
Cláudia Emanuele Carvalho de Sousa - teses.usp.br fileCláudia Emanuele Carvalho de Sousa...
120
Cláudia Emanuele Carvalho de Sousa Caracterização do eixo imune-pineal: Mecanismo de ação do controle da função pineal pela citocina pró- inflamatória TNF Characterization of immune-pineal axis: Mechanism of action of the control of pineal function by pro-inflammatory cytokine TNF São Paulo 2011
-
Upload
trinhtuong -
Category
Documents
-
view
223 -
download
2
Transcript of Cláudia Emanuele Carvalho de Sousa - teses.usp.br fileCláudia Emanuele Carvalho de Sousa...

Cláudia Emanuele Carvalho de Sousa
Caracterização do eixo imune-pineal: Mecanismo de
ação do controle da função pineal pela citocina pró-
inflamatória TNF
Characterization of immune-pineal axis: Mechanism of
action of the control of pineal function by pro-inflammatory cytokine
TNF
São Paulo
2011

Cláudia Emanuele Carvalho de Sousa
Caracterização do eixo imune-pineal: Mecanismo de
ação do controle da função pineal pela citocina pró-
inflamatória TNF
Characterization of immune-pineal axis: Mechanism of
action of the control of pineal function by pro-inflammatory cytokine
TNF
Dissertação apresentada ao Instituto de
Biociências da Universidade de São Paulo,
para a obtenção de Título de Mestre em
Ciências na Área de Fisiologia Geral.
Orientadora: Regina Pekelmann Markus
São Paulo
2011

Ficha Catalográfica
Carvalho-Sousa, Cláudia Emanuele
Caracterização do eixo- imune-pineal : mecanismo de ação do controle da função
pineal pela citocina pró-inflamatória TNF.
120 p. : il.
Dissertação (Mestrado) – Instituto de Biociências da
Universidade de São Paulo. Departamento de Fisiologia. 2011.
1. Glândula Pineal. 2. NFKB. 3. TNF. I. Pelkemann, Regina Markus. II. Título. III.
Universidade de São Paulo. Instituto de Biociências. Departamento de Fisiologia.
Comissão Julgadora:
____________________________ ____________________________
Prof(a). Dr(a). Prof(a). Dr(a).
____________________________
Prof(a). Dr(a).
Orientadora

Dedicatória
“Ainda que eu falasse as línguas dos homens e dos anjos, e não tivesse amor, seria como
o metal que soa ou como o címbalo que retine.
E ainda que tivesse o dom de profecia, e conhecesse todos os mistérios e toda a ciência, e
ainda que tivesse toda fé, de maneira tal que transportasse os montes, e não tivesse
amor, nada seria (I Coríntios 13: 1; 2)”.
À minha família que representa o amor mais profundo que posso experimentar.

A arte de ser feliz
Houve um tempo em que minha janela se abria sobre uma cidade que parecia ser feita de
giz. Perto da janela havia um pequeno jardim quase seco.
Era uma época de estiagem, de terra esfarelada, e o jardim parecia morto. Mas todas as
manhãs vinha um pobre com um balde, e, em silêncio, ia atirando com a mão umas gotas
de água sobre as plantas. Não era uma rega: era uma espécie de aspersão ritual, para que
o jardim não morresse. E eu olhava para as plantas, para o homem, para as gotas de água
que caíam de seus dedos magros e meu coração ficava completamente feliz.
Às vezes abro a janela e encontro o jasmineiro em flor. Outras vezes encontro nuvens
espessas. Avisto crianças que vão para a escola. Pardais que pulam pelo muro. Gatos que
abrem e fecham os olhos, sonhando com pardais. Borboletas brancas, duas a duas, como
refletidas no espelho do ar. Marimbondos que sempre me parecem personagens de Lope
de Vega. Ás vezes, um galo canta. Às vezes, um avião passa. Tudo está certo, no seu
lugar, cumprindo o seu destino. E eu me sinto completamente feliz.
Mas, quando falo dessas pequenas felicidades certas, que estão diante de cada janela,
uns dizem que essas coisas não existem, outros que só existem diante das minhas
janelas, e outros, finalmente, que é preciso aprender a olhar, para poder vê-las assim.
Cecília Meirelles

Agradecimentos
Meu Deus obrigada por ser comigo quando e onde precisei, por permitir que
conseguisse passar por todas etapas até aqui. Agradeço por todas pessoas que foram
usadas para me fortalecer e fazer crescer.
Aos meus pais, sempre incentivadores! Minha mãe pelo exemplo de vida, de
sabedoria, pelos cuidados, por me dar tantos motivos para seguir adiante. Meu pai pelo
carinho, pela alegria, por todos sacrifícios empregados. Às minhas amadas irmãs, por
trazerem tantas alegrias. Lina, por ser a melhor amiga que pude contar, pelos dias
maravilhosos durante a faculdade, pelos ensinamentos de vida, por sua doçura e amor.
Ticinha, você é luz em nossas vidas, seu apoio foi essencial, nossas conversas à distância
mudaram a cor do meu dia. Eduardo, um irmão que a vida me trouxe, obrigada por
deixar tantas boas novas, por tantos risos, por me lembrar da grandeza de Deus em
todas as coisas. Ao querido Miguel, que nos ensinou tanto, nosso amor foi incondicional
às realidades, um anjo que logo chegou e partiu.
À família Carvalho, pela presença constante em minha vida. Em especial a tia
Melânia, tio Pedro, tio Chico, tia Dulce e ao meu tio Marconi pelo maior presente que
pude receber, saudades eternas. Ao meu afilhado, por ser também exemplo. Aos meus
primos queridos, sempre amáveis e divertidos, pela força. Em especial ao Pedrinho e
Samuel, meus queridos, e ao André, um grande irmão.
Aos irmãos que pude escolher, meus amigos! Tenho tantos nomes na cabeça, mas
sei que alguns não são apenas nomes, fizeram parte de tudo até aqui. A vocês que
fizeram da saudade algo suportável, pela alegria, por vivências memoráveis. À minha
amiga Lívia, uma dádiva de Deus em minha vida, pelas incontáveis demonstrações de
carinho, de companheirismo, sua amizade é imprescindível, obrigada por tanta
paciência, pelos conselhos, pela força. Às minhas amigas de Abaeté, Gabi, Alana,

Marjory, Tati e Thaís, que mesmo a distância mostraram que o essencial é duradouro.
Aos amigos da Igreja Batista, pela oração e por tanto carinho. Aos colegas de faculdade
que tanto aprendi, em especial as minhas grandes e eternas amigas, Fabi, Mayna, Sarah
e Suzana, que marcaram Juiz de Fora em minha vida. Aos amigos da escola Daniel e
Vinícius, pela fidelidade. E as minhas mais recentes amigas, que compartilharam comigo
toda a caminhada na pós-graduação, Maisinha, Naty, Ketrin e Nortista, representam
minha família aqui, nossos momentos ora trágicos e cômicos, as discussões filosóficas,
científicas, angústias, quantas experiências trocadas.
Aos amigos do laboratório, companheiros da vida acadêmica. Obrigada por tanto
aprendizado, pela paciência, pelas colaborações, pelas brincadeiras. Em especial, ao
Sanseray, grande amigo e colaborador, obrigada pelo incentivo desde o curso de inverno,
por todos momentos que pude contar com você. Ao Eduardo, um exemplo pra mim,
obrigada por ser tão disponível, por me auxiliar em todas etapas deste trabalho. Ao
Pedroca, agora Professor Pedro, por tantas contribuições, por me ajudar na caminhada
do laboratório, pelas conversas produtivas, sua orientação foi essencial em todo
processo. À Professora Luciana Pinato, pela constante colaboração, pelas boas
conversas, pelos ensinamentos. À Sandrinha, entrou agora e já participou de tantas
vivências no laboratório, pelo apoio, amizade e colaboração. À professora Zulma
Ferreira, sempre disposta a ajudar, pelas contribuições nos ensaios e como não falar nas
boas risadas. À orientadora Regina P. Markus, pela acolhida, confiança, incentivo, pelas
críticas fundamentais ao longo desses dois anos, por compartilhar suas experiências e
conhecimento, pela presença sempre marcante, por ensinar em todas oportunidades.
Agradeço, por fim, o fundamental apoio financeiro e científico da FAPESP
(08/56080-4), CNPq e CAPES.

Índice
I. Introdução ................................................................................................................................ 13
1. Glândula Pineal .................................................................................................................... 13
1.1. Histórico da glândula pineal ......................................................................................... 13
1.2. Aspectos Morfofuncionais - Anatomia, Histologia, Inervação e Vascularização ......... 15
1.3. Produção de melatonina pela glândula pineal ............................................................. 19
1.4. Da secreção aos mecanismos de ação da melatonina ................................................. 25
2. Fator de Necrose Tumoral (TNF) ......................................................................................... 26
2.1. Caracterização do TNF .................................................................................................. 26
2.2. Sinalização do TNF ........................................................................................................ 28
3. Fator de Transcrição Nuclear Kappa B (NFKB) .................................................................... 32
3.1. Ativação do NFKB ......................................................................................................... 34
4. Eixo-Imune-Pineal ............................................................................................................... 38
4.1. Melatonina e defesa ..................................................................................................... 38
4.2. O tempo na resposta imune: relação entre melatonina e adaptação. ........................ 40
4.3. A construção de um novo conceito: Pineal como alvo para mediadores da resposta
imune .................................................................................................................................. 42
II. Objetivos ................................................................................................................................. 49
III. Material e Métodos ................................................................................................................ 51
1. Animais ................................................................................................................................ 51
2. Drogas e Reagentes ............................................................................................................. 51
3. Preparo de drogas ............................................................................................................... 52
4. Cultura da glândula pineal .................................................................................................. 53
5. Cultura de pinealócitos ....................................................................................................... 53
6. Teste de Viabilidade Celular ................................................................................................ 54
7. Imuno-histoquímica da glândula pineal .............................................................................. 54
7.1. Método de Revelação com DAB ................................................................................... 54
7.2. Imunofluorescência- Free Floating ............................................................................... 56
8. Imunofluorescência de cultura de pinealócitos .................................................................. 57
9. Detecção da produção de NO por microscopia confocal .................................................... 58
10. Extração de proteína nuclear ............................................................................................ 59

11. Ensaio de eletromobilidade por retardo em gel (EMSA) .................................................. 60
12. Ensaio de supershift .......................................................................................................... 61
13. Análise dos resultados ....................................................................................................... 61
IV. Resultados .............................................................................................................................. 63
1- Expressão Do Receptor TNF-R1 Nos Diferentes Tipos Celulares Da Pineal. ....................... 63
2- Variação Da Expressão Do TNF-R1 Ao Longo Do Dia .......................................................... 66
3. Pinealócitos Expressam Constitutivamente o Receptor TNF-R1 ......................................... 67
4. TNF Promove A Translocação Nuclear Do NFKB ................................................................. 69
4.1. Decurso temporal da ativação do NFKB induzida por TNF .......................................... 69
4.2. Subunidades presentes na translocação nuclear do NFKB .......................................... 71
4.3. Efeito do TNF sobre a translocação dos dímeros do NFKB .......................................... 73
5. TNF promove a degradação da proteína inibitória IKBα em pinealócitos dispersos .......... 74
6. TNF aumenta a expressão de iNOS em pinealócitos através de mecanismo dependente de
NFKB ........................................................................................................................................ 76
6.1. TNF induz a expressão da sintase de óxido nítrico induzível (iNOS) em pinealócitos
isolados ................................................................................................................................ 76
7. TNF Induz A Produção De Óxido Nítrico Em Pinealócitos Isolados ..................................... 78
V. Discussão ................................................................................................................................. 81
VI. Conclusões ............................................................................................................................. 90
VII. Resumo ................................................................................................................................. 91
VIII. Abstract ................................................................................................................................ 92
IX. Referências Bibliográficas ...................................................................................................... 93
X. Súmula Curricular .................................................................................................................. 111
XI. Anexos .................................................................................................................................. 120

Lista de abreviaturas
5-HT- serotonina
5-HTP- 5-hidroxitriptofano
AAAD- enzima descarboxilase de
aminoácidos aromáticos
AA-NAT- enzima arilalquilamina N-
acetiltransferase
AC- enzima adenilil ciclase
AMPc- monofosfato de adenosina
cíclico
AP-1- proteína ativadora 1
ATP- adenosina trifosfato
BCG- Bacilo Calmette-Guerin
CD14- cluster differentiation 14
COX- ciclooxigenase
CRE- elemento responsivo a AMPc
CREB- proteína de ligação ao elemento
responsivo a AMPc
DAB- 3, 3‟-diaminobenzidina
DAF-FM-DA- 4-amino-5-metilamino-
2´,7´- difluorofluoresceina
DAPI- 4’, 6- diamidino-2-fenilindol
EMSA- ensaio de eletromobilidade por
retardo em gel
FADD - Fas-associated DD protein
GABA- ácido gama-aminobutírico
GCS- gânglio cervical superior
GFAP- glial fibrillary acidic protein,
marcador de astrócitos
GR- receptor de glicocorticóide
HIOMT- enzima hidroxindol-O-
metiltransferase
IFN-γ- interferon gama
IKB- proteína inibitória kappa B
IKK- proteína quinase de IKB
IL- interleucina
iNOS- sintase de óxido nítrico
induzível
IP3- inositol trifosfato
LPS- lipopolissacarídeo
MAPK- mitogen-activated protein kinase
MEL- melatonina
MTT- brometo de 3-(4,5)-
dimetiltialzolil -2,5 difeniltetrazólio
NA- noradrenalina
NAS- N-acetilserotonina
NEMO- modulador essencial de NFKB

NFKB- fator nuclear kappa B
NLS- sequência de localização nuclear
NSQ- núcleos supraquiasmáticos
PACAP- peptídeo ativador de adenilil
ciclase na pituitária
PAMP- padrões moleculares associados
à patógenos
PBS- phosfate buffer saline
PDTC- pirrolidinaditiocarbamato
PKA- proteína quinase dependente de
AMPc
PKC- proteína quinase dependente de
cálcio
PLC- fosfolipase C
pM, nM, μM- picomolar, nanomolar e
micromolar, respectivamente
PVN- núcleos paraventriculares
RHD- domínio de homologia Rel
RIP1- receptor-interacting protein 1
SNC- sistema nervoso central
SP1- specificity protein 1
TACE- enzima conversora de TNF alfa
TAD- domínio de transativação
TGF-β- fator de crescimento
transformador beta
TLR4- receptor do tipo toll 4
TNF- fator de necrose tumoral
TNF-R- receptor de TNF
TPOH- triptofano hidroxilase
TRAF- TNF receptor-associated factor
μm- micrômetro

____________
Introdução

13 I. Introdução 13 I. Introdução
I. INTRODUÇÃO
1. GLÂNDULA PINEAL
11..11.. HHiissttóórriiccoo ddaa ggllâânndduullaa ppiinneeaall
A identificação da pineal como órgão distinto foi feita entre os séculos III e IV
a.C. por Herófilo. Séculos mais tarde Claudio Galeno (130-200), médico e filósofo grego
caracterizou o tecido localizado no cérebro humano como uma glândula e nomeou-a de
conarium. Este termo em latim (cone) lembra a forma de uma pinha e mais tarde o nome
desta glândula passou a ser pineal (Zrenner, 1985). Já no século XVII, René Descartes
(1596–1650), conhecido por numerosas contribuições matemáticas e filosóficas,
escreveu tratados sobre a concepção do homem e suas duas naturezas: o corpo e a alma.
Sua preocupação baseava-se em como essas duas partes interagiam, ou mesmo se
conectavam. Nessa visão, a pineal aparecia como o elo de ligação física entre o corpo e a
alma. Descartes foi o primeiro a especular que a pineal seria a sede da alma e regularia o
fluxo de espíritos nos animais, controlando funções como os movimentos corporais,
memória e imaginação. O órgão também foi chamado de corpo cervical ou epífise
cerebral, este último nome se refere à localização da pineal acima do cérebro em
contraposição à hipófise que está localizada na sela túrcica, abaixo do cérebro. Assim
como para outros órgãos e estruturas anatômicas a cunhagem do nome da glândula
refletiu o estado do conhecimento e do desenvolvimento tecnológico da época.
Os estudos científicos da glândula pineal tiveram poucos progressos até a
segunda metade do século XIX. No final desse século a anatomia, histologia,
embriologia e inervação da glândula foram extensivamente elucidadas por autores como
Ahlborn, Rabl-Ruckhardt, Graaf, Korschelt e Spencer que, em conjunto, mostraram

14 I. Introdução 14 I. Introdução
semelhanças entre a pineal de mamíferos e a epífise de vertebrados inferiores.
Posteriormente, Studnicka (1905) estabeleceu que a glândula pineal era
filogeneticamente derivada de órgãos fotorreceptores, permanecendo, no entanto, sua
função desconhecida (para revisão, Simonneaux & Ribelayga, 2003).
A primeira função fisiológica atribuída à pineal foi sugerida pela análise de um
caso clínico propondo a relação entre um tumor na pineal e o aparecimento da
puberdade precoce. Essa observação foi feita pelo médico Otto Heubner que relacionou
a alteração gonadal com o tumor da pineal. Vários estudos se seguiram na tentativa de
entender a ação dos hormônios da pineal sobre a fisiologia reprodutiva. Muitos desses
trabalhos avaliaram o efeito dos extratos da pineal sobre o trato reprodutor (Simonnet
et al., 1951; Moszkowska, 1951; Kitay & Altschule, 1954). A associação da pineal com o
sistema reprodutor ficou estabelecida desde então e passou a ser estudada por
diferentes pesquisadores.
No final da década de 1950, um grupo de pesquisadores conseguiu isolar e
caracterizar a partir de extratos de pineal bovina a indolamina N-acetil-5-
metoxitriptamina (Lerner et al., 1959), denominada melatonina por sua habilidade de
promover a agregação de melanóforos da pele de anfíbios (Lerner et al., 1958). Na década
de 1960 é estabelecida a relação entre a luz ambiental e a atividade metabólica da
glândula pineal (Wurtman et al., 1963; 1967, Quay, 1967), sendo evidenciado o ritmo
diário de serotonina (Quay, 1963) e da melatonina em pineais de ratos (Quay, 1964).
Destaca-se a partir dessa concepção o entendimento de que o fotoperíodo seria um fator
determinante na adaptação de várias espécies, regulando a fisiologia e o comportamento
reprodutivo. Nesse sentido, observou-se que a pineal é responsável pela resposta
endócrina às adaptações sazonais (Hoffman & Reiter, 1965a,b). A chegada de dias
curtos ou longos significa respostas diferentes dependendo das espécies estudadas.

15 I. Introdução 15 I. Introdução
Em resumo, pode-se perceber que a busca sobre o conhecimento da glândula
pineal é antiga. A caracterização de sua origem e estrutura e, por fim, o entendimento do
controle de sua função por fatores externos representam contribuições notáveis e
universais sobre a pineal no século XX. Outros aspectos de sua fisiologia e outras ações
de seu hormônio melatonina também tiveram destaque e desde então constituem
diversas linhas de pesquisa dos dias atuais.
11..22.. AAssppeeccttooss MMoorrffooffuunncciioonnaaiiss -- AAnnaattoommiiaa,, HHiissttoollooggiiaa,, IInneerrvvaaççããoo ee
VVaassccuullaarriizzaaççããoo
1.2.1. Estrutura e composição celular
Existe uma grande variação morfológica da glândula pineal ao longo da escala
filogenética tanto intra quanto interespecífica. Anatomicamente, é considerada parte
do epitálamo, situada dorsalmente à região caudal do diencéfalo. Na maioria dos
mamíferos a glândula pineal se localiza na porção dorsal do tronco cerebral conectada
por um pedúnculo às comissuras habenular e posterior (Møller, 1992; Vollrath, 1981).
Em humanos, a pineal desenvolve-se a partir de uma evaginação do teto do terceiro
ventrículo (Oksche, 1965) que inicia-se no segundo mês de gestação e sua localização é
mantida na vida adulta. Em roedores, a pineal migra dorso-caudalmente durante a
ontogênese e se localiza centralmente no topo do cérebro entre os hemisférios cerebrais
e o cerebelo (Vollrath, 1981; Heidbüchel & Vollrath, 1983; 1999; Møller & Baeres, 2002).
O parênquima da pineal de mamíferos consiste em cordões de células separadas
por capilares com grande espaço perivascular. Em espécies de maior porte, o
parênquima é dividido em lóbulos separados por septos de tecido conjuntivo frouxo
derivado da pia-máter. O tipo celular mais abundante na pineal de mamíferos é o
pinealócito, mas também são encontradas células da glia (Korf, 2000, Møeller & Baeres,

16 I. Introdução 16 I. Introdução
2002). Em mamíferos, apesar dos pinealócitos serem células tipicamente endócrinas,
ainda expressam proteínas características de células fotorreceptoras, tais como opsinas
(Korf et al. 1985; Huang et al., 1992), recoverina (Korf et al., 1992), e antígeno S (Korf et al.,
1985, 1990). Em vertebrados não mamíferos, os pinealócitos além de produzirem o
hormônio do escuro, melatonina, também são células fotorreceptoras (Ekstro & Meissl,
2002).
A presença da glia na pineal já foi considerada questionável e o termo célula
intersticial foi utilizado para sua denominação (Wolfe, 1965). Além disso, as células
intersticiais foram tidas como pinealócitos tipo II (Calvo & Boya, 1983). O
desenvolvimento de marcadores químicos específicos para células gliais e os estudos
ultra-estruturais confirmaram a presença peculiar de células gliais típicas no
parênquima da pineal (Luo et al., 1984). Células da região proximal do parênquima da
glândula apresentam imunoreatividade positiva para proteína fibrilar associada à
astrócitos (GFAP) (Møller et al., 1978; López- Muñoz et al., 1992; Suzuki & Kachi, 1995).
Para identificação da microglia na pineal foram utilizados os mesmos marcadores de
macrófagos: ED-1, OX-42 e OX-6 que, em conjunto, revelam a imunoreatividade positiva
em todo parênquima (Tsai & McNulty, 1997; Jiang-Shieh et al., 2003, 2005). Dessa
forma, a glia apresenta localização diferencial, os astrócitos estão localizados próximos
ao pedúnculo pineal, na porção proximal da glândula e a microglia está distribuída de
forma difusa por todo o parênquima da glândula entre os pinealócitos.
1.2.2. Vascularização
A pineal se destaca por sua intensa vascularização, já apontada nos estudos
anatômicos de Andreas Versalius (1515–1564) (para revisão, Stanley, 1994). Ela recebe
suprimento sanguíneo por pequenas ramificações arteriolares provenientes das artérias

17 I. Introdução 17 I. Introdução
coroidais. Por sua vez, cada uma das artérias origina-se da artéria cerebral posterior
(Reiter, 1981). Em ratos, a glândula também se destaca por ter um alto fluxo sanguíneo;
o maior comparado às outras glândulas do organismo (Goldman & Wurtman, 1964).
A pineal é reconhecida como um órgão circumventricular (Johnson & Gross,
1993; Duvernoy & Risold, 2007). Esses órgãos apresentam características morfológicas
que os distinguem do resto do sistema nervoso (Hofer, 1958). São estruturas localizadas
na linha média do cérebro circundando o terceiro e quarto ventrículo e que apresentam
a barreira endotelial mais frouxa devido a ausência de junções oclusivas e a presença de
capilares fenestrados (Johnson & Gross, 1993). Essas características permitem a
comunicação direta entre o sangue e líquor sendo funcionalmente semelhantes a
portões através da barreira hematoencefálica, o que levou a comparação dessas
estruturas a “janelas do cérebro” (Gross, 1987).
1.2.3. Inervação
O padrão de inervação da pineal está diretamente relacionado às diferenças
anatômicas da pineal entre as espécies devido, principalmente, à posição da pineal em
relação ao cérebro. A glândula pineal recebe uma inervação complexa com fibras
nervosas de diferentes origens. A principal fonte consiste em fibras pós-ganglionares
simpáticas provenientes do gânglio cervical superior que chegam até a pineal através do
nervo conário (Bargmann, 1943; Kappers, 1960). Está é a inervação mais compreendida e
estudada em humanos e em outros mamíferos. A atividade destas fibras pós-
ganglionares é regulada pelos núcleos supraquiasmáticos do hipotálamo (NSQ) que,
por sua vez, recebem informações diretamente de células ganglionares retinianas via
trato retino-hipotalâmico (Kappers, 1960). As terminações nervosas simpáticas contêm,
além do neurotransmissor noradrenalina, os neuromoduladores neuropeptídeo Y (NPY)

