminergic and serotonergic systems in the brains of two megachiropteran species.
Journal of Chemical Neuroanatomy, v. 40, pp. 177–195, 2010.
DELONG, Mahlon R.; WICHMANN, Thomas. Circuits and circuit disorders of the basal
ganglia. Archives of Neurology, v. 64, p. 20–24, 2007.
DONKELAAR, Hendrik J. ten; HUIZAN, Roelie de Boer-Van. Ascending projections of the
brain sten reticular formation in a nonmammalian vertebrate (the lizard Varanus
exanthematicus), with notes on the afferent connections of the forebrain. The Journal of
Comparative Neurology, v. 280, pp. 501-528, 1981.
GEFFARD, Michel; BUIJS, Ruud M.; SEGUELA, Philippe; POOL, Christiaan W.; LE
MOAL, Michel. First demonstration of highly specific and sensitive antibodies against
dopamine. Brain Research, v. 294, pp. 161-165, 1984.
GONZÁLEZ, Hugo; CONTRERAS, Francisco; PRADO, Carolina; ELGUETA, Daniela;
FRANZ, Dafne; BERNALES, Sebastián; PACHECO, Rodrigo. Dopamine receptor D3
expressed on CD4+ T cells favors neurodegeneration of dopaminergic neurons during
Parkinson's disease. The Journal of Immunology, v. 190, pp. 5048-5056, 2013.
GONZALEZ, Agustin; SMEETS, Wilhelmus. Comparative Analysis of Dopamine and
Tyrosine Hydroxylase Immunoreadivities in the Brain of Two Amphibians, the Anuran Rana
ridibunda and the urodele Pleurodeles waltlii. The Journal of Comparative Neurology, v. 303,
pp. 457-477, 1991.

58
GOOLE, Jonathan; AMIGHI, Karim. Levodopa delivery systems for the treatment of
Parkinson's disease: An overview. [Review] International Journal of Pharmaceutics, v. 380
(1), pp. 1-15, 2009.
HABER, Suzanne N. The place of dopamine in the cortico-basal ganglia circuit. [Review]
Neuroscience, v. 282, pp. 248-257, 2014.
HALASZ, Nobert; NOWYCKY, Martha.; HÖKFELT, Tomas; SHEPHERD, Gordon Murray;
MARKEY, Keith; GOLDSTEIN, Menek. Dopaminergic periglomerular cells in the turtle
olfactory bulb. Brain Res. Bull. v. 9, pp. 383-389, 1982.
HIDALGO-SANCHEZ, Matías; ALVARADO-MALLART, Rosa-Magda. Temporal
sequence of gene expression leading caudal prosencephalon to develop a midbrain/hindbrain
phenotype, Developmental Dynamics, v. 223, pp. 141–147. 2002.
HÖKFELT, Tomas; MÅRTENSSON, Roger; BJÖRKLUND, Anders; KLEINAU, Sandra;
GOLDSTEIN, Menek. Distribution maps of tyrosine-hydroxylase-immunoreactive neurons in
the rat brain. In: Björklund, Anders; Hökfelt, Tomas editors. Classical Transmitters in the
CNS. Handbook of Chemical Neuroanatomy. Part I. Volume 2. Amsterdam: Elsevier; 1984.
pp. 277–379.
IKEMOTO, Satoshi; YANG, Chen; TAN, Aaron. Basal ganglia circuit loops, dopamine and
motivation: a review and enquiry. Behavioural Brain Research, v. 290, pp. 17-31, 2015.
JACKSON, David M; WESTLIND-DANIELSSON, Anita. Dopamine receptors: molecular
biology, biochemistry and behavioural aspects. Pharmacology & Therapeutics, v. 64, p. 291–
370, 1994.
JONES, Hugh M.; PILOWSKY, Lyn S. Dopamine and antipsychotic drug action revisited.
The British Journal of Psychiatry, v. 181 (4), p. 271-275, 2002.
KIM, Tae Eun; LEE, Hack Sup; LEE, Yong Beom; HONG, Seung Hwan; ICHINOSE,
Hiroshi; KIM, Seung U; LEE, Myung Ae. Sonic hedgehog and FGF8 collaborate to induce
dopaminergic phenotypes in the Nurr1-overexpressing neural stem cell. Biochemical and
Biophysical Research Communications, v. 305, pp. 1040– 1048, 2003.
KOHNO, Milky; MORALES, Angelica M; GUTTMAN, Zoe; LONDON, Edythe. A neural
network that links brain function, white-matter structure and risky behavior. Neuroimage, v.
149, p. 15-22, 2017.
LAU, Chi-leong; WANG, Han-Cheng; HSU, Jung-Lung; LIU, Mu-En. Does the dopamine
hypothesis explain schizophrenia? Reviews in the Neuroscience, v. 24, pp. 389–400, 2013.
LAWLOR, Patricia A; DURING, Matthew J. Gene therapy for Parkinson's disease. Expert
Reviews in Molecular Medicine, v. 6, pp. 1-18, 2004.
LENT, Roberto. Cem bilhões de neurônios. São Paulo: Editora Atheneu, 2005. 786p.
LIU, Aimin; JOYNER, Alexandra L. Early anterior/posterior patterning of the midbrain and
cerebellum, Annual Review of Neuroscience, v. 24, pp. 869– 896, 2001.
LIMACHER A, BHAGWANDIN A, FUXE K, MANGER PR. Nuclear organization and
morphology of cholinergic, putative catecholaminergic and serotonergic neurons in the brain

