
Efeito do Resveratrol sobre a produção de ROS, Óxido ... - RAQUEL... · INSTITUTO DE ENSINO E...
48
INSTITUTO DE ENSINO E PESQUISA DA SANTA CASA DE BELO HORIZONTE Núcleo de Pós Graduação e Pesquisa Efeito do Resveratrol sobre a produção de ROS, Óxido Nítrico e VEGF por leucócitos de pacientes diabéticos tipo 2. RAQUEL MIRANDA GONZAGA Belo Horizonte Fevereiro/2013
Transcript of Efeito do Resveratrol sobre a produção de ROS, Óxido ... - RAQUEL... · INSTITUTO DE ENSINO E...

1
INSTITUTO DE ENSINO E PESQUISA DA SANTA CASA DE BELO HORIZONTE
Núcleo de Pós Graduação e Pesquisa
Efeito do Resveratrol sobre a produção de ROS, Óxido Nítrico e VEGF
por leucócitos de pacientes diabéticos tipo 2.
RAQUEL MIRANDA GONZAGA
Belo Horizonte
Fevereiro/2013

2
RAQUEL MIRANDA GONZAGA
Dissertação apresentada ao programa de Pós
Graduação do Instituto de Ensino e Pesquisa da Santa
Casa de Belo Horizonte, como requisito parcial para
obtenção do título de mestre em biomedicina.
Orientador: Dr. José Augusto Nogueira Machado
Belo Horizonte
Fevereiro/2013

3
AGRADECIMENTOS
Ao Dr. José Augusto Nogueira Machado pelo carinho, paciência, cuidado, generosidade em
compartilhar tanto conhecimento e por todas as oportunidades de aprendizado e crescimento
profissional.
Aos órgãos financiadores CAPES, FAPEMIG, CNPq.
À todos os professores da Pós Graduação do IEP por todos os ensinamentos.
Aos participantes da banca examinadora, por aceitarem o convite.
À todos os funcionários do IEP que me ajudaram em algum momento. Em especial à Zélia, Mariana e
Shirley pela disponibilidade e alegria.
Às coletoras do CEM pelo auxílio e disposição.
Aos doadores diabéticos e não diabéticos pela contribuição importantíssima em meu trabalho.
À todos os alunos dos laboratórios vizinhos pela ajuda empréstimos e disponibilidade.
À Fabi e Níbia pelas conversas e risadas.
Às minhas companheiras de laboratório e eternas amigas: Polly, Nanda, Paulinha, Carol, Luana e Elisa
por toda a ajuda com meus experimentos, pela disponibilidade, amizade e momentos muito especiais.
Ao Espaço de Danças Thalita Menezes, em especial à minha professora Jéssica por me proporcionar
momentos incríveis e mágicos com a dança, pelo apoio e incentivo!
Á Dayanne, Raquel Arcanjo, Aytube, Aline Morena, Gody e Mamy, Kátia, Carol, Drí, Hellen e Fabiana,
pelo apoio e torcida.
Ao meu amado Quinteto (Érika, Andrews e Raquel) pelo amor, preocupação, encontros inesquecíveis e
cuidado de verdadeiros irmãos.
À minha amada Vovó Inês pelas orações e apoio.
À todos os meus familiares que torceram por mim e acompanharam essa etapa da minha vida.
À minha segunda família (Cleusa, Antônio, Preto, Karla e meus pequenos Arthur e Yasmin) pelo apoio
e amor.
Ao meu Juninho por todo carinho, amor, amizade, compreensão e incentivo. Obrigada por me fazer
sorrir e sempre acreditar em mim! Amo você!
Aos meus pais, Eni e Luíz, meu maior exemplo, por mais uma vez se privarem de tudo, para me ajudar
a estudar, pelo amor incondicional, apoio e torcida. Vocês são meu ar, minha inspiração! Amo vocês !
À Deus, amor maior e fonte de todo conhecimento, por me capacitar e me surpreender a cada dia com
sua bondade e misericórdia.

4
“Talvez não tenha conseguido fazer o melhor,
Mas, lutei para que o melhor fosse feito.
Não sou o que deveria ser, mas graças a Deus,
Não sou o que era antes.”
Marthin Luther King

5
RESUMO
No presente trabalho avaliamos o efeito do Resveratrol em granulócitos e mononucleares
(PBMNC) de pacientes diabéticos tipo 2 (DM2) em comparação com seus controles não diabéticos
(ND). Foram estudadas as respostas oxidantes, com a dosagem de ROS e RNS, fagocitose de
partículas opsonizadas, modulação da via da Proteína Cinase C (PKC) e a produção de VEGF (fator de
crescimento endotelial) na presença deste polifenol. A produção de ROS foi avaliada com o teste de
quimiluminescência. A dosagem de óxido nítrico foi quantificada pela Reação de Griess e o VEGF foi
analisado através do método de ELISA. Nossos resultados demonstraram que na presença de
Resveratrol a produção de ROS foi inibida em granulócitos, significativamente (p<0,05) tanto em
diabéticos do tipo 2 como em não diabéticos. Resultado semelhante foi observado durante o processo
fagocítico. O Resveratrol inibe a produção de óxido nítrico de forma mais evidente em não diabéticos. A
diferença entre indivíduos diabéticos e não diabéticos foi significativa (p<0,05). O mecanismo sugere o
envolvimento da inibição da PKC pelo Resveratrol. Esta inibição foi significativa, sendo de 66% para
pacientes diabéticos e 77% em não diabéticos, quando comparado com um inibidor seletivo de PKC
(Calphostin C) que apresentou inibição de 51% para não diabéticos e 40% para diabéticos do tipo 2. A
produção de VEGF por mononucleares na presença de Resveratrol não foi alterada nos grupos
diabéticos e não diabéticos. Nossos resultados, no seu conjunto, demonstram que a presença do
Resveratrol é capaz de interferir em células da imunidade inata tanto em pacientes diabéticos tipo 2
quanto em indivíduos não diabéticos.
Palavras Chave: diabetes, granulócitos, mononucleares, fagocitose, oxido nítrico, VEGF e Resveratrol.

6
LISTA DE ESQUEMAS
Esquema 1: Alteração das vias de sinalização celular pela hiperglicemia.
Esquema 2: Representação esquemática das fases indutora e efetora no Diabetes tipo 2 .
Esquema 3: Processo de formação do Óxido Nítrico (NO) mediado pela enzima NO-sintase.
Esquema 4: Propriedades do Resveratrol no organismo humano.
Esquema 5: Absorção do Resveratrol pelo organismo humano.
Esquema 6: .Ação do Resveratrol no processo inflamatório.
Esquema 7: Obtenção de Leucócitos pela técnica de Ficoll-Hypaque.
Esquema 8: Modulação do Resveratrol pela PKC, reduzindo a produção de ROS e evitando um possível estresse oxidativo.

7
LISTA DE FIGURAS
Figura 1: Estrutura Química do Resveratrol (3,4’,5-trihidroxiestilbeno).
Figura 2: Mecanismo de ativação da PKC e possível modulação pelo Resveratrol, demonstrando seu
efeito no processo de estresse oxidativo.
Figura 3: Avaliação da produção de ROS em granulócitos de diabéticos tipo 2 e não diabéticos.
Figura 4: Avaliação da atividade fagocítica de granulócitos estimulados com partículas de zimosan
opsonizado com fragmentos C3b.

8
LISTA DE TABELAS
Tabela 1: Espécies Reativas de Oxigênio.
Tabela 2: Reagentes necessários para solução tamponada pH: 7,3.
Tabela 3: Características clínicas e bioquímicas da população em estudo.
Tabela 4: Modulação da PKC pelo RSV.
Tabela 5: Modulação da produção de NO pelo RSV.
Tabela 6: Produção de VEGF na presença de Resveratrol.

9
LISTA DE ABREVIATURAS
AGES = produtos finais de glicosilação avançada
CALPHOSTIN C = Cladosporium cladosporioides
CEP = comitê de ética em pesquisa
cm = centímetros
COX-2 = Ciclo-oxigenase-2
DAG = diacilglicerol
DM = diabetes melitus
DM1 = diabéticos tipo 1
DM2 = diabéticos tipo 2
DMSO = dimetilsulfóxido
E/C = experimento/controle
EP = erro padrão
GMPc = guanosina monofosfato cíclico
HbA1c = hemoglobina glicada
IFN-ɣ = interferon gama
IL-12 = interleucina 12
IL-2 = interleucina 2
IL-4 = interleucina 4
IMC = índice de massa corporal
IкB = inibidor do NF-kB
KH2PO4 = fosfato de potássio monobásico
L-NAME = Nω-Nitro-L-arginina cloridrato de Ester metílico
n = número de amostras
NA = não se aplica
Na2HPO4 = fosfato de sódio bibásico anidro
NaCl = cloreto de sódio
NADPH = nicotamida adenina dinucleotídeo fosfato
NF-кB = fator nuclear kappa B
NO = óxido nítrico

10
NOS = óxido nítrico sintase
NS = não significativo
PBS = tampão fosfato salino
PCR = proteína C reativa
PDB = Éster Forbol 12,13-dibutirato
PKC = proteína cinase c
q.s.p. = quantidade suficiente para
RAGE = receptor para AGE
RLU/min = unidades relativas de luz por minuto
ROS = espécies reativas de oxigênio
RSV = Resveratrol
SCM-BH = Santa Casa de Misericórdia de Belo Horizonte
TCLE = termo de consentimento livre e esclarecido
TNF-α = fator de necrose tumoral alfa
VEGF = fator de crescimento vascular endotelial

11
Sumário
1 Introdução ...............................................................................................................................12
2 Pacientes e Métodos ...............................................................................................................20
2.1 Seleção de Paciente ................................................................................................................20
2.2 Coleta de Sangue ...................................................................................................................20
2.2.1 Separação Celular: obtenção de Leucócitos ..................................................................................20
2.3 Reagentes ..............................................................................................................................22
2.4 Opsonização de Zimosan .......................................................................................................23
2.5 Teste de Viabilidade Celular: Incorporação de Azul de Trypan ..............................................23
2.6 Ensaio de quimioluminescência dependente de luminol: quantificação das espécies reativas
de oxigênio (ROS) ..................................................................................................................................24
2.6.1 Teste de Quimioluminescência ...............................................................................................24
2.6.2 Atividade Fagocítica ................................................................................................................24
2.6.3 Inibição da PKC pelo Resveratrol ............................................................................................25
2.6.4 Produção de Oxido Nítrico ......................................................................................................25
2.7 Análise da Expressão de VEGF ..............................................................................................26
2.8 Equipamentos .........................................................................................................................26
2.9 Análise Estatística ...................................................................................................................26
3 Resultados ..............................................................................................................................27
4 Discussão ................................................................................................................................39
5 Conclusão ...............................................................................................................................43
6 Sumário de Resultados ...........................................................................................................44
Referências Bibliográficas ......................................................................................................................45

