







Línguas
Páginas
Legal


Helena Alexandra António da Fonseca
Estudo in vitro da toxicidade de corantes têxteis azo
em Tetrahymena pyriformis
Departamento de Zoologia/ Antropologia
Faculdade de Ciências da Universidade do Porto
Outubro/ 2006

Helena Alexandra António da Fonseca
Estudo in vitro da toxicidade de corantes têxteis azo
em Tetrahymena pyriformis
Tese submetida à Faculdade de Ciências da Universidade do Porto
para a obtenção do grau de Mestre em Ecologia Aplicada.
Departamento de Zoologia/ Antropologia
Faculdade de Ciências da Universidade do Porto
Outubro/ 2006

AGRADECIMENTOS Gostaria de agradecer ao meu orientador, Professor Doutor Nelson Lima, da Universidade do
Minho, por me ter dado a oportunidade de realizar o trabalho de mestrado no Departamento de
Engenharia Biológica da Universidade do Minho, pela paciência e pelos conselhos científicos.
Gostaria de agradecer à orientadora Professora Doutora Maria Teresa Borges, da Faculdade de
Ciências da Universidade do Porto, pelos conselhos amigos, acompanhamento e pelo incentivo.
Queria deixar o meu obrigada à co-orientadora Doutora Ana Nicolau, da Universidade do
Minho, pela disponibilidade, apoio, compreensão, e por me ajudar a melhorar o meu
desempenho laboratorial.
Fica aqui o meu “obrigada” às colegas investigadoras da Micoteca pelo companheirismo, pela
ajuda e pela partilha de alguns momentos, que tornaram a minha estadia no laboratório ainda
mais agradável.
Queria ainda agradecer à minha amiga e colega de trabalho Magda, pela preciosa ajuda dada
na estruturação e na revisão bibliográfica; pelos conselhos “linguísticos” e pela companhia.
Finalmente, não posso deixar de agradecer aos meus pais e avó, responsáveis pelo
desencadeamento da minha inscrição no mestrado, pelo eterno apoio e compreensão… e claro
ao meu marido, Daniel, sem o qual não teria conseguido concluir o mestrado e que ao longo do
tempo sempre me ajudou a continuar e me encorajou a acabar.
A todos aqueles que, directa ou indirectamente, de alguma forma contribuíram para a
realização desta tese, o meu obrigada.

ÍNDICE GERAL
RESUMO…………………………………………………………………………………………………..v
ÍNDICE DE FIGURAS…………………………………………………………………………………..vii
ÍNDICE DE TABELAS……………………………………………………………………………….…viii
I. INTRODUÇÃO GERAL…………..……………………………………………………………………1
2. TOXICOLOGIA E ECOTOXICOLOGIA – BREVE ABORDAGEM……………………………....5
2.1. INTRODUÇÃO......................................................................................................................6 2.1.MODOS DE ACÇÃO DOS TÓXICOS ..........................................................................................6 2.3.TESTES DE TOXICIDADE AQUÁTICA........................................................................................7 2.4. RELAÇÃO QUANTITATIVA ACTIVIDADE-ESTRUTURA (QSAR) E RELAÇÃO QUANTITATIVA ACTIVIDADE-PROPRIEDADE (QPAR) ...........................................................................................8 2.5. ORGANISMOS TESTE ..........................................................................................................9 2.6. MÉTODOS DE TESTES TRADICIONAIS....................................................................................9
2.6.1. Métodos agudos ou crónicos .....................................................................................9 2.6.2. Testes modificados, rápidos ou microbiotestes..........................................................9
3. TESTES DE TOXICIDADE COM PROTOZOÁRIOS…………………………………………….11
3.1. INTRODUÇÃO....................................................................................................................12 3.2. OS CILIADOS COMO ORGANISMOS TESTE............................................................................12 3.3. TETRAHYMENA PYRIFORMIS: UMA FERRAMENTA NA TOXICOLOGIA ........................................14
3.3.1. Cultivo in vitro ..........................................................................................................14 3.3.2. Ensaios para avaliar o efeito tóxico de compostos xenobióticos. .............................15 3.3.3. Os ensaios fisiológicos toxicológicos .......................................................................15 a. Densidade celular e taxa de crescimento ...................................................................16 b. Mudanças comportamentais.......................................................................................16 c. Crescimento, mortalidade e viabilidade celulares........................................................17 3.3.4. Os ensaios bioquímicos toxicológicos......................................................................18
4. OS PROTOZOÁRIOS CILIADOS: BREVE ABORDAGEM DA SUA MORFOLOGIA, FISIOLOGIA E TAXONOMIA………………………………………………………………………….21
4.1. INTRODUÇÃO....................................................................................................................22 4.2. MORFOLOGIA, FISIOLOGIA E TAXONOMIA ............................................................................22
5. CORANTES…………………………………………………………………………………………...25
5.1. INTRODUÇÃO....................................................................................................................26 5.2. PRINCIPAIS FONTES DE POLUIÇÃO NO SECTOR TÊXTIL E OS SEUS REQUERIMENTOS AMBIENTAIS..............................................................................................................................................27 5.3. LEGISLAÇÃO RELATIVA AOS CORANTES AZO .......................................................................28 5.4. CORANTES AZO UTILIZADOS ..............................................................................................28 5.5. SÍNTESE DOS CORANTES AZO............................................................................................30 5.6. ESTUDOS FEITOS COM OS CORANTES AZO UTILIZADOS........................................................31
6. METODOLOGIA……………………………………………………………………………………...32
6.1. MICROORGANISMO ...........................................................................................................33 6.2. CONDIÇÕES DE CULTURA E EXPOSIÇÃO A CORANTES AZO ...................................................33 6.3. COMPOSTOS TÓXICOS E SUAS CONCENTRAÇÕES................................................................34 6.4. ENSAIOS DE DETERMINAÇÃO DE TOXICIDADE .....................................................................35
6.4.1. Ensaios Fisiológicos Toxicológicos..........................................................................35

a. Ensaios de Crescimento: ............................................................................................35 b. Ensaios de Morfometria: .............................................................................................37 c. Ensaios de Predação:.................................................................................................37 6.4.2. Ensaios Bioquímicos Toxicológicos .........................................................................38
6.5. TRATAMENTO ESTATÍSTICO DOS DADOS .............................................................................38
7. RESULTADOS………………………………………………………………………………………..39
7.1. CRESCIMENTO .................................................................................................................40 7.2. MORFOMETRIA .................................................................................................................44 7.3. PREDAÇÃO.......................................................................................................................51 7.4. SÍNTESE DOS RESULTADOS...............................................................................................55
7.4.1. Síntese dos resultados para cada corante azo ........................................................55 7.4.2. Síntese dos resultados para os três testes realizados: crescimento, morfometria e predação. ..........................................................................................................................58
8. DISCUSSÃO…………………………………………………………………………………………..60
9. CONCLUSÕES……………………………………………………………………………………….66
10. REFERÊNCIAS BIBLIOGRÁFICAS……………………………………………………………...70

RESUMO
O aumento da poluição ambiental e o contínuo desenvolvimento da síntese de novos químicos
desencadeou uma crescente preocupação com os possíveis efeitos desses componentes
directa ou indirectamente na saúde humana.
Recentemente, a presença de tóxicos em ambientes aquáticos têm sido uma ocorrência
comum. Os corantes do tipo azo são bastante utilizados devido ao seu baixo custo de produção
assim como ao seu grande leque de aplicações. Porém, grandes quantidades destes corantes
são libertadas para o meio ambiente através dos efluentes industriais e a sua dispersão na
Natureza acarreta vários problemas.
O controlo da poluição aquática tem tido uma importância crescente nos últimos anos. A
libertação de corantes para o ambiente constitui apenas uma pequena porção da poluição da
água, mas os corantes são visíveis em quantidades pequenas devido à sua cor. A legislação
ambiental que existe é bastante rígida, forçando as indústrias têxteis a tratar os seus efluentes.
Este tratamento é físico-químico e muito dispendioso, além de criar um problema adicional, ou
seja, a concentração dos corantes nas lamas resultantes do tratamento dos efluentes. É
urgente combater os efeitos prejudiciais dos corantes têxteis no ambiente e a sua permanência
como resíduos recalcitrantes, inestéticos, evitando, simultaneamente, os grandes prejuízos que
as indústrias sofreriam se não pudessem produzir ou utilizar este tipo de corantes.
As alterações provocadas no meio ambiente pela introdução de substâncias tóxicas, são
susceptíveis de investigação, e, assim, a ecotoxicologia surge como um recurso para um
melhor conhecimento dos mecanismos de actuação dessas substâncias no ecossistema, mas
para isso é fundamental conhecer as alterações num indivíduo isolado. No que se relaciona
com a poluição da água os protozoários são uma excelente ferramenta de trabalho para o
estudo da toxicidade e da poluição.
No presente trabalho pretendeu-se utilizar o protozoário ciliado Tetrahymena pyriformis como
bioindicador no estudo de respostas fisiológicas e bioquímicas à presença de oito corantes azo
utilizados na indústria têxtil. Para tal, utilizou-se uma bateria de ensaios fisiológicos
toxicológicos como o crescimento, a morfometria e a predação. Estes ensaios foram realizados
numa série de testes miniaturizados usando culturas axénicas de T. pyriformis, inoculadas com
soluções dos 8 corantes em diferentes concentrações (5, 25, 50 e 100 ppm), tendo como
objectivo final a colecta de dados a fim de comparar respostas quanto à presença de diferentes
corantes têxteis azo e quanto à presença de diferentes concentrações de um mesmo corante.

Pretendeu-se com esta bateria de testes estudar se os corantes com aplicação têxtil utilizados
são tóxicos para o bioindicador utilizado e em que concentração produzem tal efeito. Espera-se
que com os resultados deste estudo se possa extrapolar a influência destes compostos no meio
aquático receptor.

ÍNDICE DE FIGURAS
Figura 1: Diagrama conceptual do estudo realizado, focando vários parâmetros a serem tomados em consideração para a realização dos testes de citotoxicidade com Tetrahymena pyriformis (adaptado de NICOLAU et al., 2001). .........................................................................3
Figura 2: Tetrahymena pyriformis retirada de uma cultura axénica em meio de cultura PPY (x100)........................................................................................................................................14
Figura 3: Células de Tetrahymena pyriformis viáveis e não viáveis, utilizando a calceína/AM e EthD-1. As duas células não viáveis mostram núcleos corados de vermelho (retirada de NICOLAU et al., 2001). .............................................................................................................18
Figura 4: ...................................................................................................................................23
- Diagrama A: Morfologia da Tetrahymena pyriformis: cp – citoprocto; cv – vacuolo contráctil; fv – vacúolo digestivo; mn - macronúcleo; n - núcleo; ki – corpo basal; um e m – membrana ondulatória e membranelas do aparelho oral (oa). ....................................................................23
- Diagrama B: Fotografia de contraste de fase de uma célula de Tetrahymena pyriformis fixada com vapor de 1 % de tetróxido de ósmio (x600)........................................................................23
Figura 5: Estrutura dos oito corantes azo (adaptado de MARTINS et al., 2001). ......................29
Figura 6: Esquema da síntese dos corantes azo (retirada de MARTINS et al., 2001)...............30
Figura 7 A e B: Corantes azo em pó (A) e soluções stock de 500 ppm dos corantes azo utilizados para os ensaios de toxicidade (B)..............................................................................34
Figura 8: Crescimento de Tetrahymena pyriformis exposta a quatro concentrações de corantes azo: 5 ppm (gráfico A), 25 ppm (gráfico B), 50 ppm (gráfico C) e 100 ppm (gráfico D), em ensaios de 48 horas. Para cada tempo de amostragem são representadas a médias e os desvios padrão de quatro réplicas.............................................................................................50
Figura 9: Efeito da exposição a diferentes concentrações de corantes azo no tamanho (área) de Tetrahymena pyriformis, em ensaios de 48 horas. As concentrações usadas foram de 5 ppm (gráfico A), 25 ppm (gráfico B), 50 ppm (gráfico C), e 100 ppm (gráfico D). Para cada tempo de amostragem são representadas a médias e os desvios padrão de quatro réplicas. ..................45
Figura 10: Efeito da exposição a diferentes concentrações de corantes azo na razão (W/L) do eixo menor (W) e maior (L) de Tetrahymena pyriformis, em ensaios de 48 horas. As concentrações usadas foram de 5 ppm (gráfico A), 25 ppm (gráfico B), 50 ppm (gráfico C), e 100 ppm (gráfico D). Para cada tempo de amostragem são representadas a médias e os desvios padrão de quatro réplicas.............................................................................................47

ÍNDICE DE TABELAS
Tabela 1: Vantagens e desvantagens dos testes de toxicidade (adaptada de MITCHELL et al., 2002)…………………..……………………………………………………………………………………7
Tabela 2: Organismos principais dos quais foram isoladas estirpes e para as quais testes toxicológicos especiais foram desenvolvidos (adaptado de SAUVANT et al., 1999)……..……..13
Tabela 3: Setup do contador automático de células -Beckman Coulter Z Series - para um tubo capilar de abertura com 140 µm …….…………………………….…………………………………..36
Tabela 4: Resultados dos testes t de Student para a comparação das médias de n.º de células/ml de amostras sujeitas aos corantes com as médias dos controlos, para todas as concentrações e tempos de amostragem, em dois ensaios independentes (1 e 2); os valores apresentados representam o valor do p estatístico………………………………………………….43
Tabela 5: Resultados dos testes ANOVA para o crescimento de Tetrahymena pyriformis. A tabela faz a comparação entre médias de grupos de corantes de todas as concentrações: guaiacol-siringol (g-s); carboxílico-sulfónico(C-S); meta-para (m-p), para os respectivos tempos de amostragem. Os valores apresentados representam o valor do p estatístico………….……..44
Tabela 6: Resultados dos testes t de Student, para a comparação de médias das áreas de amostras sujeitas aos corantes com as médias dos controlos, para todas as concentrações e tempos de amostragem. Os valores apresentados representam o valor do p estatístico……….49
Tabela 7: Resultados dos testes t de Student para a comparação das médias das razões (W/L) de amostras sujeitas aos corantes com as médias dos controlos, para todas as concentrações e tempos de amostragem. Os valores apresentados representam o valor do p estatístico……….50
Tabela 8: Resultados dos testes ANOVA para a morfometria: área e razão (W/L), de Tetrahymena pyriformis. A tabela faz a comparação entre médias de grupos de corantes de todas as concentrações: guaiacol-siringol (g-s); carboxílico-sulfónico(C-S); meta-para (m-p), para os respectivos tempos de amostragem. Os valores apresentados representam o valor do p estatístico………………………………………………………………………………………………....51
Tabela 9: Número de microesferas ingeridas durante 20 minutos por 30 indivíduos de populações de Tetrahymena pyriformis expostos a diferentes concentrações de corantes azo: 5, 25, 50 e 100 ppm, analisadas durante o ensaio 1 com a duração de 48 horas. Os valores apresentados representam o valor do p estatístico…………………………………………………………………………………………………53
Tabela 10: Resultados dos testes ANOVA para a predação de Tetrahymena pyriformis. A tabela faz a comparação entre médias de grupos de corantes de todas as concentrações: guaiacol-siringol (g-s); carboxílico-sulfónico(C-S); meta-para (m-p), para os respectivos tempos de amostragem. Os valores apresentados representam o valor do p estatístico……………………55

I. INTRODUÇÃO GERAL

Os corantes azo são utilizados na tinturaria têxtil para corar e pintar fibras naturais e sintéticas,
couros e peles, na tipografia a cores, e ainda como aditivos de produtos derivados do petróleo.
São compostos xenobióticos, uma vez que contêm grupos ausentes em moléculas naturais e,
portanto, nem todos os microrganismos que normalmente se encarregam da reciclagem da
matéria orgânica lançada no ambiente possuem a "maquinaria enzimática" necessária ao
processamento de todos os tipos de grupos funcionais. O grupo funcional xenobiótico principal
nos corantes azo é a ligação azo, ou seja -N=N- (FERREIRA, 1998).
As indústrias de corantes e as indústrias têxteis são, respectivamente, as maiores produtoras e
utilizadoras de corantes azo. Produzem-se anualmente toneladas destes corantes, sendo cerca
de 10 a 15 % desses compostos lançados nos efluentes. Uma vez que estes corantes são
recalcitrantes à degradação microbiana, os efluentes destes processos industriais são
normalmente resistentes ao tratamento biológico, seja por microrganismos seja por plantas,
sendo muitas vezes os processos de descontaminação fisico-químicos a única alternativa de
tratamento para estas águas residuais (FERREIRA, 1998; MARTINS et al., 2001; MARTINS et
al., 2002; MARTINS et al., 2003).
Desta forma é importante determinar, com segurança, quais as concentrações de químicos que
se podem introduzir num ecossistema sem prejuízo para as comunidades do meio receptor.
Para tal é necessário proceder a uma série de testes de toxicidade. Existem dois objectivos
essenciais destes testes: a obtenção de dados sobre as propriedades toxicológicas de um
produto e o estabelecimento de limites de segurança à exposição de um novo produto.
Os mecanismos pelos quais os tóxicos podem afectar os organismos podem ser estudados de
duas formas principais: através da observação de características citológicas e fisiológicas numa
única população com o objectivo de construir uma tabela de vida (Figura 1) e/ou estudando as
consequências dos tóxicos na manutenção de várias populações dentro de uma comunidade
(NICOLAU et al., 2001).
O aumento da poluição ambiental e o contínuo desenvolvimento de novos químicos levou a
uma preocupação constante acerca dos potenciais efeitos, directos ou indirectos, destes
compostos na saúde humana (NICOLAU et al., 2004). Nos últimos anos foram realizadas
inúmeras pesquisas sobre a toxicidade de vários compostos importantes, mediante a utilização
de organismos teste em vários biotestes de toxicidade. A mais valia destes testes está
relacionada com a sua simplicidade e alto grau de reprodutibilidade. Por outro lado, os
organismos teste, utilizados para avaliar riscos e impactes ambientais, devem possuir uma
gama de características importantes, tais como: têm de ser eucarióticos; a sua biologia e
respostas gerais têm de ser bem conhecidas; o seu manuseamento laboratorial tem de ser

relativamente fácil; e têm que possuir um tempo de geração curto, importante para estudos de
efeitos tóxicos prolongados (NILSSON, 1989; NICOLAU et al., 2004). Os protozoários ciliados
preenchem todos estes requisitos. Para além disso, a comunidade de ciliados (que inclui muitas
vezes espécies sensíveis, resistentes ou de tolerância intermédia aos poluentes) são um meio
para identificar um possível impacte ecológico causado por poluentes antropogénicos
descarregados para as águas superficiais. Neste contexto, os protozoários ciliados tornaram-se
uma ferramenta valiosa para a detecção de mudanças ambientais e para avaliar o estado
trófico de um dado ecossistema aquático (MADONI, 2000; NICOLAU et al., 2004).
Figura 1: Diagrama conceptual do estudo realizado, focando vários parâmetros a serem tomados em
consideração para a realização dos testes de citotoxicidade com Tetrahymena pyriformis (adaptado de
NICOLAU et al., 2001).
Recentemente, o uso de protozoários ciliados em testes toxicológicos tem sido investigado e o
seu potencial em bioensaios padrão tem sido demonstrado em diversos ambientes. O
protozoário Tetrahymena pyriformis foi, por mais de quatro décadas, o organismo de eleição em
análises, na avaliação da qualidade proteica e na determinação de efeitos de várias
substâncias tóxicas (SAUVANT et al., 1999; STEFANIDOU et al., 1999; NICOLAU et al., 2001).
Tóxicos Objecto-teste Função-teste Níveis de Organização Biológica
Resposta
Etc.
Corantes têxteis azo
Protozoários
Dados
biológicos Reprodução Viabilidade
Energia e alimento
Anabolismo Catabolismo
Crescimento Predação
Manutenção
Locomoção
Factores Ecológicos
Características Citológicas e Fisiológicas

Existem vários trabalhos que relatam estudos realizados com T. pyriformis relativamente a
parâmetros como o crescimento, mortalidade, predação e ensaios bioquímicos, nomeadamente
adenosina-5-trifosfato (ATP), actividade da fosfatase ácida (ACP) e redução do 3-[4,5-dimetil-
tiazol-2-il]-2,5-difenil-tetrazóleo brometo (MTT), na presença de tóxicos. Estes ensaios são
avaliados usando culturas axénicas de T. pyriformis (DIAS et al., 1999; NICOLAU et al., 2001).
Martins et al.. (2001, 2002 e 2003) realizaram ensaios sobre a biodegradação dos corantes azo
de aplicação têxtil por fungos, nomeadamente o fungo filamentoso Phanerochaete
chrysosporium, de modo a estudar até que ponto aqueles são recalcitrantes. O trabalho que se
propõe neste projecto apresenta uma nova abordagem no estudo destes corantes,
nomeadamente da sua toxicidade em organismos modelo, como a T. pyriformis, um trabalho
inovador na medida em que não foi ainda determinada a toxicidade destes corantes em
qualquer outro organismo teste.
O objectivo deste trabalho é estudar a toxicidade de diferentes compostos químicos usados na
indústria têxtil – corantes azo – no protozoário T. pyriformis, através da realização de ensaios
toxicológicos fisiológicos, nomeadamente, crescimento, predação e morfometria. Pretende-se
avaliar se os compostos são tóxicos e, se o forem, em que concentrações surgem os efeitos
tóxicos e quais são esses efeitos, ou seja, que alterações provocam no organismo bioindicador.

