
Crescimento de plantas de arroz sob aplicação de...
60
Camila Pinho de Sousa Crescimento de plantas de arroz sob aplicação de herbicidas do grupo das imidazolinonas e sua atividade residual em plantas bioindicadoras Orientador: Dr. Marcos Antonio Bacarin Co-orientador: Dr. Jesus Juares Oliveira Pinto Pelotas, 2010 Dissertação apresentada ao Programa de Pós- Graduação em Fisiologia Vegetal da Universidade Federal de Pelotas, como requisito parcial a obtenção do título de Mestre em Ciências.
-
Upload
dinhnguyet -
Category
Documents
-
view
214 -
download
0
Transcript of Crescimento de plantas de arroz sob aplicação de...

1
Camila Pinho de Sousa
Crescimento de plantas de arroz sob aplicação de herbicidas do grupo das imidazolinonas e sua atividade
residual em plantas bioindicadoras
Orientador: Dr. Marcos Antonio Bacarin
Co-orientador: Dr. Jesus Juares Oliveira Pinto
Pelotas, 2010
Dissertação apresentada ao Programa de Pós-Graduação em Fisiologia Vegetal da Universidade Federal de Pelotas, como requisito parcial a obtenção do título de Mestre em Ciências.

Dados de catalogação na fonte: (Gladis Rejane Moran Ferreira – CRB – 10/1793)
S725c Sousa, Camila Pinho de Crescimento de plantas de arroz sob aplicação de herbicidas do grupo das imidazolinonas e sua atividade residual em plantas bioindicadoras/ Camila Pinho de Sousa. – Pelotas, 2010.
59f.
Dissertação (Mestrado) – Programa de Pós-Graduação em Fisiologia Vegetal. Instituto de Biologia. Universidade Federal de Pelotas – Pelotas, 2010, Marcos Antonio Bacarin, Orientador; co-orientador Jesus Juares Oliveira Pinto.
1. Inibidores ALS 2. Only® 3. Kifix® 4. Oryza sativa L. 5. Teste JIP I. Bacarin, Marcos Antonio (orientador) II. Título
CDU 632.08:633.18.03

2
Banca Examinadora: Prof. Marcos Antonio Bacarin
Prof. Nei Fernandes Lopes
Dr. Ariano Martins de Magalhães Júnior

3
Ao meu marido Diogo pelo seu amor incondicional que fez com que ele
me apoiasse, incentivasse e compreendesse a minha ausência durante este período.
A minha mãe Gladis pelo amor e educação dedicados durante toda a minha vida.
A minha família por sempre acreditar que eu poderia chegar muito longe e alcançar os meus objetivos.
Dedico e ofereço

4
AGRADECIMENTOS
A Deus, Senhor da minha vida, que guiou os meus passos e permitiu que este
sonho se tornasse realidade;
Ao professor Marcos Antonio Bacarin, pela orientação, aprendizado,
dedicação e amizade e, principalmente, por acreditar na minha capacidade;
Ao professor Jesus Juares Oliveira Pinto, pelos ensinamentos e pela amizade
dedicada durante todo o período da minha graduação e mestrado;
Aos amigos do Programa de Pós-graduação em Fisiologia Vegetal (PPGFV)
pela convivência e amizade, em especial a Marília Mércia, Anelise Perboni,
Emanuela Martinazzo, Janete Adamski e Sidnei Deuner;
Aos estagiários e bolsistas de iniciação científica Márcio Farias, Pablo
Valadão, Rômulo Silveira e Andrew Bortoli, pelo auxílio na condução dos
experimentos e pela amizade;
Aos professores e funcionários do PPGFV pelos ensinamentos e apoio
recebido;
A Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
pelo apoio financeiro durante o curso;
E por fim, a todos aqueles que de alguma maneira contribuíram para que este
trabalho fosse concluído.

5
“Confie no Senhor de todo o coração e não se apóie na sua própria inteligência. Lembre de Deus em tudo o que fizer, e ele lhe mostrará o
caminho certo” Provérbios 3:5-6

6
RESUMO SOUSA, Camila Pinho de. Crescimento de plantas de arroz sob aplicação de herbicidas do grupo das imidazolinonas e sua atividade residual em plantas bioindicadoras. 2010. 59f. Dissertação (Mestrado) – Programa de Pós-graduação em Fisiologia Vegetal. Universidade Federal de Pelotas, Pelotas. Este trabalho teve por objetivo: a) avaliar o efeito da aplicação de herbicidas do grupo químico das imidazolinonas sobre o crescimento de plantas e as injúrias causadas ao aparelho fotossintético de três tipos de arroz e b) avaliar o efeito residual no solo dos herbicidas imazethapyr + imazapic sobre as plantas de milho, pepino, rabanete e tomate, utilizadas como bioindicadoras. No primeiro ensaio foram utilizados os herbicidas Only® e Kifix®, utilizando-se para cada herbicida um delineamento experimental de parcelas subdivididas, em esquema fatorial 5x3, com seis repetições, sendo o fator A as doses do herbicida correspondentes a 0, 5, 10, 15 e 20 mg p. c. m-2 do produto comercial Only® e 0, 7, 14, 21 e 28 mg p. c. m-2 do produto comercial Kifix®, e o fator B o tipo de arroz (arroz cultivado cv. Puitá Inta CL, ecótipo de arroz-vermelho sensível aos herbicidas do grupo das imidazolinonas e ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do grupo das imidazolinonas). Foram avaliados os parâmetros da fluorescência da clorofila a, fitotoxicidade, altura de plantas e massa seca da parte aérea. A análise da fluorescência da clorofila a foi realizada utilizando-se um fluorômetro portátil (HandyPEA, Hanstech).Conclui-se que a cv. Puitá Inta CL é tolerante aos herbicidas Only® e Kifix®, podendo ser utilizada no Sistema de Produção Clearfield® e que o ecótipo de arroz-vermelho com suspeita de tolerância apresentou tolerância a ambos herbicidas, sofrendo baixa fitotoxicidade em comparação ao ecótipo sensível, mesmo utilizando-se doses superiores as comerciais dos produtos. A aplicação de herbicidas do grupo químico das imidazolinonas em plantas de arroz causa alterações no metabolismo fotossintético das plantas que podem ser detectadas através da avaliação da emissão da fluorescência da clorofila a transiente. O segundo experimento foi conduzido em um delineamento experimental casualizado por bloco, totalizando quatro repetições por tratamento. Os tratamentos avaliados foram o efeito residual do herbicida Only® sob as plantas de arroz CL na safra 2006/2007, nas doses de doses de 0, 10, 15 e 20 mg p. c. m-2 do produto comercial, sobre quatro espécies bioindicadoras: milho, pepino, rabanete e tomate. Foram avaliadas as variáveis: altura de plantas, massa seca da parte aérea e massa seca das raízes, sendo esta última não realizada para as plantas de milho. Os dados relativos às variáveis biométricas gerados em ambos os experimentos foram submetidos à análise da variância (p≤0,05); em sendo significativos, estes foram testados por modelos de regressão polinomial. Foi detectada atividade residual da mistura comercial dos herbicidas imazethapyr+imazapic em solo após 1100 dias da aplicação dos herbicidas, pelo uso de plantas bioindicadoras.
Palavras-chave: inibidores ALS, Only®, Kifix®, Oryza sativa L., teste JIP

7
ABSTRACT SOUSA, Camila Pinho de. Growth of rice plants under application of herbicides imidazolinone and its residual activity in bioindicators plants. 2010. 59f. Dissertação (Mestrado) – Programa de Pós-graduação em Fisiologia Vegetal. Universidade Federal de Pelotas, Pelotas. The objective of this research was: a) evaluate the effect of imidazolinone chemical group herbicides on the growth of plants and the injuries to the photosynthetic apparatus of three rice types (varieties) exposed to these and b) evaluate the effect of soil residual herbicides imazethapyr+ imazapic on corn, cucumber, radish and tomato are used as bioindicators. At first experiments we used the herbicide Only® and Kifix®, using a randomized split plot design in a factorial 5x3 with six replications, with the first factor rates corresponded to 0, 5, 10, 15 and 20 mg p. c. m-2 of commercial product Only® and 0, 7, 14, 21 and 28 mg p. c. m-2 of commercial product Kifix®, and factor B type of rice (cultivated rice cv. Puitá Inta CL, red rice ecotype sensitive to herbicides Imidazolinone and red rice ecotype with suspected herbicide tolerance imidazolinone). We evaluated the parameters of chlorophyll a fluorescence, phytotoxicity, plant height and dry mass. Analysis of chlorophyll a fluorescence was performed using a portable fluorometer (HandyPEA, Hanstech). It was concluded that cv. Puitá Inta CL is tolerant to herbicides Only® and Kifix® and can be used in the Clearfield® system and the red rice ecotype with suspected tolerance was tolerant to both herbicides, suffering a low phytotoxity compared to the sensitive ecotype, even using higher doses of the commercial products. The application of chemical herbicides of the imidazolinone group in rice plants cause changes in the photosynthetic metabolism of plants that can be detected by evaluating the emission of chlorophyll a fluorescence transient. The second experiment was conducted in a randomized block experimental design (with) four replicates per treatment. The treatments were the residual effect of herbicide Only® in rice CL plants in the 2006/2007 harvest, at 0, 10, 15 and 20 mg p. c. m-2 doses of commercial product, on four bioindicators: corn, cucumber, radish and tomato. The variables evaluated were: height, dry weight of shoot and root dry mass, with the latter doesn’t being performed for the corn plants. Data of biometric variables generated in both experiments were subjected to analysis of variance (p ≤ 0.05) were significant in these models were tested by polynomial regression. Activity was detected in the commercial mixture of residual herbicides imazethapyr +imazapic in soil 1100 days after herbicide application, by use of bioindicators plants. Keywords: ALS-inhibiting, Only®, Kifix®, Oryza sativa L., JIP-test

8
SUMÁRIO INTRODUÇÃO GERAL ............................................................................................ 09 CAPÍTULO 1 – Efeito de herbicidas do grupo das imidazolinonas sobre plantas de arroz cultivado e arroz-vermelho ...................................................................... 14
1. Introdução .................................................................................................... . 16
2. Material e Métodos ..................................................................................... ....17
3. Resultados e Discussão ............................................................................. ....19
4. Conclusões ................................................................................................. ....20
CAPÍTULO 2 – Fluorescência da clorofila a em plantas de arroz em resposta a aplicação de herbicidas do grupo das imidazolinonas ....................................... 26
1. Introdução .................................................................................................. ....27
2. Material e Métodos ..................................................................................... ....29
3. Resultados e Discussão ............................................................................. ....32
4. Conclusões ................................................................................................. ....41
CAPÍTULO 3 – Crescimento de espécies bioindicadoras do residual do herbicida Only®, semeadas em rotação com o arroz Clearfield® ........................ 42
1. Introdução .................................................................................................. ....43
2. Material e Métodos ..................................................................................... ....44
3. Resultados e Discussão ............................................................................. ....45
4. Conclusões ................................................................................................. ....49
CONSIDERAÇÕES FINAIS ...................................................................................... 50 REFERÊNCIAS ......................................................................................................... 52

9
INTRODUÇÃO GERAL O arroz (Oryza sativa L.) é um dos cereais mais cultivados no mundo, sendo
considerado alimento básico em muitos países, especialmente nos
subdesenvolvidos (MENEZES et al., 2002). O controle de plantas daninhas na
cultura do arroz irrigado é um ponto crítico para a obtenção boas produtividades. O
arroz-vermelho, em nível mundial, é considerado uma das principais plantas
daninhas do arroz cultivado. Nos Estados Unidos da América do Norte, várias
espécies são citadas como arroz-vermelho: Oryza sativa L. ssp indica, Oryza sativa ssp
japonica, Oryza rufipogon e Oryza nivarra (VAUGHAN et al., 2001), contudo, no Brasil há
referência somente à espécie Oryza sativa L. (NOLDIN et al., 2004, VILLA et al., 2006).
Os efeitos negativos da presença do arroz-vermelho em lavouras de arroz
incluem competição por espaço, recursos do meio, aumento do custo de produção,
acamamento de plantas, dificuldade de colheita e secagem dos grãos, depreciação
da qualidade do produto, hospedagem de pragas e moléstias, diminuição do valor
comercial das áreas infestadas e fonte de disseminação e reposição do banco de
sementes no solo (MENEZES et al., 2002).
O controle de arroz-vermelho por meio de herbicidas seletivos era
considerado, até pouco tempo, prática inviável pelo fato de o arroz cultivado e o
arroz-vermelho pertencerem à mesma espécie e apresentarem similaridades
morfogênicas e fisiológicas (MENEZES et al., 2009). Além disso, o controle de arroz-
vermelho por outros métodos possui eficiência limitada, sobretudo em grandes áreas
de cultivo (ROSO et al., 2010).
Pesquisadores no Centro de Agricultura da Universidade Estadual de
Louisiana geraram utilizando-se de uma mutação induzida com etil-
metanossulfonato (EMS) em sementes de arroz da linhagem AS3510 (CROUGHAN
et al., 1996) uma linhagem de arroz tolerante aos herbicidas do grupo químico das
imidazolinonas. No Brasil, a primeira cultivar de arroz resistente aos herbicidas

10 imidazolinonas, IRGA 422 CL, foi desenvolvida por retrocruzamento, utilizando-se a
linhagem 93AS3510 como fonte doadora do gene que confere tolerância ao
herbicida e a cultivar IRGA 417 como cultivar recorrente (LOPES et al., 2002).
Concomitantemente, diversas cultivares resistentes aos herbicidas imidazolinonas
foram desenvolvidas: SATOR CL, AVAXI CL, XP710 CL e SCS 115 CL, BRS
SINUELO CL, PUITA INTA CL (SOSBAI, 2010). O lançamento das cultivares
tolerantes aos herbicidas imidazolinonas permitiu que fosse desenvolvida uma
tecnologia de manejo de arroz-vermelho que associa estas cultivares a herbicidas
não seletivos ao arroz-vermelho, sendo denominada de Sistema de Produção
Clearfield®. O herbicida inicialmente recomendado para esse sistema foi o Only®,
composto pela mistura formulada dos herbicidas imazethapyr + imazapic, nas
concentrações de 75 e 25g i.a. L-1, respectivamente (SOSBAI, 2010). Atualmente, foi
lançado o herbicida Kifix®, que é composto pela mistura formulada dos herbicidas
imazapyr + imazapic, nas concentrações de 525 e 175g i.a. kg -1, respectivamente,
também podendo ser utilizado neste sistema.
No Brasil, o Sistema de Produção Clearfield® começou a ser utilizada pelos
produtores apenas na safra 2003/04. Atualmente, seis anos após a implantação, é
estimado que aproximadamente 500 mil ha da safra 2008/09 produzida no Rio
Grande do Sul, o que corresponde a aproximadamente 50% da área total cultivada,
utilizaram o Sistema de Produção Clearfield® (BASF, 2009).
Os herbicidas químicos apresentam propriedades eficazes que permitem a
sua penetração na planta daninha e translocação até o seu sítio de ação. A maioria
dos mesmos inibem enzimas e compete por sítios de ligação específicos que são
essenciais para o metabolismo das plantas. Estas moléculas são classificadas por
grupos químicos e de acordo com o mecanismo de ação. Atualmente existem 15
diferentes mecanismos de ação de herbicidas (HRAC, 2010), destacando-se os
inibidores da enzima acetolactato sintase (ALS), onde se encontram um grande
número de moléculas herbicidas utilizadas em diversas culturas, como o bispyribac-
sodium, chlorimuron, metsulfuron, imazapyr, imazapic, imazethapyr, nicosulfuron,
entre outros. Este mecanismo de ação é dividido em cinco classes de produtos
químicos: sulfoniluréias, imidazolinonas, triazolpirimidinas sulfonanilidas, pirimidil-oxi-
tiobenzoatos, e sulfonilamino-carboniltriazolinona.
Os herbicidas da família das imidazolinonas atuam nas plantas sensíveis,
inibindo a síntese da enzima ALS (EC 2.2.1.6), também conhecida como

11 acetohidroxiácido sintase (AHAS), que apresenta o nome sistemático de piruvato:
piruvato acetaldeído transferase (descarboxilativo) sendo responsável pela
biossíntese dos aminoácidos: isoleucina, leucina e valina. Estes herbicidas possuem
como principais características, elevada persistência no solo e amplo espectro de
controle de plantas daninhas em arroz híbrido ou cultivares derivadas de linhagens
mutadas (ALISTER; KOGAN, 2005).
Como resultado da inibição da síntese destes aminoácidos, várias outras
alterações ocorrem no metabolismo das plantas sensíveis, como bloqueio da síntese
de DNA, acúmulo de cetubutirato (participante do metabolismo do propanoato, da
síntese de alanina, valina, isoleucina e leucina, entre outras rotas), bloqueio da
síntese de acetil-CoA e inibição do transporte de fotossintetizados a partir das folhas
verdes (KRAEMER et al., 2009).
A sintomatologia de ação destes herbicidas inclui a paralisação no
crescimento, clorose, necrose dos meristemas apicais, definhamento e morte das
plantas sensíveis, o que ocorre entre sete e 14 dias após a aplicação dos produtos
(TAN et al., 2006), porém mudanças profundas no metabolismo das plantas ocorrem
logo após a aplicação dos herbicidas inibidores da ALS. Em espécies e biótipos
tolerantes aos herbicidas a atividade da ALS inicialmente inibida é recuperada pela
metabolização do herbicida, que pode ser por hidroxilação do anel aromático,
hidroxilação alifática, desalquilação, deesterificação, conjugação e pela ação da
enzima citocromo P450 mooxigenase (VIDAL; WINKLER, 2002)
Recentemente, foi descrito o aparecimento de ecótipos de arroz-vermelho
com resistência aos herbicidas do grupo químico das imidazolinonas. O surgimento
de plantas daninhas resistentes a herbicidas é influenciado por vários fatores, dentre
os quais pode ser destacado a utilização intensiva do mesmo herbicida ou de
herbicidas com o mesmo mecanismo de ação, aumentando a pressão de seleção do
biótipo resistente (MONQUERO; CHRISTOFFOLETI, 2001). A utilização das
cultivares resistentes aos herbicidas em uma mesma área, em conjunto com o baixo
uso de métodos preventivos de dispersão do arroz-vermelho, conduz ao surgimento
de populações de arroz-vermelho resistente aos herbicidas do grupo químico das
imidazolinonas. Em levantamento realizado em 228 lavouras do estado do RS que
possuem suspeita de plantas com resistência, foi constatado que 55,7% destas
apresentavam indivíduos resistentes aos herbicidas imazethapyr + imazapic
(MENEZES et al., 2009), indicando a grande freqüência deste problema mesmo com

12 poucos anos de utilização do Sistema de Produção Clearfield®. Além do surgimento
de plantas daninhas resistentes, outro fator de grande preocupação para os
agricultores é a persistência destes herbicidas no solo. A persistência é uma
característica de alguns grupos químicos de herbicidas, ou ainda de herbicidas
específicos, podendo ser desejável ou não, sendo positiva quando resulta em
período suficiente para manter a cultura livre da competição de plantas daninhas
(GAZZIERO et al., 1997), todavia é indesejável quando resulta em injúrias para
culturas em sucessão ou em aumento de riscos de contaminação ambiental.
A maioria das pesquisas destacam a longa persistência no solo da maioria
dos compostos do grupo químico das imidazolinonas (ALISTER; KOGAN, 2005). A
persistência dos herbicidas desse grupo químico no solo pode ser variável com o
produto (SENSEMANN, 2007), a dose (SILVA et al., 1999), a temperatura (JENKINS
et al., 2000), a matéria orgânica (KOSKINEN; HARPER, 2001), a umidade do solo
(EHLERS; LOIBNER, 2006) e outras condições ambientais, incluindo o pH do solo
(TRACY; PENNER, 2005).
O potencial dos herbicidas do grupo químico das imidazolinonas em
ocasionar fitotoxicidade às culturas semeadas em rotação que não sejam tolerantes
foi relatado (BALL et al.,2003). Estudos indicam que a presença de resíduos dos
herbicidas do grupo das imidazolinonas em solos causa injúrias em plantas de milho,
sorgo granífero, tomate e pimenta semeados em rotação com culturas que utilizaram
estes herbicidas (ALISTER; KOGAN, 2005; PINTO et al., 2009).
Devido a estes fatores, o estudo de novas técnicas que detectem alterações
no metabolismo das plantas após a aplicação de herbicidas ou devido à atividade
residual destes produtos no solo se faz necessário. Sabe-se que o aparelho
fotossintético das plantas é muito sensível ao estresse ambiental e mesmo
pequenas alterações na sua estrutura e funcionamento podem ser facilmente
detectadas (YUSUF et al., 2010). Quando a energia é absorvida pelo aparelho
fotossintético, esta pode ser dissipada na forma de fluorescência da clorofila
(KRAUSE; WEIS, 1984), sendo essa uma das formas de monitoramento da inibição
ou redução na transferência de elétrons entre os fotossistemas da planta. A redução
na dissipação da energia pelo processo fotoquímico é refletida por incremento
correspondente na fluorescência (MAXWELL; JOHNSON, 2000).
A análise da fluorescência da clorofila a é largamente utilizada para
avaliação da capacidade fotossintética alterada por estresses bióticos ou abióticos.

