Cyntia Martins Lima Integrões em Bactérias de Gram ...
77
Universidade de Aveiro Ano 2011 Departamento de Biologia Cyntia Martins Lima Integrões em Bactérias de Gram-negativo em Ambiente Hospitalar
Transcript of Cyntia Martins Lima Integrões em Bactérias de Gram ...

Universidade de Aveiro
Ano 2011
Departamento de Biologia
Cyntia Martins Lima
Integrões em Bactérias de Gram-negativo em Ambiente
Hospitalar

2
Universidade de Aveiro
Ano 2011
Departamento de Biologia
Cyntia Martins Lima
Integrões em Bactérias de Gram-negativo em Ambiente
Hospitalar
Dissertação apresentada à Universidade de Aveiro para cumprimento
dos requisitos necessários à obtenção do grau de Mestre em
Microbiologia, realizada sob a orientação científica da Professora
Doutora Sónia Alexandra Leite Velho Mendo, do Departamento de
Biologia da Universidade de Aveiro.

3
O júri
Presidente Prof.ª Doutora Adelaide Almeida
Professora Auxiliar do Departamento de Biologia da Universidade de
Aveiro
Doutora Anabela Oliveira Pereira
Investigadora em Pós-Doutoramento do CESAM
Vogais
Prof.ª Doutora Sónia Alexandra Leite Velho Mendo.
Professora Auxiliar do Departamento de Biologia da Universidade de
Aveiro
Dr. Elmano Ramalheira
Médico Patologista Clínico do Hospital Infante D. Pedro de Aveiro

4

5
Agradecimentos
Agradeço primeiramente a Deus que me permitiu estar aqui diante de tantas
dificuldades.
A Prof. Sónia Mendo, que me recebeu no seu laboratório e com muito
carinho, e deu-me força quando não as tinham.
A todos do laboratório LBM, que me acolheram e com muita paciência me
ensinaram, e quando já não tinha mais vontade de continuar essa difícil
caminhada, eles me ensinaram a ser persistente.
A minha família que mesmo longe esteve presente nas horas mais difíceis
da minha vida, apoiando-me e incentivando-me. Meus pais sempre estarão
comigo.
Obrigada pela força a todos!

6
Palavras-chave
Integrão classe 1, disseminação de genes de resistência, bactérias de Gram-negativo
multirresistentes
Resumo
O aparecimento de surtos bacterianos tem sido um dos maiores problemas nas
instituições hospitalares. O uso abusivo e indiscriminado de antibióticos tem
conduzido à emergência de bactérias multiresistentes. Estas bactérias possuem
diversos elementos genéticos móveis (plasmídeos, transposões, sequências de
inserção e integrões) que contribuem para a disseminação de genes de resistência
aos antibióticos entre diferentes espécies, por meio de um dos três mecanismos de
transferência de genes de resistência entre células bacterianas: transformação,
conjugação e transdução.
A disseminação de integrões em bactérias de Gram negativo em meio hospitalar
constituiu o objectivo do presente trabalho.
As amostras foram recolhidas nas superfícies do Serviço de Medicina II (2008) e
Serviço de Medicina Intensiva (2010), no Hospital Infante D. Pedro, Aveiro. Após
o isolamento das bactérias em meio selectivo para bactérias de Gram-negativo
(MacKonkey agar), todos os isolados foram analisados por BOX-PCR para
avaliação da clonalidade,
Foi pesquisada a presença de integrões de classe 1 e nos isolados positivos, foi
amplificada, caracterizada e sequenciada a respectiva região variável.
Os resultados mostraram a presença de integrões de classe 1 em diferentes espécies,
tais como Klebsiella oxytoca, Escherichia coli, Klebsiella pneumoniae,
Enterobacter cloacae e Stenotrophomonas maltophilia tendo sido encontrado o
mesmo tipo de integrãoem diferentes espécies.
Assim, foi possível concluir que os integrões de classe 1 podem contribuir para a
disseminação de genes de resistência, podendo por isso contribuir para a
emergência de bactérias multirresistentes, sendo este um problema de preocupação
a nível Mundial.

7
Keywords
Abstract
Class 1 integron, dissemination of resistance genes, Gram-negative bacteria
multiresistant
The emergence of bacterial outbreaks has been the main problem in several
hospitals. The excessive and indiscriminate use of antibiotics has led to the
emergence of multiresistant bacteria, which possess mobile genetic elements
(plasmids, transposons, insertion sequences and integrons) that contribute to the
dissemination of antibiotic resistance genes between different species. There are
three main mechanisms that allow transfer of resistance genes between bacterial
cells: transformation, conjugation and transduction.
The dissemination of integrons in Gram negative bacteria within the hospital
environment contribute to the dissemination of antibiotic resistance genes, and
constituted the main aim of the present work.
The samples were collected from the inanimate surfaces of the Serviço de Medicina
II 2008and Serviço de Medicina Intensiva 2010 of Hospital Infante D. Pedro,
Aveiro. After the bacterial isolation in selective media for Gram-negative bacteria
(MacKonkey agar), all isolates were selected, based on their clonality, using the
molecular technique BOX-PCR.
The presence of class 1 integrons was evaluated and in the positive isolates the
variable region was amplified sequenced and characterized.
Results showed the presence of class 1 integrons in different species, such as
Klebsiella oxytoca, Escherichia coli, Klebsiella pneumoniae, Enterobacter cloacae
and Stenotrophomonas maltophilia, and the same array was present in different
bacterial species.
It can be concluded that class 1 integrons can contribute to the dissemination of
antibiotic resistance genes, and can therefore, contribute to the emergence of multi-
resistance strains.

8
Índice
Introdução ............................................................................................................................................................. 15
1. Infecções em Ambiente Hospitalar ................................................................................................................ 16
1.1 Klebsiella ........................................................................................................................................................ 16
1.2 Escherichia coli............................................................................................................................................... 17
1.3 Pseudomonas aeruginosa................................................................................................................................ 17
1.4 Enterobacter cloacae ...................................................................................................................................... 18
1.5 Acinetobacter baumannii ................................................................................................................................ 19
2 –Antibióticos ..................................................................................................................................................... 20
3- Mecanismo de Acção do Antibiótico ............................................................................................................. 20
3.1 - Inibição da Síntese da Parede Celular Bacteriana ......................................................................................... 22
3.2 - Inibição da Síntese Proteica .......................................................................................................................... 22
3.3 - Inibição da Síntese de Ácidos Nucleicos ...................................................................................................... 23
3.4 - Ligação à Membrana Citoplasmática ............................................................................................................ 23
4. - Mecanismos de Resistência dos Antibióticos .............................................................................................. 24
4.1 - Resistência intrínseca .................................................................................................................................... 26
4.2 - Resistência Adquirida ................................................................................................................................... 26
4.3 - Inactivação e Degradação enzimática do Antibiótico ................................................................................... 27
4.4 -Alterações na estrutura do local alvo do antibiótico ...................................................................................... 27
4.5 - Efluxo do Antibiótico ................................................................................................................................... 28
4.6 – Alteração da Permeabilidade Membranar .................................................................................................... 29
5 - Disseminação dos genes de resistência aos antibióticos ............................................................................. 29
5.1 - Mutação ........................................................................................................................................................ 30
5.2 - Transformação .............................................................................................................................................. 31
5.3 - Conjugação ................................................................................................................................................... 31
5.4 - Transdução .................................................................................................................................................... 32
5.5 - Elementos Genéticos Móveis ........................................................................................................................ 32
5.5.1 - Plasmídeo ................................................................................................................................................... 33
5.5.2 - Transposões ............................................................................................................................................... 35
5.5.3 - Integrões .................................................................................................................................................... 36
5.5.3.1 - Plataforma dos integrões ......................................................................................................................... 37
5.5.3.2 - Genes Cassettes ...................................................................................................................................... 39
5.5.3.3 - Segmento Conservado 3’ ........................................................................................................................ 38
5.5.3.3 - Segmento Conservado 5’ ........................................................................................................................ 37
5.5.3.4 - Região Variável ...................................................................................................................................... 38
5.5.3.6 - Classificação dos Integrões ..................................................................................................................... 40
5.5.3.6.1 - Integrões de Classe 1 ........................................................................................................................... 41
5.5.3.6.2 - Outras Classes dos Integrões ............................................................................................................... 41
6. Objectivo .......................................................................................................................................................... 43
7 - Material e Métodos ........................................................................................................................................ 45
7.1 - Identificação dos locais de recolhas dos Isolados ......................................................................................... 45
7.2– Recolha, Selecção e Conservação dos Isolados ............................................................................................ 45
7.3– Extracção de DNA ........................................................................................................................................ 46

9
7.4 - Caracterização Fenotípica ............................................................................................................................. 46
7.4.1- Avaliação da Clonalidade dos Isolados ...................................................................................................... 46
7.4.2 – Amplificação dos Ácidos Nucleicos por PCR ( Polymerase Chain Reaction) .......................................... 47
7.4.3 – Detecção de Genes que codificam enzimas β-Lactamases ........................................................................ 48
7.4.4 – Visualização dos Produtos Amplificados .................................................................................................. 50
7.4.5 – Digestão da Região Variável ..................................................................................................................... 50
7.4.6 – Purificação dos Produtos ........................................................................................................................... 51
7.4.7 – Determinação da Sequência Nucleotídica ................................................................................................. 51
8 - Resultados e Discussão .................................................................................................................................. 53
8.1 - Serviço de Medicina II (SMII) ...................................................................................................................... 53
8.1.1 – Tipagem Molecular ................................................................................................................................... 53
8.1.2 Caracterização da Região Variável .............................................................................................................. 53
8.1.3 Presença de enzimas β-lactamases no SMII ................................................................................................. 56
8.2 - Serviço de Medicina Intensiva (SMI) ........................................................................................................... 59
8.2.1 Tipagem Molecular ...................................................................................................................................... 59
8.2.2 Caracterização da Região Variável .............................................................................................................. 61
9. Conclusão ......................................................................................................................................................... 65
10. Referências Bibliográficas ............................................................................................................................ 66

10
Índice de Figuras
Figura 1 -Mecanismos de acção exercidos pelas diferentes classes de antibióticos ........................................ 16
Figura 2- Estrutura geral dos antibióticos β-Lactâmicos ............................................................................. 17
Figura 3 -Mecanismos de Resistência Bacteriana ....................................................................................... 18
Figura 4 -Mecanismos genéticos de transferência de DNA entre bactérias ................................................... 24
Figura 5 -Esquema simplificado de um plasmídeo ..................................................................................... 31
Figura 6 -Representa o esquema de um transposão simples, associado às respectivas IS ................................ 32
Figura 7 -Representação esquemática de um Transposão Composto ............................................................ 33
Figura 8- A figura esquematiza as três regiões que constituem um integrão.................................................. 34
Figura 9- A figura representa o mecanismo de inserção dos genes cassette na região variável de um integrão . 37
Figura 10 -Prevalência de isolados bacterianos recolhidos nas diferentes superfícies do Serviço de medicina
Intensiva .................................................................................................................................................. 56
Figura 11-Percentagem de enzimas β-lactamases presentes nos isolados que apresentavam integrão classe1 .. 63

11
Listas de Abreviaturas
aac – Aminoglicosídeo acetiltransferase
aad – Aminoglicosídeo adeniltransferase
arr – Rifampicina ADP ribosil transferase
attI – Local de recombinação 5’ CS
attC – Local de recombinação 3’CS
CS – Conserved Sequence
DNA – Ácido Desoxirribonucleico
ESBL – Beta-Lactamase de Espectro Alargado
IntI – Enzima Integrase
IR – Sequência de Inserção Invertida
IS – Sequência de Inserção
MBL – Metalo –Beta-Lactamase
MDR – Drogas Multirresistente
PBP – Penicillin Binding Proteins
PCR – Reacção em Cadeia de Polimerase
qac – Gene que codifica para a resistência a Compostos de Amónio Quaternário
Qnr – Gene de Resistência as Quinolonas
sul – Gene que codifica para a resistência de sulfonamidas
Tn – Transposão
tnpA – Transposase
VR- Região Variável

12
INTRODUÇÃO

13
Introdução
O crescente problema da resistência bacteriana está difundido em todo o mundo, e o
aparecimento de surtos hospitalares como consequência do uso indevido de antibióticos
constitui uma das maiores preocupações para a saúde humana. Assim, esta problemática tem
um grande impacto na morbilidade, na mortalidade e conduz a elevados custos nos cuidados
para a saúde pública. Contudo, o uso inadequado das diferentes classes de antibióticos,
conduziu à emergência de bactérias multirresistentes, as quais apresentam um perfil de
resistência a três ou mais classes de antibióticos, sendo por isso as opções terapêuticas cada
vez mais limitadas. (Sundsfjord 2004; Sousa 2006; Ferreira 2010; Tenover 2006). Além da
aplicação dos antibióticos na medicina humana, estes compostos são igualmente utilizados na
agricultura e na agropecuária, e muitas vezes de forma incorrecta. A elevada pressão selectiva
constitui um factor importante na emergência e disseminação de estirpes bacterianas
multirresistentes, uma vez que as bactérias têm a capacidade de desenvolver ou adquirir
mecanismos de resistência, em resposta aos mecanismos de acção dos antibióticos de forma a
sobreviverem na sua presença (Martinez 2009; Senka 2008; Sousa 2006).
Sendo que independentemente das condições do meio, podemos encontrar diferentes espécies
de bactérias de Gram-negativo, principalmente no ambiente hospitalar, as quais são
responsáveis por uma grande parte das infecções bacterianas (Senka 2008).
1. Infecções no Ambiente Hospitalar
O ambiente hospitalar está colonizado por uma vasta diversidade de espécies bacterianas,
nomeadamente bactérias de Gram-negativo, as quais têm a capacidade de desenvolver ou
adquirir adaptações que lhes permitem sobreviver nas superfícies hospitalares. Esta
permanência no ambiente hospitalar constitui um grave problema, uma vez que estas estirpes
podem facilmente infectar os pacientes imunodeprimidos. Devido à pressão selectiva
antibiótica existente no ambiente hospitalar, cada vez é mais frequente a emergência de
bactérias de Gram-negativo multirresistentes associadas a infecções nosocomiais. Em muitos
casos, esta evolução está associada a bactérias comensais e oportunistas, tais como
Escherichia coli e Klebsiella pneumoniae, que rapidamente apresentam um reduzido perfil de
susceptibilidade às diferentes classes de antibióticos (Senka 2008).

14
As bactérias de Gram-negativo, geralmente apresentam um reduzido perfil de susceptibilidade
aos antibióticos devido em parte à composição da parede celular, a qual é constituída por uma
membrana interna e uma membrana externa, reduzindo assim a permeabilidade membranar
aos antibióticos (Mahamoud 2007). Também a presença de enzimas β-lactamases corresponde
a um dos mais importante mecanismo de resistência aos antibióticos, encontrados tanto em
espécies da família Enterobacteriaceae, como em espécies de Acinetobacter spp. e
Pseudomonas spp., as quais podem facilmente causar infecções nosocomiais (Peng
2007;Coque 2008).
A elevada prevalência associada a infecções e a aquisição de mecanismos de resistência a
novos antibióticos, tem levado os microrganismos a alcançar notoriedade como patogénicos
causadores de surtos de infecção hospitalar (Ardanuy 1998).
1.1 Klebsiella pneumoniae
O género Klebsiella pertence à família Enterobacteriaceae, e é composto por cinco espécies
bacterianas, sendo a espécie Klebsiella pneumoniae a mais importante (Martínez 2004).
Sendo esta uma espécie anaeróbia facultativa, pode ser encontrada em quase todos os
ambientes naturais, tais como o solo, a água e as plantas, mas pode também ser isolada
principalmente a partir do trato respiratório e do trato urinário. Actualmente, esta espécie
comensal e oportunista está na origem de uma diversidade de infecções nosocomiais, sendo
responsável por uma elevada taxa de mortalidade, e por isso é considerada uma espécie
patogénica (Baron 1996).
Esta espécie apresenta adaptações que lhe permitem sobreviver por muito tempo na pele e em
ambientes secos, tais como nas superfícies hospitalares, e têm a capacidade de conjugarem
com facilidade, o que lhes vai permitir adquirir genes que codificam resistência a diferentes
classes de antibióticos. Apesar da resistência adquirida, esta espécie é intrinsecamente
resistente a ampicilina devido à presença da enzima β-lactamase SHV-1, mas pode também
produzir enzimas plasmídicas como ampC, metalo-β-lactamases (MBL) e carbapenases
(KPC) (Pfaller 1998).
A espécie K. pneumoniae é a que produz maior variedade destas enzimas, o que poderia ser
explicado pelo facto destes microrganismos serem bons vectores / disseminadores de

