
Disserta o- P gina de rosto 1.doc) - iac.sp.gov.br · - Ao pesquisador Dr. Helvécio Della Coletta...
82
INSTITUTO AGRONÔMICO CURSO DE PÓS- GRADUAÇÃO EM AGRICULTURA TROPICAL E SUBTROPICAL DESENVOLVIMENTO E VALIDAÇÃO DE DIAGNÓSTICO MOLECULAR DE FUNGOS PATOGÊNICOS A CITROS GRAZIÉLE RAMOS SASSERON Orientador: Marcos Antônio Machado Dissertação submetida como requisito parcial para obtenção do grau de Mestre em Agricultura Tropical e Subtropical Área de Concentração em Genética, Melhoramento Vegetal e Biotecnologia. Campinas, SP 2008
Transcript of Disserta o- P gina de rosto 1.doc) - iac.sp.gov.br · - Ao pesquisador Dr. Helvécio Della Coletta...

INSTITUTO AGRONÔMICO
CURSO DE PÓS- GRADUAÇÃO EM AGRICULTURA
TROPICAL E SUBTROPICAL
DESENVOLVIMENTO E VALIDAÇÃO DE
DIAGNÓSTICO MOLECULAR DE FUNGOS
PATOGÊNICOS A CITROS
GRAZIÉLE RAMOS SASSERON
Orientador: Marcos Antônio Machado
Dissertação submetida como requisito parcial para obtenção do grau de Mestre em Agricultura Tropical e Subtropical Área de Concentração em Genética, Melhoramento Vegetal e Biotecnologia.
Campinas, SP 2008


Ficha elaborada pela bibliotecária do Núcleo de Informação e Documentação do Instituto Agronômico S252d Sasseron, Graziéle Ramos Desenvolvimento e validação de diagnóstico molecular de fungos patogênicos a citros / Graziéle Ramos Sasseron. Campinas, 2008 71 fls Orientador: Marcos Antônio Machado Dissertação (Mestrado) Genética, Melhoramento Vegetal e Biotecnologia - Instituto Agronômico
1. Citros – mancha preta 2. Espaçador interno de transcritos (ITS) 3. Detecção 4. Gomose do citros 5. Mancha marrom das tangerinas 6. Verrugose I. Machado, Marcos Antônio II. Título
CDD. 634.3

Aos meus pais, Benedito e Fátima,
por me concederem a oportunidade de estudar
e me apoiarem em todos os momentos,
DEDICO
Aos meus irmãos, Daniele e Otávio
pelo apoio e carinho,
OFEREÇO

AGRADECIMENTOS
- Ao meu orientador, Dr. Marcos Antônio Machado, pela oportunidade e confiança que
me concedeu, permitindo a realização dos meus trabalhos da pós - graduação
juntamente com o trabalho na Clínica Fitopatológica, e pelos ensinamentos ao longo
destes dois anos.
- À pesquisadora Dra. Maria Luísa P.N. Targon, pela amizade e principalmente pelo
auxílio na realização deste trabalho, sempre me apoiando e incentivando.
- Ao Carlos Ivan Aguilar - Vildoso, pela valiosa sugestão e colaboração na revisão deste
trabalho.
- Ao pesquisador Dr. Helvécio Della Coletta pelo auxílio nos momentos necessários.
- Ao Renato Reis, pela valiosa sugestão na obtenção dos isolados obtidos neste trabalho.
- Aos professores da Pós- Graduação, pelo aprendizado e conselhos fornecidos.
- Aos funcionários da PG- IAC, pelo auxílio e amizade no decorrer do curso.
- Aos meus amigos da Clínica Fitopatológica, em especial à Fabiana e à Silvia, pela
compreensão e ajuda.
- Ao meu amigo Vandeclei, pelo carinho e amizade, por sempre me ajudar na coleta de
material para a realização dos experimentos.
- A todos os funcionários do Centro APTA Citros Sylvio Moreira, que contribuíram
para a realização deste trabalho.
- Ao César C. Graf, José Eduardo M. Teófilo, Henrique Fiorese e Júnior Zanetti, que
gentilmente concederam material para a elaboração deste trabalho.
- A todos os meus colegas da Pós- Graduação, em especial ao João Guilherme e à
Eliana, que sempre me acolheram.

SUMÁRIO
RESUMO ...................................................................................................................................vABSTRACT............................................................................................................. ................vii1 INTRODUÇÃO ........................................................................................................ ............ 12 REVISÃO DE LITERATURA................................................................................. ............ 42.1 Gomose dos citros e Phytophthora spp. ................................................................ ............ 42.2 Mancha preta dos citros e Guignardia citricarpa.................................................. ............ 72.3 Mancha marrom das tangerinas e Alternaria alternata f. sp. citri......................... ............102.4 Verrugose e Elsinoe australis e Elsinoe fawcetti................................................... ............132.5 Detecção de regiões ITS em fungos....................................................................... ............152.6 qPCR (PCR quantitativo)....................................................................................... ............163 MATERIAL E MÉTODOS ...................................................................................... ............173.1 Isolamento, manutenção e conservação dos fungos Alternaria alternata f. sp. citri, Elsinoe
australis, Elsinoe fawcetti, Guignardia citricarpa e Phytophthora nicotianae........... ............173.2 Ensaio de patogenicidade dos fungos Alternaria alternata f. sp. citri, Elsinoe australis,
Elsinoe fawcetti, Guignardia citricarpa e Phytophthora nicotianae........................... ............193.3 Extração de DNA total dos fungos e das lesões .................................................... ............193.4 Oligonucleotídeos e amplificação por PCR dos fungos Alternaria alternata f. sp. citri,
Elsinoe australis, Elsinoe fawcetti, Guignardia citricarpa e Phytophthora nicotianae...........203.5 Clonagem, sequenciamento e análise das sequências............................................ ............233.6 Validação dos testes de detecção dos fungos Alternaria alternata f. sp. citri, Elsinoeaustralis, Elsinoe fawcetti, Guignardia citricarpa e Phytophthora nicotianae........... ............243.6.1 Alteraria alternata f. sp. citri.............................................................................. ............243.6.2 Elsinoe australis.................................................................................................. ............243.6.3 Elsinoe fawcetti ................................................................................................... ............243.6.4 Guignardia citricarpa ......................................................................................... ............253.6.5 Phytophthora nicotianae..................................................................................... ............253.6.6 Validação dos testes em frutos de citros infectados com Alternaria alternata f. sp. citri,
Elsinoe spp. e Guignardia citricarpa após tratamento em packing house .................. ............263.7 PCR quantitativo para Phytophthora nicotianae ................................................... ............263.7.1 Inoculação das plântulas de citros e coleta de material ...................................... ............263.7.2 qPCR................................................................................................................... ............274 RESULTADOS E DISCUSSÃO.............................................................................. ............284.1 Isolamento e manutenção dos fungos Alternaria alternata f. sp. citri, Elsinoe australis eElsinoe fawcetti. ........................................................................................................... ............284.2 Teste de patogenicidade de Alternaria alternata................................................... ............294.3 Seleção de oligonucleotídeos para amplificação dos fungos Alternaria alternata f. sp. citri,
Elsinoe australis, Elsinoe fawcetti, Guignardia citricarpa e Phytophthora nicotianae............314.3.1 Alternaria alternata f. sp. citri............................................................................ .............314.3.2 Elsinoe australis.................................................................................................. .............324.3.3 Elsinoe fawcetti ................................................................................................... .............334.3.4 Guignardia citricarpa ......................................................................................... .............344.3.5 Phytophthora nicotianae..................................................................................... .............354.3.6 Sequenciamento dos produtos da amplificação .................................................. .............364.4 Validação dos testes e determinação do limite de detecção de patógenos de citros............374.4.1 Alternaria alternata f. sp. citri............................................................................ .............374.4.2 Elsinoe australis.................................................................................................. .............404.4.3 Elsinoe fawcetti ................................................................................................... .............424.4.4 Guignardia citricarpa ......................................................................................... .............45


v
SASSERON, Graziéle Ramos. Desenvolvimento e validação de diagnóstico molecular de fungos patogênicos a citros. 2008. 71f. Dissertação (Mestrado em Genética, Melhoramento Vegetal e Biotecnologia) - Pós-Graduação - IAC.
RESUMO
O diagnóstico fitossanitário é de extrema importância no agronegócio citrícola e
envolve atividades que vão desde a produção de plantas matrizes e de material de
propagação até a exportação de frutos. Por isso, a técnica de diagnóstico deve ser
eficiente, reprodutível, segura, e sensível para garantir a qualidade dos procedimentos
de detecção dos patógenos de plantas. Este trabalho teve por objetivo desenvolver e
validar testes moleculares, baseados na PCR, para complementar e ou melhorar o
diagnóstico de fungos patogênicos a citros. Para tanto, foram utilizadas regiões ITS e
seqüências SCAR, para possibilitar a amplificação específica do DNA de diferentes
fungos e sob várias condições. O trabalho foi complementado com ensaios de PCR em
tempo real (qPCR), para monitorar a infecção do oomiceto Phytophthora nicotianae
(Breda de Haan) (= Phytophthora parasitica) (Dastur) em diferentes hospedeiros. Os
microrganismos associados a citros foram obtidos de coleções mantidas no Centro
APTA Citros "Sylvio Moreira" - IAC, e de novos isolamentos de material do Banco
Ativo de Germoplasma de citros. Com exceção de Alternaria alternata (Fr.:Fr) Keissl e
A. alternata f.sp. citri (Fr.:Fr) Keissl, os outros fungos já haviam sido testados em
ensaios biológicos de patogenicidade. A validação dos oligonucleotídeos para PCR
ocorreu usando-se como molde DNA extraído de micélio, e DNA extraído diretamente
de tecido com lesão típica da doença. Para a validação do teste de detecção de P.
nicotianae, discos foliares de lima ácida Tahiti (Citrus latifolia Tanaka), provenientes
do teste da isca, foram utilizados para extração de DNA. Para o qPCR, plântulas de
limão Cravo (Citrus limonia Osbeck), de tangerina Sunki (Citrus sunki Hort. Ex.
Tanaka) e de trifoliata (Poncirus trifoliata (L.) Raf) foram inoculadas com micélio de P.
nicotianae. Material para extração de DNA foi coletado após 1, 2, 4, 6, 8 e 10 dias e
mantido a -20 ºC. Os pares de oligonucleotídeos foram específicos e sensíveis para a
detecção de A. alternata f. sp. citri, E. australis, E. fawcetti, G. citricarpa e
Phytophthora nicotianae. Elsinoe australis (Bitancourt e Jenkis), E. fawcetti (Bitancourt
e Jenkis) e Guignardia citricarpa Kiely foram detectadas em reações de amplificação
contendo dna extraído de 1 a 50 mg de micélio, frutos ou folhas. Alternaria foi
detectada em reações contendo DNA extraído de 20 a 50 mg de micélio, de 1 a 50 mg

vi
de lesões de frutos e de 45 a 50 mg de folhas. P. nicotianae foi detectada em reações
contendo DNA extraído dos discos de folhas utilizados como iscas a partir do segundo
dia após a incubação, contrapondo aos quatro dias observados no método biológico e
aos sete dias para confirmação da presença do patógeno no diagnóstico de rotina. Em
qPCR, P. nicotianae foi detectada após um dia da inoculação em todos os hospedeiros.
Para P. nicotianae foi possível confirmar a presença do patógeno associando o método
molecular ao biológico. Entretanto, o primeiro exige um maior rigor no número de
repetições.
Palavras - Chave: espaçador interno de transcritos (ITS), detecção, gomose do citros, mancha preta dos citros, mancha marrom das tangerinas, verrugose.

vii
SASSERON, Graziéle Ramos. Development and validation of molecular diagnosis of citrus pathogenic fungi. 2008. 71f. Dissertação (Mestrado em Genética, Melhoramento Vegetal e Biotecnologia) - Pós-Graduação - IAC.
ABSTRACT
The phytosanitary diagnosis is very important to citrus agrobusiness and involves
activities from the production of mother plants and material for propagation, to fruits
exportation. Therefore, the technique of diagnosis should be efficient, reproducible, safe
and sensitive to ensure the quality of the procedures for detection of plant pathogens.
This study aimed to develop and validate molecular analysis, based on PCR, to
complement the diagnosis of citrus pathogenic fungi. ITS and SCARs were used in
order to amplify DNA from different fungi under several conditions. The work was
complemented by real-time PCR (qPCR) to evaluate the infection of the oomyceto
Phytophthora nicotianae (= Phytophthora parasitica) (Dastur) in different hosts. The
fungi isolates were from the collection of the Clinica Fitopatologica.. New isolates were
obtained from plant material collected at the Banco Ativo de Germoplasma of the
Centro APTA Citros. Except for Alternaria alternata (Fr.: Fr) Keissl and A. alternata
f.sp. citri (Fr.: Fr) Keissl, the other fungi had already been tested in trials of biological
pathogenicity. The validation of primers for PCR was done using as a template DNA
extracted from fungi mycelium, and from plant material presenting typical symptoms of
the disease. To validate Phytophthora diagnosis, leaf discs of Tahiti acid lime (Citrus
latifolia Tanaka), from the test of bait, were used for DNA extraction. For qPCR,
seedlings of Rangpur lime (Citrus limonia Osb.), Sunki mandarin (Citrus sunki Hort.
Ex. Tanaka) and trifoliate orange (Poncirus trifoliata (L.) Raf) were innoculated with
mycelium of P. nicotianae. Plant material for DNA extraction was collected at 1, 2, 4,
6, 8 and 10 days and stored at -20ºC. The pairs of oligonucleotides were specific and
sensitive to detect the fungi. Elsinoe australis, E. fawcetti and Guignardia citricarpa
Kiely were detected by PCR using as a template DNA extracted from 1 mg to 50 mg of
mycelium, fruits or leaves. Alternaria sp. was detected by PCR using DNA extracted
from 20 to 50 mg of mycelium; 1 to 50 mg of symptomatic fruits, and from 45 and 50
mg of leaves. P. nicotianae was detected by qPCR using as a template DNA extracted
from disk leaves two days after pathogen innoculation, instead of four to seven days to
confirm the presence of the pathogen. Associating the biological to the molecular

viii
method, P. nicotianae was detected, by qPCR, one day after the pathogen innoculation
in all hosts tested. However, the second method requires a higher number of repetitions.
Key words: internal transcribed spacer (ITS), PCR, detection, citrus gummosis, citrus black spot, Alternaria brown spot, citrus scab.

1
1 INTRODUÇÃO
A citricultura é uma das atividades mais importantes do agronegócio brasileiro,
gerando cerca de 400 mil empregos diretos, com faturamento médio de três bilhões de
dólares por ano (Neves et al., 2008). O País é o maior produtor do mundo de frutos
cítricos e de suco concentrado congelado, respondendo atualmente pela exportação de
US$ 1,3 bilhões (Seagri, 2008), possuindo uma área de aproximadamente 600 mil
hectares só no Estado de São Paulo (Abecitrus, 2008).
Historicamente, a citricultura nacional teve profundas mudanças devido a
problemas fitossanitários, desde a troca de porta-enxertos suscetíveis à gomose e à
tristeza, até a obrigação de produção de mudas em ambiente protegido, provocados pela
necessidade de evitar insetos vetores da clorose variegada dos citros.
A citricultura vem convivendo com um grande número de patógenos que estão
surgindo ou sendo introduzidos devido à globalização. Como exemplos do surgimento
temos o caso da morte súbita do citros e, como introdução, o caso do huanglongbing
(HLB ou greening). As doenças já estabelecidas acarretam maiores gastos de produção,
tornando-se parte do manejo da cultura. A introdução ou o surgimento de novas doenças
levam a uma quebra da estabilidade das práticas culturais.
Para evitar a disseminação de patógenos e para ter áreas isentas de novas
doenças, é necessário utilizar mudas sadias, apoiado em um adequado programa de
matrizes. Para tanto é imprescindível a realização de diagnóstico fitossanitário com
rapidez e acurácia. Na citricultura, o uso de mudas sadias vem sendo uma das medidas
adotadas para controlar a disseminação de patógenos, sendo garantido pelos programas
de certificação.
O material de propagação empregado tem que ter qualidade fitossanitária e ao
mesmo tempo ter características genéticas superiores, possibilitando produções
economicamente viáveis, otimizando o manejo com o potencial genético dos cultivares.
Assim, as culturas de propagação vegetativa têm na qualidade fitossanitária um fator
essencial para a sua competitividade, como ocorre no caso dos citros e da cana-de-
açúcar, que naturalmente acumulam patógenos, principalmente vírus e viróides.
No Brasil, as principais doenças de citros transmissíveis por material de
propagação são exocorte, xiloporose, sorose, tristeza, clorose variegada e
huanglongbing. Os agentes causais podem ser eliminados, geralmente, pelo uso de

2
clones nucelares ou micro-enxertia, o que deve ser, posteriormente, confirmado pela
indexação biológica ou molecular. No entanto, os patógenos causadores de doenças
como mancha preta (Guignardia citricarpa), cancro (Xanthomonas axonopodis pv.
citri), clorose variegada (Xylella fastidiosa), HLB (Candidatu sLiberibacter) e gomose
(Phytophthora spp.) podem contaminar mudas, durante sua produção, mesmo usando
sementes e borbulhas sadias.
A legislação sobre comercialização de mudas cítricas sadias no Estado de São
Paulo, por meio dos programas de certificação em viveiros telados, exige que elas sejam
isentas de Xylella fastidiosa, Phytophthora spp., Tylenchulus semipenetrans e
Pratylenchus spp.. Para o envio de mudas para áreas isentas da mancha preta é exigido
o laudo adicional de isenção de Guignardia citricarpa (Phyllosticta citricarpa) (São
Paulo, 2003).
A comercialização de frutos com sintomas de mancha preta, mancha marrom em
tangerinas e da verrugose em laranja doce e limão é muito prejudicada, principalmente
para a exportação de fruta in natura, pois todas elas são consideradas pragas
quarentenárias do tipo A1 em alguns países. Assim, para a exportação torna-se
necessário o atestado fitossanitário, confirmando que os frutos estão livres dos
patógenos (Mendes & Freitas, 2005). No entanto, o diagnóstico usualmente utilizado
para estas doenças baseia-se no isolamento do patógeno em meio seletivo, seguido de
sua caracterização morfológica. Esse procedimento é demorado e depende da ocorrência
de estruturas viáveis dos fungos, o que nem sempre é possível, principalmente para
frutos processados em packing-houses.
Para minimizar o tempo de execução e liberação do atestado de fitossanidade,
uma das alternativas é o uso de técnicas baseadas na PCR, que poderiam ser utilizadas
de modo complementar às usuais. Este método baseia-se na extração de DNA do
material suspeito e amplificação de regiões genômicas específicas de cada patógeno,
através da PCR (Reação em Cadeia da Polimerase). A vantagem de realizar o
diagnóstico molecular reside no tempo de execução, uma vez que, dependendo do caso
e da consistência da validação, poderá dispensar a etapa de isolamento do patógeno.
Isso é particularmente crítico quando não é possível concretizar o isolamento em função
da qualidade e da quantidade disponível do material original.
O diagnóstico para detecção de Phytophthora spp., agente causal da gomose do
citros, é realizado através do teste da isca, que se baseia no tropismo deste oomiceto por
alguns ferimentos nas folhas de variedades cítricas (Feichtenberger, 2001). Este método

3
necessita de, até uma semana para ser concluído, requerendo pessoal treinado para
identificação do esporângio por microscopia. Embora alguns estudos com a utilização
direta do material para análise (substrato ou radicelas) estejam em andamento, ainda não
existe tecnologia segura de obtenção de DNA nessas condições. Assim, a extração de
DNA a partir dos fragmentos foliares obtidos através do método da isca poderia
diminuir o tempo de execução e eliminaria a necessidade de identificação de estrutura
de reprodução.
Com o crescente aumento de informações sobre genoma de vários desses
patógenos e com as possibilidades de focalizar as seqüências ribossomais, como os
espaçadores internos dos transcritos (ITS), para o desenvolvimento de diagnóstico
molecular, é possível acelerar e melhorar a eficiência de detecção com técnicas
baseadas na PCR.
Assim, o objetivo deste trabalho foi desenvolver e validar testes para o
diagnóstico molecular de fungos patogênicos de citros através da PCR, utilizando
oligonucleotídeos de sequências de regiões ITS para detecção do oomiceto
Phytophthora nicotianae, agente causal da gomose dos citros, e dos fungos causadores
da mancha preta dos citros (Guignardia citricarpa), mancha marrom de Alternaria
(Alternaria alternata f. sp. citri) e verrugose (Elsinoe spp.). A técnica da PCR em tempo
real ou PCR quantitativo foi também utilizada no monitoramento da infecção de P.
nicotianae em três genótipos de citros com diferentes suscetibilidades à gomose.

4
2 REVISÃO DE LITERATURA
A citricultura é de suma importância econômica para o Brasil, com uma área de
600 mil hectares colhidos no Estado de São Paulo (FNP, 2006). No entanto, está sujeita
ao ataque constante de inúmeros patógenos e pragas que afetam a produtividade e
elevam os custos de produção. Dentre os patógenos pode-se citar Phytophthora spp. que
causa danos em mudas de viveiro, em plantas no campo e em frutos na pós-colheita,
Guignardia citricarpa, Alternaria alternata f. sp. citri e Elsinoe spp. que afetam
principalmente frutos, tornado-os impróprios à comercialização.
2.1 Gomose dos citros e Phytophthora spp.
A gomose dos citros, causada por Phytophthora spp., é uma das doenças de solo
mais sérias na citricultura e está distribuída em todas as regiões produtoras do mundo.
Estes oomicetos causam perdas na produção, desde regiões áridas até as tropicais
(Whiteside et al.,1993). As perdas podem ocorrer em todas as fases de produção de
citros, desde o viveiro, passando pelo pomar e até em pós-colheita. No viveiro, observa-
se tombamento, podridões do colo, raízes e radicelas (Figura 1 a), gomose, murcha e,
menos frequentemente, podridão foliar. Em pomares com plantas adultas podem ser
observadas podridões do colo, raízes e radicelas, gomose (Figura 1 b) e podridão parda
dos frutos. Em pós-colheita, a podridão parda (Figura 1 c).
O sintoma mais comum da gomose dos citros é a exsudação de goma em lesões
no tronco, que pode ocorrer, também, acima do ponto de enxertia em variedades mais
suscetíveis. Phytophthora spp. colonizam o tronco em direção aos tecidos do câmbio,
necrosando-os e, provocando distúrbios de transporte de seiva para o sistema radicular,
com conseqüente podridão nas raízes e sintomas reflexos na copa (Feichtenberger et al.,
1997). Em situações mais graves, pode ocorrer o anelamento do tronco, levando a morte
da planta.

