DIVERSIDADE DE BACTÉRIAS ENDOFÍTICAS EM · PDF file06 Padrão de...
116
UNIVERSIDADE ESTADUAL DE MONTES CLAROS DIVERSIDADE DE BACTÉRIAS ENDOFÍTICAS EM BANANEIRA ‘PRATA- ANÃ’ SUZANE ARIÁDINA DE SOUZA 2011
Transcript of DIVERSIDADE DE BACTÉRIAS ENDOFÍTICAS EM · PDF file06 Padrão de...

UNIVERSIDADE ESTADUAL DE MONTES CLAROS
DIVERSIDADE DE BACTÉRIAS ENDOFÍTICAS EM BANANEIRA ‘PRATA-
ANÃ’
SUZANE ARIÁDINA DE SOUZA
2011

SUZANE ARIÁDINA DE SOUZA
DIVERSIDADE DE BACTÉRIAS ENDOFÍTICAS EM BANANEIRA ‘PRATA-ANÃ’
Dissertação apresentada à Universidade Estadual de Montes Claros, como parte das exigências do Programa de Pós-Graduação em Produção Vegetal no Semiárido, área de concentração em Produção Vegetal, para obtenção do título de “Magister Scientiae”.
Orientadora Profª. D.Sc. Sílvia Nietsche
JANAÚBA MINAS GERAIS – BRASIL
2011

Catalogação: Biblioteca Setorial Campus de Janaúba
Souza, Suzane Ariádina de.
S729d Diversidade de bactérias endofíticas em bananeiras ‘Prata-Anã’ [manuscrito] / Suzane Ariádina de Souza – 2011.
101 p.
Dissertação (mestrado)-Programa de Pós-Graduação em Produção Vegetal no Semiárido, Universidade Estadual de Montes Claros-Janaúba, 2011.
Orientadora: Profª. D. Sc. Silvia Nietsche. 1. Bananeiras. 2. Bactérias endofíticas. 4. Musa spp. I. Nietsche, Silvia. II. Universidade Estadual de Montes Claros. III. Título. CDD. 634.772

SUZANE ARIÁDINA DE SOUZA
DIVERSIDADE DE BACTÉRIAS ENDOFÍTICAS EM BANANEIRA ‘PRATA-ANÃ’
Dissertação apresentada à Universidade Estadual de Montes Claros, como parte das exigências do Programa de Pós-Graduação em Produção Vegetal no Semiárido, área de concentração em Produção Vegetal, para obtenção do título de “Magister Scientiae”.
Aprovada em 02 de setembro de 2011.
Profª. D.Sc. Sílvia Nietsche Profª. D.Sc. Márcia Regina Costa (Orientadora) (Coorientadora)
Profª. D.Sc. Adelica Aparecida Xavier D.Sc. Luciana Nogueira Londe (DCA- UNIMONTES) (EPAMIG)
JANAÚBA MINAS GERAIS – BRASIL
2011

A Deus, minha fonte de sabedoria e amor; À minha amada mãe, Marluce; Ao meu querido irmão, Augusto; Aos meus queridos avôs Alfredo e Deza; Ao meu namorado, Fabrício. Dedico

AGRADECIMENTOS
Mais uma etapa se finda, e agora só tenho a agradecer a todos que me
ajudaram a subir mais este degrau na escada da vida.
A Deus, meu grande mestre, por ter me carregado no colo nos momentos
mais difíceis desta caminhada;
À minha mãe, pelo amor incondicional, por ser meu exemplo de vida.
Ao meu irmão, Augusto, pelo apoio e estímulo;
Aos meus avôs Alfredo e Maria Joaquina, por estarem do meu lado
sempre que precisei;
A Fabrício, pelo amor, companherismo e paciência;
À CAPES, pela bolsa concedida, e à FAPEMIG, pelo apoio financeiro,
para execução deste trabalho;
À minha orientadora, Sílvia Nietsche, minha eterna gratidão, por todos
os ensinamentos, pela paciência, pelo apoio, oportunidade e pelas palavras de
estímulo;
À professora Márcia, pela ajuda, paciência, pela amizade e pelo
agradável convívio;
À professora Adelica, pela disposição, pelos conselhos e pelas valiosas
sugestões;
Ao pesquisador Eder Jorge de Oliveira, pela ajuda nas análises
estatísticas;
À pesquisadora Luciana Londe, pela ajuda na análise dos dados e pela
disposição;
Aos meus familiares, principalmente Tia Marlene, Alcides, Murillo,
Fernando e Alcilene, pelo apoio, convivência e pela moradia durante esses anos;
Aos colegas do Laboratório de Fitopatologia: Leandro, Isaac, Josiane,
Bruna, Umberson, Lívia, Rafael, Jefferson e Maria Isabel, pela amizade e

brincadeiras, agradeço, em especial, a Acleide, pela ajuda na realização do
experimento.
Aos amigos do laboratório de Biotecnologia: Renata, Elizete, Josiane,
João, Irton, Mara, Felipe, Rayka, Gláucia e Tallyta, pela convivência e
companherismo, em especial a Nayara, Tiago e Emilly, pela ajuda na condução
deste trabalho.
Aos colegas do mestrado, pela alegria e agradável amizade. Agradeço
especialmente Poly, Irani, Joseilton, Fábio, Léo e Renata, por todo apoio, pelas
palavras de conforto e por me escutarem nas horas de angústia.
A todos os funcionários da UNIMONTES, principalmente a Dona Ana e
Dona Brasilina, pela disposição em ajudar.
Enfim, a todos que estiveram ao meu lado durante essa caminhada.
Muito Obrigada.

SUMÁRIO
RESUMO .................................................................................................. I ABSTRACT ........................................................................................... III 1 INTRODUÇÃO ..................................................................................... 1 2 REFERENCIAL TEÓRICO ................................................................ 4 3 MATERIAL E MÉTODOS ................................................................ 17 3.1 Coleta das Raízes e Isolamento das Bactérias .................................... 17 3.2 Caracterização dos isolados bacterianos com relação à resposta de Gram ......................................................................................................... 21 3.3 Teste de patogenicidade por meio da reação de hipersensibilidade ... 22 3.4 Caracterização da Diversidade via Marcador Molecular ................... 23 3.4.1. Extração de DNA ........................................................................... 23 3.4.2 Amplificação por rep-PCR .............................................................. 24 3.4.3 Amplificação pela técnica ARDRA ................................................ 25 3.4.4 Amplificação por PCR dos genes nifH ............................................ 26 3.5 Análise dos Dados Moleculares ......................................................... 27 3.6 Sequenciamento do DNA ................................................................... 27 4 RESULTADOS .................................................................................... 30 4.1 Isolamento das Bactérias .................................................................... 30 4.2 Caracterização dos isolados bacterianos com relação à resposta de Gram ......................................................................................................... 31 4.3 Teste de patogenicidade por meio da reação de hipersensibilidade ... 32 4.4 Análise de variância molecular .......................................................... 33 4.5 Sequenciamento do DNA bacteriano ................................................. 34 4.6 Caracterização da Diversidade via rep-PCR ...................................... 41 4.7 Caracterização da Diversidade pela técnica ARDRA ........................ 47 4.8 Caracterização do Gene nifH .............................................................. 56 5 DISCUSSÃO ........................................................................................ 60 6 CONCLUSÕES ................................................................................... 77 REFÊRENCIAS BIBLIOGRÁFICAS ................................................. 78 ANEXOS ............................................................................................... 100

TABELAS
01 Localização geográfica das áreas de coleta de raízes de bananeira ‘Prata-Anã’ para isolamento de bactérias endofíticas.................................
18
02 Sítio de restrição das enzimas Hinf I, MspI, NdeI e RsaI........................ 25
03 Análise de variância molecular (AMOVA), de 201 isolados bacterianos provenientes de dezesseis populações de ‘Prata-Ana’, utilizando os marcadores moleculares REP, ERIC e BOX.........................
33
04 Análise de variância molecular (AMOVA), de 201 isolados bacterianos provenientes de dezesseis populações de ‘Prata-Ana’...
33
05 Identificação das bactérias endofíticas de raízes de bananeira ‘Prata-Anã’ com base na identidade da sequência parcial do gene16S rDNA......
35
06 Frequência de bactérias endofíticas obtidos de raízes bananeira ‘Prata-Anã’coletadas em 16 áreas de cultivo no Norte de Minas Gerais e Guanambi (BA)...........................................................................................
39
07 Número de bandas monomorficas e polimorficas formadas com a utilização dos primers REP, ERIC e BOX.................................................
42
08 Distância genética entre os isolados da mesma área onde foram coletadas raízes de bananeira ‘Prata-Anã’ por meio da técnica rep PCR...........................................................................................................
45
09 Agrupamento dos gêneros e espécies isolados de raízes de bananaeira ‘Prata-Anã’ utilizando a técnica rep-PCR..................................................
47
10 Distância genética entre os isolados da mesma área onde foram coletadas raízes de bananeira ‘Prata-Anã’ utilizando os dados da técnica ARDRA.....................................................................................................
53
11 Agrupamento dos gêneros e espécies isolados de raízes de bananeira ‘Prata-Anã’ utilizando a técnica ARDRA.................................
55
12 Número de isolados e identificação de gêneros e espécies de bactérias endofíticas que amplificaram o gene nifH por área de coleta............................................................................................................
58

FIGURAS
01 Mapa indicando os municipios pertencentes ao Estado de Minas Gerais onde foram realizadas as coleta das raízes de bananeira ‘Prata-Anã’........................................................................................................................
19
02 Mapa indicando o municipio pertencente ao Estado da Bahia onde foi realizada a coleta das raízes de bananeira ‘Prata-Anã’.............................................................
19
03 Infiltração de suspensão de 108 células bacterianas para determinação da reação de hipersensibilidade (A) e sintomas característicos de reação hipersensibilidade positiva em folha de fumo (B)....................................................
23
04 Variação na coloração e na forma das bactérias isoladas de raízes de bananeira ‘Prata-Anã’...............................................................................................................
30
05 Número de unidades formadoras de colônias bacterianas (UFC/g de raiz) de bactérias endofíticas de raiz de bananeira ‘Prata-Anã’ de diferentes áreas..............
31
06 Padrão de coloração de Gram negativa (A) e Gram positiva (B) das bactérias endofíticas isoladas de bananeira ‘Prata-Anã’.........................................................
32
07 Reação de hipersensibilidade negativa em plantas de pimentão (A), feijão (B) e fumo (C)...................................................................................................................
32
08 Gel de agarose contendo produto da amplificação do DNA de isolados de bactérias endofíticas pelo iniciador BOX I. (M) marcador de peso molecular 100 pb; 6 a 146: codigos dos isolados.............................................................................
42
09 Gel de agarose contendo produto da amplificação do DNA de isolados de bactérias endofíticas pelos iniciadores ERIC I e ERIC 2. (M) marcador de peso molecular 100 pb; 18 a 199; 141 a 162: códigos dos isolados.....................................................................................................................
43
10 Gel de agarose contendo produto da amplificação do DNA de isolados de bactérias endofíticas pelos iniciadores REP I e REP 2. (M) marcador de peso molecular 100 pb; 38 a 56: código dos isolados...................................................
44
11 Dendograma de dissimilaridade obtido através da técnica rep-PCR, dos isolados de bactérias endofíticas provenientes de raízes de bananeira ‘Prata-Anã’.........................................................................................................................
46
12 Perfil de restrição da região 16S DNAr dos isolados bacterianos isolados de bananeira ‘Prata-Anã’ após a digestão com a enzima RsaI ...................................
49
13 Perfil de restrição da região 16S DNAr dos isolados bacterianos isolados de bananeira ‘Prata-Anã’ após a digestão com a enzima MspI ..................................
50

14 Perfil de restrição da região 16S DNAr dos isolados bacterianos isolados de bananeira ‘Prata-Anã’ após a digestão com a enzima HinfI.....................................
51
15 Perfil de restrição da região 16S DNAr dos isolados bacterianos isolados de bananeira ‘Prata-Anã’ após a digestão com a enzima NdeI.....................................
52
16 Dendograma de similaridade obtido através da técnica ARDRA dos isolados de bactérias endofíticas provenientes de bananeira ‘Prata-Anã’...............................
54
17 Amplificação do gene nifH de 18 bactérias endofíticas de raízes de bananeira ‘Prata-Anã’. (M) marcador de peso molecular 100 pb; 4 a 67: códigos dos isolados....................................................................................................................
57

i
RESUMO
SOUZA, Suzane Ariádina. Diversidade de bactérias endofíticas em raízes de bananeira ‘Prata-Anã’. 2011. 101 p. Dissertação (Mestrado Produção Vegetal no Semiárido)- Universidade Estadual de Montes Claros, Janaúba- MG1 A bananeira (Musa spp.) é uma das fruteiras mais cultivadas em todo território nacional. Esta cultura, assim como outras plantas, está associada a microrganismos, que podem trazer benefícios às mesmas. Dentre estes, estão às bactérias endofíticas, que habitam o interior de tecidos e órgãos vegetais sem causar prejuízos ao seu hospedeiro e sem produzir estruturas externas emergindo dos tecidos vegetais. A diversidade de microrganismos endófitos tem sido avaliada em muitas plantas, mas em banana, pouco se sabe sobre esta relação. Os objetivos do presente trabalho foram acessar a diversidade genética e identificar em nível de espécie as bactérias endofíticas presentes em raízes de bananeira ‘Prata-Anã’, por meio do uso de marcadores moleculares. O trabalho foi conduzido nos laboratórios de Fitopatologia e Biotecnologia do Campus Janaúba da Universidade Estadual de Montes Claros. Foram utilizados fragmentos de raízes de banana ‘Prata-Anã’, provenientes de quatro municípios do Norte de Minas Gerais e um da Bahia. Foi realizada uma desinfestação das raízes para isolamento das bactérias endofíticas. Os isolados bacterianos obtidos foram caracterizados com relação à resposta de Gram, e a patogenicidade foi testada pela reação de hipersensibilidade em plantas não hospedeiras, sendo usadas mudas de pimentão, feijão e fumo. A diversidade genética foi verificada utilizando as técnicas rep-PCR e ARDRA. Foram selecionados isolados pela presença do gene nifH. A identificação em níveis de gênero e espécie foi realizada por meio do sequenciamento parcial da região rDNA 16S. Foram isoladas 201 bactérias endofíticas, destas 151 foram classificadas como Gram-positivas e 50 como Gram-negativas. Nenhuma apresentou reação de hipersensibilidade nas plantas testadas. A técnica rep-PCR gerou, para cada primer: REP, ERIC e BOX, separadamente, perfis moleculares diferentes. Foram obtidos no total 50 locus passíveis de leitura, sendo que todos os fragmentos mostraram-se polimórficos. A análise de diversidade dos isolados por meio da técnica ARDRA, obtida pelos produtos de clivagem com as quatro enzimas de restrição, produziu 45 bandas polimórficas e nenhuma monomórfica. A RsaI foi a enzima que apresentou maior poder discriminatório, gerando 13 bandas diferentes, seguido de MspI com 12, HinfI com 11 e NdeI com 9 bandas.
1 Comitê de Orientação: Profª Silvia Nietsche- DCA/UNIMONTES (Orientadora); Profª Márcia Regina Costa - DCA/UNIMONTES (Coorientadora); Profª Adelica Aparecida Xavier - DCA/UNIMONTES; Pesquisadora Luciana Nogueira Londe - EPAMIG.

ii
A amplificação por PCR do gene nifH foi verificada em 40 isolados. Após o sequenciamento da região rDNA 16S dos 201 isolados bacterianos, foram obtidas 143 sequências com boa qualidade. A análise das sequências revelou que os isolados estão distribuídos em 17 gêneros bacterianos: Bacillus, Agrobacterium, Klebsiella, Paenibacillus, Lysinibacillus, Streptomyces, Artrobacter, Micrococus, Rhizobium, Sporolactobacillus, Acetobacter, Aneurinibacillus, Pantoea, Brevibacillus, Stenotrophomonas, Pseudomonas e Enterobacter, os quais incluem 16 espécies diferentes. O gênero Bacillus foi o que apresentou maior número de espécies isoladas. As bactérias identificadas nesse trabalho têm potencial para promoção de crescimento, solubilização de fosfato, controle biológico e fixação de nitrogênio. Palavras-chave: Musa spp., Bactérias endofíticas, rep-PCR, ARDRA, Bacillus sp.

iii
ABSTRACT
SOUZA, Suzane Ariádina. Diversity of endophytic bactéria in roots of ‘Prata-Anã’ banana. 2011. 101 p. Dissertation (Master’s degree in Plant Production in the Semi arid)- Universidade Estadual de Montes Claros, Janaúba- MG2 Banana (Musa spp.) is one of the most cultivated fruit trees in the country. That culture, as well as other plants, is associated with microorganisms that can benefit them. Among these microorganisms, are the endophytic bacteria, which inhabit the interior of plant tissues and organs without causing damage to their host and without producing external structures emerging from plant tissues. The diversity of endophytes microorganisms has been evaluated in many plants, but in bananas little is known about this relationship. The objectives of this study were to access the genetic diversity and identify at the species level endophytic bacteria present in roots of 'Prata-Anã' banana, through the use of molecular markers. The study was carried out in the laboratory of Plant Pathology and Biotechnology Campus Janaúba Universidade Estadual de Montes Claros. Fragments of roots of ‘Prata-Anã’ banana from four municipalities in the north of Minas Gerais and Bahia were used. It was accomplished a root desifestation in order to isolate the endophytic bacteria. The bacterial isolates were characterized as for the response of Gram, and pathogenicity was tested by the hypersensitivity reaction in non-host plants, being used red pepper, common beans and tobacco plants. The genetic diversity was verified using the ARDRA and rep-PCR techniques. Isolates were selected by the presence of the nifH gene. The identification at the genus and species levels was performed by partial sequencing of 16S rDNA. They were isolated 201 endophytic bacteria, of these 151 were classified as Gram positive and 50 as Gram negative. None showed hypersensitivity reaction in the tested plants. The rep-PCR technique generated for each primer: REP, ERIC and BOX, separately, different molecular profiles. We obtained a total of 50 readable loci, and all fragments were shown to be polymorphic. The diversity analysis of isolates by ARDRA technique, obtained through cleavage products with four restriction enzymes, produced 45 polymorphic bands and no monomorphic. The RsaI was the enzyme which presented the highest discriminatory power, generating 13 different bands, followed by MspI with 12, HinfI with 11 bands and NdeI with 9 bands. Theamplification of the nifH gene by PCR was detected in 40 isolates. After the
1 Guidance Committee: Profª Silvia Nietsche- DCA/UNIMONTES (Adviser); Profª Márcia Regina Costa - DCA/UNIMONTES (Co-Adviser); Profª Adelica Aparecida Xavier - DCA/UNIMONTES; Pesquisadora Luciana Nogueira Londe - EPAMIG.

iv
sequencing of the 16S rDNA region from 201 bacterial isolates, were obtained 143 sequences with good quality. The sequence analysis revealed that the isolates are distributed among 17 bacterial genera: Bacillus, Agrobacterium, Klebsiella, Paenibacillus, Lysinibacillus, Streptomyces, Artrobacter, Micrococus, Rhizobium, Sporolactobacillus, Acetobacter, Aneurinibacillus, Pantoea, Brevibacillus, Stenotrophomonas, Pseudomonas and Enterobacter, which include 16 different species. The genus Bacillus showed the greatest number of isolate species. The bacteria identified in this study have potential for providing growth, phosphate solubilization, biological control and nitrogen fixation. Keywords: Musa spp., Endophytic bacteria, rep-PCR, ARDRA, Bacillus sp..

1
1 INTRODUÇÃO
A bananeira (Musa spp.) é uma das fruteiras mais cultivadas em todo
território nacional. O Brasil ocupa a quarta colocação no ranking mundial, tendo
uma produção de 7.116.808 toneladas e área colhida de 510.825 mil hectares. O
Estado de Minas Gerais destaca-se como quarto maior produtor do país, com
uma produção em 2010 de 654.861 toneladas (AGRIANUAL, 2011). A região
Norte de Minas é responsável por mais de um terço da produção estadual com,
aproximadamente, 241mil toneladas em uma área de 11.600 hectares. Os
maiores produtores da fruta são os municípios de Jaíba, Janaúba, Matias
Cardoso, Nova Porteirinha e Varzelândia (ABANORTE, 2008).
O Norte de Minas Gerais é reconhecido como o maior produtor mundial
da banana do grupo Prata, uma vez que, segundo os dados da Federação da
Agricultura de Minas Gerais - FAEMG, dos 11,6 mil hectares de área plantada
com banana, 80% são da cultivar Prata-Anã (ABANORTE, 2008).
Observa-se que as plantas, incluindo a bananeira, são consideradas um
microecossistema complexo, onde diferentes nichos são explorados por uma
extensa variedade de microrganismos, dentre eles os endofíticos (AZEVEDO et
al., 2000). São considerados endofíticos todos aqueles microrganismos que
podem ou não crescer em meios de cultura, ou seja, cultivados ou não, e que
habitam o interior de tecidos e órgãos vegetais sem causar prejuízos ao seu
hospedeiro e sem produzir estruturas externas emergindo dos tecidos vegetais
(AZEVEDO e ARAÚJO, 2007). Em geral, são representados por fungos e
bactérias, que desempenham funções importantes no processo de adaptação da
planta com o meio ambiente (MENDES e AZEVEDO, 2007).
Com o aumento de informações sobre interação planta/microrganismos e
com a determinação das diferentes funções destes no interior dos vegetais, tem

2
sido dada atenção ao estudo dos mesmos, já que podem atuar no controle
biológico de inúmeras doenças, na promoção de crescimento vegetal
(HALLMANN et al., 1997; HALLMANN e BERG, 2006; RYAN et al., 2008) e
na biorremediação de áreas poluídas (NEWMAN e REYNOLDS, 2005;
GERMAINE et al., 2009). Além disso, o uso desses microrganismos pode ser
preferível ao uso de fertilizantes químicos e aos de pesticidas, não somente
devido ao menor custo, mas por contribuir com um sistema agrícola sustentável.
Dentre os microrganismos endofíticos, têm se destacado as bactérias, já
que elas estão entre os principais e mais abundantes grupos de microrganismos
que interagem com as plantas. O fato de elas serem capazes de colonizar os
tecidos internos das plantas, constitui-se em vantagens sobre outros
microrganismos que habitam partes externas, pois podem sobreviver em um
ambiente mais uniforme, além de serem menos afetadas pela temperatura,
potencial osmótico e radiação ultravioleta (LODEWYCKX et al., 2002).
Os microrganismos endofíticos têm sido isolados de raízes, nódulos,
caule, folhas e frutos em uma extensa variedade de plantas, incluindo muitas de
interesse agrícola, tais como, cana-de-açúcar (CAVALCANTE e
DOBEREINER, 1988), milho (ARAUJO et al., 2000), videira (BELL et al.,
1995), algodão (QUADT-HALLMANN et al., 1997), arroz (STOLZFUS et al.,
1997), tomate (PILLAY e NOWAK, 1997), citros (MARCON, 2002), batata
(REITER et al., 2003), trigo e sorgo (ZINNIEL et al., 2002), entre outras.
Tradicionalmente, o estudo de endofíticos foi baseado em técnicas de
isolamento em meios de cultura. Entretanto, nos últimos anos, com o avanço das
técnicas moleculares, tem sido possível investigar aqueles que não são
cultiváveis ou de natureza fastidiosa, possibilitando o estudo do material
genético a partir de amostras coletadas no ambiente (ASSUMPÇÃO et al.,
2009).

3
As técnicas moleculares têm sido utilizadas amplamente em estudos de
caracterização de bactérias endofiticas tais como, a Rep-PCR utilizando as
sequências conservadas ERIC, REP e BOX, a amplificação e restrição do rDNA
16S, bem como a clonagem e sequenciamento desse gene (LOUWS et al., 1994;
TORRES, 2005; RYAN et al., 2008). Além disso, os primers que permitem a
amplificação específica de genes codificando importantes características das
bactérias, como o gene nifH, necessários para a fixação de nitrogênio ou de
genes envolvidos na degradação de poluentes orgânicos, são utilizados para
estudar o potencial da população endofítica que participam de processos
importantes dentro da planta hospedeira (RYAN et al., 2008).
Vários trabalhos têm sido realizados com o objetivo de isolar, identificar
e estudar a diversidade de bactérias endofíticas nos tecidos de várias espécies de
plantas e suas interações (FERREIRA et al., 2008; VIDEIRA et al., 2009; LI et
al., 2010; MAGNANI et al., 2010). Há poucos relatos na literatura sobre a
caracterização de bactérias endofíticas na cultura da bananeira. As espécies:
Burkolderia cepacia (Palleroni and Holmes 1981), Azospirillum brasilense
(Tarrand et al., 1979), Azospirillum amazonense (Magalhaes et al., 1984),
Burkholderia sp. (Yabuuchi et al. 1993 emend. Gillis et al. 1995), Citrobacter
sp.( Werkman and Gillen 1932), Klebsiella variicola (Rosenblueth et al. 2004)
foram citadas por Rosenblueth e Martínez-Romero (2006) como colonizadoras
de bananeira. Já Ganem et al. (2009), trabalhando com explantes de bananeiras
tetraploides cultivares Tropical e Galil, identificaram quinze gêneros de
bactérias, destes, o gênero Klebsiella sp.( Trevisan 1885 emend. Carter et al.
1999, emend. Drancourt et al. 2001) apresentou maior frequência.
Assim, os principais objetivos do presente trabalho foram acessar a
diversidade genética e identificar em nível de espécie as bactérias endofíticas
presentes em raízes de bananeira ‘Prata-Anã’, por meio do uso de marcadores
moleculares.