15
plasmídeos, ou adquirirem genes que codificam enzimas β-lactamases de espectro estendido
(ESBLs) mais rapidamente que outras espécies pertencentes à família Enterobacteriacea
(Livermore 1995).
K. pneumoniae é uma importante causa de infecção hospitalar principalmente associada à
pneumonia decorrente da colonização de pacientes hospitalizados, sendo proporcionais aos
dias de internação, podendo ser isoladas nasofaríngea, mãos e fezes com uma frequência de
20%, 42% e 77% respectivamente. A alta taxa de colonização nosocomial pela Klebsiella
pode estar associada com o uso do antibiótico e prestação dos cuidados hospitalares. Ou seja,
pacientes que tiveram infecções por K.pneumoniae produtoras de ESBL, tiveram infecções
precedidas pela colonização do tracto gastrointestinal. A capacidade desse microrganismo em
se disseminar rapidamente, muitas vezes, pode levar aos surtos nosocomiais (Podschun 1998).
1.2 Escherichia coli
A espécie E. coli pertence ao género Escherichia, que por sua vez pertence à família
Enterobacteriaceae. Esta espécie é caracterizada como anaeróbica facultativa, oxidase
negativa e pode ser isolada predominantemente do sistema gastrointestinal do Homem e
diversos animais (Sáenz 2004).
A bactéria E. coli, juntamente com outras bactérias comensais, desempenha um papel
protector do intestino contra infecções causadas, por exemplo, por bactérias patogénicas como
a Salmonella. No entanto, a bactéria E. coli, apesar de ser considerada uma espécie comensal
e oportunista, pode tornar-se patogénica devido à aquisição de genes de resistência aos
antibióticos e ao desenvolvimento de uma série de factores de virulência (Yingst 2006).
Sendo assim, esta espécie é responsável por graves infecções intestinais, infecções urinárias,
colite hemolítica, meningite neonatal e septicémia (Obi 2004). Contudo, pode desencadear
infecções entéricas, dependendo do tipo de factor de virulência possuem: enterotoxigénicas,
enteroinvasivas, enterohemorragicas, enteropatogenicas e enteroagregativas (Guentzel 1996;
Obi 2004).
1.3 Pseudomonas aeruginosa
Esta espécie oportunista é uma das mais patogénicas e virulentas, e apresenta algumas
propriedades que a tornam um problema para as instituições hospitalares, nomeadamente

16
inúmeros factores de virulência, e uma grande capacidade de adquirirem genes de resistência
aos antibióticos (Siegel 2008, Pool 2001).
Infecções causadas por esta espécie (pneumonias, infecções do trato urinário, bacteremias e
infecções nos rins) têm sido particularmente desafiantes, uma vez que o seu perfil de
resistência tem-se tornado significativo ao longo do tempo, em consequência da pressão
selectiva existente no ambiente hospitalar (Nouér 2005).
Sendo assim, a maioria das infecções causada por P. aeruginosa ocorre no ambiente
hospitalar em pacientes submetidos a procedimentos invasivos, como o uso de sondas,
cateteres vasculares, tubo endotraqueal e cirurgias, sendo mais frequente em pacientes
internados no serviço de medicina intensiva (Mulvey 2009).
A capacidade de P. aeruginosa sobreviver e colonizar superfícies hospitalares deve-se em
grande parte à formação de biofilmes, que impedem a penetração dos antibióticos (Siegel
2008). Também o desenvolvimento e aquisição de mecanismos de resistência contribuem para
o perfil de multirresistência apresentado por esta espécie, e daí a elevada patogenicidade.
Perante estes factores, as opções de tratamento de infecções causadas por P. aeruginosa são
cada vez mais escassas e limitadas, e daí a elevada percentagem de mortalidade (Nickel
1989).
1.4 Enterobacter cloacae
A espécie E. cloacae pertence à família Enterobacteriaceae e é considerada uma
espécie oportunista, uma vez que faz parte da flora normal do trato gastrointestinal dos
humanos, mas também pode ser encontrada no ambiente (Keller 1998).
Esta espécie é intrinsecamente resistente à ampicilina e às cefalosporinas de estreito
espectro, devido à produção de cefalosporinases cromossómicas e de algumas ESBLs
codificadas por plasmídeos. Também apresenta uma elevada frequência de mutações
que lhes conferem resistência às cefalosporinas de amplo espectro e ao aztreonam, mas
o seu perfil de resistência pode também estar associado a proteínas da membrana
externa, que expulsam o antibiótico (Cornaglia 1995). No caso de isolados clínicos de
E. cloacae, a resistência ao imipenemo pode ser devida a alteração nas porinas em

17
combinação com a hiperprodução de cefalosporinases cromossomais, ou pode estar
associada directamente à produção de carbapenemases (Jeong 2003).
Sendo esta espécie comensal e oportunista pode tornar-se patogénica e infectar
pacientes imunodeprimidos. Alguns autores sugerem que infecções causadas por E.
clocae, normalmente estão associadas ao trato digestivo (Jeong 2003; Lambert-
Zechovsky 1992; Sekar 2008).
1.5 Acinetobacter baumannii
O genéro Acinetobacter é caracterizado como bacilo de Gram-negativo, ubiquitário,
aeróbio estrito, não fermentador, imóvel, catalase positiva e oxidase negativa. Até à
data, dentro deste género estão descritas trinta e uma espécies, distribuídas por diversos
habitats naturais (solo, água, vegetais, animais e humanos), mas algumas fazem parte da
flora comensal da pele e mucosas humanas (Ash 2002).
A bactéria A. baumannii é oportunista e apresenta vários factores de virulência que
permitem a sua sobrevivência em superfícies hospitalares, e a colonização de
equipamentos hospitalares, principalmente ventiladores mecânicos, máquinas de
diálises, fontes de água. Além das suas adaptações, esta espécie consegue sobreviver em
condições ambientais adversas como soluções de desinfectantes e variações de
temperatura, o que contribui para o seu potencial de patogenicidade (Bernards 2004).
Um outro mecanismo de resistência presente nestas espécies é a produção de ESBL e de
carbapenemases, que lhes conferem um alto índice de resistência aos oximino-β-
lactâmicos. Os produtores destas enzimas são cada vez mais um grave problema para as
instituições hospitalares devido à disseminação horizontal destes genes (Livermore
2007).
Sendo assim, actualmente esta espécie está frequentemente associada a surtos
hospitalares, causando graves infecções tais como septicemia, pneumonias e infecções
urinárias (Poirel 2006).

18
2 –Antibióticos
Antibióticos são compostos que têm a capacidade de interagir com as bactérias. Estes, podem
ser classificados como bactericidas, quando eliminam totalmente o microrganismo, ou como
bacteriostáticos, se apenas inibem o seu metabolismo.
O primeiro antibiótico surgiu de uma forma imprevisível e surpreendente. Após várias
experiências realizadas por Alexander Fleming, em 1928 surge a penicilina. A partir de 1940
iniciou-se o processo de comercialização dos antibióticos e sua administração terapêutica. No
decorrer do tempo, esses compostos foram importantes no controlo de surtos bacterianos, e
mostraram-se eficazes no tratamento de infecções causadas por bactérias patogénicas
(Livermore 2003; Aminov 2009).
A introdução dos antibióticos é provavelmente uma das formas mais bem sucedidas no
tratamento de infecções na história da medicina. No entanto, esta descoberta pode estar cada
vez mais ameaçada pela emergência de bactérias resistentes às diferentes classes de
antibióticos, e pela diminuição do desenvolvimento de novos compostos (Aminov 2009).
Tendo em conta que os microrganismos possuem uma enorme diversidade metabólica, as
diferentes classes de antibióticos apresentam locais de acção específicos dentro da célula
bacteriana, de modo a combatê-la. (Hancock 2007;Aminov 2009 Sousa 2006; Ferreira 2010).
Esta eficácia também está relacionada com a concentração administrada e com a estabilidade
da estrutura química, de forma a não ser facilmente alterada ou degradada (Hancock 2007;
Senka 2008).
3 - Mecanismo de Acção dos Antibióticos
O recurso aos antibióticos na medicina moderna é inevitável e essencial no combate a
infecções bacterianas. Contudo, e apesar da sua eficácia, cada vez mais surgem bactérias
resistentes a diferentes classes de antibióticos. A resposta a essa grande problemática foi e
continuará a ser a introdução de novos antibióticos, com diferentes locais de acção dentro da
célula bacteriana, de modo a combater as bactérias resistentes (Coates 2007).
Os antibióticos não exercem uma actividade biológica no organismo humano, uma vez que
apenas actuam contra células procarióticas, mas impedem o crescimento ou provocam a morte

19
dos microrganismos infectantes. As diferentes classes de antibióticos são agrupadas de acordo
com seu modo de acção na célula bacteriana, como esquematiza a Figura 1 e resume a Tabela
1. Existem diferentes mecanismos de acção, entre os quais se destacam: a inibição da síntese
da parede celular, inibição da síntese proteica, inibição da síntese de ácidos nucléicos e
interferência ao nível do metabolismo celular bacteriano (Ferreira 2010; Wright 2005; Senka
2008).
Figura 1. Mecanismos de acção exercidos pelas diferentes classes de antibióticos sobre a célula bacteriana
(Adaptada Saenz, 2004).
3.1 - Inibição da Síntese da Parede Celular Bacteriana
A estrutura rígida da parede celular permite que as bactérias mantenham uma pressão
osmótica muito elevada, devido à ligação cruzada dos seus precursores, que ocorre durante a
síntese da parede celular bacteriana e é catalisada por enzimas específicas. As classes de
antibióticos que actuam ao nível da síntese do peptidoglucano afectam a integridade da parede
celular e levam à sua lise. Este processo é controlado pelas proteínas PBP (penicilin-binding-
proteins), que interagem com os antibióticos β-lactâmicos de forma a inibir irreversivelmente
as enzimas D-D-carboxitranspeptidases. Os antibióticos β-lactâmicos (penicilinas,
Síntese da Parede Celular
β-Lactâmicos
Glicopéptidos
Síntese de DNA
Quinolonas
Síntese de RNA
Rifampicina
a
Síntese Proteica
Tetraciclina
Cloranfenicol
Aminoglicosídeo
Macrolídeos
Ma
Vias Metabólicas
Sulfonamidas

20
cefalosporinas, carbapenemos e monobactâmicos), possuem um anel β-lactâmicos activo
(Figura 2), que impede a biossíntese do peptidoglucano na sua fase parietal. Em relação aos
glicopéptidos (vancomicina e teicoplanina), estes interferem igualmente na síntese do
peptidoglucano, mas ligam-se quimicamente ao terminal D-alanina-D-alanina, bloqueando a
reacção transpeptidase (Lemaire 2008;Ferreira 2010; Senka2008; Tenover 2006).
Figura 2. Estrutura geral dos antibióticos β-Lactâmicos (http://imagem.google.com).
3.2 - Inibição da Síntese Proteica
O mecanismo da inibição da síntese de proteínas bacterianas é bastante distinto da célula
eucariótica, uma vez que as subunidades ribossomais são diferentes. Assim o alvo de acção
dos macrolídeos, das tetraciclinas e dos aminoglicosídeos é ao nível das subunidades
ribossomais 30S ou 50S, impedindo que a tradução do RNA mensageiro ocorra normalmente
(Ferreira 2010; Senka 2008; Walsh2000).
À subunidade ribossomal 30S ligam-se os aminoglicosídeos que têm a capacidade de inibir a
iniciação da síntese proteica, e as tetraciclinas que impedem a ligação codão-anticodão entre o
RNA transferência e o local A dos ribossomas. À subunidade ribossomal 50S ligam-se os
macrolídeos que interferem com a elongação da cadeia polipeptídica, e os cloranfenicóis que
impedem a acção da transpeptidase (Ferreira 2010;Senka 2008; Tenover 2006).

21
3.3 - Inibição da Síntese dos Ácidos Nucléicos
Existem classes de antibióticos que actuam ao nível da replicação e da transcrição do DNA,
de modo a que estas fases não ocorram normalmente, inibindo assim a síntese dos ácidos
nucleicos (Senka 2008; Walsh 2000).
As quinolonas interferem na acção das enzimas topoisomerase II, DNA girase e
topoisomerase IV, as quais estão envolvidas na replicação do DNA. No caso da rifampicina,
esta interfere a nível da transcrição do DNA, uma vez que tem a capacidade de se ligar à RNA
polimerase (Senka 2008; Ferreira 2010; Tenover 2006).
3.4 - Ligação à Membrana Citoplasmática
A membrana celular bacteriana tem a função de semi-permeabilidade, e permite a passagem
de iões K+ acumulados no citoplasma bacteriano. A alteração dessa permeabilidade provoca
efluxo de K+, de aminoácidos e de nucleótidos, causando a morte celular. Como exemplo, as
polimixinas têm efeito inibitório onde alteram a permeabilidade da membrana externa e
ligam-se entre os fosfolípidos. A polimixina é bastante eficaz em bactérias de Gram-negativo,
pois altera tanto a membrana citoplasmática como a membrana externa (Ferreira 2010).
Tabela 1 - Mecanismos de acção de diferentes classes de antibióticos. (Adaptada de Alanis 2005)
Mecanismos de acção Família dos Antibióticos
Inibição da Síntese
da parede celular
β-lactâmicos (penicilinas, cefalosporinas, carbapenemos;
monobactâmicos), glicopeptídicos.
Inibição da Síntese
de Proteínas
Tetaciclinas; aminoglicosídeos,
Oxazalidinonas(Linezolidas),Streptogaramicinas.
Inibição da Síntese de DNA Fluoquinolonas
Inibição e Síntese de RNA Rifampicinas
Reacções Metabólicas Sulfonamidas e Trimetropim
Desorganização da
Membrana Citoplasmática
Polimixinas

22
4. - Mecanismos de Resistência aos Antibióticos
Ao longo do tempo, cada vez mais classes de antibióticos foram utilizadas de forma
indiscriminada, quer na medicina quer na pecuária e na agricultura. Perante esta pressão
selectiva, as bactérias tentaram responder com o desenvolvimento e aquisição de mecanismos
de resistência aos antibióticos, de modo a conseguirem sobreviver na presença destes
compostos. Actualmente, sabe-se que existem espécies bacterianas intrinsecamente resistentes
a determinadas classes de antibióticos, sendo estes mecanismos inatos da própria célula. No
entanto, há uma grande diversidade de espécies que têm a capacidade de adquirir genes de
resistência, alterando assim o seu perfil de susceptibilidade aos antibióticos, conseguindo
ultrapassar a acção imposta pelos antibióticos (Alonso 2001; Lim 2009;Poole 2004;
Sundsfjord 2004Alanis 2005; Martinez 2009).
Os padrões de resistência têm sido principalmente determinados por modificações na
permeabilidade da membrana externa de proteínas específicas, a outer membrane protein
(Omp), pela presença de bombas de efluxo, e pela produção de enzimas especializadas na
hidrólise de antibióticos (Figura 3). Existem bactérias com capacidade de combinar e
expressar diferentes mecanismos de resistência, desenvolvendo um perfil de resistência
significativo, como acontece com algumas espécies de K. pneumoniae que apresentam
resistência aos carbapenemos, devido à combinação da hiperprodução de ESBLs ou
cefalosporinases, com mutações em proteínas membranares, nomeadamente as OMPK.
Assim, esta combinação de mecanismos de resistência permite-lhes uma melhor “defesa”
contra os antibióticos em causa (Mohamudha 2010; Livemore 2002; Bush 1995).

