






![5HIHUrQFLDV %LEOLRJUiILFDV - PUC-Rio · BALLOU R.H. Business Logistics Management: 3ODQQLQJ 2UJDQL]LQJ DQG &RQWUROOLQJ WKH 6XSSO\ &KDLQ. Prentice-Hall, 1999. ...](https://static.fdocumentos.com/doc/165x107/5b5f072b7f8b9a6d448d81a1/5hihurqfldv-leolrjuiilfdv-puc-ballou-rh-business-logistics-management.jpg)
Línguas
Páginas
Legal


Populações em paisagens fragmentadas: da teoria das ilhas aos modelos atuais de
metapopulação

1. A teoria da biogeografia de ilhas
2. A teoria de metapopulações
Principais tópicos

A teoria da biogeografia das ilhas(MacArthur, R.H. and Wilson, E.O. 1967. The theory of island
biogeography. Princeton Univ. Press. Ed., Princeton)
Esta teoria estava baseada em três observações:
1. Comunidades insulares são mais pobres em espécies do que as comunidades continentais equivalentes (Sc > S1);
2. Esta riqueza aumenta com o tamanho da ilha;3. Esta riqueza diminui com o aumento do isolamento da ilha.
S2
S3 S1
S4
Sc
(S1 > S2)(S3 > S4)
(S1 > S3)
(S2 > S4)

A teoria da biogeografia das ilhas(MacArthur, R.H. and Wilson, E.O. 1967. The theory of island
biogeography. Princeton Univ. Press. Ed., Princeton)
Esta teoria estava baseada também numa premissa:
1. As ilhas não funcionam como um sistema fechado:
S2
S3 S1
S4
Sc
(S1 > S2)(S3 > S4)
(S1 > S3)
(S2 > S4)

A teoria da biogeografia das ilhas(MacArthur, R.H. and Wilson, E.O. 1967. The theory of island
biogeography. Princeton Univ. Press. Ed., Princeton)
Existe um equilíbrio dinâmico entre extinção e imigração
Se
Tax
a de
imig
raçã
o(e
spéc
ies/
tem
po)
Tax
a de
ext
inçã
o(e
spéc
ies/
tem
po)
Chegada de uma espécie nova na ilha
Número de espécies0 P

A teoria da biogeografia das ilhas(MacArthur, R.H. and Wilson, E.O. 1967. The theory of island
biogeography. Princeton Univ. Press. Ed., Princeton)
Existe um equilíbrio dinâmico entre extinção e imigração
Se
Tax
a de
imig
raçã
o(e
spéc
ies/
tem
po)
Tax
a de
ext
inçã
o(e
spéc
ies/
tem
po)
Espécies com maior capacidade de locomoção chegam mais rapidamente
Número de espécies0 P
Predação e competição aumentam o risco de
extinção