4
2 REFERENCIAL TEÓRICO
A banana (Musa spp.) é explorada na maioria dos países tropicais com
produção mundial de 90,7 milhões de toneladas (FAO, 2010). Segundo dados do
Agrianual (2011), o Brasil está entre os maiores produtores mundial de banana.
Dentre os estados produtores, destaca-se Bahia, São Paulo, Santa Catarina,
Minas Gerais e Pará. Minas Gerais é quarto maior produtor, tendo em 2010, um
total de 40.373 hectares de área colhida dessa cultura. Entre as regiões mineiras,
o Norte de Minas destaca-se como maior produtora, alcançando em 2009 uma
produção de 241 mil toneladas em uma área de 11.600 hectares (ABANORTE,
2008).
Segundo Souto et al. (1997), a cultura da banana foi introduzida na
região do Norte de Minas no início da década de 80, onde os primeiros plantios
ocorreram pelos produtores do Perímetro Irrigado do Gorutuba, situado em
Porteirinha-MG, com a variedade Nanicão, que sofreu intenso ataque de
nematoides, comprometendo a sua viabilidade na região. Essa variedade foi
sendo substituída gradativamente pela ‘Prata-Anã’, devido a sua aceitação,
melhor remuneração e qualidade dos frutos. Atualmente o Norte de Minas
Gerais é o maior produtor mundial da banana 'Prata-Anã' (SEAPA, 2008).
A localização da região Norte-Mineira é favorável com relação à
distância dos mercados consumidores, pois se encontra entre os da região
Sudeste e Sul e também próximo aos da região Nordeste do Brasil, tendo o
escoamento da produção facilitado. Associadas a isto a ampliação do nível
tecnológico no sistema produtivo e a organização dos produtores tem permitido
a expansão para novos mercados, inclusive o internacional (ALMEIDA et al.,
2000).
Embora exista um número expressivo de variedades de banana no Brasil,
quando se consideram aspectos como preferência dos consumidores,

5
produtividade, porte, tolerância a pragas e doenças, resistência à seca, e ao frio,
restam poucas cultivares com potencial agronômico para serem usadas
comercialmente. As cultivares mais difundidas no Brasil são : Prata, Prata-Anã,
Pacovan (responsáveis por aproximadamente 60% da área cultivada com banana
no Brasil), do grupo AAB, e Nanica, Nanicão e Grande Naine, do grupo AAA,
utilizadas principalmente na exportação (OLIVEIRA, 1999).
A banana ‘Prata-Anã’ (Musa acuminata x Musa balbisiana), proveniente
de uma mutação da cultivar 'Branca' ocorrida no início do século no estado de
Santa Catarina, é uma das mais cultivadas na região Norte-Mineira.Essa cultivar
possui fruto de seção pentagonal, com cinco quinas bem visíveis quando verde,
de tamanho médio, com 10 a 13 cm de comprimento e 3,5 a 4,0 cm de diâmetro,
com casca fina, de cor amarelo-ouro e endocarpo de cor creme-róseo pálido
(MASCARENHAS, 1999). A cultivar é tolerante ao frio, e dispensa o uso de
escoramento, devido ao grande vigor da planta (SILVA et al., 1999).
A prevalência do cultivo de bananeiras tipo Prata, com destaque para a
'Prata-Anã', evidencia a tradição de seu cultivo e a sua aceitação comercial. É
uma cultivar exigente em nutrientes, não só por produzir grande massa
vegetativa, mas também por apresentar elevadas quantidades de elementos
absorvidos pela planta e exportados pelos frutos (SILVA et al., 1999). Além
disso, é susceptível à sigatoka-amarela, à sigatoka-negra e ao mal-do-Panamá
(DONATO et al., 2009) e, moderadamente, à broca-da-bananeira (ALVES,
2001).
Assim, nas últimas décadas, tem se investido muito em pesquisas para
disponibilizar formas alternativas de nutrientes às plantas, e em metodologias
alternativas de controle de patógenos, com enfoque no uso de microrganismos
endofíticos (KUSS et al., 2007; BARROSO e NAHAS, 2008; RYAN et al.,
2008; LUCON et al., 2009).

6
O termo ‘Endófito’ (Endophyte) tem origem Grega, em que ‘endon’
significa ‘de dentro’ e ‘phyte’ significa ‘planta’ (CARROLL, 1988). Os
microrganismos endofíticos são todos aqueles que habitam o interior de tecidos
e órgãos vegetais sem causar prejuízos ao seu hospedeiro e sem produzir
estruturas externas, emergindo dos tecidos vegetais (AZEVEDO e ARAÚJO,
2007). São divididos em dois tipos: Tipo I, que não produzem estruturas
externas à planta e Tipo II, que produzem estruturas externas à planta
(MENDES e AZEVEDO, 2007). São diferentes dos microrganismos
patogênicos, que causam doenças nas plantas, e dos epifíticos que vivem na
superfície dos vegetais. No entanto, essa distinção entre microrganismos
endofíticos, patogênicos e epifíticos é complicada, já que um microrganismo
endofítico pode se tornar patogênico conforme o genótipo do hospedeiro e as
condições de ambiente, e um epifítico pode, eventualmente, entrar em uma
planta e lá permanecer por um determinado período, causando ou não danos à
mesma (AZEVEDO, 1998).
Um dos primeiros trabalhos com bactérias endofíticas foi realizado por
Colombo (1978), no qual relatou a ocorrência de bactérias endofíticas no talo de
algas, entre sifões e dentro dos filamentos cenocíticos. O autor sugeriu a
ocorrência de um equilíbrio fisiológico entre a bactéria e o seu hospedeiro.
Também supôs que o fato dos endofíticos se encontrarem próximo aos
cloroplastos e nas zonas mais jovens dos talos, indicaria uma ligação com a
atividade fotossintética da alga e sua provável dependência de oxigênio.
A partir dos anos 80, os trabalhos se tornaram mais frequentes. Jacobs et
al. (1985), empregando métodos e técnicas mais aperfeiçoadas de estudo e
análise, enumeraram, localizaram e caracterizaram bactérias endofíticas de
raízes de beterraba utilizando técnicas de imunológia e de microscopia
eletrônica de varredura. Os autores observaram um aumento na população de

7
bactérias nas regiões centrais e periféricas da raiz, e sugeriram que as regiões de
maior ocorrência constituem uma rota de colonização das bactérias.
As bactérias endofíticas têm sido alvo de vários estudos, com diferentes
culturas, uma vez que, segundo Misaghi e Donndelinger (1990), através de
processos coevolutivos elas conseguiram manter uma relação bastante íntima
com seu hospedeiro. Essas bactérias possuem também a habilidade de se manter
dentro dos tecidos vegetais isentos da competição microbiana, o que é uma
vantagem ecológica sobre as bactérias epifíticas. Essa característica pode
acrescentar uma vantagem no aproveitamento desses microrganismos em
aplicações biotecnológicas.
Apesar da ocorrência de microrganismos endofíticos, em diferentes
plantas, pouco se sabe sobre sua identidade, diversidade e níveis populacionais
nos diferentes tecidos. Acredita-se que de cerca de 300 000 espécies de plantas
que existem na Terra, cada planta individual é hospedeira de um ou mais
endófitos (STROBEL et al., 2004), principalmente por bactérias, uma vez que
elas constituem um dos grupos mais diversos no planeta e podem compreender
mais de um milhão de espécies (KENNEDY, 1999). Vários relatos indicam que
essas bactérias existem em uma variedade de tecidos em numerosas espécies de
plantas, sugerindo uma existência onipresente na maioria, se não em todos os
vegetais superiores (LODEWYCKX et al., 2002). Apesar dessa grande
diversidade, estima-se que tenhamos conhecimento de apenas uma pequena
fração das bactérias existentes (TORSVIK et al., 2002). Muitos fatores, tais
como rotação de culturas, tipos de solo e os fitopatógenos, são conhecidos por
influenciarem a estrutura da população de bactérias endofíticas (GRANER et al.,
2003).
Acredita-se que as bactérias endofíticas se dispersam nas plantas através
das sementes, propagação vegetativa, implementos agrícolas, ventos ou insetos
(BALDANI, 1997). Uma vez presentes no solo, elas penetram na planta, via

8
zona radicular, através de aberturas que ocorrem naturalmente como resultado
do crescimento das plantas ou através de pelos radiculares (SPRENT e de
FARIA, 1988). Além de proporcionar vias de entrada, essas aberturas favorecem
a exsudação radicular e estimulam o processo de quimiotaxia e permitem a
penetração direta das bactérias nas raízes. Além disso, tais substâncias
constituem fonte de nutrientes na região da rizosfera (LODEWYCKX et al.,
2002).
A maneira de penetração também pode ser pelas aberturas naturais,
estômatos, lenticelas, hidatódios, ou por injúrias causadas por insetos e
maquinário agrícola. Além disso, essas bactérias são capazes de degradar a
parede celular das plantas pela liberação de enzimas hidrolíticas como celulases
e pectinases, sendo essa uma possível forma de penetração. Embora observações
demonstrem a possibilidade de mecanismos de penetração ativa de algumas
bactérias endofíticas, muito pouco se sabe sobre a origem e regulação dessas
enzimas (LODEWYCKX et al., 2002). Uma maneira diferente de penetração é
descrito por Ashbolt e Inkerman (1990) em cana-de-açúcar através da associação
com cochonilha, e por Kluepfel (1993) via diferentes insetos.
Então, pode-se dizer que a colonização endofítica pode ser
intracelularmente e limitada ao interior de algumas células, intercelularmente
localizada, e intra e intercelular, ao mesmo tempo, de maneira sistêmica
(STONE et al., 2000). Os diferentes mecanismos de distribuição das bactérias
endofíticas podem estar relacionados com interações com outras bactérias ou
com os diferentes mecanismos de cada microrganismo, que lhes permitem
habitar vários nichos, representado pelos tecidos e, mais especificamente, pelos
espaços intercelulares no interior de cada tecido (DI FIORI e DEL GALLO,
1995).
Mesmo colonizando sistemicamente a planta, as bactérias endofíticas
apresentam preferência de colonização por certos tecidos. Kuklinsky-Sobral et

9
al. (2004) observaram, em soja, que a densidade e a diversidade dessas bactérias
variam de acordo com o tecido, fase de desenvolvimento da planta, mudanças
sazonais e genótipo do hospedeiro. Os autores constataram que a densidade
bacteriana foi maior na raiz e menor em folhas nessa cultura. Fisher et al. (1992)
isolaram bactérias e fungos endofíticos de três tipos de tecidos (epiderme, córtex
do caule e folha) de plantas de milho sadias, e verificaram que as partes das
plantas mais próximas do solo eram mais colonizadas por bactérias que a parte
superior das plantas.
Em termos quantitativos os estudos indicam que o tecido radicular é o
mais colonizado por bactérias endofíticas. O fato de a colonização ser abundante
no tecido radicular pode refletir uma consequência da raiz ser o sítio primário de
entrada dos endofíticos nas plantas. Além do mais, isso pode explicar a estreita
relação entre rizobactérias e bactérias endofíticas (LODEWYCKX et al., 2002).
De acordo com Germida et al. (1998), a comunidade de bactérias endofíticas é
um subconjunto da comunidade do rizoplano.
Várias são as razões que despertam o interesse pelo potencial
biotecnológico das bactérias endofíticas, tais como: íntima associação com a
planta hospedeira, fixação de nitrogênio, indução de resistência à condição de
estresse, mudanças nas condições fisiológicas da planta, produção de enzimas,
drogas de interesse farmacológico e produção de reguladores de crescimento
vegetal (LODEWYCKX et al., 2002). De fato, inúmeros relatórios mostraram
que as bactérias endofíticas são capazes de controlar patógenos em plantas
(STURZ e MATHESON, 1996; DUIJFF et al., 1997; KRISHNAMURTHY e
GNANAMANICKAM, 1997), insetos (AZEVEDO et al., 2000) e nematoides
(HALLMAN et al., 1997, 1998). Em alguns casos, podem também acelerar a
emergência das plântulas, promover o estabelecimento de plantas em condições
adversas (CHANWAY, 1997) e auxiliar no crescimento do vegetal (BENT e
CHANWAY, 1998).

10
São muitos os trabalhos de isolamento de bactérias em culturas com
interesse econômico. Li et al. (2008) isolaram bactérias endofíticas de nódulos
de soja dos gêneros Pantoea spp. (Gavini et al. 1989 emend. Mergaert et al.
1993 emend. Brady et al., 2010), Serratia spp. (Bizio, 1823), Acinetobacter spp.
(Brisou and Prevot 1954), Bacillus spp.(Cohn, 1872), Agrobacterium spp.(
Conn, 1942) e Burkholderia spp.. Germida et al. (1998) e Misko e Germida
(2002) encontraram uma alta proporção de endofíticos Bacillus sp.,
Flavobacterium sp.( Bergey et al., 1923 emend. Bernardet et al. 1996),
Micrococcus sp.( Cohn 1872 emend. Stackebrandt et al., 1995 emend. WIESER
et al., 2002), Pseudomonas sp. (Migula, 1894) e Rathayibacter sp.( Zgurskaya et
al., 1993) em raízes de canola e trigo. Magnani et al. (2010) isolaram 32
bactérias endofíticas de cana-de-açúcar, dessas 18 foram extraídas do caule e 14
das folhas.
Entretanto, há poucos relatos na literatura sobre o isolamento das
bactérias endofíticas na cultura da bananeira. Weber e Freire (2003) detectaram
a presença de bactérias diazotróficas em todas as amostras de raízes e de
pseudocaules de bananeiras coletados nas diferentes localidades de
microrregiões serranas do Estado do Ceará. Populações semelhantes foram
observadas por Weber (1998) em amostras de raízes e pseudocaules de
bananeiras provenientes do Rio de Janeiro e da Bahia.
Thomas et al. (2008), avaliando a comunidade endofítica em banana
cultivar Grand Naine, identificaram isolados dos gêneros Enterobacter
(Hormaeche and Edwards, 1960), Klebsiella, Ochrobactrum (Holmes et al.,
1988), Bacillus, Pantoea (Gavini et al., 1989 emend. Mergaert et al., 1993
emend. Brady et al., 2010) ou Staphylococcus (Rosenbach, 1884). Já Ganem et
al. (2009), trabalhando com explantes de bananeiras tetraploides, registraram
quinze bactérias, oito provenientes da cultivar ‘Tropical’ e sete da cultivar
‘Galil’, pertencentes a seis gêneros diferentes. Identificaram-se um isolado como

11
Erwinia chrysanthemi (Burkholder et al., 1953), seis como Klebsiella
pneumoniae pneumoniae (Schroeter 1886; Trevisan 1887), um como
Pseudomonas huttiensis (Leifson, 1962), um como Enterobacter gergoviae
(Brenner et al., 1980), um como Enterobacter cloacae (Jordan 1890)
Hormaeche and Edwards 1960, um como Paenibacillus polymyxa (Prazmowski
1880) (Ash et al., 1994) e um como Salmonella spp.( Lignieres, 1900).
O emprego de técnicas moleculares tornou-se possível a partir dos
estudos pioneiros de Pace et al. (1986), em análises de estrutura de comunidades
microbianas utilizando as informações da sequência de nucleotídeos do gene da
subunidade 16S do RNA ribossômico (16S rDNA). Este gene tem sido muito
utilizado por estar presente em todas as bactérias, derivar de um ancestral
comum, além de ser geneticamente estável e apresentar um tamanho suficiente
para análises filogenéticas (CHENEBY et al., 2000). O gene 16S rRNA tem sido
largamente utilizado para estudos taxonômicos, pois permite a análise de
sequências tanto estreitamente relacionadas quanto entre microrganismos mais
distantes (STACKEBRANDT e GOEBEL, 1994). Nos dias atuais, vários
estudos têm sido realizados utilizando técnicas moleculares, com o intuito de
identificar e avaliar a diversidade e a filogenia de microrganismos presentes nos
diferentes ambientes.
Dentre as técnicas moleculares, destacam-se a análise de rep-PCR, que
permite a amplificação de sequências definidas da molécula de DNA, e o
método de polimorfismo no comprimento de fragmentos de restrição, ARDRA
(Amplified Ribosomal DNA Restriction Analysis) (CHUEIRE et al., 2000).
Várias técnicas utilizam a PCR como base para estudos de diversidade
de microrganismos. A rep-PCR (PCR- repetitivo) emprega a PCR para
relacionar o genótipo dos organismos, sendo usada em fingerprints genômicos
com a utilização de primers correspondentes a regiões repetitivas (rep) ao longo
do genoma (KOEUTH et al., 1995). Esses elementos repetitivos no DNA são

12
observados em bactérias, e servem como sítios para primers na amplificação do
DNA genômico (De BRUJIN, 1992). As bandas amplificadas no rep-PCR são
separadas de acordo com seu tamanho em gel de agarose, e geram um perfil que
serve como uma impressão digital específica de cada estirpe bacteriana
(LUPSKI, 1993).
Famílias de sequências repetitivas intergênicas têm sido encontradas em
procariotos e foram primeiramente identificadas em bactérias entéricas como
Escherichia coli. Três famílias de regiões repetitivas já foram descritas. São os
elementos REP (repetitive extragenic palindromic) de 35-40 pb, ERIC
(enterobacterial repetitive intergenic consensus) de 124-127 pb e o elemento
BOX de 145 pb (VERSALOVIC et al., 1991). O uso de rep-PCR tem a
vantagem de que os primers possuem sequências conhecidas e já foram
utilizados em estudos com várias bactérias Gram-negativas e Gram-positivas.
O primer BOX gera padrões de banda complexos e robustos, enquanto o
REP gera um menor nível de complexidade, porém produz padrões de bandas
reproduzíveis e diferenciadoras. O primer ERIC é mais sensível a condições
subótimas de PCR, como contaminantes na preparação do DNA; entretanto, gera
padrões altamente discriminatórios (RADEMAKER e DE BRUJIN, 1997;
RADEMAKER et al., 1998).
A técnica molecular que emprega os elementos BOX-PCR reúne várias
vantagens, uma vez que é uma técnica rápida, de execução fácil e altamente
discriminatória para espécies, produz resultados que representam bem as
análises baseadas na homologia DNA-DNA (De BRUJIN, 1992).
Esta técnica vem sendo muito utilizada para avaliar diversidade genética
de populações microbianas. Ela foi usada por Rademaker et al. (2000) para
estudar diversidade genética em população de Xanthomonas spp.(Dowson 1939
emend. Vauterin et al., 1995). Goris et al. (2003) empregaram BOX-PCR para
avaliar diversidade genética entre linhagens bacterianas que receberam o gene

13
gfp. Fernandes et al. (2003), em estudo de caracterização de rizóbios nativos em
culturas dos feijões guandu e caupi, verificaram um elevado grau de diversidade
genética entre as estirpes pela técnica BOX-PCR, mostrando eficiência para
diferenciar espécie, mas não para a diferenciação de estirpe de rizóbios.
A técnica aplicada ao estudo da diversidade microbiana, denominada
ARDRA, consiste na amplificação e posterior digestão do rDNA com enzimas
de restrição. Este método é baseado no princípio de que os sítios de restrição no
rDNA são conservados de acordo com padrões filogenéticos. Dessa forma, pode
ser utilizado o gene 16S rDNA para o estudo de grupos heterogêneos, ou a
região espaçadora entre o gene 16S e o 23S rDNA para o estudo de grupos
muito similares (RANJARD et al., 2000).
Os ácidos ribonucleicos ribossomais (rRNA) são considerados os
biopolímeros mais adequados para estudos de diversidade. Seus genes são
universalmente distribuídos, cuja molécula possui a maior conservação. Sua
variabilidade pode apresentar-se em maior ou menor extensão em diferentes
regiões da molécula (LANE et al., 1985). O valor desse método está na sua
rapidez e habilidade para avaliar diferenças entre grupos filogenéticos, efetuando
análises em vários níveis de classificação, inclusive em estudos de evolução,
gerando novos marcadores para estudos de genética de populações (ABD-
ELSALAM et al., 2003).
Recomenda-se cuidado na escolha do fragmento de rDNA a ser
amplificado e analisado. No caso da análise de diversidade de grupos de
indivíduos com elevada afinidade genética, o fragmento amplificado deve incluir
o espaço intergênico 16S-23S rDNA. Essa região intergênica apresenta maior
variabilidade não só na sua composição de bases como também no seu tamanho
ao serem comparadas as regiões gênicas 16S ou 23S (REIS JUNIOR et al.,
2004). O uso de primers específicos é vantajoso sobre aqueles não específicos
porque, em uma única reação, obtém-se um número elevado de bandas,

14
permitindo a detecção de polimorfismo suficientemente elevado para a análise
genética das bactérias (CHUEIRE et al., 2000).
Essa técnica tem sido aplicada para o estudo da diversidade microbiana
associada a vegetais ou a diferentes solos (KUKLINSKY- SOBRAL et al.,
2004), análise da diversidade genética de bactérias degradadoras de pesticidas
(DESAINT et al., 2000), caracterização de bactérias diazotróficas (CRUZ et al.,
2001), em estudos taxonômicos e filogenéticos (RODAS et al., 2003), entre
outros. Além disso, o ARDRA é mais rápido do que o sequenciamento da região
16S rDNA e menos oneroso. A maioria das caracterizações é realizada através
de um pequeno número de enzimas, amplamente utilizadas, e uma interpretação
visual do padrão de restrição (HEYNDRYCKX et al., 1996). Mocali et al.
(2003) pesquisaram a diversidade endofítica em Ulmus spp., utilizando as
técnicas de sequenciamento rDNA 16S e ARDRA, e encontraram representantes
dos gêneros Bacillus spp., Curtobacterium spp.( Yamada and Komagata 1972),
Pseudomonas spp., Stenotrophomonas spp.( Palleroni and Bradbury 1993),
Sphingomonas spp.( Yabuuchi et al., 1990 emend. Takeuchi et al., 1993 emend.
Yabuuchi et al., 1999 emend. Takeuchi et al., 2001 emend. Yabuuchi et al.,
2002 emend. Busse et al., 2003) , Enterobacter spp. e Stahlylococcus spp..
A presença do gene nifH, que codifica a unidade Fe-nitrogenase do
complexo nitrogenase, tem se tornado um marcador muito utilizado no estudo da
diversidade de endofíticos com potencial para fixar N2, em estudos
independentes do cultivo (IZQUIERDO e NUSSLEIN, 2006). Esse gene
apresenta sequências extremamente conservadas, sendo considerado um dos
genes funcionais mais antigos. As relações filogenéticas entre bactérias,
baseadas em divergência na sequência do nifH, apresentam-se de maneira
semelhante ao que ocorre com o 16S rDNA. A vantagem da utilização desse
gene para estudo de populações microbianas no ambiente é que, neste caso, se

15
trata de um gene funcional, o que permite uma correlação entre a estrutura e a
função da comunidade microbiana (ROSADO et al., 1999).
Teixeira et al. (2007), utilizando primers específicos do gene nifH para
os grupos Proteobacteria, Actinobacteria e Bacillus, verificaram que o produto
de amplificação do gene nifH foi observado somente em oito espécies
bacterianas endofíticas, de um total de 47 isolados de mandioca. Destas, algumas
Enterobacter cancerogenus (Urosevic, 1966) Dickey and Zumoff 1988,
Stenotrophomonas maltophilia (Hugh, 1981) Palleroni and Bradbury 1993 e
Pseudomonas fluorescens (Migula, 1895) apresentaram capacidade para
produção de hormônio de crescimento e para fixação de nitrogênio atmosférico,
constatado pela presença do gene nifH.
Cruz et al. (2001) identificaram oito novas bactérias fixadoras de
nitrogênio, além de Herbaspirillum seropedicae (Baldani et al., 1986 emend.
Baldani et al., 1996), H. rubrisubalbicans (Christopher and Edgerton 1930)
Baldani et al., 1996), Burkholderia brasilensis (Baldani et al., 1997) e
Burkholderia tropicalis (Reis et al., 2004), em algumas bactérias isoladas de
banana e abacaxi através da técnica ARDRA, revelando a grande diversidade de
bactérias fixadoras de nitrogênio associadas a essas culturas. Por outro lado,
Assumpção et al. (2009) isolaram bactérias endofíticas de diferentes grupos
morfológicos a partir de sementes de soja e identificaram 62 isolados por meio
do sequenciamento parcial do gene 16S rDNA. Os critérios utilizados para
selecionar os isolados para serem identificados foram morfológicos e baseados
na análise de ARDRA.
O sequenciamento do gene 16S rDNA tem auxiliado nos estudos de
diversidade genética bacteriana, pois permite a identificação das espécies
isoladas. Até 1975, a ideia de sequenciamento de cromossomos inteiros, parecia
improvável. Porém, atualmente inúmeras sequências completas de muitas
espécies já são conhecidas e registradas em bancos genéticos. A descoberta de

16
técnicas que utilizam enzimas de restrição, de clonagem gênica, de PCR e o
aprimoramento dos processos de eletroforese, com máquinas automatizadas de
sequenciamento, têm permitido as análises de DNA em larga escala (SNUSTAD
e SIMMONS, 2008).
Em bactérias, como a região 16S é altamente conservada, o
sequenciamento total ou parcial deste gene tem sido usado em estudos de
filogenia e taxonomia. Chueire et al. (2003) empregaram a técnica de
sequenciamento dessa região, com o intuito de estabelecer relações filogenéticas
de estirpes comerciais recomendadas para as culturas de soja e do feijoeiro no
Brasil. Videira et al. (2009) avaliaram a diversidade de bactérias associadas ao
arroz e registraram espécies fixadoras de nitrogênio através do sequenciamento
do gene 16S rRNA. Resultados de estudos baseados na amplificação e
sequenciamento de fragmentos dos genes de rDNA 16S mostraram que a
diversidade de microrganismos em amostras ambientais é vasta (HUNTER-
CEVERA, 1998).
Até o momento, poucas sequências do genoma de bactérias endofíticas
foram publicadas (RYAN et al., 2008). Porém, várias sequências já estão
depositadas em banco de dados, e servem como ferramenta nos estudos de
diversidade genética. Portanto, as sequências genômicas oferecem informações
valiosas sobre a biologia das bactérias endofíticas, e quanto mais sequências
tornam-se disponíveis, maior número de informações é gerado sobre a
identidade e os mecanismos envolvidos no sucesso de colonização e interação
desses microrganismos com as plantas.