18 I. Introdução 18 I. Introdução
(Schon et al., 1985; Zhang et al., 1991; Mikkelsen & Møller, 1999) e ATP (adenosina tri-
fosfato) (Mortani-Barbosa et al., 2000). A noradrenalina tem grande importância
funcional pois é o estímulo indutor da síntese de melatonina (Klein & Weller., 1970)
enquanto o ATP e o NPY atuam como co-transmissores e regulam a estimulação
adrenérgica. O ATP é co-liberado com a noradrenalina pelos terminais simpáticos e
interage com receptores do subtipo P2Y1 (Ferreira et al., 2001) potencializando a síntese
de melatonina induzida por noradrenalina (Ferreira et al., 2003). Já o NPY modula a
transmissão noradrenérgica por mecanismos pré e pós-sinápticos. A estimulação com
NPY de receptores pós-sinápticos inibe a liberação de noradrenalina (Vacas et al., 1987;
Simonneaux et al., 1994) e a formação de AMPc (Olcese, 1991; Harada et al., 1992;
Simonneaux et al., 1994). Por outro lado, embora haja redução de AMPc, o NPY mostrou
exercer efeito positivo sobre a síntese de melatonina em glândula pineal de rato (Vacas
et al., 1987, Simonneaux et al., 1994).
As técnicas neuroanatômicas mais modernas como o traçamento neuronal, a
imuno-histoquímica e a hibridização in situ tiveram grande destaque no entendimento
da inervação não simpática. Os primeiros estudos ultra-estruturais da pineal já
sugeriam a presença de fibras extra-simpáticas pela presença de botões sinápticos
contendo vesículas granulares do tipo peptidérgica e colinérgica. Outra importante
evidência anatômica foi obtida pela demonstração de que após a remoção do gânglio
cervical superior diversas fibras peptidérgicas permaneciam na pineal (Møller et al.
1996). Da mesma forma, o uso da técnica de traçamento viral confirmou a existência de
conexões neurais entre algumas áreas cerebrais e a pineal em diversas espécies de
roedores (Møller, 1999; Møller & Baeres, 2002).
As fibras centrais derivam de diferentes regiões encefálicas, especialmente do
núcleo paraventricular hipotalâmico (PVN) (Korf & Wagner 1980; Møller et al., 1990,

19 I. Introdução 19 I. Introdução
Larsen et al., 1991), folheto intergeniculado talâmico (IGL) (Mikkelsen & Møller, 1990;
Cipolla et al., 1994) e núcleo dorsal da rafe (Leander et al. 1998; Møller & Hay-Schmidt,
1998). Estas projeções terminam, em sua maior parte, na porção proximal da glândula
através das comissuras habenular e posterior (Kappers, 1960, Møller, 1978) e do
pedúnculo pineal. Também estão presentes inervações de outras áreas cerebrais e
periféricas que contribuem para liberação de uma variedade de neuropeptídeos, a citar:
substância P, PACAP (do inglês, pituitary adenylate cyclase-activating peptide), CGRP (do
inglês, calcitonin gene-related peptide), peptídeo intestinal vasoativo (VIP) e ácido gama-
aminobutírico (GABA). Em resumo, a inervação autonômica simpática e
parassimpática, bem como inervação direta proveniente do sistema nervoso central,
projetam-se sobre a pineal. Destas, o efeito funcional da inervação simpática é a mais
conhecida e estudada.
11..33.. PPrroodduuççããoo ddee mmeellaattoonniinnaa ppeellaa ggllâânndduullaa ppiinneeaall
1.3.1. Controle neural
As mudanças diárias nas condições de iluminação ambiental geram impactos em
vários aspectos da vida. Como resposta adaptativa vários processos fisiológicos e
comportamentais são rítmicos. Alguns são meramente reativos a essas condições
externas e outros são antecipatórios, controlados pelo relógio biológico interno. Em
mamíferos, o relógio biológico central é localizado nos núcleos supraquiasmáticos
(NSQ) do hipotálamo anterior. Estes núcleos geram um ritmo endógeno circadiano com
período muito próximo a 24 horas (para revisão, Reppert & Weaver, 2002). Os núcleos
supraquiasmáticos têm papel essencial na variação diária da produção de melatonina,

20 I. Introdução 20 I. Introdução
gerando o ritmo e propagando a resposta à luz detectada pela retina (para revisão
Moore, 1997; Kennaway & Wright, 2002).
A informação luminosa é percebida por meio do fotorreceptor melanopsina
presente nas células ganglionares retinianas, transformada em um sinal elétrico e
transmitida pelo trato retino-hipotalâmico aos NSQ (Provencio et al., 2000). Estes,
através de uma via polissináptica, projetam-se para o núcleo paraventricular do
hipotálamo (PVN) promovendo a liberação de GABA (Kalsbeek et al., 1999, 2000).
Portanto, os NSQ controlam o ritmo da produção de melatonina por imposição de um
sinal inibitório GABAérgico sobre o PVN durante a fase de claro, inibindo assim, a
produção de melatonina pela pineal neste ponto da via (Teclemariam-Mesbah et al.,
1999). Na fase escura do dia, o PVN é ativado a partir dos NSQ por uma via
glutamatérgica que promove a ativação de uma via polineural que chega à coluna
intermediolateral da medula espinal. Fibras pré-ganglionares simpáticas projetam-se ao
gânglio cervical superior que, por sua vez, emite neurônios pós-ganglionares simpáticos
que inervam diretamente a pineal e liberam noradrenalina (Wurtman et al., 1967) e ATP
(Mortani-Barbosa, 2000) induzindo a síntese de melatonina (revisto por Simonneaux e
Ribelayga, 2003; Perreau-Lenz et al., 2003).
1.3.2. Via biossintética da melatonina
Como mencionado anteriormente, na ausência de iluminação ambiental as fibras
simpáticas que inervam a pineal liberam os co-transmissores noradrenalina e ATP
(Mortani-Barbosa et al., 2000). A noradrenalina liberada pelos terminais simpáticos
interage com receptores pós-sinápticos dos tipos α1 e β1-adrenérgicos presentes na
membrana dos pinealócitos e o ATP promove a ativação de receptores P2Y1( Ferreira et
al., 1994, 2001). A ativação dos adrenoceptores β é essencial para a indução da síntese de

21 I. Introdução 21 I. Introdução
melatonina enquanto que a estimulação de adrenoceptores alfa, ou de receptores
purinérgicos P2Y1 potencia a resposta desencadeada por ativação dos adrenoceptores β
(Mortani-Barbosa, 2000; Klein et al., 1983).
A ativação dos receptores β1 induz a ativação da proteína G estimulatória (Gs)
que, por sua vez, ativa a enzima adenilil ciclase (AC) promovendo a formação do
segundo mensageiro monofosfato de adenosina cíclico (AMPc). O aumento de AMPc
ativa a proteína quinase dependente de AMPc (PKA) induzindo a transcrição do gene
(roedores) e/ou potenciando a atividade (roedores, ungulados e primatas) da enzima
arilalquilamina N-acetiltransferase (AA-NAT). Este é um dos passos chave para a
síntese de melatonina pois resulta no padrão de atividade rítmica desta enzima
controlado pelo ciclo claro-escuro (Klein & Weller, 1970).
Os receptores alfa adrenérgicos são acoplados à proteína Gq que, ao se ligarem a
noradrenalina, ativam a enzima fosfolipase C (PLC) promovendo aumento do cálcio
intracelular (Ca+2) via inositol trifosfato (IP3) e, consequentemente, a ativação da
proteína quinase dependente de cálcio (PKC) (Sugden et al., 1985; Vanecek et al., 1985). A
co-transmissão purinérgica também atua por mecanismo semelhante ao induzir a
ativação da enzima PLC e consequente elevação dos níveis de cálcio (Ferreira et al., 2001,
2003). Dessa forma, a estimulação dos receptores α1 e P2Y1 convergem com a resposta
β-adrenérgica por promoverem a regulação positiva da PKC sobre a adenilil ciclase e
portanto potencializando a via de sinalização necessária para transcrição da AA-NAT.
As cascatas de sinalização estão representadas em forma esquemática na figura 1.
A síntese de melatonina é iniciada pela captação do aminoácido triptofano da
corrente sanguínea. Esse precursor é hidroxilado a 5-hidroxitriptofano pela enzima
triptofano hidroxilase e é, em seguida, descarboxilado formando serotonina (5-
hidroxitriptamina, 5-HT). A serotonina é acetilada pela enzima AA-NAT à N-

22 I. Introdução 22 I. Introdução
acetilserotonina que é, por fim, convertida em melatonina (N-acetil-5-
metoxitriptamina) pela enzima hidroxindol-O-metiltransferase (HIOMT) (Axerold &
Weissbach, 1960).
1.3.3. Regulação transcricional e pós-transcricional da enzima AA-NAT
Existem diferenças pontuais na regulação da via biossintética de melatonina
entre as espécies de vertebrados. Em comum, a parte final da via neural consiste na
liberação circadiana de noradrenalina sobre a glândula exclusivamente à noite. A síntese
de melatonina é melhor estudada em roedores onde sua regulação ocorre no nível
transcricional. Nessas espécies, a resposta beta-adrenérgica na glândula promove a
ativação da via de transcrição AMP cíclico- proteína quinase A (PKA). A PKA fosforila
o fator de transcrição CREB (do inglês, cyclic-AMP regulating element binding) e a enzima
AA-NAT. CREB fosforilado se liga ao elemento CRE (do inglês, cAMP-responsive element)
do DNA no promotor do gene da AA-NAT promovendo a transcrição gênica. A
fosforilação da AA-NAT induz a sua interação com a chaperona 14-3-3 (Ganguly et al.,
2001, 2002) que impede sua degradação pelo proteassoma (Gastel et al., 1998; Ganguly et
al., 2001). Tanto o gene quanto a proteína da AA-NAT apresentam um ritmo circadiano
com níveis de transcrição 100-150 vezes maior durante a noite (Borjigin et al., 1995;
Roseboom et al., 1996; Garidou et al., 2001).
Em ungulados e primatas os níveis do RNAm do gene da AA-NAT não
apresentam diferenças significativas entre o dia e noite (Coon et al., 1995; Schomerus et
al., 2000; Johnston et al., 2004) indicando a expressão constitutiva deste gene e
sugerindo que a regulação da atividade da AA-NAT seja pós-transcricional. Durante o
dia, a proteína, assim que traduzida, é rapidamente degradada por atividade
proteassomal (Korf et al., 1998; Stehle et al., 2001). Na fase de escuro a ativação da via

23 I. Introdução 23 I. Introdução
AMPc/PKA (para revisão Ganguly et al., 2002), a fosforilação da AA-NAT e a
consequente ligação com a chaperona 14-3-3 impedem sua degradação. Como já descrito
anteriormente, a produção circadiana de melatonina é gerada pelo ritmo de
atividade/transcrição da enzima AA-NAT. Já a enzima HIOMT não apresenta ritmo
diário evidente sendo sua atividade regulada a longo termo (Axelrod et al., 1965; Sugden
& Klein, 1983,a.b; Ribelayga et al., 1997) e participando da resposta sazonal da síntese de
melatonina (Ribelayga et al., 1999, 2000).

24 I. Introdução 24 I. Introdução
FIGURA 1. VViiaass ddee ssiinnaalliizzaaççããoo ddaa bbiioossssíínntteessee ddee mmeellaattoonniinnaa nnaa ppiinneeaall ddee rraattooss.. No escuro as
fibras pós-ganglionares simpáticas liberam noradrenalina (NA) ativando os receptores β e α-
adrenérgicos. A ativação dos receptores β1 promove a ativação da adenilil ciclase (AC) que
resulta no aumento dos níveis de AMPc, ativação da proteína PKA e fosforilação o fator de
transcrição CREB, que induz a transcrição da AA-NAT (indicado nas setas verdes). A ativação
dos receptores α1 pela noradrenalina e P2Y1 pelo co-transmissor ATP induz cascatas de
sinalização que convergem com a resposta β-adrenérgica (setas vermelhas). Estas vias levam ao
aumento dos níveis de cálcio e a consequente ativação da PKC que, por sua vez, potencia a
atividade da enzima AC. A PKA também fosforila a enzima AA-NAT após sua tradução,
permitindo sua ligação à chaperona 14-3-3 e sua estabilização e ativação. AA-NAT é o fator
chave para síntese de melatonina, pois a via só prossegue após a sua tradução, como
esquematizado na parte direita da figura.

25 I. Introdução 25 I. Introdução
11..44.. DDaa sseeccrreeççããoo aaooss mmeeccaanniissmmooss ddee aaççããoo ddaa mmeellaattoonniinnaa
A melatonina é uma molécula altamente lipofílica. Ao invés de ser armazenada
na pineal após sua síntese, ela é liberada nos capilares se ligando à albumina (Cardinali
et al., 1972). A melatonina secretada tem meia vida curta na circulação de
aproximadamente 30 minutos, dependendo da espécie examinada (Cardinali et al., 1972;
Waldhauser et al., 1984). A melatonina é metabolizada primariamente no fígado onde
sofre uma reação de duas etapas, primeiro é hidroxilada, seguida da conjugação com
sulfato ou glucoronato pelo citocromo p450 formando 6-sulfatoximelatonina (para
revisão, Tan et al., 2007). Estudos recentes apontam que esse metabólito também pode
ser formando em outros locais além do fígado como o córtex cerebral, os rins e o coração
de ratos (Lahiri et al., 2004). O perfil desse metabólito na urina e no plasma reflete tanto
aspectos quantitativos como qualitativos da secreção de melatonina e serve para análise
do ritmo endógeno de melatonina (Arendt, 1986; Bojkowskl, 1987).
Por ser uma molécula lipofílica a melatonina pode atravessar passivamente a
membrana celular e dessa forma, além de atuar sobre receptores, pode regular
diretamente reações/funções no interior das células independentemente da interação
com receptores. Por outro lado, diversas ações da melatonina são mediadas por
receptores de membrana MT1 e MT2 (Reppert et al., 1994, 1995; Dubocovich, 1988;
1997). Esses receptores pertencem a família de receptores acoplados a proteína G, que
contém sete domínios transmembrânicos e são responsáveis principalmente pelos
efeitos cronobiológicos sobre o NSQ. Neste caso, o receptor MT2 atua induzindo o
deslocamento de fase enquanto o receptor MT1 age inibindo a atividade de disparo
neuronal. Esses efeitos são mediados por concentrações equivalentes as encontradas no
plasma (pM a nM) durante a noite (para revisão, Dubocovich et al., 2010).

26 I. Introdução 26 I. Introdução
Um terceiro receptor, inicialmente descrito como MT3 (Dubocovich, 1988) foi
caracterizado posteriormente como sendo a enzima quinona redutase II (Nosjean et al.,
2000). Esse receptor ainda não foi clonado e por isso e de acordo o comitê de
nomenclatura IUPHAR é grafado em itálico (Dubocovich et al., 2000). A quinona
redutase participa da proteção contra o estresse oxidativo evitando reações de
transferência de elétrons de quinonas (Foster et al., 2000). A melatonina também se liga
com menor afinidade à calmodulina (Benitez-King, 2006) assim como a receptores
nucleares da família do ácido retinóico RORa1, RORa2 e RZRb (Wiesenberg et al., 1995;
Carlberg, 2000). Parte das ações antioxidantes da melatonina são mediadas por
mecanismos dependentes de receptor, mas também incluem ações que não dependem
dessa interação constituindo da ligação direta a radicais livres e reações de transferência
de elétrons na cadeia respiratória (Poeggeler et al., 2002).
2. FATOR DE NECROSE TUMORAL (TNF)
22..11.. CCaarraacctteerriizzaaççããoo ddoo TTNNFF
O TNF também descrito como caquexina foi identificado em 1975 como a
proteína induzida por uma endotoxina e que causava a necrose hemorrágica de
sarcomas transplantados em camundongos o que resultou na denominação de “fator de
necrose tumoral” (Carswell et al., 1975). Este termo já foi utilizado para denominar duas
proteínas diferentes, TNF- α e TNF-β, mas considerando suas especificidades, em 1998
chegou-se ao consenso que TNF- α e TNF- β seriam substituídos por TNF e linfotoxina
α (LTF α), respectivamente (Tracey, 2008). No presente texto utilizaremos a
nomenclatura acordada em 1998.

27 I. Introdução 27 I. Introdução
Após sua descoberta o grande interesse sobre o TNF foi baseado na expectativa
de seu potencial como uma droga anti-cancer. Mais tarde ficou evidente que o TNF é
uma citocina altamente pleiotrópica mediando diversas atividades biológicas. Foi
originalmente identificado como o mediador humoral da resposta endotóxica sistêmica
que causava febre, hipotensão e choque (Cerami & Beutler, 1988, Tracey et al., 1988). Os
estudos desde então revelaram também que o TNF é um dos mediadores inflamatórios
mais proeminentes para o início das reações inflamatórias do sistema imune inato,
incluindo a indução da produção de outras citocinas, a ativação ou expressão de
moléculas de adesão e multiplicação celular (Wajant., 2003). A importância do papel do
TNF durante o processo inflamatório pode ser demonstrada pela perspectiva
encontrada nas terapias anti-TNF que baseiam-se na utilização de anticorpos anti-TNF
ou administração de receptores solúveis de TNF no controle da artrite reumatóide e em
outras condições inflamatórias (Mpofu et al., 2005; Tracey, 2008).
O TNF é produzido primariamente como uma proteína de 26 kDa conhecida
como pró-TNF (Kriegler et al., 1988), que é expressa na membrana plasmática onde pode
ser clivada no seu domínio extracelular pela metaloproteinase TACE (do inglês, TNF
alpha converting enzyme) (Black et al., 1997; Moss et al., 1997) resultando na forma solúvel de
TNF (17 kDa). Tanto a forma associada à membrana como a forma solúvel de TNF são
ativas apenas na conformação trimérica e possuem distintas atividades biológicas.
Diversos tipos celulares linfóides ou não podem produzir TNF, incluindo macrófagos,
células T, mastócitos, granulócitos, natural killer (NK), fibroblastos, neurônios,
queratinócitos e células musculares (revisto por Bradley, 2008). Inclusive a própria
pineal produz TNF em resposta a estimulação com lipopolissacarídeo (LPS) (da Silveira
Cruz-Machado et al., 2010).

28 I. Introdução 28 I. Introdução
O TNF não é detectável em indivíduos saudáveis, mas os níveis séricos e
teciduais são elevados em condições inflamatórias e infecciosas, e se correlacionam com
a severidade das infecções (Nurnberger et al., 1995; Waage et al., 1997; Robak et al., 1998).
22..22.. SSiinnaalliizzaaççããoo ddoo TTNNFF
Os efeitos do TNF são mediados por dois tipos de receptores, designados como
receptor de TNF tipo 1 (TNF-R1) (descrito também como TNFRSF1A, p55/60, CD120a)
e receptor de TNF tipo 2 (TNF-R2) (descrito também como TNFRSF1B p75/80,
CD120b) (Ledgerwood, 1999). Enquanto os dois receptores compartilham similaridades
no domínio extracelular que consiste em repetições de cisteína (Beutler & van Huffel,
1994), TNF-R1 e TNF-R2 diferem no domínio intracelular, na afinidade a ligantes, no
perfil de expressão celular, na cascata de sinalização e consequentemente na sua função.
As duas formas de TNF, tanto membranar (TNFm) quanto a solúvel (TNFs), podem se
ligar aos dois tipos de receptores, no entanto, algumas ligações podem ser mais
favorecidas que outras. O TNFs se liga preferencialmente ao TNF-R1 (constante de
dissociação [Kd]∼20 pM) do que ao TNFR2 (Kd∼400 pM). (Greel et al, 1998). O TNF-
R1 é constitutivamente expresso na maioria dos tecidos enquanto a expressão do TNF-
R2 é restrita, tipicamente encontrada nas células do sistema imune. Por isso, na maior
parte das células, o TNF-R1 é o principal mediador da sinalização do TNF (Thoma et al.,
1990)
Alguns dos membros da família do TNF são conhecidos por sua capacidade de
induzir a morte celular por ativar a via de sinalização apoptótica. Esses receptores,
incluindo o TNF-R1, têm em comum o chamado domínio de morte (do inglês death
domain) (DD), presente na porção intracelular da molécula (Tartaglia et al., 1993a;
Schulze-Osthoff et al., 1998) e essencial para induzir a apoptose. Em receptores não

29 I. Introdução 29 I. Introdução
estimulados, o domínio citoplasmático é pré-associado à proteína inibitória SODD (do
inglês, silencer of death domains) com o domínio de morte. Essa proteína evita a sinalização
constitutiva do TNF-R1. A ligação do TNF ao domínio extracelular do receptor induz a
dissociação da proteína inibitória SODD com o DD que fica livre para ligar-se à proteína
adaptadora TRADD (Chen et al, 2002). Esse novo complexo protéico serve de
plataforma para ligação de diferentes proteínas que contém o DD. A via de sinalização a
ser disparada pela interação entre TNF e TNF-R1 depende de qual proteína se acoplará
à TRADD (do inglês, TNFRSF1A-Associated via death domain). TRADD inicia a sinalização
recrutando duas proteínas adicionais RIP-1 (do inglês, receptor interacting protein-1), uma
serina/treonina quinase que se liga ao TRADD por seu próprio domínio de morte (Hsu
et al., 1996) permitindo a ligação ao TRAF2 (do inglês, TNFR-associated factor 2) que não
possui DD (Takeuchi et al., 1996). O complexo TRADD-RIP1-TRAF2 é internalizado
(Jones et al., 1999) e liberado do TNF-R1. A formação desse complexo resulta na ativação
de diferentes quinases: NIK (do inglês, NFKB inducing kinase), IKKα e β (do inglês, IKB
kinase α e β) (Mercurio & Manning, 1999) ou MAP quinases (Eder, 1997), ERK, JNK e
p38 quinase (Liu et al., 1996; Kelliher et al., 1998). A ativação dessas quinases promove
tanto a sobrevivência celular quanto a geração de sinais pró-inflamatórios através da
ativação do fatores de transcrição, fator nuclear kappa B (NFKB) e AP-1 (do inglês,
activator protein 1). O TNF-R1, como já abordado anteriormente, pode ainda sinalizar por
vias que levam a morte celular. Essas vias envolvem a ligação da proteína FADD (do
inglês, Fas-associated DD protein) ao TRADD e o recrutamento subsequente da pro-caspase
8 que inicia a apoptose através da clivagem e ativação da pro-caspase 3. O recrutamento
e a ativação da pro-caspase 8 pode ser inibido por cFLIP (do inglês, Cellular caspase-8

30 I. Introdução 30 I. Introdução
(FLICE)-like inhibitory protein) induzido pela via NFKB, inibindo a via apoptótica. Esta
sinalização pode ser resumida no esquema representado na figura 2.
As vias de sinalização ativadas pelo TNF-R2 são menos definidas do que aquelas
envolvidas na sinalização do TNF-R1. Sem ter o domínio de morte a interação mais
relevante após a ativação do TNF-R2 é o recrutamento de TRAF2 que promove a
ativação da via do NFKB. A ativação do TNF-R2 resulta na expressão de citocinas e
também de proteínas intracelulares regulatórias com potencial anti-apoptótico, tais
como TRAF1, TRAF2 e proteínas inibidoras de apoptose c-IAP1 e c-IAP2, (Wang et al.,
1998) além de também poder ocorrer a ativação das MAP quinases p38 e JNK (Winston
et al., 1995; Liu et al., 1996, Kelliher et al., 1998; Lee et al., 2003).
A utilização de diferentes mecanismos de sinalização pelos receptores de TNF
condiz com a habilidade de cada receptor em sinalizar uma reposta biológica distinta. A
ligação do TNF-R1 é necessária e suficiente para induzir tanto efeitos citotóxicos como
respostas pró-inflamatórias, enquanto que o TNF-R2 ativa vias inflamatórias e de
sobrevivência celular (Rothe et al., 1994, 1995). Em algumas circunstâncias as vias
podem convergir, o TNF-R2 pode promover a sinalização do TNF-R1 por aumentar a
associação do TNFs com o TNF-R1 através de um mecanismo de passagem de ligante
que ocorre, principalmente, em baixas concentrações (Slowik et al., 1993); nestes casos o
TNF-R2 captura o TNFs passando-o ao TNF-R1 (Tartaglia et al., 1993b).

31 I. Introdução 31 I. Introdução
FIGURA 2. RReepprreesseennttaaççããoo eessqquueemmááttiiccaa ddooss ccoommppoonneenntteess ddaa vviiaa ddee ssiinnaalliizzaaççããoo ddoo TTNNFF.. O TNF
possui dos tipos de receptores, TNF-R1 e TNF-R2 e no esquema são representadas as vias de
ativação desencadeadas por cada um. A ligação do TNF ao TNF-R1 resulta no recrutamento de
TRAF2, RIP1 e FADD através da proteína adaptadora TRADD. FADD recruta e ativa a caspase
8, levando à apoptose. TRAF2 e RIP1 levam à ativação dos fatores de transcrição AP-1 e NFKB
através da ativação das proteínas quinases JNK, p38 e IKK. A ligação do TNF ao TNF-R2 ativa o
recrutamento de TRAF1, 2 e 3 que, através da ligação a proteína RIP-1, pode tanto levar a
cascata da ativação do NFKB, via IKKs, ou do fator de transcrição AP-1, por meio das MAPKs
(Baseado em Baud & Karin, 2001; Wajant, 2003).

