
GENES DE VIRULÊNCIA EM Salmonella enterica · das doenças infecciosas dos ... potenciais...
43
UNIVERSIDADE FEDERAL DE GOIÁS ESCOLA DE VETERINÁRIA E ZOOTECNIA PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL Disciplina: SEMINÁRIOS APLICADOS GENES DE VIRULÊNCIA EM Salmonella enterica: Descrição, componentes estruturais e métodos de detecção Adriana Marques Faria Orientador: Prof. Dr. Guido Fontgalland Coelho Linhares GOIÂNIA 2013
Transcript of GENES DE VIRULÊNCIA EM Salmonella enterica · das doenças infecciosas dos ... potenciais...

UNIVERSIDADE FEDERAL DE GOIÁS
ESCOLA DE VETERINÁRIA E ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL
Disciplina: SEMINÁRIOS APLICADOS
GENES DE VIRULÊNCIA EM Salmonella enterica:
Descrição, componentes estruturais e métodos de detecção
Adriana Marques Faria
Orientador: Prof. Dr. Guido Fontgalland Coelho Linhares
GOIÂNIA
2013

ii
ADRIANA MARQUES FARIA
GENES DE VIRULÊNCIA EM Salmonella enterica:
Descrição, componentes estruturais e métodos de detecção
Seminário apresentado junto à
Disciplina Seminários Aplicados do
Programa de Pós-Graduação em
Ciência Animal da Escola de Veterinária e Zootecnia da
Universidade Federal de Goiás.
Nível: Doutorado
Área de concentração:
Sanidade Animal, Higiene e Tecnologia de Alimentos
Linha de Pesquisa:
Etiopatogenia, epidemiologia, diagnóstico e controle
das doenças infecciosas dos animais
Orientador:
Prof. Dr. Guido Fontgalland Coelho Linhares
Comitê de Orientação:
Prof.ª Dr.ª Maria Auxiliadora de Andrade - EVZ /UFG
Prof. ª Dr.ª Moema Pacheco Chediak Matos - EVZ /UFG
GOIÂNIA
2013

iii
SUMÁRIO
1. INTRODUÇÃO............................................................................................. 1
2. REVISÃO DE LITERATURA........................................................................
2.1 Estrutura celular bacteriana.......................................................................
3
3
2.1.1 Genótipo e fenótipo de Salmonella enterica........................................... 5
2.2 Resistência de Salmonella a agentes antimicrobianos e biocidadas......... 6
2.3 Genes de Virulência.................................................................................. 9
2.3.1 Ilhas de patogenicidade.......................................................................... 10
2.3.2 Plasmídeos............................................................................................. 12
2.3.3 Transposons ..........................................................................................
2.3.4 Integrons ................................................................................................
15
16
2.3.5 Genes cassete........................................................................................
2.4 Métodos de detecção de genes de virulência............................................
2.4.1 Reação em Cadeia da Polimerase.........................................................
2.4.2 Hibridização de ácidos nucleicos............................................................
I. Southern Blotting..........................................................................................
II. Microarray....................................................................................................
2.4.3 Outros métodos de detecção de genes de virulência ...........................
I. Fingerprinting................................................................................................
II. Tipificação de sequência de multilocus.......................................................
III. Dendograma..............................................................................................
IV.Sequência genômica...................................................................................
18
19
19
22
22
24
25
25
25
26
27
3. CONSIDERAÇÕES FINAIS......................................................................... 28
REFERÊNCIAS BIBLIOGRÁFICAS................................................................. 29