59
of the Cape porcupine (Hystrix africaeaustralis): increased brain size does not lead to
increased organizational complexity. The Journal of Chemical Neuroanatomy, v. 36, pp. 33-
52, 2008.
LÓPEZ, Jesús M.; GONZÁLEZ, Agustín. Organization of the catecholaminergic systems in
the brain of lungfishes, the closest living relatives of terrestrial vertebrates. The Journal of
Comparative Neurology, v. 525, pp. 3083-3109, 2017.
MARCOVALDI, Maria Ângela; MARCOVALDI, Guy Guani. Marine turtles of Brazil: the
history and structure of Projeto TAMAR-IBAMA. Biological Conservation, [S.l.], v. 91,
p.35-41, 1999.
MARÍN, Faustino; HERRERO, María-Trinidad; VYAS. Sheela, PUELLES, Luis. Ontogeny
of tyrosine hydroxilase mRNA expression in mid- and forebrain: neuromeric pattern and
novel positive regions. Developmental Dynamics, v. 234 (3), pp. 709-717, 2005.
MASEKO, B. C; BOURNE, J. A; MANGER, P. R. Distribution and morphology of
cholinergic, putative catecholaminergic and serotonergic neurons in the brain of the Egyptian
rousette flying fox, Rousettus aegyptiacus. Journal of Chemical Neuroanatomy, v. 34, pp.
108-127, 2007.
MEDEIROS, Hélder Henrique Alves; SANTANA, Melquisedec Abiaré Dantas; LEITE,
Mariana Dias; AQUINO, Luisa Alves Pereira; BARROS, Marília Abero Sá; GALVÃO,
Nayana Tânia; LADD, Fernando Vagner Lobo; CAVALCANTE, Judney Cley; COSTA,
Miriam Stela Maris Oliveira; CAVALCANTE, Jeferson Souza; NASCIMENTO-JÚNIOR,
Expedito Silva. The cytoarchitectonic and TH-immunohistochemical characterizationof the
dopamine cell groups in the substantia nigra, ventral tegmentalarea and retrorubral field in a
bat (Artibeus planirostris). Neuroscience Research, v. 112, pp. 37-46, 2016.
MEYLAN, Anne B.; MEYLAN, Peter A. Introduction to the evolution, life history, and
Biology of sea turtles. IUCN/SSC Marine Turtle Specialist Group Publication, [S.l.], n. 4, p.
3-5, 1999.
MONEY, Kelli M.; STANWOOD, Gregg D. Developmental origins of brain disorders: roles
for dopamine. Frontiers in Cellular Neuroscience, v. 7, p. 260, 2013.
MOONS, Lieve; Van GILS, Jelle; GHIJSELS, Els; VANDESANDE, Frans.
Immunocytochemical localization of L-dopa and dopamine in the brain pf the chicken (Gallus
domesticus). The Journal of Comparative Neurology, v. 346, pp. 97-118, 1994.
MOON, Don-Joon; MASEKO, Busisiwe C.; IHUNWO, Amadi O.; FUXE, Kjell; MANGER,
Paul R. Distribution and morphology of catecholaminergic and serotonergic neurons in the
brain of the highveld gerbil, Tatera braintsii. The Journal of Comparative Neurology, v. 34,
pp. 134-144, 2007.
MTSG. IUCN SSC Marine Turtle Specialist Group. Green turtle (Chelonia mydas).
Disponível em: https://iucn-mtsg.org/about-turtles/species/green/. Acesso em: 06 de agosto de
2017.
NAGATSU, Toshi; STJÄRNET, Lennart. Catecholamine Synthesis and Release. Advances in
Pharmacology, v. 42, pp. 1-14, 1997.