17
3 MATERIAL E MÉTODOS
3.1 Coleta das Raízes e Isolamento das Bactérias
Foram coletadas raízes, em plantas jovens que não haviam emitido
cachos, pertencentes à cultivar Prata-Anã. As coletas foram realizadas em cinco
localidades (TABELA 1, FIGURA 1 e 2), raízes com 10 a 15 cm de
comprimento foram retiradas a 15 cm de profundidade, individualizadas em
saquinhos, identificadas e posteriormente transportadas para o laboratório de
Fitopatologia da UNIMONTES, onde foi realizado o isolamento das bactérias
endofíticas. No momento da coleta aplicou-se um questionário aos produtores
para um prévio conhecimento das condições de manejo do bananal (em anexo).

18
TABELA 1. Localização geográfica das áreas de coleta de raízes de bananeira ‘Prata-Anã’ para o isolamento de bactérias endofíticas.
Área Município Estado Coordenadas geográficas de cada área
1 Janaúba Minas Gerais S15º49’51.4’’; W 43º16’12.8’’; 548 m
2 Janaúba Minas Gerais S 15º50’2.1’’; W43º19’32’’; 503 m
3 Guanambi Bahia S 14º13’30’’; W42º46’53’’; 525 m
4 Porteirinha Minas Gerais S 15º35’31.8’’; W 43º16’24.2’’; 549 m
5 Nova Porteirinha Minas Gerais S 15º41’42.8’’; W43º16’31.4’’; 530 m
6 Nova Porteirinha Minas Gerais S 15º42’25.4’’; W 43º16’18.6’’; 527 m
7 Nova Porteirinha Minas Gerais S 15º45’4.9’’; W 43º15’46.1’’; 561 m
8 Nova Porteirinha Minas Gerais S 15º43’28.3’’; W 43º15’43.7’’; 532 m
9 Nova Porteirinha Minas Gerais S15º46’57.7’’; W 43º16’43’’; 575 m
10 Nova Porteirinha Minas Gerais S 15º46’54.8’’; W 43º16’44.6’’; 552 m
11 Matias Cardoso Minas Gerais S 15º04’53.8’’;W 43º47’50.1’’; 487 m
12 Jaíba Minas Gerais S 15º15’2.9’’; W 43º55’20.5’’; 480 m
13 Jaíba Minas Gerais S 15º05’11’’; W 44º00’57.7’’; 458 m
14 Jaíba Minas Gerais S 15º17’10.8’’; W 43º48’26.1’’; 491 m
15 Jaíba Minas Gerais S 15º14’58.1’’; W 43º47’40.5’’; 478 m
16 Jaíba Minas Gerais S 15º12’49.8’’; W 43º48’36.1’’; 485 m

19
FIGURA 1. Mapa indicando os municípios pertencentes ao Estado de Minas Gerais onde foram realizadas as coletas das raízes de bananeira ‘Prata-Anã’.
FIGURA 2. Mapa indicando o município pertencente ao Estado da Bahia onde
foi realizada a coleta das raízes de bananeira ‘Prata-Anã’.
Matias Cardoso Jaíba
Porteirinha
Nova Porteirinha Janaúba
Guanambi

20
Fragmentos de raízes de bananeira livres de sintomas foram lavadas em
água corrente durante três minutos e, posteriormente, para eliminação dos
microrganismos epifíticos, 1,5 g de fragmentos de raízes foram imersos em
álcool a 70% por 1 minuto, hipoclorito de sódio (NaCLO) a 4,0% por 3 minutos
e, em seguida, foram lavados por três vezes em água destilada e esterilizada.
Posteriormente, os fragmentos foram expostos em câmara de fluxo à luz
ultravioleta durante 10 minutos, transferidos para tubos de ensaio contendo 15
mL de solução salina estéril a 0,85% e submetidos a banho de ultrassom/10 min,
na potência de, aproximadamente, 40 Kz. O procedimento de desinfecção e
banho de ultrassom das raízes foi novamente repetido nas mesmas condições
descritas anteriormente, e os fragmentos foram transferidos para tubos contendo
solução salina estéril a 0,85% e mantidos em banho de ultrassom/10 min, na
potência de aproximadamente 40 Kz. Após o banho, esses fragmentos foram
macerados em 15 mL de solução salina a 0,85% e, novamente submetidos a
banho de ultrassom /10 min.
As suspensões obtidas foram diluídas até 10-1 e, a partir dessa suspensão
diluída, uma alíquota de 0,1 mL foi plaqueada em meio Agar-Nutritivo-
Dextrose-Levedura (10 g de dextrose, 5 g de extrato de levedura, 3 g de extrato
de carne, 5 g de peptona e 18 g de ágar), Batata Dextrose Agar (200 g de batata,
20 g de dextrose e 20 g de Ágar) e “Trypic Soy Agar” (30 g de TSA), com três
repetições cada, efetuando espalhamento com auxílio de alça de Drigalski. Após
48 h de incubação, sob temperatura a 25 ± 1 ºC e fotoperíodo de 12 h, realizou-
se a contagem do número de Unidade Formadora de Colônia por grama de raiz
(UFCs/g). As colônias bem individualizadas, que apresentaram diferenças
principalmente quanto à cor e ao formato, dentro de uma mesma amostra, foram
transferidas para tubos contendo meio de melhor crescimento no isolamento
(MARIANO et al., 2000).

21
Após o segundo banho de ultrassom e antes do maceramento das raízes,
foi realizado um plaqueamento para confirmar a natureza endofítica dos
isolados. Não havendo crescimento bacteriano nessas placas dentro de 48 horas,
os isolados obtidos pela trituração das raízes foram considerados endofíticos
(NAVES et al., 2004).
Os isolados foram armazenados em tubos contendo meio de melhor
crescimento (NYDA, BDA ou TSA), e mantidos em geladeira à temperatura de
4 ºC. Também foram armazenados em eppendorf contendo água mineral
esterelizada e mantidos em temperatura ambiente. Para realização dos ensaios,
os isolados mantidos em geladeira foram multiplicados em meio de melhor
desenvolvimento.
3.2 Caracterização dos isolados bacterianos com relação à resposta de
Gram
Os isolados bacterianos foram caracterizados com relação à resposta de
Gram. Para realizar a reação, adicionou-se uma gota de suspensão de bactéria
diluída, em lâminas flambadas em álcool 92,8% e, com o auxílio de um bastão
de vidro, preparou-se o esfregaço da suspensão bacteriana. Posteriormente, o
esfregaço foi coberto com solução cristal violeta durante um minuto e lavado
rapidamente com água corrente, em seguida coberto com lugol por um minuto e
novamente lavado em água corrente. Após esse período, as lâminas foram
lavadas com álcool etílico por 30 segundos e contrastadas com safranina por um
minuto, e novamente lavadas em água corrente (MARIANO et al., 2000). As
lâminas foram secas ao ar e observadas com objetiva de imersão. As células que
apresentaram coloração violeta foram consideradas Gram-positivas e coloração
avermelhada Gram-negativas.
Para fim de comparação, os isolados foram comparados com as espécies
Enterococcus faecalis e Escherichia coli (isolados gentilmente cedidos pelo

22
laboratório de Microbiologia da Unimontes) caracterizadas como Gram-positiva
e Gram-negativa, respectivamente.
3.3 Teste de patogenicidade por meio da reação de hipersensibilidade
Para a verificação da patogenicidade dos isolados bacterianos, foi
realizado o teste de reação de hipersensibilidade em plantas não hospedeiras da
bactéria, sendo utilizadas mudas de pimentão (Capsicum annuum Mill), fumo
(Nicotiana tabacum L.) e de feijão (Phaseolus vulgaris L.) produzidas em casa
de vegetação. Os isolados bacterianos foram multiplicados em meio TSA por 24
horas. Após esse período, adicionaram-se 20 mL de solução salina a 0,85% de
NaCl estéril às placas e, com o auxílio de uma lâmina de microscopia, as
colônias foram colocadas em suspensão e transferidas para um bécher contendo
água destilada. As suspensões bacterianas obtidas foram filtradas em gaze dupla,
transferidas para tubos de ensaio e ajustadas para 0,1 de absorbância em
espectofotômetro que corresponde, aproximadamente, a 108 células/mL. As
suspensões bacterianas foram infiltradas nas folhas com auxílio de seringas
descartáveis utilizadas para aplicação de insulina, sempre tangenciando as
nervuras até que o local mudasse de cor (FIGURA 3 A). Foram inoculadas três
folhas por planta para cada isolado. Após 24 horas da inoculação, avaliou-se o
aparecimento de possíveis reações de hipersensibilidade nas folhas das plantas
não hospedeiras (ROMEIRO, 2001).
A reação de hipersensibilidade positiva é caracterizada por uma
mudança no aspecto das folhas, com o colapso do tecido vegetal ao redor do
sítio de infecção. Nos pontos de infiltração ocorre necrose das células, tendo a
aparência de folha de jornal (FIGURA 3 B).

23
FIGURA 3. Infiltração de suspensão de 108 células bacterianas para determinação da reação de hipersensibilidade (A) e sintomas característicos de reação hipersensibilidade positiva em folha de fumo (B).
3.4 Caracterização da Diversidade via Marcador Molecular
3.4.1. Extração de DNA
Para a extração de DNA, cada isolado foi cultivado em meio TSB
líquido por 24 h a 37 ºC sob agitação constante de 180 rpm. A extração de DNA
genômico das bactérias foi realizada com o auxílio de um Kit de extração de
DNA, DNeasy Blood e Tissue Handbook, fabricado pela empresa Qiagen.
Posteriormente foi realizada a quantificação do DNA, em que cada amostra
extraída foi diluída (1:100) em H2O milli-Q estéril, sendo depois colocada em
curvette de quartzo e submetida à leitura da densidade óptica a 260 nm (DO260),
para estimar a quantidade de DNA, e a 280 nm (DO280) para estimar a
contaminação com proteínas, utilizando o espectofotômetro. Após a estimação
da quantidade de DNA, todas as amostras foram ajustadas para uma
concentração de 10ng/μL. Em seguida, o DNA foi submetido a uma eletroforese
em gel de agarose a 1,2 % (60 V por aproximadamente 1 h) em tampão TBE (89
mmol/L Tris base, 89 mmol/L Tris borato, EDTA 2mmol/L pH 8,0). Para
A B

24
visualização do DNA, o gel foi corado com brometo de etídio em uma
concentração final de 0,5 μmol/ml e submetido à luz ultravioleta.
3.4.2 Amplificação por rep-PCR
Os isolados foram comparados aos perfis genômicos obtidos por rep-
PCR (PCR-repetitivo). Os primers utilizados foram: REP1R-I (5´-
IIIICGICGICATCIGGC-3´) e REP2-I (5´-ICGITTATCIGGCCTAC-3`), ERIC
1R (5´- ATGTAAGCTCCTGGGGATTCA-3´) e ERIC2 (5´-
AAGTAAGTGACTGGGGTGAGCG-3´); e BOXA1R (5´-
CTCCGGCAAGGCGACGCTGAC -3´) (LOUWS et al., 1994). Cada reação foi
composta de 2,5 µl de Tampão 10X (500 mM KCl, 100 mM Tris HCl); 0,7 µl de
MgCl2 (50 mM); 2 µl de dNTPs (2,5 mM de cada), 1 µl de cada primer (50
pmoles/µM), 0,3 µl de Taq Polimerase (5 U-1µl), 3 µl de DNA (10 ng/µl)
completando-se com água miliQ estéril para um volume de 25 µl.
As amostras foram amplificadas com os primers REP, BOX e ERIC e
submetidas ao programa de amplificação: uma etapa de desnaturação (95 ºC por
7 min), seguido de 30 ciclos (94 ºC por 1 min, 53 ºC por 1 min para anelamento
do primer BOX 1AR, 39 °C para o primer REP e 52 ºC para o ERIC, 65 ºC por
8 min, e uma etapa final de extensão (65 °C por 15 min).
Os produtos de amplificação foram separados em gel de agarose a 1,2%.
Após corrida em tampão TBE 1X, por aproximadamente 3 h a 100 V/cm, o gel
foi corado com uma solução de brometo de etídio (20 mg/ml) e fotografado sob
luz UV. Marcador de peso molecular 100bp-DNA Ladder foi utilizado para
servir como parâmetro para estimar o tamanho dos fragmentos amplificados.
A análise dos fragmentos amplificados com os primers BOX, ERIC e
REP foi baseada na presença ou ausência de bandas, atribuindo-se os valores 1 e
0, respectivamente, a cada situação.

25
3.4.3 Amplificação pela técnica ARDRA
O DNA dos isolados bacterianos endofíticos foram amplificados com os
primers FGPS 1490 5´- TGCGGCTGGATCACCTCCTT-3´ e FGPS132 5`-
CCGGGTTTCCCCATTCGG-3´. As condições de amplificação foram 0,8 µL
de dNTPs (2,0 mM de cada), 2,5 µL de tampão 10X, 0,75 µL de MgCl2
(50mM), 0,25 µL de cada primer (5 mM), 0,2 µL de Taq DNA polimerase (5 U
µL-1), 7 µL de DNA (50ng), com volume final de 25 µL . Em seguida, o
material foi colocado em termociclador e submetido ao programa de
amplificação: uma etapa de desnaturação (95 °C por 3 min), seguidos de 35
ciclos intermediários (94 °C por 1min, 60 °C por 1 min, 72 °C por 2 min), e uma
etapa final de extensão (72 °C por 3 min).
Os produtos de amplificações foram digeridos com as enzimas de
restrição que reconhecem quatro ou cinco pares de bases (TABELA 2), HinfI,
MspI, NdeI e RsaI, recomendado por Fernandes et al. (2003).
TABELA 2: Sítio de restrição das enzimas Hinf I, MspI, NdeI e RsaI.
Enzimas Sítio de restrição
Hinf I 5’...G↓ANTC...3 e 3’...CTNA ↑ G... 5’
Msp I 5´...C↓CGG...3´ e 3´...GGC↑C...5´
Nde I 5’...CA ↓ TATG...3 e 3’...GTAT ↑ AC... 5’
Rsa I 5´...GT↓AC...3´ e 3´...CA↑TG...5´
Para cada enzima foi preparada uma mistura contendo: 10µL do produto
de PCR; 2 µL do tampão específico para cada enzima (10X); 2 µL da enzima (5
U reação -1) e 18 µL de água mili-Q estéril. As misturas foram incubadas durante
16 h em banho-maria a 37 ºC. Os fragmentos foram analisados por eletroforese
em gel de agarose a 1,2%, em TBE 1X, a 100 V por 3 horas. Marcador de peso

26
molecular 100bp-DNA Ladder foi utilizado para servir como parâmetro para
estimar o tamanho dos fragmentos amplificados. Os fragmentos amplificados
foram visualizados sob luz ultravioleta e fotografados em sistema digital.
A análise dos fragmentos amplificados e digeridos foi baseada na
presença ou ausência de bandas, atribuindo-se os valores 1 e 0, respectivamente,
a cada situação.
3.4.4 Amplificação por PCR dos genes nifH
O DNA dos isolados obtidos a partir das raízes foram amplificados para
verificação da presença da região gênica nifH, segundo Teixeira et al. (2007),
utilizando o primer universal 19f F 5´-GGAATTCTGTGACCTAAAGCTGA-
3´ e 19f R 5´-AGCATACATTGCCATCATTTCACC-3´. As condições de
amplificação foram 2,0 µL de dNTPs (2,0 mM de cada), 2,5 µL de tampão 10X,
1µL de MgCl2 (50mM), 0,5 µL de cada primer (5 mM), 0,6 µL de Taq DNA
polimerase (5 U µL-1), 5 µL de DNA (50ng), com volume final de 25 µL . Em
seguida, o material foi colocado num termociclador e submetido ao seguinte
programa de amplificação: uma etapa de desnaturação (94 °C por 2 min),
seguidos de 30 ciclos intermediários (94 °C por 30 segundos, 52 °C por 30
segundos, 72 °C por 30 segundos), e uma etapa de final de extensão (72 °C por 7
min).
Os produtos de amplificação foram observados em gel de agarose 1,2% e
fragmentos de aproximadamente 270 pb foram esperados para o gene nifH.
Marcador de peso molecular 100bp-DNA Ladder foi utilizado para servir como
parâmetro para estimar o tamanho dos fragmentos amplificados.
A análise foi feita analisando a presença ou ausência da banda que
caracterizava o gene.

27
3.5 Análise dos Dados Moleculares
Os resultados obtidos com os primers tanto da técnica rep-PCR quanto
da ARDRA foram analisados com os dados em conjunto de acordo com cada
técnica, com o programa R 2.13, usando o coeficiente de similaridade de
coincidência simples (simple matching) que é dado pela fórmula S(SM) = a +
d/a + b +c + d. O valor de a corresponde ao número de bandas presentes na
unidade J e na unidade K; b é o número de bandas presentes na unidade K
mas ausentes na unidade J; c é o número de bandas presentes na unidade J mas
ausentes em K; d é o número de bandas ausentes em K e J. Portanto, a e d
representam o número de concordâncias positivas e negativas, respectivamente,
enquanto b e c representam as discordâncias (SNEATH e SOKAL, 1973).
A análise de agrupamento foi determinada pelo método UPGMA
(Unweighted Pair Group Method With Arithmetic Mean) utilizando o programa
MEGA 5 (TAMURA et al., 2011).
3.6 Sequenciamento do DNA
Os isolados foram identificados através do sequenciamento parcial da
região 16S. Inicialmente foi amplificada a região 16S utilizando os iniciadores
27 (5'-AGAGTTTGATC(AC)TGGCTCAG-3') e 1492R (5'
ACGG(CT)TACCTTGTTACGACTT-3') através da técnica de PCR. A reação
com volume final de 25 L consistiu da mistura de 2,0 L de dNTPs (2,0 mM
de cada), 2,5 µL de tampão 10X, 0,75 µL de MgCl2 (50mM), 2,5 µL de cada
primer (5 mM), 0,3 µL de Taq DNA polimerase (5 U µL-1), 5 µL de DNA
(50ng), completando o volume final com água ultra pura. Em seguida o material
foi colocado em um termociclador e submetido à amplificação: uma etapa de
desnaturação ( 94 °C por 3 min.), seguido de 30 ciclos intermediários (94 °C

28
por 30 segundos, 50 °C por 30 segundos e 72 °C por 1 min.) e uma etapa final
de extensão (72 °C por 7 min.).
Após a amplificação, os fragmentos foram analisados por eletroforese
em gel de agarose a 1,2%, em TBE 1X, a 100 V por 3 horas, posteriormente os
mesmos foram visualizados sob luz ultravioleta e fotografados em sistema
digital. Após essa etapa, os fragmentos que apresentaram 1500bp foram cortados
do gel e, com a ajuda de um kit de extração de gel (QIAquick Gel Extration kit-
Qiagen), o DNA bacteriano foi purificado, para posterior sequenciamento.
O sequenciamento das amostras foi realizado pela empresa ACTGene
Análises Moleculares LTDA (Centro de Biotecnologia, UFRGS, Porto Alegre,
RS) utilizando o sequenciador automático ABI-PRISM 3100 Genetic Analyzer
armado com capilares de 50 cm e polímero POP6 (Applied Biosystems). Os
DNAs-molde (30 a 45 ng) foram marcados utilizando-se 3,2 pmol do primer
1492R (5'-ACGG(CT)TACCTTGTTACGACTT-3') e 2 L do reagente BigDye
Terminator v3.1 Cycle Sequencing RR-100 (Applied Biosystems) em um volume
final de 10 L. As reações de marcação foram realizadas em termociclador
GeneAmp PCR System 9700 (Applied Biosystems) com uma etapa de
desnaturação inicial a 96 ºC por 3 min seguida de 25 ciclos de 96 ºC por 10 seg,
55 ºC por 5 seg e 60 ºC por 4 minutos. Após marcadas, as amostras foram
purificadas pela precipitação com isopropanol 75% e lavadas com etanol 60%.
Os produtos precipitados foram diluídos em 10 l de formamida, desnaturados a
95 ºC por 5 min, resfriados em gelo por 5 min e eletroinjetados no sequenciador
automático. Os dados de sequenciamento foram coletados utilizando-se o
programa Data Collection v1.0.1 (Applied Biosystems) com os parâmetros Dye
Set “Z”; Mobility File “DT3100POP6{BDv3}v1.mob”; BioLIMS Project
“3100_Project1”; Run Module 1 “StdSeq50_POP6_50cm_cfv_100”; e Analysis
Module 1 “BC-3100SR_Seq_FASTA.saz”.

29
Logo após o sequenciamento das amostras, as sequências foram
comparadas com as presentes no banco de dados do NCBI (Nacional Center for
Biotechnology Information) (www.ncbi.nlm.nih.gov), pelo programa BLAST
(Basic Local Alignment Search Tool) (ALTSCHUL, et al., 1997) para
nucleotídeos. Pela comparação das sequências, os isolados foram identificados
em nível de espécie quando os valores de similaridade encontrados variaram
entre 98% e 100% e menores que 98% em nível gênero.
Após a identificação dos isolados, foi avaliada a frequência de gêneros e
espécies presentes em cada área onde foram realizadas as coletas.

30
4 RESULTADOS
4.1 Isolamento das Bactérias
Foram obtidos 201 isolados de bactérias endofíticas a partir das raízes de
bananeira ‘Prata-Anã’ cultivadas em diferentes pomares localizados nos quatros
municípios pertencentes à região Norte do Estado de Minas Gerais e em um
município do Estado da Bahia, Guanambi. Não foram observadas colônias nas
placas usadas como controle de assepsia, após o segundo banho de ultrassom,
confirmando assim a natureza endofítica dos isolados obtidos.
Por isolamento das bactérias endofíticas em meio de cultura, foi possível
observar vários tipos morfológicos, apresentando diferenças principalmente
quanto a cor e formato das colônias (FIGURA 4).
FIGURA 4. Variação na coloração e na forma das bactérias isoladas de raízes de bananeira ‘Prata-Anã’.
Observou-se que dentre os meios utilizados no plaqueamento bacteriano,
o meio “Trypic Soy Agar” (TSA) favoreceu maior desenvolvimento bacteriano,
uma vez que promoveu crescimento de 53,8% dos isolados; já o Agar-Nutritivo-
Dextrose-Levedura (NYDA) com 29,8% e o Batata Dextrose Agar (BDA) com
apenas 16,4%.

