
CENTRO DE ESTUDOS GERAIS INSTITUTO DE BIOLOGIA …livros01.livrosgratis.com.br/cp065131.pdf · 2.2)...
101
MARIANA RODRIGUES PEREIRA ESTUDO DA EXPRESSÃO DE RECEPTORES DE ADENOSINA EM RETINA DISSERTAÇÃO SUBMETIDA À UNIVERSIDADE FEDERAL FLUMINENSE VISANDO A OBTENÇÃO DO GRAU DE MESTRE EM NEUROIMUNOLOGIA Orientador: Roberto Paes de Carvalho UNIVERSIDADE FEDERAL FLUMINENSE 2005 CENTRO DE ESTUDOS GERAIS INSTITUTO DE BIOLOGIA PROGRAMA DE NEUROIMUNOLOGIA
Transcript of CENTRO DE ESTUDOS GERAIS INSTITUTO DE BIOLOGIA …livros01.livrosgratis.com.br/cp065131.pdf · 2.2)...

MARIANA RODRIGUES PEREIRA
ESTUDO DA EXPRESSÃO DE RECEPTORES DE ADENOSINA EM RETINA
DISSERTAÇÃO SUBMETIDA À UNIVERSIDADE FEDERAL
FLUMINENSE VISANDO A OBTENÇÃO DO GRAU DE MESTRE EM NEUROIMUNOLOGIA
Orientador: Roberto Paes de Carvalho
UNIVERSIDADE FEDERAL FLUMINENSE 2005
CENTRO DE ESTUDOS GERAIS INSTITUTO DE BIOLOGIA PROGRAMA DE NEUROIMUNOLOGIA

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

MARIANA RODRIGUES PEREIRA
ESTUDO DA EXPRESSÃO DE RECEPTORES DE ADENOSINA EM RETINA
Trabalho desenvolvido no laboratório de Neurobiologia Celular do Departamento de Neurobiologia, Instituto de Biologia, UFF.
Orientador: Roberto Paes de Carvalho
Niterói 2005
Dissertação de Mestrado submetida à Universidade Federal Fluminense como requisito parcial para obtenção de grau de Mestre em Neuroimunologia

P 434 Pereira, Mariana Rodrigues Estudo da expressão de receptores de adenosina em retina / Mariana Rodrigues Pereira. ______ Niterói: [s.n.], 2005. 108 f.:il. Dissertação. ______ (Mestrado em Neuroimunologia). _____ Universidade Federal Fluminense, 2005. 1. Anatomia animal. 2. Adenosina. 3. Vertebrados – Visão - Fisiologia. 4. Nervo ótico. I. Título.
CDD. 591.48

ESTE TRABALHO FOI DESENVOLVIDO NO LABORATÓRIO DE NEUROBIOLOGIA CELULAR, DO PROGRAMA DE NEUROIMUNOLOGIA DO INSTITUTO DE BIOLOGIA DA UNIVERSIDADE FEDERAL FLUMINENSE SOB ORIENTAÇÃO DE ROBERTO PAES DE CARVALHO E NA VIGÊNCIA DE AUXÍLIOS CONCEDIDOS PELO CONSELHO NACIONAL DE DESENVOLVIMENTO CIENTÍFICO E TECNOLÓGICO (CNPq), COORDENAÇÃO DE APERFEIÇOAMENTO DE PESSOAL DE ENSINO SUPERIOR (CAPES) E FUNDAÇÃO DE AMPARO A PESQUISA DO ESTADO DO RIO DE JANEIRO (FAPERJ).
ii

SUMÁRIO Lista de Abreviaturas viii Resumo xi Abstract xii I) Introdução 1 1) Adenosina 1 1.1) Metabolismo 1 1.2) Transporte 3 1.3) Receptores 6 1.3.1) Distribuição dos receptores 9 1.3.2) Localização celular e subcelular no SNC 10 1.3.3) Estrutura dos receptores 11 1.3.4) Dimerização de receptores 13 2) Receptores A1 e A3 de adenosina 13 2.1) Sinalização via adenilil ciclase 13 2.2) Sinalização por fosfolipase C 15 2.3) Sinalização através de canais iônicos 16 2.4) Sinalização via MAP kinases 17 2.5) Dessensibilização 18 2.6) Principais funções 20 3) Receptores A2a e A2b de adenosina 21 3.1) Sinalização via adenilil ciclase 21 3.2) Sinalização através de fosfolipase C 22 3.3) Sinalização pelas MAP kinases 22 3.4) Dessensibilização 24 3.5) Principais funções 24 4) Modelo de estudo: a retina 26
II) Objetivos 29
vi

III) Materiais e métodos 31 3.1) Materiais 31 3.2) Métodos 32 3.2.1) Culturas complexas 32 3.2.2) Culturas de agregados 33 3.2.3) Tratamento com drogas 33 3.2.4) “Binding” em células viva 34 3.2.5) Dosagem de proteína pelo método de Bradford 35 3.2.6) Eletroforese em SDS-poliacrilamida SDS-(PAGE) 36 3.2.7) “Western Blotting” 37 3.2.8) Retirada dos anticorpos e “bloting” para actina 39 3.2.9) Análises estatísticas 40 IV) Resultados 41 4.1) Ligação de 3(H)CCPA ao receptor A1 em culturas complexas em monocamada e em culturas de agregados
41
4.2) O tratamento das culturas complexas em monocamada com análogos permeáveis de AMPc aumenta a ligação do 3(H)DPCPX ao receptor A1
43
4.3) A ativação do receptor A2a é capaz de aumentar a ligação do 3(H)DPCPX ao receptor A1
48
4.4) O bloqueio da atividade da PKA é capaz de bloquear o efeito do receptor A2a
52
4.5) O bloqueio ou a ativação do receptor A1 aumenta a sua ligação ao 3(H)DPCPX
54
4.6) O tratamento com adenosina deaminase promove aumento da ligação de 3(H)DPCPX ao receptor A1
57
V) Discussão 61 VI) Conclusões 71 VI) Referências Bibliográficas 73
vii

AGRADECIMENTOS
Em primeiro lugar a Deus pelo dom da vida e por todas as oportunidades que cruzaram o meu caminho. Ao meu orientador Roberto, pela oportunidade e apoio, pelos ensinamentos e a confiança depositada em mim. Muito obrigada! À minha co-orientadora e grande amiga Jainne, por tudo o que me ensinou, por sempre ter acreditado que eu conseguiria desenvolver esse projeto, pelos momentos de alegria e descontração no laboratório e pela força e estímulo nos momentos difíceis. Aos colegas de laboratório que são tão importantes: Alexandre, a pessoa que consegue ser mais calma do que eu. Obrigada por toda a ajuda que me deu com o Western e pelas risadas garantidas. Daniel, um amigo para toda hora. Sempre disposto a ajudar tanto com o computador quanto na bancada. Octavia, o completo oposto de mim em todos os sentidos. Acho que é por isso que nos damos tão bem. Obrigada pela força e por todas as piadas geradas sobre você. Cristiane, amiga e companheira. Pela ajuda e apoio. Márcia, pelas discussões sobre os nossos trabalhos que me ajudaram muito. Marcelo, Camila, Renato, Raquel, Luciane e Vivian pela amizade. Os amigos que passaram pelo laboratório: Igor, Rochele, Rodrigo e Vanessa. Vanusa pelos experimentos iniciais. Elaine e Luzeli pela amizade e apoio técnico. À Aninha por tudo o que me ajudou, pela paciência em me esperar todas as vezes em que me atrasei para fazermos os experimentos. Você se tornou uma verdadeira amiga nesse tempo em que trabalhamos juntas. À Paulinha pelos ensinamentos, pela amizade e apoio e pela ajuda na câmara escura. À Karin pela contribuição, interesse e disposição para ajudar. À Ana Ventura pelos conhecimentos passados e por tudo o que eu já pedi emprestado no seu laboratório. Aos professores Beth e Cláudio pela ajuda e pelas ótimas aulas. À minha irmã Ana e ao meu cunhado Max que mesmo distantes sempre me apoiaram nas minhas decisões e à minha sobrinha Carolina, que mesmo ainda não ter vindo ao mundo, já é tão importante na minha vida. Ao meu noivo Diego, pelo carinho, ajuda e companheirismo. Não tenho palavras para dizer o quanto você é importante para mim. Obrigada pelo apoio e paciência e por estar sempre presente em todos os momentos.
iii

“Mestre não é quem ensina,
mas quem de repente aprende”
Guimarães Rosa
iv

v
Dedico este trabalho à minha família: Diego, Ana, Max e Carolina.

RESUMO
A adenosina é um importante neuromodulador no SNC e suas ações
são promovidas pela ativação dos receptores A1, A2a, A2b e A3. Trabalhos
anteriores do laboratório mostraram a presença de receptores A1 e A2 em
retina intacta de pinto. Em cultura, a detecção do receptor A1 depende do
estado de agregação das células. Culturas em monocamada mostraram uma
ligação baixa ou nula do agonista A1 3(H)CCPA ou antagonista 3(H)DPCPX. Incubação prolongada com análogos permeáveis de AMPc
como SpAMPc ou 8 BrAMPc foi capaz de aumentar a detecção do
receptor. Além disso, forskolina, um ativador direto da adenilil ciclase,
também aumentou marcadamente a ligação. Estes resultados sugeriam a
modulação da detecção dos receptores A1 via ativação de receptores A2a.
Os receptores A2a de adenosina estão expressos nas retinas de
embriões a partir de 7 dias de desenvolvimento (E7) onde o nível de
expressão é alto. Em E9 o nível cai cerca de 45% e se mantém constante
até E16. Este receptor também foi encontrado em culturas em
monocamada. O tratamento das culturas com o agonista A2a DPMA
aumentou a ligação de 3(H)DPCPX e este efeito foi bloqueado
completamente por H89, um inibidor da PKA. A incubação com o
antagonista A1 DPCPX ou com o agonista CHA aumentaram a ligação de 3(H)DPCPX. Do mesmo modo, os tratamentos com adenosina deaminase,
EHNA ou inosina aumentaram a ligação.
Os resultados indicam que a ativação de receptores A2a modula a
expressão de receptores A1 nas culturas. Além disso, os dados também
sugerem a possível participação de receptores A3 ativados por inosina.
xi

ABSTRACT
Adenosine is an important neuromodulator in CNS and its actions are
promoted by activation of A1, A2a, A2b and A3 receptors. Previous studies
from our laboratory have shown that A1 e A2 receptors are present in intact
chick embryo retina. In culture, the detection of A1 receptor depends on
cell aggregation. Monolayer cultures have shown a low or undetectable
binding of A1 agonist 3(H)CCPA or antagonist 3(H)DPCPX. Chronic
treatment with the permeable cyclic AMP analogs SpcAMP or 8 BrcAMP
was able to increase receptor detection. In addition, forskolin also
significantly increased the binding. These results suggest the modulation of
A1 receptor detection via A2a receptor activation.
A2a adenosine receptors are expressed in embryonic retinas starting
at 7 days of development (E7) when the levels of receptor expression are
higher. At E9 the level decreased about 45% and remained constant up to
E16. This receptor was also finder in monolayer cultures. Treatment of
cultures with the A2a agonist DPMA increased 3(H)DPCPX binding and
this effect was completely blocked by H89, a PKA inhibitor. Treatment
with A1 antagonist DPCPX or agonist CHA increased 3(H)DPCPX
binding. In the same way, treatment with adenosine deaminase, EHNA or
inosine increased the binding.
The results indicate that the activation of A2a receptors modulates
the expression of A1 receptors in chick retina cultures. In addition, the
results suggest the participation of A3 receptors activated by inosine.
xii

LISTA DE ABREVIATURAS
8Br AMPc - 8 Bromoadenosina-3’,5’monofosfato cíclico, sal de sódio.
ADA – adenosina deaminase.
Akt – proteína kinase B.
AMP - adenosina 3’-5’ monofosfato.
AMPc - adenosina 3´-5´monofosfato cíclico.
ATP - adenosina 5´-trifosfato.
BME - meio básico de Eagle.
BSA - albumina sérica bovina.
C1 - primeiro dia de cultura.
C6 - sexto dia de cultura.
CAMKII - Proteína kinase II dependente de cálcio e calmodulina.
CCPA - 2-cloro-N6-ciclopentiladenosina.
CHA - ciclohexiladenosina.
CMF - solução sem cálcio e magnésio.
Cpm - contagens por minuto.
CREB – proteína ligante aelemento de resposta à AMPc.
DAG – diacilglicerol.
DMSO - dimetil sulfóxido.
DPCPX – 8- Ciclopentil-1,3- dipropilxantina.
DPMA - N6-[2-(3,5-Dimethoxifenil)-2-(metilfenil)-etil]adenosina.
E7 – sétimo dia de estágio de desenvolvimento embrionário.
E8 - oitavo dia de estágio de desenvolvimento embrionário.
E9 – nono dia de estágio de desenvolvimento embrionário.
E11 – décimo primeiro dia de estágio de desenvolvimento embrionário.
E14 – décimo quarto dia de estágio de desenvolvimento embrionário.
E16 – décimo sexto dia de estágio de desenvolvimento embrionário.
EHNA - eritro-9-(2-hydroxi-3-nonil) adenina.
ERK – proteína kinase ativada por sinais extracelulares.
viii

Go - proteína G sem ação sobre a enzima adenilil- ciclase.
GABA - ácido gama amino butírico.
Gi - proteína G inibitória.
mGlu1 – receptor metabotrópico de glutamato do tipo 1.
mGlu5 – receptor metabotrópico de glutamato do tipo 5.
GMPc- guanosina 3´-5´monofosfato cíclico.
GRK – kinase de receptor acoplado à proteína G.
GRK2 – kinase de receptor acoplado à proteína G do tipo 2.
Gs - proteína G estimulatória.
H89 – {N-(2-((p- Bromocinamil)amino)etil]-5-isoquinolinasulfonamida, 2HCl}.
IB-MECA – iodobenzilmetilcarboxiamidoadenosina.
IP3 – inositol trifosfato.
JNK – proteína kinase c-Jun NH2-terminal.
L-NAME - Nω – nitro-L-arginina metil éster.
MAPK – proteína kinase ativada por mitógeno.
MEK – MAP kinase kinase.
NBMPR - S-(Nitrobenzil)-6-tioinosina.
NECA – 5’ -N-Etilcarboxiamidoadenosina.
NFkB – fator nuclear kB.
NMDA – N-metil-D-aspartato.
NOS- óxido nítrico sintase.
PI3K – fosfatidil inositol 3 kinase.
PIP2 – fosfatidilinositol 4,5-bifosfato.
PLC – fosfolipase C.
PLCβ - fosfolipase C β.
PKA – proteína kinase dependente de AMPc.
PKC – proteína kinase dependente de Ca+2.
P2Y1 – receptores de purinas tipo 2Y1.
Raf – MAP kinase kinase kinase
Ras – subtipo monomérico de proteína ligante de nucleotídeos de guanina (proteína G).
RNAm – ácido ribonucléico mensageiro.
R-PIA – R-fenilisopropiladenosina.
ix

SAH - S-adenosilhomocisteína.
SAPK – proteína kinase ativada por stress.
SCH 58261 – 7-(2-feniletil)-5-amino-2-(2-furil)-pirazolo [4,3-e]-1,2,4-triazolo[1,5-
c]piramida.
SDS - dodecil sulfato de sódio.
SFB – soro fetal bovino.
Sítio P – receptor intracelular de adenosina.
SNC- sistema nervoso central.
SOS – fator de troca de nucleotídeo.
Sp cAMP - Sp-Adenosina-3’5’ monofosforotioato cíclico.
Src- proteína tirosina cinase citoplasmática purificada do sarcoma da retina de pinto
TBS - solução tampão Tris.
TBS-T - solução tampão Tris acrescida de tween-20.
Tris - tris-(hidroximetil)-aminometano.
x

I- INTRODUÇÃO
1) Adenosina
Adenosina é um importante componente do sistema purinérgico e é
encontrada em todos os tecidos apresentando um papel modulatório de
diversos processos fisiológicos. Suas principais ações descritas são:
regulação das funções cardiovasculares como vasodilatação e diminuição
da pressão sanguínea e modulação do sistema nervoso central através da
regulação da liberação de neurotransmissores (Sato et al., 2005; Cunha,
2001 para revisão). Além disso, é capaz de produzir inibição da lipólise,
broncoconstrição e citoproteção em episódios de isquemia e hipóxia ou de
stress oxidativo (Van der Graaf et al., 1999; Fan et al., 2003; Wardas, 2002
para revisão).
1.1) Produção e metabolismo
A principal via de produção de adenosina no meio extracelular vem
da atividade da enzima ecto-5’-nucleotidase que converte 5’-AMP em
adenosina. O 5’-AMP pode vir de duas fontes: da conversão de
nucleotídeos ou pela liberação de AMPc. O AMPc é liberado pelas células
por um transportador e convertido no meio extracelular em 5’-AMP por

uma ecto-fosfodiesterase (Brunton & Mayer, 1979; Henderson & Strauss,
1991).
No meio intracelular, a adenosina pode ser formada através da
quebra de 5’-AMP pela 5’- nucleotidase. Outra via já descrita ocorre por
hidrólise de S-adenosilhomocisteína (SAH) pela S-adenosilhomocisteína
hidrolase (Lloyd et al., 1988). Entretanto, essa via não possui uma grande
importância na produção deste nucleosídeo (Pak et al, 1994).
A adenosina pode ser metabolizada intracelularmente em AMP pela
adenosina kinase ou em inosina pela adenosina deaminase. Já no meio
extracelular ela é metabolizada pela ecto-adenosina deaminase (figura 1)
(Latini & Pedata, 2001 para revisão).