23
Figura 3. Mecanismos de Resistência Bacteriana. (http://imagem.google.com)
4.1 - Resistência intrínseca
A resistência intrínseca está normalmente associada a genes cromossomais ou a determinantes
das características da bactéria, independentes das mutações ou da resistência adquirida. Este
tipo de resistência ocorre naturalmente nas células bacterianas que não sofrem alterações
genéticas ou bioquímicas no genoma e, portanto desenvolvem os seus próprios mecanismos
resistência. Sendo assim, uma espécie bacteriana pode ser resistente a uma determinada classe
de antibióticos em virtude da sua composição genética (Mulvey 2009;Harbottle 2006).
Como exemplo, os bacilos de Gram-negativo são intrinsecamente resistentes à vancomicina e
teicoplanina, e espécies de Streptococcus spp. são intrinsecamente resistentes aos
aminoglicosídeos. Em ambos os casos, a resistência intrínseca está relacionada com o elevado
peso molecular das moléculas que constituem estes compostos, o que impede a sua difusão
através das porinas presentes na membrana externa (Sousa 2006; Mulvey M.2009; Alanis
2005).
4.2 - Resistência Adquirida
A resistência adquirida envolve a aquisição de um novo material genético, e como resultado a
alteração do genótipo da bactéria. Como consequência da alteração genética, o fenótipo

24
apresentado pela bactéria receptora será igualmente alterado, sendo o resultado dos genes
adquiridos, podendo exibir assim um perfil de resistência (Mulvey 2009;Harbottle 2006).
Sendo assim, a transferência horizontal de genes de resistência tem um papel importante na
disseminação de determinantes genéticos de resistência entre diferentes espécies bacterianas,
sendo este um problema mundial cada vez mais preocupante no tratamento de infecções
bacterianas (Alonso 2001; Alanis 2005). O processo de conjugação ocorre entre espécies
devido principalmente a plasmídeos conjugativos. Contudo, há outros elementos genéticos
móveis importantes na disseminação de genes de resistência aos antibióticos, tais como
transposões, sequências de inserção e integrões (Lim 2009;Poole 2004; Sundsfjord 2004;
Ferreira 2010). Actualmente, um exemplo preocupante em todo o mundo é a disseminação
horizontal de genes que codificam enzimas β-lactamases, os quais podem ser facilmente
adquiridos por diversas bactérias patogénicas (Mulvey 2009;Harbottle 2006).
As mutações cromossomais, apesar de menos frequentes, podem também ser consideradas um
processo de resistência adquirida, uma vez que a bactéria pode exibir um novo perfil de
resistência (Mulvey 2009; Martinez 2009).
4.3 - Inactivação e Degradação enzimática do Antibiótico
Actualmente, a maioria das espécies bacterianas desenvolvem ou adquirem enzimas capazes
de inactivar ou degradar a maioria das classes de antibióticos, sendo este considerado um dos
principais mecanismos de resistência. As alterações na estrutura química destes compostos
ocorrem por hidrólise, transferência de grupos químicos e mecanismos de oxidação-redução.
Estes determinantes de resistência normalmente surgem associados a elementos genéticos
móveis, o que tem contribuído para uma elevada disseminação destes genes e para um
aumento da multirresistência (Mulvey 2009; Senka2008; Ferreira 2010).
A presença de enzimas β-lactamases constitui um dos mecanismos enzimáticos mais comuns
e representativos em bactérias de Gram-negativo, tendo apresentado, ao longo dos anos, um
impacto considerável em todo o mundo (Mulvey 2009). Estas enzimas têm a capacidade de
hidrolisar o anel β-lactâmico das penicilinas, cefalosporinas e carbapenemos, inactivando-os,
impedindo assim que estes exerçam a sua acção na célula bacteriana. A grande maioria dos
genes que codificam para enzimas β-lactamases pode estar localizado no cromossoma, ou

25
associados a elementos genéticos móveis, facilitando a sua transferência entre diferentes
espécies (Mulvey 2009; Senka 2008; Tenover 2006).
Outro grupo importante na inactivação antibiótica são as enzimas transferases, que têm a
capacidade de actuar e modificar quimicamente algumas classes de antibióticos, tais como os
aminoglicosídeos, cloranfenicol, macrolídeos e rifampicina (Senka, 2008).
4.4 -Alterações na estrutura do local alvo do antibiótico
A alteração do local de acção dos antibióticos constitui uma outra forma de resistência
igualmente importante, uma vez que facilmente pode ocorrer mutações no local alvo, de
forma a impedir que o respectivo antibiótico se ligue e exerça a sua acção (Chroma 2010).
Um exemplo deste mecanismo de resistência pode ocorrer ao nível dos genes cromossomais
que codificam as enzimas DNA girase e a topoisomerase IV, o que vai provocar uma redução
na susceptibilidade às quinolonas (Mulvey 2008; Ferreira 2010).
4.5 - Efluxo do antibiótico
O sistema de efluxo é um mecanismo importante na resistência a diversas classes de
antibióticos, nomeadamente macrolídeos, tetraciclinas e quinolonas (Senka 2008). Neste tipo
de mecanismo são os transportadores activos que bombeiam compostos para fora da célula,
diminuindo a concentração do antibiótico no interior da bactéria (Zechini 2009; Martinez
2009; Chevalier 2008). Um outro exemplo é o sistema MexA-MexB-OprD presente na
espécie P. aeruginosa, que pode estar associado à resistência ao meropenemo (Martinez
2009).
Os sistemas de efluxo são agrupados em famílias de proteínas transportadoras com base na
homologia da sequência de aminoácidos, semelhança na estrutura secundária e tamanho.
Estão descritas cinco classes: Major Facilitator Superfamily (MFS), ATP-binding cassette
(ABC), Small Multidrug Resistance (SMR), Resistance nodulation-division (RND),
Multidrug and Toxic compound extrusion (MATE). Todos utilizam força motriz de protões
como fonte de energia, com excepção da família ATP-binding que utilizam energia da
hidrólise do ATP para exportação dos substratos (Poole 2004; Mahamoud 2007).

26
O sistema de efluxo da família RND é o mais importante sistema de resistência aos
antibióticos em bactérias de Gram-negativo, apresenta uma baixa especificidade de substrato,
sendo capaz de transportar uma variedade de compostos como antibióticos, biocidas, corantes,
detergentes, inibidores metálicos, hidrocarbonetos aromáticos, sais biliares, homoserinas e
lactonas associadas com o “quorum-sensing” (Poole 2004; Moreira 2004; Martinez 2009).
4.6 – Alteração da Permeabilidade membranar
A parede celular das bactérias de Gram-negativo difere da parede celular das bactérias de
Gram-positivo, devido à presença de uma membrana externa, a qual tem um papel importante
como barreira permeável que evita a entrada de substâncias nocivas, mas ao mesmo tempo
permite o fluxo de moléculas de nutrientes. Este fluxo de moléculas para o interior da célula é
assegurado por um complexo de proteínas transmembranares ou porinas, nomeadamente as
OMPs. As porinas permitem a difusão de solutos hidrófilos e hidrofílicos, de acordo com o
seu peso molecular, dimensão e estrutura (Nikaido 1994; Ferreira 2010). Em algumas
espécies, a sobreexpressão ou subexpressão de uma ou mais porinas pode estar relacionada
com a resistência a algumas classes de antibióticos (Nikaido 1994).
5 - Disseminação dos genes de resistência aos antibióticos
Como referido anteriormente, actualmente, é cada vez mais frequente a emergência de
estirpes multirresistentes aos antibióticos, principalmente devido à rápida disseminação de
genes de resistência. Esta transferência horizontal de genes ocorre devido a elementos
genéticos móveis, contribuindo para a evolução e adaptação bacteriana ao meio envolvente
(Benett 2008; Kelly 2009).
As modificações na herança genética podem ser feitas por uma de duas maneiras: i) por
mutações que alteram o DNA de uma célula pré-existente, essas alterações resultam de
alterações de bases, deleções e ou inversões no DNA e ii) por aquisição de um novo material,
através da captura de novos genes (Kelly 2009).
As bactérias apresentam três principais mecanismos de transferência de determinantes de
resistência de uma célula bacteriana para outra, tais como a transformação, a conjugação e a
transdução (Figura 4).

27
Figura 4. Mecanismos genéticos de transferência de genes entre bactérias. (A) Transformação; (B) Transdução e
(C) Conjugação (Adaptada Furuya 2006).
5.1 – Mutação
As mutações espontâneas ocorrem aleatoriamente por delecção, duplicação, inversão ou
translocação de fragmentos de DNA, durante a sua replicação, podendo por vezes não serem
reparadas por mecanismos celulares. No entanto, existem mutações induzidas por agentes
mutagénicos físicos e químicos. As mutações provocam alterações nas sequências de bases de
uma determinada região genómica, e como consequência, modificações nos níveis de
expressão dos genes, e nas funções das proteínas (Ferreira 2010;Lima e Mota 2003; Senka
2008). Assim, a resistência aos antibióticos pode ocorrer por mutações pontuais nos
nucleótidos da cadeia de DNA em crescimento e permitem produzir um fenótipo de
resistência (Senka 2008; Normark 2002).

28
5.2 - Transformação
A transformação é uma propriedade natural de algumas espécies bacterianas para captar
fragmentos de DNA do meio ambiente, os quais são incorporados no genoma bacteriano por
recombinação homóloga. Esta alteração genotípica pode conferir um novo fenótipo / perfil de
resistência à bactéria (Ferreira 2010; Kelly 2009).
As bactérias competentes que utilizam um mecanismo de captação de DNA não
selectivo podem translocar não só fragmentos de DNA cromossómicos, como
plasmídeos; sendo que a captação de plasmídeos pelas células naturalmente
competentes é menos eficaz dada a complexidade do processo (Ferreira 2010).
5.3 - Conjugação
A conjugação é um importante mecanismo e o mais comum na transferência de genes de
resistência aos antibióticos, de uma célula dadora com um determinado perfil de resistência,
para uma célula receptora, normalmente com elevada susceptibilidade. Este mecanismo é
mediado por plasmídeos conjugativos, os quais sofrem inicialmente replicação, e
posteriormente são transferidos entre bactérias através da formação de pillus sexual que
permite a ligação entre as duas células (Alanis 2005; Kelly 2009;Ferreira 2010;Lima e
Mota2003). Após a transferência do plasmídeo ou de parte dele para a célula receptora, esta
pode mantê-lo viável, ou se não o conseguir suportar este degenera (Tenover 2008; Summers
2006).
5.4 - Transdução
A transdução é mediada por bacteriófagos que permitem a transferência de material genético
entre bactérias, e por isso contribuem para a disseminação de determinantes de virulência e
para a evolução de bactérias patogénicas (Kelly 2009). A célula bacteriana é usada para a
multiplicação das partículas virais e, no final do processo, ocorre a lise da célula bacteriana
com libertação de novas partículas para o ambiente circundante (Kelly 2009;Ferreira
2010;Lima e Mota2003).
Na transdução generalizada qualquer gene da bactéria dadora pode ser transferido para a
bactéria receptora; na transdução especializada, sendo um processo mais específico, sendo

29
um processo dependente da ocorrência de um ciclo lisogênico. Regiões específicas do DNA
bacteriano unem-se covalentemente ao DNA do fago, são empacotados e transferidas. Após
um determinado período de tempo e de acordo com certos estímulos, este fago pode iniciar
um ciclo lítico (Ferreira 2010; Kelly 2009).
5.5 - Elementos Genéticos Móveis
Como referido anteriormente, a disseminação de genes de resistência aos antibióticos está
associada a elementos genéticos móveis, e é um problema crescente em todo o mundo, pois
compromete o tratamento de infecções bacterianas (Mulvey 2009; Mindlin 2006; Partridge
2002, Harbottle 2006).
Os elementos genéticos móveis são segmentos de DNA que codificam proteínas importantes
na mediação e movimentação dos fragmentos de DNA. Estes elementos podem mover-se de
um local para outro dentro do genoma de uma mesma célula bacteriana, onde se incluem os
transposões, integrões e sequência de inserção; ou podem mover-se de uma célula para outro,
pelos mecanismos já descritos. Em ambos os casos, normalmente ocorre mobilização de
genes de resistência a antibióticos (Kelly 2009;Benett 2008).
5.5.1 - Plasmídeos
Plasmídeos são moléculas circulares de DNA de cadeia dupla, extra-cromossomais, com
capacidade auto-replicativa e que contêm genes que codificam os elementos necessários para
sua expressão e mobilização. Estas estruturas não possuem genes essenciais à sobrevivência
das bactérias, mas quando presentes conferem determinadas vantagens: capacidade de
conjugar, capacidade de expressar genes que conferem resistência aos antibióticos ou que
codificam para factores de virulência, e capacidade de metabolizar alguns substratos (Bennett
2008; Clewell 2005, Harbottle 2006). Assim, é cada vez mais evidente que os plasmídeos
contribuem para a diversidade fenotípica nas comunidades bacterianas (Bahl 2007).
O tamanho dos plasmídeos é variável, e a sua complexidade genética aumenta com o
tamanho. Os plasmídeos de pequenas dimensões – plasmídeos não conjugativos – possuem
apenas informação para a sua auto-replicação, não apresentam mobilidade horizontal, mas
podem ser mobilizados para outras células por plasmídeos conjugativos que se encontrem na

30
mesma célula. Os plasmídeos de maiores dimensões – plasmídeos conjugativos – codificam
funções necessárias (genes tra) para promover a transferência horizontal de genes, e assumem
um importante papel na disseminação de genes de resistência e na problemática mundial da
multiresistência bacteriana (Bennett 2008; Clewell 2005, Ferreira 2010).
Os plasmídeos que transportam genes de resistência são denominados Plasmídeo R ou Factor
R. Estes elementos são móveis e podem transportar um ou mais genes de resistência, como
por exemplo, a metais pesados, a alguns anti-sépticos, antibióticos e desinfectantes (Clewell
2005; Harbottle 2006; Li 2005). Normalmente, estes plasmídeos podem transportar outros
elementos como integrões, transposões e/ou sequência de inserção, permitindo a combinação
de vários determinantes de resistência aos antibióticos num único elemento genético móvel
(Novais 2010).
5.5.2 - Transposões
Transposões são segmentos de DNA linear delimitados por sequências de nucleótidos
repetidas e invertidas, que podem “saltar” de um local para outro, nomeadamente do
cromossoma para o plasmídeo bacteriano, ou vice-versa. Todos os genes que se encontrem
associados a estes elementos, como por exemplo, genes de resistência aos antibióticos, são
igualmente transportados e disseminados (Mulvey 2009; Clewell 2005, Kelly 2009).
Os transposões que pertencem ao conjunto de elementos transponíveis mais simples são as
sequências de inserção (IS) (Figura 6). Normalmente movem-se de um local para outro no
cromossoma bacteriano ou para o plasmídeo, ou vice-versa (Clewell 2005; Bennett 2008,
Kelly 2009).

31
Figura 6. Representação esquemática de um transposão simples
O mecanismo de transposição ocorre devido à recombinação genética entre a sequência
nucleotídica do transposão e dos genes a ele associados, e o local de inserção. Sendo assim,
este mecanismo é mediado pela enzima transposase (tnpA) que se encontra associada ao
transposão, e flanqueada por sequências invertidas (IR) características de cada IS. Esta
enzima reconhece a sequência do local de recombinação e permite assim a inserção ou
excisão dos genes associados ao transposão (Bennett 2008, Ferreira 2010; Clewell 2005).
Além dos transposões simples, existem os transposões compostos, como representa a Figura
7, os quais apresentam duas IS a ladear um segmento de DNA (Souza 2006, Ferreira 2010).
Figura 7. Representação esquemática de um Transposão Composto.
IS IS
SRI SRI SRI SRI G1 G2
Transposão Composto
Transposase

32
5.5.3 - Integrões
Nos anos 80 foi descoberto um outro elemento que permite a disseminação de genes de
resistência aos antibióticos – o integrão – cuja integração e excisão destes genes ocorre por
um mecanismo sítio-específico (Lévesque 1995; Hall e Collis 1995).
Hall e Collis (1995) definiram os integrões como sendo elementos genéticos não móveis por
si só, mas que têm a capacidade de reconhecer e capturar genes cassette por um sistema de
recombinação sítio-específico (Peters 2001; Fluit e Schmitz 1999; Hall e Collis 1995). Os
genes cassette normalmente codificam resistência aos antibióticos, e quando integrados
podem ser expressos devido a um promotor presente na estrutura conservada do integrão.
Perante esta situação, os integrões são outros dos elementos responsáveis pela disseminação
de multirresistência em bactérias de Gram-negativo, quer no ambiente hospitalar, quer fora
deste, uma vez que normalmente se encontram associados a plasmídeos, transposões ou
sequências de inserção (Cambray 2010; Mazel 2006; Summer 2006; Bennett 2004; Fruit e
Schmitz 2004).
5.5.3.1 - Constituição dos integrões
Regra geral, um integrão é composto / flanqueado por duas regiões conservadas terminais
(5’CS e 3’CS), e uma região central constituída por uma diversidade de genes cassette que
normalmente conferem resistência às diferentes classes de antibióticos (Figura 8) (Hall e
Collis 1995; Bennett 2008).
Figura 8. A figura esquematiza as três regiões que constituem um integrão.
intI attI
Pc
P2
qacEA sulI
5’CS 3’CS
- - - - - - - - - SRI
- - - - - - -

33
5.5.3.2- Segmento Conservado 5’ (5’CS)
O 5’CS constitui o “coração” do integrão, uma vez que inclui os componentes essenciais para
o mecanismo de integração e expressão dos genes de cassette. Sendo assim, este segmento é
composto por: a) um gene que codifica a enzima integrase (intI); b) um local de recombinação
adjacente (attI) que permite o reconhecimento e integração / inserção dos genes cassette na
região variável do integrão; e c) um promotor principal, Pc (ou PAnt) que permite a expressão
dos genes cassette (Carattoli 2001; Soto 2003). Além deste promotor, neste segmento está
presente um segundo promotor, provavelmente mais fraco e orientado em sentido contrário ao
Pc, sendo este designado por promotor P2 (Fluit e Schmitz 2004; Cambray 2010).
O processo de integração ocorre devido à recombinação sítio-específico entre as sequências
homólogas dos locais específicos attI do 5’CS e attC do gene cassette (Ceccarelli 2007;
Gonzáles 2004). Quando integrados na região variável, os genes cassette podem ser expressos
pela enzima integrase (intI), a qual pertence à superfamília das tirosinas recombinases
(Mindlin 2006; Collis e Stokes 2002; Ceccarelli 2007).
O promotor Pc é constituído por duas sequências de seis nucleótidos nas posições -35 e -10
bp, e regula a expressão dos genes de resistência integrados na região variável. Estudos
anteriores revelaram algumas diferenças nos níveis de expressão dos genes cassette, e
concluíram que quanto mais próximo o gene cassette estiver da região promotora, maior será
a probabilidade deste gene ser expresso, e consequentemente maior poderá ser o nível de
resistência de acordo com o meio envolvente (Bennett 2008;Collis e Hall 1995).
5.5.3.3 - Segmento Conservado 3’ (3’ CS)
O 3’CS é composto por dois genes de resistência que se encontram fundidos: a) um gene que
confere resistência aos compostos de amónio quaternário (qacEΔ1); e b) um gene que confere
resistência às sulfonamidas (sul1) (Fluit e Schmitz 1999; Soto 2003). A montante destes
genes, por vezes surge um terceiro gene designado por insertion sequence commun region
(ISCR1), o qual permite a mobilização de genes de resistência que se encontrem a montante
deste segmento (Bennett 2008).

