
DESVENDANDO AS CÉLULAS-TRONCO: DOS SONHOS À...
130
DESVENDANDO AS CÉLULAS-TRONCO: DOS SONHOS À REALIDADE 16 a 20 de julho de 2007 Centro de Estudos do Genoma Humano Depto. de Genética e Biologia Evolutiva Instituto de Biociências Universidade de São Paulo Docentes responsáveis: Profa. Dra. Eliana Maria Beluzzo Dessen Profa. Dra. Regina Célia Mingroni-Netto
Transcript of DESVENDANDO AS CÉLULAS-TRONCO: DOS SONHOS À...

DESVENDANDO AS CÉLULAS-TRONCO: DOS SONHOS À REALIDADE
16 a 20 de julho de 2007
Centro de Estudos do Genoma Humano Depto. de Genética e Biologia Evolutiva
Instituto de Biociências Universidade de São Paulo
Docentes responsáveis: Profa. Dra. Eliana Maria Beluzzo Dessen Profa. Dra. Regina Célia Mingroni-Netto

2
INDICE Cronograma 03 Apresentação do curso 04 Portifólio como recurso de avaliação 06 Textos de apoio
As células eucarióticas e sua capacidade de diferenciação 08 Diferenciação celular e o controle do gene eucariótico 10 Sinalização celular 18 O básico sobre células-tronco 24 Clonagem 31 Reprogamação celular 36 Células-tronco: progressos científicos e o futuro das pesquisas 39 Terapia celular – o uso de células-tronco no tratamento de doenças – etapas e questões geradas 42 A polêmica das células-tronco.embrionárias 43 Desvendando as raízes do câncer 48 Células-tronco: mocinha e bandida 57 Célula-tronco por encomenda 58 Reprodução assistida 59 Medula óssea e as células-tronco hematopoiéticas 61 Transplante de medula óssea – uma terapia celular bem conhecida 63 Colcha de retalho de leis 67 O artigo 5o
Quando começa a vida? 77 . Da lei No. 11.105, de 2005, não é inconstitucional 72
Que vida, biológica ou moral? 79 Glossário 81
Atividades: Atividade de acolhimento e contrato pedagógico 85 Levantamento de conceitos 87 Estudo dirigido 1 – Identificação de conceitos relativos a células-tronco 88 Atividade 1 – Diferenciação da hemácia 90 Atividade 2 – um caso muito interessante de diferenciação celular 92 Diferenciação no protozoário Naegleria grubei 94 Diferenciação celular em tecido ósseo 96 Atividade 3 – Diferenciação celular em tecido ósseo 98 Atividade 4 – Diferenciação de células embrionárias: a formação do
trofoblasto 99 As primeiras mudanças morfológicas no embrião 101 Atividade 5 – Desenvolvimento muscular 104 Estudo dirigido 2 – Clonagem reprodutiva versus clonagem terapêutica 107 Atividade 6 – Jogo das células-tronco 108 Atividade 7 – Hematopoiese: seguindo uma via de diferenciação 118 Atividade 8 – Regras para o debate 123 Atividade 9 – Palavras cruzadas 124
Anexos: Respostas do estudo dirigido 2 – Clonagem 126 Gabarito da atividade 2 – Diferenciação no protozoário N. gruberi 126
Solução das palavras cruzadas 128 Bibliografia sobre câncer e células-tronco 129

3
CRONOGRAMA
Data Manhã
Tarde
16/07
Acolhimento inicial.Apresentação do curso. Levantamento de conceitos. Estudo dirigido sobre células-tronco.
Diferenciação celular. Oficinas. O gene eucariótico e sua regulação. Estruturação do debate sobre ética.
17/07
Sinalização celular. Oficinas. Células-tronco: definição, tipos e características. Clonagem reprodutiva versus clonagem terapêutica. Desdiferenciação celular.
Diferenças entre células-tronco embrionárias e de adulto. Jogo sobre células-tronco. Palavras cruzadas. Preparação do debate
18/07
Câncer e células-tronco Reprodução assistida Preparação do debate.
As doenças do sangue e transplantes de medula óssea. Oficina. Preparação do debate
19/07
Perspectivas da aplicação de células-tronco em doenças musculares. Filme Globo News. Discussão.
Debate: A polêmica sobre a utilização de células-tronco embrionárias em terapia humana: aspectos legais e éticos. Discussão de situações problema.
20/07
Oficina de criação: Como abordar o tema células-tronco numa feira de ciências? Início da apresentação dos grupos.
Apresentação dos grupos. Avaliação do curso.

4
APRESENTAÇÃO DO CURSO Desvendando as células-tronco: dos sonhos à realidade
Objetivos • Ampliar o universo conceitual significativo dos professores de ensino médio no que
se refere a células-tronco e suas aplicações. • Capacitar o professor de ensino médio para acompanhar de maneira crítica a
literatura de divulgação científica sobre o tema. • Utilizar o tema "células-tronco” como eixo para a integração de conceitos clássicos
da Biologia Celular e Molecular, bem como ponto de referência para a análise e discussões no campo da bioética.
• Considerar o papel do professor em sala de aula como "Agente disseminador” de temas que permeiam a discussão de valores éticos e sociais e que exigem um posicionamento crítico acerca de situações relacionadas à área de Ciências da Natureza e suas tecnologias.
Conteúdo e Metodologia de desenvolvimento
• Noções de diferenciação e sinalização celular. Relação genótipo-fenótipo. • Células-tronco: definição, potencialidade e plasticidade. Reprogramação celular. • Células-tronco embrionárias versus células-tronco de adultos. Potencialidades de
uso e aplicações experimentais. • Terapia celular. Aplicações atuais. • Clonagem terapêutica versus clonagem reprodutiva. • Câncer e células-tronco. • Problemas legais e éticos decorrentes do uso de células-tronco.
Estratégias e recursos tecnológicos
A metodologia utilizada para desenvolver o conteúdo acima relacionado é variada, com ênfase em métodos não expositivos. Além de curtas apresentações orais dialogadas e de palestras de especialistas, diversas atividades didáticas/pedagógicas serão utilizadas como facilitadores da aprendizagem: (1) estudos dirigidos; (2) atividades presenciais como jogos didáticos ou oficinas (3) Debate, (4) animações demonstrativas de fenômenos biológicos, (5) discussão de artigos publicados pela mídia leiga, etc.
Formas de acompanhamento e de avaliação dos participantes
Avaliação
A avaliação será constituída por duas ferramentas: • Avaliação formativa: elaboração de portifólio diário. O aluno deverá registrar as
atividades desenvolvidas durante o dia e o seu envolvimento nas mesmas respondendo diariamente as seguintes questões: (1) O que aprendi hoje? O que as atividades me acrescentaram? (2) O que não foi adequado? Considerar nas respostas: o conteúdo da aula, as atividades e estratégias utilizadas, o relacionamento com o grupo e a própria participação. As respostas serão por escrito e entregues no final do dia. Um portfolio pode ser definido como um conjunto de diferentes tipos de documentos (anotações pessoais, experiências de aula, trabalhos pontuais, controles de aprendizagem, conexões com outros temas, fora da escola, representações visuais, etc.) que proporciona evidências de conhecimentos que foram sendo construídas durante o aprendizado, as estratégias utilizadas para aprender e a disposição de quem o elabora para continuar aprendendo. Devido à brevidade do curso o portifolio a ser elaborado diariamente no final de cada dia será mais sucinto e registrará o desenvolvimento

5
do programa de ensino e as reflexões diárias do processo de aprendizagem de modo a que o estudante sinta a aprendizagem como algo próprio e não alienada de seus processos pessoais e coletivos. O portfolio nesse caso é entendido também como uma reconstrução de conhecimento. Ele não se caracteriza como algo descritivo, mas reflexivo. Todos os portifólios são lidos diariamente pelos docentes do curso que podem desse modo realizar um acompanhamento mais personalizado da aprendizagem de cada um dos alunos.
• Avaliação final: Cada participante deverá fazer uma breve apresentação individual com relação a: (a) Quais os conteúdos/atividades desenvolvidas durante o curso que poderiam ser levadas para a escola? Por quê? (b) De que maneira isso poderia ser feito?
Relação Nominal dos professores Prof. Dra. Eliana Maria Beluzzo Dessen (professora responsável)
Monitoria (mestrandos e doutorandos do Instituto de Biociências) Adriana Ribeiro de Oliveira Marques, Ana Carolina Susuki Dias Cintra, Fernando Nodari, Lúcia Teiceira Machado Paula Cristina Gorgueira Onofre Renato Chimaso dos Santos Yoshikawa, Silvio Ganika Higa, Vivian Lavander Mendonça

6
PORTFÓLIO COMO RECURSO DE AVALIAÇÃO (adaptado de Hernandez, Fernando. Cultura visual, mudança educativa e projeto de trabalho. Porto
Alegre, Artmed, 2000)
A avaliação é um processo inerente ao processo de construção de conhecimento. Mais do que memorizar ou recordar informações ou aplicar fórmulas para resolver
problemas, o objetivo do processo educativo se propõe a aprender a formular problemas e desenvolver a capacidade de buscar, organizar e interpretar a informação dando-lhe sentido e transformando-a em conhecimento.
A avaliação compreende três formas de recolhimento de informações: • avaliação inicial, para perceber o conhecimento prévio de estudantes ao iniciarem
o curso; • a avaliação formativa, que está na base do processo avaliador e não tem a
finalidade de controlar ou qualificar, mas ajudar estudantes a “progredir no caminho do conhecimento”, e
• a avaliação somativa, que é o processo de síntese, que “permite reconhecer se [as] os estudantes alcançaram os resultados esperados (...) e serve como passagem para dar credibilidade oficial aos conhecimentos adquiridos.”
O portfólio representa uma possibilidade alternativa de avaliação, e pode ser, para
algumas disciplinas, substituto das avaliações pontuais em forma de provas e exames. Na educação ele serve como possibilidade de indicar a trajetória de aprendizagem e
de novas formas de avaliar o desenvolvimento do conhecimento. Uma das vantagens da realização do portfólio é a de perceber o desenvolvimento do programa de ensino e a participação mais ativa de estudantes, o que permite que sintam a aprendizagem como algo próprio e não alienada de seus processos pessoais e coletivos.
O portfólio é uma forma de avaliação dinâmica realizada pelo próprio estudante e que reflete seu desenvolvimento e suas mudanças através do tempo “. Nele inclui-se a avaliação do processo, a maneira de encarar e de interpretar as experiências e os processos de aprendizagem”. Definição de um portfólio:
Podemos definir um portfólio como um conjunto de diferentes tipos de documentos (anotações pessoais, experiências de aula, trabalhos pontuais, controles de aprendizagem, conexões com outros temas, fora da escola, representações visuais, etc.) que proporciona evidências de conhecimentos que foram sendo construídas, as estratégias utilizadas para aprender e a disposição de quem o elabora para continuar aprendendo. (...) Um portfoóio não significa apenas selecionar, ordenar evidências de aprendizagem e organiza-las num formato para serem apresentadas. (...) O que caracteriza definitivamente o portfólio como modalidade de avaliação não é tanto o seu formato físico (pasta, caixa, CD-ROM, etc.), mas sim a concepção de ensino e aprendizagem que veicula“.
O portfólio não é a mera recopilação de apontamentos; mas pode ser entendido como uma reconstrução de conhecimento. Ele não se caracteriza como algo descritivo, mas reflexivo. Assim, um diário reflexivo é uma ferramenta importante para a sua realização Estabelecer as finalidades de aprendizagem por parte de cada estudante
• Cada qual explicita o que pretende chegar a aprender. • Professora explicita os objetivos. • Uma possibilidade: extrair uma frase de cada apontamento de aula (ou leitura) e
fazer um comentário reflexivo, representativo do que foi significativo.
• Incluir experiências da sala de aula e de fora dela.

7
• Pensar no grupo: o processo de aprendizagem é mais significativo se for proveitoso para todo o grupo.
• Fazer um acordo público por escrito é conveniente e, se possível, presente na sala de aula como forma permanente de compromisso compartilhado.
• Nomear as fontes relacionadas com o processo (não apenas fontes bibliográficas): as evidências de aprendizagem
• Encontrar um fio condutor que organize a seleção das evidências que farão parte do portfólio
• Ter presente as perguntas: o que aprendi? De que maneira aprendi? O portfólio é propriedade do estudante
• O trabalho realizado no portfólio é memória de aprendizagem. • Cada portfólio é criação única, pois cada qual determina que evidências e que
experiências devem ser incluídas e faz uma auto-avaliação do seu processo de formação.
• Ele é parte do processo de aprendizagem de cada aluna e cada aluno. • Ele pode tornar-se público para compartilhar com o grupo e ajudar no processo
coletivo de aprendizagem – “estudantes e docentes podem ir construindo um conhecimento compartilhado mais equilibrado” (p.170).
Os componentes do portfólio a) O propósito
• Diário reflexivo: falar sobre os temas, comentando-os, não de forma descritiva, mas de forma reflexiva - também com perguntas, questionamentos, dúvidas.
• Não é mera recopilação dos apontamentos. • Estudantes explicitam como imaginam construir o seu portfólio. • Cada exemplo selecionado para dar evidência de seu progresso deve ser
recolhido, criado e organizado de uma determinada forma para demonstrar sua avaliação. Ter presente o fio condutor mais a explicitação do porquê de ter selecionado cada evidência.

8
AS CÉLULAS EUCARIÓTICAS E SUA CAPACIDADE DE DIFERENCIAÇÃO
Regina Célila Mingroni Netto e Eliana Maria Beluzzo Dessen Nosso corpo é formado por diversos tipos de células
O nosso corpo é constituído de trilhões de células, organizadas em diversos tipos de tecidos. Todas essas células originam-se de uma única, denominada célula-ovo ou zigoto, que, por sua vez, é o resultado da união de duas outras: o espermatozóide e o óvulo. À medida que o embrião cresce, grupos de células vão se tornando diferentes em estrutura e função, em decorrência de um processo chamado de diferenciação celular (Figura 1).
Figura 1. O zigoto dá origem aos trilhões de células diferenciadas de nosso organismo. Em última análise, esse processo é controlado pelo DNA, que é o material
genético. Mas, se o DNA contém a informação genética e essa informação é a mesma em todas as células do nosso corpo, você consegue entender como é possível que as células possam ser tão diferentes? O que se tem concluído das pesquisas científicas é que as células dos tecidos se diferenciam por terem diferentes trechos da molécula de DNA, ou seja, diferentes genes em funcionamento. Assim, as modificações celulares no processo de diferenciação resultam da ativação de certos genes e da inativação de outros: cada tipo de célula possui um conjunto característico de genes ativos. Em conseqüência dessa atividade diferencial, o conjunto de proteínas codificadas pelos genes varia dependendo do tipo de célula. Por exemplo, nas células do tecido nervoso, estão ativos genes que codificam proteínas que tornam as células ramificadas e capazes de fazer sinapses. Por outro lado, nas células das glândulas salivares, devem estar ativos genes que codificam enzimas secretadas na saliva. É claro que os genes que determinam a produção das enzimas da saliva não devem estar ativos em nenhum outro tecido do corpo. A atividade
Célula nervosa
Células do sangue
Células adiposas
Células musculares
Óvulo sendo fertilizado
Fases iniciais do desenvolvimento embrionário, a partir do zigoto

9
diferencial dos genes começa a ser determinada no decorrer do desenvolvimento embrionário e persiste nos tecidos adultos.
Todas as células têm duas características importantes: o grau de diferenciação e a potencialidade. O grau de diferenciação reflete o quanto uma célula é especializada. A potencialidade é a capacidade que ela tem de originar outros tipos celulares. Quanto maior a potencialidade da célula, geralmente será menor o seu grau de diferenciação. O zigoto é a célula com a máxima potencialidade, pois ele dá origem a todos os tipos de células. Assim, ele não é especializado ou diferenciado. No outro extremo, há células com potencialidade nula, como é o caso dos glóbulos vermelhos. Durante o processo de diferenciação dessas células, elas perdem o núcleo. Perderam, conseqüentemente, a capacidade de originar células iguais a elas. Logo, não têm potencialidade.
A compreensão das diferenças de potencialidade celular é importante para o entendimento de uma série de tópicos tratados nesse volume.
Figura 2. Classificação das células de acordo com sua potencialidade.
Células-tronco totipotentes podem originar um organismo inteiro. Ex. Zigoto e primeiras células que resultam da divisão do zigoto
Células-tronco pluripotentes podem originar quase todos os tipos de
tecidos. Ex. Massa interna do blastocisto
Célula-tronco multipotente podem originar diversos tipos de tecidos. Ex. Células-tronco do adulto
CT hematopoiética
outras CT
plaquetas Glóbul
hemácias Glóbulos brancos

10
DIFERENCIAÇÃO CELULAR E O CONTROLE DO GENE EUCARIÓTICO Eliana Maria Beluzzo Dessen (adaptado de Fundamentos de Biologia Celular – Alberts e col.)
A diferenciação produz uma variedade de células especializadas em eucariotos
Durante as repetidas divisões celulares que ocorrem no zigoto unicelular transformando-o em um organismo multicelular, as células individuais sofrem diferenciação celular, isto é, tornam-se especializadas em estrutura e função. É a regulação de genes que leva a essa especialização. Genes ativos em células de uma asa de mosca em desenvolvimento, por exemplo, são expressos como proteínas que tornam as células chatas e lisas, formando uma superfície de vôo forte e fina, semelhante a plástico transparente. Noutro exemplo, as células dos olhos em desenvolvimento, outros genes estão se expressando e sintetizam as proteínas que formam lentes capazes de focalizar luz. Células especializadas podem reter todo o seu potencial genético
O zigoto possui um conjunto completo de genes que dará origem a todos os tipos de células especializadas do organismo. O que ocorre a esses genes à medida que as células se diferenciam? Uma hemácia, por exemplo, perde seu núcleo e todo seu DNA. Porém, a maioria das células diferenciadas retém o núcleo e um conjunto completo de cromossomos. Quando todos os genes ainda estão presentes as células diferenciadas retêm seu potencial de expressa-los? Uma maneira de responder a essas questões é a experimentação, ou seja, substituir o núcleo de um ovo ou zigoto pelo núcleo de uma célula diferenciada. Se genes forem perdidos ou irreversivelmente inativados durante a diferenciação, o núcleo transplantado não permitirá o desenvolvimento de um embrião normal. Experimentos pioneiros de transplantes de núcleos foram realizados pelos embriologistas Robert Briggs e Thomas King na década de 1950. Esses pesquisadores destruíram os núcleos de óvulos de sapo com luz ultravioleta (UV) e, em seguida, transplantaram no óvulo anucleado um núcleo de célula intestinal de girino. Muitos dos ovos contendo os núcleos transplantados começaram a se desenvolver, porém, poucos originaram girinos normais. Desse modo os pesquisadores foram capazes de clonar sapos – produzir cópias geneticamente idênticas – usando núcleos de células diferenciadas. Tais estudos mostraram que núcleos de células diferenciadas podem reter todo o seu potencial genético. Evidencias adicionais apareceram em 1997 com a clonagem do primeiro mamífero usando núcleos diferenciados. Nesse caso, os pesquisadores usaram choques elétricos para fundir uma célula de glândula mamária de ovelha com um óvulo do qual o núcleo havia sido retirado. O ovo começou a se dividir, foi implantado no útero de outra ovelha, e desenvolveu-se na celebrada “Dolly”. Como previsto Dolly parecia-se com sua parental feminina, a célula de mama doadora do núcleo, e não com o óvulo doador ou a mãe de aluguel. Outra indicação que a diferenciação não interfere no potencial genético é o processo natural de regeneração, ou seja, a reposição de partes perdidas do corpo. Quando uma salamandra perde uma perna, por exemplo, certas células do toco do membro se diferenciam, e então se rediferenciam para dar origem a uma nova perna. Em plantas, a habilidade de uma célula diferenciada desenvolver-se em um novo organismo é comum. A figura 1C mostra de modo esquematizado uma única célula, removida da raiz de cenoura e colocada em meio de cultura, pode começar a se dividir e originar uma planta adulta. Essa técnica pode ser usada para clonar plantas, reproduzindo centenas de milhares de organismos geneticamente idênticos a partir de células somáticas de um único indivíduo. Desse modo, é possível propagar grande número de plantas que tem características desejáveis tais como alta produtividade de frutos ou resistência a doenças. O fato de uma planta madura poder se desdiferenciar e originar

11
todos os tipos de células especializadas de uma nova planta é uma evidência que a diferenciação não necessariamente envolve mudanças irreversíveis no DNA. Cada tipo de célula diferenciada tem um padrão de expressão gênica
Se todas as células diferenciadas de um organismo contêm os mesmos genes, e todos os genes têm o potencial de ser expressar, como as células tornam-se especializadas? Como já foi dito, as grandes diferenças entre as células em um organismo resultam da expressão seletiva de genes. À medida que um embrião em desenvolvimento sofre sucessivas divisões, genes específicos são ativados em diferentes células durante diferentes períodos de tempo. Grupos de células seguem vias de desenvolvimento diversas, e cada grupo desenvolve um tipo particular de tecido. Finalmente, no organismo maduro, cada tipo de célula – nervosa ou pancreática, por exemplo, - tem um padrão diferente de genes que são expressos. A Tabela abixo ilustra padrões de expressão gênica para alguns genes em células de três diferentes tecidos especializados de um mamífero. Os genes para as enzimas da via metabólica da glicolise estão ativos em todas as células metabolicamente ativas, incluindo células do pâncreas, do cristalino e nervosas, como exemplificado. Entretanto, os genes que codificam proteínas especializadas são expressos apenas por células específicas.
Célula pancreática Célula do cristalino (embrião)
neurônio
Genes das enzimas da via
glicolítica
Funcionais Funcionais Funcionais
Gene do cristalino
Inativo Funcional Inativo
Gene da insulina
Funcional Inativo Inativo
Gene da hemoglobina
Inativo Inativo inativo
As proteínas especializadas que foram usadas como exemplo são as proteínas transparentes do cristalino, que formam a lente do olho; o hormônio insulina; e a proteína transportadora de oxigênio, hemoglobina. Note que os genes para hemoglobina não estão ativos em nenhum dos tipos celulares mostrados na figura. Eles se expressam apenas nas células que irão se desenvolver em hemácias. Os genes para insulina são ativados apenas nas células do pâncreas que produzem hormônio. As células nervosas expressam genes para outras proteínas especializadas não mostradas. Células maduras do cristalino, e as hemácias, por exemplo, atingem um grau máximo de diferenciação, pois elas, após acumularem produtos protéicos, perdem seus núcleos e, assim, todos os seus genes. Vimos então que as células eucarióticas tornam-se especializadas porque expressam apenas certos genes. Desse modo, a diferenciação celular em organismos multicelulares resulta da expressão gênica seletiva, assim como a habilidade de bactérias produzirem diferentes enzimas quando necessárias. A seguir será examinado com mais detalhe o controle da expressão gênica nos eucariotos. A transcrição é controlada por proteínas que se ligam a seqüências reguladoras de DNA
Quando comparados com os procariotos, os eucariotos enfrentam as mesmas tarefas básicas de coordenação da expressão gênica, porém de um modo muito mais complexo. Alguns genes têm que responder a mudanças nas condições fisiológicas. Muitos outros são parte de circuitos genéticos de desenvolvimento que organizam as células em tecidos e tecidos em um organismo inteiro (exceto para os eucariotos

12
unicelulares). Nesses casos, os sinais que controlam a expressão gênica são produtos de genes que regulam o desenvolvimento e não sinais do meio externo. Algumas das seqüências de DNA reguladoras são curtas, cerca de 10 pares de nucleotídeos, e atuam como um interruptor gênico, ligando ou desligando o gene, em resposta a um único sinal. Esse tipo simples de interruptor gênico predomina nas bactérias. Nos eucariotos existem longas seqüências reguladoras de DNA (algumas vezes mais do que 10.000 pares de bases) que atuam como um microprocessador molecular, respondendo a uma variedade de sinais que são por elas integrados e que determinam a taxa de início da transcrição. As seqüências de DNA reguladoras não funcionam por si só. Para que haja qualquer efeito essas seqüências devem ser reconhecidas por proteínas denominadas proteínas reguladoras que têm a capacidade de se ligarem ao DNA. É a combinação de uma seqüência de DNA e suas moléculas de proteínas associadas que atuam como interruptor no controle da transcrição. Centenas de seqüências reguladoras de DNA foram identificadas, e cada uma delas é reconhecida por uma ou mais proteínas reguladoras. As proteínas que reconhecem seqüências especificas de DNA o fazem porque a superfície da proteína ajusta-se perfeitamente na dupla hélice de DNA de maneira seqüência-específica, e assim, diferentes proteínas irão reconhecer diferentes seqüências de nucleotídeos. Na maioria dos casos, a proteína insere-se no sulco maior da dupla hélice e realiza uma série de contatos moleculares com os pares de bases. A proteína forma pontes de hidrogênio, ligações iônicas, e interações hidrofóbicas com as extremidades das bases, usualmente sem romper as pontes de hidrogênio que une os pares de bases. Embora cada contato individual seja fraco, os cerca de 20 contatos que são geralmente formados na interface DNA-proteína atuam juntos para assegurar que a interação seja altamente especifica e muito forte; de fato as interações DNA-proteínas estão entre as mais firmes e específicas interações moleculares conhecidas em biologia. Embora cada exemplo de reconhecimento proteína-DNA seja único em seus detalhes, muitas das proteínas responsáveis pela regulação gênica contêm um dos vários padrões estáveis de dobramento que formam os chamados motivos estruturais. Essas regiões da proteína que se apresentam dobradas em motivos estruturais se ajustam ao sulco maior da dupla hélice do DNA e formam associações estreitas com um curto trecho de pares de bases. A iniciação da transcrição gênica em eucariotos é um processo complexo
Os interruptores dos genes presentes em bactérias são exemplos vivos da economia e simplicidade freqüentemente observada em biologia. Em eucariotos, entretanto, um gene típico responde a muitos sinais diferentes, e sua regulação é, conseqüentemente, mais complexa. A polimerase do RNA de eucariotos necessita de fatores gerais de transcrição
Nos eucariotos, são várias as funções atribuídas aos fatores gerais de transcrição no processo de início da transcrição pela polimerase II do RNA: posicionar corretamente a polimerase no promotor, ajudar a separar as duas fitas da molécula de DNA para permitir que a transcrição se inicie e liberar a polimerase do RNA do promotor uma vez iniciada a transcrição. O termo geral refere-se ao fato desses fatores associam-se a todos os promotores transcritos pela polimerase II do RNA. Nesse aspecto, os fatores gerais de transcrição diferem dos repressores e ativadores (descritos em bactérias no texto VI) que atuam em genes ou operons específicos, e das proteínas reguladoras dos genes eucarióticos (discutidas a seguir), que também atuam apenas em genes específicos. A Figura 1 mostra um modelo de como os fatores gerais de transcrição associam-se aos promotores utilizados pela polimerase II do RNA. O processo de montagem do complexo de iniciação começa com a ligação do fator de transcrição TFIID a uma curta seqüência de DNA dupla hélice composta por nucleotídeos T e A, conhecida como

13
seqüência TATA ou TATA box. Ao ligar-se ao DNA, o fator TFIID causa uma dramática distorção local na molécula de DNA. Tal distorção funciona como um sinal para a subseqüente montagem de outras proteínas no promotor. A seqüência TATA é um componente presente em praticamente todos os promotores utilizados pela polimerase II do RNA e localiza-se cerca de 25 nucleotídeos a montante (upstream) do sítio de início da transcrição. Após a ligação do primeiro fator geral de transcrição ao DNA, outros fatores também se ligam, juntamente com a polimerase II do RNA, para formar o complexo de iniciação da transcrição. Proteínas reguladoras controlam a distância a expressão de genes eucarióticos
As bactérias utilizam proteínas reguladoras (ativadoras e repressoras) para regular a expressão de seus genes. As células dos eucariotos utilizam a mesma estratégia básica. Embora seja necessária a presença conjunta dos fatores gerais de transcrição e da polimerase do RNA para o início da transcrição in vitro (veja figura 1), dentro das células essas proteínas sozinhas não conseguem iniciar a transcrição de modo eficiente. Praticamente todos os promotores eucarióticos necessitam também de proteínas ativadoras que auxiliam a associação dos fatores gerais de transcrição e da polimerase do RNA.
Os sítios do DNA aos quais se ligam as proteínas ativadoras dos genes eucarióticos foram denominados enhancers, desde que sua presença aumenta dramaticamente a taxa de transcrição. Foi muito surpreendente para os biólogos quando, em 1979, foi descoberto que essas proteínas ativadoras podiam se ligar a segmentos muito distantes do promotor, a milhares de pares de bases. Além disso, esses ativadores eucarióticos podem influenciar a transcrição quando se ligam a montante (upstream) ou a jusante (dowstream) do gene. Como as seqüências enhancers e as proteínas ligadas a elas funcionam a distâncias tão grandes? Como elas se comunicam com o promotor? Vários modelos de “ação à distância” foram propostos, mas o mais simples deles parece se aplicar para a maioria dos casos. O segmento de DNA compreendido entre o enhancer e o promotor dobra-se permitindo que as proteínas ligadas ao enhancer fiquem em contato ou com a polimerase do RNA ou com um dos fatores gerais de transcrição ligados ao promotor (Figura 2). Desse modo, o segmento de DNA compreendido entre o enhancer e o promotor DNA atuaria como uma estrutura de ligação que aproximaria a proteína ligada ao enhancer, localizado a milhares de pares de bases, permitindo sua interação com o complexo de proteínas ligadas ao promotor. Em eucariotos, as proteínas reguladoras ligadas a seqüências reguladoras distantes do promotor podem aumentar ou então diminuir a atividade da polimerase do RNA ligada ao promotor. Uma das maneiras de ação de tais proteínas é influenciar a montagem do complexo de iniciação. Proteínas ativadoras irão facilitar a montagem do complexo enquanto repressoras impedem a montagem correta. O empacotamento do DNA em nucleossomos no promotor pode afetar a iniciação da transcrição
As proteínas da cromatina e o DNA são parceiros no controle das atividades do material genético dentro da célula. O cromossomo é um complexo nucleoprotéico intrincadamente enovelado e com muitos domínios (Figura 3), nos quais a estrutura da cromatina local está estreitamente relacionada à manutenção de genes na configuração ativa ou silenciada, e a outras atividades da célula tais como a replicação do DNA, o emparelhamento e segregação dos cromossomos, e a manutenção da integridade do telômero e centrômero. Algumas regiões do genoma (heterocromatina, telômero e centrômero) estão empacotadas com características estruturais especificas. Esse empacotamento diferencial é definido por histonas modificadas, ou pela associação de proteínas adicionais não histônicas, ou então por moléculas de RNA reguladoras, que surpreendentemente também estão implicadas na organização da cromatina. Por exemplo, o X inativo dos

14
mamíferos está enriquecido por variantes de histonas como a macro H2A, quase três vezes maior que a H2A. No centrômero dos vertebrados, uma das histonas do octâmero, a H3, é substituída pela variante CENP-A. Esta por sua vez, forma um complexo com outras proteínas do centrômero, influenciando assim o empacotamento da cromatina centromérica. Uma vez que os nucleossomos estão localizados ao longo do DNA em intervalos regulares e com pouca especificidade, é provável que eles ocorram sobre regiões promotoras. Tais nucleossomos podem ser deslocados quando a transcrição do gene é ativada, embora ainda não seja completamente entendido como ocorre esse deslocamento. Sabe-se, porém que a célula possui proteínas especializadas cuja função é deslocar nucleossomos dos promotores e liberar o caminho para a montagem dos fatores gerais de transcrição. Outra possibilidade é que, como um prelúdio para a iniciação, as histonas nas vizinhanças do promotor sejam quimicamente modificadas, um passo que desestabiliza os nucleossomos afetados.
Nucleossomos formados em seqüências reguladoras de DNA podem também interferir com a expressão gênica bloqueando a ligação de proteínas. Entretanto, nem sempre isso ocorre. Enquanto há evidencias de que algumas seqüências reguladoras são mantidas expostas em regiões livres de nucleossomos, certas proteínas reguladoras parecem capazes de ligarem-se às seqüências do DNA mesmo quando essas se encontram incorporadas em nucleossomos, possivelmente desestabilizando e desmontando parcialmente o nucleossomo nesse processo. A célula tem várias estratégias para assegurar que o inicio da transcrição ocorra num DNA empacotado em nucleossomos. Entretanto, também é claro que quanto mais compacta for a forma da cromatina (aquela encontrada em cromossomos mitóticos, cromossomos X inativos, e outras regiões da cromatina interfásica) mais resistente ela será ao inicio da transcrição. Presumivelmente, isso ocorre porque as proteínas reguladoras, os fatores gerais de transcrição, e a polimerase do RNA não podem ter acesso ao DNA quando ele está tão densamente empacotado. Genes eucarióticos são regulados por uma combinação de proteínas
Nos eucariotos, as seqüências que controlam a expressão de um gene podem se espalhar por longos segmentos de DNA. Em animais e plantas não é raro encontrar seqüências reguladoras localizadas a 50.000 pares de nucleotídeos, embora a maioria desse DNA sirva apenas como “espaçador” e não seja reconhecido por proteínas reguladoras do gene. As proteínas reguladoras de genes não funcionam individualmente para ligar ou desligar um gene. Enquanto essa idéia cabe para muitos ativadores e repressores de bactérias, a maioria das proteínas que regulam os genes dos eucariotos funcionam como parte de um comitê de proteínas reguladoras, todas necessárias para fazer com que o gene se expresse na célula certa, em resposta às condições corretas, no tempo certo, e com nível de expressão adequado. O termo controle combinatorial refere-se ao modo como grupos de proteínas trabalham juntas para determinar a expressão de um único gene. Como mostrado na figura 4, muitas proteínas diferentes ligam-se a seqüências reguladoras para influenciar o inicio da transcrição nos eucariotos. A maioria dos genes eucarióticos possui regiões reguladoras contendo numerosos sítios para ambos os tipos de proteínas: com ação ativadora e repressora. Padrões estáveis de expressão gênica podem ser transmitidos para as células filhas Embora todas as células procarióticas e eucarióticas sejam capazes de ligar e desligar genes, organismos multicelulares necessitam de mecanismos especiais de controle para gerar e manter seus diferentes tipos de células. Uma vez que uma célula de um organismo multicelular tenha se diferenciado em um tipo específico, ela geralmente irá

15
permanecer diferenciada, e se for capaz de dividir-se, toda a sua progênie será do mesmo tipo celular. Algumas células altamente especializadas nunca se dividem após a diferenciação, como por exemplo, células da musculatura esquelética e neurônios. Mas há vários outros tipos de células diferenciadas, tais como fibroblastos, células da musculatura lisa, e células do fígado (hepatócitos), que irão se dividir muitas vezes durante a vida do organismo. Todos esses tipos celulares originam, quando se dividem, apenas células como elas mesmas: células de musculatura lisa não originam células do fígado. Isso significa que as mudanças na expressão gênica dão origem as células diferenciadas devem ser lembradas e transmitidas para suas células filhas em todas as divisões subseqüentes, ao contrário das mudanças temporárias na expressão gênica que ocorrem nas células de procariotos e eucariotos. Por exemplo, nas células ilustradas na figura 13, a produção de cada proteína reguladora, uma vez iniciada, deve ser perpetuada nas células filhas em cada divisão celular. Como isso deve ocorrer? Há várias maneiras de assegurar que as células filhas “lembrem” que tipos de células espera-se que elas sejam. Uma das mais simples delas é por meio de uma alça de feedback positivo, onde uma proteína reguladora chave ativa a transcrição do gene que a codifica além de ligar genes específicos de outros tipos celulares (figura 5). Por exemplo, a proteína reguladora MyoC funciona com esse tipo de feedback. Outra maneira de manutenção do tipo celular é através da propagação fiel da estrutura da cromatina da célula parental para a célula filha mesmo com um evento de replicação entre elas. Um exemplo disto é o fenômeno da inativação do cromossomo X nos mamíferos. O evento de inativação de um dos cromossomos X, o de origem paterna ou o de origem materna, ocorre no início do desenvolvimento embrionário e, a partir daí, o mesmo cromossomo X é inativado por muitas gerações seguidas. O mecanismo molecular por meio do qual o estado da cromatina é transmitido não é ainda totalmente conhecido em detalhes
Figura 1.Início da transcrição de um gene eucariótico pela polimerase II do RNA. Para que a transcrição possa se iniciar são necessários vários fatores gerais de transcrição denominados, TFIIA, TFIIB e assim por diante. (A) o promotor contém uma seqüência de DNA denominada TATA Box, localizada a cerca de 25 nucleotídeos do sítio de início de transcrição. (B) O fator TFIID reconhece a seqüência TATA, se liga a ela, e permite a ligação do fator TFIIB. (C a E) os demais fatores de transcrição e a polimerase ligam-se ao promotor como em uma linha de montagem.
início da transcrição
RNA polimerase
transcrição

16
Figura 2. Modelo de ativação gênica à distância. Nesse exemplo, os fatores gerais de transcrição, os fatores de transcrição e a polimerase do RNA por si só não se associam eficientemente ao promotor e uma proteína reguladora ligada ao enhancer é necessária para estimular o processo de montagem do complexo de iniciação.O dobramento do DNA permite o contato entre a proteína reguladora ligada ao enhancer e o complexo de iniciação ligado ao promotor. No desenho, a linha interrompida indica a grande distancia que geralmente existe entre o enhancer e o promotor.
Figura 3. Esquema de alguns dos níveis de empacotamento da cromatina do cromossomo mitótico altamente condensado. O nível deorganização melhor compreendido é aquele em que o DNA nu associa-se às histonas formando os nucelossomos. As estruturas que correspondem aos níveis seguintes de organização são mais especulativas.
Proteína ativadora
Ligação de fatores gerais de Transcrição, polimerase do RNA, mediadores, etc.
Início da transcrição
Início da transcrição
DNA
Colar de contas
Fibra cromatínica de 30nm
A fibra se organiza em
alças
Segmento condensado
Cromossomo mitótico
Cada molécula de DNA foi empacotada em cromossomos mitóticos que é 10.000 vezes
mais curto que o DNA entendido.