60
NAGATSU, Toshiharu; HIROSHI, Ichinose. Molecular Biology of Catecholamine-Related
Enzymes in Relation to Parkinson's Disease. Cellular & Molecular Neurobiology, v. 19 (1),
pp. 57-66, 1999.
NAUMANN, Robert K.; ONDRACEK, Janie M.; REITER, Samuel; SHEIN-IDELSON,
Mark; TOSCHES, Maria Antonietta; YAMAWAKI, Tracy M.; LAURENT, Gilles. The
reptilian brain. Currente Biology, v. 25 (8), pp. 317-321, 2015.
NIEH, Edward H.; KIM, Sung.-Yon; NAMBURI, Praneeth; TYE, Kay M. Optogenetic
dissection of neural circuits underlying emotional valence and motivated behaviors. Brain
Research, v. 1511, p. 73–92, 2013.
NOAAFISHERIES. Disponível em: <https://www.seeturtles.org/>. Acesso em: 16 de julho de
2017.
O’CONNELL, Lauren A.; MATTHEWS, Bryan J.; RYAN, Michel J.; HOFMANN, Hans A.
Characterization of the dopamine system in the brain of the túngara frog, Physalaemus
pustulosus. Brain, Behavior and Evolution, v. 76, pp. 211-225, 2010.
PACHECO, Rodrigo; CONTRERAS, Francisco; ZOUALI, Moncef. The dopaminergic
system in autoimmune diseases. Frontiers in Immunology, v. 5, p. 117, 2014.
PARENT, André; POIRIER, Louis J. Occurrence and distribution of monoamine-containing
neurons in the brain of the painted turtle, Chrysemys picta. Journal of Anatomy, v. 110, pp
81-89, 1971.
PARENT, André. Monoaminergic systems of the brain. In Biology of the Reptilia (ed. Gans
C.). Academic Press, London, v. 10, pp. 247-285, 1979.
PEREZ-SANTANA, Lilian; MARÍN, Oscar; SMEETS, Wilhelmus J. A. J. Afferent
connections of the nucleus accumbens of the snake, Elaphe guttata, studied by means of in
vitro and in vivo tracing techniques in combination with TH immunohistochemistry,
Neuroscience Letters, v. 225, p. 101-104, 1997.
PIETERS, Raymond P.; GRAVETT, Nadine; FUXE, Kjell; MANGER, Paul R. Nuclear
organization of cholinergic, putative catecholaminergic and serotonergic nuclei in the brain of
the eastern rock elephant shrew, Elephantulus myurus. Journal of Chemical Neuroanatomy, v.
39, pp. 175-188, 2010.
PRADO, Carolina; BERNALES, Sebastina; PACHECO, Rodrigo. Modulation of T-cell
mediated immunity by dopamine receptor d5. Endocrine, Metabolic & Immune Disorders.
Drug Targets, v. 13, pp. 184-194, 2013.
PRAKASH, Nilima. WURST, Wolfgang. Development of dopaminergic neurons in the
mammalian brain. Cellular and Molecular Life Sciences, v. 63, p. 187-206, 2006.
REINER, Anton; Karten, Harvey J. Basal ganglionic pathways to the tectum: studies in
reptiles. The Journal of Comparative Neurology, v. 193, p. 565-589, 1980.
RIDDLE, Robert; POLLOCK, Jonathan D. Making connections: the development of
mesencephalic dopaminergic neurons. Developmental Brain Research, v. 147, p. 3 – 21. 2003.