2. TOXICOLOGIA E ECOTOXICOLOGIA – BREVE ABORDAGEM

2.1. Introdução
A noção de toxicidade existe desde o século XVI. O médico suíço Paracelsus escreveu em
1567 que “Todas as substâncias são venenosas. Não há nada que não o seja. A dose certa
diferencia um veneno de um remédio”. Dada esta longa história, podíamos esperar que
actualmente houvesse dados disponíveis para quase todos os químicos que se utilizam. No
entanto, isto é de longe verdadeiro. Existem actualmente cerca de 2500 químicos que são
importados para ou manufacturados na União Europeia em grandes quantidades (mais de 1000
toneladas/ano). No entanto, só estão disponíveis 14 % dos dados de toxicidade ambiental e
saúde humana, não existindo dados para 21 % dos químicos e apenas 3 % têm um conjunto de
dados completos (MITCHELL et al., 2002).
À escala global, o Instituto Worldwatch estima que não existe informação disponível sobre os
efeitos tóxicos de 79 % dos mais de 70000 químicos sintéticos. Se nos debruçarmos sobre o
caso da qualidade ambiental da água, a situação é bem pior (MITCHELL et al., 2002).
2.1. Modos de acção dos tóxicos
Quando se tenta prever a toxicidade, é vital diferenciar os tipos de efeito tóxico, existindo vários
modelos para descrever a toxicidade. A diferenciação fundamental é entre toxicidade reactiva
(ou específica) – associada a um mecanismo, como por exemplo uma reacção química ou a
inibição de um caminho metabólico - e não reactiva (não específica ou narcótica) – associada à
quantidade de tóxico que actua nas células.
Muitos investigadores propuseram uma classificação de tóxicos com base no seu modo de
acção, no entanto destaca-se a classificação de Verhaar et al.. (2000 in MITCHELL et al.,
2002). Estes investigadores usam uma combinação do modo de acção e a relação quantitativa
estrutura-actividade (QSAR) para definir quatro classes:
1. químicos inertes (de toxicidade básica);
2. químicos menos inertes (químicos com acidez na ligação do hidrogénio);
3. químicos reactivos (os que reagem não selectivamente com certas estruturas químicas
encontradas comummente nas células);
4. químicos activos específicos (os que exibem toxicidade devido a interacções com certos
receptores).
Em termos gerais, as concentrações com efeitos tóxicos para o segundo grupo são 5 a 10
vezes mais baixas do que para o primeiro grupo. No entanto, as concentrações para o terceiro

e quarto grupo são ainda mais baixas, ou seja, 10 e 104 vezes. Verhaar et al.. (2000 in
MITCHELL et al., 2002) validou este sistema de classificação através 176 compostos diferentes
e 964 dados independentes do Centro Europeu de Ecotoxicidade e Toxicologia de Químicos
(ECETOC).
2.3.Testes de toxicidade aquática
Uma análise sistemática da toxicidade de produtos químicos em diferentes águas naturais pode
permitir precisar a relação entre a toxicidade e parâmetros químicos. O estudo da toxicidade
realizado com diferentes organismos, permitirá definir uma cartografia da biodisponibilidade e
da toxicidade de compostos tóxicos (LE DÚ, 1993).
Assim, têm sido utilizados uma miríade de organismos modelo e tipos de testes de toxicidade. A
popularidade destes testes varia, ao longo do tempo, de acordo com a preferência dos
investigadores, das organizações, dos países, etc., estando continuamente a ser revistos,
melhorados ou até dando azo a novas criações (Tabela 1). No Reino Unido, o Direct Toxicity
Assessment (DTA) Demonstration Programme Steering Group recomendou que as formas de
vida mais simples devem ser utilizadas para os testes de ecotoxicidade, ou seja, bactérias,
plantas e invertebrados devem ser utilizados em vez dos vertebrados que, quando
seleccionados, o devem ser em números mínimos de indivíduos (MITCHELL et al., 2002).
Tabela 1: Vantagens e desvantagens dos testes de toxicidade (adaptada de MITCHELL et al., 2002).
Vantagens Desvantagens
- Holísticos
- Simples
- Baixo custo
- Baseados nos efeitos
- Envolvem condições controladas
- Não diagnósticos quando usados
sozinhos
- Não necessariamente representativos do
ambiente natural
- Não podem testar tudo
- Não podem considerar as condições
alteradas do meio
Os dados resultantes dos testes de toxicidade aquática apresentam uma grande variedade de
aplicações:
� decisões industriais para o desenvolvimento, manufactura e comercialização de um
produto;
� registo de produtos de forma a satisfazer os regulamentos de segurança;

� permissão de descargas de efluentes industriais ou municipais;
� avaliação do risco de impacte ecológico ou ambiental;
� prossecução ou defesa de actividades relacionadas com compostos químicos;
� análise da qualidade da água e protecção dos organismos aquáticos.
2.4. Relação quantitativa estrutura-actividade (QSAR) e relação
quantitativa actividade-propriedade (QPAR)
Uma QSAR é um modelo matemático que relaciona a actividade biológica (toxicidade) das
moléculas com as suas estruturas químicas. Modelos que descrevem uma relação entre a
actividade biológica de moléculas e as suas propriedades químicas ou físico-químicas são
QPAR. Quando se constrói uma relação QSAR/QPAR, o primeiro passo é usar dados de
toxicidade para estabelecer uma relação matemática entre a toxicidade química de substâncias
não testadas.
As aplicações dos QSAR/QPAR são, entre outras, estimar:
� as propriedades físico-químicas com relevância biológica como o coeficiente de partição
água-octanol, solubilidade em água e biodegradação;
� e prever pontos de ecotoxicidade;
� o potencial de bioacumulação de químicos em organismos aquáticos.
Muitos autores usaram os modelos QPAR para a previsão de toxicidade, nomeadamente, o
coeficiente água – octanol, hidrofobicidade, solublidade aquática, etc. (MITCHELL et al., 2002).
As QSAR têm sido utilizadas em ecotoxicologia como ferramentas úteis para a previsão de
toxicidade. Sabendo que os compostos químicos exibem toxicidade segundo vários
mecanismos de acção tóxica (por exemplo, narcose), o desenvolvimento e a aplicação de
QSAR evoluiu de uma abordagem meramente química para uma relacionada com os
mecanismos de acção dos compostos. A vantagem das QSAR baseadas em mecanismos é a
sua elevada predição de toxicidade. No entanto, para utilizar esta ferramenta, os mecanismos
de actuação dos compostos têm que estar bem identificados, se bem que a correcta
determinação destes mecanismos de acção não é sempre fácil, pois a toxicidade é um efeito
complicado e muitos passos envolvidos neste são pouco conhecidos e caracterizados (REN,
2003).

2.5. Organismos teste
Uma variedade de tipos de organismos teste, de vários níveis tróficos e muitos géneros
diferentes, são utilizados nos testes de toxicidade aquática. Os mais utilizados incluem plantas
(algas), animais invertebrados (crustáceos) e espécies de peixes de forma a determinar os
efeitos tóxicos nos vários níveis tróficos. Os testes realizados com microrganismos também se
têm tornado mais comuns, talvez com o objectivo de estudar as culturas das lamas activadas
das estações de tratamento de águas residuais. No geral, existem alguns critérios utilizados
para escolher o organismo: deve ser sensível ao tóxico, deve representar as populações
indígenas que são principalmente ecologicamente importantes, deve ser de fácil aquisição e de
manutenção laboratorial prática (LARSEN et al., 1997).
2.6. Métodos de testes tradicionais
2.6.1. Métodos agudos ou crónicos Os testes agudos são normalmente realizados antes dos testes crónicos, sobretudo quando se
quer avaliar a mortalidade ou a sobrevivência. A LC50 (concentração letal 50, ou seja, a
concentração que causa 50 % de mortalidade) é a mais frequentemente utilizada. A toxicidade
aguda é importante para medir a letalidade do tóxico, estimando assim a sua concentração e
potência.
Os testes crónicos de toxicidade são desenhados para avaliar os efeitos da longa exposição
aos contaminantes. O crescimento (peso e comprimento), número normal de embriões,
anormalidades morfológicas e número de descendentes são efeitos sub-letais típicos utilizados
nos testes crónicos. O benefício destes testes é que possibilitam uma acção de remediação
antes da mortalidade (MITCHELL et al., 2002).
Os testes de toxicidade sub-letais que não têm como finalidade a mortalidade, são muito
importantes para determinar a toxicidade de uma ou várias substâncias. Podem ser divididos
em três categorias:
� testes bioquímicos e fisiológicos (inibição respiratória ou enzimática);
� testes comportamentais (locomoção, territorialismo, agressão);
� testes histológicos (alterações em tecidos dos organismos estudados);
2.6.2. Testes modificados, rápidos ou microbiotestes. O tempo e o custo que envolviam os testes tradicionais de toxicidade levaram ao
desenvolvimento de testes alternativos. Como já foi mencionado anteriormente, a preocupação

com o bem-estar dos organismos teste foi também considerada nesta pesquisa. Não obstante,
os testes crónicos e agudos são relativamente dispendiosos e requerem uma contínua
manutenção dos organismos testes (MITCHELL et al., 2002). Este facto levou ao
desenvolvimento de uma gama de rápidos microbiotestes de ecotoxicidade dos quais se
destacam:
� Microtox: teste que mede as mudanças na produção de luz por bactérias luminescentes
(Vibrio fischeri) expostas a um efluente ou tóxico, sendo o tempo requerido para este
teste menos de 1 hora;
� Algaltoxkit: teste de inibição de crescimento de 72 horas, baseado na alga verde
Selenastrum capricornutum;
� Daphtoxkit: bioensaio de 24 horas de LC50 realizado em microplaca, que utiliza larvas
de crustáceos de água doce Daphnia magna e Daphnia pulex, cujo tempo de incubação
é de 24 horas.

3. TESTES DE TOXICIDADE COM PROTOZOÁRIOS

3.1. Introdução
Ultimamente, a presença de tóxicos nos ambientes aquáticos tornou-se uma ocorrência comum.
Estes são provenientes principalmente de esgotos de indústrias, que reduzem a eficiência do
tratamento biológico das águas residuais devido a fenómenos de intoxicação (NICOLAU et al.,
2001).
Diferentes organismos podem apresentar sensibilidades variáveis de acordo com os químicos;
e a sensibilidade de uma certa espécie pode variar de acordo com o tipo de composto tóxico.
Isto significa que um modelo de acção de um conjunto de químicos encontrados para uma dada
espécie pode não ser aplicado a outra espécie diferente. No entanto, a toxicidade de um
químico ou de um conjunto de químicos, no que se refere à deplecção de ambos os organismos
e espécies, pode ser demonstrado testando a comunidade da microfauna que habita as lamas
activadas (NICOLAU et al., 2001).
Os protozoários ciliados muitas vezes podem atingir densidades de 107 células/ L no tanque de
arejamento. Apresentam um papel importante no processo de purificação das águas ao
remover, através da predação, a maioria das bactérias dispersas, que causariam grande
turbidez no efluente final. São muito sensíveis às variações ambientais e foi já descoberto que
mudanças na comunidade de protozoários podem afectar toda a cadeia alimentar destes
ecossistemas artificiais, afectando também a performance biológica do tratamento de águas
residuais (NICOLAU et al., 2001).
Estes protozoários, pelo que foi anteriormente relatado e porque: 1) representam um
componente básico das comunidades microplânctónicas e microbênticas de ambientes
aquáticos, 2) apresentam ampla distribuição, 3) e são de importância ecológica significativa, ou
seja, desempenham funções chave nas cadeias tróficas aquáticas, onde medeiam o fluxo de
substâncias biológicas e de energia de um nível trófico para o seguinte; são importantes
organismos indicadores assinalando a deterioração dos ecossistemas aquáticos (LARSEN et
al., 1997; MADONI, 2000; NICOLAU et al., 2004).
3.2. Os ciliados como organismos teste Durante muito tempo, os protozoários foram extensivamente estudados, descritos e
classificados por protozoólogos. Os protozoários são células eucarióticas presentes em quase
todos os ambientes aquáticos e terrestres. O seu comportamento normal no meio ambiente
pode estar relacionado com a presença de poluentes e com a qualidade do ar, solo e água.
Este facto levou muitos toxicologistas e ecotoxicologistas a usar os protozoários como modelos

animais para o estudo de compostos xenobióticos e para a avaliação dos seus riscos para a
saúde pública (LARSEN et al., 1997; NICOLAU et al., 1999; SAUVANT et al., 1999; DIAS et al.,
2003).
Entre as várias espécies de protozoários seleccionadas para estudos ecotoxicológicos, os
ciliados são os mais usados. A tabela que se segue refere-se aos principais ciliados utilizados
em estudos de toxicidade:
Tabela 2: Organismos principais dos quais foram isoladas estirpes e para as quais testes toxicológicos
especiais foram desenvolvidos (adaptado de SAUVANT et al., 1999).
Espécies de protozoários Testes toxicológicos
Colpidium campylum Protozoários utilizados no estudo dos efeitos e interacções entre poluentes aquáticos orgânicos e não orgânicos
Euplotes vannus Euplotes harpae Paramecium putrinum Oxytricha Spirostomum ambigum Entosiphon sulcatum Stentor sp. Paramecium sp. Tetrahymena vorax Tetrahymena thermophila Tetrahymena pigmentosa
Organismos testes para a avaliação da toxicidade de iões de metais pesados em substâncias orgânicas
Tetrahymena pyriformis O mais comum modelo protista usado em estudos toxicológicos
A Tetrahymena pyriformis tem sido usada nas últimas décadas por vários estudos toxicológicos
e ecotoxicológicos, que a validam como um organismo teste ideal e como um complemento ou
alternativa a modelos animais mais complexos na investigação toxicológica (NILSSON, 1981;
NICOLAU et al., 1999; SAUVANT et al., 1999; NICOLAU et al., 2001; DIAS & LIMA, 2002;
NICOLAU et al., 2004). Além disso, foi o primeiro protozoário a ser cultivado axenicamente, ou
seja, num meio definido, livre de bactérias e outros organismos, sendo mantido em culturas
celulares apropriadas à adição de um composto que constitui, em princípio, a única alteração
nas condições de cultura. (SAUVANT et al., 1999; STEFANIDOU et al., 1999 ; NICOLAU et al.,
2001; NICOLAU et al., 2004).

Para além do uso de outros protozoários como organismos teste, a Tetrahymena pyriformis é
um dos protozoários ciliados mais utilizados para avaliar os efeitos citotóxicos de compostos
xenobióticos (SAUVANT et al., 1999). Este ciliado foi o organismo teste seleccionado para a
avaliação da qualidade das proteínas, bem como para a determinação do efeito de várias
substâncias (carcinógenos, insecticidas, fungicidas, micotoxinas, químicos orgânicos, metais
pesados, drogas farmacêuticas, etc.). A biologia desta célula eucariótica é bem conhecida,
sendo caracterizada por um curto tempo de geração que fez dela um modelo a utilizar nos
ensaios in vitro (SAUVANT et al., 1995; BONNET et al., 2005).
3.3. Tetrahymena pyriformis: uma ferramenta na toxicologia
3.3.1. Cultivo in vitro
No seu ambiente natural, Tetrahymena pyriformis alimenta-se de bactérias, mas pode ser
mantida num meio artifical, axénico e quimicamente definido (Figura 2).
Figura 2: Tetrahymena pyriformis retirada de uma cultura axénica em meio de cultura PPY (x100).
Foram já propostos meios de cultura muito similares para este organismo, mas as diferenças
entre eles podem influenciar os resultados obtidos. A temperatura das culturas pode variar entre
25 ºC a 30 ºC. A maioria dos ensaios foi realizada a uma temperatura óptima de 28 °C, com
culturas axénicas e sem agitação. Na maioria dos ensaios, as culturas de Tetrahymena

pyriformis encontravam-se em fase de crescimento exponencial, com densidades de células
entre 4 e 5x104 células/mL. De entre os muitos meios de cultura propostos, a maioria dos
investigadores utiliza meio com proteose–peptona e extracto de levedura, suplementado com
sais inorgânicos, conhecido por meio PPYS. As proporções de proteose-peptona e extracto de
levedura podem variar de 0 a 2 %.
A constituição do meio de crescimento é muito importante, uma vez que vários trabalhos
demonstram que o estado fisiológico da Tetrahymena pyriformis e o efeito tóxico de compostos
xenobióticos estão relacionados com a composição do meio de cultura, o pH e, especialmente,
com a quantidade de matéria orgânica presente (SAUVANT et al., 1999).
Para além disso, a simplicidade do seu manuseamento laboratorial é essencial para torná-las
uma alternativa viável à utilização de outros animais aquáticos, na avaliação da toxicidade de
poluentes (LARSEN et al., 1997; NICOLAU et al., 1999; SAUVANT et al., 1999; DIAS et al.,
2003).
3.3.2. Ensaios para avaliar o efeito tóxico de compostos xenobióticos
Hoje em dia, os testes biológicos são bastante desenvolvidos, porque a prática laboratorial
requer ensaios rápidos, simples e sensíveis para avaliar a toxicidade dos poluentes ambientais
(SAUVANT et al., 1995). Em toxicologia, os bioensaios agudos normalmente medem a
mortalidade relativa de concentrações crescentes de xenobióticos e condições de exposição
específicas. Até à data, a evolução da metodologia dos bioensaios é baseada em respostas
sub-letais do modelo celular.
Em Tetrahymena pyriformis, a avaliação dos efeitos citotóxicos de compostos xenobióticos
pode ser realizada utilizando vários parâmetros e ensaios metabólicos, fisiológicos ou
bioquímicos toxicológicos.
3.3.3. Os ensaios fisiológicos toxicológicos
A taxa de crescimento e as alterações morfológicas têm sido utilizadas há algumas décadas. A
diminuição do crescimento populacional é um método frequentemente usado para avaliar a
toxicidade sub-letal de compostos orgânicos e inorgânicos, que não requer especial experiência
técnica. Outros parâmetros como a mobilidade celular, padrões natatórios, taxa de fagocitose e
a análise do citoesqueleto podem ser estudados e foram propostos para a determinação do
estado fisiológico e energético de Tetrahymena pyriformis, quando em contacto com diversos
poluentes.