17
Figura 4. Seqüências reguladoras de um gene eucariótico típico. O promotor é a seqüência de DNA onde a polimerase do RNA e os fatores gerais de transcrição se ligam. As seqüências reguladoras do gene são usadas como sítios de ligação de proteínas reguladoras cuja presença no DNA afetam a taxa de início de transcrição. As seqüências reguladoras podem estar localizadas adjacentes ao promotor, muito longe dele na direção 5’ (montante) ou, a 3’do gene (jusante). Figura 5. Esquema do modo como uma alça de feedback positivo pode criar a memória celular. A proteína A é uma proteína reguladora que ativa sua célula progenitora que experimentou um sinal transitório que deu início à produção da proteína. (I) A proteína A não é normalmente produzida, pois ela é necessária para sua própria transcrição. (II) sinal transiente liga o gene A, (III) O efeito do sinal transiente é lembrado em todas as células descendentes.
Seqüências reguladoras do gene
DNA espaçador
Fatores gerais de transcrição RNA polimerase Proteínas
Reguladoras do gene
5’ Início da transcrição
I II
III

18
SINALIZAÇÃO CELULAR Adriana Ribeiro de Oliveira-Marques – adaptado de Alberts e col. 2004.
Os organismos multicelulares possuem um elaborado sistema de comunicação
celular. Tal sistema depende de: 1. moléculas-sinal extracelulares, produzidas por células para sinalizar células
vizinhas ou mais distantes 2. um elaborado sistema de proteínas que cada célula contem e que a habilita a
responder a um conjunto particular de sinais de modo célula-específico. Essas proteínas incluem:
a. proteínas receptoras de superfície celular que se ligam a molécula sinal b. uma variedade de proteínas sinalizadoras intracelulares que distribuem o
sinal para partes apropriadas da célula. Entre essas proteínas estão: quinases, fosfatases, proteínas que se ligam a GTP e proteínas que interagem com as anteriormente citadas.
c. no final de uma via de sinalização intracelular estão proteínas alvo, que são alteradas quando a via está ativa e mudam o comportamento da célula. Dependendo do efeito do sinal, essas proteínas alvo podem ser reguladoras, canal de íons, componentes da via metabólica, partes do citoesqueleto, etc
Princípios gerais da comunicação celular Para facilitar a compreensão de como ocorre a sinalização celular vamos fazer uma analogia com a transmissão de uma mensagem por telefone. Uma pessoa fala ao telefone e sua voz é convertida num sinal elétrico. O sinal é amplicado e a mensagem é carregada na forma de impulsos elétricos pelo fio do telefone. Na extremidade oposta o sinal elétrico é convertido em onda sonora que é captada pelo ouvido e finalmente expressada na forma de impulsos no encéfalo de quem a recebeu. Em passos sucessivos ao longo dessa via de comunicação, formas diferentes de sinais são usadas para representar a mesma informação: o ponto crítico na transmissão ocorre quando a informação é convertida de uma forma em outra. Esse processo de conversão é chamado transdução de sinal. Os sinais que passam entre as células são mais simples que as mensagens humanas: um tipo particular de molécula é produzido por uma célula – a célula sinalizadora – e detectada por outra – a célula alvo – por meio de proteínas receptoras, que reconhecem e respondem de modo especifico à molécula sinal. A proteína receptora realiza o primeiro passo numa série de processos de transdução na extremidade final da via de sinalização, na célula alvo, aonde o sinal extracelular que chega é convertido num sinal intracelular que direciona o comportamento da célula. Os dois pontos chave dizem respeito à recepção e a transdução do sinal. Quando os biólogos referem-se a sinalização celular, são esses dois aspectos que eles geralmente tem em mente. A seguir serão brevemente descritos os diferentes tipos de sinais que as células enviam umas para as outras. Sinais podem atuar em curta ou longa distância As células num organismo multicelular usam centenas de tipos de moléculas extracelulares para enviar sinais uma para as outras – proteínas, peptídeos, aminoácidos, esteróides, derivados de ácidos graxos e até gases dissolvidos – mas há apenas uma dezena de modos básicos de comunicação. A maneira mais pública de se comunicar é transmitir o sinal pelo corpo todo secretando na corrente sanguínea do organismo (Figura 1D). Moléculas sinais usadas dessa maneira são chamadas hormônios, e em animais, as células que produzem hormônios são chamadas células endócrinas.

19
Menos publico é o processo conhecido como sinalização parácrina. Nesse caso, as moléculas sinal difundem localmente pelo meio extracelular, relembrando as células nas vizinhanças da célula secretora: elas atuam como mediadores locais (Figura 1B). Muitas das moléculas-sinal que mediam a inflamação nos locais de infecção ou proliferação celular em cicatrização funcionam dessa maneira. Um terceiro modo de comunicação é a sinalização neuronal. Assim como na sinalização hormonal, mensagens são freqüentemente entregues em longas distancias; na sinalização neuronal, entretanto, mensagens são liberadas em linhas privadas para células individuais de modo muito rápido (Figura 1C). O axônio de um neurônio termina em junções especializadas (sinapses) nas células alvo distantes do corpo celular neuronal. Quando ativada por sinais do meio ou por outros nervos, o neurônio manda impulsos elétricos ao longo do axônio com velocidade superior a 100 metros por segundo. Chegando ao axônio terminal, os sinais elétricos intracelulares são convertidos em uma forma química extracelular: cada impulso elétrico estimula o terminal a secretar um pulso de um sinal químico chamado neurotransmissor. Neurotransmissores difundem através do estreito gap (<100nm) entre a membrana do axônio terminal e a membrana da célula alvo em menos que um mili-segundo. Um quarto modo de comunicação é o célula a célula – o mais intimo e a curta distancia de todos – não requer a liberação da molécula secretada. Ao contrário, as células fazem contato direto por meio de moléculas sinalizadoras em suas membranas plasmáticas. A mensagem é liberada pela ligação de uma molécula sinal ancorada na membrana plasmática da célula sinalizadora para uma molécula receptora embebida na membrana plasmática da célula alvo (Figura 1A) Enquanto um sinal neuronal é como uma chamada telefônica, essa sinalização dependente de contato é como uma conversa frente a frente.
Figura 1: Diferentes tipos de Sinalização. (A) dependente de contato, (B) parácrina, (C) sináptica, (D) endócrina, (E) autócrina e (F) junção do tipo Gap (modificada de Alberts e col., 2004).
No desenvolvimento embrionário, por exemplo, a sinalização dependente de contato tem um papel importante nos tecidos nos quais as células adjacentes, que são inicialmente semelhantes, têm que se tornar especializadas de modos diferentes. Assim, na linha de células que originam o tecido nervoso, células individuais têm que ser indicadas para diferenciar-se como neurônios enquanto suas vizinhas permanecem não

20
neuronais. Os sinais que controlam esse processo são transmitidos via contato célula-celula: cada futuro neurônio inibe seu vizinho imediato de diferenciar-se também como neurônio.
Cada célula responde a um número limitado de sinais. Uma célula típica de um organismo multicelular esta exposta a centenas de sinais diferentes de seu meio. Eles podem estar livres no fluido extracelular, ou ancorados na matriz extracelular, ou ligado às superfícies de células vizinhas. A célula deve responder de modo seletivo a esta mistura de sinais, desprezando alguns e reagindo a outros, de acordo com a sua função especializada. Se a célula vai reagir a uma molécula sinal depende, antes de qualquer coisa, dela possuir um receptor para esse sinal. Sem um receptor, a célula será surda ao sinal e não pode responder a ele. Produzindo apenas um numero limitado de receptores entre as centenas possíveis, a célula restringe a gama de sinais que podem afeta-la. Mas esta gama reduzida de sinais pode ainda ser usada para controlar o comportamento da célula por meio de modo complexo. A complexidade pode ser de dois tipos. Primeiro, um sinal, ligando a um tipo de proteína receptora, pode causar uma multiplicidade de efeitos na célula alvo: forma, movimento, metabolismo, expressão gênica – tudo pode ser alterado conjuntamente. O sinal de um receptor-de-superfície-da-célula é geralmente retransmitido para o interior da célula por um conjunto de outros componentes intracelulares que produz efeitos amplos. Esse sistema de retransmissão e os alvos intracelulares nos quais ele atua variam de um tipo de célula especializada para outro, de modo que células diferentes respondem de modo diferente a um mesmo sinal. Assim, quando uma célula do músculo cardíaco é exposta ao neurotransmissor acetilcolina, ela diminui a freqüência de contrações, mas quando uma glândula salivar é exposta ao mesmo sinal, ela secreta componentes da saliva (Figura 2).
Figura 2: Diferentes respostas induzidas pelo neurotransmissoraAcetilcolina. (A) célula muscular cardíaca, (B) célula musculoesquelética e (C) célula de glândula salivar (modificada de Alberts e col., 2004). O segundo tipo de complexidade existe porque numa célula típica uma coleção completa de receptores diferentes – algumas dúzias deles. Esse número é suficiente para tornar a célula simultaneamente sensível a muitos sinais extracelulares. Esses sinais, atuando juntos, podem evocar respostas que são mais do que a soma dos efeitos que cada sinal poderia causar separadamente. Os sistemas de retransmissão intracelulares dos diferentes sinais interagem, de modo que a presença de um sinal modifica a resposta de outro. Assim uma combinação de sinais pode simplesmente fazer com a célula sobreviva; outra combinação pode dirigir a célula a uma via de especialização; outra pode leva-la a dividir-se; e na ausência de qualquer sinal a célula pode ser programada para morrer. Desse modo, um número relativamente pequeno de sinais pode ser usado em diferentes combinações para fornecer controles sutis e complexos sobre o comportamento da célula.

21
Receptores retransmitem sinais por meio de vias de sinalização intracelular Antes de traçar com detalhes como uma molécula sinal particular controla o comportamento da célula, é importante saber alguns princípios gerais. A recepção de sinais começa no ponto onde um sinal originado fora da célula encontra a molécula alvo pertencente à própria célula. Em quase todos os casos a molécula alvo é uma proteína receptora, e ela é usualmente ativada por apenas um tipo de sinal. A proteína receptora desempenha o passo primário de retransmissão: ela recebe o sinal externo, e gera em resposta um novo sinal intracelular (Figura 3). Como regra, esse é apenas o primeiro evento em uma cascata subseqüente de sinais intracelulares nos processos de transdução do sinal. Neles, a mensagem é passada de um conjunto de moléculas sinalizadoras intracelulares para outro, cada um deles produzindo por sua vez o próximo sinal até que, por exemplo, uma enzima de uma via metabólica seja ativada, a expressão de um gene ativada, ou o citoesqueleto entre em funcionamento. Esse efeito final é a resposta da célula. Essas cadeias de retransmissão, ou cascatas de sinalização, de moléculas sinalizadoras intracelulares têm várias funções cruciais (Figura 3):
1. Elas transferem fisicamente o sinal de um ponto no qual ele foi recebido para a maquinaria celular que irá responder, a qual está freqüentemente localizada em alguma outra parte da célula.
2. Elas transformam o sinal numa forma molecular que é capaz de estimular uma resposta
3. Na maioria dos casos, cascatas de sinais também amplificam o sinal recebido, tornando-o mais forte, de modo que poucas moléculas sinais extracelulares são suficientes para evocar uma grande resposta.
4. as cascatas de sinalização podem distribuir o sinal de modo a que ele influencie vários processos em paralelo: em qualquer passo da via, o sinal pode divergir (separar) e ser retransmitido para um número de diferentes alvos intracelulares, criando ramos no fluxo de informação e evocando uma resposta complexa.
5. Por ultimo, cada passo nessa cascata de sinalização está aberto à interferência de outros fatores, de modo que a transmissão do sinal pode ser modulada de acordo com as condições que prevalecerem dentro ou fora da célula. Vamos inicialmente considerar algumas das vias de sinalização mais simples
antes de considerar as cascatas mais longas que retransmitem sinais dos receptores na membrana da célula nas células animais.
Algumas moléculas sinal podem cruzar a membrana plasmática Moléculas sinal extracelulares em geral são de duas classes, correspondendo a
dois tipos fundamentalmente diferentes de tipos de receptores. A primeira e maior classe de sinais consiste de moléculas que são muito grande ou muito hidrofílicas para atravessarem a membrana plasmática da célula alvo. As proteínas receptoras dessas moléculas sinais devem estar localizadas nas membranas das células alvo e retransmitir a mensagem através da membrana (Figura 4A) A segunda, e menor classe de sinais consiste de moléculas que são suficientemente pequenas e hidrofóbicas para difundir pela membrana plasmática. Para essas moléculas sinal, os receptores encontram-se no interior das células alvo e são geralmente ou proteínas reguladoras de genes (discutidas no cap 8) ou enzimas. Elas são ativadas quando a molécula sinal liga-se a elas (Figura 4B) As moléculas sinal, hidrofóbicas, mais bem conhecidas são os hormônios esteróides (inclusive cortisol, estradiol e testosterona) e o hormônio da tireóide (tiroxina). Todas elas passam através da membrana plasmática da célula alvo e liga-se a proteínas receptoras localizadas ou no citosol, ou no núcleo. Os receptores para esses hormônios são proteínas reguladoras de genes que estão presentes na forma inativa na célula não estimulada. Quando seu hormônio correspondente se liga, a proteína receptora sofre uma grande mudança conformacional que possibilita que ela

22
se ligue a seqüências reguladoras correspondentes no DNA; ela pode então promover ou a inibição ou a ativação da transcrição de um conjunto de genes. Há proteínas receptoras diferentes para cada tipo de hormônio; cada receptor atua num conjunto diferente de sítios reguladores e assim regula um conjunto diferente de genes. O papel essencial dos receptores dos hormônios esteróides é evidenciado pela dramática conseqüência de uma mutação, a que causa a falta de receptores de testosterona em humanos. O hormônio masculino testosterona atua no feto e na puberdade como um sinal para o desenvolvimento de sinais para o desenvolvimento das características sexuais secundárias. Alguns raros indivíduos são geneticamente machos (XY), mas não possuem o receptor de testosterona como resultado de uma mutação no gene correspondente: eles produzem hormônio, mas suas células não respondem a ele. A conseqüência é quem eles desenvolvem a aparência de mulheres. Essa demonstração do papel chave dos receptores de hormônio da testosterona mostra também que o receptor é necessário não em apenas um tipo de célula para mediar um efeito, mas em muitos tipos de células produzindo uma gama completa de características que distinguem os dois sexos.
Texto modificado de: Alberts, B.; Johnson, A.; Lewis, J; Raff, M.; Roberts, K.; Walter, P. - tradução: Ana Beatriz Gorini da
Veiga e col. – Biologia Molecular da Célula - Capítulo 15: Comunicação Celular - 4ª edição – Porto Alegre – Artmed, 2004
Figura 3: Diferentes tipos de proteínas de sinalização intracelular ao longo de uma rota de sinalização, desde o receptor de superfície celular até o núcleo. Nesse exemplo, uma série de proteínas sinalizadoras intracelulares conduz o sinal da molécula sinalizadora para dentro da célula, causando uma mudança na expressão gênica (no DNA). O sinal é amplificado, convertido (transduzido) e distribuído ao longo da via

23
de sinalização. No final, essa transmissão de sinais ativa ou inativa proteínas-alvo que alteram o comportamento celular, neste caso, a proteína-alvo é uma proteína reguladora de um gene.
Figura 4. A) Uma célula sinalizadora hidrofílica é incapaz de atravessar a membrana plasmática, por isso ela se liga a receptores de superfície celular, os quais geram um ou mais sinais dentro da célula-alvo. B) Algumas moléculas sinalizadoras são suficientemente pequenas e hidrofóbicas e conseguem atravessar a membrana celular e se ligar a um receptor citoplasmático (presente do citoplasma) ou a um receptor nuclear (presente no núcleo, como este exemplo) (Figura modificada de Alberts e col., 2004)

24
O BÁSICO SOBRE CÉLULAS-TRONCO
National Institutes of Health – Stem cells information - http://stemcells.nih.gov/info/basics/basics1.asp Tradução e adaptação Eliana Maria Beluzzo Dessen
Nota sobre a nomenclatura Células-tronco: (A nova fronteira da Medicina. Organizador Marco Antonio Zago e Dimas Tadeu Covas. Ed. Atheneu)
Apesar de estranha à língua portuguesa, a denominação “célula-tronco” foi adotada neste livro porque se impôs nos últimos anos na imprensa e nos meios científicos nacionais. Isso é apenas um reconhecimento dessa tendência por parte dos autores, não uma manifestação explícita de apoio a essa escolha. O termo constitui uma tradução literal do inglês “stem cell”. As línguas latinas têm expressões que descrevem melhor sua função primordial: célula madre (castelhano), cellula staminale (italiano) e céllule souche (francês). Em Portugal há uma forte tendência para utilizar as expressões célula-mãe ou célula estaminal, que estariam mais de acordo com a índole de nossa língua. CARACTERÍSTICAS DAS CÉLULAS-TRONCO
As células-tronco possuem três características gerais: (a) dividem-se dando origem a células iguais a ela, (b) são indiferenciadas e (c) podem dar origem a células especializadas ou diferenciadas.
Os cientistas estão tentando entender: (a) porque as células-tronco embrionárias têm a capacidade de proliferar
As células-tronco são
durante longos períodos, mais de um ano, em laboratório, sem diferenciarem-se e (b) quais são os fatores nos organismos vivos que normalmente regulam a proliferação e renovação dessas células. A elucidação dessas questões pode esclarecer como a proliferação é regulada durante o desenvolvimento embrionário normal e durante o processo de divisão celular alterado que leva ao câncer. Além disso, tais informações permitiriam que os cientistas conseguissem cultivar células-tronco embrionárias em laboratório com mais eficiência.
indiferenciadas
As células-tronco são capazes de se renovar por longo período. Ao contrário de células diferenciadas como musculares, sanguíneas ou nervosas, que não se dividem mais, as células-tronco replicam-se muitas vezes. Uma população inicial de células-tronco pode multiplicar-se em laboratório muitas vezes. Células com essa característica são ditas auto-renováveis. Foram necessários mais de 20 anos de pesquisas para que se aprendesse a cultivar células embrionárias sem que elas se diferenciassem espontaneamente. Assim sendo, uma importante área de estudo com células-tronco é entender os sinais, no organismo maduro, que mantém a população de células-tronco proliferando e indiferenciadas até que elas sejam necessárias para reparar em tecido. Essa informação é fundamental para que os cientistas possam cultivar grande número de células indiferenciadas no laboratório para experimentação futura.
, pois não possuem nenhuma estrutura tecido-específica que permita a realização de funções especializadas como, por exemplo, produzir saliva, contrair-se ou transmitir impulsos nervosos.
As células-tronco podem originar células especializadas. Quando células indiferenciadas originam células especializadas, o processo é chamado diferenciação. O entendimento dos sinais internos e externos da célula que desencadeiam a diferenciação é ainda incipiente. Os sinais internos são controlados pelos genes, que são os portadores das instruções para o funcionamento da célula. Os sinais externos para a diferenciação incluem substancias químicas secretadas por outras células, contato físico com células vizinhas e certas moléculas no microambiente celular
Muitas questões sobre células-tronco permanecem sem resposta. Por exemplo, os sinais internos e externos para a diferenciação são os mesmos para todas as células? Existe um conjunto de sinais que pode ser identificado como indutores de diferenciação para todos os tipos de células? Respostas a essas perguntas podem levar os
.

25
pesquisadores a encontrar novas maneiras de controlar a diferenciação de células-tronco em laboratório, uma vez que o cultivo de células ou tecidos pode ser usado para propósitos específicos inclusive terapia celular
As células-tronco encontradas em organismos não embrionários, denominadas .
células-tronco do adulto, normalmente dão origem aos tipos celulares dos tecidos nos quais residem. Por exemplo, célula-tronco de medula óssea normalmente origina células do sangue como hemácias, células brancas e plaquetas. Até recentemente se achava que células hematopoéticas - células que dão origem às células do sangue – não fossem capazes de gerar células de um tecido diferente, como células nervosas do cérebro. Entretanto, muitos experimentos realizados nos últimos anos mostraram que células-tronco de um tecido podem originar células de um tecido diferente, um fenômeno denominado plasticidade
. Um exemplo de plasticidade é a origem de neurônios, de células musculares cardíacas e de células produtoras de insulina a partir de células hematopoéticas. Assim, a possibilidade de usar células-tronco de adulto para terapias celulares tornou-se uma área de investigação muito ativa.
AS CÉLULAS-TRONCO EMBRIONÁRIAS Estágios do desenvolvimento embrionário importantes para gerar células-tronco embrionárias As células tronco-embrionárias são derivadas de embriões. No caso da espécie humana, os pesquisadores utilizam apenas células-tronco de embriões que foram obtidos a partir de óvulos fertilizados in vitro e que foram doados para fins de pesquisa científica. Os embriões dos quais as células-tronco humanas são derivados têm cerca de 4 a 5 dias e estão no estágio de blastocisto
. Os blastocistos têm três estruturas: o trofoblasto que é uma camada de células que rodeia o blastocisto, a blastocele – cavidade no interior do blastocisto e a massa interna de células, com aproximadamente 30 células, localizada numa extremidade da blastocele.
Figura 3. Fases iniciais do desenvolvimento embrionário. A - Estágio de 8 células, B – Mórula, C – início de blastocisto, D – blastocisto.
A B
C D
Zona pelúcida
Massa interna de células
Cavidade do blastocisto trofoblasto
Zona pelúcida em degeneração

26
Cultivo de células-tronco embrionárias em laboratório O cultivo de células-tronco em laboratório é chamado de cultura de células. As células-tronco embrionárias são isoladas por transferência da massa interna de células para uma placa de cultura contendo um meio nutritivo denominado meio de cultura. Nesse meio as células se dividem e se distribuem pela superfície da placa. A superfície interna da placa de cultura, geralmente, é recoberta com células epiteliais de camundongo que foram tratadas para não se dividir. Essa camada de células é denominada feeder layer
Durante vários dias, as células da camada interna do embrião proliferam e começam a superpopular a placa de cultura. Quando isso ocorre, o excesso de células é transferido para outras placas de cultura contendo meio fresco. Esse processo de replaquear as células é repetido muitas vezes, por muitos meses, é chamado de subcultura. Cada ciclo de subcultura é denominado
. Ela fornece uma superfície aderente na qual as células-tronco podem se unir. Além disso, a feeder layer libera nutrientes no meio de cultura. Recentemente, foram desenvolvidos métodos de cultivo sem a camada de células de camundongos o que corresponde a um grande avanço técnico, pois havia o risco de transmitir contaminantes (vírus e macromoléculas) do camundongo para as células humanas.
passagem. Após 6 meses, as 30 células originais da massa de células interna do blastocisto gerou milhões de células-tronco embrionárias. Tais células-tronco que proliferam em cultura por 6 meses sem se diferenciar são pluripotentes e constituem uma linhagem de células-tronco embrionárias
. Uma vez estabelecidas as linhagens elas podem ser congeladas ou usadas imediatamente em experimentos.
Testes de laboratório para identificar células-tronco embrionárias Os testes para identificação das células-tronco embrionárias estão resumidos abaixo:
• Cultivar as células em cultura para ver se permanecem indiferenciadas; • Estudar os marcadores de superfície
• Verificar a presença da proteína Oct-4, um fator de transcrição que ajuda a ligar e a desligar genes no tempo certo, um passo importante no processo de diferenciação e desenvolvimento embrionário;
que são encontrados apenas em células indiferenciadas;
• Analisar os cromossomos para detectar se estão normais; • Determinar se as células podem ser cultivadas após serem descongeladas; • Testar se células são pluripotentes: (a) manipular as células para que se
diferenciem em diferentes tecidos; (b) permitir que elas se diferenciem espontaneamente em cultura e (c) injetar as células em camundongos imuno-reprimidos para testar a formação de teratoma, um tipo de tumor benigno. Os teratomas contem uma mistura de células diferenciadas ou parcialmente diferenciadas – uma indicação de que as células-tronco embrionárias são capazes de se diferenciar em muitos tipos de células.
Estimulação de células-tronco embrionárias para diferenciar As células-tronco são estimuladas para se diferenciar por meio do acréscimo de diferentes substâncias, fatores de crescimento, ou hormônios ao meio de cultura. CÉLULAS-TRONCO DO ADULTO

27
Célula-tronco do adulto é uma célula indiferenciada localizada entre as células diferenciadas que compõem tecidos ou órgãos de um organismo. No organismo vivo, as células-tronco do adulto têm a função de manter e reparar os tecidos nos quais elas se encontram. Essas células são também chamadas células-tronco “adultas” ou células-tronco teciduais. A origem das células-tronco do adulto nos tecidos é desconhecida. Nos últimos anos, células-tronco do adulto foram encontradas em um número grande de tecidos. Esse fato levou os cientistas a se perguntarem se elas não poderiam ser usadas em transplantes de células. Na realidade, as células-tronco da medula óssea (hematopoiéticas), que dão origem às células do sangue, vem sendo usadas em transplantes a cerca de 30 anos. Certos tipos de células-tronco do adulto têm a habilidade de se diferenciarem num grande número de tipos celulares quando colocadas em ambiente apropriado. Se a diferenciação dessas células puder ser controlada em laboratório. Elas poderão vir a ser tornar a base para terapias de muitas doenças. Onde são encontradas as células-tronco de adulto? As células-tronco do adulto foram identificadas em muitos órgãos e tecidos. Um ponto importante a ser entendido sobre essas células é que há um número muito pequeno de células-tronco nos tecidos. Acredita-se que elas residam numa pequena área de cada tecido onde elas permanecem quiescentes (não se dividindo) por muitos anos até que sejam ativadas por doenças ou lesões. Os tecidos adultos nos quais células-tronco já foram localizadas são: encéfalo, medula óssea, sangue periférico, vasos sanguíneos, músculo esquelético, pele, fígado, polpa dos dentes, e tecido adiposo. É uma área de pesquisa intensa e com freqüência novos tecidos entram para essa lista. Cientistas do mundo inteiro estão tentando encontrar maneiras de crescer células-tronco do adulto em cultura e manipula-las para gerar tipos celulares específicos que possam ser usados no tratamento de lesões de diferentes tipos. Alguns exemplos de potenciais tratamentos incluem a substituição de células produtoras de dopamina no cérebro de doentes de Parkinson, células produtoras de insulina para tratamento de diabetes do tipo I e reparo do músculo cardíaco danificado por enfarto com células da musculatura do coração. O que se sabe sobre a diferenciação de células-tronco do adulto? As células-tronco do adulto entram em vias de diferenciação para formar células especializadas de diferentes tecidos nos quais elas residem (figura 4). Células-tronco do adulto de alguns tecidos podem também exibir a habilidade de formar tipos celulares de outros tecidos, um processo conhecido como plasticidade.

28
Figura 4. Diferenciação de células-tronco mesenquimais e hematopoiéticas Num organismo vivo as células-tronco podem se dividir por um longo período e originar células diferenciadas que têm formas, estruturas e funções especializadas daquele tecido. A seguir são apresentados alguns exemplos de vias de diferenciação de célula-tronco do adulto:
• Célula-tronco hematopoiética que origina todos os tipos de células do sangue: hemácias, linfócitos B e T, células killer, neutrófilos, basófilos, eosinófilos, monócitos, macrófagos e plaquetas (figura 4).
• Célula-tronco mesenquimais (estroma da medula óssea) origina uma variedade de tipos de células: células do osso (osteócitos), células da cartilagem (condrócitos), células adiposas, e outros tipos de tecido conectivo tais como os tendões,
• Células-tronco epiteliais do revestimento do trato digestório ocorrem em criptas profundas e originam vários tipos de células: células absortivas, células de Paneth e células enteroendócrinas,
• Células-tronco da camada basal da epiderme e na base do folículo do pêlo. As células-tronco epidermais dão origem aos queratinócitos, que migram para a superfície da pele e formam uma camada protetora. As células-tronco foliculares originam o folículo do pêlo e epiderme.
Um grande número de experimentos sugere que certas células-tronco de adulto são
pluripotentes. Essa habilidade de diferenciar-se em múltiplos tipos celulares é chamada plasticidade (ou transdiferenciação, por alguns autores). A lista seguinte apresenta exemplos de plasticidade das células-tronco de adulto (figura 5):
• Células-tronco hematopoiéticas podem se diferenciar em três tipos principais de células nervosas: neurônios, oligodendrócitos e astrócitos; células de músculo cardíaco; células de fígado.
• Células-tronco estromais podem se diferenciar em: células de musculatura cardíaca e esquelética
• Células-tronco de cérebro podem diferenciar em: células sanguíneas e células de músculo esquelético.

29
Figura 5. -Plasticidade das células-tronco de adulto
Pesquisas estão tentando determinar os mecanismos que conferem plasticidade às
células-tronco de adulto. Se tais mecanismos forem identificados e controlados, as células existentes em um tecido sadio podem ser induzidas a repopular e reparar um tecido doente. PERGUNTAS CHAVE SOBRE CÉLULAS-TRONCO DO ADULTO E CÉLULAS-TRONCO EMBRIONÁRIAs Questões importantes sobre as células-tronco permanecem sem resposta. Entre elas estão as que se seguem:
• Quantos tipos de células-tronco do adulto existem e em quais tecidos elas existem?
• Quais são as fontes de células-tronco do adulto no corpo? Serão elas células-tronco “remanescentes” das células-tronco embrionárias, ou elas são originadas de outro modo? Por que elas permanecem num estado indiferenciado quando as células ao seu redor foram diferenciadas?
• As células-tronco do adulto exibem plasticidade normalmente, ou elas se transdiferenciam apenas quando manipuladas pelos cientistas? Quais são os sinais que regulam a proliferação e a diferenciação das células-tronco que exibem plasticidade?
• É possível manipular células-tronco do adulto para aumentar sua proliferação de modo a produzir células suficientes para um transplante?
• Existe um tipo de célula-tronco de adulto que tenha a capacidade de gerar as células de todos os tecidos e órgãos?
• Quais os fatores que estimulam as células-tronco a migrarem para locais lesionados ou danificados?