12
1 Introdução
O Diabetes Mellitus (DM) é uma patologia multifatorial caracterizada pela falha no metabolismo
da glicose causada por falta ou por defeitos nos receptores de insulina [1]. Existem dois principais tipos
de Diabetes. O Diabetes Tipo 1 (DM1) ocorre pela destruição das células β pancreáticas, acomete
principalmente crianças e adolescentes levando a deficiência completa na produção de insulina e o
Diabetes Tipo 2 (DM2) ocorre, geralmente, por um defeito na secreção da insulina, podendo estar
associado a idade, sedentarismo, obesidade, hábitos alimentares, fatores genéticos, influências
ambientais e outras condições associadas a inflamação crônica e infecção [2]. Atualmente o DM2 atinge
também pessoas mais jovens devido ao aumento da obesidade na infância [1, 3].
Dentre os sintomas do DM pode-se destacar poliúria, polidipsia e perda de peso [4]. Estudos
apontam que em longo prazo são estabelecidas complicações características da patologia como
retinopatia, nefropatia, neuropatia e cardiopatia [5]. Todas essas complicações podem potencializar o
impacto gerado pela doença na população. O número de diabéticos tem aumentado devido ao
crescimento populacional, urbanização, obesidade e falta de atividade física. Dados do IBGE-2010
demonstram que cerca de 6% dos brasileiros são diabéticos, segundo a OMS (Organização Mundial de
Saúde) aproximadamente 10% da população do mundo possui a doença. Estudos sugerem que em
2030 essa taxa pode dobrar sendo que aproximadamente 12% da população será diabética no país[6-7].
A deficiência na produção ou a resistência à insulina dão origem à hiperglicemia que, por sua
vez, acarreta diversas complicações características da patologia. A junção de todos esses fatores
associados à hiperglicemia pode ativar e/ou alterar diversas vias de sinalização, levando a inflamação,
causando disfunção endotelial, culminando nas complicações micro e macrovasculares características
desta doença [8].
Estudos demonstram que o Diabetes está relacionado com o estresse oxidativo, condição
caracterizada pelo aumento da resposta oxidante, devido à produção de espécies reativas de oxigênio
(ROS), sem o aumento equivalente da resposta antioxidante. No Diabetes, o estresse oxidativo atua na
indução de complicações micro e macrovasculares[9]. A tabela 1 apresenta alguns exemplos de
espécies reativas de oxigênio (ROS):

13
Tabela 1: Espécies Reativas de Oxigênio
Radicais Não Radicais
Superóxido .O-2 Peróxido de Hidrogênio H2O2
Hidroxila OH. Ácido Hipocloroso HClO
Peroxila LO.2 Ozônio O3
Alkoxila LO. Oxigênio “singlet”
Hidroxiperoxila HO.2 Peróxidos Lipídicos
Fonte: Darley-Usmar e cols., 1995 [10]
O estresse oxidativo induzido pela hiperglicemia ocorre devido a ativação de vias de
sinalização celular, as quais podemos destacar: o aumento do fluxo da via dos polióis, o aumento da
formação de AGES, a ativação da Proteína Cinase C (PKC) e o aumento do fluxo da via das
hexosaminas, levando a disfunção e dano celular (Esquema 1) [11].
Esquema 1: Alteração das vias de sinalização celular pela Hiperglicemia.
A hiperglicemia altera diversas vias de sinalização, levando a ativação de moléculas sinalizadoras que por sua vez, podem
levar a alterações na função de proteínas ou até mesmo na expressão gênica, causando diversos danos e disfunções
celulares que contribuem para as complicações diabéticas.
Adaptado de King L.G. 2008[2]
No Diabetes, a ativação da via Diacilglicerol/PKC (DAG/PKC) pode modular várias funções
vasculares como a permeabilidade, contratilidade e também influenciar na produção de espécies
reativas de oxigênio (ROS)[7]. A ativação da PKC exerce importante função na patogênese da doença.
A PKC é uma serina – treonina responsável pela transdução de sinais, atuando em estímulos

14
neuronais, hormonais e fatores de crescimento[12]. No diabetes sua ativação ocorre devido ao aumento
de Diacilglicerol (DAG) estimulado pela hiperglicemia. A estimulação da via DAG/PKC pode modular
várias funções vasculares como a permeabilidade, contratilidade e também influenciar na produção de
ROS. A síntese de DAG e ativação da PKC ativam a NADPH oxidase induzindo a produção de ROS,
levando a Fase Indutora (Fase I) que por sua vez, ativa a Fase Efetora (Fase II). Essa ultima é
caracterizada pela formação de AGES (Produtos Finais de Glicosilação Avançada) por oxidases ou
rotas não enzimáticas e pela interação entre AGES e seu receptor (RAGE), formando o complexo AGE-
RAGE, que levam a ativação celular, culminando em um processo inflamatório, como descrito por
Nogueira-Machado, JA & Chaves, M.M.(Esquema 2) [1-2, 13-14].
Esquema 2: Representação esquemática das fases indutora e efetora no Diabetes tipo 2.
Adaptado de: Nogueira-Machado, J.A. & Chaves, M.M.,2008
A hiperglicemia também altera os níveis de Óxido Nítrico (NO), devido ao aumento da
produção de ROS ocorre uma redução dos níveis do mesmo. O NO é formado por uma reação
catalisada por enzimas entre o oxigênio molecular e a L-arginina podendo ter efeito benéfico ou
maléfico para o organismo. (Esquema 3). As enzimas NO-sintases (NOS) são importantes para a
biossíntese do NO[15]. Existem três isoformas, duas delas consideradas formas constitutivas, presentes
no endotélio e nos neurônios: eNOS e nNOS e a forma induzível, que pode ser expressa em
macrófagos, neutrófilos e também em células endoteliais: iNOS [16-17].

15
Esquema 3. Processo de formação do Oxido Nítrico (NO) mediado pela enzima NO-sintase.
Outro fator alterado pela patologia do Diabetes é o VEGF (Vascular endothelial growth factor)
que é um fator de crescimento responsável pela vasculogênese e angiogênese[18]. A angiogênese é o
processo de formação de novos vasos por células endoteliais que, por sua vez, revestem a parede do
vaso sanguíneo. Esse processo é dependente de reguladores que podem ser fatores de crescimento
ou citocinas. Dentre os fatores de crescimento, o VEGF possui destaque por ser um forte estimulador
deste processo.
As alterações nesses mecanismos levaram a busca por compostos capazes de auxiliar na
prevenção ou controle das complicações diabéticas que ocorrem ao longo da doença. Dentre esses
compostos podemos destacar o Resveratrol (RSV).
O Resveratrol (3,4’,5-trihidroxiestilbeno) foi identificado primeiramente em 1940 nas raízes de
Verartum grandiflorum por Takaoka[6]. O uso deste composto com função medicinal foi descrito
primeiramente no Tratado Indiano Ayurveda (2.500 A.C), no qual o suco da uva (Vitis vinifera L.) era
um componente importante do “darakchasava”, um suco fermentado conhecido por suas propriedades
cardiotônicas[19].
O principal impulso para a pesquisa com o Resveratrol veio da observação paradoxal de que
uma baixa incidência de doenças cardiovasculares pode coexistir com a ingestão de dieta rica em
gordura e consumo moderado de vinho tinto (150-300mL/dia), um fenômeno conhecido como Paradoxo
Francês[20].
O Resveratrol é um polifenol da classe dos Estilbenos presente em uma ampla variedade de
plantas, incluindo amora, amendoim, eucalipto e a casca da uva (Vitis vinifera), sua principal fonte, com
diversas atividades biológicas que incluem propriedades antioxidantes, anti-inflamatórias e
antitumorais[7]. É um composto não flavonóide derivado da fenilalanina, contém dois anéis aromáticos
com hidroxilas reativas em sua estrutura, que pode se apresentar em duas isoformas: cis e trans-
resveratrol. (Figura 1). A isomerização ocorre devido à ação da luz e a isoforma trans é a principal
responsável pelo efeito do Resveratrol em mamíferos[21].
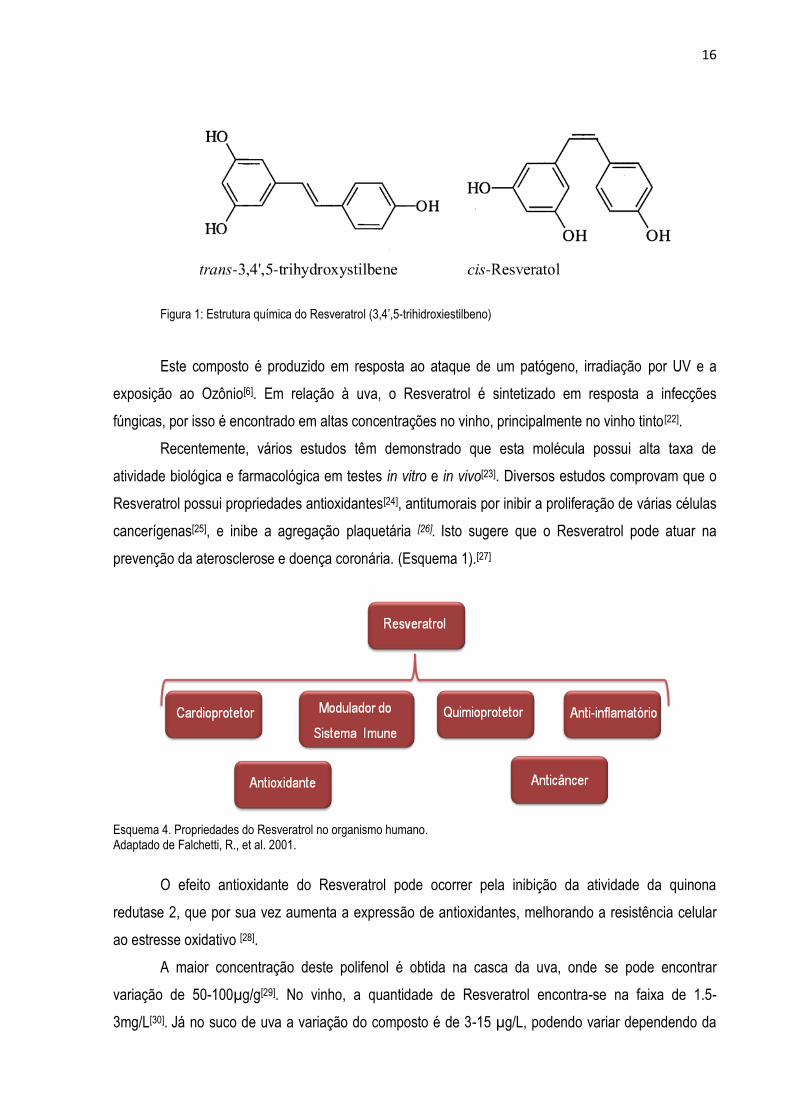
16
Figura 1: Estrutura química do Resveratrol (3,4’,5-trihidroxiestilbeno)
Este composto é produzido em resposta ao ataque de um patógeno, irradiação por UV e a
exposição ao Ozônio[6]. Em relação à uva, o Resveratrol é sintetizado em resposta a infecções
fúngicas, por isso é encontrado em altas concentrações no vinho, principalmente no vinho tinto [22].
Recentemente, vários estudos têm demonstrado que esta molécula possui alta taxa de
atividade biológica e farmacológica em testes in vitro e in vivo[23]. Diversos estudos comprovam que o
Resveratrol possui propriedades antioxidantes[24], antitumorais por inibir a proliferação de várias células
cancerígenas[25], e inibe a agregação plaquetária [26]. Isto sugere que o Resveratrol pode atuar na
prevenção da aterosclerose e doença coronária. (Esquema 1).[27]
Esquema 4. Propriedades do Resveratrol no organismo humano. Adaptado de Falchetti, R., et al. 2001.
O efeito antioxidante do Resveratrol pode ocorrer pela inibição da atividade da quinona
redutase 2, que por sua vez aumenta a expressão de antioxidantes, melhorando a resistência celular
ao estresse oxidativo [28].
A maior concentração deste polifenol é obtida na casca da uva, onde se pode encontrar
variação de 50-100µg/g[29]. No vinho, a quantidade de Resveratrol encontra-se na faixa de 1.5-
3mg/L[30]. Já no suco de uva a variação do composto é de 3-15 µg/L, podendo variar dependendo da
17
espécie de uva[21]. Segundo alguns estudos, a quantidade necessária de RSV diária para se obter
todos os seus efeitos benéficos deve ser de aproximadamente 20mg[8]. Logo, para que isso seja
possível sugere-se que a melhor maneira de ingestão do composto deve ser por cápsulas.
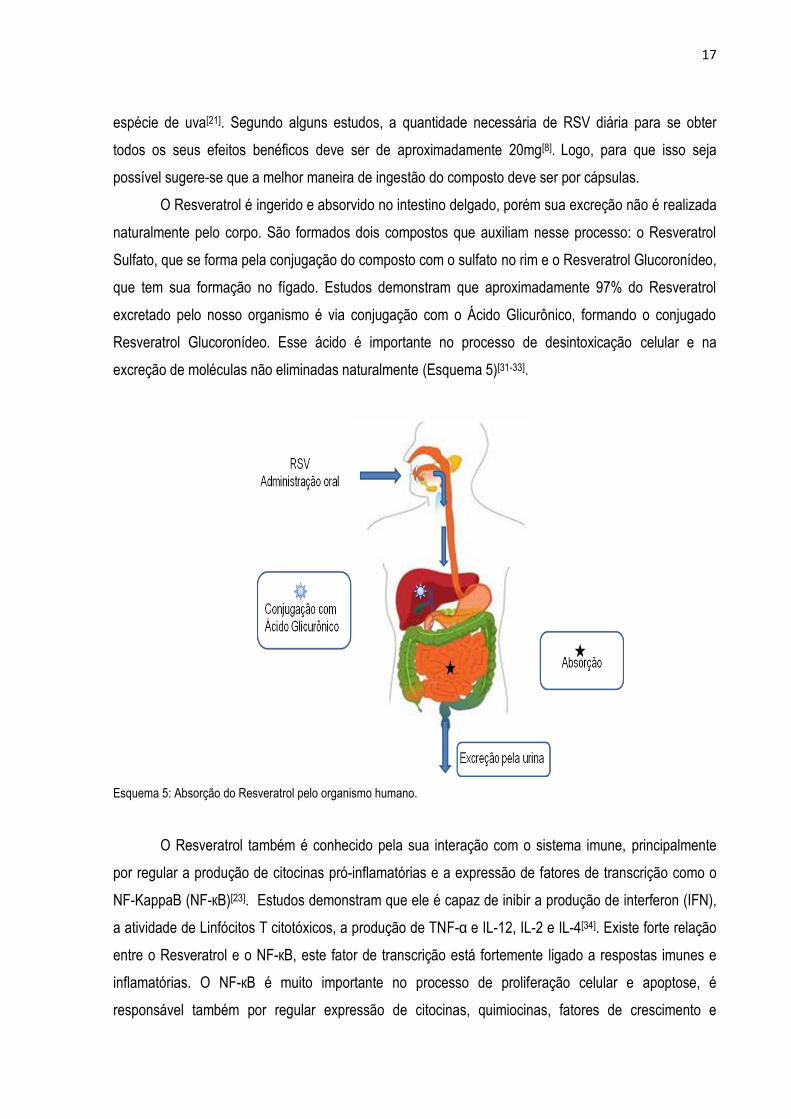
O Resveratrol é ingerido e absorvido no intestino delgado, porém sua excreção não é realizada
naturalmente pelo corpo. São formados dois compostos que auxiliam nesse processo: o Resveratrol
Sulfato, que se forma pela conjugação do composto com o sulfato no rim e o Resveratrol Glucoronídeo,
que tem sua formação no fígado. Estudos demonstram que aproximadamente 97% do Resveratrol
excretado pelo nosso organismo é via conjugação com o Ácido Glicurônico, formando o conjugado
Resveratrol Glucoronídeo. Esse ácido é importante no processo de desintoxicação celular e na
excreção de moléculas não eliminadas naturalmente (Esquema 5)[31-33].
Esquema 5: Absorção do Resveratrol pelo organismo humano.
O Resveratrol também é conhecido pela sua interação com o sistema imune, principalmente
por regular a produção de citocinas pró-inflamatórias e a expressão de fatores de transcrição como o
NF-KappaB (NF-кB)[23]. Estudos demonstram que ele é capaz de inibir a produção de interferon (IFN),
a atividade de Linfócitos T citotóxicos, a produção de TNF-α e IL-12, IL-2 e IL-4[34]. Existe forte relação
entre o Resveratrol e o NF-кB, este fator de transcrição está fortemente ligado a respostas imunes e
inflamatórias. O NF-кB é muito importante no processo de proliferação celular e apoptose, é
responsável também por regular expressão de citocinas, quimiocinas, fatores de crescimento e