iv
LISTA DE FIGURAS
FIGURA 1: Estruturas de uma célula bacteriana Gram negativa típica.
Presença de cápsula, parede celular, membrana celular,
citoplasma, nucleoide, fímbrias, flagelos, ribossomos e plasmídeo.
......................................................................................................... 2
FIGURA 2: Representação esquemática dos métodos de transferência gênica
horizontal entre bactérias. Transdução e transformação,
Conjugação; Transposons (genes saltadores) e Conjugação......... 5
FIGURA 3: Representação esquemática dos principais modos de ação de
antibióticos sobre uma célula bacteriana...................................... 7
FIGURA 4: Representação esquemática ilustrando a quantidade de genes
encontrados em Salmonella Typhi. A cada gene, é atribuída uma
cor de acordo com a função exercida na célula bacteriana............9
FIGURA 5: Representação esquemática do cromossomo de Salmonella
enterica. Ilhas de patogenicidade (SPI) são mostradas fora do
cromossomo enquanto genes de virulência e reguladores
transcricionais são mostrados na parte interna............................. 11
FIGURA 6: A: Representação esquemática de célula bacteriana, plasmídeos e
seus respectivos métodos de resistência a antimicrobianos. be –
Bomba de efluxo; ed- Enzimas degradam o antimicrobiano; el-
Enzimas se ligam ao antimicrobiano e inibem sua ação. Fonte:
Adaptada de scienceblogs.com.br. B: Representação esquemática
de vários genes e funções em um plasmídeo, cada cor representa
um gene diferente...........................................................................14
FIGURA 7: Representação esquemática de um transposon codificando um
gene bacteriano e após a ação de uma enzima específica, o
mesmo é realocado em outra posição na mesma célula bacteriana.
....................................................................................................... 15

v
FIGURA 8: Representação esquemática da conformação de integron e genes
cassete........................................................................................... 17
FIGURA 9: Representação esquemática de Reação em cadeia da polimerase
demonstrando as fases de desnaturação, anelamento, extensão e
amplificação da região de interesse.............................................. 20
FIGURA 10: Gel de agarose corado demonstrando perfil plasmidial de vários
sorotipos de Salmonella ............................................................... 21
FIGURA 11: Representação esquemática da técnica de Southern blotting
demonstrando as etapas necessárias para a sua realização.
Extração de DNA; digestão de DNA; preparo da sonda;
transferência; hibridização e detecção do DNA............................. 23
FIGURA 12: Resultado de Southern blotting de plasmídeos de virulência de
Salmonella Cholerasuis isoladas de humanos e
suínos.......................................................................................... 23
FIGURA 13: Representação esquemática de Microarray de genes de virulência e
resistência de Salmonella spp. e E. coli. Cada marcação determina um
gene padrão..................................................................................... 24
FIGURA 14: Tipificação de sequência multilocus (MLST), cada quadrado
branco condiz com a presença de um gene alvo determinado na
abscissa......................................................................................... 26
FIGURA 15: Dendograma de Salmonella Javiana. Presença de genes
semelhantes (linhas vermelhas) ou distintos a partir do peso
molecular em plasmídeos. Presença de resistência a
antimicrobianos testados (quadrado vermelho), baixa
suscetibilidade a antimicrobianos testados (quadrado verde) e
suscetibilidade plena a antimicrobianos testados (azul claro).
Presença de genes de virulência, amarelo presente, azul ausente.
....................................................................................................... 27

vi
LISTA DE QUADROS
QUADRO 1: Genes essenciais/fisiológicos e genes de virulência/adaptativos
que podem ser encontrados em Salmonella enterica............... 10