Figura 1: Vias de produção, metabolismo e transporte de adenosina (Latini & Pedata,
2001 para revisão).
1.2) Transporte
A principal via para a regulação dos níveis de adenosina ocorre
através do seu transporte bi-direcional. Os transportadores são classificados
em dois grupos: transportadores equilibrativos que transportam adenosina

de acordo com o seu gradiente de concentração e os transportadores
concentrativos que transportam a adenosina contra o seu gradiente de
concentração e de forma dependente de Na+. Além disso, os
transportadores equilibrativos são divididos em sensíveis a NBMPR
(nitrobenziltioinosina) e insensíveis a NBMPR (Jarvis & Yong, 1986). No
sistema nervoso central foi demonstrado uma grande quantidade de RNAm
para os transportadores equilibrativos, principalmente RNAm de
transportadores sensíveis a NBMPR (Anderson et al., 1999).
O transporte de adenosina tem sido bastante estudado pelo nosso
laboratório usando tanto culturas mistas, como culturas purificadas de
neurônios e culturas purificadas de glia. Todas obtidas de embriões de
galinha.
Nas culturas purificadas de neurônios, cerca de 40% dos neurônios e
praticamente todos os fotorreceptores possuem o mecanismo de transporte
de adenosina. A captação mostra-se saturável e pode ser bloqueada por
NBMPR, além de muito reduzida pelo pré-tratamento da 3(H)adenosina
com adenosina deaminase. A análise do material intracelular pós-captação
indica a presença de 35% de adenosina e 65% de inosina, hipoxantina e
nucleotídeos mais ácido úrico. Além disso, essas células também liberam
adenosina em condições basais e estimuladas por altas concentrações de
K+, de uma maneira dependente de Ca2+ (Paes-de-Carvalho et al., 1990).

Em culturas mistas de embriões de galinha de 8 dias, glutamato
estimula a liberação de purinas e esse efeito é bloqueado por antagonistas
de receptores de NMDA e kainato. Esse estímulo é mimetizado por kainato
e NMDA e bloqueado pelos seus respectivos antagonistas indicando a
participação desses dois receptores na liberação de purinas. Já se observa
liberação após adição de 1 µM de glutamato e a resposta máxima é obtida
após tratamento com 100 µM de glutamato. Experimentos com kainato e
NMDA demonstraram que o estímulo não é capaz de liberar todo o “pool”
de purinas e que esse efeito é reversível. O transporte é bloqueado por
NBMPR e inibidores de CAMKII. Além disso, é dependente de Ca2+ mas
os canais de Ca2+ do tipo L não estão envolvidos. Os resultados indicam
que a ativação de receptores ionotrópicos de glutamato promove uma
liberação de purinas dependente de cálcio que é realizado pelo
transportador equilibrativo sensível ao NBMPR. Analisando-se a natureza
do material intracelular após a captação, encontrou-se 95% de nucleotídeos.
Entretanto, a maior parte do material extracelular liberado após estímulo
com glutamato era inosina (Paes-de-Carvalho et al., 2005).
Estudando-se culturas purificadas de glia obtidas de embriões de
galinha de 11 dias, observou-se que as células eram capazes de captar e
liberar adenosina. Esse transporte é bloqueado por NBMPR (de forma
menos potente quando comparado com as culturas de células mistas) e pelo
pré-tratamento da 3(H)adenosina com adenosina deaminase. O efeito menos

potente do NBMPR nas culturas de glia quando comparado ao seu efeito
nas culturas mistas, sugere a presença dos outros transportadores não
sensíveis a essa droga em maior concentração nessas células. A captação é
bloqueada pelos inibidores de CAMKII e PKC e ligeiramente bloqueada
pelo inibidor da PKA. Já a ativação dos receptores A1 ou A2a de adenosina
não apresenta nenhum efeito na captação. Entretanto, os agonistas desses
receptores induziram um pequeno aumento na liberação de purinas. Além
disso, a própria adenosina é capaz de promover um grande efluxo de
purinas sugerindo um mecanismo de “homoexchange” nessas células de
glia. Ao contrário das culturas mistas, kainato e NMDA não estimulam a
liberação de adenosina nestas culturas de glia excluindo o envolvimento de
receptores de glutamato estimulando o transporte (Ferreira et al., resultados
não publicados).
Esses resultados indicam que tanto as células de glia quanto os
neurônios têm a maquinaria para o transporte, síntese e metabolismo de
adenosina.
1.3) Receptores
A adenosina atua via 4 subtipos de receptores já clonados: A1, A2a,
A2b e A3 (figura 2). Classicamente os receptores de adenosina são
divididos em dois grupos: os receptores A1 e A3 que estão acoplados

negativamente à enzima adenilil ciclase diminuindo os níveis intracelulares
de AMPc, e os receptores A2a e A2b que estão acoplados positivamente à
enzima adenilil ciclase promovendo um aumento dos níveis intracelulares
de AMPc (van Calker et al., 1979; Zhou et al., 1992; Pierce et al., 1992).
Entretanto, esses receptores podem se acoplar a outros tipos de proteína G
ativando diferentes vias de sinalização (Biber et al., 1997; Abracchio et al.,
1995; Palmer et al., 1995; Offermanns & Simon, 1995; Gao et al., 1999).
Figura 2: Receptores de adenosina. Os receptores A1 e A3 de adenosina estão acoplados
às proteínas Gi/Go e os receptores A2 de adenosina estão acoplados à proteína Gs
(http://www.aderis.com/img/art_adenosine.gif).

A presença de receptores A1 e A2a de adenosina foi demonstrada em
retinas de diversas espécies (Blazynski & Perez, 1991 para revisão).
Paes-de-Carvalho e de Mello em 1982 mostraram um acúmulo de
AMPc induzido por adenosina durante o desenvolvimento. Entre 8 e 14
dias, a adenosina não promoveu aumento de AMPc. Após 14 dias, as
retinas passam a promover acúmulo de AMPc tendo como resposta
máxima em 17 e 18 dias. Após a eclosão, a produção foi muito reduzida.
Também foi observado um aumento de AMPc em culturas complexas com
apenas 1 dia tendo um efeito máximo com 4 dias que foi mantido até 8 dias
de cultura.
Em 1985, Paes-de-Carvalho e de Mello mostraram que dopamina e
2-cloroadenosina produziram acúmulo de AMPc em retinas de 16 dias. Em
concentrações saturantes ou sub-saturantes das drogas, os efeitos não foram
somatórios indicando a presença de um receptor inibitório para acúmulo de
AMPc. Esse fato foi demonstrado em retinas de 12 dias onde a adenosina
promoveu inibição do acúmulo de AMPc induzido por dopamina de forma
dose-dependente com uma inibição máxima de 70%.
Alguns anos depois, foi mostrado através da técnica de “binding” a
presença de receptores A1 em retinas intactas de embriões de galinha em
desenvolvimento. Este receptor já se encontra presente com 10 dias de
desenvolvimento do embrião alcançando níveis máximos em 17 dias. Após

a eclosão do ovo, o número de receptores cai, mas tende a se estabilizar
(Paes-de-Carvalho, 1990).
1.3.1) Distribuição dos receptores
O receptor A1 de adenosina possui uma distribuição por todo sistema
nervoso central com grande concentração no córtex, hipocampo e cerebelo.
Já o receptor A2 está mais presente no striatum e no bulbo olfatório (figura
3). O receptor A3 é encontrado em baixas concentrações no hipocampo,
córtex, cerebelo e striatum (Fredholm et al., 2000 para revisão).
Figura 3: Autoradiografia mostrando a presença do
receptor A1 no córtex e hipocampo (em cima) e do
receptor A2 no striatum (em baixo)
(http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=b
nchm.figgrp.1262).

Entretanto, os receptores A1 também são expressos em outras
regiões como glândula adrenal, músculo esquelético, fígado, rins, tecido
adiposo, glândulas salivares, esôfago, pulmão e pâncreas. Os receptores
A2a são encontrados no timo, em leucócitos, em plaquetas, no coração e no
pulmão, enquanto que os receptores A2b estão em mastócitos, tecido
adiposo, glândula adrenal, rins e fígado. Já os receptores A3 encontram-se
presentes nos mastócitos, pulmão, tireóide, glândula adrenal, fígado, rins,
coração e intestino (Fredholm et al., 2001).
1.3.2) Localização celular e subcelular dos receptores no SNC
Os receptores A1 são encontrados tanto em neurônios quanto em
astrócitos, microglia e oligodendrócitos (Biber et al., 1997; Gebicke-
Haerter et al., 1996; Othman et al., 2003). Outros estudos demonstraram
que esses receptores são mais abundantes nas sinapses, tanto nos terminais
pré-sinápticos quanto nos pós-sinápticos (Tetzlaff et al., 1987). Entretanto,
a ativação do receptor A1 e o efeito produzido depende do tipo celular já
que este receptor controla a liberação de glutamato, acetilcolina e
serotonina mas não modula a liberação de GABA e noradrenalina (Cunha,
2001).
Os receptores A2a são encontrados nos neurônios, em astrócitos e
microglia (Li et al., 2001). Nos gânglios da base os receptores estão

localizados predominantemente nas espinhas dendríticas e nas regiões pós-
sinápticas (Hettinger et al., 2001; Rodrigues et al., 2005). Já os receptores
A2a do hipocampo estão mais localizados nas sinapses em particular nas
regiões pré-sinapticas (Rebola et al., 2005). Além disso, ao contrário do
receptor A1, o receptor A2a regula a liberação de glutamato, acetilcolina,
GABA e noradrenalina (Lopes et al., 2002; Rebola et al., 2002; Cunha et
al., 2000; Barraco et al., 1995).
Foi encontrado RNAm para receptor A2b em microglia
(Hammarberg et al., 2003) Além disso, esse receptor foi encontrado em
astrócitos (Allaman et al., 2003). Entretanto, mais estudos são necessários
para caracterizar a presença desse receptor em neurônios.
O receptor A3 é encontrado em neurônios (Brand et al., 2001),
astrócitos (Wittendorp et al., 2004) e microglia (Hammarberg et al., 2003)
1.3.2) Estrutura dos receptores
Os receptores de adenosina são acoplados a proteínas G. Assim
sendo, como todos os receptores metabotrópicos, eles apresentam 7
domínios transmembrana com 3 alças intracelulares e 3 alças
extracelulares. Além disso, apresentam uma cadeia carboxi terminal
intracelular e uma cadeia amino terminal extracelular (Olah et al., 1995
para revisão).

A cadeia carboxi terminal e a terceira alça intracelular possuem um
papel importante, pois elas contêm os sítios de interação do receptor com a
proteína G. Estudos onde o receptor A1 foi mutado demonstraram que o
terceiro e o sétimo domínio transmembrana contêm resíduos de
aminoácidos críticos para a ligação de agonistas e antagonistas. Além
disso, o quinto domínio transmembrana parece estar envolvido no
reconhecimento seletivo de agonistas sintetizados a partir de uma
substituição na posição 5’ da ribose da molécula de adenosina (Olah et al.,
1995 para revisão).
Já foi mostrado que o acoplamento do receptor A2a à proteína G
depende da terceira alça intracelular. Além disso, a porção amino terminal
da terceira alça intracelular é quem determina a seletividade desse receptor
à proteína Gs, enquanto que os resíduos de lisina e ácido glutâmico da
segunda alça intracelular são necessários para que haja um eficiente
acoplamento à proteína G (Olah, 1997).
Outros estudos mostram que o resíduo treonina na posição 298 da
cadeia C terminal é o responsável pela rápida dessensibilização do receptor
A2a (Palmer & Stiles, 1997).
Os receptores de adenosina, exceto o receptor A2a, possuem um
resíduo cisteína na região carboxi terminal que pode servir como um sítio
de palmitoilação envolvido na formação de uma quarta alça intracelular.