A teoria da biogeografia das ilhas(MacArthur, R.H. and Wilson, E.O. 1967. The theory of island
biogeography. Princeton Univ. Press. Ed., Princeton)
Este equilíbrio dinâmico depende da área da ilha.
Segrd
Tax
a de
imig
raçã
o(e
spéc
ies /
tem
po)
Tax
a de
ext
inçã
o(e
spéc
ies /
tem
po)
Número de espécies0 PSepq
grandepequena
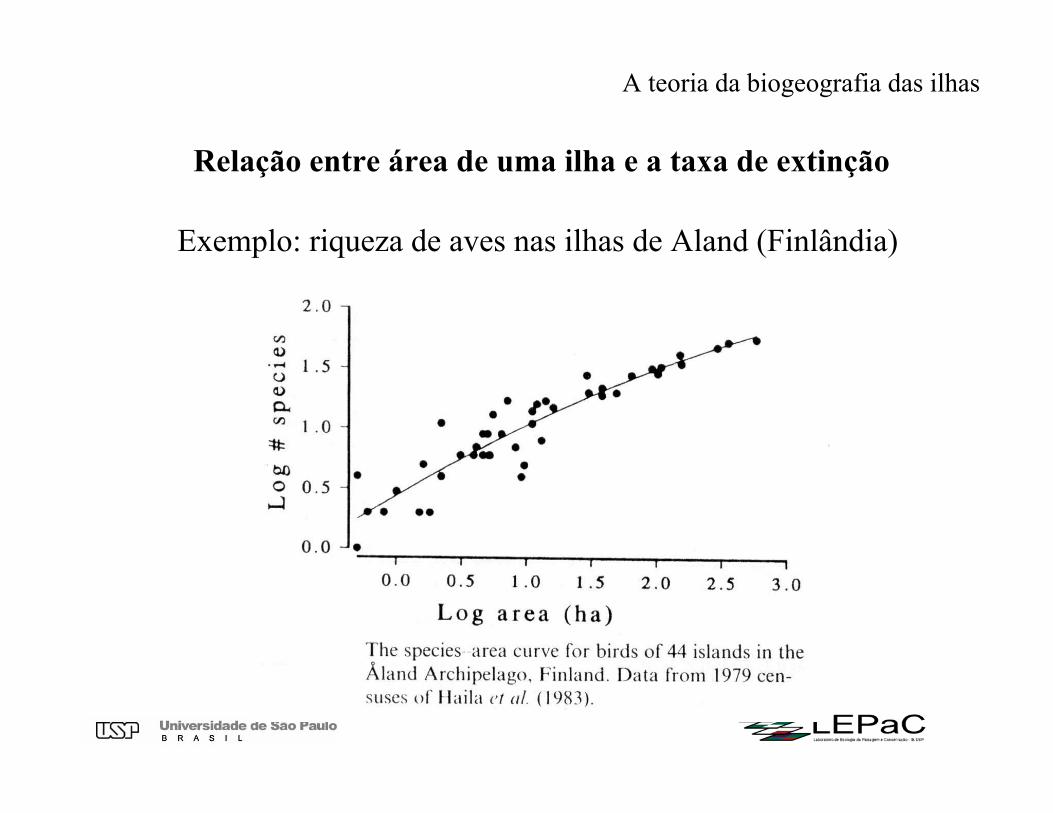
Relação entre área de uma ilha e a taxa de extinção
A teoria da biogeografia das ilhas
Exemplo: riqueza de aves nas ilhas de Aland (Finlândia)
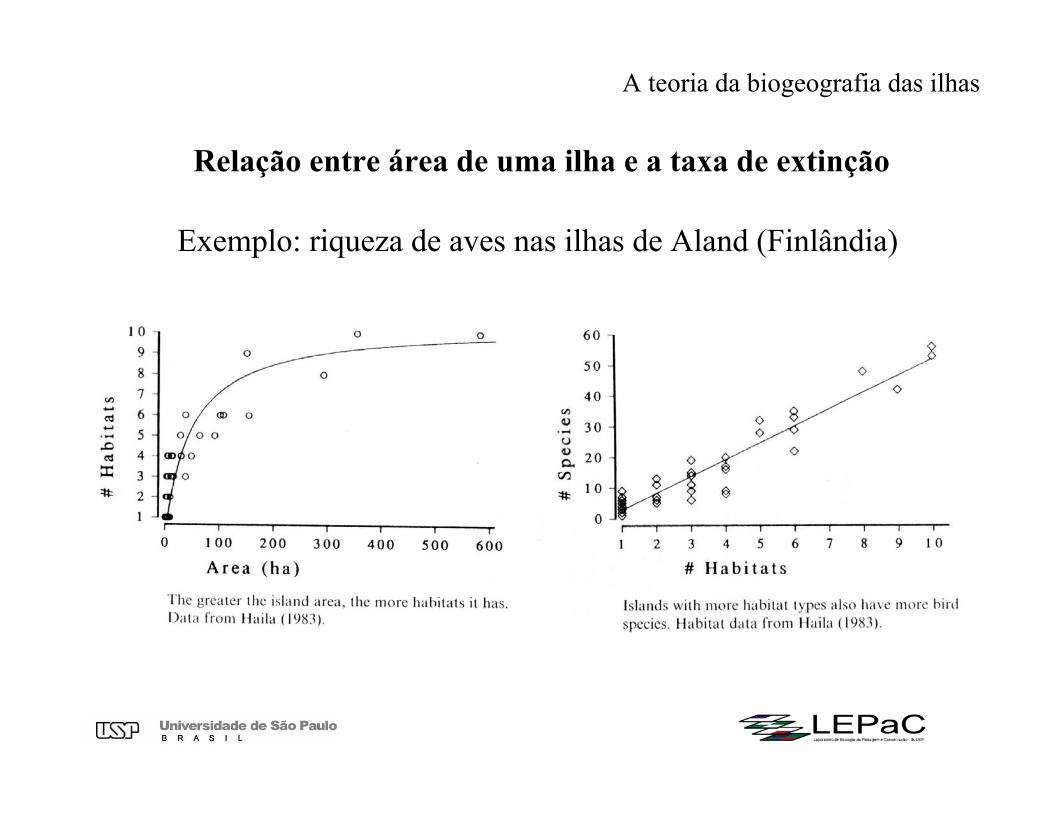
Relação entre área de uma ilha e a taxa de extinção
A teoria da biogeografia das ilhas
Exemplo: riqueza de aves nas ilhas de Aland (Finlândia)