a. Densidade celular e taxa de crescimento
Em condições óptimas, o crescimento de Tetrahymena pyriformis é caracterizado por uma fase
de crescimento logarítmica – fase log -, uma fase de crescimento pré-estacionária, e uma fase
estacionária. Na fase logarítmica, que pode durar desde algumas horas a dois dias,
dependendo do inóculo, a densidade celular aumenta logaritmicamente e o tempo de geração
ocorre entre as 3 e as 7 horas. Na fase pré-estacionária o crescimento diminui por algumas
gerações antes de entrar na última fase, a estacionária.
Durante estas fases, o pH aumenta de 6,7 para 8 em meio PPYS. Este comportamento pode
ser explorado para testes de toxicidade. Após a exposição a compostos xenobióticos, a
densidade celular e a taxa de crescimento são frequentemente propostas para especificar o
impacto tóxico das substâncias testadas; as contagens celulares são feitas automaticamente
por um contador electrónico de partículas, microscopicamente por um hemocitómetro ou por
medições de densidade óptica com espectrofotómetro. Estas contagens permitem a realização
de curvas de crescimento, que servem para calcular o tempo de geração das populações, que
depois do contacto com xenobióticos aumenta.
A densidade celular é também usada para a comparação do potencial tóxico de substâncias.
Os resultados são fornecidos em valores de IC50 (a concentração efectiva que reduz a
densidade celular a 50 % face à verificada nos controlos). É ainda de salientar que a medição
indirecta da densidade óptica é equivalente à contagem directa electrónica de partículas
quando se determina a densidade celular de culturas de Tetrahymena pyriformis, excepto nos
testes de químicos coloridos ou insolúveis (SAUVANT et al., 1999).
b. Mudanças comportamentais
Em condições normais, Tetrahymena pyriformis é caracterizada por grande mobilidade e
actividade. Na presença de compostos xenobióticos o organismo ajusta o seu comportamento
aos factores tóxicos stressantes. Assim, a sua locomoção e a sua mobilidade foram descritas
como dependentes do estado de saúde das células.
A fagocitose é um fenómeno normal na Tetrahymena pyriformis. A sua taxa é influenciada pela
temperatura, pelo pH e pela presença de partículas sólidas. Vários compostos podem interferir

inibindo ou estimulando a predação. Além disso, alguns xenobióticos podem acumular-se,
aumentando a sua toxicidade (SAUVANT et al., 1999).
O protozoário Tetrahymena pyriformis tem sido muito usado como um modelo experimental
para estudos toxicológicos. Foi já confirmado que a endocitose (fagocitose e pinocitose) ocorre
através da formação de vacúolos. A fagocitose nos protozoários é uma função celular clássica,
reflectindo o nível de energia do protozoário e representa o principal meio de defesa em
organismos que não possuem a capacidades de fabricar anticorpos. A predação foi já utilizada
em vários estudos, nomeadamente com substâncias psicotrópicas (drogas) (STEFANIDOU et
al., 1999).
c. Crescimento, mortalidade e viabilidade celulares
Os ensaios de crescimento são os mais frequentemente usados para determinar a toxicidade
sub-letal. Um ensaio de mortalidade/crescimento, através da utilização de observação
microscópica das alterações a baixa ampliação, é uma técnica de fácil reprodução e não requer
experiência técnica ou despesas operacionais (NICOLAU et al., 1999).
No entanto, estes ensaios apresentam algumas limitações, que os tornam inadequados para
alguns compostos tóxicos. O reconhecimento de células mortas torna-se muitas vezes
ambíguo, uma vez que células não móveis bem como as que possuem formas alteradas podem
ser contadas como células mortas. As observações ao microscópio de luz podem subestimar o
verdadeiro número de células viáveis, ou seja, uma contagem falsa e negativa, uma vez que
muitas células anormais não se encontram de facto mortas. Estas podem recuperar a sua forma
normal após a remoção do tóxico, crescendo normalmente nos ensaios de recuperação
(NICOLAU et al., 2001; DIAS & LIMA, 2002).
A citotoxicidade e morte celular têm sido investigadas através da utilização de marcadores
retidos pelas células vivas ou libertados de células lisadas. Os derivados fluorogénicos são
substratos para esterases, coram células vivas, e são amplamente utilizados como marcadores
citoplasmáticos e como sondas de viabilidade.
A calceína AM é um éster lipofílico não fluorescente que se difunde passivamente nas células e
é transformado em calceína fluorescente, por esterases intracelulares. Uma vez hidrolizada, a
calceína livre insolúvel nos lípidos e com alta carga negativa, é aprisionada dentro de células
com membranas intactas e torna-se verde fluorescente a um comprimento de onda de 530 nm.
Um segundo corante é então adicionado para corar as células mortas. EthD-1 (homodímero de

etídio) é um fluorocromo impermeável que passa através de membranas não funcionais e
estabelece uma ligação forte com os ácidos nucleicos de cadeia dupla, emitindo uma
fluorescência vermelha a um comprimento de onda de 617 nm. As células viáveis verdes e
células não viáveis vermelhas podem ser detectadas por microscopia de fluorescência (Figura
3). A sua quantificação e análise de dados podem ser feitas através de várias técnicas como:
microfluorometria, citometria de fluxo, microscopia laser e confocal (NICOLAU et al., 2001; DIAS
et al., 2002).
Figura 3: Células de Tetrahymena pyriformis viáveis e não viáveis, utilizando a calceína/AM e EthD-1. As
duas células não viáveis mostram núcleos corados de vermelho (retirada de NICOLAU et al., 2001).
Este novo método para avaliar a citotoxicidade foi utilizado como alternativa à contagem directa
de protozoários viáveis ao microscópio de luz. Os resultados obtidos com este método e com o
da contagem directa são semelhantes, especialmente no que diz respeito ao aumento do tempo
de exposição (NICOLAU et al., 2001).
3.3.4. Os ensaios bioquímicos toxicológicos
Ao longo de décadas, a mobilidade celular e o aspecto das células observadas ao microscópio
de luz visível têm sido utilizados como indicadores de viabilidade. No entanto, nos últimos anos,
vários testes têm sido desenvolvidos ou adaptados a estas células, nomeadamente: conteúdo
em adenosina-5-trifosfato (ATP), a actividade da fosfatase ácida (ACP), e a redução do 3-[4,5-
dimetil-tiazol-2-il]-2,5-difenil-tetrazóleo brometo (MTT), e mais recentemente, o teste bioquímico
denominado MWFP (Multiwell filter plates) para avaliar a viabilidade das células (DAYEH et al.,
2005).

Gikas et al. (1993 in NICOLAU et al., 2001; NICOLAU et al., 2004) utilizaram ensaios que
detectaram a quantidade de ATP para caracterizar a viabilidade da biomassa; Dowhanick et al.
(1994 in NICOLAU et al., 2001; NICOLAU et al., 2004); Gamborg et al. (1994 in NICOLAU et al.,
2001) utilizaram o teste da ATP para detectar microrganismos na indústria da cerveja e da
alimentação e Nicolau et al. (2004) utilizaram o teste da ATP para determinar o estado
energético de culturas de protozoários submetidos a diferentes tóxicos. Estes investigadores,
entre outros, assumiram a possibilidade de utilizar a concentração de ATP para detectar as
células viáveis de uma certa espécie.
Muitos trabalhos foram realizados utilizando a actividade da ACP de outras hidrolases para
detectar a actividade digestiva dos protozoários, uma vez que há uma íntima relação entre a
função lisossómica normal e a digestão intracelular em Tetrahymena e outros ciliados. No
entanto, apenas alguns autores sugeriram o teste da actividade da ACP para a avaliação da
toxicidade. Entre eles destaca-se a utilização da actividade da ACP por Nicolau et al. (2004),
como um indicador do estado metabólico das culturas, principalmente da função digestiva
intracelular.
Os ensaios toxicológicos convencionais são muitas vezes lentos e trabalhosos, e tornam-se
impraticáveis quando muitos compostos ou concentrações estão a ser testadas. Isto levou a um
grande interesse por ensaios colorimétricos ou fluorimétricos que podem ser miniaturizados em
microplacas de 96 poços e avaliados com espectofotómetro de microplacas ELISA. Neste
aspecto, o ensaio MTT e a utilização dos protozoários é uma excelente opção: permite
simultaneamente testes de diferentes tóxicos a várias concentrações, é rápido e não requer
experiência técnica nem despensas operacionais como outros bioensaios estandardizados
(DIAS et al., 1999). O ensaio colorimétrico MTT, baseado na redução do corante tetrazólio tem
sido extensivamente utilizado na avaliação in vitro da proliferação celular e citotoxicidade, bem
como no estudo de aditivos, micotoxinas e fármacos contra o cancro (DIAS et al., 1999;
NICOLAU et al., 2001; ZILBERG & SINAI 2006). Trata-se de um método quantitativo que
consiste na redução metabólica do corante tetrazólio, um grupo de compostos quaternários de
amónia solúveis em água, por dehidrogenases de células viáveis imobilizadas ou suspensas
num meio. O resultado deste ensaio é a produção de cristais insolúveis de formazano em água.
Os cristais formados podem ser observados ao microscópio no citoplasma celular ou extraídos
e dissolvidos com solventes orgânicos, como o dimetil sulfóxido (DMSO), sendo assim possível
a sua quantificação com um espectrofotómetro (DIAS et al., 1999; NICOLAU et al., 2001;
ZILBERG & SINAI, 2006).

Os ensaios bioquímicos colorimétricos MTT, ATP e ACP que utilizam os protozoários parecem
ser excelentes opções porque proporcionam testes simultâneos com diferentes tóxicos a várias
concentrações e são fáceis e rápidos de realizar (NICOLAU et al., 2001).

4. OS PROTOZOÁRIOS CILIADOS: BREVE ABORDAGEM DA SUA MORFOLOGIA, FISIOLOGIA E TAXONOMIA.

4.1. Introdução
A Tetrahymena pyriformis é caracterizada por possuir um ciclo de vida muito curto, o que
permite a sua fácil cultura em condições laboratoriais apropriadas. Os efeitos tóxicos de
substâncias também podem ser testados em várias gerações. Todas estas propriedades podem
explicar a utilização deste modelo biológico em numerosos estudos fisiológicos e bioquímicos
bem como em pesquisas farmacológicas e toxicológicas.
4.2. Morfologia, fisiologia e taxonomia Tetrahymena pyriformis (Ehremberg, 1830) é um ciliado de água doce pertencente ao Filo
Ciliophora, à classe Oligohymenophorea, subclasse Hymenostomia, ordem Hymenostomatida,
subordem Tetrahymenina.
O seu corpo tem um comprimento de aproximadamente 50-60 µm e uma largura de 30 µm e
tem forma de pêra, característica que lhe deu o nome do restritivo específico (Figura 4). São
possíveis várias modificações da sua forma, nomeadamente em culturas antigas ou sob stress.
A superfície da Tetrahymena pyriformis é coberta por 400 a 600 cílios ancorados par a par em
cada corpo basal que, por sua vez, está ancorado a um citoesqueleto cortical (DIAS, 1998).
Debaixo da membrana plasmática que cobre todo o organismo incluindo os cílios, há um
sistema de alvéolos achatados que se encontram agrupados em pares na base de cada cílio.
Uma densa camada de citoplasma chamada epiplasma encontra-se debaixo destes alvéolos,
que são localizados na zona de transição do corpo basal para os cílios.
O aparelho oral, localizado perto da extremidade anterior da célula, consiste em quatro
estruturas ciliares: membrana ondulatória (um) nas margens direita e posterior da cavidade
bucal, e as três membranelas da zona adoral que ficam fora da cavidade (Figura 4).
A parede da cavidade bucal em forma de funil é suportada por uma série de fitas peliculares e
por um intrincado sistema de microtúbulos originados a partir dos corpos basais. Estas fitas,
unindo as membranelas, contribuem para a formação de uma fibra citofaríngea pós-oral que se
entende ao longo da citofaringe. O citóstoma fica na base da cavidade bucal e a estrutura de
forma cónica da citofaringe abre-se no vacúolo digestivo abaixo do citóstoma.
Na zona da citofaringe, a transição da estrutura da película constituída por três camadas para a
membrana celular que envolve o vacúolo digestivo é evidente. Quando o vacúolo digestivo está
cheio, deixa a região oral e um novo vacúolo é formado. A eficiência e a taxa de formação de
vacúolos digestivos depende do ciclo celular, estado fisiológico da célula e factores ambientais

tais como pH, temperatura, etc. Foi já mostrado que muitas substâncias orgânicas e inorgânicas
podem interferir com a formação de vacúolos digestivos, daí que a capacidade de formação de
novos vacúolos é um bom indicador de condições tóxicas.
Figura 4:
- Diagrama A: Morfologia da Tetrahymena pyriformis: cp – citoprocto; cv – vacuolo contráctil; fv – vacúolo
digestivo; mn - macronúcleo; n - núcleo; ki – corpo basal; um e m – membrana ondulatória e
membranelas do aparelho oral (oa).
- Diagrama B: Fotografia de contraste de fase de uma célula de Tetrahymena pyriformis fixada com
vapor de 1 % de tetróxido de ósmio (x600).
As fases da digestão são similares a outros ciliados. A última fase é a egestão de material não
digerido, que ocorre no citopígeo. Esta é uma estrutura permanente da célula que se encontra
entre os alvéolos na superfície ventral posterior da célula. Durante a egestão, a película que se
encontra nas margens do citopígeo e a membrana do vacúolo fundem-se com a membrana
celular. Quando a defecação termina, o citopígeo fecha e o vacúolo vazio colapsa em pequenas
vesículas, cujas membranas podem ser recicladas para se tornarem membranas de outro
vacúolo digestivo.
Uma importante característica é que o vacúolo contráctil mantém constante o volume celular
eliminando fluído a uma taxa igual à entrada passiva de água numa célula hiperosmótica.
A B

Alguns iões (como o sódio) são transportados activamente através da membrana e expelidos, e
as mitocôndrias são de facto associadas a esta estrutura.
A Tetrahymena pyriformis é uma importante espécie amicronucleada, ou seja, tem apenas um
macronúcleo esférico e polipóide, com cerca de 10 µm de diâmetro. Contêm densos grânulos
de cromatina e muitos nucléolos em forma de copo. Os grânulos de cromatina, compostos por
DNA e proteínas, são visíveis em fases celulares. Na interfase, a cromatina é finamente
granulada e distribuída ao longo do macronúcleo; nesta fase, é muito activa na síntese, e
aparece escura durante a observação microscópica de contraste de fase. Durante o ciclo
celular, ocorrem mudanças regulares relacionadas com a duplicação celular e morfogénese, na
estrutura dos agregados de cromatina. A configuração da cromatina muda para a forma
condensada na fase estacionária e durante o esfomeamento; é simultânea com as mudanças
na organização nucleolar.
Na Tetrahymena, os usuais organelos citoplasmáticos são mitocôndrias, retículo
endoplasmático, complexo de Golgi, ribossomas, peroxissomas e lisossomas. As suas
estruturas espelham o estado fisiológico da célula. Durante o período de esfomeamento, na
fase estacionária, ou em reposta a certos tratamentos, ocorrem mudanças estruturais nas
células: as mitocôndrias e os peroxissomas tornam-se densos, aparecem gotas lipídicas,
vacúolos autofágicos, partículas de glicogénio e muitos corpos de inclusão, reflectindo
mudanças de metabolismo.
Perante este breve descrição, fica claro que muitas das características estruturais dos
organelos são marcadores importantes do estado físiológico da Tetrahymena e podem ser
usados para criar modelos simples do estudo da toxicidade de muitas substâncias (SAUVANT
et al., 1999).

5. CORANTES

5.1. Introdução
Os corantes têxteis azo são caracterizados pela presença de um ou mais grupos azo (-N=N-).
São o maior e mais versátil grupo de corantes, e mais de metade dos corantes anualmente
produzidos (estimados globalmente em 1994 em cerca de 1 milhão de toneladas) são corantes
azo. Presumivelmente, mais de 2000 corantes azo diferentes são utilizados para tingir vários
materiais tais como têxteis, couro, plásticos, cosméticos e alimentos (STOLZ, 2001; NOVOTNÝ
et al., 2006). Amplamente usados na indústria têxtil, a sua aplicação ascende até aos 70 % de
todos os corantes utilizados. Durante o processo de coloração dos têxteis, 2 a 50 % dos
corantes aplicados podem ser perdidos nas águas residuais industriais que posteriormente são
libertadas no meio ambiente (NOVOTNÝ et al., 2006). Existe apenas o registo de um único
grupo azo num produto natural (4,4′-dihidroxiazobenzeno); todos os outros produzidos
industrialmente são considerados compostos xenobióticos, resistindo à biodegradação (STOLZ,
2001).
A aguda toxicidade de corantes azo medida pelos critérios da União Europeia para a
classificação de substância perigosas é baixa, sendo os valores de LD50 de 250-2000 mg/Kg.
Alguns corantes azobásicos, ácidos e directos foram classificados como sendo muito tóxicos ou
apenas tóxicos para peixes, crustáceos, algas e bactérias, enquanto que os corantes azo
reactivos têm concentrações efectivas muito elevadas (níveis de CE>100 mg/L) e assim não
são considerados tóxicos para organismos aquáticos. Os corantes azo não iónicos são
geralmente classificados como tóxicos ou potencialmente tóxicos. As algas são sensíveis aos
corantes, mas o efeito inibitório está muitas vezes relacionado com a inibição da luz quando as
concentrações destes corantes são muito elevadas, em vez de um efeito directo dos corantes
(NOVOTNÝ et al., 2006).
A síntese de corantes azo está bem estabelecida e anualmente são desenvolvidos novos
corantes. Os efluentes coloridos de indústrias de coloração de tecidos e de indústrias têxteis
causam poluição visual no ambiente e são perigosos para a vida, uma vez que são resistentes
a tratamento biológico.
São considerados compostos xenobióticos recalcitrantes devido à presença de uma ligação
N=N e outros possíveis grupos que não são facilmente biodegradáveis, por exemplo o grupo
sulfónico (SO3H). Os corantes azo são tóxicos e podem tornar-se perigosos para o ambiente
através da formação de aminas aromáticas (anilinas), que são carcinogénicas ou mutagénicas.
As anilinas são formadas pela clivagem da ligação azo que, usualmente, ocorre em condições
de anaerobiose por muitas estirpes de bactérias (MARTINS et al., 2001).

Os organismos vivos normalmente não sintetizam ligações azo e grupos sulfónicos aromáticos,
sendo o conhecimento limitado sobre a biodegradação destes grupos.
Para proteger o ambiente, são necessárias estratégias de aumentem a gama de compostos
xenobióticos degradados nos tratamentos de águas residuais ou as capacidades de
degradação destes compostos por microrganismos. Tendo isto em mente, os corantes azo que
foram utilizados neste estudo incluem grupos ácidos reactivos que providenciam ligações
covalentes com as fibras têxteis, evitando a perda de grandes quantidades de corantes após o
processo de tinturaria e grupos bioacessíveis ao sistema enzimático de fungos – grupos
guaiacol e siringol (MARTINS et al., 2002).
5.2. Principais fontes de poluição no sector têxtil e os seus
requerimentos ambientais
Na década de 1970, foram relatados casos de trabalhadores responsáveis pelo controle de
qualidade de artigos têxteis, com problemas de lacrimação, tosse e reacções alérgicas da pele.
Pessoas que utilizavam roupas que não necessitavam de passagem a ferro, apareciam com
infecções na pele. No entanto, naquela altura, estes incidentes eram ignorados. Na década de
1980, principalmente no final, com o avanço da ciência e da tecnologia e com o aumento da
preocupação ambiental dos cidadãos, os relatórios acerca do potencial efeito nefasto de certos
produtos têxteis receberam grande atenção. Na indústria têxtil moderna, os processos de
coloração de têxteis e a sua rectificação são processos tipicamente químicos e a poluição
decorrente destes é uma realidade.
Generalizando, as substâncias nocivas presentes nos têxteis e produtos de vestuário incluem
as seguintes categorias:
� formaldeído, branqueador fluorescente e amaciador;
� resíduos de pesticidas; antisépticos, inibidores de bolor nas fibras de lã e algodão
(como, por exemplo, o pentaclorofenol);
� resíduos de metais pesados;
� resíduos de monómeros de corantes azo e formaldeído nas fibras químicas;
� resíduos de pesticidas e fertilizantes utilizados nas culturas de algodão.
Desde a década de 1990, muitos países adoptaram medidas e requisitos ambientais, para
restringir a utilização de químicos nefastos na produção de têxteis e vestuário. Alguns destes
requisitos são impostos através de leis e de regulamentações (Policy Research Center for
Environment and Economy, 1999).