Este sítio está envolvido com dessensibilização e internalização de
receptores (Palmer et al., 1996).
1.3.3) Dimerização de receptores
Os receptores A1 e A2a são conhecidos por formarem
homodímeros nas membranas celulares. Além disso, eles são capazes de
formar complexos com outros tipos de receptores podendo assim, em
alguns casos, modificar a atividade deles. Já foi demonstrado que o
receptor A1 forma heterodímero com receptores como o receptor P2Y1
(Yoshioka et al., 2002), receptor metabotrópico de glutamato mGlu1
(Ciruela et al., 2001) e receptor D1 de dopamina (Guinés et al., 2000). Já
o receptor A2a pode formar heterodímero com o receptor metabotrópico
de glutamato mGlu5 (Ferre et al., 2002) e com o receptor D2 de
dopamina (Canals et al., 2003).
2) Receptores A1 e A3 de adenosina
2.1) Sinalização via adenilil ciclase
Os receptores A1 e A3 podem se acoplar às proteínas Gi ou Go
inibindo a atividade da enzima adenilil ciclase (van Calker et al., 1979;

Zhou et al., 1992). Quando esses receptores são ativados por agonistas, eles
ativam as proteínas Gi ou Go que, por serem proteínas inibitórias sensíveis
à toxina pertussis, se ligam à enzima inibindo-a. Assim, a conversão de
ATP em AMPc é inibida e, consequentemente, os níveis intracelulares
desse nucleotídeo cíclico diminuem. Quando a produção desse segundo
mensageiro cai, a kinase que estaria abaixo na via de sinalização, no caso a
PKA, não é ativada (figura 4).
Figura 4: Esquema da sinalização do receptor A1 ou A3 via adenilil ciclase. Quando
ativado, o receptor inibe a atividade da enzima diminuindo a produção de AMPc e,
consequentemente, a sinalização deste segundo mensageiro (modificado de
http://www.starosta.com/technical/art/membrane_receptor.jpg).
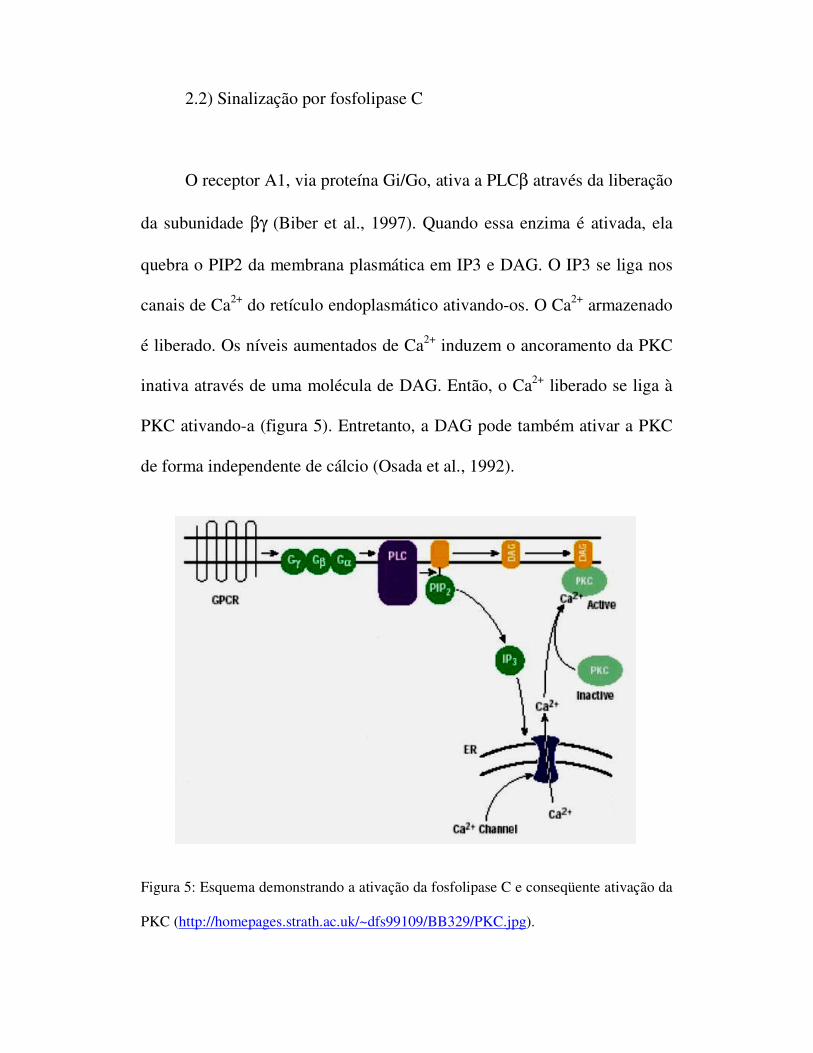
2.2) Sinalização por fosfolipase C
O receptor A1, via proteína Gi/Go, ativa a PLCβ através da liberação
da subunidade βγ (Biber et al., 1997). Quando essa enzima é ativada, ela
quebra o PIP2 da membrana plasmática em IP3 e DAG. O IP3 se liga nos
canais de Ca2+ do retículo endoplasmático ativando-os. O Ca2+ armazenado
é liberado. Os níveis aumentados de Ca2+ induzem o ancoramento da PKC
inativa através de uma molécula de DAG. Então, o Ca2+ liberado se liga à
PKC ativando-a (figura 5). Entretanto, a DAG pode também ativar a PKC
de forma independente de cálcio (Osada et al., 1992).
Figura 5: Esquema demonstrando a ativação da fosfolipase C e conseqüente ativação da
PKC (http://homepages.strath.ac.uk/~dfs99109/BB329/PKC.jpg).

Em 1997, Biber e colaboradores demonstraram que em culturas de
astrócitos o receptor A1 ativa a PLCβ através da subunidade βγ da proteína
G. Entretanto, mais subunidades βγ são necessárias para ativar a PLC do
que subunidades α para inibir a adenilil ciclase. Parece que esse efeito
depende do nível de expressão do receptor nas culturas. Somente níveis
altos do receptor A1 são capazes de estimular a PLC enquanto que a
inibição da adenilil ciclase independe do nível de expressão do receptor.
O receptor A3 também pode ativar a PLC via proteína Gi ou Go
através da liberação da subunidade βγ ou através da proteína Gq
(Abbracchio et al., 1995; Palmer et al., 1995).
2.3) Sinalização através de canais iônicos
Quando o receptor A1 é ativado, ele é capaz de promover a abertura
de canais de K+ aumentando a condutância desse íon (Gerber et al., 1989).
Outro efeito é o fechamento dos canais de Ca2+ do tipo N (Mogul et al.,
1993). O fechamento dos canais de Ca2+ e abertura dos canais de K+
contribuem para a inibição da liberação de neurotransmissores promovido
pelos receptores A1 pré-sinápticos.

2.4) Sinalização via MAP kinases
As proteínas da família das MAPK estão envolvidas com sobrevida,
proliferação e diferenciação em todos os organismos. Essa família consiste
das proteínas kinases reguladas por sinal extracelular (ERK) e das proteínas
kinases ativadas por stress (SAPK) como a P38 e a JNK (Rane et al., 2000
para revisão; Roux & Blenis, 2004 para revisão).
Essas proteínas são kinases serina/treonina normalmente ativadas via
receptores tirosina kinase (Serger & Krebs, 1995). Entretanto, receptores
acoplados a proteína G também são capazes de ativar as MAPK (Luttrell et
al., 1997 para revisão).
Em células COS-7 os receptores A1 de adenosina podem ativar
ERK1/2 através da subunidade βγ da proteína Gi ou Go (Faure et al.,
1994). Essa ativação é tempo e dose dependente (Schulte & Fredholm,
2000) e sensível a PI3 kinase (Dickenson et al., 1998).
Dados do nosso laboratório demonstram que a ativação de receptores
A1 pode estimular a fosforilação das ERK 1/2 em culturas de glia de retina
(Rodrigues et al., dados não publicados).
O receptor A3 pode ativar a via das ERK 1/2 através da subunidade
βγ da proteína Gi ou Go. A sinalização foi mostrada ser dependente de
PI3K, Ras e MEK (Schulte et al., 2002). Mais recentemente, foi
demonstrado que a ativação do receptor A3 inibe a proliferação celular

através da inibição da fosforilação das ERKs em células A375 de
melanoma humano. Nesse caso, a ativação da via PI3K/Akt pelo receptor
A3 leva a uma fosforilação de um sítio inibitório na Raf inativando-a
(Merighi et al., 2005).
2.5) Dessensibilização
O receptor A1 possui uma cinética de dessensibilização e
internalização muito lenta. Ramkumar e colaboradores em 1991 mostraram
que após 8 horas de estímulo com agonista A1, células DDT1 MF-2
apresentaram uma redução de 40% do número de receptores na membrana.
Alguns receptores quando perdem o seu sítio de palmitoilação
passam a apresentar uma cinética de dessensibilização e internalização
mais rápida (Moffett et al., 2001). Entretanto, Ferguson e colaboradores
demonstraram em 2002 que o sítio de palmitoilação do receptor A1 não
interfere no mecanismo de dessensibilização e internalização desse receptor
e que a ativação prolongada leva a uma redistribuição da arrestina-3 em
“clusters”.
Escriche e colaboradores em 2003 demonstraram que a
dessensibilização e internalização dos receptores A1 em células DDT1MF-
2 são mediados por cavéola e não por clatrina. Após 40 minutos de
estímulo com R-PIA, grande parte dos receptores está internalizado em

vesículas. Também foi mostrado que o domínio carboxi terminal do
receptor interage com a caveolina-1.
Palmer e colaboradores em 1995 mostraram que receptores A3 de
rato transfectados em células CHO, possuem uma cinética de fosforilação e
dessensibilização extremamente rápida. Após 4 minutos de exposição a 10
µM de NECA, o receptor A3 alcançou o máximo de fosforilação que se
manteve por 20 minutos na presença do agonista. Esta fosforilação envolve
GRK2. Além disso, eles demonstraram que após 10 minutos na presença de
NECA, os receptores tiveram seus sítios de ligação reduzidos (34 ± 10 %
de redução de Bmax) o que caracteriza uma dessensibilização rápida.
Dados da literatura indicam que ERK 1/2 estaria envolvida na
ativação de GRK2 e, consequentemente, participando do processo de
dessensibilização do receptor (Trincavelli, 2002). O tratamento com
inibidor das ERKs impediu a translocação para a membrana da GRK2 e
bloqueou a dessensibilização e a internalização do receptor.
Ferguson e colaboradores em 2002 mostraram que o sítio de
palmitoilação do receptor A3 é importante para o processo de
dessensibilização e internalização. Quando o sítio de palmitoilação foi
mutado, esse receptor passou a ter uma fosforilação basal provavelmente
por facilitar a ação da GRK. Além disso, a internalização ocorreu em uma
proporção mais rápida quando comparado ao não mutado.

2.6) Principais funções
Já é bem conhecido na literatura que a ativação do receptor A1 inibe
a liberação de neurotransmissores por hiperpolarizar os terminais pré-
sinápticos através da abertura de canais de K+ e pelo fechamento de canais
de Ca2+. Assim, durante episódios de isquemia e hipóxia, a ativação de
receptores A1 leva a um bloqueio da liberação de neurotransmissores,
principalmente o glutamato, promovendo um efeito neuroprotetor para as
células (Sakamoto et al., 2004).
A ativação do receptor A3 produz efeitos neuroprotetores pois foi
mostrado que o tratamento crônico com o agonista seletivo IB-MECA
diminui a morte neuronal ocorrida durante a isquemia (von Lubitz et al.,
1994). Esse efeito protetor também foi observado em culturas de miócitos
(Liu et al., 1994).
O receptor A3 também está envolvido em processos inflamatórios. O
tratamento com inosina estimula, via receptor A3, a degranulação de
mastócitos (Jin et al., 1997).

3) Receptores A2a e A2b de adenosina
3.1)Sinalização via adenilil ciclase
Os receptores A2a e A2b de adenosina estão acoplados a proteínas
Gs estimulando a produção de AMPc (van Calker, 1979). Quando esses
receptores são ativados, ocorre a ativação da enzima adenilil ciclase e
consequentemente aumento dos níveis intracelulares de AMPc. Esse
segundo mensageiro ativa a PKA que pode migrar para o núcleo da célula
ativando transcrição gênica via CREB (Gonzalez et al., 1989) (figura 6).
Figura 6: Receptor A2 ativa a enzima adenilil ciclase aumentando os níveis
intracelulares de AMPc o qual ativa PKA (modificado de
http://www.starosta.com/technical/art/membrane_receptor.jpg).

3.2) Sinalização através de fosfolipase C
Dados da literatura demonstram que receptores A2a são capazes de
ativar PLCβ quando transfectados em células COS-7 com Gα15 e Gα16.
Esse efeito não foi observado com Gαq, Gα11 ou Gα14 (Offermanns &
Simon, 1995).
O receptor A2b ativa a PLCβ via proteína Gq (Gao et al., 1999).
Entretanto, foi demonstrado que em células HEL (células de eritroleucemia
humana) a ativação do receptor A2b aumenta os níveis intracelulares de
Ca2+ de forma independente de PLC. Esse efeito é dependente de Gs que
poderia estar ativando diretamente canais de cálcio já que este efeito foi
demonstrado ser independente de AMPc (Feoktistov et al., 1994).
Nos neurônios piramidais da região CA3 do hipocampo, o receptor
A2b ativa canais de Ca2+ do tipo P mas não do tipo N. Esse mecanismo tem
sido proposto ser via ativação da adenilil ciclase já que inibidores de PKA
bloqueiam esse efeito (Feoktistov et al., 1994).
3.3) Sinalização pelas MAP kinases
Em células endoteliais, foi mostrado que a ativação de ERK 1/2 via
receptor A2a era independente de proteína Gs, AMPc, PKC e cálcio mas
dependente de Ras e MEK. Os autores sugeriram que o efeito poderia ser

via proteína G12/13 (Sexl et al., 1997). Em células HEK 293, a ativação
também é independente de Gs e AMPc mas envolve Ras e SOS. Entretanto,
em células CHO transfectadas com o receptor A2a e em células PC12, a
fosforilação é dependente de AMPc, PKA e Src (Seidel et al., 1999).
Estudos em mastócitos humanos demonstraram que o receptor A2b
ativa as três famílias de MAP kinases mas com cinéticas diferentes. A
fosforilação máxima da ERK 1/2 foi alcançada rapidamente e foi
sustentada por 30 minutos enquanto que a fosforilação da P38 foi rápida e
transiente (alcançou o nível máximo em 1 minuto e decaiu após 10
minutos). Já a fosforilação da JNK foi alcançada apenas após 10 minutos
de estímulo e voltou ao basal após 30 minutos (Feoktistov et al., 1999).
Essas cinéticas diferentes indicam que cada uma das fosforilações ocorre
por vias diferentes.
Em células HEK 293 foi mostrado que os níveis máximos de
fosforilação da ERK 1/2 são alcançados 5 minutos após estimulação com
NECA e que depende de PLC, Ras e MEK mas não de PKC (Gao et al.,
1999). Entretanto, outros autores demonstraram que a ativação das ERKs
em células CHO transfectadas com o receptor A2b era dependente de
AMPc e PI3K mas independente de PKA (Schulte & Fredholm, 2000).
Além disso, a ativação da P38 é dependente de AMPc e PKA mas
independente de PI3K.

3.4) Dessensibilização
Willets e colaboradores em 1999 demonstraram que a
desensibilização do receptor A2a é rápida e dependente de GRK2.
O receptor A2b também possui uma cinética de dessensibilização e
internalização rápida. Em células CHO transfectadas com o receptor, 50%
de dessensibilização foi obtida após uma hora de exposição ao agonista.
Além disso, a internalização se mostrou dependente de arrestina. A porção
carboxi terminal parece ser importante já que a sua deleção bloqueia tanto a
dessensibilização quanto a internalização (Matharu et al., 2001).
3.3) Principais funções
Vários estudos demonstram que o bloqueio do receptor A2a promove
a inibição da liberação de neurotransmissores principalmente o glutamato.
Esse fato é importante durante episódios de isquemia e hipóxia já que o
bloqueio do receptor A2a inibe a liberação de glutamato e confere proteção
às células (Latini et al., 1999).
Ferreira e Paes-de-Carvalho em 2001 mostraram que glutamato
promove morte celular em culturas purificadas de neurônios de retina. Esse
efeito é bloqueado por antagonistas de receptores de glutamato do tipo
AMPA/kainato e NMDA. Entretanto, a pré-incubação das culturas por 24

horas com adenosina é capaz de bloquear a morte induzida por glutamato.
Este efeito é mimetizado por agonista de receptor A2a mas não de A1.
Adenosina em presença de EHNA (inibidor de adenosina deaminase),
NBMPR e análogos permeáveis de AMPc como 8BrAMPc e SPAMPc
também bloqueiam a morte celular.
Nestas mesmas culturas foi demonstrado que a troca do meio causa
morte neuronal. Esse efeito é também bloqueado pela pré-incubação com
adenosina, NBMPR e agonista de receptor A2a, mas não por agonista de
receptor A1 (Paes-de-Carvalho et al., 2003).
Os receptores A2b participam do controle do tônus vascular. Dados
da literatura indicam que a ativação do receptor provoca vasodilatação
(Haynes et al., 1995; Rubino et al., 1995). Esse efeito parece ser
independente da produção de NO já que inibidor da NO sintase (L-NAME)
não produziu bloqueio. Entretanto, em artérias renais de ratos, o receptor
A2b também produziu vasodilatação de maneira dependente da produção
de NO (Martin & Potts, 1994).
Dados da literatura indicam que o receptor A2b pode estar envolvido
com aumento da contratilidade do miocárdio (Liang & Haltiwanger, 1995).
Entretanto, mais estudos necessitam ser feitos nessa área.