5
a b c Figura 1 - Sintomas da gomose dos citros. a. Podridão de raízes em limão ‘Cravo’; b. gomose no tronco de limão verdadeiro; c. podridão parda em laranja. Fotos: Aguilar-Vildoso.
O gênero Phytophthora pertence ao reino Stramenopila, ao filo Oomicota, à
classe Oomicetes e à ordem Pythiales. As espécies de Phytophthora capazes de causar
doenças em citros são: P. arecae (Coleman) Pethybridge, P. boehmeriae Sawada, P.
cactorum (Lebert & Cohn) Schröeter, P. capsici (Leonian) Mchau & Coffey, P.
cinnamomi Rands, P. citricola Sawada, P. citrophthora (Smith & Smith) Leonian, P.
drechsleri Tucker, P. hibernalis Carne, P. megasperma Drechsler, P. nicotianae Breda
(= P. parasitica) de Haan var. parasitica (Dastur), P. palmivora Babacauh e P. syringae
(Klebahn) Klebahn.. No Brasil, P. citrophthora e P. nicotianae são os principais agentes
da gomose dos citros (Feichtenberger, 1989). No Estado de São Paulo, P. nicotianae é
predominante (Rossetti et al., 1972; Aguilar-Vildoso, 1997).
As principais características morfológicas das espécies de Phytophthora são: 1)
micélio cenocítico; 2) produção de esporângios; 3) não é produzida vesícula na
liberação dos zoósporos; 4) quando produzem estruturas sexuais produzem um oôgonio
e um anterídio, originando o oósporo; 5) a fase vegetativa é diplóide; 6) os zoósporos
são esporos móveis, biflagelados; 7) os clamidósporos são esporos assexuais podendo
ou não estar presentes, e apresentam parede celular (Aguilar-Vildoso, 1997).
As estruturas agem diretamente na sobrevivência, patogênese e disseminação
deste gênero de oomiceto. A sobrevivência do patógeno por longos períodos no solo é
devido aos clamidósporos e principalmente aos oósporos. Os esporângios são estruturas
dilatadas em relação às hifas, com uma abertura apical, sendo formados na superfície do
solo ou em órgão afetado, sendo impotantes na identificação do patógeno, e na

6
disseminação pela produção dos zoósporos os quais utilizam a água para encontrar
novas plantas (Erwin & Ribeiro,1996).
As espécies do gênero Phytophthora estão divididas em seis grupos, que estão
distribuídos em função do tipo de esporângio e na posição do anterídio. Grupo I -
esporângio papilado e anterídio paragino (P. cactorum); Grupo II - esporângio papilado
e anterídeo anfígino (P. nicotianae e P. citrophthora); Grupo III - esporângio semi
papilado e anterídeo paragino (P. citricola); Grupo IV - esporângio semi papilado e
anterídeo anfígino (P. infestans); Grupo V - esporângio não papilado e anterídeo
paragino (P. megasperma); Grupo VI - esporângio não papilado e anterídeo anfígino (P.
cinnamomi) (Waterhouse, 1963). Baseado nesta divisão, P. citrophthora e P. nicotianae
principais espécies que ocorrem no Brasil estão incluídos no grupo II, por possuírem
esporângios papilados e anterídios anfígenos (Feichtenberger, 2001).
Para a ocorrência da infecção é necessário que ocorra temperatura favorável, que
varia para cada espécie do patógeno envolvido. No geral, a espécie P. nicotianae é
favorecida por temperaturas elevadas (30 oC) e P. citrophthora, por temperaturas mais
amenas (25 oC). Os propágulos infectam somente tecidos do câmbio do tronco quando
os ferimentos são recentes. Após 10 dias da produção do ferimento, os calos formados
impedem a penetração dos patógenos (Rossetti, 2001).
A disseminação de Phytophthora pode ocorrer, geralmente, através de mudas,
substrato e água contaminada, sendo necessário, para evitar isso, o uso de material
propagativo sadio. Devido à sua permanência por longos períodos no solo, através das
estruturas de resistência, e a disseminação deste patógeno ser muito rápida foi adotada
uma medida no Estado de São Paulo para minimizar os danos causados por este
patógeno, sendo obrigatório para a comercialização de mudas o certificado
fitossanitário.
Para o diagnóstico, o teste de rotina utilizado é o da isca com folhas cítricas. Este
método baseia-se no tropismo dos zoósporos por exsudações que ocorrem de ferimentos
de tecidos vegetais (Erwin & Ribeiro, 1996). O método é suficientemente prático para
ser usado como rotina, mas demorada cerca de uma semana, e exige longo trabalho e
observação dos fragmentos foliares em microscópio óptico. Sua principal vantagem está
associada ao fato de detectar formas vivas do oomiceto, simplicidade e necessitar de
equipamentos disponíveis em qualquer laboratório. Entretanto, há necessidade de
pessoal treinado e pode ocorrer subjetividade, dependendo das interferências do teste,
como outros microrganismos ou sujeiras no substrato (Feichtenberger, 2001).

7
Várias estratégias moleculares têm sido desenvolvidas para diferenciar espécies
de Phytophthora, incluindo o uso de RAPD (Goodwin et al., 1989; 1990), DNA
mitocondrial (Forster et al., 1988 e 1990), e DNA ribossomal (rDNA) (Lee & Taylor,
1992; 1993). A associação das regiões ITS (Internal Transcribed Spacer) do rDNA e a
PCR (Polymerase Chain Reaction) mostram-se adequadas para diferenciar espécies de
Phytophthora (Lee & Taylor, 1992).
Os estudos baseados na região ITS de espécies de Phytophthora do grupo
taxonômico IV mostram variação nesta região, possibilitando, assim, a diferenciação
entre as espécies (Tooley et al., 1997). Assim, a detecção de espécies de Phytophthora
pelo uso de técnicas moleculares, vem sendo utilizada para várias espécies deste gênero
em diferentes plantas.
Em citros, várias técnicas vêm sendo desenvolvidas para a diferenciação e
identificação de P. nicotianae e P. citrophthora. Dentre elas, o uso de sondas para
hibridização, PCR nested e a PCR, ambos baseados na região ITS, possibilitaram a
diferenciação entre as espécies do gênero Phytophthora em solo contaminado (Érsek et
al., 1994; Ippolito et al., 2002, Ippolito et al., 2004;Oliveira et al., 2008) e, em água
(Kong et al., 2003).
2.2 Mancha preta dos citros e Guignardia citricarpa
A mancha preta dos citros (MPC) ou pinta preta, causada pelo fungo Guignardia
citricarpa Kiely, (Phyllosticta citricarpa (MacAlpine) van der Aa), afeta principalmente
frutos (Feichtenberger, 1996). Em citros, além da espécie patogênica, há uma espécie
endofítica, Guignardia mangiferae A. J. Roy (P. capitalensis Hennings), que dificulta o
diagnóstico tradicional, realizado inicialmente pelo isolamento, levando à necessidade
da PCR.
Nos frutos, os sintomas preferencialmente aparecem no estágio final de
desenvolvimento, após a colheita, durante o seu armazenamento e durante o seu
transporte. Em geral, plantas velhas e estressadas estão mais sujeitas à doença, assim
como os frutos mais expostos aos raios solares e a altas temperaturas (Kotzé, 1981).
Além dos frutos, folhas, ramos e pecíolos, podem ser afetados. Porém, o maior
dano ocorre nos frutos, podendo-se observar seis tipos de sintomas da doença (Figura
02): 1) mancha preta (hard spot) é o sintoma mais típico, formando lesão bem definida e

8
circundada por uma borda de cor marrom escura a preta com pontuações pretas
correspondendo as picnídios (Figura 02 a); 2) mancha sardenta (fleckle spot) ocorre em
frutos maduros e após colheita, formando pequenas lesões definidas de coloração
avermelhada que evoluem para pardo-avermelhado (Figura 02 b), 3) mancha trincada
(cracked spot) aparece em frutos verdes, podendo atingir pequenas áreas até a totalidade
do fruto, mas são superficiais (Figura 02 c); 4) mancha virulenta (virulent spot) ocorre
pela fusão de diferentes tipos de sintomas, desde a típica até a sardenta (Figura 02 d); 5)
falsa melanose (speckled blotch ou false melanose), com lesões pequenas e definidas,
sendo semelhantes às da melanose dos citros, entretanto com pontuações satélites
(Figura 02 f); 6) mancha rendilhada (lacy spot) caracterizada por ocorrer em frutos
verdes e atingir grandes áreas do fruto, porém superficial, entretanto os produtores
denominam este sintoma de falsa melanose, criando confusão com a denominação
internacional (Figura 02 e) (Aguilar-Vildoso et al., 2002).
Em folhas, os sintomas são frequentes em limão verdadeiro (C. limon (L.)
Burm. f.), limão Cravo (C. limonia Osbeck) ou em tangerinas, principalmente na
mexerica do Rio (C. deliciosa Tenore), apresentando manchas pequenas com bordas
bem definidas e de coloração marrom-avermelhada a preto. Nos ramos verdes,
pedúnculo, pecíolo e espinhos as lesões são pequenas, com bordas pretas bem definidas
e com centro claro e no cálice do fruto, as lesões podem variar de avermelhada a
marrom-avermelhada (Aguilar-Vildoso et al., 2002).
Atualmente, G. citricarpa está inserido no reino Fungi, subreino Dycaria, do filo
Ascomicota, da classe Dothiomycetes e da ordem dos Botryosphaeriales (Hibbett et al.,
2007), estando Guignardia citricarpa na forma teleomórfica e Phyllosticta citricarpa na
forma anamórfica (Glienke-Blanco, 1999).
O fungo G. citricarpa possui pseudotécio globoso a subgloboso, os ascos são na
forma de clava, bitunicados, contendo oito ascósporos unicelulares e hialinos. Os
ascósporos possuem apêndices hialinos (capa gelatinosa) nas duas extremidades. A capa
gelatinosa permite a fixação à superfície de contato e, ao serem liberados ativamente, os
ascósporos são levados por corrente de ar a grandes distâncias (Aguilar-Vildoso et al.,
2002). A sua fase assexual (P. citricarpa) é caracterizada pela presença de um apêndice,
estrutura que se projeta do conídio, semelhante a um flagelo. Os picnidiósporos são
hialinos, unicelulares com apêndices que podem ser caducos, envoltos por uma capa
gelatinosa. Os picnídios são pequenos, globosos e pretos (Aguilar-Vildoso et al., 2002).

9
a b c
d e f Figura 02 - Sintomas de mancha preta do citros (Guignardia citricarpa) em frutos de laranja doce. a. mancha dura; b. mancha sardenta, c. mancha trincada, d. mancha virulenta, e. mancha rendilhada, e f. falsa melanose. Fonte: Aguilar-Vildoso et al. (2002).
O ciclo da doença inicia-se com a queda das folhas que entram em
decomposição, formando os corpos de frutificação conhecidos como pseudotécios. No
ciclo primário, a produção de esporos sexuais é importante para a sobrevivência do
fungo, devido à sua maior viabilidade e resistência ao ressecamento. Quando os
ascósporos entram em contato com o tecido aderem-se pela mucilagem e, na presença
de água, começam a germinar, formando o tubo germinativo e o apressório, penetrando
diretamente no tecido, sem a necessidade de aberturas naturais ou ferimentos (Aguilar-
Vildoso et al., 2002).
O início do ciclo secundário ocorre através da formação dos esporos assexuais
nos picnídios, os picnidiósporos, formados em folhas, ramos e frutos. Os picnidiósporos
emergem através de um ostíolo. A fase assexual do ciclo secundário é responsável pela
doença na planta e nas plantas vizinhas. Os picnídios são formados nas folhas dos citros,
em decomposição, junto com os pseudotécios; os esporos produzidos infectam a região
da saia da planta, pelos respingos de chuva, sempre a curtas distâncias. O ciclo da
doença é fechado quando a infecção por ascósporos ou por picnidiósporos ocorre em
novas folhas. A queda dessas folhas e sua decomposição levam à formação de novos

10
esporos sexuais (ascósporos) iniciando, assim, um novo ciclo primário (Aguilar-Vildoso
et al., 2002).
A identificação da mancha preta em frutos é realizada pela observação da
presença de picnidiósporos com um apêndice na extremidade do esporo, que se torna
caduco, sendo mais bem evidenciado ao microscópio óptico por contraste de fase. Essa
é a característica do gênero Phyllosticta, forma anamórfica do gênero Guignardia
(Aguilar-Vildoso et al., 2002).
A detecção de G. citricarpa em tecidos suspeitos sem picnídios é feita
inicialmente por isolamento e avaliação morfológica da colônia e, em seguida, por PCR.
No isolamento, qualquer meio de cultura apropriado para fungos poderá ser utilizado,
mas para uma maior eficiência, pode ser usado um meio suplementado com glicose
(Campos & Aguilar-Vildoso, 2001). Em meios de cultura comuns, as colônias são
escuras e estromáticas, enquanto que em meio semi-seletivo com glicose, são
inicialmente sem pigmentação e, posteriormente, tornam-se de coloração cinza, até
preto.
O principal problema na detecção de G. citricarpa está na existência de uma
outra espécie do gênero (G. mangiferae), considerada endofítica e não patogênica a
citros. Regiões ITS (Baayen et al., 2002; Bonants et al., 2003; Peres et al, 2007) e
alguns SCAR (Glienke-Blanco, 1999) estão sendo utilizados para desenhar
oligonucleotídeos para diferenciar as duas espécies, por PCR.
O uso da PCR em diagnósticos de rotina pode diferenciar as duas espécies, G.
citricarpa e G. mangiferae além da vantagem de utilizar pequenas quantidades de lesões
de frutos e folhas, o que reduz o tempo para a confirmação do resultado, uma vez que o
isolamento em meio de cultura torna-se desnecessário.
2.3 Mancha marrom das tangerinas e Alternaria alternata f. sp. citri
A mancha marrom de Alternaria (MM), causada pelo fungo Alternaria alternata
f. sp. citri (Fr.:Fr) Keissl patótipo tangerina, afeta frutos, folhas e ramos (Figura 03 a, b,
c) de tangerinas, tangores e tangelos e provoca queda de folhas, seca de ramos e queda
prematura de frutos, comprometendo a produtividade das plantas. As lesões da doença
em frutos reduzem também o seu valor quando comercializados in natura (Whiteside et
al., 1993) (Figura 03 a). Em outros países, como a África do Sul, Argentina, Austrália,

11
Colômbia, Cuba, Espanha, Estados Unidos, Israel, Itália e Turquia, essa doença é de
grande importância econômica. No Brasil, foi introduzida em 2001 e vem causando
elevadas perdas em cultivos comerciais de tangor Murcott (C. reticulata Blanco x C.
sinensis Osbeck) e tangerina Poncan (C. reticulata Blanco) (Feichtenberger et al.,
2005).
Nos frutos, as lesões são de coloração marrom-escuras, inicialmente deprimidas
no centro, podendo apresentar exsudatos de consistência viscosa e são circundadas por
um halo amarelado (Figura 03 a). O centro das lesões podem tornar-se corticoso e
saliente, formando pústulas que se destacam facilmente deixando orifícios na superfície.
Nas folhas jovens, as lesões são necróticas e inicialmente pequenas, de coloração
marrom ou preta, com halos amarelados ao seu redor. Com o tempo, essas lesões
aumentam de tamanho e tomam grandes áreas da superfície da folha (Figura 03 b). Em
ramos novos, as lesões são necróticas, de tamanho variável e coloração marrom-escura
(Feichtenberger et al., 2005) (Figura 03 c).
a b c Figura 03 - Sintomas da mancha marrom em citros (Alternaria alternata f. sp. citri) patótipo tangerina. a. sintomas em frutos ; b. sintomas em folhas e c. sintomas em ramos. Foto: Aguilar-Vildoso.
O gênero Alternaria spp. inclui espécies saprofíticas, endofíticas e patogênicas
(Rotem, 1994, Peever et al., 2004). Dentre estas espécies, três são descritas como
causadoras de doenças em frutos cítricos, dentre elas: 1) Mancha marrom de Alternaria
(Alternaria brown spot), afeta frutos, folhas e ramos de tangerinas, tangelos e tangores,
provocada por A. alternata f. sp. citri patótipo tangerina que produz a toxina ACT, 2)
Mancha foliar de Alternaria (Alternaria leaf spot), afeta as folhas e frutos de limões , o
agente causal é a A. alternata f. sp. citri patótipo limão, que produz a toxina ACR ou

12
ACRL e 3) Podridão negra de Alternaria (Alternaria black rot), afeta os frutos, podendo
muitas vezes ocorrer a sua queda, causada pela A. alternata que não produz toxina
(Laranjeira et al., 2005).
Alternaria alternata pertence ao reino Fungi, ao sub-reino Dycaria, ao filo
Ascomicota, classe Pleosporomycetidae e a ordem dos Pleosporales (Isshiki et al.,
2003; Thomma, 2003). Este fungo, na forma anamórfica, produz esporos que podem ter
forma e tamanho variáveis, e coloração de oliva a marrom-escura, dotados de quatro a
seis septos transversais e um ou mais longitudinais.
A espécie A. alternata contém sete patótipos, que produzem toxinas que causam
doenças em diferentes plantas, dentre elas: a toxina AF, que causa doenças em
morangos, a toxina AK, que afeta pêra, a toxina ACT, que afeta especificamente
tangerinas, tangores e tangelos (Hatta et al., 2002) e a toxina ACR ou ACRL que afeta
limões (Isshiki et al., 1997; Akimitsu et al., 2003).
No caso da mancha marrom em tangerinas, a ACT é uma micotoxina formada
pelo ácido tenuazônico que, em contato com a planta, atua na membrana plasmática
(Walton, 1996), provocando a sua permeabilização e a morte celular. Assim, o
desenvolvimento do fungo sobre os tecidos mortos, é devido a produção de um grande
número de conídios, que podem ser carregados pelo vento até a superfície de frutos ou
para outros órgãos de plantas, iniciando novas infecções. Os sintomas aparecem nos
frutos 24 a 48 horas após a infecção. A toxina produzida pelo fungo é responsável pelas
áreas necrosadas dos tecidos atacados, sendo que o tamanho das lesões e a extensão da
necrose podem variar, em função da suscetibilidade do tecido da variedade cítrica
(Feichtenberger et al., 2005).
A detecção do agente causal da mancha marrom é realizada pelo isolamento em
meio de cultura suplementado com tetraciclina e um fungicida do grupo dos
benzimidazóis, para inibir o crescimento de bactérias e outros fungos, e posteriormente,
identificação morfológica dos esporos (Aguilar-Vildoso et al., 2004). A caracterização
morfológica dos esporos é complicada, uma vez que as duas espécies de Alternaria que
afetam citros são produtoras de toxinas, a mancha marrom de Alternaria (A. alternata f.
sp. citri patótipo tangerina) e a mancha foliar de Alternaria (A. alternata f. sp. citri
patótipo limão) possuem esporos pequenos. Por isso, outros métodos estão sendo
empregados para detecção e diferenciação deste fungo, dentre eles, testes biológicos,
análises bioquímicas e métodos moleculares (Akimitsu et al., 2003).

13
O uso de métodos moleculares, baseados na PCR, pode ser utilizado em análises
de rotina, uma vez que reduziria o tempo para a identificação do patógeno, e eliminaria
o isolamento e posterior caracterização morfológica através da microscopia. A
vantagem do uso de testes moleculares para exame de rotina está na rapidez e acurácia
dos resultados. Dentre os métodos moleculares encontram-se o uso de RAPD para
estudos da estrutura genética de diferentes populações de A. alternaria (Peever et al.,
1999, Peever et al., 2002), a utilização da técnica de PCR, baseando-se em
oligonucleotídeos desenhados a partir do gene da toxina ACT, que causa doença em
tangerinas. Vários trabalhos estão sendo realizados para detecção e diferenciação de A.
alternata f. sp. citri patótipo tangerina, através de Southern blot e da PCR, com base
nestes genes (Masunuka et al,2000).
2.4 Verrugose e Elsinoe australis e Elsinoe fawcetti
A verrugose do citros é causada pelo fungo Elsinoe australis Bitancourt e
Jenkins (Sphaceloma australis Bitancourt e Jenkis) e por Elsinoe fawcetti Bitancourt e
Jenkis (Sphaceloma fawcetti Bitancourt e Jenkis). Esse último fungo é encontrado
praticamente em todas as regiões produtoras do mundo, afetando folhas, ramos, frutos e
botões florais (Figura 04a e b). Enquanto, E. australis está confinada à América do Sul e
afeta principalmente frutos de laranjas doce (Fantin, 1988) (Figura 04 c).
A verrugose da laranja Azeda afeta, limões, limas, tangerina e a própria laranja
Azeda, sendo causada por E. fawcetti que provoca lesões corticosas, cor de palha, mais
ou menos salientes, atacando frutos, folhas e ramos ainda jovens, causando deformações
salientes na casca, que vão crescendo à medida que o fruto se desenvolve (Figura 04 a,
b). Em condições favoráveis podem ocorrer pequenas pústulas pretas sobre as lesões,
onde se formam as frutificações do fungo, que garantem a sua disseminação (Laranjeira
et al., 2005).
A verrugose da laranja doce causada por E. australis afeta somente os frutos.
Essa forma da doença ocorre principalmente na América do Sul, com desenvolvimento
de lesões salientes e corticosas, semelhantes às descritas para a verrugose da laranja
Azeda. E. australis (S. australis) ataca os tecidos jovens dos frutos em formação, logo
após a queda das pétalas (Laranjeira et al., 2005) (Figura 04 c).