• Área da ilha < Área mínima necessária para a sobrevivência de uma determinada população;
• Redução da heterogeneidade do habitat;• Intensificação das competições inter e intra específicas
devido à escassez de recursos;• Extinções secundárias, devido ao desaparecimento de
espécies-chave;• Aumento dos riscos de extinções estocásticas.
Relação entre área de uma ilha e a taxa de extinção
A teoria da biogeografia das ilhas
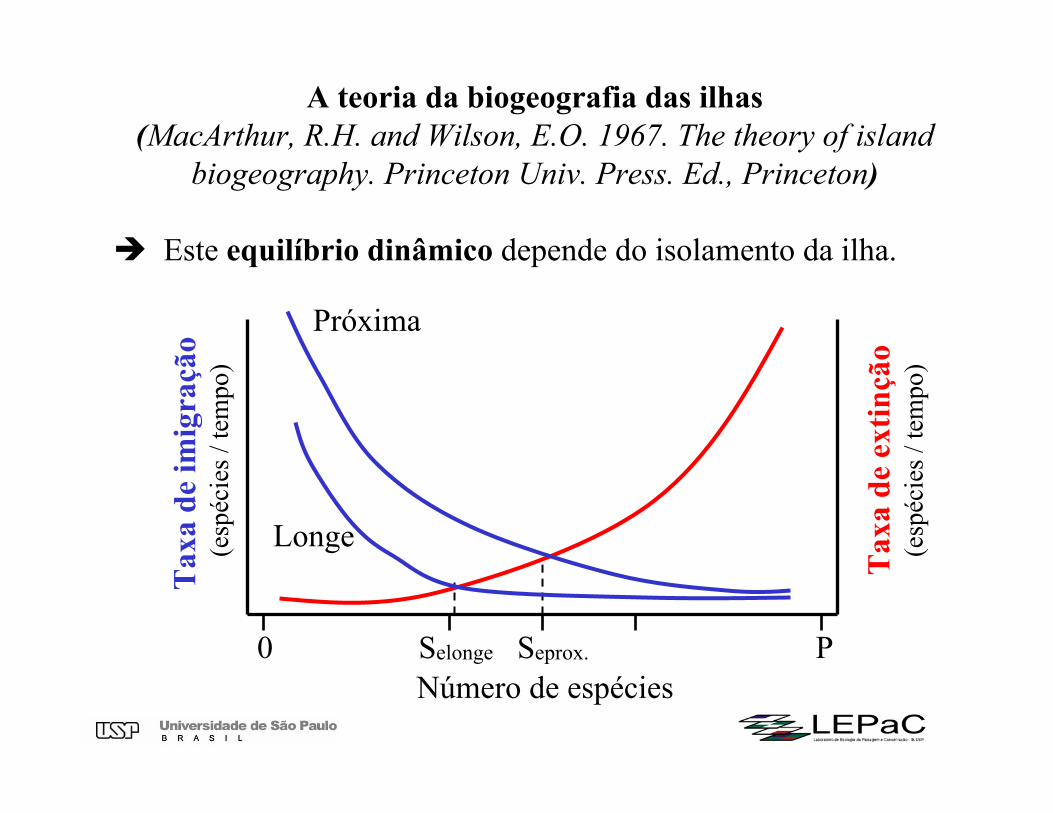
A teoria da biogeografia das ilhas(MacArthur, R.H. and Wilson, E.O. 1967. The theory of island
biogeography. Princeton Univ. Press. Ed., Princeton)
Este equilíbrio dinâmico depende do isolamento da ilha.
Selonge
Tax
a de
imig
raçã
o(e
spéc
ies /
tem
po)
Tax
a de
ext
inçã
o(e
spéc
ies /
tem
po)
Número de espécies0 PSeprox.
Longe
Próxima

Relação entre isolamento de uma ilha e a taxa de imigraçãoA teoria da biogeografia das ilhas
Exemplo: aves nas ilhas das Bahamas

A teoria da biogeografia das ilhas(MacArthur, R.H. and Wilson, E.O. 1967. The theory of island
biogeography. Princeton Univ. Press. Ed., Princeton)
A riqueza de uma comunidade insular depende de equilíbrio dinâmico entre as taxas de extinção e de imigração, que, por sua vez, são influenciadas pela área e isolamento da ilha.
Trata-se de uma teoria simples, sedutora, fácil de ser testada (robusta aos testes?) e com muitas implicações práticas.
A teoria da biogeografia das ilhas

A teoria da biogeografia das ilhas
Limites quando aplicada para habitats fragmentados
Não considera as diferenças entre espécies
Não houve tempo para ocorrer equilíbrio biogeográfico
Analogia fragmento com ilha é fraca;
Isolamento não é a melhor medida de espaçamento
Não considera a qualidade do habitat
A teoria da biogeografia das ilhas

A teoria da biogeografia das ilhas(MacArthur, R.H. and Wilson, E.O. 1967.)
A teoria da biogeografia das ilhas
Trata-se de uma teoria simples, sedutora, fácil de ser testada (robusta aos testes?) e com muitas implicações práticas.