5.3. Legislação relativa aos corantes azo
Como foi referido anteriormente, muitos países adoptaram legislação ambiental e requerimentos
restringindo o uso de químicos perigosos na produção de têxteis e vestuário. Uma das leis mais
conhecidas é a Segunda Emenda ao Acto de Protecção do Consumidor elaborada pelo governo
alemão em 1994, proibindo a utilização de corantes azo. A Quinta Emenda a este acto foi
aprovada em Abril de 1997 e, desde então, muitos outros países europeus seguiram os passos
da Alemanha. Outros países europeus, tais como a Suécia, a França e a Dinamarca formularam
legislação respeitante aos corantes azo (Policy Research Center for Environment and Economy,
1999).
Tendo em consideração que os corantes azo são muito utilizados nos processos de coloração
de têxteis e couro, é de salientar a sua capacidade de libertar algumas arilaminas que podem
causar cancro. A União Europeia, restringiu estas arilaminas nos produtos comercializados na
Europa. Assim, adoptou uma legislação relativa a corantes azo em Setembro de 2002, que se
aplicou a todos os Estados Membros a partir de 11 de Setembro de 2003. A Directiva
2002/61/EC, cobre 22 aminas e é relevante para artigos têxteis e artigos de couro que estejam
em contacto directo com a pele. Esta directiva reformulada pela 2004/21/ EC foi implementada
em Portugal pelo Decreto Lei n.º 208/2003.
A legislação portuguesa é uma transposição directa da legislação europeia, o que significa que
se rege pelos mesmos requisitos. Esta legislação proíbe a utilização de corantes azo no fabrico
de artigos têxteis ou de pele, que entrem em contacto prolongado com a pele ou com a boca. A
Directiva estabelece ainda que os produtos referidos não podem conter as 22 aminas listadas
na legislação, numa concentração acima dos 30 ppm. Se os artigos forem feitos de fibras
recicladas não podem conter mais do que 70 ppm.
5.4. Corantes azo utilizados
Os corantes têxteis azo caracterizam-se pela presença de pelo menos um grupo cromóforo azo,
mas podem conter dois, três ou, mais raramente, quatro grupos azo (-N=N-). Os grupos de
azoto têm hibridização sp2 e estão ligados normalmente a anéis aromáticos. Os corantes azo,
cujos grupos substituintes são anéis benzénicos, denominam-se homocíclicos. Se contêm um
ou mais anéis heterocíclicos, os corantes azo designam-se heterocíclicos.
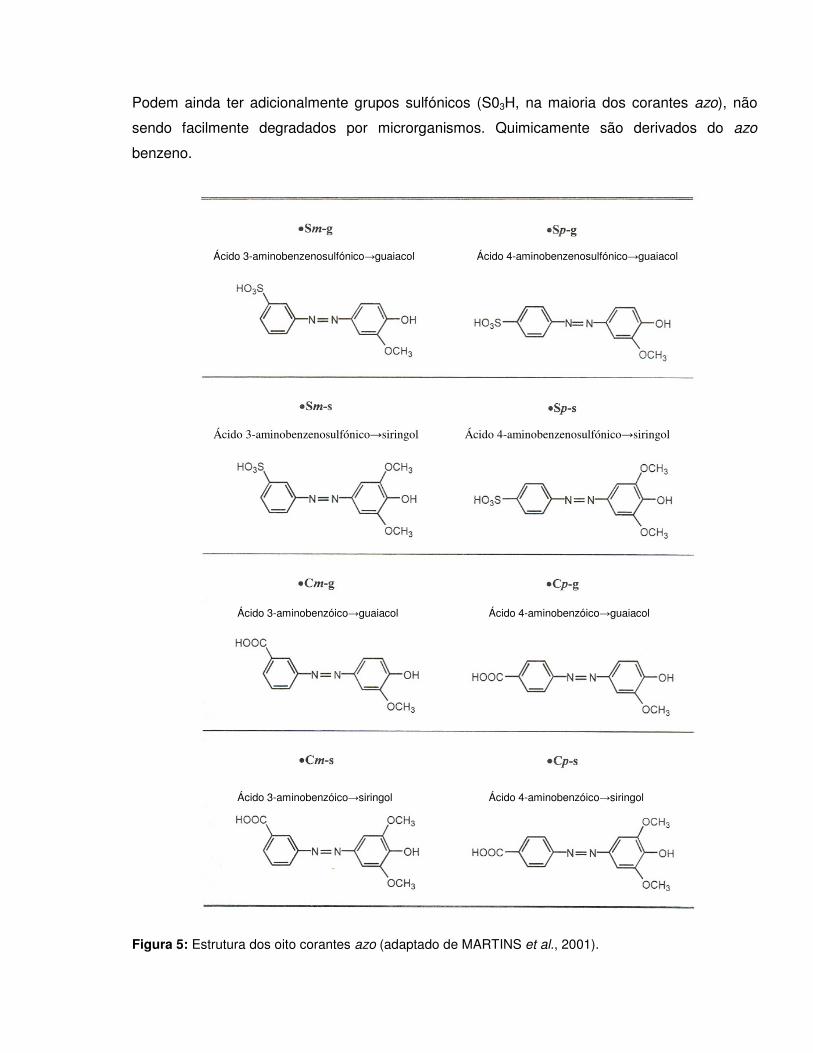
Podem ainda ter adicionalmente grupos sulfónicos (S03H, na maioria dos corantes azo), não
sendo facilmente degradados por microrganismos. Quimicamente são derivados do azo
benzeno.
Figura 5: Estrutura dos oito corantes azo (adaptado de MARTINS et al., 2001).
Ácido 3-aminobenzenosulfónico→guaiacol Ácido 4-aminobenzenosulfónico→guaiacol
Ácido 3-aminobenzenosulfónico→siringol Ácido 4-aminobenzenosulfónico→siringol
Ácido 3-aminobenzóico→guaiacol Ácido 4-aminobenzóico→guaiacol
Ácido 3-aminobenzóico→siringol Ácido 4-aminobenzóico→siringol

A nomenclatura apresentada para os corantes neste trabalho, expressando a componente
diazo e a componente de ligação dos grupos sulfónicos é comum na química têxtil para
evidenciar o processo de síntese. As siglas referem-se ao tipo de ácido (carboxílico – C ou
sulfónico – S), à sua posição no anel (meta – m ou para – p) e ao componente de ligação
(guaiacol – g ou siringol – s). Utilizaram-se assim os oito corantes (Figura 5): ácido meta-
aminobenzenosulfónico-guaiacol (Sm-g), ácido para-aminobenzenosulfónico-guaiacol (Sp-g),
ácido meta-aminobenzenosulfónico-siringol (Sm-s), ácido para-aminobenzenosulfónico-siringol
(Sp-s), ácido meta-aminobenzóico-guaiacol (Cm-g), ácido para-aminobenzóico-guaiacol (Cp-g),
ácido para-aminobenzóico-siringol (Cm-s), ácido para-aminobenzóico-siringol (Cp-s).
5.5. Síntese dos corantes azo
Oito corantes azo foram sintetizados por Ferreira (1998) por diazotação de ácidos meta ou para
aminobenzoicos ou aminosulfónicos (componentes diazo) e a sua junção com guaiacol ou
siringol (componentes de acoplamento) (Figura 6).
Figura 6: Esquema da síntese dos corantes azo (retirada de MARTINS et al., 2001).
A uma solução de componentes diazo (25 mmol) em NaOH 5 % (16 mL) e água (4 mL) foi
adicionada uma solução de NaNO2 (20 mmol) em água (4 mL). A solução resultante foi
adicionada lentamente e com agitação constante a uma mistura de HCl concentrado (4 mL) e
gelo (50 g) mantendo a temperatura entre 0 e 5 ºC, tendo-se formado um precipitado. Depois o

ácido diazotado foi adicionado à solução gelada de acoplamento (20 mmol) em NaOH a 5 %
(40 mL), com agitação. Finalmente, NaCl (40 g) foi adicionado e a mistura foi agitada
continuamente durante 30 minutos à temperatura ambiente. O corante azo precipitado foi
filtrado e lavado com etanol e éter. Os corantes sintetizados (Figura 5) foram caracterizados
utilizando espectroscopia de UV-visível, IV e H-NMR e espectroscopia de massa. A
nomenclatura apresentada, expressa o componente diazo → componente de acoplamento, é
usada na química têxtil ara sugerir o processo de síntese (MARTINS et al., 2001).
5.6. Estudos feitos com os corantes azo utilizados
Estes corantes foram sintetizados com ácido aminobenzóico e aminosulfónico como
componentes diazo e com grupos bioacessíveis como 2-metoxifenol (guaiacol) e 2,6-
dimetoxifenol (siringol) como componentes de acoplamento. Estes grupos bioacessíveis estão
presentes na estrutura da lenhina e parecem ser pontos de acesso para as enzimas linholíticas
produzidas por fungos como o Phanerochaete chrysosporium (MARTINS et al., 2001).
Foram realizados também estudos comparativos de degradação fúngica destes corantes azo
reactivos simples ou misturados (MARTINS et al., 2003); e estudos sobre a biodegradação de
corantes azo têxteis por Phanerochaete chrysosporium (MARTINS et al., 2001).

6. METODOLOGIA

6.1. Microorganismo
O organismo unicelular que serve de bioindicador para esta investigação é o protozoário
Tetrahymena pyriformis da estirpe CCAP 1630/1W, obtido da Colecção de Culturas de Estirpes
de Algas e Protozoários do Reino Unido. As células do banco de colecção foram mantidas em
cultura na estufa, a uma temperatura constante de 25 ºC em meio PPY - Proteose Peptone
Yeast Extract Médium (CCAP,1995), sendo periodicamente - cada 2 meses - sujeitas a
rejuvenescimento.
6.2. Condições de cultura e exposição a corantes azo
Todos os ensaios toxicológicos utilizaram culturas axénicas, de 18 a 24 horas, de Tetrahymena
pyriformis. As células cresceram até à fase exponencial na estufa a 25 ºC, em meio de PPY (2
% de proteose peptona e 0,25 % de extracto de levedura) a pH entre 7,0 e 7,5. Antes da
esterilização por autoclavagem, o meio foi filtrado utilizando filtros com porosidade de 0,45 µm,
de modo a eliminar partículas que pudessem interferir com a observação microscópica e
contagem electrónica de células.
Todos os ensaios foram efectuados com uma densidade inicial de cultura entre as 8000 e as
11000 células/mL. Estas densidades foram conseguidas através da concentração por
centrifugação das culturas celulares em tubos de 50 mL, numa centrífuga Sigma 4K15, a uma
velocidade de 4500 rpm, durante 2 minutos, após a qual foi retirada cerca de metade do
sobrenadante. Da cultura concentrada de células foi retirada uma amostra de 500 µL, diluída
em 1 mL posteriormente em água destilada, perfazendo um volume final de 1,5 mL num tubo
Eppendorf de 2 mL. Foram retiradas três amostras de 30 µL, às quais se adicionaram 30 µL de
formalina a 4 %; as amostras foram observadas num microscópio óptico Leica DMR a uma
ampliação de 100x e determinou-se o número total de células em cada amostra, fazendo-se
depois a média das três amostras. Para se obter densidades entre as 8000 e as 11000
células/mL, em cada amostra de 30 µL, deveríamos ter entre 250 a 400 células.
Os corantes azo estudados foram adicionados às culturas celulares nas seguintes
concentrações: 5, 25, 50 e 100 ppm, tendo sido as células não expostas ao corante usadas
como controlo. As células de T. pyriformis tratadas e não tratadas foram incubadas em tubos
Eppendorf de 2 mL, num volume total de 1,5 mL (DIAS et al., 2003). O pequeno volume destes
tubos não provoca a baixa oxigenação da cultura, uma vez que Dias et al. (2002) levaram a
cabo dois ensaios nos quais comparam as condições de crescimento de culturas celulares em
tubos Eppendorf de 2 mL e em tubos de plástico de 12 mL, encontrando uma elevada
correlação de taxa de crescimento em ambos os ensaios (r2=0,989).

Os ensaios foram realizados nas mesmas condições, sem stress adicional por esfomeamento,
competição por alimento ou mudança das condições ambientais (NICOLAU et al., 2001).
6.3. Compostos tóxicos e suas concentrações
Os oito corantes (Figuras 5 e 7), previamente descritos, foram sintetizados por investigadores
da Universidade do Minho (FERREIRA, 1998), e são os seguintes:
� ácido meta-aminobenzenosulfónico-guaiacol (Sm-g)
� ácido para-aminobenzenosulfónico-guaiacol (Sp-g)
� ácido meta-aminobenzenosulfónico-siringol (Sm-s)
� ácido para-aminobenzenosulfónico-siringol (Sp-s)
� ácido meta-aminobenzóico-guaiacol (Cm-g)
� ácido para-aminobenzóico-guaiacol (Cp-g)
� ácido meta-aminobenzóico-siringol (Cm-s)
� ácido para-aminobenzóico-siringol (Cp-s)
A
B
Figura 7 A e B: Corantes azo em pó (A) e soluções stock de 500 ppm dos corantes azo utilizados para
os ensaios de toxicidade (B).

Os corantes foram administrados às culturas celulares através de oito soluções para as quatro
concentrações de corantes estipuladas: 5, 25, 50 e 100 ppm. Para tal foram previamente
preparadas duas soluções stock de todos os corantes utilizados: solução stock A de 1000 ppm
e solução stock B de 500 ppm (Figura 7). Antes da esterilização por autoclavagem, as soluções
foram filtradas utilizando filtros com porosidade de 0,45 µm, e mantiveram-se ao abrigo da luz,
uma vez que os corantes são fotossensíveis.
As culturas celulares expostas aos corantes azo nas quatro concentrações estipuladas: 5, 25,
50 e 100 ppm, foram realizadas da seguinte forma:
� 100 ppm - 150 µL de solução stock A dos corantes + 350 µL de água destilada + 500
µL de PPY concentrado 2x + 500 µL de cultura de T. pyriformis;
� 50 ppm - 75 µL de solução stock A dos corantes + 425 µL de água destilada + 500 µL
de PPY concentrado 2x + 500 µL de cultura de T. pyriformis;
� 25 ppm - 75 µL de solução stock B dos corantes + 425 µL de água destilada + 500 µL
de PPY concentrado 2x + 500 µL de cultura de T. pyriformis;
� 5 ppm - 15 µL de solução stock B dos corantes + 485 µL de água destilada + 500 µL de
PPY concentrado 2x + 500 µL de cultura de T. pyriformis;
6.4. Ensaios de Determinação de Toxicidade
6.4.1. Ensaios Fisiológicos Toxicológicos
As culturas celulares de T. pyriformis foram expostas a corantes azo em ensaios com a duração
de 48 horas. Foram realizados dois ensaios independentes para cada concentração dos oito
corantes azo, com duas a quatro réplicas. Os corantes foram adicionados às células nas
seguintes concentrações: 5, 25, 50 e 100 ppm, as células não expostas ao corante foram
usadas como controlo. As células de T. pyriformis tratadas e não tratadas foram incubadas em
tubos Eppendorf de 2 mL, num volume total de 1,5 mL (DIAS et al., 2003), para cada tempo de
amostragem (1, 24 e 48 horas).
a. Ensaios de Crescimento:
Contagem de células no contador automático de células
Para a contagem automática das células, estas têm que estar suspensas num electrólito. A
suspensão de células é retirada, através de vácuo, por uma abertura com um diâmetro

conhecido, inserida num tubo com eléctrodos, que criam uma corrente eléctrica que atravessa a
abertura. Cada célula que passa na abertura desloca um volume igual de electrólito, causando
uma resistência na corrente eléctrica. Esta modulação é detectada pelo aparelho através de um
sinal. Estes sinais são amplificados electronicamente e contados automaticamente. O aparelho
tem que ser calibrado para as células e volumes pretendidos e é estabelecido um tamanho
superior e um inferior para eliminar as contagens acima ou abaixo destes tamanhos,
respectivamente (TEUNISSON, 1971).
Dimensões celulares e ajuste do contador automático de células
Os ajustes do aparelho - Beckman Coulter Z Series (E.U.A.) - para um tubo capilar de abertura
de 140 µm, foi realizado previamente para um volume de contagem de 1 mL. As células são
contadas de acordo com o seu volume e não com a sua forma. Antes de cada ensaio procedeu-
se à confirmação dos valores pré-definidos no setup, que se evidenciam na Tabela 3. Foram
escolhidos os valores relativos aos tamanhos mais predominantes, ou seja entre 10 e 30 µm.
As contagens foram realizadas para células com tamanhos menores do que 10 µm, tamanhos
entre 10 e 30 µm e tamanhos acima do 30 µm. Foram retiradas amostras de 1,5 mL, das
culturas controlo e das expostas aos corantes, às 1, 24 e 48 horas, as quais foram diluídas
1:10; 1:20 e 1:30, respectivamente, numa solução salina isotónica – ISOTON II (Beckman
Coulter, E.U.A.). Para cada amostra diluída fez-se a contagem das células vivas em quatro sub-
amostras de 1 mL, a partir das quais se calculou o valor médio das quatro contagens. Para
minimizar erros de contagem as diluições foram realizadas para garantir uma concentração
baixa de solução, sendo a ideal abaixo dos 5 %.
Tabela 3: Setup do contador automático de células -Beckman Coulter Z Series - para um tubo capilar de
abertura de 140 µm.
Abertura Limite de tamanho inferior Limite de tamanho superior
140 µm 10 µm 30 µm
O aparelho foi verificado periodicamente, tendo sido limpo com solução salina ISOTON II, após
longos períodos sem utilização (mais do que 15 dias). Quando as contagens davam erros de
leitura que poderiam estar associados a detritos acumulados no aparelho, procedeu-se à sua

limpeza através do esvaziamento e enchimento da abertura com ISOTON II, ou mesmo através
do esvaziamento e enchimento de todo o sistema.
b. Ensaios de Morfometria:
Com este método pretendeu-se saber qual a alteração da morfologia celular por exposição aos
vários corantes. Assim, amostras tratadas e não tratadas foram fixadas durante 1 hora em
solução NBF (Formalina Neutra Tamponizada), lavadas e coradas com azul toluidina a 0,01 %
(Sigma). Sub-amostras de 50 µL de cultura suspensa foram colocadas em lâminas e analisadas
com um microscópio óptico invertido (Nikon Diaphot 300) a uma ampliação de 100x. Procedeu-
se seguidamente à análise da imagem com uma câmara de vídeo monocromática CCD (Sony
AVCD5CE), binarizada e os dados foram analisados pelo software MATLAB 5.1 (The
Mathworks Inc., Natick) de modo a determinar a área e a razão (W/L) dos eixos mais curto (W)
e mais longo (L) das células. Os parâmetros morfológicos foram determinados através da
imagem binária final. Foi realizada uma calibração prévia, através de uma lâmina com uma rede
graduada, ajustada para um milímetro. A uma ampliação de 100x, 0,5 mm correspondiam a um
valor médio de 459 pixels no ecrã do computador. Seguindo as mesmas condições para a
captura de imagem, o equivalente entre pixels e micrómetros foi calculado segundo a equação:
1 pixel = 1, 089 µm. Em cada sub-amostra, foram analisadas aleatoriamente 100 células nos
seguintes tempos de amostragem: 1 e 48 horas (DIAS et al.. 2003).
c. Ensaios de Predação:
Estes ensaios foram realizados em tubos Eppendorf de 2 mL, por adição de microesferas
amarelo-esverdeadas fluorescentes de látex (FluoSpheres , Molecular Probes, E.U.A.) com
0,5 µm de diâmetro, às células tratadas e às células controlo. Foi utilizada uma concentração
final de aproximadamente 1,05x106 microesferas/mL em meio PPY de acordo com Dias et al..
(2003). Ensaios realizados anteriormente determinaram qual o tamanho e a concentração de
microesferas que possibilitava a melhor observação microscópica e a contagens subsequente
de microesferas predadas. Sherr et al. (1987 in NICOLAU et al., 1999) realizou experiências
com ciliados fagotróficos estuarinos e mostrou que estes ingeriam as bactérias fluorescentes a
uma taxa superior do que ingeriam as microesferas de tamanho idêntico, o que indica que este

ensaio pode subestimar a predação. No entanto, este ensaio apenas pretende avaliar a
toxicidade dos corantes azo em Tetrahymena pyriformis.
Para cada concentração e cada corante, após 20 minutos de incubação no escuro, as amostras
são mortas e fixadas pela adição de solução de NBF (10 % (v/v) de formalina em tampão
fosfato salino (PBS) de pH 7,0, a uma concentração final de 2-5 % (v/v)) durante 1 hora.
Procedeu-se depois à lavagem e ressuspensão de células em azida de sódio a 0,1 % (p/v), em
PBS, e armazenadas no escuro até à análise subsequente com um microscópio epifluorescente
(ZEISS Axioskop). Cada sub-amostra, retirada às 1, 24 e 48 horas, foi observada a uma
ampliação de 100x; quando uma célula era localizada, a luz incidente foi mudada para luz UV
(com filtro de excitação EX 450-490 nm, espelho FT 510 e filtro de barreira BA 520) e foi
contado o número de microesferas ingeridas, com uma ampliação de 1000x. Em cada sub-
amostra, retirada de cada ensaio, as 30 células foram analisadas e o número total de
microesferas ingeridas foi determinado para os dois ensaios independentes. (NICOLAU et al.,
1999; DIAS et al.. 2003).
6.4.2. Ensaios Bioquímicos Toxicológicos
6.5. Tratamento estatístico dos dados
Depois da realização dos ensaios procedeu-se à análise estatística dos dados. Em primeiro
lugar realizaram-se testes F para comparar as variâncias de duas amostras de dados
(comparou-se sempre os controlos com cada um dos corantes, para cada concentração e
tempo de amostragem). De acordo com os resultados obtidos realizaram-se posteriormente
testes t de Student para variâncias iguais e testes t de Student para variâncias desiguais, para
verificar se as médias de duas amostras eram significativamente diferentes (comparou-se
sempre os controlos com cada um dos corantes, para cada concentração e tempo de
amostragem). Utilizaram-se os testes F e os testes t de Student para a análise estatística de
todos os ensaios: crescimento, predação e morfometria. Realizaram-se também análises de
variância com factor único (ANOVA) (quando as variâncias das amostras eram iguais) e testes t
de Student para variâncias desiguais para verificar se as médias dos seguintes grupos: siringol-
guaiacol (s-g); carboxílico-sulfónico (C-S) e meta-para (m-p), eram significativamente diferentes.
Foram aceites os valores de p<0,05 como nível de significância estatística entre grupos (DIAS
et al.. 2003) e o p<0,01 foi utilizado para enfatizar o maior nível de significância estatística
obtida. Todo o tratamento estatístico foi realizado no software Microsoft Excel 2003.