30
SEMELHANÇAS ENTRE CÉLULAS-TRONCO EMBRIONÁRIAS E CÉLULAS-TRONCO DO ADULTO
Tanto as células-tronco embrionárias humanas como as células-tronco do adulto apresentam vantagens e desvantagens com relação ao seu potencial de uso em terapia celular regenerativa. É claro que as células-tronco embrionárias são pluripotentes enquanto as células-tronco do adulto são multipotentes. Assim sendo, as células-tronco do adulto são geralmente limitadas a diferenciar nos tipos de células diferentes presentes em seus tecidos ou órgãos. Entretanto, há evidencias de plasticidade, como comentado acima, aumentando assim o número de tipos celulares que elas podem originar. Grande número de células-tronco embrionárias pode ser obtido em cultura, enquanto as células-tronco de adulto são raras nos tecidos e os métodos para expandir seu número em cultura ainda não foram desenvolvidos. Essa é uma diferença importante, pois um grande número de células é necessário para as terapias regenerativas. Uma vantagem potencial do uso de células-tronco do adulto é que as células do próprio paciente podem ser expandidas e reintroduzidas. Isso significa que não haverá problemas de rejeição das células transplantadas. As células-tronco embrionárias quando injetadas em um paciente podem causar rejeição. Entretanto, ainda não foi determinado se células-tronco embrionárias humanas podem causar algum tipo de rejeição. POSSIBILIDADES DE UTILIZAÇÃO DE CÉLULAS-TRONCO HUMANAS E OS OBSTÁCULOS A SEREM VENCIDOS PARA VIABILIZAR SEU USO EM TERAPIA
Há vários entraves de ordem técnica que precisam ser vencidos para que as
células-tronco possam passar a ser empregadas rotineiramente em terapia celular. Um dos principais objetivos das pesquisas com células-tronco embrionárias humanas é a identificação de como as células indiferenciadas tornam-se diferenciadas. Os cientistas sabem ligar ou desligar determinados genes é um processo crucial. Algumas das mais sérias condições médicas como câncer e defeitos congênitos são devidos a anormalidades na divisão celular anormal e na diferenciação. Uma melhor compreensão do controle genética e molecular desses processos pode dar informações sobre como tais doenças ocorrem e sugerir novas estratégias para terapia. Os cientistas ainda não compreendem completamente os sinais que ligam ou desligam genes na diferenciação das células-tronco. Uma potencial aplicação das células-tronco é a geração de órgão e tecidos para substituir tecidos lesados e que atualmente só é possível a partir de doação de órgãos de pessoas com morte cerebral.
Para realizar as promessas de uso, os cientistas devem ser capazes de reproduzir, manipular e diferenciar as células em número suficiente para os transplantes. A seguinte lista de passos precisa ser obtida:
• Proliferar extensivamente e gerar quantidades suficientes de tecido, • Diferenciar as células no tipo celular desejado, • Garantir a sobrevivência das células no corpo do transplantado, após o
transplante, • Garantir a integração das células transplantadas no tecido do receptor, • Garantir o correto funcionamento das células durante o período de vida do
transplantado, • Evitar qualquer tipo de dano no transplantado, inclusive rejeição.

31
CLONAGEM
Mayana Zatz
A clonagem é um mecanismo comum de propagação da espécie em plantas ou bactérias. Em humanos, os clones naturais são os gêmeos idênticos que se originam da divisão de um mesmo óvulo fertilizado. A grande revolução da Dolly abriu caminho para a possibilidade de clonagem humana ao demonstrar, pela primeira vez, que era possível clonar um mamífero - isto é, produzir uma cópia geneticamente idêntica, a partir de uma célula somática diferenciada. Para entendermos porque esta experiência foi surpreendente precisamos recordar um pouco de embriologia.
Todos nós já fomos uma célula única, resultante da fusão de um óvulo e de um espermatozóide. Esta primeira célula já tem no núcleo o DNA com toda a informação genética necessária para o novo ser. No núcleo das nossas células, o DNA se organiza em pares de cromossomos e apresenta-se muito condensado. Com exceção das células sexuais — o óvulo e o espermatozóide, que têm 23 cromossomos —, em todas as outras células do corpo humano há 46 cromossomos (23 pares) em seus núcleos. As células do corpo, não sexuais, são as chamadas células somáticas.
A grande contribuição da clonagem da Dolly foi justamente a descoberta que uma célula somática de mamífero, já diferenciada, poderia ser reprogramada ao estágio inicial e voltar a ser totipotente. Os cientistas escoceses realizaram isso ao transferirem o núcleo de uma célula somática da glândula mamária de uma ovelha para um óvulo enucleado — quer dizer, de onde tinham retirado o núcleo. Surpreendentemente, o óvulo começou a comportar-se como um óvulo recém-fecundado por um espermatozóide. Clonagem reprodutiva
Para obtenção de um clone, o óvulo para o qual os cientistas transferiram o núcleo da célula somática foi inserido no útero de uma outra ovelha. Se desejássemos fazer clonagem humana reprodutiva, teríamos que retirar o núcleo de uma célula somática que, teoricamente, poderia ser de qualquer tecido de uma criança ou adulto, inserir o núcleo em um óvulo, e depois implantá-lo no útero de uma mulher, que funcionaria como barriga de aluguel. Se o óvulo se desenvolvesse teríamos um novo ser com as mesmas características físicas da criança ou adulto de quem foi retirada a célula somática. Seria como um gêmeo idêntico nascido posteriormente (Figura 6).
Já sabemos que esse processo não é fácil. Dolly só nasceu depois de 276 tentativas fracassadas. Além disso, dentre os núcleos das 277 células da “mãe de Dolly“ inseridos em um óvulo sem núcleo, 90% não alcançaram nem o estágio de blastocisto. As tentativas posteriores de clonar outros mamíferos, camundongos, porcos, bezerros, um cavalo e um veado, também mostram eficiência muito baixa e uma proporção muito grande de abortos e embriões malformados. Penta, a primeira bezerra brasileira clonada a partir de uma célula somática adulta, em 2002, morreu com um pouco mais de um mês. Ainda em 2002, foi anunciada a clonagem do “copycat”, o primeiro gato de estimação clonado a partir de uma célula somática adulta. Para chegar a isso, foram utilizados 188 óvulos que geraram 87 embriões e apenas um animal vivo. Na realidade, experiências recentes, com diferentes modelos animais, têm mostrado que a reprogramação da célula somática para um estágio embrionário, indiferenciado, que originou Dolly, é extremamente difícil.
Ian Wilmut, o cientista escocês famoso pela experiência que resultou no nascimento de Dolly, afirma que praticamente todos os animais clonados nos últimos anos a partir de células não-embrionárias estão com problemas (Rhind, 2003). Entre os diferentes defeitos dos pouquíssimos animais que nasceram vivos após inúmeras tentativas, observaram-se: placentas anormais, gigantismo em ovelhas, defeitos cardíacos em porcos, problemas pulmonares em vacas, ovelhas e porcos, problemas imunológicos, falha na produção de leucócitos, defeitos musculares em carneiros. De acordo com

32
Hochedlinger e Jaenisch (2003), os avanços recentes em clonagem reprodutiva permitem quatro conclusões importantes:
• A maioria dos clones morre no início da gestação; • os animais clonados têm defeitos e anormalidades semelhantes
independentemente da célula doadora ou da espécie; • essas anormalidades provavelmente ocorrem por falhas na reprogramação do
genoma das células somáticas; • a eficiência da clonagem depende do estágio de diferenciação da célula doadora.
De fato, a clonagem reprodutiva a partir de células embrionárias têm mostrado uma
eficiência de 10 a 20 vezes maior, provavelmente porque os genes importantes no início da embriogênese estão ainda ativos no genoma da célula doadora. (Hochedlinger e Jaenisch, 2003).
Entre todos os mamíferos já clonados, é interessante notar que a eficiência é um pouco maior em bezerros. Outro fato intrigante é não se ter notícias de macaco que tenha sido clonado. Talvez por essa razão, a cientista inglesa Ann McLaren afirma que as falhas na reprogramação do núcleo somático podem vir a se constituir em barreira intransponível para a clonagem humana. Mesmo assim, pessoas como o médico italiano Antinori e a seita dos raelianos defendem a clonagem humana, um procedimento que tem sido proibido em todos os países. De fato, em 2003, as academias de ciências de 63 países, inclusive a brasileira, formalizaram documento conjunto em que pedem o banimento da clonagem reprodutiva humana. O fato é que a simples possibilidade de clonar humanos suscita discussões éticas em todos os segmentos da sociedade: Por que clonar? Quem deveria ser clonado? Quem iria decidir? Quem será o pai ou a mãe do clone? O que fazer com os clones que nascerem defeituosos?
Na realidade o maior problema ético atual é o enorme risco biológico associado à clonagem reprodutiva.
Apesar de todos estes argumentos contra a clonagem humana reprodutiva, as experiências com animais clonados têm nos ensinado muito acerca do funcionamento celular. Por outro lado, a tecnologia de transferência de núcleo para fins terapêuticos, a chamada clonagem terapêutica, poderá ser extremamente útil para obtenção de células-tronco embrionárias. A técnica de clonagem terapêutica para obtenção de células-tronco embrionárias
Se tivermos um óvulo humano cujo núcleo foi substituído por um núcleo de célula somática e deixarmos que se divida no laboratório – não em um útero -, teríamos a possibilidade teórica de obter blastocistos e usar suas células-tronco embrionárias pluripotentes para formar diferentes células (Figura 7). Isso já foi feito em animais. Com isso, abrir-se-iam perspectivas fantásticas para futuros tratamentos. Hoje, só se consegue cultivar em laboratório células com as mesmas características do tecido de onde foram retiradas ou transformá-las em poucos tipos celulares. É importante esclarecer que, na clonagem para fins terapêuticos, os tecidos seriam produzidos apenas em laboratório. Não se trata de clonar um feto até alguns meses dentro do útero para depois lhe retirar os órgãos para transplante, como alguns acreditam.

33
Figura 6. Clonagem reprodutiva
.
Figura 7. Clonagem terapêutica

34
Aspectos éticos
As 63 academias de ciência do mundo que se posicionaram contra a clonagem reprodutiva defendem as pesquisas com células embrionárias para fins terapêuticos. Em relação aos que acham que a clonagem terapêutica pode abrir caminho para clonagem reprodutiva, devemos lembrar que existe uma diferença fundamental entre os dois procedimentos: a implantação ou não em um útero humano. Bastaria simplesmente proibir a implantação no útero para conter os abusos!
A cultura de tecidos é uma prática comum em laboratório. É realizada a partir de diversos tipos de células, sem dilemas éticos. No caso da clonagem terapêutica, a diferença seria o início da cultura a partir de óvulos que permitiriam a produção de qualquer tecido no laboratório. Ou seja: ao invés de poder produzir-se apenas um tipo de tecido, já especializado, o uso de óvulos permitiria fabricar qualquer tipo de tecido. Em relação ao risco de comércio de óvulos, esse risco seria o equivalente ao que ocorre hoje com transplante de órgãos.
Em relação ao problema da destruição de “embriões humanos”, novamente devemos lembrar que estamos falando de cultivar tecidos - ou, futuramente, órgãos -, a partir de embriões e que esses nunca serão inseridos em um útero. Sabemos que 90% dos embriões gerados em clínicas de fertilização e inseridos no útero de uma mulher não geram vida. Além disso, um trabalho recente (Mitalipova et al., 2003) mostrou que células obtidas de embriões de má qualidade, que não teriam potencial para gerar uma vida, mantêm a capacidade de gerar linhagens de células-tronco embrionárias em laboratório e, portanto, de gerar tecidos. Ao usar células-tronco embrionárias com potencial baixíssimo de gerar indivíduos para regenerar tecidos em uma pessoa condenada por uma doença letal, poderíamos interpretar que na realidade estamos criando vida. Isso é comparável ao que se faz hoje em transplante quando se retira órgãos de uma pessoa com morte cerebral (mas que poderia permanecer em vida artificialmente mantida por mais tempo).
É extremamente importante que as pessoas entendam a diferença entre clonagem reprodutiva humana, clonagem terapêutica e terapia celular com células-tronco embrionárias antes de tomar posição. Por outro lado, também não podemos acreditar que as células-tronco vão curar todas as doenças humanas. As pesquisas que estão iniciando agora serão fundamentais para responder a questões sobre o potencial das células-tronco adultas em comparação com as embrionárias, que doenças poderão ser tratadas e quais são os benefícios e riscos da terapia celular. Situação brasileira
Com a aprovação pela Câmara dos Deputados da Lei de Biossegurança, no dia 2 de março de 2005, o Brasil entra na seleta lista de países do mundo em que os cientistas podem realizar pesquisas com células-tronco embrionárias e trabalhar para encontrar tratamentos para doenças genéticas até hoje incuráveis e para lesões físicas ainda irreversíveis. Em alguns casos, as células-tronco embrionárias são a única esperança.
Os cientistas apostam muito nessas células embrionárias, pois elas são as únicas capazes de produzir todos os 216 tecidos do nosso corpo. A esperança é que inúmeras doenças, entre elas as neuromusculares, o diabetes, o mal de Parkinson e as lesões de medula possam ser tratadas pela substituição ou correção de células ou tecidos defeituosos. A terapia celular com células-tronco representa também um grande avanço nas técnicas existentes hoje de transplante de órgãos. Se as pesquisas derem os resultados esperados, deverá ser possível no futuro fabricar tecidos e órgãos em quantidade suficiente para todos. Seria o fim das longas filas de transplante de órgãos. Do mesmo modo que trocamos peças do nosso carro poderemos substituir ou corrigir a função de órgãos com defeitos. Mas para chegar lá, ainda temos que pesquisar e estudar muito.

35
As pesquisas com células-tronco do adulto, por sua vez, já foram iniciadas em pacientes cardíacos e em outras doenças como esclerose múltipla, acidente vascular ou diabetes, a maior pesquisa do mundo com pacientes cardíacos está sendo realizada no Brasil com 1.200 pessoas. Mas essas células têm algumas limitações. Hoje, elas só podem ser transformadas em células de alguns dos tecidos do corpo. Em especial, os pesquisadores sabem como transformar as células-tronco do adulto em células dos órgãos ou tecidos de onde foram retiradas: por exemplo, em células da medula óssea, que produz os componentes básicos do sangue. A terapia com células-tronco do adulto têm dado bons resultados no tratamento de leucemia. Nele, células-tronco do adulto da medula óssea e mais recentemente do cordão umbilical e da placenta, são transplantadas nos pacientes a partir de doadores compatíveis. Outra técnica utilizada ainda experimentalmente é a de auto-transplante na qual as células-tronco são retiradas e reinjetadas no paciente para o tratamento de lesões cardíacas e na recuperação do tecido nervoso de pessoas que sofreram acidentes vasculares. Mas ninguém sabe ainda se o tratamento é eficiente - por enquanto, é uma tentativa terapêutica experimental. A má notícia é que o auto-transplante não pode resolver o problema dos mais de 5 milhões de brasileiros portadores de doenças genéticas, pois o defeito está presente em todas as suas células. Para essas pessoas talvez seja necessário o uso de células-tronco obtidas de embriões. Referências: Hochedlinger K, Jaenish R (2003): Nuclear transplantation, embryonic stem cells and the potential
for cell therapy. N. Engl. Journal of Medicine 349:275-212 Mitalipova M, Calhoun J, Shin S, Wininger D et al. (2003): Human embryonic stem cells lines
derived from discarded embryos. Stem cells 21:521-526
Rhind SM, Taylor JE, De Sousa PA, King TUI, McGarry M, Wilmut I (2003): Human Cloning: can it be made safe? Nature reviews 4:855-864

36
REPROGRAMAÇÃO CELULAR Fernando Nodari
Ao longo do desenvolvimento de um embrião, suas células vão se comprometendo
e diferenciando, o que as tornam cada vez mais especializadas e cada vez menos potentes. Esses processos de desenvolvimento e diferenciação celular são acompanhados por modificações epigenéticas no DNA da célula. Essas modificações não alteram a seqüência de bases do DNA, mas são transmitidas para as células filhas após a mitose, e fazem com que genes específicos sejam expressos ou reprimidos. Como conseqüência das modificações epigenéticas cada linhagem celular tem um conjunto específico de genes expressos e reprimidos.
Aparentemente esse processo de diferenciação é irreversível, a não ser através de modificações experimentais. Fazer com que uma célula diferenciada ou comprometida volte alguns passos do caminho da diferenciação e se desdiferencie é chamado reprogramação celular. Para que isso ocorra não basta apagar as modificações epigenéticas de uma célula, é necessário, também, refazer as modificações morfológicas e funcionais características de uma célula desdiferenciada.
Dois tipos de experimentos tentam entender a reprogramação celular: a) experimentos de clonagem e b) experimentos que testam a plasticidade celular. Experimentos de clonagem
Para realizar uma clonagem são necessárias duas células: um ovócito e uma célula diferenciada como, por exemplo, uma célula diplóide da pele. A célula diferenciada é induzida a se manter no estágio G0 (Fig. 8), seu núcleo é removido e implantado em um ovócito previamente anucleado. Em seguida são aplicados dois estímulos: um deles (choque elétrico, no caso de ovelhas, e meio de cultura com estrôncio, no caso de ratos) induz a fusão do núcleo com o citoplasma do ovócito e o outro (estímulo químico) ativa o ovócito. Tais estímulos provocam a reprogramação do núcleo, ou seja, ele se desdiferencia e se o processo de reprogramação ocorrer de modo correto a célula torna-se totipotente. O genoma dessa nova célula é idêntico ao da célula diferenciada, doadora do núcleo. A eficiência da clonagem está diretamente relacionada ao estado de diferenciação da célula doadora do núcleo.
O que diferencia os dois tipos de clonagem, reprodutiva ou terapêutica, é o destino dado à nova célula. Na clonagem terapêutica, ela é induzida, em laboratório, a diferenciar-se em um tipo celular especifico, tecido ou órgão a ser usado em terapia celular. Por exemplo, um paciente portador de mal de Alzheimer poderia doar células diferenciadas de pele para que, em laboratório, elas fossem diferenciadas em células do sistema nervoso e em seguida fossem implantadas em seu cérebro. Esse tipo de terapia ainda não é realizado atualmente.
Na clonagem reprodutiva, essa nova célula é implantada num útero de uma mãe de aluguel para dar origem a um organismo com genoma idêntico ao do doador da célula diferenciada. Há barreiras técnicas na clonagem reprodutiva. Uma delas é a ausência das modificações epigenéticas desencadeadas pela maturação do ovócito e do espermatozóide e pela fertilização do ovócito que preparam o genoma dos gametas para o início correto do desenvolvimento embrionário. Em outras palavras, quanto mais diferenciada for a célula doadora do núcleo, menor será a eficiência de clonagem.
Dependendo da qualidade do processo de reprogramação três resultados são possíveis: 1. a ausência de reprogramação do genoma resulta na morte imediata do embrião; 2. a reprogramação parcial permite a sobrevivência dos clones por um certo período, porém resulta em fenótipos anormais e/ou letais em vários estágios do desenvolvimento; 3. a ocorrência de reprogramação completa, resulta em animais completamente normais. Os fenótipos de embriões clonados observados até hoje sugerem que a reprogramação completa é a exceção. Para aumentar a porcentagem de sucesso da clonagem reprodutiva é necessário tratar separadamente os dois genomas

37
parentais para induzir as modificações epigenéticas estabelecidas durante a gametogênese. Atualmente, não é possível realizar tais modificações.
As dificuldades técnicas acima mencionadas nos levam a duas perguntas muito distintas:
1. Falhas na reprogramação celular podem interferir na clonagem terapêutica? A célula-tronco embrionária clonada para fins terapêuticos não precisa ter a habilidade de refazer todo o desenvolvimento fetal e produzir um organismo completo, ela precisa, apenas, ser capaz de produzir, em laboratório, um tipo celular, um tecido ou um órgão. Assim sendo, as células cuja reprogramação não foi adequada são automaticamente descartadas caso não conseguem seguir adiante o processo de diferenciação desejado.
2. Um embrião clonado é equivalente a um embrião fertilizado? No embrião originado por clonagem o conteúdo genético é derivado de um único indivíduo e, assim sendo, esse embrião é produto de tecnologia laboratorial e não de um evento natural. Por essa razão, há pesquisadores que defendem o ponto de vista de que não há criação de uma nova vida, mas sim apenas a propagação de uma vida existente, pois não ocorreu meiose, troca genética ou concepção. Experimentos que testam a plasticidade celular
Uma das possibilidades de testar a plasticidade de células é por meio da fusão de células diferenciadas com células-tronco embrionárias. Nesse caso espera-se que fatores presentes na célula-tronco embrionária induzam a desdiferenciação do núcleo diferenciado. Esse processo produz células com características de célula-tronco embrionária, porém, tetraplóides (4n).
Em outra linha de pesquisa cultivam-se células diferenciadas em meio de cultura apropriado para células-tronco embrionárias e adicionam-se alguns dos fatores de transcrição responsáveis por manter uma célula-tronco indiferenciada. Esse procedimento induz as células diferenciadas a desdiferenciar. Essas células são capazes de produzir células das três camadas germinativas (ectoderma, mesoderma e endoderma), porém, de forma desordenada e sem controle.
Em uma terceira linha de pesquisa tenta-se demonstrar que células-tronco adultas presentes nos tecidos originados de um determinado folheto embrionário (ectoderma, por exemplo), sob condições específicas de cultura ou através de implantação em diferentes tecidos, seriam capazes de produzir células diferenciadas de um folheto embrionário diferente, além do seu próprio, Ou seja, elas passariam de pluripotentes a multipotentes.
Todas as pesquisas citadas nos itens “a” e “b” tentam entender quais são os fatores envolvidos no processo de desdiferenciação e como eles atuam, ou seja, tentam compreender como modificações epigenéticas no DNA são feitas e desfeitas. Com isso, será possível produzir células-tronco pluripotentes ou totipotentes a partir de células que podem ser encontradas no organismo adulto e utilizar essas células desdiferenciadas nas terapias em vez das polêmicas células tronco embrionárias. Porém, muitos cientistas dizem que ainda não há evidências de que as células-tronco adultas possam ser tão versáteis quanto as embrionárias.
Apesar dos cientistas já terem conseguido clonar algumas espécies de animais (exemplo, ovelhas, ratos e vacas) e já terem conseguido produzir células desdiferenciadas em laboratório, os mecanismos moleculares responsáveis pelo processo de desdiferenciação ainda são desconhecidos.Há indicações de que RNAs mensageiros e/ou proteínas citoplasmáticas (ex.: fatores de transcrição) exerçam um papel importante na reprogramação do núcleo celular.
Há, porém, muitas dificuldades de ordem técnica nesses tipos de experimentos, com isso, são necessárias análises cuidadosas dos resultados, por exemplo, atualmente são exigidas evidências de desdiferenciação (expressão de genes relacionados a pluripotência, repressão da expressão de genes característicos de uma célula diferenciada, capacidade de auto-renovação e plasticidade) para comprová-la. Isso faz com que os resultados surjam lentamente, porém, de forma confiável.

38
Até o momento essas pesquisas produziram muitos resultados promissores, porém, ainda serão necessários muitos anos para que consigamos entender como podemos reprogramar uma célula e mudar seu destino.
Figura 8: Fases do ciclo de vida celular.

39
CÉLULAS-TRONCO: PROGRESSOS CIENTÍFICOS E O FUTURO DAS PESQUISAS
Silvio Higa
O que se sabe a respeito de células-tronco embrionárias (pluripotentes)? • São obtidas de blastocisto pré-implantacionais, cerca de 5 dias após a fertilização. • Já é possível cultivar estas células in vitro por mais de dois anos sem que haja
diferenciação. • O gene da enzima telomerase está ativo o que possibilita a manutenção de longos
telômeros, isto garante a capacidade de replicação por muitas gerações. • Não possuem cromatina sexual, ou seja, ainda não ocorreu a inativação de um
dos cromossomos X.
O que se sabe a respeito de células-tronco adultas? • Podem ser encontradas no cérebro, medulas ósseas, sangue periférico, músculos
esqueléticos, tecido adiposo, tecido epitelial da pele e do tubo digestório, córnea, polpa dentária, retina, fígado e pâncreas. Portanto, são encontradas em tecidos que tem origem a partir dos três folhetos embrionários (ectoderma, mesoderma e endoderma).
• Não há evidencias de que sejam pluripotentes. • São raras. Geralmente são difíceis de serem identificadas, isoladas e purificadas.
Estima-se que na medula óssea apenas 1 em cada 10.000 ou 15.000 células seja uma célula-tronco.
• O método usado para identificá-las depende da determinação dos marcadores de superfície celular e observação de padrões de diferenciação em meios de cultura.
• Não proliferarem in vitro, por um longo período de tempo, como o fazem in vivo. • A melhor estudada até hoje é a proveniente da medula óssea: célula-tronco
hematopoiética que possui capacidade restrita de proliferar e de se manter indiferenciada em meio de cultura. Estes são os dois principais fatores que limitam seu uso em pesquisas e transplantes.
• Parece não haver diferenças qualitativas entre células-tronco adultas obtidas de medula óssea, cordão umbilical e sangue periférico.
• Algumas parecem possuir plasticidade, ou seja, capacidade de se diferenciar em células de outro tecido diferente daquele do qual se originou. Por exemplo, já foi demonstrado em humanos e ratos: células-tronco da medula óssea deram origem a neurônios, oligodentrócitos e astrócitos.
• Já foi demonstrado em ratos que uma única célula-tronco hematopoiética é capaz de reconstituir toda a medula óssea após a mesma ter sido destruída por radiação.
• É difícil - se não impossível - distinguir células-tronco adultas de células progenitoras (célula-tronco progenitora especializada).
Comparação entre células-tronco embrionárias e adultas O que elas têm em comum?
• Possuem capacidade de auto-renovação e de dar origem a células especializadas. • Os cientistas usam técnicas similares para marcar e monitorar a expressão de
certos genes afim de identificá-las. • São capazes de proliferar e de se especializar quando transplantadas para
animais cujo sistema imunológico foi suprimido. O que elas têm de diferente?
• A principal diferença está na origem, os cientistas acreditam que as embrionárias existam apenas nos embriões e que as adultas estão presentes em diversos tipos de tecidos do corpo humano.

40
• As embrionárias são pluripotentes, ou seja, podem dar origem a tecidos provenientes dos três folhetos germinativos (ectoderma, mesoderma e endoderme). Ainda não se sabe se as adultas possuem a mesma capacidade.
• Em laboratório, as embrionárias podem se multiplicar por muitas gerações sem que haja diferenciação; já as adultas, sofrem diferenciação.
• As embrionárias, quando injetadas em cobaia cujo sistema imunológico foi suprimido, geram teratomas (mistura de diferentes tipos celulares). O mesmo resultado não é observado com as adultas.
Quais são as perguntas que ainda precisam ser respondidas a respeito de células-tronco?
• Existe uma céula-tronco universal? Ou seja, existe uma célula-tronco (talvez circulando no sangue) que possa gerar células de quaisquer órgãos ou tecidos?
• Quais são as origens das células-tronco no adulto? Elas são "sobras" de células-tronco embrionárias ou elas surgem de alguma outra forma? E se a última alternativa for a verdadeira - como parece ser - exatamente como elas surgem e como elas permanecem em um estado indiferenciado enquanto todas as células ao seu redor se encontram diferenciadas?
• Quantos tipos de células-tronco adultas existem e em quais tecidos? • Células-tronco adultas podem proliferar em meio de cultura até que seja obtida a
quantidade necessária para transplante? • Quais as evidências de que células especializadas geradas a partir de transplantes
de células-tronco possam substituir células de tecidos lesados ou danificados? • A plasticidade apresentada por células-tronco adultas in vitro também ocorre in
vivo? Quais são os sinais que regulam a proliferação e diferenciação das células-tronco que apresentam plasticidade?
• Quais são os fatores responsáveis pela migração das células-tronco até os tecidos danificados?
• Quais são os controles intrínsecos que fazem uma célula-tronco se diferenciar em determinado tipo celular ao invés de outro?
• Quais os mecanismos que permitem as células-tronco embrionárias proliferarem in vitro sem que haja diferenciação?
• A partir do conhecimento dos mecanismos que regulam a especialização de células-tronco embrionárias poderão os cientistas a controlar o mesmo processo em células-tronco adultas?
• Culturas de células-tronco embrionárias são de fato homogêneas e indiferenciadas ou elas apenas parecem ser?
• Toda célula-tronco pluripotente passa pelo estágio de célula-progenitora enquanto se especializa? Em caso afirmativo, a célula progenitora poderia ser usada para transplante?
• Qual o estágio de diferenciação da célula-tronco é o melhor para o transplante? O mesmo estágio é o melhor para qualquer tipo de transplante ou varia de caso para caso?
• Qual o melhor estágio de diferenciação de uma célula-tronco para se testar drogas e toxinas?
Potenciais usos das células-tronco embrionárias
• Transplante para repor ou restaurar tecidos que foram danificados por doenças ou acidentes. Exemplos: lesões na medula espinhal, problemas cardíacos, mal de Parkinson's, diabetes, distrofia muscular Duchenne, câncer, entre outras.
• Através das técnicas de transferência nuclear poderiam ser reprogramadas para ficarem idênticas às células do receptor, evitando, assim, a rejeição.
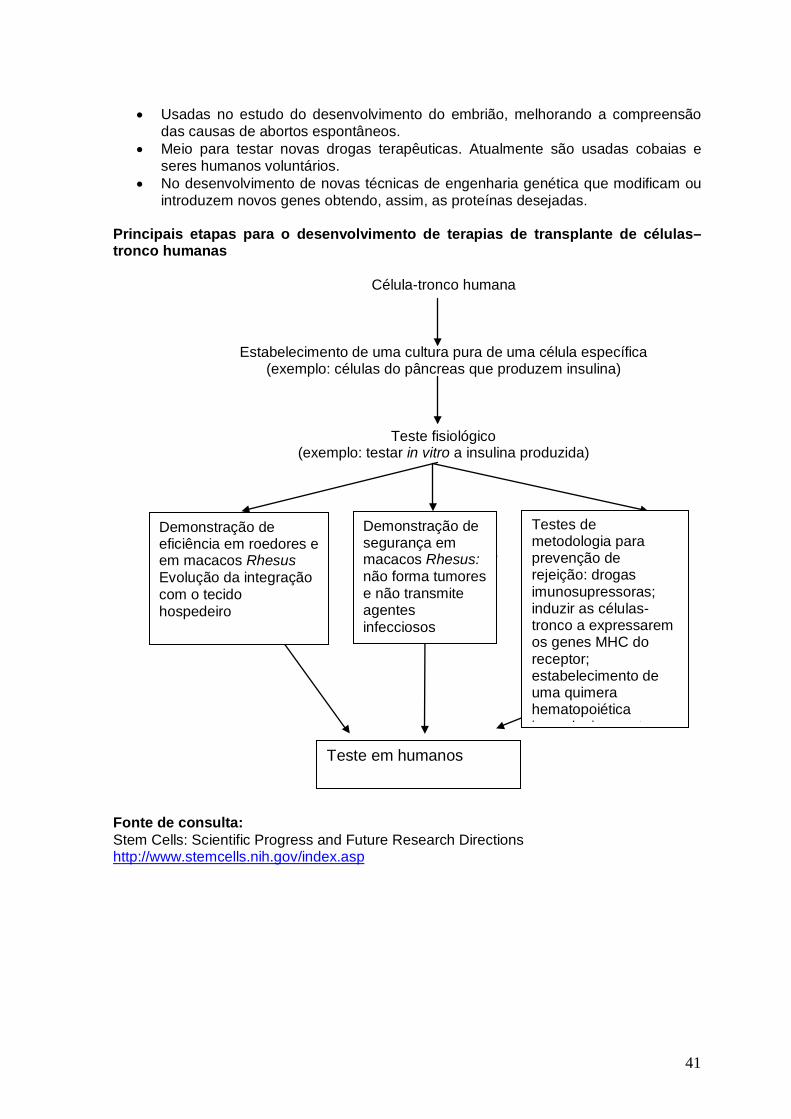
41
• Usadas no estudo do desenvolvimento do embrião, melhorando a compreensão das causas de abortos espontâneos.
• Meio para testar novas drogas terapêuticas. Atualmente são usadas cobaias e seres humanos voluntários.
• No desenvolvimento de novas técnicas de engenharia genética que modificam ou introduzem novos genes obtendo, assim, as proteínas desejadas.
Principais etapas para o desenvolvimento de terapias de transplante de células–tronco humanas
Célula-tronco humana
Estabelecimento de uma cultura pura de uma célula específica (exemplo: células do pâncreas que produzem insulina)
Teste fisiológico (exemplo: testar in vitro a insulina produzida)
Teste em humanos
Fonte de consulta: Stem Cells: Scientific Progress and Future Research Directions http://www.stemcells.nih.gov/index.asp
Demonstração de eficiência em roedores e em macacos Rhesus Evolução da integração com o tecido hospedeiro
Demonstração de segurança em macacos Rhesus: não forma tumores e não transmite agentes infecciosos
Testes de metodologia para prevenção de rejeição: drogas imunosupressoras; induzir as células-tronco a expressarem os genes MHC do receptor; estabelecimento de uma quimera hematopoiética i l i t
Teste em humanos

42
TERAPIA CELULAR - O USO DE CÉLULAS-TRONCO NO TRATAMENTO DE DOENÇAS: ETAPAS E QUESTÕES GERADAS
Antes que um novo procedimento de terapia comece a ser utilizado no tratamento de doentes são necessários os seguintes requisitos:
• uma idéia que possa ser testada com base em experimentos científicos – no caso da terapia celular, a idéia é usar células-tronco para reparar ou substituir tecidos ou órgãos danificados.
• Pesquisas e testes em laboratório – geralmente, os experimentos preliminares são feitos em placas de cultura com células humanas e de outros animais. Essa etapa inclui o desenvolvimento de diferentes linhagens de células-tronco em laboratório, a verificação da eficácia que cada tipo celular apresenta no reparo de diversos tecidos, etc.
• Estabelecimento de um modelo experimental que simula o modo de utilização em seres humanos - modelos de terapia mais refinados são testados em modelos animais. Os resultados devem ser reprodutíveis e a segurança de sua aplicação deve ser provada.
Para cada nova aplicação da técnica de terapia celular devem ser considerados:
• Quais são os benefícios? • Quais são os riscos? • Quem será beneficiado? Há potenciais riscos na aplicação? • O que significa para mim? Para minha família? Para meus conhecidos? • Por que outras pessoas podem ter opiniões diferentes das minhas?
Há três tipos de questões que devem ser consideradas quando nós pensamos em clonagem terapêutica:
• Questões éticas são aquelas que nos convidam a pensar sobre as potenciais conseqüências originadas por um determinado procedimento ou pelo emprego de alguma metodologia.
• Questões legais – tanto o público como as pessoas que fazem as leis precisam da ajuda dos cientistas para entender a técnica, seu significado e como o seu emprego deve ser regulamentado pelo governo.
• Questões sociais – dizem respeito ao impacto da aplicação da técnica na sociedade.
Link para o vídeo Globo News http://www.video.google.com/videoplay?docid=8414495255301311833

43
A POLÊMICA DAS CÉLULAS-TRONCO EMBRIONÁRIAS Trechos do livro Clonagem da ovelha Dolly às células-tronco
Lygia da Veiga Pereira. Editora Moderna, 2a
edição, reformulada, páginas 77 a 88, 2005
Apesar de tão promissoras como forma de terapia, as células-tronco embrionárias foram centro de enorme polêmica nos Estados Unidos, onde em 2001 foi proibido o desenvolvimento de novas linhagens dessas células utilizando-se verba do governo federal. Aqui no Brasil, até 2004, as pesquisas com embriões humanos não eram permitidas. Nesse ano foi criado um projeto de Lei de Biossegurança que proibia “o uso de embriões humanos como material biológico disponível”, impedindo o desenvolvimento das pesquisas com células-tronco embrionárias no país. Esse projeto de Lei foi muito criticado por várias instituições científicas, incluindo a Academia Brasileira de Ciências, a Sociedade Brasileira para o Progresso da Ciência (SBPC) e a Fundação de Amparo à Pesquisa do Estado de São Paulo (Fapesp), o que levou à criação no Senado Federal de uma emenda que permitiria o uso para a pesquisa de embriões congelados a mais de três anos. Depois de muita discussão envolvendo a sociedade, cientistas, grupos religiosos e associações de docentes, em março de 2005 esse projeto de Lei foi aprovado na Câmara dos Deputados e sancionado pelo presidente da república Luís Inácio Lula da Silva. A partir desse momento, ficou permitida no Brasil pesquisa com embriões humanos congelados a mais de três anos para extração de células-tronco embrionárias, mas a clonagem terapêutica foi proibida. Mas por que tanta polêmica em torno das células-tronco embrionárias? O grande problema é que essas células são extraídas de embriões humanos, produzidos por clonagem terapêutica ou aqueles excedentes de processos de fertilização in vitro. Durante um ciclo de fertilização invitro são gerados de 5 a 8 embriões. Desses, 2 ou 3 são transferidos para o útero da mulher. Os embriões restantes devem ser congelados para serem utilizados em uma futura tentativa de gravidez. Porém, muitas vezes o casal não quer mais engravidar, e esses embriões ficam “esquecidos” nos congeladores. Com o passar do tempo, a capacidade dos embriões congelados darem origem a um bebê vai diminuindo. São justamente esses embriões que são destruídos para se extrair as células-tronco embrionárias.Para algumas pessoas isso equivale a matar uma pessoa e por isso é inaceitável. Essa é uma questão delicada, que envolve aspectos legais, éticos, culturais e religiosos. Vale lembrar que estamos falando de um embrião de cinco dias, basicamente um conglomerado amorfo de células. Um embrião gerado no ventre de uma mulher nesse estágio do desenvolvimento possui somente 20% de chance de se implantar e se desenvolver em um bebê. Se isso já caracteriza uma vida ou não, não sei, mas é importante fazer uma distinção entre embrião e um aborto de um feto de 3-4 meses. Pela cultura judaica, por exemplo, a vida começa depois de o embrião se implantar no útero materno, e dessa forma não há nenhum problema em utilizar esses embriões para produzir células-tronco embrionárias. A definição de vida é muito complexa, mas uma coisa eu posso garantir: aquele embrião excedente trará muito mais benefícios à vida de pessoas já vivas na forma de células-tronco embrionárias do que no fundo de uma lata de lixo, onde é descartado nas clínicas de reprodução assistida, ou esquecido em um congelador, caso o casal não queira ter mais filhos. Outro argumento contra o uso de células-tronco embrionárias é o medo de que seja criado um “comércio de embriões”. Seguindo essa mesma argumentação, não deveríamos permitir transfusões de sangue nem doações de órgãos, pois também poderia ser criado comércio deles. Com uma legislação séria e vigilância, evitamos o comércio de sangue ou órgãos, permitindo que milhões de vidas sejam salvas com transplantes. Da mesma forma, precisamos discutir a melhor forma de trabalharmos com as células-tronco embrionárias sem ferir direitos nem deveres, mas proporcionando à humanidade uma nova e revolucionária forma de terapia para inúmeras enfermidades.