61
RINK, Elke; WULLIMANN, Mario F. The teleostean (zebrafish) dopaminergic system
ascending to the subpallium (striatum) is located in the basal diencephalon (posterior
tuberculum). Brain Research, v. 889, pp. 316–330, 2001.
SALAMONE, John D.; CORREA, Mercè. Motivational views of reinforcement: implications
for understanding the behavioral functions of nucleus accumbens dopamine. Behavioural
Brain Research, v. 137, pp. 3–25, 2002.
SCHULTZ, Wolfram. Updating dopamine reward signals. Current Opinion in Neurobiology,
v. 23 (2), p. 229-238, 2013.
SEETURTLES. Disponível em: < https://www.seeturtles.org/>. Acesso em: 16 de julho de
2017.
SHIH, Ming Chi; AMARO JR, Edson; FERRAZ, Henrique Ballalai; HOEXTER, Marcelo
Queiroz; GOULART, Fabricio Oliveira; WAGNER, Jairo; LIN, Li Fu; FUL, Ying Kai;
MARI, Jair Jesus; LACERDA, Acioly Luiz Tavares de; TUFIK, Sergio; BRESSAN, Rodrigo
Affonseca. Neuroimagem do transportador de dopamina na doença de Parkinson: primeiro
estudo com [99m
Tc]-TRODAT-1 e SPECT no Brasil. Arquivos de Neuro-Psiquiatria, v. 64,
pp. 628-634, 2006.
SIBLEY, David R.; MONSMA, Jr., Frederick J.; SHEN, Yong. Molecular neurobiology of
dopaminergic receptors. International Review of Neurobiology, v. 35, pp. 391-415, 1993.
SILVA, Guilherme Pereira da. O campo magnético natural terrestre como parâmetro da
movimentação oceânica de tartarugas marinhas. Tese (Doutorado) – Universidade Federal de
Pernambuco. CTG. Programa de Pós-Graduação em Oceanografia, 2013.
SMEETS, Wilhelmus J. A. J.; HOOGLAND, Piet V.; VOORN, Pieter. The distribuition of
dopamine immunoreactivity in the forebrain and midbrain of the lizard Gekko gecko: an
immunohistochemical study with antibodies against dopamine. The Journal of Comparative
Neurology , v. 253, pp. 46-60, 1986.
MEREDITH, Gloria E.; SMEETS, Wilhelmus J. A. J. Immunocytochemical analysis of the
dopamine system in the forebrain and midbrain of Raja radiate: evidence for a substantia
nigraand ventral tegmental area in cartilaginous fish. Journal of Comparative Neurology, v.
265, pp. 530-548, 1987.
SMEETS, Wilhelmus J. A. J. Distribution of dopamine immunoreactivity in the forebrain and
midbrain of the snake Python regius: a study with antibodies against dopamine. The Journal
of Comparative Neurology, v. 271, pp. 115-129, 1988.
SMEETS, Wilhelmus J. A. J.; GONZÁLEZ, Agustin. Cathecolamine systems in the brain of
vertebrates: new perspectives through a comparative approach. Brain Research Reviews, v.
33, p. 308-379, 2000.
SPÜHLER, Isabelle Ayumi; HAURI, Andreas. Decoding the dopamine signal in macaque
prefrontal cortex: a simulation study using the Cx3Dp simulator. PLOS ONE, v, 8 (8), p.
71615, 2013.
STUESSE, Sherry L.; CRUCE, William L. R.; NORTHCUTT, R. Glenn. Localization of
catecholamines in the brains of Chondrichthyes (cartilaginous fishes). In Wilhelmus J. A. J.

