






Línguas
Páginas
Legal


UNIVERSIDADE DO VALE DO ITAJAÍ MESTRADO EM CIÊNCIA E TECNOLOGIA AMBIENTAL
EFEITOS DE PERTUBAÇÕES NATURAIS E ANTRÓPICAS SOBRE AS ASSOCIAÇÕES DE CRUSTÁCEOS DECÁPODOS INFRALITORAIS NO
ESTUÁRIO DO RIO ITAJAÍ-AÇU, SC, BRASIL
ISABEL DO PRADO LEITE
ITAJAÍ 2012

UNIVERSIDADE DO VALE DO ITAJAÍ
MESTRADO EM CIÊNCIA E TECNOLOGIA AMBIENTAL
EFEITOS DE PERTUBAÇÕES NATURAIS E ANTRÓPICAS SOBRE AS ASSOCIAÇÕES DE CRUSTÁCEOS DECÁPODOS INFRALITORAIS NO
ESTUÁRIO DO RIO ITAJAÍ-AÇU, SC, BRASIL
ISABEL DO PRADO LEITE
Trabalho de Conclusão apresentado ao Programa de Pós-Graduação em Ciência e Tecnologia Ambiental, como parte dos requisitos para obtenção do grau de Mestre em Ciência e Tecnologia Ambiental
Orientador: Dr. Paulo Ricardo Pezzuto
Co-orientador: Dr. Jurandir Pereira Filho
ITAJAÍ 2012

ii
AGRADECIMENTOS
Agradeço, ao Prof. Dr. Paulo Ricardo Schwingel, mais uma vez, pela oportunidade de realizar este estudo e a Mestra em Ciência e Tecnologia Ambiental Aline Antunes pelo apoio, especialmente nas fases iniciais do projeto. Ao Prof. Dr. Paulo Ricardo Pezzuto, pela orientação, pelo tempo dedicado e pela amizade no decorrer deste trabalho. Obrigada! Ao Prof. Dr. Carlos Emílio Bemvenuti, por ter aceitado participar como membro externo da banca. Obrigada por contribuir com este trabalho! Aos professores, estagiários, bolsistas e funcionários do Laboratório de Oceanografia Biológica. Aos professores e colegas do Mestrado em Ciência e Tecnologia Ambiental. A minha família, amigos e especialmente meu namorado, pelo apoio e incentivo fornecidos durante este trabalho, principalmente na etapa final desta dissertação. Obrigada!

iii
SUMÁRIO LISTA DE FIGURAS .................................................................................................... iv
LISTA DE TABELAS .................................................................................................. vii
RESUMO ........................................................................................................................ ix
ABSTRACT ..................................................................................................................... x
1. INTRODUÇÃO ........................................................................................................... 1
2. OBJETIVOS ................................................................................................................ 9
2.1 Objetivo Geral ......................................................................................................... 9
2.2 Objetivos Específicos .............................................................................................. 9
3. ÁREA DE ESTUDO .................................................................................................... 9
4. MATERIAIS E MÉTODOS ..................................................................................... 10
4.1. Desenho amostral. ................................................................................................. 10
4.1.1. Enchentes e Dragagens. ................................................................................ 11
4.2. Amostragens. ......................................................................................................... 16
4.3. Processamento em laboratório ............................................................................... 17
4.4. Análise de Dados. .................................................................................................. 17
4.4.1 Dados Abióticos. .............................................................................................. 17
4.4.2 Dados Bióticos. ................................................................................................. 18
4.4.3 Relação entre variáveis bióticas e abióticas. .................................................... 19
4.4.4 Efeitos das enchentes e da dragagem de aprofundamento. .............................. 19
5. RESULTADOS .......................................................................................................... 20
5.1 Variáveis abióticas .................................................................................................. 20
5.2 Composição e distribuição dos crustáceos decápodos. .......................................... 24
5.2.1 Variação espacial ............................................................................................... 28
5.2.2 Variação temporal.............................................................................................. 33
5.3 Relações entre as variáveis abióticas e os crustáceos decápodos. .......................... 39
5.4 Efeitos das Enchentes. ............................................................................................ 43
5.4.1 Enchente de 2008.............................................................................................. 43
5.4.2 Enchente de 2011.............................................................................................. 46
5.5 Efeitos da Dragagem de Aprofundamento ............................................................. 48
6. DISCUSSÃO .............................................................................................................. 52
7. CONCLUSÕES .......................................................................................................... 70
8. REFERÊNCIAS BIBLIOGRÁFICAS .................................................................... 71

iv
LISTA DE FIGURAS
Figura 1. Localização dos pontos de coleta no estuário do rio Itajaí-Açu (SC) entre maio
de 2008 e novembro de 2011. ................................................................................ 11
Figura 2. Precipitação mensal acumulada no município de Itajaí – SC. A linha
representa os valores registrados no período de maio de 2008 a novembro de
2011. A área cinza do gráfico as médias históricas para os meses de janeiro a
dezembro considerando o período de janeiro de 1980 a março de 2012... ............ 13
Figura 3. Vazão diária do rio Itajaí-Açu entre abril de 2008 e dezembro de 2011. A linha
representa a descarga média diária (m³/s) e os marcadores representam as datas
em que houve coletas no estuário. ........................................................................ 13
Figura 4. Danos causados pela enchente de novembro de 2008 sobre o berço de
atracação do Porto de Itajaí. .................................................................................. 14
Figura 5. Delimitação da área do canal de acesso ao Porto Organizado de Itajaí alvo da
dragagem de aprofundamento e a adequação- bacia de evolução, canal interno e
canal externo. ........................................................................................................ 16
Figura 6. Variação dos valores médios da profundidade entre os pontos 1 a 3 e 4 a 6 das
coletas efetuadas no estuário do rio Itajaí-Açu entre maio de 2008 e novembro de
2011. Em outubro e novembro de 2008 e em setembro de 2009 não foram
realizadas amostragens.. ......................................................................................... 22
Figura 7. Variação entre os pontos 1 a 3 e 4 a 6 das variáveis abióticas da água de fundo
das coletas efetuadas no estuário do rio Itajaí-Açu entre maio de 2008 e novembro
de 2011. Em outubro e novembro de 2008 e em setembro de 2009 não foram
realizadas amostragens... ........................................................................................ 23
Figura 8. Variação da descarga fluvial durante as coletas efetuadas no estuário do rio
Itajaí-Açu entre maio de 2008 e novembro de 2011. Foi utilizada a média da
descarga no dia da coleta junto com os dois dias anteriores. Os dados de descarga
não estavam disponíveis para todas as datas onde foram efetuadas amostragens..24
Figura 9. Contribuição percentual da abundância das principais famílias de crustáceos
decápodos no rio Itajaí-Açu entre maio de 2008 e novembro de 2011. ................. 28
Figura 10. Abundância e biomassa médias nos pontos amostrados para o total de
crustáceos capturados e para as famílias Portunidae, Penaeidae e Palaemonidae. A
barra representa o erro padrão. . ............................................................................. 30

v
Figura 11. Análise MDS realizada com todas as espécies cuja abundância representou,
em ao menos um dos pontos amostrais, mais do que 3% do total. Os fatores
utilizados foram os pontos, numerados de 1 (mais a jusante) a 6 (mais a
montante)....... ........................................................................................................ 31
Figura 12. Abundância média e erro padrão das espécies nos pontos. Foram incluídas
apenas aquelas espécies que a análise SIMPER indicou como representativas nas
diferenças encontradas pela ANOSIM. . ................................................................ 32
Figura 13. Análise MDS realizada com todas as espécies cuja abundância representou,
em ao menos um dos setores (A ou B), mais do que 3% do total. Os fatores
utilizados foram os setores, A e B, que representam os pontos 1-3 e 4-6
respectivamente. . ................................................................................................... 32
Figura 14. Abundância média e erro padrão das espécies nos setores A e B. Foram
incluídas apenas aquelas espécies que a análise SIMPER indicou como
representativas nas diferenças encontradas pela ANOSIM. . ................................ 33
Figura 15. Abundância e biomassa médias nos meses amostrados para o total de
crustáceos capturados e para as famílias Portunidae, Penaeidae e Palaemonidae. A
barra representa o erro padrão. Notar as diferenças de escala de abundância e
biomassa entre as famílias. Em outubro e novembro de 2008 e em setembro de
2009 não foram realizadas amostragens.. . ............................................................ 35
Figura 16. Análise MDS realizada com todas as espécies cuja abundância representou,
em ao menos um dos anos, mais do que 3% do total. Os fatores utilizados foram
os anos, 2008-2011. . ............................................................................................. 38
Figura 17. Abundância média e erro padrão das espécies nos anos. Foram incluídas
apenas aquelas espécies que a análise SIMPER indicou como representativas nas
diferenças encontradas pela ANOSIM. . ................................................................ 39
Figura 18. Bi-plot da ACC referente às variáveis ambientais e biológicas. A análise foi
realizada utilizando todas as espécies cuja abundância representou ao menos 0,1%
do total. Das 4 variáveis ambientais testadas, o oxigênio dissolvido foi removido
do modelo por apresentar p>0,05 nas permutações de Monte Carlo. Os vetores
representam as variáveis ambientais salinidade (Sal), pH, e temperatura
(Temp)......... . ......................................................................................................... 41
Figura 19. Box-plot com a ocorrência das principais espécies em diferentes valores dos
parâmetros ambientais. A caixa representa os percentis de 25% e 75%, as barras
superiores e inferiores os limites de ocorrência que não foram considerados como

vi
outliers, as barras centrais representam a mediana, os círculos os outliers, e o
asterisco (*) representa valores extremos. . ........................................................... 42
Figura 20. Análise MDS realizada com todas as espécies cuja abundância representou,
em ao menos um setor e período, mais do que 3% do total. Os fatores utilizados
foram: “CEA” e “CEB”, dezembro de 2008 a janeiro de 2009 nos setores A e B
respectivamente; “SE1A” e “SE1B”, dezembro de 2009 a janeiro de 2010 nos
setores A e B respectivamente; “SE2A” e “SE2B”, dezembro de 2010 a janeiro de
2011 nos setores A e B respectivamente.. .............................................................. 44
Figura 21. Abundância média e erro padrão das espécies nos períodos e setores
equivalentes a Figura 20. Foram incluídas apenas aquelas espécies que a análise
SIMPER indicou como representativas nas diferenças encontradas pela
ANOSIM.... ............................................................................................................ 45
Figura 22. Análise MDS realizada com todas as espécies cuja abundância representou,
em ao menos um setor e período, mais do que 3% do total. Os fatores utilizados
foram: “CEA” e “CEB”, agosto a setembro de 2011 nos setores A e B
respectivamente; “SE1A” e “SE1B”, agosto a setembro de 2008 nos setores A e B
respectivamente; “SE2A” e “SE2B”, agosto a setembro de 2010 nos setores A e B
respectivamente.. .................................................................................................... 47
Figura 23. Abundância média e erro padrão das espécies nos períodos e setores
equivalentes a figura 22. Foram incluídas apenas aquelas espécies que a análise
SIMPER indicou como representativas nas diferenças encontradas pela
ANOSIM... ............................................................................................................. 48
Figura 24. Análise MDS realizada com todas as espécies cuja abundância representou,
em ao menos um setor e período, mais do que 3% do total. Os fatores utilizados
foram: “CD1A” e “CD1B”, março a junho de 2009 nos setores A e B
respectivamente; “SDA” e “SDB”, março a junho de 2010 nos setores A e B
respectivamente; “CD2A” e “CD2B”, março a junho de 2011 nos setores A e B
respectivamente. . ................................................................................................... 49
Figura 25. Abundância média e erro padrão das espécies nos períodos e setores
equivalentes a figura 24. Foram incluídas apenas aquelas espécies que a análise
SIMPER indicou como representativas nas diferenças encontradas pela
ANOSIM.... . .......................................................................................................... 51

vii
LISTA DE TABELAS
Tabela 1. Localização dos pontos de coleta no estuário do rio Itajaí-Açu (SC) entre maio
de 2008 e novembro de 2011. ................................................................................ 11
Tabela 2. Situação das atividades de dragagens no estuário do rio Itajaí-Açu entre maio
de 2008 a novembro de 2011. * Observações da Saída de Campo. . ..................... 14
Tabela 3. Calendário amostral indicando os meses onde foram coletadas amostras, em
quais ocasiões ocorreram dragagens (manutenção e/ou aprofundamento) e
períodos quando ocorreram enchentes e dragagens de aprofundamento do canal do
Rio Itajaí-Açu... . .................................................................................................... 15
Tabela 4. Valores médios (±DP), mínimos e máximos das variáveis ambientais nos
pontos amostrados no estuário do rio Itajaí-Açu entre maio de 2008 a novembro
de 2011. . ................................................................................................................ 22
Tabela 5. Listagem das espécies e/ou morfotipos coletadas no estuário do rio Itajaí-Açu
entre maio de 2008 a novembro de 2011.. . ........................................................... 25
Tabela 6. Abundância numérica (nº/10min) das espécies em cada ponto e sua
participação relativa na abundância total (%) no estuário do rio Itajaí-Açu entre
maio de 2008 a novembro de 2011.. . .................................................................... 26
Tabela 7. Biomassa (g/10min) das espécies em cada ponto e sua participação relativa na
biomassa total (%) no estuário do rio Itajaí-Açu entre maio de 2008 a novembro
de 2011.. . ............................................................................................................... 27
Tabela 8 Dimensões das principais espécies capturadas no estuário do rio Itajaí-Açu
entre maio de 2008 e novembro de 2011. Para os siris o tamanho representa a
largura da carapaça enquanto que para camarões refere-se ao comprimento total.
N é numero de indivíduos medidos e pesados. < representa o menor e > o maior
valor encontrado. Foi considerado o peso úmido de cada indivíduo. . .................. 27
Tabela 9. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores
do MDS representado na figura 11. No SIMPER foram incluídas todas as espécies
até atingirem 90% de representatividade nos resultados. O SIMPER não foi
considerado quando o resultado do ANOSIM teve p>0,05 (**).. .......................... 31
Tabela 10. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores
do MDS representado na figura 13. No SIMPER foram incluídas todas as espécies
até atingirem 90% de representatividade nos resultados. ....................................... 33

viii
Tabela 11. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores
do MDS representado na figura 16. No SIMPER foram incluídas todas as espécies
até atingirem 90% de representatividade nos resultados. O SIMPER não foi
considerado quando o resultado do ANOSIM teve p>0,05 (**). ........................... 38
Tabela 12. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores
do MDS representado na figura 20. No SIMPER foram incluídas todas as espécies
até atingirem 90% de representatividade nos resultados. O SIMPER não foi
considerado quando o resultado do ANOSIM teve p>0,05 (**)... ......................... 45
Tabela 13. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores
do MDS representado na figura 22. No SIMPER foram incluídas todas as espécies
até atingirem 90% de representatividade nos resultados. O SIMPER não foi
considerado quando o resultado do ANOSIM teve p>0,05 (**).. .......................... 47
Tabela 14. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores
do MDS representado na figura 24. No SIMPER foram incluídas todas as espécies
até atingirem 90% de representatividade nos resultados. O SIMPER não foi
considerado quando o resultado do ANOSIM teve p>0,05 (**).. .......................... 50

ix
RESUMO
Os estuários e as associações de crustáceos decápodos que habitam esses ecossistemas apresentam grande importância ecológica e econômica. O estuário do rio Itajaí-Açu, abriga dois grandes portos em Itajaí e Navegantes e está sujeito a muitos impactos de origem antrópica, como por exemplo, as dragagens e também eventos naturais extremos, como as enchentes. Duas enchentes de grandes proporções atingiram a região, em novembro de 2008 e setembro de 2011, e ainda não se sabe ao certo quais foram os impactos desse evento sobre a fauna estuarina. Assim, o presente estudo teve como objetivo analisar os efeitos de perturbações naturais e antrópicas sobre a estrutura e a dinâmica das associações de crustáceos decápodos infralitorais do estuário do rio Itajaí-Açu. Para isso, foram selecionados seis pontos amostrais distribuídos entre a foz deste estuário e o meandro a montante do rio Itajaí-Mirim. Foram realizadas coletas mensais desde maio de 2008 até novembro de 2011, com arrasto de fundo. Parâmetros físico-químicos também foram coletados utilizando uma sonda. Os camarões e siris foram identificados, medidos e pesados. Foram utilizadas análises descritivas e estatísticas multivariadas sobre os dados padronizados. Ao longo do estudo ocorreram dragagens de aprofundamento que aumentaram a profundidade do canal nos pontos mais externos de 10 para 14 metros. A vazão do rio Itajaí-Açu nos dias anteriores a coleta explicaram a variabilidade da salinidade, pH e oxigênio dissolvido na água de fundo do estuário. No total foram encontradas 20 espécies ou morfotipos distintos de decápodos, sendo que apenas seis contribuíram juntos com mais de 95% do número total de indivíduos capturados. Em todos os anos foi capturada uma maior quantidade de indivíduos da família Portunidae, chegando a 91% em 2008, sobretudo nos pontos intermediários do estudo. As outras duas famílias com contribuição relevante foram Penaeidae e Palemonidae. O MDS (técnica de escalonamento multidimensional não-métrico) e a ANOSIM (análise de similaridade) possibilitaram a separação do estuário em dois setores, A e B, sendo o primeiro a parte mais a juzante onde ocorre a circulação de grandes navios e as dragagens. A temperatura e a salinidade foram as principais variáveis que afetaram a distribuição dos decápodos. Logo após as enchentes foram observadas baixas abundâncias, provavelmente em função do efeito agudo desse tipo de evento extremo sobre a fauna. Entretanto, de 3 a 4 meses após a enchente de 2008 houve a recuperação da assembleia de decápodos, ressaltando a elevada resiliência desse grupo de organismos após grandes distúrbios. As dragagens podem estar afetando principalmente as espécies dominantes, mas esses efeitos são difíceis de confirmar devido ao elevado número de outros fatores que podem ter proporcionado as alterações sobre os crustáceos.
Palavras-Chave: Dragagem, enchente, macrocrustáceos, resiliência.

x
ABSTRACT
Estuaries, and the groups of decapod crustaceans that inhabit these ecosystems are of great ecological and economic importance. The estuary of the Itajaí-Açu River is home to the major ports of Itajaí and Navegantes, and is subject to heavy anthropic impact, such as the dredging operations, and extreme natural events like floods. Two major floods have affected the region, one in November 2008 and the other in September 2011, and it is still not known for certain what impacts these events have had on the fauna of the estuary. The aim of this study, therefore, is to analyze the effects of natural and anthropic disturbances on the structure and dynamic of the groups of infra-littoral decapod crustaceans of the Itajaí-Açu River estuary. We selected six sampling stations distributed from the mouth of the Itajaí-Açu River, to a meander of the Itajaí-Mirim River, further upstream. The crustaceans were collected monthly, from May 2008 to November 2011, by bottom trawling. Thephysical and chemical parameters were also analyzed, using a probe. Shrimp and crabs were identified, measured and weighed. Descriptive statistics and multivariate analyses were carried out on the standardized data. During the study, the depth of the channel increased from 10 to 14 meters due to the dredging operations. The flow of the river in the days before the samplings explained the variability of salinity, pH and dissolved oxygen in bottom water. While twenty different species were identified, 95% of the total individuals caught consisted of just six species. Each year, higher numbers of individuals of the Portunidae family were caught, reaching 91% in 2008, particularly in the intermediate stations. The other two families with relevant contribution were Penaeidea and Palemonidae. MDS (non-metric multidimensional scaling) and ANOSIM (analysis of similarity) allowed the division of the estuary into two sections, A and B, the first being the portion further downstream, where the dredging operations were more concentrated. Temperature and salinity were the main variables affecting the decapod distribution. Soon after both floods a very low abundance was observed, probably due to the acute effect of this type of extreme event on the fauna. However, three to four months after the flood of 2008 the decapod populations had recovered, highlighting the high resilience of this group of organisms following major disturbances. The dredging may be having a negative effect, particularly on the dominant species, but these effects are difficult to confirm because the changes observed may be due to many factors.
Keywords: dredging, flood, macrocrustaceans, resilience

1
1. INTRODUÇÃO
Os estuários são corpos de água costeiros onde ocorre a transição entre o
ambiente continental e marinho, apresentando assim feições desses dois ambientes,
além de suas individualidades (Knox, 2001). São assim caracterizados pela presença
mensurável de água do mar diluída em água doce, mistura que é constantemente
variável devido a efeitos de maré, vazão fluvial e chuva (McLusky, 1989; Hobbie, 2000;
Miranda et al., 2002). Entretanto, existem diferentes definições para os limites de um
estuário considerando não somente salinidade, mas também o efeito da maré, feições
morfológicas ou aspectos biológicos (Day Jr. et al., 1989). Segundo esses últimos
autores, a definição mais utilizada considera que o limite superior do estuário é o local a
montante onde ainda é percebido o efeito da amplitude de maré.
Os ambientes estuarinos apresentam, em curto prazo, uma grande variabilidade
dos parâmetros físico-químicos, o que tende a limitar o número de organismos que
habitam tipicamente esses locais (Angonesi, 2005). Entretanto, estes ambientes são
biologicamente mais produtivos do que os rios ou o oceano adjacente, pois as
características hidrodinâmicas de circulação que aprisionam altas concentrações de
nutrientes estimulam a produção primária (Miranda et al., 2002), tornando a biomassa
nesses ecossistemas bastante elevada (McLusky, 1989). Além disso, são importantes
locais de reprodução, alimentação ou desenvolvimento de espécies marinhas e fluviais,
que habitam o estuário em determinados momentos de seu ciclo de vida (Hobbie, 2000;
Veado, 2008), sendo, portanto ambientes de grande importância biológica (Medeiros,
2003).
Os estuários apresentam ainda grande relevância no contexto econômico
(Medeiros, 2003), basta observar que cerca de 2/3 das grandes cidades estão localizadas
próximas a estes ambientes (Pereira Filho et al., 2003). Essa importância se deve à
facilidade de construção de portos, marinas, e indústrias de pescado nesses locais
(Pereira Filho et al., 2003), além da preferência da população em viver junto a zona
costeira (Hobbie, 2000). A maior parte de toda a matéria originada da decomposição
intempérica dos continentes passa pelos estuários em direção aos oceanos, tornando
esses sistemas os principais fornecedores de nutrientes para a região costeira (Schettini,
2002; Pereira Filho et al., 2003). Dessa forma, além de estarem em zonas de grande
interesse econômico, os estuários são também responsáveis por sustentar diversas

2
populações de peixes e invertebrados marinhos que abastecem as indústrias de pescado
mundial.
Uma característica marcante em estuários é a forte interação entre o sedimento e
a coluna d’água, de modo que o meio bentônico apresenta um forte efeito estruturador,
regulando ou modificando a maioria dos processos físicos, químicos e biológicos nesses
ambientes (Day Jr. et al., 1989). Dentre os grupos de organismos que compõem a fauna
bentônica estuarina pode-se destacar os macrocrustáceos decápodos, representados
principalmente pelos camarões, caranguejos e siris (McLusky, 1989; Bemvenuti &
Colling, 2010).
São inúmeros os processos nos quais os macrocrustáceos decápodos estão
envolvidos em um estuário, como a predação sobre diversos grupos, o consumo de
depósitos e a necrofagia (Raffaelli et al., 1989), além da bioturbação, da regeneração de
nutrientes, ou de servirem como fonte de alimento para o necton demersal (Day Jr. et
al., 1989). No Brasil, algumas espécies importantes de siris (e. g. Callinectes danae, C.
sapidus) (Barreto et al., 2006) e de camarões como o camarão-rosa (Farfantepenaeus
brasiliensis e F. paulensis) são exploradas comercialmente. Todas essas espécies
também são exploradas no Sul do Brasil (Lana, 1996; Ferreira & D’Incao, 2008), sendo
que os organismos podem ser capturados tanto no mar como no próprio estuário.
Os macroinvertebrados bentônicos estuarinos são bastante sensíveis a mudanças
no ambiente (Sánchez-Moyano & García-Asencio, 2010), uma vez que o grupo está
intimamente ligado ao substrato, local onde os efeitos de perturbações são mais
evidenciados. Por esse motivo os macrocustáceos têm sido utilizados também como
instrumentos para se mensurar as perturbações que estão agindo sobre estes ambientes
(Bemvenuti & Rosa-Filho, 2000). Entretanto, a grande dinâmica dos estuários torna
difícil a diferenciação entre variações naturais e os efeitos de perturbações e, portanto, é
preciso um monitoramento temporal dos organismos e das variáveis ambientais para
que conclusões mais robustas possam ser alcançadas (Angonesi, 2005).
Além das variações decorrentes das mudanças sazonais nas variáveis ambientais,
eventos isolados podem vir a afetar de forma drástica a estrutura da macrofauna
bentônica estuarina. Esses eventos podem ser de origem natural ou provocados pelo
homem. Entre os eventos naturais pode-se destacar as enchentes (Eyre & Ferguson,
2006) e as secas (Pillay & Perissinotto, 2008), que podem modificar praticamente todos
os processos que ocorrem nos estuários. Em relação a impactos provocados pelo
homem, pode-se destacar a descarga de efluentes domésticos, industriais e agrícolas

3
(Bellotto et al., 2009), a construção de portos e barragens (Dauvin et al., 2010), as
dragagens e a remoção de organismos através da pesca (Miranda et al., 2002). Esses
impactos afetam o fluxo de água, sedimentos, nutrientes e matéria orgânica nos
estuários e consequentemente os macrocrustáceos bentônicos (Hobbie, 2000).
Em relação às mudanças naturais, pode-se destacar a ocorrência de enchentes
extremas que podem modificar a comunidade bentônica em curto prazo (Cardoso et al.,
2008). De fato, a entrada de água doce em excesso em um estuário pode reduzir
bastante a biomassa dos organismos bentônicos (Rozas et al., 2005), de modo que
durante e após uma enchente ocorrem mudanças rápidas e complexas nas interações
entre o bentos e o pelagial (Eyre & Ferguson, 2006). De modo geral, após um tempo o
ecossistema deveria se reestabelecer naturalmente, porém Cardoso et al. (2008) sugerem
que ambientes impactados apresentam uma menor resiliência, e assim, com os impactos
constantes que um estuário geralmente está sujeito, os efeitos de uma enchente de
grandes proporções podem ser permanentes.
Segundo IPCC (2007), eventos extremos deverão ocorrer com maior frequência
nos próximos anos em função das mudanças climáticas globais, que são aceleradas pela
atividade antrópica. De fato, o aumento de eventos extremos devido as mudanças
globais pode representar um problema para os ecossistemas estuarinos (Cardoso et al.,
2008; Pillay & Perissinotto, 2008), de modo que a realização de estudos que permitam a
tomada de medidas para predizer e/ou diminuir tais impactos são cada vez mais
necessários.
Além de todos os danos ambientais trazidos pelas enchentes como a remoção de
organismos e a modificação nos ciclos dos nutrientes (Eyre & Ferguson, 2006), é
comum ocorrerem dragagens em estuários após esses eventos para permitir que as
embarcações de grande porte voltem a utilizar o canal que recebe uma grande carga de
sedimentes durante as cheias. As dragagens são responsáveis tanto pela remoção direta
dos organismos bentônicos, como também pela ressuspensão de sedimentos (Newell et
al., 1998), esses últimos podem estar contaminados com diferentes poluentes que se
acumulam nos substratos dos estuários e, ao serem remobilizados, podem voltar a estar
biodisponíveis na coluna d’água (Bellotto et al., 2009). Somente os sedimentos em
suspensão, mesmo livres de contaminantes, já seriam um problema em estuários uma
vez que a elevada turbidez pode reduzir significativamente a abundância de espécies-
chaves da cadeia trófica, alterando a distribuição de diversos organismos, interferindo