34
Vários trabalhos têm mostrado que este segmento nem sempre se encontra com esta
disposição conservada. O que está acontecer, é que foram descritos integrões que não
apresentavam este segmento, ou seja, o gene qacEΔ1/sulI foi delectado, e no seu lugar
permaneceram genes correspondentes ao transposão Tn402 (Santos, 2010; Toleman, 2007).
5.5.3.4 - Região Variável (RV)
Esta região, como o nome indica, pode ser constituída por um tamanho e número variável de
genes cassette, os quais normalmente conferem resistência a diferentes classes de antibióticos.
Os genes cassette são integrados de forma sequencial, e sempre da posição 5’-3’, não havendo
um limite para o número de genes cassette inseridos (Hall e Collis 1995; Bennett 2004). Em
alguns casos, esta região pode não apresentar qualquer tipo de gene cassette, e por isso, diz-se
que se encontra vazia (Gillings 2008).
5.5.3.5 - Genes Cassette
Os genes cassette não apresentam promotores e podem ser encontrados sob duas formas: a)
podem-se apresentar sob a forma de uma molécula livre circular incapaz de promover por si
só a sua replicação e expressão; b) podem-se apresentar sob a forma de uma molécula linear,
fazendo parte do conteúdo genético do integrão (Michael 2004;Sabaté 2002; Bennett 2008).
Os genes cassette inseridos na região variável do integrão têm normalmente entre 500 a 1000
bp, e apresentam características estruturais específicas, tais como um único gene de
resistência e um local de recombinação de elemento base 59 (59-be ou attC). Este local de
recombinação vai ser reconhecido pelo local de recombinação attI presente no 5’CS, o que vai
permitir a mobilização dos genes cassette na região variável (Bennett 2004;Fluit e Schmitz
1999).

35
Figura 9. A figura representa o mecanismo de integração dos genes cassette na região variável de um integrão.
5.5.3.6 - Classificação dos Integrões
Os primeiros integrões descritos foram classificados como integrões de resistência, uma vez
que na sua maioria transportam genes que conferem resistência às diferentes classes de
antibióticos. Estes integrões foram divididos em três classes, as quais normalmente se
encontram associadas a plasmídeos, transposões e sequências de inserção, e apresentam um
número reduzido de genes cassette integrados na RV (Gillings 2008). Mais tarde, uma quarta
classe de integrões de resistência foi descrita, a qual foi designada como super-integrões, uma
vez que apresentam uma RV constituída por inúmeros genes cassette (Collis 2002; Ceccarelli
2007).
Actualmente estão descritas nove classes de integrões, e a sua classificação baseia-se nas
diferenças das sequências nucleotídicas dos genes que codificam a enzima integrase (Sabaté
2002).

36
5.5.3.6.1 - Integrões de Classe 1
Esta classe de integrões é uma das mais comuns entre os isolados clínicos de bactérias de
Gram-negativo, mas a sua presença em bactérias de Gram-positivo já foi descrita em
trabalhos anteriores (Collis 2002; Xu 2011). Estes integrões de resistência normalmente
apresentam um ou mais genes cassette integrados na RV, os quais conferem resistência a
diferentes classes de antibióticos, nomeadamente antibióticos β-lactâmicos, aminoglicosídeos,
cloranfenicol, rifampicina, trimetoprim/sulfametoxazol, quinolonas, entre outras (Sabaté
2002; Partridge 2002). Mais de 100 genes cassette já foram identificados em integrões de
classe 1, sendo os mais conhecidos e melhor caracterizados, associados a bactérias de Gram-
negativo multirresistentes, nomeadamente Escherichia coli, Citrobacter spp., Klebsiella spp.,
Enterobacter spp., Proteus spp., Acinetobacter spp. e Pseudomonas spp. (Peng 2007; Stokes
2006).
Tal como descrito anteriormente e representado na Figura 9, este tipo de integrões
caracterizam-se pela presença de dois segmentos conservados, o 5’CS e o 3’CS, e uma RV
(Sabaté 2002; Partridge 2002; Lévesque 1995).
Actualmente está descrito que este tipo de integrões pode estar associado a transposões
derivados do transposão Tn402, uma vez que na ausência do 3’CS, os genes a montante da
RV constituem a estrutura tni completa, formada pelos genes tniC, tniB e tniA (Sabaté 2002;
Partridge 2002; Lévesque 1995).
5.5.3.6.2 - Outras Classes de Integrões
Os integrões de classe 2 apresentam um número reduzido de genes cassette integrados na
região variável, e dois segmentos conservados (5’CS e 3’CS) (Hall e Collis 1998). Uma
diferença observada neste caso, reside na sequência nucleotídica do gene que codifica a
enzima integrase classe 2 (intI2), na qual ocorreu uma mutação no codão 179, e daí existir
uma homologia de apenas 40% com a enzima integrase classe 1 (Cambray 2010; Sabaté 2002;
Hall e Collis 1998).
Diferentes trabalhos até agora realizados, mostraram que a região variável encontrada com
maior frequência é constituída por três genes cassette (dfrA1 – sat2 – aadA1), e que este tipo

37
de integrões normalmente se encontra associado ao transposão Tn7 (Bennett 2004; Cambray
2010; Sabaté 2002).
Em relação aos integrões de classe 3, a enzima integrase (intI3) apresenta 61% de homologia
com a sequência de aminoácidos do gene intI1, e tem igualmente a capacidade de integrar
genes de resistência, mas de uma forma menos frequente (Fluit 2004; Carattoli 2001).
No caso dos integrões de classe 4, também designados como super-integrões, uma vez que
apresentam múltiplos genes cassette integrados na região RV. Collis et al. (2002) descreve a
presença de um super-integrão em estirpes de Vibrio cholerae, o qual está associado ao
cromossoma bacteriano mais pequeno. Esse integrão apresenta uma RV constituída por 179
genes cassette, mas apenas alguns conferem resistência aos antibióticos (Collis 2002;
Ceccarelli 2007).
Além destas classes de integrões, foram descritas outras classes, as quais não se encontram
muito ligadas a genes de resistência aos antibióticos, e talvez por isso raramente têm sido
encontradas em isolados clínicos. As enzimas integrases destas classes de integrões
apresentam uma homologia com o gene intI1 que varia entre 45 e 50% (Sabaté 2002).

38
Objectivo
Uma das preocupações dentro do ambiente hospitalar é o controlo e a monitorização de surtos
e de infecções bacterianas. Entre estas, as infecções causadas por bactérias de Gram-negativo
que apresentam elevada resistência aos antibióticos são particularmente preocupantes.
Assim, constituiu objectivo do presente estudo avaliar a presença de genes de resistência aos
antibióticos, associados a integrões de classe 1, em bactérias de Gram-negativo
multirresistentes no ambiente hospitalar, nomeadamente no Serviço de Medicina Intensiva e
no Serviço de Medicina II a pesquisar.

39
MATERIAL E MÉTODOS

40
7 - Material e Métodos
7.1 - Identificação dos locais de recolhas dos Isolados
As amostras bacterianas foram recolhidas em diferentes superfícies e equipamentos
hospitalares do Serviço de Medicina II e do Serviço de Medicina Intensiva, do Hospital
Infante D. Pedro em Aveiro.
No Serviço de Medicina II são internados homens e mulheres principalmente de idade
avançada e com algumas complicações. Nesta secção, os isolados bacterianos foram
recolhidos a partir das grades das camas dos pacientes. Esta recolha foi realizada em 2008,
com o intuito de comparar os resultados finais com os resultados obtidos num trabalho
idêntico realizado em 2005.
Em relação ao Serviço de Medicina Intensiva, este recebe pacientes em estado crítico e
normalmente imunodeprimidas, estando por isso mais expostas a infecções bacterianas. Este
serviço é amplo, e composto apenas por seis camas e um quarto de isolamento. Em 2010,
foram recolhidas amostras bacterianas a partir de várias superfícies hospitalares,
nomeadamente do carrinho de emergência, da maçaneta da porta de entrada, do chão antes e
depois da limpeza, das torneiras e do suporte de sabão líquido, das grelhas do ar
condicionado, da bancada de prescrição médica, da secretária e do telefone, dos teclados,
ratos e monitores de computadores e das camas. Além desta recolha, em simultâneo foi
realizada uma outra recolha a partir de diferentes equipamentos hospitalares tais como
estetoscópios, ventiladores mecânicos, monitores cardíacos, bombas de infusão e bombas de
seringas.
7.2– Recolha, Selecção e Conservação dos Isolados bacterianos
As amostras bacterianas foram recolhidas a partir das superfícies referidas anteriormente com
a ajuda de uma zaragatoa estéril humedecida em água destilada esterilizada. Após a passagem
da zaragatoa pela superfície, esta foi inoculado em 1 ml de meio líquido nutritivo de caldo de
triptona de soja (TSB) (Sigma-Aldrich, Portugal), e incubada a 37ºC durante
aproximadamente 18h. Posteriormente, e após se ter registado crescimento bacteriano, as
alíquotas foram conservadas a -80 ºC após adição de glicerol a 45%.

41
Durante o tempo de estudo, cada amostra foi descongelada, diluída (10-1
, 10-3
, 10-5
) e semeada
em meio selectivo para bactérias de Gram-negativo, designado Mackonkey agar (McK)
(OXOID, Portugal). Todas as placas foram incubadas a 37°C durante aproximadamente 18h.
Após o tempo de incubação, verificou-se que algumas placas de McK apresentavam
crescimento e outras não. No caso das placas com crescimento bacteriano, todas as colónias
isoladas foram repicadas para placas de tryptone soya agar (TSA) (Merck, Portrugal), e
incubadas nas mesmas condições anteriores. No caso das placas de McK em que não se
observou qualquer crescimento bacteriano, as diluições originais foram inoculadas em meio
em meio TSA, e incubadas nas mesmas condições, de modo a verificar se as amostras
conservadas em glicerol estariam viáveis.
7.3– Extracção de DNA
Durante o estudo, para alguns isolados, foi necessário proceder à extracção de DNA total
bacteriano. Para este fim foi utilizado o kit Genomic DNA Purification (Fermentas), e
seguidas as indicações do fabricante:
1. Inocular os colónias isoladas em TSB a 37ºC durante 24h.
2. Centrifugar (5’/1000rpm) 2 ml da suspensão bacteriana e desprezar o sobrenadante.
3. Ressuspender bem todo o pellet em 200µl de tampão TE.
4. Adicionar 400µl da solução de lise fornecida no kit, e incubar em banho seco
(5’/65ºC).
5. Adicionar 600µl de clorofórmio e centrifugar (2’/10000rpm).
6. Adicionar 800µl da solução de precipitação aquosa fornecida no kit e centrifugar
novamente (2’/10000rpm).
7. Dissolver bem o pellet em 100µl de NaCl.
8. Adicionar 300µl de etanol (EtOH) absoluto frio e deixar precipitar 10’-1h a -20ºC.
9. Descartar o EtOH, ressuspender o DNA em 100µl de água destilada estéril e conservar
a -20°C.

42
7.4 Caracterização Fenotípica
7.4.1- Avaliação da Clonalidade dos Isolados
Com o intuito de reduzir o número de isolados bacterianos, é necessário seleccionar diferentes
grupos clonais. Para este fim, utiliza-se a técnica molecular BOX-PCR, na qual é utilizado o
primer BOXA1R que permite a amplificação de regiões cromossomais específicas. Os
isolados foram seleccionados com base nos géis realizados pelo BOX PCR, tendo em conta 3
ou mais bandas diferentes. Nesta reacção foram utilizados os reagentes presentes na Tabela 3,
assim como o programa de amplificação representado na mesma tabela.
Tabela 2 – Reagentes e respectivo primer e programa de amplificação utilizados na distinção de
diferentes grupos clonais.
Primer Sequência Referência
BOXA1R 5’-CTACGGCAAGGCGACGCTGACG-3’ Versalovic 1994
Reagente Volume
25 mM MgCl2 1,5 μl Fases do PCR Programa
Buffer (5x) 2,5 μl Desnaturação inicial 95°C 7’
10 mM dNTP’s 0,25 μl Desnaturação 94°C 1’
DMSO 0,63 μl Hibridação dos primers 53°C 1’
dH2O 6,8 μl Síntese das cadeias 65°C 8’
Primer BOXA1R 0,38 μl Ciclos 35 vezes
Taq polymerase (5U/ μl) 0,06 μl Síntese final 65°C 16’
Vf = 12,5 μl Após final da reacção 15°C ∞
DNA total / suspensão 1 μl
No final desta reacção, os produtos amplificados foram separados num gel de agarose a 1%,
tal como está descrito na secção 7.4.4, o que permitiu observar diferentes perfis bacterianos e
distinguir diferentes grupos clonais.