62
Smeets & Anton Reiner (Eds.), Phylogeny and development of catecholamine systems in the
CNS of vertebrates. Cambridge: Cambridge University Press, pp. 21-47, 1994.
STRANGE, Philip G. New insights into dopamine receptors in the central nervous system.
Neurochemistry International, v. 22, p. 223-236, 1993.
SUGAM, Jonathan A.; DAY, Jeremy J.; WIGHTMAN, R. Mark; CARELLI, Regina M.
Phasic nucleus accumbens dopamine encodes risk-based decision-making behavior.
Biological Psychiatry, v. 71,pp. 199-205, 2012.
TEMPLE, John G.; MILLER, Diane B.; BARTHALMUS, George T. Differential
vulnerability of snake species to MTPT: A behavioral and biochemical comparison in
ratsnakes (Elaphe) and watersnakes (Nerodia). Neurotoxicology and Teratology, v. 24, pp.
227-233, 2002.
The IUCN Red List of Threatened Species. Version 2017.1. <www.iucnredlist.org>. Acesso
em: 05 de agosto de 2017.
UNGERSTEDT, Urban. Stereotaxic mapping of the monoamine pathways in the rat brain.
Acta Physiologica Scandinavica, v. 82, supp. 367, pp. 1–48, 1971.
URBAN, Nina B. L.; SLIFSTEIN, Mark; MEDA, Shashwath; XU, Xiaoyan; AYOUB,
Rawad; MEDINA, Olga; PEARLSON, Godfrey D.; KRYSTAL, John H.; ABI-DARGHAM,
Anissa. Imaging human reward processing with positron emission tomography and functional
magnetic resonance imaging. Psychopharmacology, v. 221, p. 67–77, 2012.
VAN DEN HEUVEL, Dianne M. A; PASTERKAMP, R. Jeroen. Getting connected in the
dopamine system. Progress in Neurobiology, v. 85, pp 75–93, 2008.
VERNEY, Catherine; ZECEVIC, Nada; PUELLES, Luis. Structure of longitudinal brain
zones that provide the origin for the substantia nigra and ventral tegmental area in human
embryos, as revealed by cytoarchitecture and tyrosine hydroxylase, calretinin, calbindin, and
GABA immunoreactions. Journal of Comparative Neurology, v. 429, pp. 22-44, 2001.
WISE, Roy A; KOOB, George F. The development and maintenance of drug addiction.
Neuropsychopharmacology, v. 39, pp. 254–262. 2014.
WU, Wen-Lian; BURNETT, Duane A.; SPRING, Richard; GRENLEE, William J.; SMITH,
Michelle; FAVREAU, Leonard; FAWZI, Ahmad; ZHANG, Hongtao; LACHOWICZ, Jean E.
Dopamine D1/D5 receptor antagonists with improved pharmacokinetics: design, synthesis,
and biological evaluation of phenol bioisosteric analogues of benzazepine D1/D5 antagonists.
Journal of Medicinal Chemistry, v. 48 (3), pp 680-693, 2005.
Reiner, Anton. The Conservative Evolution of the Vertebrate Basal Ganglia. In: Handbook of
Behavioral Neuroscience. Elsevier B.V., v. 24, pp. 63-97, 2017.
REINER, Anton; KARLE, Ellen J.; ANDERSON, Keith D.; MEDINA Loreta.
Catecholaminergic perikarya and fibers in the avian nervous system. In: SMEETS, Wilhelmus
J. A. J.; REINER, Anton (eds) Phylogeny and development of catecholamine systems in the
CNS of vertebrates. Cambridge University Press, Cambridge, pp 135–181, 1994.
WURST, Wolfgang; BALLY-CUIF, Laure. Neural plate patterning: upstream and
downstream of the isthmic organizer, Nature Reviews Neuroscience, v. 2, pp. 99– 108, 2001.

63
WYNEKEN, Jeanette. The anatomy of sea turtles. National Oceanic and Atmospheric
Administration, U.S. Department of Commerce, NOAA Technical Memorandum NMFS-
SEFSC-470, 2001.
YAMAMOTO, Kei; VERNIER, Philippe. The evolution of dopamine systems in chordates.
Frontiers in Neuroanatomy, v. 5, p. 21, mar. 2011.
YAMAMOTO, Kei; VERNIER, Philippe. The evolution of dopamine systems in chordates.
Frontiers in Neuroanatomy, v. 5, pp. 1-21, 2001.
YAN, Yiqing; JIANG, Wei; LIU, Lei; WANG, Xiaqiong; DING, Chen; TIAN, Zhigang;
ZHOU, Rongbin. Dopamine controls systemic inflammation through inhibition of NLRP3
inflammasome. Cell, v. 160, pp. 62-73, 2015.
YETNIKOFF, Leora; LAVEZZI, Heather N.; REICHARD, Rhett A.; ZAHM, Daniel S. An
update on the connections of the ventral mesencephalic dopaminergic complex. Neuroscience,
p. 23–48, 2014
WWF. Green Turtle, Malyasia. Disponível em:<https://www.worldwildlife.org/photos/green-
turtle-malyasia>. Acesso em: 03 de março de 2018.

64
AUTORIZAÇÃO DE CAPTURA, COLETA E TRANSPORTE DE MATERIAL
BIOLÓGICO
65

66

67

68
69

70
Top Related