
Evolução, Genética e Computação - DCA | FEEC ...lboccato/topico_2_inspiracao_biologica.pdf ·...
28
IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux Tópico 2 Evolução, Genética e Computação 1 Evolução, Genética e Computação 1 Base biológica 1.1 Hereditariedade (BURNS, 1983) A história da genética é fascinante principalmente pela velocidade com que se evoluiu de observações a demonstrações experimentais dos mecanismos fundamentais envolvidos. Na antiguidade, defendia-se a idéia de que o sexo era determinado pela procedência do líquido seminal no homem: se do testículo direito sexo masculino; se do testículo esquerdo sexo feminino. A mulher era vista apenas como uma “incubadeira” neste processo.
Transcript of Evolução, Genética e Computação - DCA | FEEC ...lboccato/topico_2_inspiracao_biologica.pdf ·...

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 1
Evolução, Genética e Computação
1 Base biológica
1.1 Hereditariedade (BURNS, 1983)
A história da genética é fascinante principalmente pela velocidade com que se
evoluiu de observações a demonstrações experimentais dos mecanismos
fundamentais envolvidos.
Na antiguidade, defendia-se a idéia de que o sexo era determinado pela
procedência do líquido seminal no homem: se do testículo direito sexo
masculino; se do testículo esquerdo sexo feminino. A mulher era vista apenas
como uma “incubadeira” neste processo.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 2
1672: descoberta do óvulo pelo holandês Graaf as fêmeas de mamíferos
também apresentam ovulação!
1675: descoberta do espermatozóide pelo holandês Von Leeuwenhoeck.
1866: idéias efetivas acerca da hereditariedade monge agostiniano Gregor
Mendel (República Tcheca). Ele atacou o problema de modo simples e lógico,
escolheu material adequado, concentrou-se em poucas características contrastantes,
desenvolveu um programa de cruzamentos controlados, tratou os resultados de
forma eficiente e sugeriu fatores causais (hoje chamados de genes) como sendo os
responsáveis pelos fenômenos observados.
Ninguém havia chegado tão perto da compreensão real da hereditariedade (ficou
faltando, em essência, apenas elucidar os mecanismos celulares envolvidos), mas
foram necessários mais de 30 anos para que a comunidade científica se desse conta
da importância e eficácia desses resultados.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 3
É curioso constatar que um dos fatores que dificultaram a assimilação dos
resultados de Mendel foi justamente o intenso debate reinante nos meios
científicos após a divulgação da teoria de seleção natural de Darwin.
1900: redescoberta das idéias de
Mendel de forma independente por
três pesquisadores.
1a lei de Mendel: Cada caráter é
condicionado por dois fatores. Eles
separam-se na formação dos gametas,
passando apenas um fator por gameta.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 4
1.2 Do Darwinismo ao Neo-Darwinismo
DARWIN (1859) sugeriu que, para haver evolução, é necessário que exista uma
população de indivíduos sujeita a:
1. Reprodução com herança;
2. Variação; e
3. Seleção Natural.
Embora Darwin tenha considerado essas hipóteses como suficientes para explicar
a origem das espécies, hoje elas são aceitas apenas como suficientes para explicar
os processos ecológicos.
Na época de Darwin (meados de 1800), entretanto, sabia-se muito pouco sobre os
processos de reprodução e praticamente nada sobre genética. Além disso, não
havia um bom entendimento sobre como operam os mecanismos de seleção
natural.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 5
A união da genética com diversos conceitos e teorias sobre seleção natural leva ao
que hoje é chamado de Neo-Darwinismo.
Sobre a Seleção Natural
O princípio da seleção natural indica que os indivíduos cujas variações se adaptam
melhor ao ambiente terão maior probabilidade de sobreviver e se reproduzir.
A seleção natural é probabilística, e seu alvo primário é o indivíduo, embora seu
efeito resultante vá se manifestar na espécie como um todo. A espécie é o
beneficiário final do processo evolutivo (MAYR, 1988).
Exemplo clássico: mariposa salpicada (Biston betularia)