43
7.4.2 – Amplificação dos Ácidos Nucleicos por PCR ( Polymerase Chain Reaction)
Tendo em conta o ponto anterior, de cada grupo clonal foi seleccionado um isolado
bacteriano, reduzindo desta forma o número de isolados em estudo. Todos os isolados
seleccionados foram utilizados no prosseguimento deste estudo. Sendo assim, e recorrendo à
técnica de PCR, inicialmente foi realizado um “screening” para detectar a presença da enzima
integrase de classe 1 (intI1), utilizando os primers e as condições de amplificação resumidas
na Tabela 3 e na Tabela 4, respectivamente. Posteriormente, em todos os isolados positivos
para o gene intI1, foi caracterizada a respectiva região variável (RV), utilizando os primers e
as respectivas condições de amplificação descritas nas Tabela 3 e 4. Nestes isolados foi
também testada a presença do gene tniC, como resultado da delecção do 3’CS. Neste caso foi
igualmente utilizado o primer e as condições resumidas nas Tabelas 3 e 4.
Em alguns isolados em particular, foi também testada a presença do elemento ISCR1
localizado a montante da RV do integrão de classe 1. Sendo assim, e caso a sua presença se
verificasse, uma segunda RV pode ser amplificada constituindo assim um integrão complexo
de classe 1. Neste caso foi igualmente utilizado o primer e as condições resumidas nas
Tabelas 3 e 4.
Para confirmar a eficácia da reacção de amplificação em todos os casos anteriores, foi
utilizado um controlo positivo. Sendo assim, neste caso foi utilizado um isolado de K.
pneumoniae, o qual apresenta um integrão complexo de classe 1 (Santos, 2010a).
44
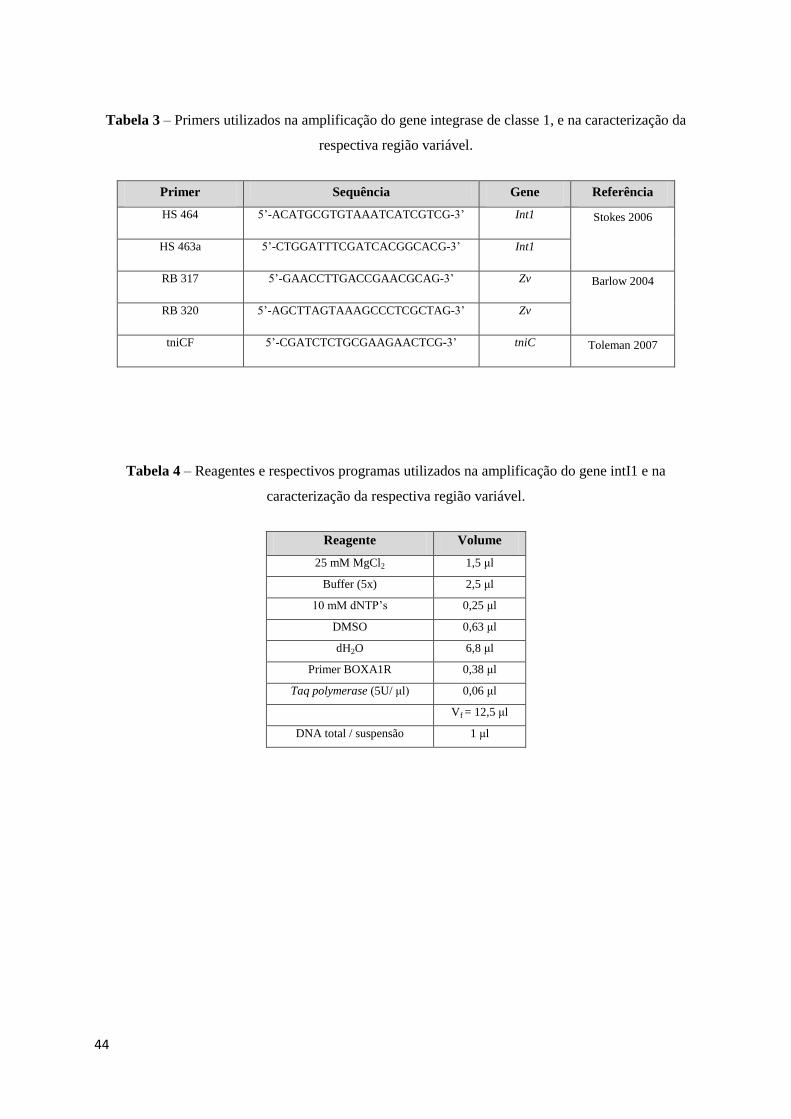
Tabela 3 – Primers utilizados na amplificação do gene integrase de classe 1, e na caracterização da
respectiva região variável.
Primer Sequência Gene Referência
HS 464 5’-ACATGCGTGTAAATCATCGTCG-3’ Int1 Stokes 2006
HS 463a 5’-CTGGATTTCGATCACGGCACG-3’ Int1
RB 317 5’-GAACCTTGACCGAACGCAG-3’ Zv Barlow 2004
RB 320 5’-AGCTTAGTAAAGCCCTCGCTAG-3’ Zv
tniCF 5’-CGATCTCTGCGAAGAACTCG-3’ tniC Toleman 2007
Tabela 4 – Reagentes e respectivos programas utilizados na amplificação do gene intI1 e na
caracterização da respectiva região variável.
Reagente Volume
25 mM MgCl2 1,5 μl
Buffer (5x) 2,5 μl
10 mM dNTP’s 0,25 μl
DMSO 0,63 μl
dH2O 6,8 μl
Primer BOXA1R 0,38 μl
Taq polymerase (5U/ μl) 0,06 μl
Vf = 12,5 μl
DNA total / suspensão 1 μl
45
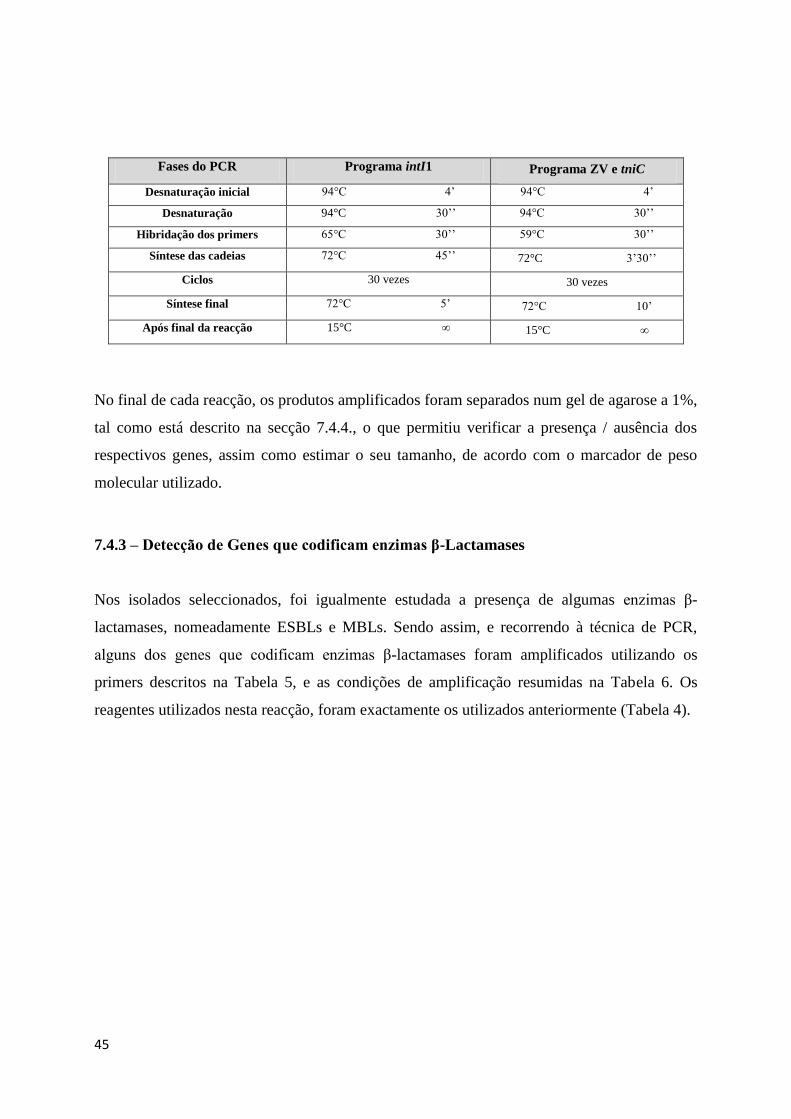
Fases do PCR Programa intI1 Programa ZV e tniC
Desnaturação inicial 94°C 4’ 94°C 4’
Desnaturação 94°C 30’’ 94°C 30’’
Hibridação dos primers 65°C 30’’ 59°C 30’’
Síntese das cadeias 72°C 45’’ 72°C 3’30’’
Ciclos 30 vezes 30 vezes
Síntese final 72°C 5’ 72°C 10’
Após final da reacção 15°C ∞ 15°C ∞
No final de cada reacção, os produtos amplificados foram separados num gel de agarose a 1%,
tal como está descrito na secção 7.4.4., o que permitiu verificar a presença / ausência dos
respectivos genes, assim como estimar o seu tamanho, de acordo com o marcador de peso
molecular utilizado.
7.4.3 – Detecção de Genes que codificam enzimas β-Lactamases
Nos isolados seleccionados, foi igualmente estudada a presença de algumas enzimas β-
lactamases, nomeadamente ESBLs e MBLs. Sendo assim, e recorrendo à técnica de PCR,
alguns dos genes que codificam enzimas β-lactamases foram amplificados utilizando os
primers descritos na Tabela 5, e as condições de amplificação resumidas na Tabela 6. Os
reagentes utilizados nesta reacção, foram exactamente os utilizados anteriormente (Tabela 4).

46
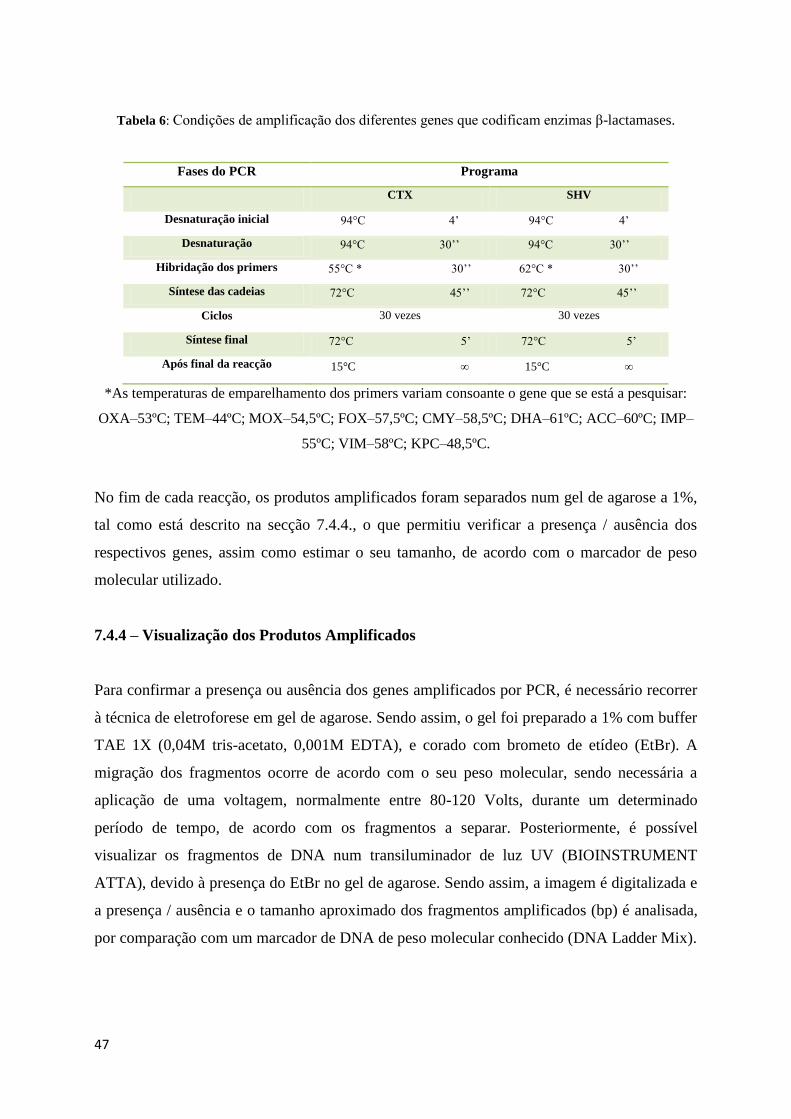
Tabela 5: Primers utilizados na pesquisa de alguns genes que codificam para enzimas β-lactamases.
Designação do
Primer
Sequência Gene a
amplificar
Tamanho do
fragmento
Referências
CTX Rv 5’-CGCAATATCATTGGTGGTGCC-
3’
CTX 538bp
Henriques et al 2006
CTX Fw 5’-
GTGCAGTACCAGTAAAGTTATGG
-3’
CTX 538bp
SHV Rv 5’-GATTGGCGGCGCTGTTATCGC-
3’
SHV 304bp
SHV Fw 5’-GCGAAAGCCAGCTGTCGGGC-
3’
SHV 304bp
OXA Rv 5’-AGTGTGTTTAGAATGGTGATC-
3’
OXA 814bp
OXA Fw 5’-
ACACAATACATATCAACTTCGC -
3’
OXA 814bp
TEM Rv 5’-TTTGGTATGGCTTCATTC-3’ TEM 425bp
TEM Fw 5’-AAAGATGCTGAAGATCA-3’ TEM 425bp
IMP Rv 5’-GTTTTAAYAAAACAACCACC-3’ IMP 232bp
IMP Fw 5’-GAATAGAGTGGATTAATTCTC-
3’
IMP 232bp
VIM Rv 5’-GCCACGTTCCCCGCAGACG-3’ VIM 475bp
VIM Fw 5’-
GATGGTGTTTGGTCGCATATCG-3’
VIM 475bp
KPC Rv 5’-CGTTGACGCCCAATCC-3’ KPC 800bp Fontana et al 2010
KPC Fw 5’-TGTCACTGTATCGCCGTC-3’ KPC 800bp
47
Tabela 6: Condições de amplificação dos diferentes genes que codificam enzimas β-lactamases.
Fases do PCR Programa
CTX SHV
Desnaturação inicial 94°C 4’ 94°C 4’
Desnaturação 94°C 30’’ 94°C 30’’
Hibridação dos primers 55°C * 30’’ 62°C * 30’’
Síntese das cadeias 72°C 45’’ 72°C 45’’
Ciclos 30 vezes 30 vezes
Síntese final 72°C 5’ 72°C 5’
Após final da reacção 15°C ∞ 15°C ∞
*As temperaturas de emparelhamento dos primers variam consoante o gene que se está a pesquisar:
OXA–53ºC; TEM–44ºC; MOX–54,5ºC; FOX–57,5ºC; CMY–58,5ºC; DHA–61ºC; ACC–60ºC; IMP–
55ºC; VIM–58ºC; KPC–48,5ºC.
No fim de cada reacção, os produtos amplificados foram separados num gel de agarose a 1%,
tal como está descrito na secção 7.4.4., o que permitiu verificar a presença / ausência dos
respectivos genes, assim como estimar o seu tamanho, de acordo com o marcador de peso
molecular utilizado.
7.4.4 – Visualização dos Produtos Amplificados
Para confirmar a presença ou ausência dos genes amplificados por PCR, é necessário recorrer
à técnica de eletroforese em gel de agarose. Sendo assim, o gel foi preparado a 1% com buffer
TAE 1X (0,04M tris-acetato, 0,001M EDTA), e corado com brometo de etídeo (EtBr). A
migração dos fragmentos ocorre de acordo com o seu peso molecular, sendo necessária a
aplicação de uma voltagem, normalmente entre 80-120 Volts, durante um determinado
período de tempo, de acordo com os fragmentos a separar. Posteriormente, é possível
visualizar os fragmentos de DNA num transiluminador de luz UV (BIOINSTRUMENT
ATTA), devido à presença do EtBr no gel de agarose. Sendo assim, a imagem é digitalizada e
a presença / ausência e o tamanho aproximado dos fragmentos amplificados (bp) é analisada,
por comparação com um marcador de DNA de peso molecular conhecido (DNA Ladder Mix).

48
7.4.5 – Digestão da Região Variável
Uma vez que vários isolados bacterianos apresentavam tamanhos muito semelhantes da RV,
uma das formas rápidas de compreender se estamos perante ou não de uma RV com a mesma
constituição, é através da digestão desta região.
Para isso, foi utilizado 1µl da enzima Taq1, 2µl do tampão da mesma e 5µl do produto
amplificado por PCR. A digestão ocorreu durante uma hora a 65ºC. Posteriormente, os
produtos digeridos foram separados por electroforese em gel de agarose, e a obtenção de um
perfil idêntico, significa que as RV apresentam a mesma constituição.
7.4.6 – Purificação dos Produtos de PCR
Os produtos amplificados por PCR foram purificados de acordo com as instruções do
fabricante do kit de purificação QlAquick (QIAGEN, Alemanha). Em alguns casos é
necessário recorrer a esta purificação antes da sequenciação, uma vez que o fragmento que se
pretende sequenciar não se encontrava puro.
7.4.7 – Determinação da Sequência Nucleotídica
Os produtos purificados foram enviados para a empresa StabVida (Oeiras Portugal) para
posterior sequenciação. A sequenciação foi efectuada num sequenciador automático ABI
3700 (Apllied Biosytems, Perkin-Elmer, Foster City, C.A).
As sequências nucleotídicas e as respectivas sequências de aminoácidos foram analisadas
recorrendo ao programa informático Blast e ClustalW (BioEdit), e comparadas com as
sequências depositadas no GenBank.

49
RESULTADOS E DISCUSSÃO

50
8 - Resultados e Discussão
8.1 - Serviço de Medicina II (SMII)
8.1.1 Tipagem Molecular
No SMII foram recolhidas e isoladas bactérias de Gram-negativo das grades das camas dos
pacientes que lá se encontravam internados. Uma vez que a população bacteriana isolada foi
numerosa, foi necessário recorrer a técnicas moleculares de tipagem genética, nomeadamente
o BOX-PCR. Portanto, a análise dos diferentes perfis genéticos permitiu identificar diferentes
grupos clonais, e por isso reduzir o número de isolados bacterianos em estudo. Deste modo,
foi apenas seleccionado um isolado bacteriano de cada grupo clonal, e de um total de 174
isolados bacterianos, 47 (27%) pertenciam a diferentes grupos clonais, os quais vão ser
utilizados no desenrolar desta primeira parte do trabalho, com o intuito de se conhecer melhor
o seu perfil de resistência aos antibióticos.
8.1.2 Caracterização da Região Variável
Numa primeira abordagem, e recorrendo à técnica molecular PCR, em todos os 47 isolados
bacterianos foi realizado um screening para detectar a presença do gene que codifica a enzima
integrase classe 1 (intI1). Foi possível verificar que dos 47 isolados apenas 11 (23%)
apresentam o respectivo gene. Assim, nos isolados positivos, onde foi posteriormente
amplificada, sequenciada e analisada a respectiva região variável (RV), tal como mostra a
Tabela 7.
De acordo com os resultados obtidos, o tamanho das RVs amplificadas varia entre 1000 e
6000 bp, e o número de genes cassette que constitui cada região varia entre 1 e 6, o que vem
comprovar o baixo número de genes cassette encontrados em integrões de classe 1.
É de realçar alguns casos, em que o mesmo arranjo de genes cassette foi encontrado em
espécies diferentes, o que pode mostrar que estes integrões de classe 1 provavelmente estão
associados a plasmídeos conjugativos e por isso podemos estar perante uma transferência
horizontal de genes de resistência aos antibióticos.
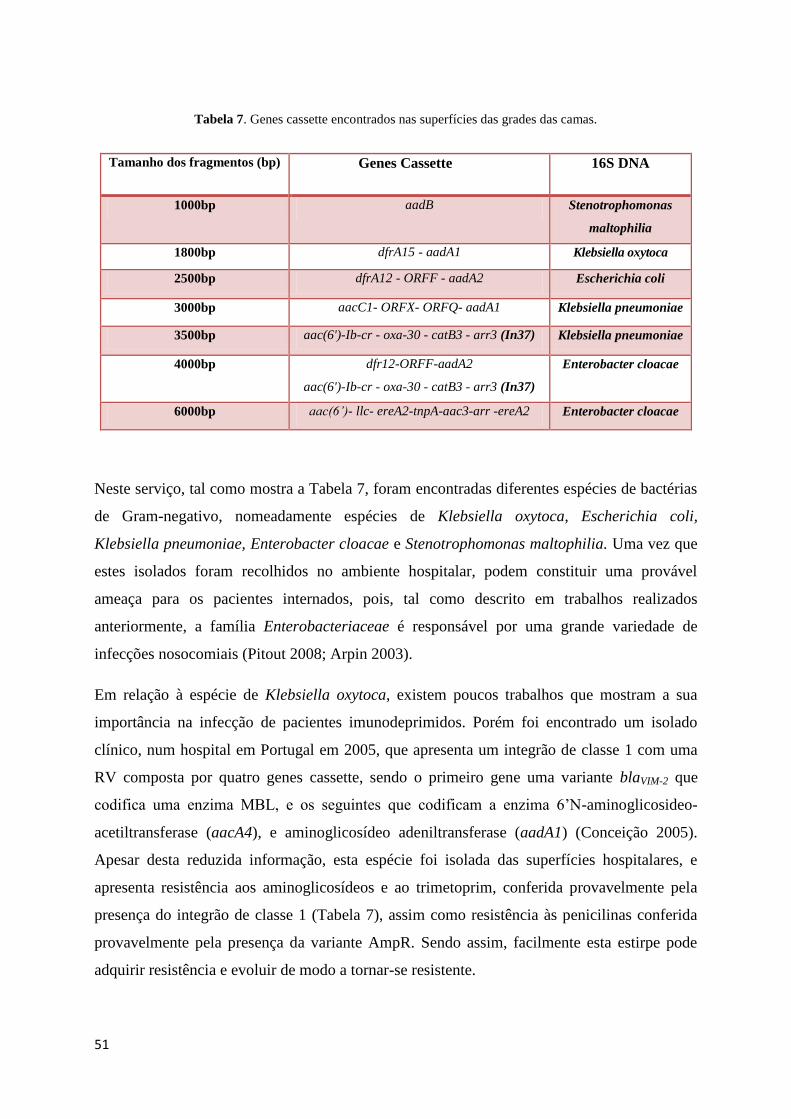
51
Tabela 7. Genes cassette encontrados nas superfícies das grades das camas.
Tamanho dos fragmentos (bp)
Genes Cassette 16S DNA
1000bp aadB Stenotrophomonas
maltophilia
1800bp dfrA15 - aadA1 Klebsiella oxytoca
2500bp dfrA12 - ORFF - aadA2 Escherichia coli
3000bp aacC1- ORFX- ORFQ- aadA1 Klebsiella pneumoniae
3500bp aac(6')-Ib-cr - oxa-30 - catB3 - arr3 (In37) Klebsiella pneumoniae
4000bp dfr12-ORFF-aadA2
aac(6')-Ib-cr - oxa-30 - catB3 - arr3 (In37)
Enterobacter cloacae
6000bp aac(6’)- llc- ereA2-tnpA-aac3-arr -ereA2 Enterobacter cloacae
Neste serviço, tal como mostra a Tabela 7, foram encontradas diferentes espécies de bactérias
de Gram-negativo, nomeadamente espécies de Klebsiella oxytoca, Escherichia coli,
Klebsiella pneumoniae, Enterobacter cloacae e Stenotrophomonas maltophilia. Uma vez que
estes isolados foram recolhidos no ambiente hospitalar, podem constituir uma provável
ameaça para os pacientes internados, pois, tal como descrito em trabalhos realizados
anteriormente, a família Enterobacteriaceae é responsável por uma grande variedade de
infecções nosocomiais (Pitout 2008; Arpin 2003).
Em relação à espécie de Klebsiella oxytoca, existem poucos trabalhos que mostram a sua
importância na infecção de pacientes imunodeprimidos. Porém foi encontrado um isolado
clínico, num hospital em Portugal em 2005, que apresenta um integrão de classe 1 com uma
RV composta por quatro genes cassette, sendo o primeiro gene uma variante blaVIM-2 que
codifica uma enzima MBL, e os seguintes que codificam a enzima 6’N-aminoglicosideo-
acetiltransferase (aacA4), e aminoglicosídeo adeniltransferase (aadA1) (Conceição 2005).
Apesar desta reduzida informação, esta espécie foi isolada das superfícies hospitalares, e
apresenta resistência aos aminoglicosídeos e ao trimetoprim, conferida provavelmente pela
presença do integrão de classe 1 (Tabela 7), assim como resistência às penicilinas conferida
provavelmente pela presença da variante AmpR. Sendo assim, facilmente esta estirpe pode
adquirir resistência e evoluir de modo a tornar-se resistente.