44
A questão do uso de embriões humanos para a pesquisa é polêmica em todo o mundo, e cada país tem sua própria forma de legislar sobre o assunto. Até 2005, o Reino Unido tinha uma legislação mais liberal, permitindo a pesquisa com embriões humanos gerados especificamente para a extração de células-tronco embrionárias, e também a clonagem terapêutica. Porém, antes de começarem as pesquisas, os grupos interessados devem submeter seus projetos a um órgão federal que fornece licença para estes estudos. Bélgica, Japão, Coréia do Sul também permitem a pesquisa com células-tronco embrionárias e a clonagem terapêutica. Já na França e no Canadá, a clonagem terapêutica é proibida, e pode-se utilizar para pesquisa somente embriões excedentes de clínicas de fertilização. Nos Estados Unidos, as pesquisas com células-tronco embrionárias, incluindo a clonagem terapêutica, são liberadas somente com o uso de dinheiro da iniciativa privada – dinheiro do governo federal não pode ser utilizado nessas pesquisa, o que limita em muito a capacidade da comunidade científica norte-americana de realizar trabalhos nessa área. O impacto da aprovação das pesquisas com embriões humanos no Brasil foi discutido no artigo publicado no jornal o Estado de São Paulo. Esperança por um fio
“Dra.Lygia, com a aprovação do Projeto de Lei de Biossegurança pela Câmara dos
Deputados, quantos pacientes sairão das filas de transplantes?” Gelei com a pergunta feita em entrevista ao vivo, no dia seguinte da aprovação do uso de embriões humanos para a extração de células-tronco embrionárias. Ela sintetiza toda a expectativa que a luta por essa aprovação gerou no último ano. Respirei fundo e respondi: “Nenhum...”. Nenhum hoje, nenhum até mesmo nos próximos anos. Mas quem sabe muitos no longo prazo, agora que podemos trabalhar com células-tronco embrionárias humanas no Brasil. Talvez um certo sensacionalismo faça parte do jogo, e tenha sido importante para mobilizar a sociedade e os parlamentares e levar à aprovação do Projeto de Lei de Biossegurança. Mas agora que a poeira baixou, quais são as reais possibilidades das células-tronco embrionárias?
As células-tronco embrionárias são o tipo mais versátil de células-tronco até hoje identificadas em mamíferos. Enquanto as células-tronco derivadas da medula óssea ou do sangue de cordão umbilical conseguem se transformar em somente alguns tecidos, as células-tronco embrionárias possuem a formidável capacidade de dar origem a todos os tecidos do corpo. Estas células não são uma novidade da ciência – desde a década de 1980 faz-se pesquisas com células-tronco embrionárias de camundongos. Trabalhando com eles, descobrimos como multiplica-las e transforma-las no laboratório em células da medula óssea, do músculo cardíaco, em neurônios, entre outras. E mais: quando transplantadas em animais doentes, estas células derivadas das células-tronco embrionárias foram capazes de aliviar os sintomas de diversas doenças, desde leucemia e doença de Parkinson até paralisia causada por trauma da medula espinhal (daí o entusiasmo do Super-Homem Christopher Reeve em relação a essas células).
Em 1998 surgiram as primeiras linhagens de células-tronco embrionárias humanas, e junto com elas a enorme expectativa de seu uso terapêutico. Porém, antes de começarmos testes clínicos injetando células-tronco embrionárias em seres humanos, temos algumas questões fundamentais que devem ser resolvidas.
A primeira diz respeito à segurança dessas células. Quando injetadas em seu estado nativo em camundongos, as células-tronco embrionárias podem formar teratomas. Assim se injetarmos essas células nos pacientes (seja ele um camundongo ou uma pessoa) temos que primeiro induzi-las no laboratório a se transformar no tipo celular que nos interessa. Caso contrário, no organismo elas se multiplicam e podem se diferenciar descontroladamente formando tumores.
Uma segunda questão importantíssima diz respeito à compatibilidade entre as células-tronco embrionárias e o paciente. Ora, em qualquer transplante é necessário

45
existir uma compatibilidade entre doador e receptor para que o órgão não seja rejeitado. O mesmo deve acontecer com um transplante de células-tronco embrionárias. Como garantir que teremos células-tronco embrionárias compatíveis com todos os pacientes? Uma forma seria criar um banco dessas células, cada uma derivada de um embrião diferente, e torcer para encontrar uma compatível com o paciente. Porém, nossa experiência com bancos de medula óssea demonstrou que isso é extremamente difícil de se conseguir.
Uma alternativa seria então criar células-tronco embrionárias “sob medida”, ou seja, geneticamente idênticas ao paciente. Com as técnicas de clonagem, podemos criar um embrião clonado do paciente, e dele extrair as células-tronco embrionárias. Estas poderiam gerar tecidos 100% compatíveis com o paciente. Essa técnica chama-se clonagem terapêutica, e foi realizada pela primeira vez em seres humanos na Coréia no início de 2004.
É importante ressaltar que, apesar da clonagem terapêutica resolver a questão da compatibilidade das células-tronco embrionárias, infelizmente ela não poderia ser usada em indivíduos com doenças genéticas. As células-tronco embrionárias geradas a partir das células destes pacientes também carregariam a doença, e por isso não seriam capazes de gerar tecidos sadios para transplante. Assim, para o tratamento de doenças genéticas com células-tronco – sejam embrionárias, da medula ou do sangue do cordão -, a melhor alternativa é conseguir um doador aparentado, que tem maior chance de ser compatível com o paciente.
E no Brasil, como andam as pesquisas com células-tronco embrionárias? Em 1999, com o financiamento da Fapesp, nosso grupo estabeleceu as primeiras linhagens de células-tronco embrionárias de camundongo totalmente “made in Brazil”, implantando a tecnologia no país e a disponibilizando para outros grupos de pesquisa. Atualmente, pelo menos cinco grupos trabalham com essas células, estudando sua capacidade de transformação em diferentes tecidos, e já estão capacitados a trabalhar com as células-tronco embrionárias humanas – só dependiam da aprovação da lei de Biossegurança. Provavelmente com toda a discussão em torno destas células, outros grupos de pesquisa se interessarão por trabalhar com elas. Para que estas pesquisas avancem no país, será fundamental um financiamento consistente por parte dos governos estaduais e do governo federal.
Quanto à clonagem terapêutica, a colaboração entre grupos que fazem clonagem animal e aqueles que trabalham com células-tronco embrionárias poderia tornar esta prática uma realidade no país. Porém, como resultado das negociações envolvidas na aprovação do Projeto de Lei de Biossegurança, este proíbe a clonagem terapêutica. Não tem problema, a conquista do direito de utilizar embriões congelados para pesquisa foi um primeiro e importantíssimo passo – quem sabe em uma segunda rodada a clonagem terapêutica possa ser renegociada?
E enquanto não podemos utiliza-las como agente terapêutico temos muito a aprender com as embrionárias. Ao desvendarmos os mecanismos envolvidos em sua capacidade de se transformar em qualquer tipo de célula, aprendemos sobre a biologia do ser humano – esses conhecimentos básicos terão ao longo prazo grandes benefícios à saúde humana.
Em conclusão, o uso terapêutico das células-tronco embrionárias ainda está longe de se tornar uma realidade, tanto no Brasil quanto no mundo. Porém, para que exista alguma chance de isso um dia acontecer, precisamos pesquisar – e foi este direito que adquirimos esta semana, passando de meros observadores do desenvolvimento de uma área promissora da medicina para jogadores muito competitivos. Afinal de contas, as pesquisas com células-tronco embrionárias de medula e de cordão umbilical no Brasil são motivo de orgulho nacional. Agora poderemos fazer o mesmo bonito com as células-tronco embrionárias.
Lygia da Veiga Pereira, O Estado de São Paulo, 6 março de 2005

46
Medicina regenerativa A clonagem terapêutica em plantas já é realizada há muito tempo: a partir de algumas células de uma planta adulta, podemos gerar no laboratório os diversos órgãos dessa planta: raízes, caule, folhas, flores, etc.
Comparando com plantas, ainda estamos no início do desenvolvimento da ciência da regeneração celular em animais. No entanto, à medida que dominarmos os mecanismos de reprogramação das células, iniciaremos uma nova era da medicina: a medicina regenerativa. Com ela, células programadas e multiplicadas em laboratório regenerarão órgãos e tecidos danificados quando re-implantadas em pacientes, acabando com as angustiantes esperas por doadores de órgãos. Um dia até, quem sabe, conheceremos tão bem os processos de diferenciação celular que nem precisaremos transplantar células. Conseguiremos induzir, por exemplo, o toco da perna de uma pessoa que sofreu uma amputação a reiniciar o desenvolvimento e regenerar aquela perna – como a lagartixa que regenera seu rabo cortado. Assim, o momento é de abrir o leque das pesquisas sobre diferenciação e reprogramação celular. Estudos com os clones animais e com os diversos tipos de células-tronco nos ajudarão a entender esses processos e dessa forma controla-los. Na tabela você pode ver o número de indivíduos nos Estados Unidos acometidos pelas mais diversas doenças que se beneficiarão dessas pesquisas. Tabela. Estimativa de afetados por doenças que se beneficiarão com pesquisa em células-tronco humanas (EUA)
Doenças Número de pessoas afetadas (em milhões)
Doenças cardiovasculares 58 Doenças auto-imunes 30 Diabetes 16 Osteoporose 10 Câncer 8,2 Alzheimer 4 Parkinson 1,5 Queimaduras severas 0,3 Trauma de medula espinhal 0,25 Defeitos de nascimento 0,15 Total 128,4
Clonagem reprodutiva versus clonagem terapêutica Mesmo sem iniciar experimentos de clonagem humana de fato, os defensores da clonagem reprodutiva já vêm causando um grande mal à humanidade. Com suas declarações de que iniciarão experimentos de clonagem com seres humanos, à revelia da opinião da comunidade científica internacional que repudia esses experimentos, eles geraram um medo da clonagem em geral, que em 2001 correu o risco de ser totalmente proibida nos Estados Unidos e também no Brasil. Mas isso não é bom? Não, pois podemos acabar proibindo a clonagem terapêutica também, o que, aliás, aconteceu no Brasil em 2005. Estamos em um momento delicado. Tecnicamente, a clonagem humana se aproxima mais e mais da realidade. Temos agora de decidir o que fazer com ela. Ao longo da história, a humanidade já esteve nessa posição algumas vezes. A descoberta da energia nuclear, por exemplo, nos proporcionou a tomografia computadorizada e a ressonância magnética e ao mesmo tempo duas explosões de bombas atômicas. Aprendemos pelo menos uma lição: o poder deve ser usado com responsabilidade. A

47
clonagem terapêutica revolucionará a medicina, proporcionando tecidos para transplantes que aliviarão as mais diversas enfermidades humanas. Por sua vez, a clonagem como forma de reprodução é comprovadamente um fracasso, e é consenso na comunidade cientifica mundial que não deve ser realizada em seres humanos. Devemos evitar a proibição cega – remanescente da época de Galileu Galilei -, que invariavelmente leva ao atraso da ciência e da melhora da qualidade de vida humana. Precisamos sim é de legislação e vigilância para nos defender da clonagem reprodutiva até que seja demonstrado que este é um método seguro de reprodução assistida, e ao mesmo tempo usufruir os potenciais de aplicações médicas da clonagem terapêutica. Vamos utilizar de forma responsável os nossos poderes da clonagem com fins exclusivamente terapêuticos, para que possamos viver as reais maravilhas deste admirável mundo novo. Curiosidade versus responsabilidade Você pode estar pensando: “Mas como resistir à curiosidade científica e não tentar fazer um clone humano?” A ciência não pode parar, imagine tudo que poderemos aprender com esses experimentos!” Deixe-me então lembra-lo de uma figura célebre pelas razões erradas na história da ciência mundial: o médico nazista Joseph Menguele. Durante a segunda guerra mundial, o Dr. Menguele realizou uma série de experimentos em seres humanos, sem nenhuma restrição ética/moral – a ciência sem limites. Com todas essa liberdade, Menguele aprendeu infinitamente menos sobre genética humana – diria até NADA – do que o frei Gregor Mendel no século XIX, trabalhando com ervilhas! Estudando essas plantas, esse, sim, ilustre personagem na história da ciência estabeleceu as leis básicas da genética e da hereditariedade que se aplicam desde a uma ervilha até ao ser humano. Assim, a ciência séria é realizada de forma ética em modelos experimentais mais simples, que nos ajudam a desvendar os mistérios da complexa e fascinante biologia humana.

48

49

50

51

52

53

54

55

56

57

58

59
REPRODUÇÃO ASSISTIDA Lúcia Machado
Adaptado de Marilena C. D. V. Corrêa e Cristiano Costa http://www.ghente.org/temas/reproducao/index.htm
No Brasil, o número de casais que procuram clínicas especializadas em Reprodução Assistida (R.A.) vem aumentando consideravelmente. Espera-se que um aumento ainda mais significativo ocorra em cidades que ofereçam, gratuitamente, em hospitais públicos este tipo de tratamento, como é o caso de São Paulo.
Reprodução Assistida é um conjunto de técnicas, utilizadas por médicos especializados, que tem como principal objetivo tentar viabilizar a gestação em mulheres com dificuldades de engravidar. Muitas vezes essas dificuldades, até mesmo a infertilidade do casal ou um de seus membros, podem trazer sérios prejuízos ao relacionamento conjugal.
As diferentes variantes técnicas do conjunto da RA podem ser reunidas em dois grupos: 1. As mais antigas e mais simples - nas quais a fecundação se dá dentro do corpo da mulher - são chamadas de inseminação artificial. Caso os gametas utilizados na inseminação sejam do próprio casal, chamamos de inseminação homóloga; caso um ou ambos os gametas sejam obtidos a partir de doadores anônimos, chamamos de inseminação heteróloga. 2. As técnicas mais modernas de inseminação artificial - nas quais a fecundação se dá fora do corpo da mulher, ou seja, pelo procedimento de fertilização in vitro.
Existem diversas variantes técnicas da fertilização in vitro, como descritas abaixo: • os gametas masculino e feminino são transferidos diretamente na tuba uterina da
mulher. Essa técnica encontra o apoio da Igreja Católica, quando os gametas utilizados são do próprio casal;
• os ovócitos são fertilizados por espermatozóides em laboratório, e após 3 a 4 dias, os blastocistos são transferidos por via vaginal, na altura das tubas uterinas;
• os espermatozóide são injetados no interior do ovócito e o embrião é transferido, por via vaginal, para a tuba uterina.
• Os espermatozóides são colocados, por via vaginal, diretamente na altura da tuba uterina.
Congelamento de Embriões
Quando a técnica empregada é a de fertilização in vitro, o médico produz um grande número de embriões a partir dos ovócitos e espermatozóides doados. Somente alguns destes embriões serão implantados no útero materno, os demais serão mantidos congelados (criopreservados), para utilização posterior, caso seja necessário.
De acordo com a Resolução 1358/92 do Conselho Federal de Medicina (CFM), os embriões criopreservados não podem ser destruídos ou descartados, devendo permanecer congelados por tempo indeterminado. O destino a ser dado a esses embriões caso ocorra divórcio, doença grave ou morte de um ou ambos os cônjuges, deve ser anunciado previamente por escrito pelo casal. Mas qual destino é possível aos Embriões Congelados?
No Brasil, a única possibilidade é a doação voluntária e anônima destes embriões para mulheres estéreis que desejam gerar um filho. A escolha do doador e da receptora do embrião é realizada pelas clínicas de reprodução assistida, visando obter a maior semelhança fenotípica possível entre a futura mãe e bebê a ser gerado.
Em outros países, entretanto, os embriões congelados passados um prazo legal pré-estabelecido podem ser utilizados para pesquisa médica. Na Inglaterra as pesquisas

60
médicas permitidas são aquelas que: venham a promover avanços no tratamento da infertilidade; promovam o desenvolvimento de novas técnicas contraceptivas; aumentem o conhecimento de doenças congênitas ou a detecção de anormalidades gênicas ou cromossômicas no embrião antes da implantação. Também é permitido o uso destes embriões para a obtenção de células-tronco.
A clonagem terapêutica de células embrionárias é permitida em países como Alemanha e EUA. Em ambos, o limite máximo permitido para o desenvolvimento do embrião in vitro é de 14 dias, sendo permitido em casos especiais o desenvolvimento do embrião até o limite de 18 dias. Lembramos que a clonagem dita terapêutica – aquela que visa a produção de células-tronco embrionárias para utilização em pesquisa - não visa em hipótese alguma a obtenção de seres humanos inteiros. Alguns problemas derivados da técnica de fertilização in vitro
• O que fazer com os embriões excedentes que não foram implantados?
• Homens doadores anônimos de espermatozóides em clínicas de fertilização podem ser acionados na justiça para reconhecimento de paternidade?
• É lícito escolher o sexo do bebe antes da implantação do embrião no útero?
• É lícito vender óvulos e espermatozóides para fertilização assistida de terceiros?
• É lícito implantar mais do que um embrião para garantir o sucesso do
procedimento?
• Uma mulher que “alugou” sua barriga para possibilitar o desenvolvimento de um embrião originado da união de ovócitos e espermatozóides de terceiros tem direito legal sobre a criança?
Link: http://www.brightcone.com/title.jsp?title=855969956 http://www.brightcone.com/title.jsp?title=769641514

61
MEDULA ÓSSEA E AS CÉLULAS-TRONCO HEMATOPOIÉTICAS Ana Carolina Susuki Dias Cintra
A medula óssea vermelha dos humanos é constituída por um tecido esponjoso e
mole (conhecido popularmente como “tutano”) localizado no interior dos ossos nos fetos e ao longo dos anos passa a ficar restrita a apenas alguns ossos nos adultos, como as costelas, vértebras, partes esponjosas de ossos curtos, extremidades de ossos longos dos membros e interior dos ossos do crânio e do esterno. Esse tecido é responsável por parte da reciclagem celular, destruição e produção de novas células que compõem o tecido sanguíneo e imunológico, como os glóbulos vermelhos, glóbulos brancos e plaquetas. As células sanguíneas estão em constante renovação, pois possuem uma vida útil curta: enquanto um eritrócito vive cerca de 120 dias, por exemplo, um trombócito vive cerca de 9 dias e um leucócito, 7 dias.
ATENÇÃO: Não confundir medula óssea com medula espinhal!! A medula espinhal é formada de tecido nervoso que ocupa o espaço dentro da coluna vertebral e tem como função transmitir os impulsos nervosos, a partir do cérebro, para todo o corpo.
Células componentes do sangue e hematopoiése:
• Eritrócito ou glóbulos vermelhos ou hemácias: responsáveis pelo transporte de
oxigênio e de pequena quantidade de gás carbônico; • Leucócitos ou glóbulos brancos:
o Neutrófilos: destroem partículas relativamente pequenas por fagocitose; o Eosinófilos: atacam parasitas e inativam substâncias que produzem
inflamações; o Basófilos; liberam anticoagulantes que previnem a coagulação do sangue,
além de liberar histamina, responsável pela inflamação; o Monócitos: originam o macrófago, que destrói partículas relativamente
grandes por fagocitose; o Linfócitos: responsáveis pela resposta imunológica, mecanismos de
defesa do organismo; • Plaquetas: importantes da coagulação do sangue.
A medula óssea possui células-tronco hematopoiéticas (CTH) e células estromais.
As CTH são um grupo de células-tronco não-embrionárias (adultas) pluripotentes, que têm a capacidade de se dividir e originar todos os tipos de células sanguíneas, com capacidade de auto-renovação (vida longa) e de se dividir sem diferenciação. Já as células estromais também possuem a capacidade de se dividir e de se diferenciar, além de ser responsável pela liberação de sinais reguladores da diferenciação das CTH.
Diferenciação: as células-tronco hematopoiéticas geram dois tipos de células progenitoras multipotentes (CPM), de vida curta:
• célula Progenitora Mielóide: responsável pela formação de células do sangue • Célula Progenitora Linfóide: responsável pela formação de células do sistema
imune Uma pequena população de células-tronco hematopoiéticas (CTH) na medula
óssea é a fonte da maioria dos diferentes tipos de células do sangue e do sistema imunológico. As CTHs residem em um micro-ambiente cercado por células estromais que fornecem importantes sinais reguladores de sua divisão e diferenciação. Quando é necessário produzir mais sangue ou células imunológicas, uma CTH se divide gerando uma cópia de vida longa, que permanece no micro-ambiente e retém sua identidade de CTH pluripotente, enquanto a outra cópia tem vida curta e é chamada progenitora multipotente (CPM). Esta, por sua vez, se divide para produzir progenitoras

62
especializadas na formação de linhagens mielóides (do sangue) e linfóides (do sistema imune).
À medida que as descendentes dessas linhagens se tornam progressivamente especializadas, vão perdendo a capacidade de se proliferar, até parar de se dividir. Nessa etapa, estão completamente diferenciadas. Apenas a célula-tronco retém o potencial proliferativo ilimitado devido a sua capacidade de auto-renovação permanente e de divisão sem diferenciação.
Figura 9. Diferenciação das células do sangue e sua potencialidade nas diferentes fases. Scientific American Brasil ano 5, no. 51, pg.38 – agosto de 2006. Doenças que afetam as células sanguíneas:
o Anemias: caracterizada pela baixa concentração de hemoglobina no sangue, e pode estar relacionada à produção insuficiente ou defeituosa de glóbulos vermelhos, ocasionada por diversos fatores. Dessa forma, o transporte de oxigênio é afetado e, conseqüentemente, menos energia sendo produzida pelas células, ocasionando fraqueza, cansaço, atraso no crescimento e sistema imunológico deficiente.
o Anemia Falciforme: doença hereditária que afeta a estrutura da hemoglobina, deformando as hemácias (originando um formato de foice) e prejudicando sua estabilidade e o seu transporte, ocasionando quadro anêmico.
o Hemofilia: doença ocasionada por defeito genético, que pode ser hereditário, na produção de fatores de coagulação sanguínea que acarreta sangramentos excessivos e lenta coagulação do sangue.
o Câncer: são muitas as formas de câncer que ocorrem nas células do sangue. Umas delas, muito conhecida, é a leucemia, que afeta as células-tronco que dão origem aos leucócitos (glóbulos brancos) e é ocasionada por mutação no DNA que leva a um descontrole no processo de divisão celular, caracterizando um tumor. Além de se dividir descontroladamente, as células cancerosas permanecem em estágio não diferenciado.
Texto adaptado de:
o Sangue – o fluido da vida. o Scientific American Brasil – ano 5, no. 51, pg.38 – agosto de 2006.

63
TRANSPLANTE DE MEDULA ÓSSEA – UMA TERAPIA CELULAR BEM CONHECIDA
Vivian Lavander Mendonça
As origens do transplante de medula óssea As primeiras tentativas científicas de utilização da medula óssea de indivíduos
saudáveis para recompor a função perdida por uma medula doente começaram no final da Segunda Guerra Mundial, com vítimas de acidentes na produção de bombas atômicas.
O transplante de medula óssea surgiu da seguinte idéia: como todas as células do sangue e do sistema imunológico são originadas a partir de células da medula dos ossos, caso haja algum dano ou problema com esse sistema em uma pessoa, pode-se substituí-lo por um sistema saudável. Na verdade, são as células-tronco hematopoéticas da medula óssea que garantem ao receptor a função antes prejudicada, de geração de todas as células do sangue.
Casos em que o transplante de medula óssea é realizado
O transplante é indicado para o tratamento de várias doenças graves que afetam as células do sangue, como anemia aplásica grave (doença em que não há formação das células sangüíneas), algumas doenças hereditárias (talassemias) e vários tipos de leucemias (leucemia mielóide aguda, leucemia mielóide crônica, leucemia linfóide aguda).
O diagnóstico da doença exige, entre outros exames, a avaliação histológica, citogenética e molecular de células da medula óssea do paciente, que são aspiradas geralmente do osso ilíaco ou do esterno (ou da tíbia, em crianças).
Antes do transplante, os pacientes devem ser tratados com altas doses de quimioterápicos e radiação para eliminar as células da medula óssea doente. Esse procedimento é delicado, pois faz com que o número de células sangüíneas seja drasticamente reduzido. O tecido sadio de um doador é então introduzido através de uma veia do receptor e as células migram para a medula óssea. Se o transplante tiver sucesso, em um mês a função da medula será restabelecida. Mesmo assim, o paciente deve receber acompanhamento médico durante um período mínimo de um ano após o transplante, para detecção e tratamento de possíveis complicações.
Condições para ser um doador de medula óssea Transplante autogênico
No caso de transplantes de células-tronco autogênicas não há barreiras imunológicas, uma vez que as próprias células do indivíduo são re-introduzidas. Para viabilizar esse procedimento é necessário que o indivíduo, apesar de doente, tenha um número suficiente de células-tronco sadias. Essa forma de transplante é utilizada principalmente em alguns casos de câncer.
São duas as principais preocupações nos transplantes autogênicos: a obtenção de um número suficiente de células sadias e a eliminação completa das células cancerosas presentes no organismo do doente. Há várias técnicas sendo desenvolvidas para essa finalidade, inclusive o uso de drogas especificamente testadas para agir sobre células cancerosas, sem efeito sobre as células sadias.
Os transplantes autogênicos geralmente são bem sucedidos, sendo o principal risco a recorrência da doença no caso de células cancerosas sobreviverem à terapia anterior ao transplante.
Transplante alogênico
Neste caso, o principal risco para o paciente é a ocorrência de rejeição ao tecido transplantado. Vamos entender por que a rejeição ocorre.
As células do sangue apresentam em sua superfície proteínas específicas, codificadas por um conjunto de genes conhecidos como Complexo Principal de

64
Histocompatilidade – MHC (Major Histocompatibity Complex). Essas proteínas funcionam como antígenos, ou seja, induzem à formação de anticorpos, se transferidas para outro organismo.
Nos seres humanos, esses antígenos são denominados HLA (Human Leukocyte Antigen) e como alguns desses genes possuem mais de 40 alelos diferentes, variam muito entre indivíduos, sendo iguais no caso de gêmeos idênticos. Quanto mais aparentados forem dois indivíduos, mais alelos do MHC eles terão em comum.
Quando células são retiradas de um doador e transplantadas em um receptor não gêmeo, vários desses antígenos HLA são diferentes. O sistema imunológico do receptor considera essas células como estranhas e tenta matá-las, e as células do doador também tentam eliminar as células do receptor. É aí que se dá o processo de rejeição.
Antes que o transplante ocorra, os tecidos do receptor e do doador em potencial devem ser analisados para verificar a compatibilidade, ou seja, o grau de semelhança, dos antígenos HLA. Porém, como não é fácil encontrar um doador compatível, muitas vezes são realizados transplantes em que a compatibilidade HLA entre doador e receptor é parcial. O grau de disparidade entre os antígenos da superfície celular do doador e do receptor vai determinar a intensidade das reações de rejeição, que podem ser minimizadas com medicamentos.
E se não for possível encontrar um doador compatível?
A probabilidade de se encontrar um doador compatível entre irmãos é de 35%, mas este número cai para 0,0001% entre doadores não aparentados.
Quando não há acesso a um doador compatível, a solução é procurar em Bancos de Doadores de Medula. Nesses bancos, voluntários de todo o mundo são cadastrados e têm seus antígenos HLA determinados. No Brasil, há um Registro Nacional de Doadores de Medula Óssea (Redome), que coordena a pesquisa de doadores e está instalado no Instituto Nacional do Câncer (Inca). De acordo com este Instituto, o número de doadores cadastrados vem crescendo, tendo mais de 300 mil em 2006, e a campanha para aumentar o registro continua para garantir que mais pessoas sejam beneficiadas por esses transplantes no país.
Obtenção de células-tronco para transplante de medula óssea As células-tronco utuilizadas em transplantes de medula óssea podem ser retiradas de três fontes diferentes:
a) da medula do doador: o doador é anestesiado e submetido a múltiplas punções, com agulhas, nos ossos posteriores da bacia. Uma pequena parte da medula é aspirada.
b) do sangue circulante:um pequeno número de células-tronco está presente no sangue circulante. A administração de certas drogas estimula a saída de parte dessas células da medula, aumentando assim seu número na circulação. É possível, então, coletar a quantidade necessária de células-tronco fazendo com que o sangue do doador circule por uma máquina que retira especificamente as células desejadas.
c) do sangue do cordão umbilical e da placenta: o cordão umbilical é um órgão que liga o feto à placenta e lhe assegura a nutrição por meio de vasos sangüíneos durante a gestação. Imediatamente após o parto, o cordão é pinçado, para impedir que o sangue contido em seu interior se perca, e o sangue é retirado com o auxílio de uma agulha. As células vermelhas do sangue são coletadas e a amostra é congelada e armazenada por até 15 anos, sem que haja perda da qualidade das células-tronco. Este é um procedimento simples e que permite a obtenção de células a partir de um órgão que em geral era descartado.

65
A B Figura 10: coleta de células-tronco hematopoiéticas para transplante: (A) da medula óssea, (B) do cordão umbilical.
O uso de células-tronco do sangue de cordão umbilical em transplantes é mais vantajoso do que o de medula óssea, por vários motivos: elas se implantam mais eficientemente, são mais tolerantes à incompatibilidade entre receptor e doador, têm disponibilidade imediata e há possibilidade de realização do transplante sem que o doador seja submetido a qualquer tipo de procedimento cirúrgico. A facilidade de coleta e da análise prévia de antígenos HLA estimulou a criação de Bancos de Sangue de Cordão Umbilical no Brasil, também coordenados pelo Redome. Esses bancos seguem normas rígidas para coleta, processamento e armazenamento dos tecidos de cordão umbilical, definidas pela Agência Nacional de Vigilância Sanitária (Anvisa), e a coleta só é realizada se a pessoa estiver ciente da gratuidade da doação e autorizar o possível descarte do material após o prazo seguro para sua utilização.
O que podemos esperar para o futuro?
Células-tronco hematopoéticas cultivadas em laboratório podem se diferenciar em células de outros tecidos, tais como fígado, intestino, pele, músculo cardíaco, e talvez, células nervosas.
Embora se saiba da existência dessas diversas possibilidades de diferenciação, a maneira como isso ocorre ainda não está clara. Por isso, pesquisadores no mundo inteiro buscam compreender os mecanismos envolvidos na diferenciação celular.
Os resultados animadores de algumas pesquisas com células-tronco levam a crer que os transplantes de medula óssea serão apenas uma das modalidades de terapia utilizando essas células. Pelas características das células-tronco, acredita-se no potencial dessas terapias para tratamento de diversas doenças, inclusive as crônico-degenerativas. Vamos apresentar brevemente alguns desses resultados animadores obtidos em pesquisas com células-tronco hematopoéticas.
A equipe do Centro de Pesquisa Gonçalo Moniz, da Fiocruz Bahia, realizou em 2003 o primeiro transplante de células de medula óssea em pacientes com insuficiência cardíaca devida à doença de Chagas, procedimento até então inédito no mundo.
Outra fonte de pesquisas interessante é o tratamento de infartos do miocárdio. Nestes casos, há morte de parte do tecido cardíaco e as células remanescentes não são capazes de reconstituir o tecido morto.
Experimentos indicam que as células-tronco hematopoéticas introduzidas são capazes de migrar para áreas doentes e de originar novas células de músculo cardíaco e de vasos sangüíneos.