13 Uma das vantagens do uso deste método é o fato da medição não ser destrutiva,
sendo possível realizar tanto em laboratório quanto em campo. Vários estudos
avaliaram a relação da fluorescência das clorofilas com a fotossíntese e o estado
fisiológico das plantas (LICHTENTHALER, 1992; FRANKART et al., 2003; YUSUF et
al., 2010).
Existem dois procedimentos para estudar a fluorescência da clorofila a,
pulso modulado (que permite a quantificação dos coeficientes de extinção) e a
cinética da emissão da fluorescência transiente (ou curva OJIP). Nas avaliações com
pulso modulado são utilizados fluorômetros de luz modulada (SCHREIBER et al.,
1996), cujas aplicações variam desde a rápida identificação de injúrias causadas ao
aparelho fotossintético, até a análise detalhada da alteração da capacidade
fotossintética da planta. A avaliação da cinética de emissão da fluorescência
transiente ou polifásica envolve equipamento com maior resolução na detecção da
intensidade da fluorescência. Com estes equipamentos pode-se identificar que a
variação entre a fluorescência inicial e máxima não é linear, mas sim polifásica
(LÁZAR, 2006). Com a elucidação do comportamento polifásico do aumento da
fluorescência, Strasser e Strasser (1995) propuseram analisá-los por meio de uma
série de parâmetros matemáticos denominado de teste JIP (STRASSER;
STRASSER, 1995; STRASSER et al., 2000). O uso correto dos parâmetros
matemáticos propostos pelo teste JIP permite quantificar o fluxo de energia
absorvido e aproveitado no sistema fotossintético, podendo também avaliar o
desempenho fotossintético de plantas (TSMILLE; STRASSER, 2008).
Este trabalho teve por objetivo avaliar o efeito da aplicação de herbicidas do
grupo químico das imidazolinonas e as injúrias causadas ao aparelho fotossintético
de três tipos de arroz expostos a estes herbicidas, além de determinar o efeito
residual no solo dos herbicidas imazethapyr + imazapic, sobre as plantas de milho,
pepino, rabanete e tomate, utilizadas como bioindicadoras.

14
CAPÍTULO 1
EFEITO DE HERBICIDAS DO GRUPO DAS IMIDAZOLINONAS SOBRE PLANTAS DE ARROZ CULTIVADO E ARROZ-VERMELHO
Effect of Herbicides Imidazolinone on Plants Cultivates Rice and Red Rice
RESUMO - Este trabalho avaliou o efeito dos herbicidas Only® e Kifix®, pertencentes ao grupo químico das imidazolinonas, em plantas de arroz cultivado e arroz-vermelho sensível e com suspeita de tolerância a estes herbicidas. Foram conduzidos dois experimentos em casa-de-vegetação no município de Capão do Leão/RS, um utilizando o herbicida Only® e outro o herbicida Kifix®. Os experimentos foram conduzidos em parcelas representadas por caixas de madeira, cada parcela foi dividida em três sub-parcelas, contendo três linhas de arroz do tipo: a) arroz cultivado cv. Puitá Inta CL, b) ecótipo de arroz-vermelho sensível a herbicidas do grupo das imidazolinonas e, c) ecótipo de arroz-vermelho com suspeita de tolerância a herbicidas do grupo das imidazolinonas. Foram realizados dois experimentos independentes, um para cada herbicida testado (Only® (imazethapyr+imazapic) e Kifix® (imazapyr+imazapic)), em ambos os e ensaios foi utilizado o esquema fatorial 5x3, sendo o fator A doses do herbicida e o fator B os tipos de arroz, com seis repetições. Para o Only® foram utilizadas as doses de 0, 5, 10, 15 e 20mg p. c. m-2
do produto comercial e para o Kifix® as doses foram 0, 7, 14, 21 e 28 mg p. c. m-2 do produto comercial , ambos acrescidos de 0,5% v/v, do adjuvante Dash®. A aplicação ocorreu quando as plantas do arroz CL estavam nos estádio V3-V4. A adição de água para a inundação das plantas ocorreu aos sete dias após a aplicação dos herbicidas. Foram avaliadas as seguintes variáveis-resposta: fitotoxicidade (aos 15, 30 e 60 dias após a aplicação dos herbicidas), altura de plantas e massa seca da parte aérea (aos 60 dias após a aplicação dos herbicidas). Os dados foram submetidos à análise da variância (p≤0,05); seguido de modelos de regressão polinomial quando significativos. Ocorreu um aumento de toxidez com o incremento da dose herbicida para a cv. Puitá Inta CL, nos dois herbicidas utilizados, porém os resultados obtidos para a altura de plantas e massa seca da parte aérea comprovam que esta toxidez não foi prejudicial para o crescimento das plantas. Foi observado um efeito semelhante dos herbicidas Only e Kifix sobre o ecótipo de arroz-vermelho sensível, onde ambos demonstraram um controle insatisfatório deste ecótipo (em torno de 50%) quando estas plantas foram expostas a aplicação das doses comerciais dos herbicidas. O ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do grupo das imidazolinonas apresentou toxicidade

15 semelhante para ambos os herbicidas testados, sendo esta crescente durante as avaliações. Porém, os resultados mostram que na aplicação da dose comercial dos produtos esta toxicidade não ultrapassou 35%. Conclui-se que os herbicidas Only® e Kifix® são eficientes no controle do arroz-vermelho sensível e que a cv. Puitá Inta CL é tolerante a ambos os herbicidas. O ecótipo de arroz-vermelho com suspeita de tolerância apresentou tolerância a ambos herbicidas, mesmo utilizando-se doses superiores as comerciais dos produtos, sendo esta tolerância maior para o herbicida Kifix®, quando comparado ao herbicida Only®. Palavras-chave: Oryza sativa, Only®, Kifix®, controle. ABSTRACT - This research evaluated effect of herbicides Only® and Kifix® in on rice plants, sensitive red rice plants and red rice with suspected tolerance to these herbicides. Two experiments were conducted in a greenhouse, at Universidade Federal de Pelotas, on Capão do Leão-RS, southern Brazil, using herbicides Only® and Kifix®. The experiment was conducted in wooden box that represented plots in a split plot experimental design. Each plot was divided in three sub-plots, wich contained three lines of: a) cultivated rice cv. Puitá Inta CL, b) ecotype red rice sensitive to herbicides of the imidazolinone group, and c) ecotype of red rice with suspected herbicide tolerance of imidazolinone group. We used a 5x3 factorial scheme, with factor A was doses of herbicide and factor B was types of rice, with six replicates. Doses of 0, 5, 10, 15 e 20mg p. c. m-2 of the commercial product Only® (imazethapyr + imazapic) and 0, 7, 14, 21 and 28 mg p. c. m-2 of the commercial product Kifix® (imazapyr + imazapic), both increased by 0.5% v / v, the adjuvant Dash® were applied in stage V3-V4 CL rice. Seven days after application plants were maintained with constant irrigation. We measured the following variables: phytotoxicity (15, 30 and 60 days after herbicide application), plant height and aboveground dry biomass (60 days after herbicide application). Data were subjected to analysis of variance (p≤0.05), followed by polynomial regression models, when significant. There was an increase in toxicity with increasing herbicide rate for cv. Puitá Inta CL in the two herbicides, but the results for plant height and dry mass show that this toxicity was not detrimental (harmful) to plant growth. We observed a similar effect of herbicides Only and Kifix on the red rice ecotype sensitivity, and both showed an unsatisfactory control of this ecotype (around 50%) when these plants were exposed to doses of the commercial application of herbicides. The red rice ecotype with suspected herbicide tolerance imidazolinone showed similar toxicity for both herbicides, and this increased during the evaluations. However, the results show that the applications rate of commercial products this toxicity has not exceeded 35%. It is concluded that the herbicides Only® and Kifix® are effective in controlling of sensitive red rice and the cv. Puitá Inta CL is tolerant to both herbicides. The red rice ecotype with suspected tolerance was tolerant to both herbicides, even using doses higher than the commercial products, and is more tolerant to Kifix® herbicide when compared to the herbicide Only®. Keywords: Oryza sativa, Only®, Kifix®, control.

16 1. INTRODUÇÃO
No Brasil, a cultura do arroz (Oryza sativa L.) tem grande importância
econômica e social, porém diversos fatores limitam o incremento da sua
produtividade, com destaque para a alta incidência de arroz-vermelho na maioria
das regiões brasileiras (NOLDIN et al., 2004) e do mundo (BURGOS et al., 2008).
No Rio Grande do Sul, as perdas na produtividade decorrentes do arroz-vermelho
foram de um milhão de toneladas na última década, ou seja, aproximadamente 20%
da produção (IRGA, 2009).
O arroz-vermelho pertence à mesma espécie do arroz cultivado e, devido às
semelhanças morfofisiológicas entre ambos, seu controle é difícil (ROSO et al.,
2010). Sua competitividade e estabelecimento como planta daninha podem ser
explicados em função da alta capacidade de produção de matéria seca; da estatura
mais elevada em especial dos ecótipos predominantes nas lavouras; e do ciclo
geralmente mais longo que a maioria das cultivares utilizadas no Rio Grande do Sul
(SCHWANKE et al., 2008). Nas perdas devido à presença do arroz daninho, estão
incluídos: custos de produção, competição por recursos do meio, redução na
produtividade de grãos, depreciação do produto colhido, além de atuar como abrigo
para pragas e fontes de inóculos de patógenos.
Devido a isto, após anos de busca por uma solução deste problema foi
desenvolvimento o Sistema de Produção Clearfield® em arroz cultivado, que
proporciona uma estratégia de manejo eficaz no controle seletivo de plantas
daninhas, pelo uso de genótipos de arroz tolerante aos herbicidas imazethapyr e
imazapic (STEELE et al., 2002). No entanto, é de se esperar que o arroz cultivado
tolerante ao herbicida imazethapyr e imazapic possua também tolerância a outros
herbicidas do mesmo grupo químico, como imazaquin e imazapyr (WEBSTER;
MASSON, 2001). Estes herbicidas pertencem ao grupo químico das imidazolinonas
e possuem como mecanismo de ação a inibição da enzima acetolactato sintase
(ALS, EC 2.2.1.6), que participa da biossíntese dos aminoácidos valina, leucina e
isoleucina (RAJGURU et al.,2005). Os sintomas das plantas sob efeito dos
herbicidas inibidores da ALS incluem paralisação do crescimento, amarelecimento
dos meristemas e redução no crescimento das raízes (TAN et al.,2006), resultando
muitas vezes na morte dos vegetais (DEVINE; PRESTON, 2000).
No Brasil, o Sistema de Produção Clearfield® começou a ser utilizada pelos
produtores apenas na safra 2003/04. Estima-se que aproximadamente 500 mil ha da

17 safra 2008/09 produzida no Rio Grande do Sul utilizaram este sistema de produção
(BASF, 2009). Os resultados de pesquisa obtidos no RS demonstram eficácia
superior a 95% no controle de arroz-vermelho quando se utiliza o Sistema de
Produção Clearfield® (VILLA et al., 2006). O incremento de produtividade da lavoura
de arroz irrigado no Estado do Rio Grande do Sul nos últimos anos tem sido
atribuído ao uso deste sistema, o qual pode ser considerado como a mais importante
ferramenta disponibilizada aos produtores para controle químico do arroz-vermelho.
Apesar de proporcionar grande vantagem para a cultura do arroz, a adoção
de genótipos resistentes a herbicidas utilizados no Sistema de Produção Clearfield®,
requer o cuidado quanto à seleção de populações de arroz-vermelho resistentes
(MENEZES et al., 2009; ROSO et al., 2010). Dentre os problemas que podem
ocorrer no Sistema de Produção Clearfield® se encontram o fluxo de genes ou a
resistência natural em plantas de arroz-vermelho, prejudicando a eficiência do
mesmo (VILLA et al., 2006). Diversos fatores podem contribuir para o surgimento
dessa resistência como: alta frequência de uso de cultivares de arroz resistentes aos
herbicidas; elevada densidade de infestação dessa planta daninha; controle
deficiente; efeito residual dos herbicidas e uso de sementes não certificadas (VILLA
et al., 2006). Nessa ótica, fica evidente que o uso contínuo do Sistema de Produção
Clearfield® nas áreas de arroz irrigado no Rio Grande do Sul pode favorecer o
desenvolvimento de populações de arroz-vermelho resistentes aos herbicidas do
grupo das imidazolinonas, devido ao uso repetido de herbicidas de um mesmo grupo
ou pertencentes a diferentes grupos, mas com o mesmo mecanismo de ação, fato
este que dificultaria o controle desta planta daninha.
O objetivo deste trabalho foi avaliar o efeito dos herbicidas Only®
(imazethapyr+imazapic) e Kifix® (imazapyr+imazapic), pertencentes ao grupo
químico das imidazolinonas, em plantas de arroz cultivado e de arroz-vermelho
ecótipo sensível e ecótipo com suspeita de tolerância a estes herbicidas.
2. MATERIAL E MÉTODOS O experimento foi conduzido em casa de vegetação da Faculdade de
Agronomia Eliseu Maciel FAEM/UFPel, no município de Capão do Leão/RS. Dois
experimentos foram realizados, um com o herbicida Only® e outro com o Kifix®. O
delineamento experimental utilizado nos dois experimentos foi de parcelas
subdivididas, em esquema fatorial 5x3, sendo o fator A dose do herbicida e o fator B

18 tipos de arroz. As parcelas foram representadas por caixas de madeira (60cm de
comprimento x 40cm de largura x 20cm de altura), forradas com lona plástica e
preenchidas com 50kg de solo (Tab. 1). Para o fator dose foram utilizadas às
dosagens de 0, 5, 10 (dose comercial recomendada), 15 e 20mg p. c. m-2 do produto
comercial Only® e 0, 7, 14 (dose comercial recomendada), 21 e 28mg p. c. m-2 do
produto comercial Kifix®, ambos acrescidos de 0,5% v/v, do adjuvante Dash®. As
parcelas foram dividida em três sub-parcelas, contendo linhas independentes dos
três tipos de arroz: a) arroz cultivado cv. Puitá Inta CL, b) ecótipo de arroz-vermelho
sensível aos herbicidas do grupo das imidazolinonas e, c) ecótipo de arroz-vermelho
com suspeita de tolerância aos herbicidas do grupo das imidazolinonas. Em cada
linha foram semeadas 25 sementes (de um dos tipos de arroz) e após a germinação
foram desbastadas mantendo-se 15 plantas por linha, estabelecendo um
espaçamento de 2cm entre plantas e de 5cm entre linhas. Cada linha foi
considerada como sendo uma repetição, totalizando seis por tratamento.
Tabela 1. Propriedades físicas e químicas do solo utilizado nos experimentos
Registro
pH água
Ca Mg Al H+Al CTC efetiva
Saturação (%) Índice SMP
cmolc/dm3 Al Bases Amostra 4,5 2,2 1 0,5 4,4 4 12 44 6
Registro
% Mat. Org. m/v
% Argila
Text. P-Mehlich mg/dm3
P-resina CTC pH 7 cmolc/dm3
K mg/dm3
Amostra 1,4 13 4 40 --- 7,9 99
A aplicação dos herbicidas foi realizada em pós-emergência, quando o arroz
estava no estádio fenológico V3-V4 (SOSBAI, 2010), correspondente a 20 dias após
a semeadura, utilizando um pulverizador costal, pressurizado a CO2, munido de
quatro pontas de pulverização do tipo leque (110.015) e à pressão constante de
210kPa. A regulagem utilizada no equipamento proporcionou a aplicação
equivalente ao volume de 150L ha-1 de calda herbicida. A irrigação por inundação foi
efetuada no sétimo dia após a aplicação dos tratamentos.
As variáveis-resposta determinadas foram: fitotoxicidade, altura de plantas e
massa seca da parte aérea (MSA). As avaliações visuais de fitotoxicidade foram
realizadas aos 15, 30 e 60 dias após a aplicação dos tratamentos (DAA), seguindo a
escala percentual, em que zero representou ausência de dano e 100% morte das
plantas (FRANS et al., 1986). A altura de plantas, ou seja, a distância entre o colo da

19 planta até o ápice da folha mais jovem, foi medida quando as plantas de arroz, na
parcela testemunha, atingiram o estádio V6 (SOSBAI, 2010) o que correspondeu
aos 80 dias após a semeadura. A seguir, as plantas foram cortadas rente ao solo e
transferidas para uma estufa de circulação forçada de ar à temperatura de 70±5ºC
até atingir massa constante, quando foram determinadas as massas em balança
analítica.
Os resultados foram submetidos à análise da variância (p≤0,05), e os dados
ajustados por modelos de regressão polinomial (MACHADO; CONCEIÇÃO, 2007). A
escolha dos modelos foi baseada na significância estatística (teste F), no ajuste do
coeficiente de determinação (R2) e no significado biológico do modelo, conforme o
proposto por Adati et al. (2006).
3. RESULTADOS E DISCUSSÃO
Nos dois herbicidas testados à análise da variância demonstrou interação
significativa entre o fator dose e cultivar para todas as variáveis avaliadas.
Os sintomas causados pelos herbicidas imidazolinonas acontecem
aproximadamente 15 DAA, apresentando inicialmente clorose das folhas jovens, e
surgimento posterior de coloração avermelhada nas nervuras da face abaxial, com
posterior evolução para necrose das folhas como descrito por Tan et al. (2006). A
expressão destes sintomas permitiu a realização das avaliações de fitotoxicidade.
Na avaliação de fitotoxicidade realizada aos 15 DAA, os resultados ajustaram-se a
modelos lineares (Fig. 1 A e B). A cv. Puitá Inta CL apresentou fitotoxicidade de 3%
e 10%, respectivamente, para as doses comerciais dos herbicida Only® (Fig. 1A) e
Kifix® (Fig. 1B). Ambos herbicidas controlaram apenas em 45% o arroz-vermelho
sensível aos herbicidas do grupo das imidazolinonas, quando aplicados na dose
comercial. Já no dobro da dose o herbicida Only® apresentou controle 10% superior
quando comparado ao herbicida Kifix®. O herbicida Only demonstrou controle de 9 e
41% do ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do
grupo das imidazolinonas, respectivamente para a dose comercial e o dobro da
dose, enquanto que o herbicida Kifix controlou em 22 e 51% este ecótipo,
respectivamente para a dose comercial e o dobro da dose.