18
moléculas de adesão celular como por exemplo, proteínas de fase aguda e mediadores inflamatórios
(COX-2 e iNOS). O processo de interferência do Resveratrol na expressão de NF-кB ainda não é
completamente conhecido, porém sabe-se que esse composto inibe a fosforilação de IkB e pode
também bloquear a expressão do gene regulador do fator de transcrição[6].
O Resveratrol pode ter característica anticâncer por possuir efeito inibitório no crescimento de
várias linhagens de células cancerosas [35-36] e por agir como um agente quimioprotetor e inibidor em
diversos eventos relacionados com a iniciação, promoção e progressão do tumor.
A capacidade antioxidante, “scavanger”[37] de radicais livres, do Resveratrol é a mais estudada,
por reduzir o acúmulo de espécies reativas de oxigênio (ROS). Essa redução pode ocorrer diretamente
ou por atuação nas vias de sinalização que produzem essas espécies.
Além disso, o Resveratrol pode modular vias de sinalização, como por exemplo, a PKC. Como
descrito anteriormente, essa via é de grande importância na patogênese do Diabetes Tipo 2, pois
quando é ativada leva a ativação da NADPHoxidase induzindo a produção de ROS, culminando em um
estresse oxidativo (Figura 2).
Figura 2: Mecanismo de ativação da PKC e possível modulação pelo Resveratrol demonstrando seu efeito no processo de estresse oxidativo. Adaptado de Nogueira – Machado, et al, 2008[1]
O RSV também está envolvido na modulação de vários fatores que medeiam a inflamação. O
processo inflamatório tem início com uma fase aguda que pode perdurar por poucos dias, mas se não é
solucionada pode se tornar uma fase crônica na qual observa-se a presença de linfócitos bem como a
proliferação de vasos sanguíneos. No esquema seguinte pode ser observado os locais de ação do RSV
no processo inflamatório (Esquema 6)[7].

19
Esquema 6 : Ação do Resveratrol no processo inflamatório.
Adaptado de la Lastra, C.A. and I. Villegas, 2005[7]
Estudos tem demonstrado forte relação do RSV com a patologia do Diabetes através da
alteração ou modulação de diversas vias ou mecanismos fundamentais para o desenvolvimento da
doença e de suas complicações, tais como produção de ROS [38], fatores de crescimento como o VEGF
[39], produção de NO, PKC [40-42] e fatores de transcrição como o NFкB [7].
Dessa forma, nosso objetivo foi avaliar o efeito do Resveratrol sobre a função de granulócitos e
mononucleares de pacientes diabéticos tipo 2 em comparação com indivíduos não diabéticos. Para
isso, abordamos especificamente os seguintes pontos:
• Comparar o efeito do Resveratrol na produção de Espécies Reativas de Oxigênio (ROS) em
pacientes diabéticos tipo 2 (DM2) e controles não diabéticos (ND),
• Avaliar o efeito do Resveratrol na fagocitose por granulócitos de ND e DM2, estimulados com
partículas de Zimosan opsonizado com fragmento C3b,
• Avaliar a possível inibição da PKC pelo Resveratrol em granulócitos de ND e DM2,
• Avaliar o efeito do Resveratrol na produção de Óxido Nítrico,
• Avaliar o efeito do Resveratrol sobre a produção de VEGF por mononucleares de pacientes
diabéticos tipo 2 em comparação com indivíduos não diabéticos.

20
2 Pacientes e Métodos
2.1 Seleção de Paciente
O presente trabalho foi aprovado pelo comitê de ética em pesquisa (CEP) da Santa Casa de
Belo Horizonte (SC-BH). Os pacientes foram selecionados no Centro de Especialidades Médicas
(CEM) da SC-BH.
Foram incluídos os doadores que concordaram em assinar o Termo Consentimento Livre e
Esclarecido (TCLE). Foram selecionados pacientes de ambos os sexos, com diagnóstico de diabetes
tipo 2 e doadores do grupo controle, sabidamente normoglicêmicos.
Foram excluídos do trabalho pacientes portadores de doenças autoimunes, câncer, demência,
gestantes, fumantes e aqueles que não concordaram em assinar o Termo de Consentimento livre e
esclarecido (TCLE).
2.2 Coleta de Sangue
Amostras de sangue venoso periférico foram coletadas dos pacientes diabéticos e dos indivíduos
controle por meio da punção venosa, em tubos vacuntainer contendo heparina como anticoagulante.
2.2.1 Separação Celular: obtenção de Leucócitos
Esquema 7: Obtenção de Leucócitos pela técnica de Ficoll-Hypaque.
Purificação de Leucócitos

21
Os leucócitos foram obtidos a partir do sangue periférico, de acordo com Bicalho, H. M. S. e
cols[43], quatro mililitros de sangue periférico heparinizado foram adicionados a seis mililitros de
gradiente duplo de Ficoll-Hypaque de densidade 1,08 (3,0ml) e 1,12 (3,0ml). Este tubo foi centrifugado
por 30 minutos a 1000 rpm, originando um anel formado por células mononucleares e outro anel
formado por granulócitos. Cada anel foi então transferido separadamente para um tubo siliconizado e
foi submetido a três lavagens em PBS. Após as lavagens, as células foram ressuspensos em um
mililitro de PBS. A contagem destas células foi realizada no microscópio utilizando uma câmara de
Neubawer e o número calculado pela seguinte fórmula: células/mL = nº/4 x diluição (100) x 104. A
viabilidade celular de cada amostra foi superior a 90%, como determinado pelo teste de exclusão
realizado por incorporação de azul de Trypan.