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 6
Antes da revolução industrial, a mariposa salpicada clara era comumente
encontrada em regiões urbanas na Grã-Bretanha. Desde 1850, a frequência de
ocorrência da forma escura em torno de regiões bastante industrializadas, como ao
redor de Manchester e Birmingham, aumentou drasticamente de 1 para 90%.
Em regiões menos industrializadas, a mariposa escura continuou sendo rara.
1.3 Terminologia biológica
A terminologia biológica a ser empregada representa uma analogia com as
entidades biológicas reais, sendo que as entidades computacionais corresponderão
invariavelmente a estruturas bem mais simples que seus equivalentes biológicos.
Célula órgão sistema organismo população comunidade
ecossistema

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 7
Célula
Órgão
Sistema
Organismo
Nível fisiológico
População
Comunidade
Ecossistema
Nível ecológico
Figura 1: Divisão hierárquica dos sistemas biológicos (de Castro & Timmis, 2002).
Célula: unidade estrutural básica dos seres vivos, que se compõe de numerosas
partes, sendo as fundamentais a parede ou membrana, o protoplasma e o núcleo. A
célula é a menor unidade de matéria viva que pode existir de maneira
independente, e ser capaz de se reproduzir. Toda célula de um mesmo organismo
contém o mesmo conjunto de um ou mais cromossomos. Nos seres humanos, cada
célula somática (não-germinativa) contém 23 pares de cromossomos.
Cromossomo: estrutura nucleoprotéica formada por uma cadeia de DNA, sendo a
base física dos genes nucleares, os quais estão dispostos linearmente. Cada espécie

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 8
apresenta um número característico de cromossomos. Quando os cromossomos são
arranjados em pares (cada cromossomo proveniente de um dos pais, embora haja
exceções), os respectivos organismos são chamados diplóides. Organismos cujos
cromossomos não se apresentam aos pares são chamados haplóides.
gene gene
gene
Figura 2: Ampliação de um organismo para enfocar o material genético. Organismo célula
cromossomo gene DNA
Genes: blocos funcionais de DNA, os quais codificam uma proteína específica. É a
denominação que damos hoje ao fator mendeliano. Cada gene está localizado em

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 9
uma posição (locus) particular do cromossomo. Quando dois genes se comportam
segundo a 1a lei de Mendel, são ditos alelos, e se encontram no mesmo locus de
dois cromossomos homólogos. Para exemplificar em termos muito simples, é
possível pensar um gene como o responsável pela definição de uma característica
do indivíduo, como a cor dos olhos. As diferentes colorações (azul, castanho etc.)
correspondem a alelos. No entanto, também existem pontos de vista muito mais
refinados (DAWKINS, 1990).
Como muitos organismos apresentam células com mais de um cromossomo, o
conjunto de todos os cromossomos compõe o material genético do organismo,
denominado genoma.
A B A A B A B B A B
Cromossomo diagramático Locus
Alelos: {A, B}
Gene
Figura 3: Ilustração pictórica de um cromossomo salientando o locus, os genes e os alelos.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 10
Epistasia: interferência funcional
entre genes localizados em
posições diferentes.
Exemplo: gene associado à
calvície total é epistático sobre os
genes de coloração de cabelo
(inibe a ação).
pleiotropia: quando um único gene
afeta múltiplas características
fenotípicas.
poligenia: quando uma única
característica fenotípica é
determinada pela interação
simultânea de muitos genes.
gene produto caráter
Figura 4: Pleiotropia e poligenia.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 11
1.4 Fenótipo x Genótipo (FOGEL, 1995)
1909: introdução dos conceitos de genótipo e fenótipo.
Genótipo: representa o conjunto específico de genes do genoma. Neste caso,
indivíduos com o mesmo genoma são ditos terem o mesmo genótipo.
Fenótipo: é a manifestação do genótipo no comportamento, fisiologia e morfologia
do indivíduo, como um produto de sua interação com o ambiente.
A seleção natural opera somente na expressão fenotípica do genótipo (MAYR,
1970; HARTL & CLARK, 1989).
Para modelar este fenômeno, ATMAR (1992) estendeu um resultado da literatura
baseado no emprego de dois espaços de estados:
1. G: espaço genotípico (representa a informação);
2. P: espaço fenotípico (representa o comportamento).
Com G e P definidos, é possível criar quatro funções, onde I representa um
conjunto de efeitos do ambiente (entrada), na forma:

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 12
f1: I G P (epigênese);
f2: P P (seleção);
f3: P G (sobrevivência genotípica);
f4: G G (variação genética).
A adaptação evolutiva ocorre a partir da aplicação iterativa dessas funções.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 13
g1
p1p2
g2g2*
f1
f2
f3
f4Espaço genotípico G
Espaço fenotípico F
Figura 5: Evolução explicada através de mapeamentos entre os espaços genotípico e
fenotípico.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 14
1.5 Adaptação evolutiva
Sob um processo de competição por recursos limitados e seleção natural,
indivíduos mais bem adaptados têm uma maior probabilidade de sobreviver e
propagar seu material genético. A adaptação (fitness) pode ser definida como a
probabilidade de sobrevivência e reprodução (viabilidade), ou então como uma
função do número de descendentes que o organismo produziu (fertilidade).
ATMAR (1994) indicou que uma medida singular da adaptação evolutiva é dada
pela habilidade de uma população antecipar seu ambiente e agir no sentido de
atender aos requisitos associados às predições realizadas.
Uma alternativa a esta definição de medida de adaptação é a idéia simplificada de
que a adaptação pode ser medida em termos de mudanças na freqüência gênica
dentro da população (FISHER, 1930). Esta definição despreza os efeitos importantes
de pleiotropia e poligenia, ao assumir a relação (um gene) (uma característica)
para descrever o relacionamento (genótipo) (fenótipo).