O que é uma metapopulação ?
Conjunto de sub-populações isoladas espacialmente em fragmentos de habitat e unidas funcionalmente por fluxos biológicos.
A teoria da metapopulações

O que é uma metapopulação ?
As populações locais correm risco de extinção: existe uma dinâmica de extinções e recolonizações locais
Tempo 1 Tempo 2Fragmentos ocupados
Fragmentos vazios
A teoria da metapopulações

• As duas premissas básicas da biologia de populações segundo a abordagem de metapopulação:
1. as populações estão estruturadas em conjuntos de populações reprodutivas locais;2. a migração entre as populações locais tem uma influência limitada na dinâmica local, permitindo principalmente o restabelecimento de populações locais extintas.
O que é uma metapopulação ?
• Estas premissas contrastam com os modelos clássicos de dinâmica de população que assumem uma estrutura de população panmítica (onde todos os indivíduos têm a mesma chance de interagir).• A abordagem de metapopulação advem da necessidade de considerar a posição dos indivíduos e das populações no espaço (espacialização da dinâmica de populações).
A teoria da metapopulações

O modelo clássico de LevinsO modelo de Levins (1969) assume que:
- há um grande número de manchas de habitat;- todas as manchas são de mesmo tamanho- todas as manchas têm o mesmo isolamento- todas as manchas estão unidas por migração- não considera-se o tamanho da população local- considera apenas se uma mancha de habitat está
ou não ocupada
P = Proporção de fragmentos ocupados = 7 / 17
A teoria da metapopulações

O modelo clássico de Levins
O modelo de Levins é uma forma simples de descrever adinâmica de ocupação das manchas de habitat (dp/dt).Nesse modelo:
-o risco de extinção é considerado constante e igualem todas as manchas = e-a taxa de extinção no tempo t é então de: e . P-a possibilidade de colonização é considerada proporcional à proporção de manchas ocupadas(fonte de colonizadores) = c . P -a taxa de colonização no tempo t é de : c . P (1 – P)
Logo:dP/dt = cP (1-P) – eP
A teoria da metapopulações

O modelo clássico de LevinsdP/dt = cP (1-P) – eP
Quando a metapopulação atinge o equilíbrio numa determinada proporção de fragmentos, P, dP/dt = 0, então:
cP (1-P) = ePc (1-P) = ec - cP = e- cP = e - ccP = c - eP= (c - e) / cP = 1 - e/c
A metapopulação deve se manter unicamente se e/c < 1, ou c > e
A teoria da metapopulações

O modelo clássico de Levins
e / c < 1 ou c > e
Isso quer dizer que uma metapopulação tende a se extinguir na
medida que:
- o tamanho médio dos fragmentos diminui
- a densidade dos fragmentos diminui (aumento do
isolamento)
A teoria da metapopulações

Um exemplo de metapopulação do tipo Levins: The Glanville fritillary (Melitaea cinxia)
metapopulation
É uma espécie deboborleta ameaçada deextinção, que desapareceu da Finlândia no final dosanos 1970, e agora ocorre, naquela região, apenas nas ilhas de Aland e emalgumas ilhas ao redor.
A teoria da metapopulações

Um exemplo de metapopulação do tipo Levins: The Glanville fritillary (Melitaea cinxia)
metapopulation
- É uma das metapopulações mais bem estudadas (trabalhos de equipe do Ilkka Hanski)
- A área de ocorrência na Finlândia é de 1200 km2 de terra, numa região de 50 por 70 km2.
- Ela tem duas plantas hospedeiras (Plantago e Veronica) que ocorrem em campos secos, que são os patches potenciais de habitat para Melitaea cinxia.
- Há 1502 manchas de campos secos na área de ocorrência.
A teoria da metapopulações

A pergunta inicial é de saber se a peristência da
espécie nestas condições de fragmentação está
condicionada pela dinâmica de metapopulações
ou pela dinâmica das populações locais.
Um exemplo de metapopulação do tipo Levins: The Glanville fritillary (Melitaea cinxia)
metapopulation
A teoria da metapopulações

Uma abordagem de metapopulação baseada no modeloclássico de Levins parece ser útil quando 4 condições sãoobedecidas: 1. O habitat ocorre em manchas distintas que podem
suportar populações reprodutoras da espécie.2. Todas as populações locais apresentam um risco elevado
de extinção, inclusive as que ocorrem nos maiorespatches.
3. As manchas de habitat não podem ser demasiadamenteisoladas para impedir a recolonização (nempróximas demais para formar uma únicapopulação).
4. As populações locais não podem ter uma dinâmicatotalmente sincrônica (se isso ocoresse, a metapopulação se extinguiria rapidamente)
Glanville fritillary (Melitaea cinxia) metapopulationA teoria da metapopulações