4
em processos como a competição e consequentemente afetando de forma negativa a
função berçário dos estuários (González-Ortegón et al., 2010).
Em relação aos efeitos da dragagem sobre a epifauna bentônica, Smith et al.
(2006) observaram que a atividade pode reduzir significativamente a diversidade e a
abundância desse grupo de organismos. Esses autores também observaram mudanças na
comunidade, como a dominância de algumas espécies com maior capacidade de
locomoção e redução na biomassa média. Porém, os efeitos negativos não se limitam ao
bentos. Newell et al. (1998) destaca que deve-se considerar as diversas populações,
comunidades e até ecossistemas que dependem da epifauna bentônica estuarina para se
sustentar, de modo que o impacto sobre esse grupo de organismos não se limitará a
determinado estuário, mas poderá se estender para outros importantes ecossistemas,
inclusive marinhos.
Da mesma forma como são complexos os impactos que agem sobre a fauna
estuarina, também são complicados os estudos realizados nesses ecossistemas. Por
estarem em locais densamente ocupados pela população humana, os estuários tendem a
sofrer interferência direta das atividades antrópicas, de modo que sua estrutura se torna
totalmente diferente a partir do momento que esses locais são ocupados (Lindegarth &
Hoskin, 2001). Assim, torna-se difícil conhecer como funcionam estuários não
impactados pelo homem, uma vez que a grande maioria já foi modificada de alguma
forma. Marques et al. (1993), entretanto, compararam dois estuários com características
semelhantes e notaram que a riqueza específica era bem inferior no estuário impactado,
da mesma forma como foi sugerido por Sánchez-Moyano & García-Asencio (2010). As
espécies presentes em estuários impactados e não impactados também são bastante
diferentes, embora a biomassa possa ser semelhante entre esses dois ecossistemas
(Marques et al., 1993). Essas diferenças sugerem que, em ambientes com distúrbios
frequentes, a fauna bentônica se adapta, de modo que apenas as espécies aptas para lidar
ou evitar o distúrbio conseguem se estabelecer em longo prazo (Newell et al., 1998).
Nesse contexto, o estado de Santa Catarina apresenta importantes estuários,
destacando-se em seu limite norte a Baia da Babitonga, no sul o estuário do Rio
Mampituba, e no litoral centro-norte o estuário do rio Itajaí- Açu, alvo deste estudo. A
fauna do estuário do rio Itajaí-Açu tem sido estudada sob vários aspectos nos últimos
anos. Resgalla Jr. (2009) verificou a importância da pluma do rio para a comunidade
zooplanctônica de toda a região, identificando um padrão de distribuição bem marcado
do zooplâncton no baixo estuário com três zonas com características distintas: na parte

5
mais a montante e na parte superior do perfil no baixo estuário, há o predomínio de
espécies límnicas e estuarinas, em baixa riqueza, densidade e biomassa; sobre a pluma
do estuário, já na plataforma continental, há o domínio de espécies costeiras,
eurihalinas, tropicais e grandes, com as maiores densidades e biomassas de todo o
perfil; já na plataforma abaixo da pluma e no fundo do perfil no baixo estuário, foi
encontrada a maior riqueza específica de todo o perfil, dominada por copépodos de
pequeno tamanho. Esses resultados são importantes para compreender a estrutura dos
estuários, uma vez que existe uma forte interação entre grupos bentônicos e
planctônicos nesses ambientes.
O Saco da Fazenda, enseada rasa situada na margem direita, próximo a
desembocadura do estuário do rio Itajaí-Açu, apesar de sofrer forte pressão antrópica
causada pelo fluxo de efluentes domésticos e resíduos sólidos, abriga uma grande
diversidade de aves, atuando como um importante local de alimentação e repouso para
bandos mistos tanto de aves marinhas costeiras como límnicas. E atua também como
fonte alternativa de alimento para aves habitantes das bordas, que durante a baixamar
excursionam pela planície de maré a procura de poliquetas e crustáceos (Branco, 2000).
A predação das aves marinhas e limícolas sobre organismos aquáticos no Saco da
Fazenda constitui mais uma importante interação que demonstra a importância do baixo
estuário do rio Itajaí-Açu para ecossistemas adjacentes, inclusive terrestres.
Outro importante grupo de organismos presentes no estuário do Itajaí-Açu é a
ictiofauna, que apresenta, em geral, menor número de espécies que outros locais onde
foram feitos estudos semelhantes na costa sul do Brasil (Hostim-Silva et al., 2002). Essa
menor riqueza em relação a outros estuários pode ser reflexo da forte pressão antrópica
que esse ecossistema está sofrendo. Entretanto, foi verificado que o estuário apresenta
importantes espécies de peixes de valor comercial, o que comprova a importância sócio-
econômica da área para as comunidades ribeirinhas e para toda a região do Vale do
Itajaí (Hostim-Silva et al., 2002; Branco et al., 2009a). Outro fator importante em
relação à ictiofauna do estuário do Itajaí-Açu e dos ecossistemas adjacentes é o elevado
percentual de juvenis que habitam esses ambientes, o que reforça a importância dos
estuários como áreas berçário e de crescimento de muitas espécies de peixes (Hostim-
Silva et al., 2002; Branco et al., 2009a).
Em um estudo realizado entre 2000 e 2005 no Saco da Fazenda, foi observada a
diminuição de peixes demersais e bentônicos, que foram sendo substituídos
gradualmente ao longo dos anos em que este ambiente passou por dragagens (Branco et

6
al., 2009a). Entretanto, apesar dessa atividade afetar de forma significativa o bentos, os
autores destacam que a melhoria dos padrões de circulação promovida pelo
aprofundamento do canal pode ter contribuído para o aumento da diversidade de peixes
nos últimos anos de estudo. Assim, pode-se destacar que o estuário do Itajaí-Açu
apresenta um importante habitat para a ictiofauna, promovendo condições para o
crescimento de organismos juvenis que podem vir a sustentar populações ribeirinhas
quando retornam ao mar. Entretanto a pressão antrópica, através da poluição, dragagens
e pesca, vem modificando a assembléia de peixes da região (Branco et al., 2009a) o que
demonstra a necessidade da preservação desse ecossistema (Hostim-Silva et al., 2002).
Em um estudo mais recente, Antunes (2010) observou o mesmo padrão que
Hostim-Silva et al. (2002) e Branco et al. (2009a) para a dominância de poucas espécies
ícticas no estuário do Rio Itajaí-Açu. Em seu estudo, Antunes (2010) observou que a
ictiofauna foi composta por 46 espécies, das quais apenas seis representaram 99% da
captura durante o período de estudo, entre maio de 2008 e julho de 2009. As espécies
mais abundantes foram predominantemente juvenis, da mesma forma como observado
por outros autores (Hostim-Silva et al., 2002; Branco et al., 2009a), ressaltando a
importância desse habitat para diversas populações de peixes.
Apesar desses estudos, ainda faltam informações sobre diferentes grupos de
organismos que habitam o estuário do rio Itajaí-Açu para melhor compreensão sobre a
ecologia desse ambiente. Entre esses grupos estão os crustáceos. Até 2009, as únicas
informações disponíveis sobre esses organismos foram obtidas no Saco da Fazenda,
onde Branco & Freitas Jr. (2009) investigaram os caranguejos e siris, observando para
esses grupos a dominância (mais de 60% da captura) de uma única espécie, sendo ela
Uca uruguayensis para os caranguejos, e Callinectes danae para siris. Esse padrão de
dominância reflete o ambiente altamente impactado, geralmente dominado por poucas
espécies que de alguma forma toleram ou evitam o estresse (Newell et al., 1998).
Mais recentemente, foi observada a ocorrência de poucas espécies de crustáceos
decápodos epibentônicos dominando o canal do estuário do rio Itajaí-Açu,
principalmente a espécie C. danae (Leite, 2010), como também foi observado no estudo
de Branco e Freitas Jr. (2009) no Saco da Fazenda. A preferência de algumas espécies
por áreas mais a montante ou mais a juzante também foi observada no estuário, porém,
em virtude das grandes variações nas variáveis físico-químicas, na grande estratificação
deste estuário e na ocorrência de um grande evento de enchente durante o estudo, os
padrões encontrados não foram conclusivos e um estudo abrangendo maior escala

7
temporal seria necessário para que a distribuição dos macrocrustáceos epibentônicos no
estuário fosse realmente conhecida (Leite, 2010).
Dentre os impactos que afetam o estuário do Itajaí-Açu pode-se destacar o
aporte de efluentes tanto domésticos e industriais, bem como a lixiviação de material
dos centros urbanos e toda a carga de matéria orgânica e poluentes que os mesmos
trazem através do rio (Bellotto et al., 2009). Carreados pela correnteza, provenientes de
regiões mais superiores do rio, há também agroquímicos da zona rural do médio e alto
Vale do Itajaí (Veado, 2008), que podem vir a se acumular no estuário por conta da
circulação restrita (Schettini & Truccolo, 2009). Ainda há atividades portuárias, com
intensa circulação de navios com cargas internacionais que podem vir a trazer espécies
invasoras através da água de lastro como, por exemplo, o siri Charybdis hellerii
(Tavares & Mendonça Jr., 2004), que foi identificado no estuário do rio Itajaí-Açu por
Leite (2010). A circulação de navios, e também de outras embarcações, promove ainda
a ressuspensão de sedimentos, efeito também observado durante o processo de
dragagem.
Em relação às dragagens, há registros da atividade no rio Itajaí-Açu desde 1895,
no entanto, estas se tornaram mais frequentes e significativas após a década de 1960,
com o incremento do comércio marítimo (Schettini, 2001). Atualmente há um programa
de dragagem de manutenção permanente no baixo estuário, realizado por uma draga de
injeção de água, com o intuito de manter o canal com profundidade adequada aos
grandes navios que atracam nos portos de Itajaí e Navegantes (Branco et al., 2009b).
Eventualmente são feitas dragagens de aprofundamento ou emergenciais para que os
portos não fiquem parados, como ocorreu após a enchente de novembro de 2008,
quando a profundidade do canal atingiu valores inferiores a 8 metros (Antunes, 2010;
Leite, 2010).
De fato, o Vale do Itajaí tem enfrentado enchentes que vêm ocorrendo há mais
de um século (Antunes, 2010). A partir da década de 1920 tem aumentado o número de
pequenas enchentes devido ao aumento da ocupação no Alto Vale do Itajaí e, para evitar
alagamentos na região, foram realizadas obras de engenharia, como represas de
contenção que foram projetadas a partir de 1957 (Porath, 2004). Além disso, são
executadas no canal obras de engenharia para a manutenção dos portos, como a fixação
do canal através de molhes e a dragagem (Branco et al., 2009b). Entretanto, mesmo
após esses esforços, eventos extremos de enchente já foram registrados no Vale do Itajaí
na década de 1980, favorecidos por um período de El Niño (Schettini & Truccolo,

8
2009), e mais recentemente, em 2008 e em 2011, resultados de pluviosidade acima da
média (Leite, 2010). Entretanto, os efeitos desses eventos sobre os ecossistemas ainda
precisam ser mais estudados, considerando os efeitos das mudanças climáticas globais
que devem torná-los cada vez mais frequentes (IPCC, 2007).
Na região Sul do Brasil já foram observadas importantes modificações no bentos
em função de anos mais secos ou chuvosos (Bemvenuti & Colling, 2010; D’Incao &
Dumont, 2010). Entretanto, eventos extremos têm grande potencial para alterar a
estrutura da epifauna bentônica, composta por peixes e invertebrados, dos quais se
destacam os siris e camarões (Pillay & Perissinotto, 2008). Esses eventos afetam,
inclusive, a resiliência destes organismos (Cardoso et al., 2008), e as enchentes que
ocorreram no rio Itajaí-Açu podem ter tido um papel fundamental na estruturação atual
do sistema bentônico estuarino. De fato, durante o evento de novembro de 2008, houve
grandes mudanças na estrutura dos macrocrustáceos (siris e camarões) do estuário do rio
Itajaí-Açu (Leite, 2010). Paralelamente, Antunes (2010) e Silva (2010) detectaram
padrões parecidos nas associações de peixes e da macrofauna bentônica,
respectivamente, para o mesmo estuário, sugerindo que os efeitos da enchente podem
afetar todo o ecossistema.
Frequentemente é difícil interpretar os efeitos de distúrbios nos estuários, uma
vez que são ambientes bastante complexos; as dinâmicas físicas, químicas e geológicas
podem interferir nos resultados e mascarar impactos da atividade humana no
ecossistema estuarino. Por essa razão, estudos em estuários devem apresentar réplicas
espaciais e temporais para que o impacto de uma perturbação seja realmente detectado
(Dauvin et al., 2010). A enchente de novembro de 2008 foi responsável pela completa
expulsão da cunha salina do estuário, resultando na alteração de todos os parâmetros
físico-químicos que são responsáveis pela distribuição dos organismos nesses ambientes
(Antunes, 2010; Leite, 2010). Schettini (2002) correlaciona a expulsão da cunha salina a
ocasiões onde a descarga fluvial é superior a 1000m³/s. Em relação aos decápodos, no
mês seguinte após o evento foi possível notar uma redução significativa na abundância e
riqueza desses organismos, com recuperação gradativa dos mesmos nos meses seguintes
(Leite, 2010). Entretanto, ainda não foi possível obter resultados totalmente conclusivos
quanto aos efeitos da enchente sobre os distintos componentes da fauna do estuário do
rio Itajaí-Açu devido ao pouco tempo amostral empregado pelos diversos autores
(Antunes, 2010; Leite, 2010; Silva, 2010), que não abrangeu uma réplica temporal, ou
seja, um novo período do ano, idêntico ao da enchente, porém sem o efeito da mesma.

9
O presente estudo visa identificar melhor tais efeitos a partir do uso de um programa
amostral de mais longo prazo.
2. OBJETIVOS 2.1. Objetivo Geral
Analisar os efeitos de perturbações naturais e antrópicas sobre a estrutura e a
dinâmica das associações de crustáceos decápodos infralitorais do estuário do rio Itajaí-
Açu, entre maio de 2008 e novembro de 2011.
2.2. Objetivos específicos
• Descrever a composição quali-quantitativa das associações de crustáceos
decápodos infralitorais no estuário do rio Itajaí-Açu, e verificar suas variações
espaço-temporais;
• Analisar as variações espaço-temporais nos parâmetros físico-químicos desse
estuário e determinar sua influência sobre a distribuição dos crustáceos
decápodos infralitorais;
• Identificar possíveis efeitos dos eventos extremos de enchente de novembro de
2008 e setembro de 2011 sobre esses organismos;
• Identificar possíveis efeitos das dragagens de aprofundamento sobre esse mesmo
grupo.
3. ÁREA DE ESTUDO
A área de estudo compreende o baixo estuário do rio Itajaí-Açu, entre os
municípios de Itajaí e Navegantes. Este estuário drena uma bacia de 15500 km², onde
estão localizados dois portos que operam cargas internacionais e uma grande frota
pesqueira (Medeiros, 2003). O rio Itajaí-Açu é responsável por cerca de 90% do aporte
fluvial no estuário, sendo os outros 10% atribuídos ao rio Itajaí-Mirim (Schettini, 2002).
A bacia do Itajaí-Açu é a maior da vertente atlântica do estado (Porath, 2004) e
apresenta grande importância econômica para a região, devido aos Portos de Itajaí e
Navegantes e a intensa atividade pesqueira (Schettini, 2001). O baixo estuário apresenta
as margens densamente ocupadas, com a desembocadura fixada por molhes e guia-
correntes. O estuário é do tipo cunha salina altamente estratificado, afetado pelo regime

10
de micro maré (Schettini, 2002). Nesse aspecto, a principal forçante hidrodinâmica no
estuário é a vazão do Rio Itajaí-Açu que, por sua vez, é bem variável ao longo do ano,
com picos em fevereiro e outubro e menores valores em abril e dezembro (Schettini &
Truccolo, 2009). Em períodos prolongados de baixa vazão a cunha salina pode penetrar
até 30 km rio acima, além de desencadear eventos de hipoxia que, quando ocorrem, se
desenvolvem principalmente entre a região mais baixa do médio estuário e a mais alta
do baixo estuário (Delfim, 2009). Em contraposição, em eventos de vazão extrema, 4 a
5 vezes maior que o normal, a cunha salina pode ser totalmente expulsa do estuário
(Schettini, 2002). Esse regime de circulação implica que praticamente todo, se não todo,
o material proveniente na bacia de drenagem fica retido no estuário quando este está sob
condições normais ou de baixa vazão (Schettini, 2001).
O sedimento no estuário do Rio Itajaí-Açu varia em resposta a descarga fluvial,
sendo, na maior parte do tempo, dominado pela fração de argila (Schettini, 2002).
Segundo esse autor, a maior porcentagem do sedimento é de origem continental,
entretanto, em períodos de baixa descarga pode ocorrer um aumento da entrada de areia
fina de origem marinha. Quando a vazão é muito elevada, a porção de silte e argila
diminui e pode ocorrer a predominância de areia.
4. MATERIAL E MÉTODOS
4.1. Desenho amostral
Os pontos amostrais foram distribuídos desde a desembocadura, o ponto 1, até
aproximadamente 10 km rio acima (ponto 6) (Tabela 1, Figura 1). Próximo à
desembocadura está localizado o Saco da Fazenda, um ecossistema com menor
circulação, porém, ligado diretamente ao rio Itajaí-Açu. O ponto 3 está na altura dos
portos de Itajaí e Navegantes, responsáveis por intenso tráfego de grandes navios, e
motivo pelo qual são necessárias dragagens periódicas. Os demais pontos estão mais
afastados da área de influência dos portos, dos quais se pode destacar o ponto 5,
próximo a desembocadura do rio Itajaí-Mirim (Figura 1).
As coletas foram realizadas mensalmente no âmbito do projeto de
monitoramento ambiental do Porto de Itajaí. Foram incluídos dados das coletas
realizadas desde maio de 2008 até novembro de 2011 com arrastos em seis pontos
amostrais entre a foz do rio e à montante da desembocadura do rio Itajaí-Mirim (Tabela
1, Figura 1). Não foi realizada coleta em setembro de 2009 devido a elevada vazão do

11
rio e excesso de chuvas na região. Também não foram efetuadas coletas nos meses de
outubro e novembro de 2008, meses onde a vazão do rio foi muito elevada devido a um
volume extremamente alto de precipitação (Figura 2), que resultou em uma grave
enchente no final do mês de novembro no litoral catarinense.
Tabela 1. Localização dos pontos de coleta no estuário do rio Itajaí-Açu (SC) entre maio de 2008 e novembro de 2011.
Ponto Amostral
Nome do Ponto Posição Geográfica
Latitude (ºS) Longitude (ºW) 1 Foz Itajaí-Açu 26º54,783' 48º38,570' 2 Cepsul 26º54,554' 48º38,949' 3 Porto Itajaí 26º53,896' 48º40,013' 4 Pepsico 26º52,904' 48º40,117' 5 Desembocadura Itajaí-Mirim 26º53,189' 48º41,263'
6 Igreja 26º51,930' 48º41,449'
Figura 1. Localização dos pontos de coleta no estuário do rio Itajaí-Açu (SC) entre maio de 2008 e novembro de 2011.
4.1.1. Enchentes e Dragagens
Durante os três anos e meio de estudo, duas enchentes de grandes proporções
atingiram a região do Vale do Itajaí: a primeira em novembro de 2008 e a segunda em
setembro de 2011. Esses eventos foram resultado de chuvas acima da média na região
(Figura 2), o que ocasionou, consequentemente, o aumento da vazão do rio Itajaí-Açu
(Figura 3). A enchente de novembro de 2008 gerou grandes prejuízos à região, inclusive
ao Porto de Itajaí (Figura 4).

12
A Superintendência do Porto de Itajaí executa constantemente dragagens de
manutenção no canal de acesso ao Porto de Itajaí e ao Porto de Navegantes
(PORTONAVE) (Tabela 2). A dragagem de manutenção é realizada a cada quinze dias
durante a maré vazante e foi realizada em dois períodos ao longo dos três anos e meio
de estudo. No primeiro período a dragagem teve início em maio de 2008 e se estendeu
até setembro de 2008, a segunda dragagem ocorreu de forma descontínua entre agosto
de 2010 e fevereiro de 2011, sendo interrompida nos meses de novembro de 2010 e
janeiro de 2011. Nas operações de manutenção foram utilizadas dragas que operam
através do sistema de injeção de água, que promove a fluidificação dos sedimentos
finos, principal constituinte do fundo (Schettini, 2002).
Dragagens de aprofundamento também são realizadas no estuário, com o
objetivo de aumentar a profundidade do calado do rio para facilitar a navegação de
navios de grande porte. Porém essas dragagens podem ser de caráter emergencial, como
a que ocorreu após a enchente de novembro de 2008. Esse evento extremo gerou o
assoreamento do canal de navegação, e assim foi necessário aumentar a profundidade
do canal para aproximadamente 10 m utilizando uma draga autotransportadora de
arrasto (hopper) com descarga pelo fundo. Essa dragagem teve início em dezembro de
2008 e se estendeu de forma descontínua até julho de 2009 (Tabela 2). Ainda em 2009
começou um estudo para uma nova dragagem de aprofundamento no estuário com
objetivo de aumentar a profundidade para 14m.
Essa nova dragagem de aprofundamento iniciou no mês de março de 2011 e se
estendeu até o termino deste estudo, sendo interrompida entre os meses de julho e
setembro. Novamente foi utilizada uma draga autotransportadora de arrasto (hopper)
com descarga pelo fundo e um total de aproximadamente 6 milhões de metros cúbicos
de sedimento foi dragado (Porto de Itajaí, 2012).
Dessa forma, o calendário amostral se encontra resumido na tabela 3, totalizando
40 campanhas amostrais, sendo 24 em meses com a ocorrência de dragagens no
estuário. Durante 13 meses ocorreram dragagens de aprofundamento do canal, além da
ocorrência de duas enchentes de grandes proporções durante o estudo (Tabela 3). A área
onde ocorreram as dragagens é limitada à região de circulação dos grandes navios que
atracam nos portos de Itajaí e Navegantes (Figura 5). Portanto, no presente estudo
somente os pontos 1, 2 e 3 foram considerados com influência direta das dragagens,
sendo o ponto 3 aquele mais próximo ao Porto de Itajaí, que está localizado a margem
direita do rio Itajaí-Açu a aproximadamente 3 km da sua foz. Assim, em algumas

análises o estuário foi dividido em setores: “A”, pontos 1,
dragagens; e “B”, pontos 4, 5 e 6
Figura 2. Precipitação mensal acumulada no município de Itajaí registrados no período de maio de 2008 para os meses de janeiro a dezembro considerando o período de janeiro de 1980 a março de 2012. Fonte:ARAÚJO, S. A. REIS, F. H. Relatório Climatológicodo Vale do Itajaí. Itajaí- SC. Laboratório de Climatologia.
Figura 3. Vazão diária do rio Itajaídescarga média diária (m³/s) e os
o estuário foi dividido em setores: “A”, pontos 1, 2 e 3 -
dragagens; e “B”, pontos 4, 5 e 6 - a montante, sem dragagens.
. Precipitação mensal acumulada no município de Itajaí – SC. A linha representa os vregistrados no período de maio de 2008 a novembro de 2011. A área cinza do gráfico as médiaspara os meses de janeiro a dezembro considerando o período de janeiro de 1980 a março de 2012. Fonte:ARAÚJO, S. A. REIS, F. H. Relatório Climatológico. Estação meteorológica automática. Universidade
SC. Laboratório de Climatologia.
. Vazão diária do rio Itajaí-Açu entre abril de 2008 e dezembro de 2011. A linha representa a descarga média diária (m³/s) e os marcadores representam as datas em que houve coletas no estuário.
13
a jusante, com
A linha representa os valores
A área cinza do gráfico as médias históricas para os meses de janeiro a dezembro considerando o período de janeiro de 1980 a março de 2012. Fonte:
. Estação meteorológica automática. Universidade
A linha representa a
marcadores representam as datas em que houve coletas no estuário.

14
Figura 4. Danos causados pela enchente de novembro de 2008 sobre o berço de atracação do Porto de Itajaí. Fonte: Porto de Itajaí (2012).
Tabela 2. Situação das atividades de dragagens no estuário do rio Itajaí-Açu entre maio de 2008 a novembro de 2011. * Observações da Saída de Campo.
Período/ Mês Situação de Dragagens
Maio até Setembro/ 2008 Dragagem de manutenção
Dezembro/2008 até Julho/ 2009 Dragagem de aprofundamento
Agosto até Outubro/ 2010 * Dragagem de manutenção
Dezembro/ 2010 * Dragagem de manutenção
Fevereiro /2011 * Dragagem de manutenção
Novembro/ 2010 e Janeiro/ 2011 * Interrupção da dragagem de manutenção
Março/ 2011 Inicio da dragagem de aprofundamento
Julho até final de Setembro/2011 Interrupção da dragagem de aprofundamento
Outubro/ 2011 Reinício da dragagem de aprofundamento
Novembro/ 2011 Dragagem de aprofundamento
Dezembro/ 2011 Término da dragagem de aprofundamento
15
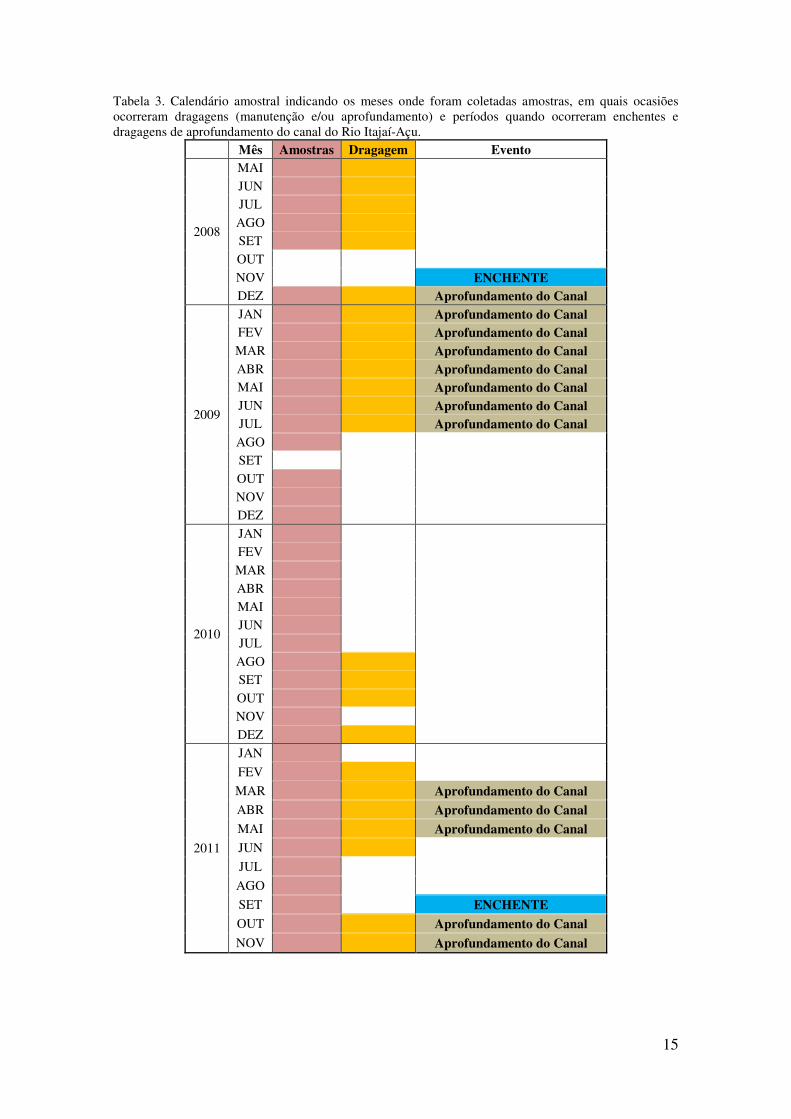
Tabela 3. Calendário amostral indicando os meses onde foram coletadas amostras, em quais ocasiões ocorreram dragagens (manutenção e/ou aprofundamento) e períodos quando ocorreram enchentes e dragagens de aprofundamento do canal do Rio Itajaí-Açu.
Mês Amostras Dragagem Evento
2008
MAI X X
JUN X X JUL X X
AGO X X SET X X
OUT
NOV ENCHENTE DEZ X X Aprofundamento do Canal
2009
JAN X X Aprofundamento do Canal FEV X X Aprofundamento do Canal MAR X X Aprofundamento do Canal ABR X X Aprofundamento do Canal MAI X X Aprofundamento do Canal JUN X X Aprofundamento do Canal JUL X X Aprofundamento do Canal AGO X SET
OUT X
NOV X DEZ X
2010
JAN X FEV X
MAR X
ABR X MAI X
JUN X JUL X
AGO X X SET X X
OUT X X
NOV X DEZ X X
2011
JAN X
FEV X X
MAR X X Aprofundamento do Canal ABR X X Aprofundamento do Canal MAI X X Aprofundamento do Canal JUN X X
JUL X
AGO X
SET X ENCHENTE OUT X X Aprofundamento do Canal
NOV X X Aprofundamento do Canal

16
Figura 5. Delimitação da área do canal de acesso ao Porto Organizado de Itajaí alvo da dragagem de aprofundamento e adequação- bacia de evolução, canal interno e canal externo. Fonte: AQUAPLAN (2009).
4.2. Amostragens
A obtenção das amostras foi realizada com uma embarcação motorizada tipo
baleeira, com 10 metros de comprimento e motor de 45 Hp. Em cada estação e data foi
realizado um arrasto de popa com uma rede de portas com malha de 20 mm no corpo e
no ensacador, medidos entre nós opostos com a malha esticada. Os arrastos tiveram
duração média de 5 ou de 10 minutos, e foram realizados em velocidade média de 2 nós.
Os crustáceos foram separados do resto da captura em cada lance, acondicionados em
sacos plásticos etiquetados e mantidos congelados em freezer até o processamento em
laboratório.
Em função de dificuldades operacionais como elevada quantidade de entulhos e
do grande volume de material capturado, em certos lances foi necessário reduzir o
tempo de arrasto e/ou quartear a captura ainda a bordo. Os tempos de arrasto
efetivamente realizados e os volumes totais e quarteados das capturas foram anotados, e
a partir daí os dados foram padronizados para números ou biomassa de indivíduos
capturados por dez minutos de arrasto.