20 (A)
Fito
toxi
cida
de (%
)
0
20
40
60
80
100
120
(B)
(C)
Fito
toxi
cida
de (%
)
0
20
40
60
80
100
120
(D)
(E)
Doses do herbicida Only® (mg p. c. m-2)0 5 10 15 20
Fito
toxi
cida
de (%
)
0
20
40
60
80
100
120
(F)
Doses do herbicida Kifix® (mg p. c. m-2)
0 7 14 21 28
Figura 1. Fitotoxicidade aos 15 (A e B), 30 (C e D) e 60 (E e F) dias após a aplicação dos herbicidas Only®(A) e Kifix®(B) nas plantas arroz cv. Puitá Inta CL (• e ------); ecótipo de arroz-vermelho sensível aos herbicidas do grupo químico das imidazolinonas (o e - - -) e ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do grupo químico das imidazolinonas (▼ e .........). UFPel, Capão do Leão/RS, 2010. Os pontos apresentados nos gráficos são os valores originais. (mg p. c. m-2)
y = -0,03+0,3x; R2= 0,81 ( • ) y = -8,28+6,6x-0,13x2; R2 = 0,85 (-°-) y = -0,18-0,28x+0,12x2; R2 = 0,99 (..▼..)
y = -0,63+1,11x-0,03x2; R2= 0,95 ( • ) y = -3,84+4,47x-0,07x2; R2 = 0,95 (-°-) y = -5,92+2,02x; R2 = 0,94 (..▼..)
y = -2+1,02x; R2= 0,90 ( • ) y = -5,3+5,46x; R2 = 0,88 (-°-) y = -7,8-3,41x; R2 = 0,93 (..▼..)
y = 0,81+0,74x; R2= 0,95 ( • ) y = 2,94+3,58x; R2 = 0,94 (-°-) y = 0,78+1,83x; R2 = 0,99 (..▼..)
y = -1,96+1,12x; R2= 0,95 ( • ) y = -3,95+5,99x; R2 = 0,84 (-°-) y = -8,98+3,99x; R2 = 0,93 (..▼..)
y = 0,35+1,82x-0,04x2; R2= 0,96 ( • ) y = -5,99+7,49x-0,13x2; R2 = 0,96 (-°-) y = 2,13+2,14x; R2 = 0,99 (..▼..)

21
A segunda avaliação de fitotoxicidade realizada aos 30 dias após a
aplicação dos herbicidas foi realizada (Fig. 1 C e D), mostra que a dose comercial de
ambos os herbicidas foi tóxica em aproximadamente 10% para a cv. Puitá Inta CL.
Os herbicidas Only® (Fig. 1C) e Kifix® (Fig. 1D), nas doses comerciais, controlaram
respectivamente, em 49 e 53%, o arroz-vermelho sensível aos herbicidas do grupo
das imidazolinonas. Os dois herbicidas demonstraram controle de 100% deste
ecótipo quando aplicados no dobro da dose comercial. O ecótipo de arroz-vermelho
com suspeita de tolerância aos herbicidas do grupo das imidazolinonas foi
controlado em 26% por ambos os herbicidas na dose comercial, e em 60 e 52%,
respectivamente pelos herbicidas Only® e Kifix® aplicados no dobro da dose.
As fitotoxicidades determinadas aos 60 DAA (Fig. 1E e 1F) foram superiores
para todos os tipos de arroz e herbicidas utilizados, em comparação as avaliações
anteriores (15 e 30 DAA). O herbicida Kifix® (Fig. 1E) causou o dobro da toxicidade
em comparação ao herbicida Only® (Fig. 1F), quando aplicados na dose comercial,
para a cv. Puitá Inta CL. No dobro da dose, a fitotoxicidade causada por ambos os
herbicidas foi de aproximadamente 21%. O controle do arroz-vermelho sensível aos
herbicidas do grupo das imidazolinonas foi de 56 e 74%, respectivamente para os
herbicidas Only® e Kifix® aplicados na dose comercial. Os herbicidas controlaram
32% do ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do
grupo das imidazolinonas, quando aplicados na dose comercial. Porém, quando
aplicado no dobro da dose comercial, o herbicida Only® controlou 71%, superando
os 62% do herbicida Kifix® na mesma dose.
As plantas da cv. Puitá Inta CL, para ambos herbicidas aplicados, possuem
menor altura do que os dois genótipos de arroz-vermelho na dose de 0g ha-1
(controle) (Fig. 2). O crescimento das plantas de arroz cv. Puitá Inta CL para os dois
herbicidas foi semelhante, ou seja, pequeno incremento na altura foi verificado com
o aumento das doses dos herbicidas testados, demonstrando que as plantas desta
cultivar mostram ser tolerantes a ambos os herbicidas. As plantas de arroz-vermelho
sensível a herbicidas do grupo das imidazolinonas sofreram decréscimo de altura a
partir da dose comercial recomendada de 58% para ambos os herbicidas utilizados
(Fig. 2A e 2B), no entanto, quando foi utilizada doses acima da comercial ocorreu
morte total de plantas para ambos herbicidas. Também, as plantas do ecótipo de
arroz-vermelho com suspeita de tolerância sofreram reduções de altura, quando
aplicada à dose comercial, de 5 e 8%, respectivamente para Only® (Fig. 2A) e Kifix®
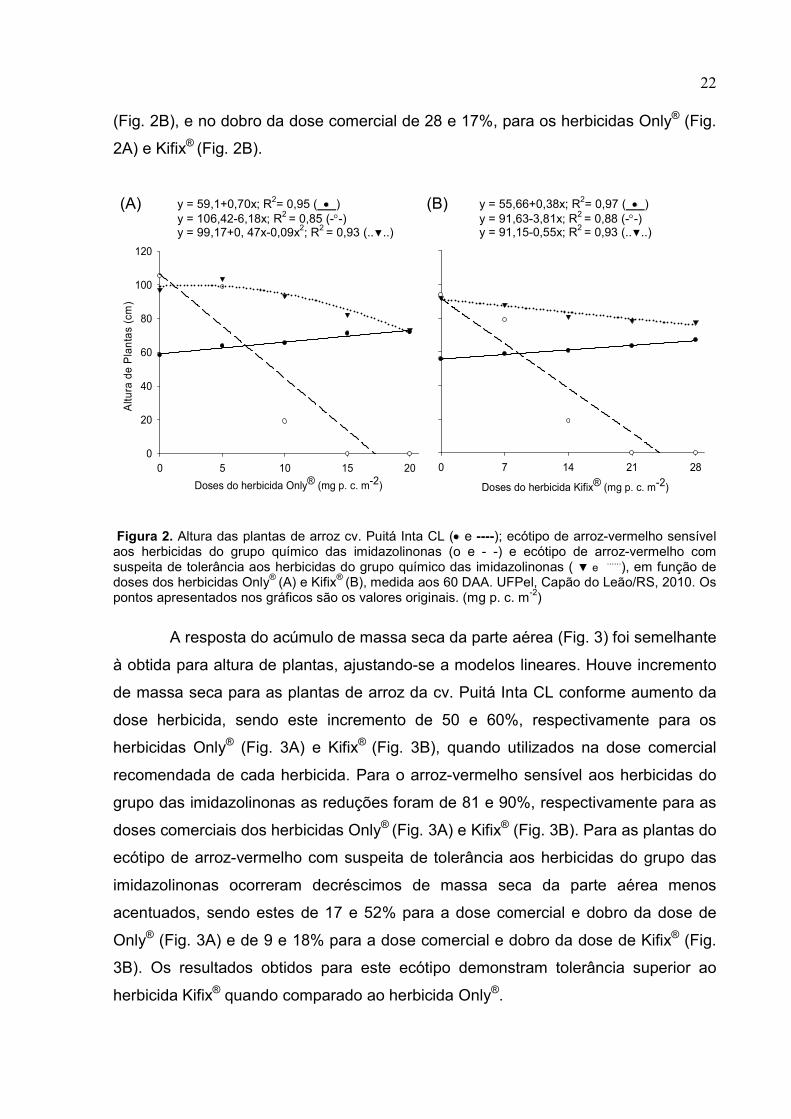
22 (Fig. 2B), e no dobro da dose comercial de 28 e 17%, para os herbicidas Only® (Fig.
2A) e Kifix® (Fig. 2B).
(A)
Doses do herbicida Only® (mg p. c. m-2)0 5 10 15 20
Altu
ra d
e Pl
anta
s (c
m)
0
20
40
60
80
100
120
(B)
Doses do herbicida Kifix® (mg p. c. m-2)
0 7 14 21 28
Figura 2. Altura das plantas de arroz cv. Puitá Inta CL (• e ----); ecótipo de arroz-vermelho sensível aos herbicidas do grupo químico das imidazolinonas (o e - -) e ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do grupo químico das imidazolinonas ( ▼ e ......), em função de doses dos herbicidas Only® (A) e Kifix® (B), medida aos 60 DAA. UFPel, Capão do Leão/RS, 2010. Os pontos apresentados nos gráficos são os valores originais. (mg p. c. m-2)
A resposta do acúmulo de massa seca da parte aérea (Fig. 3) foi semelhante
à obtida para altura de plantas, ajustando-se a modelos lineares. Houve incremento
de massa seca para as plantas de arroz da cv. Puitá Inta CL conforme aumento da
dose herbicida, sendo este incremento de 50 e 60%, respectivamente para os
herbicidas Only® (Fig. 3A) e Kifix® (Fig. 3B), quando utilizados na dose comercial
recomendada de cada herbicida. Para o arroz-vermelho sensível aos herbicidas do
grupo das imidazolinonas as reduções foram de 81 e 90%, respectivamente para as
doses comerciais dos herbicidas Only® (Fig. 3A) e Kifix® (Fig. 3B). Para as plantas do
ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do grupo das
imidazolinonas ocorreram decréscimos de massa seca da parte aérea menos
acentuados, sendo estes de 17 e 52% para a dose comercial e dobro da dose de
Only® (Fig. 3A) e de 9 e 18% para a dose comercial e dobro da dose de Kifix® (Fig.
3B). Os resultados obtidos para este ecótipo demonstram tolerância superior ao
herbicida Kifix® quando comparado ao herbicida Only®.
y = 55,66+0,38x; R2= 0,97 ( • ) y = 91,63-3,81x; R2 = 0,88 (-°-) y = 91,15-0,55x; R2 = 0,93 (..▼..)
y = 59,1+0,70x; R2= 0,95 ( • ) y = 106,42-6,18x; R2 = 0,85 (-°-) y = 99,17+0, 47x-0,09x2; R2 = 0,93 (..▼..)

23
(A)
Doses do herbicida Only® (mg p. c. m-2)
0 5 10 15 20
Mas
sa s
eca
parte
aér
ea (c
m)
0,0
0,4
0,8
1,2
1,6
2,0
(B)
Doses do herbicida Kifix® (mg p. c. m-2)
0 7 14 21 28
Figura 3. Acúmulo de matéria seca na parte aérea de plantas de arroz cv. Puitá Inta CL (• e ----); ecótipo de arroz-vermelho sensível aos herbicidas do grupo químico das imidazolinonas (o e - -) e ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do grupo químico das imidazolinonas ( ▼ e ......), em função de doses dos herbicidas Only® (A) e Kifix® (B), medida aos 60 DAA. UFPel, Capão do Leão/RS, 2010. Os pontos apresentados nos gráficos são os valores originais. (mg p. c. m-2)
Estudos mostraram o potencial de toxidez inicial às plantas de arroz quando
utilizado o herbicida imazethapyr, porém com o decorrer do tempo há uma
recuperação (VILLA et al., 2006). Além disto, o incremento da dose herbicida resulta
em aumento na fitotoxicidade de cultivares de arroz tolerante (DAL MAGRO et al.,
2006). Este aumento de toxidez com o incremento da dose herbicida foi observado
na cv. Puitá Inta CL, para os dois herbicidas utilizados. No entanto, a altura de
plantas e a massa seca da parte aérea comprovam que esta toxidez não foi
prejudicial para o crescimento das plantas. Vários autores demonstram que a altura
de plantas tolerantes, observada no momento da colheita, pode não ser afetada pela
aplicação de imazethapyr ou imazethapyr + imazapic (PELLERIN; WEBSTER, 2004;
VILLA et al., 2006). No presente experimento, o aumento das doses resultou em um
incremento de altura e massa seca da parte aérea desta cultivar, sendo este último
superior a 50% para ambos herbicidas aplicados na dose comercial, fato este que
pode ser explicado pelo controle do arroz-vermelho, que se encontrava nas linhas
paralelas a esta cultivar. Geralmente, o arroz-vermelho é mais alto, produz mais
perfilhos e apresenta maior biomassa do que o arroz cultivado (ESTORNINOS et al.,
y = 0,75+0,046x; R2= 0,98 ( • ) y = 1,86-0,2x+0,005x2; R2 = 0,89 (-°-) y = 1,67-0,01x-0,002x2; R2 = 0,91 (..▼..)
y = 0,73+0,027x; R2= 0,90 ( • ) y = 1,20-0,11x+0,003x2; R2 = 0,99 (-°-) y = 1,35-0,008x; R2 = 0,94 (..▼..)

24 2005), apresentando também uma maior eficiência na utilização do nitrogênio (N)
quando comparado ao arroz cultivado em condições competitivas, absorvendo até
60% do N do fertilizante aplicado (BURGOS et al. 2006). Essas características
fazem do arroz-vermelho, quando presente na lavoura, um concorrente mais forte
por espaço, nutrientes, água e luz do que o arroz cultivado (SHIVRAIN et al., 2009),
o que prejudica o desenvolvimento da cultura.
O efeito dos herbicidas Only e Kifix sobre o ecótipo de arroz-vermelho
sensível foi semelhante. A fitotoxicidade apresentada pelas plantas em todas as
avaliações se manteve aproximadamente estável, sofrendo pequenos acréscimos. O
controle das plantas de arroz-vermelho sensível foi superior a 50%, quando
expostas a aplicação das doses comerciais dos herbicidas, o que é possível ser
observado através das avaliações de fitotoxicidade, altura de plantas e massa seca
da parte aérea. Os herbicidas imazethapyr+imazapic nas doses de 7,5+2,5mg p.c.
m-2 aplicados em pós emergência, não apresentam controle total do arroz-vermelho,
possibilitando escape desta planta daninha (SANTOS et al., 2007). O controle foi
aumentado com o incremento das doses, chegando a 100% quando utilizadas doses
superiores a 15 e 21mg p. c. m-2, respectivamente para os herbicidas Only e Kifix.
Ainda, incrementos na dose dos herbicidas imazethapyr+imazapic aumentam o
controle de arroz-vermelho sensível (FONTANA et al., 2007).
O aumento na eficácia de controle de arroz-vermelho com incremento na
dose de imazethapyr aplicado em pós-emergência também foi observado em outros
trabalhos, atingindo controle superior a 90% (STEELE et al., 2002). Foram
observadas reduções superiores de massa seca da parte aérea quando comparadas
as reduções obtidas para altura de plantas, em ambos herbicidas aplicados na dose
comercial. Esta altura de plantas encontrada se deve ao fato de que estes herbicidas
são aplicados em pós-emergência das plantas de arroz, ou seja, quando estas já
apresentam um determinado crescimento. Porém, devido ao mecanismo de ação
destes produtos, que interrompem a formação dos aminoácidos leucina, valina e
isoleucina, através da inibição da enzima ALS sensível, ocorre uma paralisação do
crescimento das plantas, por meio da inibição da divisão celular. Entretanto, a
massa seca aérea destas plantas diminui após a paralisação deste crescimento,
pois, além disto, estes herbicidas inibem a síntese de acetil-CoA, precursor inicial da
biossíntese de ácidos graxos e do ciclo de Krebs, e inibem o transporte de
fotossintetizados (KRAEMER et al., 2009), o que obriga o vegetal a consumir as

25 suas reservas, resultando no seu definhamento, podendo levá-lo a morte,
dependendo da gravidade da injúria.
O ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do
grupo das imidazolinonas apresentou toxicidade semelhante para ambos os
herbicidas testados, sendo esta crescente durante as avaliações. Porém, os
resultados mostram que na aplicação da dose comercial dos produtos a toxicidade
não ultrapassou 35%, ou seja, valor este significativamente inferior ao encontrado
para o ecótipo sensível. Os resultados obtidos para as variáveis-resposta altura de
plantas e massa seca da parte aérea corroboram com os encontrados para
fitotoxicidade, sendo observadas reduções inferiores as encontradas para o ecótipo
sensível. Entretanto, para estas duas variáveis a atuação dos herbicidas foi diferente
em todas as doses aplicadas, sendo menores as injúrias para o herbicida Kifix, do
que para o Only. O mecanismo de resistência aos inibidores da ALS é atribuído
principalmente a uma enzima alterada (TRANEL; WRIGHT, 2002) por uma das
mutações pontuais que modificam a seqüência de aminoácidos (WRIGHT et al.,
1998). Várias mutações que conferem resistência aos herbicidas inibidores da ALS
são identificadas em várias espécies de plantas (TRANEL; WRIGHT, 2002). Outros
mecanismos de resistência incluem a expressão gênica aumentada e a alteração do
metabolismo dos herbicidas na planta (RAJGURU et al., 2005).
4. CONCLUSÕES a) A cv. Puitá Inta CL de arroz é tolerante aos herbicidas Only® e Kifix® em
doses até o dobro da recomendada;
b) Os herbicidas Only® e Kifix® são eficientes no controle do ecótipo de arroz-
vermelho sensível aos herbicidas do grupo químico das imidazolinonas, nas
doses recomendadas;
c) As plantas do ecótipo de arroz-vermelho com suspeita de tolerância aos
herbicidas do grupo químico das imidazolinonas resistem aos herbicidas
Only® e Kifix®, mesmo utilizando-se doses superiores as doses
recomendadas.