1. O habitat ocorre em manchas distintas que podem suportar populações reprodutoras da espécie.
• São os campos secos que ocorrem em pequenas manchas nas ilhas de Aland.
• Tamanho médio das manchas : 0,13 ha• Tamanho máximo das manchas: 6,8 ha• n= 1502 patches• As borboletas passam 60 a 80% do seu tempo de vida na
mancha natal, o que caracteriza um população local (e não apenas um agregado de indivíduos)
Glanville fritillary (Melitaea cinxia) metapopulation
A teoria da metapopulações

2. Todas as populações locais apresentam um risco elevado de extinção, inclusive as que ocorrem nos maiores patches. • Em 1994 havia 377 populações locais (para 1502 patches)• A maior população tinha ca 500 borboletas.• No caso de borboletas, há muitos casos de populações
com várias centenas de indivíduos que se extinguiram em apenas alguns anos.
• Logo, nenhuma das populações de Melitaea cinxia pode ser considerada estável.
Glanville fritillary (Melitaea cinxia) metapopulation
A teoria da metapopulações

3. As manchas de habitat não podem ser demasiadamente isoladas para impedir a recolonização (nem próximas demais para formar uma única população).
Glanville fritillary (Melitaea cinxia) metapopulation
A distância média ao fragmento mais próximo entre ospatches é de 240 m
A distância média dedeslocamento pela borboleta em migração éde 590 m
A distância máximaé de 3870 m
A distância máxima dedeslocamento pela borboleta emmigração é de 3050 m
A teoria da metapopulações

4. As populações locais não podem ter uma dinâmica totalmente sincrônica (se isso ocoresse, a metapo-pulação se extinguiria rapidamente)
Glanville fritillary (Melitaea cinxia) metapopulation
A teoria da metapopulações

Efeito da área e da densidade das manchas na proporção de patches ocupados
Glanville fritillary (Melitaea cinxia) metapopulation
A teoria da metapopulações

Em conclusão1. O estudo com a Melitaea cinxia evidenciou a importância
da área e densidade (isolamento) dos fragmentos na proporção de patches ocupados.
2. Não há população local estável e desta forma a manutenção de Melitaea cinxia depende de uma dinâmica (não sincrônica) de extinções e recolonizações, na qual as recolonizações têm que superar as extinções locais.
Glanville fritillary (Melitaea cinxia) metapopulation
A teoria da metapopulações

O quão comun é o modelo de metapopulação deLevins ?
A estrutura do tipo metapopulação de Levins parece ser o casode:
- 57 das 94 espécies de borboletas residentes da Finlândia (com muita incerteza)
- Insetos florestais que vivem em microhabitats, como tronco de árvores mortas (besouro)
- Daphnia em poças d’água em rochas;- Anfíbios em brejos/lagoas- Aves e pequenos mamíferos em paisagens tendo pequenos
bosquesSão poucos os casos confirmados no campo, numa área e
num período de tempo suficientemente grandes.
A teoria da metapopulações

Limites da aplicabilidade do modelo de Levins
As condições de existência de uma metapopulação clássica (tipo Levins) são muito restritivas:
1. Fragmentos pequenos de tamanho e isolamento semelhante;
2. Populações locais com dinâmicas totalmente independentes;
3. Gama de migração estreita: grande o suficiente para permitir recolonizações, mas não grande demais para influir na dinâmica local.
A teoria da metapopulações

Ampliação do conceito de metapopulação
Em 1997, o conceito de metapopulação foi ampliado para incluir outros tipos de populações fragmentadas. As condições básicas para existir uma metapopulação:
1. Existência de um espaço categórico (um mosaico enão um gradiente);2. Existência de manchas de habitats (favoráveis para aespécie) que podem ser distingüidas da matriz (habitat vsmatriz).3. As populações locais estão unidadas por migrações.
A teoria da metapopulações

Ampliação do conceito de metapopulação
A teoria da metapopulações

Ampliação do conceito de metapopulação
A teoria da metapopulações

• As metapopulações apresentam um variedade de estruturas que se organizam num contínuo indo desdes populações em desequilíbrio às “patchy populations”, ou do modelo de Levins a um modelo de continente-ilha
• A dinâmica destas populações vaira muito, havendo menor ou maior importância da dinâmica local, ou das possibilidades de recolonização.
• O modelo adequado deve ser utilizado para a população em estudo.
Ampliação do conceito de metapopulação
A teoria da metapopulações