7. RESULTADOS

Os resultados são apresentados por ordem alfabética: crescimento, morfometria e predação. Os
resultados dos ensaios de crescimento e morfometria apresentam-se em gráficos e tabelas de
significância onde se registam os valores de p estatístico. Os resultados dos ensaios de
predação apresentam-se em tabelas de médias, desvios-padrão com os valores de p.
Foram apenas considerados para a análise de resultados os valores estatisticamente
significativos para p<0,05 e p<0,01, caso estes se registassem em ambos os ensaios realizados
para cada parâmetro (estes encontram-se assinalados a cinza nas respectivas tabelas). Nas
tabelas encontram-se a itálico os valores de p>0,05.
Para cada parâmetro analisaram-se os resultados e, por último, fez-se uma síntese, dando
especial ênfase aos efeitos globais causados por cada corante azo estudado.
7.1. Crescimento
Embora se tenham realizados dois ensaios independentes, apenas se apresenta um deles
(SEWARD et al., 2001). Os resultados obtidos num de dois ensaios independentes de
crescimento são apresentados na Figura 8. Apresenta-se apenas um dos gráficos, uma vez que
as diferenças entre eles não aparentam ser muito relevantes. Em cada gráfico estão patentes
as médias de quatro réplicas retiradas em cada tempo de amostragem e o respectivo desvio-
padrão.
Conforme se observa na Figura 8 e de acordo com a Tabela 4, à medida que as concentrações
dos corantes foram aumentando, aumentaram globalmente as diferenças significativas entre os
controlos e os corantes. De uma forma geral encontram-se mais diferenças significativas nos
corantes sulfónicos do que nos corantes carboxílicos. Seguidamente será feita a análise dos
resultados relativos a cada corante, para todas as concentrações:
Cm-g: este corante não possui toxicidade para as concentrações de 5, 25 e de 100 ppm. Para
uma concentração de 50 ppm registou-se uma diferença significativa (p<0,01) entre este
corante e o controlo depois de 1 hora de exposição.
Cp-g: este corante não possui toxicidade para a concentração de 5 ppm. Já para as
concentrações de 25 e 100 ppm o corante inibiu o crescimento de Tetrahymena pyriformis às 24
e 48 horas significativamente (p<0,05). Para uma concentração de 50 ppm o corante apenas
inibiu o crescimento após 1 hora de exposição, tendo a seguir recuperado, embora se note um
crescimento menor que o controlo, no entanto não significativo estatisticamente. Em todas as

concentrações e de um modo geral este corante inibiu o crescimento de Tetrahymena
pyriformis.
A B
0
20000
40000
60000
80000
100000
120000
140000
160000
180000
200000
1 24 48
Tempo de exposição (horas)
n.º
cél
ula
s/ m
l
Controlo
Cm-g
Cp-g
Sm-g
Sp-g
Sm-s
Sp-s
Cm-s
Cp-s
C D
0
20000
40000
60000
80000
100000
120000
140000
160000
180000
1 24 48
Tempo de exposição (horas)
n.º
cél
ula
s/ m
l
Controlo
Cm-g
Cp-g
Sm-g
Sp-g
Sm-s
Sp-s
Cm-s
Cp-s
0
20000
40000
60000
80000
100000
120000
140000
1 24 48
Tempo de exposição (horas)
n.º
cél
ula
s/ m
l
Controlo
Cm-g
Cp-g
Sm-g
Sp-g
Sm-s
Sp-s
Cm-s
Cp-s
Figura 8: Crescimento de Tetrahymena pyriformis exposta a quatro concentrações de corantes azo: 5
ppm (gráfico A), 25 ppm (gráfico B), 50 ppm (gráfico C) e 100 ppm (gráfico D), em ensaios de 48 horas.
Para cada tempo de amostragem são representadas a médias e os desvios padrão de quatro réplicas.
Sm-g: este corante revelou-se mais tóxico apenas nos ensaios para a concentração de 5 ppm,
uma vez que às 24 horas este provocou uma acentuada quebra no crescimento relativamente
ao controlo e aos outros corantes. Este decréscimo foi registado igualmente às 48 horas. Para
as concentrações de 25 e 50 ppm apenas se registou diferenças significativas (p<0,01 e
p<0,05, respectivamente) em relação ao controlo após 1 hora de exposição, tendo a população
de Tetrahymena recuperado o crescimento. Para a concentração de 100 ppm houve diferenças
significativas em relação ao controlo para níveis de significância de 0,01 às 24 horas de
exposição, tendo a população recuperado seguidamente.
Sp-g: este corante revelou valores de crescimento irregulares quando comparado com o
controlo. Para a concentração de 5 ppm apenas registou um crescimento menor e
significativamente diferente para p<0,05 às 48 horas de exposição; para a concentração de 50
0 20000
40000
60000
80000
100000
120000
140000
1 24 48 Tempo de exposição (horas)
n.º
cél
ula
s/ m
l
Controlo
Cm-g Cp-g Sm-g Sp-g Sm-s Sp-s Cm-s Cp-s

ppm registou um crescimento menor e significativamente diferente para p<0,01 à 1 e para
p<0,05 às 48 horas de exposição; para a concentração de 100 ppm registou um crescimento
maior e significativamente diferente para p<0,01 às 24 horas de exposição.
Sm-s: este corante revelou um comportamento irregular similar ao corante Sp-g, uma vez que
para a concentração de 5 ppm apenas registou um crescimento menor e significativamente
diferente para p<0,01 às 48 horas de exposição; para a concentração de 50 ppm registou um
crescimento menor e significativamente diferente para diferente para p<0,01 à 1 e para p<0,05
às 24 horas de exposição; para a concentração de 100 ppm registou um crescimento maior e
significativamente diferente para p<0,01 às 24 horas de exposição.
Sp-s: este corante registou para a concentração de 5 ppm um crescimento menor e
significativamente diferente para p<0,01 após 48 horas de exposição; para a concentração de
25 ppm este corante revelou um crescimento menor que o controlo significativamente diferente
(p<0,05) após 1 hora de exposição; para a concentração de 50 ppm registou um crescimento
menor e significativamente diferente para diferente para p<0,05 após 24 horas de exposição;
para a concentração de 100 ppm registou um crescimento maior e significativamente diferente
para p<0,05 após 1 hora de exposição e p<0,01 após 24 horas.
Cm-s: este corante condicionou globalmente para todas as concentrações um crescimento
inferior relativamente ao controlo, no entanto apenas na concentração de 100 ppm consegue
obter valores de crescimento menores e significativamente diferentes do controlo. Desta forma
às 24 horas os valores mostram-se estatisticamente diferentes para p<0,01 e às 48 horas para
um p<0,05. A seguir ao corante Cm-g é o que apresenta menos diferenças significativas
relativamente aos valores de crescimento do controlo.
Cp-s: este corante influenciou um crescimento irregular para todas as concentrações: às 48
horas de exposição para as concentrações de 5 e 25 ppm o crescimento foi antagónico, ou
seja, inibiu o crescimento significativamente (p<0,01) e aumentou o crescimento
significativamente (p<0,05), respectivamente. Para a concentração de 100 ppm o corante
mostrou-se significativamente diferente do controlo para um valor de p<0,05.
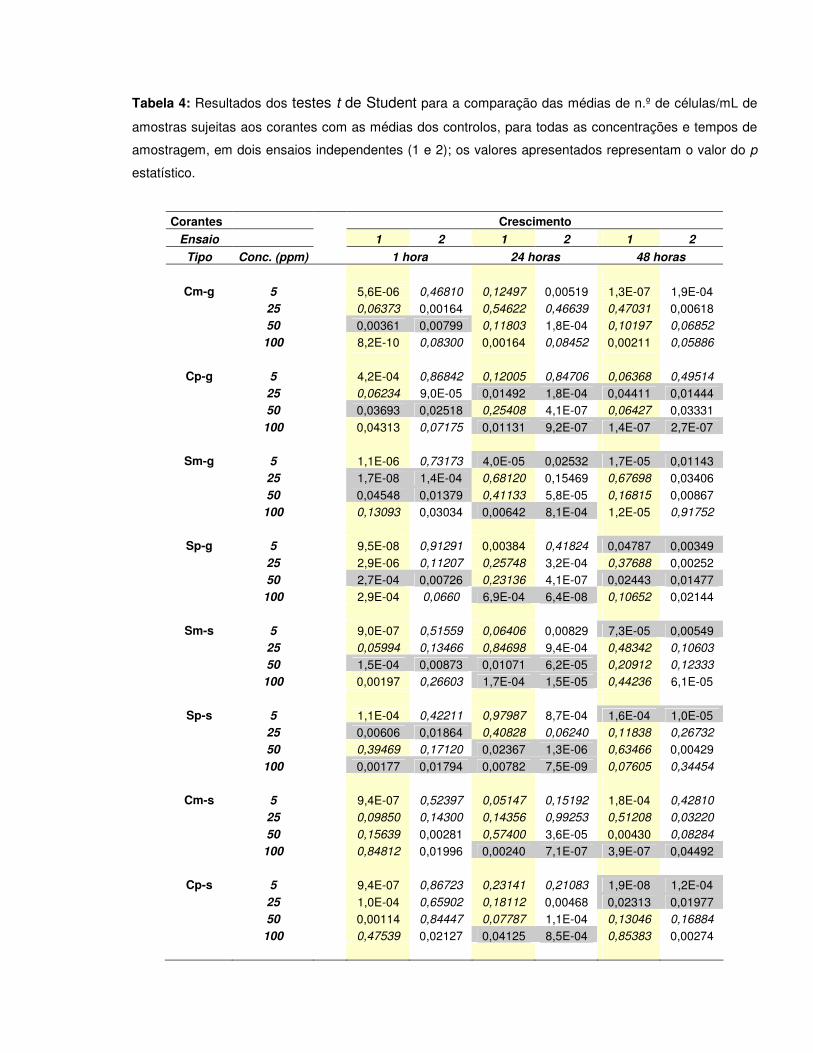
Tabela 4: Resultados dos testes t de Student para a comparação das médias de n.º de células/mL de
amostras sujeitas aos corantes com as médias dos controlos, para todas as concentrações e tempos de
amostragem, em dois ensaios independentes (1 e 2); os valores apresentados representam o valor do p
estatístico.
Corantes Crescimento
Ensaio 1 2 1 2 1 2
Tipo Conc. (ppm) 1 hora 24 horas 48 horas
Cm-g 5 5,6E-06 0,46810 0,12497 0,00519 1,3E-07 1,9E-04 25 0,06373 0,00164 0,54622 0,46639 0,47031 0,00618 50 0,00361 0,00799 0,11803 1,8E-04 0,10197 0,06852
100 8,2E-10 0,08300 0,00164 0,08452 0,00211 0,05886
Cp-g 5 4,2E-04 0,86842 0,12005 0,84706 0,06368 0,49514
25 0,06234 9,0E-05 0,01492 1,8E-04 0,04411 0,01444 50 0,03693 0,02518 0,25408 4,1E-07 0,06427 0,03331 100 0,04313 0,07175 0,01131 9,2E-07 1,4E-07 2,7E-07
Sm-g 5 1,1E-06 0,73173 4,0E-05 0,02532 1,7E-05 0,01143 25 1,7E-08 1,4E-04 0,68120 0,15469 0,67698 0,03406 50 0,04548 0,01379 0,41133 5,8E-05 0,16815 0,00867 100 0,13093 0,03034 0,00642 8,1E-04 1,2E-05 0,91752
Sp-g 5 9,5E-08 0,91291 0,00384 0,41824 0,04787 0,00349
25 2,9E-06 0,11207 0,25748 3,2E-04 0,37688 0,00252 50 2,7E-04 0,00726 0,23136 4,1E-07 0,02443 0,01477 100 2,9E-04 0,0660 6,9E-04 6,4E-08 0,10652 0,02144
Sm-s 5 9,0E-07 0,51559 0,06406 0,00829 7,3E-05 0,00549 25 0,05994 0,13466 0,84698 9,4E-04 0,48342 0,10603
50 1,5E-04 0,00873 0,01071 6,2E-05 0,20912 0,12333
100 0,00197 0,26603 1,7E-04 1,5E-05 0,44236 6,1E-05
Sp-s 5 1,1E-04 0,42211 0,97987 8,7E-04 1,6E-04 1,0E-05 25 0,00606 0,01864 0,40828 0,06240 0,11838 0,26732
50 0,39469 0,17120 0,02367 1,3E-06 0,63466 0,00429 100 0,00177 0,01794 0,00782 7,5E-09 0,07605 0,34454
Cm-s 5 9,4E-07 0,52397 0,05147 0,15192 1,8E-04 0,42810
25 0,09850 0,14300 0,14356 0,99253 0,51208 0,03220
50 0,15639 0,00281 0,57400 3,6E-05 0,00430 0,08284
100 0,84812 0,01996 0,00240 7,1E-07 3,9E-07 0,04492
Cp-s 5 9,4E-07 0,86723 0,23141 0,21083 1,9E-08 1,2E-04 25 1,0E-04 0,65902 0,18112 0,00468 0,02313 0,01977 50 0,00114 0,84447 0,07787 1,1E-04 0,13046 0,16884
100 0,47539 0,02127 0,04125 8,5E-04 0,85383 0,00274

Relativamente aos resultados do teste ANOVA (registados na Tabela 5), quanto à comparação
entre médias de grupos de corantes de todas as concentrações: guaiacol-siringol (g-s);
carboxílico-sulfónico (C-S); meta-para (m-p), para os respectivos tempos de amostragem,
apenas se registou uma diferença significativa. Os resultados mostram que no ensaio 1
realizado para o crescimento há uma diferença significativa entre os grupos de corantes com
guaiacol e com siringol.
Tabela 5: Resultados dos testes ANOVA para o crescimento de Tetrahymena pyriformis. A tabela faz a
comparação entre médias de grupos de corantes de todas as concentrações: guaiacol-siringol (g-s);
carboxílico-sulfónico (C-S); meta-para (m-p), para os respectivos tempos de amostragem. Os valores
apresentados representam o valor do p estatístico.
7.2. Morfometria
Embora se tenham realizado dois ensaios independentes, apenas se apresenta um deles
(SEWARD et al., 2001). Os resultados obtidos num de dois ensaios independentes de
morfometria são apresentados nas Figuras 9 e 10. Apresenta-se apenas um dos gráficos, uma
vez que as diferenças entre eles não aparentam ser muito relevantes.
Estão representados nas Figuras 9 e 10, respectivamente, os efeitos da exposição a diferentes
concentrações de corantes azo na área e na razão W/L de Tetrahymena pyriformis, em ensaios
de 48 horas, para concentrações de 5, 25, 50 e 100 ppm. Em cada gráfico estão patentes as
médias de quatro réplicas retiradas em cada tempo de amostragem e o respectivo desvio-
padrão.
Grupos de Ensaio Crescimento
corantes 1 hora 24 horas 48 horas
g-s 1 0,97894 0,86530 0,03806 2 0,72822 0,68213 0,06867
C-S 1 0,42627 0,61737 0,33407
2 0,69345 0,14303 0,10550
m-p 1 0,38404 0,82828 0,98973
2 0,38410 0,93030 0,09308

A B
0
200
400
600
800
1000
1200
1 48
Tempo de exposição (horas)
Áre
a ( µµ µµ
m)
Controlo
Cm-g
Cp-g
Sm-g
Sp-g
Sm-s
Sp-s
Cm-s
Cp-s
0
200
400
600
800
1000
1200
1 48
Tempo de exposição (horas)
Áre
a ( µµ µµ
m)
Controlo
Cm-g
Cp-g
Sm-g
Sp-g
Sm-s
Sp-s
Cm-s
Cp-s
C D
0
200
400
600
800
1000
1200
1 48
Tempo de exposição (horas)
Áre
a ( µµ µµ
m)
Controlo
Cm-g
Cp-g
Sm-g
Sp-g
Sm-s
Sp-s
Cm-s
Cp-s
0
200
400
600
800
1000
1200
1 48
Tempo de exposição (horas)Á
rea
( µµ µµm
)
Controlo
Cm-g
Cp-g
Sm-g
Sp-g
Sm-s
Sp-s
Cm-s
Cp-s
Figura 9: Efeito da exposição a diferentes concentrações de corantes azo no tamanho (área) de
Tetrahymena pyriformis, em ensaios de 48 horas. As concentrações usadas foram de 5 ppm (gráfico A),
25 ppm (gráfico B), 50 ppm (gráfico C), e 100 ppm (gráfico D). Para cada tempo de amostragem são
representadas a médias e os desvios padrão de quatro réplicas.
De acordo com a Figura 9 e com a Tabela 6, nota-se de forma geral que as diferenças entre as
áreas dos controlos e as áreas dos corantes para ambos os tempos de amostragem são
escassas. À medida que aumentam as concentrações dos corantes, foram-se verificando mais
diferenças significativas globais quando comparamos as médias dos corantes com as dos
controlos, sendo essas diferenças acentuadas com o aumento da concentração e para as
amostragens realizadas após 48 horas de exposição. Apresenta-se, de seguida, a análise dos
resultados relativos a cada corante, para todas as concentrações:
Cm-g: este corante apresentou, apenas, uma diferença relativa ao controlo às 48 horas, para
uma concentração de 100 ppm. Verificou-se que houve um aumento significativo da área
(p<0,05) das células de Tetrahymena pyriformis relativamente ao controlo.