14
b a b c
Figura 04 - Sintomas de verrugose. a. fruto de limão Cravo, b. folha de limão Cravo, c. fruto de laranja Pêra.
O gênero Elsinoe pertence ao reino Fungi, ao sub-reino Dycaria, ao filo
Ascomiceto, e a ordem Myriangiales e da família Elsinoaceae (Swart et al., 2001, Kim
et al., 2007, Hibbett et al., 2007). Esses fungos são raramente encontrados na fase
perfeita. Os ascomas são pulvinados, circulares e elípticos, compostos de tecido
pseudoparenquimático contendo vários lóculos com ascos. Os ascos são subglobosos a
elípticos, com parede espessada no topo, contendo oito ascósporos. Os ascósporos são
hialinos, oblongo elíptico, com 1 a 3 septos, usualmente constritos no septo central
(Laranjeira et al., 2005). O acérvulo é epidérmico a sub-epidérmico, composto de
pseudoparênquima hialino a marrom pálido. Os conídios são hialinos, unicelulares,
elipsoidais (Sivanesan & Critchett, 1974b; Fantin & Kimati, 1989b; Laranjeira et al.,
2005).
Para a detecção de Elsinoe spp. foi desenvolvido um meio semi-seletivo para o
isolamento de Elsinoe de lesões de verrugose (Fantin, 1988; Fantin & Kimati, 1989a).
Atualmente, métodos moleculares estão sendo empregados para detectar e diferenciar
espécies de Elsinoe que afetam citros. Estes métodos estão sendo realizados pelo uso de
análises da região ITS do DNA ribossomal (Tan et al., 1996) e por RAPD (Hyun et al.,
2007).
A diferenciação e detecção de Elsinoe spp. são de suma importância para o
controle da doença e principalmente para impedir a sua entrada em áreas livres, uma vez
que E. australis é uma praga quarentenária do tipo A1 em alguns países. A vantagem do
diagnóstico molecular é a rapidez e acurácia na detecção e diferenciação do fungo, o
que não ocorre no isolamento, pois neste procedimento é necessário um longo tempo
para o crescimento do fungo e nem sempre se consegue obter o isolado.

15
2.5 Detecção de regiões ITS em fungos
A utilização da PCR e de outras técnicas moleculares para caracterizar fungos é
possível devido à presença de seqüências específicas. As regiões mais utilizadas em
sistemática são as que transcrevem os RNA ribossômicos (rDNA), porque encontram-se
em seqüência e com grande número de repetições (‘tandem’), além de não sofrerem
seleção, comportando-se como neutros, podendo-se estimar o tempo da divergência
entre os taxons. No diagnóstico, vêm sendo utilizadas as regiões do rDNA, pelo grande
número de cópias e pelo grande número de espécies que possuem suas seqüências em
bancos de dados. Essas regiões são caracterizadas pela alternância de seqüências
transcritas e não transcritas. As regiões não transcritas são a ITS e a IGS. A ITS possui
duas regiões denominadas de ITS-1, amplificadas pelos oligonucleotídeos universais
ITS1 e ITS2, que se encontram entre o 18S e o 5.8S; já a ITS2, que encontra-se entre o
5.8S e o 25-28S, é amplificada pelo ITS3 e ITS4. O fragmento total do ITS é obtido
pelo uso do ITS1 e ITS4, amplificando juntamente toda a região 5.8S, variando de 600 a
800 pares de bases. As características que marcam está região como conveniente para a
identificação molecular são: i. toda a região ITS é frequentemente entre 600 a 800 pares
de bases e pode ser amplificado com oligonucleotídeos universais, ii. multicópias
naturais de repetição de rDNA fazem a região ITS fácil para amplificar pequenos
fragmentos e iii. estudos vêm demonstrando que regiões ITS são frequentemente
variáveis entre a distinção morfológica das espécies de fungos (Gardes & Bruns, 1993).
Vários testes moleculares vêm sendo utilizados para a identificação de fungos,
incluindo o uso de fragmentos de DNA mitocondrial e ribossomal rDNA. Além da
identificação de fungos, as regiões ITS vêm sendo utilizadas para estudos de filogenia,
através do uso da técnica de PCR. Nos ascomicetos, a morfologia e similaridade na
maioria das classes estão intimamente relacionadas, mas a similaridade pode envolver
grupos não relacionados. Na PCR, com o uso de oligonucleotídeos obtidos de regiões
ITS, é possível promover novas fontes de dados para a reconstrução filogenética deste
grupo de fungos. Algumas regiões do 26S do gene de rDNA são pouco conservadas e
freqüentemente com grandes mutações, esta região tem servido como marcadores
potenciais para diferenciar fungos e populações (Tan et al., 1996).
PCR utilizando oligonucleotídeos para amplificar a região ITS do rDNA tem
diferenciado alguns patógenos como Phytophthora, Pythium, Verticillium, Fusarium
entre outros. As regiões ITS-1 e ITS-2 vêm sendo utilizadas para o diagnóstico das

16
doenças causadas por oomicetos; a região ITS2 é o principal foco de estudo para as
espécies de Phytophthora spp. (Tooley et al., 1997); e para Guignardia citricarpa vêm
sendo realizados testes moleculares, através de amplificação das regiões ITS-1 e ITS-2
do rDNA (Bonants et al., 2003).
2.6 qPCR (PCR quantitativo)
PCR quantitativo em tempo real (qPCR) é um método que foi desenvolvido com
o objetivo de determinar o número de moléculas alvo que servem de molde para
amplificação pela DNA polimerase. A diferença entre o qPCR e o PCR comum é que o
primeiro método quantifica o produto de PCR em cada ciclo, utilizando uma curva
exponencial, fazendo com que a quantidade de material inicial seja determinada. No
segundo método o resultado corresponde à fase de saturação da reação (Gachon et al.,
2004).
A principal vantagem do qPCR é a rapidez em fornecer dados confiáveis. Este
método é altamente sensível na detecção de DNA e RNA, devido à combinação de
amplificação pelo PCR e o sistema de detecção, podendo em qualquer caso verificar a
especificidade com a PCR por eletroforese, curva de dissociação e dados do
sequenciamento. Outra vantagem desta técnica é que pode ser utilizada para estudos
com pequena quantidade de material inicial (Freeman et al., 1999).
Atualmente existem três estratégias para detectar os produtos de qPCR (Bustin,
2000) e todas utilizam corantes fluorescentes. O método SyBR green utiliza corantes
fluorescentes que se ligam às moléculas de DNA de dupla fita, enquanto que a
Molecular beacon e Taqman baseiam-se na hibridização de sondas fluorescentes que se
ligam ao amplicon específico (Bustin, 2000). Os métodos mais utilizados em plantas são
o SyBR green e o Taqman (Gachon & Saindrenan, 2004).
O método SyBR green é mais versátil porque seu agente é intercalante e liga-se a
qualquer tipo de seqüência, além de permitir que vários genes possam ser detectados em
uma mesma amostra (Bustin, 2000).
O qPCR vem sendo utilizado em estudos com plantas na quantificação e
detecção de DNA exógeno, no diagnóstico e na quantificação de patógenos ou
microrganismos asssociados com plantas (Mayer et al., 2003) e na seleção de genótipos
resistentes, como Verticillium albo-atrum em alfafa (Larsen et al., 2007) .

17
Além dessas aplicações, e por ser um teste rápido e seguro, qPCR vem
auxiliando no diagnóstico de fungos como: Phoma tracheiphila (Licciardello, 2006),
Guignardia citricarpa (Gent-Pelzer et al., 2007), Phytophthora spp. (Ippolito et al.,
2004; Hayden et al., 2006; Yan & Liou, 2006; Bilodeau et al., 2007) e de bactérias
como a do cancro cítrico (Xanthomonas axonopodis pv. citri) (Mavrodieva et al., 2004).
Alguns estudos estão sendo feitos para a diagnose de oomicetos através dos três
métodos de qPCR, Taqman (Lee & Taylor., 1993), SyBR green e Molecular beacon
(Tyagi & Kramer, 1996), para identificar e detectar P. ramorum em amostras
contaminadas com o patógeno.
Em citros, vários trabalhos vêm sendo realizados para detecção e diferenciação
de Phytophthora spp. Seqüências de oligonucleotídeos específicos para P. nicotianae e
P. citrophthora foram modificados para obter oligonucleotídeos do tipo Scorpion, que
possui uma alça (harpin), sendo possível com esta técnica diferenciar e detectar as duas
espécies deste gênero que afetam citros (Ippolito et al., 2004). O qPCR também está
sendo utilizado na seleção de genes do controle interno nas diferentes fases do ciclo de
vida de P. nicotianae (ciclo assexual, ciclo sexual e fase de patogênese) (Yan & Liou,
2006).
3 MATERIAL E MÉTODOS
3.1 Isolamento, manutenção e conservação dos fungos Alternaria alternata f. sp.
citri, Elsinoe australis, Elsinoe fawcetti, Guignardia citricarpa e Phytophthora
nicotianae
Os isolados de Phytophthora nicotianae (IAC 01/95) obtido de substrato
contaminado, Guignardia citricarpa (IAC 13/96) obtido de lesões típicas de mancha
preta em frutos de limão Verdadeiro (C. limon), coletados em Mogi Guaçu/SP e
Guignardia mangiferae (IAC 41/99) obtido de folhas assintomáticas de tangerina
Poncan (Citrus reticulata Blanco) do Banco de Matrizes do Centro APTA Citros Sylvio
Moreira, Cordeirópolis/SP utilizados no trabalho, fazem parte da coleção mantida na
Clínica Fitopatológica de Citros do Centro APTA Citros Sylvio Moreira,
Cordeirópolis/SP. Os isolados de Phytophthora citrophthora (LRS 44/87) obtido de
substrato contaminado e o isolado de Alternaria alternata f. sp. citri patótipo limão

18
(LRS 26/04) isolado a partir de lesões em folhas sintomáticas, coletada no município de
Capão Bonito/SP foram cedidos pela Unidade de Pesquisa e Desenvolvimento de
Sorocaba - Instituto Biológico, Sorocaba/SP.
Os fungos foram mantidos em meio cenoura-ágar (CA) (200g de cenoura, 20g
de ágar por litro de meio de cultura), a 25 °C; os isolados de P. nicotianae e P.
citrophthora foram repicados a cada 15 dias, de G. mangiferae e G. citricarpa a cada 30
dias e de A. alternata a cada 20 dias. Para a conservação de P. nicotianae usou-se o
método de Castellani modificado por Figueiredo (1967) e para a de Guignardia spp. e
A. alternata em tubo inclinado com meio cenoura-ágar e cobertos com óleo mineral
após crescimento.
A. alternata f. sp. citri foi isolado de lesões de mancha marrom de folhas (IAC
01/06) e frutos (IAC 02/06) de tangor Murcott (Citrus reticulata Blanco x Citrus
sinensis L. Osb.), coletados no Banco Ativo de Germoplasma do Centro APTA Citros
Sylvio Moreira, Cordeirópolis/SP. O isolado de A. alternata foi obtido a partir de lesão
em caule de limão Cravo (IAC 03/06) (Citrus limonia Osbeck.).
Para o isolamento de A. alternata f. sp. citri, lesões características em frutos
foram cortadas em pequenos pedaços, com bisturi, desinfestadas superficialmente com
álcool etílico comercial (96%) por 1 minuto e hipoclorito de sódio a 2% por 2 minutos,
em seguida, foram lavadas com água esterilizada e transferidas para placas de Petri com
meio cenoura-ágar com antibióticos (200g de cenoura, 20g de ágar por litro de meio de
cultura, suplementado para 100 µg/mL de carbendazim (Derosal) e 50 µg/mL de
tetraciclina) (adaptado de Aguilar-Vildoso et al., 2004). Para o isolamento de A.
alternata foi observada uma lesão característica de Alternaria no caule de limão Cravo,
apresentando esporos. A lesão foi homogeneizada em tubo Eppendorf com 1 mL com
água estéril para liberação dos esporos e, em seguida foram plaqueados 100 µL da
suspensão em meio cenoura-ágar com antibiótico (tetraciclina) e incubadas a 25°C.
Estes isolados foram mantidos em meio cenoura-ágar, em BOD a 25 °C, e repicados a
cada 20 dias. A conservação foi feita em tubo inclinado com meio cenoura-ágar e óleo
mineral.
O isolamento de E. australis foi realizado a partir de frutos de laranja Pêra
(Citrus sinensis L. Osbeck) apresentando sintomas típicos. O isolado de E. fawcetti foi
obtido de frutos de limão Cravo (Citrus limonia Osbeck) com sintomas típicos de
verrugose. Os frutos foram lavados em água corrente, borrifados com álcool (96%) e
hipoclorito de sódio comercial a 2%. As lesões foram raspadas sobre três meios de

19
cultura diferentes, ágar - ágar (20g de ágar por litro de meio de cultura), cenoura - ágar
(200g de cenoura, 20g de ágar por litro de meio de cultura) e BDA (39g de BDA
MERCK por litro de água destilada), suplementado com 100 µg/mL de carbendazim
(Derosal), 100 µg/mL de pentacloronitrobenzeno (PCNB) e 50 µg/mL de tetraciclina
(adaptado de Fantin, 1988).O crescimento desse fungo foi lento, o que propicia o
crescimento de outros microrganismos.
3.2 Ensaio de patogenicidade dos fungos Alternaria alternata f. sp. citri, Elsinoe
australis, Elsinoe fawcetti, Guignardia citricarpa e Phytophthora nicotianae
Para os isolados de Phytophthora spp. (IAC 01/95 e LRS 44/87), Guignardia
spp. (IAC 13/96) e Alternaria alternata de limão (LRS 26/04) não foi realizado o teste
de patogenicidade, pois a sua patogenicidade já tinha sido comprovada em trabalhos
anteriores. O teste de patogenicidade foi realizado para os isolados de A. alternata e
Elsinoe spp. obtidos durante a realização deste trabalho, entretanto, para Elsinoe spp.
não foi possível comprovar a patogenicidade pelo uso do teste biológico, uma vez, que
este fungo é de difícil esporulação.
No teste de patogenicidade para A. alternata foi preparada uma suspensão de
esporos de Alternaria alternata f. sp. citri e outra com o de Alternaria alternata. Para
tanto, foram adicionados 10 mL de água destilada esterilizada em placas de Petri com
colônias de A. alternata f. sp. citri ou A. alternata em meio CA, com 25 dias. As
colônias foram suspensas com auxílio de alça de Drygalsky.
Para a inoculação, ramos de mudas sadias de tangor Murcott (suscetível) foram
colocados em câmara úmida. Como controle resistente foi utilizado ramos de laranjeira
Pêra. Cada folha do ramo foi inoculada com 10 µL da suspensão com micélio e esporos
do fungo e mantidas a 27°C até o aparecimento do sintoma.
3.3 Extração de DNA total dos fungos e das lesões
A extração de DNA foi realizada com algumas modificações das instruções da
Promega para o Wizard Genomic DNA Purification Kit, Manual Técnico nº 050, tanto
de micélio, quanto de tecido foliar e de lesões em frutos. As alterações realizadas são

20
descritas após o protocolo geral. A extração de DNA foi realizada a partir de 50 mg de
micélio, lesões em folhas ou frutos coletados no campo. O material foi triturado com
nitrogênio líquido e transferido para tubos Eppendorf. Em seguida, foram adicionados
600 µL de ‘Nuclear Lise Solution’, foi feita uma agitação em Vortex e os tubos foram
incubados em banho-maria a 65 °C por 15 minutos. Foram adicionados 3 µL de ‘Rnase
Solution’, e os tubos foram incubados a 37 °C durante 15 minutos. Em seguida foram
adicionados 200 µL de ‘Protein Precipition Solution ’ e foi feita uma centrifugação a
13.000 rpm por 10 minutos. O sobrenadante foi transferido para um novo tubo
Eppendorf contendo 600 µL de isopropanol à temperatura ambiente. Após
homogeneização, foi feita uma centrifugação a 13.000-16.000 rpm durante 5 minutos, à
temperatura ambiente. O sobrenadante foi descartado e o sedimento lavado com 600 µL
de etanol 70% à temperatura ambiente. Após centrifugação, o DNA foi seco a 55°C por
10 a 15 minutos, suspenso em 50 µL de ‘DNA Rehidration Solution’ e, em seguida,
estocado a -20°C.
As modificações que foram introduzidas na metodologia básica para extração de
DNA foram para melhorar a qualidade do DNA e incluíram as seguintes etapas:
- Nos discos foliares colonizados por Phytophthora nicotianae obtidos a partir
do teste da isca, o material foi triturado sem nitrogênio líquido, apenas com a ‘Nuclear
Lise Solution’.
- Nas lesões de Alternaria alternata retiradas de frutos e folhas, no final do
protocolo de extração, após a adição de 50 µL de ‘DNA Rehidration Solution’, foram
colocadas no tubo Eppendorf de 1,5 mL duas pérolas de vidro e foi feita uma incubação
de 2°C a 10°C até completa dissolução do DNA. Após a dissolução, as pérolas de vidro
foram transferidas para outro tubo contendo 50 µL de ‘DNA Rehidration Solution’ e
essa solução foi utilizada para PCR.
3.4 Oligonucleotídeos e amplificação por PCR dos fungos Alternaria alternata f. sp. citri, Elsinoe australis, Elsinoe fawcetti, Guignardia citricarpa e Phytophthora
nicotianae
Os oligonucleotídeos utilizados para a amplificação de DNA de A. alternata e
Elsinoe spp. foram desenhados a partir de seqüências disponíveis no Genbank
(http://www.ncbi.nlm.nih.gov). A posição e orientação das regiões amplificadas estão

21
indicadas na Figura 05. Os oligonucleotídeos para detecção de P. nicotianae, G.
citricarpa e A. alternata f. sp. citri foram obtidos de Tooley et al.(1997); Ippolito et al.
(2004); Glienke-Blanco. (1999); Bonants et al. (2003); Peres et al. (2007); Masunuka et
al. (2000), respectivamente (Tabela 01). O desenho dos oligonucleotídeos também foi
realizado para E. australis e E. fawcetti, com o auxílio do programa Jellyfish
(http:\jellyfish.labvelocity.com/index.jhtml.url).
As reações de amplificação foram preparadas usando como molde DNA extraído
de micélio de culturas puras e de frutos e folhas sintomáticas para as respectivas
doenças. Para cada condição de amplificação, as reações foram otimizadas
considerando-se temperatura de anelamento, concentração de DNA e magnésio e
número de ciclos. A Tabela 02 apresenta as informações sobre as condições das reações
e ciclos da PCR.
Tabela 01 - Especificações dos pares de oligonucleotídeos usados para detecção de Alternaria
alternata f. sp. citri, Elsinoe australis, Elsinoe fawcetti, Guignardia citricarpa, Guignardia
mangiferae e Phytophthora nicotianae.
Patógeno Primer Seqüência Tamanho do
amplicon (pb)
Referência ou Nr. do
Acesso
AAP+ 5’ TTTGATGCTTAGAAGTAGTTTATTTG 3’
AAP- 5’ AGAGTTCAGGAAATGACTGCAG 3’
571 AY295065
ACTT1-F2 5’ CACAGGCTATCTTCACATGC 3’
Alternaria alternata f.
sp. citri
ACTT1-R2 5’ CGTAGCTCCATAGCATTGCT 3’ 781 MASUNAKA et al., 2000
CSP+ 5’ GTTTCTTTTCCTCCGCTTATTG 3’
CSP- 5’ GTAGGTGAACCTGCGGAAGGAT 3’ 636 U28059
Els 156-175F 5’ GGAGGACCGAATCAACTCTG 3’
Els554-573R 5’ CAGCGGGTATTCCTACCTGA 3’ 418 U28057
Els156-175F 5’ GGAGGACCGAATCAACTCTG 3’
Els503-522R 5’ AGGTTTCACGGCAGAGGTTA 3’ 377 U28057
Els156-175F 5’ GGAGGACCGAATCAACTCTG 3’
Elsinoe australis
Els 253-272R 5’ATTTCGCTGCGTTCTTCATC 3’ 117 U28057
CSP+ 5’ GTTTCTTTTCCTCCGCTTATTG 3’
CSPi 5’ TTGACGGCGGAGGGTCACCGGTG 3’ 525 AF097577
Els183-202F 5’ TGATTGGTAAATGACGCTC 3’ Elsinoe fawcetti
Els374-393R 5’TGATTGGTGAAATGACGCTC 3’ 210 U28058
GCP1 5’ AAGTGTGAGTGTCGAAGGTGG 3’
GCP2 5’GACGACTCGGCTTTTCTACGGC 3’ 373 GLIENKE-BLANCO, 1999
GCF3 5’AAAAAGCCGCCCGACCTACCT3’
GCR7 5’TGTCCGGCGGCCAG3’ 490 BONANTS et.al, 2003
GCN 5’CTGAAAGGTGATGGAAGGGAGG3’
Guignardia citricarpa
GCMR 5’CATTACTTATCGCATTTCGCTGC3’ 300 PERES et. al, 2007
GCF2 5’TAACTTCTATTGAAAGGTTCCAGAGT3’ Guignardia
mangiferae GCR4 5’TCAGGACTTCACAAAATGAATTCTT3’ 210 BONANTS et.al, 2003
ITS3 5’ GCATCGATGAAGAACGCAGC 3’
PINIC1 5’ ATTCAAAAGCCAAGCCACCG 3’ 455 TOOLEY et al.,1997
Pn5B 5’GAACAATGCAACTTATTGGACGTTT3’
Phytophthora
nicotianae
Pn6 5’AACCGAAGCTGCCACCCTAC3’ 120 IPPOLITO et al., 2004
22
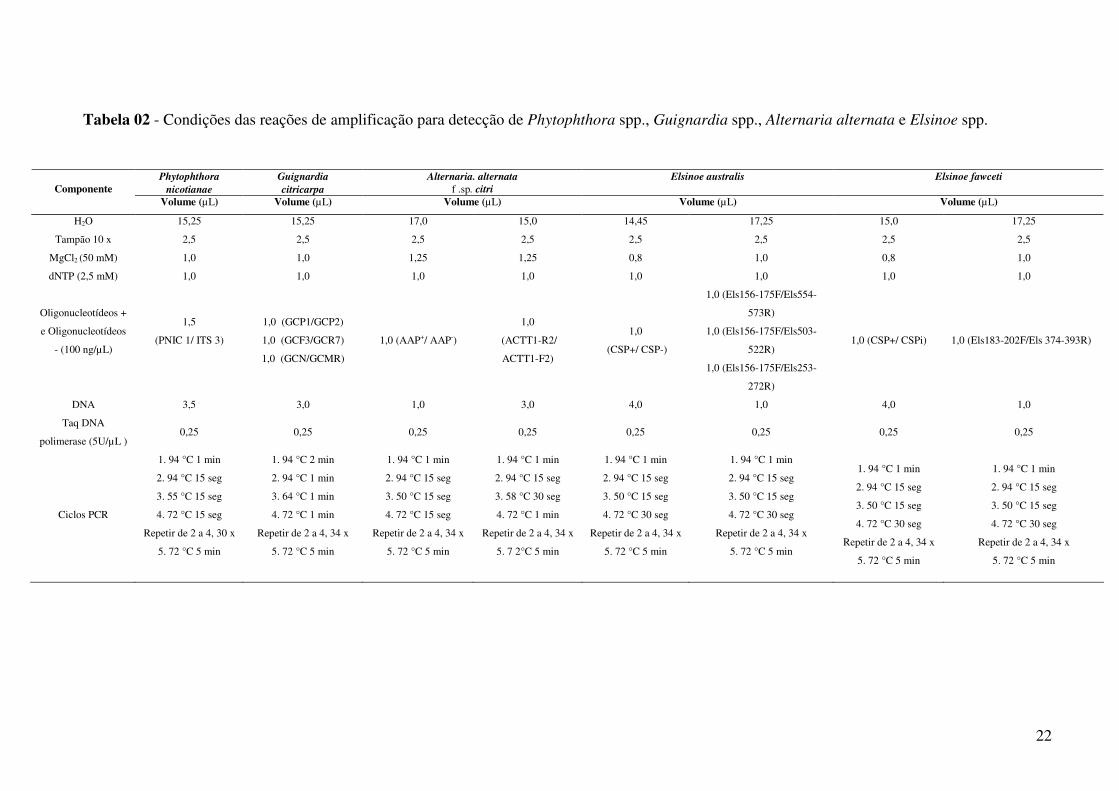
Tabela 02 - Condições das reações de amplificação para detecção de Phytophthora spp., Guignardia spp., Alternaria alternata e Elsinoe spp.
Phytophthora Guignardia Alternaria. alternata Elsinoe australis Elsinoe fawceti
nicotianae citricarpa f .sp. citri Componente Volume (µL) Volume (µL) Volume (µL) Volume (µL) Volume (µL)
H2O 15,25 15,25 17,0 15,0 14,45 17,25 15,0 17,25
Tampão 10 x 2,5 2,5 2,5 2,5 2,5 2,5 2,5 2,5
MgCl2 (50 mM) 1,0 1,0 1,25 1,25 0,8 1,0 0,8 1,0
dNTP (2,5 mM) 1,0 1,0 1,0 1,0 1,0 1,0 1,0 1,0
Oligonucleotídeos +
e Oligonucleotídeos
- (100 ng/µL)
1,5
(PNIC 1/ ITS 3)
1,0 (GCP1/GCP2)
1,0 (GCF3/GCR7)
1,0 (GCN/GCMR)
1,0 (AAP+/ AAP-)
1,0
(ACTT1-R2/
ACTT1-F2)
1,0
(CSP+/ CSP-)
1,0 (Els156-175F/Els554-
573R)
1,0 (Els156-175F/Els503-
522R)
1,0 (Els156-175F/Els253-
272R)
1,0 (CSP+/ CSPi) 1,0 (Els183-202F/Els 374-393R)
DNA 3,5 3,0 1,0 3,0 4,0 1,0 4,0 1,0
Taq DNA
polimerase (5U/µL ) 0,25 0,25 0,25 0,25 0,25 0,25 0,25 0,25
Ciclos PCR
1. 94 °C 1 min
2. 94 °C 15 seg
3. 55 °C 15 seg
4. 72 °C 15 seg
Repetir de 2 a 4, 30 x
5. 72 °C 5 min
1. 94 °C 2 min
2. 94 °C 1 min
3. 64 °C 1 min
4. 72 °C 1 min
Repetir de 2 a 4, 34 x
5. 72 °C 5 min
1. 94 °C 1 min
2. 94 °C 15 seg
3. 50 °C 15 seg
4. 72 °C 15 seg
Repetir de 2 a 4, 34 x
5. 72 °C 5 min
1. 94 °C 1 min
2. 94 °C 15 seg
3. 58 °C 30 seg
4. 72 °C 1 min
Repetir de 2 a 4, 34 x
5. 7 2°C 5 min
1. 94 °C 1 min
2. 94 °C 15 seg
3. 50 °C 15 seg
4. 72 °C 30 seg
Repetir de 2 a 4, 34 x
5. 72 °C 5 min
1. 94 °C 1 min
2. 94 °C 15 seg
3. 50 °C 15 seg
4. 72 °C 30 seg
Repetir de 2 a 4, 34 x
5. 72 °C 5 min
1. 94 °C 1 min
2. 94 °C 15 seg
3. 50 °C 15 seg
4. 72 °C 30 seg
Repetir de 2 a 4, 34 x
5. 72 °C 5 min
1. 94 °C 1 min
2. 94 °C 15 seg
3. 50 °C 15 seg
4. 72 °C 30 seg
Repetir de 2 a 4, 34 x
5. 72 °C 5 min