4) Modelo de estudo: a retina
A retina apresenta uma organização das células em camada. Ela
consiste da camada das células ganglionares que contém os corpos
celulares das células ganglionares. Em seguida aparece a camada
plexiforme interna que contém as sinapses das células ganglionares com as
células bipolares, amácrinas e horizontais. Logo abaixo, está a camada
nuclear interna que contém os corpos celulares das células bipolares,
horizontais e amácrinas. Depois encontra-se a camada plexiforme externa
que contém as sinapses entre as células da camada nuclear interna com os
fotoreceptores localizados na camada nuclear externa. Ao longo de todas as
camadas encontram-se a células da glia de Müller (figura 7).

Figura 7: Esquema mostrando a disposição da retina em camadas (http://wbmo.mpimf-
heidelberg.mpg.de/~teuler/Pics/retina1.jpg).
A retina é um modelo muito usado para estudos neuroquímicos. Ela
contém todos os sistemas de neurotransmissores e transdução de sinais
encontrados em outras partes do sistema nervoso central. Além disso, ela é
de fácil obtenção e é possível fazer culturas usando embriões de galinha.
No nosso laboratório, fazemos três tipos de culturas a partir de embriões de

galinha: culturas mistas (contendo neurônios e células de glia), culturas
purificadas de neurônios e culturas purificadas de glia.
Assim, neste trabalho, nós utilizamos culturas mistas para estudar a
presença dos receptores A1 e a modulação de sua detecção pelos receptores
A2a. Além disso, demonstramos a expressão dos receptores A2a durante o
desenvolvimento e em culturas em monocamada de retinas de embriões de
pinto.

II- OBJETIVOS
Dados anteriores do nosso laboratório mostraram a detecção de
receptores A1 em retinas intactas durante o desenvolvimento de embriões
de galinha (Paes-de-Carvalho, 1990). Entretanto, em culturas de retinas não
foi demonstrado se esses receptores estão presentes ou não e como a sua
expressão pode ser regulada. Apesar do acúmulo de AMPc induzido por
dopamina poder ser inibido por altas concentrações de adenosina nas
culturas através da interação com os chamados sítios P intracelulares, esta
inibição não pôde ser observada com os agonistas de receptores A1 (Paes-
de-Carvalho, 1987, tese).
Em relação ao receptor A2a, resultados prévios mostram que
agonistas de receptores A2a promovem acúmulo de AMPc em retinas
intactas. O acúmulo aumenta com o desenvolvimento do embrião atingindo
um máximo em 17 dias. Resultados similares foram obtidos em culturas de
retina (Paes-de-Carvalho & de Mello, 1982). Entretanto, não se tem
nenhuma informação sobre a expressão desses receptores durante o
desenvolvimento e em culturas de retinas de embrião de galinha.
Assim, os objetivos do nosso trabalho foram:
1- caracterizar a presença dos receptores A1 de adenosina em culturas
complexas;

2- verificar se a ativação de receptores A2a de adenosina podem regular
a detecção do receptor A1 nas culturas;
3- estudar a expressão dos receptores A2a de adenosina durante o
desenvolvimento e em culturas complexas de embrião de galinha.

III- MATERIAIS E MÉTODOS
3.1) Materiais
Ovos de galinha fertilizados da espécie White Leghorn foram obtidos
de um aviário local. Adenosina deaminase foi obtida da Calbiochem
(California, Estados Unidos). 8 Bromoadenosina-3’,5’monofosfato cíclico,
sal de sódio (8BrAMPc), Sp-Adenosina-3’5’monofosforotioato cíclico
(SpAMPc), forskolina e N-[2-(p-bromocinamilamino)etil]-5-
isoquinolinesulfonamida.2HCl (H89) foram obtidos da Biomol Res. Labs.
(Pensilvânia, Estados Unidos). N6-ciclohexiladenosina (CHA), 8-
ciclopentil-1,3-dipropilxantina (DPCPX), N6-[2-(3,5-dimetoxifenil)-2-(2-
metilfenil)-etil]adenosina (DPMA), inosina, eritro-9-(2-hidroxi-3-
nonil)adenina-HCl (EHNA), penicilina, streptomicina, glutamina, albumina
sérica bovina (BSA) e dimetil sulfóxido (DMSO) foram obtidos da
Sigma/RBI Chem.Co. (Missouri, Estados Unidos). 5-amino-7-(2-feniletil)-
2-(2-furil)-pirazolo[4,3-e]-1,2,4-triazolo[1,5-c]pirimidina (SCH 58261), foi
uma gentileza de Rodrigo A. Cunha (Universidade de Coimbra, Coimbra,
Portugal) cujo composto foi doado por Scott Weiss (Vernalis, Inglaterra).
O anticorpo anti-A2a e peptídeo controle anti-A2a foram obtidos da
Chemicon International (California, Estados Unidos). Os anticorpos anti-
actina e o secundário anti-cabra foram doados pela professora Elizabeth

Giestal de Araujo (Universidade Federal Fluminense, Niterói) foram
obtidos da Santa Cruz (California, Estados Unidos). [propil-3H]8-
ciclopentil-1,3-dipropil-xantina ([3H]DPCPX), atividade específica de 130
Ci/mmol, anticorpo secundário anti-rabbit, glicina, SDS, Tris, acrilamida
PAGE, metilenobisacrilamida, mercaptoetanol, hyperfilm e kit ECL foram
obtidos da Amersham Pharmacia Biotech UK Limited (Buckinghamshire,
Inglaterra). [3H]-2-cloro-N6-ciclopentiladenosina ([3H]CCPA), de atividade
específica de 30 – 60 Ci/mmol, foi obtido da PerkinElmer (Boston, Estados
Unidos). BME, tripsina e soro fetal bovino (SFB) foram obtidos da GIBCO
(Nova York, Estados Unidos). Os outros reagentes utilizados eram de
pureza analítica.
3.2) Métodos
3.2.1) Culturas complexas
Retinas de embriões de 8 dias foram dissecadas em CMF (salina sem
Ca2+ e Mg2+) e incubadas por 15 minutos em 0,1% de tripsina a 37oC. Em
seguida, a solução foi retirada e as retinas dissociadas em pipeta Pasteur
com BME contendo 3% de soro fetal bovino (SFB), 100 U/mL de
penicilina, 100 µg/mL de streptomicina e 2mM de glutamina. As células
foram plaqueadas em placas de 24 poços de 16 mm de diâmetro ou em

placas de 40 mm de diâmetro na densidade de 5 x 106 células por mL. As
culturas foram incubadas a 37oC por 6 dias em atmosfera de 5% de CO2 e
95% de ar. O meio de cultura foi trocado no dia seguinte ao preparo da
cultura e depois a cada 48 horas.
3.2.2) Culturas de agregados
Retinas de embriões de 8 dias foram dissecadas em CMF e
tripsinizadas por 15 minutos em 0,1% de tripsina a 37oC. Após a retirada da
solução, as retinas foram dissociadas em pipeta Pasteur com BME
contendo 3% SFB. Então, as células foram adicionadas em frasco de
Erlenmeyer e mantidas em agitação no incubador orbital a 37oC em
ambiente de 5% de CO2 e 95% de ar por 6 dias. A troca do meio foi feita
no dia seguinte ao preparo e a cada 48 horas. Metade do volume de meio
contido no frasco de Erlenmeyer era retirado e se acrescentava o mesmo
volume de meio fresco.
3.2.3) Tratamento com drogas
Os tratamentos ocorreram em C1 e foram mantidos até C6 no caso de
drogas como SpAMPc (20 µM), 8BrAMPc (100 µM), forskolina (5 µM),
DPMA (100 nM), SCH (100 nM), H89 (5 µM), DPCPX (100 nM), CHA

(100 nM). Além disso, SCH ou H89 foram adicionados 30 minutos antes
da adição do DPMA aos poços.
No caso de drogas como ADA (0,1 U/mL), EHNA (10 µM) e inosina
(10 µM) os tratamentos ocorreram em C5 e foram mantidos até C6.
3.2.4) “Binding” em célula viva
Foram feitas incubações no volume final de 250 µL por poço de 16
mm de diâmetro a 37oC por 1 hora. No sexto dia de cultura, o meio foi
retirado e cada poço lavado 2 vezes com 400 µL de Hanks’. Em seguida,
todos os poços foram incubados com Hanks’ + adenosina deaminase 1
U/mL por 10 minutos a 37oC. Após isso, CHA (1mM) foi adicionado a
alguns poços para a determinação da ligação inespecífica por mais 10
minutos a 37 oC. Logo após, foi adicionado 3(H)DPCPX (50 nM) em todos
os poços e as células incubadas por 1 hora a 37oC. Terminada a incubação,
a solução foi retirada, cada poço lavado com Hanks’ e, em seguida, foi
adicionado água Mili-Q para a lise das células. A placa foi congelada por
24 horas. O material lisado foi colocado em filtros que foram secos em
estufa por 30 minutos a 100oC. Os filtros secos foram colocados nos vials
contendo líquido de cintilação e levados para contagem no cintilador. O

resultado foi expresso em cpm (cintilações por minutos). A proteína
contida na placa foi dosada pelo método de Lowry et al., 1951.
3.2.5) Dosagem de proteína pelo método de Bradford
As amostras de proteína de retinas intactas de embriões de 7, 9, 11,
14 e 16 dias ou de culturas complexas de embriões de 8 dias foram
recolhidas em tampão de amostra [SDS (0,3g), glicerol (1mL), 2-
mercaptoetanol (0,1mL) tris (0,5M, pH 6,8-1,25mL) e H2O (10mL)] e a
dosagem de proteína foi realizada de acordo com o método de Bradford.
Foram colocados em tubos de vidros, 2µL das amostras e, em seguida,
foram acrescentados 3mL de reagente de Bradford. Para realização da
curva padrão, foram adicionados nos tubos de vidro 0, 2, 4, 6 e 10 µg de
BSA (1mg/mL). Em seguida, foram adicionados 2µL do tampão de
amostra SDS (usado nos experimentos) e ainda 3mL de reagente de
Bradford. Posteriormente, as misturas foram agitadas em vortex e a leitura
da absorbância foi realizada no comprimento de onda de 595nm (Bradford,
1976).

3.2.6) Eletroforese em SDS-Poliacrilamida SDS-(PAGE)
Depois de 6 dias em placas de 40 mm , o meio de cultura das células
foi removido e as células lavadas com solução salina Hanks’ duas vezes.
Uma solução tampão SDS-PAGE foi adicionada a cada placa. As células
foram retiradas da placa, fervidas por 5 minutos a 1000C e estocadas a -
200C, a fim de solubilizar e desnaturar todas as proteínas celulares. No caso
das retinas intactas, os embriões foram dissecados em CMF e as retinas
lavadas em Hanks’ para a retirada da salina. Em seguida, foram
adicionadas em solução tampão SDS-PAGE, fervidas por 5 minutos a
100ºC e estocadas a -20ºC. Após a dosagem da proteína, foi aplicado no
topo do gel de poliacrilamida-SDS a 10%, volume de amostra equivalente a
60µg de proteína. O procedimento para a eletroforese em gel de
poliacrilamida foi baseado na técnica descrita por Laemmli et al., 1970. O
gel de separação, foi preparado para um volume de 9 ml e uma
concentração final de acrilamida de 10g%. O gel de concentração foi
preparado para um volume de 3 ml e uma concentração final de acrilamida
de 4,5g%. O tampão de corrida utilizado (1000 ml) tinha em sua
composição Tris (25 mM), glicina (192 mM) e SDS (0,1 g%). Durante a
corrida a amperagem foi mantida constante em 10 mA por 3 horas.

3.2.7) “Western Blotting”
O gel foi retirado da placa onde ocorreu a polimerização e em
seguida foi posto em contato com uma membrana de PVDF para onde as
proteínas eram transferidas. A transferência das proteínas para membranas
PVDF foi baseada na técnica descrita por Towbin et al., 1979.
Imediatamente após a corrida, o gel foi incubado em tampão de
transferência junto com a membrana por 30 minutos (Hybond-P,
Amersham), após esta ter sido hidratada por 10 segundos em metanol a
100%. Cinco folhas de papel de filtro, nas dimensões do gel a ser
transferido, foram também incubadas em tampão de transferência (solução
contendo Tris, 25 mM; glicina, 192 mM; metanol, 10% (V/V)). Em
seguida, o sistema de transferência foi montado de acordo com as
instruções do fabricante. Sobre a grade do aparelho foram colocados, nesta
ordem: 2 folhas de papel de filtro, membrana de PVDF, gel, 2 folhas de
papel de filtro. Durante a montagem, tinha-se o cuidado de remover todas
as bolhas entre os componentes do sistema. A transferência foi realizada a
4ºC durante 1 hora em voltagem constante de 45 V. Após a transferência,
as membranas foram mantidas a 4ºC em TBS (Tris, 50mM; NaCl,150mM)
até a incubação com anticorpos específicos.
A membrana de PVDF foi incubada inicialmente em solução tampão
TBS-T (TBS contendo 0,1% de Tween 20) contendo leite em pó desnatado,

5 g%, durante 1 hora e 30 minutos à temperatura ambiente, com agitação
suave. Com este procedimento era prevenida a ligação de anticorpos a
sítios não específicos na membrana de PVDF. Após este período, a
membrana foi lavada uma vez por cinco minutos com tampão TBS-T. Em
seguida, a membrana foi incubada “overnight” a 4ºC com o anticorpo anti-
A2a (anticorpo policlonal anti-rato feito em coelho) diluído na proporção
de 1:2500. Todos os anticorpos foram diluídos em solução tampão TBS-T
contendo leite em pó desnatado (5g%).
A membrana, previamente incubada com anticorpo primário foi
lavada em solução TBS-T três vezes por 10 minutos e então incubada com
o anticorpo secundário (anti-coelho diluído na proporção de 1:3000 em
solução TBS-T, conjugados à peroxidase, contendo leite em pó desnatado
5g%) por uma hora, à temperatura ambiente. Em seguida, as membranas
eram lavadas da seguinte maneira: 2 lavagens com TBS-T por 10 minutos e
por último uma lavagem com TBS apenas por mais 10 minutos.
A imunodetecção foi promovida pela reação quimioluminescente do
sistema ECL para western blotting. Após a incubação da membrana com
uma mistura de soluções de detecção por 4 minutos a temperatura
ambiente, o excesso do reagente foi retirado com cuidado e a membrana
era, então, envolvida em filme de PVC e exposta imediatamente a filme
Hiperfilm, por curtos intervalos de tempo, que era revelado para obtenção
de uma imagem autorradiográfica adequada.

3.2.8) Retirada dos anticorpos e “blotting” para actina
Após a revelação as membranas foram lavadas em TBS. Em seguida,
foram incubadas com glicina 0,2M pH=2,2 e deixadas em agitação
constante em “shaker” para a retirada da ligação dos anticorpos primário e
secundário. Depois, elas foram lavadas em TBS e incubadas em solução
tampão TBS-T contendo leite em pó desnatado, 5 g%, durante 1 hora e 30
minutos à temperatura ambiente para bloquear as ligações inespecíficas.
Após o bloqueio, as membranas foram incubadas com o anticorpo primário
anti-actina na diluição de 1:300 (anticorpo policlonal anti-coelho feito em
cabra) “overnight”. Depois, as membranas foram lavadas em TBS-T e
incubadas com o anticorpo secundário conjugado a peroxidase na
proporção de 1:5000 (anticorpo policlonal anti-cabra) por 1 hora. Logo
após, as membranas foram lavadas 2 vezes com TBS-T por 10 minutos e 1
vez em TBS por 10 minutos. Em seguida, as membranas eram incubadas
com ECL e reveladas do mesmo modo já descrito.

3.2.9) Análises Estatísticas
As análises estatísticas foram realizadas por ANOVA seguido pelo
teste de Newman-Keuls usando o software Graph Pad Prism. Os resultados
foram expressos como média ± EPM (erro padrão da média) para n ≥ 3 ou
como média ± desvio da média para n = 2.