66
Figura 11 Tratamento do coração contendo células danificadas por infarto. As células-tronco são introduzidas com o auxilio de seringa e cateter.
No Instituto do Coração (Incor) de São Paulo, estão sendo realizadas aplicações de células-tronco em pacientes com insuficiência cardíaca, causada por doença de Chagas, hipertensão ou de origem desconhecida. Duas técnicas diferentes têm sido utilizadas: a aplicação de células-tronco isoladas da medula óssea e a utilização de um hormônio que estimula a liberação das células-tronco da medula óssea do próprio paciente para a circulação sanguínea – dali as células migram para as áreas lesadas.
As células-tronco também têm sido utilizadas em pesquisas para tratamento de doenças auto-imunes, tais como a artrite reumatóide e o lúpus eritematoso sistêmico. Uma equipe do Hospital das Clínicas da Faculdade de Medicina de Ribeirão Preto, da USP (HCFMRP-USP), empregou células-tronco de medula óssea, retiradas do próprio paciente e submetidas à quimioterapia, para transplante. A quimioterapia destrói as células defeituosas do sistema imune. Os resultados foram animadores e estão sendo agora comparados com os resultados obtidos com a terapia convencional, que não envolve células-tronco.
Apesar do entusiasmo dos cientistas e da sociedade com os resultados positivos da terapia celular, é importante lembrar que ainda são necessárias muitas pesquisas (compostas de diversas etapas até o estabelecimento de um novo procedimento médico), financiamentos e discussões no campo político, ético e legal. Este é o cenário ideal para o desenvolvimento dessa nova forma de terapia.
Texto adaptado de: Sangue – fluido da vida. Parte 3 – Terapia celular: sonhos e realidade -Departamento de Genética e Biologia
Evolutiva – Centro de Estudos do Genoma Humano - Eliana Maria Beluzzo Dessen, Maria Gabriela Guimarães Ribeiro dos Santos, Rodrigo Venturoso Mendes da Silveira e Regina Célia Mingroni Netto
Outras fontes consultadas Revista Pesquisa Fapesp - Células tronco – Marcos Pivetta – Ed. 110; Abril 2005. Páginas da Internet: Com Ciência. Artigo: Células-tronco – Pesquisa brasileira em CT já apresenta resultados. Luciene Zanchetta.
10/02/2004 (http://www.comciencia.br) Inca – Instituto Nacional do Câncer. 15/04/2007 (http://www.inca.gov.br)
catéter Seringa contendo células-tronco
catéter Balão do catéter
Zona da borda Área enfartada

67

68

69

70

71

72
O ARTIGO 5° DA LEI N° 11.105, DE 2005, NÃO É INCONSTITUCIONAL Reginaldo Minaré -Advogado e Diretor Jurídico da ANBio.
www.anbio.org.br
No dia 30 de maio de 2005 o Procurador-Geral da República ajuizou Ação Direta
de Inconstitucionalidade – Adin n° 3510 – alegando ser inconstitucional o artigo 5° da Lei n° 11.105/05, que permite, cumpridas determinadas exigências, a utilização de células-tronco embrionárias em pesquisas e terapias.
A Adin 3510, cujo relator é o Ministro Carlos Brito, tem por assunto tema de fundamental relevância, principalmente para aqueles que são portadores de determinadas doenças ainda incuráveis e para os que serão em futuro breve,
Na ação proposta, o Procurador-Geral tenta demonstrar que o artigo 5° da Lei n° 11.105/05 é incompatível com o que está disposto no caput do artigo 5° da Constituição Federal – CF e no artigo 1° inciso III também da CF.
já conta com diversas instituições admitidas na qualidade de amicus curiae e foi tema de audiência pública no Supremo Tribunal Federal - STF, e atualmente está com o Ministro relator para análise da transcrição dos argumentos apresentados na audiência pública realizada no dia 20/04/2007.
Dispõe o artigo 5° da Lei 11.105/05:
• "Art. 5º É permitida, para fins de pesquisa e terapia, a utilização de células-tronco
embrionárias obtidas de embriões humanos produzidos por fertilização in vitro e não utilizados no respectivo procedimento, atendidas as seguintes condições:
I – sejam embriões inviáveis; ou II – sejam embriões congelados há 3 (três) anos ou mais, na data da publicação desta Lei, ou que, já congelados na data da publicação desta Lei, depois de completarem 3 (três) anos, contados a partir da data de congelamento.
§1º Em qualquer caso, é necessário o consentimento dos genitores. § 2º Instituições de pesquisa e serviços de saúde que realizem pesquisa ou terapia
com células-tronco embrionárias humanas deverão submeter seus projetos à apreciação e aprovação dos respectivos comitês de ética em pesquisa.
§ 3º É vedada a comercialização do material biológico a que se refere este artigo e sua prática implica o crime tipificado no art. 15 da Lei no 9.434, de 4 de fevereiro de 1997". Dispõe o artigo 5° caput e artigo 1° inciso III, ambos da CF:
• "Art. 5º Todos são iguais perante a lei, sem distinção de qualquer natureza,
garantindo-se aos brasileiros e aos estrangeiros residentes no País a inviolabilidade do direito à vida, à liberdade, à igualdade, à segurança e à propriedade, nos termos seguintes:
• Art. 1º A República Federativa do Brasil, formada pela união indissolúvel dos
Estados e Municípios e do Distrito Federal, constitui-se em Estado Democrático de Direito e tem como fundamentos:
III - a dignidade da pessoa humana;"

73
Contestar os argumentos do Procurador-Geral e defender a constitucionalidade do artigo 5° da Lei 11.105/05 é, ao mesmo tempo, a realização de um exercício profundo da hermenêutica cujos argumentos e conclusões devem ser apresentados dentro de uma retórica precisa e objetiva.
Poder-se-ia também dizer que a contestação da Adin está dentro do campo de uma discussão política e ética, que se realiza na sociedade global e não apenas na sociedade brasileira. Todavia, acredito que, no Brasil, o debate político e ético já foi realizado no âmbito do Congresso Nacional, onde diversas audiências públicas sobre o tema foram elaboradas, a sociedade participou e os representantes eleitos pelo povo, respeitando todas as regras do procedimento democrático estabelecido pela CF, votaram, em sua grande maioria, pela aprovação da redação do artigo 5° da Lei 11.105/05. Assim, e considerando que a CF em seu artigo 102 estabelece que ao STF compete a guarda da Constituição, entendo que a análise do tema neste fórum deve ser realizada de forma precisa e objetivando responder se o artigo 5° da Lei 11.105/05 afronta o artigo 5° caput da CF ou o princípio da dignidade humana previsto no inciso III do artigo 1° da CF. Na ação proposta, argumenta o Procurador-geral que pelo fato da vida humana iniciar no momento da concepção e a Constituição garantir a inviolabilidade do direito à vida, de todo brasileiro e estrangeiro residente no Brasil, a proteção deste bem jurídico deve ser realizada, de forma absoluta e dogmática, a partir do momento da concepção, não permitindo qualquer flexibilização.
Que a vida está presente no momento da fecundação é algo que não contexto, pois não conheço argumento convincente na biologia que afirme o contrário. Todavia, considero pertinente o questionamento feito por Umberto Eco, - na obra Em que crêem os que não crêem? -, que pergunta, para ilustrar sua proposta de reflexão sobre até onde é possível retroceder, se vida e humanidade já não estão no sêmen, e podemos dizer também no óvulo, antes mesmo do encontro entre os 23 cromossomos masculinos com os 23 cromossomos femininos? Para problematizar seu questionamento, Umberto Eco propõe a reflexão sobre o que dizer a respeito do desperdício de sêmen por parte de um adolescente em tentação?
Estas questões colocadas são pertinentes, pois nos convida a refletir com profundidade sobre qual o significado conceitual da palavra vida no contexto do artigo 5° da CF, e a intensidade do alcance do artigo.
Fazendo uma análise literal do texto constitucional, fica claro que a CF objetivou garantir a vida de brasileiros e estrangeiros residentes no País. Assim, não resta dúvida que o constituinte de 1988 não tinha a intenção de retroceder ao ponto de atingir aquele adolescente em tentação lembrado por Umberto Eco. Também não nos parece razoável atribuir ao constituinte a idéia de que, com a redação que aprovou, tinha a intenção de considerar um embrião congelado importado da Nova Zelândia como um estrangeiro residente no País.
Por outro lado, compreendendo ser o comando contido no caput do artigo 5° da Constituição o estabelecimento de um princípio, imperioso se faz estabelecer o entendimento sobre qual seria a intensidade da proteção necessária para garantir que o princípio do direito à vida não fosse violado pela legislação e, também, refletir sobre situações onde outros princípios constitucionais com ele concorriam.
Afirmar que para garantir a inviolabilidade do princípio do direito à vida seria necessária uma proteção absoluta e inflexível, inclusive para embriões congelados e inviáveis para a reprodução humana, sem dúvidas seria uma argumentação simplista e até falaciosa. Pois, fazendo uma afirmação nesse sentido, seria difícil depois justificar a constitucionalidade do aborto em caso de gravidez oriunda de estupro, que é um procedimento garantido pelo Código Penal, o aborto no caso de anencefalia do feto, que já é uma prática autorizada em muitos casos pelo Poder Judiciário, onde caudalosa é a jurisprudência nesse sentido, e até mesmo os critérios utilizados para assegurar a preferência pela vida da gestante em casos onde a preservação das vidas do feto e da gestante não são compatíveis. Tudo isso, sem falar que a própria Constituição já

74
relativizou a proteção ao direito à vida ao permitir que, em caso de guerra declarada, seja adota a pena de morte.
Diante do que até aqui foi argumentado, resta claro que interpretar o caput do artigo 5° da CF como comando para uma proteção absoluta e inflexível do direito à vida não é um exercício razoável da hermenêutica e, conseqüentemente, a retórica fica desprovida de fundamento sólido e convencedor.
Efetivamente, ao aprovar a redação do artigo 5° da Lei 11.105/05, o Congresso Nacional em nada violou o princípio de proteção do direito à vida. O que fez o Parlamento foi legitimar um interesse da sociedade, pelo menos o interesse da maioria representada, que considera que em nada viola o princípio do direito à vida o fato de permitir a realização de pesquisas com célula-tronco embrionária para tentar descobrir a cura ou tratamento para melhorar as condições de vida de crianças, adolescentes, jovens e idosos, que são acometidos por doenças incuráveis até o momento ou com poucas possibilidades de tratamento.
Além disso, o Parlamento não atuou de forma irresponsável no momento que legislou. O artigo 5° da Lei 11.105/05 tem uma redação que permite o uso de célula-tronco embrionária de forma contida, muito criteriosa e amparada por instrumentos coercitivos que podem ser utilizados em qualquer situação que configurar a prática de um excesso ou banalização. Modo de proceder que de forma alguma torna banal o uso de embriões humanos.
Segundo dispõe o artigo 5° da Lei 11.105/05, só é permitido o uso de células-tronco embrionárias obtidas de embriões humanos produzidos por fertilização in vitro e não utilizados no respectivo procedimento; e que sejam oriundas de embriões congelados há 3 (três) anos ou mais até a data da publicação da Lei ou que, já congelados na data da publicação da Lei, completaram 3 (três) anos, contados a partir da data de congelamento, ou que sejam inviáveis. Exige ainda o referido artigo 5º em análise que, em qualquer caso, o uso de embriões congelados deve se precedido do consentimento dos genitores, e que os protocolos de pesquisas sejam aprovados pelos Comitês de Ética em Pesquisa - CEP.
Para garantir que os critérios estabelecidos sejam cumpridos, a própria Lei 11.105/05, em seu artigo 24, criminalizou o uso de embriões humanos em desacordo com as regras estabelecidas em seu artigo 5°. Já para garantir que a permissão não sirva para fomentar a criação de um mercado de embriões e de células-tronco embrionárias, o próprio artigo 5º proíbe, em seu § 3°, a comercialização do material biológico afirmando que sua prática configura o crime tipificado no artigo 15 da Lei 9.434/97, lei que dispõe dobre a remoção de órgãos, tecidos e partes do corpo humano para fins de transplante e tratamento.
Evidente, portanto, que o artigo 5° da Lei 11.105/05, em nada viola o artigo 5° caput da CF. Logo, o argumento de inconstitucionalidade não pode ser legitimo para retirá-lo do ordenamento jurídico brasileiro.
Já a defesa da tese de que a preservação da dignidade da pessoa humana estaria ameaçada com a manutenção do artigo 5° da Lei 11.105/05 no ordenamento jurídico pátrio, sem dúvida trilha o caminho da contramão do consenso estabelecido pela maioria. O que ficou demonstrado com a deliberação do Congresso Nacional é que a aprovação da matéria disciplinada nos termos do mencionado artigo 5° aprimora o respeito ao princípio da dignidade da pessoa humana. Inclusive, foi esta a tese que aglutinou o consenso da maioria no procedimento de legitimação pela democracia, e não aquela defendida pelo Procurador-Geral, que é exatamente a tese contrária que foi derrotada no Parlamento e que não contou com o veto do Chefe do Poder Executivo.
Locução cristaliza na maioria das línguas, mas pouco comentada e definida, o significado conceitual da expressão "dignidade humana" é um daqueles conceitos que imaginamos conhecer muito e ter pleno domínio, mas que ao sermos convidados a comentá-lo e explicá-lo percebemos que nossa convicção não é tão clara como acreditávamos. Oriunda do latim dignitas, a palavra dignidade significa valor, distinção,

75
princípio ao qual está baseado o proceder que enseja respeito, e corresponde à tradução feita por Âncio Boécio e os escolásticos da palavra grega aksióma – axioma -, que segundo Aristóteles significa a proposição primeira de que parte a demonstração, o princípio basilar que deve ser necessariamente possuído por quem queira aprender o que quer que seja.
Na modernidade, Immanuel Kant, na obra Fundamentação da Metafísica dos Costumes, procura demonstrar que o ser humano possui um valor em si mesmo, uma dignidade, e constrói o famoso imperativo prático: "Age de tal maneira que uses a humanidade, tanto na tua pessoa como na pessoa de qualquer outro, sempre e simultaneamente como fim e nunca simplesmente como meio". Para que o ser humano identifique a limitação que esse imperativo prático impõe às suas ações, Kant propõe a seguinte reflexão: "Age sempre segundo aquela máxima cuja universalidade como lei possas querer ao mesmo tempo". Segundo Kant, essa é a fórmula para extrair ou identificar uma vontade boa. Em nosso momento histórico, esse imperativo prático kantiano é muito citado como significado da expressão "dignidade da pessoa humana".
A idéia de uma dignidade inerente a todos os membros da família humana se encontra na base da Carta Internacional dos Direitos Humanos, e no Brasil o artigo 1°, inciso III, da CF afirma que a República Federativa do Brasil se constitui em Estado Democrático de Direito e tem como um de seus fundamentos a dignidade da pessoa humana.
O Procurador-Geral, para desenvolver sua tese, levanta a existência de conflito de princípios constitucionais, argumentando que o uso de células-tronco embrionárias, por violar o princípio do direito à vida, necessariamente viola o princípio da dignidade humana. Fica claro, em seus argumentos, que o Procurador coloca o princípio do direito à vida em patamar absolutamente superior ao princípio da dignidade da pessoa humana, não aceitando que em nenhum momento essa ordem hierárquica seja invertida.
Não admite, por exemplo, que em nome do respeito ao princípio da dignidade humana, a milhares de crianças portadoras de distrofias musculares sejam-lhes dada a oportunidade da ciência atuar na tentativa de encontrar cura para uma doença que deteriorará seus músculos e as levará à morte precoce.
Não admite, também, que em nome do respeito ao princípio da dignidade humana, a milhares de idosos, atuais e futuros, que trabalharam para construir as famílias e o Estado, e que são portadores de doenças degenerativas, sejam-lhes garantido o direito de ver os cientistas trabalhando para encontrar a cura da doença que comprometerá integralmente o gozo de sua velhice.
Evidente, portanto, que ao aprofundar a reflexão sobre o que se pretende realizar com o desenvolvimento de pesquisas com as células-tronco embrionárias, não dá para querer que a pretensão do Procurador-geral que propôs a ação de inconstitucionalidade seja elevada à universalidade como uma lei que posso querer que seja também aplicada a mim e ao meu próximo.
Cabe observar ainda que o argumento de que as pesquisas com células-tronco oriundas de outras fontes, como cordão umbilical, poderiam substituir o uso das células-tronco embrionárias no processo de busca de cura para os males acima mencionados, também não justifica a pretensão do Procurador-Geral. Visto que, sendo possível lançar mão também do uso de célula-tronco embrionária, modalidade de pesquisa que é considerada por significativo número de cientistas com mais apta a atingir objetivo tão nobre, não é razoável privar a sociedade da liberdade de pelo menos tentar.
Assim, resta indubitável que a pretensão que objetiva colocar o princípio do direito à vida em patamar absolutamente superior ao princípio da dignidade da pessoa humana, e dogmatizar uma hierarquia de princípios constitucionais nestes termos, não pode ser acolhida, visto não possuir a razoabilidade que se espera ver aplicada em processo que, em seu âmago, é suscitado conflito de princípios constitucionais. Principalmente, quando neste momento histórico a interpretação dos sistemas jurídicos não mais se baseia apenas no ideal clássico da ciência oriunda da física aristotélica, pois com ele concorre o

76
próprio relativismo ou incertezas da física moderna e o probabilismo da ciência experimental.
Neste caso específico, garantir, com regras claras e razoáveis, a liberdade para que a sociedade possa pesquisar a cura para milhares de pessoas é, sem dúvida, uma alternativa mais próxima ao respeito do princípio da dignidade humana do que aquela que objetiva proibir a pesquisa com células-tronco embrionárias, oriundas de embriões inviáveis para reprodução ou congelados a mais de três anos, em nome do respeito ao princípio do direito à vida.
Caso seja acolhida a tese do combativo Procurador-Geral, resta indubitável que não só o fundamento do Estado Democrático de Direito previsto no inciso III do artigo 1° da CF estaria sendo violado, como também o parágrafo primeiro desse mesmo artigo 1° da CF, que dispõe: "Todo o poder emana do povo, que o exerce por meio de representantes eleitos ou diretamente, nos termos desta Constituição".
Para concluir, cabe lembrar que aquele que, por motivo de crença, não concordar com o procedimento estabelecido pelo artigo 5° da Lei 11.105/05, tem todo direito de não autorizar o uso de seus embriões e, inclusive, não fazer uso quando desenvolvida uma técnica baseada nesta pesquisa.

77
Revista Pesquisa Fapesp QUANDO COMEÇA A VIDA?
23/04/2007 -
Fábio de Castro
Agência FAPESP – As células-tronco embrionárias humanas devem ser utilizadas em pesquisas científicas? A importância, dúvidas e a complexidade da questão são tão grandes que, pela primeira vez, o Supremo Tribunal Federal (STF) realizou uma audiência pública sobre um assunto em julgamento na casa.
Na audiência, realizada em Brasília na última sexta-feira (20/4), 34 cientistas apresentaram posições favoráveis e contrárias ao uso das células-tronco. O objetivo era fornecer subsídios científicos para os 11 ministros que compõem o STF.
Em março de 2005, as pesquisas com células-tronco embrionárias humanas foram aprovadas no Brasil, no âmbito da Lei de Biossegurança. Em maio do mesmo ano, no entanto, o então procurador-geral da República, Cláudio Fonteles, entrou no STF com uma Ação Direta de Inconstitucionalidade contra o artigo a respeito das pesquisas, sob a alegação de que estudos do gênero “ferem o direito de embriões”.
O pedido de Fonteles foi acatado no fim de 2006 pelo ministro do STF Carlos Ayres Britto, que foi relator do caso. Para decidir a questão, os ministros precisarão, segundo Britto, discutir quando a vida humana começa.
O relator convocou então a audiência, para a qual convidou 18 cientistas. Outros 11 foram chamados pela Procuradoria Geral da República. Quatro foram convidados pela presidência da República e um pela Confederação Nacional dos Bispos do Brasil (CNBB).
Para Ayres Britto, do ponto de vista técnico não existe na Constituição um conceito claro de quando começa a vida. O subsídio oferecido pela comunidade científica, segundo ele, permitiria aos ministros do STF formular “um conceito operacional de vida”.
Para alguns dos cientistas presentes na audiência, a vida começa na fecundação. Outros alegaram que ela surge apenas no terceiro ou quarto dia, quando ocorre a nidação – processo em que a célula migra para o útero materno. Um terceiro grupo defendeu que o embrião só pode ser considerado vivo quando acontece a formação do sistema nervoso e que questões éticas que envolvem o tema impediram, até agora, o avanço de pesquisas na área.
A geneticista Mayana Zatz, do Centro de Estudos do Genoma Humano da Universidade de São Paulo (USP), destacou a importância de que a legislação permita as pesquisas com células-tronco embrionárias humanas, que, segundo ela, são hoje as únicas com potencial para recuperar certas doenças neurológicas incuráveis.
Para Mayana, a possibilidade de serem desenvolvidas pesquisas com tais células definirá, no futuro, a existência ou não de tratamento para inúmeras doenças degenerativas que atingem a população. Segundo ela, a célula-tronco embrionária só se tornaria um feto por meio da intervenção humana, já que, para isso, ela tem de ser inserida no útero.
“O que é eticamente mais correto: preservar um embrião congelado, mesmo sabendo que a probabilidade de ele gerar um ser humano é praticamente zero, ou doá-lo para pesquisas que poderão resultar em futuros tratamentos?”, questionou.
De acordo com a cientista, 7 mil doenças genéticas degenerativas atingem mais de 5 milhões de crianças nascidas de pais normais no Brasil. “Toda célula é vida, um coração a ser transplantado é vivo, mas não é um ser humano. Estamos defendendo que, da mesma maneira que um indivíduo em morte cerebral doa órgãos, um embrião congelado possa doar suas células”, disse. Muita discussão, pouca conclusão
Patrícia Pranke, professora da Universidade Federal do Rio Grande do Sul e diretora-presidente do Instituto de Pesquisa com Célula-Tronco, falou na audiência no STF que só a partir do quarto dia o embrião (blastocisto) pode ser implantado no útero, o único ambiente em que poderá se desenvolver. Segundo ela, os embriões ou são implantados no útero ou são congelados. “O próprio congelamento diminui a possibilidade de o embrião se desenvolver depois. Por que não doá-los para pequisa?”, disse.
Para Lúcia Braga, da Rede Sarah de Hospitais de Reabilitação, a pergunta correta a ser feita é: qual destino será dado aos embriões que não chegam a ser implantados no útero? “Podemos ficar aqui dias discutindo quando a vida começa, sem chegar a uma conclusão”, afirmou.

78
Já para a professora-adjunta do Departamento de Biologia Celular da Universidade de Brasília (UnB) Lenise Martins, a vida humana começa na fecundação. Segundo ela, todo ser vivo tem fases diferentes durante o seu ciclo de vida.
Como exemplo, ela utilizou o desenvolvimento da lagarta e da borboleta, que são um mesmo animal em fases diferentes de um mesmo ciclo de vida. “O indivíduo não precisa começar a manifestar sua sabedoria para ser considerado humano. O embrião humano já é da espécie Homo sapiens mesmo que não possa ainda aprender”, afirmou.
O médico Marcelo Vacari Mazzenoti, da Universidade Federal de São Paulo (Unifesp), especializado em crianças com má-formação, também defendeu que a vida humana começa na fecundação e afirmou que a utilização de células-tronco embrionárias humanas não é necessária para a medicina atual.
“Podemos utilizar células-tronco adultas em diversas situações, como [no estudo de tratamentos contra] doença de Chagas, doenças auto-imunes, acidentes vasculares cerebrais, lesões de medula espinhal e doenças genéticas, dentre outras. Já com relação à utilização de células tronco embrionárias, não há fato objetivo e concreto que confirme a sua utilidade”, defendeu.
Mazzenoti mencionou que há 72 aplicações clínicas descritas com o uso de células-tronco adultas e nenhuma aplicação descrita de células-tronco embrionárias humanas. “Não é preciso interromper a vida para trabalhar com células-tronco.”
A professora da Universidade Federal do Rio de janeiro (UFRJ) Cláudia Maria de Castro Batista defendeu a autonomia do embrião humano. Para ela, a vida humana é um processo contínuo, coordenado e progressivo que começa a partir da fecundação do óvulo pelo espermatozóide.
“Uma vez que o óvulo é fecundado, forma-se a primeira célula do Homo sapiens e todo um programa de fertilização é disparado. O direito à vida e à integridade física desde o primeiro momento da existência é o princípio de igualdade que deve ser respeitado”, afirmou.
Lílian Piñero Eça, do Instituto de Pesquisas com Células-Tronco (IPCTron), fez uma exposição sobre o diálogo entre o embrião humano e sua mãe. A cientista defendeu que duas a três horas depois da fecundação, após o encontro do espermatozóide com o óvulo, o embrião já se comunica com a mãe.
“Pelo menos cem neurotransmissores são emitidos pelo embrião para os 75 trilhões de células existentes no corpo da gestante, que começa a sofrer mudanças hormonais”, disse. Segundo a pesquisadora, essa é a forma de o embrião “falar” para o corpo da mãe se preparar para a gravidez.
O coordenador da Divisão de Medicina Óssea da Faculdade de Medicina de Ribeirão Preto (FMRP) da USP, Júlio César Voltarelli, questionou o que considera o principal argumento por parte dos que são contra o uso das células de embriões: que não seriam necessárias uma vez que benefícios clínicos poderiam ser conseguidos com as células adultas.
Para Voltarelli, a utilização somente de células-tronco adultas não é suficiente para tratar várias doenças auto-imunes em estágio precoce. Além disso, no caso da esclerose lateral amiotrófica, por exemplo, 95% dos pacientes morrem até os 4 anos de idade. “Só a utilização de células-adultas não é suficiente nesses casos. Precisamos utilizar células-tronco embrionárias”, disse. Saiba mais lendo a página do Centro de Estudos do Genoma Humano: http://www.genoma.ib.usp.br
• É constitucional pesquisar células-tronco a partir de embriões? Erickson Gavazza Marques http://www.genoma.ib.usp.br/noticias/pdf/rep-erickson_marques070420.pdf
• STF assiste a disputa ideológica pela “vida” – Laura Capriglione - http://www.genoma.ib.usp.br/noticias/pdf/rep-laura_capriglione70421.pdf
• Decisão suprema – Luis Carlos Azevedo - http://www.genoma.ib.usp.br/noticias/pdf/art-luiz_azevedo070504.pdf

79
São Paulo, quinta-feira, 03 de maio de 2007 Que vida, biológica ou moral? Oscar Vilhena Vieira
POUCO TEMPO antes de deixar o comando do Ministério Público Federal, o então
procurador-geral Claudio Fonteles propôs uma ação direta de inconstitucionalidade contra dispositivos da Lei de Biossegurança que autorizam a pesquisa, para fins terapêuticos, com células de embriões inviáveis para fertilização.
Argumenta o ex-procurador-geral da República que a lei é inconstitucional, pois violaria o direto à vida, bem como o princípio da dignidade humana -ambos entrincheirados em nossa Constituição. O raciocínio é simples. A vida começa com a fecundação. O direito à vida é protegido pela Constituição. Logo, fazer pesquisa com células embrionárias é atentar contra a dignidade da vida humana.
O raciocínio do ex-procurador-geral é tão cartesiano quanto incorreto. O primeiro equívoco do ex-procurador-geral é não reconhecer que o debate
colocado ante o Supremo Tribunal Federal é de natureza prevalentemente moral, e não de natureza "estritamente científica", como propõe em artigo nesta Folha de S.Paulo ("Tendências/Debates", 26/4).
A questão fundamental, portanto, não é quando começa a vida biológica, mas sim que grau de proteção jurídica deve ser conferido à vida em cada etapa de seu desenvolvimento.
Reconhecer que o embrião tem vida não significa que estejamos dispostos a equipará-lo moral e juridicamente a uma pessoa. Seria como comparar uma semente de jacarandá encontrada no chão da floresta com uma árvore centenária que protegemos com nossa legislação ambiental.
A dor de ver uma semente sendo comida por um passarinho não é equiparável àquela de ver uma árvore derrubada por um raio, como nos lembra o filósofo Michael Sandel.
Todos que já perderam uma pessoa querida sabem o que significa a morte de um ser humano, e esta não pode ser comparada com o não-desenvolvimento de um embrião, ainda mais quando falamos de um embrião que se encontra fora do útero e é inviável para fertilização.
Essa distinção no valor atribuído a cada uma das diversas etapas de evolução da vida já é feita pelo nosso direito e, até onde sei, jamais foi questionada pelo ex-procurador-geral da República Claudio Fonteles.
Nosso Código Penal permite, por exemplo, o aborto quando houver risco de vida para a mãe. Ou seja, na ponderação feita pelo legislador, ele deu mais importância à vida da mãe do que à expectativa de vida do feto -e é razoável que assim tenha feito.
Importante destacar, por outro lado, que a pesquisa autorizada pela Lei de Biossegurança se resume apenas àqueles embriões que foram produzidos fora do útero materno para fins de fertilização, mas que não se demonstraram viáveis para esse mesmo fim, seja por um problema de natureza fisiológica, seja porque, depois de três anos congelados, não mais podem ser implantados com segurança em um útero materno.
Ou seja, estamos falando de embriões que não possuem nenhuma expectativa de evoluir para a condição humana. A equiparação mecânica feita pelo ex-procurador-geral é, portanto, destituída de sentido.
Isso não quer dizer que o embrião não tenha valor e que não deva ser protegido. Antes o contrário. Ele tem valor e devemos protegê-lo. Porém, essa proteção deve ser distinta daquela proteção que conferimos às pessoas.

80
É exatamente isso o que faz a Lei de Biossegurança. Ela proíbe a pesquisa com qualquer embrião que seja viável. Mais do que isso, proíbe qualquer pesquisa que não tenha fins terapêuticos, portanto, humanitários.
O terceiro aspecto preocupante do argumento levado a cabo pelo ex-procurador-geral da República é a sua omissão em relação à dignidade e à própria vida de milhões de pessoas humanas que sofrem doenças graves e letais, como Parkinson, diabetes, doenças coronárias ou lesões de medula, que poderiam ser beneficiadas com o progresso nas pesquisas com células-tronco.
Ao elevar o embrião inviável à condição de ser humano, o sofrimento de milhares de seres humanos reais está sendo relegado à mais absoluta irrelevância. E essa não parece ser uma escolha moralmente adequada por quem luta em favor da vida.
OSCAR VILHENA VIEIRA, 41, advogado, mestre em direito pela Universidade de Colúmbia (EUA) e doutor em ciências políticas pela USP, é professor de direito constitucional da Escola de Direito da Fundação Getúlio Vargas e diretor jurídico da Conectas Direitos Humanos. É autor, entre outras obras, de "Direitos Fundamentais: uma Leitura da Jurisprudência do STF".

81
GLOSSÁRIO Bioética - O estudo dos problemas éticos suscitados pelas pesquisas biológicas e pelas suas aplicações por pesquisadores, médicos, etc. Blastocisto – embrião, em fase de pre-implantação no útero, com cerca de 150 células produzidas por divisão celular após a fertilização. O blastocisto é uma esfera formada por uma camada externa de células (o trofoblasto), uma cavidade (a blastocele), contendo em seu interior um conjunto de células denominado massa interna de células. Célula-tronco - célula capaz de se dividir por períodos indefinidos sem dar origem a células especializadas. Célula-tronco do adulto - célula indiferenciada encontrada em tecidos que pode renovar-se a si mesma (com certas limitações) e se diferenciar em todas as células especializadas do tecido do qual foi originada. Célula-tronco do adulto é também denominada célula-tronco adulta, ou tecidual. Células-tronco de cordão umbilical – células-tronco coletadas do cordão umbilical imediatamente após o nascimento podem produzir todos os tipos de células sanguíneas. As células-tronco de cordão são normalmente usadas para tratar pacientes com câncer ou outras doenças do sangue, após sofreram quimioterapia para destruir sua própria medula óssea. Célula-tronco embrionária - célula indiferenciada encontrada no embrião que tem o potencial para se diferenciar numa ampla variedade de células especializadas. Célula-tronco hematopoiética – célula-tronco que origina todas as células do sangue: hemácias, glóbulos brancos (todos os tipos) e plaquetas.
Célula pluripotente: célula capaz de gerar os três tipos de células germinativas (ectoderme, mesoderme e endoderme), ou seja, tem o potencial para se desenvolver nos mais de 200 tipos celulares conhecidos do corpo humano.
Célula-progenitora: derivada da célula-tronco e que dará origem a células diferencidas.
Célula totipotente: capaz de dar origem aos tecidos que formarão o embrião. Ex.: zigoto.
Célula-tronco unipotente: capaz de dar origem a uma única linhagem de células diferenciadas. Clonagem reprodutiva – o objetivo da clonagem reprodutiva é a criação de um animal idêntico ao doador do núcleo da célula somática. O embrião é implantado no útero e desenvolve em um ser vivo. O primeiro animal a ser criado por clonagem reprodutiva foi a ovelha Dolly, nascida no Instituto Roslin na Escócia, em 1996. Clonagem terapêutica – a meta da clonagem terapêutica é a criação de células perfeitamente compatíveis com as do paciente no qual elas serão injetadas. Nessa técnica os cientistas combinam o núcleo de uma célula somática do paciente com uma célula ovo da qual o núcleo foi retirado. Essa nova célula, ao se dividir, origina células-tronco embrionárias que são coletadas e usadas para gerar tecidos que são compatíveis com o organismo do paciente, isto é, o tecido formado não causará rejeição quando transplantado. Clone – geração de cópias idênticas de uma molécula, célula, ou organismo.