23
a
b c
d e
Figura 05 - Localização da seqüência dos oligonucleotídeos. a. Alternaria alternata, b. Elsinoe australis, c. Elsinoe fawcetti, d. Guignadia citricarpa e e. Phytophthora
nicotianae. As setas indicam o sentido da amplificação 5’-3’.
3.5 Clonagem, sequenciamento e análise das sequências
Os fragmentos amplificados foram purificados e ligados no vetor pGEM-T
(Promega), de acordo com as informações do fabricante. Células de Escherichia coli
DH5α foram transformadas pelo método da solução de RbCl/CaCl2 (Hanahan, 1983).
As colônias brancas foram repicadas em 200 µL do meio líquido Circle Grow
(Molecular Biology Certified Bacterial Growth Media - QBiogene - Bio 101 Systems -
USA) contendo 8% (v/v) de glicerol e 100 µg/mL de ampicilina, em microplacas de 96
poços, incubadas durante a noite a 37 oC e armazenadas a -80 ºC. Para o
sequenciamento, foi feita a extração de DNA de plasmídeos pelo método boiling (Marra
& Milgroom, 1999). As reações de sequenciamento foram preparadas de acordo com as
instruções da Applied Biosystems para o DNA sequencing Kit Big Dye Terminator
cycle sequencing ready reaction v3 and v3.1. O sequenciamento foi feito no ABI 3730
DNA Analyzers. Alternativamente, o sequenciamento foi feito a partir do produto de
PCR purificado. As seqüências geradas foram avaliadas quanto à qualidade utilizando o
programa Phred-Phrap-Consed e os contigs montados com CAP3 (Gordon, 2004). Para
confirmação dos contigs, inicialmente foi feita análise comparativa através do Blastn
com seqüências depositadas no GenBank (www.ncbi.nlm.nih.gov).

24
3.6 Validação dos testes de detecção dos fungos Alternaria alternata f. sp. citri,
Elsinoe australis, Elsinoe fawcetti, Guignardia citricarpa e Phytophthora nicotianae
3.6.1 Alteraria alternata f. sp. citri
A validação do teste para detecção de A. alternata f. sp. citri foi realizada por
PCR, usando como molde DNA extraído de micélio do isolado IAC 02/06 e DNA total
extraído de diferentes quantidades de lesões de folhas e frutos. Para PCR foi utilizado o
par de oligonucleotídeos ACTT1-F2/ACTT1-R2. Como controle negativo foi utilizado
o isolado IAC 01/06, de A. alternata que não produz a toxina ACT. A determinação do
limite de detecção foi realizada usando como molde DNA extraído de 1 mg a 50 mg de
lesões em folhas, frutos e micélio. Também para avaliação da correção entre peso fresco
e área de lesão foi realizada regressão linear e calculado o coeficiente de determinação.
3.6.2 Elsinoe australis
Para a validação do teste da detecção de E. australis foi utilizado DNA extraído
a partir de micélio e lesões em frutos. O isolado utilizado foi o IAC 04/06 e o controle
negativo utilizado foi o isolado IAC 05/06 de E. fawcetti. Os pares de oligonucleotídeos
utilizados foram: Els156-175F/Els503-522R e Els156-175F/Els554-573R. A
determinação do limite de detecção foi realizada usando como molde DNA extraído de
1 mg a 50 mg de lesões em frutos e micélio. Do mesmo modo, aplicou-se regressão
linear para avaliar a correlação entre peso fresco e área de lesão.
3.6.3 Elsinoe fawcetti
Também se utilizou DNA extraído de micélio, lesões em folhas e frutos na
validação de Elsinoe fawcetti. O isolado utilizado foi o IAC 05/06 e o controle negativo
foi o isolado IAC 04/06 de E. australis. Foi utilizado o par de oligonucleotídeos Els183-
202F/Els374-393R. As mesmas faixas de concentração de DNA total entre 1 mg e 50
mg foram utilizadas para a validação dos pares de oligonucleotídeos.

25
3.6.4 Guignardia citricarpa
A validação do teste para detecção de Guignardia citricarpa foi realizada por
PCR com DNA extraído de micélio do isolado IAC 13/96 e DNA extraído diretamente
de lesões de folhas e frutos. Para avaliação da correção entre peso fresco e área de lesão
foi realizada regressão linear e calculados o coeficiente de determinação. As
informações sobre os pares de oligonucleotídeos utilizados estão na Tabela 01. Foi
utilizado como controle negativo o isolado de G. mangiferae IAC 41/99. Para
determinar o limite de detecção foram preparadas reações contendo DNA extraído de 1
mg a 50 mg, seja de micélio seja de tecido vegetal de folhas e frutos.
3.6.5 Phytophthora nicotianae
A validação do teste para detecção de Phytophthora nicotianae foi realizada
associando-se o teste biológico (teste da isca) ao teste por PCR. Para tanto, foi utilizado
o isolado IAC 01/95. Em copos plástico descartáveis translúcidos foram colocados dois
discos de cultura com micélio de P. nicotianae, de aproximadamente 0,8 cm de
diâmetro e 100 mL de água destilada. Em seguida, foram colocados 50 discos de 0,6 cm
de folhas de lima ácida Tahiti (Citrus latifolia Tanaka). Os copos foram mantidos: i.
fechados com tampa plástica semi-translúcida, ii. fechados com papel alumínio; e iii.
sem tampas, permitindo a total passagem de luz e incubados em BODs com fotoperíodo
de 12 horas claro/escuro a 15 ºC, 20 ºC, 25 ºC, 30 ºC e 35 ºC. Para cada tratamento
foram feitas sete repetições. Após 2, 4, 6, 8 e 10 dias da instalação do experimento, três
discos de folhas de cada repetição foram retirados para montagem das lâminas,
observação e contagem dos esporângios em microscópio óptico.
Para o teste molecular, foi feita a extração de DNA a partir de sete discos de
folhas, uma de cada repetição por tratamento. Para a PCR foi utilizado o par de
oligonucleotídeos ITS3/PINIC1. Como controle negativo, foi utilizado o DNA do
isolado LRS 44/87 de P. citrophthora (Smith & Smith) Leonian. Para determinar o
limite de detecção foram preparadas reações contendo DNA extraído a partir de 1 mg
até 50 mg de micélio.

26
3.6.6 Validação dos testes em frutos de citros infectados com Alternaria alternata f. sp. citri, Elsinoe spp. e Guignardia citricarpa após tratamento em packing house
Também foram realizadas validações para frutos com mancha preta dos citros,
mancha marrom de tangerinas e verrugose de laranja doce e azeda que receberam
tratamento em packing house. Para determinar o limite de detecção foram preparadas
reações contendo DNA extraído de 1 mg a 50 mg de lesões de frutos.
3.7 PCR quantitativo para Phytophthora nicotianae.
3.7.1 Inoculação das plântulas de citros e coleta de material.
O isolado utilizado para a inoculação foi o de Phytophthora nicotianae IAC
01/95, que foi inoculado em frutos de limão Siciliano (Citrus limon (L.) Burm) para
reativar a sua patogenicidade. Para a inoculação os frutos foram lavados em água
corrente e com o auxílo de um palito foi feito um ferimento. Em seguida, colocou-se o
micélio do patógeno no ferimento e cobriu com esparadrapo. Para o reisolamento do
patógeno, após 15 dias, pedaços do albedo do fruto foram retirados com o auxílio de um
bisturi e foi realizada a desinfestação superficial com álcool etílico comercial (96%) por
1 minuto e com hipoclorito de sódio a 2% por 2 minutos. Em seguida, o material foi
colocado em placas de Petri contendo meio de cultura cenoura-ágar, suplementado com
50 µg/mL de rifampicina e 50 µg/mL de ampicilina.
A inoculação foi realizada em plântulas de tangerina Sunki, Poncirus trifoliata e
limão Cravo, inserindo-se palitos colonizados com micélio do oomiceto a 3 cm da base
da planta. O local da inoculação foi coberto com fita adesiva. Cada plântula foi coberta
com um saco plástico para manter a umidade. O material foi colocado em câmara de
germinação com temperatura de 24 °C, durante 6 horas no escuro e 27 °C durante 18
horas no claro. Após 24 horas foram retirados 30 mg da casca de cinco plântulas de
cada variedade em estudo e estes foram armazenados a -20 °C. O material foi coletado
durante 10 dias, a intervalos de 24 horas, com cinco repetições.

27
3.7.2 qPCR
Foi feita a extração de DNA de três repetições de cada material inoculado com
P. nicotianae, coletado após 1, 2, 4, 6, 8 e 10 dias da inoculação. Foi utilizado protocolo
de extração de DNA descrito em 3.3. Em seguida, foi feita a quantificação do DNA em
gel de agarose 1%. A presença do patógeno foi confirmada por PCR, usando dois pares
de oligonucleotídeos, ITS3/PINIC1 e Pn5-B/Pn6. O segundo par de oligonucleotídeos
foi selecionado para ser utilizado em qPCR por amplificar um fragmento menor.
Como controles endógenos foram testados os genes da β - tubulina e da β -
actina, que se mostraram adequados para o trabalho. Optou-se pelo uso do gene da β -
actina no restante da avaliação. Os oligonucleotídeos utilizados foram: F - 5’
TACAACGAGCTTCGTGTTGC 3’ e R - 5’ CTTCTCTCTGTTGGCCTTGG 3’.
A avaliação da eficiência de amplificação foi realizada em reações contendo
diluições seriais de DNA nas concentrações de 100 ng/µL, 10 ng/µL, 1 ng/µL, 0,1 ng/µL
e 0,01 ng/µL, extraído de tecido infectado com P. nicotianae. A eficiência foi verificada
pela relação linear entre o ciclo em que a fluorescência foi detectada acima do limite
mínimo (Ct) e o log da concentração de DNA. Conforme a concentração do DNA
diminui, aumenta o número de ciclos necessários para a detecção da fluorescência (Ct).
Por meio do valor da inclinação (slope) da curva foi calculada a eficiência da
amplificação pela fórmula:
E (eficiência de amplificação) = 10 (-1/slope) -1
Foram preparadas quatro reações de cada ponto de diluição do DNA, compostas
por: 2 µL de DNA (das diluições seriais), 1 µL de cada oligonucleotídeo e 12,5 µL do
SYBR green PCR Master Mix (Applied Biosystems), ajustando o volume final para 25
µL com água Milli-Q. O ciclo utilizado foi: 50 °C por 2 minutos, 95 °C por 10 minutos,
40 vezes 95 °C por 15 segundos, 60 °C por 1 minuto. Foi utilizado o ABI PRISM 7000
SDS versão 1.1. (Applied Biosystems). Uma boa eficiência de amplificação foi
considerada acima de 97% .
Tecido sadio, sem o patógeno, foi utilizado como controle negativo e também
como normalizador. As reações e avaliações foram realizadas no ABI PRISM 7000
SDS versão 1.1 por análises de quantificação relativa. A detecção dos produtos foi

28
medida através da fluorescência emitida pelo marcador SYBR green que se intercala na
dupla fita de DNA.
Para a normalização das amostras foi utilizado o controle endógeno (CE) β-
actina. Os resultados foram normalizados com o valor do Ct obtido do CE, e o Ct foi
definido como o primeiro ciclo de amplificação na qual a fluorescência do produto de
PCR é detectada acima do limite mínimo. A equação utilizada para a normalização foi:
∆Ct = Ct (gene alvo) – Ct (controle endógeno)
O aumento da concentração de DNA do gene alvo para cada condição foi
calculado por meio da fórmula:
∆∆Ct = ∆Ct da amostra - ∆Ct do calibrador
O calibrador é o valor de Ct obtido para uma amostra específica; o aumento dos
níveis de expressão é obtido sempre em relação ao calibrador específico utilizado.
Para todas as reações de qPCR foi feita a curva de dissociação para a verificação
de amplificações inespecíficas, que podem ocorrer devido a contaminações.
4 RESULTADOS E DISCUSSÃO
4.1 Isolamento e manutenção dos fungos Alternaria alternata f. sp. citri, Elsinoe
australis e Elsinoe fawcetti
Os isolados de Alternaria alternata f. sp. citri associados a citros foram
denominados IAC 01/06, IAC 02/06 e IAC 03/06, foram originados de lesões da
mancha marrom em folhas e frutos de tangor Murcott e de lesão em caule de limão
Cravo, respectivamente (Figura 06 a, b). Os isolamentos não apresentaram dificuldades;
foi feita a desinfestação superficial e uso de meio semi-seletivo. Independente da
origem, as colônias dos três isolados não apresentaram diferença na coloração,
apresentaram crescimento rápido e atingiram a borda das placas.
O isolamento de Elsinoe spp. apresentou maior dificuldade. No entanto, foram
obtidos dois isolados desse gênero: IAC 04/06 e IAC 05/06 (Figura 06 c, d). O fungo,

29
nas condições avaliadas, foi isolado somente em meio BDA. Os dois fungicidas
testados, pentacloronitrobenzeno (PCNB) e carbendazim (Derosal), mostraram-se
apropriados, pois inibiram o crescimento de fungos secundários, mas não interferiram
no crescimento de Elsinoe spp. Este fungo desenvolvou-se lentamente em meio de
cultura. Quanto ao aspecto cultural, o isolado de E. australis apresentou colônia de
coloração escura com bordas creme (Figura 06 c); o isolado de E. fawcetti apresentou
coloração creme, ligeiramente róseo (Figura 06 d), à semelhança dos isolamentos
realizados por Fantin (1988). As bordas eram ligeiramente irregulares e recortadas, com
o diâmetro variável ao redor da colônia, que apresentam aspecto estromático.
A manutenção realizada para A. alternata f. sp. citri a cada 20 dias em meio de
cultura cenoura - ágar e para Elsinoe spp. a cada 30 dias em meio de cultura BDA,
juntamente com o armazenamento em BOD a 25°C foram adequados para o
desenvolvimento dos isolados.
a b c d Figura 06 - Aspecto cultural dos fungos isolados de tecidos cítricos com sintomas típicos. a. Alternaria alternata f. sp. citri (IAC 02/06); b. Alternaria alternata sem toxina (IAC 03/06); c. Elsinoe australis (IAC 04/06); d. Elsinoe fawcetti (IAC 05/06).
4.2 Teste de patogenicidade de Alternaria alternata
A técnica de inoculação em folhas de ramos destacados foi satisfatória para a
comprovação da patogenicidade e identificação dos isolados de Alternaria alternata
(Tabela 03 e Figura 07), como reportado por Aguilar-Vildoso et al. (2004). Após 72
horas da inoculação, os dois isolados obtidos de lesões corticosas da mancha marrom
provocaram lesões somente nas folhas de tangor Murcott e não nas de laranja Pêra, o
que confirmou ser A. alternata f. sp. citri patótipo tangerina (Figura 07 a). Este
comportamento de patogenicidade diferencial entre os isolados originados da mancha

30
marrom e do isolado originado da podridão negra era esperado para poder discriminar
entre A. alternata e A. alternata f. sp. citri. As lesões ocorreram devido à produção da
toxina ACT pelo fungo isolado de folhas (IAC 01/06) e frutos (IAC 02/06).
Nas folhas de ramos destacados inoculados com o isolado de Alternaria
alternata que não produz toxina (IAC 03/06), não houve desenvolvimento de lesões em
nenhuma das duas variedades avaliadas (Figura 07 b). Este comportamento confirma
que este último isolado de A. alternata é avirulento a tecidos foliares intactos, não
produz a toxina ACT, e para colonizar necessita de lesões para penetrar nos tecidos
(Thomma, 2003).
O teste de patogenicidade também pode ser utilizado como um método de
diagnóstico na detecção de A. alternata f. sp. citri (Akimitsu et al., 2003), uma vez que
este ensaio não requer laboratório com alta infraestrutura, podendo ser realizado
facilmente. Porém, para a utilização deste método é necessário, antes, realizar o
isolamento do patógeno em meio de cultura. Como leva alguns dias para o crescimento,
torna-se um método de detecção mais demorado.
Figura 07 - Aspecto das folhas de tangor Murcott inoculadas com a suspensão de esporos de Alternaria alternata associados a citros após 72 horas em câmara úmida. a. lesão causada pelo isolado IAC 02/06; b. nenhuma lesão provocada pelo isolado IAC 03/06.
a b
31
Tabela 03 - Formação de lesões em folhas de brotações de tangor Murcott e laranja Pêra, após 72 horas de inoculação com suspensão de esporos de colônias de Alternaria sp. associadas a citros.
Formação de lesões em
folhas
Isolado de
Alternaria sp.
Origem Sintoma Tangor
Murcott
Laranja
Pêra
IAC01/06 Tangerina Poncan – folha Mancha marrom + -
IAC02/06 Tangerina Poncan – fruto Mancha marrom + -
IAC03/06 Limão Cravo- caule Podridão Negra - -
4.3 Seleção de oligonucleotídeos para amplificação dos fungos Alternaria alternata
f. sp. citri, Elsinoe australis, Elsinoe fawcetti, Guignardia citricarpa e Phytophthora
nicotianae
4.3.1 Alternaria alternata f. sp. citri
O par de oligonucleotídeos AAP+/AAP- não diferenciou os isolados de
Alternaria, enquanto que o ACTT1-F2/ACTT1-R2 diferenciou A. alternata de A.
alternata f. sp. citri patótipo tangerina. Este par de oligonucleotídeos também
diferenciou a Alternaria alternata f. sp. citri patótipo de limão da A. alternata f. sp. citri
patótipo tangerina (Figura 08 b). O par AAP+/AAP- amplificou um fragmento de
aproximadamente 571 pares de bases, e foi inespecífico para a diferenciação das
espécies ditas saprófitas ou oportunistas da patogênica e produtora da toxina ACT
(Figura 08 a).
O par de oligonucleotídeos ACTT1-F2/ACTT1-R2, específico da região do gene
que codifica a toxina ACT liberada pelo fungo, amplificou um fragmento de 781 pares
de bases apenas quando se utilizou DNA de A. alternata f. sp. citri patótipo de
tangerina, mostrando ser específico (Figura 08 b).
No caso do par AAP+/AAP- a baixa especificidade explica-se por amplificarem
DNA da região ribossomal, útil no estudo de sistemática de fungos, mas com baixo
nível de variabilidade intraespecífica. Portanto, no caso de Alternaria o diagnóstico
baseou-se na amplificação de uma região responsável pela produção da toxina