17
Simultaneamente, foram registrados dados físico-químicos da água de fundo
com um Multianalisador portátil (Water Checker U-10 Horiba), sendo registrados dados
de salinidade, pH, oxigênio dissolvido (mg/l) e temperatura da água (°C). Em algumas
ocasiões foi utilizada a Sonda multiparâmetros (Sonda multiparamétrica marca YSI,
modelo 6600 V2) para obtenção dos dados. Não foi possível determinar a profundidade
local precisamente, mas foram registradas as profundidades máximas atingidas pelo
Multianalisador ou pela Sonda, antes de tocarem o fundo.
Além dos meses em que a elevada vazão do rio não permitiu a amostragem, no
ponto 1 em junho de 2009 e em fevereiro, junho e novembro de 2011, e no ponto 2 em
junho de 2011, não foi possível a coleta de crustáceos devido à grande quantidade de
entulho que não possibilitou que a rede fosse puxada para dentro da embarcação. Esses
eventos foram, portanto, desconsiderados em todas as análises.
4.3. Processamento em laboratório
Foram elaboradas planilhas para as amostragens biológicas de Brachyura e
camarões. A identificação dos exemplares de Brachyura ao menor nível taxonômico
possível foi realizada de acordo com Melo (1996). Com o auxílio de um paquímetro e
uma balança digital com precisão de 0,01mm e 0,01g, respectivamente, foram
registrados a largura da carapaça (distância entre os espinhos laterais) e o peso úmido
total de cada organismo, sendo considerados apenas os exemplares íntegros. Os sexos
foram determinados por meio do exame da forma do abdômen.
Em laboratório os camarões foram identificados a partir de D’Incao (1995) e
Costa et al. (2003) e para cada exemplar foi registrado o sexo por meio da presença do
petasma (machos) ou telicum (fêmeas). Foram tomados o comprimento da carapaça (em
mm) e o valor do peso total (em g), utilizando um ictiômetro e uma balança digital,
respectivamente.
4.4. Análise de Dados
4.4.1 Dados Abióticos
Os dados foram analisados empregando estatística descritiva para verificar a
variação da profundidade e das variáveis abióticas (salinidade, temperatura, pH e
oxigênio dissolvido) da água de fundo no tempo e espaço. Para isso foram utilizadas
tabelas dos valores médios, mínimos e máximos desses parâmetros entre os pontos
amostrais e gráficos com valores médios dos pontos 1 a 3 e 4 a 6 ao longo dos meses

18
onde houve coletas. Foram gerados ainda gráficos com a descarga média de três dias e
diária para o rio Itajaí-Açu.
4.4.2 Dados Bióticos
A partir dos dados originais, foram calculados os valores de biomassa e
abundância das espécies nas seis estações durante cada um dos meses de amostragem
nos três anos e meio de estudo. Os dados foram padronizados em função do tempo
padrão de arrasto de 10 minutos e do quarteamento eventualmente realizado no campo.
Todas as análises foram realizadas a partir dessa tabela já padronizada.
Os dados foram analisados em princípio empregando estatística descritiva, onde
foram utilizadas tabelas para verificar a contribuição em termos de biomassa e
abundância dos decápodos capturados em cada ponto. Foram analisados também,
gráficos da contribuição percentual da abundância das principais famílias capturadas
nos anos de estudo. E com auxilio de gráficos de média e erro padrão, a variação mensal
e entre os pontos tanto para biomassa como para abundância do total de crustáceos e das
principais famílias capturadas.
Para efetuar a comparação da estrutura da fauna entre os pontos e entre os anos
foi utilizada a técnica de escalonamento multidimensional não-métrico (MDS). Para
execução dessa análise foram excluídas as espécies que não atingiram pelo menos 3%
da abundância total em nenhum ponto ou ano. Foi utilizado o coeficiente de
similaridade de Bray-Curtis e uma variável dummy foi introduzida para contornar o
excesso de zeros contidos na planilha de dados, assim como para reduzir o peso dado às
espécies raras (Clarke & Warwick, 1994). Para verificar a significância das diferenças
observadas no MDS foram realizadas análises de similaridade (ANOSIM). Em todos os
casos em que a ANOSIM foi significativa (p<0,05), a análise de contribuição percentual
de similaridade (SIMPER) foi utilizada para identificar as espécies responsáveis pelas
diferenças significativas encontradas nos grupos definidos no MDS. Essas análises
(MDS, ANOSIM e SIMPER) foram realizadas com o auxílio do software PRIMER
versão 6.1.6. (Clarke & Gorley, 2006). Para complementar a análise do MDS foram
gerados gráficos da abundância média das espécies representativas no SIMPER quando
o ANOSIM foi significativo.

19
4.4.3 Relação entre variáveis bióticas e abióticas
Uma Análise de Correspondência Canônica (ACC) foi utilizada para verificar a
relação das espécies com as variáveis abióticas salinidade, pH, temperatura e oxigênio
dissolvido. Foram utilizadas espécies que contribuíram com mais de 0,1% do total
capturado. Permutações de Monte Carlo foram executadas para testar a significância
(p<0,05) de cada fator ambiental, sendo somente os fatores significantes (no caso
salinidade, pH e temperatura) incluídos passo a passo (forward selection) no modelo
gerado (Leps & Smilauer, 2003). Para essa análise foi utilizado o software Canoco for
Windows 4.5 (ter Braak & Smilauer, 2002).
Gráficos do tipo Box-plot foram utilizados para verificar em quais condições de
salinidade, pH, temperatura e oxigênio dissolvido as espécies analisadas na ACC foram
capturadas.
4.4.4 Efeitos das enchentes e da dragagem de aprofundamento
Para avaliar os efeitos das enchentes de novembro de 2008 e setembro de 2011
sobre a composição e abundância das espécies de crustáceos, foi utilizada a técnica de
escalonamento multidimensional não-métrico (MDS). Para a enchente de novembro de
2008 foram utilizados os dados de abundância dos meses imediatamente seguintes à
enchente (dezembro/2008 e janeiro/2009) e do mesmo período dos anos seguintes
(dezembro/2009 e janeiro/2010; dezembro/2010 e janeiro/2011). Os fatores utilizados
para essa análise foram: “CE”, período após enchente, dezembro de 2008 e janeiro de
2009; “SE1”, período sem enchente, dezembro de 2009 e janeiro de 2010; e “SE2”,
período sem enchente, dezembro de 2010 e janeiro de 2011. Como os organismos
presentes nas porções mais externas e internas do estuário poderiam responder de forma
diferenciada às enchentes, os fatores foram divididos ainda nos setores “A” e “B”.
A abordagem utilizada para a enchente de setembro de 2011 foi semelhante à de
2008, entretanto, como na amostra do mês de agosto de 2008 os efeitos da enchente já
podiam ser observados (elevada vazão e baixa salinidade), para essa análise foram
considerados os dados referentes aos meses de agosto e setembro dos anos 2008, 2010 e
2011 (em 2009 não houve amostragem em setembro). Assim, os fatores utilizados para
o MDS da enchente de 2011 foram: “SE1”, período sem enchente, agosto e setembro de
2008; “SE2”, período sem enchente, agosto e setembro de 2010; e “CE”, período com
enchente, agosto e setembro de 2011. Da mesma forma que as demais análises, os
fatores foram divididos nos setores “A” e “B”. Ressalta-se que ao contrário do

20
observado em 2008 quando não havia condições de navegação, em 2011 as amostragens
puderam ser realizadas durante o período de enchente, permitindo o exame do eventual
efeito imediato do evento sobre a fauna.
A análise MDS também foi utilizada para verificar as eventuais alterações sobre
as associações de crustáceos ocorridas pela dragagem de aprofundamento. Para isso,
foram selecionados os períodos entre os meses de março e junho dos anos 2009, 2010 e
2011. Nos anos 2009 e 2011 (para o período supracitado) estava ocorrendo de forma
contínua a dragagem de aprofundamento do canal do rio Itajaí-Açu, enquanto que em
2010 a dragagem havia sido interrompida (Tabela 3). Assim, no MDS foram incluídos
os fatores: “CD1”, com dragagem em 2009; “SD”, sem dragagem em 2010; e “CD2”,
com dragagem em 2011. Além disso, como os pontos 1 a 3 estão diretamente sujeitos às
dragagens e os pontos 4 a 6 não sofrem impactos diretos dessa atividade, o estuário foi
dividido em setores “A” e “B”.
Em todos os casos em que foi realizado o MDS, foram selecionadas apenas as
espécies que contribuíram com mais de 3% da abundância total nos períodos com e sem
dragagem e com e sem enchentes. Além disso, para todo MDS, Análises de
Similaridade (ANOSIM; p<0,05) foram utilizadas para testar a significância das
diferenças observadas entre os fatores. A Análise de Contribuição Percentual de
Similaridade (SIMPER) foi utilizada para identificar as espécies responsáveis pelas
diferenças significativas apontadas pelo ANOSIM. Essas análises (MDS, ANOSIM e
SIMPER) foram realizadas com o auxílio do software PRIMER versão 6.1.6. Assim
como na análise dos dados bióticos, foram gerados gráficos da abundância média das
principais espécies que contribuíram significativamente na análise do MDS (Clarke &
Warwick, 1994; Clarke & Gorley, 2006).
5. RESULTADOS
5.1 – Variáveis abióticas
A profundidade média foi maior nos pontos externos (1, 2 e 3) e menor nos
pontos internos (4, 5 e 6), com uma diminuição gradual do ponto 4 para o ponto 6. A
maior profundidade (14,0m) foi observada no ponto 3, em frente ao Porto e a menor
profundidade (3,0m) ocorreu nos pontos 4, 5 e 6 (Tabela 4). A profundidade média foi
maior nos meses de dezembro de 2008 a agosto de 2009 e entre abril e novembro de
2011 nos pontos externos, e menor entre os meses de maio a setembro de 2008 nos

21
pontos internos (Figura 6). Observou-se um aumento gradativo da profundidade em
todo o estuário do início desse estudo em 2008 até o término do mesmo em 2011
(Figura 6).
A salinidade apresentou maiores valores próximo a foz (ponto 1) e menores
valores em direção ao seu interior (ponto 6) (Tabela 4). A variabilidade desse parâmetro
foi elevada (0-34) e o desvio padrão foi maior nos pontos 4, 5 e 6 (Tabela 4). Em geral,
a salinidade foi mais baixa no verão, nos meses de janeiro a março e maior no inverno,
entre julho e setembro. A salinidade média nos pontos 4-6 foi menor que 25 em
aproximadamente metade das amostras e, em nove ocasiões, foi igual a zero. Já os
pontos 1-3 apresentaram salinidade média mais estável, ocorrendo em 82,5% das
amostras valores entre 25 e 34 (Figura 7).
Os registros da variação espacial do pH médio da água de fundo seguiu o mesmo
padrão de distribuição dos valores da salinidade média, com maiores valores próximo a
foz, decrescendo para o interior do estuário, porém, a amplitude de variação entre os
pontos foi menor no pH. O desvio padrão foi maior nos pontos 4-6, que em algumas
ocasiões apresentaram valores inferiores 5,5 (Tabela 4). As ocasiões onde o pH foi
menor coincidiram com os eventos de baixa salinidade. Observou-se, por fim, uma
tendência de diminuição no pH ao longo do estudo como um todo (Figura 7).
A temperatura média da água de fundo apresentou comportamento similar entre
os pontos. Porém, foi observada uma pequena diferença nos valores da temperatura dos
pontos externos para os mais internos (Tabela 4). A temperatura da água apresentou
variação sazonal, com valores máximos no verão (26,5ºC) e mínimos (15,3ºC) durante o
inverno (Tabela 4 e Figura 7). O menor valor de temperatura foi observado no inverno
de 2010.
A concentração de oxigênio dissolvido (OD) da água de fundo apresentou ampla
variação entre os pontos e entre os meses de amostragem. O oxigênio dissolvido não
mostrou um padrão regular de distribuição entre os pontos, atingindo seu menor valor
(0,6mg/l) no ponto 3 (Tabela 4). Foram observados valores elevados no inverno de 2008
e de 2011 e menores entre fevereiro de 2009 a março de 2010 (Figura 7). Em algumas
ocasiões os valores médios nos pontos 1-3 apresentaram uma grande diferença para os
valores observados nos pontos 4-6, sendo maiores no setor mais interno do estuário.
A descarga média do rio Itajaí-Açu apresentou picos em março e abril de 2010,
no verão de 2011, e em agosto e setembro de 2011, quando foi registrada uma grande
enchente na região (Figura 8). Os dados da descarga para o final de 2009 e entre março

e maio de 2011 estavam indisponíveis e não foram incluídos.
novembro de 2008 não houve coleta e, por esse motivo, as informações para esses dois
meses também não foram mos
de descarga no final de 2008
diárias sobre essa variável.
Tabela 4. Valores médios (±DP), mínimos e máximos das variáveis ambientais nos pontos amostrestuário do rio Itajaí-Açu entre maio de 2008 a novembro de 2011.
Figura 6. Variação dos valores médios no estuário do rio Itajaí-Açu entre maio de 2008 e novembro de 2011. Em outubro e novembro de 2008 e em setembro de 2009 não foram reali
Variável 1
Profundidade (m)
10,1 (±1,7) (4,7 – 13,3)
10,3 (±1,8)(6,5
Salinidade 30,0 (±5,9) (3,0 – 34,4)
28,4 (±7,3)(0,2
pH 7,8 (±0,4) (6,8 – 8,7)
Temperatura (ºC)
21,6 (±2,7) (16,8 – 26,2)
21,6 (±2,8)(16,8
OD (mg/l) 5,6 (±1,7) (1,7 – 9,2)
estavam indisponíveis e não foram incluídos. Nos meses de outubro e
novembro de 2008 não houve coleta e, por esse motivo, as informações para esses dois
meses também não foram mostradas na Figura 8. Nota-se, entretanto, valores elevados
final de 2008, quando se observa a figura 2, que apresenta informações
Valores médios (±DP), mínimos e máximos das variáveis ambientais nos pontos amostrçu entre maio de 2008 a novembro de 2011.
dos valores médios da profundidade entre os pontos 1 a 3 e 4 a 6 das coletaAçu entre maio de 2008 e novembro de 2011. Em outubro e novembro de 2008 e
em setembro de 2009 não foram realizadas amostragens.
Ponto 2 3 4 5
10,3 (±1,8) (6,5 – 13,9)
10,3 (±1,6) (7,6 – 14,0)
7,8 (±1,5) (3,0 – 10,0)
7,4 (±1,8)(3,0 – 10,0)
28,4 (±7,3) (0,2 – 34,4)
25,2 (±8,9) (0,0 – 34,0)
20,1 (±12,4) (0,0 – 33,9)
16,7 (±12,9)(0,0 - 33,6)
7,7 (±0,5) (6,4 – 8,7)
7,6 (±0,6) (5,7 – 8,7)
7,4 (±0,8) (5,3 – 8,4)
7,3 (±0,8)(5,1 – 8,7)
21,6 (±2,8) (16,8 – 26,1)
21,5 (±2,8) (16,8 – 25,7)
21,4 (±2,9) (15,4 – 26,0)
21,4 (±3,0)(15,3 – 26,5)
5,3 (±1,7) (1,5 – 8,7)
4,9 (±1,9) (0,6 – 8,9)
5,6 (±1,8) (1,9 – 10,0)
5,8 (±1,8)(2,6 – 9,5)
22
Nos meses de outubro e
novembro de 2008 não houve coleta e, por esse motivo, as informações para esses dois
se, entretanto, valores elevados
que apresenta informações
Valores médios (±DP), mínimos e máximos das variáveis ambientais nos pontos amostrados no
entre os pontos 1 a 3 e 4 a 6 das coletas efetuadas
Açu entre maio de 2008 e novembro de 2011. Em outubro e novembro de 2008 e
6 7,4 (±1,8)
10,0) 7,3 (±1,6)
(3,3 – 10,6) 16,7 (±12,9)
33,6) 15,1 (±11,9) (0,0 - 33,3)
7,3 (±0,8) 8,7)
7,1 (±0,8) (5,0 - 8,4)
21,4 (±3,0) 26,5)
21,4 (±3,0) (15,3 – 26,4)
5,8 (±1,8) 9,5)
5,4 (±1,8) (3,1 – 9,5)
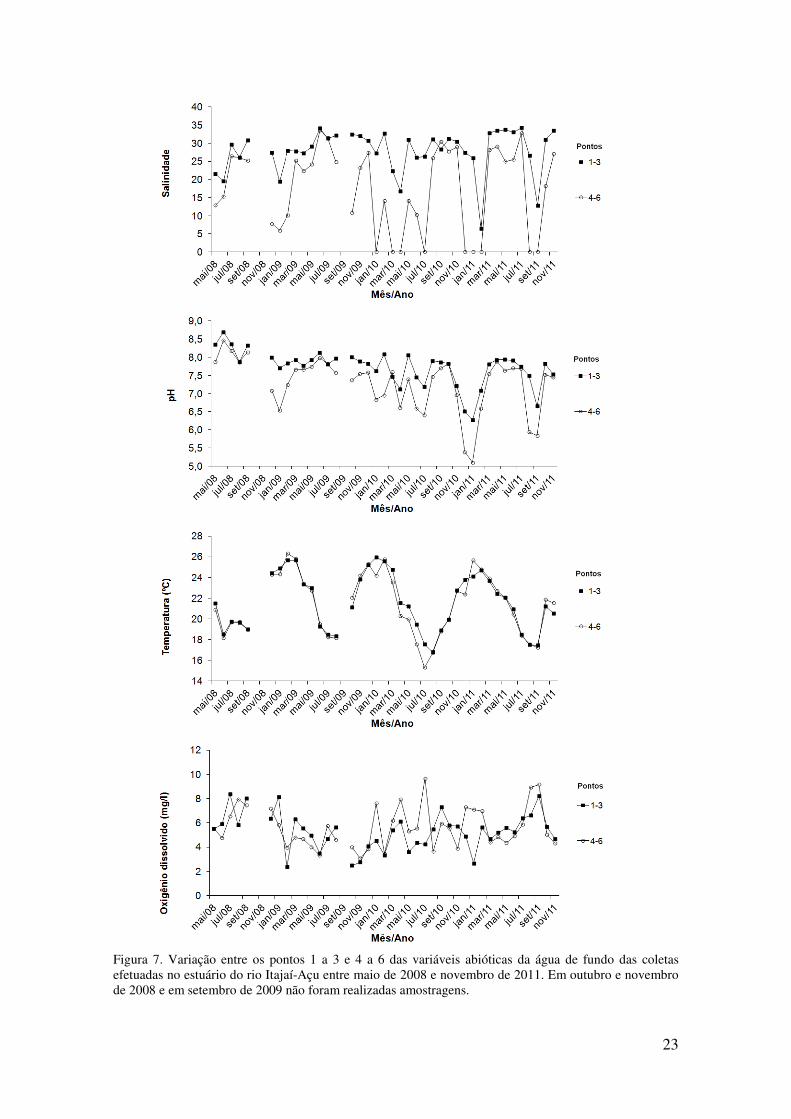
Figura 7. Variação entre os pontos 1 a 3 e 4 a 6efetuadas no estuário do rio Itajaíde 2008 e em setembro de 2009 não foram reali
7. Variação entre os pontos 1 a 3 e 4 a 6 das variáveis abióticas da água de fundoefetuadas no estuário do rio Itajaí-Açu entre maio de 2008 e novembro de 2011. Em outubro e novembro de 2008 e em setembro de 2009 não foram realizadas amostragens.
23
das variáveis abióticas da água de fundo das coletas
Açu entre maio de 2008 e novembro de 2011. Em outubro e novembro

24
Figura 8. Variação da descarga fluvial durante as coletas efetuadas no estuário do rio Itajaí-Açu entre maio de 2008 e novembro de 2011. Foi utilizada a média da descarga no dia da coleta junto com os dois dias anteriores. Os dados de descarga não estavam disponíveis para todas as datas onde foram efetuadas amostragens.
5.2 – Composição e distribuição dos crustáceos decápodos
Durante os três anos e meio de amostragens foram coletadas 20 espécies ou
morfotipos de crustáceos decápodos no estuário do rio Itajaí-Açu (Tabela 5) totalizando
3636 exemplares que contribuíram para uma biomassa total de 58.719,8g (Tabelas 6 e
7). A espécie Callinectes danae contribuiu com aproximadamente 48% da abundância e
58% da biomassa, Xiphopenaeus kroyeri com 29% da abundância e 4% da biomassa, e
Callinectes sapidus com 7% da abundância e 32% da biomassa. Nenhuma outra espécie
teve participação superior a 3% em termos de biomassa, e apenas as espécies
Callinectes ornatus e Nematopalaemon schmitti, além das três supracitadas,
contribuíram com mais de 3% do total capturado. Apenas 6 espécies em termos de
número e 4 em termos de biomassa contribuíram, juntas, com mais de 95% do total
capturado.
As famílias Penaeidae e Portunidae contribuíram juntas com 90,84% do total
(Tabela 6), sendo que a primeira foi a que mais contribuiu em números de indivíduos
em cada ano amostrado, chegando a 91% em 2008 (entre maio a dezembro deste ano)
(Figura 9). Penaeidae foi a segunda família com a maior contribuição percentual nos
anos, exceto em 2010. Nesse ano Palaemonidae teve um incremento acentuado no
número de indivíduos e passou da terceira para segunda posição em termos de
abundância. Outras famílias contribuíram com um máximo de 4,35% em 2011 (Figura
9).

25
Diferentes medidas foram tomadas de todas as espécies capturadas, entretanto
apenas sete espécies tiveram um número mínimo de 10 indivíduos diferentes pesados e
medidos (Tabela 8). Dentre as espécies do gênero Callinectes, C. sapidus apresentou os
indivíduos com maiores tamanhos e biomassas individuais, e C. ornatus os menores
(Tabela 8). O camarão rosa (F. paulensis) foi a espécie de camarão que apresentou
maior biomassa média e X. kroyeri a menor (Tabela 8).
Tabela 5. Listagem das espécies e/ou morfotipos coletados no estuário do rio Itajaí-Açu entre maio de 2008 a novembro de 2011.
Ordem/Infraordem/Família Nome científico Ordem Decapoda Infraordem Penaeidea
Família Penaeidae Xiphopenaeus kroyeri (Heller, 1862) Farfantepenaeus paulensis (Pérez Farfante, 1967) Artemesia longinaris Bate, 1888 Litopenaeus schmitti (Bunkenroad, 1936) Infraordem Caridea Caridea sp. Família Palaemonidae Palaemoninae sp.1 Palaemoninae sp.2 Nematopalaemon schmitti (Holthuis,1950) Macrobrachium acanthurus (Wiegmann, 1836) Macrobrachium olfersi (Wiegmann, 1836) Macrobrachium sp. Família Hippolytidae Hippolytidae sp. Exhippolysmata oplophoroides (Holthuis, 1948) Infraordem Brachyura Família Portunidae
Callinectes danae Smith, 1869 Callinectes bocourti A. Milne Edwards, 1879 Callinectes ornatus Ordway, 1863 Callinectes sapidus Rathbun, 1896 Charybdis hellerii A. Milne Edwards, 1867 Família Xanthidae Xanthidae sp. Família Majidae Libinia ferreirae Brito Capello, 1871