Sm-g: este corante registou, apenas, uma diferença significativa após 1 hora de exposição,
para uma concentração de 50 ppm. Verificou-se que houve um aumento significativo da área
(p<0,05) das células tratadas relativamente ao controlo.
Sp-g: este corante registou já mais diferenças, tendo sido verificadas sempre às 48 horas de
exposição e para as concentrações de 25, 50 e 100 ppm. A área das células expostas a este
corante aumentaram significativamente relativamente às células do controlo, para um valor de
p<0,05 e para um valor de p<0,05 relativamente à maior concentração testada (100 ppm).
Sm-s: este corante registou diferenças significativas relativamente ao controlo, apenas para
tempos de exposição de 48 horas, nas concentrações de 25 ppm e 100 ppm. A área das
células expostas a este corante aumentaram significativamente na concentração de 25 ppm
(p<0,05) e na concentração de 100 ppm (p<0,01), mostrando que a área das células aumentou
relativamente à área das células do controlo.
Sp-s: este corante registou, apenas, uma diferença significativa após 48 horas de exposição,
para uma concentração de 5 ppm. Verificou-se que houve um aumento significativo da área
(p<0,01) das células tratadas relativamente às células do controlo.
Cp-s: este corante apresentou duas diferenças relativas ao controlo às 48 horas, para as
concentrações de 50 e 100 ppm. Verificou-se que houve um aumento significativo da área
(p<0,01) das células de Tetrahymena pyriformis relativamente ao controlo.
Cp-g e Cm-s: estes corantes não registaram diferenças significativas para qualquer
concentração utilizada e para cada tempo de amostragem.
Da análise acima referida, podemos verificar que em todos os casos de diferenças significativas
relativamente às áreas celulares, quando comparados com as células do controlo, os corantes
provocaram um aumento das áreas celulares.
De acordo com a Figura 10 e com a Tabela 7, pode-se referir que as diferenças entre a razão
eixo menor/eixo maior (W/L) nas células tratadas e nas células não tratadas são ainda menos
do que as verificadas para as áreas das células. Neste caso verificam-se mais diferenças na
concentração de 50 ppm do que nas restantes. Apresenta-se, de seguida, relativamente ao
parâmetro razão (W/L), a análise dos resultados relativos a cada corante, para todas as
concentrações:

A B
00,10,20,30,40,50,60,70,80,9
1
1 48
Tempo de exposição (horas)
Raz
ão
(W
/L)
Controlo
Cm-g
Cp-g
Sm-g
Sp-g
Sm-s
Sp-s
Cm-s
Cp-s
0
0,1
0,2
0,3
0,4
0,50,6
0,7
0,8
0,9
1
1 48
Tempo de exposição (horas)
Raz
ão (
W/L
)
Controlo
Cm-g
Cp-g
Sm-g
Sp-g
Sm-s
Sp-s
Cm-s
Cp-s
C D
0
0,10,2
0,30,4
0,5
0,60,7
0,80,9
1
1 48
Tempo de exposição (horas)
Raz
ão (
W/L
)
Controlo
Cm-g
Cp-g
Sm-g
Sp-g
Sm-s
Sp-s
Cm-s
Cp-s
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
1 48
Tempo de exposição (horas)
Raz
ão (
W/L
)
Controlo
Cm-g
Cp-g
Sm-g
Sp-g
Sm-s
Sp-s
Cm-s
Cp-s
Figura 10: Efeito da exposição a diferentes concentrações de corantes azo na razão (W/L) do eixo menor
(W) e maior (L) de Tetrahymena pyriformis, em ensaios de 48 horas. As concentrações usadas foram de
5 ppm (gráfico A), 25 ppm (gráfico B), 50 ppm (gráfico C), e 100 ppm (gráfico D). Para cada tempo de
amostragem são representadas a médias e os desvios padrão de quatro réplicas.
Cp-g: este corante registou, apenas, uma diferença significativa após 1 hora de exposição, para
uma concentração de 50 ppm. Verificou-se que houve uma diminuição significativa (p<0,01) na
razão (W/L) relativamente ao controlo, indicando uma forma celular mais alongada do que a
apresentada pelas células controlo.
Sm-g: este corante marcou, uma diferença significativa após 48 horas de exposição, para uma
concentração de 50 ppm. Verificou-se que houve uma diminuição significativa (p<0,01) na razão
(W/L) relativamente ao controlo, indicando uma forma celular mais alongada do que a
considerada normal (controlo).
Sp-g: este corante apresentou de forma semelhante ao corante Cp-g apenas uma diferença
significativa após 1 hora de exposição, para uma concentração de 50 ppm. Verificou-se
igualmente que houve uma diminuição, embora menos significativa (p<0,05) na razão (W/L)
relativamente ao controlo, indicando uma forma celular mais alongada do que a apresentada
pelas células do controlo.

Sp-s: este corante registou o inverso do que tem sido analisado, ou seja, verificou-se que na
concentração mais elevada testada (100 ppm) e no maior tempo de exposição (48 horas) este
corante provocou um arredondamento celular significativamente superior (p<0,05) ao registado
para o controlo. A toxicidade neste caso teve um efeito diferente na forma das células
relativamente ao que foi registado para os restantes corantes analisados.
Cm-s: este corante provocou na concentração de 25 ppm, às 48 horas, um significativo
alongamento das células (p<0,05) comparativamente ao controlo. Este efeito já foi registado
anteriormente noutros corantes para a concentração de 50 ppm.
Cm-g, Sm-s e Cp-s: estes corantes não registam diferenças significativas relativamente à
razão (W/L) para qualquer concentração utilizada e para cada tempo de amostragem.
Quando se analisa a Tabela 8, relativa aos resultados do teste ANOVA, quanto à comparação
entre médias de áreas e razões W/L de grupos de corantes de todas as concentrações,
guaiacol-siringol (g-s); carboxílico-sulfónico (C-S); meta-para (m-p), para os respectivos tempos
de amostragem, verifica-se que existem várias diferenças significativas dentro destes grupos
em ambos os ensaios realizados (ensaios 1 e 2). Relativamente ao parâmetro área das células
de Tetrahymena pyriformis verifica-se diferenças significativas, à 1 hora em dois grupos
distintos e em ambos os ensaios: guaiacol-siringol (g-s) para um valor de p<0,01 e meta e para
(m-p), assumindo valor de p<0,05. Esta diferença mantém-se no apenas no ensaio 1, em
ambos os grupos, após 48 horas de exposição. Quanto ao parâmetro morfométrico - razão W/L
– nota-se que existem diferenças significativas entre os seguintes grupos de corantes:
� grupo guaiacol-siringol (g-s) às 48 horas, em ambos os ensaios, para valores de p<0,05;
� grupo carboxílico-sulfónico (C-S) à 1 hora, em ambos os ensaios para valores de
p<0,05;
� grupo meta-para (m-p) em ambos os tempos de amostragem para valores de p<0,01.
Analisando os dois parâmetros (área e razão W/L) simultaneamente pode-se constatar que:
� os efeitos tóxicos entre os grupos de corantes guaiacol-siringol (g-s) são
significativamente diferentes consoante o parâmetro morfométrico analisado, uma vez
que se manifestaram efeitos tóxicos relativamente às áreas após uma hora de exposição
e relativamente à razão W/L após 48 horas de exposição;

Tabela 6: Resultados dos testes t de Student, para a comparação de médias das áreas de amostras
sujeitas aos corantes com as médias dos controlos, para todas as concentrações e tempos de
amostragem. Os valores apresentados representam o valor do p estatístico.
Corantes Morfometria (áreas)
Ensaio 1 2 1 2
Tipo Conc. (ppm) 1 hora 48 horas
Cm-g 5 0,28886 0,51031 0,74846 0,00134 25 0,10094 3,48E-05 2,21E-05 0,19843
50 0,23728 0,34881 2,24E-09 0,00021 100 2,85E-06 0,20680 1,2E-05 0,01215
Cp-g 5 0,68665 0,54153 0,02412 0,22624
25 0,17857 3,24E-06 9,52E-06 0,75207
50 0,16146 0,00112 1,15E-04 0,21841
100 0,30942 0,40945 3,07E-04 0,11054
Sm-g 5 0,40184 0,71977 0,37552 0,68360
25 0,06750 3,89E-11 8,66E-09 0,96133
50 0,03296 0,00312 0,00434 0,05165
100 2,33E-05 0,63412 0,08838 0,97663
Sp-g 5 0,26047 0,25131 0,77413 0,10075
25 0,47751 5,1E-08 0,04927 0,01584 50 0,42635 7,09E-05 0,01337 0,00634 100 4,08E-07 0,46771 3,27E-14 2,35E-04
Sm-s 5 0,77607 0,84544 0,31957 0,01287 25 0,46570 1,04E-05 0,00254 0,03119 50 0,21733 0,33529 4,74E-07 0,08863
100 0,25708 0,44162 7,69E-06 0,00241
Sp-s 5 0,55630 0,71570 0,00794 0,00133 25 0,31324 5,01E-09 0,18832 0,66165
50 0,00435 0,19519 7,56E-05 0,89713
100 0,11012 1,59E-07 2,28E-05 0,05781
Cm-s 5 0,38537 0,70568 0,54583 0,14174
25 0,89844 1,46E-11 0,72850 0,23987
50 0,35935 0,16046 9,43E-08 0,87239
100 0,23636 0,23390 1,21E-04 0,54992
Cp-s 5 0,60599 0,25483 0,26225 0,13232
25 0,56325 2,8E-07 5,36E-06 0,44375
50 0,60605 0,58697 9,2E-07 3,06E-05 100 0,00227 0,21646 2,33E-05 1,99E-07

Tabela 7: Resultados dos testes t de Student para a comparação das médias das razões (W/L) de
amostras sujeitas aos corantes com as médias dos controlos, para todas as concentrações e tempos de
amostragem. Os valores apresentados representam o valor do p estatístico.
Corantes Morfometria (razões W/L)
Ensaio 1 2 1 2
Tipo Conc. (ppm) 1 hora 48 horas
Cm-g 5 0,24458 0,11882 0,28730 0,00205
25 0,19959 0,00035 0,71012 0,24045
50 4,11E-16 0,66816 4,49E-14 0,31062
100 0,29990 0,88772 0,62682 0,00960
Cp-g 5 0,34405 0,08526 9,61E-05 0,25837
25 0,25465 9,21E-05 0,22754 0,26474
50 4,97E-16 1,9E-04 1,76E-30 0,05881
100 0,48773 0,03614 0,13260 0,06472
Sm-g 5 0,39019 0,49394 0,32690 0,21387
25 0,89062 0,00035 0,71939 0,78671
50 8,04E-24 0,34352 1,76E-26 5,17E-04
100 0,01188 0,03614 0,52408 0,3015
Sp-g 5 0,05897 0,23569 0,06387 0,53330
25 0,73316 0,00351 0,01021 0,37961
50 0,02304 0,03602 5,75E-14 0,14326
100 0,00333 0,21755 0,06663 0,00131
Sm-s 5 0,42999 0,00628 0,80016 0,25515
25 0,52053 9,04E-05 0,52464 0,02429
50 0,00294 0,21182 5,63E-18 0,05968
100 0,27859 0,09511 0,69467 0,74046
Sp-s 5 0,23663 0,24567 3,02E-05 0,59953
25 0,19131 0,06357 0,00061 0,59448
50 3,63E-05 0,75412 8,24E-26 0,18879
100 0,23185 5,85E-07 0,03751 8,28E-10
Cm-s 5 0,05909 0,01899 0,49730 0,13825
25 0,87785 0,00076 0,01929 0,03335
50 1,51E-07 0,15417 3,74E-14 0,71050
100 0,05611 0,04421 0,13507 0,65896
Cp-s 5 0,37666 0,01195 0,31039 0,17670
25 0,35705 0,00110 0,25601 0,26000
50 0,86316 0,77352 1,98E-13 0,05640
100 0,00214 0,77373 0,75515 2,41E-04

� os efeitos tóxicos entre os grupos de corantes com grupos carboxílicos e com grupos
sulfónicos (C-S) são significativamente mais evidentes após uma exposição inicial a
estes corantes;
� os efeitos tóxicos entre grupos de corantes com orientação meta ou para (m-p) são
significativamente diferentes durante todo o tempo de exposição, com mais realce após
1 hora de exposição das Tetrahymena pyriformis aos corantes.
Tabela 8: Resultados dos testes ANOVA para a morfometria: área e razão (W/L), de Tetrahymena
pyriformis. A tabela faz a comparação entre médias de grupos de corantes de todas as concentrações:
guaiacol-siringol (g-s); carboxílico-sulfónico (C-S); meta-para (m-p), para os respectivos tempos de
amostragem. Os valores apresentados representam o valor do p estatístico.
Grupos de Ensaio Morfometria (áreas)
Grupos
de Ensaio Morfometria (razões W/L)
corantes 1 hora 48 horas corantes 1 hora 48 horas
g-s 1 5,62E-10 5,99E-29 g-s 1 0,05896 0,04285 2 3,99E-05 0,05741 2 0,46016 4,3E-06
C-S 1 0,98416 0,10891 C-S 1 0,01341 0,08348
2 0,46010 0,00141 2 4,89E-10 0,09542
m-p 1 0,04135 0,04199 m-p 1 1,27E-06 4,67E-06
2 0,04413 0,33845 2 6,44E-14 0,00168
7.3. Predação
Os efeitos das diferentes concentrações dos tóxicos na predação de Tetrahymena pyriformis
estão representados na Tabela 9. Apresentam-se os resultados em forma de tabela, onde estão
patentes as médias das microesferas ingeridas em cada ensaio, os desvios padrão e os níveis
de significância para p<0,05 e p<0,01 para os testes t de Student realizados para comparar as
médias dos vários corantes com as dos controlos.
Embora se tenham realizados dois ensaios independentes, apenas se apresenta um deles
(SEWARD et al., 2001) em forma de tabela, na qual se optou por colocar juntamente com os
dados das médias das microesferas ingeridas e dos desvios padrão, os níveis de significância
adoptados.

Ao comparar os resultados em ambos os ensaios e os valores de p obtidos aquando do
tratamento estatístico t de Student a fim de apurar diferenças entre os corantes e os controlos
para cada concentração e para cada tempo de amostragem, pode-se concluir que os ensaios
são diferentes. Devido a este facto serão apenas consideradas para análise dos resultados as
diferenças significativas encontradas em ambos os ensaios e será apresentado apenas o
ensaio 1, visto ser o que apresenta mais consistência nos resultados. De forma geral, nota-se
que em todas as concentrações existe uma tendência para a diminuição na ingestão de
microesferas ao longo das 48 horas. Os valores dos desvios-padrão indicam também que em
cada célula observada variou muito o número total de microesferas ingeridas. De uma forma
global e salvaguardando algumas pontuais excepções, repara-se que o número de
microesferas ingeridas nos três tempos de amostragem aumentou à medida que se aumentou a
concentração dos corantes estudados. Apresenta-se de seguida a análise dos resultados
relativos a cada corante, para todas as concentrações:
Cm-g: este corante apresentou um aumento global da predação nos três tempos de
amostragem de acordo com o aumento da concentração do corante. Por exemplo, à 1 hora as
médias de microesferas ingeridas nas concentrações de 5, 25, 50 e 100 ppm foram 7,90; 7,60;
11,63 e 10,67; respectivamente. Notou-se em ambos os ensaios uma ingestão de microesferas
à 1 hora na concentração de 100 ppm significativamente menor (p<0,05) do que a do controlo.
Cp-g: este corante revelou um decréscimo global da predação principalmente com o aumento
das concentrações até aos 50 ppm. Nesta concentração a ingestão de microesferas à 1 hora foi
significativamente menor (p<0,01) do que a do controlo. Para uma concentração de 100 ppm
houve uma estimulação aparente da ingestão de microesferas, dado que passou de um valor
de 7,23 para 11,60 microesferas ingeridas a uma concentração de 50 e de 100 ppm,
respectivamente.
Sm-g e Sm-s: estes corantes, de uma forma global, registaram um aumento da ingestão de
microesferas com o aumento da concentração, no entanto não se registaram diferenças
significativas relativamente ao controlo.
Sp-g: este corante apresentou um aumento da ingestão de microesferas com o aumento da
concentração até 100 ppm, sendo estas diferenças significativamente menores do que o
controlo para o tempo de amostragem de 48 horas, para as concentrações de 25 e 100 ppm, o
que pode significar um efeito mais tóxico naquela concentração.
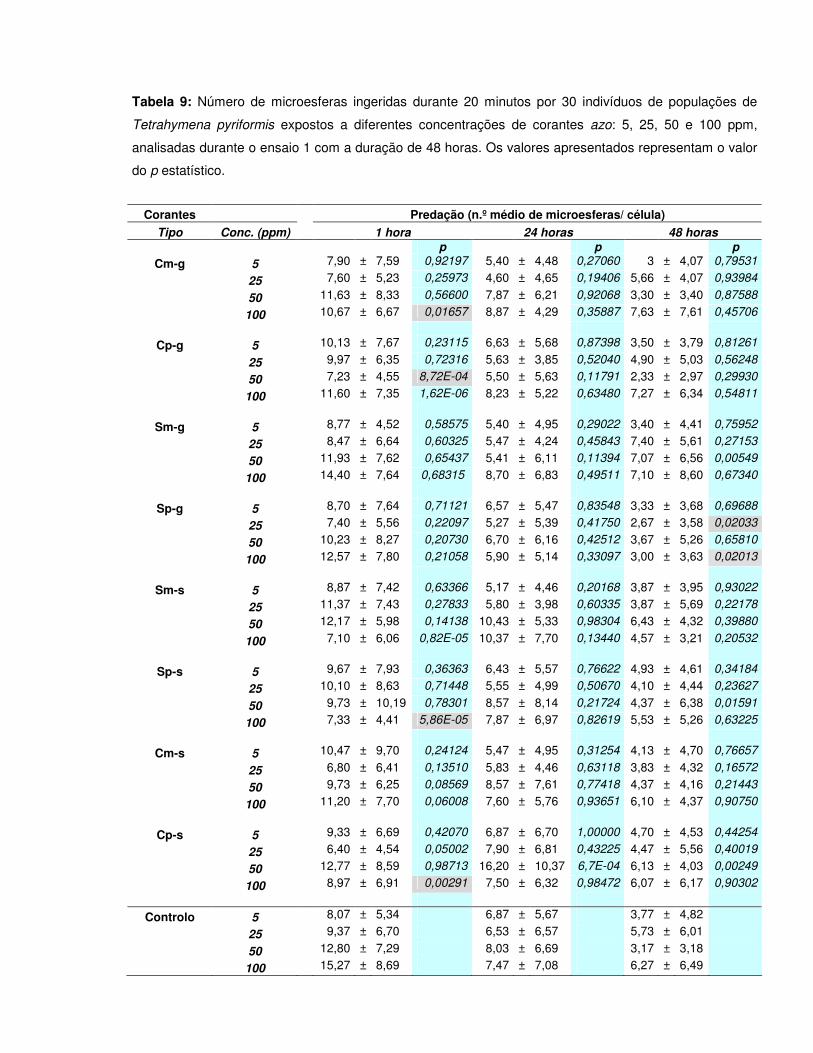
Tabela 9: Número de microesferas ingeridas durante 20 minutos por 30 indivíduos de populações de
Tetrahymena pyriformis expostos a diferentes concentrações de corantes azo: 5, 25, 50 e 100 ppm,
analisadas durante o ensaio 1 com a duração de 48 horas. Os valores apresentados representam o valor
do p estatístico.
Corantes Predação (n.º médio de microesferas/ célula)
Tipo Conc. (ppm) 1 hora 24 horas 48 horas p p p
Cm-g 5 7,90 ± 7,59 0,92197 5,40 ± 4,48 0,27060 3 ± 4,07 0,79531
25 7,60 ± 5,23 0,25973 4,60 ± 4,65 0,19406 5,66 ± 4,07 0,93984
50 11,63 ± 8,33 0,56600 7,87 ± 6,21 0,92068 3,30 ± 3,40 0,87588
100 10,67 ± 6,67 0,01657 8,87 ± 4,29 0,35887 7,63 ± 7,61 0,45706
Cp-g 5 10,13 ± 7,67 0,23115 6,63 ± 5,68 0,87398 3,50 ± 3,79 0,81261
25 9,97 ± 6,35 0,72316 5,63 ± 3,85 0,52040 4,90 ± 5,03 0,56248
50 7,23 ± 4,55 8,72E-04 5,50 ± 5,63 0,11791 2,33 ± 2,97 0,29930
100 11,60 ± 7,35 1,62E-06 8,23 ± 5,22 0,63480 7,27 ± 6,34 0,54811
Sm-g 5 8,77 ± 4,52 0,58575 5,40 ± 4,95 0,29022 3,40 ± 4,41 0,75952
25 8,47 ± 6,64 0,60325 5,47 ± 4,24 0,45843 7,40 ± 5,61 0,27153
50 11,93 ± 7,62 0,65437 5,41 ± 6,11 0,11394 7,07 ± 6,56 0,00549
100 14,40 ± 7,64 0,68315 8,70 ± 6,83 0,49511 7,10 ± 8,60 0,67340
Sp-g 5 8,70 ± 7,64 0,71121 6,57 ± 5,47 0,83548 3,33 ± 3,68 0,69688
25 7,40 ± 5,56 0,22097 5,27 ± 5,39 0,41750 2,67 ± 3,58 0,02033
50 10,23 ± 8,27 0,20730 6,70 ± 6,16 0,42512 3,67 ± 5,26 0,65810
100 12,57 ± 7,80 0,21058 5,90 ± 5,14 0,33097 3,00 ± 3,63 0,02013
Sm-s 5 8,87 ± 7,42 0,63366 5,17 ± 4,46 0,20168 3,87 ± 3,95 0,93022
25 11,37 ± 7,43 0,27833 5,80 ± 3,98 0,60335 3,87 ± 5,69 0,22178
50 12,17 ± 5,98 0,14138 10,43 ± 5,33 0,98304 6,43 ± 4,32 0,39880
100 7,10 ± 6,06 0,82E-05 10,37 ± 7,70 0,13440 4,57 ± 3,21 0,20532
Sp-s 5 9,67 ± 7,93 0,36363 6,43 ± 5,57 0,76622 4,93 ± 4,61 0,34184
25 10,10 ± 8,63 0,71448 5,55 ± 4,99 0,50670 4,10 ± 4,44 0,23627
50 9,73 ± 10,19 0,78301 8,57 ± 8,14 0,21724 4,37 ± 6,38 0,01591
100 7,33 ± 4,41 5,86E-05 7,87 ± 6,97 0,82619 5,53 ± 5,26 0,63225
Cm-s 5 10,47 ± 9,70 0,24124 5,47 ± 4,95 0,31254 4,13 ± 4,70 0,76657
25 6,80 ± 6,41 0,13510 5,83 ± 4,46 0,63118 3,83 ± 4,32 0,16572
50 9,73 ± 6,25 0,08569 8,57 ± 7,61 0,77418 4,37 ± 4,16 0,21443
100 11,20 ± 7,70 0,06008 7,60 ± 5,76 0,93651 6,10 ± 4,37 0,90750
Cp-s 5 9,33 ± 6,69 0,42070 6,87 ± 6,70 1,00000 4,70 ± 4,53 0,44254
25 6,40 ± 4,54 0,05002 7,90 ± 6,81 0,43225 4,47 ± 5,56 0,40019
50 12,77 ± 8,59 0,98713 16,20 ± 10,37 6,7E-04 6,13 ± 4,03 0,00249
100 8,97 ± 6,91 0,00291 7,50 ± 6,32 0,98472 6,07 ± 6,17 0,90302
Controlo 5 8,07 ± 5,34 6,87 ± 5,67 3,77 ± 4,82
25 9,37 ± 6,70 6,53 ± 6,57 5,73 ± 6,01
50 12,80 ± 7,29 8,03 ± 6,69 3,17 ± 3,18
100 15,27 ± 8,69 7,47 ± 7,08 6,27 ± 6,49