32
responsável pela patogenicidade. Assim, para patógenos, em que as toxinas são
determinantes na patogênese ou na diferenciação de hospedeiros, deve ser realizado o
desenho dos oligonucleotídeos no gene ou nos genes relacionados com a via biosintética
da toxina em particular.
a b
Figura 08 - Padrão eletroforético das reações de amplificação para a detecção de Alternaria alternata f. sp. citri com DNA extraído de micélio com os pares de oligonucleotídeos. a. par de oligonucleotídeos AAP+/AAP-e b. par de oligonucleotídeos ACTT1-F2/ACTT1-R2. 1. Alternaria alternata f. sp. citri, 2 Alternaria alternata f. sp. citri patótipo limão, 3. Alternaria alternata f. sp. citri patótipo tangerina. M. 1Kb Plus DNA Ladder.
4.3.2 Elsinoe australis
O par de oligonucleotídeos CSP+/CSP- que amplificou um fragmento de 636
pares de bases mostrou ser inespecífico, pois amplificou um fragmento quando DNA de
E. fawcetti foi utilizado como molde. Os pares de oligonucleotídeos Els156-
175F/Els554-573R (Figura 09 b) e Els156-175F/Els503-522R (Figura 09 c)
amplificaram fragmentos de 418 e 377 pares de bases, respectivamente, sendo estes
oligonucleotídeos específicos para detecção de E. australis. O par de oligonucleotídeos
Els156-175F/Els253-272R não amplificou fragmento, não sendo apropriado para a
detecção desse patógeno.
Com estes resultados, dois pares de oligonucleotídeos mostraram-se específicos
para a detecção de E. australis: Els156-175F/Els554-573R e Els156-175F/Els503-522R.
Na comparação realizada com o par de oligonucleotídeo Els156-175F e Els503-522R no
850
M 1 2 3 pb
650
100 100
850
650
M 1 2 3 pb

33
Genebank foi comprovada a homologia de seqüências de Elsinoe, depositadas no banco
de dados, com e-value de 0,033 e 100% de máxima identificação (dados não
mostrados).
a b c Figura 09 - Padrão eletroforético das reações de amplificação para a detecção de Elsinoe australis a partir de DNA extraído de micélio e os pares de oligonucleotídeos: a. CSP+/CSP-: 1. controle negativo (Guignardia citricarpa), 2 e 3. Elsinoe fawcetti (IAC 05/06), 4 e 5. Elsinoe australis (IAC 04/06), b. Els156-175F/Els554-573R e c. Els156-175F/Els503-522R. 1 e 2. controle negativo Elsinoe fawcetti (IAC 05/06), 3 e 4. Elsinoe
australis (IAC 04/06) e M. 1 Kb Plus DNA Ladder.
4.3.3 Elsinoe fawcetti
Inicialmente foi utilizado o par de oligonucleotídeos CSP+/CSPi (Figura 10 a),
que amplificou um fragmento de 525 pares de bases. No entanto, esse par de
oligonucleotídeos também amplificou um fragmento do tamanho esperado quando foi
usado DNA de Guignardia citricarpa como molde, mostrando ser inespecífico para a
detecção e diferenciação deste fungo. O par de oligonucleotídeos Els183-202F/Els374-
393R (Figura 10 b), que amplificou um fragmento de 210 pares de bases, foi específico,
podendo ser utilizado para a detecção de E. fawcetti.
A análise in silico dos pares de oligonucleotídeos Els183-202F/Els374-393R
apresentou e-value de 0,033 e 100% de máxima identificação (dados não mostrados),
comprovando assim a sua especificidade na diferenciação das duas espécies de Elsinoe
avaliadas nesse estudo.
M 1 2 3 4 5 pb
100
650
850
M 1 2 3 4 pb
100
650
850
M 1 2 3 4 pb
100
650
850

34
a b
Figura 10 - Padrão eletroforético das reações de amplificação para a detecção de Elsinoe fawcetti com DNA extraído de micélio e os oligonucleotídeos: a. CSP+/CSPi: 1. controle negativo (Guignardia citricarpa), 2 e 3. Elsinoe australis (IAC 04/06), 4 e 5. Elsinoe fawcetti (IAC 05/06), b. Els183-202F/Els374-393R. 1 e 2. controle negativo, Elsinoe australis (IAC 04/06), 3 e 4. Elsinoe fawcetti (IAC 05/06). M. 1 Kb Plus DNA Ladder.
4.3.4 Guignardia citricarpa
Todos os oligonucleotídeos avaliados amplificaram fragmento de tamanho
esperado quando o DNA do isolado patogênico IAC 13/96 de G. citricarpa foi utilizado
como molde (Figura 11). Foram amplificados fragmentos de tamanho esperado, sendo
373 pb para o par GCP1/GCP2 (Figura 11 b) (Glienke-Blanco, 1999), 470 pb para
GCF3/GCR7 (Figura 11 c) (Bonants et al., 2003), 300 pb para GCN/GCMR (Figura 11
d) (Peres et al., 2007).
Neste teste inicial também foi utilizado o par de oligonucleotídeos GCF2/GCR4
(Figura 11 a) que amplificou um fragmento de 210 pb (Bonants et al., 2003) quando
DNA do controle negativo foi utilizado como molde (G. mangiferae) (Figura 11 a). Os
três pares de oligonucleotídeos específicos de G. citricarpa não amplificaram
fragmentos quando DNA de G. mangiferae foi utilizado como molde. A importância
desse diagnóstico está no fato da espécie endofítica apresentar ampla ocorrência em
plantas de citros e, desse modo, poder ser isolada e identificada erroneamente como G.
citricarpa. A identificação precisa e segura de G. citricarpa é extremamente importante,
uma vez que a mancha preta dos citros é uma praga quarentenária A1 em vários países.
No Brasil há regiões isentas e que são exportadoras de frutos cítricos, como exemplo
100
650
300
M 1 2 3 4 5 pb
650
300
100
M 1 2 3 4 pb
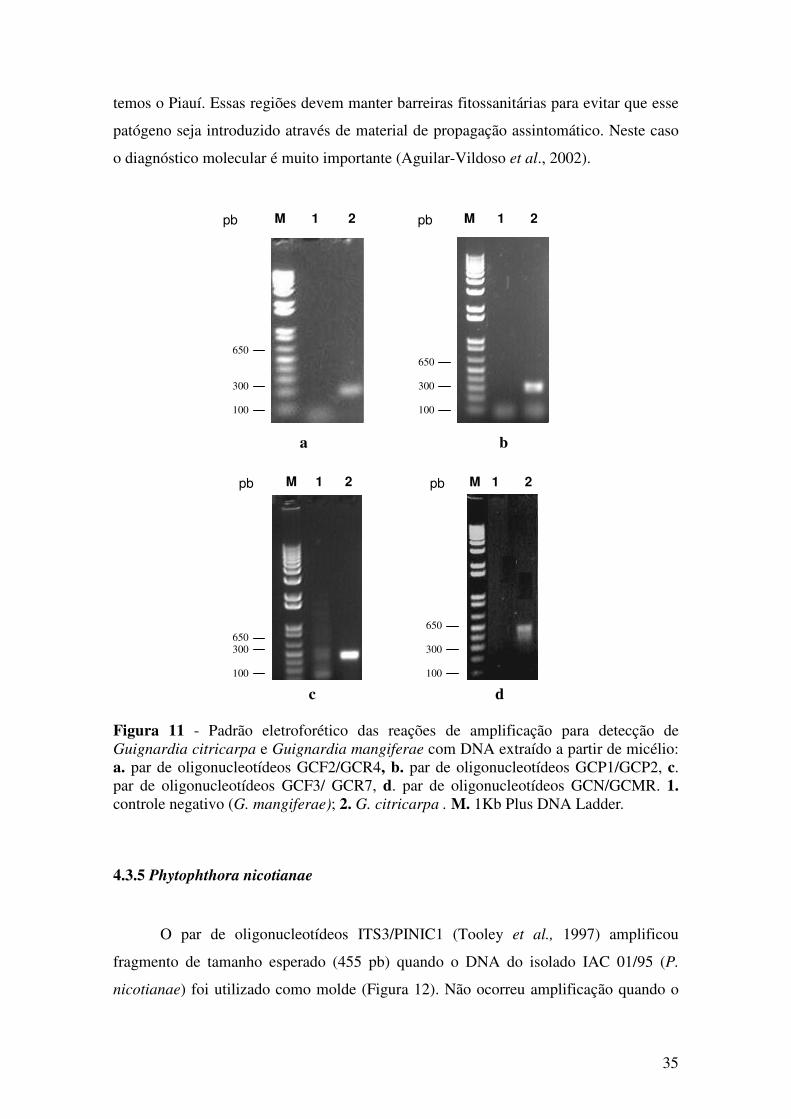
35
temos o Piauí. Essas regiões devem manter barreiras fitossanitárias para evitar que esse
patógeno seja introduzido através de material de propagação assintomático. Neste caso
o diagnóstico molecular é muito importante (Aguilar-Vildoso et al., 2002).
a b
c d Figura 11 - Padrão eletroforético das reações de amplificação para detecção de Guignardia citricarpa e Guignardia mangiferae com DNA extraído a partir de micélio: a. par de oligonucleotídeos GCF2/GCR4, b. par de oligonucleotídeos GCP1/GCP2, c. par de oligonucleotídeos GCF3/ GCR7, d. par de oligonucleotídeos GCN/GCMR. 1. controle negativo (G. mangiferae); 2. G. citricarpa . M. 1Kb Plus DNA Ladder.
4.3.5 Phytophthora nicotianae
O par de oligonucleotídeos ITS3/PINIC1 (Tooley et al., 1997) amplificou
fragmento de tamanho esperado (455 pb) quando o DNA do isolado IAC 01/95 (P.
nicotianae) foi utilizado como molde (Figura 12). Não ocorreu amplificação quando o
M 1 2 pb
100
300
650
M 1 2 pb
100
300
650
M 1 2 pb
100
650 300
M 1 2 pb
100
300
650

36
DNA de P. citrophthora (LRS 44/87) foi utilizado como molde (Figura 12). Assim, esse
par de oligonucleotídeos mostrou-se ser específico para diferenciação entre as espécies
de Phytophthora predominantes no Brasil. Deve ser destacado, no entanto, que se P.
palmivora for detectada no Brasil, este tipo de diagnóstico não pode ser empregado sem
validação prévia, ou terão que ser desenvolvidos outros oligonucleotídeos para essa
espécie.
Figura 12 - Padrão eletroforético dos produtos das reações de amplificação com os oligonucleotídeos ITS3/PINIC1, a partir de DNA extraído de micélio: 1. de Phytophthora citrophthora (LRS 44/87); 2 e 3. Phytophthora nicotianae (IAC 01/95), M. 1Kb DNA Ladder plus.
4.3.6 Sequenciamento dos produtos da amplificação
O Anexo 1 apresenta as seqüências dos fragmentos amplificados de cada
patógeno. Cada seqüência obtida foi analisada através do Blastn do banco de dados do
NCBI (www.ncbi.nlm.nih.gov). O par de oligonucleotídeos GCP1/GCP2 utilizado na
detecção de Guignardia citricarpa, por se tratar de um SCAR (Sequence Characterized
Amplified Regions), foi confirmado através da seqüência original descrita por Glienke-
Blanco (1999). Com esta análise foi confirmado que o fragmento amplificado
correspondia à seqüência do DNA do patógeno. Os valores de e-value e a máxima
identificação encontrados para cada fragmento estão descritos na Tabela 04.
M 1 2 3 pb
100
650
300

37
Tabela 04 - Resultados das análises realizadas in silico para a confirmação do fragmento de DNA correspondente à seqüência dos patógenos de citros.
Patógeno Oligonucleotídeos E-value Identidade máxima
Nr. Do Acesso
Alternaria alternata ACTT1-F2/ACTT1-R2 0,0 98% AB034586.1 Els 156-175F / Els 554-573R 7e-129 92% U28057.1 Elsinoe australis Els 156-175F / Els 503-522R 3e-142 95% U28057.1
Elsinoe fawcetti Els183-202F/ Els374-393R 3e-75 98% U28057.1 GCF3/GCR7 1e-162 90% AF374371.1 Guignardia citricarpa
GCMR/GCN 5e-122 99% AF374371.1 Phytophthora
nicotianae
ITS-3 / PINIC-1 0,0 99% EF426559.1
4.4 Validação dos testes e determinação do limite de detecção de patógenos de citros.
4.4.1 Alternaria alternata f. sp. citri
Foram amplificados fragmentos de tamanho esperado quando se utilizou DNA
extraído de 20 mg a 50 mg de micélio, de DNA extraído de 1 mg a 50 mg de lesões de
frutos e de DNA extraído de 45 mg e 50 mg de folhas com o par de oligonucleotídeos
ACTT1-F2/ACTT-R2 (Figura 13 a, b, c).
Na regressão linear entre o peso das lesões (mg) e a área das lesões (cm2) foi
obtido o coeficiente de 0,0568 mg/cm2 de lesão (R2 = 0,964) e 0,0701 mg/cm2 de lesão
(R2 = 0,9864) para as lesões de frutos e de folhas, respectivamente (Tabela 05).
Na validação realizada com o par de iniciadores ACTT1-F2/ACTT-R2, foi
possível detectar e diferenciar a A. alternata f. sp. citri causadora da mancha marrom
das outras espécies de A. alternata que afetam citros. Este par de oligonucleotídeos
mostrou ser sensível a até 1 mg de tecido lesionado em frutos, o equivalente a uma lesão
de aproximadamente 0,0491 cm2 (Tabela 05). Entretanto, para DNA extraído de folhas
este limite foi a partir de 45 mg, o equivalente a uma área de 3,0628 cm2 (Tabela 05).
De acordo com Masunaka et al. (2000) este par de oligonucleotídeos é baseado
no gene AKT1 e AKT2, genes estes envolvidos na biosíntese da toxina ACT. Em seu
trabalho o autor conseguiu diferenciar as espécies de Alternaria que afetam citros,
diferenciando principalmente a A. alternata f. sp. citri que afeta tangerinas da A.

38
alternata f. sp. citri que afeta limões, sendo que estas duas espécies produzem uma
toxina responsável pela sua patogênese.
a
b
c
Figura 13 - Padrão eletroforético de reações de amplificação para detecção de Alternaria alternata f. sp. citri com o par de oligonucleotídeos ACTT1-F2/ACTT1-R2. a. DNA extraído a partir de micélio. 1. controle negativo (A. alternata), 2. 1 mg, 3. 5 mg, 4. 10 mg, 5. 15 mg, 6. 20 mg, 7. 25 mg, 8. 30 mg, 9. 35 mg, 10. 40 mg, 11. 45 mg e 12. 50 mg. b. DNA extraído a partir de lesões em frutos. c. DNA extraído a partir de lesões em folhas: 1. 1 mg, 2. 5 mg, 3. 10 mg, 4. 15 mg, 5. 20 mg, 6. 25 mg, 7. 30 mg, 8. 35 mg, 9. 40 mg, 10. 45 mg e 11. 50 mg, M. 1Kb Plus DNA Ladder.
300
100
M 1 2 3 4 5 6 7 8 9 10 11 12 pb
650
M 1 2 3 4 5 6 7 8 9 10 11 pb
100
300
650
M 1 2 3 4 5 6 7 8 9 10 11 pb
100
300
650

39
Tabela 05 - Correspondência do peso fresco (mg) e área (cm2) de lesões de frutos e folhas com mancha marrom da tangor Murcott, utilizados para a validação do par de oligonucleotídeos para diagnóstico de Alternaria alternata f.sp. citri.
Peso fresco (mg)
Área da lesão de fruto (cm2)
Área da lesão de folha (cm2)
1 0,0491 0,1257 5 0,1197 0,5145
10 0,3317 0,9053 15 0,6104 1,1978 20 0,8478 1,5531 25 1,1325 1,8751 30 1,8877 2,1421 35 2,1862 2,3973 40 2,3845 2,7487 45 2,5768 3,0628 50 2,8752 3,4261
Neste trabalho, foram confirmados os resultados de Masunaka et al. (2000),
conseguindo diferenciar as três espécies de A. alternata: Alternaria alternata f. sp. citri
que afeta tangerina, responsável pela toxina ACT, Alternaria alternata f. sp. citri que
afeta limões, responsável pela toxina ACRL ou ACR e a Alternaria alternata, que não
produz toxina. Entretanto, além de diferenciar as espécies de A. alternata também foi
verificada a sensibilidade do par de iniciadores ACTT1-F2/ACTT1-R2, quando
utilizado frutos de tangor Murcott colonizado pelo patógeno, sendo o limite a partir de
uma lesão.
Porém, para se obter estes limites de detecção, foram realizadas modificações no
método de extração, devido à ação da toxina, que necrosa o tecido. Para solucionar esta
interferência no método de extração foram utilizadas pérolas de vidro, os quais
conseguem eliminar os interferentes na reação, uma vez, que o vidro tem afinidade com
o DNA.
Em outros trabalhos realizados com marcadores moleculares também foi
possível à diferenciação das espécies de A. alternata que afetam citros. Peever et al.
(1999) e Peever et al., (2002) conseguiram diferenciar as espécies de Alternaria que
afetam citros, porém em seus trabalhos as análises foram realizadas por RAPD. Pryor et
al. (2003) também realizaram trabalhos para diferenciarem isolados de Alternaria, com
o uso de marcadores de microssatélites.
A utilização de marcadores moleculares para diferenciar as espécies de
Alternaria é uma boa alternativa quando se pretende realizar estudos em nível de taxon,

40
entretanto em análises de rotina, este método torna-se inviável, uma vez que é mais
trabalhoso, sendo necessário um tempo maior de execução, além de o custo ser mais
elevado e nem sempre o laboratório possuir infraestrutura para a realização desta
técnica.
Com os resultados obtidos neste trabalho observou-se que a utilização da técnica
de PCR para ser utilizado em análises de rotina permite uma maior rapidez e segurança,
uma vez que o método atual realizado pelo isolamento do patógeno requer um tempo
maior na sua execução e necessita de um profissional qualificado para realizar a
caracterização morfológica, uma vez que esta diferenciação baseia-se no tamanho do
esporo e outras características do fungo. Com o uso do par de oligonucleotídeos
ACTT1-F2/ACTT1-R2 e do protocolo de extração utilizando pérolas de vidro é possível
realizar diagnóstico de rotina com maior rapidez e segurança.
4.4.2 Elsinoe australis
A validação do teste para detecção de Elsinoe australis foi feita por PCR com os
pares de oligonucleotídeos Els156-175F/Els503-522R e Els156-175F/Els554-573R e
DNA extraído de micélio e lesões em frutos (Figura 14 e 15). Os dois pares de
oligonucleotídeos amplificaram os fragmentos do tamanho esperado, seja com DNA
extraído de micélio seja com DNA total extraído de lesões (Figura 14 a e b e Figura 15 a
e b). Com a regressão linear realizada entre o peso das lesões (mg) e a área das lesões
(cm2) obteve- se o coeficiente de 0, 0509 mg/cm2 de lesão (R2 = 0,9981) (Tabela 06).
Com estes resultados foi possível comprovar a especificidade dos pares de
oligonucleotídeos para serem utilizados em diagnóstico de rotina para E. australis,
causadora da verrugose da laranja doce.
A grande vantagem de se utilizar a PCR, particularmente para esse fungo, é a
versatilidade do método face às dificuldades de isolamento em meio seletivo.

41
Tabela 06 - Correspondência entre o peso fresco (mg) e área (cm2) de lesões de laranjas com verrugose utilizados para a validação dos oligonucleotídeos para diagnósticos de Elsinoe australis.
Peso fresco (mg)
Área da lesão (cm2)
1 0,0707 5 0,2376
10 0,5301 15 0,7519 20 0,9895 25 1,2605 30 1,4981 35 1,7455 40 1,9850 45 2,3482 50 2,6034
a
b Figura 14 - Padrão eletroforético de reações de amplificação para detecção de Elsinoe
australis com o par de oligonucleotídeos Els156-175F/Els 554-573R. a. DNA extraído a partir de micélio. 1. controle negativo (Elsinoe fawcetti), 2. 1 mg, 3. 5 mg, 4. 10 mg, 5. 15 mg, 6. 20 mg, 7. 25 mg, 8. 30 mg, 9. 35 mg, 10. 40 mg, 11. 45 mg e 12. 50 mg. b. DNA extraído a partir de lesões em frutos: 1. 1 mg, 2. 5 mg, 3. 10 mg, 4. 15 mg, 5. 20 mg, 6. 25 mg, 7. 30 mg, 8. 35 mg, 9. 40 mg, 10. 45 mg e 11. 50 mg, M. 1Kb Plus DNA Ladder.
650
M 1 2 3 4 5 6 7 8 9 10 11 12 pb
300
100
650
M 1 2 3 4 5 6 7 8 9 10 11 pb
100
300

42
a
b
Figura 15 - Padrão eletroforético de reações de amplificação para detecção de E.
australis com o par de oligonucleotídeos Els156-175F/Els503-522R. a. DNA extraído a partir de micélio. 1. controle negativo (E. fawcetti IAC 05/06), 2. 1 mg, 3. 5 mg, 4. 10 mg, 5. 15 mg, 6. 20 mg, 7. 25 mg, 8. 30 mg, 9. 35 mg, 10. 40 mg, 11. 45 mg e 12. 50 mg. b. DNA extraído a partir de lesões em frutos: 1. 1 mg, 2. 5 mg, 3. 10 mg, 4. 15 mg, 5. 20 mg, 6. 25 mg, 7. 30 mg, 8. 35 mg, 9. 40 mg, 10. 45 mg e 11. 50 mg, M. 1Kb plus DNA Ladder.
4.4.3 Elsinoe fawcetti
A validação do teste para a detecção de Elsinoe fawcetti por PCR foi feita com o
par de oligonucleotídeos Els183-202F/Els374-393R, que amplificou fragmento de
tamanho esperado quando se utilizou DNA extraído de 1 mg a 50 mg de micélio como
molde e DNA extraído de 1 mg a 50 mg de lesões em frutos e folhas de limão Cravo
(Figura 16 a, b, c, Tabela 07). A regressão linear foi realizada entre o peso das lesões
(mg) e a área das lesões (cm2), obtendo-se o coeficiente de 0,0482 mg/cm2 de lesão
(R2 = 0,9917) para as lesões retiradas de frutos; para as lesões retiradas de folhas o
coeficiente foi de 0,0485 mg/cm2 de lesão (R2 = 0,999).
650
100
300
M 1 2 3 4 5 6 7 8 9 10 11 pb
M 1 2 3 4 5 6 7 8 9 10 11 12 pb
650
300
100