26
Tabela 6. Abundância numérica (nº/10min) das espécies em cada ponto e sua participação relativa na abundância total (%) no estuário do rio Itajaí-Açu entre maio de 2008 a novembro de 2011.
Espécies Ponto 1 Ponto 2 Ponto 3 Ponto 4 Ponto 5 Ponto 6 Total %
Callinectes danae 126 128 368 407 312 396 1738 47,8
Xiphopenaeus kroyeri 547 112 94 194 78 23 1048 28,8
Callinectes sapidus 4 6 27 20 142 45 244 6,7
Nematopalaemon schmitti 72 95 2 16 2 16 203 5,6
Callinectes ornatus 104 33 4 4 0 6 151 4,2
Farfantepenaeus paulensis 10 10 31 8 12 31 102 2,8
Macrobrachium acanthurus 0 0 0 4 58 0 62 1,7
Libinia ferreirae 8 2 0 4 6 0 20 0,6
Macrobrachium sp. 0 0 0 2 6 4 12 0,3
Palaemoninae sp1 0 0 0 0 4 6 10 0,3
Exhippolysmata oplophoroides 10 0 0 0 0 0 10 0,3
Callinectes bocourti 4 0 3 0 0 0 7 0,2
Macrobrachium olfersi 0 0 0 0 4 2 6 0,2
Palaemoninae sp2 0 0 0 0 6 0 6 0,2
Charybdis hellerii 4 0 0 0 0 0 4 0,1
Xantidae sp. 0 0 0 0 0 4 4 0,1
Caridea sp. 1 0 1 2 0 0 4 0,1
Litopenaeus schmitti 2 0 1 0 0 0 3 0,1
Artemesia longinaris 2 0 0 0 0 0 2 0,1
Hippolytidae sp. 0 0 0 1 0 0 1 0,0
Total 894 386 531 662 630 533 3636
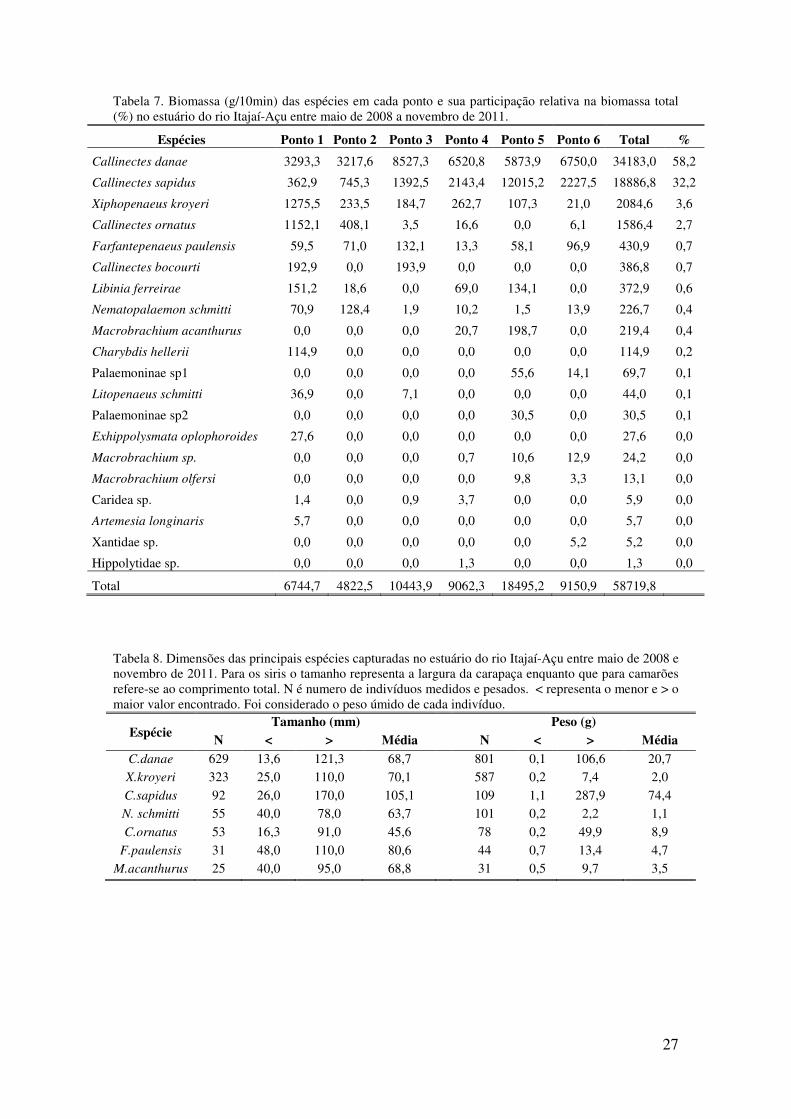
27
Tabela 7. Biomassa (g/10min) das espécies em cada ponto e sua participação relativa na biomassa total (%) no estuário do rio Itajaí-Açu entre maio de 2008 a novembro de 2011.
Espécies Ponto 1 Ponto 2 Ponto 3 Ponto 4 Ponto 5 Ponto 6 Total %
Callinectes danae 3293,3 3217,6 8527,3 6520,8 5873,9 6750,0 34183,0 58,2
Callinectes sapidus 362,9 745,3 1392,5 2143,4 12015,2 2227,5 18886,8 32,2
Xiphopenaeus kroyeri 1275,5 233,5 184,7 262,7 107,3 21,0 2084,6 3,6
Callinectes ornatus 1152,1 408,1 3,5 16,6 0,0 6,1 1586,4 2,7
Farfantepenaeus paulensis 59,5 71,0 132,1 13,3 58,1 96,9 430,9 0,7
Callinectes bocourti 192,9 0,0 193,9 0,0 0,0 0,0 386,8 0,7
Libinia ferreirae 151,2 18,6 0,0 69,0 134,1 0,0 372,9 0,6
Nematopalaemon schmitti 70,9 128,4 1,9 10,2 1,5 13,9 226,7 0,4
Macrobrachium acanthurus 0,0 0,0 0,0 20,7 198,7 0,0 219,4 0,4
Charybdis hellerii 114,9 0,0 0,0 0,0 0,0 0,0 114,9 0,2
Palaemoninae sp1 0,0 0,0 0,0 0,0 55,6 14,1 69,7 0,1
Litopenaeus schmitti 36,9 0,0 7,1 0,0 0,0 0,0 44,0 0,1
Palaemoninae sp2 0,0 0,0 0,0 0,0 30,5 0,0 30,5 0,1
Exhippolysmata oplophoroides 27,6 0,0 0,0 0,0 0,0 0,0 27,6 0,0
Macrobrachium sp. 0,0 0,0 0,0 0,7 10,6 12,9 24,2 0,0
Macrobrachium olfersi 0,0 0,0 0,0 0,0 9,8 3,3 13,1 0,0
Caridea sp. 1,4 0,0 0,9 3,7 0,0 0,0 5,9 0,0
Artemesia longinaris 5,7 0,0 0,0 0,0 0,0 0,0 5,7 0,0
Xantidae sp. 0,0 0,0 0,0 0,0 0,0 5,2 5,2 0,0
Hippolytidae sp. 0,0 0,0 0,0 1,3 0,0 0,0 1,3 0,0
Total 6744,7 4822,5 10443,9 9062,3 18495,2 9150,9 58719,8
Tabela 8. Dimensões das principais espécies capturadas no estuário do rio Itajaí-Açu entre maio de 2008 e novembro de 2011. Para os siris o tamanho representa a largura da carapaça enquanto que para camarões refere-se ao comprimento total. N é numero de indivíduos medidos e pesados. < representa o menor e > o maior valor encontrado. Foi considerado o peso úmido de cada indivíduo.
Espécie Tamanho (mm) Peso (g)
N < > Média N < > Média C.danae 629 13,6 121,3 68,7 801 0,1 106,6 20,7 X.kroyeri 323 25,0 110,0 70,1 587 0,2 7,4 2,0
C.sapidus 92 26,0 170,0 105,1 109 1,1 287,9 74,4 N. schmitti 55 40,0 78,0 63,7 101 0,2 2,2 1,1
C.ornatus 53 16,3 91,0 45,6 78 0,2 49,9 8,9
F.paulensis 31 48,0 110,0 80,6 44 0,7 13,4 4,7 M.acanthurus 25 40,0 95,0 68,8 31 0,5 9,7 3,5

Figura 9. Contribuição percentual da abundância das principais famílias de crustáceos decápodos no rio Itajaí-Açu entre maio de 2008 e novembro de 2011.
5.2.1- Variação espacial
Em média, a abundância dos crustáceos
ponto 1, onde foi observado o maior valor,
Penaeidae (Figura 10). Entretanto
variação similar a biomassa média de Portunidae,
menores no ponto 2. A abundância média dos Portunidae foi relativamente similar entre
os pontos, sendo maior nos pontos mais a montante do estuário e menor próximo
foz. A família Penaeidae foi a que apresentou o padrão de
da abundância total, apesar de ter contribuindo significativamente
biomassa dessa família também foi maior no ponto 1 e menor no ponto 6.
abundância como a biomassa média de Palaemonidae
distribuição irregular entre os pontos, apresentando maiores valores de abundânc
biomassa no ponto 5 e chegando a
Através do MDS foi possível verificar uma
amostras utilizando os pontos como fator
concentrados na parte inferior
(Figura 11). A ANOSIM comprovou esse resulta
significativas entre os pontos adjacentes
diante, e indicando diferenças (p<0,05) entre os pontos mais afastado
Além disso, as estações 3 a 6 pareceram ser mais similares entre si
SIMPER indicou que as espécies
Contribuição percentual da abundância das principais famílias de crustáceos decápodos no rio Açu entre maio de 2008 e novembro de 2011.
ndância dos crustáceos foi similar entre os pontos
foi observado o maior valor, reflexo da elevada contribuição da família
. Entretanto, a biomassa média total apresentou padrão de
variação similar a biomassa média de Portunidae, com maiores valor
A abundância média dos Portunidae foi relativamente similar entre
os pontos, sendo maior nos pontos mais a montante do estuário e menor próximo
A família Penaeidae foi a que apresentou o padrão de distribuição
da abundância total, apesar de ter contribuindo significativamente apenas no ponto 1. A
biomassa dessa família também foi maior no ponto 1 e menor no ponto 6.
abundância como a biomassa média de Palaemonidae foram extremamente baixa
distribuição irregular entre os pontos, apresentando maiores valores de abundânc
biomassa no ponto 5 e chegando a praticamente zero no ponto 3 (Figura
Através do MDS foi possível verificar uma leve gradação na distribuição das
amostras utilizando os pontos como fatores. Os pontos 1, 2 e 3 tenderam a ficar
concentrados na parte inferior do gráfico e os pontos 4, 5 e 6 mais na
A ANOSIM comprovou esse resultado, não mostrando diferenças
s pontos adjacentes, por exemplo, pontos 1 e 2, 2 e 3 e assim por
diante, e indicando diferenças (p<0,05) entre os pontos mais afastados uns
lém disso, as estações 3 a 6 pareceram ser mais similares entre si
cou que as espécies C. danae, C. sapidus e X. kroyeri foram
28
Contribuição percentual da abundância das principais famílias de crustáceos decápodos no rio
similar entre os pontos exceto no
contribuição da família
a biomassa média total apresentou padrão de
valores no ponto 5 e
A abundância média dos Portunidae foi relativamente similar entre
os pontos, sendo maior nos pontos mais a montante do estuário e menor próximo à sua
stribuição mais similar ao
apenas no ponto 1. A
biomassa dessa família também foi maior no ponto 1 e menor no ponto 6. Tanto a
mente baixas, com
distribuição irregular entre os pontos, apresentando maiores valores de abundância e
amente zero no ponto 3 (Figura 10).
gradação na distribuição das
tenderam a ficar mais
na parte superior
do, não mostrando diferenças
, por exemplo, pontos 1 e 2, 2 e 3 e assim por
s uns dos outros.
lém disso, as estações 3 a 6 pareceram ser mais similares entre si (Tabela 9). O
foram, em geral, as

29
principais responsáveis pelas diferenças observadas (Tabela 9). Observando a
abundância média dessas espécies nos pontos é possível verificar que C. danae ocorreu
principalmente na zona intermediária e no interior do estuário, assim como C. sapidus,
que apresentou pico de abundância no ponto 5. X. kroyeri foi observado principalmente
no ponto 1. C. ornatus e N. schmitti, que também foram indicadas no SIMPER,
ocorreram principalmente nos pontos 1 e 2, enquanto M. acanthurus só foi observado
no ponto 4 e principalmente no ponto 5 (Figura 12).
Assim como observado na distribuição espacial de alguns parâmetros
ambientais, o MDS utilizando os pontos como fator mostrou que existe um padrão de
agrupamento diferente das espécies entre os pontos mais internos e externos do estuário.
Considerando, portanto, os pontos agrupados em setores A (1, 2 e 3) e B (4, 5 e 6), um
novo MDS foi feito onde nota-se a tendência de divisão entre os mesmos (Figura 13), a
qual foi significativamente comprovada pela ANOSIM (Tabela 10). C. danae e C.
sapidus apresentaram maior abundância no setor B, e X. kroyeri e N. schmitti no setor A
(Figura 14).

30
Figura 10. Abundância e biomassa médias nos pontos amostrados para o total de crustáceos capturados e para as famílias Portunidae, Penaeidae e Palaemonidae. A barra representa o erro padrão.

31
Figura 11. Análise MDS realizada com todas as espécies cuja abundância representou, em ao menos um dos pontos amostrais, mais do que 3% do total. Os fatores utilizados foram os pontos, numerados de 1 (mais a jusante) a 6 (mais a montante). Tabela 9. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores do MDS representado na figura 11. No SIMPER foram incluídas todas as espécies até atingirem 90% de representatividade nos resultados. O SIMPER não foi considerado quando o resultado do ANOSIM teve p>0,05 (**).
Pontos ANOSIM
SIMPER (Dissimilaridade de Bray- Curtis) R p
1 X 2 0,002 0,344 **
1 X 3 0,059 0,012 C. danae (36,35%); X. kroyeri (29,98%); N. schmitti
(9,77%); C. sapidus (9,29%); C. ornatus (9,14%)
1 X 4 0,062 0,013 C. danae (40,39%); X. kroyeri (31,79%); N. schmitti
(10,65%); C. ornatus (7,80%)
1 X 5 0,117 0,003 C. danae (32,95%); X. kroyeri (25,17%); C. sapidus (19,63%); N. schmitti (9,24%); C. ornatus (6,82%)
1 X 6 0,091 0,004 C. danae (34,27%); X. kroyeri (26,31%); C. sapidus (16,34%); N. schmitti (12,31%); C. ornatus (8,69%)
2 X 3 0,017 0,141 ** 2 X 4 0,018 0,103 **
2 X 5 0,092 0,008 C. danae (39,92%); C. sapidus (21,36%); X. kroyeri (16,57%); N. schmitti (10,0%) C. ornatus (5,10%)
2 X 6 0,051 0,015 C. danae (42,43%); C. sapidus (17,40%); X. kroyeri
(16,57%); N. schmitti (13,63%) 3 X 4 -0,011 0,768 ** 3 X 5 0,027 0,096 ** 3 X 6 -0,001 0,419 **
4 X 5 0,035 0,043 C. danae (49,27%); C. sapidus (24,91%); X. kroyeri
(13,41%); M. acanthurus (6,95%) 4 X 6 0,006 0,251 ** 5 X 6 0,017 0,128 **
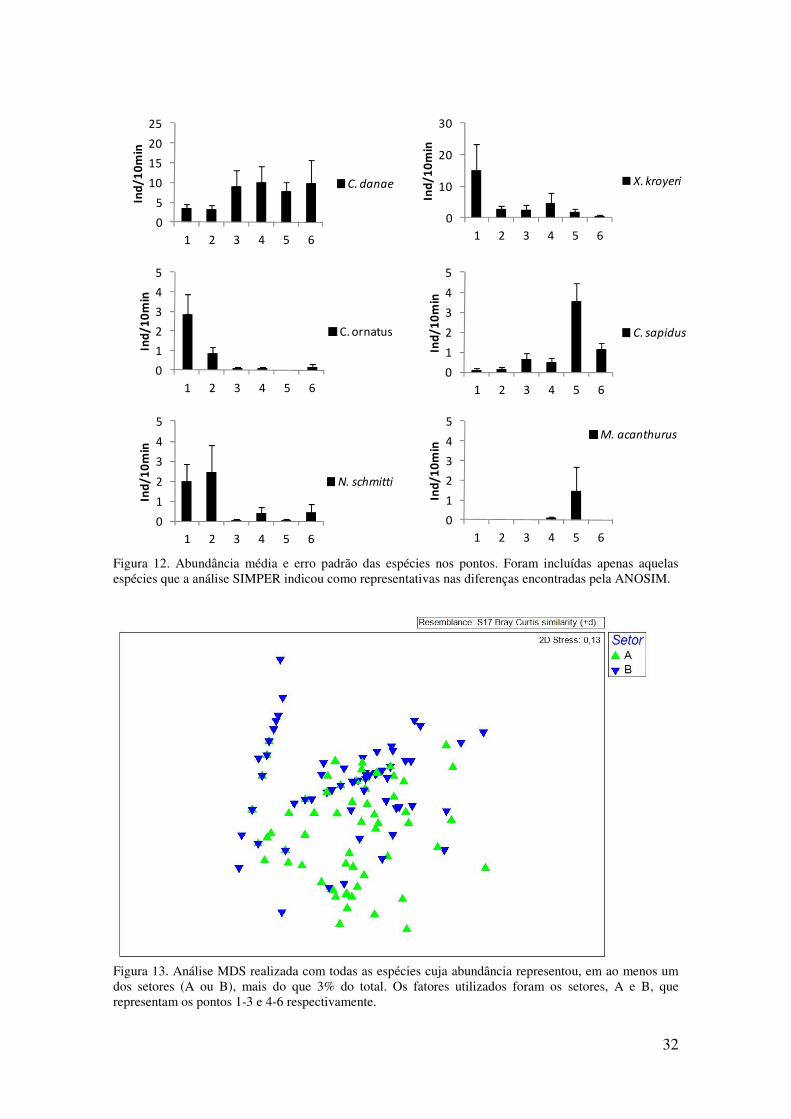
32
Figura 12. Abundância média e erro padrão das espécies nos pontos. Foram incluídas apenas aquelas espécies que a análise SIMPER indicou como representativas nas diferenças encontradas pela ANOSIM.
Figura 13. Análise MDS realizada com todas as espécies cuja abundância representou, em ao menos um dos setores (A ou B), mais do que 3% do total. Os fatores utilizados foram os setores, A e B, que representam os pontos 1-3 e 4-6 respectivamente.
0
5
10
15
20
25
1 2 3 4 5 6
Ind/10min
C. danae
0
10
20
30
1 2 3 4 5 6
Ind/10min
X. kroyeri
0
1
2
3
4
5
1 2 3 4 5 6
Ind/10min
C. ornatus
0
1
2
3
4
5
1 2 3 4 5 6
Ind/10min
C. sapidus
0
1
2
3
4
5
1 2 3 4 5 6
Ind/10min
N. schmitti
0
1
2
3
4
5
1 2 3 4 5 6
Ind/10min
M. acanthurus

Tabela 10. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores do MDS representado na figura 13. No SIMPER foram incluídas todas as espécies até atingirem 90% de representatividade nos resultados.
Setor ANOSIM
R
A x B 0,032
Figura 14. Abundância média e erro padrão das espécies nos aquelas espécies que a análise SIMPER indicou como representativas nas diferenças encontradas pela ANOSIM.
5.2.2- Variação temporal
A variação temporal da abundância e
estuário foi similar ao longo do
dos indivíduos nas amostragens, o primeiro entre agosto e setembro de 2008 e o
segundo entre fevereiro de 2009 e jul
principalmente da elevada
a abundância e biomassa média total permaneceram em patamares inferiores e
relativamente homogêneos,
aumentos na captura (Figura
a abundância média total foi muito baixa, com valores próximo
maior abundância total foi
principalmente de Portunidae. Para esta família a abundâ
meses de março e julho, com picos nos anos de 2008 e 2009.
ocorreu preferencialmente
. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores do MDS No SIMPER foram incluídas todas as espécies até atingirem 90% de
representatividade nos resultados. ANOSIM
SIMPER (Dissimilaridade de Brayp
2 0,002 C. danae (45,29%); X. kroyeri (20,1(17,70%); N. schmitti (8,78%)
. Abundância média e erro padrão das espécies nos setores A e B. Foram incluídas apenas aquelas espécies que a análise SIMPER indicou como representativas nas diferenças encontradas pela
A variação temporal da abundância e da biomassa média dos crustáceos no
similar ao longo do estudo. Nota-se dois períodos marcantes da ocorrência
dos indivíduos nas amostragens, o primeiro entre agosto e setembro de 2008 e o
reiro de 2009 e julho de 2009 (Figura 15). Ambos tiveram reflexo
captura das espécies de Portunidae. A partir desses períodos
abundância e biomassa média total permaneceram em patamares inferiores e
relativamente homogêneos, com três ocasiões, porém, onde foram observados pequenos
(Figura 15). Nos meses seguintes às enchentes de 2008 e
ncia média total foi muito baixa, com valores próximos ou iguais
maior abundância total foi registrada em julho de 2009, com contribuição
idae. Para esta família a abundância foi sempre maior entre os
julho, com picos nos anos de 2008 e 2009. A família Penaeid
preferencialmente entre fevereiro e abril, com abundâncias significativas nesse
33
. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores do MDS No SIMPER foram incluídas todas as espécies até atingirem 90% de
SIMPER (Dissimilaridade de Bray- Curtis)
7%); C. sapidus
. Foram incluídas apenas aquelas espécies que a análise SIMPER indicou como representativas nas diferenças encontradas pela
biomassa média dos crustáceos no
marcantes da ocorrência
dos indivíduos nas amostragens, o primeiro entre agosto e setembro de 2008 e o
mbos tiveram reflexo
. A partir desses períodos
abundância e biomassa média total permaneceram em patamares inferiores e
onde foram observados pequenos
s de 2008 e de 2011,
s ou iguais a zero. A
em julho de 2009, com contribuição
sempre maior entre os
A família Penaeidae
, com abundâncias significativas nesse

34
mesmo período de 2009, onde, inclusive, teve uma grande contribuição para a
abundância média total. A família Palaemonidae foi a de menor representatividade no
total, ocorrendo de forma isolada nos meses de janeiro e maio de 2010, entre setembro
de 2010 e janeiro de 2011, e com um pico em março de 2011, onde sua abundância foi
equivalente a 43% do total capturado. Nessa família as diferenças na flutuação da
abundância e da biomassa média foram mais evidentes (Figura 14).
Tendo os anos como fatores, o MDS mostrou dois agrupamentos distintos: as
amostras de 2008 e 2009 ficaram principalmente na parte central do gráfico e as
amostras de 2010 e 2011 tenderam a se distribuir mais à direita do mesmo (Figura 16).
A ANOSIM acusou a inexistência de diferenças entre 2008 e 2009 e entre 2010 e 2011.
Entre os dois grupos, entretanto, a diferença foi significativa (Tabela 11). A diferença
entre os anos mais distantes foi devido principalmente à elevada abundância média de
C. danae em 2008 e 2009 e forte queda nos anos seguintes (Tabela 11, Figura 17).
Outras espécies indicadas pelo SIMPER foram X. kroyeri, C. ornatus e F. paulensis
com maior abundância nos dois primeiros anos, e N. schmitti e C. sapidus com maior
abundância em 2010 e 2011 (Figura 17).
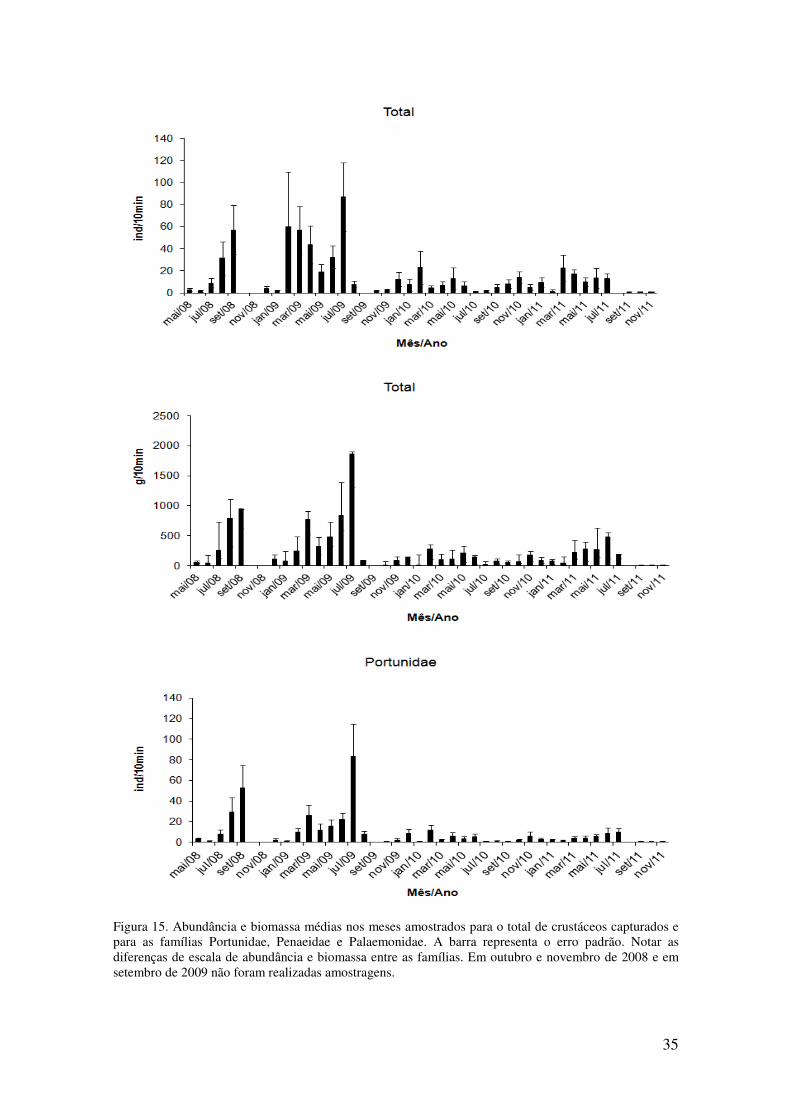
35
Figura 15. Abundância e biomassa médias nos meses amostrados para o total de crustáceos capturados e para as famílias Portunidae, Penaeidae e Palaemonidae. A barra representa o erro padrão. Notar as diferenças de escala de abundância e biomassa entre as famílias. Em outubro e novembro de 2008 e em setembro de 2009 não foram realizadas amostragens.