Sp-s: este corante registou uma ingestão de microesferas não consistente com o aumento da
concentração, nota-se que a ingestão relativamente à concentração não foi regular. No entanto,
para a concentração mais elevada (100 ppm) após 1 hora de exposição, a diferença na
ingestão de microesferas por Tetrahymena tinha sido significativamente diferente da ingestão
realizada pelas células do controlo no mesmo tempo de amostragem.
Cm-s: este corante, à semelhança do anterior, não apresentou uma ingestão de microesferas
consistente com o aumento da concentração, nota-se que a ingestão relativamente à
concentração não foi regular. De qualquer forma, para as concentrações e tempos de
amostragem efectuados não se registaram diferenças significativas entre os valores de ingestão
para as células tratadas quando comparadas com as células do controlo.
Cp-s: este corante, não apresentou igualmente uma predação concordante que aumentasse ou
diminuísse progressivamente com o aumento da sua concentração. Nota-se que a ingestão
relativamente à concentração não foi regular. No entanto, para a concentração mais elevada
(100 ppm) após 1 hora de exposição, a diferença na ingestão de microesferas pelas células
(8,97) tinha sido significativamente diferente da realizada pelas células do controlo (15,27) no
mesmo tempo de amostragem.
De uma forma geral, neste ensaio de predação consegue-se apurar que existem mais valores
significativamente mais baixos de ingestão de microesferas após 1 hora de exposição, nas
concentrações mais altas de corantes (100 ppm – 4 casos e 50 ppm – 1 caso), o que nos indica
um efeito inibitório da predação logo a pós a exposição ao corante. No entanto, como este
efeito não é significativamente diferente nos tempos de amostragem seguintes, 24 e 48 horas,
deduz-se que as células adaptaram-se ao corante e recuperaram a sua capacidade predatória
normal.
Quando se analisa a Tabela 10, relativa aos resultados do teste ANOVA, quanto à comparação
do parâmetro predação entre grupos de corantes de todas as concentrações: guaiacol-siringol
(g-s); carboxílico-sulfónico (C-S); meta-para (m-p), para os respectivos tempos de amostragem,
verifica-se que existem várias diferenças significativas dentro destes grupos, mas nunca em
simultâneo em dois ensaios realizados (ensaios 1 e 2). Verificam-se diferenças significativas
entre os seguintes grupos de corantes:
� grupo guaiacol-siringol (g-s) às 24 horas, no ensaio 1 e às 48 horas no ensaio 2, para
valores de p<0,01;

� grupo carboxílico-sulfónico (C-S) às 24 horas e às 48 horas no ensaio 2, para valores de
p<0,05.
Estes resultados da ANOVA não são significativos para concluir acerca da toxicidade entre
grupos de corantes.
Tabela 10: Resultados dos testes ANOVA para a predação de Tetrahymena pyriformis. A tabela faz a
comparação entre médias de grupos de corantes de todas as concentrações: guaiacol-siringol (g-s);
carboxílico-sulfónico (C-S); meta-para (m-p), para os respectivos tempos de amostragem. Os valores
apresentados representam o valor do p estatístico.
Grupos de Ensaio Predação
corantes 1 hora 24 horas 48 horas
g-s 1 0,37430 0,00026 0,78654
2 0,06157 0,42672 0,00267
C-S 1 0,36020 0,13578 0,58373
2 0,15224 0,03309 4,57E-05
m-p 1 0,73364 0,09951 0,20538
2 0,38224 0,34185 0,56802
7.4. Síntese dos Resultados
7.4.1. Síntese dos resultados para cada corante azo
� O corante Cm-g não possui toxicidade significativa relativamente aos parâmetros
avaliados. No ensaio de crescimento apenas se registaram duas diferenças
significativas (p<0,01) entre este corante e o controlo depois de 1 hora de exposição,
para uma concentração de 50 ppm e entre este corante e o controlo às 48 horas, para
uma concentração de 100 ppm (Figura 8 e Tabela 4). Na morfometria verificou-se um
aumento significativo da área (p<0,05) das células de Tetrahymena pyriformis
relativamente ao controlo (Figuras 9 e 10; e Tabela 6). Apresentou um aumento global
da predação nos três tempos de amostragem de acordo com o aumento da

concentração do corante. Notou-se em ambos os ensaios uma diminuição significativa
da ingestão de microesferas à 1 hora na concentração de 100 ppm relativamente ao
controlo (Tabela 9).
� O corante Cp-g inibiu significativamente o crescimento de Tetrahymena pyriformis às 24
e 48 horas, para as concentrações de 25 e 100 ppm. Para uma concentração de 50 ppm
o corante inicialmente inibiu o crescimento, mas depois a Tetrahymena pyriformis
conseguiu recuperar (Figura 8 e Tabela 4). Relativamente à morfometria, apresentou
apenas uma diferença significativa na área celular após 1 hora de exposição, para uma
concentração de 50 ppm. Verificou-se que houve uma diminuição significativa na razão
W/L relativamente ao controlo, indicando uma forma celular mais alongada (Figuras 9 e
10; e Tabela 6). Revelou um decréscimo global da predação principalmente com o
aumento das concentrações até aos 50 ppm. Nesta concentração a ingestão de
microesferas à 1 hora foi significativamente menor do que a do controlo. Para uma
concentração de 100 ppm houve uma estimulação aparente da ingestão de microesferas
(Tabela 9).
� O corante Sm-g revelou ser mais tóxico na concentração mais baixa (5 ppm), para os
ensaios de crescimento. Para as concentrações de 25 e 50 ppm apenas se registou
diferenças significativas em relação ao controlo após 1 hora de exposição, tendo a
população de Tetrahymena recuperado o crescimento. Para a concentração de 100 ppm
houve diferenças significativas em relação ao controlo, às 24 horas de exposição, tendo
a população posteriormente recuperado (Figura 8 e Tabela 4). Quanto à morfometria,
para uma concentração de 50 ppm, registou-se um aumento significativo na área celular,
após 1 hora de exposição, bem como uma diminuição significativa na razão W/L
relativamente ao controlo, indicando uma forma celular mais alongada do que a
considerada normal (Figuras 9 e 10; e Tabela 6).
� O corante Sp-g revelou valores de crescimento irregulares tanto superiores como
inferiores quando comparado com o controlo, em todas as concentrações (Figura 8 e
Tabela 4). Relativamente à morfometria e especificamente relativamente à área celular,

este corante registou já mais diferenças, tendo sido verificadas sempre às 48 horas de
exposição e para as concentrações de 25, 50 e 100 ppm. Quanto à razão W/L este
corante apresentou apenas uma diferença significativa após 1 hora de exposição, para
uma concentração de 50 ppm, indicando uma forma celular mais alongada do que a
apresentada pelas células do controlo (Figuras 9 e 10; e Tabela 6). Registou ainda um
aumento, embora não significativo da ingestão de microesferas com o aumento da
concentração (Tabela 9).
� O corante Sm-s revelou valores de crescimento irregulares tanto superiores como
inferiores quando comparado com o controlo, em todas as concentrações (Figura 8 e
Tabela 4). Relativamente ao parâmetro área celular este corante registou diferenças
significativas relativamente ao controlo, apenas para tempos de exposição de 48 horas,
nas concentrações de 25 e 100 ppm, mostrando que a área das células aumentou
relativamente à área das células do controlo (Figuras 9 e 10; e Tabela 6). Quanto à
predação verificou-se um aumento da ingestão de microesferas com o aumento da
concentração até 50 ppm, sem que se evidenciassem diferenças significativas
relativamente ao controlo. Quando na concentração de 100 ppm obteve uma ingestão
inferior à registada para 50 ppm, o que pode significar um efeito mais tóxico naquela
concentração máxima (Tabela 9).
� O corante Sp-s revelou valores de crescimento irregulares tanto superiores como
inferiores quando comparado com o controlo, em todas as concentrações (Figura 8 e
Tabela 4). Relativamente à área celular, parâmetro morfométrico, o corante registou
apenas um aumento significativo da área das células tratadas relativamente às células
do controlo, após 48 horas de exposição, para uma concentração de 5 ppm.
Relativamente à razão W/L verificou-se que na concentração mais elevada de 100 ppm,
às 48 horas, este corante provocou um arredondamento celular significativamente maior
ao registado para o controlo, o que indica toxicidade. Neste caso a toxicidade teve um
efeito diferente na forma das células, relativamente ao que foi registado para os
restantes corantes analisados (Figuras 9 e 10; e Tabela 6). Quanto à predação este
corante registou uma ingestão de microesferas variável e irregular, de acordo com o
aumento da concentração (Tabela 9).

� O corante Cm-s obteve sempre um crescimento inferior relativamente ao controlo; no
que respeita apenas à concentração de 100 ppm, conseguiu obter valores de
crescimento menores e significativamente diferentes do controlo (Figura 8 e Tabela 4).
Quanto à área celular este corante provocou na concentração de 25 ppm, às 48 horas,
um significativo alongamento das células comparativamente ao controlo (Figuras 9 e 10;
e Tabela 6). Relativamente à predação, não apresentou uma ingestão de microesferas
consistente e significativa com o aumento da concentração, nota-se que a ingestão
relativamente à concentração foi irregular (Tabela 9).
� O corante Cp-s provocou nos parâmetros crescimento e predação valores variáveis e
muito irregulares para todas as concentrações testadas e tempos de amostragem
(Figura 8 e Tabelas 4 e 9). Relativamente ao parâmetro área celular verificou-se um
aumento significativo da área de Tetrahymena pyriformis relativamente ao controlo, às
48 horas, para as concentrações de 50 e 100 ppm (Figuras 9 e 10; e Tabela 6).
7.4.2. Síntese dos resultados para os três testes realizados: crescimento, morfometria e predação.
No que diz respeito aos testes ANOVA realizados para todos os parâmetros, nos quais se
compararam médias de grupos de corantes de todas as concentrações: guaiacol-siringol (g-s);
carboxílico-sulfónico (C-S); meta-para (m-p), para os respectivos tempos de amostragem
registou-se o seguinte:
1. No parâmetro crescimento (Tabela 5) os resultados mostram que apenas no ensaio 1
realizado se verificou uma diferença significativa entre os grupos de corantes com
guaiacol e siringol.
2. No parâmetro morfometria (área e razão W/L) verifica-se que existem várias diferenças
significativas dentro destes grupos em ambos os ensaios realizados (ensaios 1 e 2)
(Tabelas 6, 7 e 8):
� quanto à área de Tetrahymena pyriformis verificam-se diferenças significativas à 1
hora nos grupos guaiacol-siringol (g-s) e meta e para (m-p);

� quanto à razão W/L nota-se que existem diferenças significativas entre os seguintes
grupos de corantes: guaiacol-siringol (g-s) às 48 horas; carboxílico-sulfónico (C-S) à
1 hora; e meta e para (m-p) em ambos os tempos de amostragem;
� analisando os dois parâmetros (área e razão W/L) simultaneamente nota-se que os
efeitos tóxicos dentro do grupo carboxílico-sulfónico (C-S) são significativamente
mais evidentes após uma exposição inicial a estes corantes e que os efeitos tóxicos
dentro do grupo meta e para (m-p) são significativamente diferentes durante todo o
tempo de exposição, particularmente após 1 hora de exposição das células aos
corantes (Tabelas 6, 7 e 8).
3. No parâmetro predação verificou-se que a ingestão de microesferas foi mais baixa após
1 hora exposição, nas concentrações de 50 e 100 ppm, indicando um efeito inibitório da
predação logo após a exposição ao corante. Às 24 e 48 horas, registou-se a ausência
de inibição o que indica uma possível adaptação celular ao corante. Quanto à
comparação da predação entre grupos de corantes de todas as concentrações:
guaiacol-siringol (g-s); carboxílico-sulfónico (C-S); meta e para (m-p) para os respectivos
tempos de amostragem, verifica-se que existem várias diferenças significativas dentro
destes grupos, mas nunca em simultâneo em dois ensaios realizados (ensaios 1 e 2).
Desta forma, e como foi dito anteriormente, os resultados do teste ANOVA não são
significativos para concluir acerca da toxicidade entre estes grupos de corantes.

8. DISCUSSÃO

O estudo da toxicidade de corantes azo é importante uma vez que estes correspondem a mais
de metade dos corantes anualmente produzidos (STOLZ, 2001; NOVOTNÝ et al., 2006);
representam o maior e mais versátil grupo cuja aplicação da indústria têxtil ascende até aos 70
%; e são compostos recalcitrantes e resistentes à biodegradação, que podem ser libertados
para o meio ambiente através dos efluentes industriais, provocando vários problemas
ambientais no meio aquático receptor (FERREIRA, 1998; NOVOTNÝ et al., 2006).
Moawad et al (2003) referiu que estudos para avaliar a toxicidade dos corantes têxteis
realizados por Sharma & Sobti (2000), Rajaguru et al (2001), Wang et al (2002), concluíram que
os efeitos tóxicos dependiam muito da concentração e que todos os corantes são tóxicos em
concentrações mais elevadas.
Salienta-se o facto de terem sido empregues quatro concentrações 5, 25, 50 e 100 ppm de
corantes azo e nenhuma delas ter tido notórios efeitos citotóxicos. Estas concentrações foram
empregues tendo em vista duas razões:
� o facto de os corantes azo presentes em produtos téxteis não poderem libertar aminas
numa concentração acima dos 30 ppm ou dos 70 ppm, caso os artigos sejam feitos de
fibras recicladas, de acordo com a Directiva Europeia 2004/21/ EC implementada em
Portugal pelo Decreto Lei n.º 208/2003 (única legislação existente que indica valores de
concentração permitidos, relativos a corantes azo). As concentrações de 30 e 70 ppm
estão abrangidas pelas utilizadas neste estudo e por este facto pode-se inferir quanto à
toxicidade destes corantes para esta gama de concentrações;
� a necessidade de conhecer os efeitos tóxicos sub-letais de corantes azo, em células
eucarióticas, em concentrações mais baixas particularmente inferiores a 30 ppm.
Os resultados relativos à toxicidade em todos os parâmetros analisados são globalmente
irregulares e heterogéneos, o que pode ser explicado por várias razões:
� as concentrações utilizadas, por serem concentrações sem efeitos tóxicos muito
notórios, tiveram implicações durante a fase de tratamento estatístico dos resultados,
uma vez que se registaram diferenças entre os dois ensaios realizados para cada
parâmetros testado. Assim sendo, decidiu-se a análise dos resultados estatísticos
semelhantes para ambos os ensaios, o que se revelou inconclusivo relativamente à
toxicidade dos corantes quando comparados com o controlo;

� as condições das culturas celulares utilizadas para os testes de toxicidade variarem ao
longo do tempo, tendo favorecido o crescimento em alguns ensaios em detrimento de
outros;
� o facto de se ter trabalhado com concentrações baixas que podem corresponder a
limiares de toxicidade e, assim sendo, os resultados são muito heterogéneos. E nestes
casos as diferentes respostas dependem sobretudo do estado fisiológico individual dos
organismos teste, podendo haver indivíduos mais resistentes do que outros; o aumento
das concentrações dos tóxicos causaram uma inibição ou estimulação das respostas
fisiológicas de T. pyriformis;
� a variação da densidade inicial da cultura, embora ténue, pode ter afectado a taxa de
crescimento devido a limitação de espaço e/ou alimento e ter influenciado na resposta
fisiológica aos tóxicos.
Nos estudos toxicológicos apenas um bioensaio não é o suficiente para dar uma imagem global
dos efeitos num organismo teste. Desta forma, esta abordagem utilizando uma bateria de três
testes toxicológicos fisiológicos, permitiu uma recolha de informação mais abrangente acerca da
toxicidade de corantes têxteis azo, em Tetrahymena pyriformis. Os ensaios bioquímicos são
importantes como complemento aos ensaios fisiológicos, desta forma o conteúdo em ATP, a
actividade da ACP e a redução do MTT podiam ter representado boas alternativas aos
tradicionais testes toxicológicos (NICOLAU et al.. 2001). No entanto, não foram utilizados neste
estudo, uma vez que as soluções que contém os corantes são coloridas, interferindo assim com
os resultados destes testes colorimétricos.
O parâmetro no qual, quando comparadas todas as concentrações, se notou um aumento
global da toxicidade de todos os corantes, foi o crescimento. Inicialmente, optou-se por realizar
a contagem de células ao microscópio; no entanto verificou-se que esta era muito demorada e
sujeita a erros (especialmente quando se trabalha com concentrações celulares elevadas)
(TEUNISSON, 1971), e os resultados obtidos foram inconclusivos e muito díspares entre
ensaios para a mesma concentração. Assim, optou-se por realizar as contagens celulares
relativas aos ensaios de crescimento através do contador automático de células, o que
melhorou os resultados obtidos, uma vez que para uma dada concentração, corante e tempo de
amostragem não havia discrepância nas médias das sub-amostras realizadas. Algumas
discrepâncias na contagem deveram-se possivelmente à sedimentação das células em
suspensões mais densas durante as contagens.