22
2.3 Reagentes
I. PBS solução Salina Tamponada (pH:7.3)[44]
Tabela 2: Reagentes necessários para Solução Salina Tamponada (PBS) em pH 7.3.
Reagentes Quantidade (gramas)
Na2HPO4 8.12
KH2PO4 1.35
NaCl 8.0
H2O destilada q.s.p. 1 litro
II. Luminol (5 – amino - 2,3 - dihydro – 1,4 phthalozinedione) – SIGMA Co.
Foi pesada uma quantidade 1,77 gramas de Luminol e esta foi diluída em 1,0mL de dimetilsulfóxido
(DMSO) obtendo-se então uma concentração de 10-2M. Para chegar a concentração de uso 10-4M, foi
realizada uma diluição em PBS.
III. Solução para separação de Granulócitos
-Monopaque: Gradiente de separação de células mononucleares, com densidade igual a 1.08%,
preparado pela Bion.
-Leucopaque: Gradiente de separação de granulócitos, com densidade igual a 1.12%, também
preparado pela Bion.
IV. Solução para separação de Mononucleares.
-Monopaque: Gradiente de separação de células mononucleares, fornecido pela Bion.
V. Azul de Trypan - SIGMA Co.
O reagente foi usado à 1% diluído em PBS.
VI. Resveratrol–( 3,4′,5-Trihydroxy-trans-stilbene,5-[(1E)-2-(4-Hydroxyphenyl)ethenyl]-1,3-benzenediol)
SIGMA Co.
O RSV foi reconstituído primeiramente com 1mL de DMSO, formando o primeiro estoque. A seguir o
estoque que estava a 1 molar foi diluído para uma concentração de 1000µg, com solução de PBS, que
foi novamente diluído para se obter uma concentração de uso de 10µg/mL. Foram feitas alíquotas
desta diluição que foram armazenadas a -20ºC.

23
VII. PDB - Phorbol 12,13-dibutyrate SIGMA Co.
O reagente foi diluído em DMSO e em seguida foram realizadas diluições para alcançar a concentração
de uso 1x10-4.
VIII. L-NAME - Nω-Nitro-L-arginine methyl ester hydrochloride SIGMA Co.
O reagente foi reconstituído com DMSO e diluído para uma concentração de 1mM.
IX. Calphostin (Calphostin C - Cladosporium cladosporioides) SIGMA Co
Para obter a concentração de uso 0,1µM foi realizado uma diluição de 1:10 do estoque primário 1mM.
2.4 Opsonização de Zimosan
Para a produção de Zimosan opsonizado (Zc3b) foi coletado sangue venoso periférico humano
em tubos vacuntainer sem anticoagulante, e armazenados em geladeira por 2 horas. Após o repouso, o
sangue foi então, centrifugado a 3000 rpm por 15 minutos, formando no tubo 2 fases, sendo a primeira
de cor amarelada translúcida correspondente ao soro. Para cada mL de soro foi acrescentado 50µl de
Zimosan (13mg), formando uma nova solução que foi incubada a 37ºC por 2 horas. Após este processo
o reagente estava pronto para uso.
2.5 Teste de Viabilidade Celular: Incorporação de Azul de Trypan
O teste de viabilidade celular foi realizado com o reagente Azul de Trypan a 1%. Onde as
células, contadas em câmara de Neubawer, que incorporaram o Azul de Trypan foram consideradas
mortas. As amostras utilizadas no experimento continham, pelo menos, 90% de células vivas.

24
2.6 Ensaio de quimioluminescência dependente de luminol: quantificação das espécies
reativas de oxigênio (ROS)
Com o ensaio de quimioluminescência foi possível avaliar, indiretamente, a atividade da
NADPH-oxidase, que é uma enzima responsável pela geração de ROS durante o processo de
fagocitose das células. Em testes in vitro, esta luminosidade pode ser ampliada, usando-se reagentes
químicos que, ao reagirem com as espécies reativas de oxigênio (ROS), passam a emitir uma
luminescência maior.
2.6.1 Teste de Quimioluminescência
Inicialmente, para avaliar o perfil antioxidante do Resveratrol, foi realizada uma leitura basal
onde foi adicionado ao tubo específico para o luminômetro, 100µL de granulócitos (1x106 /mL), 400µL
de PBS e por fim 200µL de Luminol 10-4. Esta leitura teve uma duração de 30 minutos, onde a cada
minuto o aparelho registra um valor. Após a leitura basal foi adicionado 100µL de Resveratrol (10µg)
ao mesmo tubo, sendo novamente realizada uma leitura por 30 minutos.
2.6.2 Atividade Fagocítica
Para avaliar a atividade fagocítica, foi realizada uma leitura basal contendo 100µL de
granulócitos (1x106 /mL), 400µL de PBS e por fim 200µL de Luminol 10-4 incubada por 30 minutos.
Após este tempo foi adicionado ao tubo 30µL de Zimosan opsonizado com partículas C3b e realizado
uma nova leitura de 30 minutos. Após o final desta leitura, por fim, foi acrescentado 100µL de
Resveratrol (10µg) e realizada a leitura por 30 minutos.

25
2.6.3 Inibição da PKC pelo Resveratrol
Para avaliar a atuação do Resveratrol na PKC foi realizada inicialmente uma leitura basal,
como já demonstrada nos tópicos acima, realizada a leitura de 15 minutos e logo após, acrescentado
ao tubo 30µL de PDB para ativar a produção de ROS pela via de interesse, realizada a leitura de 30
minutos e acrescentado logo após 100µL de Calphostin C, um inibidor da via, foi então realizada mais
uma leitura de 30 minutos. Em outro tubo foi realizado uma nova leitura basal, com as mesmas
quantidades e reagentes citados anteriormente, foi realizado a leitura de 15 minutos e logo após foi
adicionado novamente o PDB sendo então feita mais uma leitura de 30 minutos e por fim adicionado
100µL Resveratrol e realizado uma leitura de 30 minutos. Este experimento teve a finalidade de
analisar se o Resveratrol poderia inibir a produção de ROS PKC. Um próximo experimento foi feito
onde após a leitura basal foi acrescentado 100µL de Calphostin C e após os 30 minutos de leitura foi
adicionado 30µL de PDB. Em um novo tubo foi adicionado após a leitura basal 100µL Resveratrol e
após a sua leitura foi acrescido 30µL de PDB. Estes dois últimos experimentos foram realizados para
observar se o Resveratrol e o Calphostin C poderiam inibir totalmente a PKC, mesmo ela sendo ativada
após o acréscimo de um inibidor já conhecido (Calphostin C) e um possível inibidor como o
Resveratrol.
2.6.4 Produção de Oxido Nítrico
A produção de Oxido Nítrico foi avaliada em sobrenadante de cultura celular (18 horas em
estufa de 5% CO2/37ºC) de granulócitos, pelo método de Griess, na presença/ausência de um inibidor
(L-NAME) e presença/ausência de Resveratrol.
Método de Griess: Os sobrenadantes da cultura de granulócitos (18 horas em estufa de 5%
CO2/37ºC) foram analisados quanto à produção de nitrito pela reação de Griess, como medida da
produção de óxido nítrico[45]. Para medir a produção de nitrito, alíquotas de 100uL das amostras foram
incubadas com 100uL dos reagentes (50uL da solução de sulfanilamida 1% e 50uL de solução de N-
naphthylethylenediamine dihydrocloride 0,1% em 2,5% de H3PO4) à temperatura ambiente por 10
minutos. A leitura foi realizada no espectrofotômetro com absorbância de 570nm.

26
2.7 Análise da Expressão de VEGF
A análise da produção de VEGF foi realizada em sobrenadante de cultura celular (18 horas em
estufa de 5% CO2/37ºC) de granulócitos, na presença e ausência de Calphostin C, LPS e Resveratrol.
A dosagem foi feita de acordo com as indicações do fabricante do kit: VEGF (Human) ELISA Kit – Enzo
Life Sciences (ADI-900-080).
2.8 Equipamentos
Luminômetro (Ciba-Corning – Magic Lite Analizer)
Espectrofotômetro (Stat Fax 2100; UV mini - 1240 – Shimadzu)
Centrífugas (Centrifugador Excelsa Fanem Ltda; Bio Eng – 6000)
Banho Maria (Hemoquímica)
Estufa de CO2
Microscópio (Edu VU 1300 – WESCO)
2.9 Análise Estatística
A análise estatística foi feita utilizando o teste “t” de Student, sendo p < 0,05 considerado como
estatisticamente significativo.
27
3 Resultados
Neste trabalho os resultados serão apresentados em forma de perguntas buscando seguir uma
sequência lógica para alcançar o objetivo geral. Abaixo estão apresentadas as principais características
das populações estudadas:
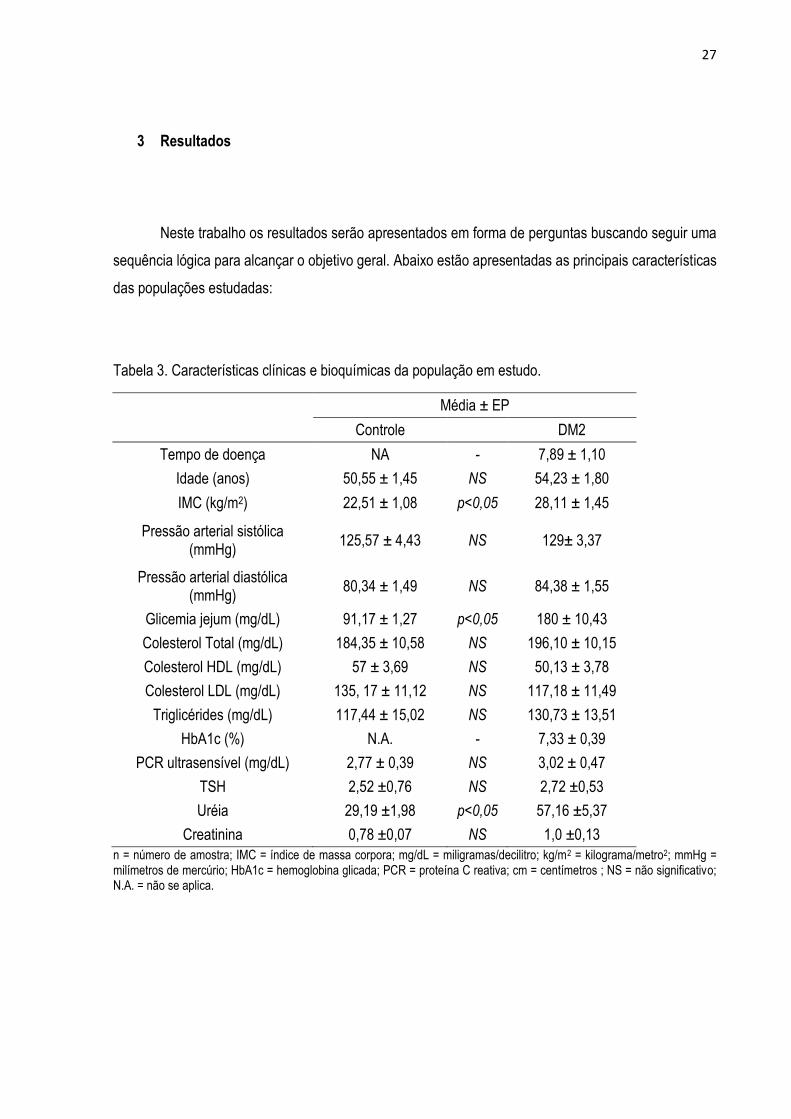
Tabela 3. Características clínicas e bioquímicas da população em estudo.
Média ± EP
Controle DM2
Tempo de doença NA - 7,89 ± 1,10
Idade (anos) 50,55 ± 1,45 NS 54,23 ± 1,80
IMC (kg/m2) 22,51 ± 1,08 p<0,05 28,11 ± 1,45
Pressão arterial sistólica (mmHg)
125,57 ± 4,43 NS 129± 3,37
Pressão arterial diastólica (mmHg)
80,34 ± 1,49 NS 84,38 ± 1,55
Glicemia jejum (mg/dL) 91,17 ± 1,27 p<0,05 180 ± 10,43
Colesterol Total (mg/dL) 184,35 ± 10,58 NS 196,10 ± 10,15
Colesterol HDL (mg/dL) 57 ± 3,69 NS 50,13 ± 3,78
Colesterol LDL (mg/dL) 135, 17 ± 11,12 NS 117,18 ± 11,49
Triglicérides (mg/dL) 117,44 ± 15,02 NS 130,73 ± 13,51
HbA1c (%) N.A. - 7,33 ± 0,39
PCR ultrasensível (mg/dL) 2,77 ± 0,39 NS 3,02 ± 0,47
TSH 2,52 ±0,76 NS 2,72 ±0,53
Uréia 29,19 ±1,98 p<0,05 57,16 ±5,37
Creatinina 0,78 ±0,07 NS 1,0 ±0,13
n = número de amostra; IMC = índice de massa corpora; mg/dL = miligramas/decilitro; kg/m2 = kilograma/metro2; mmHg = milímetros de mercúrio; HbA1c = hemoglobina glicada; PCR = proteína C reativa; cm = centímetros ; NS = não significativo; N.A. = não se aplica.