31
A densidade de bactérias endofíticas isoladas nos meios de cultura
utilizados variou entre as áreas de coletas. Na área 12 observaram 13x104 UFC/g
de raiz, predominantemente dentro do gênero Bacillus spp. e na área 8,
quantificou-se a menor densidade de UFC/g de raiz, aproximadamente 1x104,
porém com maior diversidade comparativamente à área 12 (FIGURA
5).
0
1
2
3
4
5
6
7
8
9
10
11
12
13
14
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Áreas de coleta
UF
Cx1
0-1/g
de
raiz
(x10
-4)
FIGURA 5. Número de unidades formadoras de colônias bacterianas (UFC/g de raiz) de bactérias endofíticas de raiz de bananeira ‘Prata-Anã’ de diferentes áreas.
4.2 Caracterização dos isolados bacterianos com relação à resposta de
Gram
Foi determinado o padrão de coloração de Gram de todas as bactérias
isoladas, sendo 150 classificadas como Gram-positivas e 51 como Gram-
negativas (FIGURA 6). A área 1, localizada em Janaúba, foi a que apresentou

32
maior número de bactérias Gram-negativas. Por outro lado, na área 2 e 5 foram
amostradas somente bactérias Gram-positivas.
FIGURA 6. Padrão de coloração de Gram-negativa (A) e Gram-positiva (B) das bactérias endofíticas isoladas de bananeira ‘Prata-Anã’. 4.3 Teste de patogenicidade por meio da reação de hipersensibilidade
Até os cinco dias após a inoculação, nenhum dos 201 isolados
bacterianos coletados apresentou reação de hipersensibilidade nas plantas de
fumo (Nicotiana tabacum L.), pimentão (Capsicum annuum Mill) e feijão
(Phaseolus vulgaris L.) (FIGURA 7) descartando a hipótese dos isolados
bacterianos serem fitopatogênicos.
FIGURA 7. Reação de hipersensibilidade negativa em plantas de pimentão (A), feijão (B) e fumo (C).
A B
A B C
33
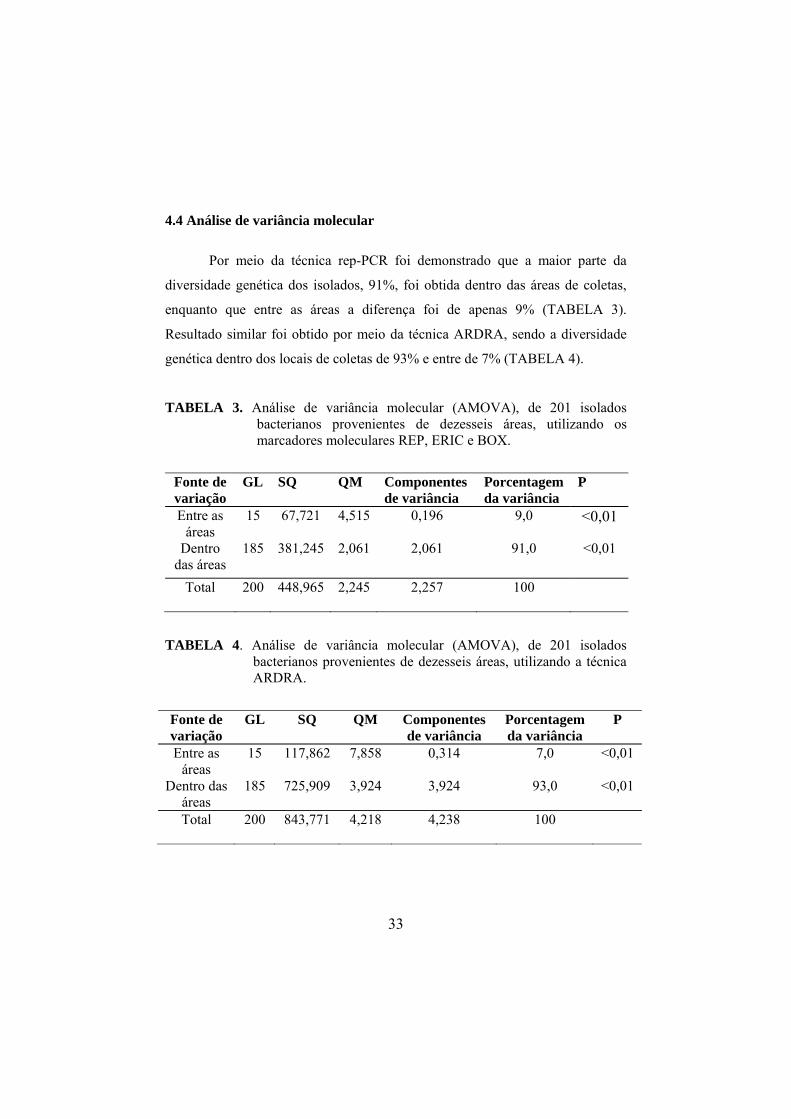
4.4 Análise de variância molecular
Por meio da técnica rep-PCR foi demonstrado que a maior parte da
diversidade genética dos isolados, 91%, foi obtida dentro das áreas de coletas,
enquanto que entre as áreas a diferença foi de apenas 9% (TABELA 3).
Resultado similar foi obtido por meio da técnica ARDRA, sendo a diversidade
genética dentro dos locais de coletas de 93% e entre de 7% (TABELA 4).
TABELA 3. Análise de variância molecular (AMOVA), de 201 isolados bacterianos provenientes de dezesseis áreas, utilizando os marcadores moleculares REP, ERIC e BOX.
Fonte de variação
GL SQ QM Componentes de variância
Porcentagem da variância
P
Entre as áreas
15 67,721
4,515
0,196
9,0 <0,01
Dentro das áreas
185 381,245
2,061
2,061
91,0 <0,01
Total 200 448,965
2,245 2,257
100
TABELA 4. Análise de variância molecular (AMOVA), de 201 isolados bacterianos provenientes de dezesseis áreas, utilizando a técnica ARDRA.
Fonte de variação
GL SQ QM Componentes de variância
Porcentagem da variância
P
Entre as áreas
15 117,862
7,858
0,314
7,0 <0,01
Dentro das áreas
185 725,909
3,924
3,924
93,0 <0,01
Total 200 843,771
4,218 4,238
100

34
4.5 Sequenciamento do DNA bacteriano
As sequências consenso foram comparadas com sequências do banco de
dados público GenBank através do programa BLASTn (NCBI
www.ncbi.nih.gov). A análise dos dados apresentou valores de similaridade
variando entre 90% e 100% (TABELA 5). Dois isolados apresentaram 100% de
similaridade com as espécies de B. pumilus (Meyer e Gottheil, 1901) e B.
safensis (Satomi et al., 2006), e 90% foram encontrados entre um isolado e o
gênero Pseudomonas sp.
Após o sequenciamento parcial da região rDNA 16S dos 201 isolados
bacterianos, foram obtidas 143 sequências com boa qualidade. A análise das
sequências revelou que os isolados estão distribuídos por 17 gêneros
bacterianos: Bacillus, Agrobacterium (Conn, 1942), Klebsiella, Paenibacillus,
Lysinibacillus, Streptomyces, Artrobacter, Micrococus (Cohn 1872 emend.
Stackebrandt et al., 1995 emend. Wieser et al., 2002), Rhizobium,
Sporolactobacillus, Acetobacter (Beijerinck, 1898), Aneurinibacillus (Shida et
al., 1996 emend. Heyndrickx et al., 1997), Pantoea, Brevibacillus (Shida et al.,
1996), Stenotrophomonas, Pseudomonas e Enterobacter, os quais incluem 16
espécies diferentes (TABELA 6). Os diferentes gêneros e espécies de bactérias
endofíticas encontradas neste trabalho foram isolados de forma aleatória nas
áreas de coletas, e em função do bulk das amostras não foi possível detectar se
nos casos de isolamento de mais de uma espécie na mesma área, se estas eram
oriundas da mesma planta, e, portanto se tais espécies e/ou gêneros ocorriam
concomitantemente no mesmo nicho ecológico.

35
TABELA 5. Identificação das bactérias endofíticas de raízes de bananeira ‘Prata-Anã’ com base na identidade da sequência parcial do gene16S rDNA.
Isolado E value 3* Identidade ** Organismo mais
relacionado *** Teste de Gram
Acesso no Genbank ****
1 0.0 98% Bacillus pumilus + HM006706.1 3 7.e -78 98% Bacillus.pumilus + JF738118.1 4 0.0 98% Bacillus subtilis + AY741264.1 5 7. e -119 99% Bacillus pumilus + HQ218993.1 6 2. e -47 98% Bacillus subtilis + JF926531.1 7 0.0 98% Agrobacterium tumefaciens - GU784794.1 9 1.e -116 98% Bacillus subtilis + AY741264.1
10 1.e -136 98% Bacillus pumilus + GQ917222.1 11 5.e-172 97% Bacillus sp. + HQ218993.1 12 4.e-116 98% Bacillus pumilus + GQ917222.1 14 0.0 99% Bacillus pumilus + HQ218993.1 15 4.e-127 98% Bacillus pumilus + GQ917222.1 16 4.e-111 97% Bacillus sp. + AJ550463.1 17 2.e -109 97% Bacillus sp. + JF802184.1 18 6.e-68 96% Bacillus sp. + EU977790.1 23 3.e -179 98% Klebsiella pneumoniae - JN201948.1 24 0.0 98% Bacillus thuringiensis + JF947357.1 25 0.0 98% Bacillus cereus + GU451184.1 26 0.0 98% Bacillus methylotrophicus + HM209756.1 27 0.0 97% Bacillus sp. + HQ256520.1 28 1.e-163 96% Paenibacillus sp. + EF178460.1 30 0.0 98% Bacillus axarquiensis + JF414764.1 32 4.e -76 94% Bacillus sp. + HQ003450.1 33 4.e -60 94% Bacillus sp. + GQ34O503.1 34 1.e-131 98% Bacillus pumilus + JN215511.1 35 2.e-88 94% Bacillus sp. + GQ340516.1 36 4.e-49 96% Lysinibacillus sp. + JF700466.1 37 2.e -146 96% Bacillus sp. + JN215502.1 38 3.e -97 96% Stenotrophomonas sp. - EU931559.1 39 3.e-72 94% Streptomyces sp. + FJ951435.1 40 7.e-99 97% Bacillus sp. + GQ340516.1 41 3.e-71 98% Bacillus subtilis + EF101707.1 42 2.e-89 96% Bacillus sp. + JN082266.1 43 3.e-61 91% Bacillus sp. + GU188935.1 44 4.e -173 98% Bacillus amyloliquefaciens + GU122948.1 45 0.0 98% Lysinibacillus sphaericus + JN215512.1 46 0.0 99% Bacillus pumilus + FJ236809.1 47 1.e -147 97% Bacillus sp. + FJ611939.1 48 0.0 98% Bacillus subtilis + AY741264.1 49 0.0 98% Bacillus licheniformis + EU366371.1
...continua...
3 * Valor E: probabilidade de se encontrar aleatoriamente o mesmo alinhamento entre duas sequências. **Identidade: porcentagem de identidade entre a sequência do isolado de banana e o organismo relacionado. ***Organismo mais relacionado: organismo que possui a sequência com a qual a sequência parcial do gene 16S rDNA do isolado de banana apresentou maior homologia. ****Acesso no GenBank: número de acesso da sequência do organismo relacionado.
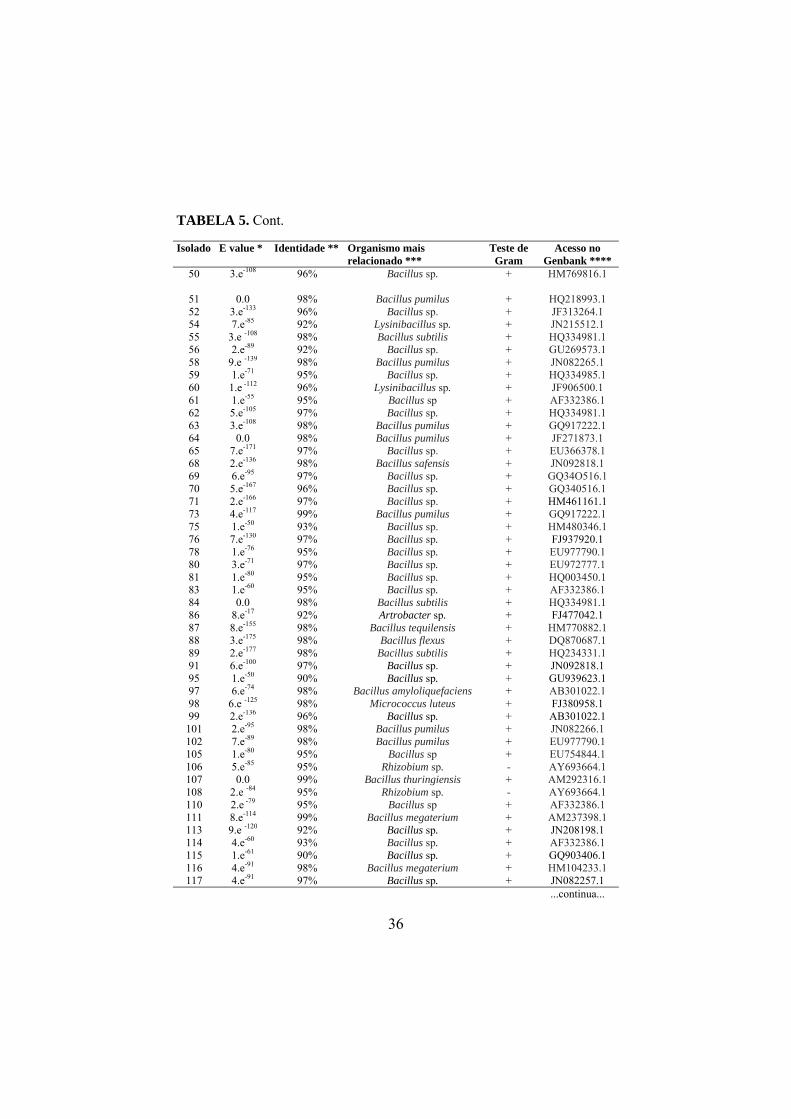
36
TABELA 5. Cont. Isolado E value * Identidade ** Organismo mais
relacionado *** Teste de Gram
Acesso no Genbank ****
50 3.e-108 96% Bacillus sp. + HM769816.1
51 0.0 98% Bacillus pumilus + HQ218993.1 52 3.e-133 96% Bacillus sp. + JF313264.1 54 7.e-85 92% Lysinibacillus sp. + JN215512.1 55 3.e -108 98% Bacillus subtilis + HQ334981.1 56 2.e-89 92% Bacillus sp. + GU269573.1 58 9.e -139 98% Bacillus pumilus + JN082265.1 59 1.e-71 95% Bacillus sp. + HQ334985.1 60 1.e -112 96% Lysinibacillus sp. + JF906500.1 61 1.e-55 95% Bacillus sp + AF332386.1 62 5.e-105 97% Bacillus sp. + HQ334981.1 63 3.e-108 98% Bacillus pumilus + GQ917222.1 64 0.0 98% Bacillus pumilus + JF271873.1 65 7.e-171 97% Bacillus sp. + EU366378.1 68 2.e-136 98% Bacillus safensis + JN092818.1 69 6.e-95 97% Bacillus sp. + GQ34O516.1 70 5.e-167 96% Bacillus sp. + GQ340516.1 71 2.e-166 97% Bacillus sp. + HM461161.1 73 4.e-117 99% Bacillus pumilus + GQ917222.1 75 1.e-50 93% Bacillus sp. + HM480346.1 76 7.e-130 97% Bacillus sp. + FJ937920.1 78 1.e-76 95% Bacillus sp. + EU977790.1 80 3.e-71 97% Bacillus sp. + EU972777.1 81 1.e-80 95% Bacillus sp. + HQ003450.1 83 1.e-60 95% Bacillus sp. + AF332386.1 84 0.0 98% Bacillus subtilis + HQ334981.1 86 8.e-17 92% Artrobacter sp. + FJ477042.1 87 8.e-155 98% Bacillus tequilensis + HM770882.1 88 3.e-175 98% Bacillus flexus + DQ870687.1 89 2.e-177 98% Bacillus subtilis + HQ234331.1 91 6.e-100 97% Bacillus sp. + JN092818.1 95 1.e-50 90% Bacillus sp. + GU939623.1 97 6.e-74 98% Bacillus amyloliquefaciens + AB301022.1 98 6.e -125 98% Micrococcus luteus + FJ380958.1 99 2.e-136 96% Bacillus sp. + AB301022.1 101 2.e-95 98% Bacillus pumilus + JN082266.1 102 7.e-89 98% Bacillus pumilus + EU977790.1 105 1.e-80 95% Bacillus sp + EU754844.1 106 5.e-85 95% Rhizobium sp. - AY693664.1 107 0.0 99% Bacillus thuringiensis + AM292316.1 108 2.e -84 95% Rhizobium sp. - AY693664.1 110 2.e -79 95% Bacillus sp + AF332386.1 111 8.e-114 99% Bacillus megaterium + AM237398.1 113 9.e -120 92% Bacillus sp. + JN208198.1 114 4.e-60 93% Bacillus sp. + AF332386.1 115 1.e-61 90% Bacillus sp. + GQ903406.1 116 4.e-91 98% Bacillus megaterium + HM104233.1 117 4.e-91 97% Bacillus sp. + JN082257.1
...continua...
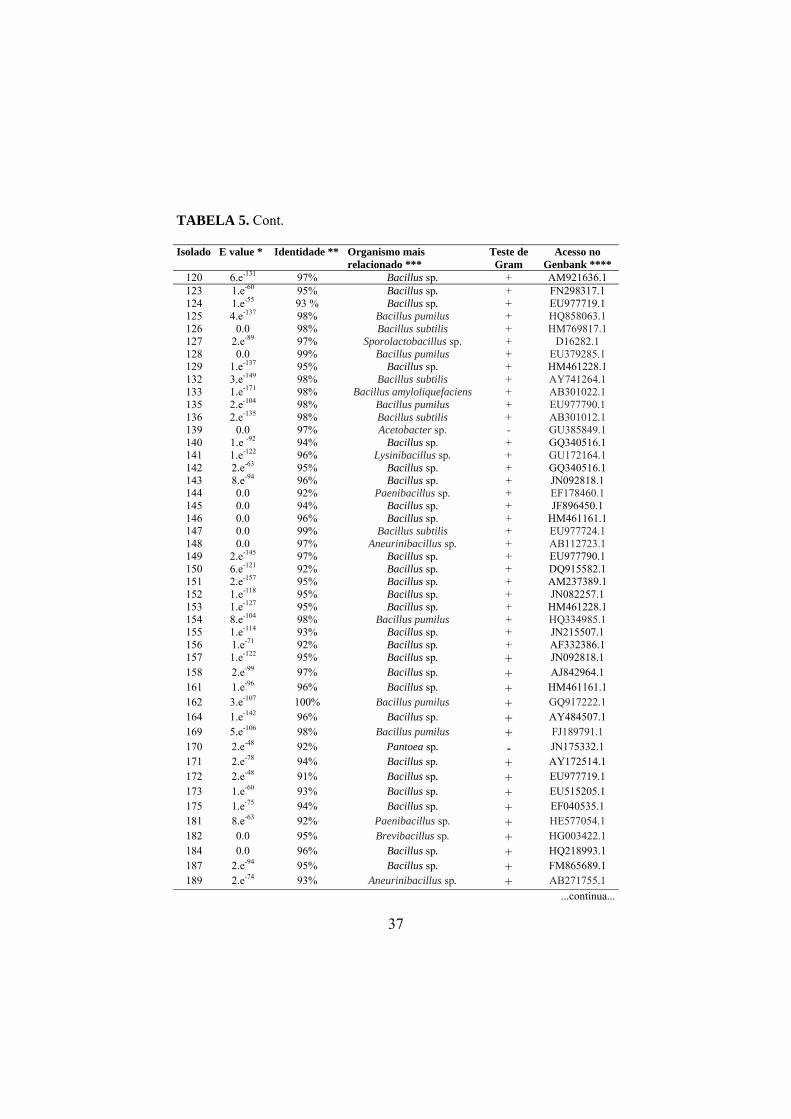
37
TABELA 5. Cont. Isolado E value * Identidade ** Organismo mais
relacionado ***Teste de Gram
Acesso no Genbank ****
120 6.e-131 97% Bacillus sp. + AM921636.1 123 1.e-60 95% Bacillus sp. + FN298317.1 124 1.e-55 93 % Bacillus sp. + EU977719.1 125 4.e-137 98% Bacillus pumilus + HQ858063.1 126 0.0 98% Bacillus subtilis + HM769817.1 127 2.e-89 97% Sporolactobacillus sp. + D16282.1 128 0.0 99% Bacillus pumilus + EU379285.1 129 1.e-137 95% Bacillus sp. + HM461228.1 132 3.e-149 98% Bacillus subtilis + AY741264.1 133 1.e-171 98% Bacillus amyloliquefaciens + AB301022.1 135 2.e-104 98% Bacillus pumilus + EU977790.1 136 2.e-135 98% Bacillus subtilis + AB301012.1 139 0.0 97% Acetobacter sp. - GU385849.1 140 1.e -92 94% Bacillus sp. + GQ340516.1 141 1.e-122 96% Lysinibacillus sp. + GU172164.1 142 2.e-63 95% Bacillus sp. + GQ340516.1 143 8.e-94 96% Bacillus sp. + JN092818.1 144 0.0 92% Paenibacillus sp. + EF178460.1 145 0.0 94% Bacillus sp. + JF896450.1 146 0.0 96% Bacillus sp. + HM461161.1 147 0.0 99% Bacillus subtilis + EU977724.1 148 0.0 97% Aneurinibacillus sp. + AB112723.1 149 2.e-145 97% Bacillus sp. + EU977790.1 150 6.e-121 92% Bacillus sp. + DQ915582.1 151 2.e-157 95% Bacillus sp. + AM237389.1 152 1.e-118 95% Bacillus sp. + JN082257.1 153 1.e-127 95% Bacillus sp. + HM461228.1 154 8.e-104 98% Bacillus pumilus + HQ334985.1 155 1.e-114 93% Bacillus sp. + JN215507.1 156 1.e-71 92% Bacillus sp. + AF332386.1 157 1.e-122 95% Bacillus sp. + JN092818.1 158 2.e-99 97% Bacillus sp. + AJ842964.1 161 1.e-96 96% Bacillus sp. + HM461161.1 162 3.e-107 100% Bacillus pumilus + GQ917222.1 164 1.e-142 96% Bacillus sp. + AY484507.1 169 5.e-106 98% Bacillus pumilus + FJ189791.1 170 2.e-48 92% Pantoea sp. - JN175332.1 171 2.e-78 94% Bacillus sp. + AY172514.1 172 2.e-48 91% Bacillus sp. + EU977719.1 173 1.e-60 93% Bacillus sp. + EU515205.1 175 1.e-75 94% Bacillus sp. + EF040535.1 181 8.e-63 92% Paenibacillus sp. + HE577054.1 182 0.0 95% Brevibacillus sp. + HG003422.1 184 0.0 96% Bacillus sp. + HQ218993.1 187 2.e-94 95% Bacillus sp. + FM865689.1 189 2.e-74 93% Aneurinibacillus sp. + AB271755.1
...continua...
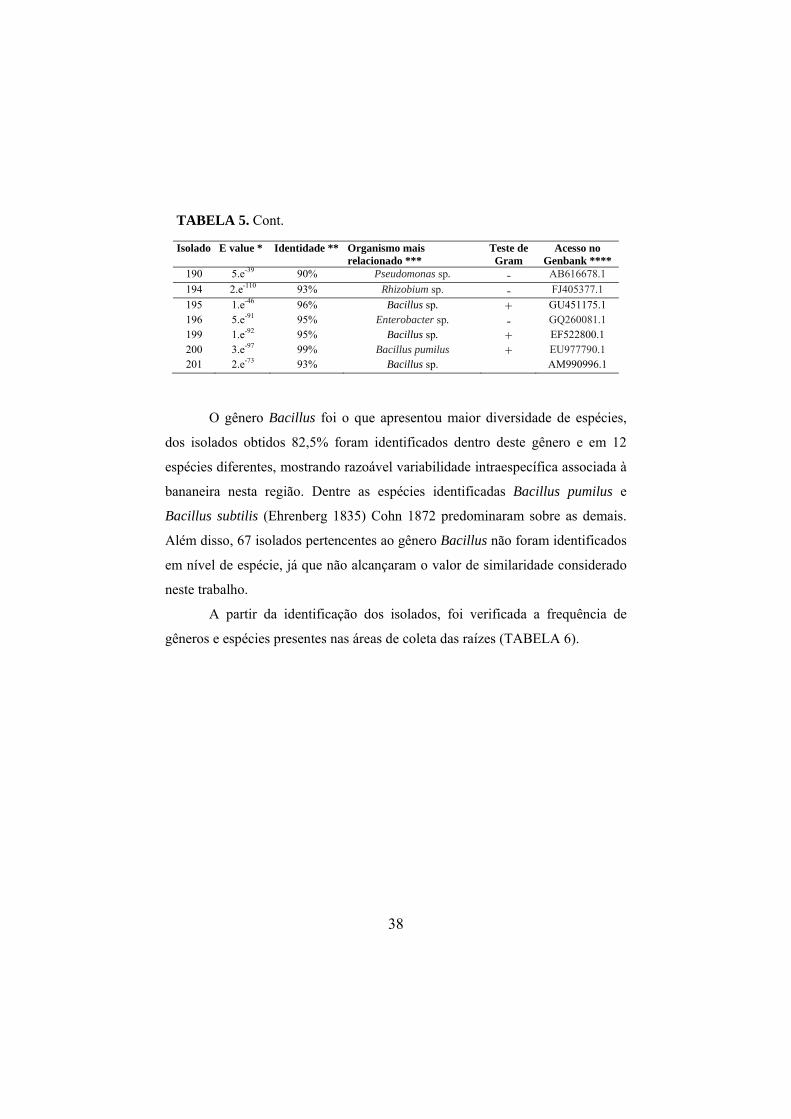
38
TABELA 5. Cont. Isolado E value * Identidade ** Organismo mais
relacionado *** Teste de Gram
Acesso no Genbank ****
190 5.e-39 90% Pseudomonas sp. - AB616678.1
194 2.e-110 93% Rhizobium sp. - FJ405377.1
195 1.e-46 96% Bacillus sp. + GU451175.1 196 5.e-91 95% Enterobacter sp. - GQ260081.1 199 1.e-92 95% Bacillus sp. + EF522800.1 200 3.e-97 99% Bacillus pumilus + EU977790.1 201 2.e-73 93% Bacillus sp. AM990996.1
O gênero Bacillus foi o que apresentou maior diversidade de espécies,
dos isolados obtidos 82,5% foram identificados dentro deste gênero e em 12
espécies diferentes, mostrando razoável variabilidade intraespecífica associada à
bananeira nesta região. Dentre as espécies identificadas Bacillus pumilus e
Bacillus subtilis (Ehrenberg 1835) Cohn 1872 predominaram sobre as demais.
Além disso, 67 isolados pertencentes ao gênero Bacillus não foram identificados
em nível de espécie, já que não alcançaram o valor de similaridade considerado
neste trabalho.
A partir da identificação dos isolados, foi verificada a frequência de
gêneros e espécies presentes nas áreas de coleta das raízes (TABELA 6).