1. INTRODUÇÃO
A crescente problemática de agentes patogênicos resistentes e com
acentuada virulência é uma realidade mundial nos dias de hoje. Bactérias da
Família Enterobacteriaceae representam e demandam muitos estudos com o
intuito de entender os seus métodos de ação e o porquê da sua presença ser tão
deletéria aos homens e animais.
Quando se estuda os patógenos bacterianos, é possível descobrir que
estes possuem múltiplos fatores que em conjunto são essenciais à capacidade de
infectar e causar injúria sobre seus respectivos hospedeiros. Na verdade, a
sobrevivência da bactéria parece depender de um balanço entre muitos produtos
de genes atuando de maneira simultânea. A ausência ou presença de certos
genes está totalmente relacionada a diversos mecanismos essenciais ou
adaptativos às bactérias. Tais estruturas podem ser encontradas em plasmídeos
ou intimamente no próprio cromossomo da bactéria. Alguns genes possuem
funções fisiológicas para o microrganismo, enquanto outros determinam fatores
de virulência que são fundamentais à sobrevivência, invasão e permanência da
bactéria em condições adversas no hospedeiro e no ambiente (1).
O gênero Salmonella é composto por bactérias muito importantes na
área de veterinária e de saúde pública, pois é responsável por doença em animais
e humanos, e extensas perdas econômicas na produção animal. Também é
extensivamente estudada em termos de fisiologia, genética e estrutura celular,
sendo uma das bactérias patogênicas melhor caracterizadas em termos de
virulência. A virulência da Salmonella, e seus respectivos sorotipos, é multifatorial
e complexa, incluindo presença de estruturas como fímbrias e flagelos, bem como
sua habilidade de penetrar e replicar nas células epiteliais (2).
Cada característica da bactéria é determinada por um gene específico,
seja a própria respiração celular ou resistência a antimicrobianos. O estudo de
genes que conferem resistência e características que denominam as bactérias
como virulentas tem como intuito entender a função de cada gene, tornando
possível também determinar o meio de aquisição deste entre as bactérias (3).
Com os avanços nos métodos de diagnóstico, é possível determinar
toda a sequência genética de microrganismos e populações bacterianas de
maneira geral (4). O estudo de métodos para detecção da presença de fatores e

2
estruturas determinantes de virulência em Salmonella constitui um campo
promissor de pesquisa com diversas técnicas sendo utilizadas com relativo
sucesso (5).
Para compreender a virulência de bactérias e sua importância é válido
entender quais seriam os mecanismos de ação, como surgiram e como fazer o
seu diagnóstico. Logo, pretende-se fazer uma revisão de literatura sobre a
importância dos genes de virulência em Salmonella enterica, descrevendo suas
estruturas e os métodos de detecção mais utilizados.

3
2. REVISÃO DE LITERATURA
2.1 Estrutura celular bacteriana
As bactérias são organismos relativamente simples, unicelulares de
diferente conformação estrutural, onde o seu material genético não está envolto
por uma membrana nuclear e, portanto, são seres procariotos (pré-núcleo) (6). De
maneira geral, uma célula bacteriana típica é composta de uma cápsula, parede
celular, membrana celular, citoplasma contendo material nuclear e estruturas
acessórias como flagelo, fímbrias e plasmídeos (Figura 1) (2).
FIGURA 1: Estruturas de uma célula bacteriana Gram negativa típica. Presença de cápsula, parede celular, membrana celular, citoplasma, nucleoide, fímbrias, flagelos, ribossomos e plasmídeo. Fonte: Adaptado de TORTORA 6 et al. (2012).
A presença de estruturas como cápsula, fímbrias e plasmídeos é
importante na evolução de bactérias, visto que determinam maior capacidade de
sobrevivência no ambiente, motilidade e características de adaptação e
resistência, respectivamente. A partir do conhecimento de algumas características
estruturais de bactérias é possível determinar particularidades que são
fundamentais na determinação da patogenicidade e também são auxiliares no
diagnóstico laboratorial de infecções (2).
O genótipo de uma bactéria representa as propriedades potenciais de
um determinado organismo por meio da composição genética que codifica todas
as suas características particulares. Ainda, representa as características
potenciais delineadas por uma coleção de genes presentes em seu DNA. Quanto
CITOPLASMA
RIBOSSOMOS
NUCLEOIDE
FLAGELO
PLASMIDEO CÁPSULA
PAREDE CELULAR
MEMBRANA
PLASMÁTICA
FÍMBRIAS