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 15
Sistemas naturais em evolução são fortemente pleiotrópicos e altamente
poligênicos (HARTL & CLARK, 1989). Além disso, a seleção natural atua sobre uma
coleção de características fenotípicas, e não sobre uma única característica isolada.
Como conseqüência, um gene não pode ser considerado como uma unidade
individual de seleção, pelo fato de não ser possível atribuir a um gene isolado um
valor significativo para o seu nível de adaptação.
Logo, a evolução se refere mais apropriadamente a mudanças em adaptação e
diversidade, e não diretamente a mudanças na freqüência gênica.
Além disso, a evolução é oportunista, ou seja, características críticas serão
intensamente otimizadas, enquanto características menos críticas podem sofrer
uma grande variedade de tratamentos alternativos.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 16
1.6 Reprodução assexuada e sexuada
Embora os detalhes da evolução biológica ainda não estejam completamente
compreendidos, existem alguns aspectos fundamentados em fortes evidências
experimentais:
1. a evolução é um processo que opera sobre cromossomos e não sobre
organismos;
2. a seleção natural é o mecanismo que relaciona cromossomos com a eficiência
da entidade que eles representam, permitindo assim que organismos mais bem
adaptados ao meio ambiente tenham uma taxa maior de reprodução;
3. o processo evolutivo se dá durante o estágio reprodutivo. Dentre os fenômenos
que podem ocorrer neste estágio é possível citar a mutação e a recombinação
genética (crossover);
4. o processo evolutivo não possui memória direta.
IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 17
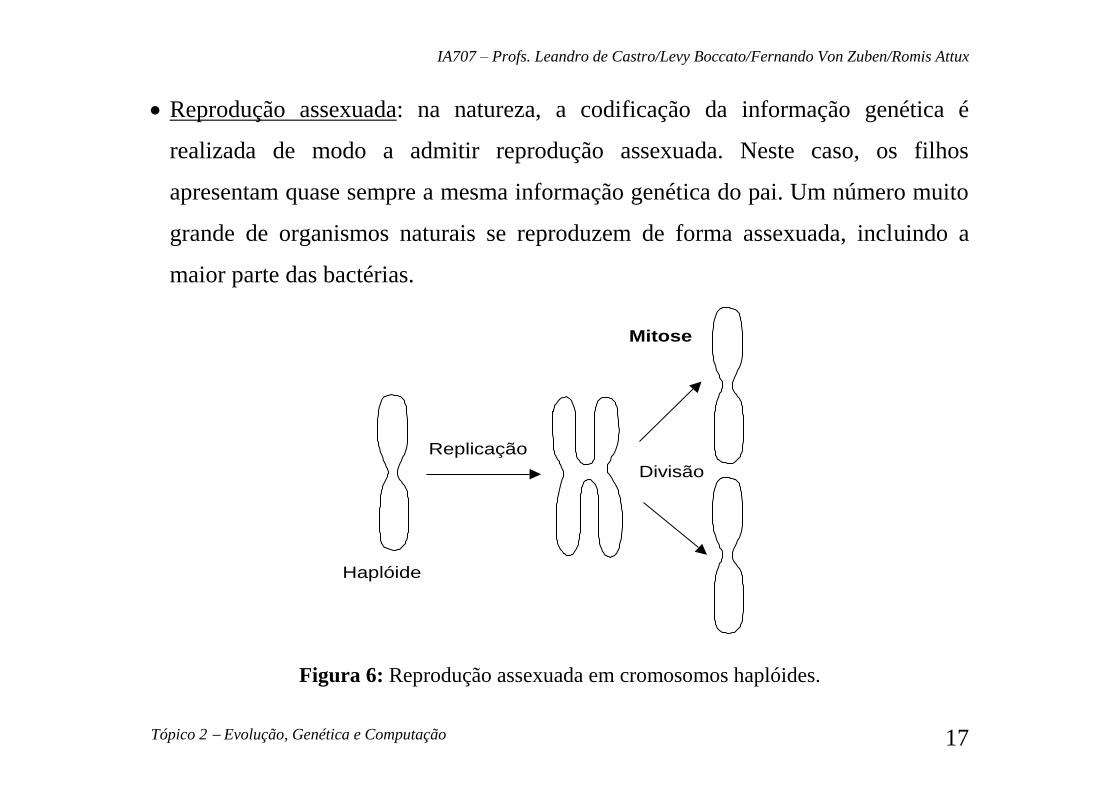
Reprodução assexuada: na natureza, a codificação da informação genética é
realizada de modo a admitir reprodução assexuada. Neste caso, os filhos
apresentam quase sempre a mesma informação genética do pai. Um número muito
grande de organismos naturais se reproduzem de forma assexuada, incluindo a
maior parte das bactérias.
Replicação
Haplóide
Divisão
Mitose
Figura 6: Reprodução assexuada em cromosomos haplóides.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 18
Reprodução sexuada: neste caso, os filhos apresentam uma informação genética
que corresponde a uma combinação da informação genética de seus pais. Na
reprodução sexuada diplóide, cada pai fornece uma célula haplóide denominada
gameta, formada por cromossomos resultantes da recombinação dos pares de
cromossomos existentes nos respectivos pais. Os cromossomos presentes nos
gametas fornecidos pelos pais se emparelham para gerar uma nova célula diplóide.
Replicação
Diplóide
Separação dos homólogos
Meiose
Ovo
Esperma
Zigoto diplóide
Gametas haplóides