32 I. Introdução 32 I. Introdução
Muitas linhagens celulares co-expressam os dois receptores, embora o TNF-R2
esteja preferencialmente expresso em células hematopoiéticas. Esses receptores
apresentam um padrão de expressão mais limitado in vivo, sendo regulado pelas
condições fisiopatológicas do tecido. Em condições de injúria ou isquemia por exemplo,
a expressão desses receptores pode mudar de intensidade e localização variando
conforme o tecido. Esse tipo de variação resulta de mudanças no nível da síntese desses
receptores bem como na liberação desses receptores da membrana. Alguns estímulos
que incluem o próprio TNF aumentam a expressão do TNF-R2 através de ativação
transcricional enquanto que o TNF-R1 sofre comumente uma regulação negativa
(Winzen et al., 1992; Kalthoff et al., 1993).
3. FATOR DE TRANSCRIÇÃO NUCLEAR KAPPA B (NFKB)
O NFKB foi inicialmente descrito como fator regulador do gene que transcrevia
a cadeia leve kappa da imunoglobulina de linfócitos B (Sen & Baltimore, 1986) o que
resultou no termo fator nuclear kappa B. Desde essa descoberta houve grande interesse
em entender os estímulos que o ativavam, os diversos genes e as funções biológicas que
esse fator regula. A função mais relevante do NFKB foi demonstrada no sistema imune
onde a expressão dos genes regulados por ele é essencial para as respostas da imunidade
inata e adquirida (Li et al., 2002; Bonizzi & Karin, 2004).
Originalmente descoberto no núcleo de linfócitos (Sen & Baltimore, 1986), o
fator nuclear kappa B é expresso de maneira ubíqua em todos os tipos celulares. A
família de fatores de transcrição NFKB inclui um grupo de proteínas conservadas desde
a Drosófila até o homem e que possuem em comum uma sequência de 300 aminoácidos
denominada domínio de homologia Rel (RHD, do inglês, Rel homology domain)
(Hoffmann et al., 1999). Esse domínio que nomeia o grupo, também conhecido como

33 I. Introdução 33 I. Introdução
família Rel, contem regiões funcionais cruciais localizadas na porção N-terminal da
proteína , responsável pela ligação ao DNA, dimerização, interação com as proteínas
inibitórias IKBs (também conhecidas como NFKBI) e direcionamento nuclear.
A família Rel consiste em cinco genes Nfkb1, Nfkb2, Rela, C-rel e Relb os quais
codificam sete proteínas: p105, p50 (ou NFKB1), p100, p52 (ou NFKB2), p65 (ou RelA),
c-Rel e RelB. p50 é gerada através da clivagem de p105, e a p52 a partir da proteína p100.
Essas proteínas são funcionais como dímeros e podem se apresentar como homodímeros
ou heterodímeros produzindo 15 possíveis combinações de dímeros. Neste trabalho será
utilizado a nova nomenclatura para a família NFKB assim como para as proteínas
regulatórias, de acordo com HUGO Gene Nomenclature Committe at the European
Bioinformatics Institute (disponível em http //www.genenam es.org).
As subunidades do NFKB ainda apresentam diferenças importantes em sua
expressão, regulação e funcionalidade. As proteínas RelA, c-Rel e RelB possuem o
domínio de transativação C-terminal (TAD, do inglês, transcription activation domain)
necessário para transcrição gênica sendo, portanto, consideradas as subunidades
ativadoras de expressão gênica. Já as subunidades p52 e p50 não tem o domínio TAD e
por isso são por definição repressoras da transcrição gênica. Por isso, a ligação dos
homodímeros dessas subunidades, a exemplo de p50/p50, ao sítios kappa B leva a
repressão da expressão gênica. Essas subunidades dependem da interação com outros
fatores para regular positivamente a transcrição (Hayden & Ghosh, 2008). Por isso,
heterodímeros de p50 ou p52 com as demais subunidades onde o TAD é presente atuam
como ativadoras da expressão gênica. A expressão dessas moléculas também varia em
relação ao tipo celular. As subunidades RelA e p50 estão presentes praticamente em
todos tipos celulares estudados, enquanto c-Rel, RelB e p52 tem expressão restrita a
células hematopoiéticas (revisto por Liou & Hsia, 2003).

34 I. Introdução 34 I. Introdução
RelB preferencialmente forma heterodímeros com p100 assim que é processada
em p52 (Dobrzanski et al., 1995) enquanto que RelA e c-Rel formam
predominantemente heretodímeros com p50 (Karin & Ben-Neriah, 2000).
Diferentemente dos outros membros RelB possui um domínio chamado zíper de leucina
(LZ) na porção N-terminal que tem função transcricional importante. Esse domínio
também confere especificidade adicional a RelB, incluindo sua heteromerização com
p52. Existem também modificações pós-transcricionais em partes diferentes das
subunidades de NFKB que incluem fosforilações e acetilações que modulam a ligação do
DNA e atividade transcricional (Perkins, 2006).
33..11.. AAttiivvaaççããoo ddoo NNFFKKBB
Em células não-estimuladas os dímeros do NFKB são retidos no citoplasma em
virtude de sua associação com a família de proteínas inibitórias IKB (Karin & Neriah,
2000). A interação com as proteínas IKB mascara a sequência de localização nuclear
(NLS, do inglês, nuclear localization sequence) do NFKB impedindo sua translocação ao
núcleo (Hayden & Ghosh, 2004). A família IKB consiste em nove membros IKBα, iKBβ,
BCL-3, iKBε e iKBγ além das proteínas precursoras p100 e p105 que possuem em
comum repetições múltiplas de anquirinas que medeiam a ligação com os dímeros do
NFKB (Hatada et al., 1992).
Os IKBs têm estrutura similar mas possuem diferenças em relação a sua
preferência de ligação e sua regulação transcricional pelos membros do NFKB. O
heterodímero clássico RelA/p50 é majoritariamente regulado pelo IKBα. Já o iKBε
regula preferencialmente os dímeros RelA/RelA e c-Rel/RelA (para revisão, Whiteside
& Israel, 1997). O papel do IKBβ é menos conhecido mas também se associa a RelA/p50.
Além de diferentes perfis de ligação aos dímeros, as três formas principais de IKB tem

35 I. Introdução 35 I. Introdução
diferentes cinéticas (Hoffmann et al., 2002). A proteína IKBα, por exemplo, é degradada
mais rapidamente em resposta a estímulos como o próprio TNF ou sua síntese.
Muitos dos sinais extracelulares que resultam na ativação da via de transcrição
NFKB disparam vias de transdução de sinal que culminam na ativação de um mesmo
alvo molecular. Essas vias se iniciam principalmente a partir da ativação de receptores
como o TNF-R1, cada qual com seus mecanismos intrínsecos de regulação, mas
convergindo para fosforilação das quinases IKKs (do inglês, iKB kinase).
Consequentemente, esse é um passo importante de regulação para determinar a
resposta de ativação do NFKB a vários estímulos. As IKKS consistem de um complexo
trimérico de quinases que compreende as subunidades catalíticas IKKα, IKKβ e a
subunidade IKKγ ,também conhecida como NEMO (do inglês, NFKB essential modulator),
que tem papel estrutural e regulatório. As proteínas das famílias NFKB e IKB e o
complexo de IKKs estão representadas na figura 3.

36 I. Introdução 36 I. Introdução
Família Rel
Família IKB
Complexo IKK
FIGURA 3. OOss mmeemmbbrrooss ddaass ffaammíílliiaass NNFFKKBB,, IIKKBB ee IIKKKK. A) Membros da família NFKB: RelA
(p65), RelB, c-Rel, p50/p105 (NFKB1) e p52/p100 (NFKB2). As subunidades p50 e p52 (não
mostrada) são derivadas do precursor mais proteínas P105 e P100, respectivamente. Todos os
membros da família NFKB contêm um domínio N-terminal Rel homologia (RHD) que medeia a
ligação do DNA e a dimerização e contém o domínio de localização nuclear. As proteínas RelA,
RelB e c-Rel, contêm na porção C-terminal o domínio de ativação transcricional (TAD). B) As
proteínas inibitórias IKBs consistem em IKBα, IKBβ, IKBε e BCL-3 mais as proteínas da família
NFKB, p105 e p100, que possuem o domínio de repetição de anquirinas (ANK) na porção C-
terminal. C) As três subunidades do complexo IKKs são mostradas: as subunidades catalíticas
IKKα e IKKβ e a subunidade reguladora NEMO, também conhecido como IKKγ. DD, região de
homologia com um domínio de morte; HLH, hélice-alça-hélice; LZ, domínio de zíper de leucina;
NDB, NEMO-binding domain obrigatórios; PEST, domínio rico em prolina.

37 I. Introdução 37 I. Introdução
O TNF, a IL-1β e o LPS disparam a via de sinalização do NFKB chamada clássica
ou canônica que culmina com a translocação nuclear do dímero RelA/p50. O
acoplamento do receptor com seus ligantes resulta na ativação do complexo de IKKs
que fosforila o IKBα nas serinas 32 e 36 promovendo sua poliubiquitinação. Quando
ubiquitinado o IKBα é degradado pelo proteassoma 26S expondo,portanto, o sítio NSL
de RelA e induzindo a translocação nuclear do dímero RelA/p50 (para revisão,
Rothwarf & Karin 1999; Ghosh & Karin, 2002). O complexo IKK também fosforila os
TADs de RelA e c-Rel podendo aumentar a sua atividade transcricional (Perkins, 2006).
Na via clássica IKKβ é a subunidade catalítica responsável pela atividade de IKK na
maioria dos tipos celulares. A deleção de IKKα em células que expressam IKKβ não
afeta a atividade clássica de IKK (Hu et al., 1999). A via alternativa depende somente da
subunidade IKKα a qual fosforila p100 induzindo o processamento de p52 e
subsequente ativação de RelB/p52. Essa via é ativada em resposta a uma parte dos
ligantes da família TNF incluindo CD40l, LTαβ, BAFF, RANKL e TWAK, e sua
regulação se difere da via clássica.
A rápida ativação do NFKB é essencial para uma resposta imune eficiente, mas
essa resposta não pode perdurar e precisa ser finalizada no tempo apropriado para
evitar o dano tecidual ou maiores complicações. O processo inflamatório descontrolado
é indesejado, além de aumentar o risco de câncer e doenças auto-imunes. O controle da
ativação do NFKB é feito por dois mecanismos principais. Um deles constitui de uma
alça de retroalimentação negativa que evolve a síntese de novas proteínas iKBs. Assim
que os dímeros de NFKB translocam ao núcleo um dos genes alvo é o gene Ikbα que
codifica o IKBα (Sun et al., 1993). Os novos IKBα sintetizados são direcionados ao
núcleo como monômeros e se associam aos dímeros p50/RelA levando à sua inativação e
exportação para o citoplasma. Outro mecanismo que contribui para a finalização da

38 I. Introdução 38 I. Introdução
resposta do NFKB é a degradação das subunidades de NFKB ligadas ao DNA pelo
proteassoma (para revisão, Vallabhapurapu & Karin, 2009).
4. EIXO-IMUNE-PINEAL
44..11.. MMeellaattoonniinnaa ee ddeeffeessaa
A melatonina, originalmente descrita como hormônio da glândula pineal,
também é produzida por outras estruturas. Esta molécula, altamente conservada ao
longo da escala filogenética, atuava inicialmente como antioxidante e bloqueadora de
processos enzimáticos relacionados com o estresse oxidativo sendo a função de defesa a
principal ação da melatonina em organismos mais primitivos (Hardeland & Fuhrberg,
1996, Reiter et al., 2000). Em vertebrados, a melatonina acumula funções de defesa com
função cronobiótica, isto é, marcadora do escuro (Hardeland & Fuhrberg, 2006; Tan et
al., 2010).
Ao longo das últimas duas décadas um grupo substancial de pesquisadores tem
demonstrado o papel regulatório da melatonina sobre as funções de defesa (Nelson &
Demas, 1997; Gilad et al., 1998; Maestroni, 1998; Hotta et al., 2000; Garcia et al., 2001;
Andrew & Hotchkiss, 2002; Guerrero & Reiter, 2002; Carrillo-Vico et al., 2003; Pontes
et al., 2006; Srinivasan et al., 2008). Os estudos iniciais sobre sua ação imunomodulatória
baseavam-se na retirada da glândula pineal e na observação de diversos parâmetros
imunológicos, incluindo principalmente os órgãos linfóides. A supressão da imunidade,
atrofia do timo e a redução de células imunes no baço após a pinealectomia representam
evidências marcantes do papel regulador exercido pela melatonina no sistema imune
(Csaba & Barath, 1975; Maestroni et al., 1986; Brainard et al., 1988; Jankovic et al., 1994).

39 I. Introdução 39 I. Introdução
Um importante aspecto dos efeitos da melatonina é que estes são observados em
uma ampla faixa de concentrações. Esta variabilidade implica que suas ações sejam
ligadas, ou não, à ritmicidade circadiana. A melatonina pode ser produzida em outros
locais além da pineal, tendo sido descrita no trato gastrointestinal (Bubenik, 2002), na
retina (Zmijewski et al., 2009), células epidérmicas (Slominski et al., 1996, 2002) e células
imunocompetentes (Carrilo-Vico et al., 2004; Pontes et al., 2006, 2007). Nestas condições
a melatonina é produzida em altas concentrações, na faixa de μM – mM, enquanto a
concentração plasmática noturna está na faixa de pM – nM. A produção extrapineal
pode ocorrer de maneira rítmica como acontece na retina ou constitutiva como acontece
no trato gastrointestinal, mas, em nenhum desses locais, a melatonina secretada parece
contribuir para variação rítmica do hormônio na circulação, sendo este padrão
dependente exclusivamente da produção pineal. Estudos mostram que tanto os níveis
de melatonina noturnos como a produzida localmente produzem efeitos regulatórios
sobre o sistema de defesa.
Os efeitos citoprotetores da melatonina são mediados por sua capacidade de
atuar como anti-oxidante (Reiter et al., 2002, Bilici et al., 2002, Zhang et al., 2004) e por
inibir vias de sinalização que desencadeiam respostas inflamatórias (Gilad et al., 1998;
Maestroni, 1998; Lissoni, 1999; Reiter et al., 2000; Skwarlo-Sonta et al., 2003, Markus et
al., 2010). Esses efeitos são obtidos em altas concentrações de melatonina e incluem sua
ações diretas como “scavenger” de radicais livres (Poeggeler et al., 2002) ou como
indutora do aumento da atividade de enzimas antioxidantes (para revisão Luchetti et al.,
2010).
Outra abordagem dos efeitos da melatonina está em sua ação sobre a resposta
imune inata, exibindo padrões diferentes de regulação. Em altas concentrações, a
melatonina possui efeitos imuno-estimulatórios. Ela promove aumento da transcrição

40 I. Introdução 40 I. Introdução
do gene de diversas citocinas pro-inflamatórias em células mononucleares (Garcia-
Maurino et al., 1997; Liu et al, 2001; Carrillo-Vico et al., 2003), aumento da atividade
fagocítica dessas mesmas células (Pontes et al., 2006) e da apresentação de antígenos
(Pioli et al., 1993), além do aumento da proliferação de linfócitos T e atividade de células
linfóides. Todos esses efeitos são relevantes para a montagem da resposta inflamatória,
por isso nestas condições a melatonina tem propriedades pró-inflamatórias. Por outro
lado, em concentrações compatíveis com as encontradas na circulação a melatonina tem
efeitos contrários, de inibição da resposta inflamatória. A concentração plasmática de
melatonina reduz de forma significativa a adesão e rolamento de células
imunocompetentes à camada endotelial e o aumento da permeabilidade vascular
induzidos por agentes pró-inflamatórios em ratos (Lotufo et al., 2001, 2006). Suas ações
devem ser entendidas de acordo com o contexto fisiológico e fisiopatológico, por isso
restringir seu caráter ao efeito pró ou anti-inflamatório pode ser equivocado. As suas
propriedades bem marcadas como sua ação citoprotetora e imunomodulatória podem
predizer a sua aplicação e investigação em diversas condições e doenças.
44..22.. OO tteemmppoo nnaa rreessppoossttaa iimmuunnee:: rreellaaççããoo eennttrree mmeellaattoonniinnaa ee aaddaappttaaççããoo..
Os mecanismos biológicos evoluíram de forma a permitir a antecipação das
mudanças ambientais associadas ao tempo. Essas adaptações são cruciais para
sobrevivência, principalmente para espécies que convivem com mudanças drásticas no
ambiente. Essas espécies se deparam com desafios de disponibilidade de alimento e
baixas temperaturas que fazem do inverno, um período particularmente difícil para se
reproduzir e sobreviver. Nesse sentido, os indivíduos deveriam também prever
condições imunologicamente desafiantes.

41 I. Introdução 41 I. Introdução
Diferentemente de algumas funções fisiológicas, a função imune foi tida por
muitos anos como relativamente constante ao longo das estações (Sheldon & Verhulst,
1996), inclusive as flutuações no número de morte e doenças ao longo do tempo foram
mais atribuídas a virulências dos agentes causadores do que necessariamente a
mudanças na própria imunidade (Nelson et al., 2002). A manutenção da função imune é
considerada energeticamente elevada, as cascatas de atividade mitótica, a montagem do
processo inflamatório, a febre e a produção de fatores imunorregulatórios requerem
energia substancial (Demas et al., 1997). Diante disso, todos esses processos devem ser
altamente regulados para que a imunidade não perca efetividade diante de situações
desfavoráveis como nas mudanças sazonais do ambiente. Considerando que certas
atividades como a reprodução podem ser reduzidas durante períodos desfavoráveis,
outras como a função imune não pode ser reduzida sem graves comprometimentos para
a sobrevivência (Nelson et al., 2002).
Picos sazonais no tamanho dos órgãos linfáticos geralmente ocorrem no final do
outono ou no início do inverno e mínimos sazonais são observados antes do início da
reprodução. Dados de campo sugerem que a função imunológica e processos de doença
são reforçados durante o inverno (Nelson & Demas, 1997; Hotchkiss & Nelson, 2002).
Nos seres humanos também foram descritas mudanças sazonais na função imune.
Citocinas pró-inflamatórias como interferon (IFNγ) e INFα e IL6 estão aumentadas
durante o inverno em comparação com o verão (Katila et al., 1993; Maes et al., 1994).
As mudanças sazonais na função imune são determinadas pelo fotoperíodo e
mediadas pela duração da secreção de melatonina (Nelson & Demas, 1995, 1996; Nelson
& Drazen, 2000; Prendergast et al, 2004). De forma geral, o reforço da função imune
provê a antecipação do organismo frente aos estressores recorrentes que comprometem
a imunidade (Nelson et al., 2002). Em conjunto, todas as mudanças exercidas pela

42 I. Introdução 42 I. Introdução
melatonina no sistema imune são significativas para uma resposta imune integrada e
relevante para sobrevivência. Sobre a mesma lógica, o sistema imune pode atuar
diretamente sobre a função pineal. Essas ações refletem-se em ajustes sobre a
melatonina circulante de modo a permitir a regulação da função imune (Nelson &
Drazen, 1999; Skwarlo-Sonta et al., 2003).
Em modelo de inflamação crônica através da injeção de Bacilo Calmette-Guerin
(BCG) em patas de camundongo foi observado um ritmo diário da lesão granulomatosa
induzida. O edema da pata era menor à noite e maior durante o dia. Nesse caso a
extirpação da pineal aboliu o ritmo do edema que era restaurado após a administração
de melatonina durante a fase escura (Lopes et al., 1997). Pode-se entender que a natureza
da resposta imune é também modificada por uma variação diária das concentrações de
melatonina. Entende-se que o tempo também é determinante sobre a função imune em
uma escala muito menor,ou seja, o ciclo dia/noite também implica em alterações no
perfil de resposta inflamatória.
44..33.. AA ccoonnssttrruuççããoo ddee uumm nnoovvoo ccoonncceeiittoo:: PPiinneeaall ccoommoo aallvvoo ppaarraa mmeeddiiaaddoorreess
ddaa rreessppoossttaa iimmuunnee
Em mamíferos, a glândula pineal é responsável pela transdução do fotoperíodo
ambiental em sinalização hormonal. O escuro captado pela retina e processado pelo
sistema nervoso é traduzido através da produção e liberação do hormônio melatonina.
Portanto, esta indolamina é também conhecida como o hormônio do escuro (Reiter,
1993).
Os dados da literatura demonstram que melatonina é uma molécula diretamente
relacionada à resposta imune. Os estudos caracterizaram seu papel regulatório em
diferentes células imunes e a sua relação com a adaptação sazonal e processos de defesa
(para revisão, Guerrero & Reiter, 2002). Sobre a mesma lógica o sistema imune pode

43 I. Introdução 43 I. Introdução
atuar diretamente sobre a função pineal. Essas ações refletiram-se em ajustes sobre a
melatonina circulante de modo a permitir a regulação da função imune (Nelson &
Drazen, 1999; Skwarlo-Sonta et al., 2003). Nesse sentido, a própria pineal passa a ser
também alvo de mediadores imunológicos. Consistindo nessa concepção diversos
estudos mostraram que citocinas e outros sinalizadores da inflamação afetam a função
pineal (Mucha et al. 1994; Withyachumnarnkul et al., 1990a, 1990b; Zylinska et al., 1995,
Tsai et al., 2001; Ferreira et al., 2005; Jiang-Shieh et al., 2005; Fernandes et al., 2006; 2009;
da Silveira Cruz-Machado et al., 2010). Essas relações apontam para a existência de uma
comunicação bidirecional entre o sistema imune e a glândula (Skwarlo-Sonta et al, 2003;
Markus et al., 2007,). A proposta do Eixo-Imune-Pineal tem como pressuposto essa
interação e traz o conceito de que a comunicação recíproca entre a glândula pineal e o
sistema imune é essencial para a montagem da resposta inflamatória. As diversas
abordagens que contribuíram para construção dessa proposta serão apresentadas a
seguir. O presente trabalho tem como precursor resultados que constituem parte dessa
concepção e deram suporte para sua elaboração.
No estudo conduzido por Lopes e colaboradores (1997), além da observação de
que a lesão induzida por BCG na pata de camundongos apresentava ritmo diário e este
era alterado após pinealectomia e restaurado com melatonina exógena, foi mostrado
ainda que o ritmo da excreção do metabólito da melatonina, a 6-sulfatoximelatonina,
era perdido após remoção da adrenal (Lopes et al., 2001). Esses dados sugeriam pela
primeira vez a participação da adrenal no controle da função pineal. Foi demonstrada a
importância dos glicocorticóides na manutenção do ritmo diário da lesão
provavelmente pela indução do pico noturno de melatonina nessas condições. Essa
interação foi confirmada posteriormente quando demonstrou-se que a corticosterona

44 I. Introdução 44 I. Introdução
potencia a síntese de melatonina in vitro (Ferreira et al., 2005) e in vivo (Fernandes et al.,
2009).
Diversos mediadores da resposta imune também demonstraram exercer efeitos
sobre a função pineal. Foi mostrado que a citocina interleucina 1 beta (IL-1β) é expressa
constitutivamente na glândula pineal e aumentada após tratamento com noradrenalina,
IFNγ e LPS (Tsai & McNulty, 1999). Estudo do mesmo grupo também mostrou que o
RNAm da IL-6 e do fator de crescimento beta (TGF-β) são transcritos na glândula
pineal e regulados positivamente pela noradrenalina (Tsai et al., 2001a). O efeito dessas
citocinas sobre a atividade da glândula foi avaliado através da análise do conteúdo ou
síntese de precursores e da própria melatonina. Foi observado que o IFNγ pode induzir
tanto o aumento como a redução da melatonina em pineais cultivadas dependendo das
concentrações de IFNγ e do nível de estimulação adrenérgica utilizada
(Withyachumnarnkul et al. 1990b). Os fatores estimulares de colônias granulocitárias
(GM-CSF e G-CSF) e citocinas secretadas por células imunocompetentes ativadas
potenciam a produção de melatonina in vivo e in vitro na pineal de rato (Zylińska et al.,
1995). Também foi avaliado a participação da microglia na resposta da pineal à essas
citocinas. IL-1β, TNF e INFγ modulam o conteúdo de 5-HT em culturas de glândulas
pineais de rato (Tsai et al., 2001b).
Os efeitos inibitórios da melatonina noturna sobre a migração de leucócitos à
camada endotelial (Lotufo et al., 2001, 2006) implicariam que, durante a noite, a
montagem da resposta seria retardada por ação desse hormônio. Para haver o
recrutamento celular adequado, independente da hora do dia, os níveis de melatonina
circulantes devem ser reduzidos. Essa hipótese foi confirmada pela demonstração de
que agentes envolvidos na fase de montagem da resposta promovem efeitos negativos
sobre a produção de melatonina. O TNF inibe transientemente a transcrição do gene

45 I. Introdução 45 I. Introdução
que codifica a enzima AA-NAT e também a produção do precursor da melatonina, a N-
acetilserotonina, em glândulas pineais de ratos em cultura (Fernandes et al., 2006). O
estudo in vivo com um modelo de inflamação em humanos também mostrou a mesma
relação. Mulheres com mastite, um processo inflamatório não-infeccioso, apresentam
altos níveis de TNF e têm supressão do ritmo de melatonina (Pontes et al., 2006). No
mesmo contexto de uma resposta pró-inflamatória, a injeção de LPS promove a redução
dos níveis plasmáticos de melatonina em ratos (Tamura et al., 2010) e inibição da
produção de N-acetilserotonina e melatonina induzida por noradrenalina em pineais em
cultura (da Silveira Cruz-Machado et al., 2010). Observou-se ainda que em quadros de
infecção generalizada, caracterizada por altos níveis de TNF, há supressão do ritmo de
melatonina (Mundigler et al., 2002)
A caracterização dos mecanismos envolvidos na regulação da função pineal por
diversos fatores inflamatórios permitiu sugerir que o fator NFKB está diretamente
evolvido nas respostas observadas. Esse fator é constitutivamente expresso na glândula
pineal e apresenta ritmo diário, com máximo acúmulo nuclear no final da fase de claro e
rápida redução após o apagar das luzes (Cecon et al., 2010). A ativação da via NFKB por
fatores pró-inflamatórios como o TNF e LPS bloqueia (Fernandes et al., 2006, da Silveira
Cruz-Machado et al., 2010), enquanto que a inibição dessa via por corticosterona
potencia a síntese de melatonina induzida por noradrenalina em glândulas pineais em
cultura (Ferreira et al., 2005). A pineal também mostrou ser instrumentalizada para
responder à esses fatores. Já foram detectados em pinealócitos receptores para LPS,
TLR4 e CD14 (da Silveira Cruz-Machado et al., 2010) e para glicocorticóides (GR)
(Ferreira et al., 2005).
Os achados sobre a função pineal e processos de injúria indicam a comunicação
bidirecional do sistema imune com a glândula pineal. Essa concepção antes era proposta

46 I. Introdução 46 I. Introdução
do ponto de vista que presumia a existência de um mecanismo de retroalimentação para
regulação da melatonina circulante, considerando-se seu papel imunomodulatório
(Nelson & Drazen, 1999; Tsai et al., 2001b; Skwarlo-sonta et al., 2003). A proposta do
Eixo-Imune-Pineal considera a existência de mecanismos comuns de regulação da
produção de melatonina frente a condições desafiantes e relevante para uma resposta
inflamatória. No estado de higidez do organismo a melatonina é sintetizada
ritmicamente pela pineal. Diante de uma situação adversa como uma injúria, a pineal é
sinalizada a interromper a produção de melatonina para permitir a montagem da
resposta inflamatória. Por outro lado, na periferia, células imunocompetentes seriam
estimuladas e passariam a produzir grandes quantidades de melatonina que atuaria de
maneira parácrina, exercendo seu efeito antioxidante e anti-inflamatório, contribuindo
para finalização da resposta. Essa situação aconteceria de maneira transiente, de modo a
admitir que o animal resolva uma infecção ou lesão tecidual de maneira eficiente,
quando esses níveis assim seriam restaurados. O catabolismo dos mediadores e aumento
de glicocorticóides circulantes após a fase de resolução contribuiriam positivamente
para o retorno da síntese de melatonina pela glândula pineal (Markus et al., 2007).