28
Os dados acima mostram diferença significativa para o IMC, os níveis de glicemia em jejum
entre os pacientes diabéticos tipo 2 e seu respectivo controle não diabético, pôde ser observado
também que a hemoglobina glicosilada (HbA1c) encontra-se em porcentagem superior ao valor normal
em DM2 e também uma alteração dos níveis de uréia.
Assim procuramos responder nossa primeira pergunta:
29
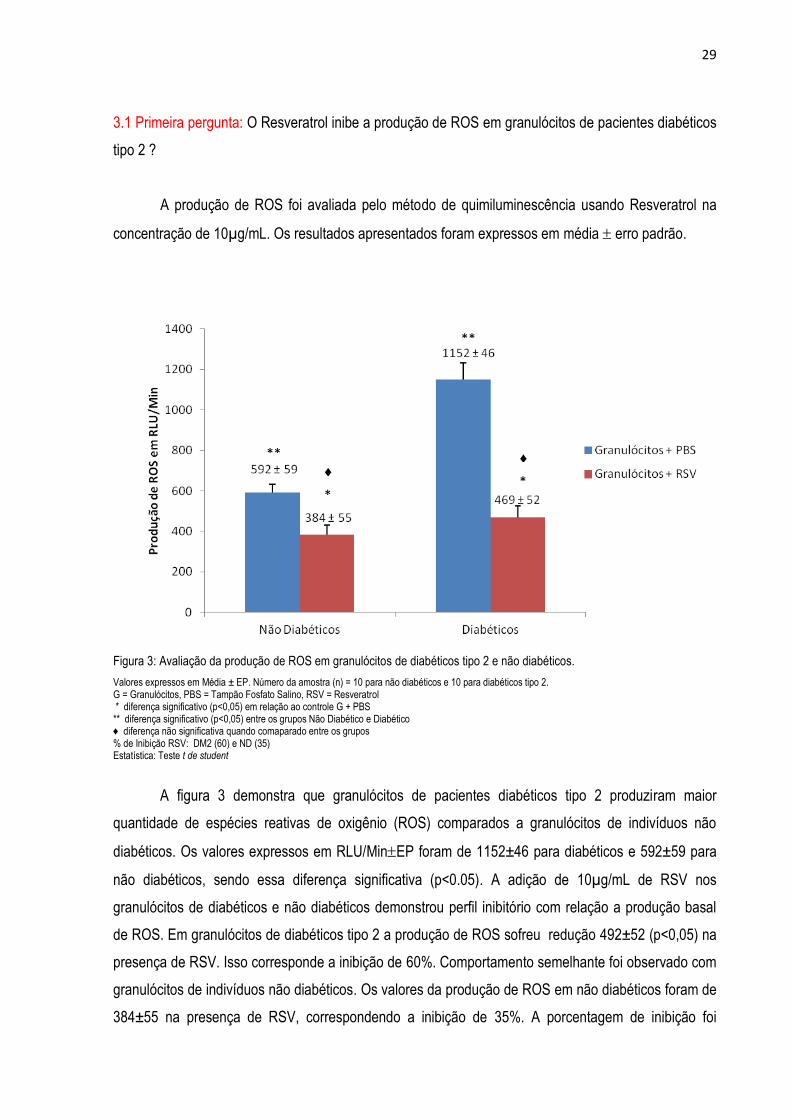
3.1 Primeira pergunta: O Resveratrol inibe a produção de ROS em granulócitos de pacientes diabéticos
tipo 2 ?
A produção de ROS foi avaliada pelo método de quimiluminescência usando Resveratrol na
concentração de 10µg/mL. Os resultados apresentados foram expressos em média erro padrão.
Figura 3: Avaliação da produção de ROS em granulócitos de diabéticos tipo 2 e não diabéticos.
Valores expressos em Média ± EP. Número da amostra (n) = 10 para não diabéticos e 10 para diabéticos tipo 2. G = Granulócitos, PBS = Tampão Fosfato Salino, RSV = Resveratrol * diferença significativo (p<0,05) em relação ao controle G + PBS ** diferença significativo (p<0,05) entre os grupos Não Diabético e Diabético ♦ diferença não significativa quando comaparado entre os grupos % de Inibição RSV: DM2 (60) e ND (35) Estatística: Teste t de student
A figura 3 demonstra que granulócitos de pacientes diabéticos tipo 2 produziram maior
quantidade de espécies reativas de oxigênio (ROS) comparados a granulócitos de indivíduos não
diabéticos. Os valores expressos em RLU/MinEP foram de 1152±46 para diabéticos e 592±59 para
não diabéticos, sendo essa diferença significativa (p<0.05). A adição de 10µg/mL de RSV nos
granulócitos de diabéticos e não diabéticos demonstrou perfil inibitório com relação a produção basal
de ROS. Em granulócitos de diabéticos tipo 2 a produção de ROS sofreu redução 492±52 (p<0,05) na
presença de RSV. Isso corresponde a inibição de 60%. Comportamento semelhante foi observado com
granulócitos de indivíduos não diabéticos. Os valores da produção de ROS em não diabéticos foram de
384±55 na presença de RSV, correspondendo a inibição de 35%. A porcentagem de inibição foi
* *
**
**
♦ ♦

30
semelhante tanto para o grupo controle quanto para o grupo DM2, não havendo diferença significativa
entre os grupos. Este experimento foi realizado com granulócitos não estimulados o que nos levou a
questionar se granulócitos previamente ativados exibiriam o mesmo comportamento. Dessa forma,
ativamos os granulócitos com partículas opsonizadas de Zimozan, sabidamente conhecido como
indutor no processo fagocítico, que ativa a via metabólica produtora de ROS. Com base nesta
suposição, fizemos a nossa segunda pergunta.

31
3.2 Segunda Pergunta: O Resveratrol inibe a fagocitose de partículas opsonizadas em granulócitos de
pacientes diabéticos tipo 2 e não diabéticos?
A fagocitose foi avaliada pelo método de quimioluminescência, através da produção de ROS,
na presença de partículas opsonizadas de zimosan, que induzem a produção de ROS pela via
alternativa do complemento.
Figura 4: Avaliação da atividade fagocítica de granulócitos estimulados com partículas de zimosan opsonizado com fragmentos C3b. Valores expressos em relação E/C = experimento expresso em RLU/ Controle expresso em RLU. Os valores da produção de ROS em granulócitos ativados com partículas de ZC3b expressos em RLU/Min foram: Não Diabéticos = 1777 ± 166 e Diabéticos = 2516 ± 203 e para granulócitos ativados + Resveratrol: Não Diabéticos = 302 ± 12 e Diabéticos = 629 ± 62. Número da amostra (n) = 10 para não diabéticos e 10 para diabéticos tipo 2. G= Granulócitos, PBS= Tampão Fosfato Salino, RSV= Resveratrol, ZC3b = Partículas de Zimosan opsonizado com fragmentos C3b. * experimento significativo (p<0.05) em relação ao controle G + PBS ** = experimento significativo (p<0,05) entre os grupos Não Diabético e Diabético # = experimento não significativo comparado entre os grupos % de Inibição DM2(44) ND(81) Estatística: Qui-quadrado
Na figura 4 observamos maior produção de ROS por granulócitos de não diabéticos na
presença de ZC3b comparado com o grupo diabético. Os valores expressos em E/C ± EP foram, 3.24
± 0.36 para não diabéticos e 2.22 ± 0.018 para diabéticos, sendo esta diferença significativa
(p<0,05). Ao adicionar o RSV na concentração de 10µg/mL foi observada redução da produção de
ROS para ambos os grupos. Nossos resultados sugerem que o RSV inibe a atividade fagocítica de
granulócitos ativados ou sem nenhum estímulo.
Para verificarmos a atuação do Resveratrol na via de sinalização da PKC, importante via de
sinalização para o DM2 capaz de induzir a produção de ROS, realizamos um experimento ativando
*
*
**
**
#
#

32
esta via com Éster de Forbol (PDB), ativador conhecido da PKC, e inibindo-a com Calphostin C, inibidor
da PKC, ou com Resveratrol. Neste contexto formulamos a terceira pergunta.

33
3.3 Terceira Pergunta: O Resveratrol inibe a PKC?
Para avaliar a possível atividade inibitória do Resveratrol sobre a PKC, analisamos a produção
de ROS pelo método de quimioluminescência dependente de Luminol na presença de PDB, Calphostin
C e Resveratrol. Os valores obtidos foram expressos em RLU/min ± Erro padrão.
Tabela 4: Modulação da PKC pelo Resveratrol.
EXPERIMENTOS
Produção de ROS (RLU/Min ± EP)
Não Diabético
%
Inibição
Diabético (DM2)
%
Inibição
p
1) GRANULÓCITOS + PBS 1620 ± 599 - 3033 ± 864 p<0,05
2) GRANULÓCITOS + PDB 2702 ± 387 - 7214 ± 351 p<0,05
3) GRANULÓCITOS + RSV 588 ± 49 63% 469 ± 30 84% NS
4) GRANULÓCITOS + CALPHOSTIN C 475 ± 146 70% 866 ± 16 71% p<0,05
5) GRANULÓCITOS + PDB + RSV 624 ± 35 77% 2434 ± 672 66% p<0,05
6) GRANULÓCITOS + PDB+CALPHOSTIN C 1332 ± 375 51% 4287 ± 254 40% p<0,05
7) GRANULÓCITOS + CALPHOSTIN C + RSV 176 ± 22 62% 522 ± 55 73% p<0,05
Valores expressos em Média de unidade de luz/minuto ± Erro Padrão
Número da amostra (n) = 10 para não diabéticos e 10 para diabéticos tipo 2.
DM2= Diabeticos tipo 2; ND= Não Diabéticos; NS= Não Significativo; *p<0.05; PDB = Phorbol 12,13-dibutyrate ; RSV = resveratrol
p<0,05: experimento 2 vs experimento 1; 3 vs 1; 4 vs 2; 5 vs 1; 6 vs 2; 7 vs 6; para ND e DM2.
p<0,05: experimento 4 vs 8 para ND
Estatística: Teste t de student
Os resultados da tabela 4 demonstram que a produção basal de ROS em granulócitos de
diabéticos (3033 ± 864) foi significativamente maior (p<0,05) comparado com não diabéticos (1620 ±
599). Na presença de PDB houve aumento da produção de ROS em ambos os grupos, em diabéticos a
produção foi de 7214 ± 351 e em não diabéticos 2702 ± 387. No entanto, a produção de ROS
estimulada com PDB foi significativamente maior (p<0,05) nas células de pacientes diabéticos. Em
ambos os experimentos, com e sem estímulo, podemos observar que os granulócitos de diabéticos
produziram quantidade maior de ROS do que as células de não diabéticos.
Ao analisar a ação do Resveratrol observamos que houve redução significativa (p<0,05) da
produção de ROS por granulócitos de diabéticos e não diabéticos, comparado com a leitura basal,
entretanto não houve diferença significativa (p<0,05) entre os grupos. Com esses dados verificamos
que o Resveratrol pode inibir a produção de ROS mesmo em célula não ativada.