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 19
Durante a reprodução sexuada, os cromossomos estão sujeitos à recombinação.
Recombinação ou crossover: consiste na troca aleatória de material genético entre
dois cromossomos.
Figura 7: Recombinação genética (crossover) entre dois cromossomos.
1.7 Mutação
Tanto na reprodução assexuada como na sexuada, o processo de cópia do material
genético dos pais está sujeito a erros, denominados mutações. Uma mutação
corresponde então a uma troca aleatória, no processo de cópia do material
genético, de um nucleotídeo por outro.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 20
Nucleotídeo: unidade elementar do DNA.
Figura 8: Ilustração de diferentes tipos de mutação.
A mutação é uma conseqüência inescapável da existência de sistemas que se
reproduzem continuamente em um universo com diferencial de entropia positivo,
sendo um mecanismo importante de implantação da diversidade entre os
indivíduos.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 21
Em estágios iniciais do processo evolutivo natural, as taxas de mutação podem ter
sido muito maiores. O fato de que as taxas de mutação atuais nos organismos vivos
são muito baixas na maioria dos casos observados pode ser devido a estes
organismos, como espécie, terem atingido um estado de estacionariedade em
relação às condições ambientais atuais. Neste caso, o nível de evolução orgânica
atual não deve servir de referência para nenhum tipo de implementação
computacional de algoritmo evolutivo.
Geralmente, a recombinação deve ter probabilidades de ocorrência muito maior
que a mutação.
Observação: em implementação computacional, à primeira geração de indivíduos é
geralmente imposto um determinado nível de diversidade.
Teoricamente, as taxas ótimas de mutação são inversamente proporcionais ao
número de variáveis de decisão envolvidas e diretamente proporcionais à distância
do ótimo. A probabilidade de ocorrência de mutação deve ser inversamente

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 22
proporcional ao tamanho da população (DE JONG, 1975). Existem estratégias que
incorporam a definição da taxa de mutação ao próprio genótipo.
2 A notação computacional
Obs: as informações contidas nesta seção podem não ser genéricas e também
podem referir-se ao caso de algoritmos evolutivos específicos.
Um cromossomo haplóide geralmente corresponde a uma estrutura de dados (e.g.,
uma cadeia de bits) que representa um candidato à solução (indivíduo de uma
população) de um problema.
Um gene corresponde a um único bit, ou, então, a um pequeno bloco de bits
adjacentes que codificam um elemento particular da solução candidata. Por
exemplo, em otimização de funções multiparamétricas, o conjunto de bits utilizado
para codificar um parâmetro específico pode ser considerado um único gene.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 23
Em uma cadeia de bits, um alelo é 0 ou 1. Para alfabetos maiores, mais alelos
podem ser considerados para cada posição (locus).
A grande maioria das aplicações de algoritmos evolutivos emprega codificação
haplóide e baseada em um único cromossomo. Neste caso, o genótipo passa a
corresponder à configuração de bits do cromossomo.
1 0 0 0 1 1 0 0 1 0
Cromossomo utilizando representação binária
Locus
Alelos: {0,1}
Gene
Figura 9: Estrutura de dados (cadeia de bits ou binária) de um algoritmo evolutivo.
2.1 Superfície de adaptação
Uma vez definido o processo de codificação (representação) para um dado
problema, o objetivo é buscar uma solução desejada dentre todos os possíveis
candidatos à solução (elementos do espaço de busca).

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 24
Genótipo: uma solução potencial de um dado problema pode ser codificada na
forma de uma ou mais cadeias cromossômicas (estrutura de dados).
Fenótipo: a decodificação de um ou mais cromossomos leva a um ponto na
superfície de adaptação, definida no espaço de busca.
Espaço de busca normado: coleção de candidatos à solução do problema + uma
norma (operador que estabelece a noção de vizinhança).
Nota: os algoritmos evolutivos geralmente assumem que existe alguma correlação
entre candidatos à solução que sejam vizinhos (segundo a norma estabelecida).
Assumindo representação haplóide e baseada em um único cromossomo, e
considerando genes correspondentes a um único bit, o número de possíveis
candidatos à solução em função do comprimento do cromossomo é dado nas
figuras a seguir:

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 25
0 2 4 6 8 100
200
400
600
800
1000
1200
comprimento
candidatos
10 12 14 16 18 200
2
4
6
8
10
12x 10
5
comprimento
candidatos
Superfície de adaptação: foi originalmente definida pelo biólogo Sewell Wright,
em 1931, como a representação do nível de adaptação de todos os genótipos.
Observe que aqui é realizada uma avaliação diretamente sobre o genótipo, sem
considerar o fenótipo.
Exemplo: suponha que cada genótipo é uma cadeia de bits de comprimento p, e
que a distância entre dois genótipos é dada pela distância de Hamming, ou seja, o
número de posições cujos bits são diferentes. Também suponha que a cada
genótipo pode ser atribuído um nível de adaptação. Neste caso, a superfície de