47 I. Introdução 47 I. Introdução
FIGURA 4. OO EEiixxoo--iimmuunnee--ppiinneeaall: Em sujeitos saudáveis o NSQ controla a produção de
melatonina transduzindo a informação ambiental luminosa no pico noturno de melatonina em
concentrações que reduz a transmigração de leucócitos para os tecidos. Em uma situação de
injúria, moléculas produzidas no início da inflamação são percebidas pela glândula pineal
sinalizando a supressão da síntese de melatonina. A ausência do pico noturno de melatonina
permite a migração adequada de leucócitos e portanto uma montagem eficiente da resposta. Os
leucócitos já nos locais de injúria passam a produzir melatonina de forma parácrina de maneira
a resolver o processo inflamatório.

____________
Objetivos

49 II. Objetivos 49 II. Objetivos
II. OBJETIVOS
Objetivo desta dissertação foi demonstrar que algumas das moléculas mais
relevantes para desencadear a resposta ao TNF estão presentes em glândulas pineais em
cultura e em pinealócitos isolados de ratos. Neste sentido também analisamos o efeito
funcional da ativação da via de sinalização do TNF em pinealócitos. Para isso avaliamos:
1. A presença dos receptores TNF-R1 nos diferentes tipos celulares da pineal
2. A presença do receptor TNF-R1 em cultura de pinealócitos e o efeito do TNF
sobre a expressão do receptor.
3. A translocação nuclear do fator NFKB em glândulas pineais em cultura.
4. As subunidades do NFKB presentes em glândulas pineais estimuladas ou não
com TNF.
6. O efeito do TNF sobre a expressão da proteína inibitória IKBα em
pinealócitos em cultura.
5. O efeito do TNF sobre a expressão de iNOS e produção de óxido nítrico

____________
Material e Métodos

51 III. Material e Métodos 51 III. Material e Métodos
III. MATERIAL E MÉTODOS
1. ANIMAIS
Foram utilizadas ratos (Rattus novergicus) fêmeas pré-púberes da linhagem Wistar
criadas no biotério do Instituto de Biociências da Universidade de São Paulo. Os
animais foram alojados em gaiolas de polipropileno mantidas em sala com temperatura
e umidade controladas e adequadas à espécie. O ciclo claro-escuro empregado foi de 12
h claro/ 12 h escuro, luz ligada às 06:30 h. Os animais receberam ração e água ad libitum
durante todo procedimento experimental. Os animais foram mortos por decapitação
sem administração de anestésicos antes do apagar das luzes (ZT10 – ZT12) (ZT –
Zeitgeber time – ZT zero corresponde ao momento do ascender das luzes). Os
experimentos foram conduzidos de acordo com os princípios da ética em
experimentação animal, e aprovado pela Comissão de Ética em Uso de Animais em
Experimentação (CEUA) do Instituto de Biociências (protocolo 081/2008).
2. DROGAS E REAGENTES
As drogas e os reagentes utilizados são descritos a seguir conforme a empresa de
procedência.
Abcam (Cambridge, MA, EUA): anticorpos anti-TNF-R1 (ab19139) e anti-ED1
(ab31630)
Calbiochem (Darmstadt, Alemanha): NP-40 e 1400W.
Gibco BRL (Grand Island, NY, EUA): penicilina, estreptomicina e glutamina.

52 III. Material e Métodos 52 III. Material e Métodos
Invitrogen Life Technology (Calrsbad, CA, EUA): ditiotreitol (DTT), fluoreto de
fenilmetilsulfonil (PMSF), anticorpo anti-coelho conjugado ao fluoróforo FITC, T4
polinucleotídeo quinase, DMEM (do inglês, Dulbecco’s modified Eagle médium) soro fetal
bovino, e 4’ 6-diamidino-2-fenilindol (DAPI), 4-amino-5-metilamino-2´,7´-
difluorofluoresceina (DAF-FM-DA).
Santa Cruz Biotechnology (Santa Cruz, Califórnia, EUA): (Santa Cruz, Califórnia,
EUA): anticorpos da via NFKB: anti-p-50 (sc-114X); anti-RelA (sc-109X); anti-p52 (sc-
298X); anti-c-Rel (sc-70X); anti-RelB (sc-226X); anti-BCL-3 (sc-185X); anti-IKBα
(sc371) e anti-iNOS (sc-651).
Sigma-Aldrich (Saint Louis, Missouri, EUA): HEPES, meio de cultura BGJb,
bisacrilamida (N, N’-metilenobisacrinamida; anticorpo anti-GFAP conjugado ao fluoróforo
Cy3, poli(dIdC), pirrolidinaditiocarbamato (PDTC), 3-(4,5-dimetil-2-tiazol)-2,5-
difenil-brometo de tetrazolium (MTT); fator de necrose tumoral (TNF), tripsina;
inibidor de tripsina e albumina bovina fração V (BSA).
Perkin Elmer (Boston, MA, EUA): Easy Tides Adenosine 5’-triphosphate, [γ-32P].
Promega (Madison, MA, EUA): Oligonucleotídeo consenso para NFKB (5’-
AGTTGAGGGGACTTTCCCAGGC-3’) e AP-1 (5'-CGTTGATGAGTCAGCCGG AA-
3').
3. PREPARO DE DROGAS
O TNF foi preparado em alíquotas na concentração de 100 µg/mL em água
deionizada purificada e os estoques foram congelados a -20 °C. No dia do experimento
as alíquotas foram diluídas em meio de cultura DMEM (cultura de células) ou BGJb
(cultura de glândula pineal).

53 III. Material e Métodos 53 III. Material e Métodos
O indicador de NO, DAF-FM DA, foi diluído em DMSO (100%) e estocado em
alíquotas de 1 mM a -20 °C. No momento do uso as alíquotas foram diluídas em solução
salina (mM ; NaCl 150, KCl 5, CaCl2 2, NaHCO3 15, C6H12O6 11).
As demais drogas listadas anteriormente foram, quando necessário, diluídas em
água deionizada purificada por sistema Milli-Q (Millipore®, Billerica, MA, EUA).
4. CULTURA DA GLÂNDULA PINEAL
A cultura das glândulas foi realizada segundo protocolo descrito por Parfitt e
col. (1976) e modificado por Ferreira e col. (1994). Após a decapitarão do animal as
pineais foram retiradas do crânio rapidamente e cultivadas em placas de cultura de 24
poços. Foi adicionada uma pineal por poço, contendo 200 µl de meio de cultura BGJb
acrescido de glutamina (2 mM), penicilina (100 U/ml) e estreptomicina (100 μg/mL) e
mantidas a 37 °C em atmosfera com 95% de saturação de O2 e 5% de CO2 por 48 horas
antes de serem submetidas aos protocolos experimentais O meio de cultura foi trocado
a cada 24 h e 1h antes da administração de TNF.
5. CULTURA DE PINEALÓCITOS
A cultura de pinealócitos foi realizada segundo protocolo descrito por Ferreira e
col. (2003). Glândulas pineais foram retiradas, cortadas em pequenos pedaços e
submetidas à dissociação enzimática com tripsina (0,25 %, 37 °C, 15 min) seguida por
dispersão mecânica na presença de inibidor de tripsina (0,3% em solução contendo, em
mmol/L, NaCl 120, KCl 5, NaHCO3 25, KH2PO4 1,2, glicose 12 e 0,1% w/v albumina
bovina). Logo após a dispersão a suspensão foi centrifugada (1000 g, 15 min), descartado
o sobrenadante e as células foram ressuspensas em meio de cultura DMEM acrescido de
soro fetal bovino (10 %, inativado por calor) e penicilina (100 U/mL). Dessa suspensão

54 III. Material e Métodos 54 III. Material e Métodos
foi retirada uma alíquota para contagem do número de células utilizando a câmara de
Neubauer e a viabilidade das células foi estimada pela técnica de exclusão por azul de
Trypan. Após o cálculo de viabilidade e número de células por poço, as células foram
semeadas em placas cobertas previamente com poli-L-lisina (0,5 a 1 x 105 céls./poço). A
cultura foi mantida em estufa (37 °C em atmosfera com 95% de saturação de O2 e 5% de
CO2 ) por 18 horas antes do início dos tratamentos.
6. TESTE DE VIABILIDADE CELULAR
A viabilidade celular foi avaliada pelo ensaio colorimétrico de MTT. Os
pinealócitos foram semeados na densidade de 5 × 104 células/poço em placas de 96 poços
(150 mL) e tratados ou não com TNF (80 ng/mL, 2h) e, em seguida, incubados com
MTT (0,5 mg/mL) por 4h a 37 °C. A inibição da ligação DNA-NFKB e da atividade da
enzima sintase de óxido nítrico induzida (iNOS) foi obtida através da incubação das
células com PDTC (25 µM) ou 1400W( 1 µM), respectivamente. O corante formazan foi
dissolvido com dimetilsulfóxido (DMSO). As placas foram lidas em espectrofotômetro
de microplacas (Spectramax 250, Molecular Devices, CA, EUA) a 540 nm. A viabilidade
celular foi definida de acordo com o percentual de densidade óptica de cada grupo em
relação ao grupo controle.
7. IMUNO-HISTOQUÍMICA DA GLÂNDULA PINEAL
77..11.. MMééttooddoo ddee RReevveellaaççããoo ccoomm DDAABB
Para obtenção dos tecidos, os animais foram anestesiados através de injeção
intramuscular de ketamina (160 mg/kg) acrescido de xilazina (40 mg/kg) e perfundidos
transcardialmente com aproximadamente 150 mL de solução salina, seguida de

55 III. Material e Métodos 55 III. Material e Métodos
aproximadamente 300 mL de solução salina acrescida de paraformaldeído (PAF, 4%) a
4°C (pH 9,5). Cada pineal foi então removida e crioprotegida no mesmo fixador
enriquecido com 20% de sacarose por 12 h, seguindo-se por solução de sacarose 30% em
tampão fosfato-salino (PBS, pH 7.4, mmol/L: NaCl 137, KCl 2,7, Na2HPO4 2 H2O 8,1,
KH2PO4 1,76) a 4 °C por 24 h adicionais. As pineais foram então incluídas em meio para
congelamento (Tissue-Tek, Sakura Finetek, Torrence, CA, EUA), rapidamente
congeladas em gelo seco e estocadas a -80 °C. A criomicrotomia foi realizada em
criostato e os cortes com espessura de 10 μm foram montados em lâminas para
microscopia com aderência eletrostática (Star Frost, Alemanha). Após secarem em
temperatura ambiente, as lâminas foram estocadas a -20 °C até a realização da imuno-
histoquímica. Seguindo o protocolo de imuno-histoquímica, após fixação em
paraformaldeído 4% por 30 min, os cortes foram incubados com glicina (0,1 M) por 5
min, seguido por incubação com solução de bloqueio (3% de albumina e 0,01% de
saponina em PBS) por 30 min em temperatura ambiente. A biotina endógena foi
bloqueada com o kit comercial avidin-biotin blocking kit (Vector, SP2001, Burlingame,
CA, EUA) por 30 min. Seguiu-se então, a incubação com o anticorpo produzido em
coelho anti-TNF-R1 (1:500, Abcam) diluído em solução de bloqueio. O anticorpo
primário foi incubado por 16 h a 4 °C, seguindo-se de incubação com o anticorpo
secundário anti-coelho conjugado a biotina (1:200, Abcam) por 1 h a temperatura
ambiente. O complexo avidina-biotina foi marcado com kit comercial (Vectastain Elite
Kit, Vector, PK-605, Burlingame, CA, EUA) e a atividade da peroxidase foi revelada com
3, 3’-diaminobenzidina (DAB, substrate kit for peroxidase, Vector, SK-4100, Burlingame,
CA, EUA). Após o ensaio de imuno-histoquímica as lâminas foram contra-coradas com
azul de metileno.

56 III. Material e Métodos 56 III. Material e Métodos
77..22.. IImmuunnoofflluuoorreessccêênncciiaa-- FFrreeee FFllooaattiinngg
Cada pineal foi removida do crânio e fixada em paraformaldeído 4%
suplementado com 20% de sacarose a 4 °C por 3 dias seguido de crioproteção em
solução de sacarose 30% em PBS a 4 °C por 18h adicionais. As pineais foram então
incluídas em meio para congelamento (Tissue-Tek, Sakura Finetek, Torrence, CA,
EUA), rapidamente congeladas em gelo seco e estocadas a -80 °C. Os cortes foram
realizados em criostato com espessura de 30 μm. Os cortes foram primeiramente
lavados em PBS (3X 10 min) seguido por incubação com solução de bloqueio (2% de
albumina e 0,3% de Triton X em PBS) a temperatura ambiente por 60 min. Foi então
realizada a incubação com os anticorpos primários: produzido em coelho anti-TNF-R1
(1:100, Abcam), produzido em camundongo anti-ED-1 (1:100, Abcam) e/ou produzido
em camundongo anti-GFAP conjugado ao fluoróforo Cy3 (1:2000, Sigma-Aldrich) e
produzido em camundongo anti-TNF-R2 diluídos em solução de bloqueio por 48 h a
4˚C em constante agitação. Após esta etapa, os cortes foram novamente lavados em PBS
por 30 min, seguindo-se da incubação dos anticorpos secundários fluorescentes
apropriados: anti-coelho conjugado ao FITC (1:200, Sigma-Aldrich) e/ou anti-
camundongo conjugado ao Cy3 (1:200, Jackson ImmunoResearch, West Grove, PA,
USA) por 90 min em temperatura ambiente. O núcleo foi marcado com DAPI (30 μM, 5
min , Invitrogen) diluído em PBS e incubado em temperatura ambiente.
Os controles negativos foram realizados com a omissão do anticorpo primário e
com a incubação dos cortes com o anticorpo secundário correspondente. A marcação foi
observada por microscopia confocal (Zeiss, LSM510, Baden-Wurttemberg, Alemanha)
sendo utilizado o laser HeNe 543 para excitação do fluoróforo Cy3 no comprimento de
onda de 543 nm e filtro de emissão em 560 nm. Para a excitação do fluoróforo FITC
utilizou-se laser argônio no comprimento de onda de 488 nm e filtro de emissão em 505.

57 III. Material e Métodos 57 III. Material e Métodos
O laser enterprise (UV) foi utilizado para excitação do marcador DAPI no comprimento
de onda de 364 nm e a emissão verificada com filtro de 435-485 nm.
8. IMUNOFLUORESCÊNCIA DE CULTURA DE PINEALÓCITOS
Para o ensaio de imunocitoquímica os pinealócitos foram semeados em placas de
8 poços (Lab-Tek chamber slide, Nalge Nunc International, Nova York, EUA). Após o
período de execução do protocolo experimental, as células foram lavadas 2X em PBS e
em seguida fixadas com paraformaldeido 4% em PBS por 15 minutos. Após a fixação as
células foram lavadas novamente em PBS e permeabilizadas com solução de saponina
0,5 % em PBS e por último incubadas com solução de bloqueio (1% BSA e 0,3 M de
glicina em PBS). Após essa sequência as células foram incubadas overnight (18 h, 4°C).
em câmara úmida com os anticorpos primários: produzido em coelho anti-TNF-R1
(1:100), produzido em coelho anti-IKBα (1:50) e/ou produzido em coelho anti-NOS2
conjugado ao fluoróforo TRITC (1:25). Após a incubação com os anticorpos primários,
as células foram lavadas com PBS (2X) e em seguida incubadas com o anticorpo
secundário anti-coelho conjugado ao fluoróforo Texas Red (1:400, Abcam) por 1 h em
temperatura ambiente. O núcleo foi marcado com DAPI (300 μM, 5 min) a temperatura
ambiente.
A marcação foi analisada por microscopia confocal (Zeiss, Baden-Wurttemberg,
Alemanha) e visualizada por diferentes objetivas. O laser HeNe 543 foi utilizado para
excitação dos fluorofóros Texas Red e TRITC (excitação em 543 nm e filtro de emissão
em 650 e 560-615 nm, respectivamente). Para visualização do DAPI foi utilizado o laser
enterprise (UV), excitação no comprimento de onda de 364 nm com filtro de emissão
em 435–485 nm. Para cada ensaio foi realizada a análise de 3 imagens correspondentes a
campos diferentes com, aproximadamente, 7 células por campo. A quantificação da

58 III. Material e Métodos 58 III. Material e Métodos
fluorescência foi realizada através do programa público Image J (National Institute of
Health, USA, download em http://rsb.info.nih.gov/ij).
9. DETECÇÃO DA PRODUÇÃO DE NO POR MICROSCOPIA
CONFOCAL
A produção de NO foi detectada por microscopia confocal utilizando o
indicador intracelular DAF-FM DA. O DAF-FM DA reage com o NO-2 (formado durante
a oxidação espontânea do NO) e essa reação emite fluorescência (Kojima et al., 1998).
A detecção do NO nos pinealócitos foi realizada de acordo com protocolo
descrito por Tamura e col. (2009). Os pinealócitos foram cultivados em lamínulas de
vidro contidas em placas próprias para microscopia confocal mantidas por 18 h em
estufa (200 µL DMEM, 37°C, 5% CO2,). Após esse período o meio foi substituído por
solução salina contendo, em mM: NaCl 150, KCl 5, CaCl2 2, NaHCO3 15, C6H12O6 11 e
suplementada com L-arginina (100 M), pH 7,4. Seguiu-se o tratamento experimental e
nos últimos 50 minutos de tratamento as células foram carregadas com o indicador de
NO DAF-FM DA (5 M). Após o tratamento, as células foram lavadas com solução
salina para retirar o excesso do probe e levadas para análise dos níveis de NO por
microscopia confocal. O ensaio foi realizado em ambiente com baixa luminosidade
desde a incubação com DAF-FM DA até a obtenção das imagens. Os registros foram
feitos com a objetiva de imersão em óleo (40X), a excitação realizada com laser argônio
(488 nm) e a emissão 515-530 nm. Foram obtidas 4 imagens de campos selecionados
aleatoriamente de cada placa, contendo pelo menos 7 células viáveis. A quantificação da
fluorescência emitida pelo DAF-FM DA foi realizada através do programa Image J. Ao final
dos experimentos as células foram tratadas com o doador de NO nitroprussiato de sódio
(1 mM) para verificar a eficácia da detecção de NO. O efeito dos diferentes tratamentos

59 III. Material e Métodos 59 III. Material e Métodos
no aumento de NO induzido por TNF foi avaliado tendo sido o valor da média
(unidades arbitrárias) obtida no grupo controle considerado como 100%.
10. EXTRAÇÃO DE PROTEÍNA NUCLEAR
Após cultivo e tratamentos as glândulas pineais foram armazenadas em freezer -
80° C e depois processadas para extração nuclear de proteínas. Cada glândula pineal foi
homogeneizada em 100 μL de tampão de lise (HEPES, pH 7,5, 10 mM; KCl 10 mM;
EDTA 0,1 mM; glicerol 10%, DTT, 1 mM, PMSF, 0,1 mM) com utilização de um
homogenizador e pistilo apropriados. Em seguida, foi adicionado 12,5 μL do detergente
NP-40 (10%), as amostras foram agitadas rapidamente com auxílio do vortex e
incubadas em gelo por 15 min. Após esse período foram centrifugadas (12000 g, 1 min,
4°C) e o sobrenadante, correspondente a fração citoplasmática do extrato, descartado.
O pellet resultante foi então lavado com 50 μL do tampão de lise e centrifugado (12000 g,
1 min, 4°C) sendo o sobrenadante novamente descartado. O pellet nuclear formado foi
ressuspenso em 20 μL do tampão de extrato nuclear (HEPES, pH 7,5, 10 mM; KCl 5 M;
EDTA 1 mM; glicerol 10%, DTT 1 mM, PMSF 0,1 mM) ficando 15 min em gelo sob
agitação. Após centrifugação (20.000 g, 5 min, 4°C) o sobrenadante (extrato nuclear)
foi aliquotado e estocado a -20 °C até o ensaio.
A quantificação da concentração de proteína dos extratos foi realizada a 280 nm
através do espectrofotômetro ND-1000 (Nanodrop, Wilmington, DE, EUA).

60 III. Material e Métodos 60 III. Material e Métodos
11. ENSAIO DE ELETROMOBILIDADE POR RETARDO EM GEL
(EMSA)
O EMSA determina o acúmulo nuclear do fator NFKB. O ensaio consiste na
reação de ligação de proteínas do extrato nuclear com a sequência consenso específica
para o fator de transcrição NFKB marcada com o radioisótopo 32P.
Para os ensaios foi utilizado 6 μg de proteína do extrato nuclear de pineais
incubadas na ausência ou presença de TNF (30 ng/mL) por 5 a 60 minutos. Seguiu-se
incubação destas, com tampão de ensaio (Tris-HCl 10 mM pH 7,5, MgCl2 1 mM, NaCl
50 mM, DTT 0,5 mM, EDTA 0,5 mM, poli (dIdC) 1 μg e glicerol 4%) por 20 min em
temperatura ambiente. Na sequência, as amostras foram incubadas por 30 min em
temperatura ambiente com 40000 cpm de sonda de oligonucleotídeo dupla-fita
correspondente ao consenso para NFKB (5’-AGTTGAGGGGACTTTCCCAGGC-3’,
Promega, Madison, WI, EUA) marcado com radioisótopo. A marcação da sonda foi
realizada pela incubação do oligonucleotídeo consenso para NFKB com 1 μL de [γ32P]-
ATP (atividade específica 3000 Ci/mmol) na presença da enzima T4 polinucleotídeo
quinase (10 U). Esta enzima substitui os fósforos do oligonucleotídeo por fósforo
radioativo e o que não é incorporado é removido por coluna sefadex G-25 (GE
Healthcare Life Sciences). Em seguida, foi adicionado 2 μL de tampão de corrida (Tris-
HCl 250 mM pH 7,5, glicerol 4%, azul de bromofenol 0,2%). Os complexos proteína-
DNA foram analisados em gel de poliacrilamida não-desnaturante, 6% de
acrilamida:bisacrilamida (37,5:1) por eletroforese (150V, 1h30min) em tampão 0,25 x de
Tris-borato/EDTA (TBE). Finalizada a corrida, o gel foi seco a vácuo, exposto ao filme
BIOMAX-MS (Kodak, Rochester, NY, EUA) em cassete apropriado por 48 h a -80˚C.
Seguiu-se a revelação por imersão do filme em solução reveladora por 5 min (Kodak,
Rochester, NY, EUA) e em solução fixadora por 10 min (Kodak, Rochester, NY, EUA).
As autoradiografias foram quantificadas através do programa Image J.