82
Cultura de células – crescimento de células in vitro em um meio artificial para experimentos em laboratório. Diferenciação: é o processo através do qual uma célula não especializada torna-se especializada. Ectoderma – folheto embrionário mais externo formado por células derivadas da camada interna do blastocisto, origina o sistema nervoso, órgãos do sentido, pele e estruturas relacionadas. Embrião – em humanos, o organismo em desenvolvimento a partir da fertilização até o final da oitava semana de gestação, quando então passa a ser chamado de feto. Endoderma – folheto embrionário com posição mais interna formado por células derivadas da camada interna do blastocisto, origina os pulmões e outras estruturas respiratórias e os órgãos do aparelho digestório. Ética - Estudo dos juízos de apreciação referentes à conduta humana suscetível de qualificação do ponto de vista do bem e do mal, seja relativamente a determinada sociedade, seja de modo absoluto. Feed layer – Células usadas em co-cultura para manter as células-tronco embrionárias. Para o cultivo de células-tronco embrionárias, são incluídos na cultura fibroblastos de embrião de camundongo ou humanos que foram tratados de modo a impedir sua divisão. Fertilização in vitro – técnica que une ovócito e espermatozóide em laboratório. Gene – unidade funcional de herança e que corresponde a um segmento de DNA nos cromossomos. O gene é uma unidade de transcrição. In vitro – denominação em latim para “dentro de vidro”, ou em “tubo de ensaio” em experimentos de laboratório, um meio artificial, fora do organismo. Marcadores de superfície – proteínas presentes na superfície externa da célula e que são únicas para determinados tipos celulares. Elas podem ser detectadas por meio de anticorpos e outros métodos de detecção. Massa interna de células – grupo de células dentro do blastocisto. Essas células dão origem ao embrião e finalmente ao feto. A partir dessas células são geradas as linhagens de células-tronco embrionárias. Meio de cultura – liquido que cobre as células numa placa de cultura e que contem nutrientes para alimentar as células. O meio pode também incluir outros fatores adicionados para produzir mudanças nas células. Mesoderma – folheto embrionário com posição mediana formado por células derivadas da camada interna do blastocisto, origina os ossos, músculos, tecido conectivo, rins e estruturas relacionadas. Micro ambiente – moléculas e compostos tais como nutrientes e fatores de crescimento no fluido que rodeia a célula num organismo ou no laboratório e que tem um importante papel na determinação das características da célula. Passagem – um ciclo de crescimento celular e proliferação em cultura.

83
Plasticidade: é a capacidade de uma céula-tronco adulta de um tecido gerar uma célula(s) especializada(s) de um outro tecido. Por exemplo, já foi demonstrado in vitro que células-tronco hematopoiéticas são capazes de gerar neurônios. Sinais – fatores internos e externos que controlam as mudanças na estrutura e função das células Terapia celular ou medicina regenerativa – tratamento no qual as células-tronco são induzidas em tipos celulares específicos necessários para reparar tecidos danificados ou substituir células que foram destruídas. Teratoma – os cientistas comprovam se eles conseguiram obter uma linhagem de células-tronco embrionárias injetando essas células em camundongo com o sistema imune reprimido. Uma vez que elas não podem ser destruídas pelo sistema imune do camundongo, elas sobrevivem e formam um tumor benigno, com muitas camadas, denominado teratoma. Mesmo que a formação de tumores não seja normalmente desejada, nesse teste, os teratomas servem para verificar a capacidade das células-tronco de originar todos os tecidos celulares. Isso porque os teratomas contem todos os tipos celulares derivados das três camadas germinativas do embrião. Trofoblasto – tecido extra embrionário responsável pela implantação, desenvolvimento em placenta, e controle das trocas de oxigênio e matabólitos entre a mãe e o embrião. Figura 1. Termos chave usados no debate da plasticidade das células-tronco.
Célula- tronco
transdiferenciação Célula progenitora
Célula madura
diferenciação desdiferenciação
transdiferenciação

84
ATIVIDADES

85
ATIVIDADE DE ACOLHIMENTO E CONTRATO PEDAGÓGICO Curso: Desvendando as células-tronco: dos sonhos à realidade Nome da Oficina: “ÔNIBUS, uma viagem para...................... Metodologia: Oficina introdutória para o curso: Construção de um ônibus para dar início a uma viagem (curso) que será iniciada no dia 16 às 9:00 horas e chegará ao local de destino no dia 20 às 16:30 horas. Material: 1. Desenho de um ônibus grande. 2. Etiquetas adesivas:
2.1. Motor: retangulares 2x1cm 2.2. Parafusos: retangulares 0,5 x 0,5 cm aproximadamente 2.3. Carcaça: quadradas 2x2 2.4. Bagagem: 8x5 cm com borda colorida (desenho da alça da mala) 2.5. Combustível: retangulares 5x3cm. Recortar duas Gotas deixando o meio
unido. 2.6. Passageiros e motoristas: 2 cm de diâmetro branca ou colorida não metálica 2.7. Pedras do caminho – retangulares 5x3 ou quadradas 5x5 cm para serem
recortadas em formas de pedras com espaço para escrita no meio. 3. Dois fachos de luz para os faróis = avaliação. 4. Um envelope retangular padrão para ser colado em cima do ônibus onde serão
colocadas as malas com as bagagens. 5. 8 envelopes onde serão colocadas cada uma das diferentes etiquetas conforme as
metáforas. Os envelopes deverão ser identificados com o numero da ordem de acordo com a dinâmica pré-estabelecida e a metáfora correspondente.
6. Papel com o destino do ônibus. 7. 1 rolo de fita adesiva para colagem do envelope e dos faróis. 8. Crachás com lugar para colocar os nomes: Caso alguma atividade prevista durante o
curso for realizada por grupos, os grupos poderão ser identificados com esferas adesivas coloridas nos crachás.
Cronograma da atividade: Sugestões para a organização prévia dos professores:
1. Distribuir entre os participantes, na entrada para a sala de aula um crachá (em branco) e um envelope contendo todas as pecas de etiquetas que serão usadas durante a oficina.
2. Comunicar aos mesmos que deverão ter um lápis ou caneta na mão para os registros.
3. A cada situação, esclarecer qual a etiqueta a ser usada. 4. Distribuir, previamente entre os professores as diferentes funções: distribuição dos
crachás, colagem das etiquetas, recolhimento dos dados que serão entregues no momento da oficina, coordenador da fala introdutória, confecção do ônibus, etc.
Tempo total: 50 minutos Introdução: - 5 min. - Apresentação da idéia da viagem e construção do transporte para que ela
aconteça. - Explicar que a participação, no momento da construção sera de apenas uma fração
pequena do grupo (6 pessoas) porem que após o termino todos deverão colocar as

86
suas contribuições nos respectivos lugares para que possam ser conhecidas tanto pelos alunos como pelos professores.
- Explicar que o ônibus permanecera no local do curso para que todos tenham a oportunidade de conhecer o que foi escrito por todos.
- Definir ou colar o destino. Sugestão de pergunta para o inicio da atividade: - O que é necessário para que um ônibus possa ser usado para uma viagem? - Inicio da construção do ônibus:
o 3 min. Motor: as pecas do motor deverão ser colocadas na hora enquanto vão sendo lidas pelo monitor responsável; sugerimos que seja o conteúdo do curso previamente escritos nas pecas.
o 2 min. Parafusos: habilidades que os alunos e professores deverão
desenvolver para que a viagem aconteça; previamente escritas e coladas na hora unindo as pecas do motor.
o 5 minutos. Combustível: energia que mobilizara os participantes para a
viagem/curso. Cada participante devera escrever um grande desejo e um grande medo em relação a viagem que terá inicio = expectativas. 2 min. Para escrever e 3 min. para a colagem e leitura (se assim desejarem os participantes)
o 5 min. Carcaça: Limites ou regras construídas coletivamente. 3 min para
escrever e 2 min. para ver se todos estão de acordo.= validação das regras. Nesse momento os professores tb podem contribuir com sugestões.
o 3 min. Pedras do caminho: - problemas que poderão aparecer durante a
viagem/curso.
o 3 min. Faróis e espelhos retrovisores: Avaliação continua.
o 3 min. Bagagem: Sugestão: “O que na minha vida me motivou a fazer este curso ou participar desta viagem?”
o 5 min. finais.- Subida no ônibus; definição de papeis e partida. Cada
passageiro devera colocar o seu nome no crachá, desenhar a sua “cara” na etiqueta esférica e ir colocando –se em algum lugar no ônibus e falando o seu nome.

87
Levantamento de conceitos 1. Dentre as frases seguintes a que melhor define um gene é
a) uma unidade de transcrição na molécula de DNA b) uma molécula de DNA em um cromossomo c) uma cadeia simples de DNA que codifica um polipeptídio d) um segmento da molécula de DNA que codifica um polipeptídio e) não tenho certeza da resposta
2. Na célula eucariótica a atividades dos genes é regulada principalmente pelo controle
a) do início da transcrição b) da tradução c) do processamento das proteínas d) do splicing do pré-mRNA e) não sei responder
3. Comparando-se o zigoto de uma determinada espécie com uma célula muscular do organismo adulto verifica-se que
a) a seqüência de DNA muda dependendo do tipo de diferenciação celular b) a morfologia dos dois tipos de células é diferente c) o genoma dos dois tipos celulares é diferente d) o padrão de genes ativos e o mesmo e) não sei responder
4. Dois tipos celulares diferentes de um mesmo organismo, por exemplo, células nervosas e células musculares
a) apresentam uma coleção de proteínas diferentes b) possuem os mesmos genes em atividades c) apresentam genomas com diferentes seqüências de DNA d) diferenciaram-se somente durante o desenvolvimento embrionário e) não sei responder
5. Células-tronco são
a) todas as células presentes em um embrião em qualquer fase do desenvolvimento b) células presentes apenas em embriões de vegetais e animais c) células com potencialidade para originar diferentes tipos celulares d) células com alto grau de diferenciação e) não sei responder
6. Durante a diferenciação celular
a) células-tronco dividem-se com muita freqüência b) todos os genes do genoma estão em funcionamento c) células indiferenciadas originam células especializadas d) células diferenciadas transformam-se em células especializadas e) não sei responder

88
ESTUDO DIRIGIDO 1 IDENTIFICAÇÃO DE CONCEITOS RELATIVOS A CÉLULAS-TRONCO
Classe dividida em grupos de 6. Leia o texto acima. A compreensão do texto depende do entendimento de vários conceitos nele incluídos. Durante sua leitura grife pelo menos 15 desses conceitos. Os conceitos deverão ser classificados em uma das seguintes categorias: (1) conceitos básicos relacionados ao tema, (2) ética, (3) procedimentos experimentais. Após a leitura compare os conceitos selecionados por você durante a leitura com a seleção feita pelos demais colegas. Questões para discutir em grupo:
a) Nas células-tronco qual o alvo do estímulo proveniente das células vizinhas?
b) Qual é a diferença entre célula-tronco embrionária e de adulto? c) Qual a pergunta da equipe de pesquisadores para a realização do
experimento? d) Faça um esquema do experimento realizado pela equipe de Gerd
Hasenfuss. e) Qual o resultado obtido e o que ele comprova?
SÓ PARA ELES: CÉLULAS-TRONCO ADULTAS QUE SE COMPORTAM COMO EMBRIONÁRIAS PODEM DERRUBAR OBSTÁCULO À PESQUISA - MARINA VERJOVSKY - CIÊNCIA HOJE ON-LINE - 31/03/2006
Um estudo alemão pode representar o fim dos dilemas éticos enfrentados na pesquisa sobre células-tronco embrionárias. Essas células, capazes de se diferenciar em todos tecidos do organismo, são apontadas como o futuro da medicina regenerativa. No entanto, sua obtenção requer o sacrifício de embriões, proibido em muitos países e criticado por diversos setores da sociedade.
Agora, a equipe de Gerd Hasenfuss, da Universidade Georg-August, em Göttingen, mostrou que há no testículo de camundongos células-tronco adultas que, dependendo do meio usado para cultivo, adquirem propriedades das células embrionárias, ou seja, a capacidade de gerar qualquer tecido do organismo. Caso células do mesmo tipo sejam encontradas em humanos – como acreditam os autores do estudo –, um grande obstáculo à pesquisa na área cairá por terra.
No estudo publicado em 24 de março no site da revista Nature
Os pesquisadores constataram que, nessas condições, as células se comportavam como as células-tronco ditas pluripotentes, ou seja, elas eram capazes de se transformar em vários tipos celulares, como músculos, gordura, ossos, cartilagem ou tecido neuronal e intestinal.
, a equipe de Hasenfuss verificou que, quando essas células-tronco estão nos testículos dos camundongos, produzem espermatozóides devido ao estímulo de outras células locais. A equipe alemã resolveu testar o que aconteceria na ausência desse estímulo: para isso, as células foram injetadas na pele de camundongos ou cultivadas in vitro.
No entanto, os pesquisadores não conseguiram encontrar esse tipo celular no ovário de roedoras – eles ainda não sabem se elas realmente não existem ou se a equipe não foi capaz de identificá-las. Por ora, este avanço só é válido para os animais do sexo masculino. Potencialidades:
As células-tronco são importantes para a medicina porque podem ser úteis no tratamento de doenças cardíacas, leucemia, Alzheimer e Parkinson, além de permitirem a

89
reconstituição de medula óssea e de tecidos queimados ou destruídos sem risco de rejeição, caso o doador seja o próprio paciente.
No Brasil, o Instituto do Coração (Incor), vinculado ao Hospital das Clínicas da Universidade de São Paulo (USP), vem estudando o tratamento da insuficiência cardíaca com o transplante de células-tronco adultas extraídas da medula óssea dos pacientes e injetadas no músculo cardíaco. Os resultados preliminares indicam que se trata de um procedimento seguro.
Porém, o fato das células-tronco adultas não apresentarem a plasticidade das embrionárias restringe a sua utilização em alguns tratamentos, pois nenhuma delas foi capaz, até o momento, de se diferenciar em certos tipos celulares, como por exemplo, as células produtoras de insulina.
O trabalho do grupo alemão é importante justamente por reunir as qualidades de ambos os tipos de células-tronco. “As células adultas que apresentam plasticidade são um grande passo para acabar com os problemas éticos que vêm sendo enfrentados atualmente”, avalia o cardiologista José Eduardo Krieger, vice-coordenador do projeto que estuda células-tronco adultas no Incor. “Se esta nova descoberta se reproduzir em humanos pode ser um avanço importante para a pesquisa de reparação de órgãos humanos”.

90
ATIVIDADE 1. DIFERENCIAÇÃO DA HEMÁCIA Renato Chimaso dos Santos Yoshikawa
Objetivo da atividade: Visualizar a diferenciação da hemácia, estabelecendo a relação entre o sinal celular e o fenótipo. Material por grupo: Massa de modelar de 6 cores; contas de diferentes cores e tamanhos, 1m de fio colorido; 4 envelopes contendo as cartas de sinais. Metodologia: Modelagem de uma célula com as estruturas mais relevantes para o processo de diferenciação da hemácia.
a) Montagem inicial Modelar um PROERITROBLASTO de acordo com as seguintes características:
• É a maior célula da linhagem celular que forma a hemácia; represente-o com 10 cm de diâmetro;
• Apresenta os elementos característicos de uma célula que sintetiza intensamente proteínas; represente 10 poli ribossomos;
• O núcleo é esférico e central; tem um ou dois grandes nucléolos, a cromatina é pouco condensada;
• Ao redor do núcleo, encontram-se seis mitocôndrias; • Há pequena produção de hemoglobina; representar 10 moléculas de hemoglobina.
b) Em cada uma das quatro etapas do processo de diferenciação, cada grupo deverá escolher três cartas de sinais pertinentes. Quando possível, o efeito de cada sinal deve ser modelado em massinha.
• Selecionar do envelope I as três cartas de sinais que deverão ser aplicados à
célula modelada inicialmente para que se transforme no ERITROBLASTO BASÓFILO.
• Selecionar do envelope II as três cartas de sinais que deverão ser aplicados à célula para que se transforme no PRÉ-RETICULÓCITO.
• Selecionar do envelope III as três cartas de sinais que deverão ser aplicados à célula para que se transforme no RETICULÓCITO.
• Selecionar do envelope IV as três cartas de sinais que deverão ser aplicados à célula para que se transforme na HEMÁCIA. Abaixo estão representadas as quatro etapas de diferenciação celular para a
transformação de um PROERITROBLASTO em HEMÁCIA:

91
II. Pré-reticulócito
III
IV.
polirribossomo
mitocôndria hemoglobina
I. Eritroblasto basófilo

92
ATIVIDADE 2 UM CASO MUITO INTERESSANTE DE DIFERENCIAÇÃO CELULAR
Vivian Lavander Mendonça - Adaptado de Differentiation in the Amoeboflagellate Naegleria, disponível em http://7e.devbio.com/article.php?ch=12&id=10
Quando consideramos o processo de diferenciação celular, logo pensamos no desenvolvimento dos animais, que apresentam centenas de tipos celulares diferentes geradas a partir de uma única célula, o zigoto. Existe, no entanto, um caso muito interessante de diferenciação celular que ocorre em um ser unicelular: trata-se do protozoário Naegleria gruberi. A célula de N. gruberi é mais comumente encontrada na forma amebóide, locomovendo-se por meio de pseudópodes entre as partículas do solo. Os pseudópodes também são usados na captura de bactérias, que constituem seu alimento. Em algumas situações, como após uma chuva, torna-se mais difícil a captura das bactérias, que ficam dispersas na água que passa a ocupar os espaços entre as partículas do solo. Em pouco mais de uma hora, N. gruberi se transforma em uma célula alongada e portadora de dois longos flagelos. Com eles, os protozoários locomovem-se com agilidade no meio líquido e localizam rapidamente seu alimento. Observe as principais mudanças no fenótipo desse protozoário no gráfico abaixo:
Figura. Diferenciação em indivíduos de Naegleria gruberi. Indivíduos com célula amebóide foram cultivados em meio contendo bactérias; no tempo 0, foi adicionada água no meio, simulando o que ocorre após uma chuva. Após 80 minutos, quase 100% da população apresenta a forma flagelada. Para que se formem flagelos, a célula deve sintetizar diversas proteínas, como a tubulina, que compõe o corpúsculo basal e os microtúbulos. Essa proteína está ausente nos indivíduos com fenótipo amebóide, o que significa que os genes envolvidos estão “desligados” nessas células. Como a célula “sabe” que deve ativar modificar a expressão de seus genes e sintetizar as proteínas dos flagelos? Temos aí um indício de que existe um processo de sinalização entre a célula e o meio externo. A presença ou ausência de determinadas substâncias no meio extracelular pode ser detectada pela membrana plasmática e pode desencadear uma série de reações químicas no citoplasma. Essa cascata de reações pode gerar moléculas capazes de
A = início da síntese de tubulina
B = síntese intensa de tubulina
C = organizam-se os corpúsculos basais e em seguida os flagelos tornam-se visíveis; célula se torna arredondada
D = célula se torna alongada

93
entrar no núcleo da célula e ativar/desativar certos genes. No caso do protozoário, a mudança no meio externo desencadeia um processo que leva à ativação dos genes que comandam a síntese das proteínas flagelares. Assim, a mudança na forma da célula resulta de modificações na expressão dos genes de cada indivíduo. Utilizando marcadores de RNAm, pesquisadores observaram que, após a transcrição dos genes que codificam a tubulina, as moléculas de RNAm se dirigem para a periferia da célula e se agrupam em uma região determinada, onde ocorrerá a síntese das subunidades que vão compor a proteína e onde, em seguida, surgirão os flagelos. O citoesqueleto participa desse processo de transporte e ancoragem das moléculas de RNAm em pontos específicos do citoplasma. Temos aí um indício de que existe também uma sinalização interna, responsável pela orientação das substâncias dentro da célula. Após a síntese protéica, moléculas de tubulinas organizam-se formando microtúbulos. Outras proteínas também participam da estrutura do flagelo. Assim, podemos destacar três eventos fundamentais no processo de diferenciação celular:
• a sinalização entre a célula e o meio externo a ela, resultando em mudanças na expressão gênica; • a sinalização interna, da qual participa o citoesqueleto, resultando em orientação de moléculas dentro da célula; • a organização das proteínas após a síntese protéica, resultando em mudanças no fenótipo da célula.
Adaptado de Differentiation in the Amoeboflagellate Naegleria, disponível em http://7e.devbio.com/article.php?ch=12&id=10 Outras fontes de consulta: Han, J. W.; Park, J. H.; Kim, M.; Lee, J. 1997. mRNAs for microtubule proteins are
specifically colocalized during the seqüencial formation of basal body, flagella and cytoskeletal microtubules in the differentiation of Naegleria gruberi. The Journal of Cell Biology, vol. 137: 871-879.
Walsh, C. 1984. Synthesis and assembly of the cytoskeleton of Naegleria gruberi flagellates. The Journal of Cell Biology, vol. 98: 449-456.

94
ATIVIDADE - DIFERENCIAÇÃO NO PROTOZOÁRIO NAEGLERIA GRUBERI
Desafio da atividade: Representar as etapas de diferenciação do protozoário Naegleria gruberi a partir da escolha de sinais apropriados à diferenciação celular.
Número de participantes: organizar-se em grupos de no máximo 6 pessoas.
Materiais:
• Massa de modelar (6 cores); • Total de 28 Cartas de Sinais, organizadas em 4 conjuntos (A, B, C e D); • Guia de figuras.
Montagem inicial:
Modele um indivíduo amebóide de Naegleria gruberi de acordo com as seguintes características:
a. O núcleo é esférico; b. O citoplasma apresenta as organelas características de uma célula eucariótica
animal – por simplificação, represente apenas os ribossomos, as mitocôndrias e as vesículas, que compreendem: fagossomos, lisossomos, vacúolos digestivos;
c. No estágio amebóide o núcleo e as organelas citoplasmáticas não apresentam posições fixas na célula;
d. Represente o indivíduo amebóide capturando uma bactéria. Procedimento
Para cada etapa (A, B, C e D), o grupo deve escolher três cartas de sinais a serem aplicados na célula. Discutindo em conjunto, julguem quais sinais são necessários para a diferenciação celular.
O efeito de cada sinal deve ser representado em massinha pelo grupo. Guia de figuras
Fotografias ao microscópio óptico; preparação com lugol:
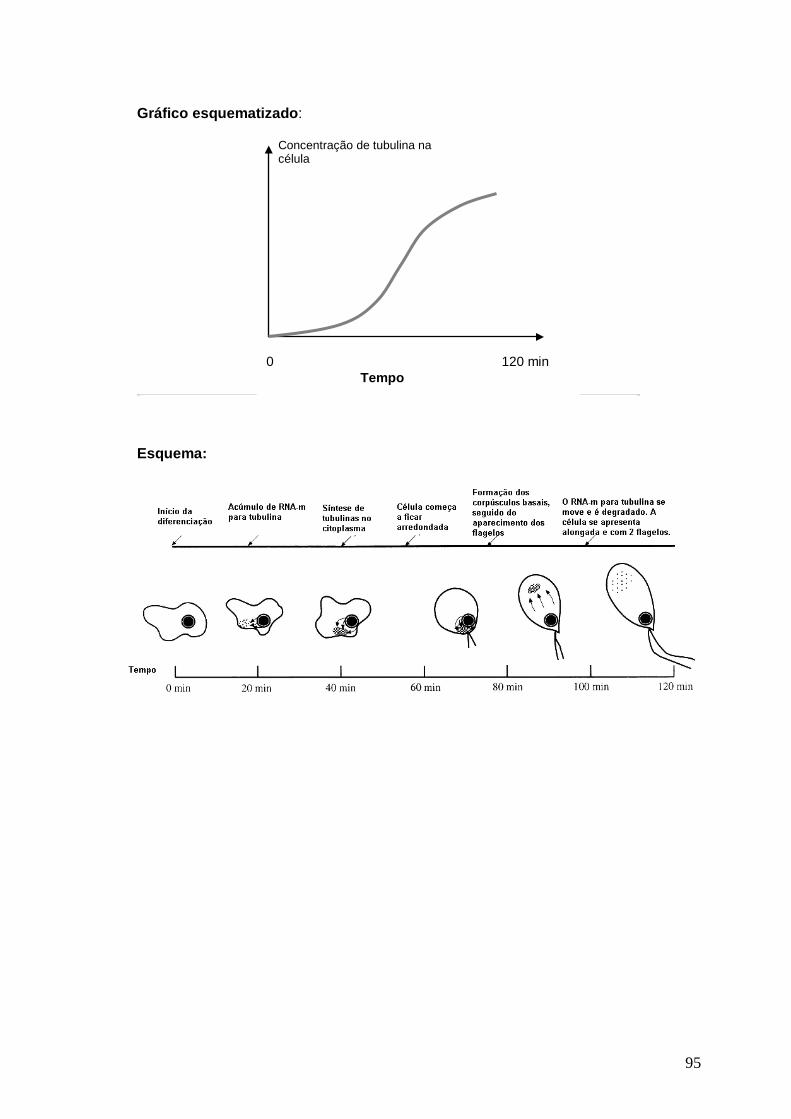
95
Gráfico esquematizado:
Esquema:
Concentração de tubulina na célula
0 120 min Tempo

96
DIFERENCIAÇÃO CELULAR EM TECIDO ÓSSEO Renato Chimaso dos Santos Yoshikawa
A Figura 1 apresenta a organização do tecido ósseo: imersos numa matriz calcificada encontram-se os osteócitos, ou seja, as células ósseas diferenciadas. Outros tipos celulares: as células precursoras mesenquimais e os osteoblastos (célula osteogênica, não completamente diferenciada, que sintetiza a parte orgânica da matriz óssea) estão localizadas na região não calcificada desse tecido.
Figura 1: Distribuição das células precursoras mesenquimais, osteoblastos e osteócitos no tecido ósseo.
No processo de osteogênese, isto é, de formação do osso, os osteócitos e os
osteoblastos originam-se por diferenciação celular a partir de células precursoras mesenquimais (Figura 2).
Figura 2: Via de diferenciação celular em que uma célula precursora mesenquimal origina osteoblasto e osteócito. O osteoblasto produz a matriz não calcificada composta principalmente por colágeno tipo I. O osteócito é uma célula em forma de estrela.
Na diferenciação osteoblástica, são expressos genes que são considerados
marcadores fenotípicos de osteoblasto, como o gene que produz o colágeno tipo I, substância característica da matriz orgânica do osso.
Para que haja a diferenciação celular osteoblástica, é necessária a presença de um fator de transcrição específico dos osteoblastos, o fator Cbfa1 (core binding factor a1), o mais precoce e específico marcador da osteogênese. Proteínas morfogênicas do osso (BMP) e o TGF (transforming growth factor) induzem a expressão de Cbfa 1.
célula precursora mesenquimal osteoblasto osteócito
células precursoras de osteoblasto
osteoblasto
matriz não calcificada
matriz calcificada
osteócito

97
Em comparação com o osteoblasto, o osteócito apresenta uma forma estrelar, aspecto conferido graças à presença de projeções celulares finas denominadas processos dendríticos.
Estudos de biologia molecular identificaram nesses processos dendríticos a proteína E11 que desempenha papel fundamental na formação dessas estruturas.
A proteína E11 interage com outras proteínas, a CD44 e a ezrina, formando um complexo que ativa a organização de filamentos de actina e miosina em regiões da célula em que se formarão os processos dendríticos (Figura 3).
Figura 3: papel da proteína E11 na formação do processo dendrítico em osteócito. Note que a proteína CD44, ligada à E11, ativa a proteína ezrina, que atua na interação entre miosina e actina, organizando o citoesqueleto.
Nas células precursoras mesenquimais o gene da proteína E11 está inativo e,
assim sendo, os processos dendríticos não estão presentes no fenótipo dessas células. A ativação do gene E11 ocorre somente na presença de proteínas específicas
como a Sp1 e Ap1, que atuam como fatores de transcrição para o gene E11. Por sua vez, tais proteínas foram sintetizadas a partir da ativação dos seus respectivos genes, o que acontece, por exemplo, graças a sinais mecânicos oriundos do meio ambiente e recebidos pelo osteócito em estágio inicial de desenvolvimento. Referências:
Harris, S. E., Bonewald, L. F., Harris, M. A., Sabatini, M., Dallas, S., Feng, J. Q., Ghosh-Choudhury, N.,
Wozney, J., e Mundy, G. R. 1994. Effects of transforming growth factor beta on bone nodule formation and expression of bone morphogenetic protein 2, osteocalcin, osteopontin, alkaline phosphatase, and type I collagen mRNA in long-term cultures of fetal rat calvarial osteoblasts. J. Bone Miner. Res. 9: 855–863.
Karsenty, G. Transcriptional control of osteoblast differentiation. Endocrinoly 142: 2731-2733. Kato, Y., Windle J. J., Koop, B. A., Mundy, G. R., Bonewald, L. F. 1997. Establishment of an osteocyte-like cell
line, MLO-Y4. J. Bone Miner. Res. 12: 2014–2023. Zhang, K., Barragan-Adjemian, C., Ye, L., Kotha,S., Dallas, M., Lu, Y., Zhao, S., Harris, M., Harris, S. E., Feng,
J. Q. e Bonewald, L. F. 2006. E11/gp38 Selective Expression in Osteocytes: Regulation by Mechanical Strain and Role in Dendrite Elongation. Molecular and Cellular Biology, p. 4539–4552.
actina
miosina
E11
CD44
ezrina
processo dendrítico

98
ATIVIDADE 3 DIFERENCIAÇÃO CELULAR EM OSSO
Introdução
No osso, os osteoblastos são as células que originam osteócitos. Uma característica morfológica própria dos osteócitos é a presença de processos dendríticos, projeções celulares longas e finas. Objetivo da atividade: A partir de um osteoblasto, formar um osteócito, ou seja, uma célula com processos dendríticos, entendendo como se dá a regulação gênica necessária para essa diferenciação celular. Materiais Tipo de material O que representa
Pano Membrana plasmática 4 fios de telefone Filamentos de actina 4 pregadores Proteína acoplada à membrana Tabuleiro circular Núcleo 15 peças vermelhas Proteínas 8 cartelas quadradas DNA; RNAm; RNAt; RNAr Montagem inicial
• Dispor o pano ao redor do tabuleiro circular simulando uma célula com citoplasma e núcleo: essa célula é o osteoblasto.
• Espalhar pelo citoplasma do osteoblasto as peças (proteínas), os fios e as cartelas.
Procedimento: Siga as instruções abaixo:
1. Encontre a única proteína que se liga ao promotor (I) do gene Sp1. 2. Essa proteína, uma vez ligada ao promotor do gene Sp1, ativa sua
transcrição. Identifique o produto da transcrição desse gene dentre as cartelas. Coloque-o na área tracejada (II).
3. Continuando o processo de expressão desse gene, forma-se outro produto, que se dirige ao citoplasma (III). Escolha-o dentre as peças.
4. Esse último produto deve se ligar agora ao promotor do gene E11. Houve realmente o encaixe da peça, isto é, a ligação (IV)? Se não, escolha outra peça até achar aquela que se liga ao promotor do gene E11.
5. A ligação dessa proteína no promotor do gene E11 ativa sua transcrição. O produto da transcrição deve ser identificado dentre as cartelas quadradas (V).
6. Os transcritos são traduzidos dando origem a proteína E11, simbolizada pelo pregador (VI).
7. As moléculas da proteína E11 aderem-se à membrana e junto com outras moléculas orienta os filamentos de actina. Prenda os filamentos de actina e forme os processos dendríticos (VII).
Considerações finais Acredita-se que os osteócitos “tocam-se” graças aos processos dendríticos, o que estabelece uma comunicação entre essas células. A comunicação é fundamental para os osteócitos, uma vez que eles se encontram imersos em uma matriz calcificada. Sem os processos dendríticos, a comunicação celular ficaria comprometida.