43
a
b
b
c
Figura 16- Padrão eletroforético de reações de amplificação para detecção de Elsinoe
fawcetti com o par de oligonucleotídeos Els183-202F/Els374-393R. a. DNA extraído a partir de micélio: 1. controle negativo (Elsinoe australis), 2. 1 mg, 3. 5 mg, 4. 10 mg, 5. 15 mg, 6. 20 mg, 7. 25 mg, 8. 30 mg, 9. 35 mg, 10. 40 mg, 11. 45 mg e 12. 50 mg. b. DNA extraído a partir de lesões em frutos e c. DNA extraído a partir de lesões em folhas: 1. 1 mg, 2. 5 mg, 3. 10 mg, 4. 15 mg, 5. 20 mg, 6. 25 mg, 7. 30 mg, 8. 35 mg, 9. 40 mg, 10. 45 mg e 11. 50 mg, M. 1Kb Plus DNA Ladder.
650
100
300
M 1 2 3 4 5 6 7 8 9 10 11 12 pb
650
100
300
M 1 2 3 4 5 6 7 8 9 10 11 pb
650
100
300
M 1 2 3 4 5 6 7 8 9 10 11 pb

44
Tabela 07 - Correspondência entre o peso fresco (mg) e área (cm2) de lesões de frutos e folhas com verrugose de limão Cravo utilizados para a validação dos oligonucleotídeos para diagnóstico de Elsinoe fawcetti.
Peso fresco (mg)
Área da lesão de fruto (cm2)
Área da lesão de folha (cm2)
1 0,0707 0,0707 5 0,1492 0,2376
10 0,4438 0,4948 15 0,6362 0,6931 20 0,8659 0,9523 25 1,1173 1,1781 30 1,3470 1,4314 35 1,7102 1,6769 40 1,9732 1,9557 45 2,2579 2,2188 50 2,4621 2,4466
Com esses resultados, foi verificada a sensibilidade do teste para detecção do
patógeno usando DNA extraído direto de lesões em frutos, além de diferenciar as duas
espécies de verrugose de citros. Com esta validação, foi possível detectar o fungo em
lesões de frutos com uma área de 0,0707 cm2. A partir destes dados pode-se observar a
eficiência e sensibilidade dos oligonucleotídeos para serem usados na PCR como
diagnóstico de rotina, uma vez que o método rotineiramente utilizado baseia-se no
isolamento e caracterização morfológica do patógeno. Entretanto, o isolamento deste
fungo é trabalhoso e nem sempre é feito com sucesso, pois para tanto é necessário
utilizar lesões no início da infecção. Além do mais esse fungo apresenta crescimento
lento, o que propicia o desenvolvimento de outros microrganismos no meio de cultura.
Timmer et al. (1996) e Tan et al. (1996) descrevem a dificuldade de diferenciar
as duas espécies de Elsinoe, uma vez que E. australis possui conídio hialino e seu
tamanho é inferior ao de E. fawcetti, sendo esta característica aparentemente utilizada
em vários trabalhos para diferenciar estas duas espécies. Entretanto, em seu trabalho o
autor não conseguiu observar esta diferença entre os conídios, mas foi possível
diferenciar as duas espécies através da coloração da cultura. Segundo Timmer et al.
(1996) análises de restrinção do espaço interno transcrito (ITS) do DNA ribossomal
indicam que E. fawcetti e E. australis pertencem a grupos diferentes, sendo possível a
amplificação por PCR dos segmentos da região ITS, diferenciando as duas espécies
deste fungo que afetam citros. Em estudos mais recentes Hyun et al. (2001), Hyun et
al.(2007) utilizando pares de oligonucleotídeos baseados em seqüência de regiões ITS,

45
conseguiram diferenciar estas duas espécies utilizando vários tipos de isolados e lesões
de frutos. São vários os trabalhos realizados para caracterizar e diferenciar E. australis e
E. fawcetti. Estes trabalhos nos comprovam que estes dois isolados pertecem a grupos
diferentes, entretanto, ainda não se sabe se existe diferença entre patógenos do mesmo
grupo de diferentes regiões (Hyun et al., 2001).
A importância de um diagnóstico sensível e rápido pode ser avaliada pela
importância dada pela European and Mediterranean Plant Protection Organization
(EPPO) na sexta reunião da EPPO Diagnostic for regulated pests (fungi) realizada em
2006 na Holanda. Neste evento foi expresso o interesse de desenvolvimento de
protocolos para detecção de quatro fungos em especial, entre os quais E. fawcetti e E.
australis (EPPO, 2008).
4.4.4 Guignardia citricarpa
O par de oligonucleotídeos GCP1/GCP2 amplificou fragmentos de tamanho
esperado tanto quando foi utilizado DNA extraído de micélio, como DNA extraído de
lesões em frutos e folhas (Figura 17 a, b, c). O par de oligonucleotídeos GCF3/GCR7
amplificou fragmentos de tamanho esperado quando se utilizou em PCR DNA extraído
de 25 mg a 40 mg de micélio; de 1 mg a 50 mg de lesões de frutos; e de 40 mg a 50 mg
de lesões em folhas (Figura 18 a, b, c). Com o par de oligonucleotídeos GCN/GCMR,
foram obtidos fragmentos de tamanho esperado quando se utilizou DNA extraído de 1
mg a 50 mg de micélio, de 1 mg a 50 mg de lesões de frutos e de 1 mg a 30 mg para
lesões de folhas (Figura 19 a, b, c).
Ao realizar a regressão linear passando pela origem entre o peso (mg) e a área
das lesões (cm2) obteve-se o coeficiente de 0,010 mg/cm2 de lesão (R2 = 0,9946), para
as lesões retiradas de frutos, enquanto que para folhas o coeficiente foi de 0,0494
mg/cm2 de lesão (R2 = 0,9897) (Tabela 08).
Na validação com os três pares de oligonucleotídeos GCP1/GCP2, GCF3/GCR7
e GCN/GCMR foi possível a detecção do patógeno e a diferenciação entre as duas
espécies de Guignardia spp. (G. citricarpa e G. mangiferae). No entanto, com o par de
oligonucleotídeos GCP1/GCP2 e GCMR/GCN, o limite de detecção na PCR foi de
DNA total extraído de 1 mg a 50 mg de micélio e de lesões de frutos.

46
a
b
c
Figura 17 - Padrão eletroforético de reações de amplificação para detecção de Guignardia citricarpa com o par de oligonucleotídoes GCP1/GCP2. a. DNA extraído a partir de micélio: 1. controle negativo (Guignardia mangiferae), 2. 1 mg, 3. 5 mg, 4. 10 mg, 5. 15 mg, 6. 20 mg, 7. 25 mg, 8. 30 mg, 9. 35 mg, 10. 40 mg, 11. 45 mg e 12. 50 mg. b. DNA extraído a partir de lesões de frutos e c. DNA extraído a partir de lesões em folhas: 1. 1 mg, 2. 5 mg, 3. 10 mg, 4. 15 mg, 5. 20 mg, 6. 25 mg, 7. 30 mg, 8. 35 mg, 9. 40 mg, 10. 45 mg e 11. 50 mg, M. 1Kb Plus DNA Ladder.
M 1 2 3 4 5 6 7 8 9 10 11 12 pb
650
300
100
M 1 2 3 4 5 6 7 8 9 10 11 pb
650
300
100
M 1 2 3 4 5 6 7 8 9 10 11 pb
650
300
100

47
a
b
c
Figura 18 - Padrão eletroforético de reações de amplificação para detecção de Guignardia citricarpa com o par de oligonucleotídeos GCF3/GCR7. a. DNA extraído a partir de micélio: 1. controle negativo (Guignardia mangiferae), 2. 1 mg, 3. 5 mg, 4. 10 mg, 5. 15 mg, 6. 20 mg, 7. 25 mg, 8. 30 mg, 9. 35 mg, 10. 40 mg, 11. 45 mg e 12. 50 mg. b. DNA extraído a partir de lesões de frutos e c. DNA extraído a partir de lesões em folhas: 1.1 mg, 2. 5 mg, 3. 10 mg, 4. 15 mg, 5. 20 mg, 6. 25 mg, 7. 30 mg, 8. 35 mg, 9. 40 mg, 10. 45 mg e 11. 50 mg, M. 1Kb Plus DNA Ladder.
M 1 2 3 4 5 6 7 8 9 10 11 12 pb
650
300
100
M 1 2 3 4 5 6 7 8 9 10 11 pb
650
300
100
M 1 2 3 4 5 6 7 8 9 10 11 pb
650
300
100

48
a
b
b
c
Figura 19 - Padrão eletroforético de reações de amplificação para detecção de Guignardia citricarpa com o par de oligonucleotídeosGCN/GCMR. a. DNA extraído a partir de micélio: 1. controle negativo (Guignardia mangiferae ), 2. 1 mg, 3. 5 mg, 4. 10 mg, 5. 15 mg, 6. 20 mg, 7. 25 mg, 8. 30 mg, 9. 35 mg, 10. 40 mg, 11. 45 mg e 12. 50 mg, b. DNA extraído a partir de lesões em frutos e c. DNA extraído a partir de lesões em folhas. 1.1 mg, 2. 5 mg, 3. 10 mg, 4. 15 mg, 5. 20 mg, 6. 25 mg, 7. 30 mg, 8. 35 mg, 9. 40 mg, 10. 45 mg e 11. 50 mg, M 1Kb Plus DNA Ladder.
M 1 2 3 4 5 6 7 8 9 10 11 pb
100
650
300
M 1 2 3 4 5 6 7 8 9 10 11 pb
650
300
100
M 1 2 3 4 5 6 7 8 9 10 11 12 pb
100
300
650

49
Tabela 08 - Correspondência entre o peso fresco (mg) e área (cm2) de lesões de frutos e folhas com mancha preta dos citros, utilizadas na validação dos oligonucleotídeos para o diagnóstico de Guignardia citricarpa.
Peso fresco
(mg) Área da lesão de fruto (cm2)
Área da lesão de folha (cm2)
1 0,0314 0,0962 5 0,0706 0,3377
10 0,2120 0,5929 15 0,2827 0,8443 20 0,3534 1,0406 25 0,4241 1,2920 30 0,5654 1,5375 35 0,6361 1,6789 40 0,7068 2,0127 45 0,7775 2,2050 50 0,9189 2,3464
De acordo com Glienke-Blanco et al. (2002), a detecção com o uso do primeiro
par de oligonucleotídeos foi eficiente na detecção e diferenciação do patógeno quando
foi utilizado DNA extraído a partir do micélio. Neste trabalho, foi possível detectar o
fungo com DNA extraído a partir de 1 mg de micélio e para lesões de manha dura o
limite de detecção foi de apenas uma lesão. Porém o fator limitante na utilização deste
par de oligonucleotídeos para diagnóstico de rotina não está somente na sensibilidade
dos oligonucleotídeos, mas principalmente no método de extração de DNA, pois
durante a extração é necessário romper a parede celular para a liberação dos esporos,
neste caso dos picnidiósporos. Para o segundo par de oligonucleotídeos, o limite de
detecção confirmou sua sensibilidade com DNA extraído de apenas uma lesão, como
descrito por Peres et al. (2007). Já na validação em lesões de folhas, o limite de
detecção foi de DNA extraído de 1 mg a 30 mg de lesões, o que sugere que a partir
desta área ocorra a interferência de algum fator que esteja inibindo a sensibilidade dos
iniciadores ou da reação de amplificação.
O limite de detecção com o uso do par de oligonucleotídeos GCF3/GCR7 para
micélio foi a partir de 25 mg micélio e para lesões em frutos, foi a partir de 1 mg.
Porém, durante a validação, que variou de 1 mg a 50 mg, ocorreram algumas falhas de
amplificação, o que permite concluir que este limite de detecção pode variar. Entretanto
no trabalho realizado por Bonants et al. (2003) com este par de oligonucleotídeos, o
limite de detecção só foi obtido com 20 lesões, o que dificultou o uso destes

50
oligonucleotídeos, pois nem sempre é possível obter este número de lesões em frutos de
citros, principalmente frutos para a exportação, mas por meio da comparação entre o
método atual, realizado por isolamento com a PCR, o autor em seu trabalho observou
uma eficência de 40% a mais na detecção de Guignardia quando utilizou o método
molecular (PCR), além do tempo de execução do teste, que no primeiro caso levou
cerca de duas semanas para obter o resultado.
Estes resultados demonstram a eficiência da utilização de PCR em diagnóstico
de rotina, sendo este mais rápido e capaz de diferenciar e detectar as duas espécies de
Guignardia spp. Meyer et al. (2006), em seu trabalho conseguiram, com a utilização de
pares de iniciadores baseados na região ITS diferenciar e detectar G. citricarpa e G.
mangiferae, com apenas DNA extraído a partir de uma lesão. Segundo os autores, para
a realização de diagnósticos de rotina é necessário obter um DNA de boa qualidade,
juntamente com pares de iniciadores específicos para o fungo, sendo possível detectar
em muitos casos, as duas espécies de Guignardia em um único material.
4.4.5 Phytophthora nicotianae
Através dessa validação, foi observado que P. nicotianae pode ser detectada por
PCR a partir do 2º dia após a incubação dos discos de folhas a 25 °C, 30 °C e 35 °C,
independente das condições de cobertura ou não dos copos. Os resultados também
mostraram que a melhor faixa de temperatura para a realização do ensaio, isto é, para
maior produção de esporângio, foi de 25 °C a 30 °C. Nessa faixa de temperatura a
produção de esporângios pode ser observada em microscopia a partir do 4º dia da
incubação dos discos de folhas em todos os tratamentos (Tabela 09). DNA total extraído
do micélio do oomiceto foi também utilizado nos testes de validação (Figura 20).
Na validação com discos foliares obtidos do teste da isca, foi possível comparar
a eficiência dos dois testes. A grande vantagem da técnica de PCR é a rapidez do
resultado, pois a partir do 2º dia de incubação do patógeno foi possível detectá-lo por
PCR. Entretato, algumas falhas que ocorrem no teste da isca também ocorrem na PCR,
pois nem sempre o disco de folha de limão retirado do teste está colonizado com o
patógeno, o que leva a falso negativo, sendo necessário, neste caso, aumentar o número
de amostragem do material. Outro fator observado foi a especificidade do par de
oligonucleotídeos ITS3/PINIC1, que mostrou-se sensível e específico, tanto para DNA

51
total de tecido colonizado, quanto de micélio. A validação a partir de DNA extraído de
diferentes quantidades de micélio do patógeno foi realizada para estimar a sensibilidade
e o potencial de uso em isolamentos sem meio seletivo, onde existe a possibilidade da
quantidade de micélio ser limitante, além das dificuldades de purificação devido à
presença de outros microrganismos.
O diagnóstico por PCR detectou 93,6% de amostras positivas, enquanto que o
ensaio biológico detectou somente 80,6%. O ensaio molecular detectou 19,4% das
amostras não detectadas pelo método biológico, enquanto que o biológico detectou
6,4% que não foi pelo molecular. Na temperatura de 35 °C houve a colonização dos
discos pelo patógeno apenas nos primeiros dias. No entanto, com o decorrer do tempo, a
taxa de colonização decaiu, uma vez, que a temperatura ideal para colonização é de 25
°C a 30 °C.
Figura 20 - Padrão eletroforético de reações de amplificação para detecção de Phytophthora nicotianae (IAC 01/95) com o par de oligonucleotídeos ITS3/PINIC1 com DNA extraído a partir de micélio: 1. controle negativo (P. citrophthora - LRS 44/87), 2. 1 mg, 3. 5 mg, 4. 10 mg, 5. 15 mg, 6. 20 mg, 7. 25 mg, 8. 30 mg, 9. 35 mg, 10. 40 mg, 11. 45 mg e 12. 50 mg de DNA total, M. 1Kb Plus DNA Ladder.
Atualmente, vários trabalhos estão sendo realizados utilizando métodos
moleculares para a detecção e diferenciação das espécies de Phytophthora. Érsek et al.
(1994) e Tooley et al. (1997), através da técnica de PCR, detectaram e diferenciaram
espécies de Phytophthora em solanáceas, por extração de DNA realizada em tecido e
solo colonizado com este patógeno. Ristaino et al. (1998) utilizando oligonucletídeos
específicos e enzimas de restrição separaram seis espécies de Phytophthora utilizando
como molde em PCR DNA extraído de micélio de cada patógeno.
M 1 2 3 4 5 6 7 8 9 10 11 12 pb
650
300
100

52
Tabela 09 - Comparação do resultado do teste biológico (isca) com o teste molecular por PCR em diferentes tempos de colonização de Phytophthora nicotianae com diferentes tipos de cobertura.
Tratamentos Ensaio Biológico Teste Molecular
Dias Dias
Cobertura Temperatura 2 4 6 8 10 2 4 6 8 10
Com tampa translúcida
15 °C - - - - + - + - + -
20 °C - + + + + - + + + +
25 °C - + + + + + + + + +
30 °C - + + + + + + + + +
35 °C - + - - - - + - - -
Sem tampa translúcida
15 °C - - - + + - + - + +
20 °C - + + + + + - + + +
25 °C + + + + + + + + + +
30 °C + + + + + + + + + +
35 °C - + + + + + - - + +
Com papel alumínio
15 °C - - + + + + + + + +
20 °C - + + + + + - + + +
25 °C - + + + + + + + + +
30 °C + + + + + + + + + +
35 °C - + - - - - + + - -
- = ausência do patógeno + = presença do patógeno
Em citros várias técnicas estão sendo utilizadas além da PCR, dentre elas a
nested PCR e o qPCR. Ippolito et al. (2002) em seu trabalho conseguiram diferenciar as
duas espécies em solo e raiz colonizados pelo patógeno, através de nested PCR, além da
utilização de solo e raízes para a extração, trabalhos também estão sendo realizados com
água utilizada para a irrigação (Kong et al., 2003).
Por meio destes trabalhos, conclui-se que o uso de técnicas moleculares para a
realização de diagnóstico de rotina é uma das alternativas para reduzir o tempo de
execução e para diferenciar as espécies de Phytophthora. Entretanto, na maioria dos
trabalhos, a extração foi realizada a partir de DNA extraído de solo ou raízes, o que
dificulta a sua utilização como rotina, pois solo e raízes podem conter outros

53
microrganismos. Além disso, é utilizada pequena quantidade de material, que neste
caso, nem sempre há uma grande quantidade do oomiceto na amostragem, o que poderia
levar a falsos negativos. Entretanto, a associação do teste molecular com o teste da isca
seria uma alternativa para a utlização de PCR em diagnóstico de rotina. Neste trabalho
foi possível observar a sensibilidade do par de oligonucleotídeos ITS3/PINIC1, como
descrito por Tooley et al. (2000), nas validações realizadas a partir de DNA extraído de
micélio e dos discos foliares colonizados com o patógeno.
4.4.6.Validação dos testes em frutos de citros infectados com Alternaria alternata f. sp. citri, Elsinoe spp. e Guignardia citricarpa após tratamento em packing house
Na validação dos testes para detecção dos patógenos em frutos após tratamento
em packing house, com exceção de Alternaria alternata e Elsinoe australis, todos os
outros patógenos foram detectados em reações com DNA extraído a partir de 1 mg de
tecido (Figuras 21 a 24).
Para Alternaria alternata a amplificação de fragmento específico deu-se em
reações com DNA extraído de 10 mg a 35 mg de tecido de casca de fruto, o que leva a
concluir que a sensibilidade do teste para detecção desse patógeno é menor quando o
fruto recebe tratamento em packing house (Figura 21). Neste caso, a detecção do
patógeno só poderá ser realizada quando tiver uma área mínima de 0,3317 cm2 para
extração de DNA.
Figura 21 - Padrão eletroforético de reações de amplificação para detecção de Alternaria alternata f. sp. citri patótipo tangerina com DNA extraído a partir de lesões de frutos de citros de pós- colheita, usando o par de oligonucleotídeos ACTT1-F2/ACTT1-R2. 1. 1 mg, 2. 5 mg, 3. 10 mg, 4. 15 mg, 5. 20 mg, 6. 25 mg, 7. 30 mg, 8. 35 mg, 9. 40 mg, 10. 45 mg e 11. 50 mg, M. 1Kb Plus DNA Ladder.
pb M 1 2 3 4 5 6 7 8 9 10 11
650
100
300

54
Na detecção de E. australis a amplificação de fragmento específico deu-se em
reações contendo DNA extraído a partir de 5 mg de lesões retiradas de frutos após
tratamento em packing house e os dois pares de iniciadores selecionados. Neste caso foi
mínima a interferência do tratamento realizado (Figura 22 a, b).
a a
a
b
Figura 22 - Padrão eletroforético de reações de amplificação para detecção de Elsinoe
australis com DNA extraído a partir de lesões de frutos de citros de pós- colheita. a. Par de oligonucleotídeos Els156-175F/Els554-573R. b. Els156-175F/Els503-522R: 1. 1 mg, 2. 5 mg, 3. 10 mg, 4. 15 mg, 5. 20 mg, 6. 25 mg, 7. 30 mg, 8. 35 mg, 9. 40 mg, 10. 45 mg e 11. 50 mg, M. 1Kb Plus DNA Ladder.
Para E. fawcetti a amplificação de fragmento específico deu-se em reações
contendo DNA extraído a partir de 1 mg de lesões retiradas de frutos após tratamento
em packing house, não ocorrendo interferência deste (Figura 23).
100
M 1 2 3 4 5 6 7 8 9 10 11 11
pb
300
650
M 1 2 3 4 5 6 7 8 9 10 11 pb
650
300
100

55
Figura 23 - Padrão eletroforético de reações de amplificação para detecção de Elsinoe
fawcetti com DNA extraído a partir de lesões de frutos de citros de pós- colheita. 1. 1 mg, 2. 5 mg, 3. 10 mg, 4. 15 mg, 5. 20 mg, 6. 25 mg, 7. 30 mg, 8. 35 mg, 9. 40 mg, 10. 45 mg e 11. 50 mg, M. 1Kb plus DNA Ladder.
Na detecção de Guignardia citricarpa em reações usando DNA extraído de 1 a
50 mg de frutos após tratamento em packing house e os pares de oligonucleotídeos
testados anteriormente, foi observado que ocorreu amplificação de fragmento de
tamanho esperado em todos os pontos. Este resultado demonstra que o tratamento
realizado em condições de packing house não interfere com o teste (Figura 24 a, b, c).
Estes resultados levam a concluir que o tratamento realizado nos frutos em
packing house não interfere na detecção do patógeno. Apenas no caso de A. alternata é
que houve signficativa interferência, embora ainda seja possível realizar diagnóstico de
rotina, pois a quantidade de lesão necessária ainda é pequena.
650
M 1 2 3 4 5 6 7 8 9 10 11 pb
300 100