52
No caso da espécie E. cloacae, tem sido descrita uma situação diferente mas preocupante,
uma vez que na maior parte das vezes, esta bactéria para além de expressar o gene blaESBL,
também pode expressar uma enzima β-lactamase do tipo AmpC indutíveis e/ou adquiridas, o
que dificulta a detecção da produção de ESBLs e o controle de disseminação (Jacoby 2009).
Neste trabalho, foi também descrito pela primeira vez em todo o Mundo, numa estirpe de
Enterobacter cloacae, o integrão de classe 1 In37. No entanto, este arranjo já foi descrito
anteriormente na Argentina e na China, respectivamente em espécies bacterianas como K.
pneumoniae e E.coli. Nestes isolados, o integrão In37 surge associado a uma sequência de
inserção, o elemento ISCR1, que estabelece a ligação entre este arranjo e o conjunto de genes
de resistência presentes a montante, neste caso o gene qnrB6, constituindo assim um integrão
complexo. Este trabalho também mostrou que o In37 se encontra associado a um plasmídeo
(Quiroga, 2007; Rodríguez-Matínez 2007).
Tendo por base outros trabalhos , e no seguimento do presente trabalho, foi igualmente
pesquisada a presença do elemento ISCR1, o qual tem uma importante função na mobilização
de genes de resistência por um mecanismo designado “rolling cycle transposition” (Toleman
2006). Assim, após a amplificação do respectivo gene ISCR1, verificou-se a sua presença, e
por isso estamos igualmente perante um integrão complexo, uma vez que a montante surge
uma segunda RV, neste caso composta pelo gene qnrS1 (Toleman 2006).
Estudos realizados por Wu e seus colaboradores (Wu 2007), descreveram igualmente um
integrão complexo In37 na China, numa estirpe de E. coli, onde estava presente pela primeira
vez ambos os genes qnrA e qnrB no mesmo plasmídeo, tal como a variante blaDHA-1, uma
AmpC β-lactamase que confere resistência a cefamicina e cefalosporina de amplo espectro
(Wu 2007;Kim 2009; Pitout 2008). O gene qnr tem sido encontrado em plasmídeos de
tamanhos que variam de 54 a 180kb, aproximadamente. No contexto genético do
determinante qnrA é caracterizado pela presença de uma estrutura inicialmente descrita por
Hall e Strokes (1990), conhecida com “ integrão complexo do tipo sul1” (Hall e Stokes 1990).
A frequência dos isolados produtores de ESBL varia muito de região para região, sendo mais
alta na América Latina. Como exemplo podemos citar que a estirpe K. pneumoniae produtora
de ESBL nos Estados Unidos não ultrapassa 5%, na Europa varia entre 20-25% e na América
Latina é de 35-40%. O SENTRY (programa de monitoramento de patógenos e resistência em

53
infecções hospitalares e adquiridas na comunidade) mostra que na América do Sul a E.coli e
K. pneumoniae são os agentes mais prevalentes das Enterobacteriaceas nos isolados de 2002
seguidos pelo Enterobacter (Fedler 2006).
As estirpes de Enterobacter são consideradas como patógenos emergentes nas infecções
hospitalares, ocupando o terceiro lugar em todo o Mundo, sendo mais prevalentes nas
infecções do tracto urinário, tecidos moles e septicemias (Fedler 2006).
Estudos anteriores realizados em diferentes países sobre integrões classe 1 provenientes de
E.coli, mostraram a presença de RV com diferentes arranjos de genes cassette, nomeadamente
genes que conferem resistência aos aminoglicosídeos (aadA1, aadA2, aadA5, aadB, aaC4),
cloranfenicol (cmla, catB8), trimetoprim (dfrA1, dfrA5, dfrA7, dfrA12, dfrA17), lincosamida
(linf) e alguns genes desconhecidos (orfD, OrfF). O tipo mais comum de gene cassette
presente em E. coli é o gene aadAseguido pelo gene dfrA ( eputien 2010; Chang 2006, Lee
2001).
8.1.3 Presença de enzimas β-lactamases no SMII
Nos últimos 20 anos, inúmeros antibióticos β-lactâmicos têm sido desenvolvidos com o
propósito principal de combater a acção hidrolítica das enzimas β-lactamases. A utilização
indiscriminada e inadequada dos antibióticos também tem favorecido a selecção e evolução
de bactérias produtoras de ESBLs e de MBLs, nomeadamente bactérias da família
Enterobacteriaceae. Uma vez que estas enzimas normalmente surgem associadas a
plasmídeos, as bactérias produtoras destas enzimas são as grandes responsáveis pela
ocorrência e disseminação mundial de resistência aos antibióticos β-lactâmicos,
principalmente no ambiente hospitalar, dentro do qual cada vez mais têm surgido novas
variantes destas enzimas (Peña 1998; Coque, 2008).
Após a realização do screening para estudar a presença de diferentes genes que codificam
enzimas β-lactamases nos 11 isolados positivos para o gene intI1, destaca-se a prevalência das
enzimas β-lactamases do tipo TEM 29% (n=4) e OXA 26% (n=4), seguidos do tipo SHV 21%
(n=3), CTX-M 14% (n=2) e VIM 7% (n=1) (Figura 11). Neste trabalho a variante IMP não foi
encontrada em nenhum dos isolados. Estes resultados vem uma vez mais mostrar que as
enzimas ESBL são cada vez mais prevalentes na família Enterobacteriacea, provavelmente

54
devido à sua localização plasmídica, tendo sido inicialmente derivadas da variante TEM-1,
TEM-2 ou SHV-1 (Arpin 2003).
Figura 11: Percentagem de enzimas β-lactamases presentes nos isolados que apresentavam integrão classe 1.
Estudos realizados anteriormente em Portugal demonstraram a prevalência das enzimas CTX-
M-15, TEM-1 e OXA-30 em isolados da E.coli (Machado 2006; Mendonça 2006). Neste
trabalho, e após a sequenciação dos respectivos genes, foi possível detectar igualmente a
presença da variante TEM-1 e CTX-M-15 em estirpes de E. coli, mas o mesmo não aconteceu
com a variante OXA-30, a qual surgir apenas associada ao integrão In37, em estirpes de K.
pneumoniae e E. cloacae.
A emergência de enzimas β-lactamases torna-se preocupante, uma vez que se trata de
bactérias comensais (E. coli, K. pneumoniae e E. cloacae), que, principalmente devido à
localização plasmídica destas enzimas, rapidamente se tornam resistentes e muitas vezes
patogénicas, podendo estar na origem de surtos hospitalares.
Verificou-se que a associação dos genes que codificam a enzima blaOXA-30 com o gene
aac(6’)-Ib-cr, que codifica uma enzima capaz de inactivar a acção de duas classes de
antibióticos em simultâneo, os aminoglicosídeos e as fluoquinolonas, tendo demonstrado que
aqueles genes eram disseminados num plasmídeo comum (Machado 2006; Mendonça 2006).
29%
21% 14%
29%
7%
TEM
SHV
CTX-M
OXA
VIM

55
As usuais formas de transmissão que incluem a disseminação clonal de estirpes produtoras de
ESBL, ou a disseminação através de genes produtores de ESBLs carregados por plasmídeos e
que são transmitidos entre diferentes géneros de Enterobactérias (Rice 1993).
Enzimas como SHV, TEM e CTX-M são apontadas como responsáveis pelo aumento da
resistência às penicilinas e cefalosporinas de espectro alargado, pois conferem resistência a
praticamente todos os antibióticos β-lactâmicos, excepto carbapenemos e cefamicinas, que
são inibidos por outras β-lactamases (Coque 2008).
O género Klebsiella é um dos que prevalece no ambiente clínico, e é uma espécie muito
virulenta entre os isolados clínicos, e frequentemente responsáveis por graves infecções
hospitalares e surtos. Sabe-se porém que esta espécie produz uma variedade de enzimas β-
lactamases plasmídicas, o que permite uma rápida disseminação destas enzimas. O
internamento prolongado no ambiente hospitalar pode levar a aquisição de graves infecções
por estirpes de K. pneumoniae portadoras de ESBL (Mulvey 2009;Livermore 1995).
Um outro estudo realizado em 1996 num hospital na Holanda em enfermarias de neurologia e
neurocirurgia, mostrou a incidência de multiresistência a vários antibióticos em
Enterobacteriacea, em que 28% dos pacientes foram infectados ou colonizados por Klebsiella
oxytoca, estirpes resistentes a pelo menos três classes de antibióticos incluindo o trimetoprim-
sulfamethoxazol, mostrando o papel importante dos integrões classe 1 no transporte de genes
de resistência (Leverstein-van Hall 2002).
8.2 - Serviço de Medicina Intensiva (SMI)
8.2.1 Tipagem Molecular
Inicialmente, o objectivo principal deste trabalho era detectar a presença da espécie
Acinetobacter baumannii nas superfícies e equipamentos hospitalares do SMI, devido às
graves infecções causadas por esta espécie. Para além de tentar detectar o foco de infecção
desta espécie, este serviço foi escolhido por ser um dos mais críticos, uma vez que é composto
por pacientes imunodeprimidos, e bastante expostos à aquisição de possíveis infecções
nosocomiais. Assim, para além do estado de saúde debilitado e de um tempo de internamento
maior, estes pacientes estão constantemente sujeitos a procedimentos invasivos, tais como
cateteres intravenoso, sondas vesicais e ventilação mecânica, e por isso estão na presença de

56
uma elevada pressão selectiva de antibióticos, o que os expõe ainda mais às bactérias que
colonizam esses equipamentos.
Os isolados do SMI, foram recolhidos num total de 34 superfícies e equipamentos
hospitalares: 20 superfícies (59%) não apresentavam crescimento de colónias bacterianas em
meio selectivo para bactérias de Gram-negativo (MacConkey agar), e por isso, apenas 14
superfícies (41%) apresentavam crescimento bacteriano neste mesmo meio. Perante este
resultado, podemos inferir que uma parte das superfícies e equipamentos hospitalares deste
serviço não estavam colonizadas por bactérias de Gram-negativo, mas sim por outro tipo de
microrganismo, uma vez que em meio TSB ocorreu crescimento.
Na continuação deste trabalho, apenas foram consideradas as superfícies e equipamentos
hospitalares colonizados por bactérias de Gram-negativo, sendo as restantes descartadas. Uma
vez que foram analisadas várias superfícies, foi obtida uma população bacteriana bastante
numerosa, pelo que é necessário recorrer a técnicas moleculares de tipagem genética, de modo
a reduzir a população em estudo. Neste caso, e após a análise de todos os perfis genéticos, foi
possível concluir que estávamos perante uma espécie bacteriana, pois o grau de similaridade
entre os diferentes perfis era muito elevado. Sendo assim, e após a sequenciação do gene 16S
rDNA, concluiu-se ser a espécie E coli. Sendo este um microrganismo comensal, colonizador
da flora intestinal, pode facilmente desenvolver / adquirir resistência e factores de virulência,
devido ao ambiente selectivo em que se encontra, podendo tornar-se uma espécie igualmente
preocupante (Branger 2005).
Com tudo isto, é possível concluir que entre os isolados bacterianos de Gram-negativo
recolhidos no ambiente hospitalar deste serviço, não foi encontrada a espécie A.baumannii.
A elevada prevalência de resistência entre isolados clínicos da E.coli encontradas num estudo
de 1995 e 2001 parece ser um sinal alarmante de que os antibióticos necessitam ser utilizados
de forma adequada, especialmente em infecções do trato urinário e intra-abdominal, onde
mostra um aumento na taxa de bacteremia por E.coli. Peña e seus colaboradores (1995),
referem que era pouco comum a resistência a fluoroquinolonas em E. coli, mas que com o uso
frequente da ciprofloxacina e norfloxacina para tratamento de pacientes com neutropenia e
cirrose, houve um aumento da resistência nestas estirpes (Peña 1995; Cheong 2001).