39
TABELA 6. Frequência de bactérias endofíticas obtidas de raízes bananeira ‘Prata-Anã’coletadas em 16 áreas de cultivo no Norte de Minas Gerais e Guanambi (BA).
Espécies/ Gêneros identificados Frequência / (área)
Acetobacter sp. 01/(16)
Agrobacterium tumefaciens 01/(1)
Aneurinibacillus sp. 01/(2); 01/(11);
Artrobacter sp. 01/(9)
Bacillus amyloliquefaciens 01/(3); 01/(9); 02/(16)
Bacillus axarquiensis 01/(3)
Bacillus cereus 01/(5)
Bacillus flexus 01/(8)
Bacillus licheniformis 01/(6)
Bacillus megaterium 02/(10)
Bacillus methylotrophicus 01/(6)
Bacillus pumilus 04/(1); 01/(2); 02/(3); 01/(4); 03/(5); 03/(7); 02/(8);
01/(9); 02/(11); 01/(12); 01/(13); 01/(14); 01/(15);
Bacillus safensis 02/(3)
Bacillus sp. 06/(1); 04/(2); 08/(3); 03/(4); 03/(5); 06/(6); 01/(7);
03/(8); 03/(9); 03/(10); 06/(11); 02/(12); 07/(13);
05/(14); 02/(15); 05/(16)
Bacillus subtilis 03/(1); 03/(4); 01/(6); 01/(8); 02/(11); 02/(16)
Bacillus tequilensis 01/(15)
Bacillus thuringiensis 01/(5); 01/(10)
Brevibacillus sp. 01/(14);
Enterobacter sp. 01/(10)
Klebsiella pneumoniae 01/(1)
Lysinibacillus sp. 01/(4); 01/(6); 01/(7); 01/(11);
Lysinibacillus sphaericus 01/(4)
Micrococcus luteus 01/(8)
Paenibacillus sp. 01/(1); 01/(15); 01/(16)
Pantoea sp. 01/(15)
Pseudomonas sp 01/(12)
Rhizobium sp 01/ (4); 01/(6); 01/(10);

40
Sporolactobacillus sp. 01/(11)
Stenotrophomonas sp. 01/(4)
Streptomyces sp. 01/(4)
De forma geral, o gênero Bacillus foi isolado em todas as áreas
amostradas, e em algumas áreas a diversidade intraespecífica foram maiores
como ocorreu na área 3, onde foi possível detectar quatro diferentes espécies de
bactérias associadas à bananeira, das quais citam-se: B. pumilus, B. safensis, B.
amyloliquefaciens (Priest et al. 1987 emend. Wang et al., 2008) e B.
axarquiensis (Ruiz-García et al., 2005). Nas áreas 5 e 13 e na área localizada em
Guanambi, foram encontradas apenas bactérias pertencentes ao gênero Bacillus,
demonstrando baixa diversidade de gêneros.
O maior número de espécies foi observado nas áreas 1 e 4. Na área 1
identificaram-se os gêneros Paenobacillus sp., Klebsiella sp., Bacillus spp. e a
espécie Agrobacterium tumefaciens (Smith and Townsend 1907) Conn 1942, e
na área 4 os gêneros Stenotrophomonas sp., Streptomyces sp., Lysinibacillus
sphaericus (Meyer and Neide 1904) Ahmed et al., 2007), Rhizobium sp. e
Bacillus spp..
Observou-se que as espécies encontradas podem estar associadas com o
manejo das áreas. Em 81% das áreas de coleta há incorporação de matéria
orgânica realizada por meio da adição de esterco como medida adicional do
manejo da cultura. Entretanto, nas áreas 8 e 11 onde se pratica o manejo
orgânico, verificou-se alta diversidade interespecífica. Na área 8 foram
identificadas as espécies B. Subtilis, B pumillus, B flexus (Priest et al., 1989) e
Micrococus luteus, enquanto na área 11 quatro gêneros diferentes: Bacillus spp.,
Aneurobacillus sp., Lysinibacillus e Sporolactobacillus sp..

41
4.6 Caracterização da Diversidade via rep-PCR
A diversidade genética de 201 isolados de bactérias endofíticas foi
avaliada por meio da análise comparativa entre os padrões eletroforéticos,
gerados pela amplificação de sequências conservadas e repetitivas do DNA
genômico, que possibilitou a construção de dendrograma com base na
dissimilaridade dos padrões eletroforéticos determinados pela análise conjunta
dos marcadores moleculares.
A técnica rep-PCR gerou, para cada primer: REP, ERIC e BOX,
separadamente, perfis moleculares diferentes. Foram obtidos no total 50 locos
passíveis de leitura, sendo que todos os fragmentos mostraram-se polimórficos
(TABELA 7).
TABELA 7. Número de bandas monomórficas e polimórficas formadas com a utilização dos primers REP, ERIC e BOX pela técnica rep-PCR.
Primer No bandas monomórficas No bandas polimórficas
REP 0 11
ERIC 0 18
BOX 0 21
Os dados obtidos indicam que o primer BOX (FIGURA 8) apresentou
maior polimorfismo do que os outros dois primers ERIC e REP (FIGURAS 9 e,
10).
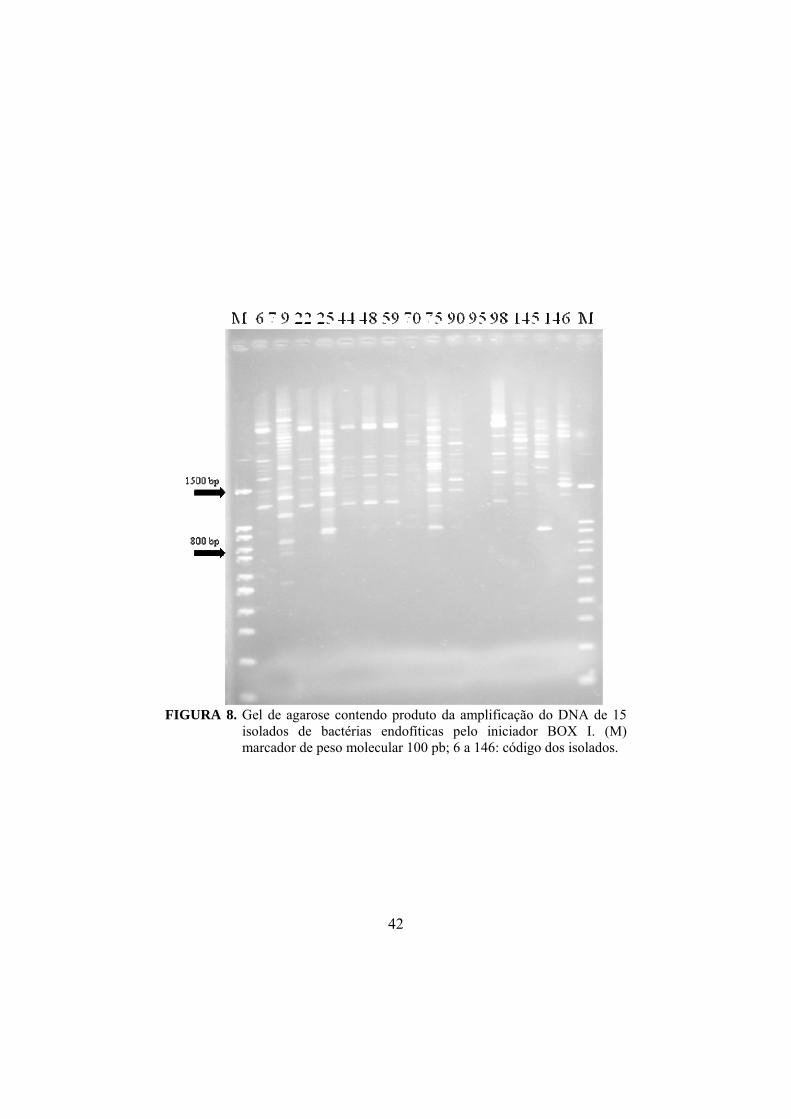
42
FIGURA 8. Gel de agarose contendo produto da amplificação do DNA de 15
isolados de bactérias endofíticas pelo iniciador BOX I. (M) marcador de peso molecular 100 pb; 6 a 146: código dos isolados.

43
FIGURA 9. Gel de agarose contendo produto da amplificação do DNA de isolados de bactérias endofíticas pelos iniciadores ERIC I e ERIC 2. (M) marcador de peso molecular 100 pb; 18 a 199: código dos isolados

44
FIGURA 10. Gel de agarose contendo produto da amplificação do DNA de
isolados de bactérias endofíticas pelos iniciadores REP I e REP 2. (M) marcador de peso molecular 100 pb; 38 a 56: código dos isolados
Utilizando os dados em conjuntos dos três primers (REP, ERIC e BOX),
constatou-se que a distância genética entre as áreas de coleta variou de 3,0 % a
12,9%. Os indivíduos da área 16, coletados no município de Jaíba, apresentaram
a menor distância genética, enquanto que os isolados coletados no município de
Guanambi apresentaram a maior distância genética (TABELA 8, FIGURA 11).
Não foi observado padrão de amplificação específico para os isolados

45
pertencentes à mesma área de coleta, já que apresentaram uma distribuição
desuniforme.
TABELA 8. Distância genética entre os isolados da mesma área onde foram
coletadas raízes de bananeira ‘Prata-Anã’ por meio da técnica rep-PCR.
Área Município da Coleta Distância genética (%)
1 Janaúba 11,0
2 Janaúba 7,8
3 Guanambi 12,9
4 Porteirinha 10,1
5 Nova Porteirinha 6,9
6 Nova Porteirinha 9,9
7 Nova Porteirinha 8,0
8 Nova Porteirinha 9,9
9 Nova Porteirinha 8,5
10 Nova Porteirinha 6,9
11 Matias Cardoso 5,5
12 Jaíba 12,4
13 Jaíba 4,0
14 Jaíba 4,5
15 Jaíba 9,5
16 Jaíba 3,0

46
Legenda:
Janaúba (1) Nova Porteirinha (9) Janaúba (2) Nova Porteirinha (10)
Guananbi (3) Matias Cardoso (11) Porteirinha (4) Jaíba (12) Nova Porteirinha (5) Jaíba (13) Nova Porteirinha (6) Jaíba (14) Nova Porteirinha (7) Jaíba (15) Nova Porteirinha (8) Jaíba (16)
FIGURA 11. Dendrograma de dissimilaridade obtido através da técnica rep-PCR dos isolados de bactérias endofíticas provenientes de raízes de bananeira ‘Prata-Anã’.

47
Analisando o dendrograma, verificou-se a formação de seis grupos
(TABELA 9). Entretanto, não foi possível correlacionar os indivíduos com as
respectivas áreas de coleta, visto que estão distribuídos de forma aleatória pelo
dendrograma.
TABELA 9. Agrupamento dos gêneros e espécies isolados de raízes de bananaeira ‘Prata-Anã’ utilizando a técnica rep-PCR.
Grupos Gêneros/ Espécies identificados em cada grupo
I Acetobacter sp, Aneurinibacillus sp , B amyloliquefaciens, B sp., B megaterium, B pumilus, B subtilis, Brevibacillus sp., Paenibacillus sp., Sporolactobacillus sp..
II B. amyloliquefaciens, B megaterium, B methylotrophicus,., B. pumilus, B. safensis, B sp., B. subtilis, B. truringiensis, Rhizobium sp..
III Artrobacter sp., Aneurinbcacillus sp., B. axarquiensis, B. flexus, B.pumilus, B. safensis, B. sp., B. subtilis, B. tequilensis, B. truringiensis, Klebsiella pneumoniae, Lysinibacillus sp., Lysinibacillus sphaericus, Paenibacillus sp., Stenotrophomonas sp., Streptomyces sp..
IV B amyloliquefaciens, B pumilus, B sp., B. subtilis, Lysinibacillus sp., Pantoea sp., Pseudomonas sp., Rhizobium sp..
V B amyloliquefaciens, B. cereus, B. licheniformis, B pumilus, B sp, B. subtilis, Enterobacter sp., Lysinibacillus sp., Micrococus luteus.
VI Agrobacterium tumefaciens
4.7 Caracterização da Diversidade pela técnica ARDRA
A análise de diversidade dos isolados por meio da técnica ARDRA,
obtida pelos produtos de clivagem com as quatro enzimas de restrição, produziu
45 bandas polimórficas e nenhuma monomórfica. A RsaI foi a enzima que
apresentou maior poder discriminatório, gerando 13 bandas diferentes, seguida

48
de MspI com 12 , HinfI com 11 bandas e NdeI com 9 bandas (FIGURAS 12, 13,
14 e 15).

49
FIGURA 12. Perfil de restrição da região 16S DNAr dos isolados bacterianos isolados de bananeira ‘Prata-Anã’ após a digestão com a enzima RsaI (B).
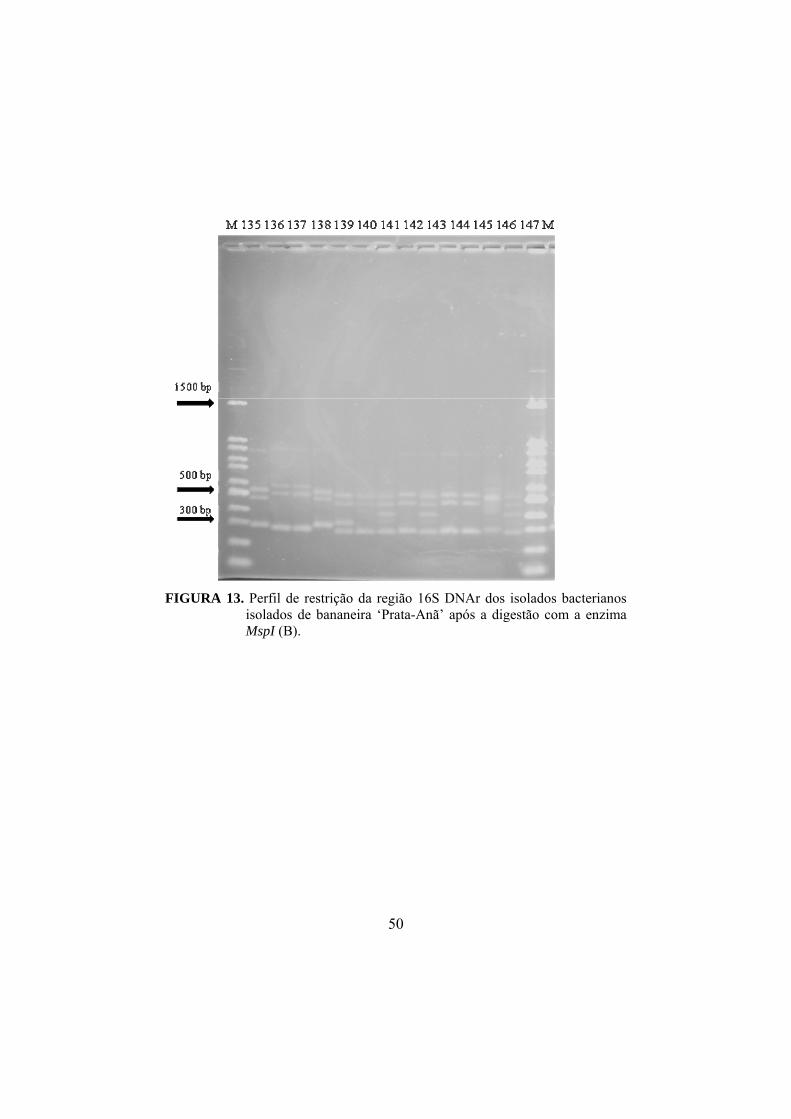
50
FIGURA 13. Perfil de restrição da região 16S DNAr dos isolados bacterianos isolados de bananeira ‘Prata-Anã’ após a digestão com a enzima MspI (B).

51
FIGURA 14. Perfil de restrição da região 16S DNAr dos isolados bacterianos isolados de bananeira ‘Prata-Anã’ após a digestão com a enzima HinfI.

52
FIGURA 15. Perfil de restrição da região 16S DNAr dos isolados bacterianos isolados de bananeira ‘Prata-Anã’ após a digestão com a enzima NdeI.
A distância genética observada entre as áreas de coleta variou de 12,9%
a 20,3%. Os indivíduos coletados na área 7, município de Nova Porteirinha,
apresentaram a menor distância genética. Por outro lado, os isolados coletados
na área 13 são os mais distantes geneticamente (TABELA 10, FIGURA 16).

53
TABELA 10. Distância genética entre os isolados da mesma área onde foram coletadas raízes de bananeira ‘Prata-Anã’ utilizando os dados da técnica ARDRA.
Área Município de coleta Distância genética (%)
1 Janaúba 19,2
2 Janaúba 19,2
3 Guananbi 19,4
4 Porteirinha 19,7
5 Nova Porteirinha 14,6
6 Nova Porteirinha 15,6
7 Nova Porteirinha 12,9
8 Nova Porteirinha 15,2
9 Nova Porteirinha 17,9
10 Nova Porteirinha 14,8
11 Matias Cardoso 17,4
12 Jaíba 18,4
13 Jaíba 20,3
14 Jaíba 17,5
15 Jaíba 18,9
16 Jaíba 15,3
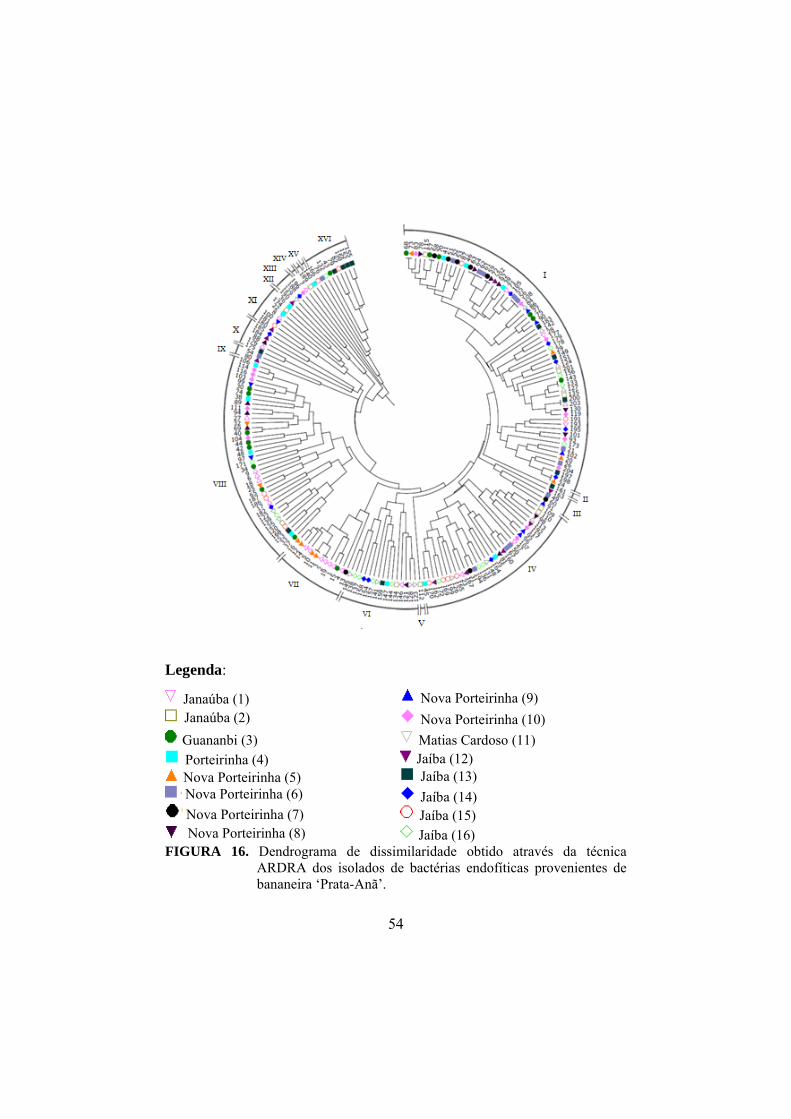
54
Legenda:
Janaúba (1) Nova Porteirinha (9) Janaúba (2) Nova Porteirinha (10)
Guananbi (3) Matias Cardoso (11) Porteirinha (4) Jaíba (12) Nova Porteirinha (5) Jaíba (13) Nova Porteirinha (6) Jaíba (14) Nova Porteirinha (7) Jaíba (15) Nova Porteirinha (8) Jaíba (16)
FIGURA 16. Dendrograma de dissimilaridade obtido através da técnica ARDRA dos isolados de bactérias endofíticas provenientes de bananeira ‘Prata-Anã’.

55
Foram formados dezesseis grupos baseados nas distâncias genéticas
entre os isolados (TABELA 11), em que cada grupo foi formado por espécies e
gêneros diferentes.
TABELA 11. Agrupamento dos gêneros e espécies isolados de raízes de bananeira ‘Prata-Anã’ utilizando a técnica ARDRA.
Grupos Gêneros/ Espécies identificados em cada grupo
I Artrobacter sp., B amyloliquefaciens, B. methylotrophicus, B. pumilus, B. safensis , B sp., B. subtilis, B. tequilensis, B. thuringiensis, Paenibacillus sp., Sporolactobacillus sp., Streptomyces sp., Rhizobium sp..
II B.sp.. III B. licheniformis, B.sp., Lysinibacillus sp.. IV Aneurinibacillus sp., B amyloliquefaciens, B. pumilus, B.sp., B.
subtilis, B. thuringiensis, Klebsiella pneumoniae, Lysinibacillus sphaericus, Micrococus luteus, Pantoea sp., Rhizobium sp..
V Isolado não identificado VI B. pumilus, B.sp, B. subtilis, Lysinibacillus sp., Paenibacillus sp.. VII B. pumilus, B.sp., B. subtilis. VIII Acetobacter sp., B amyloliquefaciens, B. axarquiensis, B cereus, B.
megaterium,B. pumilus, B.sp., B. subtilis, Brevibacillus sp., Stenotrophomonas sp., Rhizobium sp.,
IX B.sp., Lysinibacillus sp.. X B. flexus, B.sp., XI Lysinibacillus sp., Paenibacillus sp.. XII B.sp., Enterobacter sp., Pseudomonas sp.. XIII Agrobacterium tumefaciens XIV Aneurinibacillus sp.. XV B.sp.. XVI B.sp., B. subtilis.

56
4.8 Caracterização do Gene nifH
A amplificação por PCR do gene nifH foi verificada em 40 dos 201
isolados bacterianos provenientes de raízes de bananeira ‘Prata-Anã’ (TABELA
12), pois uma banda com tamanho de 270 bp foi amplificada (FIGURA 17).

57
FIGURA 17. Amplificação do gene nifH de 18 bactérias endofíticas de raízes de bananeira ‘Prata-Anã’. (M) marcador de peso molecular 100 pb; 4 a 67: códigos dos isolados.

58
TABELA 12. Número de isolados e identificação de gêneros e espécies de bactérias endofíticas que amplificaram o gene nifH, por área de coleta.
Área Origem N °de isoladoscom gene nifH
Gêneros/ Espécies
1 Janaúba 5 B. sp., B. subtilis, Klebsiella pneumoniae, Paenibacillus sp..
2 Janaúba 1 Isolado não identificado
3 Guananbi 3 B. sp..
4 Porteirinha 3
Lysinibacillus sphaericus, Stenotrophomonas sp., Streptomyces sp.
5 Nova Porteirinha 2 B. cereus, B. truringiensis.
6 Nova Porteirinha 6
B. licheniformis, B. pumilus, B. sp.,
Rhizobium sp..
7 Nova Porteirinha 2 B. pumilus
8 Nova Porteirinha 3 B. flexus, B. pumilus
9 Nova Porteirinha 2 Artrobacter sp..
10 Nova Porteirinha 2 B. sp..
11 Matias Cardoso 2 B. subtilis, Sporolactobacillus sp..
12 Jaíba 2 Isolados não identificados
13 Jaíba 0 -
14 Jaíba 0 -
15 Jaíba 2 B. tequilensis.
16 Jaíba 5
B. amyloliquefaciens, B. sp., B.
subtilis, Paenibacillus sp..
16 TOTAL 40

59
Verificou-se que apenas duas áreas de coleta não apresentaram isolados
com reações de PCR positivas para o gene nifH. A distribuição de isolados com
a presença desse gene ocorreu com frequência diferente entre os municípios
amostrados. A área 6 localizada em Nova Porteirinha apresentou maior número
de isolados com a presença desse gene. Além disso, observou-se que todas as
áreas localizadas na cidade de Nova Porteirinha apresentaram isolados com a
presença do gene para fixação de nitrogênio
Foi constatado que 40% dos isolados que apresentaram o gene nifH
pertencem ao gênero Bacillus spp. Os demais isolados pertencem aos gêneros:
Stenotrophomonas sp., Streptomyces sp. (Waksman and Henrici 1943),
Lysinibacillus sp.(Ahmed et al., 2007), Artrobacter sp.( Conn and Dimmick
1947 emend. Koch et al., 1995), Sporolactobacillus sp. (Kitahara and Suzuki
1963), Paenibacilllus sp, Rhizobium sp.(Frank 1889 emend. Young et al., 2001)
e a espécie Klebisiella pneumoniae.