61 III. Material e Métodos 61 III. Material e Métodos
12. ENSAIO DE SUPERSHIFT
As subunidades de NFKB presentes nos extratos nucleares de glândulas pineais
tratadas ou não com TNF (30 a 90 ng/mL, 5 min) foram identificadas através do ensaio
de EMSA. Foram utilizados 6 μg de proteína de um pool de 4 extratos de cada grupo
experimental. Neste ensaio foram adicionados 2 µg/mL anticorpos específicos para
subunidades de NFKB: RelA, p50, p52, c-Rel, RelB, e/ou Bcl3 por 45 minutos antes da
incubação dos extratos com a sonda consenso para NFKB-32P. As demais etapas que se
seguiram foram realizadas conforme o protocolo de EMSA descrito anteriormente.
13. ANÁLISE DOS RESULTADOS
Os resultados estão apresentados como média ± erro padrão da média (e. p. m).
O ensaio de EMSA foi quantificado pelo programa Image J. As comparações entre as
médias foram obtidas por análise de variância (ANOVA) de uma via seguida de pós-
teste Newman-Keuls utilizando o programa GraphPad Prism 5. Os grupos foram
considerados significativamente diferentes quando apresentavam probabilidade de
ocorrência ao acaso menor que 5 % (p<0,05).

____________
Resultados

63 IV. Resultados 63 IV. Resultados
IV. RESULTADOS
1- EXPRESSÃO DO RECEPTOR TNF-R1 NOS DIFERENTES TIPOS
CELULARES DA PINEAL.
A glândula pineal é composta de diferentes tipos celulares, os pinealócitos, que
representam a maioria das células do parênquima (90%) e células da glia. A expressão
do receptor TNF-R1 nos diferentes tipos celulares foi determinada pelos ensaios de
dupla imunofluorescência e análise por microscopia confocal.
Para marcação do receptor TNF-R1 utilizamos o anticorpo primário específico
seguido do secundário fluorescente (em verde, FITC). Verificamos uma
imunorreatividade positiva para TNF-R1 em todo parênquima da glândula (figura 5; A e
D). Para identificar os tipos celulares da pineal, utilizamos anticorpos específicos anti-
GFAP, marcador de astrócitos e anti-ED-1, marcador de microglia (ambos em vermelho,
Cy3). As células imuno-positivas para GFAP mostraram ser restritas à região proximal
da glândula, adjacentes ao pedúnculo pineal. Já a imunorreatividade para o marcador
ED-1 mostrou estar difusa ao longo do parênquima.
A sobreposição das imagens com dupla marcação mostra co-localização de TNF-
R1 com astrócitos (figura 5C) e microglia (figura 5E). A análise da imagem em objetiva
maior (figura 6) revela que grande parte das células do parênquima pineal tem
imunorreatividade positiva para TNF-R1 (figura 6, A e D). Praticamente todas as células
que imunoreagem para GFAP também marcam para TNF-R1 (figura 6, C), revelando a
expressão constitutiva do receptor em astrócitos. Em relação a co-localização com
microglia (figura 6, F) apenas parte das células marcadas com anti ED-1 apresentam
imunorreatividade positiva para TNF-R1 (figura 6, F, indicado pela seta), sugerindo que

64 IV. Resultados 64 IV. Resultados
nem todas células da microglia expressam TNF-R1. Como a pineal é formada por 5-10%
de células da glia a marcação para TNF-R1 em células adjacentes (figura 6, F, círculo)
não reativas para marcadores gliais sugere a expressão do receptor em pinealócitos.
FIGURA 5. IImmaaggeennss rreepprreesseennttaattiivvaass ((44XX)) ddaa eexxpprreessssããoo ddee TTNNFF--RR11 eemm sseeççõõeess ddee ggllâânndduullaa
ppiinneeaall ((3300 µµmm)). A marcação positiva de TNF-R1 em todo parênquima pineal é mostrada em
verde (A e D). As células da glia são marcadas em vermelho, astrócitos (GFAP, B) ou microglia
(ED-1, E). As células positivas para GFAP são restritas à região proximal perto do pedúnculo
pineal (B), enquanto as células ED-1-positivas estão dispersas no parênquima (E). Sobreposição
das imagens indica a marcação dupla do TNF-R1 e GFAP (C) ou ED-1 (F). Barra da escala = 200
µm.

65 IV. Resultados 65 IV. Resultados
FIGURA 6. IImmaaggeennss eemm mmaaiioorr oobbjjeettiivvaa((6600XX)) ddaa eexxpprreessssããoo ddoo TTNNFF--RR11 eemm sseeççõõeess ddee ppiinneeaall ddee
rraattooss ((3300 µµmm)).. TNF-R1 é marcado em verde, com imunorreatividade positiva ao longo do
parênquima pineal (A, C e D, F). As células da glia foram marcadas em vermelho com GFAP
para astrócitos e ED-1 para microglia. A microglia apresenta um padrão difuso de distribuição
ao longo do parênquima (E). A marcação em azul representa os núcleos emitida pelo DAPI. As
imagens amarelas mostram a colocalização positiva do TNF-R1 em astrócitos (C) e da microglia
(F). A seta cheia (F) indica células da microglia (vermelho) não positivas para o TNF-R1, e na
cabeça de seta a co-localização positiva entre microglia e TNF-R1 (amarelo). O círculo envolve
um pinealócito, que expressa o TNF-R1 (verde). Barra de escala = 20 µm.

66 IV. Resultados 66 IV. Resultados
2- VARIAÇÃO DA EXPRESSÃO DO TNF-R1 AO LONGO DO DIA
Tendo confirmado que a glândula pineal de ratos expressa constitutivamente o
receptor TNF-R1, na próxima etapa foi avaliada se esta expressão apresenta ritmicidade
na pineal. Para isso, foi utilizado cortes de pineais de ratos obtidas por eutanásia nos
ZTs 2, 10, 14, 18 e 22 (ZT 0 corresponde ao ascender das luzes, 6: 30 h). Através do
método de imunoperoxidase foi possível observar que a expressão deste receptor varia
ao longo das 24 horas, sendo a imunoreatividade maior no ZT10 (figura 7).
FIGURA 7. VVaarriiaaççããoo ddaa iimmuunnoorrrreeaattiivviiddaaddee ppaarraa TTNNFF--RR11 aaoo lloonnggoo ddoo ddiiaa eemm sseeççõõeess ddee ggllâânndduullaa ppiinneeaall.. O
slide mostra a marcação do TNF-R1 em tempos diferentes (ZT 0 corresponde ao ascender das luzes, 6: 30
h). A expressão de TNF-R1 na glândula pineal varia ao longo do dia, sendo a expressão máxima na fase de
clara (ZT 10). O controle negativo foi realizado com a omissão do anticorpo primário. Barra de escala = 50
µm.

67 IV. Resultados 67 IV. Resultados
3. PINEALÓCITOS EXPRESSAM CONSTITUTIVAMENTE O
RECEPTOR TNF-R1
Demonstramos a expressão constitutiva do receptor TNF-R1 em cortes sagitais
da glândula pineal e verificamos que tanto astrócitos como a microglia expressam o
receptor de TNF. Como comentado anteriormente, a existência de células adjacentes
não reativas aos marcadores de células da glia sugeriu que os pinealócitos expressam
constitutivamente esse receptor.
Para confirmar a presença do TNF-R1 em pinealócitos isolamos essas células em
cultura e analisamos a expressão por imunofluorescência. Verificamos que os
pinealócitos apresentam constitutivamente imunorreatividade para o TNF-R1 e que
esta é reduzida ao incubarmos as células com TNF (80 ng/mL) (figura 8). A incubação
com TNF entre 60 e 180 minutos promoveu a redução em aproximadamente 60% da
intensidade de fluorescência (44,18% ± 3.1; basal = 100%), sugerindo a diminuição do
número de receptores expressos nos pinealócitos. O teste de viabilidade celular pelo
ensaio do MTT mostrou que as concentrações de TNF utilizadas não induziram a morte
celular.

68 IV. Resultados 68 IV. Resultados
FIGURA 8. EExxpprreessssããoo ddee TTNNFF--RR11 eemm ppiinneeaallóócciittooss iissoollaaddooss..
A) Imagens representativas da imunofluorescência em pinealócitos cultivados
mostrando a expressão constitutiva do TNF-R1 (marcada em vermelho, à esquerda).
B) Quantificação da fluorescência em pinealócitos estimulados (barras cinza) ou não
(barra branca) com o TNF (80 ng / mL) por 30 a 180 min (basal =100%). Os dados estão
expressos como a média ± e.p.m. n = 4 culturas independentes. Em cada poço foram contadas 21
células escolhidas aleatoriamente em três campos diferentes. * p<0,05 vs TNF 0. Barra da escala=
20 µm.

69 IV. Resultados 69 IV. Resultados
4. TNF PROMOVE A TRANSLOCAÇÃO NUCLEAR DO NFKB
A via do NFKB é considerada central para o processo inflamatório. Essa via
também é ativada pelo TNF através da ativação dos receptores de membrana TNF-R em
diversos tipos celulares. Verificamos se o TNF promove a ativação desse fator na
glândula pineal de rato por meio da quantificação do acúmulo nuclear do NFKB e de
suas subunidades. Para tanto foi utilizado o ensaio de eletromobilidade em gel (EMSA)
e o ensaio de supershift, respectivamente.
44..11.. DDeeccuurrssoo tteemmppoorraall ddaa aattiivvaaççããoo ddoo NNFFKKBB iinndduuzziiddaa ppoorr TTNNFF
Primeiramente avaliamos o tempo de ativação da translocação nuclear do NFKB
induzida por TNF (30 ng/mL, 0, 5, 10, 15, 30 e 60 minutos). Os extratos nucleares de
cada amostra foram incubados com o oligonucleotídeo específico para o NFKB marcado
radiotivamente com γ32P. A sonda específica para o NFKB revelou dois complexos (C1
e C2) nos extratos nucleares da pineal. Observamos, através da quantificação
densitométrica do gel revelado que o TNF aumenta a translocação nuclear de C1 após 5
minutos de incubação e que os níveis nucleares deste fator de transcrição retornam aos
níveis basais após 15 minutos de incubação com TNF (Figura 9).

70 IV. Resultados 70 IV. Resultados
FIGURA 9.. DDeeccuurrssoo tteemmppoorraall ddaa ttrraannssllooccaaççããoo ddoo NNFFKKBB iinndduuzziiddaa ppoorr TTNNFF eemm ggllâânndduullaass
ppiinneeaaiiss ddee rraattoo.
A) Imagem representativa de EMSA da quantidade de NFKB nuclear nos diferentes
tempos de incubação com TNF.
(B) Quantificação densitométrica do NFKB. Os valores são representados pela média ±
e.p.m de 3-4 glândulas por ponto após a incubação com TNF 30 ng/mL em diferentes tempos. *
p<0,05 vs 0 min.

71 IV. Resultados 71 IV. Resultados
44..22.. SSuubbuunniiddaaddeess pprreesseenntteess nnaa ttrraannssllooccaaççããoo nnuucclleeaarr ddoo NNFFKKBB
Observamos que o TNF promove o aumento do acúmulo nuclear do NFKB após
5 minutos de incubação. A identificação das subunidades do NFKB presentes nos
complexos formados foi obtida através do ensaio de supershift. Para isso foi utilizado um
pool de extratos de glândulas tratadas ou não com TNF (30 ng/mL, 5 min) nos quais
foram incubados anticorpos específicos para as diferentes subunidades: p50, RelA, p52,
c-Rel, RelB e Bcl-3. Quando se adiciona um anticorpo com a proteína de interesse
forma-se um complexo maior que promove um retardo ainda maior na migração dos
extratos no gel o que nomeamos de shift , sendo neste caso um supershift.
Tanto no grupo não estimulado como no tratado com TNF, apenas o complexo
C1 apresentou um deslocamento (shift) quando incubado com os anticorpos específicos
para cada subunidade da família NFKB (Figura 10). Extratos nucleares de glândulas não
estimuladas tiveram retardo para p50 e RelA mas não para os outros anticorpos
testados. A incubação com anticorpo anti-p50 promove o deslocamento de todo o
complexo, enquanto que RelA promove um deslocamento parcial de C1. Um gel
demonstrativo é mostrado na figura 10A. É interessante notar que para p50 observamos
duas bandas de retardo (figura 10B, indicado pela primeira seta) enquanto que para
RelA só há uma banda (figura 10B, segunda seta). Portanto, podemos sugerir que na
glândula pineal em cultura (48h) tanto os dímeros p50/p50 e p50/RelA são presentes no
extrato nuclear.

72 IV. Resultados 72 IV. Resultados
FIGURA 10. EEnnssaaiioo ddee SSuuppeerrsshhiifftt ddee ggllâânndduullaass ppiinneeaaiiss eemm ccuullttuurraa..
A) Autoradiografia representativa mostrando o supershift para p50 e RelA em pineais
não-estimuladas.
B) Gráfico obtido a partir do analisador de gel do Programa Image J. Os gráficos
mostram, em cada linha, a banda de retardo (supershift, SS) correspondente de cada coluna do gel
representativo. O primeiro e o segundo picos correspondem respectivamente a C2 e C1. Cada
ponto foi obtido de um pool de 3-4 glândulas

73 IV. Resultados 73 IV. Resultados
44..33.. EEffeeiittoo ddoo TTNNFF ssoobbrree aa ttrraannssllooccaaççããoo ddooss ddíímmeerrooss ddoo NNFFKKBB
O efeito do TNF sobre a translocação dos dímeros do NFKB foi avaliado pela
incubação das glândulas por 5 minutos em concentrações crescentes (10, 30, 60, 90
ng/mL). TNF induz a translocação nuclear de ambos os dímeros (p50/p50 e p50/RelA)
(Figura 11). Concentrações mais baixas são capazes de aumentar a translocação nuclear
de p50/RelA, enquanto que as concentrações mais altas promovem a translocação de
ambos os dímeros.
FIGURA 11. EEffeeiittoo ddoo TTNNFF ssoobbrree aa ttrraannssllooccaaççããoo ddaass ssuubbuunniiddaaddeess ddoo NNFFKKBB ppaarraa oo nnúúcclleeoo ddee
ppiinneeaaiiss eemm ccuullttuurraa..
A) Gel representativo do supershift para RelA de glândulas estimuladas ou não com o
TNF (10-90 ng / mL) por 5 min.
B) Gel representativo do supershift para p50 de glândulas estimuladas ou não com o TNF
(10-90 ng/mL) por 5 min. Os gráficos abaixo representam a análise densitométrica dos géis
mostrados acima feita pelo software Image J. Cada ponto foi obtido a partir de um pool de 3-4
glândulas.

74 IV. Resultados 74 IV. Resultados
5. TNF PROMOVE A DEGRADAÇÃO DA PROTEÍNA INIBITÓRIA
IKBα EM PINEALÓCITOS DISPERSOS
Na maioria dos tipos celulares a ativação do complexo NFKB é regulada por uma
família de proteínas citoplasmáticas conhecidas como inibidoras do NFKB, NFKBI
(antes citadas como IKBs). Essas proteínas inibitórias retêm o complexo NFKB no
citoplasma, impedindo sua translocação ao núcleo e, portanto, sua associação com DNA
(Rice & Ernst, 1993). A ativação do NFKB depende da fosforilação do IKBα e
subsequente ubiquitinação que resulta na degradação do IKBα pelo proteassoma. A
liberação do NFKB permite sua translocação para o núcleo onde ele atua na transcrição
de vários genes.
Para avaliar a ativação do fator NFKB em pinealócitos isolados observamos a
expressão da proteína inibitória IKBα através de estudos imunocitoquímicos. A
expressão constitutiva da proteína é considerada como 100% do valor da fluorescência
detectada. Verificamos que o TNF (80 ng/mL) induz uma diminuição significativa (de
100% ± 3,5 para 30,1 % ± 2,0) da imunorreatividade para proteína inibitória IKB-α após
10 minutos de incubação. A redução em relação a imunofluorescência basal é mantida
dos 15 aos 60 minutos. A partir dos 15 minutos a fluorescência relativa tende a aumentar
significativamente em relação aos 10 minutos, porém não restaurando os níveis basais. A
diminuição da fluorescência após estimulação com TNF sugere a degradação da
proteína e a consequente ativação do NFKB em pinealócitos isolados (figura 12).

75 IV. Resultados 75 IV. Resultados
FIGURA 12. EExxpprreessssããoo ddoo IIKKBBαα eemm ppiinneeaallóócciittooss iissoollaaddooss..
A) Imagens representativas da imunocitoquímica mostrando a expressão do IKBα
(marcado em vermelho) em cultura de pinealócitos na ausência (esquerda) ou presença de TNF
(80 ng/mL, por 10 min; imagem à direita). Em azul estão representados os núcleos dos
pinealócitos marcados com DAPI. Barra da escala = 10 µm.
B) Quantificação da fluorescência em pinealócitos estimulados (barra cinza) ou não
(barra branca) com TNF (80 ng/mL) de 1 a 60 min. Dados são apresentados como média ± e.p.m.
(basal =100%). n = 3 experimentos independentes. *p < 0.05 vs. TNF 0.

76 IV. Resultados 76 IV. Resultados
6. TNF AUMENTA A EXPRESSÃO DE INOS EM PINEALÓCITOS
ATRAVÉS DE MECANISMO DEPENDENTE DE NFKB..
Uma vez que TNF induz a translocação nuclear dos dímeros p50/RelA, utilizamos
como medida funcional a expressão da enzima iNOS e NO em pinealócitos isolados.
66..11.. TTNNFF iinndduuzz aa eexxpprreessssããoo ddaa ssiinnttaassee ddee óóxxiiddoo nnííttrriiccoo iinndduuzzíívveell ((iiNNOOSS)) eemm
ppiinneeaallóócciittooss iissoollaaddooss
A expressão de iNOS foi verificada por imunofluorescência em pinealócitos
dispersos. Pinealócitos isolados foram incubados com concentrações crescentes de TNF
(30, 60, 90 ng/mL, por 120 min). Os resultados demonstram que o TNF (120 min.) induz o
aumento concentração-dependente da imunorreatividade para iNOS, sendo
estatisticamente significativo nas concentrações de 60 e 90 ng/mL (figura 13A).
A enzima iNOS é a isoforma de sintase de NO que depende de indução
transcricional. O promotor da enzima iNOS possui sítio de ligação tanto para o NFKB como
para o fator de transcrição AP-1 sendo que este último varia entre indução e repressão (Xie
et al., 1994, Lowestein et al, 1993, Chu et al.,1998). A ativação do NFKB é essencial para
indução da transcrição de iNOS por IL-1 e LPS (Di Marco et al, 2005). Para avaliar se o
efeito da indução de iNOS por TNF em pinealócitos isolados é dependente da ativação do
fator NFKB utilizamos o inibidor sintético da via NFKB, pirrolidina ditiocarbamato
(PDTC), (Schreck et al., 1992). Verificamos que o PDTC (25 M), pré-incubado por 30
minutos em pinealócitos isolados, inibe a expressão de iNOS induzida por TNF (80ng/mL,
120 min) (Figura 13, B)

77 IV. Resultados 77 IV. Resultados
FIGURA 13. EExxpprreessssããoo ddee iiNNOOSS iinndduuzziiddaa ppoorr TTNNFF eemm ppiinneeaallóócciittooss eemm ccuullttuurraa.
A) Imagens representativas da imuno-histoquímica para iNOS (marcação em vermelho)
em pinealócitos estimulados ou não com TNF (30-90 ng/mL por 120 min). Os núcleos dos
pinealócitos estão marcados em azul com DAPI.
B) Quantificação da fluorescência relativa da imunorreatividade para iNOS nas
diferentes concentrações de TNF (30, 60 e 90 ng/mL). (basal =100%). *p < 0.05 vs. TNF 0.
C) Quantificação da fluorescência relativa da imunorreatividade para iNOS em
pinealócitos incubados com TNF (80 ng/mL, 120 min) e com PDTC (25µM). Os dados são
expressos como média ± e.p.m. n = 3 experimentos independentes para cada parâmetro *p < 0.05
vs. TNF 0 (barra branca); **p < 0.05 quando comparado o grupo estimulado ou não com TNF na
presença ou ausência do PDTC.

78 IV. Resultados 78 IV. Resultados
7. TNF INDUZ A PRODUÇÃO DE ÓXIDO NÍTRICO EM
PINEALÓCITOS ISOLADOS
Tendo em vista que o TNF é capaz de induzir o aumento da expressão de iNOS em
pinealócitos isolados, analisamos a habilidade das células estimuladas por esta citocina em
produzir óxido nítrico. Para tanto utilizamos um método de detecção in situ do óxido
nítrico através do marcador fluorescente DAF-FM DA e analisamos por microscopia
confocal.
Para avaliar o efeito do TNF sobre a produção de NO foi utilizada a dose
submáxima desta citocina necessária para a indução de iNOS (80 ng/ml). Observamos
que pinealócitos incubados por 2 horas com TNF tiveram aumento significativo da
intensidade de fluorescência em relação ao basal (para 174,8% ± 8,6% vs basal)
indicando o acúmulo de NO (Figura 14). Com o objetivo de verificar se produção de NO
é mediada pela indução de iNOS, pinealócitos foram pré-incubados com o inibidor
específico da enzima, 1400W (1 µM, 50 min). Constatamos que a produção de NO
induzida por TNF é bloqueada quando as células são previamente tradadas com o
inibidor de iNOS (90,5% ± 5,7 vs grupo tratado com TNF).

79 IV. Resultados 79 IV. Resultados
FIGURA 14. TTNNFF iinndduuzz aa pprroodduuççããoo ddee NNOO eemm ppiinneeaallóócciittooss eemm ccuullttuurraa..
A) Imagens representativas do acúmulo de NO induzido por TNF em pinealócitos
isolados. As imagens mostram a fluorescência do marcador de NO DAF-FM em pinealócitos
sem marcador (SALINA), células incubadas apenas com marcador (salina mais DAF-FM) e
células carregadas com marcador e estimuladas com TNF (80 ng/mL). Escala de 30 µm (barra
verde).
B) Os dados estão apresentados como média ± e.p.m da porcentagem de fluorescência
emitida pelo marcador de NO (basal =100%). n = 3 experimentos independentes. *p < 0,05 vs.
TNF 0 (barra branca); **p < 0,05 quando comparado o grupo estimulado ou não com TNF na
presença ou ausência do 1400W.

____________
Discussão

81 V. Discussão 81 V. Discussão
V. DISCUSSÃO
A regulação da produção de melatonina pela glândula pineal era considerada
exclusivamente dependente da ativação da via simpática pela alternância claro/escuro,
ou pela inibição da mesma por fibras GABAérgicas estimuladas pela luz (Simmoneaux
& Ribelayga, 2003). Desta forma, a pineal era o único órgão que não respondia
diretamente a estímulos endógenos, mas sim à variação da iluminação ambiental. No
entanto, alguns autores sugeriam que a glândula pineal poderia ser alvo de citocinas e
hormônios, de tal forma que a produção noturna de melatonina seria regulada pelo
estado de higidez do animal (Skwarlo-Sonta et al., 2003, Nelson et al., 2002).
A citocina pró-inflamatória TNF é capaz de suprimir a via biossintética da
melatonina induzida por noradrenalina (Fernandes et al., 2006). Uma correlação
negativa entre melatonina e TNF em pacientes septicêmicos (Mundigler et al., 2002),
portadores de infarto do miocárdio (Dominguez-Rodriguez et al., 2004) e parturientes
portadoras de mastite (Pontes et al., 2006), reforça a hipótese que o TNF é capaz de
suprimir a produção noturna de melatonina. Por outro lado, os glicocorticóides, que são
potentes agentes anti-inflamatórios, potenciam a produção de melatonina tanto in vitro
quanto in vivo (Ferreira et al., 2005, Couto-Moraes et al., 2009; Fernandes et al., 2009).
Já é sabido que a glândula pineal expressa receptores de glicocorticóides, que
inibindo a via de sinalização NFKB, potenciam a produção de melatonina induzida por
noradrenalina (Ferreira et al., 2005; Fernandes et al., 2009). Neste trabalho foram
determinados os mecanismos moleculares envolvidos na resposta induzida por TNF,
sobre a glândula pineal, dando subsídio para o entendimento de como esta citocina
pode inibir a produção noturna de melatonina.