IV- RESULTADOS
4.1) Ligação de 3(H)CCPA ao receptor A1 em culturas complexas em
monocamada e em culturas de agregados.
Resultados anteriores do laboratório mostram que em culturas
complexas em monocamada que apresentam as células bem dissociadas e
sem grumos de células (figura 8A), a ligação de 3(H)CCPA (agonista de
receptor A1 de adenosina) aos receptores A1 é muito baixa ou nula.
Entretanto, quando as culturas em monocamada apresentam células pouco
dissociadas e com muitos grumos de células (figura 8B), a ligação do
3(H)CCPA é bem expressiva (Hang & Paes-de-Carvalho, resultados não
publicados). Esses resultados sugerem que a ligação do receptor A1
depende do estado de agregação das células em cultura.

A) B)
Figura 8: A) Culturas complexas em monocamada apresentando células bem
dissociadas (cultura não agregada) onde a ligação do 3(H)CCPA é baixa. B) Culturas
complexas em monocamada apresentando células não dissociadas (cultura agregada)
onde a ligação do 3(H)CCPA é alta. A barra representa 100 µm (A) e 200 µm (B).
Para confirmar esses resultados foram feitos experimentos de
“binding” em culturas complexas em monocamada não agregadas e em
culturas de agregados. Nas culturas de agregados o meio é mantido sob
agitação constante. Assim, as células tendem a se associar novamente
formando pequenos grumos de células em camadas de forma muito
semelhante às camadas da retina. Os resultados obtidos mostram que a
ligação do 3(H)CCPA ao receptor A1 é maior nas culturas de agregados do
que nas culturas em monocamada (figura 9).

0
10
20
30
40
**Lig
açã
o3(H
)CC
PA
(fm
ole
s/m
g p
tn)
Monocamada Agregado
Figura 9: Ligação de 3(H)CCPA ao receptor A1 em culturas complexas em
monocamada ou de agregado de embriões de 8 dias. As culturas em C6 foram incubadas
com 10 nM de 3(H)CCPA na presença ou ausência de 100 µM de CHA. Os resultados
representam a média ± EPM de 3 experimentos realizados em duplicatas (p < 0,01).
4.2) O tratamento das culturas complexas em monocamada com análogos
permeáveis de AMPc aumenta a ligação do 3(H)DPCPX ao receptor A1.
Em experimentos subseqüentes utilizamos o antagonista 3(H)DPCPX
para estudar a expressão dos receptores A1 nas culturas.
Para verificar quais os possíveis mecanismos de regulação da
expressão do receptor A1, incubamos as culturas em monocamada com
análogos permeáveis de AMPc (SpAMPc ou 8BrAMPc). Tanto o SpAMPc

(figura 10) quanto o 8BrAMPc (figura 11) foram capazes de aumentar a
ligação do 3(H)DPCPX ao receptor A1 em experimentos de “binding” em
célula viva. Pode-se observar uma certa variação dos níveis controle de
“binding”, provavelmente devido a pequenas variações no estado de
agregação das células nas culturas. No entanto, os análogos de AMPc
invariavelmente produziam aumento marcante da detecção dos receptores.
Resultados obtidos em experimentos de “binding” em
homogeneizados de células tratadas com SpAMPc ou 8BrAMPc foram
muito similares aos de célula viva (resultados não mostrados).
Além disso, o tratamento com forskolina que ativa diretamente a
enzima adenilil ciclase também foi capaz de aumentar a ligação do receptor
em experimentos de “binding” em célula viva (figura 12). Assim, a via
desse segundo mensageiro parece estar envolvida na regulação da
expressão do receptor A1.
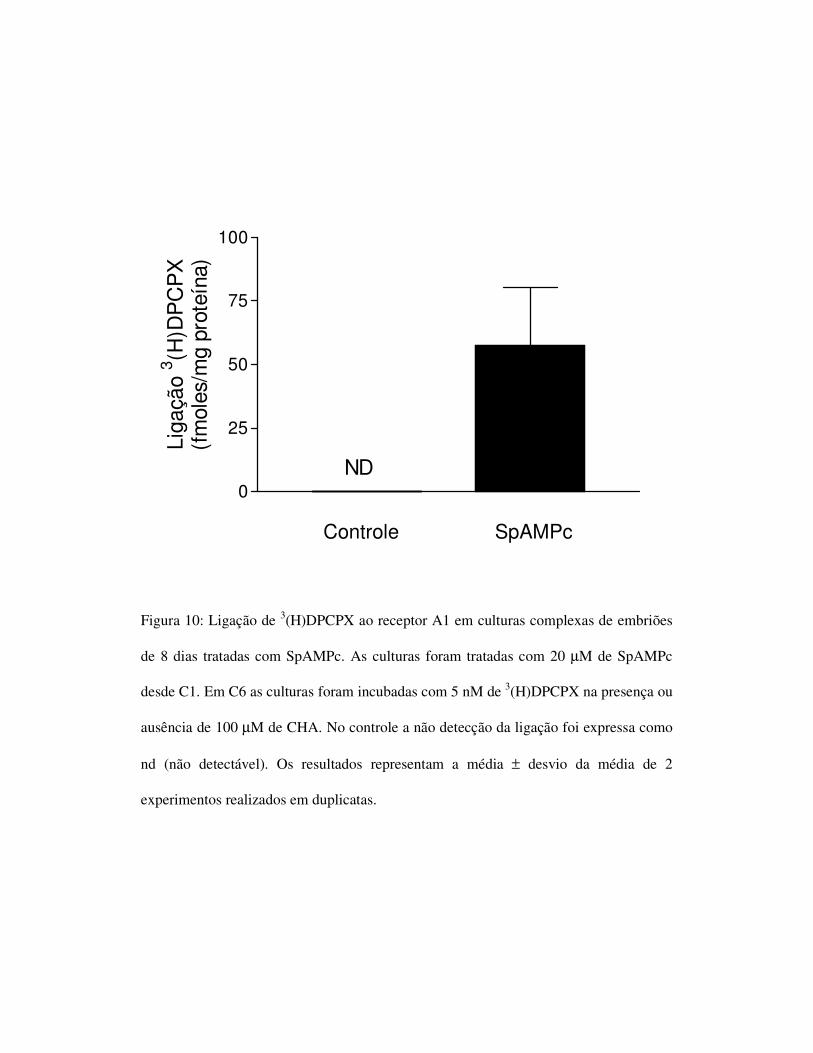
Controle SpAMPc
Figura 10: Ligação de 3(H)DPCPX ao receptor A1 em culturas complexas de embriões
de 8 dias tratadas com SpAMPc. As culturas foram tratadas com 20 µM de SpAMPc
desde C1. Em C6 as culturas foram incubadas com 5 nM de 3(H)DPCPX na presença ou
ausência de 100 µM de CHA. No controle a não detecção da ligação foi expressa como
nd (não detectável). Os resultados representam a média ± desvio da média de 2
experimentos realizados em duplicatas.
0
25
50
75
100
ND
Lig
açã
o3(H
)DP
CP
X(f
mole
s/m
g p
rote
ína)
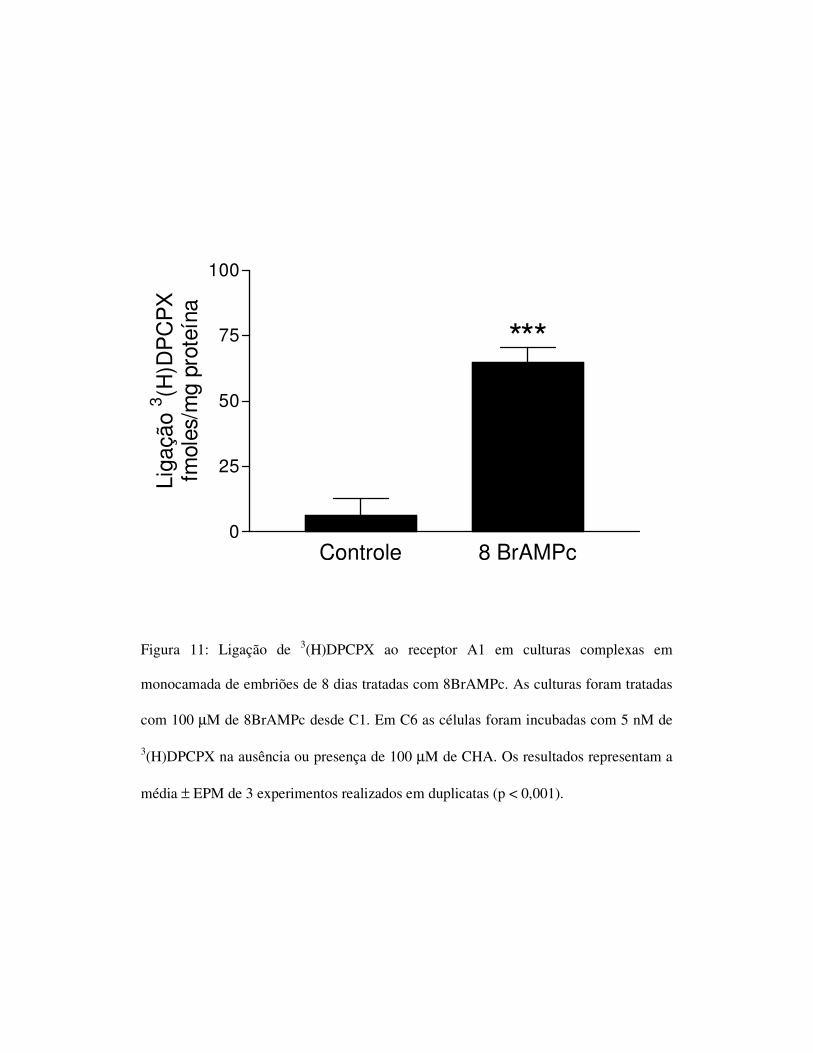
Controle 8 BrAMPc0
25
50
75
100
***
Lig
açã
o3(H
)DP
CP
Xfm
ole
s/m
g p
rote
ína
Figura 11: Ligação de 3(H)DPCPX ao receptor A1 em culturas complexas em
monocamada de embriões de 8 dias tratadas com 8BrAMPc. As culturas foram tratadas
com 100 µM de 8BrAMPc desde C1. Em C6 as células foram incubadas com 5 nM de
3(H)DPCPX na ausência ou presença de 100 µM de CHA. Os resultados representam a
média ± EPM de 3 experimentos realizados em duplicatas (p < 0,001).
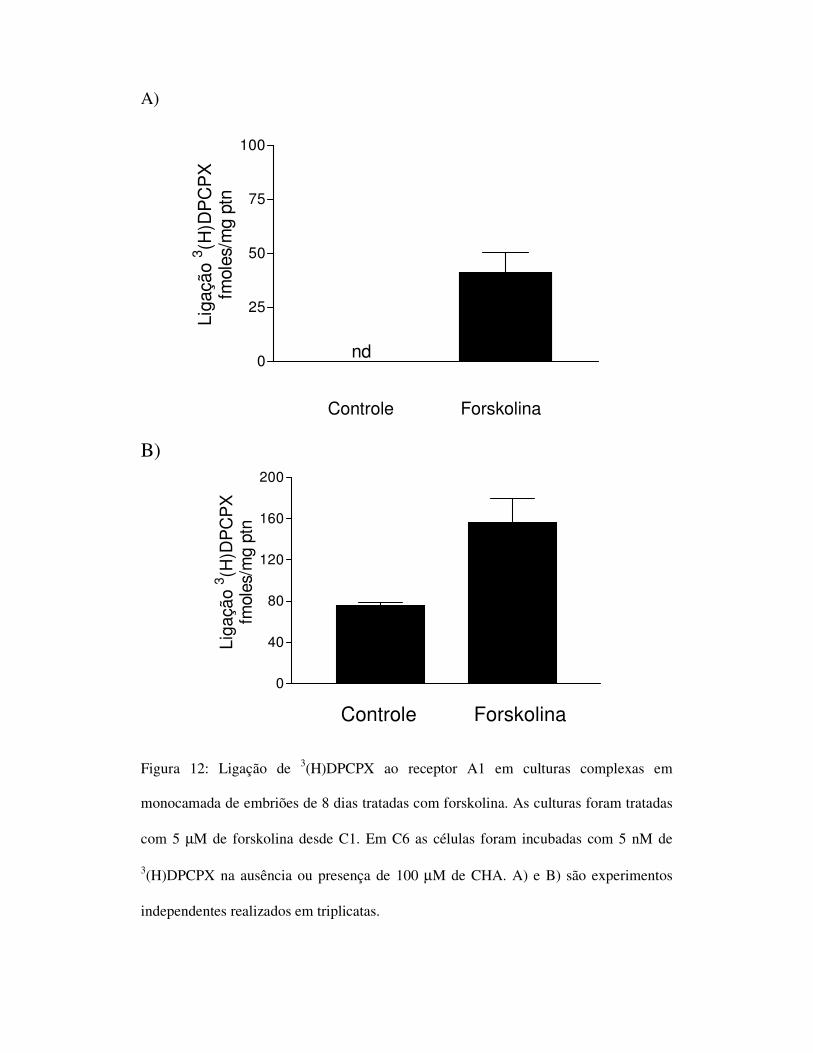
A)
0
25
50
75
100
Lig
açã
o3(H
)DP
CP
Xfm
ole
s/m
g p
tn
nd
Controle Forskolina
B)
0
40
80
120
160
200
Lig
açã
o3(H
)DP
CP
Xfm
ole
s/m
g p
tn
Controle Forskolina
Figura 12: Ligação de 3(H)DPCPX ao receptor A1 em culturas complexas em
monocamada de embriões de 8 dias tratadas com forskolina. As culturas foram tratadas
com 5 µM de forskolina desde C1. Em C6 as células foram incubadas com 5 nM de
3(H)DPCPX na ausência ou presença de 100 µM de CHA. A) e B) são experimentos
independentes realizados em triplicatas.

4.3) A ativação do receptor A2a é capaz de aumentar a ligação do
3(H)DPCPX ao receptor A1.
Já que o tratamento com análogos permeáveis de AMPc aumentaram
a ligação do 3(H)DPCPX, o receptor A2a também poderia regular a
detecção do receptor A1 pois a sua ativação leva ao aumento do níveis
intracelulares de AMPc.
Resultados prévios do laboratório demonstraram que a ativação do
receptor A2a era capaz de aumentar os níveis intracelulares de AMPc tanto
em retina intacta quanto em culturas complexas em monocamada (Paes-de-
Carvalho & de Mello, 1982). Além disso, resultados obtidos com a técnica
de Western Blot também mostram a presença da proteína do receptor A2a
tanto em retina intacta de embrião de galinha quanto nas culturas
complexas (figura 13). Durante o desenvolvimento os níveis de expressão
do receptor A2a estão mais altos em E7. Entretanto, em E9 os níveis estão
mais baixos permanecendo constantes até E14 e diminuindo em E16.