Modelos de dinâmica de metapopulação
Nicholas Gotelli & Walter KelleyA general model of metapopulation
dynamicsOikos 68: 36-44
1993
A formulação geral de um modelo de metapopulação é:
dP/dt = C – E
C: colonizaçãoE: extinção
dP/dt = g(P) (1-P) - h(P) P
A teoria da metapopulações

Modelos de dinâmica de metapopulação
No modelo de LevinsA colonização depende dos recursos regionais (P) e aextinção não
onde:c é uma constante de colonização e é uma constante de extinção
No equilíbrio: P= 1 – e/c cP
cP cPe
e
e
dP/dt = cP (1-P) - e P
A teoria da metapopulações

Modelos de dinâmica de metapopulação Modelo de Hanski (1982)
(= modelo de Levins efeito de resgate)• Efeito de resgate: diminuição dos riscos de extinção devido a um aumento no P• A colonização e a extinção dependem de recursos regionais (P)
dP/dt = cP (1-P) - eP (1-P) = cP (1-P) - eP + eP2
cP
cP cP
e (1-P)
No equilíbrio: c = e para qualquer Pse c > e então P tende a 1 se c < e então P tende a 0 eP2
eP2e (1-P)
A teoria da metapopulações

Modelos de dinâmica de metapopulaçãoModelo de Boorman & Levitt (1973)
(“source-sink metapopulation” “mainland-island metapopulation”)• A colonização depende de uma fonte externa de propágulos (um “continente”) e a extinção também é independente dos patches vizinhos.
dP/dt = c (1-P) - eP
No equilíbrio: P = c / (e+c)se e= 0, P=1; se e>0, P<1 se c=0, P=0se c=1, P<1
Análogo ao modelo de equilíbrio dinâmico de MacArthur & Wilson (1967)
c
c
c
e
e
e
A teoria da metapopulações

Modelos de dinâmica de metapopulaçãoModelo de Gotelli (1991)
(É o modelo de Boorman & Levitt (1973) com “rescue effect”)• A colonização depende de uma fonte externa de propágulos (um “continente”) e a extinção dependente dos patches vizinhos.
dP/dt = c (1-P) - eP (1-P)
No equilíbrio: P = c /ese c > e, P= 1se c<e, P<1se c=0, P= 0
(modelo contraditório, pois se há influênciados patches vizinhos na extinção, deveriahaver também na colonização)
c
c
c
eP2
eP2
e (1-P)
e (1-P)
A teoria da metapopulações

Modelos de dinâmica de metapopulaçãoA general metapopulation model (Gotelli & Kelley 1993)
dP/dt = g(P) (1-P) - h(P) P
g(P) = α + βPα: contribuição de propágulos
externos (p= 0)β: indicador da força de colonização
h(P)= γ − δPγ: medida intrínsica de extinção de
um fragmentoδ: “rescue effect”
α
0 1
1β
γ
0 1
δ
1
colonização
extinçãoP
P
A teoria da metapopulações
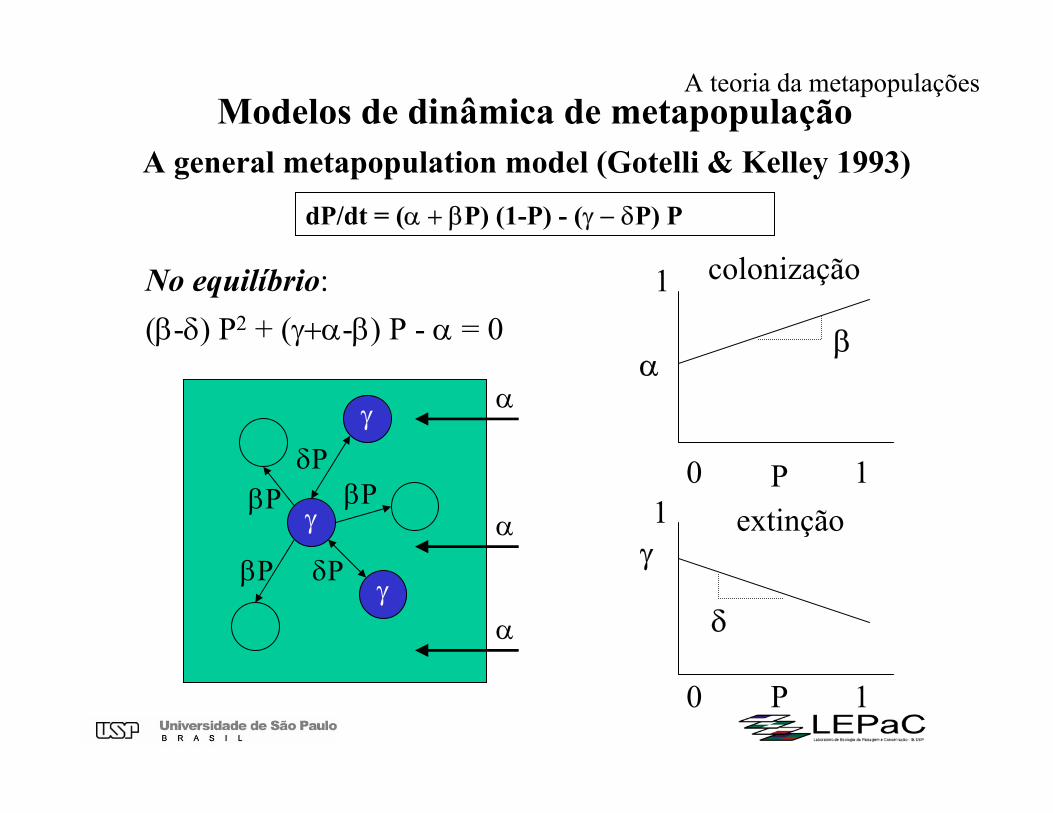
Modelos de dinâmica de metapopulaçãoA general metapopulation model (Gotelli & Kelley 1993)
dP/dt = (α + βP) (1-P) - (γ − δP) P
α
0 1
1
β
γ
0 1
δ
1
No equilíbrio: (β-δ) P2 + (γ+α-β) P - α = 0
βP
γ
colonização
extinção
α
α
α
δPβP
βP P
P
γ
γ
δP
A teoria da metapopulações