26
CAPÍTULO 2
FLUORESCÊNCIA DA CLOROFILA a EM PLANTAS DE ARROZ EM RESPOSTA A APLICAÇÃO DE HERBICIDAS DO GRUPO DAS IMIDAZOLINONAS
Chlorophyll a Fluorescence in Rice Plants in Response to Application of Herbicides
of Group Imidazolinone
RESUMO - Este estudo objetivou avaliar as injúrias causadas ao aparelho fotossintético de três tipos de arroz, expostos à aplicação de herbicidas do grupo químico das imidazolinonas. O trabalho foi conduzido em casa-de-vegetação UFPel/FAEM, Capão do Leão/RS. Foram utilizados os herbicidas Only® e Kifix®, utilizando-se para ambos um delineamento experimental de parcelas subdivididas, em esquema fatorial 3x3, com seis repetições, sendo o fator A as doses dos herbicidas correspondentes a 0 (testemunha sem aplicação), 10 (dose comercial recomendada) e 20mg p. c. m-2 do produto comercial Only® e 0 (testemunha sem aplicação), 14 (dose comercial recomendada) e 28mg p. c. m-2 do produto comercial Kifix®, e o fator B o tipo de arroz (arroz cultivado cv. Puitá Inta CL, ecótipo de arroz-vermelho sensível aos herbicidas do grupo das imidazolinonas e ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do grupo das imidazolinonas). Foram avaliados os parâmetros da fluorescência da clorofila a nas plantas aos 30 dias após a aplicação dos herbicidas, utilizando-se um fluorômetro portátil (HandyPEA, Hansatech). As medidas foram realizadas em folhas totalmente expandidas, previamente adaptadas ao escuro, no período da manhã. Verificou-se que a cv. Puitá Inta CL tolera doses comerciais dos herbicidas Only® e Kifix®, demonstrando a capacidade de se recuperar das injúrias iniciais ao seu metabolismo fotossintético, o que pode ser observado na segunda avaliação. Para o ecótipo de arroz-vermelho sensível foi verificado, já na dose comercial dos dois herbicidas, um aumento de DI0/RC e uma redução de PIABS,total, o que demonstra a elevada sensibilidade deste ecótipo ao herbicida, já na dose comercial aplicada. Nas avaliações do ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do grupo químico das imidazolinonas, foi comprovada a tolerância deste ecótipo para ambos os herbicidas aplicados na dose comercial, sendo esta tolerância mantida para doses mais elevadas do herbicida Kifix®. A aplicação de herbicidas do grupo químico das imidazolinonas em plantas de arroz causa alterações no metabolismo fotossintético das plantas que podem ser detectadas através da avaliação da emissão da fluorescência da clorofila a transiente, sendo este método

27 útil na detecção de resistência e/ou tolerância de plantas de arroz-vermelho a estes herbicidas. Palavras-chave: inibidores ALS, Oryza sativa L., fotossistemas, teste JIP ABSTRACT: This study investigated the injuries to the photosynthetic apparatus of three types of rice exposed to application of imidazolinone chemical group herbicide. Research was conducted in a greenhouse, at Universidade Federal de Pelotas, Capão do Leão/RS. We used the herbicide Only® and Kifix®, using a split-plot experimental design in a 3x3 factorial schem, with six replications, with the first factor being the herbicide rates of 0 (control), 10 (recommended rate) and 20mg p. c. m-2 of commercial product Only® and 0 (control), 14 (recommended rate) and 28mg p. c. m-
2 of commercial product Kifix®, and factor B type of rice (grown rice cv. Puitá Inta CL, red rice ecotype sensitive to herbicides Imidazolinone and red rice ecotype with suspected herbicide tolerance Imidazolinone). We evaluated the parameters of chlorophyll fluorescence in plants at 30 days after herbicide application, using a portable fluorometer (HandyPEA, Hanstech). Measurements were performed in fully expanded leaves, previously dark adapted, in the morning. It was found that cv. Puitá Inta CL tolerate doses commercial of herbicide Only® and Kifix®, this cultivar has demonstrating the ability to recover from initial photosynthetic metabolism injury, which can be observed in the second evaluation. For the red rice ecotype sensitivity was found already at a dose of two commercial herbicides, an increase of DI0/RC and a partial reduction PIABS,total, which demonstrates the high sensitivity of this ecotype to the herbicide, since the recommended rate applied. Evaluations of red rice ecotype with suspected herbicide tolerance to the imidazolinone chemical group, was shown an ecotype tolerance for both herbicides applied at recommended rate, which is maintained tolerance to higher doses of the herbicide Kifix®. The application of herbicides of the imidazolinone chemical group in rice plants causes changes in the photosynthetic metabolism of plants that can be detected by evaluating the emission of chlorophyll a fluorescence transient being the method useful for detecting resistance and/or tolerance of plants red rice to these herbicides.
Keywords: ALS-inhibiting, Oryza sativa L., photosystems, JIP-test
1. INTRODUÇÃO Acetolactato sintase (ALS, EC 2.2.1.6), também conhecida como
acetohidroxiácido sintase (AHAS), é a primeira enzima na via de biossíntese dos
aminoácidos essenciais de cadeia ramificada, leucina, isoleucina e valina (LE et al.,
2005). Esta enzima catalisa a condensação de duas moléculas de piruvato, para
formar acetolactato na rota metabólica da biossíntese da leucina e da valina, ou
catalisa a condensação de uma molécula de piruvato com uma molécula de 2-
cetobutirato, para formar 2-aceto-2-hidroxibutirato (AHB) como primeiro passo na
biossíntese da isoleucina (CHIPMAN et al., 2005). No final da década de 70, foi
descoberta a primeira classe herbicida capaz de inibir esta enzima, as sulfoniluréias.
Atualmente, a ALS é o local de ação para mais de 50 herbicidas que constituem

28 cinco classes estruturalmente distintas de produtos químicos: estas classes são:
sulfoniluréias, imidazolinonas, triazolpirimidinas sulfonanilidas, pirimidil-oxi-
tiobenzoatos, e sulfonilamino-carboniltriazolinona (HEAP, 2009). Devido à sua baixa
toxicidade em mamíferos, as doses de aplicação extremamente baixas, a alta
eficácia e o amplo espectro de ação, os herbicidas inibidores da ALS são
intensamente utilizados para controlar um amplo espectro de plantas daninhas em
diversas culturas (TAN et al., 2005).
Os sintomas visuais de fitotoxicidade incluem o acúmulo de antocianinas,
clorose foliar, e a necrose dos pontos de crescimento (TAN et al., 2006) aparecem
vários dias ou semanas após a aplicação destes herbicidas, porém mudanças
profundas no metabolismo das plantas ocorrem logo após a aplicação dos
herbicidas inibidores da ALS. Dentre estas mudanças estão a paralisação do
crescimento, que ocorre dentro de horas após a aplicação (GASTON et al., 2003), a
inibição da mitose e síntese de DNA, e diminuição na translocação de
fotoassimilados para os pontos de crescimento da planta (VILLA et al., 2006;
KRAEMER et al., 2009).
Embora a fotossíntese não seja considerada um alvo primário de herbicidas
inibidores da ALS, são detectadas alterações nas respostas de fluorescência da
clorofila a em plantas tratadas com estes herbicidas (SILVA et al., 2009). Além disso,
a inibição de reações metabólicas não envolvidas diretamente na fotossíntese pode
modificar a síntese de intermediários utilizados nesse processo, interferindo na
fotossíntese e conseqüentemente na emissão de fluorescência (XIA et al., 2006).
Em função da aplicação de herbicidas, mesmo quando os sintomas ainda não são
visíveis, podem ocorrer injúrias causadas ao aparelho fotossintético, reduzindo a
capacidade fotossintética das plantas (BARBAGALLO et al., 2003).
A mensuração de fluorescência das clorofilas é um método rápido, simples e
não invasivo, sendo largamente utilizada no entendimento dos mecanismos da
fotossíntese, bem como na avaliação da capacidade fotossintética alterada por
estresses bióticos ou abióticos pelos quais as plantas possam passar. A redução na
dissipação da energia pelo processo fotoquímico da fotossíntese é refletida por um
incremento correspondente na fluorescência, a qual pode ser aferida em folhas
intactas (MAXWELL; JOHNSON, 2000), sendo uma das formas de monitoramento
da inibição ou redução na transferência de elétrons entre os fotossistemas.

29
A avaliação da cinética de emissão da fluorescência da clorofila a permite o
cálculo de parâmetros de fluorescência que caracterizam o funcionamento do
aparelho fotossintético (YUSUF et al., 2010). Esta avaliação pode ser descrita
utilizando-se o teste JIP (STRASSER; STRASSER, 1995; TSIMILLI-MICHAEL;
STRASSER, 2008), quantificando o fluxo de energia que passa pelos fotossistemas
e avalia o desempenho fotossintético de plantas (STRASSER & STRASSER, 1995;
STRASSER et al., 2004, TSIMILLI-MICHAEL; STRASSER, 2008).
Em virtude da escassez de informações sobre o efeito das imidazolinonas no
aparelho fotossintético determinado pelo método da fluorescência das clorofilas, este
trabalho teve por objetivo empregar esta metodologia para avaliar as injúrias
causadas na maquinaria fotossintética de três tipos de arroz, expostos à aplicação
de herbicidas do grupo químico das imidazolinonas.
2. MATERIAL E MÉTODOS O experimento foi conduzido em casa de vegetação da Faculdade de
Agronomia Eliseu Maciel FAEM/UFPel, no município de Capão do Leão/RS. Foram
utilizados os herbicidas Only® e Kifix®, utilizando-se para ambos um delineamento
experimental de parcelas subdivididas, em esquema fatorial 3x3. O fator A avaliou
doses dos herbicidas, correspondentes a 0 (controle), 10 (dose comercial
recomendada) e 20mg p. c. m-2 do produto comercial Only® e 0 (controle), 14 (dose
comercial recomendada) e 28 mg p. c. m-2 do produto comercial Kifix®, ambos
acrescidos de 0,5% v/v, do adjuvante Dash®. O fator B avaliou tipos de arroz: a)
arroz cultivado cv. Puitá Inta CL, b) ecótipo de arroz-vermelho sensível aos
herbicidas do grupo das imidazolinonas e, c) ecótipo de arroz-vermelho com
suspeita de tolerância aos herbicidas do grupo das imidazolinonas. As parcelas
foram representadas por caixas de madeira (60cm de comprimento x 40cm de
largura x 20cm de altura), forradas com lona plástica e preenchidas com 50kg de
solo. As parcelas foram dividida em três sub-parcelas, contendo linhas
independentes dos três tipos de arroz. Na linha foram semeadas isoladamente 25
sementes de cada tipo de arroz, e após a germinação foram desbastadas mantendo-
se 15 plantas por linha, estabelecendo um espaçamento de 2cm entre plantas e de
5cm entre linhas. Cada linha foi considerada como sendo uma repetição, totalizando
seis por tratamento.

30
A aplicação dos herbicidas foi realizada em pós-emergência, quando o arroz
encontrava-se no estádio fenológico V3-V4 (SOSBAI, 2010), identificado pela
formação do colar na quarta folha do colmo principal (COUNCE et al., 2000), o que
correspondeu a aproximadamente 20 dias após a semeadura, utilizando-se um
pulverizador costal, pressurizado a CO2, munido de quatro pontas de pulverização
do tipo leque (110.015) e à pressão constante de 210kPa. A regulagem utilizada no
equipamento proporcionou a aplicação equivalente ao volume de 150L ha-1 de calda
herbicida. A irrigação por inundação iniciou-se no sétimo dia após a aplicação dos
tratamentos.
Os parâmetros da cinética de emissão da fluorescência transiente da
clorofila a das plantas foram avaliados aos 15 e 30 dias após a aplicação dos
herbicidas (DAA). O monitoramento da emissão da fluorescência da clorofila a foi
realizado utilizando um fluorômetro portátil (HandyPEA, Hanstech, King’s Lynn,
Norkfolk, UK). As medições foram realizadas no terço médio de folhas jovens
completamente expandidas, no período da manhã, e realizadas 20 minutos após a
adaptação das folhas ao escuro. A emissão de fluorescência foi induzida em uma
área de 4 mm de diâmetro da folha pela exposição da amostra a um pulso de luz
saturante numa intensidade de 3.000μmol m-2 s-1. As intensidades de fluorescência
foram determinadas a 50, 100, 300μs, 2 (FJ) e 30 (FI) ms e FM (fluorescência
máxima). A partir das intensidades de fluorescência foram calculados os parâmetros
estabelecidos pelo Teste JIP (STRASSER; STRASSER, 1995), sendo que a
cuidadosa interpretação dos parâmetros medidos e calculados a partir deste teste
fornece numerosas informações dos fluxos de energia através do FSII em diferentes
níveis (STRASSER et al., 2004).
Os parâmetros de fluorescência da clorofila fornecem informações sobre
propriedades biofísicas da conversão fotossintética de energia e transporte de
elétrons. Pelo Teste JIP, diversas expressões biofísicas, tais como o índice de
vitalidade, mudanças fenomenológicas, razões de fluxo, rendimentos e fluxos
específicos podem ser calculadas.
Dentre os parâmetros propostos por Strasser e Strasser (1995) podem ser
destacados os que descrevem: a) a atividade específica por centro de reação (RC)
como sendo: ABS/RC - o qual pode ser uma medida do tamanho aparente do
sistema antena ou o fluxo de absorção por RC; TR0/RC – representa a máxima taxa
pela qual um éxciton é capturado pelo RC resultando em uma redução da

31 plastoquinona (QA
-); ET0/RC – demonstra a reoxidação da QA- via transporte de
elétrons em um RC ativo; DI0/RC - que é a razão de dissipação total de energia de
excitação não capturada do total de RC, sendo a dissipação neste caso à perda de
energia na forma de calor; RE0/RC - redução do aceptor final de elétrons no lado do
aceptor de elétrons do FSI por RC; b) os rendimentos energéticos ou taxas de fluxo:
φP0 = TR0/ABS = FV/FM - rendimento quântico máximo fotoquímico primário; φE0 =
ET0/ABS - o rendimento quântico de transporte de elétrons de QA- para o
intersistema de aceptores de elétrons; φD0 - descreve o rendimento quântico para
dissipação de energia; φR0 = RE0/ABS - rendimento quântico de transporte de
elétrons de QA- para o aceptor final de elétrons do FSI; c) as eficiências: ψ0 =
ET0/TR0 – eficiência com que um éxciton capturado no RC pode mover um elétron
de QA- para o intersistema de aceptores de elétrons; ρ0 – eficiência com que um
éxciton capturado no RC pode mover um elétron dentro da cadeia de transporte de
elétrons de QA- para os aceptores finais de elétrons do FSI; δ0 – eficiência com que
um elétron pode mover o intersistema de aceptores de elétrons reduzidos para o
aceptor final de elétrons do FSI (STRASSER e STRASSER, 1995; STRASSER et
al., 2004).
Um dos parâmetros que permite a interpretação completa do sistema de
absorção e aproveitamento da energia na cadeia de transporte de elétrons da
fotossíntese e descreve o efeito de um estresse sobre o aparato fotossintético é o
índice de performance fotossintético o qual representa a combinação de três
componentes parciais, que descrevem: a) a quantidade de RC fotossintético
(RC/ABS); b) o fluxo máximo de energia que atinge o RC do FSII [φP0/(1-φP0)]; e c)
transporte de elétrons no início da iluminação [Ψ0/(1-Ψ0)], conforme equação abaixo:
PI��� �RC
ABS � �
φ��1 � φ��
� � �Ψ�
1 � �� �
RCABS
�TR�
DI��
ET�1 � ET�
Contudo, Tsimilli-Michael e Strasser (2008), introduziram mais um
componente nesta fórmula que descreve as reações de oxi-redução que ocorrem no
lado aceptor de elétrons do FSI [δ0/(1- δ0)], propondo o termo índice de performance
total (PIABS,total) o qual é descrito pela fórmula:

32
PI���,����� � PI��� � �δ�
1 � �
A criteriosa interpretação destes parâmetros permite a inferência sobre a
funcionalidade da cadeia de transporte de elétrons da fotossíntese em resposta a
um fator que cause direta ou indiretamente qualquer perturbação, seja na absorção,
captura e transferência de energia entre os fotossistemas.
3. RESULTADOS E DISCUSSÃO Em todas as plantas, a intensidade de emissão de fluorescência da clorofila a
mostrou uma típica curva em função do tempo.
A aplicação do herbicida Only® (Fig. 1A) na dose comercial provocou nas
plantas de arroz cv. Puitá Inta CL, aos 15 DAA uma elevação em: ABS/RC, DI0/RC,
RE0/RC, φD0 e δ0, associado com uma redução em φE0 e Ψ0 resultando no
decréscimo em PIABS. Contudo após 30 DAA praticamente todos os parâmetros de
fluorescência foram similares ao controle, demonstrando uma recuperação nos
danos iniciais ao aparato fotossintético causado por este herbicida. Contudo, a dose
de 20mg p. c.m-2 de produto comercial, causou aos 15 DAA as mesmas variações
nos parâmetros de fluorescência, além da elevação em φR0 e ρ0 o que resultou no
incremento acentuado em PIABS,total. Diferentemente da dose comercial, a aplicação
do dobro da dose manteve ou mesmo acentuou as variações nos parâmetros de
fluorescência resultando em decréscimo acentuado nos dois índices de performance
(PIABS e PIABS,total) aos 30 DAA, indicando marcante dano ao aparato fotossintético.