A)
E7 E9 E11 E14 E16
B)
E7 E9 E11 E14 E16
C)
Figura 13: A) Imunobloting para receptor A2a em retina intacta durante o
desenvolvimento. A figura representa o resultado de um experimento separado de dois
que apresentam o mesmo resultado. B) Quantificação do imunobloting representado em
A para receptor A2a em razão da actina. C) Imunobloting para receptor A2a em cultura
complexa de 6 dias. A figura representa o resultado de um experimento separado de
dois que apresentam o mesmo resultado.
A2a (55 KD) →
Actina (43 KD) →
0.0
0.4
0.8
1.2
Razã
o A
2A/A
ctina
(DO
)
A2a (55 KD) →

Para confirmar a especificidade do anticorpo anti-A2a, usamos um
peptídeo controle contra o anticorpo usado. Como resultado, o peptídeo
controle inibiu a ligação do anticorpo ao receptor A2a tanto na amostra de
rato (controle positivo do experimento) quanto na amostra de embrião de
pinto (figura 14).
A) B)
Rato (P70) Pinto (E7) Rato(P70) Pinto (E7)
Figura 14: Imunobloting para receptor A2a. A) Imunobloting de amostras de colículo de
rato de 70 dias (P70) e embrião de pinto de 7 dias. B) Bloqueio da marcação com o
peptídeo controle (diluição 1:2500) contra o anticorpo anti-A2a. O peptídeo controle foi
pré-incubado durante 30 minutos antes da incubação com o anticorpo anti-A2a.
Esses resultados demonstram a presença do receptor A2a e o seu
acoplamento à proteína Gs levando a um aumento dos níveis intracelulares
de AMPc no nosso modelo de estudo e suportam a hipótese de que a
ativação desse receptor poderia regular a expressão do receptor A1 nas
nossas culturas.
A2a (55 KD) →
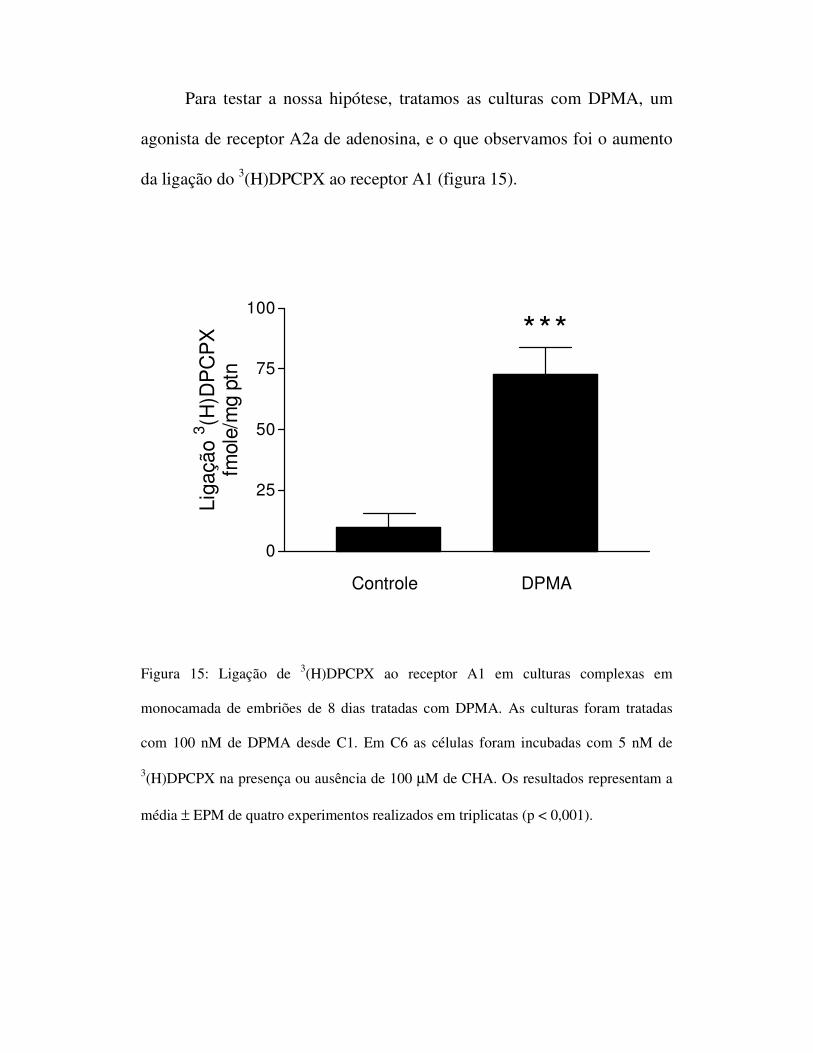
Para testar a nossa hipótese, tratamos as culturas com DPMA, um
agonista de receptor A2a de adenosina, e o que observamos foi o aumento
da ligação do 3(H)DPCPX ao receptor A1 (figura 15).
0
25
50
75
100
**
Lig
açã
o3(H
)DP
CP
Xfm
ole
/mg p
tn
*
Controle DPMA
Figura 15: Ligação de 3(H)DPCPX ao receptor A1 em culturas complexas em
monocamada de embriões de 8 dias tratadas com DPMA. As culturas foram tratadas
com 100 nM de DPMA desde C1. Em C6 as células foram incubadas com 5 nM de
3(H)DPCPX na presença ou ausência de 100 µM de CHA. Os resultados representam a
média ± EPM de quatro experimentos realizados em triplicatas (p < 0,001).

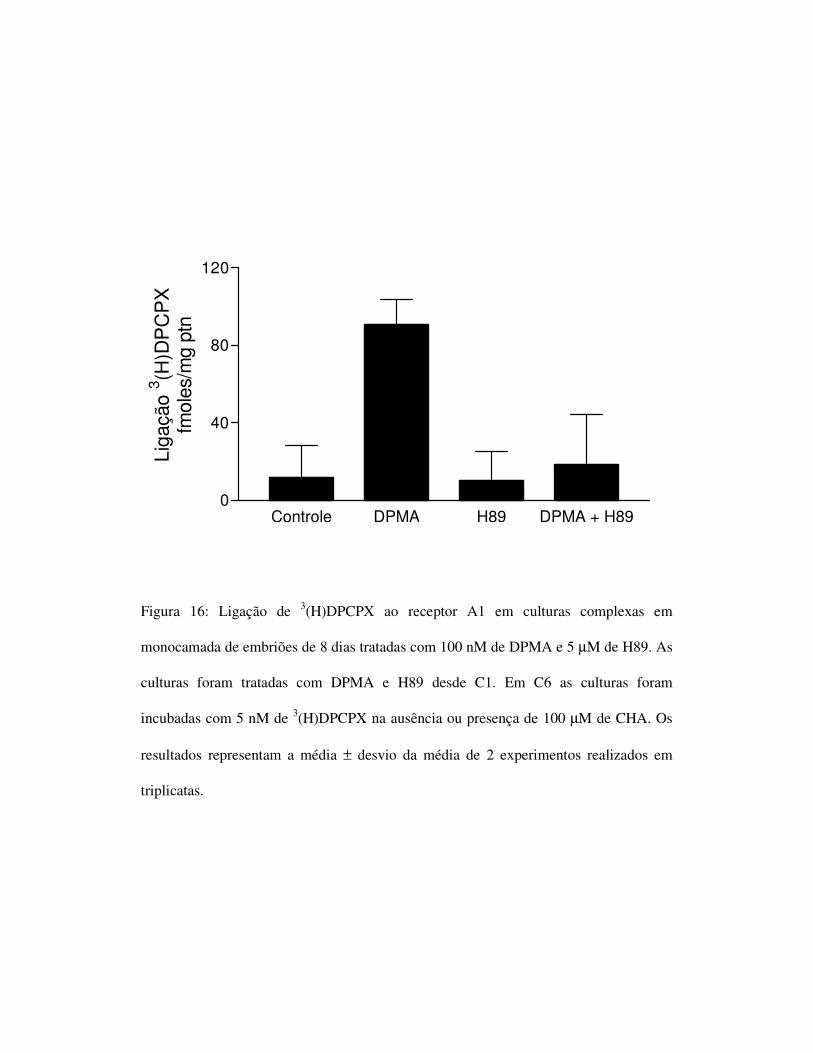
4.4) O bloqueio da atividade da PKA é capaz de bloquear o efeito da
ativação do receptor A2a.
Para verificar se a ativação do receptor A2a estaria levando a um
aumento dos níveis intracelulares de AMPc e este levando a uma ativação
da PKA e conseqüentemente aumentando a ligação do 3(H)DPCPX ao
receptor A1, inibimos a atividade da PKA com H89. A inibição da enzima
levou ao bloqueio do aumento da ligação promovida pelo DPMA
confirmando o envolvimento da PKA nessa via (figura 16).
Controle DPMA H89 DPMA + H890
40
80
120
Lig
ação
3(H
)DP
CP
Xfm
ole
s/m
g p
tn
Figura 16: Ligação de 3(H)DPCPX ao receptor A1 em culturas complexas em
monocamada de embriões de 8 dias tratadas com 100 nM de DPMA e 5 µM de H89. As
culturas foram tratadas com DPMA e H89 desde C1. Em C6 as culturas foram
incubadas com 5 nM de 3(H)DPCPX na ausência ou presença de 100 µM de CHA. Os
resultados representam a média ± desvio da média de 2 experimentos realizados em
triplicatas.

4.5) O bloqueio ou a ativação do receptor A1 aumenta a sua ligação ao
3(H)DPCPX.
Aumento ou diminuição do número de receptores de
neurotransmissores é considerado como um dos principais mecanismos de
adaptação da célula alvo à presença crônica de antagonistas ou agonistas
destes receptores. Para verificar se os receptores A1 também sofriam esse
tipo de regulação nas nossas culturas, resolvemos incubá-los com o
antagonista DPCPX ou o agonista CHA por 5 dias.
O bloqueio do próprio receptor A1 com o antagonista DPCPX
aumenta a ligação do 3(H)DPCPX ao receptor A1 provavelmente por um
mecanismo de “up regulation” do receptor (figura 17).

Controle DPCPX0
25
50
75
100
125
Lig
ação
3(H
)DP
CP
Xfm
ole
s/m
g p
tn
***
Figura 17: Ligação de 3(H)DPCPX ao receptor A1 em culturas em monocamada de
embriões de 8 dias tratatadas com DPCPX. As culturas foram tratadas com 100 nM de
DPCPX desde C1. Em C6 as células foram incubadas com 5 nM de 3(H)DPCPX na
presença ou ausência de 100 µM de CHA. Os resultados representam a média ± EPM de
três experimentos realizados em triplicatas (p < 0,001).
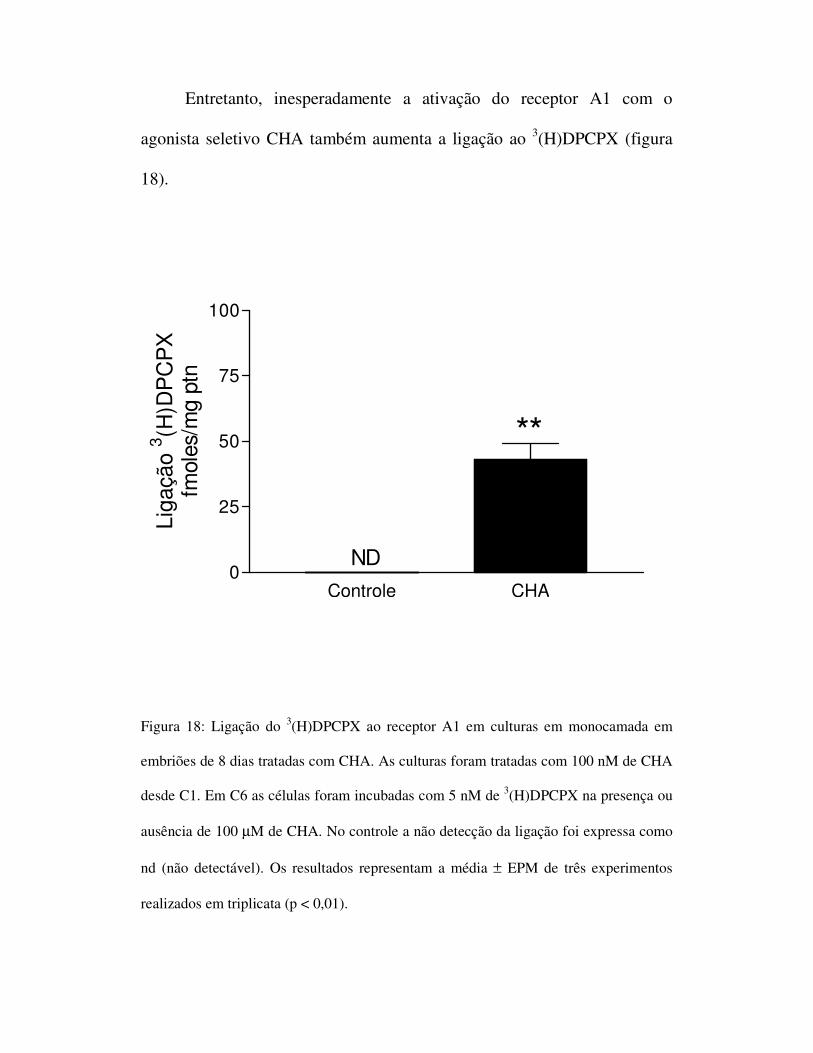
Entretanto, inesperadamente a ativação do receptor A1 com o
agonista seletivo CHA também aumenta a ligação ao 3(H)DPCPX (figura
18).
Controle CHA0
25
50
75
100
ND
**
Lig
ação
3(H
)DP
CP
Xfm
ole
s/m
g p
tn
Figura 18: Ligação do 3(H)DPCPX ao receptor A1 em culturas em monocamada em
embriões de 8 dias tratadas com CHA. As culturas foram tratadas com 100 nM de CHA
desde C1. Em C6 as células foram incubadas com 5 nM de 3(H)DPCPX na presença ou
ausência de 100 µM de CHA. No controle a não detecção da ligação foi expressa como
nd (não detectável). Os resultados representam a média ± EPM de três experimentos
realizados em triplicata (p < 0,01).

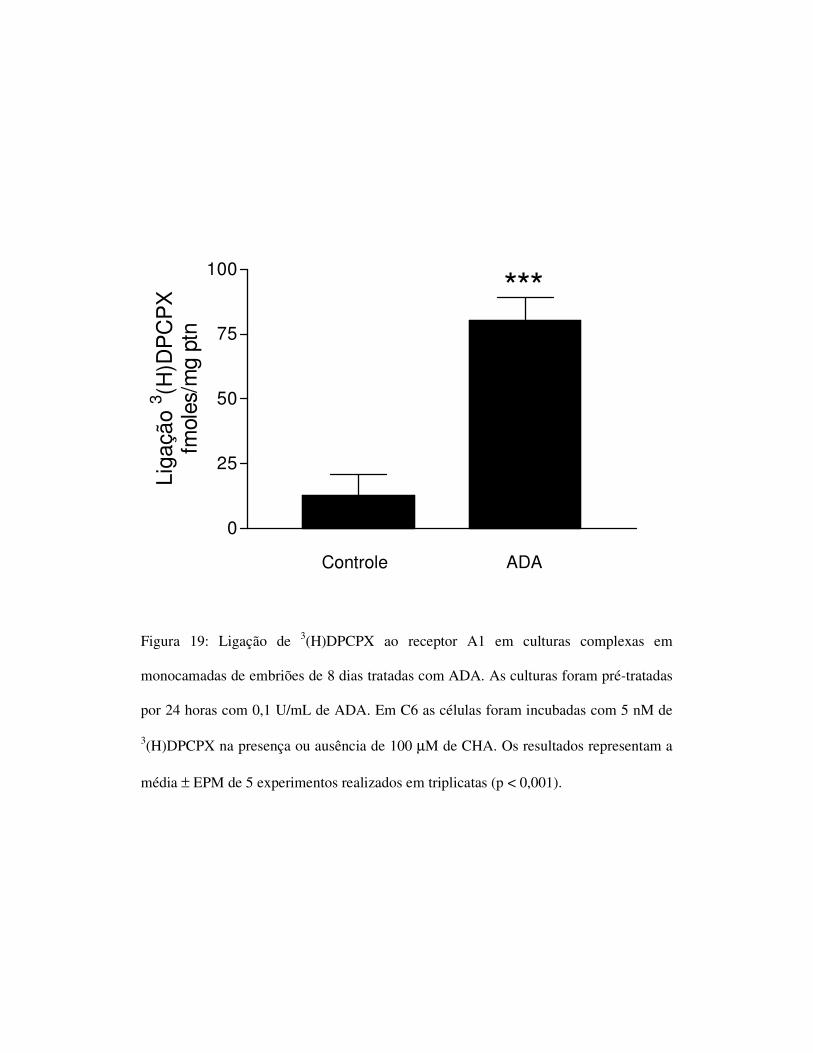
4.6) O tratamento com adenosina deaminase promove aumento da ligação
de 3(H)DPCPX ao receptor A1.
O fato das culturas complexas em monocamada apresentarem pouca
ligação, ou até mesmo nenhuma ligação do 3(H)DPCPX ao receptor A1,
poderia ser causado por um mecanismo de dessensibilização promovido
pela adenosina endógena presente no meio extracelular. Para verificar esta
possibilidade, as culturas foram pré-tratadas com adenosina deaminase
(ADA) para metabolizar a adenosina endógena. Como resultado,
observamos um aumento da detecção do receptor (figura 19).
0
25
50
75
100
Lig
açã
o3(H
)DP
CP
Xfm
ole
s/m
g p
tn
***
Controle ADA
Figura 19: Ligação de 3(H)DPCPX ao receptor A1 em culturas complexas em
monocamadas de embriões de 8 dias tratadas com ADA. As culturas foram pré-tratadas
por 24 horas com 0,1 U/mL de ADA. Em C6 as células foram incubadas com 5 nM de
3(H)DPCPX na presença ou ausência de 100 µM de CHA. Os resultados representam a
média ± EPM de 5 experimentos realizados em triplicatas (p < 0,001).