36
Figura 15 (continuação). Abundância e biomassa médias nos meses amostrados para o total de crustáceos capturados e para as famílias Portunidae, Penaeidae e Palaemonidae. A barra representa o erro padrão. Notar as diferenças de escala de abundância e biomassa entre as famílias. Em outubro e novembro de 2008 e em setembro de 2009 não foram realizadas amostragens.
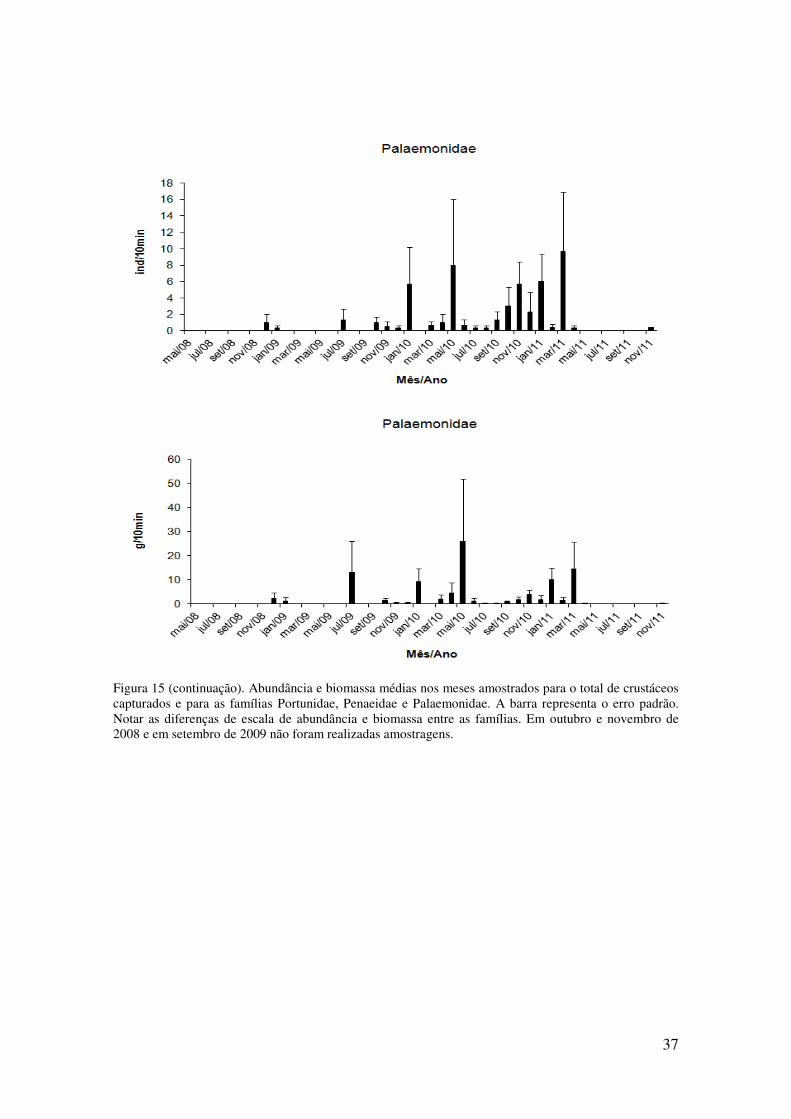
37
Figura 15 (continuação). Abundância e biomassa médias nos meses amostrados para o total de crustáceos capturados e para as famílias Portunidae, Penaeidae e Palaemonidae. A barra representa o erro padrão. Notar as diferenças de escala de abundância e biomassa entre as famílias. Em outubro e novembro de 2008 e em setembro de 2009 não foram realizadas amostragens.
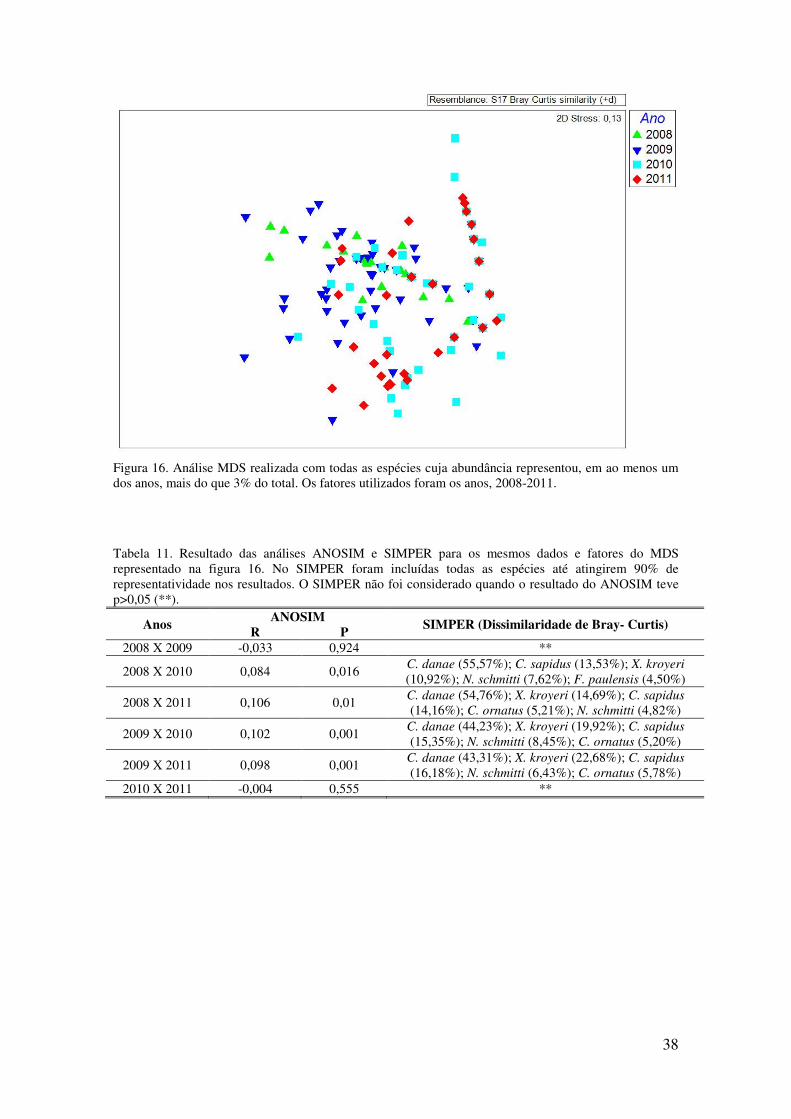
38
Figura 16. Análise MDS realizada com todas as espécies cuja abundância representou, em ao menos um dos anos, mais do que 3% do total. Os fatores utilizados foram os anos, 2008-2011. Tabela 11. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores do MDS representado na figura 16. No SIMPER foram incluídas todas as espécies até atingirem 90% de representatividade nos resultados. O SIMPER não foi considerado quando o resultado do ANOSIM teve p>0,05 (**).
Anos ANOSIM
SIMPER (Dissimilaridade de Bray- Curtis) R P
2008 X 2009 -0,033 0,924 **
2008 X 2010 0,084 0,016 C. danae (55,57%); C. sapidus (13,53%); X. kroyeri (10,92%); N. schmitti (7,62%); F. paulensis (4,50%)
2008 X 2011 0,106 0,01 C. danae (54,76%); X. kroyeri (14,69%); C. sapidus (14,16%); C. ornatus (5,21%); N. schmitti (4,82%)
2009 X 2010 0,102 0,001 C. danae (44,23%); X. kroyeri (19,92%); C. sapidus (15,35%); N. schmitti (8,45%); C. ornatus (5,20%)
2009 X 2011 0,098 0,001 C. danae (43,31%); X. kroyeri (22,68%); C. sapidus (16,18%); N. schmitti (6,43%); C. ornatus (5,78%)
2010 X 2011 -0,004 0,555 **

39
Figura 17. Abundância média e erro padrão das espécies nos anos. Foram incluídas apenas aquelas espécies que a análise SIMPER indicou como representativas nas diferenças encontradas pela ANOSIM.
5. 3. Relações entre as variáveis abióticas e os crustáceos decápodos
Das espécies encontradas, 13 contribuíram com mais de 0,1% da abundância
total (Tabela 6). A relação dessas espécies com as variáveis Salinidade, pH,
Temperatura e Oxigênio Dissolvido foi verificada por meio de uma Análise de
Correspondência Canônica (ACC). Através de permutações de Monte Carlo concluiu-se
que o parâmetro oxigênio dissolvido não foi significativo (p>0,05) para o modelo e,
portanto, essa variável não foi incluída na ACC (Figura 18).
Entre as espécies de portunídeos, C. danae, C. ornatus e C. bocourti ficaram
próximas na ACC (Figura 18). Elas ocorreram quase que exclusivamente em maiores
salinidades e pH (especialmente as duas últimas espécies), temperatura entre 18 e 25ºC
e condições intermediárias de oxigênio dissolvido entre 4 e 6 mg/l (Figura 19). Somente
C. sapidus apresentou-se deslocado das demais espécies de Portunidae, associado a
valores de pH e salinidade mais baixos (Figura 18). De fato, ao contrário das outras
espécies do gênero, C. sapidus ocorreu em um amplo espectro de salinidade e pH,
0
5
10
15
20
25
2008 2009 2010 2011
Ind/10min
C. danae
0
5
10
15
20
25
2008 2009 2010 2011
Ind/10min
X. kroyeri
0,0
0,5
1,0
1,5
2,0
2,5
2008 2009 2010 2011
Ind/10min
C. sapidus
0,0
0,5
1,0
1,5
2,0
2,5
2008 2009 2010 2011
Ind/10min
C. ornatus
0,0
0,5
1,0
1,5
2,0
2,5
2008 2009 2010 2011
Ind/10min
F. paulensis
0,0
0,5
1,0
1,5
2,0
2,5
2008 2009 2010 2011
Ind/10min
N. schmitti

40
enquanto em relação a temperatura e oxigênio dissolvido, teve ocorrência similar aos
demais portunídeos (Figura 19).
Em relação à família Penaeidae, F. paulensis ficou posicionada no centro da
ACC, sem relação com nenhuma variável ambiental. A posição de X. kroyeri indica
uma tendência de associação à temperatura, principalmente em condições de maior pH e
salinidade (Figura 18). Ambas as espécies foram capturadas quase que exclusivamente
em condições de elevada salinidade e pH, com exceção de algumas amostras onde
foram capturados indivíduos de X. kroyeri em salinidades baixas (Figura 19). Embora
tenham ocorrido em diversas temperaturas, X. kroyeri foi capturado com mais
frequência em temperaturas mais elevadas (acima de 22ºC) enquanto F. paulensis foi
capturado principalmente em menores temperaturas (abaixo de 20ºC) (Figura 19).
Ambas as espécies ocorreram com maior frequência em valores intermediários de
oxigênio dissolvido, próximo a 5 mg/L (Figura 18).
Com exceção de Palaemoninae sp2 e N. schmitti, todas as espécies de
Palaemonidae, principalmente aquelas pertencentes ao gênero Macrobrachuim, ficaram
em posição oposta aos eixos da salinidade e pH no gráfico da ACC, indicando uma forte
correlação negativa com esses parâmetros. Palaemoninae sp. 1, ocorreu em salinidades
entre 0 e 10, pH relativamente baixo, e principalmente em condições de elevada
temperatura e oxigênio dissolvido (Figura 19). Macrobrachium sp. e N. schmitti
ocorreram em um grande espectro de valores de oxigênio dissolvido e temperatura; M.
acanthurus e Palaemoninae sp2 foram encontrados em diferentes condições de
temperatura e, esta última, também em amostras com maiores valores de salinidade e
pH. Por fim, N. schmitti ocorreu exclusivamente em amostras com salinidades elevadas
e pH entre 6,5 e 8,5 (Figura 19).
Além das famílias supracitadas, apenas a família Majidae teve participação
superior a 0,1% do total capturado, representada por L. ferreirae. Essa espécie
apresentou-se separada de todas as demais na ACC, ocorrendo em posição oposta ao
vetor que representa a temperatura e deslocada em direção aos vetores da salinidade e
pH (Figura 18). Ela foi capturada em salinidade bem elevada, com pH próximo a 8,0,
temperatura de aproximadamente 19ºC e em altos valores de oxigênio dissolvido
(Figura 19).

41
Figura 18. Bi-plot da ACC referente às variáveis ambientais e biológicas. A análise foi realizada utilizando todas as espécies cuja abundância representou ao menos 0,1% do total. Das 4 variáveis ambientais testadas, o oxigênio dissolvido foi removido do modelo por apresentar p>0,05 nas permutações de Monte Carlo. Os vetores representam as variáveis ambientais salinidade (Sal), pH, e temperatura (Temp).
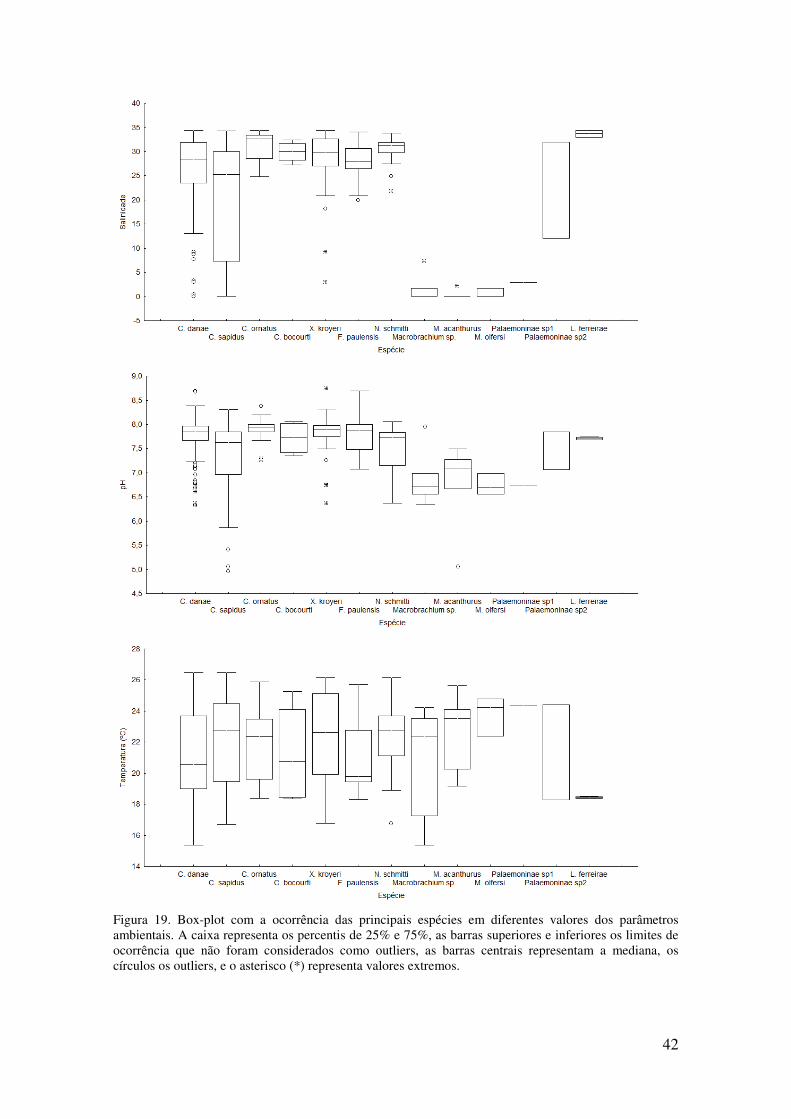
42
Figura 19. Box-plot com a ocorrência das principais espécies em diferentes valores dos parâmetros ambientais. A caixa representa os percentis de 25% e 75%, as barras superiores e inferiores os limites de ocorrência que não foram considerados como outliers, as barras centrais representam a mediana, os círculos os outliers, e o asterisco (*) representa valores extremos.

43
Figura 19 (continuação). Box-plot com a ocorrência das principais espécies em diferentes valores dos parâmetros ambientais. A caixa representa os percentis de 25% e 75%, as barras superiores e inferiores os limites de ocorrência que não foram considerados como outliers, as barras centrais representam a mediana, os círculos os outliers, e o asterisco (*) representa valores extremos.
5.4. Efeitos das Enchentes
5.4.1. Enchente de 2008
A enchente que ocorreu em novembro de 2008 foi responsável por grandes
prejuízos econômicos na região do vale do Itajaí, e foi ocasionada por chuvas acima da
média no período. Devido a forte vazão do rio, não houve amostragens em outubro e
novembro de 2008 e, para verificar possíveis efeitos da enchente sobre os crustáceos
foram examinados os dois meses seguintes ao evento (dezembro de 2008 e janeiro de
2009).
Como já evidenciado anteriormente, verificou-se uma grande queda na
abundância e biomassa dos crustáceos logo após a enchente de 2008, com recuperação
nos meses seguintes (Figura 15). Nos demais anos a abundância foi baixa nos meses de
dezembro e janeiro, e um MDS foi necessário para verificar se realmente houve
impactos decorrentes das enchentes.
Como já ressaltado, para a análise com o MDS, ANOSIM e SIMPER, as
amostras foram separadas em setores A e B e comparadas entre o período logo após a
enchente (dezembro de 2008 e janeiro de 2009, “CE”- com enchente) e os mesmos
períodos nos anos seguintes (2009/2010, “SE1”; 2010/2011, “SE2”- sem enchente 1 e
2). Assim, as amostras foram agrupadas em CEA, CEB, SE1A, SE1B, SE2A, SE2B.
O MDS da enchente de 2008 não apresentou nenhum padrão claro, indicando
ausência de um efeito significativo do evento nos dois meses seguintes ao mesmo

44
(Figura 20 e Tabela 12). A ANOSIM revelou diferença significativa apenas para dois
grupos: CEB x SE1A; e SE1A x SE2B (Tabela 12). O SIMPER indicou que as espécies
do gênero Callinectes foram as principais responsáveis por essas diferenças. C. sapidus
e C. danae foram as únicas espécies coletadas em situação de pós-enchente. Não foram
capturados indivíduos de C. ornatus e C. danae nas amostras agrupadas como CEB e
como SE2B, e indivíduos de C. sapidus em SE1A (Figura 21). X. kroyeri e N. schmitti
também foram indicados pelo SIMPER (Tabela 12).
Figura 20. Análise MDS realizada com todas as espécies cuja abundância representou, em ao menos um setor e período, mais do que 3% do total. Os fatores utilizados foram: “CEA” e “CEB”, dezembro de 2008 a janeiro de 2009 nos setores A e B respectivamente; “SE1A” e “SE1B”, dezembro de 2009 a janeiro de 2010 nos setores A e B respectivamente; “SE2A” e “SE2B”, dezembro de 2010 a janeiro de 2011 nos setores A e B respectivamente.

45
Tabela 12. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores do MDS representado na figura 20. No SIMPER foram incluídas todas as espécies até atingirem 90% de representatividade nos resultados. O SIMPER não foi considerado quando o resultado do ANOSIM teve p>0,05 (**).
Grupos ANOSIM
SIMPER (Dissimilaridade de Bray- Curtis) R p
CEA x CEB -0,041 0,727 ** CEA x SE1A 0,217 0,087 ** CEA x SE1B -0,03 0,513 ** CEA x SE2A 0,105 0,182 ** CEA x SE2B 0,001 0,424 **
CEB x SE1A 0,243 0,041 C. danae (38,12%); C. sapidus (17,61%); F. paulensis
(15,95%); N. schmitti (15,10%)X. kroyeri (13,21%) CEB x SE1B 0,013 0,307 ** CEB x SE2A 0,164 0,182 ** CEB x SE2B -0,058 0,545 **
SE1A x SE1B 0,033 0,288 ** SE1A x SE2A -0,057 0,621 **
SE1A x SE2B 0,236 0,039 C. danae (33,06%); C. sapidus (23,71%); N. scmitti
(14,02%); F. paulensis (12,77%); X. kroyeri (12,15%) SE1B x SE2A 0,047 0,275 ** SE1B x SE2B 0,016 0,346 ** SE2A x SE2B 0,174 0,188 **
Figura 21. Abundância média e erro padrão das espécies nos períodos e setores equivalentes a Figura 20. Foram incluídas apenas aquelas espécies que a análise SIMPER indicou como representativas nas diferenças encontradas pela ANOSIM.
0
5
10
15
20
25
Ind/10min
X. kroyeri
0
5
10
15
20
25
Ind/10min
N. schmitti
0
2
4
6
8
Ind/10min
C. danae
0
2
4
6
8
Ind/10min
C. sapidus
0
2
4
6
8
Ind/10min
C. ornatus

46
5.4.2. Enchente de 2011
Uma nova enchente ocorreu em setembro de 2011, novamente resultado de um
período de maior pluviosidade (Figura 1). Para analisar os efeitos desse evento foram
considerados os meses de agosto e setembro de 2011 como meses com enchente, isso
porque a partir de agosto a vazão do rio já estava bastante elevada e a salinidade era
equivalente à zero em boa parte do estuário. Deste modo, e ao contrário da análise
anterior, nesta foi avaliado o efeito da enchente durante a ocorrência do evento.
Examinando a variação temporal dos crustáceos verificou-se que em agosto e
setembro de 2011 a abundância e biomassa foram praticamente zero (Figura 15).
Da mesma forma que em 2008, para a análise com o MDS, ANOSIM e SIMPER
as amostras foram separadas em setores A e B, e comparadas entre os períodos com
enchente (2011, “CE”) e sem enchente (2008, “SE1”; 2010, “SE2”). Assim, as amostras
foram agrupadas em CEA, CEB, SE1A, SE1B, SE2A, SE2B para estas análises.
O MDS para a enchente de 2011 mostrou os pontos do primeiro período sem
enchente (2008) agrupados no mesmo lado do gráfico e praticamente todos os pontos do
período com enchente ficaram sobrepostos pois não foram capturados indivíduos na
maioria das amostras nesse período (Figura 22). A ANOSIM revelou diferenças
significativas do primeiro período sem enchente para os demais períodos. Os pontos
agrupados em CEA e CEB se diferenciaram dos pontos SE1A e SE1B, mas não foram
diferentes entre si. Essas diferenças foram principalmente explicadas, de acordo com o
SIMPER, pelas abundâncias de C. danae e X. kroyeri (Tabela 13). Ambas as espécies
estiveram praticamente ausentes no período com enchente, C. danae ocorreu em
abundância mais elevada em 2008 (SE1A e SE1B), e X. kroyeri ocorreu principalmente
nos pontos mais externos nos períodos sem enchente (SE1A e SE2A).

47
Figura 22. Análise MDS realizada com todas as espécies cuja abundância representou, em ao menos um setor e período, mais do que 3% do total. Os fatores utilizados foram: “CEA” e “CEB”, agosto a setembro de 2011 nos setores A e B respectivamente; “SE1A” e “SE1B”, agosto a setembro de 2008 nos setores A e B respectivamente; “SE2A” e “SE2B”, agosto a setembro de 2010 nos setores A e B respectivamente.
Tabela 13. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores do MDS representado na figura 22. No SIMPER foram incluídas todas as espécies até atingirem 90% de representatividade nos resultados. O SIMPER não foi considerado quando o resultado do ANOSIM teve p>0,05 (**).
Grupos ANOSIM
SIMPER (Dissimilaridade de Bray- Curtis) R p
SE1A x SE1B -0,021 0,439 **
SE1A x SE2A 0,611 0,002 C. danae (68,72%); X. kroyeri (17,14%); N. scmitti
(6,92%)
SE1A x SE2B 0,573 0,004 C. danae (75,70%); X. kroyeri (9,97%); C. sapidus
(7,44%) SE1A x CEA 0,606 0,004 C. danae (82,85%); X. kroyeri (8,12%)
SE1A x CEB 0,733 0,002 C. danae (83,32%); X. kroyeri (7,84%)
SE1B x SE2A 0,981 0,002 C. danae (79,89%); X. kroyeri (10,83%)
SE1B x SE2B 0,931 0,002 C. danae (90,57%)
SE1B x CEA 0,933 0,002 C. danae (94,75%)
SE1B x CEB 1 0,002 C. danae (94,89%)
SE2A x SE2B 0,198 0,061 **
SE2A x CEA 0,315 0,061 **
SE2A x CEB 0,344 0,061 **
SE2B x CEA -0,015 0,697 **
SE2B x CEB 0,117 0,061 **
CEA x CEB 0 1 **
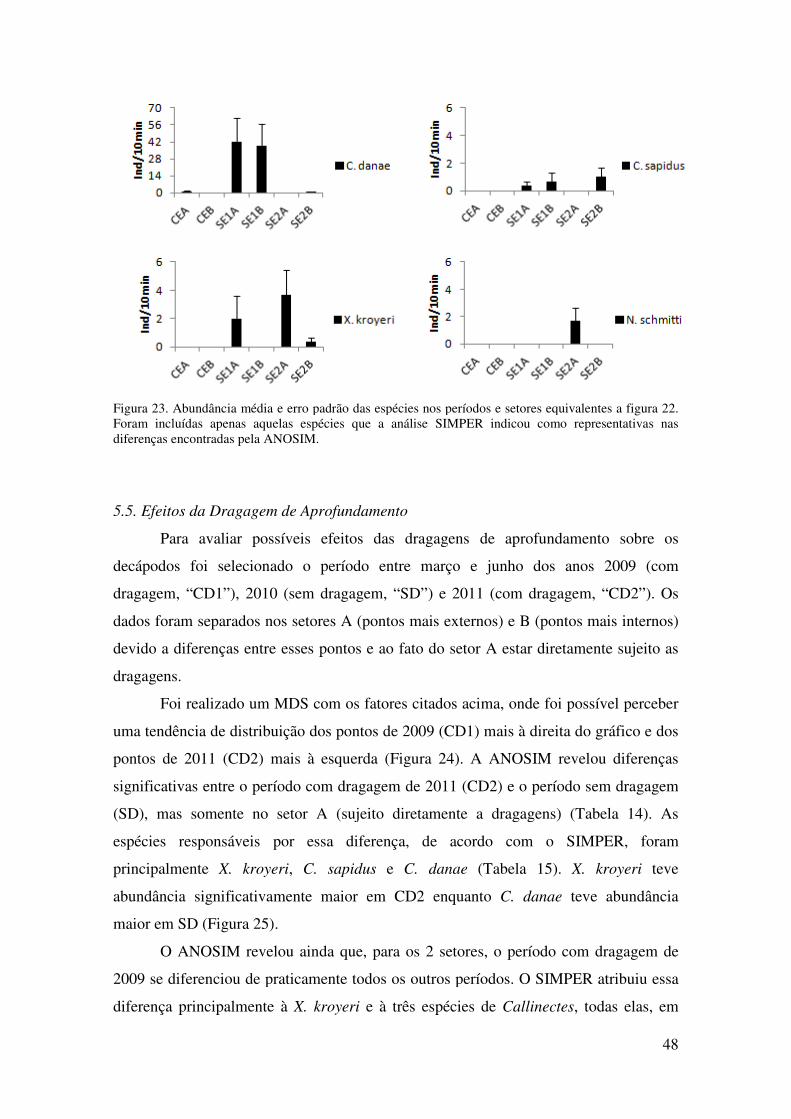
Figura 23. Abundância média e erro padrForam incluídas apenas aquelas espécies que a análise SIMPER indicou como representativas nas diferenças encontradas pela ANOSIM.
5.5. Efeitos da Dragagem de Aprofundamento
Para avaliar possíveis efeitos das dragagens de aprofund
decápodos foi selecionado o período entre março e
dragagem, “CD1”), 2010 (sem dragagem, “SD”) e 2011 (com dragagem, “CD2”). Os
dados foram separados nos setores A (pontos m
devido a diferenças entre esses pontos e ao fato do setor A estar diretamente sujeito as
dragagens.
Foi realizado um MDS com os fatores citados acima, onde foi possível perceber
uma tendência de distribuição dos
pontos de 2011 (CD2) mais à esquerda
significativas entre o período com dragagem de 2011
(SD), mas somente no setor A (sujeito direta
espécies responsáveis por essa diferença, de acordo com o SIMPER, foram
principalmente X. kroyeri
abundância significativamente maior em CD2
maior em SD (Figura 25).
O ANOSIM revelou ainda que, para os 2 setores, o período com dragagem de
2009 se diferenciou de praticamente todos os outros períodos.
diferença principalmente à
. Abundância média e erro padrão das espécies nos períodos e setores equivalentes a Foram incluídas apenas aquelas espécies que a análise SIMPER indicou como representativas nas diferenças encontradas pela ANOSIM.
de Aprofundamento
possíveis efeitos das dragagens de aprofundamento sobre os
ecionado o período entre março e junho dos anos 2009 (com
dragagem, “CD1”), 2010 (sem dragagem, “SD”) e 2011 (com dragagem, “CD2”). Os
dados foram separados nos setores A (pontos mais externos) e B (pontos mais internos)
devido a diferenças entre esses pontos e ao fato do setor A estar diretamente sujeito as
Foi realizado um MDS com os fatores citados acima, onde foi possível perceber
uma tendência de distribuição dos pontos de 2009 (CD1) mais à direita
mais à esquerda (Figura 24). A ANOSIM revelou
significativas entre o período com dragagem de 2011 (CD2) e o período sem dragagem
, mas somente no setor A (sujeito diretamente a dragagens) (Tabela
espécies responsáveis por essa diferença, de acordo com o SIMPER, foram
X. kroyeri, C. sapidus e C. danae (Tabela 15). X. kroyeri
significativamente maior em CD2 enquanto C. danae teve abund
O ANOSIM revelou ainda que, para os 2 setores, o período com dragagem de
2009 se diferenciou de praticamente todos os outros períodos. O SIMPER atribuiu essa
diferença principalmente à X. kroyeri e à três espécies de Callinectes
48
ão das espécies nos períodos e setores equivalentes a figura 22. Foram incluídas apenas aquelas espécies que a análise SIMPER indicou como representativas nas
amento sobre os
junho dos anos 2009 (com
dragagem, “CD1”), 2010 (sem dragagem, “SD”) e 2011 (com dragagem, “CD2”). Os
ais externos) e B (pontos mais internos)
devido a diferenças entre esses pontos e ao fato do setor A estar diretamente sujeito as
Foi realizado um MDS com os fatores citados acima, onde foi possível perceber
mais à direita do gráfico e dos
ANOSIM revelou diferenças
e o período sem dragagem
mente a dragagens) (Tabela 14). As
espécies responsáveis por essa diferença, de acordo com o SIMPER, foram
X. kroyeri teve
teve abundância
O ANOSIM revelou ainda que, para os 2 setores, o período com dragagem de
O SIMPER atribuiu essa
ctes, todas elas, em