60
5 DISCUSSÃO
Estudos entre plantas e bactérias endofíticas têm demonstrado a grande
importância dessa interação nos processos de adaptação das plantas aos diversos
ecossistemas bem como no incremento da sanidade e qualidade do solo. Pouco
se sabe sobre a relação dessas bactérias com a cultura da banana, sobre as
espécies colonizadoras e ainda sobre os benefícios instituídos por essa
associação nessa cultura.
No presente trabalho, foram isoladas e caracterizadas 201 bactérias
endofiticas provenientes de raízes de bananeira ‘Prata-Anã’, por meio da
associação de técnicas morfológicas e moleculares.
A densidade de bactérias endofíticas nos tecidos das raízes variou de
1x104 a 13x104 Unidades Formadoras de Colônias (UFC). Conforme Bandara et
al. (2006), bactérias endofíticas habitam as plantas, numa densidade entre 103 a
106 unidades formadoras de colônia (UFC) por grama de tecido fresco. Em
Araucaria angustifólia, o isolamento de bactérias apresentou valores de
aproximadamente 104 UFC por grama de raiz fresca (RIBEIRO e CARDOSO,
2011). Kuklinski-Sobral (2003) observou que a densidade bacteriana isolada de
plantas de soja sofre influência de fatores como cultivar, variações sazonais,
tratos culturais, estádio de desenvolvimento do hospedeiro e tecido amostrado.
Embora no presente trabalho as coletas tenham sido realizadas a partir do
mesmo tecido, raízes, e de uma mesma cultivar, Prata-Anã, as variações de
densidade obtidas se devem provavelmente às diferenças edafoclimáticas e ao
manejo cultural aplicado nas diferentes áreas de coleta.
Dos 201 isolados bacterianos coletados no presente estudo, 75% foram
classificados como Gram-positivas. Germida et al. (1998) isolaram bactérias
endofíticas de raízes de canola e observaram que a comunidade era dominada
por gêneros Gram-positivos. Todavia, Ikeda (2010) isolou 414 bactérias

61
endofíticas de raízes de milho, das quais analisou 217 quanto à morfologia,
classificando 77% como bacilos Gram-negativos e 23% como cocos Gram-
positivos. De acordo com Kobayashi e Palumbo (2000), tanto as bactérias
endofíticas Gram-positivas quanto as Gram-negativas foram isoladas de vários
tipos de tecidos de numerosas espécies de plantas.
Os resultados observados na literatura têm indicado que as bactérias
isoladas a partir de explantes contaminados de bananeira apresentam uma maior
proporção de bactérias do tipo Gram-negativas. Isso provavelmente ocorre
devido àquelas bactérias serem mais facilmente isoladas dos tecidos vegetais, de
fácil manipulação e passíveis de abordagens genéticas. Outro fator que pode ter
influenciado o maior isolamento de bactérias Gram-negativas é o meio utilizado
no isolamento. Existem diferenças dos meios de cultura quanto aos
microrganismos selecionados; o meio Mac Conkey, por exemplo, contém sais
biliares e cristal violeta que são inibidores do crescimento de bactérias Gram-
positivas, já o meio MAS é mais utilizado para crescimento destas bactérias
(ANVISA 2010).
Devido à importância das bactérias Gram-positivas, em diversas
interações benéficas têm sido estudados protocolos de detecção e diferentes
formas de isolamento destas bactérias, visto que as mesmas possuem
características interessantes que as distinguem dos seus homólogos Gram-
negativos (FRANCIS et al., 2010).
Estudos conduzidos com diferentes cultivares de bananeira indicaram
presença significativa de bactérias do tipo Gram-negativa. Nietsche et al. (2006),
ao caracterizarem os isolados de bactérias endofíticas provenientes de explantes
de bananeiras ‘Prata-Anã’, ‘FHIA 18’ e ‘SH 3640’, classificaram-nas como
bactérias do tipo Gram-negativas, com valores de 62,5%, 30% e 14% dos
isolados, respectivamente. Ganem (2009) isolou quinze bactérias de explantes
das bananeiras “Tropical” e “Galil 18” e 100% foram classificadas como

62
bactérias Gram-negativas; entretanto, neste estudo o meio utilizado no
isolamento foi BDA. Outro estudo publicado por Thomas et al. (2008)
demonstraram que as bactérias isoladas a partir de explantes contaminados da
cultivar Grand Naine eram predominantemente Gram-negativas.
Nos trabalhos citados anteriormente foram utilizados meios para
crescimentos de explantes, que possuem grande quantidade de vitaminas e
nutrientes. No trabalho de Thomas et al. (2008) foi utilizado o meio TSA,
provavelmente ambos favorecem maior desenvolvimento de bactérias Gram-
negativas.
Os resultados negativos para o teste de hipersensibilidade dos 201
isolados bacterianos sugerem a ausência de bacterias fitopatogênicas; entretanto,
este não deve ser utilizado como único parâmetro para descartar a
patogenicidade dos isolados. De acordo com Kobayashi e Palumbo (2000),
alguns gêneros de fitobactérias podem ser identificados como típicas bactérias
endofíticas, sem nenhum sintoma de doença na planta associada. Estudos
mostram que o estágio de desenvolvimento do hospedeiro e do patógeno,
mudanças ambientais e as respostas de defesa do hospedeiro (SCHULZ e
BOYLE, 2005), bem como fatores genéticos, estruturais e fisiológicos dos
mesmos, determinam se um microrganismo se comporta como endofítico ou
fitopatogênico (HEUER et al., 2002; ANDREOTE et al., 2008; KOGEL et al.,
2006). No presente estudo foi identificada bactéria da espécie Agrobacterium
tumefaciens que, apesar de ser descrita como fitopatogênica em outras plantas
hospedeiras, não apresentou sintomas de HR em plantas não hopedeiras,
comportando-se como bactéria endofítica em bananeira.
Sabe-se que a capacidade de as bactérias fitopatogênicas induzirem a
reação de hipersensibilidade reside na presença dos genes hrp. Esses genes são
responsáveis pelo crescimento de bactérias em plantas hospedeiras e em plantas
não hospedeiras pela reação de hipersensibilidade. Os genes hrp estão muito

63
próximos uns dos outros, formando um agregado, em que todos são necessários
para o crescimento da bactéria no hospedeiro e indução de hipersensibilidade em
plantas não hospedeiras (BERGAN FILHO et al., 1995). Assim, as plantas não
hospedeiras podem ser indicadoras de patogenicidade. Neste estudo não houve
reação de HR para as bactérias endofíticas as quais foram consideradas não
fitopatogênicas. Todavia, no caso de espécies fitopatogênicas, para confirmação
dos resultados é necessário realizar inoculação na cultura hospedeira e
identificar a espécie envolvida na interação (ROMEIRO, 1973).
A análise de variância molecular revelou baixa proporção da
diversidade, tanto por meio da técnica de rep-PCR quanto da ARDRA, entre os
locais de coleta. Entretanto, valores significativos foram obtidos dentro dos
locais de coleta. Esse resultado indica a existência de diferenças entre os
ambientes analisados os quais devem estar influenciando a evolução da
população das bactérias colonizadoras.
Resultados similares foram obtidos por Nakatani et al. (2009)
trabalhando com Xanthomonas axonopodis (Starr and Garces 1950 emend.
Vauterin et al., 1995) pv. Passiflora. Os autores verificaram que a maior parte
da variabilidade genética dos isolados está dentro das populações, 89,4%,
embora existam diferenças significativas entre as populações, 10,6%. Santos
(2009), analisando isolados de Bacillus cereus (Frankland and Frankland 1887)
provenientes de alimentos e do solo, observou através da análise de variância
molecular maior proporção de variação genética dentro dos grupos de isolados
(92,22%), enquanto que o valor entre os grupos foi de 7,78%.
As análises das sequências utilizando o programa BLASTn mostraram
alta diversidade de bactérias em raízes de bananeira ‘Prata-Anã’, pois foram
identificados gêneros distintos de bactérias. Vários estudos têm verificado
interação de diferentes bactérias com seus hospedeiros, com intuito de conhecer
as funções desempenhadas pelas mesmas.

64
A utilização prática de microrganismos endofíticos no controle biológico
de patógenos depende de inúmeros fatores relacionados à interação de bactérias
endofíticas-patógenos-planta, pois a competição existente entre eles pode reduzir
a eficiência do controle inviabilizando a sua utilização (AZEVEDO et al.,
2000). Assim, a identificação e o estudo de novas substâncias, a partir de
endófitos, que apresentam potencial para controle de fitopatógenos, bem como
para o crescimento das plantas, podem levar à descoberta de novos hormônios,
antibióticos e fungicidas mais eficientes para controle das doenças e para o
desenvolvimento das plantas.
As espécies Bacillus pumilus e Bacillus subtilis foram isoladas em
81,25% e 37,5% das áreas de coletas respectivamente, sendo os principais
representantes do gênero Bacillus. Esses dois microrganismos também foram
encontrados colonizando o mesmo nicho ecológico em quatro áreas. Estes
resultados indicam que provavelmente existe uma relação de simbiose entre eles.
De acordo com Raupach e Kloepper (1998), a mistura dessas duas espécies e
Curtobacterium flaccumfaciens (Hedges 1922) Collins and Jones 1984 aplicadas
nas sementes estimulou o crescimento de plântulas de pepino e reduziu os
sintomas causados por patógenos. Em geral, há um certo grau de especificidade
entre gêneros encontrados e seus hospedeiros, entretanto, estas interações são
pouco conhecidas. Dessa forma, é possível que várias espécies se
interrelacionem e possuam relações mutualísticas, no entanto, mais estudos
precisam ser realizados no intuito de entender como ocorre esse processo nas
diferentes culturas.
Verificou-se que muitos gêneros, como Bacillus e algumas espécies
descritas neste estudo, foram também encontrados em outros trabalhos de
isolamento de bactérias endofíticas. Ferreira (2008), após identificação da
comunidade endofítica das raízes de eucalipto, por meio do sequenciamento
parcial do 16S rDNA, identificaram-se Psychrobacter sp. (Juni and Heym

65
1986), Bacillus cereus, Bacillus pumilus, Lysinibacillus sphaericus, Rhizobium
tropici (Martínez-Romero et al., 1991), Rahnella aquatilis (Izard et al., 1981),
Erwinia persicina (Hao et al., 1990), Stenotrophomonas maltophilia (Hugh
1981) Palleroni and Bradbury 1993, como espécies colonizadoras. Os gêneros
Bacillus, Enterobacter e Pantoea foram identificados como endofíticos em
diversas plantas, tais como citros, cana-de-açúcar e soja (GURTLER et al.,
2005; MEDRANO e BELL, 2007). Espécies pertencentes aos gêneros:
Alcaligenes (Castellani and Chalmers 1919), Azospirillum (Tarrand et al., 1979
emend. Falk et al., 1985), Bacillus, Enterobacter, Herbaspirillum (Baldani et al.,
1986 emend. Baldani et al., 1996), Klebsiela, Pseudomonas, Burkholderia,
Paenibacillus, Acetobacter e a família Rhizobiaceae têm sido observadas em
associação com plantas de arroz, milho, sorgo, trigo, soja, feijão, tomate,
algodão e cana-de-açúcar (DI CELLO et al., 1997; BURDMAN et al., 1997;
BALDANI et al., 1997; REINHOLD-HUREK; HUREK, 1998; HALLMANN et
al., 1997; BALDANI et al.,2000).
O gênero Bacillus foi encontrado com maior frequência em todas as
áreas amostradas. De acordo com Hallmann et al. (1997), este gênero foi
relatado em vários trabalhos, como a principal bactéria endofítica de
determinadas plantas. As espécies: Bacillus cereus, Bacillus subtilis Bacillus
megaterium (Bary, 1884), e Bacillus pumilus já foram descritas como
endofíticas (BAI et al., 2002; ELVIRA-RECUENCO e VAN VUURDE, 2000;
ARAÚJO et al., 2002). Bactérias deste gênero já foram isoladas em explantes
contaminados de banana (BRAGA et al., 2001; ODUTAYO et al., 2007).
A diversidade de gêneros amostrados neste estudo ratifica que existe
uma alta variabilidade genética de bactérias endofíticas interagindo com a
cultura da bananeira. Essas interações são indispensáveis para os processos de
estabelecimento e crescimento das plantas. Bactérias pertencentes ao gênero
Bacillus, por exemplo, são classificadas como promotoras de crescimento, com

66
destaque à produção de auxinas e giberelinas por Bacillus pumilus, fixadoras de
nitrogênio, solubilizadoras de fosfato (FORCHETTI et al., 2007; STURZ;
NOWAK, 2000). Além disso, são capazes de induzir resistência sistêmica nos
vegetais (JUNG et al., 2006; ONGENA; JACQUES, 2008), e atuam no
biocontrole de doenças de plantas, demonstrando grande potencial para
utilização na agricultura, com a redução do uso de produtos químicos no
controle de pragas (LAÇAVA et al., 2004). A espécie Bacillus subtilis já foi
descrita como sendo capaz de incrementar a nodulação de raízes e promover o
crescimento de plantas de feijoeiro (LAZZARETTI e MELO, 2005).
Foram identificados dois isolados como pertencentes à espécie Bacillus
thuringiensis (Berliner, 1915). Resultado semelhante foi descrito por Teixeira et
al. (2007), que encontraram essa bactéria colonizando endofiticamente partes de
plantas de mandioca. Segundo o mesmo autor, a principal função relacionada a
esta espécie é o controle biológico, além de a mesma apresentar atividade
inseticida (ROSAS-GARCÍA, 2009). A espécie Bacillus megaterium e bactérias
do gênero Arthrobacter foram descritas com capacidade de reduzir metais
pesados e aumentar a disponibilidade de ferro para os vegetais (VALENCIA-
CANTERO et al., 2007). Espécies de Bacillus cereus têm a capacidade de suprir
doença através da produção de antibióticos (EMMERT e HANDELSMAN ,
1999).
De todos os isolados recuperados nas áreas de coleta, o gênero Bacillus
esteve presente em todas elas. Isso pode estar relacionado à especificidade deste
gênero pelo meio de isolamento utilizado neste estudo. Teixeira et al (2007),
analisando isolamento de bactérias endofíticas em mandioca, identificaram o
gênero Bacillus em todas as áreas amostradas, utilizando o meio TSA na
recuperação das bactérias. Conforme Vieira e Nahas (2000), fatores como
temperaturas, diluição e meios de cultura, isoladamente ou associados,
influenciam o crescimento de Bacillus spp.

67
Apesar da predominância de isolados do gênero Bacillus, outros gêneros
com potencial de utilização foram também isolados. Espécies do gênero
Paenibacillus sp. têm sido caracterizadas em função de sua capacidade de
produzir hormônios indutores de crescimento da planta, como auxinas e
citocininas, enzimas como quitinases, amilases, proteases, e antibióticos, e
também são solubilizadoras de fosfatos orgânicos (COELHO et al., 2009). Da
mesma forma, foi relatado que a espécie Paenibacillus polymyxa aumenta o
crescimento de plantas através da produção de citocinina e auxina (LAL e
TABACCHIONI, 2009).
Outro gênero encontrado como bactéria colonizadora de raízes de
bananeira ‘Prata-Anã’ foi o Stenotrophomonas. Representantes deste gênero são
potenciais produtores de ácido indol acético (AIA) (PARK et al., 2005) e atuam
na biodegradação de contaminantes, como 4-nitroanilina (QURESHI et al.,
2007).
Vários estudos descrevem as principais funções desempenhadas por
bactérias endofíticas. Espécies do gênero Enterobacter (Hormaeche and
Edwards, 1960) e Pseudomonas (Migula, 1894) são capazes de fixar nitrogênio
e solubilizar fosfato (ASIS e ADACHI, 2003; VERMA et al., 2001; DALTON
et al., 2004; RODRIGUEZ e FRAGA, 1999), o que aumenta o crescimento e a
produtividade das plantas. Já o gênero Streptomyces exerce uma atividade de
biocontrole sobre fungos fitopatogênicos (PARK et al., 2002). Diversos gêneros
bacterianos têm sido descritos como produtores de AIA e solubilizadores de
fosfato, entre eles, Acetobacter, Agrobacterium e Pantoea tem sido relatadas
como estimuladoras do crescimento vegetal (PATTEN; GLICK, 1996;
RODRIGUEZ; FRAGA, 1999). Além disso, um grupo diversificado de bactérias
Gram-positivas como Arthrobacter, Micrococcus, Bacillus, Rhodococcus (Zopf,
1891), Mycobacterium (Lehmann and Neumann 1896), Streptomyces e
Corynebacterium (Lehmann and Neumann 1896 emend. Bernard et al., 2010) é

68
capaz de produção de auxina, que pode estimular a absorção de nutrientes e
proliferação de raízes (TSAVKELOVA et al, 2006;. SPAEPEN et al., 2007).
Isolados do gênero Rhizobium foram identificados neste estudo, os
mesmos são utilizados como biofertilizante, uma vez que algumas espécies
podem suplementar a ausência de fertilizante nitrogenado através da promoção
do crescimento e da fixação de nitrogênio (CHOUDHURY e KENNEDY,
2004). Isso mostra que a interação plantas/Rhizobium é indispensável para
ambos os organismos, pois é classificada como uma relação simbiótica em que
ambos os organismos são beneficiados.
A espécie Klebsiella pneumoniae identificada neste estudo já foi descrita
em 50% dos isolamentos associados à cultivar Tropical (GANEM et al., 2009).
Espécies do gênero Klebsiella são fixadoras de nitrogênio e são consideradas
como microrganismo endofítico associado a raízes de plantas (SHIOMI et al.,
2006 ). A espécie Micrococus luteus foi isolada neste estudo, e pesquisas com
microrganismos pertencentes à família Micrococcineae revelam que os mesmos
apresentam potencialidades para propósitos biotecnológicos, bem como para
usos farmacêuticos e de biocontrole (AZEVEDO et al., 2000). Já a bactéria
Agrobacterium tumefaciens, apesar de já ter sido descrita como fitopatogênica,
apresenta potencial para o controle biológico de doenças fúngicas e bacterianas
(ARAUJO, 2000).
Foram constatadas diferenças na diversidade de gêneros encontrados nas
diferentes áreas de coletas. A área 4 é a mais diversa tendo sido isolados cinco
gêneros diferentes. Estes resultados podem estar relacionados à presença de
condição favorável para o estabelecimento de algumas bactérias endofíticas
nessa área. Fatores como o sistema agrícola utilizado, o uso ou não de
agroquímicos, genótipo da planta, estádio fenológico e tipo de solo
(DALMASTRI et al., 1999; FROMIN et al., 2001), provavelmente, influenciam
de alguma forma a diversidade e riqueza das populações de bactérias, sendo

69
responsáveis pela diferenças entre as espécies encontradas nas diferentes áreas
de coletas. Resultados que corroboram estas informações foram observados por
Kuklinsky-Sobral et al. ( 2004) que verificaram que a presença de diferentes
espécies de bactérias endofíticas na cultura da soja depende
do genótipo, da idade da planta, do tecido da amostra e também da época de
isolamento. O tipo de solo determinou diferenças na população endofítica
isoladas de trigo (CONN e FRANCO, 2004).
Contrastando os dados coletados com a aplicação dos questionários nas
áreas de coleta, notou-se que, na maioria das áreas, o bananal tem a mesma
idade e os tratos culturais são praticamente os mesmos. Um fato interessante da
área onde houve mais diversidade é que há incorporação de matéria orgânica,
porém isto não explica a diversidade, pois em outras áreas também é praticado
tal técnica.
Verifica-se que são raros os estudos que relacionam a comunidade
endofítica dos vegetais com manejo agrícola e tipos de solos. Seghers et al.
(2004), comparando sistemas de cultivo orgânico e tradicional, observaram
diferenças na comunidade de microrganismos endofíticos nas raízes de milho.
Os autores relataram, porém, que não fica claro pelos resultados obtidos se as
diferentes práticas agrícolas afetaram diretamente a comunidade endofítica das
raízes, ou indiretamente, por meio de mudanças na comunidade total de
microrganismos do solo.
Quando se compara a diversidade genética gerada através da técnica
Rep-PCR, observa-se que o primer Box permitiu a formação de um maior
número de bandas polimórficas, apresentando maior poder discriminatório entre
os isolados bacterianos. Segundo Van Berkum (1999), as regiões BOX, por
estarem associadas a graus elevados de polimorfismos, tem participação em
processos de evolução adaptativa, mediando a interação dos microrganismos
com ambientes inóspitos ou adversos.

70
Apesar dos diferentes níveis de diversidade observados entre as
populações de isolados, nenhum primer utilizado com a técnica rep-PCR
apresentou bandas que relacionasse os isolados com suas áreas de coleta. Costa
et al. (2007), ao estudarem a diversidade molecular de Ralstonia solanacearum
(Smith 1896) Yabuuchi et al., 1996, perceberam, por meio dos perfis
eletroforéticos gerados por BOX-PCR, um alto grau de polimorfismo; no
entanto não foi observado padrão de amplificação específico para os isolados
pertencentes aos dois ecossistemas utilizados no trabalho (várzea e terra-firme)
ou às diferentes biovares. Entretanto, Silva et al. (2007) verificaram através da
caracterização molecular por rep-PCR que a diversidade genética entre os
isolados de Xanthomonas albilineans (Ashby 1929) Dowson 1943 apresentaram
certa relação com a região de origem.
Em geral, a utilização dos oligonucleotideos rep-PCR tem permitido a
caracterização de bacterias Gram-positivas e Gram-negativas, devido a
complexidade dos produtos gerados pela amplificação por PCR. Como a técnica
é baseada na amplificação de regiões repetitivas do genoma, ela tem sido
amplamente utilizada, por ser considerada de fácil execução, principalmente
para análise de grandes populações bacterianas (MEHTA et al., 2002). De
acordo com Versalovic et al. (1998), as sequências repetitivas estão por todo o
genoma, por isso a técnica de rep-PCR tem revelado ser potencialmente capaz
de caracterizar o genoma completo, demonstrando poder de diferenciação
semelhante ou melhor que outros métodos para análise de microrganismos.
Quando se comparam os valores de distância genética gerada pelos
dados de rep-PCR, entre áreas localizadas dentro do mesmo município, Nova
Porteirinha demonstrou o menor nível de variação. Uma das razões se deve,
provavelmente, à pequena distância geográfica média de 8,3Km entre as áreas
nas quais as amostras foram obtidas. No entanto, no município de Jaíba, a
diferença entre as distâncias genéticas foi superior, com uma variação de 9,4

71
pontos percentuais entre a menor e a maior área. Seguindo o mesmo raciocíonio,
pode-se considerar que a maior variação deve estar associada à maior distância
entre as áreas de coleta, que foi de 16,5 Km. Além disso, fatores como manejo e
tipo de solo podem influenciar a comunidade bacteriana, favorecendo assim o
desenvolvimento de diferentes microrganismos.
A análise do dendograma formado através da matriz de distância com os
dados dos primers da técnica de rep-PCR demonstra que não houve nenhum
critério de agrupamento. O grupo III apresentou maior diversidade de isolados,
uma vez que possui representantes de oito gêneros e de nove espécies diferentes.
Segundo Assumpção (2009), a diversidade e a estrutura de comunidades
microbianas variam com o tamanho da amostra, pois, quanto maior o tamanho
da amostra, maior a probabilidade de serem amostradas espécies raras.
Analisando a estrutura dos grupos formados, observa-se que apesar de
ter sido feito o isolamento em apenas uma área nos municípios de Porteirinha e
Matias Cardoso, seus representantes estão distribuídos por quatro e cinco
grupos, respectivamente, demonstrando que nesses locais a diversidade é alta.
O grupo V, apesar de ser formado por indivíduos de diferentes áreas de
coleta, apresentou menor variabilidade genética entre os isolados dos quais
muitos pertencem ao mesmo gênero. Como a técnica utilizada é baseada na
amplificação de regiões repetitivas do genoma, pode ter acontecido que mesmo
sendo diferentes, a região amplificada foi a mesma para todos os isolados, não
permitindo a diferenciação entre eles. Todavia, Antunes (2010) descreve que o
padrão de amplificação dos diferentes primers não é idêntico, uma vez que
podem ser formados diferentes números de bandas por oligonucleotídeo, e
também a condição de anelamento e a prevalência ou distribuição dos elementos
repetitivos podem variar em cada isolado.
Os resultados obtidos pela análise conjunta dos primers demonstraram
diferenças entre os isolados. Conforme Rademaker et al. (2000), a combinação

72
dos dados de REP, ERIC e BOX-PCR evidentemente é mais significante e
consistente, pois o número total de amplicons é consideravelmente aumentado.
Além do mais, o genoma é mais amplamente coberto, visto que algumas regiões
podem ter mais cópias de um determinado elemento do que outras.
Nota-se que fatores genéticos e ecológicos também influenciam a
diversidade genética dos microrganismos. Segundo Arber (2000), as principais
estratégias genéticas responsáveis pelo surgimento e amplificação da diversidade
genética são mutação, recombinação e aquisição de sequências de DNA de
outros organismos, as chamadas transferências horizontais gênicas. Já os fatores
ecológicos estão ligados à oportunidade de colonizar um hospedeiro e à
competição entre os isolados (SPIERS et al., 2000).
A técnica ARDRA foi eficiente e identificou variação entre os isolados
bacterianos avaliados, tanto pelo polimorfismo apresentado quanto pelas
estimativas de distância genética. Resultados similares foram obtidos por
diversos autores em diferentes culturas. Cruz et al. (2001), trabalhando com
bactérias endofíticas fixadoras de nitrogênio isoladas a partir de plantas de
bananeira e abacaxizeiro, observaram que as enzimas apresentaram variação nos
tamanhos dos fragmentos gerados, cuja enzima HaeIII foi capaz de gerar dez
tipos de perfis, seguida por AluI com sete tipos, e HinfI e RsaI com cinco tipos
cada.
Em relação às distâncias genéticas geradas entre as áreas, constatou-se
que aquelas localizadas no mesmo município apresentaram-se próximas
geneticamente, possivelmente por estarem a uma distância geográfica também
próxima. Dessa forma, as áreas mais próximas geograficamente revelaram
valores similares de distância genética, logo, quanto menor a distância
geográfica, maior a chance de isolar indivíduos semelhantes geneticamente.
O grande número de grupos formados no presente estudo confirma o
potencial da técnica nos estudos visando à avaliação da variabilidade bacteriana.

73
Michaud et al. (2004) analisaram a diversidade de linhagens isoladas de
Brevibacterium spp (Breed, 1953), agrupando os mesmos em 52 grupos, usando
apenas a enzima de restrição AluI.
Nao houve nenhum critério para a formação dos agrupamentos; todavia,
o isolado 161 identificado como B. sp., o isolado 112 que não foi identificado, o
7 identificado como Agrobacterium tumefaciens, o 189 como Aneurinibacillus
sp, 140 como B. sp apresentam características peculiares que os distinguem dos
demais, sendo os únicos representantes dos grupos II, V, XIII, XIV e XV
respectivamente. Dentre os dezesseis grupos formados, o grupo IV é
considerado o mais biodiverso, já que possui isolados de sete gêneros e de sete
espécies diferentes. As variações registradas entre os dezesseis grupos se devem
ao fato de a técnica gerar perfis de bandas diferentes, e estas alterações revelam
a diversidade entre os e dentro dos grupos bacterianos, agrupando os indivíduos
de genótipos semelhantes (MOCALI et al., 2003; KUKLINSKYSOBRAL et al.,
2004).
Ao analisar os dendrogramas depois da identificação das espécies, foi
possível observar que o gênero Bacillus está distribuído por todos os grupos,
pois possuem maior número de representantes. O isolado 7 identificado como
Agrobacterium tumefaciens em ambos os dendrogramas formou um grupo
isolado, já que é uma espécie que apresenta características bem diferentes das
outras. Os gêneros Pseudomonas e Enterobacter pertencem ao mesmo grupo,
isso porque apresentam semelhanças, ambos são Gram-negativos e estão
agrupados na mesma divisão gammaproteobactéria. A maioria dos gêneros da
divisão Firmicutes está agrupada próximo ou no mesmo grupo, confirmando que
as técnicas moleculares foram eficientes na formação dos agrupamentos
filogenéticos, baseando-se nas diferentes características dos isolados.