Para verificar se o efeito produzido pela adenosina deaminase é
dependente da sua atividade metabólica, tratamos as culturas com EHNA
(inibidor da atividade metabólica da adenosina deaminase).
Interessantemente, EHNA sozinho produziu aumento da detecção do
receptor A1 (figura 20).
0
25
50
75
100
125
150
Lig
açã
o3(H
)DP
CP
Xfm
ole
s/m
g p
tn
**
Controle EHNA
Figura 20: Ligação de 3(H)DPCPX ao receptor A1 em culturas complexas em
monocamadas de embriões de 8 dias tratadas com EHNA. As culturas foram pré-
tratadas por 24 horas com 10 µM de EHNA. Em C6 as células foram incubadas com 5
nM de 3(H)DPCPX na presença ou ausência de 100 µM de CHA. Os resultados
representam a média ± EPM de 3 experimentos realizados em triplicatas (p < 0,01).

O principal metabólito produzido pela deaminação da adenosina pela
adenosina deaminase é a inosina. Surpreendentemente, o tratamento das
células por 24 horas com inosina também aumenta a ligação do receptor A1
(figura 21) de forma muito semelhante ao efeito produzido pela adenosina
deaminase. Esse resultado mostra que, provalvelmente, o efeito produzido
pela adenosina deaminase é promovido pela inosina formada pelo
metabolismo e não pela retirada da adenosina endógena.
0
25
50
75
100
Lig
ação
3(H
)DP
CP
Xfm
ole
s/m
g p
tn
Controle Inosina Figura 21: Ligação de 3(H)DPCPX ao receptor A1 em culturas complexas em
monocamadas de embriões de 8 dias tratadas com inosina. As culturas foram pré-
tratadas por 24 horas com 10 µM de inosina. Em C6 as células foram incubadas com 5
nM de 3(H)DPCPX na presença ou ausência de 100 µM de CHA. Os resultados
representam a média ± desvio da média de 2 experimentos realizados em triplicatas.

VII- REFERÊNCIAS BIBLIOGRÁFICAS
ABBRACCHIO, M.P., BRAMBILLA, R., CERUTI, S., KIM, H.O.,
VON LUBTIZ, D.K., JACOBSON, K.A. & CATTABENI, F. (1995). G-
protein-dependent activation of phospholipase C by adenosine A3 receptors
in rat brain. Mol. Pharmacol. 48, 1038-1045.
ALLAMAN, I., LENGACHER, S., MAGISTRETTI, P.J. &
PELLERIN, L. (2003). A2b receptor activation promotes glycogen
synthesis in astrocytes through modulation of gene expression. Am. J.
Physiol. 284, C696-704.
ANDERSON, C.M., XIONG, W., GEIGER, J.D., YOUNG, J.D.,
CASS, C.E., BALDWIN, S.A. & PARKINSON, F.E. (1999).
Distribution of equilibrative, nitrobenzylthioinosine-sensitive nucleoside
transporters (ENT1) in brain. J. Neurochem.73, 867-873.
BARRACO, R.A., HELFMAN, C.C., GOODWIN, B.P. &
ANDERSON, G.F. (1995). Evidence for presynaptic adenosine A2a
receptors associated with norepinephrine release and their desensitization
in the rat nucleus tractus solitarius. J. Neurochem. 65, 1604-1611.

BIBER, K., KLOTZ, K.N., BERGER, M., GEBICKE-HÄRTER, P.J.
& VAN CALCKER, D. (1997). Adenosine A1 receptor-mediated
activation of phospholipase C in cultured astrocytes depends on the level of
receptor expression. J. Neurosci. 17, 4956-4964.
BLAZYNSKI, C. & PEREZ, M.T. (1991). Adenosine in vertebrate
retina: localization, receptor characterization, and function. Cell Mol.
Neurobiol. 11, 463-484.
BRADFORD, M.M. (1976). A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the principle of
protein-dye binding. Anal. Biochem. 72, 248-254.
BRAND, A., VISSIENNON, Z., ESCHKE, D. & NIEBER, K. (2001).
Adenosine A1 and A3 receptors mediate inhibition of synaptic transmission
in rat cortical neurons. Neuropharmacology 40, 85-95.
BREDT, S.D. & SNYDER, S.H. (1990). Isolation of nitric oxide synthase,
a calmodulin-requering enzyme. Proc. Natl. Acad. Sci. 87, 682-685.
BRUNTON L.L. & MAYER, S.E. (1979). Extrusion of cyclic AMP from
pigeon erythrocytes. J. Biol. Chem. 254, 9714-9720.

CANALS, M., MARCELLINO, D., FANELLI, F., CIRUELA, F., DE
BENEDETTI, P., GOLDBERG, S.R., NEVE, K., FUXE, K., AGNATI,
L.F., WOODS, A.S., FERRÉ, S., LLUIS, C., BOUVIER, M. &
FRANCO, R. (2003). Adenosine A2a-dopamine D2 receptor-receptor
heteromerization: qualitative and quantitative assessment by fluorescence
and bioluminescence energy transfer. J. Biol. Chem. 278, 46741-46749.
CIRUELA, F., SAURA, C., CANELA, E.I., MALLOL, J., LLUIS, C.
& FRANCO, R. (1996). Adenosine deaminase affects ligand-induced
signalling by interacting with cell surface adenosine receptors. FEBS
Letters. 380, 219-223.
CIRUELA, F., SAURA, C., CANELA, E.I., MALLOL, J., LLUIS, C.
& FRANCO, R. (1997). Ligand-induced phosphorylation, clustering, and
desensitization of A1 adenosine receptors. Mol. Pharmacol. 52, 788-797.
CIRUELA, F., ESCRICHE, M., BURGUEÑO, J., ANGULO, E.,
CASADÓ, V., SOLOVIEV, M.M., CANELA, E.I., MALLOL, J.,
CHAN, W.Y., LLUIS, C., MCLLHINNEY, J.R.A. & FRANCO, R.
(2001). Metabotropic glutamate 1α and adenosine A1 receptors assemble
into functionally interacting complexes. J. Biol. Chem. 276, 18345-18351.

COSSENZA, M. & PAES-DE-CARVALHO, R. (2000). L-arginine
uptake and release by cultured avian retinal cells: differential cellular
localization in relation to nitric oxide synthase. J. Neurochem. 74, 1885-
1894.
CUNHA, R.A. & RIBEIRO, J.A. (2000). Purinergic modulation of
[3H]GABA release from rat hippocampal nerve terminals.
Neuropharmacology 39, 1156-1167.
CUNHA, R.A. (2001). Adenosine as a neuromodulator and as a
homeostatic regulator in the nervous system: different roles, different
sources and different receptors. Neurochem. Int. 38, 107-125.
DE MELLO, M.C.F., VENTURA, A.L.M., PAES-DE-CARVALHO,
R., KLEIN, W.L. & DE MELLO, F.G. (1982). Regulation of dopamine-
and adenosine-dependent adenylate cyclase systems of chicken embryo
retina cells in culture. Proc. Natl. Acad. Sci. 79, 5708-5712.
DICKENSON, J.M., BLANK, J.L. & HILL, S.J. (1998). Human
adenosine A1 receptor and P2Y2-purinoceptor-mediated activation of the
mitogen-activated protein kinase cascade in transfected CHO cells. Br. J.
Pharmacol. 124, 1491-1499.

DIXON, A.K., WIDDOWSON, L. & RICHARDSON, P.J. (1997).
Desensitisation of the adenosine A1 receptor by the A2a receptor in the rat
striatum. J. Neurochem. 69, 315-321.
DUNWIDDIE, T.V., DIAO, L., KIM, H.O., JIANG, J.L. &
JACOBSON, K.A. (1997). Activation of hippocampal adenosine A3
receptors produces a desensitization of A1 receptor-mediated responses in
rat hippocampus. J. Neurosc. 17, 607-614.
ESCRICHE, M., BURGUEÑO, J., CIRUELA, F., CANELA, E.I.,
MALLOL, J., ENRICH, C., LLUÍS, C. & FRANCO, R. (2003). Ligand-
induced caveolae-mediated internalization of A1 adenosine reeptors:
morphological evidence of endosomal sorting and receptor recycling. Exp.
Cell Res. 285, 72-90.
FAN, M., QIN, W. & MUSTAFA, S.J. (2003). Characterization of
adenosine receptor(s) involved in adenosine-induced bronchoconstriction
in an allergic mouse model. Am. J. Physiol. Lung Cell Mol. Physiol. 284,
L1012-1019.

FAURE, M., VOYNO-YASENETSKAYA, T.A. & BOURNE, H.R.
(1994) cAMP and βγ subunits of heterotrimeric G proteins stimulate the
mitogen-activated protein kinase pathway in COS-7 cells. J. Biol. Chem.
269, 7851-7854.
FEOKTISTOV, I, MURRAY, J.J. & BIAGGIONI, I. (1994). Positive
modulation of intracellular Ca2+ levels by adenosine A2b receptors,
prostacyclin, and prostaglandin E1 via a cholera toxin-sensitive mechanism
in human erythroleukemia cells. Mol. Pharmacol. 45, 1160-1167.
FEOKTISTOV, I. & BIAGGIONI, I. (1997). Adenosine A2b receptors.
Pharmacol. Rev. 49, 381-402.
FEOKTISTOV, I., GOLDSTEIN, A.E. & BIAGGIONI, I. (1999). Role
of p38 mitogen-activated protein kinase and extracellular signal-regulated
protein kinase kinase in adenosine A2B receptor-mediated interleukin-8
production in human mast cells. Mol. Pharmacol. 55, 726-734.

FERGUSON, G., WATTERSON, K.R. & PALMER, T.M. (2002).
Subtype-specific regulation of receptor internalization and recycling by the
carboxyl-terminal domains of the human A1 and rat A3 adenosine
receptors: consequences for agonist-stimulated translocation of arrestin3.
Biochemistry 41, 14748-14761.
FERRÉ, S., KARCZ-KUBICHA, M., HOPE, B.T., POPOLI, P.,
BURGUENO, J., GUTIERREZ, M.A., CASADÓ, V., FUXE, K. &
FRANCO, R. (2002). Synergistic interaction between adenosine A2A and
glutamate mGlu5 receptors: implications for striatal neuronal function.
Proc. Natl. Acad. Sci. 97, 8606-8611.
FERREIRA, J.M. & PAES-DE-CARVALHO, R. (2001). Long-term
activation of adenosine A2a receptors blocks glutamate excitotoxicity in
cultures of avian retinal neurons. Brain Res. 900, 169-176.
FRANCO, R. (2000). Dopamine D1 and adenosine A1 receptors form
functionally interacting heteromeric complexes. Proc. Natl. Acad. Sci. 97,
8606-8611.

FREDHOLM, B.B., ARSLAN, G., HALLDNER, L., KULL, B.,
SCHULTE, G. & WASSERMAN, W. (2000). Structure and function of
adenosine receptors and their genes. Naunyn Schmiedebergs Arch.
Pharmacol. 362, 364-374.
FREDHOLM, B.B., IJZERMAN, A.P., JACOBSON, K.A., KLOTZ,
K.N. & LINDEN J. (2001). International union of pharmacology. XXV.
Nomenclature and classification of adenosine receptors. Pharmacol. Rev.
53,527-552.
GAO, Z., CHEN, T., WEBER, M.J. & LINDEN, J. (1999). A2b
adenosine and P2Y2 receptors stimulate mitogen-activated protein kinase
embryonic kideney-293 cells: cross-talk between cyclic AMP and protein
kinase C pathways. J. Biol. Chem. 274, 5972-5980.
GEBICKE-HAERTER, P.J., CHRISTOFFEL, F., TIMMER, J.,
NORTHOFF, H., BERGER, M. & VAN CALCKER, D. (1996). Both
adenosine A1- and A2-receptors are required to stimulate microglial
proliferation. Neurochem. Int. 29, 37-42.

GERBER, U., GREENE, R.W., HAAS, H.L. & STEVENS, D.R. (1989).
Caracterization of inhibition mediated by adenosine in hippocampus of the
rat in vitro. J. Physiol. 417, 567-578.
GOLDBERG, S.R., LLUIS, C., FRANCO, R. & CIRUELA, F. (2002).
Synergistic interaction between adenosine A2a and glutamate mGlu5
receptors: implications for striatal neuronal funstion. Proc. Natl. Acad. Sci.
99, 11940-11945.
GONZALEZ, G.A., YAMAMOTO, K.K., FISCHER, W.H., KARR,
D., MENZEL, P., BIGGS, W.3RD, WALE, W.W. & MONTMINY,
M.R. (1989). A cluster of phosphorylation sites on the cyclic AMP-
regulated nuclear factor CREB predicted by its sequence. Nature 337, 749-
752.
GUINÉS, S., HILLION, J., TORVINEN, M., LE CROM, S, CASADÓ,
V., CANELA, E.I., RONDIN, S., LEW, J.Y., WATSON, S., ZOLI, M.,
AGNATI, L.F., VERNIER, P., LLUIS, C., FERRÉ, S., FUXE, K. &
FRANCO, R. (2000). Dopamine D1 and adenosine A1 receptors form
functionally interacting heteromeric complexes. Proc. Natl. Acad. Sci. 97,
8606-8611.

HAMMARBERG, C., SCHULTE, G. & FREDHOLM, B.B. (2003).
Evidence for functional adenosine A3 receptors in microglia cells. J.
Neurochem. 86, 1051-1054.
HAYNES, J. Jr., OBIAKO, B, THOMPSON, W.J. & DOWNEY, J.
(1995). Adenosine regulates the survival of avian retinal neurons and
photoreceptors in culture. Am. J. Physiol. 268, H1862-1868.
HENDERSON, G.B. & STRAUSS, B.P. (1991). Evidence for cAMP and
cholate extrusion in C6 rat glioma cells by a common anion efflux pump. J.
Biol. Chem. 266, 1641-1645.
HETTINGER, B.D., LEE, A., LINDEN, J. & ROSIN, D.L. (2001).
Ultrastructural localization of adenosine A2a receptor suggests multiple
cellular sites for modulation of GABAergic neurons in rat striatum. J.
Comp. Neurol. 431, 331-346.
JARVIS, S.M. & YOUNG, J.D. (1986). Nucleoside transport in rat
erythrocytes: two components with differences in sensitivity to inhibition
by nitrobenzylthioinosine and p-chloromercuriphenyl sulfonate. J. Membr.
Biol. 93, 1-10.