49
geral, em maior abundância em 2009. Da mesma forma, os pontos do setor A em 2011
(CD2A) se diferenciaram de todos os demais pontos em outros anos (Tabela 14). O
SIMPER atribuiu essa variação principalmente a X. kroyeri e C. danae, porém outras 4
espécies contribuíram com pelo menos 7% das diferenças nos resultados (Tabela 14). C.
danae esteve ausente em CD2A, X. kroyeri teve abundância mediana, e N. schmitti e C.
ornatus tiveram abundância alta em comparação com os demais setores.
Figura 24. Análise MDS realizada com todas as espécies cuja abundância representou, em ao menos um setor e período, mais do que 3% do total. Os fatores utilizados foram: “CD1A” e “CD1B”, março a junho de 2009 nos setores A e B respectivamente; “SDA” e “SDB”, março a junho de 2010 nos setores A e B respectivamente; “CD2A” e “CD2B”, março a junho de 2011 nos setores A e B respectivamente.
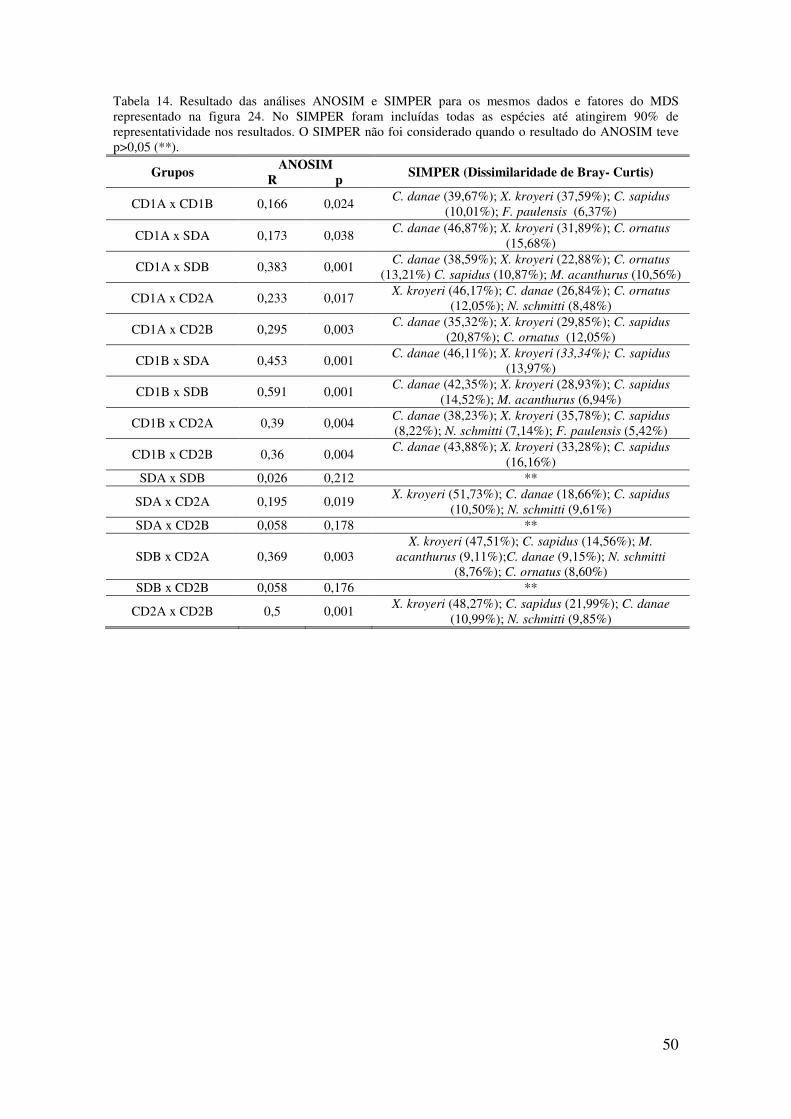
50
Tabela 14. Resultado das análises ANOSIM e SIMPER para os mesmos dados e fatores do MDS representado na figura 24. No SIMPER foram incluídas todas as espécies até atingirem 90% de representatividade nos resultados. O SIMPER não foi considerado quando o resultado do ANOSIM teve p>0,05 (**).
Grupos ANOSIM
SIMPER (Dissimilaridade de Bray- Curtis) R p
CD1A x CD1B 0,166 0,024 C. danae (39,67%); X. kroyeri (37,59%); C. sapidus
(10,01%); F. paulensis (6,37%)
CD1A x SDA 0,173 0,038 C. danae (46,87%); X. kroyeri (31,89%); C. ornatus
(15,68%)
CD1A x SDB 0,383 0,001 C. danae (38,59%); X. kroyeri (22,88%); C. ornatus
(13,21%) C. sapidus (10,87%); M. acanthurus (10,56%)
CD1A x CD2A 0,233 0,017 X. kroyeri (46,17%); C. danae (26,84%); C. ornatus
(12,05%); N. schmitti (8,48%)
CD1A x CD2B 0,295 0,003 C. danae (35,32%); X. kroyeri (29,85%); C. sapidus
(20,87%); C. ornatus (12,05%)
CD1B x SDA 0,453 0,001 C. danae (46,11%); X. kroyeri (33,34%); C. sapidus
(13,97%)
CD1B x SDB 0,591 0,001 C. danae (42,35%); X. kroyeri (28,93%); C. sapidus
(14,52%); M. acanthurus (6,94%)
CD1B x CD2A 0,39 0,004 C. danae (38,23%); X. kroyeri (35,78%); C. sapidus (8,22%); N. schmitti (7,14%); F. paulensis (5,42%)
CD1B x CD2B 0,36 0,004 C. danae (43,88%); X. kroyeri (33,28%); C. sapidus
(16,16%) SDA x SDB 0,026 0,212 **
SDA x CD2A 0,195 0,019 X. kroyeri (51,73%); C. danae (18,66%); C. sapidus
(10,50%); N. schmitti (9,61%) SDA x CD2B 0,058 0,178 **
SDB x CD2A 0,369 0,003 X. kroyeri (47,51%); C. sapidus (14,56%); M.
acanthurus (9,11%);C. danae (9,15%); N. schmitti (8,76%); C. ornatus (8,60%)
SDB x CD2B 0,058 0,176 **
CD2A x CD2B 0,5 0,001 X. kroyeri (48,27%); C. sapidus (21,99%); C. danae
(10,99%); N. schmitti (9,85%)

Figura 25. Abundância média e erro padrForam incluídas apenas aquelas espécies que a análise SIMPER indicou como representativas nas diferenças encontradas pela ANOSIM.
Abundância média e erro padrão das espécies nos períodos e setores equivalentes a
Foram incluídas apenas aquelas espécies que a análise SIMPER indicou como representativas nas diferenças encontradas pela ANOSIM.
51
ão das espécies nos períodos e setores equivalentes a figura 24. Foram incluídas apenas aquelas espécies que a análise SIMPER indicou como representativas nas

52
6. DISCUSSÃO
No estuário do rio Itajaí-Açu circulam grandes navios cargueiros que dependem
de uma profundidade adequada para poderem atracar nos portos de Itajaí e Navegantes.
Após a enchente que ocorreu no final de 2008 foi realizada uma dragagem emergencial
de aprofundamento para possibilitar o reinício das atividades dos portos. Essa dragagem
começou em dezembro de 2008 e foi concluída em julho de 2009, elevando a
profundidade na parte externa do estuário à aproximadamente 11 metros. A
profundidade nos pontos mais internos (4-6) se manteve em patamares inferiores, pois a
circulação dessas grandes embarcações está restrita aos pontos 1-3. Em março de 2011
teve início uma nova dragagem com o objetivo de aumentar a profundidade do canal de
acesso para 14 metros, explicando o aumento na profundidade observado a partir desse
mês. Isso se torna relevante uma vez que mudanças na profundidade do estuário podem
promover diversas alterações nos meios biótico e abiótico através da modificação dos
padrões de circulação (Torres, 2000).
O estuário do rio Itajaí-Açu é do tipo cunha salina, ou seja, em grande parte da
sua extensão é esperada a ocorrência de duas massas de água bem distintas: uma de
origem continental na superfície; e uma de origem marinha junto ao substrato (Schettini,
2002). Conforme a vazão do rio diminui, a cunha salina se estabelece cada vez mais a
montante e, em algumas ocasiões, a influência marinha pode ser percebida até 30 km rio
acima (Schettini, 2002). Além da vazão, que aumenta conforme a pluviosidade na bacia
hidrográfica, alterações na batimetria podem causar mudanças na penetração da cunha
salina (Torres, 2000). Assim, no baixo-estuário deverá ser observada uma massa de
água marinha próxima ao substrato sempre que a cunha salina estiver bem estabelecida.
Da mesma forma, conforme a vazão aumenta, a cunha salina é empurrada cada vez mais
a jusante, como observado por Schettini (2002). Valores próximos a zero foram
observados no setor mais interno nesse estudo sempre que a descarga foi superior a
500m³/s e, quando superior a 1000m³/s, salinidades inferiores a 20 foram observadas
inclusive nos pontos localizados próximos a foz.
A salinidade é um dos principais parâmetros que afetam a distribuição dos
organismos nos estuários (Day Jr. et al., 1989), e sua variabilidade é muito relevante na
distribuição espaço-temporal dos decápodos em estuários dominados por rios (Peterson
& Ross, 1991). No presente estudo foi observada salinidade média menor nos meses de
verão, período mais chuvoso e com maiores níveis de vazão. Também os pontos mais a
montante apresentaram maior variabilidade, justamente em função do padrão de

53
circulação que atua no estuário. Períodos prolongados de baixa vazão podem explicar os
valores elevados de salinidade média (>30) observados inclusive no ponto 6, entretanto
não se pode descartar a hipótese de um possível efeito da maré. Medeiros (2003) afirma
que, apesar da baixa amplitude, a maré pode afetar a intrusão da cunha salina no
estuário do rio Itajaí-Açu.
Os valores de pH e de salinidade são maiores no mar adjacente do que no rio
Itajaí-Açu e, portanto, esses parâmetros devem variar de forma semelhante conforme as
massas de água marinha e continental se misturam no estuário. A água do mar é
naturalmente tamponada e é difícil perceber grandes variações no seu pH, entretanto,
mesmo considerando apenas os pontos mais externos da área de estudo, foi observada
uma tendência de redução no pH ao longo do tempo. As dragagens podem causar
reduções pontuais no pH, porém esse efeito é percebido apenas em curto espaço de
tempo e não na escala temporal de meses (Ohimain et al., 2008). Apesar de não ter sido
observado o mesmo para a salinidade, a redução do pH ao longo do estudo pode ser
resultado de uma maior mistura de águas continentais ocasionada pelo aumento da
vazão, principalmente em 2011.
Um fator importante regulando a distribuição de crustáceos decápodos e peixes
bentônicos é a temperatura da água (Kodama et al., 2010). No presente estudo esse
parâmetro apresentou o comportamento sazonal esperado, com maiores valores no
verão e menores no inverno. Em 2010 a temperatura da água no inverno foi inferior aos
demais anos, provavelmente em função de fatores meteorológicos ou oceanográficos de
grande escala. Embora não significativas, foram observadas pequenas diferenças na
temperatura entre os pontos mais externos e mais internos, que podem ser explicadas
pela maior flutuação da temperatura em águas continentais do que em águas marinhas:
nos oceanos a temperatura tende a ser mais constante devido à sua menor relação
superfície/volume.
O oxigênio dissolvido (OD) é também um importante fator que atua sobre o
megabentos (Kodama et al., 2010). O padrão de variação desse parâmetro foi marcado
por ocorrências pontuais de elevada ou de baixa concentração, observadas
principalmente nos pontos internos e externos, respectivamente. Essas variações no OD
no Itajaí-Açu podem ser explicadas pelos processos de decomposição da matéria
orgânica e nitrificação (Kuroshima & Bellotto, 2009). Após eventos de elevada vazão a
quantidade de sedimento em suspensão é extremamente elevada, geralmente
acompanhada de matéria orgânica proveniente do continente (Schettini, 2001). Quando

54
esse material sedimenta é esperada uma redução no oxigênio dissolvido devido à
decomposição da matéria orgânica, o que de fato pode ter ocorrido em algumas
ocasiões, como outubro e novembro de 2009, após um pico de vazão verificado no mês
de setembro. Acompanhando eventos de descarga fluvial muito acima da média foram
observados picos de oxigênio dissolvido provavelmente devido à redução da
importância relativa do aporte antrópico no estuário: nessas condições ocorre a
diminuição do tempo de residência da água e o aumento da capacidade de diluição do
sistema. Em fevereiro de 2009 houve a redução pontual do oxigênio dissolvido no ponto
3, bem em frente ao porto de Itajaí, provavelmente devido às obras no berço de
atracação que foi danificado durante a enchente de novembro de 2008. Além disso, por
diversas vezes, os menores valores de oxigênio dissolvido ocorreram em períodos onde
é esperada uma menor solubilidade dos gases na água, ou seja, quando a temperatura
esteve mais elevada. Por fim, devido à suspensão de sedimentos finos (geralmente ricos
em matéria orgânica), as dragagens também podem provocar a redução do oxigênio
dissolvido (Torres, 2000; Erftemeijer & Lewis, 2006; Ohimain et al., 2008), o que
explicaria, em parte, a maior frequência de situações de baixas concentrações
verificadas nos pontos 1-3.
Estuários são importantes ecossistemas com elevada dinâmica e produtividade
(Day Jr. et al., 1989). No entanto, apesar de serem produtivos, a ampla flutuação das
suas variáveis ambientais e as condições de estresse a que os organismos estão
submetidos tornam esses ecossistemas, em geral, locais com baixa riqueza e elevada
dominância de espécies (Day Jr. et al., 1989; Angonesi, 2005; Costa-Dias et al., 2010).
Foram encontradas 20 espécies/morfotipos de crustáceos decápodos durante os três anos
e meio de estudo. Branco e Freitas Jr. (2009) fizeram uma análise quali-quantitativa de
crustáceos decápodos no Saco da Fazenda, ambiente adjacente ao baixo-estuário do
Itajaí-Açu, entre 2000 e 2005. Em seu trabalho os autores encontraram 10 espécies, sete
iguais às do presente estudo. Portanto, comparando os dois trabalhos, ocorreram 13
espécies exclusivamente no canal do estuário e três no Saco da Fazenda. Das vinte
espécies amostradas, 14 foram encontradas na primeira fase de amostragem, executada
por Leite (2010) entre maio de 2008 e julho de 2009. Existe, portanto, uma dinâmica na
ocorrência de espécies no canal do estuário do Itajaí-Açu, que difere do que é observado
no Saco da Fazenda, mesmo sendo ecossistemas muito próximos e intimamente
relacionados.

55
No presente estudo foi observada a ocorrência ocasional de algumas espécies em
abundâncias bastante reduzidas, entre elas, o siri Charybdis helleri, invasor e original do
Indo-Pacífico (Mantellato & Garcia, 2001), que totalizou apenas 4 indivíduos. Existe
risco desse siri competir com espécies nativas de Callinectes, além de ser hospedeiro
potencial do vírus WSSV (Síndrome da Mancha Branca) (Tavares & Mendonça Jr.,
2004). Isso mostra que o estuário do rio Itajaí-Açu está sujeito a bioinvasão,
principalmente se for considerada a grande quantidade de navios que atracam nos portos
existentes em suas margens.
Neste estudo, apenas 6 espécies foram responsáveis por mais de 95% da
abundância, sendo que Callinectes danae contribuiu com aproximadamente 48%,
Xiphopenaeus kroyeri com 29%, e Callinectes sapidus com 7%. Em termos de
biomassa a dominância foi ainda mais marcante devido ao maior peso individual de C.
danae e C. sapidus que, juntas, contribuíram com mais de 90% da biomassa total
amostrada. Essa dominância de poucas espécies de decápodos foi também observada
por Leite (2010) no baixo estuário do rio Itajaí-Açu e por Branco e Freitas Jr. (2009) no
Saco da Fazenda. Esse padrão de dominância também foi observado na região para
peixes por Hostim-Silva et al. (2002) e Antunes (2010), em cujo trabalho apenas 6
espécies contribuíram com 99% da captura entre maio de 2008 e julho de 2009 nas
mesmas estações de amostragem do presente estudo. Segundo a última autora, isso pode
estar associado ao acúmulo de perturbações antrópicas no sistema, como as dragagens e
alterações nas margens do estuário, ou ainda a perturbação natural, incluindo as
enchentes, que podem gerar alterações imediatas sobre a abundância das principais
espécies.
Estuários podem ser ainda locais importantes de crescimento e desenvolvimento
de crustáceos decápodos de interesse comercial (Willians, 1984). Dentre os camarões
capturados neste trabalho, F. paulensis e X. kroyeri são duas das espécies de maior
importância econômica no Sudeste e Sul do Brasil (D’Incao et al., 2002). Além dessas,
A. longinaris e L. schmitti também são capturadas pela pesca artesanal e/ou industrial
(D’Incao et al., 2002), apesar de terem sido coletadas de forma ocasional neste estudo.
A pesca de camarões apresenta importante papel econômico e social para a região onde
o estuário do rio Itajaí-Açu está localizado (Branco & Verani, 2006). Também presente
nas capturas obtidas durante as operações de pesca comercial de X. kroyeri na região
(Branco & Verani, 2006; Bail et al., 2009; Roedel, 2009), os siris do gênero Callinectes,
principalmente C. sapidus, podem ser utilizados na alimentação humana e apresentam

56
grande relevância econômica (Willians, 1974; Severino-Rodrigues et al., 2001;
Mendonça et al., 2010). Deste modo, verifica-se que existem várias espécies com
importância econômica frequentando o baixo-estuário do rio Itajaí-Açu.
Ao analisar a contribuição das principais famílias nos diferentes anos foi
possível obervar que, independente do ano, Penaeidae e Portunidae responderam por
mais de 70% da abundância total, indicando que essas são as famílias de maior
relevância para o estudo. Entretanto, nota-se que houve mudanças na contribuição das
principais famílias e espécies nos três anos e meio de estudo, principalmente com o
aumento da participação de Palaemonidae em 2010 e 2011. Branco e Freitas Jr. (2009)
também observaram alterações na composição de espécies ao longo de cinco anos no
Saco da Fazenda. Esses autores verificaram que, embora em todos os anos a espécie
dominante tenha sido C. danae, as espécies que ocuparam a segunda e a terceira posição
em abundância variaram ao longo dos anos. A dinâmica interanual da composição e
abundância das principais espécies não ocorre apenas no estuário do Itajaí e no Saco da
Fazenda. Na Baía Sul em Florianópolis, uma área costeira com características
estuarinas, Freitas Jr. et al. (2010) verificaram, por exemplo, que Litopenaeus schmitti
foi a espécie mais abundante em 1997 (48%), porém ocupou apenas a quarta posição em
2001 (8%). Outro exemplo é o estudo de Kodama et al. (2010) que analisaram, por 26
anos, o megabentos na Baía de Toquio, no Japão. Nesse trabalho os autores notaram que
as mudanças interanuais na comunidade foram sempre decorrentes de alguma alteração
nas condições ambientais. Assim, a substituição de espécies ao longo dos anos é algo
que ocorre normalmente em vários ecossistemas, e não apenas no estuário do Itajaí-Açu.
Além disso, em geral, essas variações interanuais são reflexos de alguma mudança ou
impacto sofrido pelo ecossistema, como possíveis efeitos da enchente, dragagem, ou
ainda mudanças nos padrões de circulação. No caso do Itajaí-Açu, nos dois últimos anos
a vazão durante as coletas apresentou valores bem mais elevados do que nos dois
primeiros, o que justificaria a alteração na composição e abundância das espécies, com a
maior participação de espécies de água doce (basicamente Palaemonidae) e a redução de
espécies estuarinas e principalmente marinhas.
Verificando a distribuição espacial dos crustáceos ao longo do estuário, nota-se
que a maior abundância média foi observada próximo a foz (ponto 1), reflexo
principalmente da contribuição dos peneídeos, sobretudo de X. kroyeri. Essa espécie é
marinha e independe do estuário para o seu desenvolvimento (Branco et al., 1999;
Castro et al., 2005) e, portanto, a maior abundância junto a foz deve ser reflexo das

57
maiores abundâncias de X. kroyeri no mar adjacente. Nos pontos mais internos (3 a 6)
houve a predominância da família Portunidae, com abundância elevada de Callinectes
danae e C. sapidus. C. ornatus foi capturado principalmente próximo a foz, o que pode
ser explicado pelo fato da espécie preferir maiores salinidades (Willians, 1974).
A variação espacial da biomassa diferiu em relação à distribuição da abundância.
Isso se deve ao fato dos siris apresentarem massa individual muito maior que os
camarões. Assim, ao contrário da abundância, a biomassa média foi maior em uma
região mais interna do estuário, no ponto 5, reflexo da elevada presença de Portunidae e
principalmente de C. sapidus. De fato, foram capturados indivíduos de elevado porte
dessa espécie, tanto que sua biomassa representou 32,2% do total enquanto a sua
abundância não superou 6,7%. Os indivíduos de C. danae apresentaram tamanho
semelhante aos capturados por Branco e Freitas Jr. (2009) no Saco da Fazenda.
Entretanto, tamanhos maiores de C. sapidus foram encontrados no presente estudo na
zona de canal, sugerindo que exemplares menores da espécie devam preferir as áreas
mais rasas, como as do Saco da Fazenda, como habitat.
A variação espacial na distribuição das espécies foi confirmada pelo MDS e
ANOSIM, que indicaram diferenças significativas entre os pontos mais externos (1-3,
setor A) e mais internos (4-6, setor B). O SIMPER indicou que C. danae, C. sapidus, X.
kroyeri foram as principais responsáveis pelas diferenças observadas. As espécies que
frequentam normalmente o estuário, como C. danae e C. sapidus (Willians, 1974),
ocorreram em maiores abundâncias no setor B, enquanto que o setor A foi caracterizado
pela presença de espécies predominantemente marinhas como X. kroyeri (Branco et al.,
1999). Além disso, M. acanthurus ocorreu unicamente no setor B, por ser uma espécie
típica de salinidades baixas ou ainda de águas continentais (Willians, 1984).
A separação em diferentes setores é constantemente utilizada em estudos em
estuários por facilitar a análise e a observação de variações sazonais e/ou entre distintas
áreas (Jones et al., 1990; Peterson & Ross, 1991). Nesse estudo ficaram evidentes as
diferenças entre os pontos mais internos e externos do estuário, com o setor B
apresentando menores valores de salinidade, pH e profundidade, maior variação na
salinidade, maiores abundâncias de C. danae e C. sapidus, menor abundância de X.
kroyeri e a ocorrência de M. acanthurus e outros Palemonidae. Além disso, um fator
determinante na separação dos setores, além das diferenças supracitadas, foi o fato dos
pontos localizados no setor A estarem situados na área de circulação dos navios e,

58
portanto, estarem diretamente sujeitos aos processos de dragagem e outros possíveis
impactos promovidos pela atividade portuária.
Em relação à variação temporal da abundância, foram observados baixos valores
nos meses de dezembro e janeiro, com grande aumento e eventuais picos nos meses de
fevereiro, março e abril. Resultado semelhante já foi verificado no Saco da Fazenda,
com abundância mínima em dezembro e valores máximos entre janeiro e abril (Branco
& Freitas Jr., 2009). Outro exemplo onde esse padrão foi observado é o estudo realizado
apenas com siris ao longo de um ano no estuário da Baía da Babitonga, São Francisco
do Sul, SC, onde foi observada uma redução muito grande na captura dos organismos
nos meses de verão de 2003-2004 (Pereira, 2006). Entretanto, isso não se repete no
trabalho realizado na Baía Sul em Florianópolis, onde foram observados picos de
abundância da carcinofauna (siris e camarões) sempre no verão, com abundância
mínima no inverno (Freitas Jr. et al., 2010). Considerando que a Baía Sul, apesar de
apresentar características estuarinas, é um ecossistema marinho-costeiro, pode-se
presumir que o padrão distinto de abundância sazonal observado por Freitas Jr. et al.
(2010) decorra da sua área de estudo não estar situada no interior de um estuário
propriamente dito. De fato, a variação na abundância de decápodos no interior de
estuários e na região costeira adjacente, principalmente dos siris do gênero Callinectes,
deve estar relacionada com a migração sazonal desses organismos motivada pela
reprodução (Willians, 1974). Em seu livro esse autor propõe que em C. sapidus, e
possivelmente em todas as demais espécies do gênero, o desenvolvimento das larvas
ocorre na região costeira, com o posterior deslocamento dos juvenis para se
desenvolverem em indivíduos adultos no interior do estuário. Isso foi confirmado mais
recentemente no estudo de Posey et al. (2005), que estudaram a distribuição de juvenis
de C. sapidus em estuários dominados por rios na Carolina do Norte, EUA. Os autores
constataram que juvenis de C. sapidus dependem de áreas com baixas salinidades para
terem uma maior sobrevivência nesse tipo de estuário. Já no Brasil, Branco & Masunari
(2000) estudaram a ecologia reprodutiva de C. danae na Lagoa da Conceição, em
Florianópolis. Segundo estes autores, o estuário funciona como uma área de
crescimento, reprodução e desova para a espécie. Assim, os estuários representam
importantes áreas para a reprodução de crustáceos decápodos e, por esse motivo, as
oscilações na abundância da carcinofauna ao longo do ano no estuário do Rio Itajaí-Açu
devem estar relacionadas com os padrões reprodutivos das diferentes espécies.