57
Dentre as enterobactérias, a E.coli tem sido a espécie mais prevalente em infecções
hospitalares em todo o mundo ocupando o segundo lugar depois da S.aureus. Contudo, na
América Latina, a E.coli ocupa o primeiro lugar na lista dos patógenos mais frequentes em
infecções hospitalares principalmente associada à infecções do tracto urinário (Fedler 2006).
Geralmente os pacientes internados nos cuidados intensivos estão mais expostos a factores de
risco, nomeadamente a bactérias patogénicas multiresistentes, tais como Acinetobacter spp.,
E.coli, Pseudomonas aeruginosa, Staphylococcus aureus (Lópencoz-Pueyo 2010).
Podem ser citados alguns estudos que mostraram a elevada incidência de infecções graves e
surtos no SMI, causados por bactérias de Gram-positivo, sendo a maioria desencadeadas por
S. epidermidis e S. aureus, que fazem parte da flora normal da pele. Essas bactérias cada vez
mais tornam se resistente à linezolida, devido o aumento do uso abusivo da linezolida e
posterior disseminação clonal (Kelly 2008; López-Pueyo 2010). Um outro estudo no SMI no
Brasil mostra a elevada prevalência de pneumonias nosocomiais associadas com ventilação
mecânica, sendo que grande parte destas pneumonias são desencadeadas por P.aeruginosa
(52,0%) resistente a imipenemo, e (65,4%) por S.aureus resistente à oxacilina (Rocha 2008).
Mulvey et al (2009) relata ainda que actualmente a taxa de surtos causados por
Staphylococcus aureus resistente a meticilina (MRSA) são cada vez maiores em todo o
mundo. MRSA é considerado um dos patogénios responsáveis por graves infecções
principalmente em tracto respiratório e corrente sanguínea e afectam principalmente pacientes
com internamento prolongado (Mulvey 2009).
Ainda demonstrando que infecções e surtos causados por bactérias de Gram-positivos estão
cada vez mais presentes nos cuidados intensivos, um outro estudo realizado em Espanha
(García et al. 2010) relata um surto causado por Staphylococcus aureus resistentes a
linezolida (LRSA). A linezolida é um dos antibióticos de amplo espectro utilizado para
tratamento de várias infecções nos serviços intensivos bem como para as infecções causadas
MRSA. Porém devido a pressão selectiva exercida sobre á linezolida, as estirpes estão mais
resistente e a opção terapêutica para essas infecções está cada vez mais difícil e limitada. O
autor relata que o surto causado pelo gene cfr pode ser encontrado em algumas espécies como
o E.aureus e E. epidermidis. Sugere ainda que esse gene possui algumas vantagens de
sobrevivência, não somente pelo uso inadequado da linezolida, mas também a transferência

58
por plasmídeo a partir de um microrganismo como Staphylococcus coagulase-negativa.
(Sánchez García 2010).
8.2.2 Caracterização da Região Variável
Tal como descrito no ponto anterior, neste serviço, apenas foram encontrados isolados da
espécie E. coli, os quais foram o objecto de estudo desta segunda parte do trabalho.
Inicialmente for realizado o screening para estudar a presença do gene intI1, e nos isolados
positivos para este gene foi caracterizada a respectiva RV, a qual apresentava
aproximadamente com o mesmo tamanho (cerca de 2000 bp) em todos os isolados. Sendo
assim, e de uma forma rápida, foi realizada a digestão destes fragmentos com a enzima TaqI,
a qual revelou um perfil de digestão igual em todos os fragmentos, mostrando que estamos
perante a mesma RV. Assim, apenas um destes fragmentos foi sequenciado, e tal como
resume a Tabela 8, a RV destes isolados é constituída por dois genes cassette, um que confere
resistência ao trimetoprim (dfrA17), e um outro que confere resistência aos aminoglicosídeos
(aadA5).
Tabela 8. Genes cassette encontrados nas superfícies hospitalares do serviço de medicina intensiva.
Tamanho dos fragmentos (bp)
Genes Cassettes 16S DNA
2000 bp dfrA17 – aadA5 Escherichia coli
Este arranjo genético tem sido descrito por todo o mundo em várias espécies de bactérias de
Gram-negativo, tais como Shigella spp., K. pneumoniae, e P. aeruginosa, mas é na espécie de
E.coli que se verifica uma elevada prevalência deste integrão de classe 1 (Lee 2001; Chang
2007).
Espécies bacterianas resistentes ao trimetoprim e aos aminoglicosídeos, têm sido as mais
encontradas em todo o mundo, uma vez que estes antibióticos foram e continuam a ser uma
das opções muito utilizadas, e a presença destes genes em integrões, normalmente associados
a plasmídeos favorece a disseminação horizontal destes genes ( eputien 2010).

59
Chang et al (2007) demonstra em seu estudo comparativo um aumento da prevalência do
integrão classe 1 em E.coli tanto em Taiwan como em outros países na Europa. Em Taiwan
devido a pressão selectiva exercida sobre uso de antibióticos na veterinária, se tornou alta a
taxa de genes que confere resistência aos antibióticos como ampicilina, gentamicina,
cefalotina e sulfametaxasol/ trimetoprim nos isolados clínicos. Neste estudo mostra a
estabilidade e as mudanças do integrão classe 1 num período de 10 anos. A presença do gene
confere resistência aos aminoglicosídeos (aadA5) e trimetoprim (dfrA17) no integrão de
classe 1 na E.coli é muito comum, tanto em humanos como em animais. Porém acreditava-se
que com a diminuição do uso clínico do aminoglicosídeo e trimetoprim, a presença desses
genes diminuiriam gradualmente. A pesquisa revela que a disseminação pode estar associada
aos plasmídeos em humanos ou animais em diferentes áreas, sendo o papel do integrão
essencial para essa disseminação (Chang 2007).
Ainda um estudo na Coreia investigou o gene dfr e sua elevada prevalência no integrão de
classe 1 em E.coli nos isolados urinários resistentes ao trimetoprim. A pressão selectiva e os
elementos genéticos móveis como as plasmídeos, transposões e integrões contribuíram para
uma geração de mutantes DHFR (inibidores da diidrofolato redutase) menos sensíveis ao
trimetoprim. Na investigação a taxa de resistência do trimetoprim nos isolados de E. coli nas
infecções urinárias foi de 63.1%. Essa elevada taxa de resistência ao trimetoprim e outros
antibióticos se tornou um grande problema devido ao uso indiscriminado do antibiótico. A
prevalência maior foi dos genes dfrA17 e dfrA12, e os demais dfrA1, dfrA2 e dfrA5 também
foram detectados, sendo que a transferência é feita por plasmídeos. Na investigação mostra
que os genes dfrA1,dfrA12 e dfrA17 foram transferidos 31 dos 72 isolados em E.coli
resistentes ao trimetoprim, sendo os elementos genéticos móveis um mecanismo importante
na propagação dos genes. Em outros países, o gene dfrA1e dfrA14 são os mais prevalentes
nos isolados de infecção urinária. O estudo ainda sugere que o gene dfr inseridos no integrão
pode ter implicações importantes na divulgação adicional da resistência ao trimetoprim (Lee
2001).
Tenover (2006) relata ainda que E.coli pode causar graves infecções, resistente a diversos
antibióticos entre os quais os aminopenicilinas como amoxicilinas, ampicilinas e
cefalosporinas de baixo espectro. Onde essa resistência pode ser adquirida por plasmídeos e
codificada por enzimas β-lactamases do tipo TEM-1, TEM-2, ou SHV que hidrolisam e

60
inactivam esses antibióticos. Ainda algumas estirpes de E.coli podem desenvolver resistência
às cefalosporinas de 3ª geração e aos monobactâmicos (aztreonam), como resultado das
mutações de enzimas tipo TEM, SHV e CTX-M.
A elevada taxa de resistência aos antibióticos, associadas a diversos factores de virulência,
converte a família Enterobacteriacea, nomeadamente isolados de E. coli e de K. pneumoniae,
num problema muito relevante no ambiente hospitalar, uma vez que é responsável por uma
grande variedade de infecções nosocomiais desde infecções urinárias, gastrointestinais,
pneumonias e septicemias (Coque 2008).

61
CONCLUSÃO

62
9. Conclusão
Perante os resultados obtidos nos dois serviços hospitalares, é possível concluir que:
- o ambiente hospitalar está colonizado por vários microrganismos, entre os quais
bactérias de Gram-negativo, comensais e oportunistas, capazes de desenvolver e adquirir
resistência aos antibióticos e por isso capazes de causar infecções nosocomiais em pacientes
imunodeprimidos.
- nos isolados recolhidos foram encontrados diferentes genes de resistência, tais como
genes que conferem resistência aos aminoglicosídeos, trimetoprim, cloranfenicol, rifampicina
e eritromicina associados a integrões de classe 1, mas também foram amplificados genes que
conferem resistência aos antibióticos β-lactâmicos, tais como enzimas β-lactamases, as quais
estão normalmente associadas a plasmídeos e por vezes associadas a integrões de classe 1.
- dos integrões de classe 1 encontrados, a maioria já se encontra descrita contudo, o
integrão de classe 1 In37 foi descrito no presente trabalho, pela primeira vez, numa espécie de
Enterobacter cloacae.
- foram encontrados os mesmos arranjos genéticos em espécies diferentes, o que pode
ser o resultado da transmissão horizontal de genes, uma vez que os integrões normalmente
estão associados a elementos genéticos móveis, principalmente plasmídeos conjugativos.
Perante estes resultados constata-se que é preocupante a sobrevivência e a disseminação
bacteriana em superfícies e equipamentos hospitalares. Estes isolados podem adquirir genes
de resistência a antibióticos e tornar-se multiresistentes, não só devido à possível ocorrência
de transferência horizontal de genes, mas também devido à elevada pressão selectiva
antibiótica presente no ambiente hospitalar.
Assim, o ambiente hospitalar pode ser definido como um reservatório de bactérias comensais
e oportunistas, na maior parte das vezes multirresistentes e até patogénicas, e como um meio
de disseminação horizontal de genes de resistência. Torna-se por isso necessário controlar e
monitorizar o ambiente hospitalar, com o intuito de impedir a disseminação de bactérias para
os pacientes, bem como travar a evolução do perfil de resistência bacteriana aos antibióticos,
o que por sua vez irá reduzir a taxa de mortalidade causada por diversas infecções
nosocomiais.

63
10. Referências Bibliográficas
Alanis, A. J. (2005) Resistance to Antibiotics: Are we in the Post-Antibiotic Era? Archives of
Medical Research. 36: 697-705.
Alonso, A.; Sánchez, P.; Martínez, J. (2001) Environmental Selection of antibiotic resistance
genes. Environmental Microbiology. 3(1): 1-9.
Aminov, R. (2009) The role of antibiotics and antibiotic resistance in nature. Environmental
Microbiology. 11(12): 2970-2988.
Ardanuy, C.; Liñares, J.; Domínguez, M.; Hernández-Allés, S.; Benedí, V.; Martínez-
Martínez, L. (1998) Outer Membrane Profiles of Clonally Related Klebsiella pneumoniae
Isolates from Clinical and Activities of Cephalosporins and Carbapenems. Antimicrobial
Agents and Chemotherapy. 42(7):1636-1640.
Arpin, C.; et al. (2003) Extended- Spectrum β-lactamase- Producing Enterobacteriaceae in
Community and Private Health Care Center. Antimicrobial Agents and Chemotherapy. 47
(11): 3506-3514.
Ash, R.; Mauck, B.; Morgan M. (2002) Antibiotic Resistance of Gram-negative Bacteria in
Rivers, United States. Emerging Infectious Diseases. 8(7) 713-716.
Bahl, M.; Hansen, L.; Goesmann, A.; Sorensen, S. (2007) The multiple antibiotic resistance
IncP-1 Plasmid pKJK5 Isolated from soil environment is phylogenetically divergent from
members of the previously established β and sub-groups. Plasmid. 58: 31-43.
Barlow, R.; Pemberton, J. (2004) Isolation and Characterization of Integron-Containing
Bacteria without Antibiotic Selection. Antimicrobial Agents Chemotherapy. 48(3): 838-842.

64
Bennett, P. (2004) Genomic plasticity: Insertion Sequence Elements, Transposons and
Integrons and DNA Rearrangement. Methods in Molecular Biology. 266: 71-113.
Bennett, P. (2008) Plasmid encoded antibiotic resistance: acquisition and transfer of antibiotic
resistance genes in bacterial. British Journal of Pharmacology. 153: S347- S357.
Bernards, A.; Harinck et al.(2004) Persistent Acinetobacter baumannii? Look inside your
medical equipament. Infection Control and Hospital Epidemiologists of America. 25(11):
1002-1004.
Branger, C.; et al. (2005) Genetic Background of Escherichia coli and Extended-spectrum β-
lactamics type. Emerging Infectious Diseases. 11(1): 54- 61.
Brown, K.; Kulis, J.; Thompson, B.; Champ, T.; Mawhinney, D. (2006) Occurrence of
antibiotics in hospital, residential, and dairy effluent, municipal wastewater, and the Rio
Grande in New México. Science of the Total Environment. 366: 772-783.
Bush, K.; Jacob, G.; Medeiros, A. (1995) A functional Classification Scheme for β-lactamase
and Its Correlation With Molecular Structure. Antimicrobial Agents Chemotherapy. 1211-
1233.
Cambray, G.; Guerout, A.; Mazel, D. (2010) Integrons. The Annual Review of Genetics.
44(1): 141-1465.
Carattoli, A. (2001) Importance of Integrons in the Diffusion of Resistance. Veterinary
Research. 32(3-4): 243-259.
Ceccarelli, D.; Colombo, M. (2007) Role, Circulation and Molecular Characterization of
Integrons and ICEs in Clinical and Environmental Vibrio. Communicating Current Research
and Education Topics and Trends in Applied Microbiology. 220-229.

65
Chang, L.; Chang, T.; Chang, C. (2007) Variable Gene Cassette Patterns of Class 1 Integron-
associated Drug-resistant Escherichia coli in Taiwan. The Kaohsiung Journal of Medical
Science. 23(6): 273 - 280.
Cheong H-L; Yoo C-W; Sohn W-J; et al.(2001) Bacteremia Due to Quinolone-Resistant
Escherichia coli in a Teaching Hospital in South Korea. Clinical Infectious Diseases.33 48-
53.
Chevalier. (2008) Identification and evolution of drug efflux pump clinical Enterobacter
aerogenes strains isolated in 1995 and 2003. PLoS One. 3(9): 320.
Chroma, M.; Kolar, M. (2010) Genetic Methods for Detection of Antibiotic Resistance: Focus
on Extended-Spectrum β-Lactamases. Biomedical Paper of the Medical Faculty of the
University Palacky Olomouc Czechoslovakia Repub. 154(4): 289-296.
Clewell, D. (2005) Antibiotic Resistance Plasmids in Bacteria. Encyclopedia of Life Sciences.
Coates, A.; Hu, Y. (2007) Novel approaches to developing new antibiotics for bacterial
infections. Britishi Journal of Pharmacology. 152: 1147-1154.
Coenen, S.; Muller, A.; Monnet, D.; et al. (2007) European Surveillance of Antimicrobial
Consumption (ESAC): outpatient antibiotic use in Europe. Quality and Safety in Health Care.
16(6): 440-445.
Collis, C.; Hall, R. (1995) Expression of Antibiotic Resistance Genes in the Integrated
Cassettes of Integrons. Antimicrobiol Agents and Chemotherapy. 39(1): 155-162.
Collis, C.; Kim, M.; Stokes, H.; Hall, R. (2002a) Integron-encoded intI integrases
preferentially recognize the adjacent cognate attI site in recombination with a 59-be site.
Molecular Microbiology. 46(5): 1415-1427.

66
Collis, C.; Kim, M.; Partridge, S.; Stokes, H.; Hall, R. (2002b) Characterization of the Class 3
Integron and the Site-Specific Recombination System it Determines. Journal of Bacteriology.
184(11): 3017-3026.
Conceição, T.; Brízio, A.; Duarte, A. (2005) First Isolation of blaVIM-2 in Klebsiella oxytoca
Clinical Isolates from Portugal. Antimicrobial Agents and Chemotherapy. 49(1): 476.
Coque, T.; Baquero, F.; Cantón, R. (2008) Increase Prevalence of ESBL-producing
Enterobacteriaceae in Europe. Eurosurveillance. 13(47): 1-11.
Cornaglia, G.; Russell, K.; Satta, G.; Fontana, R. (1995) Relative Importance of Outer
Membrane Permeability and Group 1 β-Lactamase as Determinants of Meropenem and
Imipinem Activities against Enterobacter cloacae. Antimicrobial Agents and Chemothepary.
39(2): 350-355.
Costa, S.; Woodcock, J.; Gil, M.; Wise, R.; Barone, A.; Caiaffa, H.; Levin, A. (1999) Outer-
Membrane Proteins Pattern and detection of β-lactamase in clinical isolates of imipenem-
resistance Acinetobacter baumannii from Brazil, International Journal of Antimicrobial
Agents. 13: 175-182.
Fedler, K.; Biedenbach, D.; Jones, R. (2006) Assessment of pathogen frequency and
resistance patterns among pediatric patient isolates: report from the 2004 SENTRY
Antimicrobial Surveillance Program on 3 continents. Diagnostic Microbiology Infectious
Disease. 56(4): 427-436.
Fluit, A.; Schmitz, F. (1999) Class 1 Integrons, Gene Cassette, Mobility and Epidemiology.
European Journal of Clinical Microbiology & Infectious Diseases. 18(11): 761-770.
Fluit, A.; Schmitz, F. (2004) Resistance Integrons and Super-integrons. European Journal of
Clinical Microbiology & Infectious Diseases. 10(4): 272-288.

67
Furuya, E.; Lowy, F. (2006) Antimicrobial-resistant bacteria in the community setting. Nature
Medicine. 4: 36-45.
Gillings, M.; Boucher, Y.; Labbate, M.; Holmes, A.; Krishnan, S.; Holley, M.; Stokes, H.
(2008) The Evolution of de class 1 Integrons and Rise of Antibiotic Resistance. Journal of
Bacteriology. 190(14): 5095-5100.
Gonzáles, G.; Mella, S.; Zemelman, R.; Bello, H.; Domíngez, M. (2004) Integrones y
Cassettes Genéticos de Resistencia: Estructura y rol frente a los antibacterianos. Revista
Médica do Chile. 132 (5): 619-626.
Guentzel, M. (1996) Escherichia, Klebsiella, Enterobacter, Serratia, Citrobacter, and
Proteus. Medical Microbiology. 4th
Edition Chapter 26.
Hall, R.; Stokes, H. (1990) The structure of a partial duplication in the integron of plasmid
pDGO100. Plasmid. 23(1): 76-79.
Hall, R.; Collis, C. (1995) Mobile Gene Cassettes and Integrons: capture and spread of genes
by site-specific recombination. Molecular Microbiology. 15(4): 593-600.
Hancock, R. (2007) The complexities of antibiotic action. Molecular Systems Biology. 3:
142.
Harbottle, H.; Thakur, S.; Zhao, S.; White, D. (2006) Genetics of Antimicrobial resistance.
Animal Biotechnology. 17(2): 111–124.
Henriques, I.; Fonseca, F.; et al. (2006) Occurrence and diversity of integrons and β-
lactamase genes among ampicillin-resistant isolates from estuarine waters. Research in
Microbiology. 157(10): 938-947.
Jacoby, G. (2009) AmpC beta-Lactamase. Clinical Microbiology Reviews. 22(1): 161-182.