99
ATIVIDADE 4 - DIFERENCIAÇÃO DE CÉLULAS EMBRIONÁRIAS: A FORMAÇÃO DO TROFOBLASTO
Vivian Lavander Mendonça Introdução:
• Dois grupos, organizados lado a lado, participarão desta oficina cada um recebendo um kit de materiais.
• O objetivo da oficina é compreender essencialmente o papel dos fatores de transcrição na regulação da atividade dos genes e a relação entre o que acontece no nível gênico com a diferenciação de uma célula. Vale ressaltar que o processo de diferenciação é bastante complexo e o processo aqui utilizado foi simplificado. No decorrer da oficina os participantes também utilizarão conhecimentos básicos de biologia celular e molecular.
Materiais:
• modelo de célula (blastômero), que servirá de “tabuleiro”, constituído de base de pano representando o citoplasma e os limites da célula (onde estaria a membrana plasmática), e a prancha representando o núcleo com trechos de dois cromossomos (A e B) em seu interior (ao modelo não obedece às proporções reais entre as estruturas);
• peças em cartolina com as opções: DNA, RNAr (RNA ribossômico), RNAt (RNA transportador) e RNAm (RNA mensageiro);
• peças em resina com diferentes contornos; • peças tubulares com região de encaixe; • clipes coloridos; • texto de apoio “As primeiras mudanças morfológicas no embrião”, disponível na
página 101 da apostila (consultar durante o jogo se necessário; recomendamos a leitura do texto ao término da oficina).
Regras da oficina:
1. Disponha a prancha-núcleo na base de pano que representa o citoplasma da célula. Disponha clipes coloridos em uma região do citoplasma. Esses clipes representam estímulos que uma célula pode enviar para uma vizinha.
2. No núcleo, inicia-se a transcrição do gene S, localizado no cromossomo A. Localize a peça que indica corretamente qual o produto da transcrição e coloque-a no quadro amarelo correspondente.
3. Após a transcrição do gene S, ocorre a tradução no citoplasma, com a formação da proteína S.
4. A proteína S atravessa a membrana nuclear e, dentro do núcleo, liga-se à região promotora do gene CDH-1, localizada no cromossomo B. Sabendo disso, identifique a peça que representa a proteína S e coloque-a no devido lugar.
5. A proteína S ligada à região promotora do gene CDH-1 impede a transcrição desse gene.
6. Aproxime seu blastômero do da outra equipe. Quando os limites se tocarem, envie os estímulos para seu vizinho e receba os dele. Os estímulos vão até o núcleo e impedem a transcrição do gene S. Represente esta etapa e represente as conseqüências desse bloqueio, retirando todos os transcritos do gene S e todas as proteínas S de sua célula.
7. Agora a região promotora do gene CDH-1 está desbloqueada, e o gene pode ser transcrito. Localize a peça que indica corretamente qual o produto da transcrição e coloque-a no quadro amarelo correspondente.
8. Após a transcrição do gene CDH-1, ocorre a sua tradução no citoplasma, com a formação da proteína E-caderina. Essa proteína vai para a membrana

100
plasmática do blastômero, na região de contato com a célula vizinha. Represente esta etapa utilizando a peça adequada.
9. Agora o seu blastômero vai aderir ao blastômero do outro grupo através das E-caderinas. Represente esta etapa e também o achatamento da célula manipulando a base de pano.
10. O desenho abaixo representa um blastocisto, em corte transversal. Identifique no desenho abaixo em qual das camadas – 1, 2, 3 ou 4 – o seu blastômero estaria e justifique a escolha:

101
AS PRIMEIRAS MUDANÇAS MORFOLÓGICAS NO EMBRIÃO
O início do desenvolvimento embrionário nos vertebrados caracteriza-se por uma
fase de segmentação, que envolve uma série de divisões mitóticas a partir do zigoto, seguida da formação da blástula (ou blastocisto, no caso dos mamíferos) e da gástrula. Nesses dois últimos estágios, além das divisões mitóticas ocorrem rearranjos espaciais das células do embrião e o início da diferenciação dos folhetos embrionários, que em outro momento darão origem aos diversos tecidos e órgãos. Vamos analisar aqui alguns dos fatores envolvidos na determinação do arranjo das células no embrião logo no início de seu desenvolvimento, tomando como exemplo o embrião de camundongo, o mais estudado entre as espécies de mamíferos. Formação do trofoblasto: a primeira modificação na arquitetura celular do embrião Após a fecundação, o zigoto inicia a primeira divisão mitótica, originando duas células morfologicamente idênticas chamadas blastômeros. Cada blastômero sofre novas divisões, chegando ao estágio de mórula (amora, em Latim). Os blastômeros mantêm-se unidos uns aos outros, formando uma estrutura mais compacta. A figura a seguir resume esses eventos iniciais da vida embrionária:
Figura 1. Início da segmentação no embrião.
A partir do estágio de 16 células, os blastômeros mais superficiais sofrem um
achatamento progressivo e passam a expressar em suas membranas plasmáticas estruturas de adesão típicas de células epiteliais, como as zônulas de adesão, as zônulas de oclusão e as junções comunicantes (gap junctions). Essas células modificadas formam o primeiro epitélio do embrião, o trofoblasto.
As células do trofoblasto migram determinando o aparecimento de uma cavidade interna, a blastocele. As células embrionárias que não sofrem essas transformações passam a compor a massa celular interna ou botão embrionário. O embrião apresentando trofoblasto, massa celular interna e blastocele é chamado blastocisto.
Figura 2: Esquema de blastocisto, representado em corte. (1) Massa celular interna; (2) cápsula protetora; (3) trofoblasto; (4) blastocele. Repare a diferença morfológica entre as células da massa interna e da massa externa do embrião.

Adesão entre células: muito além de uma simples união Observa-se que, à medida que o número de células aumenta na mórula, a
formação de estruturas de adesão também aumenta, como indica a figura a seguir. O resultado é a compactação da mórula.
Figura 3. Mudanças na forma e na ocorrência de estruturas de adesão de membrana plasmática em células da mórula.
A formação das junções celulares nos blastômeros está relacionada com a
presença de diversas proteínas que se ligam à membrana plasmática e ao citoesqueleto. Uma dessas proteínas é a E-caderina (“E” de epitelial), codificada pelo gene CDH-1 e fundamental para a formação das zônulas de adesão.
Figura 4. Detalhe da adesão entre duas células vizinhas com a participação da E-caderina.
O zigoto herda de sua mãe moléculas de E-caderina, pois sua expressão ocorre no óvulo. Após a primeira divisão do zigoto, são os próprios blastômeros que sintetizam a proteína, indicando que o gene CDH-1 está ativado.
Para que o gene CDH-1 permaneça ativo, um outro gene, o Snail, deve estar bloqueado. Quando o gene Snail, localizado em outro cromossomo, não está bloqueado, seu transcrito é traduzido em uma proteína que bloqueia a transcrição do gene CDH-1.

103
Aparentemente, existe uma comunicação entre os blastômeros mais externos da mórula. Graças a essa comunicação ocorre liberação de fatores que bloqueiam o gene Snail e como conseqüência o gene Cdh-1 é ativado. Assim, quando uma célula se desenvolve tendo contato íntimo com células vizinhas a expressão de E-caderina aumenta e há formação das junções entre as células, o que resulta no aumento da adesão entre os blastômeros, na compactação da mórula e depois na formação das células do trofoblasto. Nas células que dão origem à massa interna do embrião o gene Snail está ativado e a expressão de E-caderina é reduzida em relação às células do trofoblasto.
Observe novamente a figura 3. É possível perceber que não ocorre apenas um aumento no número de junções entre as células; também ocorre a polarização de células embrionárias, sendo possível distinguir as regiões basal, laterais e o apical. Cada uma dessas áreas apresenta estruturas distintas na membrana plasmática: microvilos na região apical, zônulas de oclusão e de adesão e junções comunicantes na região lateral, estruturas de adesão na região basal. O citoesqueleto também se reorganiza, mudando o formato dessas células, que passam a ser achatadas. As células embrionárias que sofrem essas modificações durante a formação do blastocisto passam a formar o trofoblasto.
A relação entre adesão celular e expressão de E-caderina também se verifica nas células do organismo adulto. A redução da expressão de E-caderina em células tumorais humanas já foi verificada em certos tipos de tumores invasivos de pele, cabeça, pulmão, mama, tireóide, estômago, intestino e ovário. Essa redução é um indício de malignidade, pois as células tumorais estão mais desagregadas e podem, sob influência de outros fatores, se espalhar para outros tecidos do corpo.
O que apresentamos aqui, embora de modo bastante simplificado, mostra que o contato entre células vizinhas possui um efeito sinalizador e que a comunicação intercelular é um fator fundamental na regulação do processo de diferenciação. Mostra, ainda, que os sinais recebidos por uma célula resultam em diferenciação porque a atividade de certos genes é alterada.
Fontes de consulta:
• Alberts et al. Molecular Biology of the Cell. Garland Science, 2002. • Jamora, C. et al. Links between signal transduction, transcription and adhesion in epithelial
bud development. Nature (2003) 422: 317-322. • University of Washington - Gene Reviews:
www.ncbi.nlm.nih.gov/books/bv.fcgi?call=bv.View..ShowTOC&rid=gene.TOC • Eurekah Bioscience Database – www.eurekah.com/
Questões:
a) Faça um esquema mostrando a relação funcional entre os genes Cdh-1 e Snail no zigoto e após a primeira divisão célular.
b) Qual a conseqüência morfológica da ativação do gene Xdh-1?

104
ATIVIDADE 5 - DIFERENCIAÇÃO MUSCULAR Paula Cristina Gorgueira Onofre
Objetivo da atividade:
Entender como se dá a diferenciação muscular e quais são suas etapas. Materiais
• Massinha de modelar de uma única cor • Miçangas vermelhas, azuis e verdes • Pedaço de papel plastificado representando o “tecido” • Cartão plastificado representando o “núcleo” • Envelope com as “proteínas”
Montagem inicial
• Retire o material do envelope e disponha-o sobre a bancada.
Procedimento Siga as instruções abaixo e vá respondendo as perguntas a medida que manipula o
material: 1. Com a massa de modelar modele mioblastos (célula precursora do fibroblasto)
com aproximadamente 3 cm de diâmetro e coloque-os dentro da área no tecido. Um novo mioblasto deve ser colocado a cada 5 segundos.
2. Quando um mioblasto tocar cinco ou mais, “ligue” o gene MyoD no “núcleo”.
• Qual o estímulo para esta ativação? • O que é produzido com a ativação do gene MyoD? • Em quantas etapas? • Marque a ativação do MyoD nos mioblastos usando a miçanga vermelha.
Nesta etapa a célula já está comprometida com a via de diferenciação muscular.
3. Repita os passos anteriores até que sejam produzidos MyoD em 3 ou mais
células. Neste ponto, no núcleo, remova o inibidor do gene 2 e faça o MyoD atuar. • O que acontece? Qual o papel funcional do MyoD? • Marque as células com o gene 2 ativo utilizando o percevejo azul e
alongue-as no formato de um fuso, mantendo-as na mesma posição no tecido.
4. No núcleo, faça o outro fator de transcrição atuar nos genes específicos
musculares. • O que acontece? • O que é produzido? • Marque a ativação dos outros genes com um percevejo verde e funda estas
células marcadas duas a duas. Nesta etapa a célula é chamada de miotubo, está completamente diferenciada e torna-se contrátil.

105
Texto de apoio DESENVOLVIMENTO MUSCULAR
O músculo esquelético dos vertebrados é derivado de estruturas mesodérmicas segmentadas, chamadas somitos (figura 1). A diferenciação muscular é precedida por muitos passos, durante os quais as células precursoras migram, proliferam e finalmente são determinadas pela expressão de fatores miogênicos.
O principal fator miogênico é o MyoD. Esta proteína é um fator de transcrição que leva a ativação de diversos genes, responsáveis pela modificação da célula precursora mesodermal em outra, já determinada, chamada de mioblasto. Os mioblastos já apresentam comprometimento na via de diferenciação muscular, porém ainda não expressam as proteínas do músculo diferenciado e são capazes de proliferar.
Quando submetidos a alguns estímulos específicos os mioblastos param de proliferar e prosseguem na via de diferenciação. O MyoD passa a ativar outros fatores de transcrição (como a miogenina), que vão agir apenas nos genes das células de músculo. Após uma cascata de eventos regulatórios passam a expressar miosina e outras proteínas musculares constituintes dos sarcômeros organizados, fundem-se, alongam-se e assim passam a constituir as estruturas do músculo propriamente dito, chamadas miotubos (figura 2).
Após o nascimento, algumas células permanecem determinadas, porém não diferenciadas no tecido muscular (no estágio de mioblasto). São as chamadas células satélite, responsáveis pela regeneração muscular ao longo da vida do indivíduo. Quando são necessárias, como no caso de lesões musculares, as células satélite passam pelas mesmas etapas de diferenciação já descritas acima e podem restaurar as fibras musculares, tanto pela sua fusão nos locais lesionados (causando hipertrofia) quanto pela formação de novas fibras (causando hiperplasia). Uma característica das fibras regeneradas é a posição dos núcleos celulares, que se localizam centralmente, enquanto que nas fibras normais os núcleos localizam-se nas periferias do sincício (figura 3).
Figura 1: Cortes transversais mostrando o desenvolvimento da camada mesodérmica. Dias após o desenvolvimento: A. 17. B. 19. C. 20. D. 21. A camada fina do mesoderma dá origem aos somitos.

106
Figura 2. Etapas da diferenciaç
ão muscular.
Figura 3. Processo de regeneração muscular
Referências: http://connection.lww.com/Products/sadler/imagebank.asp http://departments.oxy.edu/biology/Franck/Bio130S_2002/bio130_april26_lecture_devbio.htm http://www.yorku.ca/thawke/images.html Buckingham, M; Bajard, L.; Chang, T; et al. The formation of skeletal muscle: from somite to limb. J. Anat. (2003) 202, pp
59-68. Brand-Saberi, B.; Christ, B. Genetic and epigenetic control of muscle development in vertebrates. Cell tissue res (1999)
296:199-212
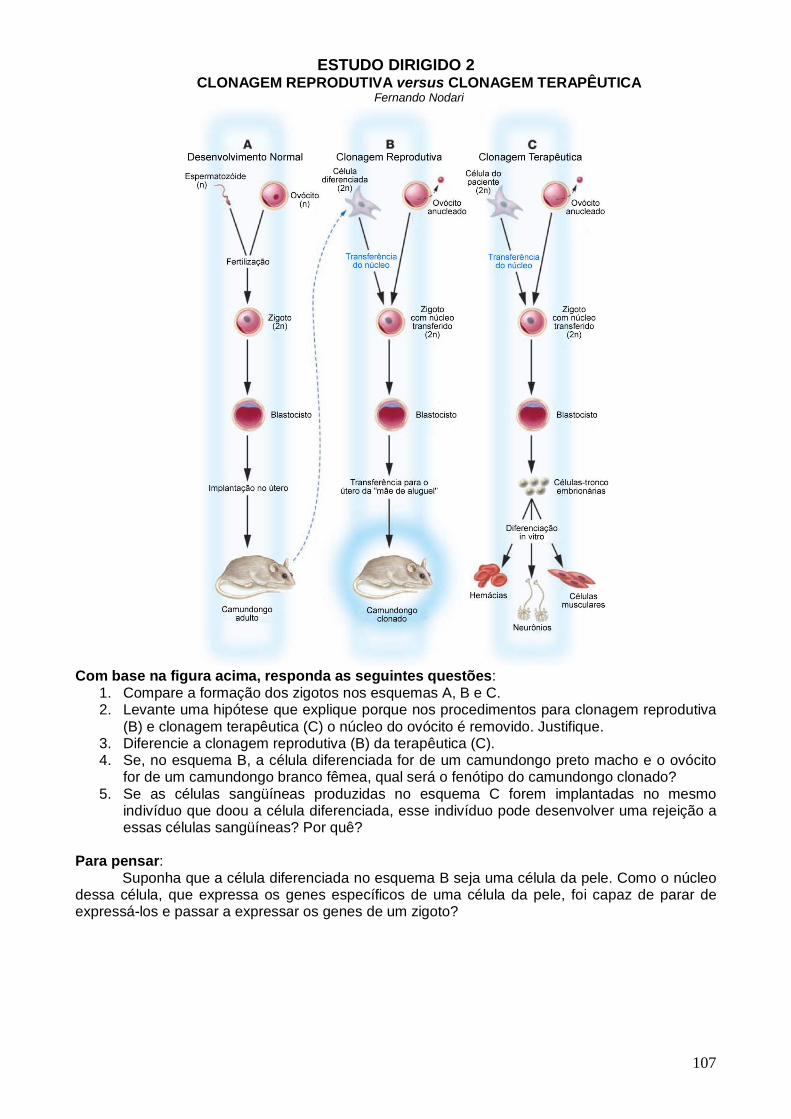
107
ESTUDO DIRIGIDO 2 CLONAGEM REPRODUTIVA versus CLONAGEM TERAPÊUTICA
Fernando Nodari
Com base na figura acima, responda as seguintes questões:
1. Compare a formação dos zigotos nos esquemas A, B e C. 2. Levante uma hipótese que explique porque nos procedimentos para clonagem reprodutiva
(B) e clonagem terapêutica (C) o núcleo do ovócito é removido. Justifique. 3. Diferencie a clonagem reprodutiva (B) da terapêutica (C). 4. Se, no esquema B, a célula diferenciada for de um camundongo preto macho e o ovócito
for de um camundongo branco fêmea, qual será o fenótipo do camundongo clonado? 5. Se as células sangüíneas produzidas no esquema C forem implantadas no mesmo
indivíduo que doou a célula diferenciada, esse indivíduo pode desenvolver uma rejeição a essas células sangüíneas? Por quê?
Para pensar:
Suponha que a célula diferenciada no esquema B seja uma célula da pele. Como o núcleo dessa célula, que expressa os genes específicos de uma célula da pele, foi capaz de parar de expressá-los e passar a expressar os genes de um zigoto?

108
ATIVIDADE 6 – JOGO DA CÉLULA-TRONCO
Silvio Higa Para jogar com tabuleiro:
1. Cada jogador escolhe uma peça identificada por cor diferente. 1. Inicia o jogo a pessoa que obtiver o maior número ao jogar um dado. 2. O primeiro jogador retira uma carta do monte e lê a pergunta (inclusive as alternativas)
para o jogador que se encontra à sua direita. 3. Se o jogador acertar, avança seis casas, mas se errar, recua duas. O jogador pode optar
por passar a vez, dando a chance para o próximo a sua direita responder. Este jogador também pode passar a vez. Se nenhum jogador acertar a resposta a carta utilizada volta para o final do monte.
4. O segundo jogador retira uma nova carta e reinicia a rodada. 5. Vence aquele que chegar primeiro no final do tabuleiro.
Para jogar sem tabuleiro: 1. Sortear a pessoa que iniciará o jogo. 2. O primeiro jogador deve retirar uma carta do monte e ler (inclusive as alternativas) para
o jogador que se encontra à sua direita. 3. Se o jogador acertar, fica com a carta, se errar, a carta é eliminada do jogo. 4.. O jogo termina após o final do tempo previamente determinado pelo grupo ou quando
terminarem as cartas. 6. Vence aquele que acumular maior quantidade de cartas.
Observações:
• O tabuleiro encontra-se nas páginas 109 e 110 • Os enunciados das cartas encontram-se nas páginas 111 a 117
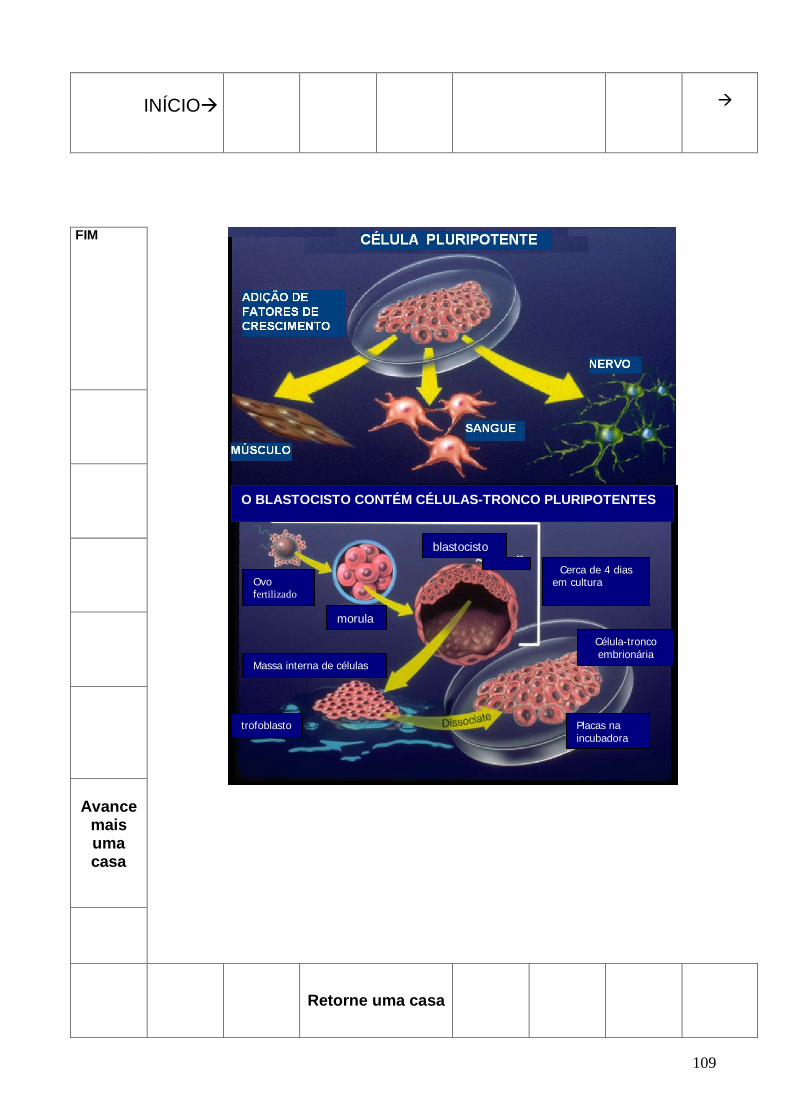
109
INÍCIO
FIM
Avance
mais uma casa
Retorne uma casa
O BLASTOCISTO CONTÉM CÉLULAS-TRONCO PLURIPOTENTES
Ovo fertilizado
morula
blastocisto
Cerca de 4 dias em cultura
Placas na incubadora
Célula-tronco embrionária
Massa interna de células
trofoblasto

110
Avance mais uma
casa
Avance mais duas casas
Retorneuma casa
Avance mais uma
casa

111
Cartas do jogo
Célula capaz de formar todos os tipos celulares Exemplos: zigoto e blastômeros. a) Totipotente. b) Pluripotente. c) Multipotente.
Célula capaz de gerar os três tipos de células germinativas (ectoderma, mesoderma e endoderma), ou seja, tem o potencial para se desenvolver nos mais de 200 tipos celulares conhecidos do corpo humano. a) Célula-tronco adulta. b) Pluripotente. c) Multipotente.
Célula capaz de formar vários tipos celulares. Exemplo: célula-tronco adulta. a) Totipotente. b) Pluripotente. c) Multipotente
Célula capaz de formar poucos tipos celulares. Exemplo: célula precursora mielóide. a) Multipotente b) Oligopotente. c) Nulipotente.
Célula incapaz de formar outros tipos celulares. Exemplo: hemácia. a) Multipotente b) Oligopotente. c) Nulipotente.
Célula capaz de se dividir por períodos indefinidos sem dar origem a células especializadas. a) pluripotente. b) oligopotente. c) nulipotente.
Célula indiferenciada encontrada em tecidos adultos que pode renovar-se a si mesma (com certas limitações) e se diferenciar em todas as células especializadas do tecido do qual foi originada. a) Célula-tronco adulta. b) Célula-tronco embrionária. c) zigoto
Célula indiferenciada encontrada no embrião que tem o potencial para se diferenciar numa ampla variedade de células especializadas. a) Célula-tronco adulta. b) Célula-tronco embrionária. c) Zigoto.
É o processo através do qual uma célula não especializada torna-se especializada. a) Mitose. b) Diferenciação. c) Transcrição.
É a capacidade de uma célula-tronco adulta de um tecido gerar células especializadas de um outro tecido. Exemplo: células-tronco hematopoiéticas podem gerar neurônios. a) Transcrição. b) Diferenciação. c) Plasticidade.
São obtidas de blastocisto pré-implantacionais, cerca de 5 dias após a fertilização. a) Célula-tronco embrionária ou celular pluripotente. b) Célula totipotente ou zigoto. c) Célula unipotente ou nulipotente.
Podem ser encontradas no cérebro, medulas ósseas, sangue periférico, músculos esqueléticos, tecido epitelial da pele e do tubo digestório, córnea, polpa dentária, retina, fígado e pâncreas. a) Célula-tronco adulta. b) Célula-tronco embrionária. c) Blastômero.

112
São obtidas de blastocisto pré-implantacionais, cerca de 5 dias após a fertilização.
a) Célula-tronco embrionária ou celular pluripotente. b) Célula totipotente ou zigoto. c) Célula unipotente ou nulipotente.
Podem ser encontradas no cérebro, medulas ósseas, sangue periférico, músculos esqueléticos, tecido epitelial da pele e do tubo digestório, córnea, polpa dentária, retina, fígado e pâncreas.
a) Célula-tronco adulta. b) Célula-tronco embrionária c) Blastômero.
Existe uma célula-tronco adulta universal, ou seja, capaz de dar origem a qualquer tecido? a) Sim, existe. b) Não existe. c) Os cientistas ainda não
sabem responder esta pergunta.
Célula diferenciada de um tecido assume característica de células de outro tecido a) Diferenciação. b) Desdiferenciação c) Transdiferenciação.
É uma célula pluripotente. a) Célula-tronco embrionária. b) Célula-tronco adulta. c) Zigoto.
É uma celula multipotente. a) Célula-tronco embrionária. b) Célula-tronco adulta. c) Zigoto.
É uma célula totipotente. a) Célula-tronco embrionária. b) Célula-tronco adulta. c) Zigoto.
São obtidas de blastocistos pré-implantacionais, cerca de 5 dias após a fertilização. a) Linhagens de células-tronco embrionárias.
b) Linhagens de células-tronco adultas.
c) Linhagens de células multipotentes.
Existe uma célula-tronco adulta universal, ou seja, capaz de dar origem a qualquer tecido?
a) Sim, existe. b) Não existe c) Os cientistas ainda não sabem responder esta pergunta.
Possui enzima telomerase que possibilita a manutenção de longos telômeros, isto garante a capacidade de replicação por muitas gerações.
a) Célula pluripotente. b) Célula oligopotente. c) CélulanNulipotente.
São raras e, geralmente, difíceis de serem identificadas, isoladas e purificadas. a) Células-tronco embrionárias. b) Células-tronco adultas. c) Células nulipotentes.
Já foi demonstrado que uma única célula-tronco hematopoiética é capaz de reconstituir toda a medula óssea após a mesma ter sido destruída por radiação. A afirmativa é: a) falsa. b) verdadeira, mas só foi
demonstrado em ratos. c) verdadeira e já foi
demonstrado em ratos e humanos.

113
Em laboratório, as células-tronco __1__ podem multiplicar-se por muitas gerações sem que haja diferenciação; já as __2__ , sofrem diferenciação. 1 e 2 podem ser substituídos por a) adultas e embrionárias. b) embrionárias e adultas. c) adultas e pluripotentes.
Já foi demonstrado que células-tronco adultas da medula óssea são capazes de dar origem a neurônios. A este fenômeno dá-se o nome de a) diferenciação. b) desdiferenciação c) plasticidade.
Células-tronco embrionárias, quando injetadas em cobaias cujo sistema imunológico foi suprimido, geram teratomas (mistura de diferentes tipos celulares). A afirmativa é: a) verdadeira. b) falsa, pois o fenômeno foi
observado com células tronco adultas.
c) falsa, pois geram tumores, mas não geram teratomas.
Já foi demonstrado que uma única célula-tronco hematopoiética é capaz de reconstituir toda a medula óssea após a mesma ter sido destruída por radiação. A afirmativa é: d) falsa. e) verdadeira, mas só foi
demonstrado em ratos.
Células-tronco adultas e embrionárias possuem a capacidade de auto-replicação e de dar origem a células especializadas. A afirmativa é: a) totalmente falsa. b) totalmente verdadeira. c) parcialmente verdadeira.
Células-tronco adultas e embrionárias são capazes de proliferar e de se especializar quando transplantadas para animais cujo sistema imunológico foi ... a) estimulado. b) suprimido. c) mantido.
As células-tronco no adulto são "sobras" das células-tronco embrionárias? a) Sim. b) Não. c) Os cientistas ainda não
sabem responder esta pergunta.
A plasticidade apresentada pelas células-tronco in vitro também ocorre in vivo? a) Sim. b) Não. c) Os cientistas ainda não
sabem responder esta pergunta.
Existe uma célula-tronco adulta universal, ou seja, capaz de dar origem a qualquer tecido?
a) Sim, existe. b) Não existe c) Os cientistas ainda não sabem responder esta pergunta.
Os cientistas ainda NÃO sabem: (1) quais são os fatores responsáveis pela migração das células-tronco até os tecidos danificados, (2) quais são os controles intrínsecos que fazem uma célula-tronco se diferenciar em determinado tipo celular ao invés de outro. As afirmativas 1 e 2 são: a) ambas verdadeiras. b) ambas falsas. c) verdadeira e falsa,
respectivamente.
Os cientistas ainda NÃO sabem: (1) quais são os mecanismos que permitem as células-tronco embrionárias proliferarem in vitro sem que haja diferenciação, (2) qual estágio de diferenciação da célula-tronco é o melhor para transplante. As afirmativas 1 e 2 são: a) ambas verdadeiras. b) ambas falsas. c) verdadeira e falsa, respectivamente.
NÃO podemos considerar como potencial uso das células-tronco: a) transplante para restaurar
lesões na medula espinhal. b) meio para testar novas
drogas terapêuticas. c) cura para Síndrome de
Down.

114
As técnicas de transferência nuclear podem reprogramar uma célula de tal forma que fique idêntica às células do receptor, evitando, assim, a rejeição. A frase está relacionada com a) clonagem reprodutiva. b) clonagem terapêutica. c) transgênicos.
As células do sistema nervoso têm origem a partir de qual folheto embrionário? a) Ectoderma. b) Mesoderma. c) Endoderma.
As células da epiderme e seus anexos (pêlos, glândulas, ) tem origem a partir de qual folheto embrionário? a) Ectoderma. b) Mesoderma. c) Endoderma
.
. As células que constituem o sangue têm origem a partir de qual folheto embrionário?
a) Ectoderma b) Mesoderma. c) Endoderma
As células musculares têm origem a partir de qual folheto embrionário? a) Ectoderma. b) Mesoderma. c) Endoderma.
As células que constituem os ossos têm origem a partir de qual folheto embrionário?
a) Ectoderma. b) Mesoderma. c) Endoderma.
As células que constituem o epitélio do tubo digestório têm origem a partir de qual folheto embrionário?
a) Ectoderma. b) Mesoderma. c) Endoderma.
As células que constituem o pâncreas e o estomago têm origem a partir de qual folheto embrionário?
a) Ectoderma. b) Mesoderma. c) Endoderma.
Estágio do desenvolvimento embrionário que sucede a mórula. É uma bola oca de células com uma cavidade interna, a blastocele, delimitada por uma camada celular.
a) Blástula. b) Gástrula. c) Nêurula.
Estágio do desenvolvimento embrionário que sucede a blástula. Apresenta uma cavidade, o arquêntero que se comunica com o exterior através do blastóporo. a) Blástula. b) Gástrula. c) Nêurula.
Estrutura maciça de células. Fase anterior à blástula. a) Mórula. b) Gástrula. c) Nêurula.

115
Cada uma das divisões celulares que ocorrem no ovo, nas primeiras fases do desenvolvimento embrionário. a) Clivagem. b) Clasmocitose. c) Diapedese.
Estágio do desenvolvimento embrionário que sucede a gástrula. a) Mórula. b) Blástula. c) Nêurula.
Nome dado a qualquer um dos três tecidos embrionários que darão origem aos diversos tecidos e órgãos do animal adulto. a) Blástula. b) Folheto germinativo. c) Ectoderma.
Molécula que contém simultaneamente grupos funcionais amina e ácido carboxílico.
a) aminoácido b) carboidrato c) nucleotídeo.
O DNA e o RNA são formados por a) aminoácidos b) carboidratos c) nucleotídeos
No seu interior existe grande quantidade de DNA.
a) ribossomo b) lisossomo c) núcleo.
No seu interior atua a enzima RNA polimerase.
a) ribossomo b) lisossomo c) núcleo
No seu interior ocorre a transcrição.
a) ribossomo b) lisossomo c) núcleo
Organela responsável pela síntese e transporte de proteínas.
a) reticulo endoplasmático liso
b) Retículo endoplasmático rugoso.
c) Complexo golgiense.
Organela responsável pela síntese protéica e que possui uma subunidade grande e outra pequena.
a) Mitocôndria. b) Núcleo c) Ribossomo.
Organela na qual ocorre o encontro do códon com o anticódon, possui uma subunidade grande e outra pequena. a) Mitocôndria. b) Núcleo. c) Ribossomo.
Organela que pode ser encontrada aderida ao retículo endoplasmático rugoso, dentro das mitocôndrias e livre no hialoplasma. a) Lisossomo b) Peroxissomo c) Ribossomo.
Organela responsável pela síntese de ácidos graxos, esteróides e fosfolipídios. a) Retículo endoplasmático
liso. b) Retículo endoplasmático
rugoso. c) Complexo golgiense.
Organela que armazena enzimas digestivas e que irá se juntar ao fagossomo ou pinossomo.
b) Lisossomo c) Fagossomo d) Peroxissomo.
Organela que tem origem a partir do complexo golgiense e que está relacionada à autofagia (digestão de partículas da própria célula). a) Lisossomo. b) Fagossomo. c) Peroxissomo.

116
Originado a partir da fagocitose e que irá se junta ao lisossomo formando o vacúolo digestivo.
a) Lisossomo. b) Fagossomo. c) Peroxissomo.
Organela que armazena catalase, enzima responsável pela degradação da água oxigenada. a) Lisossomo. b) Fagossomo. c) Peroxissomo.
Organela formada por bolsas membranosas achatadas e que recebe vesículas do RER. a) Reticulo endoplasmático. b) Complexo golgiense. c) Centríolo .
Conjunto de fibras e túbulos protéicos que dá sustentação e define a forma da célula. Relacionado também com a movimentação dos cromossomos durante a divisão celular. a) Enzimas. b) Fibras do fuso
Conjunto de fibras protéicas que permitem a contração muscular, o movimento amebóide, o movimento dos cílios e flagelos. a) Enzimas. b) fibras do fuso. c) Citoesqueleto.
Organela que origina lisossomos, vesículas de secreção o acrossomo do espermatozóide.
a) Reticulo endoplasmático b) Complexo golgiense. c) Centríolo.
Local no interior da célula onde ocorrem diversos fenômenos como glicólise, por exemplo a) Mitocôndria. b) Hialoplasma. c) Ribossomo.
Organela no interior da qual ocorre a respiração celular a) Retículo endoplasmático. b) Ribossomo. c) Mitocôndria.
Organela com DNA circular a) Retículo endoplasmático. b) Ribossomo. c) Mitocôndria.
Constituída por lipídeos e proteínas, é semipermeável e isola a célula do ambiente externo. a) Enzimas. b) Membrana plasmática. c) Carioteca.
Organela responsável pela formação de cílios, flagelos e das fibras do fuso. a) Centríolo. b) Ribossomo. c) Lisossomo.
No interior desta organela ocorre o ciclo de Krebs e a cadeia respiratória com produção de ATP, gás carbônico e água. a) Retículo endoplasmático. a) Ribossomo. b) Mitocôndria.
Cada uma das formas que um gene pode apresentar. a) Homólogo. b) Análogo. c) Alelo.
Cada um dos dois filamentos do cromossomo duplicado que se mantêm unidos pelo centrômero. a) Cromátides. b) DNA. c) Cromossomos.
Nome dado ao cromossomo X inativado e condensado das fêmeas de mamíferos. a) Cromatina sexual. b) Cromátide-irmã. c) Cariótipo .