82 V. Discussão 82 V. Discussão
Detectamos por técnicas de imuno-histoquímica e imunocitoquímica a
marcação com anticorpos específicos para receptores TNF-R1 e TNF-R2 tanto em
pinealócitos quanto em células da glia. Vale ressaltar que a expressão de TNF-R1 é
difundida entre diferentes tecidos, tais como glandulares, musculares, neurais e células
imunocompetentes, enquanto que TNF-R2 tem sido encontrado preferencialmente em
células do sistema imune (Hehlgans & Männel, 2002). Para que fosse avaliada a
localização celular dos receptores TNF-R1 nas células da glândula pineal, inicialmente
caracterizamos os tipos celulares presentes usando imunomarcadores específicos para
astroglia e microglia.
Os estudos imuno-histoquímicos confirmaram que os astrócitos estão restritos à
região proximal, ou pedúnculo por onde penetram artéria, veia e fibras nervosas (Luo et
al., 1984, Zang et al., 1985, Schröder & Malhotra, 1987; Berger & Hedger, 2000; Jiang-
Shieh et al., 2003). Já a microglia difunde-se por todo o parênquima glandular.
A imunoreatividade positiva para TNF-R1 pode ser co-localizada com a dos
marcadores GFAP para astrócitos e ED-1 para microglia. Todas as células marcadas com
anticorpos contra GFAP (astrócitos) apresentaram dupla marcação para TNF-R1,
sugerindo fortemente uma expressão constitutiva destes receptores na astroglia. Por
outro lado, apenas algumas das células imunorreativas para ED-1 (microglia)
apresentavam colocalização para TNF-R1. Esta marcação parcial da microglia pode ser
dependente da existência de diferentes tipos de células, como sugerido em estudo com
anticorpos mais seletivos (Jiang-Shieh et al., 2003). Desta forma, podemos considerar
que nem todas as células que compõe a microglia expressam receptores para TNF-R1.
Receptores que reconhecem moléculas sinalizadoras de patógenos (PAMPS) já
foram detectados na glândula pineal. Foi mostrada a expressão constitutiva dos
receptores TLR-4 e CD-14 que reconhecem o principal componente da parede de

83 V. Discussão 83 V. Discussão
bactérias gram-negativas (LPS) (da Silveira Cruz-Machado et al., 2010). A expressão
desses receptores tem sido relatada na literatura como restrita a células da microglia e
também astroglia. Já na pineal, foi verificado que além dos astrócitos e da microglia os
pinealócitos também expressam o receptor TLR-4. A expressão constitutiva desses
receptores nos órgãos circumventriculares estudados é crucial para reação imune inata
no sistema nervoso central (Laflamme & Rivest, 2001). A detecção do LPS circulante
promove uma rápida resposta pró-inflamatória, primeiro nesses órgãos e
posteriormente é perceptível ao longo do parênquima cerebral (Rivest, 2003). Assim
que a glia é estimulada o LPS circulante é capaz de causar uma rápida ativação da
transcrição dos genes que codificam a proteína CD14 como, por exemplo, os das
citocinas IL-1β, IL-6 e TNF gerando um sinal amplificador da resposta pró-inflamatória
também em outras regiões encefálicas. A presença constitutiva desses receptores na
pineal amplia o entendimento de que a pineal é também regulada pelo estado
fisiopatológico do animal estando instrumentalizada a reconhecer moléculas específicas
de patógenos.
Os dados da literatura para o receptor de TNF-R1 na glia ainda são esparsos e
não permitem fazer a mesma análise. No entanto, seria muito especulativo imaginar que
o mesmo ocorreria para o TNF-R1 na pineal, visto que o efeito de TNF em culturas de
pineal é máximo após 12 horas de incubação, sendo que após 24 ou 48 horas não mais é
observada a supressão da produção de N-acetilserotonina induzida por noradrenalina
(Fernandes et al., 2006).
A imunoreatividade para o TNF-R1 no parênquima da pineal foi observada tanto
em células que imunorreagiam ao ED-1, quanto as que não apresentavam dupla-
marcação, sugerindo que os pinealócitos também apresentavam imunorreatividade para
o TNF-R1. A presença destes receptores em pinealócitos foi confirmada em estudo

84 V. Discussão 84 V. Discussão
imunocitoquímico realizado com pinealócitos dispersos. Esta constatação deve ser
destacada aqui. A presença constitutiva do receptor TNF-R1 também nos pinealócitos
indica que estas células podem responder diretamente ao TNF. Isso sugere que
independente da ativação de células gliais, as células produtoras de melatonina podem
detectar e responder ao TNF presente na circulação. A especulação que se fazia até
então era que as células gliais, a se destacar a microglia, seriam as células moduladoras
da resposta pineal às citocinas. De fato foi mostrado que a presença da microglia em
cultura de pinealócitos influencia o padrão de resposta às citocinas IL-1β, IFN-γ e ao
TGFβ, mas em relação ao TNF, interessantemente, a resposta é a mesma independente
da constituição de microglia (Tsai et al., 2001b). Este trabalho corrobora com os nossos
dados que mostram a presença de receptores de TNF-R1 também nos pinealócitos,
sugerindo a resposta direta do TNF nestas células.
Após caracterizar a presença dos receptores, nosso objetivo foi avaliar a
funcionalidade dos mesmos. Vale ressaltar que o agonista endógeno para TNF-R1 é o
TNF solúvel, enquanto TNF-R2 interage com a forma ligada à membrana (Grell et al.,
1998; McCoy e Tancey, 2008).
A ligação do TNF com o receptor TNF-R1 promove a ativação da cascata de
sinalização que leva à translocação nuclear do fator de transcrição NFKB (Tracey et al.,
2008). Aqui verificamos que o TNF induz ao aumento da translocação nuclear do NFKB
em pineais em cultura. Este aumento ocorre de maneira rápida, após 5 minutos de
estimulação, e transiente, retornando aos níveis basais em 15 minutos. A cinética da
ativação do NFKB pelo TNF é descrita na literatura por ser rápida e temporalmente
regulada (Hoffmann et al., 2002; Werner et al., 2008). As alças de retroalimentação
negativa controladas pelas proteínas IKBs são determinantes para rápida finalização da
resposta do NFKB. Tão logo o NFKB transloca ao núcleo após a estimulação com TNF

85 V. Discussão 85 V. Discussão
ele induz a transcrição de novas proteínas IKBα que, por sua vez se ligam aos dímeros
do NFKB sequestrando-os para o citoplasma na forma inativa (Werner et al., 2008). Já o
tempo de indução da resposta ao TNF é variável e depende do estado basal de ativação
do NFKB e das quantidades de IKBα disponíveis no núcleo (Lipniack et al., 2004).
Em pineais de ratos em cultura por 48 horas obtidas de animais eutanasiados às
16h horas, encontramos a presença dos dímeros de NFKB p50/p50 e p50/RelA. Isto
contrastou com os resultados obtidos com pineais de ratos não cultivadas (Cecon et al.,
2010), ou cultivadas por 48 horas, mas obtidas de animais mortos às 9h horas (da
Silveira Cruz-Machado et al., 2010). Nestes casos apenas dímeros p50/p50 foram
observados no núcleo. Este dímero é constitutivamente ativado na pineal apresentando
um ritmo diário, com maior translocação nuclear no final da fase de claro e queda
abrupta no início da fase de escuro (Cecon et al., 2010). Uma vez que p50/p50 não possui
o domínio de transativação (Hayden & Ghosh, 2008), ele atuaria como repressor da
transcrição gênica o que explicaria seus baixos níveis durante a noite, permitindo a
produção de melatonina. De fato a inibição do NFKB constitutivo potencia a síntese de
melatonina induzida por noradrenalina em pineais em cultura (Ferreira et al., 2005).
A estimulação das glândulas pineais com TNF aumentou o conteúdo nuclear
tanto do homodímero p50/p50 quanto do heterodímero p50/RelA. O aumento da
translocação nuclear de p50/p50 vai ao encontro dos achados da regulação desse dímero
sobre da produção de melatonina, uma vez que o TNF inibe a via biossintética da
melatonina (Fernandes et al., 2006). Além disso, estudos in silica indicam a presença de
elementos kappa B no promotor do gene da enzima AA-NAT, que é chave na via
biossintética da melatonina (Markus et al., 2007). A ligação de um dímero de NFKB
desprovido de domínio de transativação como é o caso de p50/p50 (Karin & Ben-
Neriah, 2000) resultaria na inibição da transcrição do gene da AA-NAT.

86 V. Discussão 86 V. Discussão
A estimulação com TNF também promove o aumento da translocação de
p50/RelA. A ativação da translocação nuclear deste dímero está ligada à via clássica do
NFKB (Karin & Ben-Neriah, 2000; Perkins, 2007), que é essencial para indução da
transcrição de uma variedade de genes essenciais para a montagem da resposta
inflamatória. Genes que regulam a expressão de moléculas de adesão, citocinas,
quimiocinas e enzimas tais como a sintase de óxido nítrico induzida (iNOS) e a
ciclooxigenase II (COX II), são alguns cuja transcrição é induzida pelo dímero p50/Rel
A (para revisão, Lawrence, 2009).
Desta forma, o aumento do conteúdo nuclear de p50/RelA em glândulas pineais
em cultura após estimulação com TNF deveria resultar na expressão de proteínas
envolvidas na montagem de respostas inflamatórias. De fato, a ativação desses dímeros
induz a produção de TNF, observada após incubação de glândulas pineais com LPS que
também é temporalmente coincidente com o aumento da expressão do receptor TNF-
R1 em pinealócitos isolados (da Silveira Cruz-Machado et al., 2010). Interessante
destacarmos aqui, que o TNF produzido localmente possa também exercer efeitos
parácrinos na pineal amplificando o sinal gerado inicialmente pelo LPS para promoção
da inibição da síntese de melatonina. No presente trabalho, escolhemos avaliar a
expressão da enzima iNOS quando as culturas de pinealócitos eram estimuladas com
TNF.
Vale ressaltar que são conhecidas três isoformas da enzima NOS, sendo duas
constitutivas e uma induzida. A pineal expressa a enzima constitutiva neuronal, e
apesar de ainda não ser entendida a sua função, sabe-se que sua expressão tem
regulação fotoperiódica (Jacobs et al., 1999).
Nossos dados demonstram, que o TNF é capaz de induzir ao aumento da
imunorreatividade para iNOS, sugerindo a indução desta enzima. Além disso, esta
indução é dependente da concentração de TNF e antagonizada por inibidores da ligação

87 V. Discussão 87 V. Discussão
de NFKB com seu elemento cognato no promotor dos genes controlados por este fator
de transcrição. Portanto, fica pela primeira vez demonstrado que há indução de iNOS
em pineais ativadas por TNF e que é controlada pela via de transcrição NFKB. A
produção de NO foi também demonstrada em pinealócitos estimulados com TNF, e a
incubação com o inibidor da enzima iNOS bloqueia esse efeito indicando que o acúmulo
de NO é gerado a partir de sua atividade. Vale ressaltar que a ativação do NFKB medeia
a inibição da produção de melatonina, e que doadores de NO reduzem a produção de
melatonina em glândulas pineais (Maronde et al., 1995). Esses dados podem sugerir que
o efeito do TNF sobre a produção de óxido nítrico via iNOS seja relacionada à resposta
inibitória na glândula. Esta especulação indica que a ativação da via NFKB através do
recrutamento de dímeros diferentes promove respostas transcricionais negativas e
positivas mas convergindo para um mesmo efeito.
Em suma, os resultados apresentados aqui, demonstram a base molecular para
considerar as células da pineal alvos para o TNF e, neste contexto, amplia o nosso
entendimento de que a pineal é parte integrante do sistema de alerta do organismo para
detecção de injúrias (figura 15). Podemos ressaltar que ativação do fator NFKB é
essencial para resposta ao TNF na glândula pineal. Confirmamos aqui, que este mesmo
estímulo pode ativar cascatas regulatórias diferentes. Enquanto a ativação dos dímeros
p50/p50 explicam o efeito inibitório desse fator sobre a produção de melatonina, o
acúmulo de p50/RelA nos permite especular que a ativação da transcrição de genes
também é um mecanismo relevante para resposta imune inata da pineal. Nossos
resultados corroboram com dados anteriores do nosso grupo que mostram ser a pineal
apta a responder a estímulos injuriantes. Portanto, corroboram a existência do eixo-
imune-pineal, ativo durante a montagem de uma resposta inflamatória.

88 V. Discussão 88 V. Discussão
FIGURA 15. RReepprreesseennttaaççããoo eessqquueemmááttiiccaa ddoo eeffeeiittoo dduupplloo ddoo TTNNFF eemm ppiinneeaallóócciittooss. O TNF ativa
o TNF-R1, levando à interiorização do receptor e a ativação da via NFKB. Esta ativação é
mediada pela degradação do IKBα, que depende de sua fosforilação (Neumann e Naumann,
2007). Na sequência observou-se a translocação nuclear dos dímeros p50/p50 e p50/RelA. É
sabido que p50/p50 atua inibindo a transcrição gênica enquanto p50/RelA é capaz de ativar a
transcrição de genes. Foi mostrado previamente que o TNF inibe a transcrição do gene Aa-nat,
que codifica a proteína-chave para a síntese de melatonina (Fernandes et al., 2006). Além disso,
aqui demonstramos que a translocação nuclear de p50/RelA induz a transcrição da iNOS, uma
enzima clássica de respostas inflamatórias. Sugerimos que o mecanismo de ação do TNF,
divulgado no presente trabalho, é a base molecular para a compreensão da inibição da síntese de
melatonina, que ocorre no início de uma resposta imune.

____________
Conclusões

90 VI. Conclusões
VI. CONCLUSÕES
1. Glândulas pineais de rato expressam de maneira constitutiva o receptor TNF-
R1. Este receptor é expresso nos três principais tipos celulares da pineal, em
pinealócitos, e nas células gliais, astrócitos e microglia.
2. A expressão do receptor TNF-R1 no parênquima da glândula pineal varia ao
longo dia, sendo maior no ZT 10.
3. Pinealócitos expressam o receptor TNF-R1. Na presença de TNF a expressão
do TNF-R1 é reduzida após 60 minutos de incubação.
4. Os dímeros do NFKB p50/p50 e p50/RelA são constantemente ativados em
pineais em cultura (retiradas no ZT 10). A estimulação com TNF promove o aumento da
translocação dos mesmos.
5. A proteína inibitória IKBα é expressa em pinealócitos em cultura. A presença
do TNF promove a redução da expressão dessa proteína, sugerindo sua degradação.
Portanto podemos concluir que o NFKB é ativado por TNF em pinealócitos em cultura.
6. O TNF induz a expressão da enzima iNOS em pinealócitos isolados. Vimos
que essa indução é dependente da ativação do fator NFKB e leva a produção de NO.

91 VII. Resumo
VII. RESUMO
O TNF atua diretamente sobre a pineal inibindo a via biossintética da
melatonina. De forma semelhante o LPS, potente ativador da resposta imune inata, tem
efeito inibitório sobre a produção de melatonina e seu precursor. A supressão da síntese
noturna de melatonina é admitida de forma a permitir a montagem da resposta
inflamatória, tanto durante o dia como à noite. Dados do grupo tem demonstrado que o
NFKB é um fator constitutivamente expresso na glândula pineal e que o ritmo diário de
translocação pode ser determinante para o início da síntese de melatonina. A inibição de
sua ativação potencia a síntese de melatonina enquanto que fatores que classicamente
promovem a sua ativação inibem esta produção. Nesta dissertação, verificamos as
moléculas envolvidas no reconhecimento e na de sinalização desta citocina na pineal.
Demonstramos que a pineal está apta a responder ao TNF, células gliais da pineal e as
células secretoras, os pinealócitos, expressam constitutivamente o receptor TNF-R1. O
TNF promove a translocação nuclear dos dímeros p50/p50 e p50/RelA do NFKB em
glândulas pineais em cultura. Confirmamos a ativação desta via em pinealócitos pela
análise da proteína inibitória IKBα. Vimos que a ativação desse fator é essencial para
expressão da iNOS e a produção de NO induzida por TNF em pinealócitos isolados.
Além disso, mostramos que o TNF promove a redução da expressão do receptor TNF-
R1 na membrana de pinealócitos. Em resumo, mostramos neste trabalho que a pineal
está instrumentalizada a responder ao TNF, e que os pinealócitos são alvos diretos para
o seu reconhecimento. Confirmamos a relevância do fator NFKB na resposta da pineal
em situações de injúria, ampliando o conceito de que a pineal está apta a detectar
moléculas do processo inflamatório.

92 VIII. Abstract
VIII. ABSTRACT
TNF acts directly on the pineal gland by inhibiting the biosynthesis of
melatonin. Similarly, LPS, a potent activator of the innate immune response has an
inhibitory effect on the production of melatonin and its precursor. The suppression of
nocturnal melatonin synthesis is admitted to allow the full mounting of the
inflammatory response both during the day and night. Data from our laboratory have
shown that the NFKB is constitutively expressed in the pineal gland and that the daily
rate of translocation can be determinant for the onset of melatonin synthesis. Inhibition
of their activation potentiates melatonin synthesis whereas factors that promote its
activation classically inhibit this production. In this dissertation we find the molecules
involved in recognition and signaling of this cytokine in the pineal. We demonstrated
that the pineal is responsive to TNF, the pineal glial cells and secretory cells, the
pinealocytes, constitutively express the TNF-R1. TNF promotes nuclear translocation
of p50/RelA and p50/p50 NFKB dimers in the pineal glands in culture. We confirmed
the activation of this pathway in pinealocytes by analyzing the inhibitory protein IKBα.
We have seen that the activation of this factor is essential for iNOS expression and NO
production induced by TNF in isolated pinealocytes. Furthermore we show that TNF
promotes the reduction of the expression of TNF-R1 receptor on the membrane of
pinealocytes. In summary, we show here that the pineal is instrumented to respond to
TNF and that the pinealocytes are direct targets for its recognition. We confirm the
relevance of NFKB in the pineal response in situations of injury extending the concept
that the pineal is able to detect molecules in the inflammatory process and respond
accordingly.

93 IX. Referências Bibliográficas
IX. REFERÊNCIAS BIBLIOGRÁFICAS
ARENDT, J. (1995). Melatonin and the Mammalian Pineal Gland Chapman and Hall, London.
AXELROD, J. & WEISSBACH, H. (1960). Enzymatic O-methylation of N-
acetylserotonin to melatonin. Science 131: 1312. AXELROD, J. et al. (1965). Control of hydroxyindole-O- methyltransferase activity in
the rat pineal gland by environmental lighting. J Biol Chem. 240: 949–954. BARGMANN, W. (1943). Die Epiphysis cerebri. In Handbuch der mikroskopischen
Anatomie des Menschen, vol. 6:4 (ed. W. von Mollendorff), pp. 309–502. Berlin: Springer.
BENITEZ-KING, G. (2006). Melatonin as a cytoskeletal modulator: implications for
cell physiology and dis-ease. J Pineal Res. 40: 1–9. BEUTLER, B. & VAN HUFFEL, C. (1994). Unravelling function in the TNF ligand and
receptor families. Science 264: 667 – 668. BILICI, D. et al. (2002). Protective effect of melatonin in carrageenan-induced acute
local inflammation. Pharmacol. Res. 46: 133–139.
BLACK, R.A. (2002). Tumor necrosis factor-α converting enzyme. Int. J. Biochem. Cell. Biol. 1: 1 – 5.
BLACK, R.A. et al. (1997). A metallo-proteinase disintegrin that releases tumour-necrosis factor-α from cells. Nature 385: 729 – 733
BOJKOWSKI, C.J. et al. (1987). Melatonin secretion in humans assessed by measuring its metabolite, 6-sulfatoxymelatonin. Clin Chem. 33(8): 1343-8.
BONIZZI, G. & KARIN, M. (2004). The two NF-κB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 25(6): 280-8.
BORJIGIN, J. et al. (1995) Diurnal variation in mRNA encoding serotonin N-
acetyltransferase in pineal gland. Nature 378: 783–785.
BRADLEY, J.R. (2008). TNF-mediated inflammatory disease. J Pathology: 149-160.
BRAINARD, G.C et al. (1988). Neuroendocrine regulation of immune parameters. Photoperiod control of the spleen in Syrian hamster. Ann. N.Y. Acad. Sci. 540: 704-706.

94 IX. Referências Bibliográficas
BRENNAN, F.M., et al. (1989). Inhibitory effect of TNF alpha antibodies on synovial cell interleukin-1 production in rheumatoid arthritis. Lancet. 2:244−247.
BUBENIK, G.A. (2002). Gastrointestinal melatonin: localization, function, and clinical
relevance. Dig Dis Sci. 47: 2336–2348.
CALVO, J. & BOYA (1983). Postnatal development of cell types in the rat pineal gland. J.Anat. 137: 185-191.
CARDINALI, D.P. et al. (1972). Binding of melatonin to human and rat plasma proteins, Endocrinology 91: 1213–1218.
CARLBERG, C. (2000) Gene regulation by melatonin. Ann N Y Acad Sci. 917: 387–396. CARLOTTI, F. et al. (2000). Dynamic shuttling of nuclear factor kappa B between the
nucleus and cytoplasm as a consequence of inhibitor dissociation. J Biol Chem. 275: 41028-41034.
CARRILO-VICO, A. et al. (2004). Evidence of melatonin synthesis by human
lymphocytes and its physio- logical significance: possible role as intracrine,
autocrine and/ or paracrine substance. FASEB J. 18: 537–539.
CARRILO-VICO, A. et al. (2005). Human lymphocyte-synthesized melatonin is
involved in the regulation of the interleukin-2/interleukin-2 receptor system. J Clin
Endocrinol Metab 90: 992–1000.
CARSWELL, A. et al. (1975). An endotoxin-induced serum factor that causes necrosis of
tumors. Proc Natl Acad Sci USA 72: 3666-3670.
CECON, E. et al., (2010). Daily variation of constitutively activated nuclear factor kappa B (NFKB) in rat pineal gland. Chronobiol Int. 27: 52-67.
CERAMI, A. & BEUTLER, B. (1988). The role of cachectin/TNF in endotoxic shock and
cachexia. Immunol Today. 9(1): 28-31.
CHEN, et al, (2002). NF-R1 signaling: a beautiful pathway. Science. 296(5573): 1634-5.
CIPOLLA-NETO, J. et al., (1994). The Effects of Intergeniculate Leaflet Lesion on The Metabolism of the Rat Pineal Gland. Adv Pineal Res. 8: 95-10.
COON, S.L. et al. (1995). Pineal serotonin N-acetyltransferase: expression cloning and
molecular analysis. Science. 270: 1681-1683. COUTO-MORAES, R. et al., (2009). The immune-pineal axis: stress as a modulator of
pineal gland function. Ann N Y Acad Sci. 1153: 193-202.

95 IX. Referências Bibliográficas
CSABA, G. & BARATH, P. (1975). Morphological changes of thymus and the thyroid
gland after postnatal extirpation of pineal body. Endocrinol. Exp. 9: 59-67.
DA SILVEIRA CRUZ-MACHADO, S. et al. (2010). TLR4 and CD14 receptors expressed in rat pineal gland trigger NFKB pathway. J Pineal Res. 49: 183–192.
DEMAS, G.E. et al. (1997). Metabolic costsof mounting an antigen-stimulated immune
response in adult and aged C57BL/6J mice. Am. J. Physiol. 273: 1631–1637. DOBRZANSKI, P. et al. (1995). Specific inhibition of RelB/p52 transcriptional activity
by the C-terminal domain of p100. Oncogene 10: 1003–7. DOMINGUEZ-RODRIGUEZ, A. et al. (2004). Light/dark patterns of interleukin-6 in
relation to the pineal hormone melatonin in patients with acute myocardial
infarction. Cytokine 26(2): 89-93.
DUBOCOVICH, M.L. et al. (1997). Melatonin receptor antagonists that differentiate between the human Mel1a and Mel1b recombinant subtypes are used to assess the pharmacological profile of the rabbit retina ML1 presynaptic heteroreceptor. Naunyn-Schmiedeberg’s. Archives of Pharmacology. 355 (3): 365–375
DUBOCOVICH, M.L. et al. (2000). Melatonin receptors. In The IUPHAR Compendium
of Receptor Characterization and Classification, 2nd edn. (IUPHAR, ed.), pp. 271–277. IUPHAR Media, London.
DUBOCOVICH, M.L. et al. (2010). International Union of Basic and clinical
Pharmacology. LXXV. Nomenclature, classification and pharmacology of G protein-coupled melatonin receptors. Pharmacol Rev. (in press).
DUVERNOY, H.M. & RISOLD, P.Y. (2007). The circumventricular organs: an atlas of
comparative anatomy and vacularization. Brain Res Rev. 56: 119-147. EDER, J. (1997). Tumor necrosis factor and Il-1 signaling: do MAPKK kinases connect it
all? Trends Pharmacol. Sci. 18: 319-322.
EKSTRÖM, P. & HILMAR, M. (2003). Evolution of photosensory pineal organs in new light: the fate of neuroendocrine photoreceptors. Phil. Trans. R. Soc. Lond. 358 (1438): 1679-700.
FERNANDES, P.A. et al. (2009). Local corticosterone infusion enhances nocturnal
pineal melatonin production in vivo. J Neuroendocrinol. 21: 90-97. FERNANDES, P.A. et al. (2006). Effect of TNF-alpha on the melatonin synthetic
pathway in the rat pineal gland: basis for a “feedback” of the immune response on circadian timing. J Pineal Res. 41: 344-250.
FERREIRA, Z.S. & MARKUS, R.P. (2001). Characterization of P2Y(1)-like receptors in
cultured rat pineal glands. Eur J Pharmacol. 415: 51-156.