56
a
b
c
Figura 24 - Padrão eletroforético de reações de amplificação para detecção de Guignardia citricarpa com DNA extraído a partir de lesões de frutos de pós- colheita de citros. a. GCP1/GCP2, b. GCMR/GCN e c. GCF3/GCR7 1. 1 mg, 2. 5 mg, 3. 10 mg, 4. 15 mg, 5. 20 mg, 6. 25 mg, 7. 30 mg, 8. 35 mg, 9. 40 mg, 10. 45 mg e 11. 50 mg, M. 1Kb Plus DNA Ladder.
4.5 qPCR
Com o uso do PCR em tempo real foi possível fazer uma quantificação relativa
de DNA de Phytophthora nicotianae em diferentes tempos após a inoculação do
650
100
300
M 1 2 3 4 5 6 7 8 9 10 11 pb
650
100
300
pb M 1 2 3 4 5 6 7 8 9 10 11
650
100
300
M 1 2 3 4 5 6 7 8 9 10 11 pb

57
patógeno em três diferentes hospedeiros. Foi observado um aumento na concentração de
DNA de P. nicotianae entre quatro dias a seis dias após a inoculação, com queda
acentuada a partir desse ponto. Nota-se que no genótipo mais resistente (Poncirus
trifoliata) o aumento na concentração de DNA do patógeno ocorre aos seis dias após a
inoculação, enquanto que em limão Cravo (moderadamente resistente) e tangerina
Sunki (altamente suscetível) esse aumento ocorre aos quatro dias após a inoculação
(Figura 25 a, b, c).
Estes dados confirmam os resultados obtidos por Ippolito et al. (2004) que,
utilizaram o mesmo par de oligonucleotídeos modificados para oligonucleotídeos
Scorpion e detectaram, diferenciaram e quantificaram P. nicotanae e P. citrophothora
em amostras de solo e raízes. Segundo os autores, o PCR convencional tem contribuído
para a identificação, detecção e controle desses patógenos, entretanto a grande
desvantagem deste método são as contaminações que podem ocorrer durante o processo.
Entretanto, no PCR em tempo real é possível monitorar estas contaminações, que
podem ocorrer, principalmente em análises em grande escala.
Outros trabalhos como de Hayden et al. (2006), vêm comparando o método do
qPCR com outros métodos utilizados correntemente em tecidos colonizados por P.
ramorum. Esses autores constataram a eficiência do qPCR, principalmente quando
utilizou uma concentração pequena de DNA.
Como controle endógeno foi utilizado o gene da β-actina. De acordo com Yan et
al. (2006), a β-actina pode ser utilizada como controle endógeno, entretanto, este não é
recomendado para expressão da variabilidade dos diferentes estágios do oomiceto, mas
neste trabalho foi utilizado apenas com o intuito de detectar e fazer a quantificação
relativa do patógeno. A utilização de apenas um controle interno neste trabalho foi
suficente para acurácia dos dados. O valor do Ct tem um potencial para ser utilizado na
detecção quantitativa de Phytophthora, sendo estes resultados importantes no controle
deste oomiceto (Ippolito et al., 2004). No presente trabalho, foi possível verificar
através do Ct quando ocorre a máxima concentração de DNA do patógeno em diferentes
hospedeiros.
Com a utilização da curva de dissociação foi possível verificar que não
ocorreram contaminações durante o processo (Figura 26 a, b, c). A eficiência de
amplificação foi verificada com cinco diluições realizadas, sendo a eficiência de
amplificação de 97% (Figura 27).
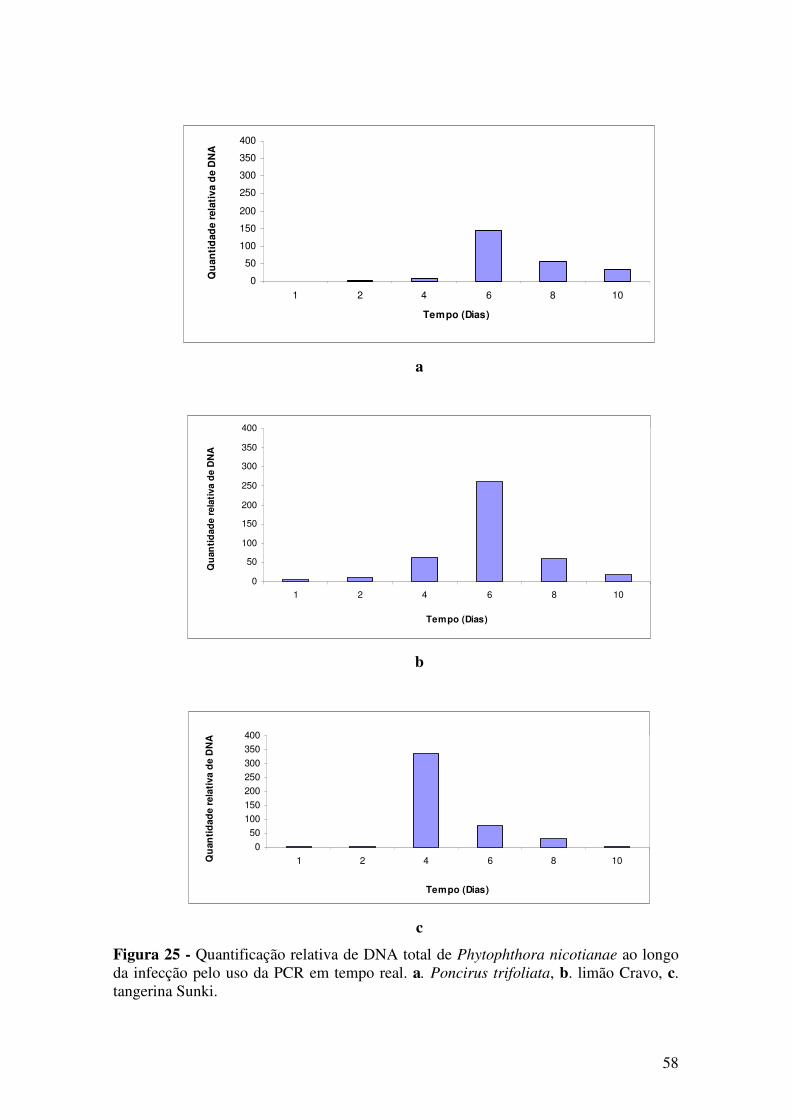
58
0
50
100
150
200
250
300
350
400
1 2 4 6 8 10
Tempo (Dias)
Qu
an
tid
ad
e r
ela
tiva d
e D
NA
a
0
50
100
150
200
250
300
350
400
1 2 4 6 8 10
Tempo (Dias)
Qu
an
tid
ad
e r
ela
tiva d
e D
NA
b
0
50
100
150
200
250
300
350
400
1 2 4 6 8 10
Tempo (Dias)
Qu
an
tid
ad
e r
ela
tiva d
e D
NA
c
Figura 25 - Quantificação relativa de DNA total de Phytophthora nicotianae ao longo da infecção pelo uso da PCR em tempo real. a. Poncirus trifoliata, b. limão Cravo, c. tangerina Sunki.

59
a
b
c
Figura 26 - Curva de dissociação realizada para verificar a existência de contaminantes. a. Poncirus trifoliata, b. limão Cravo, c. tangerina Sunki inoculados com Phytophthora
nicotianae. Ao lado de cada curva, padrão eletroforético de qPCR.

60
Figura 27 - Curva padrão baseada na diluição de DNA extraído a partir de tecido de citros infectado com Phytophthora nicotianae.
O qPCR mostrou ser sensível e específico, podendo ser utilizado em trabalhos de
detecção do patógeno diretamente de material infectado. Ippolito et al., (2004)
utilizaram os mesmos oligonucleotídeos para a detecção de P. nicotianae pelo uso de
qPCR. Os autores concluíram que existe uma alta correlação entre a concentração de
propágulos do patógeno no material infectado, tanto artificialmente quanto
naturalmente, e o Ct em qPCR.

61
5 CONCLUSÕES
Os resultados obtidos permitem concluir que: a) Para detecção de Phytophthora nicotianae em citros o ensaio biológico ainda é o
método mais seguro de detecção. A detecção por PCR associada ao ensaio biológico é
possível, mas exige um rigor de repetições que pode inviabilizar seu uso rotineiro.
b) Para a detecção de Alternaria alternata f. sp. citri patótipo tangerina, o par de
oligonucleotídeos ACTT1-F2/ACTT1-R2 é específico e sensível, porém atenção
especial deve ser dada ao método de extração de DNA.
c) Os dois pares de iniciadores GCP1/GCP2 e GCN/GCMR diferenciam e detectam
Guignardia citricarpa, sendo possível diagnosticar o patógeno em pequenas
quantidades de lesões de mancha dura de frutos cítricos.
d) Os oligonucleotídeos resultantes deste trabalho são específicos e capazes de
diferenciar Elsinoe australis (Els56-175F/Els503-522R, Els156-175F/554-573R) e
Elsinoe fawcetti (Els83-202F/Els374393R). Estes pares de oligonucleotídeos são
sensíveis mesmo quando a quantidade de material utilizado é pequena.
e) Mesmo após tratamento em packing house, os patógenos de citros poder ser
detectados por PCR em frutos com sintomas de mancha preta, verrugose e mancha
marrom.
f) Com a PCR em tempo real (qPCR) é possível monitorar a infecção de Phytophthora
nicotianae em diferentes genótipos de citros.

62
6 REFERÊNCIAS
ABECITRUS - ASSOCIAÇÃO BRASILEIRA DOS EXPORTADORES DE CÍTRICOS, http://www.abecitrus.com.br/clipping.asp#clip12 (28 janeiro 2008).
AGUILAR-VILDOSO, C. I. Caracterização genética de estirpes de Phytophthora
nicotianae, isoladas de plantas cítricas no Estado de São Paulo. 1997. 98p. Dissertação (Mestrado). Escola Superior de Agricultura “Luiz de Queiroz” Universidade de São Paulo, Piracicaba.
AGUILAR-VILDOSO, C.I.; RIBEIRO, J.G.B.; FEICHTENBERGER, E.; GOES. A.; SPÓSITO. M.B. Manual técnico de procedimentos da mancha preta dos citros. Brasília, 2002, 72p.
AGUILAR-VILDOSO, C.I.; ROSSI, M.L.; SASSERON, G.R.; NOGUEIRA, N.L. Isolamento, Caracterização e Patogenicidade de Alternaria alternata citri em tangerineiras. In. CONGRESSO PAULISTA DE FITOPATOLOGIA, 2004. Instituto Agronômico-IAC, Campinas - SP, 2004. Summa Phytopathologica, Botucatu, v. 30, p.109, 2004.
AKIMITSU, K.; PEEVER, T.L.; TIMMER, L.W. Molecular, ecological and evolutionary approaches to understanding Alternaria diseases of citrus. Plant Pathology, Oxford, v. 4, n. 6, p. 435-446, 2003.
BAAYEN, R.P.; BONANTS, P.J.M.; VERKLEY,G.; CARROLL,G.C; AA, H.A. van der; WEERDT, M. de; BROUWERSHAVEN, I.R. vander; SCHUTTE, G.C.; MACCHERON, W.; BRANCO, C.G.; AZEVEDO, J.L. Non pathogenic isolates of the citrus black spot fungus, Guignardia citricarpa, identified as a cosmopolitan endophyte of woody plants, G. mangiferae (Phyllosticta capitalensis). Phytopathology, Sta. Paul, v. 92, p. 88-93, 2002.
BILODEAU, G.J.; LÉVESQUE, C.A.; COCK, A.W.A.M.; DUCHAINE, C.; BRIÉRE,S.; URIBE, P.; MARTIN, F.N.; HAMELIN, R.C. Molecular detection of Phytophthora ramorum by real-time polymerase chain reaction using Taqman, SyBR green and Molecular Beacons. Phytopathology, Sta. Paul, v. 97, n. 5, p. 632-642, 2007.
BONANTS, P. J. M.; CARROLL, G. C.; WEERDT, M.; BROUWERSHAVEN, I.R.; BAAYEN, R.P. Development and validation of a fast PCR-basead detection method for pathogenic isolates of the citros black spot fungus, Guignardia citricarpa. Plant Pathology, Oxford, v. 109, p. 503-513, 2003. BUSTIN, S.A. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. Journal of Molecular Endocrinology, Bristol, v. 25, n. 2, p. 169-193, 2000. Review.

63
CAMPOS, S.B.; AGUILAR-VILDOSO, C.I. Efeito da glicose e sacarose no crescimento micelial de Guignardia citricarpa e Phytophthora nicotianae. Summa Phytopathologica, Botucatu, v. 27, n. 1, p. 97, 2001.
EPPO. Report of thr 6th meeting on EPPO diagnostics protocols for regulated, http://www.eppo.org/MEETINGS/2006_meetings/diag_fungi.htm (09 de março, 2008).
ÉRSEK, T.; SCHOELZ, J. E.; ENGLISH, J. T. PCR amplification of species-specific DNA sequences can distinguish among Phytophthora species. Applied and Environmental Microbiology, Washington, v. 60, n. 7, p. 2616-2621, 1994.
ERWIN, D. C.; RIBEIRO, O. K. Phytophthora Diseases Worldwide. APS Press: St. Paul. 1996. 562p.
FANTIN, G.M. Caracterização morfológica, fisiológica e patogência de espécies de Elsinoe em citros (Citrus spp). 1988. 90p. Dissertação (Mestrado) - Escola Superior de Agricultura ¨Luiz de Queiroz¨- Universidade de São Paulo, Piracicaba.
FANTIN, G. M.; KIMATI, H. Método de isolamento de Elsinoe australis e E. fawcetti a partir de frutas e folhas de citros com verrugose. In: XII Congresso Paulista de Fitopatologia, 1989a, Araras-SP. Summa Phytopathologica, 1989. v. 15. p. 22.
FANTIN, G. M.; KIMATI, H. Caracterização morfológica, fisiológica e patogênica de espécies de Elsinoe em citros. In: XX Congresso Brasileiro de Fitopatologia, 1989, Recife-PE. Fitopatologia Brasileira, 1989b. v. 14. p. 116.
FEICHTENBERGER, E. Doenças induzidas por fungos do gênero Phytophthora em citros e seu controle. Laranja, Cordeirópolis, v.10, n.2, p.359-378. 1989a.
FEICHTENBERGER, E. Mancha preta do Citros no Estado de São Paulo. Laranja, Cordeirópolis, v. 17, n. 1, p. 93-108,1996.
FEICHTENBERGER, E.; MÜLLER, G.W.; GUIRADO, N. Doenças dos citros (Citrus spp.). In: KIMATI, H. et al. (eds.). Manual de Fitopatologia. v. 2. Doenças das plantas cultivadas. 3. ed. São Paulo: Ceres. p. 261-296, 1997.
FEICHTENBERGER, E. Doenças incitadas por Phytophthora em citros. In: LUZ, E.D.M.N.; SANTOS, A.F.; MATSUOKA, K.; BEZERRA, J.L (eds). Doenças causadas por Phytophthora no Brasil. Campinas: Livraria Rural. p. 283-342, 2001.
FEICHTENBERGER, E.; SPÓSITO, M B.; PIO, R. M; CASTRO, J.L. Seleção de tangerinas e híbridos de citros para a tolerância à Mancha Marrom de Alternaria
(Alternaria alternata Keissler). Citricultura atual. Cordeirópolis, n. 45, v.8, p. 08-10, 2005.

64
FIGUEIREDO, M. B. Aplicação do método Castellani para conservação de fungos patogêncios em plantas. Biológico, São Paulo, v. 33 p. 9-13, 1967.
FNP. Anuário da Agricultura Brasileira. Agrianual 2007. São Paulo, v.1, outubro 2006, 516p.
FORSTER, H.; KINSHERF, T. G.; LEONG, S. A.; MAXWELL, D.P. Estimation of relatedness between Phytophthora species by analises of mitochondrial DNA. Mycologia, Philadelphia, v. 80, p. 466-478, 1988.
FORSTER, H.; OUDEMANS, P.; COFFEY, M.D. Mitochondrial and nuclear DNA diversity within six species of Phytophthora. Mycologia, Philadelphia, v. 14, p. 18-31, 1990.
FREEMAN, W. M.; WALKER, S. J.; VRANA, K. E. QuantitativeRT-PCR: pitfalls and potential. BioTechiques, Natick, v. 26, n. 1, p. 112-125, 1999.
GACHON, C.; MINGAN, C.; CHARRIER, B. Real Time PCR: what relevance to plant studies? Journal of Experimental Botany, Cambridge, v. 55, n. 402, p. 1445-1454, 2004.
GACHON, C.; SAINDRENAN, P. Real–time PCR monitoring of fungal development in Arabidopisis thaliana infected by Alternaria brassicicola and Botrytis cinerea. Plant Physiology and Biochemistry, Paris, v. 42, n. 5, p. 367-371, 2004.
GARDES, M.; BRUNS, T.D. ITS primers with enhanced specificity for basidiomycetes- application to the identification of mycorrhizae and rusts. Molecular Ecology, Oxford, v. 2, p. 113-118, 1993.
GENT-PELZER, M. P.E.; BROUWERSHAVEN, I.R.; KOX, L. F. F.; BONANTS, P. J. M. A TaqMan PCR method for routine diagnosis of the quarantine fungus Guignardia
citricarpa on citrus fruit. Plant Pathology, Oxford, v. 155, p. 357-363, 2007.
GLIENKE - BLANCO, C. Guignardia citricarpa Kiely: Análise genética cariotípica e interação com o hospedeiro. 1999. 200p. Tese (Doutorado)-Escola Superior de Agricultura ¨Luiz de Queiroz¨, Universidade de São Paulo, Piracicaba.
GLIENKE-BLANCO, C.; AGUILAR-VILDOSO, C.I.; VIEIRA, M. L. C.; BARROSO, P. A. V.; AZEVEDO, J. L. Genetic variability in the endophytic fungus Guignardia
citricarpa isolated from citrus plants. Genetics and Molecular Biology, Ribeirão Preto, v. 25, n. 2, p. 251-255, 2002.
GOODWIN, P. H.; KIRKPATRICK, B.C.; DUNIWAY, J. M. Cloned DNA probes for identification of P. nicotianae. Phytopathology, St. Paul, v. 79, p. 716-721. 1989.

65
GOODWIN, P. H.; ENGLISH, J. T.; NEHER, D. A.; DUNIWAY, J. M.; KIRKPATRICK, B. C. Detection of Phytophthora nicotianae from soil and host tissue with a species-specific DNA probe. Phytopathology, St. Paul, v. 23, n. 3, p. 277-281, 1990. GORDON, D. Viewing and editing assembled sequences using consed, current Protocols in Bioinformatics, In: Baxevanis, A. D. & Davison, D. B. (eds,) John Wiley & Co, New York, 2004.
HANAHAN, D. Studies on transformation on Escherichia coli with plasmids. Journal of Molecular Biology, London, v. 166, p. 557-580, 1983.
HATTA, R.; ITO, K.; YOSHITSUGU, H.; TANAKA, T.; TANAKA, T.; YAMAMOTO, M.; AKIMITSU, K.;TSUGE, T. A conditionally dispensable choromosome controls host-specific pathogenicity in the fungal plant pathogen Alternaria alternata. Genetics, Pittsburgh, v. 161 p. 59-70, 2002.
HAYDEN, H.; IVORS, K.; WILKINSON, C.; GARBELOTTO, M. TaqMan Chemistry for Phytophthora ramorum detection and quantification, with a comparison of diagnostic methods. Phytopathology, Sta. Paul, v. 96, n. 8, p. 846-854, 2006.
HIBBETT, D.S.; BINDER, M.; BISCHOFF, J. F.; BLACKWELL, M.; CANNON, P.F.; ERIKSSON, O.E.; HUHNDORF,T. J.; JAMES, T., KIRKD; P.M., CKING, R.L.; THORSTEN, H.; LUMBSCH, F.; MATHENY, P. B.; MCLAUGHLIN, D.J.; POWELL, J.; REDHEAD, S.; SCHOCH, C.L.; SPATAFORA, J.W.; STALPERS, R.V.; AIME, M.C.; APTROOT, A.; BAUER, R.; BEGEROW, D.;BENNY, G.L.; CASTLEBURY, L.A.; CROUSI, P.W.; DAI, Y.C.; GAMS, D.M.; GEISER, G.W.; GRIFFITH, C.G.; HAWKSWORTH, D.L.; HESTMARK, G.; HOSAKA, K.; HUMBER, R.A.; HYDE, K.D.; IRONSIDE, J.E.; URMAS, K.; KURTZMAN, C.P.; LARSSON, K.H.; LICHTWARDT, R.; LONGCORE, J.; DLIKOWSKA, J.M.; OBERWINKLER, F.; PARMASTO, E.; REEB, V.; ROGERS, J.D.; ROUX, C.; RYVARDEN, L.; SAMPAIO, J.P.; SCHU, A.; SUGIYAMA, J.; THORN, R.G.; TIBELL, L.; UNTEREINER, W.A.; WALKER, C.; WANG, Z.; WEIR, A.; WEISS, M.; WHITE, M.M.; WINKA, K.; ZHANG, Y. N. A higher-level phylogenetic classification of the Fungi.Research, Philadelphia, v. 111, p. 509–547, 2007. HYUN, J. –W.; TIMMER, L. W.; LEE, S. –C.; YUN, S. –H.; KO, S. –W.; KIM, K. –S. Pathological characterization and molecular analyses of Elsinoe isolates causing scab disease of citrus in Jeju Island in Korea. Plant Disease, Sta. Paul, v. 85, n. 9, p. 1013-1017, 2001. HYUN, J.-W.; PERES, N. A.; YI, S. Y.; TIMMER, L.W.; KIM, K.S.; KWON, H. M.; LIM, H. C. Development of PCR assays for the identification of species and pathotypes of Elsinoe causing scab on citrus. Plant Disease, Sta. Paul, v. 91, n. 7, p. 865-870, 2007.