34
Ao adicionar o Calphostin C, inibidor da PKC, observamos redução significativa (p<0,05) da
produção de ROS por granulócitos quando comparado com sua leitura basal e também quando
comparado entre os grupos. Os valores obtidos após acrescentar o inibidor foram: 475 ± 146 para não
diabéticos e 866 ± 16 para diabéticos. Verificamos que a inibição da produção de ROS por Calphostin
C foi significativamente semelhante em granulócitos de pacientes diabéticos e não diabéticos quando
comparado com seu respectivo controle.
Para observar uma possível inibição do Resveratrol na PKC, os granulócitos foram ativados com
PDB e logo após a leitura foi adicionado o Resveratrol. Observamos inibição significativa (p<0,05) do
Resveratrol na PKC comparado com a leitura basal em ambos os grupos. Os valores obtidos foram:
leitura basal 2702 ± 387 em não diabéticos e 7214 ± 351 em pacientes diabéticos e ao acrescentar o
Resveratrol: 624 ± 35 para não diabéticos e 2434 ± 672 para diabéticos. Como controle desse último
experimento, os granulócitos foram ativados com PDB e logo após foi adicionado o Calphostin C que
inibiu significativamente (p<0,05) a produção de ROS em células de pacientes diabéticos e não
diabéticos comparado com a leitura basal.
O mesmo protocolo foi realizado com o Resveratrol, no qual ao acrescentar o polifenol foi
encontrada redução significativa (p<0,05) da produção de ROS quando comparado com o controle do
experimento e quando comparado entre os grupos. Os valores representados em RLU/Min±EP foram:
176 ± 22 para não diabéticos e 522 ± 55 para diabéticos. Com isso podemos sugerir que o Resveratrol
é um inibidor eficaz da produção de ROS pela PKC.

35
3.4 Quarta Pergunta: O Resveratrol pode atuar na modulação de Óxido Nítrico?
Para avaliar o comportamento do RSV na produção de Oxido Nítrico entre pacientes Diabéticos
e Não diabéticos, foi realizada inicialmente cultura de granulócitos por 18 horas à 37ºC onde o
composto foi incubado com ou sem o inibidor de NO (L-NAME). Após o tempo de incubação
quantificamos o NO pelo Método de Griess.
Tabela 5: Modulação da produção de NO pelo Resveratrol.
EXPERIMENTOS
Produção de NOS(µMol)
ND DM2 p
1) GRANULÓCITOS + MEIO 8,37 ± 0,95 9,2 ± 0,92 NS
2) GRANULÓCITOS + RSV 4,7 ± 0,90 5,8 ± 1,43 p<0,05
3) GRANULÓCITOS + L-NAME 5,5 ± 0,67 3,8 ± 1,22 p<0,05
4) GRANULÓCITOS + RSV + L-NAME 3,0 ± 1,00 5,6 ± 1,29 p<0,05
Valores expressos em Média ± Erro Padrão
Número da amostra (n) = 10 para não diabéticos e 10 para diabéticos tipo 2.
DM2= Diabeticos tipo 2; ND= Não Diabéticos; NS= Não Significativo; L-NAME = Nω-Nitro-L-arginine methyl ester hydrochloride; RSV = Resveratrol
p<0,05: experimento 2 vs experimento 1; 3 vs 1; 4 vs 2 para ND vs ND
p<0,05: experimento 2 vs experimento 1; 3 vs 1 para DM2 vs DM2
N.S: experimento 4 vs 2 para DM2 vs DM2
Estatística: Teste t de student
Na tabela 5 podemos observar que houve redução significativa (p<0,05) da produção de NO ao
adicionar o Resveratrol em células de pacientes diabéticos e não diabéticos, quando comparado com a
leitura basal e quando comparado entre os grupos estudados. Os valores basais encontrados foram:
8,37 ± 0,95 para não diabéticos e 9,2 ± 0,92 para diabéticos. Os valores encontrados ao adicionar o
Resveratrol foram: 4,7 ± 0,90 em não diabéticos e 5,8 ± 1,43 em diabéticos. A inibição
de NO pelo Resveratrol se mostrou menor em pacientes diabéticos tipo 2.
Ao adicionar L-NAME, inibidor da NOS, observamos redução significativa da produção de NO em
ambos os grupos, quando comparado com seu controle e também quando comparado entre os dois
grupos. Com o objetivo de verificar o potencial inibitório do RSV foi realizado outro experimento onde o
polifenol foi incubado juntamente com o inibidor L-NAME. Verificamos que em não diabéticos houve
redução significativa (p<0,05) da produção de NO comparando com o experimento onde foi incubado
apenas o Resveratrol. Já em pacientes diabéticos não observamos redução significativa da produção

36
de NO. O L-NAME inibiu a produção de NO em maior escala em pacientes diabéticos, porém na
presença do Resveratrol esta inibição não foi potencializada.

37
3.5 Quinta Pergunta: O Resveratrol pode atuar na produção de VEGF?
Para avaliar o comportamento do RSV na produção de VEGF e se existe relação entre este
fator de crescimento e a PKC entre pacientes Diabéticos e Não diabéticos, realizamos uma cultura de
mononucleares por 72 horas à 37ºC na qual o composto foi incubado na presença ou ausência de
Calphostin C, PDB e LPS (controle positivo). Após o término da cultura celular foi recolhido o
sobrenadante para dosagem de VEGF pelo método de ELISA.
Tabela 6: Produção de VEGF na presença de Resveratrol.
EXPERIMENTOS
Concentração de VEGF (pg/mL ± EP)
Não Diabético
Diabético (DM2)
p
1) MONONUCLEAR + PBS 29,0 ± 9,0 7,0 ± 1,0 p<0,05
2) MONONUCLEAR + PDB 48,0 ± 10,0 6,9 ± 1,0 p<0,05
3) MONONUCLEAR + RSV 22,0 ± 3,0 4,0 ± 0,5 p<0,05
4) MONONUCLEAR + CALPHOSTIN C 12,0 ± 10,1 4,0 ± 0,9 p<0,05
5) MONONUCLEAR+ RSV + PDB 89,0 ± 20,0 7,0 ± 0,7 p<0,05
6) MONONUCLEAR +CALPHOSTIN C+ PDB 35,0 ± 18,0 9,0 ± 1,0 p<0,05
7) MONONUCLEAR+ RSV + CALPHOSTIN C 11 ± 4,0 4,0 ± 1,0 NS
8) MONONUCLEAR + LPS 28,0 ± 10,0 11,0 ± 1,5 NS
Valores expressos em Média ± Erro Padrão
Número da amostra (n) = 10 para não diabéticos e 10 para diabéticos tipo 2.
DM2= Diabeticos tipo 2; ND= Não Diabéticos; NS= Não Significativo; p<0.05: significativo; RSV = Resveratrol; PDB: Phorbol 12,13-dibutyrate
p<0,05: experimento 2 vs experimento 1; 5 vs 3; para ND vs ND
p<0,05: experimento 4 vs experimento 6; 8 vs 1 para DM2 vs DM2
N.S: experimento 2 vs 1; 3 vs 1; 4 vs 1; 6 vs 4; 7 vs 3; 8 vs 1 para ND vs ND
N.S: experimento 2 vs 1; 3 vs 1; 4 vs 1; 5 vs 3; 7 vs 3 para DM2 vs DM2
Estatística: Teste t de student
Na tabela 6 observamos que ao adicionar o PDB, ativador seletivo da PKC, não houve aumento
diferença significativa (p<0,05) da produção de VEGF em ambos os grupos estudados quando
comparados com seus respectivos controles.Os valores obtidos foram: 29,0 ± 9,0 na leitura basal e
48.0 ± 10.0 com a presença de PDB e 7,0 ± 1,0 na leitura basal e 6,9 ± 1,0 na presença de PDB para
pacientes diabéticos. Ao adicionar o Resveratrol foi possível observar uma redução da produção de
VEGF nos dois grupos estudados, porém essa diferença não foi significativa quando comparada com
seu respectivo controle. Ao comparar entre os grupos, diabéticos e não diabéticos foi possível

38
encontrar diferença significativa (p<0,05) onde no grupo de diabéticos essa produção foi menor. Ainda
com o objetivo de verificar uma interação da PKC com a produção de VEGF, foi adicionado o
Calphostin, inibidor da PKC, onde não observamos uma redução significativa (p<0,05) da produção de
VEGF em indivíduos não diabéticos e pacientes diabéticos. Todavia, ao comparar os valores obtidos
entre os grupos de diabéticos e não diabéticos verificamos uma diferença significativa (p<0,05) sendo a
produção de VEGF maior em não diabéticos. Mesmo na presença do Resveratrol, ao adicionar o PDB,
indivíduos não diabéticos apresentaram um aumento significativo (p<0,05) da produção de VEGF
quando comparado com seu controle. Este resultado sugere que o RSV pode potencializar a ação do
PDB em não diabéticos. Esta diferença também foi significativa quando comparada entre os grupos
estudados. Em outro experimento foi adicionado o Calphostin juntamente com o PDB, como resultado
foi obtido um discreto aumento da produção de VEGF em ambos os grupos, porém essa diferença não
foi significativa quando comparado com seus controles. Entretanto, ao comparar entre os grupos foi
possível observar uma diferença significativa (p<0,05) com aumento maior no grupo de não diabéticos.
Ao adicionar o Resveratrol juntamente com o Calphostin não foi observada nenhuma ação dos
compostos na produção de VEGF quando comparado com seus respectivos controles e entre os
grupos estudados, demonstrando que o Resveratrol não atua na inibição da produção de VEGF pela
atividade da PKC. Por fim, foi adicionado o LPS atuando como um controle positivo do experimento,
não havendo diferença significativa entre os grupos e quando comparado com seus controles. Os
resultados no seu conjunto demonstram que o Resveratrol foi capaz de reduzir a produção de VEGF
pela modulação negativa da PKC.