96 IX. Referências Bibliográficas
FERREIRA, Z.S. et al. (1994). Presence of P2-purinoceptors in the rat pineal gland. Br J
Pharmacol. 112: 107-110. FERREIRA, Z.S. et al. (2005). Corticosterone modulates noradrenaline-induced
melatonin synthesis through inhibition of nuclear factor kappa B. J Pineal Res. 38: 182-188.
FERREIRA, Z.S. et al. (2003). P2Y(1) receptor activation enhances the rate of rat
pinealocytes-induced extracellular acidification via calcium-dependent mechanism. Pharmacology. 69: 33-37.
FOSTER, C.E. et al. (2000) Structures of mammalian cytosolic quinone reductases. Free
Radic Biol Med 29: 241–245. GANGULY, S. et al. (2001). Role of a pineal cAMP-operated arylalkylamine N-
acetiltransferase/14-3-3-binding switch in melatonin synthesis. Proc Natl Acad Sci USA. 98: 8083-8088.
GANGULY, S. et al. (2002). Control of melatonin synthesis in the mammalian pineal
gland: the critical role of serotonin acetylation. Cell Tissue Res. 309: 127-137. GARCIA, C.R.S. et al. (2001). Tertian and Quartan Fevers: Temporal Regulation in
Malarial Infection. J Biol Rhy. 16: 436-443. GARCIA-MAURINO, S. et al. (1997). Melatonin enhances IL-2, IL-6, and IFN-gamma
production by human circulating CD4þ cells: A possible nuclear receptor-mediated mechanism involving T helper type 1 lymphocytes and monocytes. J Immunol 159: 574-581.
GARIDOU, M.L. et al. (2001).In vivo observation of a non-noradrenergic regulation of
arylalkylamine N-acetyltransferase gene expression in the rat pineal complex.
Neurosci 105(3): 721-9.
GASTEL, J.A. et al. (1998). Melatonin production: proteasomal proteolysis in serotonin N-acetyltransferase regulation. Science. 279: 1358-1360.
GHOSH, S. & KARIN. (2002). Missing pieces in the NF-kappaB puzzle. Cell 109: 81–
96.
GHOSH, S. & HAYDEN MS. (2008). New regulators of NF-kappaB in inflammation. Nat Rev Immunol. 8: 837-848.
GILAD, E. et al. (1998). Melatonin inhibits expression of the inducible isoform of nitric
oxide synthase in murine macrophages: role of inhibition of NFkappaB activation.
FASEB J. 12: 685–693.

97 IX. Referências Bibliográficas
GOLDMAN, H. & WURTMAN, R.J., (1964). Flow of blood to the pineal body of the rat, Nature 203: 27–88.
GREEL, M. et al. The type 1 receptor (CD120a) is the high-affinity receptor for soluble
tumor necrosis factor. Proc. Natl. Acad. Sci. 95: 570–575, 1998. GROSS, P.M. & WEINDI, A. (1987) Peering through the windows of the brain. j Cereb.
Blood Flow Metabol. 7: 662-671 GUERRERO, J.M. & REITER, R.J. (2002). Melatonin–immune system relationships,
Curr. Top. Med. Chem. 2 :167–17.
HARADA, Y. et al. (1992). Neuropeptide Y inhibits beta-adrenergic agonist- and vasoactive intestinal peptide-induced cyclic AMP accumulation in rat pinealocytes through pertussis toxin-sensitive G protein J Neurochem. 59(6): 2178-83
HARDELAND, R. & FUHRBERG, B. (1996). Ubiquitous melatonin – presence and
effects in unicells, plants and animals. Trends Comp. Biochem. Physiol. 2: 25–45.
HARDELAND, R. et al. (2006). “Melatonin.” Inter J Biochem & Cell Biol 38(3): 313-6.
HARDELAND, R. (2005). Antioxidative protection by melatonin- Multiplicity of mechanisms from radical detoxification to radical avoidance. Endocr. 27: 119–130.
HAYDEN, M.S & GHOSH, S. (2008). Shared principles in NF-κB signaling. Cell 132:
344–362.
HAYDEN, M.S. & GHOSH, S. (2004). Signaling to NF-κB. Genes Dev. 18: 2195–2224 HEHLGANS, T. & MÄNNEL, D.N. (2002). The TNF – TNF Receptor System. Biol.
Chem. 383: 1581 - 1585.
HEIDBÜCHEL, U. & VOLLRATH, L. (1983). Pineal complex of rats: Effects of
superficial pinealectomy on the deep pineal. Acta Anat. 117: 165- 169.
HOFER, H. (1958). Zur morphologie der circumventriculären Organe des
Zwischenhirnes der Säugetiere. Verh. Dtsch. Zool. Ges. 202–251.
HOFFMANN, A. et al. (2002). The IκB-NF-κB signaling module: temporal control and
selective gene activation. Science. 298: 1241–45
HOFFMANN, A. & BALTIMORE, D. (2006). Circuitry of nuclear factor kappaB
signaling. Immunol Rev. 210: 171-186.
HOTCHKISS, A.K & NELSON, RJ. (2002). Melatonin and Immune Function: Hype or
Hypothesis? Crit Rev. in Immunol. 22(5&6): 351-371

98 IX. Referências Bibliográficas
HOTTA, C.T. et al. (2003). Melatonin and N-acetyl-serotonin cross the red blood cell membrane and evoke calcium mobilization in malarial parasites. Brazi J Med Biol Res 36(11): 1583-7.
HSU, H. et al. (1995). The TNF receptor 1- associated protein TRADD signals cell death
and NF-κB acti- vation. Cell 81: 495-504.
HSU, H et al. (1996). TNF- dependent recruitment of the protein kinase RIP to the TNF
receptor-1 signalling complex. Immunity. 4: 387–396.
HU, H. et al. (1999). Abnormal morphogenesis but intact IKK activation in mice lacking
the IKKα subunit of IκB kinase. Science 284: 316–20
JACOBS, R.A. et al. (1999). Pineal nitric oxide synthase, but not heme oxygenase, mRNA is suppressed by continuous exposure to light. Brain Res Mol Brain Res. 70(2): 264-72.
JANKOVIC, B.D. et al. (1994). Pineal gland and immune system. Immune functions in
the chick embryo pinealectomised at 86 h of incubation. Ann. N.Y. Acad. Sci. 719: 398–
409.
JIANG-SHIEH, Y.F. et al. (2003). Regional heterogeneity in immunoreactive
macrophages/microglia in the rat pineal gland. J Pineal Res. 35: 45-53.
JIANG-SHIEH, Y.F. et al. (2005). Responses of microglia in vitro to the gram-positive bacterial component, lipoteichoic acid. J Neurosci Res. 82(4): 515-24.
JIANG-SHIEH, Y.F. et al. (2005). Reactive changes of interstitial glia and pinealocytes
in the rat pineal gland challenged with cell wall components from gram-positive
and -negative bacteria. J Pineal Res 38: 17-26.
JOHNSON, A.K. & GROSS, P.M. (1993) Sensory circumventricular organs and brain
homeostatic pathways. FASEB J. 7: 678-86.
JOHNSTON, J.D. et al. (2004). Rhythmic melatonin secretion does not correlate with the expression of arylalkylamine N-acetyltransferase, inducible cyclic AMP early repressor, period 1 or cryptochrome1 mRNA in the sheep pineal. Neurosci. 124: 789-795.
JONES, SJ. et al. (1999). TNF recruits TRADD to the plasma membrane but not the
trans-Golgi network, the principal subcellular location of TNF-R1. J Immunol 162(2): 1042–1048.
KALSBEEK, A. et al. (1999). GABA release from suprachiasmatic nucleus terminals is
necessary for the light-induced inhibition of nocturnal melatonin release in the rat.
Neurosci. 91(2): 453-61.

99 IX. Referências Bibliográficas
KALSBEEK, A. et al. (2000). Melatonin sees the light: blocking GABA-ergic
transmission in the paraventricular nucleus induces daytime secretion of
melatonin. Eur J Neurosci. 12(9): 3146-54.
KALTHOFF, H. et al. 1993. Tumor necrosis factor (TNF) up-regulates the expression of
p75 but not p55 TNF receptors, and both receptors mediate, independently of each
other, up-regulation of transforming growth factor alpha and epidermal growth
factor receptor mRNA. J Biol Chem 268(4):2762
KAPPERS, J.A. (1960). The development, topographical relations and innervation of the
epiphysis cerebri in the albino rat. Z. Zellforsch. 52: 163–215.
KARIN, M. & BEN-NERIAH, Y. (2000). Phosphorylation meets ubiquitination: the control of NF-κB activity. Annu. Rev. Immunol. 18: 621–63.
KATILA, H. et al. (1993). Is there a seasonal variation in the interferon producing
capacity of healthy subjects? J Interferon Res. 13: 233–234. KELLIHER, MA. et al. (1998). The death domain kinase RIP mediates the TNF-induced
NF-κB signal. Immunity 8: 297 – 303.
KENNAWAY, D.J & WRIGHT, H. (2002). Melatonin and circadian rhythms. Curr Topi Medic Chem. 2(2): 199-209.
KITAY, J.I. & ALTSCHULE, M.D. (1954). Effects of pineal extract administration on ovary weight in rats. Endocrinol. 65: 782-784.
KLEIN, D.C. & WELLER, J.L. (1970). Indole metabolism in the pineal gland: a circadian
rhythm in N-acetyltransferase. Science 169: 1093-1095. KLEIN, D.C. et al. (1983). Postsynaptic alpha-adrenergic receptors potentiate the beta-
adrenergic stimulation of pineal serotonin N-acetyltransferase. Proc Natl Acad Sci U S A. 80: 599-603.
KNIGHT, D. et al. (1993). Construction and initial characterization of a mouse-human
chimeric anti-TNF antibody. Mol Immunol. 30: 1443−1453. KORF, H.W. & WAGNER, U. (1980). Evidence for a nervous connection between the
brain and the pineal organ in the guinea pig. Cell Tissue Res 209: 505–510.
KORF, H.W. et al. (1998). The pineal organ, its hormone melatonin, and the
photoneuroendocrine system. Adv. Anat. Embryol. Cell Biol. 146: 1–100.
KORF, H.W. et al. (1985a). Opsin-like immunoreaction in the retinae and pineal organs of four mammalian species. Cell Tissue Res 242: 645–648.

100 IX. Referências Bibliográficas
KORF, HW. et al. (1985b) Immunocytochemical demonstration of retinal S-antigen in the pineal organ of four mammalian species. Cell Tissue Res. 239: 81–85.
KORF, H.W. et al. (1992). Recoverin in pineal organs and retinae of various vertebrate
species including man. Brain Res 595: 57–66. KRIEGLER, M. et al. (1988). A novel TNF/cachectin is a cell surface cytotoxic
transmembrane protein: ramifications for the complex physiology of TNF. Cell. 53:
45-53.
LAFLAMME, N. & RIVEST, S. (2001). Toll-like receptor 4: the missing link of the
cerebral innate immune response triggered by circulating gram- negative bacterial
cell wall components. FASEB J. 15: 155–63.
LAHIRI, D.K. et al. (2004). Age-related changes in serum melatonin in mice: higher
levels of combined melatonin and 6-hydroxymelatonin sulfate in the cerebral
cortex than serum, heart, liver and kidney tissues. J Pineal Res. 36: 217–223
LARSEN, P.J. et al., (1991). Efferent projections from the periventricular and medial
parvocellular subnuclei of the hypo- thalamic paraventricular nucleus to
circumventricular organs of the rat. A Phaseolus vulgaris leucoagglutinin (PHA-L)
tracing study. J Comp Neurol. 306: 462–479.
LAWRENCE, T. (2009). The nuclear factor NF-kappaB pathway in inflammation.Cold Spring Harb Perspect Biol. (6): a001651.
LEANDER, P. et al. (1998). Neuronal projections from the median and dorsal raphe
nucleus to the suprachiasmatic nucleus and the deep pineal gland of the Golden
Hamster (Mesocricetus auratus). A combined in vivo tracing and
immunohistochemical neurotransmitter study. J Comp Neurol. 399: 73–93.
LEDGERWOOD, A. (1999). Recent advances in the molecular basis of TNF signal
transduction. Lab Invest 79(9): 1041–1050.
LEE, T.H. et al. (2003). The death domain kinase RIP1 is essential for tumor necrosis factor alpha signaling to p38 mitogen-activated protein kinase. Mol Cell Bi. 23: 8377-8385.
LERNER, A.B. et al. (1959). Structure of melatonin. J Am Chem Soc. 81: 6084-6085. LERNER, A.B. et al. (1958). Isolation of melatonin, the pineal gland factor that lightens
melanocytes. J Am Chem Soc. 80: 2587. LI, Q. et al. (2002). NF-kB regulation in the immune system. Nat. Rev. Immunol. 2: 725–
734.

101 IX. Referências Bibliográficas
LIOU, H.C. & HSIA, C.Y. (2003). Distinctions between c-Rel and other NF-κB
proteins in immunity and disease. Bioessays. 25: 767-780.
LIPNIACK, T. et al. (2004). Mathematical model of NF-kappaB regulatory module.” J Theor Biol. 228(2): 195-215.
LISSONI, P. (1999). The pineal gland as a central regulator of cytokine network.Neuro
Endocrinol Lett. 20(6): 343-349.
LIU, F. et al. (2001). Pineal indoles stimulate the gene expression of immunomodulating cytokines. J. Neural. Transm. 108: 397-405.
LOPES, C. et al. (1997). Circadian rhythm in experimental granulomatous inflammation
is modulated by melatonin. J Pineal Res. 23: 72-78. LOPES, C. et al. (2001). Interaction between the adrenal and the pineal gland in chronic
experimental inflammation induced by BCG in mice. Inflamm Res. 50: 6-11. LÓPEZ-MUÑOZ, F. et al. (1992). Immunohistochemical localization of glial fibrillary
acidic protein (GFAP) in rat pineal stalk astrocytes. Histol. Histopathol. 7: 643-6. LOTUFO, C.M.C. et al. (2001). Melatonin and N-acetylserotonin inhibit leucocyte
rolling and adhesion torat microcirculation. Eur J Pharmacol. 430: 351-357. LOTUFO, C.M.C. et al. (2006). Melatonin effect on endothelial cells reduces vascular
permeability increaseinduced by leukotriene B4. Eur J Pharmacol. 534: 258-263.
LUCHETTI, F. et al. (2010). Melatonin signaling and cell protection function. FASEB J 3603-3624.
LUO, Z.R. et al. (1984). Ultrastructural characterization of glial cells in the rat pineal gland with special reference to the pineal stalk. Anat Rec. 210: 663-674.
MAES, M. et al. (1994). Seasonal variation in peripheral blood leukocyte subsets and in
serum interleukin-6, and soluble interleukin 2 and 6 receptor concentrations in normal volunteers. Experientia 50: 821-829.
MAESTRONI, G.J.M. et al. (1998). The photoperiod transducer melatonin and the immune-hematopoietic system.J Photochem Photobio. B, 43(3): 186-92.
MARKUS, R.P. et al. (2007). The immune-pineal axis: a shuttle between endocrine and paracrine melatonin sources. Neuroimmunomodulation. 14: 126-133.
MARKUS, R.P. et al. (2010). Is modulation of nicotinic acetylcholine receptors by
melatonin relevant for therapy with cholinergic drugs? Pharmacol Ther. 126: 251-262.

102 IX. Referências Bibliográficas
MARONDE, E. et al. (1995). The effect of NO-donors in bovine and rat pineal cells: stimulation of cGMP and cGMP-independent inhibition of melatonin synthesis. J Neuroendocrinol. 7(3): 207-14.
MCCOY, M.K. & TANCEY, M.G. (2008). TNF signaling inhibition in the CNS:
implications for normal brain function and neurodegenerative disease. J Neuroinfl. 5:
45.
MERCURIO, F. & MANNING, A.M. (1999). Multiple signals con- verging on NF-κB.
Curr. Opin. Cell. Biol. 11: 226 – 232.
MIKKELSEN, J.D & MØLLER, M. (1990). A direct neural projection from the
intergeniculate leaflet of the lateral geniculate nucleus to the deep pineal gland of
the rat, demonstrated with Phaseolus vulgaris leucoagglutinin. Brain Res. 520: 342–
346.
MØLLER, M. (1992). Fine structure of the pinealopetal innervation of the mammalian pineal gland. Microsc Res Tech. 21: 188-204.
MØLLER, M. (1997).Peptidergic cells in the mammalian pineal gland. Morphological
indications for a paracrine regulation of the pinealocyte. Biol Cell 89:561–567.
MØLLER, M. (1999). Introduction to mammalian pineal innervation. Microsc Res Tech
46: 235–238.
MØLLER, M. & BAERES, F.M. (2002). The anatomy and innervation of the
mammalian pineal gland. Cell Tissue Res. 309: 139-150.
MØLLER, M. & HAY-SCHMIDT, A. (1998). Direct neuronal projection from the dorsal
raphe nucleus to the pineal complex of the rat: a Phaseolus vulgaris-
leucoagglutinin in vivo neuronal tracing study. J Pineal Res. 25: 19–23.
MØLLER, M. et al. (1978). Immunohistochemical demonstration of S100 protein and GFA protein in interstitial cells of rat pineal gland. Brain. Res. 140: 1-13.
MØLLER, M. et al. (1990). Evidence for a direct neuronal projection from the
hypothalamic paraventricular nucleus to the pineal complex of the rat: an anterograde study by use of Phaseolus vulgaris leucoagglutinin (PHA- L). Adv Pineal Res. 4: 1-8.
MØLLER, M. et al. (1996). The chemical neuroanatomy of the mammalian pineal gland:
neuropeptides. Neurochem Int. 28: 23-33. MOORE, R.Y. (1997). Circadian rhythms: basic neurobiology and clinical applications.
Annu Rev Med. 48: 253– 266. MORTANI-BARBOSA, E.J. et al. (2000). Purinergic and noradrenergic cotransmission
in the rat pineal gland. Eur J Pharmacol. 401: 59-62.

103 IX. Referências Bibliográficas
MOSS, M.L. et al. (1997). Cloning of a disintegrin metalloproteinase that processes
precursor tumour-necrosis factor-α. Nature 385: 733 – 736. MOSZKOWSKA, O. (1951). Effect of epiphyseal extracts on young adult female guinea
pigs. Seances Soc Biol Fil. 145(11-12): 845-7.
MUCHA, E. et al. (1994). Effect of interleukin-1 on in vivo melatonin secretion by the
pineal gland in rats. In: Advances in Pineal Research. G.J.M. Maestroni, A. Conti,
and R.J. Reiter, eds. John Libbey&Co. London, pp. 177–181.
MUNDIGLER, A. et al. (2002). Impaired circadian rhythm of melatonin secretion in
sedated critically ill patients with severe sepsis. Crit Care Med. 30: 536-540.
NELSON, R.J & DEMAS, G.E. (1996).Seasonal changes in immune function. Q Rev Biol 71: 511– 548.
NELSON, R.J., DEMAS, G.E.; KLEIN, S.L. (1998). Photoperiod mediation of seasonal
breeding and immune function in rodents: A multifactorial approach. Amer. Zool. 38: 226-237.
NELSON, R.J & DRAZEN, D.L. (1999). Melatonin mediates seasonal adjustments in
immune function. Reprod Nutr Dev. 39(3): 383-98.
NELSON, R.J. & DRAZEN, D.L. (2000) Melatonin mediates seasonal changes in immune function. Ann N Y Acad Sci .917:404-415.
NELSON, R.J, et al. (2002). Seasonal Patterns of Stress, Immune Function, and Disease.
Cambridge: Cambridge University Press, NEUMANN, M. & NAUMANN, M. (2007). Beyond IkappaBs: alternative regulation of
NF-kappaB activity. FASEB J. 21(11): 2642-54. NOSJEAN, O. et al. (2000). Identification of the melatonin binding site MT3 as the
quinone reductase 2. J Biol Chem. 275: 31311–31317. NURNBERGER, W. et al. (1995). Definition of a new score for severity of generalized
Neisseria meningitidis infection. Eur J Pediatr. 154(11): 896–900. OKSCHE, A. (1965) Survey of the development and comparative morphology of the
pineal organ. Prog. Brain Res. 10: 3-29. OLCESE, J. (1991). Neuropeptide Y: an endogenous inhibitor of norepinephrine-
stimulated melatonin secretion in the rat pineal gland. J Neurochem. 57(3): 943-7.
PERKINS, N.D. (2006). Post-translational modifications regulating the activity and
function of the nuclear factor κB pathway. Oncogene. 25: 6717–30.

104 IX. Referências Bibliográficas
PERREAU-LENZ, S. et al. (2003). Suprachiasmatic control of melatonin synthesis in
rats: inhibitory and stimulatory mechanisms. Euro Jour of Neurosci. 17: 221-228.
PIOLI, C. et al. (1993). Melatonin increases antigen presentation and amplifies specific
and non-specific signals for T-cell proliferation. Int. J. Immunopharmac. 15: 463–468.
POEGGELER, B. et al. (2002). Melatonin’s unique radical scavenging properties – roles of its functional substituents as revealed by a comparison with its structural analogs. J. Pineal Res. 33: 20-30.
PONTES, G.N. et al. (2006). Injury switches melatonin production source from
endocrine (pineal) to paracrine (phagocytes) - melatonin in human colostrum and
colostrum phagocytes. J Pineal Res. 41: 136-141.
PONTES, G.N. et al. (2007). Pineal melatonin and the innate immune response: the
TNF-alpha increase after cesarean section suppresses nocturnal melatonin
production. J Pineal Res. 43: 365-371.
PRENDERGAST, B.J. et al. (2004). Photo-period controls the induction, retention, and
retrieval of antigen-specific immunological memory. Am J Physiol Regul Integr Comp
Physiol. 286: 54–60.
PROVENCIO, I. et al. (2000). A novel human opsin in the inner retina. J Neurosci. 20:
600-605.
QUAY, W.B. (1963). Circadian rhythm in rat pineal serotonin and its modifications by estrous cycle and photoperiod. Gen Comp Endocrinol. 14: 473-479.
QUAY, W.B. (1964). Circadian and estrous rhythms in pineal melatonin and 5-hydroxy
indole-3-acetic acid. Proc Soc Exp Biol Med. 115: 710-713. REITER, R.J. (1973). Comparative physiology: pineal gland. Annu Rev Physiol. 35: 305-
328.
REITER, R.J. (1981). The mammalian pineal gland: structure and function, Am. J. Anat.
162: 287–313.
REITER, R.J. (1991). Pineal melatonin: cell biology of its synthesis and of its
physiological interactions. Endocr Rev. 12: 151-180.
REITER, R.J. et al. (2000). Melatonin and its relation to the immune system and inflammation. Ann. N.Y. Acad. Sci. 917: 376-386.

105 IX. Referências Bibliográficas
REITER, R.J et al. (2007). Melatonin defeats neurally-derived free radicals and reduces
the associated neuromorphological and neurobehavioral damage. J Physiol Pharmacol.
58: 5-22.
REITER, R.J et al. (2008). Melatonin in relation to the "strong" and "weak" versions of
the free radical theory of aging. Adv Med Sci. 53: 119-129.
REITER, R.J. et al. (2009). Light-mediated perturbations of circadian timing and cancer
risk: a mechanistic analysis. Integr Cancer Ther. 8: 354-360.
REITER, R.J. et al. (2009). Melatonin and reproduction revisited. Biol Reprod. 81: 445-
456.
REPPERT, S.M. et al (1994) Cloning and characterization of a mammalian melatonin
recep- tor that mediates reproductive and circadian responses. Neuron. 13: 1177–1185.
REPPERT, S.M. et al. (1995) Molecular characterization of a second melatonin receptor
expressed in human retina and brain: the Mel1b melatonin receptor. Proc Natl Acad
Sci USA. 92: 8734– 8738.
REPPERT, S.M. & WEAVER, D.R. (2002). Coordination of circadian timing in
mammals. Nat. 418: 935-941.
RIBELAYGA, C et al. (1997). Adrenergic and peptidergic regulations of hydroxyindole-
O-methyltransferase activity in rat pineal gland. Brain Res. 777: 247-250.
RIBELAYGA, C. et al. (1998). Ontogenesis of hydroxyindole-O-methyltransferase gene
expression and activity in the rat pineal gland. Brain Res Dev Brain Res. 110:235-239.
RIBELAYGA, C. et al. (1999) Photoperiodic control of the rat pineal arylalkylamine-N-
acetyltransferase and hydroxyindole-O-methyltransferase gene expression and its
consequence on melatonin synthesis. J Biol Rhyth. 14: 105–115.
RIBELAYGA, C. et al. (2000). HIOMT drives the photoperiodic changes in the
amplitude of the melatonin peak of the Siberian hamster. Am J Physiol Regul Integr
Comp Physiol. 278: 1339-1345.
ROBAK, T., GLADALSKA, A., STEPIEN, H. (1998). The tumour necrosis factor family
of receptors/ligands in the serum of patients with rheumatoid arthritis. Eur Cytokine
Netw: 9(2): 145–154.