66
IPPOLITO, A.; SCHENA, L.; NIGRO, F. Detection of Phytophthora nicotianae and P.
citrophthora in citrus roots and soils by nested PCR. Plant Pathology, Oxford, v. 108, p. 855-868, 2002.
IPPOLITO, A.; SCHENA, L.; NIGRO, F.; LIGORIO, V.S.; YASEEN, T. Real-time detection of Phytophthora nicotianae and P. citrophthora in citrus roots and soil. Plant Pathology, Oxford, n. 110, p. 833-834, 2004.
ISSHIKI, A.; AKIMITSU, K.; NISHIO, K.; TSUKAMOTO, M.; YAMAMOTO. Purification and characterization of an endopolygalacturose from the rough lemon pathotype of Alternaria alternata, the cause of citrus brow spot disease. Plant Pathology, Oxford, v. 51, p. 155-167, 1997. ISSHIKI, A.; OHTANI, K.; KYO, M.; YAMAMOTO, H.; AKIMITSU, K. Green fluorescente detection of fungal colonization and endopolygalacturonase gene expression in the interaction of Alternaria citri with citrus. Phytopathology, Sta. Paul, v. 93 - p. 768-773, 2003.
Jellyfish - Sequence Analysis Software versão 3.3.1, http:\jellyfish.labvelocity.com/index.jhtml.url (10 de Janeiro, 2008).
KIM, K. W.; HYUN, J. - W. Nonhost - associated proliferation of intrahyphal hyphae of citrus scab fungus Elsinoe fawcetti: refining the perception of cell-within-a-cell organization. Science, Washington, v. 38, p. 565-571, 2007.
KONG, P.; HONG, C.; JEFFERS, S. N.; RICHARDSON, P. A. A species – specific polymerase chain reaction assay for rapid detection of Phytophthora nicotianae in irrigation water. Phytopathology, Sta. Paul, v. 93, n. 7, p. 822-831, 2003.
KOTZÉ, J.M. Epidemiology and control of citrus black spot in South Africa. Plant Disease, Sta. Paul, v. 65, p. 945-950, 1981.
LARANJEIRA. F.F.; AMORIM, L.; BERGAMIN FILHO, A.; AGUILAR-VILDOSO, C. I.; COLETTA-FILHO, H.D. Fungos, Procariotos e doenças abióticas. In: Mattos Junior, D.; DE NEGRI, J. D.; PIO, R. M & POMPEU JUNIOR, J. (eds).Citros, Cordeirópolis, p. 223-277, 2005.
LARSEN, R. C.; VANDEMARK, G.J.; HUGHES, T. J.; GRAU. C.R. Development of a Real-Time Polymerase Chain Reaction Assay for Quantifying Verticillium albo-atrum DNA in Resistant and Susceptible Alfalfa. Phytopathology, Sta Paul, v. 97, n. 11, p. 1519-1525, 2007.
LEE ,S. B.; TAYLOR, J. W. Phylogeny of five fungus - like protoctistan Phytophthora species, inferred from internal transcribed spacers of ribosomal DNA. Molecular Biology Evolution, Oxford, v. 9, p. 636-653, 1992.

67
LEE, S. B.; TAYLOR, J. W. Detection of Phytophthora species by oligonucleotide hybridization to amplified ribosomal DNA spacers. Phytopathology, Sta. Paul, v. 83, p. 177-181, 1993. LICCIARDELLO, G. Identification and detection of Phoma tracheiphila, causal agent of citrus Mal Secco disease, by real - time polymerase chain reaction. Plant Disease, Sta. Paul, v. 90, n. 12, p. 1523-1530, 2006. MARRA, R.E.; MILGROOM, M. G. PCR amplification of the mating-type idiomorphs in Cryphonectria parasitica. Molecular Ecology, Oxford, v. 8, p. 1947-1950, 1999. MAYER, Z.; BAGNARA, A.; FARBER, P.; GEISEN, R. Quantification of the copy number of nor -1, a gene of the aflotoxin biosynthetic patway by real - time PCR, and its correlation to the cfu of Aspergillus flavusin foods. International Journal of Food Microbiology, Amsterdam, v. 82, n. 2, p. 143-151, 2003. MASUNAKA, A.; TANAKA, A.; TSUGE, T.; PEEVER, T.L.; TIMMER, L.W.; YAMAMOTO, M.; YAMAMOTO, H.; AKIMITSU, K. Distribution and characterization of AKT homologs in the tangerine pathotype of Alternaria alternata. Phytopathology, Sta Paul, v. 90, n. 7, p. 762-768, 2000.
MAVRODIEVA, V.; LEVY, L.; GABRIEL, D. W. Improved sampling methods for real-time polymerase chain reaction diagnosis of citrus canker from field samples. Phytopathology, Sta Paul, v. 94, n. 1, p. 61-68, 2004.
MEYER, L.; SANDERS, G. M.; JACOBS, R.; KORSTEN, L. A one - day sensitive method to detect and distinguish between the citrus black spot pathogen Guignardia
citricarpa and the endophyte Guignardia mangiferae. Plant Disease, Sta. Paul, v. 90, n. 1, p. 97-101, 2006.
MENDES, M.A.S.; FREITAS, V.M. Espécies invasoras para a citricultura. Comunicado Técnico 130, Brasília, 2005.
NCBI - National Center for Biotechnology Information, http://www.ncbi.nlm.nih.gov/ (10 janeiro 2008).
NEVES, M.F., LOPES, F.F.; TROMBIN, V.G.; AMARO, A. A.; NEVES, E.M.; JANK, M.S. Caminhos para a citricultura. http://www.agrosoft.org.br/?q=node/25297 (28 janeiro 2008).
OLIVEIRA, M.S; MEDAU, R.; URASHIMA, A.S. Seleção de “OLIGONUCLEOTÍDEOS” para identificação de espécies de Phytophthora em citros. In. CONGRESSO PAULISTA DE FITOPATOLOGIA, 2008. Campinas - SP, 2008. Summa Phytopathologica, Botucatu, v. 34, p.36-37, 2008.rdenadoria de Assistência Técnica Integral (CATI),

68
PERES, N.A.; HARAKAVA, R.; CARROL, G. C.; TIMMER, L. W.; ADASKAVEG, J. E. Comparison of molecular procedures for detection and identification of Guignardia citricarpa and G. mangiferae. Plant Disease, Sta. Paul, v. 91, n. 5, p. 525-53, 2007. PEEVER, T. L.; CANILHOS, Y.; OLSEN, L.; IBAÑEZ, Y.-C. L.; TIMMER, L. W. Population genetic struture and host specificity of Alternaria spp. causing brown spot of Minneola tangelo and Rough Lemon in Florida. Phytopathology, Sta. Paul, v. 88, n. 10, p. 851-860, 1999.
PEEVER, T. L.; IBAÑEZ, A.; AKIMITSU, K.; TIMMER, L. W. Worldwide phylogeography of the citrus brown spot pathogen, Alternaria alternata. Phytopathology, Sta. Paul, v. 92, n. 7, p. 794-802, 2002.
PEEVER, T. L.; CARPENTER, L. B.; TIMMER, L. W. Molecular systematics of citrus - associated Alternaria species. Mycologia, Philadelphia, v. 96, n.1, p. 119-134, 2004.
PRYOR, B.; MATHERON, M.; FIGULI, P. Characterization of Alternaria isolates associated with Alternaria rot of citrus, www.azda.gov/CDP//New CBC\AILRC\2003Research\2003-04.pdf- (02 de março, 2008).
RISTAINO, J. B.; MADRITCH, M.; TROUT, C. L.; PARRA, G. PCR amplification of ribosomal DNA for species identification in the plant pathogen genus Phytophthora. Applied and Environmental Microbiology, Washington, v. 64, n. 3, p. 948-954, p. 1998. ROSSETTI, V.V.; MELO, O.F.; PENTEADO, L.B.K.; OLIVEIRA, D. Estudo das espécies e cepas de Phytophthora isoladas de pomares cítricos no Estado de São Paulo, e sua patogenia. In: Congresso Brasileiro de Fruticultura, Anais, SBF, p. 505-526, 1972.
ROSSETTI, V.V. Manual ilustrado de doenças dos citros. Piracicaba: FEALQ/Fundecitrus, 2001. 207 p.
ROTEM, J. The genus Alternaria biology, epidemiology and pathogenicity. Sta. Paul, APS Press, 1994.
SÃO PAULO, Portaria CDA-15, de 15 de outubro de 2003. Estabelecer normas para a produção de mudas cítricas certificadas no Estado de São Paulo. Diário Oficial do Estado de São Paulo, SP, 16 outubro 2003, Seção 1, v. 113, n. 197.
SEAGRI - SECRETARIA DA AGRICULTURA, IRRIGAÇÃO E REFORMA AGRÁRIA, http://www.seagri.ba.gov.br/noticias.asp?qact=view¬id=9761 (28 janeiro 2008).
SIVANESAN, A; CRITCHETT, C. Elsinoe fawcetti, CMI Descriptions of Pathogenic Fungi and Bacteria, Kew, v.438, 2p., 1974.

69
SWART, L.; CROUS, P.W.; KANG, J.C.; MCHAU, G.R.A.; PASCOE, I.; PALM, M. E. Differentiation of species of Elsinoe associated with scab disease of proteaceae based morphology, symptomatology, and ITS sequence phylogeny. Mycologia, Philadelphia, v. 93, n. 2, p. 366-379, 2001.
TAN, M.K.; TIMMER, L. W.; BROADBENT, P.; PRIEST, M.; CAIN, P. Differentiation by molecular analyses of Elsinoe spp. causing scab diseases of citrus and ITS epidemiological implications. Phytopathology, Sta. Paul, v. 86, n. 10, p. 1039-1044, 1996. THOMMA, B. P. H. J. Alternaria spp.: from general saprophyte to specific parasite. Plant Pathology, Oxfor, v. 4, n. 4, p. 225-236, 2003.
TIMMER, L. W.; PRIEST, M.; BROADBENT, P.; TAN, M. –K. Morphological and pathological characterization of species of Elsinoe causing scab of citrus. Phytopathology, Sta. Paul, v. 86, n. 10, p. 1032-1033, 1996.
TOOLEY, P.W.; BUNYARD, M.M CARRAS; HATZILOUKAS, E. Development of PCR oligonucleotídeos from internal transcribed space region 2 for detection of Phytophthora species infecting potatoes. Applied and Environmental Microbiology, Washington, v. 63, n. 4, p. 1467-1475, 1997.
TYAGI, S.; KRAMER, F.R. Molecular beacons: probes that fluoresce upon hybridization. Nature Biotechnology, London, v. 14, p. 303-308, 1996.
WALTON, J.D. Host-selectivetoxins: Agents of compatibility. The Plant Cell, Rockville, v. 8, p. 1723-1733, 1996.
WATERHOUSE, G. M. Key to the species of Phytophthora de Bary. Mycologia, Philadelphia, v. 92, p. 22, 1963.
WHITESIDE, J.O.; GARNSEY, S.M.; TIMMER, L.W. Compendium of citrus diseases. APS Press:Sta. Paul, 1993. 80p.
YAN, H. Z.; LIOU, R.F. Selection of internal control genes for real-time quantitative RT-PCR assays in the oomycete plant pathogen Phytophthora nicotianae. Science, Washington, v. 43, p. 430-438, 2006.

70
7 ANEXO
7. Anexo 1- Seqüências dos fragmentos amplificados a partir dos oligonucleotídeos selecionados para detecção dos patógenos.
Patógeno Oligonucleotídeos Sequência 5’-3’
Phytophthora nicotianae
ITS-3 / PINIC-1
1 GCATCGATGA AGAACGCAGC GAACTGCGAT ACGTAATGCG AATTGCAGGA TTCAGTGAGT 61 CATCGAAATT TTGAACGCAT ATTGCACTTC CGGGTTAGTC CTGGAAGTAT GCCTGTATCA 121 GTGTCCGTAC ATTAAACTTG ACTTTCTTCC TTCCGTGTAG TCGGTGGAGG AGATGTCAGA 181 TGTGAAGTGT CTTGCGATTG GTCTTCGGAC CGGCTGCGAG TCCTTTTAAA TGTACTAAAC 241 TGAACTTCTC TTTGCTCGAA AAGTGGTGGC GTTGCTGGTT GTGAAGGCTG CTATTGTGGC 301 AAATTGGCGA CTGGTTTGTC TGCTGCGGCG TTAATGGAAG AGTGTTCGAT TCGTGGTATG 361 GTTGGCTTCG GCTGAACAAT GCACTTATTG GACGTTTTTC CTGCTGTGGC GTGATGGACT 421 GGTGAACCAT AGCTCGGTGG CTTGGCTTTT GAATA
GCP1/GCP2
1 ACAGGTGCTG AAGTGTGAGT GTCGAAGGTG AAATTGAGGT GAGGTTGACG GCGTCTGCGG 61GAGAGGCGAG TAGCGGGTGC TGTAGAAGTT GTATGATCCG CGGTGCGCGT TGGGGTACAG 121GAGAGTGGAA GGAGACAGCG CTGCAACAGG GAAGGTTAGA CTTTCAATTG AGCCGGAGCT 181GGAGCGGTTC ACCCCGAATC ACGTGCATTC AGGCGACTTT CGGATCATCA ATCACCCGGA 241TTTTAGATGG GTGCTCTGGA AATAGGAGTG AAGAAATGTA GGTTATTACT ATCTGCCGTA 301GAAAAGCGAG TCGTCAATGA TTATACCAAT CGTGACTATG ATACCTCATG AAGCCGCCCC 361ATGCAGCACCTGT
GCMR/GCN
1 TGAAATACGT AATCCTGAAA GGTGATGGAA GGGAGGCCTT AAAAAAGCCG CCCGACCTAC 61 CTTCACACCC TTGTGTATCT ACCATGTTGC TTTGGCGGGC CGACCCGGTT TTGACCCGGG 121 CGGTCGGCGC CCCCAGCCTA GTCTCTAGGC CAGGACGCCT GGCTAAGTGC CCGCCAGTAT 181 ACAAAACTCC AGCGATTATT CTGTGTAGTC CTGAGAATTC ATTTAATGAA ATAAAACTTT 241 CAACAACGGA TCTCTTGGTT CTGGCATCGA TGAAGAACGC AGCGAAATGC GATAAGTAAT 301 GTGAATTGCA GAATTCAGTG AATCATCGAA TCTTTGAACG CACATTGCGC CCCCTGGCAT 361 TCCGGGGGGC ATGCCTGTTC GAGCGTCATT TCAACCCTCA AGCTCTGCTT GGTATTGGGC 421 GACGTCCGCT GCCGGACGCG CCTGGAAGAC CTCGGCGACG GCGTCTCAGC CTCGAGCGTA 481 GTAGTAAAAT ATCTCGCTTT GGAGGAGGGG GGCGCTGGCC GCCGGACAAT CGACCTTCGG 541 TCACTATTTT TCCAAGGTTG ACCTCGGATC AGGTAGGGAT ACCCGCTGAA CT
Guignardia citricarpa
GCF3/GCR7
1 TGAAATACGT AATCCTGAAA GGTGATGGAA GGGAGGCCTT AAAAAAGCCG CCCGACCTAC 61CTTCACACCC TTGTGTATCT ACCATGTTGC TTTGGCGGGC CGACCCGGTT TTGACCCGGG 121 CGGTCGGCGC CCCCAGCCTA GTCTCTAGGC CAGGACGCCT GGCTAAGTGC CCGCCAGTAT 181 ACAAAACTCC AGCGATTATT CTGTGTAGTC CTGAGAATTC ATTTAATGAA ATAAAACTTT 241 CAACAACGGA TCTCTTGGTT CTGGCATCGA TGAAGAACGC AGCGAAATGC GATAAGTAAT 301 GTGAATTGCA GAATTCAGTG AATCATCGAA TCTTTGAACG CACATTGCGC CCCCTGGCAT 361 TCCGGGGGGC ATGCCTGTTC GAGCGTCATT TCAACCCTCA AGCTCTGCTT GGTATTGGGC 421 GACGTCCGCT GCCGGACGCG CCTGGAAGAC CTCGGCGACG GCGTCTCAGC CTCGAGCGTA 481 GTAGTAAAAT ATCTCGCTTT GGAGGAGGGG GGCGCTGGCC GCCGGACAAT CGACCTTCGG 561 TCACTATTTT TCCAAGGTTG ACCTCGGATC AGGTAGGGAT ACCCGCTGAA CT
Continua ...

71
Continuação ... Patógeno Oligonucleotídeos Sequência 5’-3’
Alternaria alternata f. sp. citri
ACTT1-F2/ACTT1-R2
1 CACAGGCTAT CTTCACATGC AACACGCTAT TGAAAACAGC TGTGAAAGCT TCTCAGAAAG 61 TCGGGATCCC TCTGGCAAAC GTCTTTTTAA CCGATGCGCC TGGTTCCTAC CGGCCGGATA 121 ACGTCTACCC CTTTGAAGAG ATAGACAATA TAGTGCGTAC AGCGAAGTCC AGTCTCCCGC 181 TGCTTCGACT TGGGAGAGGA CAAGGTTCCA GCACTCCAGC ATATATATGT TTCTCAAGTG 241 GAACATCTGG CGCGCAAGTA CGTCTCTAGA TAAATACTCA CAATGCGCGA GATTTTCTGA 301 CCAAACTGCC AGAAACCTGT GCTACTCTCC CACCAGGGCA TCATTGCGAA CATTCTACAG 361 ATCAACACGT TTGAGCATTT TAGGCAGAGG GGCCCAAATG TCTCGCTGTG TATCCTGCCA 421 CTCGCTCACA GCTACGGACT GGTATGTGTT GCGTACAGTA CTTTGTACCG GGGAGACAGG 481 CTTGCCGTCT TACCCTCTTC CGACGTCGAA GACCTTTTTA GTATTATTGA GAAACTTCAA 541 ATAAATACGC TGTACCTTGT ACGTATACAT GCAACATTCT TAATCGCTGT ATAGTATGCT 601 CAAACCCCGT GATGTAGGTG CCTACCCTCG TTTCAAGGAT TCTAAGTGGT GGGAAAGCAG 661 GGGGACACGA TCTCAGCTGT GTCAAAGAAG TTTATACAGG TGGTGCACCA CTGCACCCGA 721 CGCTAGAGGA ACATGTCCTC CGAGACCACC CTACGTGGAA GATAAAGCAA TGCTATGGAG 781 CTACG
Elsinoe australis
Els 156-175F / Els 554-573R
1 AAAAAATATA AAGAAAAAAA CAAAATTATA TTTAAAATAA GTATAAATCA TATAAAGAGA 61 TCAAAAAAGG AATTAAAACC AAAAATCATA TGAATTTAAA GAATATTAAT TAAAATAATA 121 GATATTATAA TTATTTATAT ATCTTTTAAT AAATATGGAC AGGGCATATA AGGTATTATA 181 TAGTGAAAAA CACAATACAT AGAAATCTTA TAAAATAAAA TTAAAAAATA AAAAATATAG 241 AAAATAGATC GATTAAAAGG AAGAGCCAGA TGCACGCAAT CAAACACGAA TCTTGCAGAA 301 GTTTAGATTT TACATATACA AGTAGCTATA TAAGGTGGAA GACAAACCGA CCTCGGGAAC 361 TATTCGGATA TTTTGATTAA TAATGATCTG AAAGTAGGAT TGGGTTAGTA ATTACGTAAG 421 GAATTATAGA TGAAATAATA ATAATAAATA ATTGGAGGAC CGAATCAACT CTGTTTTTAA 481 CCATAATGTC GGAGTACTTT TGTACAATCA AATTAAAACT TTCAACAACG GATCTCTTGG 541 TTCTGGCATC GATGAAGAGC GCAGCGAAAT GCGATAAGTA ATGTGAATTG CAGAATTCAG 601 TGAATCATCG AATCTTTGAA CGCACATTGC GCCCCTTGGT ATTC CGAGGG GCATGCCTGT 661 TCGAGAGTCA TTTCACCAAT CAAGCCCCGC TTGGTATTAG GCGCCTGGCC GGCCCCCCCG 721 TGGCCGGCCC GCCCGAAATG CATCGGCGAG GCACCGACCC CCGGCGTGTT AGAATTTCGA 781 AACTTCCGG
Els 156-175F/ Els 503-522R
1 GTAGGTGAAC CTGCGGAAGG ATCATTAACG AGGAAGGGTT TCCGAAAGGA GCCCGAACTC 61 CCCACCCTTT GCTGTTGCGA ATCTGTTGCT TCGGCGGGCC CGCCCCCCCA TCCCTGGGGG 121 GGCGGCCGGG CCTCACCGCC CGGACAGCGC CCGCC GGAGGACCGAATCAA CTCTGTTTTT 181 AACCATAATG TCGGAGTACT TTTGTACAAT CAAATTAAAA CTTTCAACAA CGGATCTCTT 241 GGTTCTGGCA TCGATGAAGA ACGCAGCGAA ATGCGATAAG TAATGTGAAT TGCAGAATTC 301 AGTGAATCAT CGAATCTTTG AACGCACATT GCGCCCCTTG GTATTCCGAG GGGCATGCCT 361 GTTCGAGCGT CATTTCACCA ATCAAGCCCC GCTTGGTATT AGGCGCCTGG CCGCCCCCCC 421 CGTGGCCGGC CCGCCCGAAA TGCATCGGCG AGGCACCGAC CCCCGGCGTG TTAGAATTTC 481 GAAACGTCAG GAGCACCGGT GTTAACCTCT GCCGTGAAAC CTTTCAAACT ATTTTTCTAA 541 GGTTGACCTC GGATCAGGTA GGAATACCCG CTGAACTTAA GCATATCAAT AAGCGGAGGA 601 AAAGAAAC
Elsinoe .fawcetti
Els183-202F Els374-393R
1 GTAGGTGAAC CTGCGGAAGG ATCATTAACG AGGTAGGGCC TCCTTCGGGA GCCCGAACTC 61 CCCACCCTTT GCTGTTGCGA ATCACGTTGC TTCGGCGGGA CCTCCCCCCC TCGAGAGGGG 121 GGCGGGGGGC CGGGCCACCG CGCCCGGACA GCGCCCGCCG GGGGACCGAA CCAACTCTTG 181 TC TTGTGAAA CCTTTGCAGT CGGAGTACAA CCGTACAATC AAATCAAAAC TTTCAACAAC 241 GGATCTCTTG GTTCTGGCAT CGATGAAGAA CGCAGCGAAA TGCGATAAGT AATGTGAATT 301 GCAGAATTCA GTGAATCATC GAATCTTTGA ACGCACATTG CGCCCCTTGG TATTCCGAGG 361 GGCATGCCTG TTC GAGCGTC ATTTCACCAA TC AAGCCCCG CTTGGTATTG GGTGCGACAG 421 CCCGCCCCCC GTGGCCGGCC CGCCCGAAAT GCATCGGCGA GGCACCGACC CCCGGCGTGT 481 TAGAATTTCG AAACGTCAGG AGCACCGGTG ACCCTCCGCC GTCAACCCGG ACCCCTCCCC 541 CCCGCGGGCG GGGCCCACCC CCCTTCAAGG TTGACCTCGG ATCAGGTAGG AATACCCGCT


![1 - Tradução - O que é o externismo - Katalin Farkas [Luiz Helvécio Marques Segundo]](https://static.fdocumentos.com/doc/165x107/55cf9cc9550346d033ab0b99/1-traducao-o-que-e-o-externismo-katalin-farkas-luiz-helvecio-marques.jpg)