59
Quanto à variação da família Penaeidae, representada principalmente por X.
kroyeri, foi verificada abundância mais elevada entre fevereiro e abril. Provavelmente
esse é o período em que a espécie ocorre em maior abundância no mar adjacente, pois,
como previamente discutido, esta é uma espécie marinha. De fato, X. kroyeri foi mais
abundante nos meses de outono na Baía de Tijucas (Pezzuto et al., 2008), na região
costeira junto a desembocadura do Itajaí-Açu (Branco et al., 1999), e na Armação do
Itapocoroy, em Penha (Branco & Verani, 2006), todas situadas no litoral catarinense.
Ainda, foi observada a variação temporal da família Palaeomonidae,
representada principalmente por M. acanthurus, uma espécie de águas continentais,
geralmente encontrada próximo ou no interior de estuários, e N. schmitti, espécie
marinha de regiões costeiras rasas (Ferreira, 2009). M. acanthurus, assim como nesse
trabalho, foi capturado em baixa abundância e de forma ocasional e descontínua por
Branco & Freitas Jr. (2009) no Saco da Fazenda. N. schmitti não foi identificada nos
estudos realizados na região costeira junto a desembocadura do Itajaí-Açu (Branco et
al., 1999) e no Saco da Fazenda (Branco & Freitas Jr., 2009). Na Armação do
Itapocoroy a espécie foi capturada apenas entre julho de 2006 e junho de 2007, último
ano de um total de 10 anos de monitoramento da carcinofauna local (Roedel, 2009). As
baixas abundâncias de N. schmitti registradas neste estudo, portanto, devem ser reflexo
da sua baixa ocorrência na região costeira adjacente ao estuário.
O MDS e a ANOSIM também indicaram um agrupamento das amostras em dois
períodos: 2008 e 2009 e em 2010 e 2011. Entre as espécies de siris responsáveis por
essas diferenças, C. danae e C. ornatus foram mais abundantes em 2008 e 2009,
enquanto que C. sapidus foi mais abundante em 2010 e 2011. Nos dois últimos anos foi
observada uma maior oscilação da salinidade e, conforme constatado na literatura, C.
sapidus é bastante tolerante a esse tipo de estresse (Ogburn, et al., 2007). Assim, a
grande flutuação da salinidade pode ter limitado a ocorrência de C. danae permitindo
que uma espécie mais tolerante dominasse o ambiente já que cada espécie deve ocupar
um nicho diferente. X. kroyeri também foi responsável pelas diferenças entre os dois
períodos. Essa espécie ocorre naturalmente em maiores abundâncias nos meses de
outono e, particularmente em 2009 pode ter ocorrido um maior recrutamento desse
camarão na região.
A relação entre as variáveis abióticas e as principais espécies capturadas foi
verificada através de uma análise de correspondência canônica (ACC) e gráficos Box-
plot. Sabe-se que todas as espécies de Callinectes encontradas no presente estudo

60
podem tolerar um amplo espectro de salinidade e temperatura (Willians, 1974). Isso
explicaria o fato de não terem sido observadas espécies do gênero longe do centro na
ACC. Entretanto, C. sapidus se correlacionou de forma distinta com as variáveis
ambientais, sendo a única do gênero a ocorrer em condições de menor salinidade (em
valores próximo a 5). Isso pode ser explicado pelo fato dessa espécie ser a mais
tolerante a valores extremos dos parâmetros ambientais, como já verificado na
bibliografia (Willians, 1974; Ogburn, et al., 2007).
Norse (1978) analisou a tolerância de diversas espécies do gênero Callinectes a
valores baixos de salinidade. Com dados de campo o autor verificou que existe a
dominância de diferentes espécies do gênero conforme o gradiente de salinidade: C.
bocurti e C. sapidus são as espécie mais tolerantes a baixas salinidades e tendem a ser
relativamente mais abundantes quando os valores médios de salinidade são inferiores a
10; C. danae ocorre em maior abundância relativa em salinidades entre 10 e 33; e C.
ornatus é a espécie que tende a predominar em salinidades superiores a 33. O autor
ainda realizou experimentos de tolerância a baixas salinidades e observou que C.
sapidus e C. bocurti apresentaram 100% de taxa de sobrevivência a água do mar diluída
a 10%, enquanto que C. danae e C. ornatus apresentaram taxas de sobrevivência de
50% e 3% respectivamente, indo ao encontro dos resultados obtidos em campo. No
presente estudo C. sapidus, C. danae e C. ornatus ocorreram predominantemente em
valores crescentes de salinidade (medianas de 25, 28 e 33 e mínimas de 0, 13 e 25,
respectivamente), conforme sugerido por Norse (1978). Em contrapartida, foram
coletados apenas sete indivíduos de C. bocurti durante o presente estudo, sempre em
períodos de salinidade elevada e, assim, a diferença para os resultados apresentados por
Norse (1978) pode ser apenas um efeito do acaso ou de uma menor abundância geral da
espécie na região.
Camarões da família Penaeidae têm sua distribuição influenciada por diversos
fatores ambientais como a temperatura, o tipo de sedimento, a disponibilidade de
alimento e a salinidade (Branco & Verani, 1998; Costa et al., 2004; Lopes, 2008). No
presente estudo F. paulensis ocorreu preferencialmente em valores de temperatura entre
19 e 23ºC e salinidades entre 26 e 31, porém sem mostrar relação óbvia com essas
variáveis, indicando que outros fatores podem estar afetando a sua abundância no Itajaí-
Açu. Para D'Incao & Dumont (2010), juvenis e pré-adultos de F. paulensis habitam
preferencialmente enseadas rasas e protegidas no interior do estuário da Lagoa dos
Patos, RS. Ainda segundo os autores, exemplares adultos dessa espécie são encontrados

61
na plataforma continental, em profundidades entre 40 e 80 metros. Garcia et al. (1996),
concluíram que áreas vegetadas são os habitats preferenciais de F. paulensis dentro da
Lagoa do Patos. O baixo estuário do Itajaí-Açu não apresenta mais praticamente
nenhuma área vegetada. As margens do estuário são quase totalmente tomadas por
construções e apresenta uma larga zona de canal, o que não oferece proteção como as
várias enseadas rasas (sacos) onde F. paulensis é encontrado na Lagoa dos Patos. Isso
explicaria a baixa captura dessa espécie ao longo de todo o presente estudo.
Confirmando tal fato, Branco e Freitas Jr. (2009) verificaram que o camarão-rosa foi o
terceiro decápodo em termos de abundância no Saco da Fazenda em dois dos cinco anos
do estudo, possivelmente pelo fato dessa enseada ser mais abrigada, rasa e apresentar
algumas áreas vegetadas servindo, portanto, de abrigo para F. paulensis.
X. kroyeri ficou deslocada dos demais decápodos na ACC, tendo sido encontrada
primariamente em condições de maiores temperatura, pH e salinidade. Assim como
observado para F. paulensis, o camarão sete-barbas também apresenta distribuição e
abundância significativamente relacionadas com as oscilações da temperatura e
salinidade da água de fundo (Branco et al., 1999). Esses autores verificaram, na foz do
Rio Itajaí-Açu, que X. kroyeri apresenta flutuações sazonais, sendo os maiores índices
de captura obtidos durante o outono e os menores na primavera. Como previamente
discutido, essa espécie entra no estuário apenas acompanhando a cunha salina, e as
maiores abundâncias serão reflexo da quantidade de indivíduos na região marinha
adjacente. Assim, os fatores que estiverem afetando a região costeira em determinado
período podem ser muito mais importantes para a abundância de X. kroyeri no estuário
do que outras variáveis que atuem diretamente nesse ecossistema.
A distribuição do camarão N. schmitti depende de alguns fatores ambientais,
entre eles salinidade, temperatura e oxigênio dissolvido (Almeida et al., 2007). Essa
espécie ocorre ao longo da costa do Brasil e da Guiana e é considerada marinha (Costa
et al., 2000). Por esse motivo, a sua ocorrência dentro de um estuário deve estar
relacionada com a penetração da cunha salina, o que de fato ocorreu no presente estudo,
onde N. schmitti foi observado restritamente em condições de salinidade superior a 25.
As espécies do gênero Macrobrachium são exclusivamente de águas
continentais ou estuarinas (Tamburus et al., 2012). M. acanthurus é conhecido
popularmente como pitu, camarão canela ou camarão de água doce. Habita corpos de
água doce e salobra que se comunicam com o mar e é uma espécie com grande
potencial para cultivo (Valenti et al., 1989). M. acanthurus foi encontrado por Branco e

62
Freitas Jr. (2009) no Saco da Fazenda, sendo considerada uma espécie de ocorrência
ocasional pelos autores. No presente estudo, a captura de três espécies distintas do
gênero foi fortemente relacionada a condições de salinidade extremamente baixas.
Camarões desse gênero provavelmente não ocorrem normalmente no baixo-estuário do
Itajaí-Açu que, na maior parte do tempo, apresenta valores médios ou altos de
salinidade na água de fundo.
O caranguejo Libinia ferreirae (popularmente conhecido como caranguejo
aranha) pertence à família Majidae e é comum na região costeira de Santa Catarina,
Paraná, São Paulo e Rio de Janeiro, dentre outros (Braga et al., 2005; Winter &
Masunari, 2006; Costa & Beneditto, 2009). No presente estudo, a espécie foi capturada
em apenas uma ocasião (julho de 2011), principalmente nos pontos próximos a foz.
Nessa coleta, a salinidade estava alta (>32) em todos os seis pontos amostrados, o que
pode ter favorecido a entrada de alguns indivíduos no estuário acompanhando a cunha
salina. Essa espécie está presente na região costeira adjacente (Armação do Itapocoroy),
mas provavelmente em baixas abundâncias já que foi capturado apenas em dois dos
cinco anos do estudo de Roedel (2009), e sempre em abundâncias inferiores a 0,5% do
total.
Vários tipos de eventos extremos, incluindo as enchentes, podem causar grandes
alterações nas comunidades que habitam tanto o estuário como regiões costeiras
adjacentes (The Mermex Group, 2011). Nesse trabalho os autores fizeram uma grande
revisão bibliográfica para a região costeira do Mar Mediterrâneo, mostrando que os
impactos causados por eventos extremos como enchentes ou tempestades podem ser de
grande relevância, tanto no meio biótico como abiótico. As enchentes podem promover
a ocorrência de eventos de elevada turbidez, que foram estudados por González-Ortegón
et al. (2010). Ao estudar peixes, decápodos e suas presas em um estuário no sudoeste da
Espanha, esses autores constataram que esse tipo de evento pode prejudicar a função
berçário do ecossistema estuarino. Em um estudo na baía de Chesapeake, na costa leste
dos EUA, foram observadas alterações na macrofauna bentônica após a passagem do
furacão Isabel (Hughes et al., 2009). Outro furacão que ocorreu nos EUA, o Katrina, fez
com que águas de salinidade alta prevalecessem por meses em uma região entre-marés
no estado de Louisiana onde, normalmente, prevalecem águas continentais (Piazza &
Peyre, 2009). Esse impacto, segundo os autores, proporcionou grandes alterações no
nécton até que a situação do estuário voltasse às condições pré-furacão. Esses exemplos
ilustram as grandes modificações que podem ocorrer por conta de tempestades ou

63
enchentes. Entretanto, devido à imprevisibilidade de tais eventos, são poucos os estudos
que realmente conseguem atestar os impactos deles decorrentes (Hughes et al., 2009).
Sabe-se, entretanto, que a entrada de água doce é um dos principais fatores que afetam o
epibentos nos estuários (Jones et al., 1990) e que, espécies que não toleram baixas
salinidades podem não ter tempo de reagir a grandes aportes de água doce sendo,
portanto, excluídas (Paperno & Brodie, 2004).
Durante o presente estudo foram registrados duas grandes enchentes, uma em
novembro de 2008 e outra em setembro de 2011. Essas duas ocorrências, em períodos
semelhantes do ano, ofereceram uma oportunidade única de verificar os efeitos de um
evento natural extremo sobre os decápodos. Durante e/ou logo após as enchentes foram
capturadas quantidades mínimas de crustáceos. Isso pode ser um efeito agudo desses
eventos extremos que poderia ser explicado por pelo menos três hipóteses: (a) a fauna
pode ter sido deslocada para o mar devido a forte vazão ou ainda se movido
voluntariamente; (b) os organismos podem ter se espalhado para as áreas alagadas pelo
rio ou (c) devido à rápida mudança nas condições do estuário, pode ter havido
mortalidade da fauna. Apesar da aparente redução provocada pela enchente de 2008,
pode ser que a baixa abundância verificada nos meses de dezembro de 2008 e janeiro de
2009 seja, na verdade, um padrão que ocorra normalmente no estuário do Itajaí-Açu,
embora possa ter sido potencializado pelas enchentes. De fato, o MDS e a ANOSIM
não mostraram diferenças significativas nas assembleias quando considerados os meses
de dezembro e janeiro de 2008-2009, 2009-2010 e 2010-2011, provavelmente, em
função dos baixos valores observados em todos os anos analisados, incluindo aqueles
onde não houve enchente. Abundâncias mínimas de crustáceos decápodos também
foram observadas em dezembro e janeiro em estudos realizados no Saco da Fazenda
(Branco & Freitas Jr., 2009) e em áreas próximas a desembocadura do estuário (Bail et
al., 2009), o que reforçaria a hipótese de uma ocorrência naturalmente reduzida desses
organismos na região, durante tais períodos. Apenas C. danae e C. sapidus foram
coletados nos dois meses seguintes a enchente dentro do estuário. Mesmo em condições
adversas, a probabilidade de capturar C. danae deverá ser sempre alta no estuário do
Itajaí-Açu. Isso porque a espécie, além de ser dominante no local, também apresenta
abundância relativa elevada nos ecossistemas adjacentes (Bail et al., 2009; Branco &
Freitas Jr., 2009). Já C. sapidus, como previamente discutido, é uma espécie bastante
tolerante a flutuações nas variáveis ambientais e adaptada a condições adversas,
incluindo baixas salinidades (Willians, 1974).

64
Em relação à redução na abundância provocada pela enchente de 2011, as duas
principais espécies afetadas foram X. kroyeri e C. danae, ambas praticamente ausentes
durante o evento. X. kroyeri não deve ter migrado para outras áreas ou ter sofrido
mortalidade, mas sim evitado o estuário por não frequentar, naturalmente, locais com
baixos valores de salinidade (Castro et al., 2005). Já a ausência de C. danae, poderia ser
reflexo da mortalidade elevada dessa espécie quando sujeita a salinidades baixas por
longos períodos de tempo (Pereira-Barros, 1987 apud Branco & Masunari, 2000).
Entretanto, é provável que a espécie tenha migrado para a região costeira, pois os siris
apresentam grande capacidade de locomoção. De fato, nos meses seguintes a enchente
de 2008 foram observadas grandes quantidades de C. danae com tamanhos médios
equivalentes aos do período anterior ao evento. Tal fato sugere que não deva ter havido
mortalidade de C. danae durante as enchentes de 2008, mas sim um deslocamento para
outras áreas com um posterior retorno dos mesmos indivíduos ao estuário.
Nas duas primeiras coletas após a enchente de novembro de 2008 foram
capturadas quantidades mínimas de decápodos. Entretanto, três a quatro meses após o
evento, sua abundância aumentou substancialmente, atingindo os maiores valores de
todo o estudo ainda em 2009. Essa capacidade de se recuperar após uma perturbação
denomina-se resiliência (Begon et al., 1996). Para Odum (1983), o ecossistema pode
apresentar dois tipos de estabilidade: (a) Estabilidade de resistência, que consiste na
capacidade de resistir a perturbações, mantendo intactos sua estrutura e funcionamento;
(b) Estabilidade da elasticidade, que consiste na capacidade de se recuperar quando o
sistema é desequilibrado por uma perturbação. Segundo esse autor, dificilmente esses
dois tipos de estabilidade se desenvolvem ao mesmo tempo. Dungan & Livingston
(1982) concluíram que estuários poluídos, devido ao grande número de perturbações,
sofrem frequentes alterações na estrutura da assembleia de decápodos. Segundo esses
autores, estuários menos impactados deverão apresentar maior resistência a grandes
perturbações que possam ocorrer. Dessa forma, apesar dos crustáceos terem se
recuperado em 2008/2009, pode ser que em um evento posterior isso não ocorra, uma
vez que o estuário do rio Itajaí-Açu está sujeito a um grande número de outros impactos
que podem ir além da capacidade de recuperação da comunidade. Vale ressaltar que o
pico de abundância nos meses seguintes a enchente pode ser um efeito secundário do
evento. Rozas et al. (2005) estudou o efeito de entradas de água doce, provocadas
artificialmente por estruturas como barragens, em um sistema estuarino em Louisiana,
EUA. Dentre os resultados observados, os autores concluíram que a incorporação dos

65
nutrientes de origem continental pode ter aumentado a produtividade primária e
secundária do sistema estuarino como um todo, além de fornecer material detrítico
abundante, que poderia ser utilizado como alimento e/ou refúgio pelas espécies
amostradas no presente estudo (Leite, 2010).
Como as amostragens se encerraram dois meses após as enchentes de 2011, não
é possível, para esse evento, afirmar se os crustáceos migraram para outras áreas como
parece ter ocorrido em 2008 ou se sofreram mortalidade, como evidenciado em algumas
situações reportadas na literatura. Paperno & Brodie (2004) estudaram o efeito da
entrada de água doce sobre a comunidade de peixes e crustáceos decápodos em um
estuário na Flórida, EUA. Esses autores concluíram que, quando grandes quantidades de
água doce chegam ao estuário em um curto espaço de tempo, pode ser observada a
mortalidade de espécies estuarinas, mesmo aquelas vágeis, por não terem tempo de se
ajustar a rápida mudança na salinidade. Sendo assim, não se pode descartar a hipótese
de, apesar da resiliência, ocorrerem alterações permanentes na fauna estuarina em
função dos impactos não naturais que se acumulam no estuário ao longo do tempo
(Antunes, 2010).
Ressalta-se que, como consequência das mudanças climáticas globais, enchentes
extremas devem se tornar mais frequentes (IPCC, 2007) e, dessa forma, podem ocorrer
mudanças em longo prazo na comunidade do estuário caso não haja tempo suficiente
para a fauna se recuperar entre dois eventos distintos. A proximidade temporal entre as
duas enchentes abordadas nesse estudo, intercaladas ainda com dragagens de
manutenção e de aprofundamento, além de outros impactos crônicos vividos pelo
estuário do Itajaí-Açu podem indicar uma situação de vulnerabilidade da biota a
alterações mais drásticas no futuro. Isso porque o relatório do IPCC (2007) sugere que
diversos eventos extremos, como precipitação acima da média e secas prolongadas,
provavelmente ocorrerão com maior frequência nos próximos anos. Assim, as
observações feitas para as enchentes de 2008 e 2011 poderão ser importantes não
apenas para descrever quais foram os impactos proporcionados por esses dois eventos,
mas também para verificar, em um futuro próximo, consequências das mudanças
climáticas globais sobre estuários.
Outros fatores, que não os parâmetros ambientais medidos ou a enchente podem
estar influenciando a fauna de decápodos. A disponibilidade de alimento pode ser uma
das principais variáveis que controlam a abundância desses crustáceos, como
constataram Heyns & Froneman (2010) em seu estudo com o decápodos epibentônicos

66
em um estuário na África do Sul. Assim, o aumento na abundância durante o ano de
2009 pode ser consequência de um enriquecimento do estuário em termos de
produtividade. Vale destacar que esse aumento na captura ocorreu não apenas para os
decápodos (Leite, 2010), mas também para a ictiofauna do baixo-estuário do Itajaí-Açu
(Antunes, 2010). Isso sugere que diferentes grupos de organismos podem ter se
aproveitado do momento pós-enchente. Em relação à macrofauna do estuário, Silva
(2010) notou que houve a recuperação da comunidade seis meses após a enchente, mas
não foram encontrados valores elevados de abundância em 2009 como ocorreu para os
peixes e para os crustáceos decápodos.
A dieta de C. sapidus, C. danae, C. ornatus e X. kroyeri varia de acordo com a
disponibilidade de alimento, fato que deve se repetir em muitas outras espécies de
decápodos (Laughlin, 1982; Branco, 1996; Branco & Mortiz Jr., 2001; Mantelatto &
Christofoletti, 2001; Oliveira et al., 2006). C. sapidus pode predar diferentes
invertebrados bentônicos estuarinos e, na Lagoa dos Patos, onde é a espécie mais
abundante de Callinectes, seus principais itens alimentares são detritos e Erodona
mactroides, um molusco filtrador comum no local (Oliveira et al., 2006). C. danae, um
importante predador estuarino (Branco, 1996) raramente ingere material de origem
vegetal, se alimentando principalmente de macroinvertebrados, como crustáceos e
poliquetas (Branco & Verani, 1997). C. ornatus é um omnívoro oportunista, tendendo a
se alimentar de crustáceos e moluscos (Mantelatto & Christofoletti, 2001). No trabalho
de Stoner & Buchanan (1990), C. danae foi a espécie com dieta menos variada,
consumindo uma maior quantidade de presas de maior porte em comparação a C.
sapidus e C. ornatus. Embora generalistas na fase adulta, os juvenis de Callinectes
podem ser suscetíveis à indisponibilidade de alimento (Laughlin, 1982). Além disso,
diferente da fase adulta, os juvenis das diferentes espécies do gênero possuem
preferência por alimentos distintos (Stoner & Buchanan, 1990). Para C. ornatus, por
exemplo, observa-se que fêmeas ovígeras tendem a predar sobre organismos com
textura mais suave, como peixes, lulas ou camarões; juvenis se alimentam
principalmente de organismos sésseis e sedentários; conforme o tamanho aumenta, os
adultos, tanto machos como fêmeas, passam a consumir uma maior quantidade de
plantas, crustáceos e peixes (Mantelatto & Christofoletti, 2001). A alimentação dos
crustáceos decápodos é, portanto, generalista, varia muito de acordo com o estágio de
desenvolvimento do indivíduo, e é importante na distribuição espaço-temporal do
grupo, principalmente de indivíduos juvenis. Dessa forma, as alterações proporcionadas

67
pela enchente sobre a ictiofauna (Antunes, 2010) ou sobre a macrofauna (Silva, 2010)
podem ter influenciado a assembleia de decápodos, uma vez que esses dois grupos de
organismos podem fazer parte da dieta de siris do gênero Callinectes (e.g. Mantelatto &
Christofoletti, 2001). De fato, um estudo com C. sapidus em um estuário na Flórida,
EUA, concluiu que a resposta das presas às forçantes ambientais pode ser mais
importante do que o efeito das mesmas variáveis sobre os decápodos predadores
(Laughlin, 1982).
Outro tipo de impacto que pode estar afetando a fauna bentônica no estuário do
Itajaí são as dragagens, sobretudo aquelas de grandes proporções com o objetivo de
aumentar a profundidade do canal de acesso aos portos. O efeito da dragagem de
aprofundamento sobre os decápodos foi avaliado nesse estudo utilizando o MDS e a
ANOSIM. Essas análises revelaram diferenças significativas entre um período com
dragagem (em 2011) e outro sem dragagem (2010), porém somente no setor A (mais
externo). Tais diferenças sugerem um efeito direto da dragagem de aprofundamento
sobre a fauna, já que apenas a área do setor A está sujeita ao tráfego de grandes
embarcações e, portanto, é onde preferencialmente ocorrem as operações de
aprofundamento do canal.
Uma espécie responsável pelas diferenças nos períodos com e sem dragagem foi
X. kroyeri. No período sujeito a dragagem foi observada uma maior abundância desse
camarão do que no período sem dragagens. Esse incremento em X. kroyeri pode ser um
efeito da maior penetração da cunha salina em 2011 porque, como já visto, esse
camarão é marinho e entra no estuário acompanhando a cunha salina (Castro et al.,
2005). De fato, em alguns meses entre março e junho de 2010 (sem dragagem), foram
observados valores baixos de salinidade média, o que não ocorreu nos anos com
dragagens. A maior salinidade no ano com dragagem pode ter sido uma consequência
de mudanças no padrão de circulação do estuário promovidas pelo aumento na
profundidade, conforme discutido anteriormente.
Ainda em relação às mudanças ocorridas em períodos com e sem dragagens, foi
observado que, em 2011, a abundância de C. danae, principalmente no setor A, foi
mínima. Nesse período e setor ocorreu a dragagem de aprofundamento que pode ter
reduzido a biomassa desta espécie. C. danae dominou o estuário durante praticamente
todo o estudo e, portanto, é aquela espécie mais sujeita a mostrar impactos negativos.
Isso ocorre porque, quando um impacto acontece sobre diversas populações ao mesmo
tempo, aquela com maior número de indivíduos apresentará maior probabilidade de ser

68
atingida. Grandes reduções nessa espécie de Callinectes também foram observadas
analisando os efeitos das enchentes. Dauvin et al. (2010) estudaram a assembleia de
decápodos antes, durante e depois da construção de um porto em um estuário na França.
Os autores observaram que, em geral, houve uma diminuição na biomassa da espécie
dominante e aumento na riqueza durante dragagens. Entretanto, devido às mudanças
hidrológicas que também ocorreram no estuário, os autores concluíram que é difícil
afirmar quais mudanças realmente foram proporcionadas pela construção do porto. De
fato, devido ao grande número de variáveis que atuam sobre a fauna estuarina, é difícil
determinar qual o efeito de um impacto sobre a comunidade (Hughes et al., 2009;
Dauvin et al., 2010). Portanto, analisando os dois períodos contínuos de dragagens
(2009 e 2011), é possível perceber uma redução na abundância em 2011, entretanto
foram observadas grandes abundâncias em 2009, o que deixa dúvidas quanto ao real
impacto proporcionado por essa atividade. Nesse aspecto, torna-se importante a
aquisição de longas séries temporais antes e após os eventos para determinar o que
realmente causou as alterações (Hughes et al., 2009).
A assembleia de decápodos no estuário durante a dragagem de 2011 foi diferente
quando comparada com aquela encontrada no mesmo período dos demais anos de
estudo. Nessa ocasião foi identificada uma grande diminuição na abundância do siri C.
danae, o que não ocorreu com, por exemplo, C. ornatus. É comum que durante uma
perturbação uma espécie seja mais afetada do que outra. Kaiser & Spencer (1996),
analisando o impacto da rede de arrasto sobre a infauna em um estuário no Reino
Unido, constatou que as espécies mais abundantes foram, justamente, aquelas mais
afetadas pelo distúrbio. Assim, mesmo sendo uma espécie bastante tolerante a
variabilidade nos parâmetros ambientais, C. danae parece ser um dos decápodos com
maior susceptibilidade a grandes impactos no baixo-estuário do Itajaí-Açu. Com a
redução da espécie dominante novos nichos podem ser abertos e espécies antes raras
podem se tornar importantes na comunidade. Isso estaria de acordo com a maior
participação de outras famílias e espécies registradas durante os períodos com dragagem
em 201l, quando a abundância de C. danae esteve reduzida. Entretanto, assim que as
condições estiverem adequadas, a espécie mais comum tende a ocupar rapidamente os
espaços disponíveis, como discutido por Pinotti et al. (2011) em seu estudo com a
macrofauna na Lagoa dos Patos, RS.
A fauna bentônica estuarina apresenta elevada resiliência e, apesar dos
potenciais impactos decorrentes das enchentes e dragagens, é esperada uma rápida re-

69
colonização após o distúrbio (Bemvenuti et al., 2005). No entanto, muitos impactos que
atingem os organismos do baixo-estuário do Itajaí-Açu são de origem antrópica e se
acumulam ao longo do tempo o que poderia dificultar a recuperação após eventos de
maior magnitude como as grandes dragagens de aprofundamento ou as enchentes
(Antunes, 2010). Assim, o fato de terem sido observadas baixas abundâncias em 2011
pode ser não apenas efeito apenas da dragagem em si, mas de sucessivos impactos dos
quais a fauna ainda não teve tempo de se recuperar. Com uma elevada frequência de
perturbações a riqueza de espécies tende a ser baixa, de forma que nunca há tempo
suficiente para a comunidade atingir o equilíbrio (Connell, 1978). Em uma situação
oposta, se o tempo entre as perturbações for muito longo, a competição por recursos fará
com que a riqueza diminua novamente. A condição em que deve ser observado o maior
número de espécies é, portanto, quando existirem distúrbios em uma frequência ou
intensidade moderada (Connell, 1978). Diante da variabilidade naturalmente elevada
das suas condições ambientais, das alterações antrópicas promovidas em suas margens,
da promoção de sucessivas dragagens de aprofundamento e também da ocorrência de
enchentes de amplitudes variáveis, incluindo aquelas de proporções catastróficas, é
possível que o estuário do Itajaí-Açu esteja em uma situação sucessional próxima ao
primeiro caso, ou seja, submetido a uma grande frequência de perturbações, inclusive de
grande intensidade, o que explicaria a baixa riqueza de espécies e elevada dominância
observadas tanto para a fauna de decápodos como de peixes.
Por fim, como as enchentes são eventos imprevisíveis, é difícil que sejam
encontradas réplicas durante um estudo. Nesse aspecto, trabalhos envolvendo grandes
escalas temporais podem ser importantes para que os efeitos desses eventos sejam
percebidos. Já em relação às dragagens, por ocorrerem de forma contínua em muitos
estuários, metodologias envolvendo réplicas espaciais e temporais podem ser mais
eficientes na observação do efeito dessa atividade. Dentro do estuário do Itajaí-Açu, por
estar sujeito tanto a enchentes como dragagens, além de forte pressão antrópica nas
margens, uma abordagem como a do presente estudo, envolvendo o máximo de réplicas
temporais possíveis, parece ser a mais adequada visando a obtenção de respostas ainda
mais conclusivas sobre o efeito de tais perturbações sobre as comunidades locais.

70
7. CONCLUSÕES
• A vazão do rio Itajaí-Açu é a principal forçante afetando as variáveis salinidade,
pH e oxigênio dissolvido na água de fundo do estuário.
• A temperatura e a intrusão da cunha salina são os fatores mais relevantes para a
distribuição das espécies de decápodos no local.
• A ocupação das margens do Itajaí-Açu proporciona um diminuição da função
berçário do canal do estuário e, nessas condições, o Saco da Fazenda pode ser
um importante refúgio para algumas espécies. Ainda sim, espécies de
importância econômica são encontradas com frequência no baixo-estuário.
• O baixo-estuário é marcado pela dominância de poucas espécies que conseguem
tolerar as condições adversas desse ecossistema. Ocasionalmente espécies
marinhas ou de águas continentais são observadas no estuário, dependendo
principalmente da intrusão ou expulsão da cunha salina em determinados
setores.
• As enchentes causaram uma grande redução na abundância e biomassa de
crustáceos decápodos no baixo estuário do Itajaí-Açu. Entretanto, a elevada
resiliência desses organismos resultou em rápida recuperação da assembleia três
a quatro meses após o evento ocorrido em novembro de 2008.
• As dragagens podem estar afetando negativamente a fauna estuarina,
principalmente as espécies dominantes, ao mesmo tempo que pode propiciar
maior acesso à espécies marinhas para dentro do canal, mas esses efeitos são
difíceis de confirmar devido a grande quantidade de outros fatores que podem
ter proporcionado as alterações observadas no presente estudo.