1. A teoria da biogeografia de ilhas
2. A teoria de metapopulações
3. A transição e o novo desafio: uma teoria da paisagem
Principais tópicos
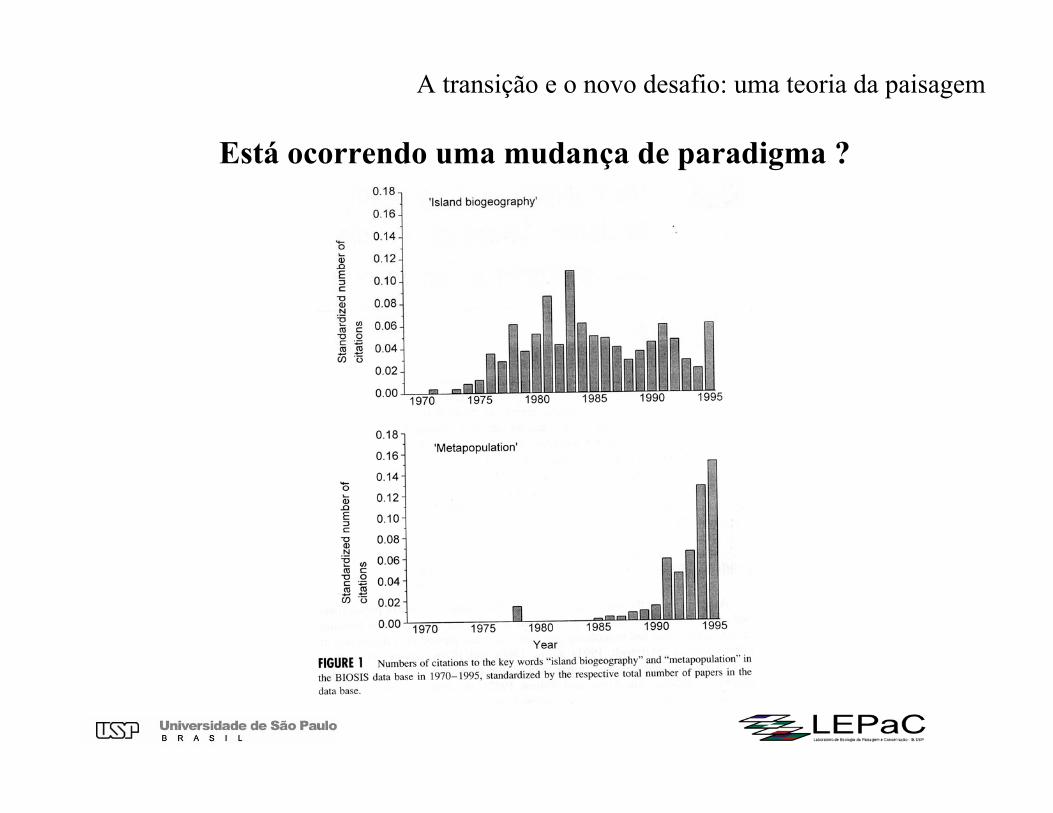
Está ocorrendo uma mudança de paradigma ?
A transição e o novo desafio: uma teoria da paisagem