39
4 Discussão
No presente trabalho avaliamos a capacidade antioxidante do Resveratrol. Nossos resultados
iniciais demonstraram que o Resveratrol inibiu a produção de ROS tanto em granulócitos de diabéticos
quanto em granulócitos de indivíduos não diabéticos.
Diversos estudos demonstram que o estresse oxidativo, originado pelo aumento da produção
de ROS sem o equivalente aumento da resposta antioxidante, tem papel importante na patogênese do
Diabetes e suas complicações [46]. No Diabetes, a hiperglicemia pode causar alterações em diversas
vias de sinalização levando a progressão das complicações diabéticas[1].
Em condições fisiológicas, nosso organismo possui eficientes mecanismos de defesa
antioxidante capaz de combater o excesso de radicais livres, mantendo a homeostase e integridade
celular. Em pacientes diabéticos, devido ao excesso de glicose, a capacidade antioxidante está
prejudicada, não sendo possível a remoção completa dos radicais livres [47-49]. Dessa forma, a
ineficiência dos sistemas antioxidantes permite a instalação do estresse oxidativo.
Atualmente tem crescido a busca por compostos naturais que possam auxiliar no controle de
patologias, como o Diabetes. O Resveratrol, um polifenol naturalmente encontrado em plantas, tem
sido estudado por possuir propriedades anti-inflamatórias[7], antioxidantes [50], dentre outras, podendo
agir diretamente com o ROS ou interagir com vias enzimáticas que participam da formação dessas
espécies reativas de oxigênio [51].
A figura 3 demonstra nitidamente que na presença do Resveratrol houve redução da produção
de ROS em granulócitos de diabéticos e não diabéticos. Esta redução pode ter ocorrido por uma
modulação do composto sobre vias de sinalização que auxiliam nesse processo de aumento de
espécies reativas de oxigênio, dentre elas podemos destacar a NADPH oxidase.
Os nossos resultados concordam com Spanier, G. et al [41] que demonstraram que o
Resveratrol modulou a NADPH oxidase, reduzindo a formação de ROS intracelular. Goldberg, D.M.et
al. [31] também demonstram que o Resveratrol diminuiu a produção de ROS, possivelmente por inibir a
translocação de subunidades regulatórias da NADPH oxidase na membrana.
Segundo Kovacic & Somanathan[52], o Resveratrol é relatado por inibir a produção de ROS
como peróxido de hidrogênio, superóxido e radical hidroxil evitando assim danos pró–oxidativos.
Sharma S. et al [53] demonstraram, em seus experimentos, que o Resveratrol possui um efeito benéfico
na modulação do estresse oxidativo em vários tecidos afetados pela hiperglicemia, incluindo células
neuronais[54], renais[53], endoteliais[55] e células β – pancreáticas [56].

40
Silan C. [57] observou em seu trabalho que o Resveratrol possui efeito protetor contra o estresse
oxidativo em diferentes tecidos e em condições patológicas como doenças cardiovasculares, resposta
inflamatória, câncer e diabetes. Harikumar KB et al [58] demonstraram que o Resveratrol se comporta
como “scavenger” de radicais livres e modulador enzimático.
Observamos que o Resveratrol foi capaz de inibir a produção de ROS em granulócitos durante
a fagocitose de partículas opsonizadas. Na figura 4, os resultados expressos pela relação
Experimento/Controle (E/C), demonstraram que ao ativar os granulócitos com ZC3b houve aumento da
produção de ROS em ambos os grupos estudados. Na presença de Resveratrol estas células
apresentaram perfil de redução dessa produção.
Com base nesses resultados sugerimos que o Resveratrol exerce ação inibitória no processo
de fagocitose medido pela produção de ROS. Leiro et al. [59] demonstraram que o Resveratrol reduz a
capacidade fagocítica de células de ratos. Este dado é comparável aos nossos resultados (Figura 4).
Em contrapartida, Vetvicka et al [60] demonstraram aumento da capacidade fagocítica ao utilizar o
Resveratrol, em células peritoneais de ratos. Esses dados, no seu conjunto, sugerem que o efeito do
Resveratrol possa ser dependente do tipo celular estudado.
Neste trabalho investigamos também a possível modulação do Resveratrol sobre a atividade
da PKC. Sabe-se que esta via está fortemente relacionada com a patologia do Diabetes. Nogueira-
Machado et al[1] descreveram que uma das rotas mais importantes para geração de ROS na patologia
do diabetes envolve a via DAG-PKC.
Os resultados da tabela 4 demonstram que o Resveratrol modulou negativamente a produção
de ROS pela via da Proteína Quinase C. O RSV inibiu a produção de ROS após ativação dessa via
com PDB, com esses resultados sugerimos que após a ativação da PKC, o Resveratrol pode inibir a
atividade da via, inibindo consequentemente a ativação da NADPH oxidase. Dessa forma, ocorre a
diminuição da produção de ROS evitando o estresse oxidativo. O esquema 8, demonstra as duas
possibilidades de modulação direta e indireta da produção de ROS pelo Resveratrol.
Kim Y. et al [61] demonstraram que o Resveratrol inibe a atividade da PKC. Concordantes
também com nossos resultados, os estudos de Jang M. et al [25] também demonstraram que o
Resveratrol inibiu a atividade da PKC em células de camundongos. Outros estudos também
demonstram que a atividade da PKCα e PKCβ pode ser potencialmente inibida pelo Resveratrol
envolvendo o PDB e a ligação de DAG[13].

41
Esquema 8: Modulação do Resveratrol pela PKC, reduzindo a produção de ROS e evitando um possível estresse oxidativo. Adaptado de Nogueira – Machado, et al, 2008
O Resveratrol também é conhecido por modular a produção de oxido nítrico. Em nossos
experimentos observamos que o composto modulou negativamente a produção de NO em diabéticos e
não diabéticos.
Concordantes com nossos resultados foram os achados de Freng Yh et al [14] que observaram
em seu trabalho que o RSV foi capaz de reduzir a produção de NO em macrófagos de camundongo.
Fujii H et al.[62] observaram que o Resveratrol reduz a produção de NO em modelo animal usando
célula renal suína.
Dados semelhantes foram encontrados por Wadsworth, T. L. et al. [63-64] que demonstraram
redução de iNOS em macrófagos de camundongos ao adicionar o polifenol. Bem como Matsuda et al
[65] que encontraram em macrófagos peritoneais de camundongo ativados por LPS, redução da
produção de NO pelo Resveratrol.
Entretanto, Wallerath T. et al [40] demonstraram que o Resveratrol pode induzir a produção de
NO em células de camundongos. Assim como Spanier G et al.[41] que demonstraram perfil ativador de
NO pelo Resveratrol em células da aorta de camundongos.
Com base nesses achados e nos nossos resultados, podemos sugerir que a modulação da
produção de NO pelo Resveratrol pode ocorrer de forma negativa ou positiva dependendo da célula
utilizada.
Isto torna o estudo e as conclusões sobre a ação efetiva do RSV mais complexa. Contudo, a
ação é coerente. Em células associadas à imunidade inata o Resveratrol mostrou efeito inibitório

42
sugerindo modulação do processo inflamatório, visto que o NO é ativador da produção de citocinas pró-
inflamatórias, tal como o INF-gama. Por outro lado, o aumento da produção de NO em células
endoteliais pode estar associado à sua ação benéfica para o sistema circulatório, visto que o NO tem
ação vasodilatadora. Neste contexto, avaliamos o efeito do RSV na produção de VEGF por células
mononucleares.
Os nossos resultados demonstram que o Resveratrol não atuou na inibição da produção de
VEGF, tanto em pacientes diabéticos quanto em não diabéticos. Sugerindo então que a modulação da
produção de VEGF não é realizada pela atividade da PKC. Estudos feitos por Tang em 2008 [66]
demonstraram que o Resveratrol inibe a produção de VEGF em células endoteliais de camundongos.
Ao contrário dos nossos resultados, foi relatado que em células cardíacas de camundongos, o
Resveratrol pode atuar como cardioprotetor, levando ao aumento de VEGF, como demonstrado por
Fuduka S. et al.[67] e D'Amico M. et al.[68].
Em resumo, os nossos resultados mostraram que o Resveratrol atuou na redução da produção
de ROS e Óxido Nítrico de forma semelhante em pacientes diabéticos e não diabéticos, demonstrando
sua capacidade antioxidante e possível ação moduladora da atividade da PKC. Na produção de VEGF
foi possível observar que o Resveratrol não atuou na inibição desse fator de crescimento pela PKC.

43
5 Conclusão
Os nossos resultados em seu conjunto, demonstram que o Resveratrol em testes in vitro promoveu em
células de pacientes diabéticos e não diabéticos a inibição de ROS, fagocitose, síntese de Óxido
Nítrico, uma possível atuação na regulação negativa do sistema PKC/NADPH oxidase e demonstrou
não atuar na inibição de VEGF pela atividade da PKC.

44
6 Sumário de Resultados
1. O Resveratrol reduziu significativamente (p<0,05) a produção de ROS em indivíduos não
diabéticos e diabéticos tipo 2.
2. O Resveratrol exerceu uma ação inibitória, significativa (p<0,05), no processo de fagocitose
medido pela produção de ROS e demonstrou uma maior eficiência em indivíduos não
diabéticos.
3. O Resveratrol foi capaz de modular negativamente a PKC em ambos os grupos demonstrando
uma inibição maior em não diabéticos.
4. O Resveratrol demonstrou a capacidade de inibir a produção de oxido nítrico em ambos os
grupos, porém sendo mais eficiente em não diabéticos.
5. O Resveratrol não atuou na inibição da produção de VEGF através da atividade da PKC em
pacientes diabéticos e não diabéticos.

45
Referências Bibliográficas
1. Nogueira-Machado, J.A. and M.M. Chaves, From hyperglycemia to AGE-RAGE interaction on the cell surface: a dangerous metabolic route for diabetic patients. Expert Opin Ther Targets, 2008. 12(7): p. 871-82.
2. King, G.L., The role of inflammatory cytokines in diabetes and its complications. J Periodontol, 2008. 79(8 Suppl): p. 1527-34.
3. Tillu, D.V., et al., Resveratrol engages AMPK to attenuate ERK and mTOR signaling in sensory neurons and inhibits incision-induced acute and chronic pain. Mol Pain, 2012. 8: p. 5.
4. Hung, L.M., et al., Cardioprotective effect of resveratrol, a natural antioxidant derived from grapes. Cardiovasc Res, 2000. 47(3): p. 549-55.
5. Haider, U.G., et al., Resveratrol suppresses angiotensin II-induced Akt/protein kinase B and p70 S6 kinase phosphorylation and subsequent hypertrophy in rat aortic smooth muscle cells. Mol Pharmacol, 2002. 62(4): p. 772-7.
6. Ignatowicz, E. and W. Baer-Dubowska, Resveratrol, a natural chemopreventive agent against degenerative diseases. Pol J Pharmacol, 2001. 53(6): p. 557-69.
7. de la Lastra, C.A. and I. Villegas, Resveratrol as an anti-inflammatory and anti-aging agent: mechanisms and clinical implications. Mol Nutr Food Res, 2005. 49(5): p. 405-30.
8. Boscolo, P., et al., Effects of resveratrol on lymphocyte proliferation and cytokine release. Ann Clin Lab Sci, 2003. 33(2): p. 226-31.
9. King, G.L. and M.R. Loeken, Hyperglycemia-induced oxidative stress in diabetic complications. Histochem Cell Biol, 2004. 122(4): p. 333-8.
10. Darley-Usmar, V., H. Wiseman, and B. Halliwell, Nitric oxide and oxygen radicals: a question of balance. FEBS Lett, 1995. 369(2-3): p. 131-5.
11. Brownlee, M., Biochemistry and molecular cell biology of diabetic complications. Nature, 2001. 414(6865): p. 813-20.
12. Koya, D. and G.L. King, Protein kinase C activation and the development of diabetic complications. Diabetes, 1998. 47(6): p. 859-66.
13. Slater, S.J., et al., Inhibition of protein kinase C by resveratrol. Biochim Biophys Acta, 2003. 1637(1): p. 59-69.
14. Feng, Y.H., J.P. Zou, and X.Y. Li, Effects of resveratrol and ethanol on production of pro-inflammatory factors from endotoxin activated murine macrophages. Acta Pharmacol Sin, 2002. 23(11): p. 1002-6.
15. Wu, J.M. and T.C. Hsieh, Resveratrol: a cardioprotective substance. Ann N Y Acad Sci, 2011. 1215: p. 16-21.
16. Marletta, M.A., Nitric oxide synthase: aspects concerning structure and catalysis. Cell, 1994. 78(6): p. 927-30.
17. Moncada, S., R.M. Palmer, and E.A. Higgs, Nitric oxide: physiology, pathophysiology, and pharmacology. Pharmacol Rev, 1991. 43(2): p. 109-42.
18. Gelinas, D.S., et al., Immediate and delayed VEGF-mediated NO synthesis in endothelial cells: role of PI3K, PKC and PLC pathways. Br J Pharmacol, 2002. 137(7): p. 1021-30.
19. Paul, B., et al., Occurrence of resveratrol and pterostilbene in age-old darakchasava, an ayurvedic medicine from India. J Ethnopharmacol, 1999. 68(1-3): p. 71-6.
20. Renaud, S. and M. de Lorgeril, Wine, alcohol, platelets, and the French paradox for coronary heart disease. Lancet, 1992. 339(8808): p. 1523-6.
21. Soleas, G.J., E.P. Diamandis, and D.M. Goldberg, Resveratrol: a molecule whose time has come? And gone? Clin Biochem, 1997. 30(2): p. 91-113.
22. Pace-Asciak, C.R., et al., Wines and grape juices as modulators of platelet aggregation in healthy human subjects. Clin Chim Acta, 1996. 246(1-2): p. 163-82.