74
As enzimas utilizadas demonstraram alto grau de polimorfismo dos
genes ribossomais, gerando perfis de amplificação variáveis. De acordo com
Andreote (2007), os genes de rDNA utilizados são considerados cronômetros
evolutivos, pois são genes que apresentam uma taxa de mutação mínima, cujo
produto da expressão é funcional e essencial para a vida do organismo. Além
disso, estes genes apresentam sítios diferenciais de evolução rápida ou lenta, o
que permite a avaliação das relações filogenéticas entre organismos em
diferentes distâncias filogenéticas.
O estudo da presença do gene nifH demonstrou que 20% das bactérias
endofíticas avaliadas no presente estudo podem agir como fixadoras de
nitrogênio. Perin (2007) observou que dos 80 isolados obtidos de cana-de-
açúcar, 80% apresentaram o gene nifH. Videira et al. (2009), avaliando a
diversidade de bactérias fixadoras de nitrogênio associadas a plantas de arroz,
isolaram 46 bactérias e todas amplificaram o gene nifH.
Uma das explicações da presença do gene nifH pode estar associada ao
tecido dos quais os isolados foram obtidos, raízes, habitats de preferência destes
indivíduos. Chelius e Tripplet (2000) e Miche et al. (2006) demonstraram a
expressão do gene nifH de Klebsiella pneumoniae no interior de raízes de milho,
e de Azoarcus sp. em raízes de arroz, respectivamente.
Apesar de 40 isolados bacterianos apresentarem o gene nifH neste
estudo, não se pode afirmar que eles sejam potenciais fixadores de nitrogênio,
pois a simples existência deste gene não representa, necessariamente, atividade
da enzima nitrogenase (DEAN e JACOBSON, 1992). Por outro lado, sua
presença no genoma é uma forte evidência de que a bactéria pode ser fixadora de
nitrogênio. Logo, a presença do gene nifH, que codifica a unidade Fe-
nitrogenase do complexo nitrogenase, tem se tornado um marcador muito
utilizado no estudo da diversidade de endofíticos com potencial para fixar N2,
em estudos independentes de cultivo (ISQUIERDO e NUSSLEIN, 2006).

75
Observou-se que isolados bacterianos expressando o gene nifH
estiveram presentes em 14 áreas de coleta. Porém, sua presença foi marcante em
todas as áreas do município de Nova Porteirinha. Provavelmente, fatores como
tipo de solo, mudanças climáticas ou até mesmo os tratos culturais das áreas
influenciaram a permanência dessas bactérias. Roesch et al. (2007) avaliaram a
diversidade de bactérias diazotróficas endofíticas associadas a plantas de milho,
e detectaram diferenças na distribuição das sequências do gene nifH entre
regiões, em que a região sul do Rio Grande do Sul apresentou menor
diversidade em comparação com ao norte do Estado. Segundo os autores, essas
diferenças podem estar relacionadas ao teor de argila dos solos e à distribuição
de chuvas das regiões amostradas. Além do mais, nesses ambientes pode haver
maior ocorrência de bactérias diazotróficas, visto que o estudo da sequência do
gene nifH é normalmente empregado para caracterizar a diversidade genética
dessas bactérias, o que explicaria este resultado.
A análise da presença do gene nifH foi realizada em 40 isolados. Destes
a maioria foi identificada como pertencentes ao gênero Bacillus, que já foi
relatado em vários estudos como potencial fixador de nitrogênio (RÓZCKI et
al., 1999; RAYMOND et al., 2004). O gênero Rhizobium também apresentou a
presença desse gene, o que já era esperado, uma vez que é classificado como
fixador de nitrogênio que são capazes de formar nódulos em associação com
leguminosas (YOUNG, 1996). A espécie Klebsiella pneunomiae apresentou a
banda característica do gene nifH. Segundo Arnold et al. (1988), os primeiros
genes envolvidos na fixação do nitrogênio foram identificados e estudados
utilizando se a bactéria Klebsiella pneumoniae. Dois gêneros da divisão
Actinobacteria e o Stenotrophomonas também possuem o gene nifH. Resultados
semelhantes foram encontrados por Assumpção (2008) que identificou quatro
isolados dessa mesma divisão, com potencial para fixação de nitrogênio. Já
Teixeira et al. (2007) isolaram o gênero Stenotrophomonas de mandioca, e o

76
mesmo apresentou o gene nifH, porém, segundo os mesmos autores, não há
relatos desse organismo como diazotrófico em outras culturas.
Portanto, as bactérias identificadas neste estudo têm potencial para
promoção de crescimento, solubilização de fosfato, controle biológico e fixação
de nitrogênio em bananeira. De acordo com Nowak (1998), a aplicação de
bactérias endofíticas em plantas provenientes de cultura de tecidos pode
favorecer o desenvolvimento de espécies micropropagadas, como a banana, pois,
ocorreria um enriquecimento biológico das mudas, além de evitar a competição
dessas bactérias com outras que colonizam a planta no ambiente. Assim, após o
estudo e confirmação dessas funções nesta cultura, as bactérias poderão ser
utilizadas dentro de um pacote tecnológico de produção de mudas
micropropagadas.

77
6 CONCLUSÕES
As caracterizações morfológica e molecular dos isolados endofíticos
colonizadores de raízes de bananeira ‘Prata-Anã’ revelam alta
diversidade genética.
Os marcadores REP, ERIC e BOX-PCR em conjunto demonstram
diversidade genética entre os isolados estudados, sendo que o BOX-PCR
apresenta maior polimorfismo.
Na análise de ARDRA, a endonuclease RsaI apresenta maior grau de
polimorfismo. Esta técnica possui alto poder discriminatório, e forma 16
grupos bem distintos de isolados bacterianos.
As análises das sequências parciais de nucleotídeos da região 16S rDNA
permitem separar os 143 isolados em 17 gêneros diferentes .O gênero
Bacillus é o que apresenta maior frequência nas áreas de coletas.
Quarenta isolados apresentam reações de PCR positivas para o gene
nifH.

78
REFÊRENCIAS BIBLIOGRÁFICAS
ABANORTE - Associação dos Bananicultores do Norte de Minas Gerais. Disponível em: <http://www.abanorte.com.br/contact-info>. Acesso em: 05 jan. 2008.
ABD-ELSALAM, K.A. et al. PCR identification de Fusarium genus based on nuclear ribosomal-DNA sequence data. African Journal of Biotechnology, Nairobi, v. 2, p. 82-85, 2003.
AGENCIA NACIONAL DE VIGILÂNCIA SANITÁRIA (ANVISA). Descrição dos meios de cultura empregados nos exames microbiológicos. Disponível em: www.anvisa.gov.br/servicosaude/microbiologia. Acesso em 08 de agosto de 2011.
AGRIANUAL . Anuário da agricultura brasileira. São Paulo: FNP Consultoria e Comércio, p 170-180, 2011.
ALMEIDA, C. O. de; SOUZA, J. da S.; CORDEIRO, Z. J. M. Aspectos socioeconômicos. In: CORDEIRO, Z. J. M. Banana produção: aspectos técnicos. Brasília: Embrapa Comunicação para transferência de tecnologia, 2000, p. 10-11.
ALTSCHUL, S.F. et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research, Oxford, v. 25, p. 3389-402, 1997.
ANDREOTE, F. D. Fatores determinantes na composição da comunidade bacteriana associada às Plantas. 2007. 201 p. Tese (Doutorado em Genética e melhoramento de plantas)- Escola Superior de Agricultura Luiz de Queiroz, Piracicaba, 2007.

79
ANDREOTE, F. D. et al. Transgenic tobacco revealing altered bacterial diversity in rizosphere during early plant development. Antonie van Leeuwenhoek, Amsterdam, v. 93, p. 415-424, 2008.
ANTUNES, J. E. L. Diversidade genética e eficiência simbiótica de isolados de rizóbios nativos em feijão-fava( Phaseolus lunatos L.). 2010. 108 p. Dissertação (Mestrado em Agronomia) - Universidade Federal do Piauí, Piauí. 2010.
ARAUJO, W. L. A comunidade bacteriana endofítica de citros e sua interação com Xylella fastidiosa, agente causal da Clorose Variegada dos Citros (CVC). 2000. 131 p. Tese (Doutorado em Microbiologia) - Universidade de São Paulo (USP), Piracicaba, 2000.
ARAÚJO, J. M.; SILVA, A. C.; AZEVEDO, J. L. Isolation of endophytic actinomycetes from roots and leaves of maize (Zea mays L.). Brazilian Archives of Biology Technology, Piracicaba, v. 43, p. 447-451, 2000.
ARAÚJO, W. L. et al. Diversity of endophytic bacterial populations and their interactions with Xylella fastidiosa in Citrus plants. Applied and Environmental Microbiology, Netherlands, v.68 , p.4906-4914, 2002.
ARBER, W. Genetic variation: molecular mechanism and impact on microbial evolution. FEMS Microbiology Reviews, Amsterdam, v. 24, p. 1-2, 2000.
ARNOLD, W. et al. Nucleotide sequence of a 24206 base pair DNA fragment carrying the entire nitrogen fixation cluster of Klebsiella pneumoniae. Journal of Molecular Biology, Londres, vol.230, p. 714-738, 1988.
ASHBOLT, N. J.; INKERMAN, P.A. Acetic acid bacterial biota of the pink sugar cane mealybug Saccharococcus sacchari, and its environs. Applied and Environmental Microbiology, Washington, v. 56, p.707–712, 1990.

80
ASIS, C. A.; ADACHI, K. Isolation of endophytic Pantoea agglomerans and nondiazotrophic Enterobacter asburiae from sweetpotato stem in Japan. Letters in Applied Microbiology, Oxford , v. 38, p. 19-23, 2003.
ASSUMPÇÃO, L.C. Diversidade da comunidade bacteriana endofítica de sementes de soja e o seu potencial biotecnológico. 2008. 93 p. Dissertação (Mestrado em Microbiologia Agrícola) - Universidade de São Paulo Escola Superior de Agricultura Luiz de Queiroz. Piracicaba, 2008.
ASSUMPÇÃO, L. C. et al. Diversidade e potencial biotecnológico da comunidade bacteriana endofítica de sementes de soja. Pesquisa Agropecuária Brasileira, Brasília, v. 44, p. 503, 2009.
AZEVEDO, J. L. Microrganismos endofíticos. In: MELO, I. S.; AZEVEDO, J. L. (Ed.) Ecologia microbiana. Jaguariúna: EMBRAPA, 1998. p. 117-137.
AZEVEDO, J. L.; ARAÚJO, W. L.; MACHERONI Jr, W. Importância dos microrganismos endofíticos no controle de insetos. In: MELO, I. S.; AZEVEDO, J. L. Controle Biológico.1°. Jaguariúna: EMBRAPA - Meio Ambiente, 2000. p. 57-59.
AZEVEDO, J. L.; ARAUJO, W. L. Diversity and applications of endophytic fungi isolated from tropical plants. In: GANGULI, B. N.; DESHMUKH, S. K. Fungi: multifacetated microbes. Boca Raton: CRC Press, Anamaya Publishers, 2007. p. 189-207.
BAI, Y. et al. Isolation of plantgrowth- promoting Bacillus strains from soybean root nodules. Canadian Journal of Microbiology, Ottawa , v. 48, p. 230-238, 2002.
BALDANI, J. L. et al.. Recent advances in BNF with non-legume plants. Soil Biology Biochemistry, Oxford, v. 9, p. 911-922, 1997.

81
BALDANI, J. I. et al.. Biological nitrogen fixation (BNF) in non-leguminous plants: the role of endophytic diazotrophs. Seropédica: Embrapa Agrobiologia, 2000. p. 397-400.
BANDARA, W. M. M. S.; SENEVIRATNE, G.; KULASOORIYA, S. A. Interactions among endophytic bacteria and fungi: effects and potentials. Journal of Biosciences, Bangalore, v. 31, p. 645-650, 2006.
BARROSO, C. B.; NAHAS, E. Solubilização do fosfato de ferro em meio de cultura. Pesquisa Agropecuária Brasileira, Brasília, v. 43, p. 529-535, 2008.
BELL, C. R. et al.. Endophytic bacteria in grapevine. Canadian Journal of Microbiology, Ottawa, v. 41, p. 46-53, 1995.
BENT, E.; CHANWAY, C. P. The growth-promoting effects of a bacterial endophyte on lodgepole pine are partially inhibited by the presence of other rhizobacteria. Canadian Journal of Microbiology, Ottawa, v. 44, p. 980-988, 1998.
CAMARGO, L. E. A. Análise genética da resistência e da patogenicidade. In: BERGAN FILHO, A.; KIMATI, H.; AMORIN, L. Manual de fitopatologia. 3. ed. São Paulo: Ave Maria Ltda, 1995. p. 470-483.
BRAGA, M. F; LISEI DE SA, M. E; MUSTAFÁ, P. C. Avaliação de um protocolo para multiplicação in vitro da bananeira (Musa sp.) cv. caipira (AAA). Revista Brasileira de Fruticultura, Jaboticabal, v. 23, n. 2, p. 215-219, 2001.
BURDMAN, S.; KIGEL, J.; OKON, Y. Effects of Azospirillum brasilense on nodulation and growth of common bean (Phaseolus vilgaris L.). Soil Biology & Biochemistry, Rehovot, v.29, p.923-929, Jul 1997.
CARROLL, G. Fungal endophytes in stems and leaves: from latent pathogen to mutualistic symbiont. Ecology, Brooklyn, v. 69, p. 2-9, 1988.

82
CAVALCANTE, V. A.; DÖBEREINER, J. A new acid-tolerant nitrogen-fixing bacterium associated with sugarcane. Plant and Soil, Dordrecht, v.108, p. 23-31, 1988.
CHANWAY, C. P. Inoculation of tree roots with PGPR soil bacteria: an emerging technology for reforestation. Forest Science, Bethesda, v. 43, p. 99-112, 1997.
CHOUDHURY, A. T. M. A.; KENNEDY, I. R. Prospects and potentials for systems of biological nitrogen fixation in sustainable rice production. Bio Fertil Soils, Berlin, v. 39, p. 219-277, 2004.
CHELIUS, M.K.; TRIPLETT, E.W. Immunolacalization of dinitrogenase reductase produced by Klebsiella pneumoniae in association with Zea mays L. Applied and Environmental Microbiology, Washington, v. 66, p.783-787, 2000.
CHENEBY, D. et al. 16S rDNA analysis for characterization of denitrifying bacteria isolated from three agricultural soils. FEMS Microbiology Ecology, Amsterdam, v. 34, p. 121-128, 2000.
CHUEIRE, L. M. O. et al. Identificação das estirpes de Bradyrhizobium e Rhizobium utilizadas em inoculantes comerciais para as culturas da soja e do feijoeiro pela técnica de PCR com "primers" aleatórios ou específicos. Agricultura Tropical, Cuiabá, v. 4, p. 80-95, 2000.
CHUEIRE, L. M. O. et al. Classificacao taxonomica das estirpes de rizobio recomendadas para as culturas da soja e do feijoeiro baseada no sequenciamento do gene 16S rRNA. Revista Brasileira de Ciência do Solo, Viçosa, v. 27, p. 833-840, 2003.
COELHO, M. R. R. et al. Método Molecular Para Estudos Ecológicos de Bactérias Diazotróficas do Gênero Paenibacillus em Amostras Ambientais. Sete Lagoas: Embrapa Milho e Sorgo, 2009. 27 p. (Boletim de Pesquisa e Desenvolvimento, Sete Lagoas).

83
COLOMBO, P. M. Occurrence of endophytic bacteria in Siphonous algae. Phycologia, Padova, v.17, p.148-151, 1978.
CONN, V. M.; FRANCO, C. M. M. Analysis of the endophytic actinobacterialpopulation in the roots of wheat (Triticum aestivum L.) by terminal restriction fragment length polymorphism and sequencing of 16S rRNA clones. Applied And Environmental Microbiology, Washington, v. 70, p. 1787-1794, 2004.
COSTA, S. B.; FERREIRA, M. A. S. V.; LOPES, C. A. Diversidade patogênica e molecular de Ralstonia solanacearum da região amazônica brasileira. Fitopatologia Brasileira, Brasília, v. 32, p. 285-294. 2007.
CRUZ, L. M. et al. 16S ribosomal DNA characterization of nitrogen-fixing bacteria isolated from banana (Musa spp.) and pineapple (Ananas comosus (L.) Merril). Applied and Environmental Microbiology, Washington, v.67, p. 2375-2379, 2001.
DALTON, D. A. et al. Endophytic nitrogen fixation in dune grasses (Ammophila arenaria and Elymus mollis) from Oregon. FEMS Microbiology Ecology, Amsterdam, v. 49, p. 469-479, 2004.
DALMASTRI, C. et al. Soil type and maize cultivar affect the genetic diversity of maize root associated Burkholderia cepacia populations. Microbial Ecology, Oslo, v. 38, p. 273-284, 1999.
DEAN DR, JACOBSON MR. Biochemical genetics of nitrogenase. In: STACY G, BURRIS RH, EVANS HJ. Biological nitrogen fixation. New York: Chapman e Hall, p. 763-834, 1992.
De BRUJIN, F. J. Use of repetitive (repetitive extragenic palindromic and enterobacterial repetitive intergenic consensus) sequences and the polymerease chain reaction to fingerprint the genomes of Rizhobium meliloti isolates and other soil bacteria. Applied and Environmental Microbiology, Washington, v. 58, p. 2180-2187, 1992.

84
DESAINT, S. et al. Genetic diversity of carbofuran-degrading soil bacteria. FEMS Microbiology Ecology, Cumbria, v. 34, p.173-180 2000.
DI CELLO, F. et al. Biodiversity of a Burkholderia cepacia population isolated from the maize rhizosphere at different plant growth stages. Applied and Environmental Microbiology, Washington, v.63, p. 4485-4493, 1997.
DI FIORI, S.; DEL GALLO, M. Endophytic bacteria: their possible role in the host plant. In: FENDRIK, I. et al. Azospirillum VI and Related Microorganisms. pp. 1 ed. Berlin: Springer-Verlag, 1995.
DONATO, S. L. R. et al. Comportamento fitotécnico da bananeira ‘Prata-Anã’ e de seus híbridos. Pesquisa Agropecúaria Brasileira, Brasília, v. 44, p. 1608-1615, 2009.
DUIJFF, B. J.; GIANINAZZI-PEARSONAND, V.; LEMANCEAU, P. Involvement of the outer membrane lipopolysaccharides in the endophytic colonization of tomato roots by biocontrol Pseudomonas fluorescens strain WCS417r. New Phytologist, Grã-Bretanha, v.135, p. 325-334,1997.
ELVIRA-RECUENCO, M.; VAN VUURDE, J. W. L. Natural incidence of endophytic bactéria in peacultivars under field conditions. Canadian Journal of Microbiology, Netherlands, v. 46, p. 1036-1041, 2000.
EMMERT, E. A. B.; AND HANDELSMAN, J. Biocontrol of plant disease: a (Gram) positive perspective. FEMS Microbiol Letters, Amsterdam, v. 171, p.1–9, 1999.
FAO. Food and Agricultural Organization. Faostat. Disponível em: http://www.faostat.fao.org/site/567/default.aspx. Acesso em: 03 Mar. 2010.
FERNADES, M. F.; FERNANDES, R. P. M.; HUNGRIA, M. Caracterização genética de rizóbios nativos dos tabuleiros costeiros eficientes em culturas do guandu e caupi. Pesquisa Agropécuaria Brasileira, Brasília, v. 38, n. 8, p. 911-920, 2003.

85
FRANCIS, I.; HOLSTERS, M.; VEREECKE, D. The Gram-positive side of plant–microbe interactions. Environmental Microbiology, Oxford, v.12, p. 1-12, 2010.
FERREIRA, A. et al. Diversity endophytic bactéria from Eucalyptus species seeds and colonization of seedlings by Pantoea agglomerans. FEMS Microbiol Letters, Ireland, v. 287, p. 8-14, 2008.
FISHER, P. J.; PETRINI, O.; LAPPIN SCOTT, H. M. The distribution of some fungal and bacterial endophytes in maize (Zea mays L.). New Phytologist, Oxford, v.122, p. 299-305, 1992.
FORCHETTI, G. et al . Endophytic bacteria in sunflower (Helianthus annuus L.): isolation, characterization, and production of jasmonates and abscisic acid in culture medium. Applied Microbiology and Biotechnology, Heidelberg, v. 76, p. 1145-1152, 2007.
FROMIN, N. et al. The genotypic diversity of Pseudomonas brassicacearum populations isolated from roots of Arabidopsis thaliana: influence of plant genotype. FEMS Microbial Ecology, Oslo, v. 37, p. 21-29, 2001.
GANEN, S.T. S. et al. Microbial Contamination in Explante of Banana Cultivars ‘Galil 18’ and ‘Tropical’. In: INTERNATIONAL SYMPOSIUM ON IN VITRO CULTURE AND HORTICULTURAL BREEDING, 4., 2009,Australia. Proceedings..., Australia: Acta Horticulturae, 2009, p. 341-344.
GERMAINE, K. J. et al. Bacterial endophyte-mediated naphthalene phytoprotection and Phytoremediation. FEMS Microbiology Lett, Ireland, v. 296, p. 226-234, 2009.
GERMIDA, J. J. et al. Diversity of root-associated bacteria associated with field-grown canola (Brassica napus L.) and wheat (Tritricum aestivum L.). FEMS Microbiology Ecology, Amsterdam, v. 26, p. 43-50, 1998.

86
GORIS, J. et al. Diversity of activated sludge bacteria receiving the 3-chloroaniline-degradative plasmid pC1gfp. FEMS Microbiology Ecology, Amsterdam, v. 46, p. 221-230, 2003.
GRANER, G. et al. A study on microbial diversity in different cultivars of Brassica napusin relation to its wilt pathogen, Verticillum longisporum. FEMS Microbiol Letters, Ireland, v. 224, p. 269–276, 2003.
GURTLER, J. B.; KORNACKI, J. L; BEUCHAT, L. R. Enterobacter sakazakii: a coliform of increased concern to infant health. International Journal of Food Microbiology, Amsterdam, v.104, p. 1-3, 2005.
HALLMANN, J.; BERG, G. Spectrum and population dynamics of bacterial root endophytes. Soil Biology, Berlim, v. 9, p.15-31, 2006.
HALLMANN, J. et al. Bacterial endophytes in agricultural crops. Canadian Journal of Microbiology, Ottawa, v. 43, p. 895-914, 1997.
HALLMANN, J. et al. Interactions between Meloidogyne incognita and endophytic bacteria in cotton and cucumber. Soil Biology and Biochemistry, Elmsford, v. 30, p. 925–937, 1998.HEUER, H. et al. Effects of T4 lysozyme release from transgenic potato roots on bacterial rhizosphere communities are negligible relative to natural factors. Applied and Environmental Microbiology, Baltimore, v. 68, p. 1325-1335, 2002.
HEYNDRICKX, M. et al. Applicability of combined amplified ribosomal DNA restriction analysis (ARDRA) patterns in bacterial phylogeny and taxonomy. Journal of Microbiological Methods, Milão, v. 26, p. 247-259, 1996.
HUNTER-CEVERA, J. C. The value of microbial diversity. Current Opinion in Microbiology, New York, v. 1, p. 278-285, 1998.
IKEDA, A. C. Caracterização morofofisiológica e genética de bactérias endofíticas isoladas de raízes de diferentes genótipos de milho (Zea mays

87
L.). 2010. 100 p. Dissertação (Mestrado em Ciências Biológicas) – Universidade Federal do Paraná, Curitiba, 2010.
IZQUIERDO, J. A.; NUSSLEIN, K. Distribution of extensive nifH gene diversity across physical soil microenvironments. Microbial Ecology, Oslo, v. 51, p. 441-452, 2006.
JACOBS, M. J.; BUGBEE, W. M.; GABRIELSON, D. A. Enumeration, location, and characterization of endophytic bacteria within sugar beet roots. Canadian Journal of Botany, Ottawa, v. 63, p. 1262-5, 1985.
JUNG, W. J. et al. Treatment of Paenibacillus illinoisensis suppresses the activities of antioxidative enzymes in pepper roots caused by Phytophthora capsici infection. World Journal of Microbiology and Biotechnology, Dordrecht, v. 22, p. 901-907, 2006.
KENNEDY, A. C. Bacterial diversity in agroecosystems. Agriculture Ecosystems and Environment, Amsterdam, v. 74, p. 65-76, 1999.
KLUEPFEL, D. A. The behavior and tracking of bacteria in the rhizosphere. Annual Review of Phytopathology, Palo Alto, v. 31, p.441–472, 1993.
KOBAYASHI, D; PALUMBO, J. Bacterial endophytes and their effects on plants and uses in agriculture. Microbial endophytes, New York, v. 2, p. 199, 2000.
KOEUTH, T.; VERSALOVIC, J.; LUPSKI, J. R. Differential subsequence conservation of interspersed repetitive Streptococcus pneumoniae BOX elements in diverse bacteria. Genome Research, Baltimore, v. 5, p. 408, 1995.
KOGEL, K. H.; FRANKEN, P.; HUCKELHOVEN, R. Endophyte or parasite what decides?. Current Opinion in Plant Biology, London, v. 9, p. 358–363, 2006.