A morfometria foi, dos métodos utilizados, o mais sensível na determinação da toxicidade entre
grupos de corantes, uma vez que os resultados foram semelhantes em ambos os ensaios
realizados e se registaram bastantes diferenças dentro dos grupos de corantes siringol-
guaiacol, carboxílico-sulfónico e meta-para, para os dois tempos de amostragem (1 e 48 horas).
O menos sensível foi o parâmetro crescimento uma vez que apenas se registou uma diferença
e apenas num dos ensaios realizados para este parâmetro.
Os resultados apresentados por este trabalho extrapolam que os protozoários desempenham
um papel muito importante na predação e no controle da densidade populacional de bactérias
(COX et al., 1999; HAHN et al., 2001). Estes corantes não apresentam uma toxicidade evidente,
portanto, podem não colocar em perigo a sobrevivência dos protozoários e a eficácia de todo o
processo de tratamento de águas residuais industriais.
Ao contrário do que refere Nicolau et al. (2001), os ensaios de predação foram os mais
trabalhosos e demorados. No início da avaliação deste parâmetro considerou-se a hipótese de
realizar a predação através do método automático (com análise de imagem) de acordo com
Dias et al. (2003); no entanto verificou-se que este método seria ainda mais moroso e
complicado, para o estudo de uma grande gama de corantes e de concentrações. Em
contrapartida, como foram contadas as microesferas a olho nú, pode ter ocorrido subestimação
no número de microesferas devido principalmente à sua intensa fluorescência e ao cansaço
ocular após muito tempo seguido de contagens ao microscópio (DIAS et al., 2003). A fagocitose
nos protozoários é uma função celular clássica, reflectindo o nível de energia do protozoário. A
diminuição da fagocitose pela Tetrahymena pode ser explicada devido ao decréscimo de
energia nestes protozoários, que está relacionada com a energia da própria célula
(STEFANIDOU et al., 1999). Assim, os casos em que houve uma significativa diminuição da
predação, devido à influência de um tóxico, mostram que as células diminuíram a sua energia e
o investimento desta na capacidade fagocítica foi muito menor.
Em todos os ensaios realizados não podemos isolar um efeito tóxico característico de
determinado corante ou grupo de corantes. Relativamente ao crescimento, os corantes que
registaram mais diferenças significativas relativamente aos controlos foram o Cp-g, o Sm-g e
Sp-s. Alguns corantes estimularam a predação quando em concentrações mais elevadas (Sp-s,
Cp-s, Cp-g, Cm-g, Sm-s) e de uma forma global aumentaram a área celular - parâmetro
morfométrico - de acordo com o aumento da concentração dos corantes (Sp-g, Sm-s, Cp-s, Cm-
g). Nestes ensaios apurou-se a toxicidade de um corante individualmente e o ideal seria a

avaliação da toxicidade da mistura de corantes, o que realmente acontece em ambientes
aquáticos sujeitos a descargas de efluentes industriais têxteis.
A hormesis é fenómeno de relação dose-resposta caracterizado pela estimulação em doses
baixas e pela inibição a doses elevadas. Foi já observada em vários estudos e é conhecida por
ser independente do agente químico, do modelo biológico e/ou do parâmetro toxicológico
usado. É ainda um conceito ainda pouco apreciado e reconhecido, que pode mudar a forma
como a toxicologia é estudada/ abordada. Esta teoria explora o facto da natureza fundamental
da dose resposta não ser linear mas em forma de U. O princípio por trás destas respostas
implica que baixas concentrações de algumas substâncias tóxicas podem produzir um efeito
benéfico quando em doses baixas (CALABRESE, 2002; CALABRESE & BALDWIN, 2003).
A estimulação é observada (nem sempre) seguindo uma resposta inicial inibitória. Dependendo
do parâmetro utilizado, a resposta hormética é muitas vezes caracterizada por ser um U
invertido (no caso do crescimento ou sobrevivência), um J (no caso de doença). Estas
respostas toxicológicas em forma de U são baseadas em estudos reais e podem ser
generalizadas. Foram já descritos 5000 exemplos de respostas horméticas independentes de
classes de substâncias químicas, modelo biológico ou testes toxicológicos utilizado
(CALABRESE, 2002; CALABRESE & BALDWIN, 2003).
Tendo por base esta teoria podemos verificar que em todos os ensaios realizados existem, para
cada corante, respostas toxicológicas que graficamente adoptam com a forma de U normal, de
U invertido ou de J. Estas respostas são mais evidentes nos ensaios de morfometria e de
predação. Relativamente à morfometria os resultados obtidos, ou seja, o alongamento celular
como resposta à toxicidade dos corantes pode ser entendido como uma resposta benéfica e
portanto hormética. No que diz respeito à predação podemos verificar que a resposta das
células ao aumento da concentração de cada corante é muito diferente. Podemos verificar que
por exemplo, relativamente ao corante Cp-g e Cm-s houve uma estimulação da predação na
concentração de 5 ppm, uma diminuição deste parâmetro nas concentrações intermédias (25 e
50 ppm) e novamente a estimulação da predação na concentração mais elevada – 100 ppm.
Estas respostas podem ser consideradas horméticas, na medida que correspondem a um
gráfico em forma de U, tendo em conta o aumento a concentração do corante. Da mesma forma
o corante Sm-s apresenta uma resposta hormética, mas neste caso em forma de U invertido,
para a mesma gradação de concentrações.
Apesar da utilização de protistas, em especial de ciliados, ter ganho um interesse crescente por
muitos investigadores, não foram encontradas muitas referências da sua utilização na avaliação

da toxicidade de corantes azo, utilizando uma bateria de testes toxicológicos. Assim, este
estudo colmata um pouco essa falta de dados.
Não é possível afirmar que a Tetrahymena pyriformis é especialmente sensível a um
determinado grupo de substâncias. Tal como os outros modelos biológicos, os efeitos
toxicológicos induzidos por substâncias orgânicas e inorgânicas variam imenso, nomeadamente
valores de IC50 dependendo também das suas características físico-químicas e dos seus
mecanismos de acção (SAUVANT, 1999). Desta forma, este ciliado poderá ter-se revelado mais
resistente do que outro organismo modelo para o tipo de corantes utilizados, não se notando
evidentes efeitos tóxicos.

9. CONCLUSÕES

A Tetrahymena pyriformis é uma célula eucariótica, de fácil e rápido crescimento em condições
axénicas e revelou-se ser um modelo ideal para este estudo ecotoxicológico. A maioria dos
estudos realizados apontam para a validação deste protozoário como um modelo, e concluem
que a Tetrahymena pyriformis pode ser considerada como um complemento ou alternativa a
outros modelos animais na investigação toxicológica. (SAUVANT et al., 1999). Este trabalho
reforça a importância dos ciliados como organismos bioindicadores e estimula a utilização do
protozoário Tetrahymena pyriformis como uma ferramenta na avaliação da toxicidade de
substâncias novas ou já existentes, em meio aquático.
Embora não se possa afirmar seguramente que a Tetrahymena pyriformis é especialmente
sensível a um determinado grupo de substâncias, através deste estudo pode-se concluir que
este ciliado é mais resistente aos efeitos tóxicos dos corantes azo utilizados, pelo que seria
importante a realização de um futuro estudo similar utilizando outros organismos testes, de
forma a comprovar a similaridade das respostas tóxicas.
Nos estudos toxicológicos apenas um bioensaio não é o suficiente para dar uma imagem global
dos efeitos num organismo teste. Desta forma, esta abordagem utilizando uma bateria de três
testes toxicológicos fisiológicos, permitiu uma recolha de informação mais abrangente acerca da
toxicidade de corantes azo, em Tetrahymena pyriformis. Espera-se que esta combinação de
ensaios possa representar uma forma sensível de testar tipos similares de agentes, bem como
o seu possível efeito em sistemas alvo de uma grande variedade de organismos. No entanto,
seria importante, no futuro, realizar trabalhos nos quais se pudesse ampliar a bateria de testes
de modo a comprovar os resultados obtidos nestes estudos de toxicidade.
Os ensaios de crescimento mais frequentemente usados para determinar a toxicidade sub-letal
realizados através da utilização de um contador automático de células revelaram-se ser muito
mais fiáveis do que os mesmos realizados através da contagem ao microscópio. De facto, o
ensaio de crescimento foi o menos demorado, uma vez que foi utilizado um contador de células.
O crescimento é um ensaio de fácil reprodução, não requerendo experiência técnica ou
despesas operacionais (NICOLAU et al., 1999). Foi utilizado o equipamento Beckman Coulter Z
Series, para as contagens celulares relativas ao crescimento. Embora este aparelho seja
utilizado para determinar em laboratório a concentração de eritrócitos, leucócitos e plaquetas de
humanos ou de animais, pode ser calibrado para protozoários, como a Tetrahymena pyriformis,
revelando ser uma ferramenta essencial nos ensaios de crescimento.

Pode-se concluir igualmente que os ensaios de predação com Tetrahymena pyriformis, ao
contrário do referido por Nicolau et al. (1999; 2001), foram os mais trabalhosos e demorados.
Seriam ainda mais morosos se tivessem sido realizados através do método automático - com
análise de imagem. Esta conclusão retirou-se do facto do presente estudo utilizar uma grande
gama de corantes (oito no total) e de concentrações (quatro no total).
Todos estes aspectos levam a concluir que a miniaturização dos testes de toxicidade e a
utilização dos protozoários ciliados constituem uma boa alternativa a outros testes de toxicidade
mais laboriosos e dispendiosos.
Com este trabalho podemos concluir que quando se utilizam concentrações baixas de tóxicos,
podemos ter respostas fisiológicas dos organismos modelo que podem ser entendidas, na
toxicologia tradicional, como irregulares e não conclusivas. No entanto, tendo em conta a teoria
da hormesis, podemos concluir que muitas das respostas deste estudo, nomeadamente no que
diz respeito ao parâmetro predação, são caracterizadas por adoptar a forma de U normal, de U
invertido ou de J. Estas respostas podem ser consideradas horméticas tendo em conta o
aumento a concentração do corante, as respostas obtidas e as concentrações baixas utilizadas.
As águas residuais de indústrias têxteis constituem uma ameaça para o ambiente em várias
partes do mundo (NILSSON et al., 2006). Os corantes têxteis são resistentes à degradação e
causam poluição visual. São perigosos porque podem manter no meio por períodos de tempo
prolongados podendo ter uma acção tóxica, mutagénica e cancerígena nos seres vivos
aquáticos. Os corantes azo são considerados recalcitrantes à degradação microbiológica mas
pouco se sabe acerca da sua toxicidade. A avaliação da ecotoxicidade dos corantes é
importante para ajudar a reduzir o efeito perigoso destes químicos (MOAWAD et al., 2003).
Desta forma, o trabalho realizado foi importante e inovador, uma vez é o primeiro e único estudo
sobre a toxicidade deste tipo de corantes têxteis azo utilizados em organismos teste. Até à data
apenas foram realizados estudos sobre a biodegradação destes corantes azo por fungos
linholíticos.
Seria ainda importante realizar futuras investigações com os mesmos corantes têxteis, no
entanto, noutras vertentes das quais se enumeram as seguintes: 1) a utilização de uma gama
mas alargada de concentrações superiores; 2) a realização da combinação de vários corantes
de forma a estudar os efeitos tóxicos das misturas; 3) a aplicação/ adequação, como já foi
referido, de mais testes toxicológicos diversificados (não apenas com carácter fisiológico); e 4) a

propensão destes corantes para a formação de aminas e a avaliação do efeito cancerígeno e
mutagénico destas sobre as células de um organismo bioindicador, como a Tetrahymena
pyriformis.

10. REFERÊNCIAS BIBLIOGRÁFICAS

BONNET, J.L.; GUIRAUD, P.; DUSSER, M.; KADRI, M.; LAFFOSSE, J.; STEIMAN, R. &
BOHATIERA, J. (2005) Assessment of anthracene toxicity toward environmental eukaryotic
microorganisms: Tetrahymena pyriformis and selected micromycetes main taxonomic groups.
Ecotoxicology and environmental safety. 60: 87-100.
CALABRESE, E. J. & BALDWIN, L. A. (2003) HORMESIS: The Dose-Response Revolution
Annu. Rev. Pharmacol. Toxicol. 2003. 43:175–97
CALABRESE, E. J. (2002) Reflections in Mutation Research - Hormesis: changing view of the
dose-response, a personal account of the history and current status. Mutation Research. 511:
181–189.
COX H. H. J., NGUYEN T. T. & DESHUSSES M. A. (1999) Predation of bacteria by the
protozoa Tetrahymena pyriformis in toluente-degrading cultures. Biotechnology Letters. 21: 235-
239.
DAYEH. V. R.; GROMINSKY S.; DEWITTE-ORR S. J.; SOTORNIK D.; YEUNG C. R.; LEE
L.E.J.; LYNN D. H. & BOLS, N. C. (2005) Comparing a ciliate and a fish cell line for their
sensitivity to several classes of toxicants by the novel application of multiwell filter plates to
Tetrahymena. Research in Microbiology. 156: 93-103.
DIAS N. (1998) Aplicação de Técnicas Fluorescentes no Estudo de Estruturas Biológicas de
Tetrahymena pyriformis. Tese de Mestrado em Engenharia Biológica. Universidade do Minho.
DIAS N., AMARAL A. L., FERREIRA, E. C. & LIMA N. (2003) Automated image analysis to
improve bead ingestion toxicity test counts in the protozoan Tetrahymena pyriformis. Letters in
Applied Microbiology. 37: 230-233.
DIAS N. & LIMA N. (2002) A comparative study using a fluorescence-based and a direct-count
assay to determine cytotoxicity in Tetrahymena pyriformis. Research in Microbiology. 153: 313-
322.
DIAS N., MORTARA R. & LIMA N. (2003) Morphological and physiological changes in
Tetrahymena pyriformis for the in vitro cytotoxicity assessment of Triton X-100. Toxicology in
Vitro. 17: 357-366.

DIAS, N., NICOLAU, A., CARVALHO, G.S., MOTA, M. & LIMA, N., (1999) Miniaturization and
application of the MTT assay to evaluate metabolic activity of protozoa in the presence of
toxicants. Journal of Basic Microbiology. 392: 103–108.
Directive 2002/61/EC of the European Parliament and of the Council. Official Journal of the
European Communities, L243/16 - L243/16.
FERREIRA I. (1998) Síntese e biodegradação de corantes azo pelo Phanerochaete
chrysosporium. Tese de Mestrado em Ciências do Ambiente. Universidade do Minho.
HAHN M. W. & HÖFLE M. G. (2001). MiniReview: Granzing of protozoa and its effect on
populations of aquatic bacteria. FEMS Microbiology Ecology. 35: 113-121.
LARSON, J.; SCHULTZ, T. W.; RASMUSSEN, L.; HOOFTMAN, R. & PAULI, W. (1997)
Progress in an ecotoxicological standard protocol with protozoa: results from a pilot ringtest with
Tetrahymena pyriformis. Chemosphere. 35: 1023-1041.
LE DÛ A. (1993) Modulation of copper toxicity by the chemical parameters of river water
Fresenius. Environmental. 2: 629-634.
MADONI P. (2000) The acute toxicity of nickel to freshwater ciliates. Environmental Pollution.
109: 53-59.
MARTINS M.; FERREIRA, I. C.; SANTOS, I. M.; QUEIROZ & LIMA N. (2001) Biodegradation of
bioacessible textile dyes by Phanerochaete chrysosporium. Journal of Biotecnology. 86: 91-98.
MARTINS M.; QUEIROZ M; SILVESTRE A. & LIMA N. (2002) Relationship of chemical
structures of textile dyes on the pre-adaptation medium and the potentialities of their
biodegradation by Phanerochaete chrysosporium. Research in Microbiology. 153: 361-368.
MARTINS M.; LIMA N.; SILVESTRE A. & QUEIROZ M. (2003) Comparative studies of fungal
degradation of single or mixed bioacessible reactive azo dyes. Chemosphere. 52: 967-973.

MITCHELL E., BURGESS J. & STUETZ R. (2002) Review: Developments in ecotoxicity testing.
Reviews in Envirnomental Science & Biotecnology. 1: 169-198.
MOAWAD, H.; ABD EL-RAHIM, W. M. & KHALAFALLAH, M. (2003) Evaluation of biotoxicity of
textile dyes using two bioassays. Journal of Basic Microbiology. 43: 218-229.
NICOLAU A., DIAS N., MOTA M. & LIMA N. (2001) Mini-review: Trends in the use of protozoa in
the assessment of wastewater treatment. Research in Microbiology. 152: 621-630.
NICOLAU A., MOTA M. & LIMA N. (1999) Physiological responses of Tetrahymena pyriformis
to copper, zinc, cycloheximide and Triton X-100. FEMS Microbiology Ecology. 30: 209-216.
NICOLAU A., MOTA M. & LIMA N. (2004) Effect of different toxic compounds on ATP content
and acid phosphatase activity in axenic cultures of Tetrahymena pyriformis. Ecotoxicology and
Environmental Safety. 57: 129-135.
NILSSON J. R. (1981) Effects of copper on phagocytosis in Tetrahymena. Eur. J. Protistol.. 25:
2-25.
NILSSON J. R. (1989) Tetrahymena in Citotoxicology: with special reference to effects of heavy
metals and selected drugs. Eur. J. Protistol.. 25: 2-25.
NILSSON, I.; MÖLLER A.; MATTIASSON, B.; RUBINDAMAYUGI M. S. T. & WELANDER U.
(2006) Decolorization of synthetic and real textile wastewater by use of white-rot fungi. Enzyme
and Microbial Technology. 38: 94-100.
NOVOTNÝ C., DIAS N., KAPANEN A., MALACHOVÁ K., VÁNDROVCOVÁ M., ITÄVAARA M. &
LIMA N. (2006) Comparative use of bacterial, algal and protozoan tests to study toxicity of azo-
and anthraquinone dyes. Chemosphere. 63: 1436-1442.
Policy Research Center for Environment and Economy (1999) The 3rd Meeting of the 2nd
Phase of CCICED Impacts of Environmental Standards and Requirements in EU Countries on
China’s Textile Industry.

REN, S. (2003) Ecotoxicity prediction using mechanism and non-mechanism based QSAR: a
preliminary study. Chemosphere. 53: 1053-1065.
ROBINSON, T.; MCMULLAN, G.; MARCHANT, R. & NIGAM, P. (2001) Review paper:
Remediation of dyes in textile effluent: a critical review on current treatment technologies with a
proposed alternative. Bioresource Technology. 77: 247-255.
SAUVANT, M.P.; PEPIN D.; BOHATIER, J. & GROLIERE C.A. (1995) Microplate Tecnique for
Screening and Assessing Cytotoxicity of Xenobiotics with Tetrahymena pyriformis. Ecotoxicity
and Envirnomental Safety. 32: 159-165.
SAUVANT, M. P.; PEPIN, D. & PICCINNI E. (1999) Tetrahymena pyriformis: a tool for
toxicological studies - a review. Chemosphere. 38: 1631-166.
SEWARD, J. R.; SINKS, G. D. & SCHULTZ, T. W. (2001) Reproductibility of toxicity across
mode of toxic action in the Tetrahymena population growth impairment assay. Aquatic
Toxicology. 53: 33–47.
STEFANIDOU, M.; ALEVISOPOULOS, C.; MARAVELIAS, C.; LOUTSIDIS, C. & KOUTSELINIS,
A. (1999) Phagocytosis of the protozoon Tetrahymena pyriformis as an endpoint in the
estimation of cocaine salt and cocaine freebase toxicity. Addiction Biology. 4 449-452.
STOLZ A.( 2001) Basic and applied aspects in the microbial degradation of azo dyes. Applied
Microbiology Biotechnology. 56: 69–80.
TEUNISSON, D. J. (1971) Elutriation and Coulter Counts of Tetrahymena pyriformis Grown in
Peanut and Cottonseed Meal Media. Applied Microbiology. 21: 878-887.
ZILBERG D. & SINAI T. (2006) Optimization and validation of a colorimetric assay for
Tetrahymena sp. Survival. Research in Microbiology. 157: 355-359.
ZSUZSA D., MADARÁSZ B. & LASZLO V. (1999) Study of histamine effects on phagocytosis
and enzyme secretion of Tetrahymena pyriformis. Acta Biologica Hungarica. 50: 325-334.
Top Related