71
8. REFERÊNCIAS BIBLIOGRÁFICAS Almeida, A. C.; Fransozo, V.; Furlan, M. & Giovannetti, N. 2007. Distribuição ecológica do camarão Nematopalaemon schmitti (Holthuis, 1950) (Crustacea, Decapoda, Caridea) no litoral norte do estado de São Paulo, Brasil. Anais do VIII Congresso de Ecologia do Brasil.
Angonesi, L. G. 2005. Dinâmica de curto prazo da macrofauna bentônica em uma enseada estuarina da Lagoa dos Patos: efeitos antrópicos e mecanismos de persistência e resiliência. Tese de doutorado. Fundação Universidade Federal do Rio Grande. 163 p. Antunes, A. 2010. Impactos da enchente, dragagem de aprofundamento e dinâmica estuarina sobre a ictiofauna no estuário do rio Itajaí-Açu, SC, Brasil. Tese de mestrado. Ciência e Tecnologia Ambiental - Universidade do Vale do Itajaí. 102 p.
AQUAPLAN. 2009. Relatório de impacto ambiental: rima da dragagem para aprofundamento do canal de acesso e da bacia de evolução do porto organizado de Itajaí, SC. SEP/PR e Superintendência do Porto de Itajaí. 412 p. Araújo, S. A. & Reis, F. H. 2012. Relatório Climatológico. Estação meteorológica automática. Universidade do Vale do Itajaí. Itajaí. Laboratório de Climatologia. Bail, G. C.; Branco, J. O.; Freitas Júnior, F.; Lunardon-Branco, M. J. & Braun, J. R. R. 2009. Fauna acompanhante do camarão sete barbas, na Foz do Rio Itajaí-Açú e sua contribuição na diversidade de crustáceos e peixes do ecossistema Saco da Fazenda. In: Branco, J. O.; Lunardon-Branco, M. J. & Bellotto, V. R. 2009. Estuário do Rio Itajaí-Açú, Santa Catarina: caracterização ambiental e alterações antrópicas. UNIVALI. 284-312 p. Barreto, A. V.; Batista-Leite, L. M. A. & Aguiar, M. C. A. 2006. Maturidade sexual das fêmeas de Callinectes danae (Crustacea, Decapoda, Portunidae) nos estuários dos rios Botafogo e Carrapicho, Itamaracá, PE, Brasil. Série Zoologia. 96 (2):141-146 p. Begon, M.; Harper, J. L. & Townsend, C. R. 1996. Ecology: individuals, populations and communities. Blackwell Science. 1068 p. Bellotto, V. R.; Kuroshima, K. N. & Cecanho, F. 2009. Poluentes no ambiente estuarino e efeitos da atividade de dragagem, 105-126p. In: Branco, J. O.; Lunardon-Branco M. J. & Bellotto V. R. 2009. Estuário do rio Itajaí-Açu, Santa Catarina: Caracterização Ambiental e Alterações Antrópicas. UNIVALI. 105-126 p. Bemvenuti, C. E. & Colling, L. A. 2010. As comunidades de macroinvertebrados bentônicos. In: Seeliger, U. & Odebrecht, C. 2010. O estuário da Lagoa dos Patos: um século de transformações. FURG. 101-114 p. Bemvenuti, C. E. & Rosa-Filho, J. S. 2000. Estrutura e dinâmica das associações de macroinvertebrados bentônicos dos ambientes estuarinos do Rio Grande do Sul: um estudo de caso. In: Avaliação e ações prioritárias para a zona costeira e marinha, 2000, Porto Seguro. Workshop... Porto Seguro: PROBIO.

72
Bemvenuti, C. E.; Angonesi, L. G. & Gandra, M. S. 2005. Effects of dredging operations on soft bottom macrofauna in a harbor in the Patos Lagoon Estuarine region of southern Brazil. Braz. J. Biol. 65(4): 573-581 p. Braga, A.A.; Fransozo, A.; Bertini, G. & Fumis, P.B. 2005. Composição e abundância dos caranguejos (Decapoda, Brachyura) nas regiões de Ubatuba e Caraguatatuba, Litoral Norte Paulista, Brasil. Biota Neotropica. 5(2): 34 p. Branco, J. O. 1996. Variações sazonais e ontogênicas na dieta natural de Callinectes danae Smith, 1869 (Decapoda, Portunidae) na Lagoa da Conceição, Florianópolis, SC. Arq. Biol. Tecnol. 39 (4): 999-1012 p. Branco, J. O. & Verani, J. R. 1997. Dinâmica da alimentação natural de Callinectes danae, Smith (Decapoda, Portunidae) na Lagoa da Conceição, Florianópolis, Santa Catarina, Brasil. Revta Bras. Zool. 14 (4): 1003-1018 p. Branco, J. O. & Verani, J. R. 1998. Estudo populacional do camarão-rosa Penaeus
paulensis Perez Farfante (Natantia, Penaeidae) na Lagoa da Conceição, Santa Catarina, Brasil. Revta. Bras. Zool. 15(2): 353-364 p. Branco, J. O.; Lunardon-Branco, M. J.; Souto, F. X. & Guerra, C. R. 1999. Estrutura populacional do camarão sete-barbas Xiphopenaeus kroyeri (Heller, 1862), na foz do rio Itajaí-Açú, Itajaí, SC, Brasil. Braz. Arch. Biol. Technol. 42(1): 115-126 p. Branco, J. O. 2000. Avifauna associada ao estuário do Saco da Fazenda, Itajaí, SC. Revta. Bras. Zool. 17(2): 387-394 p. Branco, J. O. & Masunari, S. 2000. Reproductive ecology of the blue crab, Callinectes
danae Smith, 1869 in the Conceição Lagoon System, Santa Catarina Isle, Brazil. Revta. Bras. Biol. 60(1): 17 – 27 p. Branco, J. O. & Mortiz Jr., H. C. 2001. Alimentação natural do camarão sete-barbas (Xiphopenaeus kroyeri), na Armação do Itapocoroy, Penha, SC. Revta Bras. Zool. 18 (1): 53-61 p. Branco, J. O. & Verani, J. R. 2006. Pesca do camarão sete-barbas e sua fauna acompanhante, na Armação do Itapocoroy, Penha, SC. In: Branco, J. O. & Marenzi, A. W. C. Bases ecológicas para um desenvolvimento sustentável: estudos de caso em Penha, SC. UNIVALI. 153-170 p. Branco, J. O. & Freitas Jr., F. 2009. Análise quali-quantitativa dos crustáceos no ecossistema Saco da Fazenda, Itajaí, SC. In: Branco, J. O.; Lunardon-Branco M. J. & Bellotto V. R. 2009. Estuário do rio Itajaí-Açu, Santa Catarina: Caracterização Ambiental e Alterações Antrópicas. UNIVALI. 180 – 206 p. Branco, J. O.; Freitas Jr., F.; Verani, J. R. & Hostim-Silva, M. 2009a. Ictiofauna do Saco da Fazenda, Itajaí, SC. In: Branco, J. O.; Lunardon-Branco M. J. & Bellotto V. R. 2009. Estuário do rio Itajaí-Açu, Santa Catarina: Caracterização Ambiental e Alterações Antrópicas. UNIVALI. 207- 226 p.

73
Branco, J. O.; Lunardon-Branco M. J. & Bellotto V. R. 2009b. Estuário do rio Itajaí-Açu, Santa Catarina: Caracterização Ambiental e Alterações Antrópicas. UNIVALI. 312 p. Cardoso, P. G.; Raffaelli, D.; Lilleb, A.I., Verdelhos, T. & Pardal, M. A. 2008. The impact of extreme flooding events and anthropogenic stressors on the macrobenthic communities dynamics. Est., Coast. Shelf Sci. 76: 553 – 565 p. Castro, R. H.; Costa, R. C.; Fransozo, A. & Mantelatto, F. L. M. 2005. Population structure of the seabob shrimp Xiphopenaeus kroyeri (Heller, 1862) (Crustacea: Penaeoidea) in the littoral of São Paulo, Brazil. Scientia Marina. 69 (1): 105-112 p. Clarke, K. R. & Warwick, R. M. 1994. Change in marine communities: an approach to statistical analysis and interpretation. Plymouth Marine Laboratory. 144 p. Clarke, K. R. & Gorley, R. N. 2006. PRIMER v6: User Manual/Tutorial. PRIMER-E: Plumouth. 190 p. Connell, J. H. 1978. Diversity in tropical rain forests and coral reefs: high diversity of trees and corals is maintained only in a nonequilibrium state. Science. 199: 1302-1310 p. Costa, R. C.; Fransozo, A.; Mantelatto, F. L. M. & Castro, R. H. 2000. Occurrence of shrimp species (Crustacea: Decapoda: Natantia: Penaeidea and Caridea) in Ubatuba Bay, Ubatuba, SP, Brazil. Proceedings of the Biological Society of Washington. 113(3):776-781 p. Costa, R. C.; Fransozo, A.; Melo, G. A. S. & Freire, F. A. M. 2003. Chave ilustrada para identificação dos camarões Dendrobranchiata do litoral norte do estado de São Paulo, Brasil. Biota Neotropica. 3(1): 12 p. Costa, R. C.; Fransozo, A. & Pinheiro, A. P. 2004. Ecological distribution of the shrimp Pleoticus muelleri (Bate, 1888) (Decapoda: Penaeoidea) in southeastern Brazil. Hydrobiologia. 529: 195–203 p. Costa, I. D. & Beneditto, A. P. M. 2009. Caracterización preliminar de los invertebrados bentónicos capturados accidentalmente en la pesca de camarones en el norte del estado de Río de Janeiro, sudeste de Brasil. Lat. Am. J. Aquat. Res. 37(2): 259-264 p. Costa-Dias, S.; Freitas, V.; Sousa, R. & Antunes, C. 2010. Factors influencing epibenthic assemblages in the Minho Estuary (NW Iberian Peninsula). Marine Pollution Bulletin 61: 240-246 p. Dauvin, J. C.; Alizier, S.; Vallet C. & Ruellet T. 2010. Does the Port 2000 harbour construction have an effect on the Seine estuary suprabenthic community? Estuarine, Coastal and Shelf Science. 86: 42–50 p. Day Jr., J. W.; Hall, C. A. S.; Kemp, W. M & Yáñez-Arancibia, A. 1989. Estuarine ecology. John Wiley. 558 p.

74
Delfim, R. 2009. Avaliação da hipoxia e da qualidade de água no estuário do Rio Itajaí-Açú- SC. Monografia. Universidade do Vale do Itajaí. 85 p. D’Incao, F. 1995. Taxonomia, padrões distribucionais e ecológicos dos Dendrobranchiata (Crustacea: Decapoda) do Brasil e Atlântico Ocidental. Tese de Doutorado. Ciências na área de zoologia – Universidade Federal do Paraná. 365 p. D’Incao, F.; Valentini, H. & Rodrigues, L. F. 2002. Avaliação da pesca de camarões nas regiões sudeste e sul do Brasil. 1965-1999. Atlântica. 24(2): 103-116 p. D’Incao, F. & Dumont, L. F. C. 2010. A comunidade de crustáceos decápodes. In: Seeliger, U. & Odebrecht, C. 2010. O estuário da Lagoa dos Patos: um século de transformações. FURG. 117-122 p. Dugan, P. J & Livingston, R. J. 1982. Long-term Variation of Macroinvertebrate Assemblages in Apalachee Bay, Florida. Est., Coast., Shelf Sci.14: 391-403 p. Erftemeijer, P. L. A. & Lewis, R. R. R. 2006. Environmental impacts of dredging on seagrasses: A review. Marine Pollution Bulletin 52: 1553–1572 p. Eyre, B. D. & Ferguson, A. J. P. 2006. Impact of a flood event on benthic and pelagic coupling in a sub-tropical east Australian estuary (Brunswick). Est., Coast., Shelf Sci. 66: 111-122 p. Ferreira, L. S. & D’Incao, F. 2008. Crescimento de Callinectes sapidus (Crustacea, Decapoda, Portunidae) no estuário da laguna dos Patos, RS, Brasil. Série Zoologia. 98(1):70-77 p. Ferreira, R. S. 2009. Taxonomia e padrões distribucionais dos camarões marinhos e estuarinos da família Palaemonidae (Crustacea: Decapoda: Caridea) do litoral brasilerio. Tese de mestrado. Universidade Federal do Rio Grande. 160 p. Freitas, Jr. F.; Christoffersen, M. L. & Branco, J. O. 2010. Monitoring of carcinofauna abundance and diversity during eight years of expressway construction in Santa Catarina, Brazil. Lat. Am. J. Aquat. Res. 38(3): 461-473 p. Garcia, A. M.; Vieira, J. P.; Bemvenuti, C. E. & Geraldi, R. M. 1996. Abundância e diversidade da assembléia de crustáceos decápodos dentro e fora de uma pradaria de Ruppia maritima L. no estuário da Lagoa dos Patos (RS-Brasil). NAUPLIUS. 4:113-128 p. González-Ortegón, E.; Subida, M. D.; Cuesta, J. A.; Arias, A. M.; Fernández-Delgado C. & Drake, P. 2010. The impact of extreme turbidity events on the nursery function of a temperate European estuary with regulated freshwater inflow. Est., Coast. Shelf Sci. 87: 311–324 p. Heyns, E & Froneman, W. 2010. Spatial and temporal patterns in the hyperbenthic community structure in a warm temperate southern African permanently open estuary. Est., Coast. Shelf Sci. 88: 105-115 p.

75
Hobbie, J. E. 2000. Estuarine science: a synthetic approach to research and practice. Island Press, Washington, 539 p. Hostim-Silva, M.; Vicente, M. J. D.; Figna, V. & Andrade, J. P. 2002. Ictiofauna do Rio Itajaí- Açu, Santa Catarina, Brasil. Notas Tec. Facimar. 6: 127-135 p. Hughes, C.; Richardson, C. A.; Luckenbach, M. & Seed, R. 2009. Difficulties in separating hurricane induced effects from natural benthic succession: Hurricane Isabel, a case study from Eastern Virginia, USA. Est., Coast. Shelf Sci. 85: 377-386 p. INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE (IPCC). 2007. Climate Change 2007 – Summary for Policymakers: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the IPCC. IPCC, Cambridge University Press, Cambridge, 2007. Disponível em: <http://www.ipcc.ch/>. Acesso em: 23 de outubro de 2010. Jones, K. K.; Simenstad, C. A.; Higley, D. L. & Bottom, D. L. 1990. Community structure, distribution, and standing stock of benthos, epibenthos, and plankton in the Columbia River Estuary. Prog. Oceanog. 25: 211-241 p. Kaiser, M. J. & Spencer, B. E. 1996. The effects of beam-trawl disturbance on infaunal communities in different habitats. Journal of Animal Ecology. 65: 348-358 p. Knox, G. A. 2001. The ecology of seashores. CRC Press, New Zealand, 577 p. Kodama, K.; Oyama, M.; Lee, J.; Kume, G.; Yamaguchi, A.; Shibata, Y.; Shiraishi, H.; Morita, M.; Shimizu, M. & Horiguchi, T. 2010. Drastic and synchronous changes in megabenthic community structure concurrent with environmental variations in a eutrophic coastal bay. Progress in Oceanography. 87: 157–167 p. Kuroshima, K. N. & Bellotto, V. R. 2009. Dinâmica dos nutrientes inorgânicos e orgânicos na foz do rio Itajaí-Açú. In: Branco, J. O.; Lunardon-Branco M. J. & Bellotto V. R. 2009. Estuário do rio Itajaí-Açu, Santa Catarina: Caracterização Ambiental e Alterações Antrópicas.UNIVALI. 93 – 104 p. Lana, P. C. 1996. O bentos da costa brasileira: avaliação critíca e levantamento bibliográfico (1858-1996). Rio de Janeiro, FEMAR. Laughlin, R. A. 1982. Feeding habits of the blue crab, Callinectes sapidus Rathbun, in the Apalachicola Estuary, Florida. Bulletin of Marine Science. 32(4): 807-822 p. Leite, I. P. 2010. Variação espacial e temporal da composição e abundância relativa dos macrocrustáceos epibentônicos do estuário do Itajaí-Açu, SC. Monografia. Centro de Ciências Tecnológicas da Terra e do Mar – UNIVALI. 34 p. Leps, J. & Smilauer, P. 2003. Multivariate analysis of ecological data using CANOCO. Cambridge University Press. 269 p.

76
Lindegarth, M. & Hoskin, M. 2001. Patterns of distribution of macro-fauna in different types of estuarine, soft sediment habitats adjacent to urban and non-urban areas. Est., Coast. Shelf Sci. 52: 237-247 p. Lopes, M. 2008. Ecologia populacional dos camarões-rosa, Farfantepenaeus
brasiliensis (Latreille, 1817) e F. paulensis (Pérez-Farfante, 1967) (Decapoda: Dendrobranchiata: Penaeidae) em áreas de berçário do litoral norte de São Paulo. Tese de mestrado. Universidade Estadual Paulista. 204 p. Mantelatto, F. L. M. & Christofoletti, R. A. 2001. Natural feeding activity of the crab Callinectes ornatus (Portunidae) in Ubatuba Bay (São Paulo, Brazil): influence of season, sex, size and molt stage. Marine Biology. 138: 585-594 p. Mantelatto, F. L. M. & Garcia, R. B. 2001. Biological aspects of the nonindigenous Portunid crab Charybdis hellerii in the western tropical south atlantic. Bull. Mar. Sci. 68(3):469 – 477 p. Marques, J. C.; Maranhão, P. & Pardal, M. A. 1993. Human impact assessment on the subtidal macrobenthic community structure in the Mondego Estuary (Western Portugal). Est., Coast. Shelf Sci. 37: 403-419 p. McLusky, D.S. 1989. The estuarine ecosystem. 2nd Edition. Blackie Academic and Professional. 215 p. Medeiros, A. D. 2003. A Influência da Maré e da Batimetria sobre a intrusão salina no estuário do Rio Itajaí-Açu.. Tese de mestrado. COPPE - Universidade Federal do Rio de Janeiro. 76 p. Melo, G.A.S. 1996. Manual de Identificação dos Brachyura (Caranguejos e Siris) do Litoral Brasileiro. São Paulo, Plêiade/FAPESP. 603 p. Mendonça, J. T.; Verani, J. R. & Nordi, N. 2010. Evaluation and management of blue crab Callinectes sapidus (Rathbun, 1896) (Decapoda - Portunidae) fishery in the Estuary of Cananéia, Iguape and Ilha Comprida, São Paulo, Brazil. Braz. J. Biol.70(1): 37-45 p. Miranda, L. B.; Castro, B. M. & Kjerfve, B. 2002. Princípios de oceanografia física de estuários. São Paulo, Editora da Universidade de São Paulo. 424 p. Newell, R. C.; Seiderer, L. J. & Hitchcock, D. R. 1998. The impact of dredging works in coastal Waters: a review of the sensitivity to disturbance and subsequent recovery of biological resources on the sea bed. Oceanography and Marine Biology: an Annual Review. 36:127-78 p. Norse, E. A. 1978. An experimental gradient analysis: hyposalinity as an “upstress” distributional determinant for Caribbean Portunid Crabs. Biol. Bull. 155: 586-598 p. Odum, E. P. 1983. Ecologia. Editora Guanabara. 434 p.

77
Ogburn, M. B.; Jackson, J. L. & Forward Jr, R. B. 2007. Comparison of low salinity tolerance in Callinectes sapidus Rathbun and Callinectes similis Williams postlarvae upon entry into an estuary. J. Exp. Mar. Biol. Ecol. 343–350 p. Ohimain, E. I.; Imoobe, T. O. & Bawo, D. D. S. 2008. Changes in Water Physico-Chemical Properties Following the Dredging of an Oil Well Access Canal in the Niger Delta. World Journal of Agricultural Sciences. 4 (6): 752-758 p. Oliveira, A.; Pinto, T. K.; Santos, D. P. D. & D’Incao, F. 2006. Dieta natural do siri-azul Callinectes sapidus (Decapoda, Portunidae) na região estuarina da Lagoa dos Patos, Rio Grande, Rio Grande do Sul, Brasil. Sér. Zool. 96 (3): 305-313 p. Paperno, R. & Brodie, R. B. 2004. Effects of environmental variables upon the spatial and temporal structure of a fish community in a small, freshwater tributary of the Indian River Lagoon, Florida. Est., Coast. Shelf Sci. 61: 229 -241 p. Pereira Filho, J.; Spillere, L. C. & Schettini, C. A. F. 2003. Dinâmica de nutrientes na região portuária do estuário do Rio Itajaí-Açu, SC. Atlântica. 25(1): 11-20 p. Pereira, M. J. 2006. Estrutura populacional do gênero Callinectes na Baía da Babitonga, São Francisco do Sul, SC. Tese de mestrado. Universidade do Vale do Itajaí. 61 p. Peterson, M. S. & Ross, S. T. 1991. Dynamics of littoral fishes and decapods along a coastal river-estuarine gradient. Est., Coast. Shelf Sci. 33: 467-483 p. Pezzuto, P. R.; Alvarez-Perez, J. A. & Wahrlich, R. 2008.The use of the swept area method for assessing the seabob shrimp Xiphopenaeus kroyeri (Heller, 1862) biomass and removal rates based on artisanal fisheryderived data in southern Brazil: using depletion models to reduce uncertainty. Lat. Am. J. Aquat. Res. 36(2):245-257 p. Piazza, B. P. & Peyre, M. K. L. 2009. The effect of Hurricane Katrina on nekton communities in the tidal freshwater marshes of Breton Sound, Louisiana, USA. Est., Coast. Shelf Sci. 83: 97–104 p. Pillay, D. & Perissinotto, R. 2008. The benthic macrofauna of the St. Lucia Estuary during the 2005 drought year. Est., Coast. Shelf Sci. 77: 35-46 p. Pinotti, R. M.; Colling, L. A. & Bemvenuti, C. E. 2011. Temporal dynamics of deep infralittoral macrobenthic fauna inside a subtropical estuarine environment. Braz. J. Aquat. Sci. Technol. 15(1): 26-41 p. Porath, S. L. 2004. A paisagem de rios urbanos: A presença do rio Itajaí-Açu na cidade de Blumenau. Tese de Mestrado. Programa de Pós-Graduação em Arquitetura e Urbanismo -Universidade Federal de Santa Catarina.150 p. Porto de Itajaí. Porto de Itajaí. Disponível em: <http://www.portoitajai.com.br>. Acesso em: 04 fev. 2012. Posey, M. H.; Alphin, T. D.; Harwell, H. & Allen, B. 2005. Importance of low salinity areas for juvenile blue crabs, Callinectes sapidus Rathbun, in river-dominated estuaries

78
of southeastern United States. Journal of Experimental Marine Biology and Ecology. 319: 81-100 p. Raffaelli, D.; Conacher, A.; McLachlan, H. & Emes, C. 1989. The role of epibenthic crustacean predators in na estuarine food web. Est., Coast. Shelf Sci. 28: 149- 160 p. Resgalla Jr., C. 2009. Zooplâncton do estuário do Rio Itajaí-Açu e zona costeira adjacente. In: Branco, J. O.; Lunardon-Branco M. J. & Bellotto V. R. 2009. Estuário do rio Itajaí-Açu, Santa Catarina: Caracterização Ambiental e Alterações Antrópicas. UNIVALI. 171-179 p. Roedel, T. 2009. Carcinofauna acompanhante na pesca artesanal do camarão sete-barbas, na Armação do Itapocoroy, Penha, SC: avaliação e gestão. Tese de mestrado. Ciência e Tecnologia Ambiental - Universidade do Vale do Itajaí. 98 p. Rozas, L. P.; Minello, T. J.; Munuera-Fernández, I.; Fry, B. & Wissel, B. 2005. Macrofaunal distributions and habitat change following winterespring releases of freshwater into the Breton Sound estuary, Louisiana (USA). Est. Coast. Shelf Sci. 65: 319-336 p. Sánchez-Moyano, J. E. & García-Asencio, I. 2010. Crustacean assemblages in a polluted estuary from South-Western Spain. Marine Pollution Bulletin. 60: 1890–1897 p. Schettini, C. A. F. 2001. Dinâmica de sedimentos finos no estuário do rio Itajaí-Açu, SC. Tese de Doutorado. Universidade Federal do Rio Grande do Sul. 107 p. Schettini, C. A. F. 2002. Caracterização física do estuário do rio Itajaí-Açu, SC. Revista Brasileira de Recursos Hídricos. 7(1):123-142 p. Schettini, C. A. F. & Truccolo, E. C. 2009. Circulação do baixo estuário do rio Itajaí. In: Branco, J. O.; Lunardon-Branco M. J. & Bellotto V. R. 2009. Estuário do rio Itajaí-Açu, Santa Catarina: Caracterização Ambiental e Alterações Antrópicas.UNIVALI. 13 – 26p. Severino-Rodrigues, E. S.; Pita, J. B. & Graça-Lopes, R. 2001. Pesca artesanal de siris (Crustacea, Decapoda, Portunidae) na região estuarina de Santos e São Vicente (SP), Brasil. Boletim do Instituto de Pesca. 27 (1): 7-19 p. Silva, E. S. 2010. Efeito da elevação abrupta da vazão no Rio Itajaí-Açu, decorrente da enchente de 2008, sobre a macrofauna bentônica do estuário do Rio Itajaí-Açu. Monografia. Centro de Ciências Tecnológicas da Terra e do Mar - UNIVALI. 33 p. Smith, R.; Boyd, S. E.; Rees, H. L.; Dearnaley, M. P. & Stevenson, J. R. 2006. Effects of dredging activity on epifaunal communities e Surveys following cessation of dredging. Est. Coast. Shelf Sci. 70: 207-223 p. Stoner, A. W. & Buchanan, B. A. 1990. Ontogeny and overlap in the diets of four tropical Callinectes species. Bulletin of Marine Science. 46(1): 3-12 p.

79
Tamburus, A. F.; Mossolin, E. C. & Mantelatto, F. L. 2012. Population and reproductive aspects of Macrobrachium acanthurus (Wiegmann, 1836) (Crustacea: Palaemonidae) from north coast of São Paulo state, Brazil. Braz. J. Aquat. Sci. Technol. 16(1): 9-18 p. Tavares, M. & Mendonça Jr., J. B. 2004. Introdução de crustáceos decápodes exóticos no Brasil: uma roleta ecológica. In: Silva, J. S. V. & Souza, R. C. C. L. 2004. Água de Lastro e Bioinvasão. Interciência. 59- 76 p. ter Braak, C. J. F. & Šmilauer, P. (2002). CANOCO Reference Manual and CanoDraw for Windows User's Guide: Software for Canonical Community Ordination (version 4.5). Microcomputer Power (Ithaca NY, USA). 500 p. The MerMex Group. 2011. Marine ecosystems’ responses to climatic and anthropogenic forcings in the Mediterranean. Progress in Oceanography. 91: 97–166 p. Torres, R. J. 2000. Uma análise preliminar dos processos de dragagem do porto de Rio Grande, RS. Tese de Mestrado. Fundação Universidade Federal do Rio Grande. 190 p. Valenti, W. C.; Mello, J. T. C. & Lobão, V. L. 1989. Fecundidade em Macrobrachium
acanthurus (Wiegmann, 1838) do rio Ribeira do Iguape (Crustacea, Decapoda, Palaemonidae). Revista Brasileira de Zoologia. 6(1): 9-15 p. Veado, L. 2008. Variação espaço-temporal do zooplâncton do baixo estuário do rio Itajaí-Açu, SC. Tese de Mestrado. Universidade do Vale do Vale do Itajaí. 71 p. Williams, A. B. 1974. The swimming crabs of the genus Callinectes (Decapoda: Portunidae). Fishery Bulletin. 72(3): 685-798 p. Williams, A. B. 1984. Shrimps, lobsterns, and crabs of the Atlantic Coast of the Eastern United States, Maine to Florida. Smithsonian Institution Press. 550 p. Winter , V. C. & Masunari, S. 2006. Macroepizoísmo em Libinia ferreirae (Crustacea, Brachyura, Majidae). Sér. Zool. 96 (2):135-140 p.
Top Related