106 IX. Referências Bibliográficas
ROSEBOOM, P.H. et al. (1996). Melatonin synthesis: analysis of the more than 150-fold
nocturnal increase in serotonin N-acetyltransferase messenger ribonucleic acid in
the rat pineal gland. Endocrinol. 137: 3033-3045.
ROTHE, M. et al. (1995). The TNFR2-TRAF signaling complex contains two novel
proteins related to baculoviral inhibitor of apoptosis proteins. Cell 83: 1243-1252.
ROTHE, M et al. (1994). A novel family of putative signal transducers associated with
the cytoplasmic domain of the 75 kDa tumor necrosis factor receptor. Cell 78: 681-
692.
ROTHWARF, D.M. & KARIN, M. (1999). The NF-kB activation pathway: a paradigm
in information transfer from membrane to nucleus. Sci. STKE. 1999(5)
SCHOMERUS, C. et al. (2000). Selective adrenergic/cyclic AMP-dependent switch-off of proteasomal proteolysis alone switches on neural signal transduction: an example from the pineal gland. J Neurochem. 75: 2123-2132.
SCHON, F. et al. (1985) Neuropeptide Y innervation of the rodent pineal gland and
cerebral blood vessel. Neurosci Letters. 57: 65-71. SCHRÖDER, H & MALHOTRA, S.K. (1987).Characterization of rodent pineal
astrocytes by immunofluorescence microscopy using a monoclonal antibody (J1-
31). Cell Tissue Res. 248(3): 607-10
SCHULTZE-OSTHOFF, K. et al. (1998). Apoptosis signalling by death receptors. Eur. J.
Biochem. 25: 439-459.
SEN, R. & BALTIMORE, D. (1986). Inducibility of κ immunoglobulin enhancer-
binding protein Nf-κB by a posttranslational mechanism. Cell 47: 921–928.
SHELDON, B.C. & VERHULST, S. (1996). Ecological immunology: costly parasite defenses and trade-offs in evolutionary ecology. Tree 11: 317–321.
SIMONNEAUX, V. & RIBELAYGA, C. (2003). Generation of the melatonin endocrine
message in mammals: a review of the complex regulation of melatonin synthesis by
norepinephrine, peptides, and other pineal transmitters. Pharmacol Rev. 55: 325-395.
SIMONNEAUX, V. et al. (1994). Presynaptic and postsynaptic effects of neuropeptide Y
in the rat pineal gland. J Neurochem.62: 2464-71.
SIMONNET, V. et al. (1951). Effect of pineal gland on the ovary of the young rat. Ann
Endocrinol. 12(2): 202-5.
SKWARLO-SONTA, K et al. (2003). Bidirectional communication between the pineal gland and the immune system. Can J Physiol Pharmacol. 81: 342-349

107 IX. Referências Bibliográficas
SLOMINSKI, A. et al. (2002). Serotoninergic and melatoninergic systems are fully
expressed in human skin. FASEB J. 16: 896-898.
SLOMINSKI, A. et al. (1996). Metabolism of serotonin to N-acetylserotoninmelatonin,
and 5-methoxytryptamine in hamster skin culture. J Biol Chem. 271: 12281-12286.
SLOWIK, M.R. et al. (1993). Tumor necrosis factor activates human endothelial cells
through the p55 tumor necrosis factor receptor but the p75 receptor contributes to
activation at low tumor necrosis factor concentration. AmJ Pathol 143(6): 1724–
1730.
SRINIVASAN, J. et al. (2008). Immunomodulation by melatonin: its significance for seasonally occurring diseases. Neuroimmuno 15(2): 93-101.
STEHLE, J.H. et al. (2001). Analysis of cell signaling in the rodent pineal gland deciphers
regulators of dynamic transcription in neural/endocrine cells. Eur J Neurosci. 14: 1-9.
STUDNICKA, F.K. (1905). Parietalorgane. In Lehrbuch der vergleich- enden
mikroskopischen anatomie der wirbeltiere, part 5 (ed. A. Oppel). Jena, Germany:
Gustav Fischer.
SUGDEN, D. & KLEIN, D.C (1983a). Beta-adrenergic receptor control of rat pineal
hydroxyindole-O-methyltransferase. Endocrinol 113: 348–353.
SUGDEN, D. & KLEIN, D.C (1983b). Adrenergic stimulation of rat pineal
hydroxyindole- O-methyltransferase. Brain Res 265: 348–351.
SUGDEN, D. & KLEIN, D.C. (1985). Development of the rat pineal alpha 1-
adrenoceptor.Brain Res. 325(1-2): 345-8.
SUGDEN, L.A. et al. (1985). Alpha-adrenergic potentiation of beta-adrenergic
stimulation of rat pineal N-acetyltransferase. Studies using cirazoline and fluorine
analogs of norepinephrine. Biochem Pharmacol. 33(24): 3947-50.
SUGDEN, L.A. et al. (1987). Alpha 1-adrenoceptor activation elevates cytosolic calcium
in rat pinealocytes by increasing net influx. J Biol Chem. 262: 741-745.
SUN, S.C. et al. (1993). NF-κB controls expression of inhibitor IκB α: evidence for an
inducible autoregulatory pathway. Science 259: 1912–15.
SUZUKI, T. & KACHI, T. (1995). Immunohistochemical studies on supporting cells in
the adrenal medulla and pineal gland of adult rat, especially on S-100 protein, glial
fibrillary acidic protein and vimentin. Kaibogaku Zasshi. 70: 130-9.

108 IX. Referências Bibliográficas
TAKEUCHI, M. et al. (1996). Anatomy of TRAF2. Distinct domains for nuclear factor-
κB activation and association with tumor necrosis factor signalling proteins. J Biol
Chem. 271: 19935–19942.
TAN, D.X. et al. (2010). The changing biological roles of melatonin during evolution:
from an antioxidant to signals of darkness, sexual selection and fitness. Biol Rev
Camb Philos Soc. 85(3): 607-23
TAN, D.X. et al. (2007). One molecule, many derivatives: a never-ending interaction of
melatonin with reactive oxygen and nitrogen species? J Pineal Res. 42: 28-42.
TARTAGLIA, L.A. et al. (1993a). A novel domain within the 55 kd TNF receptor signals
cell death. Cell 74: 845–853
TARTAGLIA, L.A. et al. (1993b). Ligand passing: the 75-kDa tumor necrosis factor
(TNF) receptor recruits TNF for signalling by the 55-kDa TNF receptor. J Biol
Chem. 268(25): 18542–18548
TECLEMARIAM-MESBAH, R. et al. (1999). Anatomical demonstration of the
suprachiasmatic nucleus-pineal pathway. J Comp Neurol. 406: 171-182.
THOMA, B. et al. (1990). Identification of a 60-kD tumor necrosis factor (TNF) receptor
as the major signal transducing component in TNF responses. The Journal of
experimental medicine. 172: 1019-23.
TRACEY, D. et al. (1988). Cachectin: a hormone that triggers acute shock and chronic cachexia. J Infect Dis. 157(3): 413-20.
TRACEY, D. et al. (2008). Tumor necrosis factor antagonist mechanisms of action: a
comprehensive review. Pharmacol Ther. 117: 244-279.
TSAI, S.Y. & MCNULTY, J.A. (1997). Microglia in the pineal gland of the neonatal rat:
characterization and effects on pinealocyte neurite length and serotonin content.
Glia. 20: 243-253.
TSAI, S.Y. & MCNULTY, J.A. (1999). Interleukin-1beta expression in the pineal gland
of the rat. J Pineal Res. 27: 42-48.
TSAI, S.Y. et al. (2001b). Microglia play a role in mediating the effects of cytokines on
the structure and function of the rat pineal gland. Cell Tissue Res. 303:423-431.
TSAI, S.Y. et al. (2001a). TGF-beta1 and IL-6 expression in rat pineal gland is regulated
by norepinephrine and interleukin-1beta. Histol Histopathol. 16: 1135-1141.

109 IX. Referências Bibliográficas
VACAS, M.I. et al. (1987). In vitro effect of neuropeptide Y on melatonin and
norepinephrine release in rat pineal gland. Cell Mol Neurobiol. 7(3): 309-15.
VALLABHAPURAPU, S. & KARIN, M. (2009). Regulation and function of NF-kappaB
transcription factors in the immune system. Annu Rev Immunol. 27: 693-733.
VANECEK, J. et al. (1985). Atypical synergistic a1- and b-adrenergic regulation of
adenosine 30, 50-monophosphate and guanosine 30, 50-monophosphate in rat
pinea- locytes. Endocrinol. 116: 2167–2173.
VOLLRATH, L. (1981). The pineal organ. In: Handbuch der mikroskopischen Anatomie
des Menschen. Vol VI/7. Oksche A. and Vollrath L. (eds). Springer. Berlin,
Heidelberg, New York. pp 1-665
WAAGE, A., HALSTENSEN, A., ESPEVIK., T. (1987). Association between tumour
necrosis factor in serum and fatal outcome in patients with meningococcal disease.
Lancet. 1(8529): 355–357.
WAJANT, H. (2003). Tumor necrosis factor signaling. Cell Death Differ. 10: 45-65.
WALDHAUSER, F. et al. (1984). Bioavailability of oral melatonin in humans.
Neuroendocronol. 39: 307-313
WANG, C.Y. et al. (1998). NF-kappaB antiapoptosis: induction of TRAF1 and TRAF2
and c-IAP1 and c-IAP2 to suppress caspase-8 activation. Science. 281(5383): 1680-3.
WERNER, S.L. et al. (2008). Encoding NF-kappaB temporal control in response to TNF: distinct roles for the negative regulators IkappaBalpha and A20.Gen & Devt. 22(15): 2093-101.
WHITESIDE, S.T. & ISRAEL, A. (1997). IκBproteins: structure, function and
regulation. Semin. Cancer Biol. 8: 75–82
WIESENBERG, I. et al. (1995). Transcriptional activation of the nuclear receptor RZR
alpha by the pineal gland hor- mone melatonin and identification of CGP 52608 as
a synthetic ligand. Nucleic Acids Res, 23: 327–333.
WINSTON, B.W. et al. (1995). Tumor necrosis factor alpha rapidly activates the
mitogen-activated protein kinase (MAPK) cascade in a MAPK kinase kinase-
dependent, c-Raf-1-independent fashion in mouse macrophages. Proc Natl Acad Sci
USA 92: 1614-1618.
WINZEN, R. et al. (1992). Selective decrease in cell surface expression and mRNA level
of the 55-kDa tumor necrosis factor receptor during differentiation of HL-60 cells
into macrophage-like but not granulocyte-like cells. J Immunol. 148(11): 3454–3460

110 IX. Referências Bibliográficas
WITHYACHUMNARNKUL, B., et al. (1990a). Changes in indole metabolism in organ
cultured rat pineal glands induced by interferon-gamma. J. Pineal. Res. 8: 313–322.
WITHYACHUMNARNKUL, B. et al. (1990b). Interferon gamma modulates melatonin
production in rat pineal glands in organ culture. J. Interferon Res. 10: 403–411.
WOLFE, D.E. (1965). The epiphyseal cell: An eletron-microscopic study of its
intercellular relationships and intracelulluar morphology in the pineal body of the
albino rat. Prog. Brain Res. 10: 332-386.
WURTMAN, R.J et al. (1963). Photic And Neural Control Rhythm Gland Of In The Pineal Gland: Control By Light. Science. 157: 487-492
WURTMAN, R.J. et al. (1967). Photic and neural control of the 24-hour norepinephrine
rhythm in the rat pineal gland. J Pharmacol Exp Ther. 157: 487-492.
ZHANG, S. et al. (1991). Tyrosine hydroxylase- and neuropeptide Y-immunoreactive
nerve fibers in the pineal complex of untreated rats and rats following removal of
the superior cervical ganglia. Cell Tissue Res. 265(1): 63-71.
ZHANG, S. et al. (2004). Effect of melatonin on the generation of nitric oxide in murine macrophages. Eur. J. Pharmaco. 501: 25-30.
ZMIJEWSKI, M.A. et al. (2009). The melatonin-producing system is fully functional in
retinal pigment epithelium (ARPE-19). Mol Cell Endocrinol. 307: 211-216
ZRENNER, C. Theories of pineal function from classical antiquity to 1900: a history, in: R.J. Reiter (Ed.), Pineal Res Rev III, Alan R. Liss, New York, 1985, pp. 1–40.
ZYLIŃSKA, K. et al. (1995). Effect of granulocyte-macrophage colony stimulating factor
and granulocyte colony stimulating factor on melatonin secretion in rats in vivo and in vitro studies. J Neuroimmunol. 56: 187–190

111 X. Súmula Curricular
X. SÚMULA CURRICULAR
Nome: Cláudia Emanuele Carvalho de Sousa
Naturalidade: Abaeté-MG
Data de Nascimento: 16/12/1986
Email: [email protected]
FORMAÇÃO ACADÊMICA
Mestrado
Ciências- Fisiologia Geral
Instituição: Universidade de São Paulo (USP), Instituto de Biociências
Tema: Caracterização do Eixo imune-pineal: Mecanismo de ação da citocina pró-
inflamatória TNF no controle da função pineal
Laboratório de Cronofarmacologia
Período: 12/2008 - 03/2011
Graduação
Licenciatura em Ciências Biológicas
Instituição: Universidade Federal de Juiz de Fora (UFJF)
Juiz de Fora - MG
Período 03 /2005 - 12 /2008
2° grau
Colégio Cenescista Nossa Senhora de Fátima
Abaeté - MG
2002 – 2004

112 X. Súmula Curricular
1º grau
Colégio Cenecista Nossa Senhora de Fátima
1998 - 2001
Escola Frederico Zacarias
Abaeté - MG
1994 - 1997
MONITORIA
1. Disciplina: Zoologia dos Invertebrados I
Profª Sueli de Souza Lima
Departamento de Zoologia do Instituto de Ciências Biológicas
Universidade Federal de Juiz de Fora
Período: 2º sem./2006 (bolsista) e 1° sem./2007(s/bolsa)
2. Disciplina: Biofísica
Prof. Sérgio Luiz Schmidt
Departamento de Fisiologia do Instituto de Ciências Biológicas
Universidade Federal de Juiz de Fora
Período: 2º sem./2006 (s/bolsa)
3. Programa de Preparação Pedagógica - Fisiologia para o Ensino Médio (PAE)
Profª. Responsável: Regina Pekelmann Markus
Departamento de Fisiologia
Universidade de São Paulo
Período: 1º sem./2010

113 X. Súmula Curricular
PRODUÇÃO CIENTÍFICA
Trabalhos Apresentados em eventos
1. STUTZ-REIS, Sarah, STUTZ-REIS, Suzana, SOUSA, Cláudia Emanuele Carvalho
de, MARTINS-NETO, Rafael Gioia. A evolução da musicalidade dos grilos. XXIX
Semana da Biologia, Juiz de Fora, MG, 2006.
2. SOUSA, Cláudia Emanuele Carvalho de, STUTZ-REIS, Sarah, STUTZ-REIS,
Suzana MARTINS-NETO, Rafael Gioia. The colour patterns in fossils: a probable
unpalatable butterfly from São Paulo Oligocene. XXIV Encontro Anual de Etologia,
Brasília, DF, 2006.
3. SOUSA, Cláudia Emanuele Carvalho de, STUTZ-REIS, Sarah, STUTZ-REIS,
Suzana, MARTINS-NETO, Rafael Gioia. Insect/flora interactions in the brazilian
permian: possible coleopteran (Insecta) perforations in fossil stem from Itapetininga,
SP (Irati formation, Paraná basin). XXIV Encontro Anual de Etologia, Brasília, DF,
2006.
4. STUTZ-REIS, Suzana, STUTZ-REIS, Sarah, SOUSA, Cláudia Emanuele Carvalho
de, MARTINS NETO, Rafael Gioia. Paleotanatosis: a new and fertile field of animal
behavior research. XXIV Encontro Anual de Etologia, Brasília, DF, 2006.
5. STUTZ-REIS, Sarah, SOUSA, Cláudia Emanuele Carvalho de, MARTINS-NETO,
Rafael Gioia. “Death Behavior” (Thanatoethology) in fishes from the Santana Formation
(Lower Cretaceous, Northeast Brazil). XXIV Encontro Anual de Etologia, Brasília, DF,
2006.
6. MARTINS-NETO, Rafael Gioia; SOUSA, Cláudia Emanuele Carvalho de.
Paleobiomecânica de gafanhotos (Insecta, Orthoptera) da formação Santana como
indicativo de distintos nichos ecológicos. Congresso de Ecologia do Brasil, Caxambu,
MG, 2007.
7. SOUSA, Cláudia Emanuele Carvalho de, CRUZ-MACHADO, Sanseray da Silveira.
Ritmos biológicos: um enfoque sobre o sistema de temporização circadiana dos

114 X. Súmula Curricular
mamíferos. XXXI Semana da Biologia da UFJF e XIV Mostra de Produção Científica,
Juiz de Fora, MG, 2008.
8. SOUSA, Cláudia Emanuele Carvalho de, TEIXEIRA, Lívia Clemente Motta,
TOSTES, Fabiana, PETERS, Vera Maria. Influência do índice sexual da ninhada no
desenvolvimento pós-natal de crias de ratos Wistar. 12ª Semana Temática da Biologia,
IB-USP, São Paulo, SP, 2009.
9. SOUSA, Cláudia Emanuele Carvalho de, FERNANDES, Pedro Augusto Magno,
TAMURA, Eduardo Koji, PETRILLI, Camila Lopes, MARKUS, Regina Pekelmann.
“TNF induces iNOS expression by pineal cells through a mechanism dependent of
NFkB. 41º Congresso Brasileiro de Farmacologia e Terapêutica Experimental, Ribeirão
Preto, SP, 2009.
10. CARVALHO-SOUSA, Cláudia Emanuele, CRUZ-MACHADO, Sanseray da
Silveira, PINATO, Luciana, TAMURA, Eduardo Koji, FERNANDES, Pedro Augusto
Carlos Magno, MARKUS, Regina Pekelmann. Pineal gland expresses TNFR1 in a
constitutive and rhythmic manner. XV Meeting of the Brazilian Society for Cell Biology,
São Paulo, SP, 2010.
11. CRUZ-MACHADO, Sanseray da Silveira, PINATO, Luciana, CARVALHO-SOUSA,
Cláudia Emanuele, TAMURA, Eduardo Koji, MARKUS, Regina Pekelmann. Evidences
for glia-pinealocytes interaction in cultured rat pineal gland. XV Meeting of the
Brazilian Society for Cell Biology, São Paulo, SP, 2010.
12. CRUZ-MACHADO, Sanseray da Silveira, PINATO, Luciana, CARVALHO-
SOUSA, Cláudia Emanuele, TAMURA, Eduardo Koji, FERREIRA, Zulma Silva,
MARKUS, Regina Pekelmann. Signaling transduction pathway involved in LPS-
Induced suppression of melatonin production by rat pineal gland. 42º Congresso
Brasileiro de Farmacologia e Terapêutica Experimental, Ribeirão Preto, SP, 2010.
13. PETRILLI, Camila Lopes, CARVALHO-SOUSA, Cláudia Emanuele, MUXEL,
Sandra Márcia MARKUS, Regina Pekelmann, FERREIRA, Zulma Silva. PY1 receptors
stimulation on rat pineal glands effects on nuclear factor kappa B pathway (NFKB) and
inducible nitric oxide synthase (iNOS). 42º Congresso Brasileiro de Farmacologia e
Terapêutica Experimental, Ribeirão Preto, SP, 2010.

115 X. Súmula Curricular
14. CARVALHO-SOUSA, Cláudia Emanuele, CRUZ-MACHADO, Sanseray da
Silveira, PINATO, Luciana, TAMURA, Eduardo Koji, FERNANDES, Pedro Augusto
Carlos Magno, MARKUS, Regina Pekelmann. Pineal gland is instrumented to be an
integral player of innate immune response. 42º Congresso Brasileiro de Farmacologia e
Terapêutica Experimental, Ribeirão Preto, SP, 2010
.
Publicações
1. SOUSA, Cláudia Emanuele Carvalho de, CRUZ-MACHADO, Sanseray da Silveira,
TAMURA, Eduardo Koji. Os ritmos circadianos e a reprodução em mamíferos. Boletim
do Centro de Biologia da Reprodução. 27 (1/2): 15-20, 2008.
2. SOUSA, Cláudia Emanuele Carvalho de, TEIXEIRA, Lívia Clemente Motta,
TOSTES, Fabiana, PETERS, Vera Maria. Proporção sexual da ninhada e
desenvolvimento ponderal de crias de ratos Wistar: dados preliminares. Revista
Interdisciplinar de Estudos Experimentais - Animais e Humanos. 1 (2): 82-83, 2010.
3. CRUZ-MACHADO, Sanseray da Silveira, CARVALHO-SOUSA, Cláudia
Emanuele, TAMURA, Eduardo Koji, et al. TLR4 and CD14 receptors expressed in rat
pineal gland trigger NFKB pathway. Journal of pineal research. 49 (2): 183-92, 2010.
Submetido: Claudia E Carvalho-Sousa, Sanseray S Cruz-Machado, Eduardo K Tamura, Pedro
ACM Fernandes, Luciana Pinato, Sandra M Muxel, Érika Cecon, Regina P. Markus.
Molecular basis for defining pineal gland and pinealocytes as targets for tumor necrosis
factor (TNF). Frontiers in Cellular Endocrinology (Dezembro, 2010).
PREMIAÇÃO
1. 3º lugar na categoria Melhor Trabalho de Iniciação Científica
Evento: XXIV Encontro anual de Etologia, Brasília, 2006
Trabalho: Paleotanatosis: a new and fertile field of animal behavior research

116 X. Súmula Curricular
2. Menção Honrosa
Evento: Mostra Científica da Universidade Federal de Juiz de Fora
Projeto de Iniciação Científica: Paleoartropodofauna do Cretáceo Brasileiro
3. Menção Honrosa
Evento: 41º Congresso Brasileiro de Farmacologia e Terapêutica Experimental, Ribeirão
Preto, 2009.
Trabalho: TNF induces iNOS expression by pineal cells through a mechanism
dependent of NFkB
4. Menção Honrosa: Semifinalista na Categoria Jovem Pesquisador-Mestrado.
Evento: XV Meeting of the Brazilian Society for Cell Biology, São Paulo, 2010.
Trabalho: Pineal gland expresses TNFR1 in a constitutive and rhythmic manner
ORGANIZAÇÃO DE EVENTOS /ATIVIDADES COMPLEMENTARES
1. Comissão Organizadora da I Mostra de Paleobiodiversidade
Duração: 16 a 20 de outubro de 2006
Instituto de Ciências Biológicas
Universidade Federal de Juiz de Fora
4. Aula ministrada no VI Curso de Inverno: Tópicos Avançados de Fisiologia
Tema: Introdução à Cronobiologia
Instituto de Biociências
Julho/2009
5. Comissão Organizadora do VII Curso de Inverno: Tópicos em Fisiologia
Comparativa

117 X. Súmula Curricular
Duração: 5 a 23 de julho, 2010.
CURSOS
Cursos de curta duração
1. Curso de processamento de imagens com o programa Image J
XV Meeting of the Brazilian Society for Cell Biology
Período: 21-23 de julho
Carga Horária: 18h
2.Writing a Scientific Paper: Theory and Practice
Sociedade Brasileira de Farmacologia e terapêutica experimental
18 a 21 de outubro de 2010
Cursos de Longa Duração
2. IV Curso de Inverno- Tópicos em Fisiologia Comparativa
Departamento de Fisiologia do Instituto de Biociências
Universidade de São Paulo (USP)
Duração: 10 a 27/07/2007
Carga Horária: 80h
3. XVII Curso de Verão em Fisiologia e Farmacologia “Prof.Wilson Teixeira
Beraldo”
Programa de Pós-Graduação em Ciências Biológicas: Fisiologia e Farmacologia
Universidade Federal de Minas Gerais (UFMG)
Duração: 14 a 25/01/2008
Carga Horária: 80h

118 X. Súmula Curricular
4. Curso “Desenho Experimental e Determinação Amostral com Animais de
Laboratório”
Centro de Biologia da Reprodução
Universidade Federal de Juiz de Fora
Dra. Adela Rosenkranz- Universidade de Buenos Aires, Argentina
Carga Horária: 40h
ESTÁGIOS
1. Estágio Curricular I em Zoologia/ Iniciação Científica
Projeto: Paleoartropodofauna do Cretáceo Brasileiro
Orientador: Rafael Gioia Martins Neto
Programa de Pós - Graduação em Biologia e Comportamento Animal
Universidade Federal de Juiz de Fora
Período: 10/2005 a 10/2007
2. Bolsista do Programa de Treinamento Profissional em Biologia da Reprodução
Projeto: Treinamento Profissional em boas práticas aplicadas às atividades de Biotério
do Centro de Biologia da Reprodução.
Orientadora: Vera Maria Peters
Centro de Biologia da Reprodução
Universidade Federal de Juiz de Fora
Período: 04/2007 a 03/2008
3. Estágio Curricular II em Biologia da Reprodução
Tema: Fatores que interferem no desenvolvimento pós-Natal de crias de ratos Wistar
Orientadora: Vera Maria Peters
Centro de Biologia da Reprodução

119 X. Súmula Curricular
Universidade Federal de Juiz de Fora
Período: 03/2008 – 07/2008
4. Estágio de Férias em Cronofarmacologia
Projeto: Estudo do Eixo Imune-Pineal
Orientadora: Regina P. Markus
Laboratório de Cronofarmacologia
Departamento de Fisiologia - IB
Universidade de São Paulo-USP
Período: Fevereiro de 2008; Julho de 2008
5. Bolsista do Programa de Treinamento Profissional em Morfologia
Projeto: Treinamento Profissional em Práticas e Técnicas em Histologia
Orientador: Rogério Estevam Farias
Departamento de Morfologia
Universidade Federal de Juiz de Fora
Período: 05/2008 a 12/2008

120 XI. Anexos
XI. ANEXOS

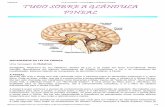

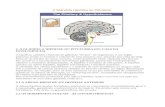









![Estudo morfológico da glândula pineal no cão€¦ · RESUMO Gomes, L. A. Estudo morfológico da glândula pineal no cão.[Morphological study of the pineal gland of the dog]. 2003.](https://static.fdocumentos.com/doc/165x107/605a7c483e82b651c15c1ac6/estudo-morfolgico-da-glndula-pineal-no-co-resumo-gomes-l-a-estudo-morfolgico.jpg)