88
KRISHNAMURTHY, K.; GNANAMANICKAM, S. S. Biological control of sheath blight of rice: induction of systemic resistance in rice by plant-associated Pseudomonas spp. Current science, Bangalore, v.72, p. 331-334, 1997.
KUKLINSKY-SOBRAL, J. A comunidade bacteriana endofítica e epifítica de (Glycine max) e estudo da interação endófitos-planta. 2003. 189 p. Doutorado (Tese em Genética e Melhoramento de plantas) – Universidade de São Paulo, Piracicaba, 2003.
KUKLINSKY-SOBRAL, J. et al. Isolation and characterization of soybean associated bacteria and their potential for plant growth promotion. Environmental Microbiology, Oxford, v. 6, p. 1244-1251, 2004.
KUSS, A.V. et al. Fixação de nitrogênio e produção de ácido indolacético in vitro por bactérias diazotróficas endofíticas. Pesquisa Agropecuária Brasileira, Brasília, v. 42, p. 1459-1465, 2007.
LACAVA, P.T. et al. Interaction between endophytic bacteria from citrus plants and the phytopathogenic bacteria Xylella fastidiosa, causal agent of citrus-variegated chlorosis. Letters in Applied Microbiology, Oxford, v. 39, p. 55-59, 2004.
LAL, S.; TABACCHIONI, S. Ecology and biotechnological potential of Paenibacillus polymyxa: a minireview. Indian Journal of Microbiology, Chandigarh, v. 49, p. 2-10, 2009.
LANE, D.L. et al. Rapid determination of 16S ribossomal RNA sequences for phylogenetic analyses. Proceedlings of National Academy os Sciences, Washington, v. 82, p. 6955- 6959, 1985.
LAZZARETTI, E. ; MELO, I. S. Influência de Bacillus subtilis na promoção de crescimento de plantas e nodulação de raízes de feijoeiro. Jaguariúna: EMBRAPA Meio Ambiente, 2005. 21 p. (Boletim Técnico 28)

89
LI, J. H. et al. Genetic diversity and potential for promotion of plant growth detected in nodule endophytic bacteria of soybean grown in Heilonjiang province of China. Soil Biology and Biochemestry, Oxford, v. 40, p. 238-246, 2008.
LI, Y.H. et al. Endophytic bacterial diversity in roots of Phragmites australis in constructed Beijing Cuihu Wetland (China). FEMS Microbiol Letters, Ireland, v. 309, p. 84-93, 2010.
LODEWYCKX, C. et al. Endophytic Bacteria and their potential applications. Critical Reviews in Plant Sciences, Boca Raton, v. 21, p. 583-606, 2002.
LOUWS, F. J. et al. Specific genomic fingerprints of phytopathogenic Xanthomonas and Pseudomonas pathovars and strains generated with repetitive sequences and PCR. Applied and Environmental Microbiology, Michigan, v. 60, p. 2286-2295, 1994.
LUCON, C. M. M. et al. Bioprospecção de isolados deTrichoderma spp. para o controle de Rhizoctonia solani na produção de mudas de pepino. Pesquisa Agropecuária Brasileira, Brasília, v. 44, p. 225-232, 2009.
LUPSKI, J. R. Molecular epidemiology and its clinical application. The Journal of the American Medical Association, Chicago, v. 270, p. 1323-1329, 1993.
MAGNANI, G. S. et al. Diversity of endophytic bacteria in Brazilian sugarcane. Genetics and Molecular Research. Ribeirão Preto, v. 9, p. 250-258, 2010.
MARCON, J. Isolamento e caracterização genética de actinomicetos endofíticos de Citrus spp. e interação com Xyllela fastidiosa. 2002.150 p. Dissertação (Mestrado em Biotecnologia) - Universidade de São Paulo, São Paulo, 2002.
MARIANO, R. L. R. et al. Isolamento de bactérias para testes de antagonismo. In: MARIANO, R.L.R. Manual de práticas em Fitobacteriologia. Recife: Rosa de Lima Ramos Mariano. 2000. p.115-119.

90
MASCARENHAS, G. C. C. Banana: Comercialização e Mercados. Informe Agropecuário, Belo Horizonte, v. 20, p. 97-108, 1999.
MEDRANO, E.G; BELL, A. A . Role of Pantoea agglomerans in opportunistic bacterial seed and boll rot of cotton (Gossypium hirsutum) grown in the field. Journal of Applied Microbiology, Oxford, v.102, p.134-143, 2007.
MEHTA, A; GONCALVES, E. R.; ROSATO, Y. B. Classificacao das bactérias fitopatogenicas utilizando técnicas baseadas em DNA. In: SERAFINI, L. A.; BARROS, N. M.; AZEVEDO, J. L. Biotecnologia: avanços na agricultura e na agroindústria. Caxias do Sul: EDUCS, 2002. p. 133-163.
MENDES, R.; AZEVEDO, J. L. Valor biotecnológico de fungos endofíticos isolados de plantas de interesse econômico. In: MAIA, L. C.; MALOSSO, E.; YANO-MELO A.M. Micologia: avanços no conhecimento.1ed. Recife: Ed. Universitária da UFPE, 2007. p.129-140.
MICHAUD, L. et al. Biodiversity of cultivable pyschorotrophic marine bacteria isolated from Terra Nova Bay (Ross Sea. Antarctica). FEMS Microbiology Letters, Ireland, v. 230, p. 63-71, 2004.
MICHE, L. et al. Up regulation of jasmonate-inducible defense proteins and differential colonization of roots of Oryza sativa cultivars with the endophyte Azoarcus sp. Molecular Plant-Microbe Interactions, Saint Paul, v.19, p. 502–511, 2006.
MISAGHI, I. J.; DONNDELINGER, C. R. Endophytic bacteria in symptom-free cotton plants. Phytopathology, Lancaster, v. 80, p. 808-811, 1990.
MISKO, A. L.; GERMIDA, J. J. Taxonomic and functional diversity of pseudomonads isolated from the roots of field-grown canola. FEMS Microbiology Ecology, Amsterdam, v. 42, p. 399-407, 2002.

91
MOCALI, S.et al. Fluctuation of bacteria isolated from elm tissues during different seasons and from ifferent plant organs. Research in Microbiology, Paris, v.154, p.105-114, 2003.
NAVES, R. L.; CAMPOS, V. P.; SOUZA, R. M. Filtrados de culturas bacterianas endofíticas na motilidade, mortalidade e eclosão de juvenis de segundo estádio de Meloidogyne javanica. Fitopatologia Brasileira, Brasília, v. 29, p. 384-388, 2004.
NAKATANI, A. K.; LOPES, R.; CAMARGO, L. E. A. Variabilidade genética de Xanthomonas Axonopodis Pv. Passiflorae. Summa Phytopathologica, Botucatu, v. 35, p. 116-120, 2009.
NEWMAN, L. A.; REYNOLDS, C. M. Bacteria and phytoremediation: new uses for endophytic bacteria in plants. Trends in Biotechnology, Amsterdam, v.23, p.6-8, 2005.
NIETSCHE, S. et al. Estabelecimento In Vitro de Explantes de Três Cultivares de Bananeira. Ciência Rural, Santa Maria, v. 36, n. 3, p. 989-991, 2006.
NOWAK, J. Benefits of in vitro “biotization” of plant tissue cultures with microbial inoculants. In Vitro Cellular and Developmental Biology Plant, Columbia, v. 34, p.122-130, 1998.
ODUTAYO, O. I. et al. Sources of microbial contamination in tissue culture laboratories in southwestern Nigeria. African Journal of Agricultural Research, Nigéria, v. 2, n. 3, p. 67-72, 2007.
OLIVEIRA, S. S. et al. A Cultura da banana: Aspectos técnicos, sócio-econômicos e agroindustriais. EMBRAPA, Brasília, DF, 1999. p. 85-105.
ONGENA, M.; JACQUES, P. Bacillus lipopeptides: versatile weapons for plant disease biocontrol. Trends in Microbiology, Cambridge, v. 16, p. 115-125, 2008.

92
PACE, N. R. et al.. The analysis of natural microbial populations by ribosomal RNA sequences. Advances in Microbial Ecology, Washington, v. 9, p. 1-55, 1986.
PARK, J. O. et al. Pathogenesis of Streptoverticillium albireticuli on Caenorhabditis elegans and its antagonism to soil-borne fungal pathogens. Letters in Applied Microbiology, Oxford, v. 35, p. 361-365, 2002.
PARK, M. et al. Isolation and characterization of diazotrophic growth promoting bacteria from rhizosphere of agricultural crops of Korea. Microbiological Research, Amsterdam, v. 160, p. 127-133, 2005.
PATTEN, C.L. ; GLICK, B.R.. Bacterial biosynthesis of indole-3-acetic acid. Can J Microbiol, Ottawa, v. 42, p. 207–220, 1996.
PERIN, L. Estudo da comunidade de bactérias diazotróficas do gênero Burkholderia em associação com cana-de-açúcar e descrição de Burkholderia silvatlantica. 2007. 103 p. Tese ( Doutorado em Ciências) - Universidade Federal Rural do Rio de Janeiro. Seropédica, 2007.
PILLAY, V. K.; NOWAK, J. Inoculum density, temperature and genotype effectson in vitro growth promotion and epiphytic and endophytic colonization of tomato (Lycopersicum esculentum L.) seedlings inoculated with a pseudomonad bacterium. Canadian Journal of Microbiology, Ottawa, v. 43, p. 354- 361, 1997.
QUADT-HALLMANN, A.; HALLMANN, J.; KLOEPPER, J.W. Bacterial endophytes in cotton: localization and interaction with other plant-associated bacteria. Canadian Journal of Microbiology, Otawa, v. 43, p. 254-259, 1997.
QURESHI, A.; VERMA, V.; KAPLEY, A.; PUROHIT, H. J. Degradation of 4-nitroaniline by Stenotrophomonas strain HPC 135. International Biodeterioration & Biodegradation, Oxford, v. 60, p. 215–218, 2007.

93
RADEMAKER, J. L. W.; DE BRUIJN, F. J. Caracterization and classification of microbes by rep-PCR genomic fingerprinting and computer assisted pattern analysis. In: ANOLLES, G. G.; GRESSOHOFF, P. M. DNA markers: protocols, applications e overviews. New York: J. Willey e sons, 1997. p. 151-171.
RADEMAKER, J. L. W.; LOUWS, F. J.; DE BRUIJN, F. J. Characterization of the diversity of ecologically important microbes by rep-PCR genomic fingerprinting. In: AKKERMANS, A. D. L.; VAN ELSAS J. D.; DE BRUIJN F. J. Molecular microbial ecology manual. 1 ed. Dordrecht: Kluwer, 1998. p. 1-26.
RADEMAKER, J. L. W. et al.. Comparison of AFLP and rep-PCR genomic fingerprints with DNA-DNA homology studies: Xanthomonas as a model system. International Journal of Systematic and Evolutionary Microbiology, Reading, v. 50, p. 665-677, 2000.
RANJARD, L.; POLY, F.; NAZARET, S. Monitoring complex bacterial communities using culture-independent molecular techniques: application to soil environment. Research in Microbiology, Paris, v.151, p.167-177, 2000.
RAUPACH, G. S.; KLOEPPER, J. W. Mixtures of plant growth-promoting rhizobacteria enhance biological control of multiple cucumber pathogens. Phytopathology, Auburn, v. 88, p. 1158-1164, 1998.
RAYMOND, J. et al.. The natural history of nitrogen fixation. Molecular Biology and Evolution, Oxford, v. 21, p. 541-554, 2004.
REIS JUNIOR, F. B. et al.. Identificação de isolados de Azospirillum amazonense associados a Brachiaria spp., em diferentes épocas e condições de cultivo e produção de fitormônio pela bactéria. Revista Brasileira de Ciência do Solo, Viçosa-MG, v. 28, p. 103-113, 2004.
REINHOLD-HUREK, B.; HUREK, T. Interactions of gramineous plants with Azoarcus spp. And other diazotrophs: identification, localization, and

94
perspectives to study their function. Critical Reviews in Plant Sciences, Boca Raton, v. 17, p. 29-54, 1998.
REITER, B. et al. Endophytic nifH gene diversity in African sweet potato. Canadian Journal of Microbiology, Ottawa, v. 9, p. 549-555, 2003.
RIBEIRO, C. M., CARDOSO, E. J. B. N.. Isolation, selection and characterization of root-associated growth promoting bacteria in Brazil Pine (Araucaria angustifolia). Microbiological Research, Piracicaba, v. 1, p. 1-10, 2011.
ROESCH, L. F. W. et al.. Diversidade de bactérias diazotróficas endofíticas associadas a plantas de milho. Revista Brasileira Ciência do Solo,Viçosa-MG, v. 31, p. 1367-1380, 2007.
RODAS, A. M.; FERRER, S. PARDO, I. 16S-ARDRA, a tool for identification of lactic acid bacteria isolated from grape must and wine. Systematic and Applied Microbiology, Stuttgart, v. 26, p. 412-422, 2003.
RODRÍGUEZ, H.; FRAGA, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotecnology Adances, Oxford, v. 17, p. 319-339, 1999.
ROMEIRO, R. S. Reação de hipersensibilidade induzida por bactérias fitopatogênicas. Revista Seiva, Viçosa-MG, v. 33, p.13-40, 1973.
ROMEIRO, R.S. Métodos em bacteriologia de plantas. 1. ed.. Viçosa: UFV, 2001. 279 p.
ROSADO, A. S.; DUARTE, G. F.; MENDONÇA-HAGLER, L. C. A moderna microbiologia do solo: Aplicação de técnicas de biologia molecular. In: SIQUEIRA, J. O. et al. Inter-relação fertilidade, biologia do solo e nutrição de plantas. Viçosa: SBCS, Lavras: UFLA/DCS, 1999. p. 429-448.

95
ROSAS-GARCIA, N. M. Biopesticide production from Bacillus thuringiensis: an environmentally friendly alternative. Recent Patents Biotechnol, Sharjah, v. 3, p. 28-36, 2009.
ROSENBLUETH, M.; MARTINEZ-ROMERO, E. Bacterial endophytes and their interactions with hosts. The American Phytopathological Society, Cuernavaca, v. 19, p. 827–837, 2006.
RÓZYCKI, H. et al. Diazotrophic bacteria in root-free soil and the root zone of pine (Pinus sylvestris L.) and oak (Quercus robur L.). Applied Soil Ecology, Amsterdam, v. 12, p. 239-25, 1999.
RYAN, R.P. et al.. Bacterial endophytes: recent developments and applications.FEMS Microbiol Letters, Ireland, v. 278, p. 1–9, 2008.
SANTOS, C. A. Ocorrência e diversidade de fatores de virulência em isolados brasileiros de Bacillus cereus provenientes de alimentos e do solo. 2009. 68 p. Dissertação (Mestrado em Genética e biologia molecular) - Universidade Estadual de Londrina, Londrina, 2009.
SCHULZ, B.; BOYLE, C. The endophytic continuum. Mycological Research, Cambridge, v.109, p. 661-686, 2005.
SEAPA. Secretária do Estado de Agricultura, Pecuária e Abastecimento. Disponível em: www.mg.gov.br. Acesso em: 20 de jan de 2010.
SEGHERS, D. et al.. Impact of agricultural practices on the Zea mays L. endophytic community. Applied and Environmental Microbiology, Oxford, v. 70, p. 1475-1482, 2004.
SHIOMI, F. H. et al. Bioprospecting endophytic bacteria for biological control of coffee leaf rust. Scientia Agrícola, Piracicaba, v. 63, n. 1, p. 32-39, 2006.

96
SILVA, S. de O. et al.. Cultivares. In: ALVES, E. J. A cultura da banana: aspectos técnicos, socioeconômicos e agroindustriais. 2. ed. Brasília: EMBRAPA-SPI, 1999. p.85-106.
SILVA, M. S.; BEDENDO, I. P.; CASAGRANDE, M. V. Caracterização molecular e patogênica de isolados de Xanthomonas albilineans (Ashby) Dowson, agente causal da escaldadura das folhas da cana-de-açúcar. Summa Phytopathologica, São Paulo, v. 33, n. 4, p. 341-347, 2007.
SNEATH, P. H.; SOKAL, R. R. Numerical taxonomy: The principles and practice of numerical classification. San Francisco: W.H. Freeman, 1973. 573 p.
SNUSTAD, D. P.; SIMMONS, M. J. Fundamentos de GENÉTICA. 4. ed. Rio de Janeiro: Guanabara Koogan, 2008. 903 p.
SOUTO, R. F. et al.. Sistema de Produção para a Cultura da Banana-Prata-Anã, no Norte de Minas. Belo Horizonte: EPAMIG, 1997. 32 p. (Boletim Técnico 48).
SPAEPEN, S.; VANDERLEYDEN, J.; REMANS, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiology Reviews, Amsterdam, v. 31, p. 425-448, 2007.
SPIERS, A. J.; BUCKLING, A.; RAINEY, P. The causes of pseudomonas diversity. Microbiology, Reading, v.146, p.2345-2350, 2000.
SPRENT, J. I.; FARIA, S. M.. Mechanismos of infection of plants by nitrogen fixing organisms. Plant and soil, Dordrecht, v. 110, p. 157-165, 1988.
STACKEBRANDT, E.; GOEBEL, B. M. Taxonomic note: a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. International Journal of Systematic Bacteriology, Washington, v. 44, p. 846-849, 1994.

97
STOLTZFUS, J. R. et al. Isolation of endophytic bacteria from rice and assessment of their potential for supplying rice with biologically fixed nitrogen. Plant and Soil, Netherlands, v. 194, p. 25-36, 1997.
STONE, J. K; BACON, C. W.; WHITE, J. F. An overview of endophytic microbes: endophytic microbes: endophytism defined. In: BACON, C.W.; WHITE, J.F. Microbial Endophytes. 1. ed. New York: Marcel Dekker. 2000. p. 3-29.
STROBEL, G. et al.. Natural products from endophitic microorganisms. Journal Natural Products, Washington, v. 67. p. 257- 268, 2004.
STURZ, A.V.; MATHESON, B. G. Populations of endophytic bacteria which influence host-resistance to Erwinia-unduced bacterial soft rot in potato tubers. Plant Soil, Dordrecht, v. 184,p. 265-27, 1996.
STURZ, A.V.; NOWAK, J. Endophytic communities of Rhizobacteria and the strategies required creating yield-enhancing associations with crops. Applied Soil Ecology, Amsterdam, v. 15, p. 183-190, 2000.
TAMURA, K. et al. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Molecular Biology and Evolution, Oxford, v. 10, p. 2731-2739, 2011.
TEIXEIRA, M. A. et al. Microrganismos endofíticos de mandioca de áreas comerciais e etnovariedades em três estados brasileiros. Pesquisa Agropecária Brasileira, Brasília, v. 42, n. 1, p. 43-49, 2007.
THOMAS, P. et al. Identification of culturable and originally non-culturable endophytic bacteria isolated from shoot tip cultures of banana cv. Grand Naine. Plant Cell, Tissue and Organ Culture, Rockville, v. 93, p. 55-63, 2008
TORRES, A. R. Diversidade genética de enterobactérias endofíticas de diferentes hospedeiros e colonização de Catharantus roseus por endófitos

98
expressando o gene gfp. 2005. 137 p. Tese (doutorado em Genética e melhoramento de plantas) - Universidade de São Paulo, Piracicaba. 2005.
TORSVIK, V.; OVREAS, L.; THINGSTAD, T. F. Prokaryotic diversity- magnitude, dynamics, and controlling factors. Science, Norway, v. 10, n. 296, p. 1064-1066, 2002.
TSAVKELOVA, E. A. et al. Microbial producers of plant growthstimulators and their practical use: a review. Applied Biochemistry and Microbiology, New York, v. 42, p. 117-126, 2006.
VALENCIA-CANTERO, E. et al.. Role of dissimilatory fermentativeiron-reducing bacteria in Fe uptake by common bean (Phaseolus vulgarisL.) plants grown in alkaline soil. Plant Soil, Netherlands, v. 291, p. 263-273, 2007.
VAN BERKUM, P. Short sequence repeats in microbial pathogenesis and evolution. Cellular and Molecular Life Sciences, Basel, v. 56, p. 729-734, 1999.
VERMA, S. C.; LADHA, J. K.; TRIPATHI, K. Evaluation of plant growth promoting and colonization ability of endophytic diazotrophic from deep water rice, Journal of Biotechnology, Amsterdam, v. 91, p. 127-141, 2001.
VERSALOVC, J.; KOEUTH, T.; LUPSKI, J. R. Distribuion of repetitive DNA sequencies in eubacteria and application to fingerprinting of bacterial genomes. Nucleic Acids Reserarch, Houston, v. 19, p. 6823-6831, 1991.
VERSALOVIC, J.; de BRUIJN, F. J.; LUPSKI, J. R. Repetitive sequence-based PCR (rep. PCR) DNA fingerprinting of Bacterial Genomes. In: de BRUIJN, F. J.; LUPSKI, J. R.; WEINSTOCK, G. M. (Ed.) Bacterial Genomes: Physical Structure and Analysis. New York: Chapman and Hall, 1998. P. 437-454.
VIDEIRA, S. S. et al. Ocorrence and diversity of nitrogen-fixing Sphingomonas bacteria associated with rice plants grown in Brazil. FEMS Microbiol Letters, Ireland, v. 293, p. 11-19, 2009.

99
VIEIRA, F. C. S; NAHAS, E. Qauntificação de bactérias totais e esporuladas no solo. Scientia Agricola, Piracicaba, v. 57, p. 539- 545, 2000.
WEBER, O. B. Ocorrência e caracterização de bactérias diazotróficas em bananeiras (Musa spp.) e abacaxizeiros (Ananas comosus (L) Merril) e seus efeitos no crescimento de mudas micropropagadas. 1998. 192 p. Tese (Doutorado em Microbiologia do Solo) – Universidade Federal Rural do Rio de Janeiro- UFRRJ, Seropédica, RJ, 1998.
WEBER, O. B.; FREIRE, F. das C. O. Contribuição de bactérias diazotróficas na cultura da bananeira: perspectivas de utilização na produção integrada. Jaguariúna: Embrapa Meio Ambiente, 2003. 29 p. (Boletim de Pesquisa e Desenvolvimento).
YOUNG, J. P. W. Phylogeny and taxonomy of rhizobia. Plant and Soil, Netherlands, v. 186, p. 45-52, 1996.
ZINNIEL, D. K. et al. Isolation and characterization of endophytic colonizing bacteria from agronomic crops and prairie plants. Applied and Environmental Microbiology, Nebraska, v. 69, p. 2198-2208, 2002.

100
ANEXOS
Questionário para produtores de banana ‘Prata-Anã’
Nome do Responsável Técnico: _______________________________ Fazenda:__________________________________ Telefone_________________ 01- Tempo de atividade agrícola: ________anos (média) 02- Cultura anteriormente plantada na área___________________________ 03- Número de hectares que explora:________hectares (média) 04- Idade do bananal________ anos (média) 05- Tem alguma formação profissional na área agrícola? ( ) Sim ( ) Não 06- Costuma mandar fazer análise de solo? ( ) Sim ( ) Não
6.1 - Se respondeu sim, com que regularidade? ____________ 6.2 - Se respondeu não, porquê? (assinale as que acha verdadeiras ( )Não sabe para que servem ( ) Sabe para que servem mas não acha importante
Outra, qual?____________ 07- Faz incorporação de matéria orgânica? ( )Sim ( ) não Qual frequência e dose aplicada/cova?____________________ Qual a fonte?______ 08- Costuma mandar fazer análise foliar? ( ) Sim ( ) não Se respondeu sim, com que regularidade? ____________________ 09- Qual adubo utiliza?____________________ 10- Quantidade de adubo que utiliza? kg por hectare do adubo _________ 11 - Como escolhe os adubos que usa? ( ) Conselho do vendedor ( ) Conselho de vizinhos ou amigos ( ) Conselho de técnicos ( ) Por meio da consulta de publicações sobre a matéria ( ) Iniciativa própria ( ) Outra forma .Qual? ______________ 12- Como irriga a banana?( ) Pivot ( ) Gotejador ( ) Microaspersão ( )Outro sistema Qual_______________ 13- Faz uso de fertilizante? ( ) sim ( ) não Se respondeu sim, qual tipo?_________________ Com que regularidade?___________ 14- Faz uso de fertilizante foliares? ( ) sim ( ) não Se respondeu sim, qual tipo?_________________ Com que regularidade?_____ 15- Qual (s) principais problemas fitossanitários: Mal do Panamá ( ) Sim ( ) Não Se sim, qual a porcentagem de área afetada?

101
Quando surgiu o primeiro foco?______ Antes do cultivo havia a doença? ________ ( ) Sigatoka amarela Qual índice é usado para começar com o controle?________ ( ) Nematoides ( ) Moleque de Bananeira 16-Qual(s) produto é usado para manejo fitossanitário? ( )Fungicida Sistêmicos ( ) Fungicida Protetores ( ) Herbicida pré-emergentes ( ) Herbicida pós emergentes ( ) Nematicidas organofosforados ( ) Nematicidas carbonatos ( ) Inseticidas ( ) Outro. Qual__________ 17- Qual é a produção por hectare?__________ População de plantas/ha__________ 18- Qual a qualidade da banana? ( ) 1° ( ) 2° 19- Faz corte de banana, quantas vezes por mês? ( )1 ( )2 ( )3 ( )4 ( )5
Mais de 5( ) .Quanto?___ 20- A produção é direcionada para qual mercado? ( ) Internacional ( ) Regional
( ) Local ( )Outro .Qual____________