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 26
adaptação vai ser um mapeamento de dimensão p+1, que associa a cada genótipo
no espaço de dimensão p um nível de adaptação definido no eixo p+1.
No contexto de superfícies de adaptação, a evolução é um processo responsável
pela movimentação da população nesta superfície, a partir de operadores genéticos
como mutação e recombinação, no sentido de conduzir os indivíduos para as
regiões de máximo local. Neste caso, a seleção natural pode ser vista como um
processo que conduz à maturação ou aumento da adaptação média da população.
Obs: a idéia de evolução a partir do deslocamento da população ao longo de uma
superfície de adaptação fixa é biologicamente irreal, porque um indivíduo da
população não pode receber um valor de adaptação independente dos demais.
O nível médio de adaptação de um indivíduo em um ambiente não-estacionário
varia em função da dinâmica do ambiente e das interações deste indivíduo e de
outros com o ambiente e entre si.

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 27
3 Da Biologia Evolutiva para a Computação Evolutiva
Biologia
(Genética)
Algoritmos
Evolutivos
Cromossomo Estrutura de dados
Gene Um ou mais atributos ocupando uma determinada posição da
estrutura de dados
Locus Posição ocupada por um gene em uma estrutura de dados
Alelo Diferentes valores (dados) que um gene pode assumir
Crossover Troca de porções entre duas ou mais estruturas de dados
Mutação Variação aleatória em uma estrutura de dados individual
Fitness Valor que quantifica a qualidade relativa (nível de adaptação) de
um indivíduo dada a função de avaliação
Seleção Processo que permite a sobrevivência e reprodução dos
indivíduos mais aptos em detrimento dos menos aptos
Genótipo Estrutura de dados que representa uma solução candidata a um
determinado problema
Fenótipo Decodificação de um ou mais cromossomos que leva a um ponto
da superfície de adaptação, definida no espaço de busca

IA707 – Profs. Leandro de Castro/Levy Boccato/Fernando Von Zuben/Romis Attux
Tópico 2 Evolução, Genética e Computação 28
4 Bibliografia
ATMAR, W. On the Rules and Nature of Simulated Evolutionary Programming. Proc. of the First Ann. Conf. On Evolutionary
Programming, pp. 17-26, 1992.
ATMAR, W. Notes on the Simulation of Evolution. IEEE Trans. on Neural Networks, vol. 5, no. 1, pp. 130-148, 1994.
BERGSON, H. A Evolução Criadora, Editora da Unesp, 2010.
BURNS, G.W. The Science of Genetics - An Introduction of heredity. 1983.
DARWIN, C. The Origin of Species, John Murray, 1859 (Penguin Classics, 1985).
DAWKINS, R. The Selfish Gene, Oxford University Press, 1990.
DE CASTRO, L. N. & TIMMIS, J. I.. Artificial Immune Systems: A New Computational Intelligence Approach, Springer
Verlag, 2002 C
DE JONG, K.A. An analysis of the behavior of a class of genetic adaptive systems. Ph.D. Thesis, The University of Michigan,
1975.
FISHER, R.A. The Genetical Theory of Natural Selection. Clarendon Press, 1930.
FOGEL, D.B. Evolutionary Computation: Toward a New Philosophy of Machine Intelligence, IEEE Press, 1995.
HARTL, D.L., CLARK, A.G. Principles of Population Genetics. Sinauer, 1989.
MAYR, E. Populations, Species, and Evolution. Belknap Press, 1970.
MAYR, E. Toward a New Philosophy of Biology: Observations of an Evolutionist. Belknap Press, 1988.
MITCHELL, M. An Introduction to Genetic Algorithms. MIT Press, 1996.


