33
Figura 1. Efeito das doses do herbicida Only® (A) e Kifix® (B) sob os parâmetros da fluorescência da clorofila a, obtidos através do teste JIP, (centro radarplot = 0,0, máximo = 2,5) em relação ao padrão de comportamento - controle (linha cheia = 1,0), para a cv. Puitá Inta CL, aos 15 e aos 30 dias após a aplicação dos herbicidas. UFPel, Capão do Leão/RS, 2010. (mg p. c. m-2)
0.0
0.5
1.0
1.5
2.0
2.5ABS/RC
TRo/RC
ETo/RC
DIo/RC
REo/RC
φPo
φEo
φDo
φRo
ψo
δo
ρ0
PI ABS
PI ABS, total
controle
10 mg p. c. m-2 - 15 DAA
10 mg p. c. m-2 - 30 DAA
20 mg p. c. m-2 - 15 DAA
20 mg p. c. m-2 - 30 DAA
(A)
0.0
0.5
1.0
1.5
2.0
2.5ABS/RC
TRo/RC
ETo/RC
DIo/RC
REo/RC
φPo
φEo
φDo
φRo
ψo
δo
ρ0
PI ABS
PI ABS, total
controle 14 mg p. c. m-2 - 15 DAA14 mg p. c. m-2 - 30 DAA28 mg p. c. m-2 - 15 DAA28 mg p. c. m-2 - 30 DAA
(B)

34
Da mesma maneira, a aplicação do herbicida Kifix® (Fig. 1B) na dose comercial
(14mg p. c. m-2 de produto comercial) induziu as mesmas alterações verificadas para
o herbicida Only® nas plantas de arroz cv. Puitá Inta CL, aos 15 DAA. Contudo aos
30 DAA não houve reversão dos danos ao aparato fotossintético, destacando o
incremento em PIABS,total, resultado da manutenção de valores elevados dos
parâmetros associados ao fluxo de elétrons entre o intersistema de transporte de
elétrons e o aceptor final de elétrons do FSI (δ0 e φR0). O mesmo fato ocorreu com a
dose de 28mg p. c. m-2 do produto comercial, porém houve intensificação dos danos
tanto aos 15 quanto aos 30 DAA.
Os parâmetros de fluorescência para as plantas do ecótipo de arroz-vermelho
com suspeita de tolerância aos herbicidas do grupo das imidazolinonas estão na
Figura 2. O herbicida Only® (Fig. 2A), após 15 dias da aplicação da dose de 10mg p.
c. m-2 provocou elevação nos índices que descrevem a performance fotossintética
(PIABS e PIABS,total), contudo aos 30 DAA foi verificado uma inversão nestes
parâmetros, isto é, uma redução, indicando um comprometimento da absorção e
aproveitamento da energia luminosa resultado de um dano ao aparelho
fotossintético. Em contrapartida, a dose de 20mg p. c. m-2 de produto comercial do
mesmo produto causou um distúrbio completo no fluxo de energia e de elétrons na
cadeia fotossintética intensificada com os dias após a aplicação. Diferentemente do
herbicida Only®, a aplicação do Kifix® (Fig. 2B) na dose comercial (14mg p. c. m-2)
provocou alteração na eficiência fotoquímica aos 15 DAA, notadamente pela queda
nos índices de performance, contudo as plantas apresentam uma recuperação após
30 DAA do herbicida, demonstrando uma capacidade de superação dos danos
fotossintéticos iniciais. Porém, a dose mais elevada (28mg p. c. m-2) não
proporcionou acentuadas alterações nos parâmetros de fluorescência, destacando-
se leve queda nos índices de performance (PIABS e PIABS,total), tanto aos 15 quanto
aos 30 DAA, demonstrando que estas plantas tem capacidade, ainda incerta, de
metabolizar altas doses dos princípios ativos deste herbicida.

35
Figura 2. Efeito das doses do herbicida Only® (A) e Kifix® (B) sob os parâmetros da fluorescência da clorofila a, obtidos através do teste JIP, (centro radarplot = 0,0, máximo = 2,5) em relação ao padrão de comportamento - controle (linha cheia = 1,0), para o ecótipo de arroz-vermelho com suspeita de tolerância aos herbicidas do grupo das imidazolinonas, aos 15 e aos 30 dias após a aplicação dos herbicidas. UFPel, Capão do Leão/RS, 2010. (mg p. c. m-2)
0.0
0.5
1.0
1.5
2.0
2.5ABS/RC
TRo/RC
ETo/RC
DIo/RC
REo/RC
φPo
φEo
φDo
φRo
ψo
δo
ρ0
PI ABS
PI ABS, total
controle
10 mg p. c. m-2 - 15 DAA
10 mg p. c. m-2 - 30 DAA
20 mg p. c. m-2 - 15 DAA
20 mg p. c. m-2 - 30 DAA
(A)
0.0
0.5
1.0
1.5
2.0
2.5ABS/RC
TRo/RC
ETo/RC
DIo/RC
REo/RC
φPo
φEo
φDo
φRo
ψo
δo
ρ0
PI ABS
PI ABS, total
controle 14 mg p. c. m-2 - 15 DAA14 mg p. c. m-2 - 30 DAA28 mg p. c. m-2 - 15 DAA28 mg p. c. m-2 - 30 DAA
(B)

36
A resposta das plantas do ecótipo de arroz-vermelho susceptível aos
herbicidas do grupo químico das imidazolinonas (Only® e Kifix®) está representada
na Fig. 3. O herbicida Only® (Fig. 3A) gerou completa desorganização no sistema
fotossintético das plantas, estimado pelos parâmetros de fluorescência,
independente da dose, sendo o dano intensificado com o tempo após a aplicação do
herbicida. Também, este fato ocorreu com o herbicida Kifix® (Fig. 3B), o qual
inicialmente (15 DAA) na dose de 14mg p. c. m-2 de produto comercial, provocou
dano ao aparelho fotossintético, contudo houve uma recuperação para níveis
próximos ao controle aos 30 DAA, indicando uma possível tolerância destas plantas
a este herbicida. Na dose de 28mg p. c. m-2 de produto comercial, aos 15 DAA,
foram observadas injúrias ao aparelho fotossintético destas plantas, sendo estas
agravadas na avaliação realizada aos 30 DAA, fato este constatado pela drástica
elevação em: ABS/RC, DI0/RC e φD0, associado com uma redução em φE0, φR0 e Ψ0,
que ocasionaram praticamente decréscimo total dos índices de performance.
O herbicida Only® (imazethapyr+imazapic) é utilizado no Sistema de Produção
Clearfield® de produção de arroz, que consiste na utilização de genótipos de arroz
tolerante aos herbicidas imazethapyr e imazapic (VILLA et al., 2006). As cultivares
utilizadas neste sistema (CL) diferenciam-se entre si em relação à mutação do gene
ALS (TAN et al., 2005). Estas mutações reduzem a ligação da enzima ALS com a
maioria dos tipos de inibidores, o que resulta em pouca ou nenhuma alteração na
função enzimática (TRANEL; WRIGHT, 2002). Estudos com plantas de arroz da cv.
Puitá Inta CL demonstram alta tolerância aos herbicidas do grupo das
imidazolinonas quando comparada a outras cultivares CL (ROSO, 2010). Os
resultados obtidos neste trabalho demonstram uma sensibilidade inicial da cv. Puitá
Inta CL, porém recuperável ao longo do tempo, quando exposta a dose
recomendada do herbicida. Outras pesquisas demonstraram a capacidade de
recuperação das cultivares tolerantes aos efeitos negativos de imazethapyr (VILLA
et al., 2006). Porém os resultados deste trabalho sugerem que esta tolerância ocorra
apenas até a dose comercial do herbicida Only®, pois em doses superiores a esta,
as plantas desta cultivar não tiveram a capacidade de se recuperar das injúrias
causadas ao seu aparato fotossintético.
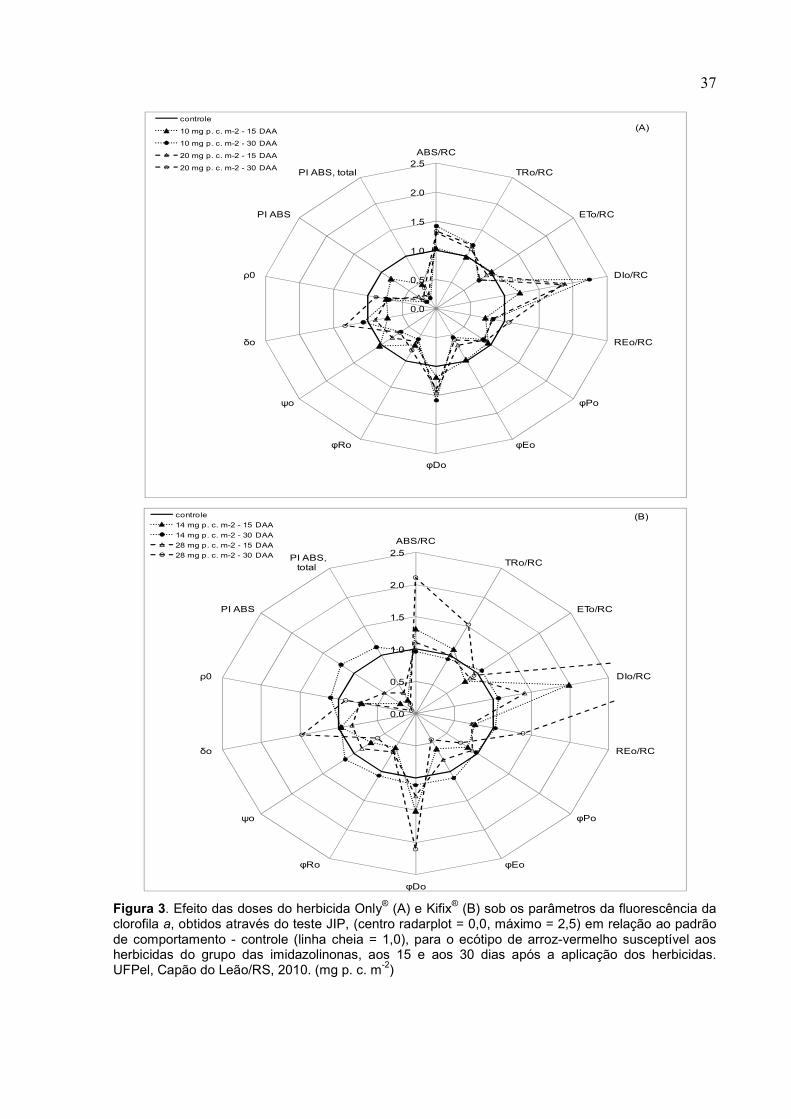
37
Figura 3. Efeito das doses do herbicida Only® (A) e Kifix® (B) sob os parâmetros da fluorescência da clorofila a, obtidos através do teste JIP, (centro radarplot = 0,0, máximo = 2,5) em relação ao padrão de comportamento - controle (linha cheia = 1,0), para o ecótipo de arroz-vermelho susceptível aos herbicidas do grupo das imidazolinonas, aos 15 e aos 30 dias após a aplicação dos herbicidas. UFPel, Capão do Leão/RS, 2010. (mg p. c. m-2)
0.0
0.5
1.0
1.5
2.0
2.5ABS/RC
TRo/RC
ETo/RC
DIo/RC
REo/RC
φPo
φEo
φDo
φRo
ψo
δo
ρ0
PI ABS
PI ABS, total
controle
10 mg p. c. m-2 - 15 DAA
10 mg p. c. m-2 - 30 DAA
20 mg p. c. m-2 - 15 DAA
20 mg p. c. m-2 - 30 DAA
(A)
0.0
0.5
1.0
1.5
2.0
2.5ABS/RC
TRo/RC
ETo/RC
DIo/RC
REo/RC
φPo
φEo
φDo
φRo
ψo
δo
ρ0
PI ABS
PI ABS, total
controle 14 mg p. c. m-2 - 15 DAA14 mg p. c. m-2 - 30 DAA28 mg p. c. m-2 - 15 DAA28 mg p. c. m-2 - 30 DAA
(B)

38
As plantas possuem mecanismos de desintoxicação de moléculas de
herbicidas inibidores da ALS como a hidroxilação do anel aromático, a hidroxilação
alifática, a desalquilação, a deesterificação, a conjugação e a ação da enzima
citocromo P450 mooxigenase (VIDAL; WINKLER, 2002). Entretanto, mediante a
aplicação de doses elevadas de herbicidas estes mecanismos podem não ser
suficientes para inativar estas moléculas, resultando em injúrias as plantas. No
ecótipo sensível aos herbicidas, foi verificado que estas plantas responderam de
forma muito semelhante nas duas doses testadas, sofrendo elevados danos ao seu
aparato fotossintético. No que concerne ao período de avaliação, aos 30 DAA o
prejuízo observado nas plantas foi superior ao encontrado aos 15 DAA, o que
comprova que este ecótipo não conseguiu se recuperar após ser submetido à ação
do herbicida Only®.
A biossíntese dos aminoácidos valina, leucina e isoleucina, é inibida pela
aplicação deste herbicida, ocorre principalmente em tecidos jovens, sendo assim, a
morte das plantas ocorre de forma muito lenta e os primeiros sintomas aparecem
nos tecidos meristemáticos (ZHOU et al., 2007). Os tecidos maduros possuem
maiores reservas de proteínas que podem ser catabolizadas para aminoácidos,
sendo comum demorar mais tempo para o efeito fitotóxico destes herbicidas serem
observados nestes tecidos (SHANER; SINGH, 1993). Entre os sintomas que
demonstram a injúria sofrida por estas plantas aos 30 DAA destaca-se a elevada
dissipação de energia na forma de calor, que é resultado da falta de capacidade da
planta em utilizar para o transporte de elétrons a energia absorvida pelos centros de
reação, que pode ser explicada pela grande redução da capacidade de realizar
fotossíntese destas plantas, o que é demonstrado pelos índices de performance
obtidos.
O conhecimento sobre as características das plantas daninhas e a
documentação das respostas que estas plantas apresentam após a aplicação dos
herbicidas é de grande relevância para se evitar o surgimento de novos biótipos
resistentes (CHRISTOFFOLETI; LÓPEZ-OVEREJO, 2004). Mundialmente, têm sido
desenvolvidos diversos métodos para diagnosticar a resistência em várias espécies
daninhas (SIMINSZKY et al., 2005). Neste trabalho foram utilizadas sementes
coletadas de plantas escapes de áreas onde foi utilizado o Sistema de Produção
Clearfield® no Rio Grande do Sul, devido a isto este ecótipo levantou a suspeita de
ser tolerante aos herbicidas do grupo químico das imidazolinonas. O presente

39 trabalho comprovou a tolerância deste ecótipo até a dose comercial do herbicida
Only®, não ocorrendo alterações significativas nos parâmetros avaliados. Porém,
para doses superiores a comercial, as injúrias ao metabolismo fotossintético destas
plantas podem resultar em prejuízo ao seu crescimento e desenvolvimento,
entretanto estas injúrias são inferiores as encontradas para o ecótipo sensível
quando submetido à mesma dose do herbicida. No entanto, há mudança
conformacional na estrutura da enzima ALS, que aumenta a ligação do cofator
tiamina difosfato e, consequentemente, incrementa a atividade da enzima,
contribuindo assim para redução da sua sensibilidade ao herbicida imazethapyr
(SALES et al., 2008).
O herbicida Kifix® foi lançado para uso comercial na safra 2008/2009,
surgindo como alternativa de herbicida para o Sistema de Produção Clearfield®,
possuindo em sua formulação as moléculas de imazapyr+imazapic. O
comportamento das plantas de arroz cv. Puitá Inta CL demonstrou a sua tolerância
ao herbicida Kifix®, inclusive para até duas vezes a dose recomendada. Esta cultivar
apresentou um incremento das performances das reações de oxi-redução do FSI
quando exposta a este herbicida, porém este incremento pode não significar uma
maior produção de NADPH, pois os elétrons que chegam a ferredoxina podem ser
destinados a rotas alternativas dependendo da necessidade do vegetal, por
exemplo, nas assimilações de nitrogênio inorgânico e enxofre, mas também na
regulação do ciclo de assimilação de CO2 (FUKUYAMA, 2004).
A elevação dos níveis deste parâmetro pode ser atribuída a necessidade do
vegetal em metabolizar as moléculas do herbicida, podendo resultar numa
aceleração do metabolismo fotossintético destas plantas, fato este que também
pode ser observado pelo aumento do fluxo de elétrons capazes de reduzir os
aceptores finais de elétrons do FSI. A injúria inicial causada pelo herbicida Kifix® ao
ecótipo de arroz-vermelho sensível foi similar para as duas doses testadas, porém
na segunda avaliação foi observado que as plantas foram capazes de tolerar a dose
comercial deste herbicida, o que pode resultar em plantas escape nas lavouras de
arroz irrigado, criando assim uma grande possibilidade do surgimento de plantas de
arroz-vermelho resistentes a este herbicida (VILLA et al.,2006). Contudo, ao expor
este ecótipo ao dobro da dose do herbicida, as injúrias ao seu aparato fotossintético
foram irreversíveis, resultando praticamente em uma inatividade dos centros de
reação e das reações da cadeia de transporte de elétrons da fotossíntese, gerando

40 assim uma grande dissipação de calor. Além disso, ocorreu aumento do tamanho
aparente do sistema antena, provavelmente, devido à necessidade das plantas em
aumentar a sua capacidade de absorver energia, pois estas estavam dissipando
grande parte da sua energia de excitação. Foi observado também, que embora
estas plantas tenham reduzido drasticamente a vitalidade do seu aparato
fotossintético, ocorreu um aumento dos níveis das reações de oxi-redução do FSI, o
que pode ser explicado pelo fluxo de elétrons pseudocíclico que atuam em certas
condições fisiológicas quando o O2 recebe os elétrons oriundos da ferredoxina no
lugar do NADP+, através de uma reação não-enzimática (FRIDLYAND; SCHEIBE,
1999). Esta reação é conhecida como reação de Mehler e pode ter importante papel
na proteção dos vegetais sob condições de estresse (HAUPT-HERTING; FOCK,
2002). O resultado desta reação é a formação de espécies reativas de oxigênio,
que podem ser geradas com a destruição do sistema de transporte de elétrons
durante condições de estresse (SOARES; MACHADO, 2007), e que em grande
quantidade no vegetal podem ocasionar a peroxidação lipídica, vazamento do
conteúdo celular, necrose dos tecidos e a morte do vegetal (YEONG-JENE et al.,
2008).
Ao se considerar os dois herbicidas, é possível sugerir que a cv. Puitá Inta
CL de arroz tolera doses mais elevadas do herbicida Kifix®, em comparação ao
herbicida Only®, porém quando submetida às doses comerciais dos dois herbicidas,
esta cultivar demonstrou a capacidade de se recuperar das injúrias iniciais ao seu
metabolismo fotossintético, sem prejudicar o seu desenvolvimento. O ecótipo de
arroz-vermelho sensível demonstrou uma maior tolerância ao herbicida Kifix®,
quando considerada apenas a dose comercial. Em doses superiores a comercial os
danos ao aparato fotossintético das plantas deste ecótipo foram maiores quando
expostas ao herbicida Kifix®. Nas avaliações do ecótipo de arroz-vermelho com
suspeita de tolerância aos herbicidas do grupo químico das imidazolinonas, foi
comprovada a tolerância deste ecótipo para ambos os herbicidas aplicados na dose
comercial, sendo esta tolerância mantida para doses mais elevadas do herbicida
Kifix®.