JIN, X., SHEPHERD, R.K., DULING, B.R. & LINDEN, J. (1997).
Inosine binds to A3 adenosine receptors and stimulates mast cell
degranulation. J. Clin. Invest. 100, 2849-2857.
LAEMMLI, U.K. (1970). Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227, 680-685.
LATINI, S., BORDONI, F., CORRADETTI, R., PEPEU, G. &
PEDATA, F. (1999). Effect of A2A adenosine receptor stimulation and
antagonism on synaptic depression induced by in vitro ischemia in rat
hippocampal slices. Br. J. Pharmacol. 128, 1035-1044.
LATINI, S. & PEDATA, F. (2001). Adenosine in the central nervous
system: release mechanisms and extracellular concentrations. J.
Neurochem. 79, 463-484.
LI, X.X., NOMURA, T., AIHARA, H., & NISHIZAKI, T. (2001).
Adenosine enhances glial glutamate efflux via A2a adenosine receptors.
Life Sci. 68, 1343-1350.

LIANG, B.T. & HALTIWANGER, B. (1995). Adenosine A2a and A2b
receptors in cultured fetal chick heart cells. High- and low-affinity coupling
to stimulation of myocyte contractility and cAMP accumulation. Circ. Res.
76, 242-251.
LIU, G.S., RICHARDS, S.C., OLSSON, R.A., MULLANE, K.,
WALSH, R.S. & DOWNEY, J.M. (1994). Evidence that adenosine A3
receptor may mediate the protection afforded by preconditioning in the
isolated rabbit heart. Cardiovasc. Res. 28, 1057-1061.
LOPES, L.V., CUNHA, R.A., KULL, B., FREDHOLM, B.B. &
RIBEIRO, J.A. (2002). Adenosine A2a receptor facilitation of
hippocampal synaptic transmission is dependent on the tonic A1 receptor
inhibition. Neuroscience 112, 319-329.
LOWRY, O.H., ROSEBROUGH, N.J., FARR, A.L. & RANDALL,
R.J. (1951). Protein measurement with the Folin phenol reagent. J. Biol.
Chem. 193, 265-275.
LLOYD, H.G., DEUSSEN, A., WUPPERMANN, H. & SCHRADER, J.
(1988). The transmethylation pathway as a source for adenosine in the
isolated guinea-pig heart. Biochem. J. 252, 489-494.

LUTTRELL, L.M., VAN BIESEN, T., HAWES, B.E., KOCH, W.J.,
KRUEGER, K.M., TOUHARA, K. & LEFKOWITZ, R.J. (1997). G-
protein-coupled receptors and their regulation: activation of the MAP
kinase signaling pathway by G-protein-coupled receptors. Adv. Second
Messenger Phosphoprotein Res. 31, 263-277.
MARTIN, P.L. & POTTS, A.A. (1994). The endothelium of the rat renal
artery plays an obligatory role in A2 adenosine receptor-mediated
relaxation induced by 5'-N-ethylcarboxamidoadenosine and N6-
cyclopentyladenosine. J. Pharmacol. Exp. Ther. 270, 893-899.
MATHARU, A.L., MUNDELL, S.J., BENOVIC, J.L. & KELLY, E.
(2001). Rapid agonist-induced desensitization and internalization of the
A2b adenosine receptor is mediated by serine residue close to the COOH
terminus. J. Biol. Chem. 276, 30199-30207.
MERIGHI, S., BENINI, A., MIRANDOLA, P., GESSI, S., VARANI,
K., LEUNG, E., MACLENNAN, S. & BOREA, P.A. (2005). A3
adenosine receptor activation inhibits cell proliferation via
phosphatidylinositol 3-kinase/Akt-dependent inhibition of the extracellular
signal-regulated kinase 1/2 phosphorylation in A375 human melanoma
cells. J. Biol. Chem.280, 19516-19526.

MEYER-FRANKE, A., WILKINSON, G.A., KRUTTGEN, A., HU,
M., MUNRO, E., HANSON, M.G.JR., REICHARDT, L.F. &
BARRES, B.A. (1998). Depolarization and cAMP elevation rapidly recruit
TrkB to the plasma membrane of CNS neurons. Neuron 21, 681 – 693.
MOFFETT, S., ROUSSEAU, G., LAGACE, M. & BOUVIER, M.
(2001). The palmitoylation state of the beta(2)-adrenergic receptor
regulates the synergistic action of cyclic AMP-dependent protein kinase
and beta-adrenergic receptor kinase involved in its phosphorylation and
desensitization. J. Neurochem. 76, 269-279.
MOGUL, D.J., ADAMS, M.E. & FOX, A.P. (1993). Differential
activation of adenosine receptors decrease N-type but potentiates P-type
Ca2+ current in hippocampal CA3 neurons. Neuron 10, 327-334.
NIE, Z., MEI, Y., FORD, M., RYBAK, L., MARCUZZI, A., REN, H.,
STILES, G.L. & RAMKUMAR, V. (1998). Oxidative stress increases A1
adenosine receptor expression by activating nuclear factor kB. Mol.
Pharmacol. 53, 663-669.

OFFERMANNS, S. & SIMON, M.I. (1995). G alpha 15 and G alpha 16
couple a wide variety of receptors to phospholipase C. J. Biol. Chem. 270,
15175-15180.
OLAH, M.E., REN, H. & STILES, G.L. (1995). Adenosine receptors:
protein and gene structure. Arch. Int. Pharmacodyn. Ther. 329, 135-150.
OLAH, ME. (1997). Indentification of A2a adenosine receptor domains
involved in selective coupling to Gs. Analysis of chimeric A1/A2a
adenosine receptors. J. Biol. Chem. 272, 337-344.
OSADA, S., MIZUNO, K., SAIDO, T.C., SUZUKI, K., KUROKI &
OHNO, S. (1992). A new member of the protein kinase C family, nPKC
theta, predominantly expressed in skeletal muscle. Mol. Cel. Biol. 12,
3930-3938.
OTHMAN, T., YAN, H. & RIVKEES, S.A. (2003). Oligodendrocytes
express functional A1 adenosine receptors that stimulate cellular migration.
Glia 44, 166-172.

PAES-DE-CARVALHO, R. & DE MELLO, F.G. (1982). Adenosine-
elicited accumulation of adenosine 3’, 5’-cyclic monophosphate in the
chick embryo retina. J. Neurochem. 38, 493-500.
PAES-DE-CARVALHO, R. & DE MELLO, F.G. (1985). Expression of
A1 adenosine receptor modulating dopamine-dependent cyclic AMP
accumulation in the chick embryo retina. J. Neurochem. 44, 845-851.
PAES-DE-CARVALHO, R., BRAAS, K.M., SNYDER, S.H. &
ADLER, R. (1990) Analysis of adenosine immunoreactivity, uptake, and
release in purified cultures of developing chick embryo retinal neurons and
photoreceptors. J. Neurochem. 55, 1603-1611.
PAES-DE-CARVALHO, R. (1990). Development of A1 adenosine
receptors in the chick embryo retina. J. Neurosci. Res. 25, 236-242.
PAES-DE-CARVALHO, R., MAIA, G.A. & FERREIRA, J.M. (2003).
Adenosine regulates the survival of avian retinal neurons and
photoreceptors in culture. Neurochem. Res. 28, 1583-1590.

PAES-DE-CARVALHO, R., DIAS, B.V., MARTINS, R.A., PEREIRA,
M.R., PORTUGAL, C.C. & LANFREDI, C. (2005). Activation of
glutamate receptors promotes a calcium-dependent and transporter-
mediated release of purines in cultured avian retinal cells: possible
involvement of calcium/calmodulin-dependent protein kinase II.
Neurochem. Int. 6, 441-451.
PAK, M.A., HAAS, H.L., DUCKING, U.K. & SCHRADER, J. (1994).
Inhibition of adenosine kinase increases endogenous adenosine and depress
neuronal activity in hippocampal slices. Neuropharmacology 33, 1049-
1053.
PALMER, TM., GETTYS, T.W. & STILES, G.L. (1995). Differential
interaction with and regulation of multiple G-proteins by the rat A3
adenosine receptor. J. Biol. Chem. 270, 16895-16902.
PALMER, T.M., BENOVIC, J.L. & STILES, G.L. (1996). Molecular
basis for subtype-specific desensitization of inhibitory adenosine receptors.
Analysis of a chimeric A1-A3 adenosine receptor. J. Biol. Chem. 271,
15272-15278.

PALMER, T.M. & STILES, G.L. (1997). Identification of an A2a
adenosine receptor domain specifically responsible for mediating short-
term desensitization. Biochem. 36, 832-338.
PIERCE, KD., FURLONG, T.J., SELBIE, L.A. & SHINE, J. (1992).
Molecular cloning and expression of an adenosine A2b receptor from
human brain. Biochem. Biophys. Res. Commun. 187, 86-93.
RAMKUMAR, V., OLAH, M.E., JACOBSON, K.A. & STILES, G.L.
(1991). Distinct pathways of desensitization of A1- and A2-adenosine
receptors in DDT1 MF-2 cells. Mol. Pharmacol. 40, 639-647.
RANE, S.G. & REDDY, E.P. (2000). Janus kinases: components of
multiple signaling pathways. Oncogene 19, 5662-5679.
REBOLA, N., OLIVEIRA, C.R. & CUNHA R.A. (2002). Transducing
system operated by adenosine A2a reecptors to facilitate acetylcholine
release in the rat hippocampus. Eur. J. Pharmacol. 454, 31-38.
REBOLA, N., CANAS, P., OLIVEIRA, C.R. & CUNHA, R.A. (2005).
Different synaptic and subsynaptic localization of adenosine A2a receptors
in the hippocampus and striatum of the rat. Neuroscience 132, 893-903.

RODRIGUES, R.J., ALFARO, T.M., REBOLA, N., OLIVEIRA, C.R.
& CUNHA, R.A. (2005). Co-localization and functional interaction
between adenosine A2a and metabotropic group 5 receptors in
glutamatergic nerve terminals of the rat striatum. J. Neurochem. 92, 443-
441.
ROUX, P.P. & BLENIS, J. (2004). ERK and P38 MAPK-activated
protein kinases: a family of protein kinases with diverse biological
functions. Microbiol. Mol. Biol. Rev. 68, 320-344.
RUBINO, A., RALEVIC, V. & BURNSTOCK G. (1995). Contribution
of P1-(A2b subtype) and P2-purinoceptors to the control of vascular tone in
the rat isolated mesenteric arterial bed. Br. J. Pharmacol. 115, 648-652.
SAKAMOTO, K., YNOKI, Y., KUWAGATA, M., SAITO, M.,
NAKAHARA, T. & ISHII, K. (2004). Histological protection against
ischemia-reperfusion injury by early ischemic preconditioning in rat retina.
Brain Res. 1015, 154-160.

SATO, A. TERATA, K, MIURA, H. TOYAMA, K., LOBERIZA,
F.R.JR., HATOUM, O.H., SAITO, T., SAKUMA, I. & GUTTERMAN,
D.D. (2005). Mechanism of vasodilation to adenosine in coronary arteioles
from patients with heart disease. Am. J. Physiol. Heart Circ. Physiol. 288,
H1633-1640.
SAURA, C., CIRUELA, F., CASADO, V., CANELA, E.I., MALLOL,
J., LLUIS, C. & FRANCO, R. (1996). Adenosine deaminase interacts
with A1 adenosine receptors in pig brain cortical membranes. J.
Neurochem. 66, 1675-1682.
SCHULTE, G. & FREDHOLM, B.B. (2000). Human adenosine A1, A2a,
A2b and A3 receptors expressed in chinese hamster ovary cells all mediate
the phosphorylation of extracellular-regulated kinase 1/2. Mol. Pharmacol.
58, 477-482.
SCHULTE, G. & FREDHOLM, B.B. (2002). Signalling pathway from
the human adenosine A3 receptor expressed in chinese hamster ovary cells
to the extracellular signal-regulated kinase 1/2. Mol. Pharmacol. 62, 1137-
1146.

SCHULTE, G. & FREDHOLM, B.B. (2003). Signalling from adenosine
receptors to mitogen-activated protein kinases. Cellular Signalling. 15,
813-827.
SEIDEL, M.G., KLINGER, M., FREISSMUTH, M. & HOLLER, C.
(1999). Activation of mitogen-activated protein kinase by the A2a-
adenosine receptor via a rap1-dependent and via a p21(ras)-dependent
pathway. J. Biol. Chem. 274, 25833-25841.
SERGER, R. & KREBS, E.G. (1995). The MAPK signaling cascade.
FASEB. 9, 726-735.
SEXL, V., MANCUSI, G., HOLLER, C., GLORIA-MAERCKER, E.,
SCHUTZ, W. & FREISSMUTH, M. (1997). Stimulation of the mitogen-
activated protein kinase via the A2a-adenosine receptor in primary human
endothelial cells. J. Biol. Chem. 272, 5792-5799.
SHAHRAKI, A. & STONE, T.W. (2004). Blockade of presynaptic
adenosine A1 receptor responses by nitric oxide and superoxide in rat
hippocampus. Eur. J. of Neurosci. 20, 719-728.

TETZLAFF, W., SCHUBERT, P. & KREUTZBERG, G.W. (1987).
Synaptic and extrasynaptic localization of adenosine binding sites in the
hippocampus. Neuroscience 21, 869-875.
TOWBIN, H., STACHELIN, T. & GORDON J. (1979). Eletrophoretic
transfer of proteins from polyacrylamide gels to nitrocellulose sheets:
procedure and some applications. Proc. Natl. Acad. Sci. 76, 4350-4354.
TRINCAVELLI, M.L., TUSCANO, D., MARRONI, M., KLOTZ,
K.N., LUCACCHINI, A. & MARTINI, C. (2002). Involvement of
mitogen protein kinase cascade in agonist-mediated human A3 adenosine
receptor regulation. Bioch. et Biophy. Acta. 1591, 55-62.
VAN CALKER, D., MÜLLER, M. & HAMPRECHT, B. (1979).
Adenosine regulates via two different types of receptors, the accumulation
of cyclic AMP in cultured brain cells. J. Neurochem. 33, 999-1005.
VAN DER GRAAF, P.H., VAN SCHAICK E.A., VISSER S.A., DE
GREEF, H.J., IJZERMAN, A.P. & DANHOF, M. (1999). Mechanism-
based pharmacokinetic-pharmacodynamic modeling of antilipolytic effects
of adenosine A(1) receptor agonists in rats: precondiction of tissue-
dependent efficacy in vivo. J. Pharmacol. Exp. Ther. 290, 702-709.

VON LUBITZ, D.K., LIN, R.C., POPIK, P., CARTER, M.F. &
JACOBSON, K.A. (1994). Adenosine A3 receptor stimulation and
cerebral ischemia. Eur. J. Pharmacol. 263, 59-67.
YOSHIOKA, K., HOSODA, R., KURODA, Y. & NAKATA, H. (2002).
Hetero-oligomerization of adenosine A1 receptors with P2Y1 receptors in
rat brains. FEBS Lett. 531, 299-303.
WARDAS, J. (2002). Neuroprotective role of adenosine in the CNS. Pol.
J. Pharmacol. 54, 313-326.
WILLETS, J.M., PARENT, J.L., BENOVIC, J.L. & KELLY, E.
(1999). Selective reduction in A2 adenosine receptor desensitization
following antisense-induced suppression of G protein-coupled receptor
kinase 2 expression. J. Neurochem. 73, 1781-1789.
WITTENDORP, M.C., BODDEKE, H.W. & BIBER, K. (2004).
Adenosine A3 receptor-induced CCL2 synthesis in cultured mouse
astrocytes. Glia 46, 410-418.

ZHOU, Q.Y., LI, C., OLAH, M.E., JOHNSON, R.A., STILES, G.L. &
CIVELLI, O. (1992). Molecular cloning and characterization of an
adenosine receptor: the A3 adenosine receptor. Proc. Natl. Acad. Sci. 89,
7432-7436.
http://www.aderis.com/img/art_adenosine.gif
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=bnchm.figgrp.1262
http://www.starosta.com/technical/art/membrane_receptor.jpg
http://homepages.strath.ac.uk/~dfs99109/BB329/PKC.jpg
http://wbmo.mpimf-heidelberg.mpg.de/~teuler/Pics/retina1.jpg

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo


