Por que a ecologia de metapopulações éimportante ?
1. A fragmentação está levando à formação de paisagens contendo apenas manchas pequenas de habitat.2. Populações pequenas são altamente susceptíveis de serem extintas.A persistência de espécies nestas paisagens depende de uma dinâmica regional.
• A teoria das ilhas sempre considera que existe uma fonte estável (continente) e está mais interessada na dinâmica local.•A análise da dinâmica de metapopulações permite focalizar esta dinâmica regional.
A teoria da metapopulações

Está ocorrendo uma mudança de paradigma ?Possíveis razões
1. Crescente descrença de extinções locais em massa (muitas espécies permanecem em pequenos fragmentos).
2. Aplicação crescente da genética de populações na biologia da conservação (problemas de endocruzamento, deriva genética) maior atenção para populações e espécies.
3. Escala de análise: a biogeografia de ilhas foi muito voltada para explicar padrões em escalas muito amplas (nacionais, continentais), enquanto a teoria de metapopulações estámais voltada para a escala de paisagem, onde ocorrem as alterações e o manejo da paisagem.
Mas, premissas das duas teorias são muito semelhantes : não há por que rejeitar uma e aceitar a outra.
A transição e o novo desafio: uma teoria da paisagem

Teoria das ilhas Teoria demetapopulação
Objeto de pesquisa Riqueza deespécies
Presença/ausênciade uma espécie
Objeto de estudo Ilhas verdadeiras Fragmentos dehabitat
Principais fatoresconsiderados
Área e isolamento Área e isolamento
Áreas fontes Continente Todos osfragmentos
A transição e o novo desafio: uma teoria da paisagem
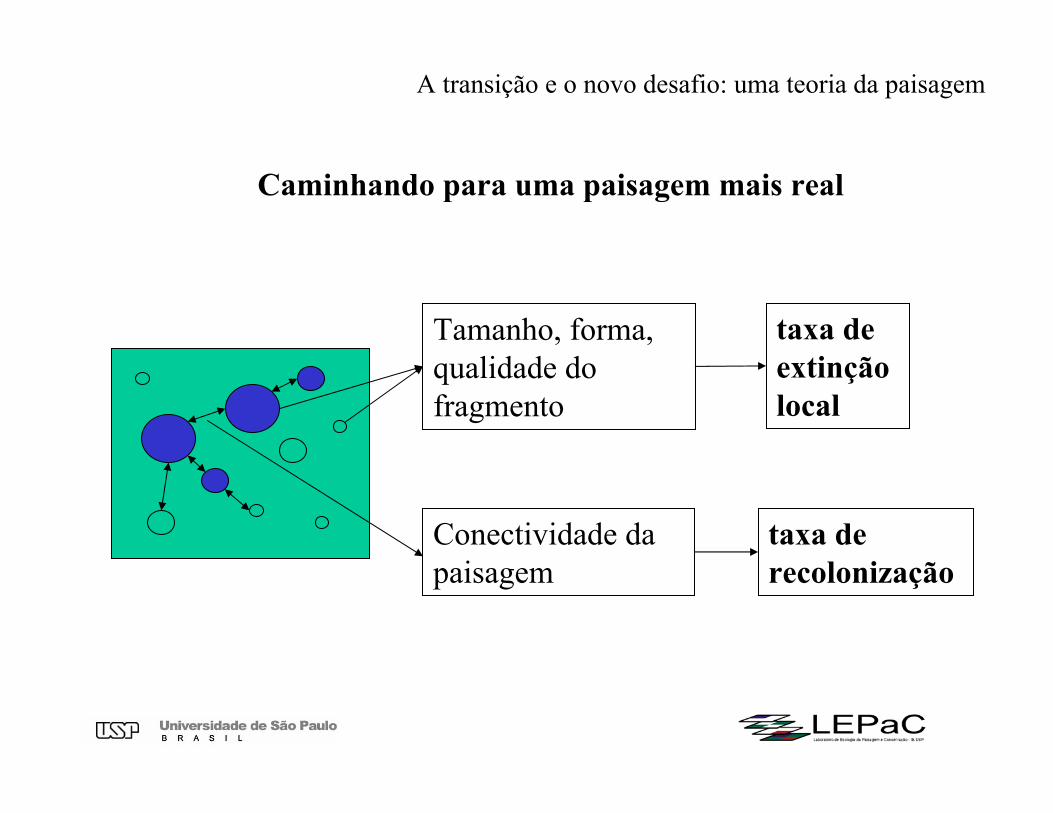
Tamanho, forma, qualidade dofragmento
taxa de recolonização
taxa de extinção local
Conectividade da paisagem
Caminhando para uma paisagem mais real
A transição e o novo desafio: uma teoria da paisagem
Top Related