41 4. CONCLUSÕES
a) A aplicação de herbicidas do grupo químico das imidazolinonas em plantas de
arroz causa alterações no processo fotossintético das plantas detectadas por
meio da emissão da fluorescência da clorofila a transiente;
b) Os parâmetros que melhor indicam a sensibilidade das plantas a herbicidas
do grupo químico da imidazolinonas são os índices de performance (PIABS,total
e PIABS) e a razão de dissipação total de energia de excitação não capturada
do total de centros de reação (DI0/RC);
c) A avaliação da emissão da fluorescência da clorofila a transiente é um
método que pode ser utilizado na detecção a campo de resistência e/ou
tolerância de plantas de arroz-vermelho a herbicidas do grupo químico das
imidazolinonas;

42
CAPÍTULO 3
CRESCIMENTO DE ESPÉCIES BIOINDICADORAS DO RESIDUAL DO HERBICIDA ONLY®, SEMEADAS EM ROTAÇÃO COM ARROZ CLEARFIELD®
Growth of the Residual Herbicide Only® Bioindicators Sown in Rotation with
Clearfield® Rice
RESUMO - Foi avaliado, por meio de bioensaio, o efeito residual do herbicida Only® (imazethapyr+imazapic) sobre as plantas de milho, pepino, rabanete e tomate, semeadas em solo, no qual foi aplicado há 1100 dias o herbicida. O estudo foi realizado em casa de vegetação no município de Capão do Leão/RS. O delineamento experimental foi o de casualização por bloco, totalizando quatro repetições por tratamento. Os tratamentos avaliados foram o residual do herbicida Only® aplicado sob as plantas de arroz CL na safra 2006/2007, nas doses de doses de 0, 10, 15 e 20mg p. c. m-2 do produto comercial, acrescido de 0,5% do adjuvante Dash®. Foram semeadas sob as parcelas 15 sementes de cada espécie bioindicadora, sendo estas desbastadas para 10 plantas após a germinação. As parcelas foram mantidas com irrigação conforme a necessidade das plantas. Após 60 dias da data da semeadura, foram avaliadas as variáveis: altura de plantas, massa seca da parte aérea e massa seca das raízes, sendo esta última não realizada para as plantas de milho. Os dados gerados foram submetidos à análise da variância (p≤0,05); em sendo significativos, estes foram testados por modelos de regressão polinomial. Conclui-se que há presença de atividade residual da mistura comercial dos herbicidas (imazethapyr+imazapic) do grupo das imidazolinonas em solo após 1100 dias da aplicação dos herbicidas. Além disso, as plantas de milho, pepino, rabanete e tomate podem ser utilizadas como espécies bioindicadoras de atividade residual da mistura comercial dos herbicidas (imazethapyr+imazapic), de forma que o rabanete e o tomate são mais sensíveis a presença do produto quando comparados ao milho e ao pepino. Palavras-chave: Only®, persistência, residual. ABSTRACT - This research determined, by bioassay, the residual effect of herbicide Only® (imazethapyr+imazapic) in corn, cucumber, radish and tomato sowed 1100 days after application of herbicide. Research was conducted in a greenhouse, at Universidade Federal de Pelotas, Capão do Leão-RS. The experimental design was

43 by block randomization, resulting in four replicates. The treatments were the residual herbicide applied under Only® rice CL plants in the 2006/2007 at doses of doses of 0, 10, 15 and 20mg p. c. m-2 of the commercial product, plus 0.5% of adjuvant Dash®. Were sown in plots 15 seeds of each species bioindicators, which were thinned to 10 plants after germination. The plots were maintained as needed for irrigation of plants. After 60 days of date of sowing, were measured for variables plant height, aboveground dry biomass and root dry biomass, with the latter not being performed for the corn plants. Data were subjected to analysis of variance (p ≤ 0.05), if significant, they were tested by polynomial regression models. We concluded that there was residual activity of the herbicide Only® (imazethapyr+imazapic), in a greenhouse, for a period of 1100 DAA, negatively affecting plant height and aboveground dry biomass of this species. In addition, corn, cucumber, radish and tomato can be used as bioindicators of residual activity of the commercial mixture of herbicides (imazethapyr + imazapic). The radish and tomato being more sensitive to the presence of the product, when compared to corn and cucumber. Keywords: Only®, persistence, residue. 1. INTRODUÇÃO
Nas lavouras de arroz irrigado do Rio Grande do Sul, a principal planta
daninha encontrada é o arroz-vermelho que possui diferentes ecótipos distribuídos
pelo estado (NOLDIN et al., 2004) com características morfofisiológicas muito
similares com o arroz cultivado. Portanto, há grande dificuldade de controle desta
espécie daninha nas lavouras de arroz irrigado, o que levou a diversas pesquisas
que resultaram no desenvolvimento do Sistema de Produção Clearfield®. Este
sistema possibilita o controle químico do arroz-vermelho em lavouras de arroz
cultivado, por meio do uso de herbicidas do grupo químico das imidazolinonas em
cultivares de arroz tolerantes. No Brasil, o herbicida recomendado para esse sistema
é o Only®, composto pela mistura formulada dos herbicidas imazethapyr + imazapic,
nas concentrações de 7,5 e 2,5mg p. c. m-2, respectivamente (SOSBAI, 2010).
Dentre as principais características dos herbicidas desse grupo é a
prolongada atividade residual no solo (LOUX; REESE, 1993), cuja persistência
atinge até 540 dias após a sua aplicação (PINTO et al., 2009)
A persistência no solo é uma característica de alguns grupos químicos de
herbicidas, ou ainda de herbicidas específicos, podendo ser desejável ou não. Esta
persistência é positiva quando resulta em período suficiente para manter a cultura
livre da competição de plantas daninhas (GAZZIERO et al., 1997). Todavia, é
indesejável quando resulta em injúrias para culturas em sucessão ou rotação ou
mesmo podendo causar aumento de riscos de contaminação ambiental.

44
A utilização de espécies não-tolerantes pode ser comprometida caso o
intervalo entre a aplicação de imazethapyr e a semeadura da cultura em rotação não
seja observado (WILLIAMS et al., 2002). Nos EUA, onde o Sistema de Produção
Clearfield® foi desenvolvido, recomenda-se a utilização do herbicida imazethapyr por
dois anos consecutivos, deixando o solo em repouso por, no mínimo, um ano.
Apesar da orientação de estudos realizados no Brasil para que seja respeitada a
mesma recomendação dos EUA, muitos produtores acabam usando a tecnologia por
mais de dois anos consecutivos (AVILA et al., 2010). Além disso, deve-se considerar
a diversidade de fatores relacionados à velocidade de degradação de um herbicida,
o que dificulta estimar o tempo necessário para sua dissipação após a aplicação,
uma vez que, este passa a ter no ambiente um conjunto de condições e de variáveis
que podem influenciar sua decomposição.
Sabe-se que além da utilização do Sistema de Produção Clearfield®,
também vem sendo empregada como técnica eficiente de controle do arroz-
vermelho a rotação do arroz irrigado com sorgo, milho ou soja, pois essas espécies
requerem condições diferenciadas de manejo de água e solo e utilizam herbicidas
diferentes dos aplicados no arroz, fatores que favorecem a redução da dominância
de espécies concorrentes (GOMES; CHRISTOFFOLETI et al., 2008).
O objetivo deste trabalho foi determinar, por meio de bioensaio, o efeito
residual da mistura comercial dos herbicidas imazethapyr+imazapic nas plantas de
milho, pepino, rabanete e tomate semeadas 1100 dias após a aplicação (DAA) dos
herbicidas imazethapyr+imazapic.
2. MATERIAL E MÉTODOS O experimento foi realizado em casa de vegetação, pertencente à
FAEM/UFPel, localizada no município de Capão do Leão/RS. O delineamento
experimental foi o de casualização por bloco, totalizando quatro repetições por
tratamento. Os tratamentos avaliados foram o residual do herbicida Only®, aplicado a
campo, sob as plantas de arroz CL na safra 2006/2007, nas doses de doses de 0
(controle), 10, 15 e 20mg p. c. m-2 do produto comercial, acrescido de 0,5% do
adjuvante Dash®. Ao término desta safra o solo foi coletado do campo e
acondicionado nas parcelas, que receberam um esquema de sucessão/rotação de
culturas descrito por Pinto et al., 2009. Na safra 2009/2010 foram semeadas sob as
parcelas 15 sementes de cada espécie bioindicadora: milho (Zea mays), pepino

45 (Cucumis sativus), rabanete (Raphanus sativus) e tomate (Solanum lycopersicum), sendo
estas desbastadas para 10 plantas após a germinação. As parcelas são
representadas por caixas de polietileno (60cm de comprimento x 40cm de largura x
20cm de estatura), cada uma contendo 50 kg de solo do tipo Planossolo Háplico
Eutrófico solódico (EMBRAPA, 2006), sendo mantidas com irrigação conforme a
necessidade das plantas.
Foram determinadas a altura de plantas e a massa seca da parte aérea
(MSA) de todas as espécies e massa seca das raízes (MSR) das olerícolas. A altura
de plantas foi avaliada 60 dias após a semeadura das bioindicadoras, medindo-se a
distância entre o colo da planta até o ápice da folha mais jovem. Após, foi realizada
a separação da parte aérea e da raiz das plantas coletadas. Estas partes foram
acondicionadas em sacos de papel separadamente, sendo transferidas para uma
estufa de circulação forçada de ar em temperatura de 70±5ºC até atingir massa
constante.
Os dados gerados foram submetidos à análise da variância (p≤0,05), em
sendo significativos foram ajustados por modelos de regressão polinomial
(MACHADO; CONCEIÇÃO, 2007). A escolha dos modelos baseou-se na
significância estatística (teste F), no ajuste do coeficiente de determinação (R2) e no
significado biológico do modelo (ADATI et al., 2006).
3. RESULTADOS E DISCUSSÃO Os dados da análise obtidos para altura de plantas apresentaram diferença
significativa no efeito residual herbicida entre as doses de Only® (Fig. 1). Os
resultados relativos à altura de plantas (Fig. 1) encontram-se representados
equações lineares acompanhadas do seu respectivo coeficiente de determinação
demonstrando que os dados obtidos ajustam-se ao modelo linear utilizado. A altura
de plantas, comparativamente ao tratamento controle, sofreu redução significativa de
18, 38, 45, 50%, respectivamente para milho (Fig. 1A), pepino, rabanete e tomate
(Fig. 1B) na presença de residual da dose de 10mg p. c. m-2 de
(imazethapyr+imazapic), contudo para a dose de 20mg p. c. m-2 de
(imazethapyr+imazapic), as reduções nesta variável-resposta foram de 35, 77, 90,
100%, respectivamente para as mesmas espécies.
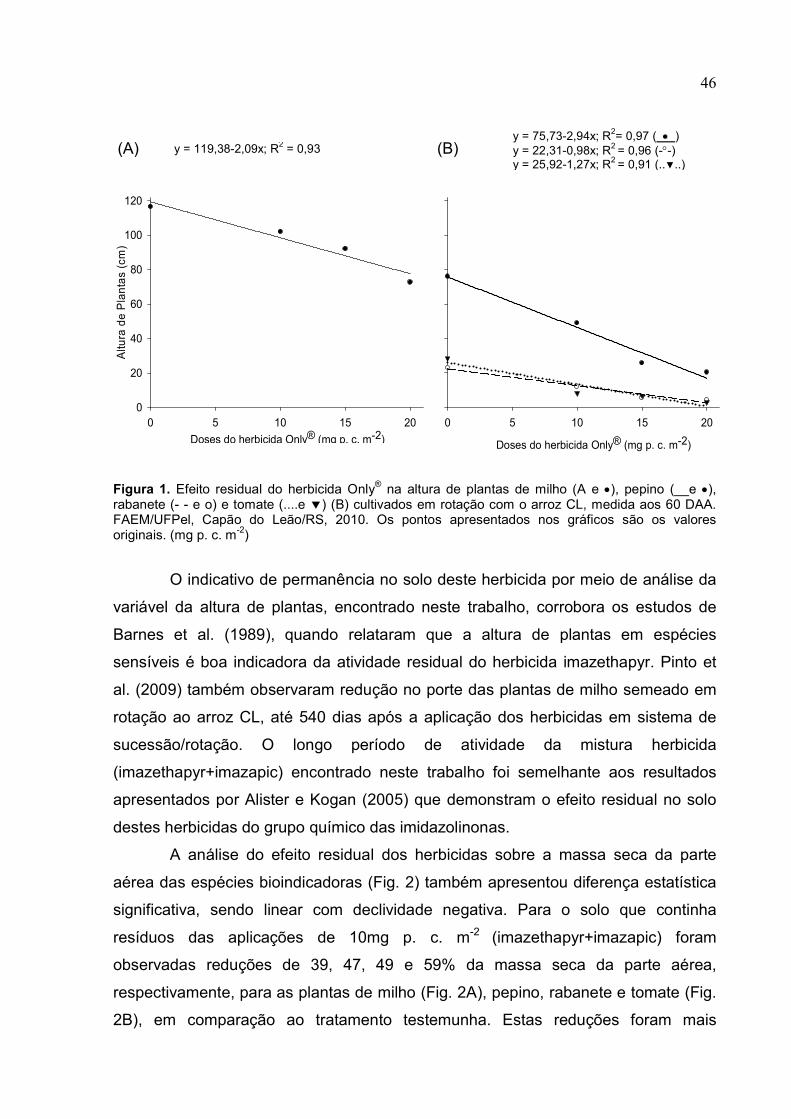
46
(A)
Doses do herbicida Only® (mg p. c. m-2)0 5 10 15 20
Altu
ra d
e Pl
anta
s (c
m)
0
20
40
60
80
100
120
(B)
Doses do herbicida Only® (mg p. c. m-2)
0 5 10 15 20
Figura 1. Efeito residual do herbicida Only® na altura de plantas de milho (A e •), pepino (__e •), rabanete (- - e o) e tomate (....e ) (B) cultivados em rotação com o arroz CL, medida aos 60 DAA. FAEM/UFPel, Capão do Leão/RS, 2010. Os pontos apresentados nos gráficos são os valores originais. (mg p. c. m-2)
O indicativo de permanência no solo deste herbicida por meio de análise da
variável da altura de plantas, encontrado neste trabalho, corrobora os estudos de
Barnes et al. (1989), quando relataram que a altura de plantas em espécies
sensíveis é boa indicadora da atividade residual do herbicida imazethapyr. Pinto et
al. (2009) também observaram redução no porte das plantas de milho semeado em
rotação ao arroz CL, até 540 dias após a aplicação dos herbicidas em sistema de
sucessão/rotação. O longo período de atividade da mistura herbicida
(imazethapyr+imazapic) encontrado neste trabalho foi semelhante aos resultados
apresentados por Alister e Kogan (2005) que demonstram o efeito residual no solo
destes herbicidas do grupo químico das imidazolinonas.
A análise do efeito residual dos herbicidas sobre a massa seca da parte
aérea das espécies bioindicadoras (Fig. 2) também apresentou diferença estatística
significativa, sendo linear com declividade negativa. Para o solo que continha
resíduos das aplicações de 10mg p. c. m-2 (imazethapyr+imazapic) foram
observadas reduções de 39, 47, 49 e 59% da massa seca da parte aérea,
respectivamente, para as plantas de milho (Fig. 2A), pepino, rabanete e tomate (Fig.
2B), em comparação ao tratamento testemunha. Estas reduções foram mais
y = 119,38-2,09x; R2 = 0,93 y = 75,73-2,94x; R2= 0,97 ( • ) y = 22,31-0,98x; R2 = 0,96 (-°-) y = 25,92-1,27x; R2 = 0,91 (..▼..)
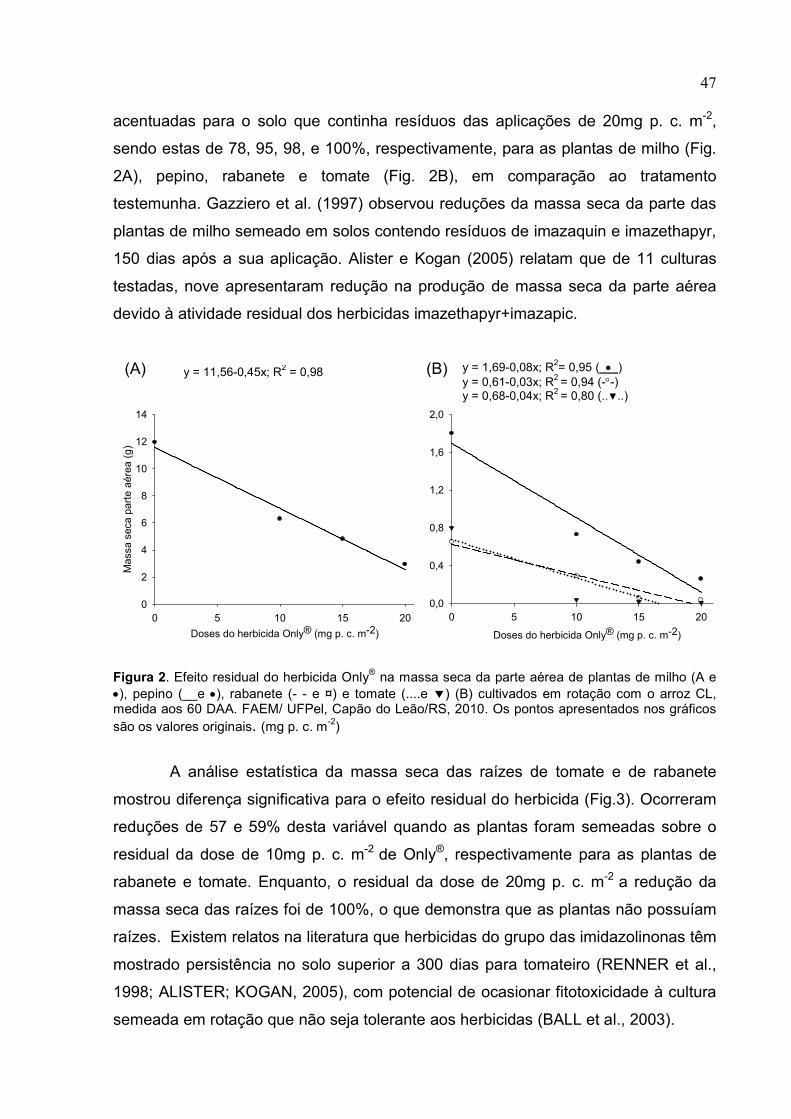
47 acentuadas para o solo que continha resíduos das aplicações de 20mg p. c. m-2,
sendo estas de 78, 95, 98, e 100%, respectivamente, para as plantas de milho (Fig.
2A), pepino, rabanete e tomate (Fig. 2B), em comparação ao tratamento
testemunha. Gazziero et al. (1997) observou reduções da massa seca da parte das
plantas de milho semeado em solos contendo resíduos de imazaquin e imazethapyr,
150 dias após a sua aplicação. Alister e Kogan (2005) relatam que de 11 culturas
testadas, nove apresentaram redução na produção de massa seca da parte aérea
devido à atividade residual dos herbicidas imazethapyr+imazapic.
(A)
Doses do herbicida Only® (mg p. c. m-2)0 5 10 15 20
Mas
sa s
eca
parte
aér
ea (g
)
0
2
4
6
8
10
12
14
(B)
Doses do herbicida Only® (mg p. c. m-2)
0 5 10 15 200,0
0,4
0,8
1,2
1,6
2,0
Figura 2. Efeito residual do herbicida Only® na massa seca da parte aérea de plantas de milho (A e •), pepino (__e •), rabanete (- - e ¤) e tomate (....e ) (B) cultivados em rotação com o arroz CL, medida aos 60 DAA. FAEM/ UFPel, Capão do Leão/RS, 2010. Os pontos apresentados nos gráficos são os valores originais. (mg p. c. m-2)
A análise estatística da massa seca das raízes de tomate e de rabanete
mostrou diferença significativa para o efeito residual do herbicida (Fig.3). Ocorreram
reduções de 57 e 59% desta variável quando as plantas foram semeadas sobre o
residual da dose de 10mg p. c. m-2 de Only®, respectivamente para as plantas de
rabanete e tomate. Enquanto, o residual da dose de 20mg p. c. m-2 a redução da
massa seca das raízes foi de 100%, o que demonstra que as plantas não possuíam
raízes. Existem relatos na literatura que herbicidas do grupo das imidazolinonas têm
mostrado persistência no solo superior a 300 dias para tomateiro (RENNER et al.,
1998; ALISTER; KOGAN, 2005), com potencial de ocasionar fitotoxicidade à cultura
semeada em rotação que não seja tolerante aos herbicidas (BALL et al., 2003).
y = 11,56-0,45x; R2 = 0,98 y = 1,69-0,08x; R2= 0,95 ( • ) y = 0,61-0,03x; R2 = 0,94 (-°-) y = 0,68-0,04x; R2 = 0,80 (..▼..)
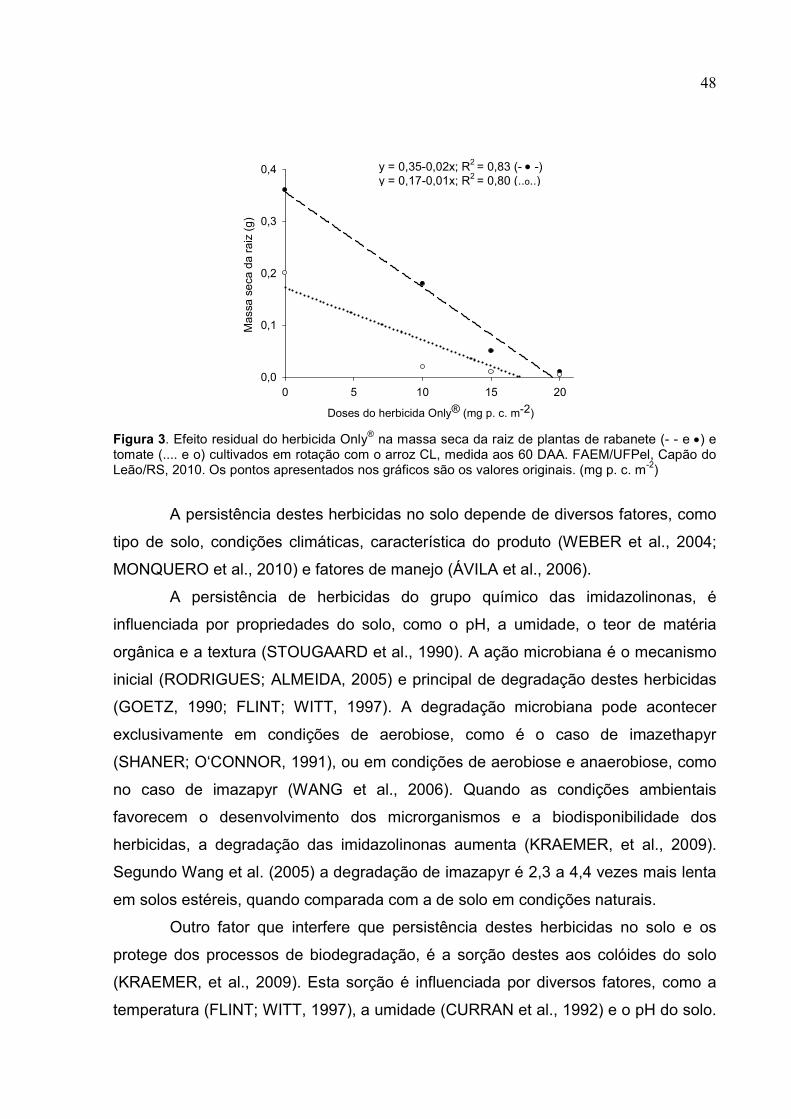
48
Doses do herbicida Only® (mg p. c. m-2)
0 5 10 15 20
Mas
sa s
eca
da ra
iz (g
)
0,0
0,1
0,2
0,3
0,4
Figura 3. Efeito residual do herbicida Only® na massa seca da raiz de plantas de rabanete (- - e •) e tomate (.... e o) cultivados em rotação com o arroz CL, medida aos 60 DAA. FAEM/UFPel, Capão do Leão/RS, 2010. Os pontos apresentados nos gráficos são os valores originais. (mg p. c. m-2)
A persistência destes herbicidas no solo depende de diversos fatores, como
tipo de solo, condições climáticas, característica do produto (WEBER et al., 2004;
MONQUERO et al., 2010) e fatores de manejo (ÁVILA et al., 2006).
A persistência de herbicidas do grupo químico das imidazolinonas, é
influenciada por propriedades do solo, como o pH, a umidade, o teor de matéria
orgânica e a textura (STOUGAARD et al., 1990). A ação microbiana é o mecanismo
inicial (RODRIGUES; ALMEIDA, 2005) e principal de degradação destes herbicidas
(GOETZ, 1990; FLINT; WITT, 1997). A degradação microbiana pode acontecer
exclusivamente em condições de aerobiose, como é o caso de imazethapyr
(SHANER; O‘CONNOR, 1991), ou em condições de aerobiose e anaerobiose, como
no caso de imazapyr (WANG et al., 2006). Quando as condições ambientais
favorecem o desenvolvimento dos microrganismos e a biodisponibilidade dos
herbicidas, a degradação das imidazolinonas aumenta (KRAEMER, et al., 2009).
Segundo Wang et al. (2005) a degradação de imazapyr é 2,3 a 4,4 vezes mais lenta
em solos estéreis, quando comparada com a de solo em condições naturais.
Outro fator que interfere que persistência destes herbicidas no solo e os
protege dos processos de biodegradação, é a sorção destes aos colóides do solo
(KRAEMER, et al., 2009). Esta sorção é influenciada por diversos fatores, como a
temperatura (FLINT; WITT, 1997), a umidade (CURRAN et al., 1992) e o pH do solo.
y = 0,35-0,02x; R2 = 0,83 (- • -) y = 0,17-0,01x; R2 = 0,80 (..o..)