46
23. Fremont, L., Biological effects of resveratrol. Life Sci, 2000. 66(8): p. 663-73. 24. Fauconneau, B., et al., Comparative study of radical scavenger and antioxidant properties of
phenolic compounds from Vitis vinifera cell cultures using in vitro tests. Life Sci, 1997. 61(21): p. 2103-10.
25. Jang, M., et al., Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science, 1997. 275(5297): p. 218-20.
26. Pace-Asciak, C.R., et al., The red wine phenolics trans-resveratrol and quercetin block human platelet aggregation and eicosanoid synthesis: implications for protection against coronary heart disease. Clin Chim Acta, 1995. 235(2): p. 207-19.
27. Falchetti, R., et al., Effects of resveratrol on human immune cell function. Life Sci, 2001. 70(1): p. 81-96.
28. Buryanovskyy, L., et al., Crystal structure of quinone reductase 2 in complex with resveratrol. Biochemistry, 2004. 43(36): p. 11417-26.
29. Jeandet, P., et al., Metabolic engineering of yeast and plants for the production of the biologically active hydroxystilbene, resveratrol. J Biomed Biotechnol, 2012. 2012: p. 579089.
30. Romero Perez, P., et al., [Epididymal adenomatoid tumor. Review of the national literature and report of a case]. Actas Urol Esp, 1996. 20(9): p. 839-43.
31. Goldberg, D.M., J. Yan, and G.J. Soleas, Absorption of three wine-related polyphenols in three different matrices by healthy subjects. Clin Biochem, 2003. 36(1): p. 79-87.
32. Vitaglione, P., et al., Bioavailability of trans-resveratrol from red wine in humans. Mol Nutr Food Res, 2005. 49(5): p. 495-504.
33. Walle, T., et al., High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab Dispos, 2004. 32(12): p. 1377-82.
34. Gao, X., et al., Immunomodulatory activity of resveratrol: suppression of lymphocyte proliferation, development of cell-mediated cytotoxicity, and cytokine production. Biochem Pharmacol, 2001. 62(9): p. 1299-308.
35. Elattar, T.M. and A.S. Virji, The effect of red wine and its components on growth and proliferation of human oral squamous carcinoma cells. Anticancer Res, 1999. 19(6B): p. 5407-14.
36. Surh, Y.J., et al., Resveratrol, an antioxidant present in red wine, induces apoptosis in human promyelocytic leukemia (HL-60) cells. Cancer Lett, 1999. 140(1-2): p. 1-10.
37. Lopez-Velez, M., F. Martinez-Martinez, and C. Del Valle-Ribes, The study of phenolic compounds as natural antioxidants in wine. Crit Rev Food Sci Nutr, 2003. 43(3): p. 233-44.
38. Gresele, P., et al., Effects of resveratrol and other wine polyphenols on vascular function: an update. J Nutr Biochem, 2011. 22(3): p. 201-11.
39. Lin, M.T., et al., Inhibition of vascular endothelial growth factor-induced angiogenesis by resveratrol through interruption of Src-dependent vascular endothelial cadherin tyrosine phosphorylation. Mol Pharmacol, 2003. 64(5): p. 1029-36.
40. Wallerath, T., et al., Resveratrol, a polyphenolic phytoalexin present in red wine, enhances expression and activity of endothelial nitric oxide synthase. Circulation, 2002. 106(13): p. 1652-8.
41. Spanier, G., et al., Resveratrol reduces endothelial oxidative stress by modulating the gene expression of superoxide dismutase 1 (SOD1), glutathione peroxidase 1 (GPx1) and NADPH oxidase subunit (Nox4). J Physiol Pharmacol, 2009. 60 Suppl 4: p. 111-6.
42. Lotito, S.B. and B. Frei, Dietary flavonoids attenuate tumor necrosis factor alpha-induced adhesion molecule expression in human aortic endothelial cells. Structure-function relationships and activity after first pass metabolism. J Biol Chem, 2006. 281(48): p. 37102-10.
43. Bicalho, H.M.S., et al., A simple techinique simultaneous human leukocyte separation. J Immunol Methods, 1981. v. 40: p. p. 115-116.
.

47
44. Lima, O.A., Silva, D. W., Imunologia imunopatologiaalergia. Editora Guanabara Koogan S. A., 1970.
45. Green, L.C., et al., Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal Biochem, 1982. 126(1): p. 131-8.
46. Folli, F., et al., The role of oxidative stress in the pathogenesis of type 2 diabetes mellitus micro- and macrovascular complications: avenues for a mechanistic-based therapeutic approach. Curr Diabetes Rev, 2011. 7(5): p. 313-24.
47. Maxwell, S.R., et al., Antioxidant status in patients with uncomplicated insulin-dependent and non-insulin-dependent diabetes mellitus. Eur J Clin Invest, 1997. 27(6): p. 484-90.
48. Santini, S.A., et al., Defective plasma antioxidant defenses and enhanced susceptibility to lipid peroxidation in uncomplicated IDDM. Diabetes, 1997. 46(11): p. 1853-8.
49. Maxwell, S., et al., Comparison of antioxidant activity in lipoprotein fractions from insulin-dependent diabetics and healthy controls. Atherosclerosis, 1997. 129(1): p. 89-96.
50. Chang, C.C., et al., Resveratrol retards progression of diabetic nephropathy through modulations of oxidative stress, proinflammatory cytokines, and AMP-activated protein kinase. J Biomed Sci, 2011. 18(1): p. 47.
51. Valko, M., H. Morris, and M.T. Cronin, Metals, toxicity and oxidative stress. Curr Med Chem, 2005. 12(10): p. 1161-208.
52. Kovacic, P. and R. Somanathan, Multifaceted approach to resveratrol bioactivity: Focus on antioxidant action, cell signaling and safety. Oxid Med Cell Longev, 2010. 3(2): p. 86-100.
53. Sharma, S., et al., Resveratrol, a polyphenolic phytoalexin, attenuates diabetic nephropathy in rats. Pharmacology, 2006. 76(2): p. 69-75.
54. Kumar, A., et al., Effects of resveratrol on nerve functions, oxidative stress and DNA fragmentation in experimental diabetic neuropathy. Life Sci, 2007. 80(13): p. 1236-44.
55. Ungvari, Z., et al., Resveratrol attenuates mitochondrial oxidative stress in coronary arterial endothelial cells. Am J Physiol Heart Circ Physiol, 2009. 297(5): p. H1876-81.
56. Palsamy, P. and S. Subramanian, Ameliorative potential of resveratrol on proinflammatory cytokines, hyperglycemia mediated oxidative stress, and pancreatic beta-cell dysfunction in streptozotocin-nicotinamide-induced diabetic rats. J Cell Physiol, 2010. 224(2): p. 423-32.
57. Silan, C., The effects of chronic resveratrol treatment on vascular responsiveness of streptozotocin-induced diabetic rats. Biol Pharm Bull, 2008. 31(5): p. 897-902.
58. Harikumar, K.B. and B.B. Aggarwal, Resveratrol: a multitargeted agent for age-associated chronic diseases. Cell Cycle, 2008. 7(8): p. 1020-35.
59. Leiro, J.M., et al., The anti-inflammatory activity of the polyphenol resveratrol may be partially related to inhibition of tumour necrosis factor-alpha (TNF-alpha) pre-mRNA splicing. Mol Immunol, 2010. 47(5): p. 1114-20.
60. Vetvicka, V. and J. Vetvickova, Combination of glucan, resveratrol and vitamin C demonstrates strong anti-tumor potential. Anticancer Res, 2012. 32(1): p. 81-7.
61. Kim, Y.A., et al., Resveratrol inhibits inducible nitric oxide synthase and cyclooxygenase-2 expression in beta-amyloid-treated C6 glioma cells. Int J Mol Med, 2006. 17(6): p. 1069-75.
62. Fujii, H., et al., Protective effect of grape seed polyphenols against high glucose-induced oxidative stress. Biosci Biotechnol Biochem, 2006. 70(9): p. 2104-11.
63. Tsai, S.H., S.Y. Lin-Shiau, and J.K. Lin, Suppression of nitric oxide synthase and the down-regulation of the activation of NFkappaB in macrophages by resveratrol. Br J Pharmacol, 1999. 126(3): p. 673-80.
64. Wadsworth, T.L. and D.R. Koop, Effects of the wine polyphenolics quercetin and resveratrol on pro-inflammatory cytokine expression in RAW 264.7 macrophages. Biochem Pharmacol, 1999. 57(8): p. 941-9.
65. Matsuda, H., et al., Effects of stilbene constituents from rhubarb on nitric oxide production in lipopolysaccharide-activated macrophages. Bioorg Med Chem Lett, 2000. 10(4): p. 323-7.

48
66. Tang, F.Y., E.P. Chiang, and Y.C. Sun, Resveratrol inhibits heregulin-beta1-mediated matrix metalloproteinase-9 expression and cell invasion in human breast cancer cells. J Nutr Biochem, 2008. 19(5): p. 287-94.
67. Fukuda, S., et al., Resveratrol ameliorates myocardial damage by inducing vascular endothelial growth factor-angiogenesis and tyrosine kinase receptor Flk-1. Cell Biochem Biophys, 2006. 44(1): p. 43-9.
68. D'Amico, M., et al., High glucose induces ventricular instability and increases vasomotor tone in rats. Diabetologia, 2001. 44(4): p. 464-70.




![documentos.camaraaduanera.cl Adj. RES. EX. N... · o Resveratrol 98%: El resveratrol de nombre IUPAC 5-[(E)-2-(4-hydroxyphenyl) ethenyl] benzene- 1,3-diol o también conocido como](https://static.fdocumentos.com/doc/165x107/5ebb24413812114a0d2f747a/adj-res-ex-n-o-resveratrol-98-el-resveratrol-de-nombre-iupac-5-e-2-4-hydroxyphenyl.jpg)