117
Cada um dos longos filamentos de DNA presentes no núcleo de células eucarióticas. a) Corpúsculo de Barr. b) Genoma. c) Cromossomo.
Um par de cromossomos geneticamente equivalentes, presente em uma célula diplóide. a) Cromátides-irmãs. b) Cromossomos homólogos. c) Cromatina sexual.
Região cromossômica pela qual as cromátides-irmãs mantêm-se unidas.
a) Centríolo. a) Centrômero. b) Nucléolo.
Ácido nucléico constituído por desoxirribose, fosfato e bases nitrogenadas. Forma uma molécula filamentosa com cadeia dupla e arranjo helicoidal. a) DNA. b) RNA mensageiro. c) RNA transportador.
É a substância que forma os genes. a) DNA. b) RNA mensageiro. c) RNA transportador.
Constituição genética de um indivíduo que, em interação com o ambiente, determina suas características (fenótipo). a) Cariótipo. b) Fenótipo. c) Genótipo.
Segmento de molécula de DNA que contém uma instrução gênica codificada para a síntese de uma proteína. a) Códon. b) Anticódon. c) Gene.
Revestimento formado por moléculas de glicídios frouxamente entrelaçadas, situados externamente à membrana plasmática de células animais. a) Parede celular de celulose. b) Glicocálix. c) Desmossomos.
Característica ou conjunto de características físicas, fisiológicas ou comportamentais de um ser vivo. a) Cariótipo. b) Fenótipo. c) Genótipo.

118
ATIVIDADE 6 - HEMATOPOIESE: SEGUINDO UMA VIA DE DIFERENCIAÇÃO
Materiais:
• um tabuleiro com a indicação de três regiões: medula óssea, sangue (circulação sanguínea) e outro tecido;
• uma chave de classificação descrevendo tipos de células sanguíneas em diferentes níveis de diferenciação, reconstituindo vias de diferenciação durante a hematopoiese;
• um dado de seis faces; • peças representando células do sangue em diferentes níveis de diferenciação.
Regras da atividade:
1. Iniciar com a leitura da primeira página da chave de classificação. Localizar a peça que representa a célula-tronco hematopoética multipotente inicial e colocá-la no tabuleiro na região correspondente à medula óssea.
2. Lançar o dado quando solicitado e seguir as instruções da chave, indo para a página indicada.
3. De uma página para outra, ocorre um evento de diferenciação, resultando em um determinado tipo celular. Localizar esse tipo celular entre as peças disponíveis e arranjar no tabuleiro no local adequado.
4. Ao final da oficina espera-se obter uma seqüência de tipos celulares representando uma via de diferenciação hematopoiética.
Tempo estimado: 30 minutos Observação: a chave de classificação encontra-se a seguir.

119
Chave de classificação
Pág. 1 Célula-tronco hematopoética (CT-H) Principal característica: é uma célula-tronco multipotente, pois tem o potencial de gerar linhagens de células que darão origem aos diversos tipos de células do sangue, como as hemácias, os granulócitos, os linfócitos, os monócitos e os megacariócitos (de onde se originam as plaquetas). A célula-tronco hematopoética multipotente origina por mitose células-filhas que seguem dois destinos: umas permanecem como células-tronco, mantendo a população dessas células na medula óssea vermelha, e outras se tornam células progenitoras, que também são multipotentes mas com potencialidade reduzida em relação à célula-tronco inicial. Você é uma CT-H que responde a um fator A, presente no meio extracelular. O fator A estimula a divisão mitótica em células-tronco. Para saber o seu destino após a divisão, lance o dado e:
• se sair um número par, vá para a página 2; • se sair um número ímpar, vá para a página 3.
Pág. 2 2. Você continua na medula óssea como célula-tronco, mantendo o mesmo nível de potencialidade do estágio anterior. Lance o dado e:
• se sair um número par, continue nesta página e lance o dado novamente; • se sair um número ímpar, vá para a página 3.
Pág. 3 3. Você se transformou em uma célula-tronco progenitora multipotente. Dois fatores estão presentes no meio extracelular: o fator B, que determina a redução da atividade mitótica, e o IL-3, que é um fator de crescimento celular. Para saber a qual dos fatores você é capaz de responder, lance o dado e:
• se sair 1 ou 2, vá para a página 2; • se sair 3 ou 4, vá para a página 4; • se sair 5 ou 6, vá para a página 5.
Pág. 4 4. Você respondeu ao fator B, reduzindo sua atividade mitótica, e não respondeu ao fator IL-3. Tornou-se uma célula-tronco linfóide multipotente. Dois fatores – E e F – estão presentes no meio extracelular e atuam promovendo vias de determinação distintas. Uma célula determinada torna-se mais comprometida com um destino de diferenciação, reduzindo sua potencialidade. Para saber a qual dos fatores de determinação você é capaz de responder, lance o dado e:
• se sair 1 ou 2, você não responde aos estímulos → vá para a página 2; • se sair 3 ou 4, vá para a página 6; • se sair 5 ou 6, vá para a página 7.
Pág. 5 5. Você respondeu ao fator B, reduzindo sua atividade mitótica, e também respondeu ao fator IL-3, tornando-se uma célula-tronco mielóide multipotente. Dois fatores – G e H – estão presentes no meio extracelular e atuam promovendo vias de determinação distintas. Uma célula determinada torna-se mais comprometida com um destino de diferenciação, reduzindo sua potencialidade. Para saber a qual dos fatores de determinação você é capaz de responder, lance o dado e:
• se sair 1 ou 2, você não responde aos estímulos → vá para a página 2; • se sair 3 ou 4, vá para a página 12; • se sair 5 ou 6, vá para a página 13.
Pág. 6 6. Você respondeu ao fator E, que é um estímulo de determinação e também estimula sua ida ao timo. Você se tornou, então, uma célula progenitora de linfócito T e migrou para o timo. Para saber o que vai acontecer com você chegando ao timo, vá para a página 10. Pág. 7 7. Você respondeu ao fator F, que é um estímulo de determinação. Você se tornou, então, uma célula progenitora de linfócito B, que permanece na medula óssea. Para saber o que vai acontecer com você, vá para a página 8. Pág. 8 8. Existem fatores de diferenciação presentes na medula óssea e você respondeu a eles, tornando-se a célula precursora de linfócitos B ou linfoblasto-B. Em seguida, você recebeu estímulos para se dividir e estímulos para diferenciação de todas as células-filhas. Para saber o que vai acontecer com você e com suas células-filhas, vá para a página 9. Pág. 9 9. Você respondeu a fatores de diferenciação, sofreu um processo de maturação e se tornou um linfócito B. Você não responde mais aos estímulos de divisão e não se multiplica mais. Você é liberado para a corrente sangüínea.

120
Para saber qual a sua morfologia e função quando vai para o sangue, vá para a página 25. Pág. 10 10. No timo, você entrou em contato com fatores de diferenciação. Você respondeu à presença deles, tornando-se a célula precursora de linfócitos T ou linfoblasto-T. O timo também libera um hormônio chamado timosina. Para saber se o que vai acontecer com você após a liberação de timosina, vá para a página 11. Pág. 11 11. Você apresentava receptores em sua membrana plasmática que reconhecem o hormônio timosina e, como resposta à ligação entre os receptores e as moléculas de timosina, você sofreu diferenciação tornando-se um linfócito T. Para conhecer a sua morfologia e função, vá para a página 24. Pág. 12 12. Você respondeu ao fator G, que é um estímulo de determinação. Você se tornou, então, uma célula progenitora granulocítica/monocítica da medula óssea. Dois fatores – I e J – estão presentes no meio extracelular e atuam promovendo vias de determinação distintas. Uma célula determinada torna-se mais comprometida com um destino de diferenciação, reduzindo sua potencialidade. Para saber o que vai acontecer com você, lance o dado e:
• se sair 1 ou 2, você não responde aos estímulos → vá para a página 2; • se sair 3 ou 4, vá para a página 14; • se sair 5 ou 6, vá para a página 15.
Pág. 13 13. Você respondeu ao fator H, que é um estímulo de determinação. Você se tornou, então, uma célula progenitora megacariocítica/eritrocítica da medula óssea. Dois hormônios presentes no sangue atuam promovendo vias de determinação distintas na medula óssea: são os hormônios trombopoetina e eritropoetina. Uma célula determinada torna-se mais comprometida com um destino de diferenciação, reduzindo sua potencialidade. Para saber o que vai acontecer com você, lance o dado e:
• se sair 1 ou 2, você não responde aos hormônios → vá para a página 2; • se sair 3 ou 4, vá para a página 20; • se sair 5 ou 6, vá para a página 21.
Pág. 14 14. Você respondeu ao fator I e se tornou uma célula determinada: o mieloblasto granulocítico. Em seguida, você recebeu estímulos para o início da diferenciação. Para saber qual a via de diferenciação que vai seguir, lance o dado e:
• se sair 1, 2 ou 3, vá para a página 17; • se sair 4 ou 5, vá para a página 18; • se sair 6, vá para a página 19.
Pág. 15 15. Você respondeu ao fator J e se tornou uma célula determinada: o promonócito. Em seguida, você recebeu estímulos para se dividir e estímulos para diferenciação de todas as células-filhas. Para saber o que vai acontecer nessa diferenciação, vá para a página 16. Pág. 16 16. Após duas divisões mitóticas, você aumentou de volume e se tornou uma célula diferenciada rica em lisossomos. Você agora recebe o nome de monócito. Para saber como é sua morfologia e função, vá para a página 29. Pág. 17 17. Você se tornou um mielócito neutrófilo, uma célula que acumula em seu citoplasma grande quantidade de grânulos específicos e lisossomos. Você não se divide mais. O processo de diferenciação celular continua. Você é finalmente liberado para o sangue. Para saber o resultado desse processo, vá para a página 31. Pág. 18 18. Você se tornou um mielócito eosinófilo, uma célula que acumula em seu citoplasma grande quantidade de grânulos acidófilos (que se coram quando a célula é submetida a tratamento com corantes ácidos). Você não se divide mais. O processo de diferenciação celular continua. Você é finalmente liberado para o sangue. Para saber o resultado desse processo, vá para a página 32. Pág. 19 19. Você se tornou um mielócito basófilo, uma célula que acumula em seu citoplasma grande quantidade de grânulos de histamina. Você não se divide mais. O processo de diferenciação celular continua. Você é finalmente liberado para o sangue. Para saber o resultado desse processo, vá para a página 33. Pág. 20

121
20. A trombopoetina é sintetizada pelo fígado, liberada na corrente sangüínea e ao passar pela medula óssea difunde para o tecido hematopoético. Você respondeu à presença do hormônio trombopoetina, pois apresentava os receptores para esse hormônio na membrana plasmática. Como resposta a essa ligação, você se tornou uma célula precursora de megacariócito (ou megacarioblasto). Você vai entrar em uma via de diferenciação. Para saber o que vai acontecer, vá para a página 22. Pág. 21 21. A eritropoetina é sintetizada pelos rins, liberada na corrente sangüínea e ao passar pela medula óssea difunde para o tecido hematopoético. Você respondeu à presença do hormônio eritropoetina, pois apresentava os receptores para esse hormônio na membrana plasmática. Como resposta a essa ligação, você se tornou uma célula precursora de eritrócito ou pró-eritroblasto. Você não se divide mais, a cromatina apresenta-se condensada e no citoplasma tem início a síntese da proteína hemoglobina. Você vai entrar em uma via de diferenciação. Para saber o que vai acontecer, vá para a página 23. Pág. 22 22. Você se tornou um megacariócito: uma célula grande, que apresenta granulações no citoplasma. Você está sintetizando proteínas que vão futuramente atuar como fatores de coagulação sangüínea. As granulações do citoplasma começam a se desprender, gerando fragmentos de célula que são liberados no sangue. Para saber o que acontece a esses fragmentos, vá para a página 35. Pág. 23 23. Você se tornou um eritroblasto. Seu tamanho está reduzido em relação ao pró-eritroblasto e você está produzindo hemoglobina intensamente no citoplasma. Em cerca de 24 horas, você sofre transformações intensas e passa a ser chamado de reticulócito. Nesse estágio, que dura cerca de 3 dias, o núcleo é deslocado para a periferia da célula até ser eliminado. Para saber o resultado dessas transformações, vá para a página 34. Pág. 24 Linfócitos T Do timo, os linfócitos T são liberados para a circulação sangüínea e podem migrar para diversos tecidos do corpo quando ocorre uma infecção. Após o combate à infecção, eles podem voltar ao sangue, circulando continuamente. Os linfócitos T não sintetizam anticorpos, mas apresentam receptores na membrana plasmática que detectam antígenos específicos. Assim, um determinado linfócito T possui apenas um tipo de receptor, que reconhece um único tipo de antígeno. Os linfócitos T podem ser classificados em subtipos de acordo com sua ação após o reconhecimento do antígeno específico: existem os citotóxicos, capazes de destruir as células que apresentam o antígeno; os auxiliares, que ativam linfócitos B; e os supressores, que inibem a liberação de anticorpos pelos linfócitos B quando necessário. Pág. 25 Linfócitos B Você passou para a circulação sangüínea no estado maduro, ou seja, com anticorpos já expostos na membrana plasmática. Do sangue, você também pode ir para o sistema circulatório linfático, passando pela linfa, por linfonodos e pelo baço. Um linfócito B produz apenas um tipo específico de anticorpo. No sangue, moléculas de um antígeno são reconhecidas pelos anticorpos que estão na sua membrana plasmática, resultando em uma resposta. Para descobrir que resposta é essa, vá para a página 26. Pág. 26 Você reconheceu a presença de seu antígeno específico no organismo. Com a ligação antígeno – anticorpo, você aumenta de volume e divide-se por mitose, por diversas vezes. Ao final do processo, formam-se numerosas células idênticas. Para saber o que acontece em seguida, lance o dado e:
• se sair um número par, vá para a página 27; • se sair um número ímpar, vá para a página 28.
Pág. 27 PLASMÓCITO Ao final das sucessivas divisões, você faz parte de um grupo de linfócitos B que se transformam em plasmócitos. O plasmócito é uma célula que produz anticorpos intensamente, liberando-os na corrente sangüínea. Esses anticorpos reconhecem os antígenos específicos e “chamam a atenção” de macrófagos, neutrófilos e eosinófilos, que vão para o local da infecção e eliminam as bactérias que possuem o tal antígeno. Pág. 28 Ao final das sucessivas divisões, você faz parte de um grupo de linfócitos B que se tornam células B de memória. Você não participará do combate à infecção bacteriana que está ocorrendo no organismo neste momento. Você terá vida longa no organismo, circulando pelo sangue e pela linfa, e ficará de prontidão para sintetizar rapidamente seus anticorpos específicos se o organismo entrar novamente em contato com as bactérias que possuem o antígeno. Pág. 29 MONÓCITO

122
Os monócitos têm o núcleo oval ou em forma de ferradura. O citoplasma é rico em lisossomos, que aparecem em preparações como numerosos grânulos. As mitocôndrias também são numerosas e o complexo golgiense, muito desenvolvido, já que ali são formados os lisossomos. Você é um monócito que acabou de ser liberado da medula óssea para o sangue. Para saber o seu destino, lance o dado e:
• se sair um número par, vá para página 30; • se sair um número ímpar, significa que você circulará pelo sangue por 2 horas. Lance novamente o dado. Você pode circular pelo sangue por até 8 horas.
Obs.: se você permanecer circulando no sangue por mais de 8 horas, vá direto para a pág. 30. Pág. 30 MACRÓFAGO Os monócitos são células precursoras de macrófagos. Quando os monócitos saem dos vasos sanguíneos e se dirigem aos tecidos, completam sua maturação, tornando-se macrófagos. Nessa maturação, a célula aumenta de volume e torna-se ainda mais rica em lisossomos. Os macrófagos apresentam núcleo grande e em forma de ferradura. Eles participam do mecanismo de defesa do organismo por fagocitose. Podem ficar entre as células de um tecido durante meses, e são ativados na presença de células ou partículas estranhas ao organismo. Nessas situações, emitem pseudópodes e fagocitam as partículas estranhas, ao mesmo tempo em que liberam substâncias que desencadeiam uma resposta inflamatória. Com a resposta inflamatória, mais macrófagos são atraídos para o local infectado. Pág. 31 NEUTRÓFILO Os neutrófilos são células maduras (diferenciadas) também conhecidas como leucócitos polimorfonucleares, porque a forma do núcleo pode variar, apresentando de dois a cinco lóbulos. No citoplasma, os neutrófilos apresentam grande quantidade de lisossomos e de grânulos específicos. Os neutrófilos fazem parte do sistema de defesa do organismo contra invasão de microrganismos. Enquanto estão circulando no sangue, são esféricos, mas em contato com um substrato sólido emitem pseudópodes e podem fagocitar os invasores. Após a fagocitose, ocorre a digestão dos invasores pelas enzimas lisossômicas. No entanto, em uma infecção nem todas as bactérias são mortas e algumas podem atacar os neutrófilos, “escapando” da ação fagocitária dos neutrófilos. Nessas situações, forma-se um líquido amarelado que contém bactérias, neutrófilos mortos e líquido extracelular: o pus. Pág. 32 EOSINÓFILOS Os eosinófilos são células maduras (diferenciadas) que constituem cerca de 5% do total de leucócitos. Possuem núcleo com dois lóbulos e citoplasma cheio de grânulos ovais acidófilos, além de grande quantidade de lisossomos. Os eosinófilos fagocitam e digerem complexos de antígenos-anticorpos, principalmente aqueles formados em processos de alergia. O número de eosinófilos no sangue aumenta durante reações alérgicas e em infecções por vermes parasitas, como o esquistossomo. Pág. 33 BASÓFILOS Os basófilos são células maduras (diferenciadas) que se caracterizam por um núcleo volumoso e de forma irregular. O citoplasma é rico em grânulos maiores do que os dos outros granulócitos. Esses grânulos contêm heparina e histamina, uma substância que atrai eosinófilos e neutrófilos. Quando ocorre uma infecção, a presença de determinados anticorpos na circulação sangüínea faz com que os basófilos liberem o conteúdo dos grânulos. A conseqüência é a migração de eosinófilos e neutrófilos para o local, o que intensifica o combate aos causadores da infecção. Os basófilos constituem menos de 1% dos leucócitos do sangue. Pág. 34 HEMÁCIAS ou ERITRÓCITOS ou GLÓBULOS VERMELHOS Assim que os reticulócitos são liberados na corrente sangüínea, passam a ser chamados de eritrócitos, ou glóbulos vermelhos. Os eritrócitos são células anucleadas com formato de disco bicôncavo, com cerca de 7µm de diâmetro. O citoplasma acumula moléculas de hemoglobina e possui pequena quantidade de ribossomos e mitocôndrias. Sua função é o transporte de oxigênio e, em menor proporção, de gás carbônico. Pág. 35 PLAQUETAS As plaquetas correspondem a fragmentos de citoplasma do megacariócito. Elas são fundamentais para o processo de coagulação sangüínea. A coagulação sanguínea bloqueia lesões em vasos, impedindo a ocorrência de hemorragias. Sob estímulo das células lesadas, as plaquetas se agregam auxiliando na formação do coágulo. Além disso, as plaquetas liberam fatores de coagulação que participam de uma cascata de reações junto com outros fatores presentes no plasma sanguíneo. Ao final desse processo, forma-se a proteína fibrina. As moléculas de fibrina se agrupam formando uma rede que aprisiona hemácias, glóbulos brancos e plaquetas, formando assim o coágulo. O coágulo pára o fluxo de sangue na região lesada do vaso.

123
ATIVIDADE 8 - REGRAS PARA O DEBATE A polêmica sobre a utilização de células-tronco embrionárias em terapia humana
• 8 grupos de 7 a 8 membros cada • Haverá um mediador • Cada grupo fará o papel de um dos seguintes membros da sociedade:
1. cientista 2. jurista 3. doente passível de ser tratado com células-tronco embrionárias 4. jornalista 5. religioso 6. político 7. dono de uma clinica de reprodução assistida 8. mulher que vai se submeter a reprodução assistida
• Cada grupo prepara 7 perguntas: uma para um dos demais grupos participantes. Por exemplo, o grupo 1 (o do cientista) preparará perguntas para os grupos 2, 3, 4, 5, 6, 7 e 8.
• Cada grupo deve eleger um relator, que se sentará à mesa para participar do debate. Os demais membros poderão dar assessoria, se necessário.
• Os grupos terão tempo livre para preparar suas participações • O debate ocorrerá como se segue:
o Inicialmente, cada relator terá no máximo 3 minutos para apresentar o seu posicionamento.
o Na primeira rodada de perguntas, a ordem de formulação das perguntas será sorteada, assim como também será sorteado quem deverá responder. Por exemplo, o grupo 1 (o do cientista) sorteou o número 6 e, assim sendo, fará uma pergunta para o relator do grupo 6 (que representa o político).
o Na segunda rodada de perguntas, a ordem de fazer perguntas será sorteada, mas o participante poderá escolher quem irá responder. Por exemplo, sorteado o número 2 o jurista fará uma pergunta para quem lhe aprouver.
o Pequeno intervalo o Durante o intervalo, um grupo de jurados fará uma seleção de perguntas
interessantes, feitas por todos os grupos e que ainda não foram usadas. O mediador fará as perguntas para os participantes do debate.
Terminado o debate, a classe será dividida em grupos e participará de uma atividade de discussão das seguintes situações:
1. Tenho um filho com morte cerebral constatada. Devo doar seus órgãos para transplante? 2. Tenho um filho de 9 anos gerado por FIV. Tenho embriões excedentes congelados. No entanto,
perdi o útero recentemente. Meu filho desenvolve quadro de doença grave e incurável, sem doadores compatíveis para transplante. (2a
3. Tenho um filho de nove anos com doença grave e incurável, sem doadores compatíveis para transplante. (a) É lícito fazer FIV, selecionar um embrião compatível com o filho atual, implantar e gerar uma criança para ser doadora do irmão mais velho?
) É lícito usar esses embriões para obter células-tronco e realizar uma terapia baseada em células?
4. Fiz fertilização “in vitro” e atualmente estou grávida de sêxtuplos. Os médicos afirmam que nessa situação corro risco de vida, assim como os embriões. É lícito fazer a redução, ou seja, eliminar alguns dos embriões?Tenho um filho de nove anos com doença grave e incurável, sem doadores compatíveis para transplante. (a) •É lícito realizar clonagem terapêutica, obter um blastocisto com células geneticamente idênticas às do meu filho para realizar uma terapia baseada em células –tronco? (b) É permitido pela lei brasileira?

124
ATIVIDADE 9 - PALAVRAS CRUZADAS CÉLULAS-TRONCO Silvio Higa

125
HORIZONTAIS 1h – O sistema nervoso tem origem a partir deste folheto embrionário. 2h - Sinônimo de plasticidade. 3h – Sinônimo de divisão celular. 4h – “Vida” em inglês. 5h - ___ trinca de bases nitrogenadas codifica um aminoácido. 6h – Composto por células-tronco. 7h – Camada de tecido do globo ocular responsável pela sustentação. 8h – No estômago é ácido e no duodeno é básico. 9h – Multiplica uma amostra de DNA milhares de vezes em poucos minutos. 10h - Neste local é produzida a maior parte do oxigênio que respiramos. 11h – Quantas bases nitrogenadas compõem um códon? 12h – Sinônimo de ato sexual. 13h – Posição ocupada por um gene no cromossomo. 14h – Sintetizado nas cristas mitocondriais. 15h – Camada mais interna da pele, firmemente unida à epiderme. 16h – Elemento químico relacionado a um tipo de transporte ativo. 17h – Iniciais da organela que sintetiza e transporta proteínas. 18h – As quatro bases nitrogenadas do DNA. 19h – Faz parte da pele e é constituída por densa rede de tecido conjuntivo. 20h – Cada uma das formas que um gene pode apresentar. 21h - As quatro bases nitrogenadas do RNA. 22h – Gera duas células-filhas com o mesmo número de cromossomos da célula-mãe. 23h – Célula capaz de formar poucos tipos celulares. 24h – Produto da transcrição. 25h – Resultante das primeiras clivagens dos blastômeros. 26h - É a capacidade de uma céula-tronco adulta de um tecido gerar células especializadas de um outro tecido. 27h – Fase do desenvolvimento embrionário no qual já é possível observar notocorda e tubo neural. 28h – Nos animais forma gametas e nos vegetais forma esporos. 29h - Elemento químico relacionado a um tipo de transporte ativo. 30h – Organela na qual ocorre a síntese protéica. 31h – Tecido atualmente muito utilizado como fonte de células-tronco adultas. VERTICAIS 1v - Célula capaz de formar todos os tipos celulares. Exemplos: zigoto e blastômeros. 2v – Organela que tem origem a partir do complexo golgiense. 3v – Possui sistema nervoso difuso. 4v – Vertebrado homeotérmico e com diafragma. 5v – Fase anterior à blástula. 6v - Célula incapaz de formar outros tipos celulares. Exemplo: hemácia. 7v – Um dos filamentos que compõe o micélio. 8v - É o processo através do qual uma célula não especializada torna-se especializada. 9v - Têm grande importância econômica como alimento e para a produção de fertilizantes, fibras, ágar, potassa e iodo. 10v – Célula totipotente. 11v – São células pluripotentes. 12v – Flor de monocotiledôneas. 13v – Marsupial. 14v – A célula-tronco embrionária é uma célula ... 15v – Macromolécula compota por nucleotídeos. 16v – A joaninha é um tipo de ... 17v – Elemento químico presente tanto no nucleotídeo como no aminoácido. 18v - A célula-tronco adulta é uma célula ... 19v – Produzido na medula óssea. 20v – Vertebrado com hemácias anucleadas. 21v – Principal excreta nitrogenado dos mamíferos. 22v – Fatores internos e externos que controlam modificações na estrutura e função da célula. 23v – Deixa o núcleo através dos poros da carioteca. 24v - Técnica de transferência nuclear que reprograma células para fiquem idênticas às do receptor, evitando, assim, a rejeição. 25v – “Célula-tronco” em inglês. Solução – página 128

126
ANEXOS Respostas do Estudo dirigido 2 – Clonagem reprodutiva versus clonagem terapêutica
1. o esquema A o zigoto é proveniente de uma fecundação, ou seja, ele é formado por dois núcleos haplóides (o núcleo do ovócito e do espermatozóide) que se fundiram e formaram um núcleo diplóide. Já os zigotos dos esquemas B e C são provenientes de transferência nuclear, ou seja, eles recebem apenas um núcleo, que, por ser de uma célula diferenciada, já é diplóide.
2. O núcleo do ovócito é removido nos procedimentos de clonagem (B e C) porque, neste mesmo ovócito, será implantado um núcleo diplóide. Se o núcleo do ovócito não fosse removido, formaríamos um embrião triplóide, o que seria inviável.
3. A diferença da clonagem reprodutiva (B) e da terapêutica (C) está em o que é feito com o blastocisto. Na clonagem reprodutiva (B), ele é implantado num útero maduro para dar origem a um indivíduo idêntico ao doador da célula diferenciada. Na clonagem terapêutica (C), as células-tronco embrionárias do blastocisto são cultivadas em laboratório para dar origem a algum tipo celular ou tecido ou órgão específico.
4. Como a célula diferenciada doa o núcleo, ou seja, doa o DNA, o fenótipo do camundongo clonado deve ser o mesmo do camundongo doador do núcleo. Com isso, o camundongo clonado seria um macho preto.
5. Se as células sangüíneas produzidas no esquema C forem implantadas no mesmo indivíduo que doou a célula diferenciada, esse indivíduo não desenvolverá uma rejeição a essas células sangüíneas. Porque rejeição ocorre quando o organismo de uma pessoa identifica um órgão ou um tecido como um invasor e passa a atacá-lo; como as células sangüíneas produzidas por clonagem terapêutica têm o mesmo genoma das células da pessoa que vai recebê-las, as novas células sangüíneas seriam reconhecidas como se fossem da própria pessoa e não como invasoras.
Para pensar: resposta no texto “Reprogramação Celular”.
CARTAS E GABARITO DA ATIVIDADE 1 - DIFERENCIAÇÃO DA HEMÁCIA CARTAS DE SINAIS: I. Eritroblasto basófilo
1. Diminuir o volume celular e perder duas mitocôndrias. 2. Ativar a produção de receptores de membrana para transferrina, que é uma proteína
transportadora de ferro. Após se combinarem, o complexo receptor-transferrina penetra no citoplasma por endocitose.
3. Perder o núcleo 4. Manter a célula inalterada 5. Expressar os genes responsáveis pela síntese de hemoglobina. 6. Desativar todos os processos de transcrição da célula 7. Perder todos os polirribossomos
II. Pré-reticulócito
1. Diminuir o volume celular e perder três mitocôndrias. 2. Condensar a cromatina e perder alguns polirribossomos. 3. Perder o núcleo 4. Formar uma projeção celular onde se situa o núcleo. 5. Desativar todos os processos de transcrição da célula 6. Expressar o gene que produz colágeno tipo I 7. Perder todos os polirribossomos

127
III. Reticulócito
1. Manter a célula inalterada 2. Sintetizar hemoglobina. 3. Expressar o gene que produz colágeno tipo I 4. O núcleo deve ser expelido com uma porção delgada de citoplasma ao seu redor. 5. Diminuir o volume celular e perder duas mitocôndrias 6. Desenvolver três projeções citoplasmáticas. 7. Perder uma mitocôndria
IV. Hemácia
1. Fragmentar a célula, produzindo as plaquetas 2. Sintetizar hemoglobina para completar a maturação do eritrócito. 3. Manter a célula inalterada 4. Manter a configuração do citoesqueleto como na etapa anterior 5. Configurar uma célula bicôncava. 6. Conferir uma nova organização do citoesqueleto, tornando-o contrátil; com isso, a célula
pode atravessar capilares sinusóides, penetrando, assim, na corrente sangüínea. 7. Produzir um novo núcleo Gabarito (I) 1-2-5, (II) 1-2-4, (III) 2-4-6 e (IV) 2-5-6.
CARTAS E GABARITO DA ATIVIDADES 2 - DIFERENCIAÇÃO NO PROTOZOÁRIO NAEGLERIA GRUBERI
Etapa A 1. Diminuir o volume celular e nuclear 2. No núcleo, iniciar síntese de RNA-mensageiro para tubulina. O RNam vai para o citoplasma. 3. Desativar todos os processos de transcrição da célula 4. Manter a forma amebóide. 5. Inibir os genes responsáveis pela síntese de tubulinas 6. Expressar o único gene responsável pela síntese do flagelo 7. No citoplasma, organizam-se polirribossomos e inicia-se síntese de tubulina. 8. Condensar a cromatina Gabarito: 2 – 4 – 7
Etapa B 1. Acumular tubulina em região específica do citoplasma. 2. Condensar a cromatina e perder alguns ribossomos 3. Perder o núcleo 4. Estimular a divisão celular 5. Expressar os genes responsáveis pela síntese de tubulinas. 6. Manter forma amebóide, mas reduzir emissão de pseudópodes. 7. Cessar síntese de tubulinas no citoplasma 8. Inibir atividade dos genes responsáveis pela síntese de tubulinas Gabarito: 1 – 5 - 6
Etapa C 1. Manter a célula inalterada 2. Formar dois corpúsculos basais. 3. Estimular a divisão celular 4. Tomar forma arredondada. 5. Perder todos os ribossomos

128
6. Fagocitar bactérias 7. Iniciar formação do flagelo, a partir da organização de tubulinas e outras proteínas em microtúbulos. 8. Reduzir o número de mitocôndrias Gabarito: 2 – 4 – 7
Etapa D 1. Tomar forma alongada. 2. Estimular a divisão celular 3. Manter a célula inalterada 4. Emitir pseudópodes 5. Organizar tubulinas e outras proteínas para alongamento dos flagelos. 6. Degradar mitocôndrias 7. Perder o núcleo 8. Posição fixa do núcleo na base da célula. Gabarito: 1 – 5 – 8

129
Solução das palavras cruzadas

130
BIBLIOGRAFIA sobre câncer e células-tronco
Bapat, S. A. 2006. Evolution of cancer stem cells. Seminars in Cancer Biology Bishop. J. M. 1997. Cancer: what should be done? Science 278 (5340): 995. Bissel, M. J. & M. A. LaBarge. 2005. Are tumor stem cells also regulated by the microenvironment?
Cancer Cell 7 (1): 17-23. Clarke, M. F. & M. W. Becker. 2006. O lado maligno das células-tronco. Scientific American Brasil
51: 38-46. Clarke, M. F. & M. Fuller. 2006. Stem cells and cancer: two faces of Eve. Cell 124: 1111-1115. Díaz, B. & E. Moreno. 2005. Competitive nature of cells. Exper. Cell Res. 306: 317-322. Fearon, E. R. 1997. Human cancer syndromes: clues to the origin and nature of cancer. Science,
278: 1043-1050. Gibbs, W. W. 2003. Desvendando as raízes do câncer. Scientific American Brasil 15: 38-47. Gibbs, W. W. 2007. As raízes do mal. Ciência e Saúde 3:26-34. Gisselsson, D. Cancer cells: Differentiation block or developmental back-tracking? 2006. Seminars
in cancer biology. Houghton, J. M., Morozov, A., Smirnova, I. & Wangh, T. C. 2006. Stem cells and cancer. Seminars
in cancer Biology. Huff, C. A., Matsui, W. H., Smith, B. Douglas & Jones, R. J. 2006. Strategies to eliminate cancer
stem cells: clinical implications. Eur. J. of Cancer 42: 12193-1297. Keith, W. N. 2006. Cancer stem cells: opportunities for novel diagnostics and drug discovery. Eur.
J. of Cancer 42: 1195-1196. Krtolica, A. 2005. Stem cell: balancing aging and cancer. Int. J. Biochem. Cell Biol. 37: 935-941. Look, A. T. 1997. Oncogenic transcription factors in the human acute leukemias. Science 278
(5340): 1059-1064. Nguyen, D. X. & Joan Massagué. 2007. Genetic determinants of cancer metastasis. Nature
Review/Genetics 8: 341-352. Ponder, B. 1997. Genetic testing for cancer risk. Science 278 (5340): 1050-1054. Rossi, D. J. & I. L. Weissman. 2006. Cell 125: 229-231. Stewart, R., Stojkovic & M. Lako. 2006. Mechanisms of sef-renewal in human embryonic stem
cells. Eur. J. Cancer 42: 1257-1272. Vaidya, J. S. 2006. An alternative model of cancer cell growth and metastasis. Int. J. of Surgery: 1-
3. Wang, J. C. Y. & J. E. Dick. 2005. Cancer stem cells: lessons from leukemia. Trends in Cell
Biology 15 (9): 494-501. Weinberg, R. A. 1996. How cancer arises. Scientific American 275 (3): 32-41.


