49 Basham e Lavy (1987) e Baughman e Shaw (1996) demonstraram que imazethapyr
e imazaquin foram mais persistentes em solos frios e secos do que em solos
aquecidos e úmidos. Solos com boas condições de umidade promovem dissipação
mais rápida dos herbicidas imidazolinonas (LOUX; REESE, 1993). Sabe-se que para
solos de clima temperado, a sorção destes herbicidas está negativamente
correlacionada com o pH da solução do solo (RENNER et al., 1988; STOUGAARD et
al., 1990; LOUX; REESE, 1993). Isso é esperado, pois os herbicidas do grupo das
imidazolinonas que predominam na forma aniônica (-COO-) em pH de 7±2 são
repelidos pelas cargas negativas dos colóides minerais e orgânicos do solo
(MONQUERO et al., 2010). Aichele e Penner (2005) afirmaram que a dissipação de
imazaquin, imazethapyr e imazamox diminuiu quando o pH desceu de 7,0 para 5,0,
devido ao aumento na sorção com a conseqüente redução da biodisponibilidade.
Devido a estes fatores, há a recomendação de um intervalo de 300 dias
entre a aplicação de herbicidas imidazolinonas e a semeadura do milho em rotação
(RODRIGUES; ALMEIDA, 2005). Porém, este intervalo poderá ser superior a 1100
dias, dependendo das condições climáticas e do solo, como observado no presente
trabalho.
A longevidade do efeito residual destes herbicidas no solo é prolongada para
as maiores doses aplicadas (FLECK; VIDAL, 1994), corroborando com os resultados
encontrados neste trabalho, onde foi observado que o incremento das doses de
imazethapyr+imazapiC, reduziu drasticamente o crescimento das plantas de milho,
rabanete, pepino e tomate.
4. CONCLUSÕES
Há presença de atividade residual da mistura comercial dos herbicidas
(imazethapyr+imazapic) do grupo das imidazolinonas em solo após 1100 dias da
aplicação dos herbicidas, estabelecida pela redução de componentes de
crescimento de plantas bioindicadoras como milho, pepino, rabanete e tomate;
a) O tomate e o rabanete possuem maior sensibilidade ao produto quando
comparados ao pepino e ao milho;
b) O milho em rotação com a cultura do arroz que utiliza o Sistema de
Produção Clearfield® pode sofrer redução na altura e no acúmulo de massa
seca da parte aérea das plantas.

50
CONSIDERAÇÕES FINAIS
A utilização do Sistema de Produção Clearfield® de produção de arroz
proporciona um controle efetivo de ecótipos de arroz-vermelho sensíveis aos
herbicidas Only® e Kifix®, utilizados neste sistema. Contudo, é necessária uma
atenção especial ao surgimento de plantas escapes (não controladas pelos
herbicidas) desta planta daninha nas áreas cultivadas com estes herbicidas. Os
resultados deste trabalho verificaram a tolerância de plantas escapes de arroz-
vermelho a estes herbicidas mesmo em doses superiores a recomendada, o que
pode resultar no surgimento de biótipos de arroz-vermelho resistentes aos
herbicidas do grupo químico das imidazolinonas.
O conhecimento do funcionamento do aparato fotossintético das plantas de
arroz mediante a aplicação de herbicidas do grupo das imidazolinonas é uma
ferramenta que pode ser utilizada para verificar alterações no metabolismo destas
plantas mesmo antes destas serem visíveis. A detecção destas alterações é
facilmente realizada através da avaliação da emissão da fluorescência da clorofila a
transiente, sendo os índices de performance (PIABS,total e PIABS) e a razão de
dissipação total de energia de excitação não capturada do total de centros de reação
(DI0/RC), os parâmetros que melhor indicam a sensibilidade das plantas de arroz a
herbicidas do grupo químico da imidazolinonas. Esta técnica, por ser um método
rápido e não-invasivo, pode ser empregada em avaliações a campo de resistência
e/ou tolerância de plantas de arroz-vermelho aos herbicidas do grupo químico das
imidazolinonas.
Além disso, a escolha de culturas utilizadas em rotação com o arroz
cultivado pelo Sistema de Produção Clearfield® deve ser realizada com cuidado,
devido à persistência dos herbicidas do grupo das imidazolinonas no solo. A
presença de atividade residual da mistura comercial dos herbicidas
(imazethapyr+imazapic) em solo foi observada até 1100 dias após a aplicação dos

51 produtos. A identificação da atividade residual de herbicidas do grupo das
imidazolinonas pode ser facilmente realizada através da utilização de espécies
bioindicadoras como o milho, o pepino, o rabanete e o tomate, sendo as duas
últimas mais sensíveis a estes herbicidas.
O manejo da persistência destes herbicidas é um ponto fundamental para a
longevidade das lavouras de arroz que utilizam o Sistema de Produção Clearfield®.
Técnicas como a sucessão e/ou rotação de culturas, o parcelamento das doses
recomendadas dos herbicidas em pré e pós-emergência, a alternância de diferentes
mecanismos de ação herbicida, o adequado manejo do solo e da água e os
cuidados com doses excessivas de herbicidas, podem reduzir em grande parte os
prejuízos que a aplicação contínua deste sistema pode causar.
Um manejo eficaz do surgimento de resistência de plantas daninhas e da
persistência dos herbicidas imidazolinonas no solo contribui para o aumento das
produções das lavouras de arroz irrigado, e permiti que o Sistema de Produção
Clearfield® se mantenha como uma importante ferramenta para o controle do arroz-
vermelho em áreas infestadas.

52
REFERÊNCIAS
ADATI, C.; OLIVEIRA, V. A.; KARAM, D. Análise matemática e biológica dos modelos de estimativa de perdas de rendimento na cultura devido à interferência de plantas daninhas. Planta Daninha, v.24, p.1-12, 2006. AICHELE, T. M.; PENNER, D. Adsorption, desorption, and degradation of imidazolinones in soil. Weed Technology, v. 19, p. 154-159, 2005. ALISTER, C.; KOGAN, M. Efficacy of imidazolinone herbicides applied to imidazolinone-resistant maize and their carryover effect on rotational crops. Crop Protection, v. 24, p. 375-379, 2005. ARIAS, D. M.; RIESEBERG, L. H. Gene flow between cultivated and wild sunflowers. Theoretical and Applied Genetics, v. 89, p. 655–660, 1994. AVILA, L. A.; MASSEY, J. H.; SENSEMAN, S. A.; ARMBRUST, K. L.; LANCASTER, S. R.; McCAULEY, G. N.; CHANDLER, J. M. Imazethapyr aqueous photolysis, reaction quantum yield, and hydroxyl radical rate constant. Journal of Agricultural and Food Chemistry, v. 54, p. 2635-2639, 2006. AVILA, L. A.; TELÓ, G. M.; FERREIRA, R. B.; MARCHESAN, E.; MACHADO, S. L. O.; ROSSATO, T. L.; CEZIMBRA, D. M.; RIGÃO JR., G. Retorno da produção de arroz irrigado com cultivares convencionais após o uso do Sistema Clearfield®. Planta Daninha, v. 28, p. 123-129, 2010. BALL, D. A.; YENISH, J. P.; ALBY, T. Effect of imazamox soil persistence on dryland rotational crops. Weed Technology, v. 17, p. 161-165, 2003. BARBAGALLO, R. P. et al. Rapid, noninvasive screening for perturbations of metabolism and plant growth using chlorophyll fluorescence imaging. Plant Physiology, v. 132, p. 485-496, 2003. BARNES C. J.; GOETZ, A. J.; LAVY, T. L. Effects of imazaquin residues on cotton (Gossipium hirsitum). Weed Science, v. 37, p. 820-824, 1989. BASF. RS: Basf propõe valor único do Clearfield. Disponível em: http://www.paginarural.com.br/noticias_detalhes.php?id=107808. Acessado em: 21/05/2009. BASHAM, G. W.; LAVY, T. L. Microbial and photolytic dissipation of imazaquin in soil. Weed Science, v. 35, p. 865-870, 1987. BAUGHMAN, T. A.; SHAW, D. R. Effect of wetting/drying cycles on dissipation patterns of bioavailable imazaquin. Weed Science, v. 44, p. 380-382, 1996. BURGOS, N. R.; NORMAN, R. J.; GEALY, D. R.; BLACK, H. L. Competitive N uptake between rice and weedy rice. Field Crops Research, v. 99, p. 96-105, 2006

53 BURGOS, N. R.; NORSWORTHY, J. K.; SCOTT, R. C.; SMITH, K. L. Red rice status after five years of Clearfield® rice technology in Arkansas. Weed Technology, v. 22, p. 200–208, 2008. CHIPMAN, D. M.; DUGGLEBY, R. G.; TITTMANN, K. Mechanisms of acetohydroxyacid synthases. Current Opinion in Chemical Biology, v. 9, p. 475–481, 2005.
CHRISTOFFOLETI, P. J.; LOPÉZ OVEJERO, R. F. L. Definições e situação da resistência de plantas daninhas aos herbicidas no Brasil e no mundo. In: CHRISTOFFOLETI, P. J. Aspectos de resistência de plantas daninhas a herbicidas. Campinas: HRAC-BR, 2004. p. 3-22 CONAB. Arroz – Brasil. Série Histórica de: área, produtividade e produção. Disponível em: http://www.conab.gov.br. Acessado em: 21/05/2009. COUNCE, P. A.; KEISLING, T. C.; MITCHELL, A. J. A. Uniform, objective, and adaptative system for expressing rice development. Crop Science, v. 40, p. 436-443, 2000. CROUGHAN, T.P.; UTOMO, H, S.; SANDERS, D. E.; BRAVERMAN, M. P. Herbicide-resistant rice offers potential solution to red rice problem. Louisiana Agriculture, v. 39, p. 10-12, 1996. CURRAN, W. S.; LOUX, M. M.; LIEBEL, R. A.; SIMMONS, F. W. Photolysis of imidazolinone herbicides in aqueous solution and on soil. Weed Science, v. 40, p. 143-148, 1992. DAL MAGRO, T.; PINTO, J. J. O.; AGOSTINETTO, D.; GALON, L.; REZENDE, A. L.; LAZAROTTO, C. A. Suscetibilidade de cultivares de arroz irrigado (Oryza sativa) à deriva simulada do herbicida imazethapyr + imazapic. Planta Daninha, v. 24, p. 751-759, 2006. DEVINE, M. D.; PRESTON, C. The molecular basis of herbicide resistance. Pages 72–104, In: COBB, A. H.; KIRKWOOD, R. C. eds. Herbicides and Their Mechanisms of Action. Sheffield, Great Britain: Sheffield Academic, 2000. EHLERS, G. A. C.; LOIBNER, A. P. Linking organic pollutant (bio) availability with geosorbent properties and biomimetic methodology: A review of geosorbent characterisation and (bio) availability prediction. Environmental Pollution, v. 141, p. 494-512, 2006. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA [EMBRAPA], Centro Nacional de Pesquisa de Solos. Sistema Brasileiro de Classificação de Solos. Rio de Janeiro: EMBRAPA Solos, 2006. 306p. ESTORNINOS, L. E.; GEALY, D. R.; GBUR, E. E.; TALBERT, R. E.; MCCLELLAND, M. R. Rice and red rice interference. II. Rice response to population densities of three red rice (Oryza sativa) ecotypes. Weed Science, v. 53, p. 683–689, 2005.