68
Jeong, S.; Lee, K.; Chong, Y.; Yum, J.; Lee, S.; Choi, H.; Kim, J.; Park, K.; Han, B.; Lee, S.;
Jenog, T. (2003) Characterization of a new integron containing VIM-2, a metallo-β-lactamase
gene cassette, in clinical isolate of Enterobacter cloacae. Journal of Antimicrobial
Chemoterapy. 51: 397-400.
Keller, R.; Pedroso, M.; Ritchmann, R.; Silva, R. (1998) Occurrence of Virulence-Associated
Properties in Enterobacter cloacae. American Society for Microbiology. 66(2): 645-649.
Kelly, S.; Collins, J.; Maguire, M.; Gowing, C.; Flanagan, M.; Donnelly, M.; Murphy, P.
(2008) An outbreak of colonization with linezolid-resistant Staphylococcus epidermidis in an
intensive therapy unit. Journal of Antimicrobial Chemotherapy. 61: 901-907.
Kelly, B.; Verpermann, A.; Bolton, D. (2009) The Role of Horizontal Gene Transfer in the
evolution of selected food borne Bacterial pathogens. Food and Chemical Toxicology. 47:
951-968.
Kim, S.; Park, Y.; Yu, J.; Kim, Y.; Han, K. (2009) Prevalence and characteristics of aac(6’)-
Ib-cr in AmpC-producing Enterobacter cloacae, Citrobacter freundii, and Serratia
marcences: a multicenter study from Korea. Diagnostic Microbiology and Infectious Disease.
63: 314-318.
Lambert-Zechovsky, N.; Bingen, E.; Denamur, E.; Brahimi, N.; Brun, P.; Mathieu, H.; Elion,
J. (1992) Molecular Analysis Provides Evidence for the Endogenous Origin of Bacteremia
and Meningitis Due to Enterobacter cloacae in an Infant. Clinical Infectious Diseases. 15(1):
30-32.
Lee, C.; Oh, J.; Cho, J.; Park, J.; Kim, J.; Seol, S.; ChoD, T. (2001) The prevalence of
Trimethoprim-resistance-conferring di-hydrofolate reductase genes in urinary isolates of
Escherichia coli in Korea. Journal of Antimicrobial Chemotherapy. 47: 599-604.

69
Lemaire, S.; Fuda, C.; Bambeke, F.; Tulkens, P.; Mobashery, S. (2008) Restoration of
Susceptive of Methicillin-resistant Staphilococcus aureus to β-Lactam Antibiotics by Acidic
pH. The Journal of Biological Chemistry. 238(19): 12769-12776.
Leverstein-van Hall, M.; Box, A.; Blok, H.; Paaw, A.; Fluit, A.; Verhoef, J. (2002) Evidence
of Extensive Interspecies Transfer of Integron-Mediated Antimicrobial Resistance Genes
among Multidrug-Resistant Enterobacteriaceae in Clinic Setting. The Journal of Infectious
Disease. 186: 49-56.
Lévesque, C.; Piché, L.; Larose, C.; Roy P. (1995) PCR Mapping of Integrons Reveals
Several Novel Combinations of Resistance Genes. Antimicrobial Agents and Chemotherapy.
39(1): 185-191.
Li, X. (2005) Quinolone Resistance in Bacteria: emphasis on plasmid-mediated mechanism.
International Journal of Antimicrobial Agents. 25(6): 453-463.
Lim, K.; Yasin, R.; Yeo, C.; Puthucheary, S.; Thong, K. (2009) Characterization of Multidrug
Resistant ESBL-Producing Escherichia coli Isolates From Hospitals Malaysia. Journal of
Biomedicine and Biotechnology.
Lima, N.; Mota M. (2003) Biotecnologia Fundamentos e Aplicações. Lidel. 120-133.
Livermore, D. (1995) β-Lactamases in Laboratory and Clinical resistance. Clinical
Microbiology. 8: 557-585.
Livermore, D. (2002) Multiple Mechanism of Antimicrobial Resistance in Pseudomonas
aeruginosa: Our worst Nightmare? Clinical Infectious Diseases. 34: 634-640.
Livermore, D. (2003) Bacterial Resistance: Origins, Epidemiology and Impact. Clinical
Infectious Diseases. 36(1): S11-23.

Livermore, D.; Cantón, R.; Gniadkowski, M.; Nordmann, P.; Rossolini, G.; Arlet, G.; Ayala,
J.; Coque, T.; Ken-Zdanowicz, I.; Luzzaro, F.; Poriel, L.; Woodford, N. (2007) CTX-M:
changing the face of ESBLs in Europe. Journal of Antimicrobial Chemotherapy. 59: 165-174.
López-Pueyo, M.; Bercellina-Gaite, F.; Amaya-Villar, R.; Guarnacho-Montero, J. (2010)
Multiresistência antibiótica em unidades críticos. Medicina Intensiva. 35(1): 41-53.
Machado, E.; Coque, T.; Souza, J.; Peixe L. (2006) Dissemination in Portugal of CTX-M-15,
OXA-1 and TEM-1 Producing Enterobacteriaceae Strains Containing the aac(6’)-Ib-cr gene,
which encodes an aminoglycoside and fluoroquinolone modifying enzyme. Antimicrobial
Agents and Chemotherapy. 50(9): 3220-3221.
Mahamoud, A.; Chevalier, J.; Alibert-Franco, S.; Kern, W.; Pàges, J. (2007) Antibiotic Efflux
pumps in Gram-negative Bacteria: the Inhibitor response strategy. Journal of Antimicrobial
Chemotherapy. 59: 1223-1229.
Martínez, J.; Martínez, L.; Rosenblueth, M.; Silva, J.; Martínez-Romero, E. (2004) How are
Genes Sequence Analyses Modifying Bacterial Taxonomy? The case of Klebsiella.
International Microbiology. 7: 267-268.
Martinez, J. (2009a) The role of natural environments in the evolution of resistance traits in
pathogenic bacteria. Proceedings of the Royal Society. 276: 2521-2530.
Martinez, J.; Sánchez, M.; Solano, M.; Hernandez, A.; Garmendia, L.; Fajardo, A.; Ortega, C.
(2009b) Functional role bacterial multidrug efflux in microbial natural ecosystems. FEMS
Microbiology Reviews. 33(2): 430- 49.
Mazel, D. (2006) Integrons: Agents of bacterial evolution. Nature Reviews Microbiology.
4(8): 608-20.

Mendonça, N.; Louro, D.; Castro, A.; Diogo, J.; Caniça, M. (2006) CTX-M-15, OXA-30 and
TEM-1-producing Escherichia coli in two Portuguese regions. Journal of Antimicrobial
Chemotherapy. 57: 1014-1016.
Michael, C.; Gillings, M.; Holmes, A.; Hughes, L.; Andrew, N.; Holley, M.; Stokes, H.
(2004) Mobile Genes Cassettes: A Fundamental Resource for Bacterial Evolution. The
American Naturalist. 164(1).
Mindlin, S.; Petrova, M.; Bass, I.; Gorlenko, Z. (2006) Origin, Evolution, and Migration of
Drug Resistance Genes. Russian Journal of Genetics. 42(11): 1495-1511.
Mohamudha, P.; Harish, B.; Parija, S. (2010) AmpC Beta-Lactamases Among Gram-negative
Clinical Isolates From a Tertiary Hospital, South India. Brazilian journal of Microbiology. 41:
596-602.
Moreira, M.; Souza, E.; Moraes, C. (2004) Multidrug Efflux Systems in Gram-negative
Bacteria. Brazil Journal of Microbiology. 35: 19-28.
Mulvey, M.; Bryce, E.; Boyd, D.; et al. (2004) Ambler class A Extended-Spectrum Beta-
Lactamase-producing Escherichia coli and Klebsiella spp. in Canadian Hospitals.
Antimicrobial Agents and Chemotherapy. 48(4): 1204-1214.
Mulvey, M.; Simor, A. (2009) Antimicrobial resistance in hospitals: How concerned should
we be? Canadian Medical Association Journal. 180(4): 408-415.
Murray, P.; Rosenthal, K.; Pfaller, M. (2009) Medical Microbiology. 6th
MOSBY pp. 303-
307.
Nikaido, H.; Vaara, M. (2003) Molecular Basis of Bacterial Outer Membrane Permeability.
Revisited Microbiology and Molecular Biology. American Society for Microbiology. 67: 593-
656.

72
Normark, B.; Normark, S. (2002) Evolution and Spread of Antibiotic Resistance. Journal of
Internal Medicine. 252(2): 91-106.
Nouér, S.; Nucci, M.; Oliveira, M.; Piffano, F.; Pellegrino, C.; Moreira B. (2005) Risk factor
for acquisition of multidrug resistant Pseudomonas aeruginosa producing SPM metallo-beta-
lactamase. Antimicrobial Agents Chemotherapy. 3663-3667.
Novais, A.; Baquero, F.; Machado, E.; Cantón, R.; Peixe, L.; Coque, T. (2010) International
Spread and Persistence of TEM-24 is Caused by the confluence of highly penetration
Enterobacteriaceae clones and an IncA/C2 plasmid containing Tn1696::Tn1 and IS5075-
Tn21. Antimicrobial Agents and Chemotherapy. 54(2): 825-834.
Obi, C.; Green, E.; Bessong, P.; Villiers, B.; Hoosen, A.; Igumbor, E.; Potgieter, N. (2004)
Gene encoding virulence Gene encoding virulence makers among Escherichia coli isolates
from diarrheic stool samples and river sources in rural Venda communities of South Africa.
Water SA. 30(1): 37-42.
Partridge, S.; Brown, H.; Hall, R. (2002) Characterization and Movement of the Class 1
Integron Known as Tn2521 and Tn1405. Antimicrobial Agents and Chemotherapy. 46(5):
1288-1294.
Peña, C.; Albareda, J.; Pallares, R.; Pujol, M.; Tubau, F.; Ariza, J. (1995) Relationship
between Quinolone Use and Emergence of Ciprofloxacin-resistant Escherichia coli in
Bloodstream Infections. Antimicrobial Agents and Chemotherapy. 39(2): 520-524.
Peña, C.; Pujol, M.; Ardanuy, C.; Ricart, A.; Pallares, R.; Ariza, J; Gudiol, F. (1998)
Epidemiology and successful control of a large outbreak due to Klebsiella pneumoniae
producing extended-spectrum beta-lactamases. Antimicrobial Agents and Chemotherapy.
42(1): 53-58.

73
Peng, C.; Lee, M.; Fu, H.; Chen, Y.; Hsu, C. (2007) Characterization of Class 1 Integrons and
Antimicrobial Resistance in CTX-M-3 Producing Serratia marcencens Isolates from Southern
Taiwan. Japanese Journal of Infectious Diseases. 60: 250-256.
Peters, E.; Leverstein-van, M.; Box, A.; Verhoef, J.; Fluit, A. (2001) Novel Gene Cassettes
and Integrons. Antimicrobial Agents and Chemotherapy. 45(10): 2961-2964.
Pfaller, M.; Jones, R.; Doern, G.; Kugler, K. (1998) Bacterial Pathogens Isolated from
Patients with Bloodstream Infection: Frequencies of Occurrence and Antimicrobial
Susceptibility Patterns from SENTRY antimicrobial Surveillance Program (United States and
Canada, 1997). Antimicrobial Agents and Chemotherapy. 42(7): 1762-1770.
Pitout, J.; Church, D.; Gregson, D. (2008) Surveillance for plasmid-mediated quinolone
resistance in Enterobacteriaceae within the Calgary Heath Region, Canada: the emergence of
aac(6’)-Ib-cr. Journal of Antimicrobial Chemotherapy. 61: 999-1002.
Podschun, R.; Ullman, U. (1998) Klebsiella spp. as nosocomial pathogens: epidemiology,
taxonomy, typing methods and pathogenicity factors. Clinical Microbiology Review. 11(4):
589-603.
Poirel, L; Nordmann, P. (2006) Carbapenem resistance in Acinetobacter baumannii:
mechanisms and epidemiology. European Journal Clinical Microbiology Infectious Disease.
12(9): 826-836.
Poole, K. (2001) Multidrug Efflux Pumps and Antimicrobial Resistance in Pseudomonas
aeruginosa and Related Organisms. Journal Molecular Microbiology Biotechnology. 3(2):
255-264.
Poole, K. (2004) Efflux-mediated multiresistance in Gram-negative Bacteria. Clinical
Microbiol Infection. 10: 12-26.

74
Recchia, G.; Hall, R. (1997) Origins of the Mobile Gene Cassettes Found in Integrons. Trends
in Microbiology. 5(10): 389-394.
Rice, L.; Carias, L.; Etter, L.; Shales, D. (1993) Resistance to Cefoperazone-Sulbactam in
Klebsiella pneumoniae. Evidence for enhanced resistance resulting from the coexistence for
two different resistance mechanisms. Antimicrobial Agents and Chemotherapy. 37(5): 1061-
1065.
Rocha, L.; Vilela, C.; Cezário, R.; Almeida, A.; Filho, P. (2008) Ventilator-Associated
Pneumonia in an Adult Clinical-Surgical Intensive Care Unit of a Brazilian University
Hospital: Incidence, Risk Factors, Etiology, and Antibiotic Resistance. The Brazilian Journal
of Infectious Diseases. 12(1): 80-85.
Rodriguéz-Martínez, J.; Velasco, C.; García, I.; Cano, M.; Martínez-Martínez, L.; Pascual, A.
(2007) Characterization of integrons containing the Plasmid-mediated
quinolone resistance gene qnrA1 in Klebsiella pneumoniae. International Journal of
Antimicrobial Agents. 29: 705-709.
Sabaté, M.; Prats, G. (2002) Estructura y Funcion de los Integrons. Enfermedades Infecciosas
y Microbiología clínica. 20(7): 341-345.
Sáenz, Y. (2004) Caracterización fenotípica e genotípica de la resistência a antibióticos en
cepas de Escherichia coli nos patógenos de alimentos y de la microflora intestinal de
humanos y animales. Dissertação de Doutorado na área de Bioquímica e Biologia Molecular.
Universidade de La Rioja.
Sánchez, M.; Torres, M.; Morales, G.; et al. (2010) Clinical Outbreak of Linezolid-Resistant
Staphylococcus aureus in an Intensive Care Unit. Journal of American Medical Association.
303(22): 2260-2264.

75
Sahly, H.; Aucken, H.; Benedi, V.; Forestien, C.; Fussing, V.; Hansen, D. (2004) Impairment
of Respiratory burst in polymorphonuclear leukocytes by ESBL strains of Klebsiella
pneumoniae. European Journal Clinical Microbiology Infectious Disease. 23: 20-26.
Santos, C.; Caetano, T.; Ferreira, S. and Mendo, S. (2010a). A novel complex class 1 integron
found in a Klebsiella pneumoniae isolate from Portugal. Clinical Microbiology and Infection.
17(7): 1036-1039.
Santos, C.; Caetano, T.; Ferreira, S. and Mendo, S. (2010b). First description of blaIMP-8 in a
Pseudomonas mendocina isolated at the Hospital Infante D. Pedro, Aveiro, Portugal.
Research in Microbiology. 161(4): 305-307.
Santos, C.; Caetano, T.; Ferreira, S. and Mendo, S. (2010c). Tn5090-like class 1 integron
carrying blaVIM-2 in a Pseudomonas putida strain from Portugal. Clinical Microbiology and
Infection. 16(10): 1558-1561.
Sekar, V.; Santiago, T.; Vijayan, K.; Alavandi, S.; Stalin, V.; Rajan, J.; Sanjuktha, M.;
Kalaimani, N. (2008) Involvement of Enterobacter cloacae in the mortality of the fish, mogul
cephalous. Letters in Applied Microbiology. 1-6.
Senka, D., u kovi ., Kos B. (2008) Antibiotic Resistance Mechanisms in Bacteria:
Biochemical and Genetic Aspects. Food Technol. Biotechnol. 46(1): 11-21.
eputien , V.; Povilonis, .; Ru auskas, M.; Pavilonis, A.; Su ied liene, E. (2010) Prevalence
of Trimethoprim resistance genes in Escherichia coli isolates of human and animal origin in
Lithuania. Journal of Medical Microbiology. 59: 315-322.
Siegel, R. (2008) Emerging Gram-negative Antibiotic Resistance: Daunting Challenges,
Declining sensitivities, and Dire Consequences. Respiratory Care. 53(4): 471-479.

76
Soto, S.; Lobato, M.; Mendoza, M. (2003) Class 1 Integron-Borne Gene Cassettes in
Multidrug-Resistant Yersinia enterocolitica Strains of Different Phenotypic and Genetic
Types. Antimicrobiol Agents and Chemotherapy. 47(1): 421-425.

77