54 FLECK, N.G., VIDAL, R.A. Injúria potencial de herbicidas de solo ao girassol. III, Imazaquin e imazethapyr. Planta Daninha, v.12, p.39-43, 1994. FLINT, J. L.; WITT, W. W. Microbial degradation on imazaquin and imazethapyr. Weed Science, v. 45, p. 586-591, 1997. FONTANA, L. C.; AGOSTINETTO, D.; PINTO, J. J. O.; ROSENTHAL, M. D.; RIGOLI, R. P.; FIGUEREDO, S.S. Controle de arroz-vermelho (Oryza sp.) com o herbicida nicosulfuron ou a mistura formulada de imazethapyr+imazapic. Planta Daninha, v. 25, p. 783-790, 2007. FRANKART, C.; EULLAFFROY, P.; VERNET, G. Comparative effects of four herbicides on non-photochemical fluorescence quenching in Lemna minor. Environmental and Experimental Botany, v. 49, p. 159-168, 2003. FRANS, R.; CROWLEY, H. Experimental design and techniques for measuring and analyzing plant responses to weed control practices. In: SOUTHERN WEED SCIENCE SOCIETY. Research methods in weed science. 3.ed. Clemson: p. 29-45, 1986. FRIDLYAND, L.E.; SCHEIBE, R. Controlled distribution of electrons between acceptors in chloroplasts: a theoretical consideration. Biochimica et Biophysica Acta, v. 1413, p. 31-42, 1999. FUKUYAMA, K. Structure and function of plant-type ferredoxins. Photosyntesis Research, v. 81, p. 289–301, 2004. GASTON, S.; RIBAS-CARBO, M.; BUSQUETS, S.; BERRY, J. A.; ZABALZA, A.; ROYUELA, M. Changes in mitochondrial electron partitioning in response to herbicides inhibiting branched-chain amino acid biosynthesis in soybean. Plant Physiology, v. 133, p. 1351–1359, 2003. GAZZIERO, D. L. P.; KARAN, D.; VOLL, E.; ULBRICH, A. Persistência dos herbicidas imazaquin e imazethapyr no solo e os efeitos sobre plantas de milho e pepino. Planta Daninha, v. 15, p. 162-169, 1997. GOETZ, A. J.; LAVY, T. L.; GBUR JR, E. E. Degradation and field persistence of Imazethapyr. Weed Science, v. 38, p. 421-428, 1990. GOMES JR., F. G.; CHRISTOFFOLETI, P. J. Biologia e manejo de plantas daninhas em áreas de plantio direto. Planta Daninha, v. 26, p. 789-798, 2008. HAUPT-HERTING, S.; FOCK, H.P. Oxygen exchange in relation to carbon assimilation in water-stressed leaves during photosynthesis. Annals of Botany, v. 89, p. 851–859, 2002. HEAP, I. International Survey of Herbicide Resistant Weeds. http://www.weedscience.com. Acessado: 21/05/2009.

55 HERBICIDE RESISTANCE ACTION COMMITTEE [HRAC]. Classification of Herbicides According to Mode of Action. Disponível em: www.plantprotection.org/hrac/. Acessado em: 15/06/2010. INSTITUTO RIOGRANDENSE DO ARROZ [IRGA]. Arroz irrigado no RS — área, produção e rendimento. Disponível em: http://www.irga.rs.gov.br. Acessado em: 21/05/2009. JENKINS, S.R.; WEHTJE, G. R.; MORGAN, J.M.; BOLLINGER, A. F.; YOUNG, D. G. Temperature effects on retention of atrazine and imazapyr soils. Water Air Soil Pollution, v.118, p.169-178, 2000. KOSKINEN, W. C.; HARPER, S. S. The retention process: mechanisms. In: CHENG, LEON, V. W. J.; CARL, R. F. A bioassay technique detects imazethapyr leaching and liming-dependant activity. Weed Technology, v. 15, p. 1-6, 2001. KRAEMER, A. F.; MARCHESAN, E.; AVILA, L. A.; MACHADO, S. L. O.; GROHS, M. Destino ambiental dos herbicidas do grupo das imidazolinonas. Planta Daninha, v. 27, p. 629-639, 2009. KRAUSE, G. H.; WEIS, E. Chlorophyll fluorescence as a tool in plant physiology. II. Interpretation of fluorescence signals. Photosynthesis Research, v. 5, p. 139-157, 1984. LAZÁR, D. The polyphasic chlorophyll a fluorescence rise measured under high intensity of exciting light. Functional Plant Biology, v. 33, p. 9–30, 2006. LE, D. T.; YOON, M. Y.; KIM, Y. T.; CHOI, J. D. Roles of three wellconserved arginine residues in mediating the catalytic activity of tobacco acetohydroxy acid synthase. Journal of Biochemistry, v. 138, p. 35–40, 2005. LICHTENTHALER, H. K. The Kautsky effect: 60 years of chlorophyll fluorescence induction kinetics. Photosynthetica, v. 27, p. 45-55, 1992. LOPES, M. C. B.; ROSSO, A. F.; LOPES, S. I. G.; CARMONA, P. S.; LEITES, A.; ULBRICH, A.; LOUZANO, L. C. IRGA 422CL: A nova cultivar desenvolvida pelo programa de melhoramento genético do Instituto Rio Grandense do Arroz para o Sistema de Produção Clearfield®. In: CONGRESSO BRASILEIRO DE ARROZ IRRIGADO, 3.; REUNIÃO DA CULTURA DO ARROZ IRRIGADO, 25., 2003, Balneário Camboriú. Anais... Itajaí: EPAGRI, 2003. p. 3-5. LOUX, M. M.; REESE, K. D. Effect of soil type and pH on persistence and carryover of imidazolinone herbicides. Weed Technology, v. 7, p. 452-458, 1993. MACHADO, A. A.; CONCEIÇÃO, A. R. WinStat – Sistema de Análise Estatística para Windows versão 1.0. Universidade Federal de Pelotas, 2007. MAXWELL, K.; JOHNSON, G. N. Chlorophyll fluorescence: a practical guide. Journal of Experimental Botany, v. 51, p. 659-668, 2000.

56 MENEZES, V. G.; MARIOT, C. H. P.; KALSING, A.; GOULART, I. C. G. R. Arroz-vermelho (Oryza sativa) resistente aos herbicidas imidazolinonas. Planta Daninha, v. 27, p. 1047-1052, 2009. MENEZES, V. G., AGOSTINETTO, D., FLECK, N. G.; SILVEIRA, C. A. Caracterização de biótipos de arroz-vermelho em lavouras de arroz do estado do Rio Grande do Sul. Planta Daninha, v. 20, p. 221-227, 2002. MONQUERO, P. A.; CHRISTOFFOLETI, P. J. Manejo de populações de plantas daninhas resistentes aos herbicidas inibidores da acetolactato sintase. Planta Daninha, v. 19, p. 67-74, 2001. MONTEALEGRE, F.; VARGAS, J. P. Efecto de algunas practicas culturales sobre la población de arroz rojo y los rendimientos del arroz comercial. Arroz, v. 38, n. 359, p. 19-14, 1989. NOLDIN, J. A.; YOKOYAMA, S.; STUKER, H.; RAMPELOTTI, F. T.; GONÇALVES, M. I. F.; EBERHARDT, D. S.; ABREU, A.; ANTUNES, P.; VIEIRA, J. Desempenho de populações híbridas F2 de arroz-vermelho (Oryza sativa) com arroz trangênico (O. sativa) resistente ao herbicida amônio-glufosinate. Planta Daninha, v. 22, p. 381-395, 2004. PELLERIN, K. J.; WEBSTER, E. P. Imazethapyr at different rates and timings in drill- and water-seeded imidazolinone-tolerant rice. Weed Technology, v. 18, p. 223- 227, 2004. PINTO, J. O.; NOLDIN J. A.; MACHADO, A.; PINHO, C. F.; ROSENTHAL, M. D.; DONIDA, A.; GALON, L.; DURIGAN, M. Milho (Zea mays) como espécies bioindicadora da atividade residual de (imazethapyr + imazapic). Planta Daninha, v.27, p. 1005-1014, 2009. RAJGURU, S. N.; BURGOS, N. R.; SHIVRAIN, V. K.; STEWART, J. M. Mutations in the red rice ALS gene associated with resistance to imazethapyr. Weed Science, v. 53, p. 567–577, 2005. RENNER, K. A.; MEGGITT, W. F.; PENNER, D. Effect of soil pH on imazaquin and imazethapyr adsorption to soil and phytotoxicity to corn (Zea mays). Weed Science, v. 36, p. 78-83, 1988. RENNER, K. A. et al. Effect of tillage application method on corn (Zea mays) response to imidazolinone residues in soil. Weed Technology, v. 12, p. 281-285, 1998. RODRIGUES, B. N.; ALMEIDA, F. S. Guia de herbicidas. 5.ed. Londrina: Edição dos Autores, 2005. 591 p. ROSO, A. C.; MEROTTO JR., A.; DELATORRE, C. A. Bioensaios para diagnóstico da resistência aos herbicidas imidazolinonas em arroz. Planta Daninha, v. 28, p. 411-419, 2010.

57 SALES, M. A.; SHIVRAIN, V. K.; BURGOS, N. R.; KUK, Y. I. Amino acid substitutions in the acetolactate synthase gene of red rice (Oryza sativa) confer resistance to imazethapyr. Weed Science, v. 56, p. 485–489, 2008. SCHREIBER, U.; KUHL, M.; KLIMANT, I.; REISING, H. Measurement of chlorophyll fluorescence within leaves using a modified PAM fluorometer with a fiber-optic microprobe. Photosynthesis Research, v. 47, p. 103-109, 1996. SCHWANKE, A. M. L., NOLDIN, J. A., ANDRES, A., PROCÓPIO, S. O.; CONCENÇO, G. Caracterização morfológica de ecótipos de arroz daninho (Oryza sativa) provenientes de áreas de arroz irrigado. Planta Daninha, v. 26, p. 249-260, 2008. SENSEMANN, S. A. Herbicide Handbook. 9.ed. Lawrence: WSSA, 2007. 458p. SHANER, D. L.; O‘CONNOR, S. The imidazolinones herbicides. Boca Raton: CRC Press, 1991. 289p SHANER, D. L.; SINGH, B. K. Phytotoxicity of acetohydroxyacid synthase inhibitors is not due to accumulation of 2-ketobutyrate and/or 2-aminobutyrate, Plant Physiology, v. 103, p. 1221–1226, 1993. SHIVRAIN, V. K.; BURGOS, N. R., GEALY, D. R.; SMITH, K. L.; SCOTT, R. C.; MAUROMOUSTAKOS, A.; BLACK, H. Red Rice (Oryza sativa) Emergence Characteristics and Influence on Rice Yield at Different Planting Dates. Weed Science, v. 57, p. 94–102, 2009. SILVA, A. A.; OLIVEIRA JR., R. S.; COSTA, E. R.; FERREIRA, L. R. Efeito residual no solo dos herbicidas imazamox e imazethapyr para as culturas de milho e sorgo. Planta Daninha, v. 17, p. 345-354, 1999. SILVA, C. M. M.; GOMES, M. M. A.; FREITAS, S. P. Interferência de herbicidas, associados a um análogo de brassinosteróide, no aparato fotossintético de mudas de Eucalyptus grandis. Planta Daninha, v. 27, p. 789-797, 2009. SIMINSZKY, B.; COLEMAN, N. P.; NAVEED, M. Denaturing high-performance liquid chromatography efficiently detects mutations of the acetolactate synthase gene. Weed Science, v. 53, p. 146-152, 2005. SOARES, A. M. S.; MACHADO, O. L. T. Defesa de plantas: Sinalização química e espécies reativas de oxigênio. Revista Trópica, v. 1, p. 9-19, 2007SANTOS, F. M.; MARCHESAN, E.; MACHADO, S. L. O.; VILLA, S. C. C.; AVILA, L. A.; MASSONI, P. F. S. Controle químico de arroz-vermelho na cultura do arroz irrigado. Planta Daninha, v. 25, p. 405-412, 2007. SOCIEDADE SUL BRASILEIRA DE ARROZ IRRIGADO [SOSBAI]. Arroz irrigado: recomendações Técnicas da Pesquisa para o Sul do Brasil. Porto Alegre: Sosbai, 2010, 188p.

58 SOUSA, R. O.; PERALBA, M. C. R.; MEURER, E. J. Short caain organic, acid dynamics in solucion of flooded soil treated wth riegrass residues. Communications in Soil Science and Plant Analysis, v. 33, p. 779-78, 2002. STEELE, G. L.; CHANDLER, J. M.; McCAULEY, G. N. Control of red rice (Oryza sativa) in imidazolinone-tolerant rice (Oryza sativa). Weed Technology, v. 16, p. 627-630, 2002. STOUGAARD, R. N.; SHEA, P. J.; MARTIN, A. R. Effect of soil type and pH on adsorption, mobility, and efficacy of imazaquin and imazethapyr. Weed Science, v. 38, p. 67-73, 1990. STRASSER, B.J.; STRASSER, R.J. Measuring fast fluorescence transients to address environmental question: the JIP test. In: MATHIS, P. (Ed.), Photosynthesis: From Light to Biosphere, vol. V. Kluwer Academic Publisher, The Netherlands, p. 977–980, 1995. STRASSER, R.J.; SRIVASTAVA, A.; TSIMILLI-MICHAEL, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In: YUNUS, M.; PATHRE, U.; MOHANTY, P. (Eds.), Probing Photosynthesis: Mechanisms, Regulation and Adaptation. Taylor and Francis, London, UK, p. 445–483, 2000. STRASSER, R.J.; TSIMILLI-MICHAEL, M.; SRIVASTAVA, A. Analysis of the Chlorophyll a fluorescence transient. In: Papageorgiou, C., Govindjee, (Eds.), Chlorophyll Fluorescence: A Signature of Photosynthesis. Springer, Netherlands, p. 321–362, 2004. TAN, S.; EVANS, R. R.; SINGH, B. K. Herbicidal inhibitors of amino acid biosynthesis and herbicide-tolerant crops. Amino Acids, v. 30, p. 195-204, 2006. TAN, S.; EVANS, R. R.; DAHMER, M. L.; SINGH, B. K.; SHANER, D. L. Imidazolinone-tolerant crops: history, current status and future. Pest Management Science, v. 61, p. 246-257, 2005. TSSIMILLI-MICHAEL, M; STRASSER, R.J. In vivo assessment of plants vitality: applications in detecting and evaluating the impact of Mycorrhization on host plants. In: VARMA, A. (Ed.), Mycorrhiza: State of the Art, Genetics and Molecular Biology, Eco-Function, Biotechnology, Eco-Physiology, Structure and Systematics, 3ed, Springer, Dordrecht, 2008, pp. 679–703. TRACY M. A.; PENNER D. Adsorption, desorption, and degradation of Imidazolinones in Soil. Weed Technology, v. 19, p. 154-159. 2005. TRANEL, P. J.; WRIGHT, T. R. Resistance of weeds to ALS-inhibiting herbicides: what have we learned? Weed Science, v. 50, p. 700–712, 2002. VAUGHAN, L. K.; OTTIS, B. V; SNELLER, C.; CHANDLER, J. M.; PARK, W. D. Is all red rice found in commercial rice really Oryza sativa? Weed Science, v. 49, p. 468-476, 2001.

59 VIDAL, R. A.; WINKLER, L. M. Resistência de plantas daninhas: seleção ou indução a mutação pelos herbicidas inibidores da acetolactato sintase (ALS). Pesticidas: Revista de Ecotoxicologia e Meio Ambiente, v. 12, p. 31-42, 2002. VILLA, S. C. C., MARCHEZAN, E.; AVILA, L. A.;MASSONI, P. F. S.; TELO, G. M.; MACHADO, S. L. O.; CAMARGO, E. R. Arroz tolerante a imidazolinonas: controle do arroz vermelho, fluxo gênico e efeito residual do herbicida em culturas sucessoras não tolerantes. Planta Daninha, v. 24, p. 761-768, 2006. WANG, X.; WANG, H.; FAN, D. Degradation and metabolism of imazapyr in soils under aerobic and anaerobic conditions. International Journal of Environmental Analytical Chemistry, v. 86, p. 541-551, 2006. WEBER, B. J.; WILKERSON, G. G.; REINHARDT, C. F. Calculating pesticide sorption coefficients (Kd) using selected soil properties. Chemosphere, v. 55, p. 157-166, 2004. WEBSTER, E. P.; MASSON, J. A. Acetolactate synthaseinhibiting herbicides on imidazolinone-tolerant rice. Weed Science , v. 49, p. 652-657, 2001. WILLIAMS, B. J.; STRAHAN, R.; WEBSTER, E. P. Weed management systems for Clearfield Rice. Louisiana Agriculture, v. 45, p. 16-17, 2002. WRIGHT, T. R.; BASCOMB, N. F.; STURNER, S. F.; PENNER, D. Biochemical mechanism and molecular basis of ALS-inhibiting herbicide resistance in sugarbeet (Beta vulgaris) somatic cell selections. Weed Science, v. 46, p. 13–23, 1998. XIA, X. J.; HUANG, Y. Y.; WANG, L.;HUANG, L. F.; YU, Y.L.; ZHOU,Y.H.; YU, J. K. Pesticides-induced depression of photosynthesis was alleviated by 24-epibrassinolide pretreatment in Cucumis sativus L. Pesticide Biochemistry and Physiology, v. 86, p. 42-48, 2006. YEONG-JENE, C.; YI-XUAN, W.; MOU-YEN, C.; CHING-YUH, W. Role of Antioxidative System in Paraquat Resistance of Tall Fleabane (Conyza sumatrensis). Weed Science, v. 56, p. 350–355, 2008. YUSUF, M.A.; KUMAR, D.; RAJWANSHI, R.; STRASSER, R.J.; TSIMILLI-MICHAEL, M.; GOVINDJEE; SARIN, N.B. Overexpression of γ-tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurements. Biochimica et Biophysica Acta, v. 1797, p. 1428-1438, 2010.
ZHOU, Q.; LIU, W.; ZHANG, Y.; LIU, K. K. Action mechanisms of acetolactate synthase-inhibiting herbicides. Pesticide Biochemistry and Physiology, v. 89, p. 89–96, 2007.


