4
ao fenótipo, refere-se às propriedades verdadeiramente expressas, resultado
direto da manifestação do genótipo. Em termos moleculares, o fenótipo
representa a coleção de proteínas da qual provém a maior parte das
características da célula (6).
As bactérias podem transferir seu material genético de duas maneiras:
vertical e horizontal. Na transferência gênica vertical, os genes são passados de
um microrganismo para seus descendentes. Na transferência gênica horizontal,
os genes podem ser adquiridos de outros microrganismos da mesma geração.
Esse fenômeno envolve uma célula doadora que contribui parte de seu genoma
para uma célula receptora, que pode ser de uma espécie ou até mesmo de um
gênero diferente (7). Após a transferência, parte do DNA da doadora é
incorporado ao DNA da receptora, que passa a ser denominada de recombinante
(8).
Alguns genes adquiridos horizontalmente podem proporcionar efeitos
deletérios à célula bacteriana que os recebeu. Dessa maneira, essa bactéria será
eliminada da população ao qual está inserida, da mesma forma que mutações
deletérias são perdidas, já que são eventos prejudiciais a sobrevivência do
microrganismo. Por outro lado, genes que conferem ao patógeno vantagem
seletiva em relação ao hospedeiro têm o potencial de serem rapidamente
espalhados dentro da população bacteriana (9).
Para que haja a transferência gênica horizontal existem três
mecanismos a citar: transdução, transformação e conjugação (6) (Figura 2). Além
disso, existem mecanismos adicionais para a modificação genética das bactérias,
os chamados elementos genéticos móveis, cuja presença é associada a genes
codificadores de funções que podem proporcionar vantagem para a sobrevivência
dos microrganismos (10). Plasmídeos, transposons e integrons são os elementos
genéticos móveis de maior interesse na genética bacteriana (11).

5
FIGURA 2: Representação esquemática dos métodos de transferência gênica horizontal entre bactérias. Transdução e transformação, Conjugação; Transposons (genes saltadores) e Conjugação. Fonte: www.studentconsult.com66.
2.1.1 Genótipo e fenótipo de Salmonella enterica
A família Enterobacteriaceae é constituída por membros bastante
semelhantes entre si, apresentando parede celular típica de organismos Gram
negativos constituída interna e externamente por membranas separadas por
glicopeptídeo, com a presença de várias proteínas que diferenciam de acordo
com gênero, espécie e subespécie. O gênero Salmonella é bastante estudado
dentro desta família, onde seus representantes são espécies que apresentam
motilidade, havendo poucas variantes imóveis. Resistem à dessecação e ao
congelamento, podendo sobreviver no ambiente em períodos que variam de
meses a anos (12).
A variação genética em Salmonella sp. está relacionada à codificação
de estruturas como os lipopolissacarídeos, flagelos e fímbrias, bem como a
TRANSDUÇÃO E
TRANSFORMAÇÃO
CONJUGAÇÃO CONJUGAÇÃO

6
expressão de genes de virulência específicos que alteram a fisiologia celular ou
protegem o patógeno das defesas do hospedeiro. Portanto, a pressão seletiva
para promover o polimorfismo genético responsável por variação antigênica
desses microrganismos é consequência direta da presença das estruturas de
superfície, já que são alvos do sistema imune do hospedeiro (13).
A variabilidade fenotípica observada em cepas de Salmonella enterica
é determinada por mecanismos biológicos implícitos em seu genoma e sua
repercussão na epidemiologia da doença causada por este patógeno é
frequentemente objeto de estudos (14).
Nesse sentido, a recombinação genética, que se refere à troca de
genes entre duas moléculas de DNA para formar novas combinações em um
cromossomo, é considerada a fonte da variação evolutiva da maioria dos
procariotos (6). Estima-se que aproximadamente 25% do genoma de uma
bactéria são obtidos de outra célula, podendo esta ser da mesma espécie ou não.
Além disso, esse evento possui papel essencial na preservação da integridade
genética por meio da reparação de possíveis falhas no DNA bacteriano (8).
Em determinadas espécies de bactérias os genes codificadores de
funções específicas são altamente propensos à recombinação e transferência
gênica horizontal de modo que antígenos similares podem ser encontrados em
cepas distantemente relacionadas (15). No gênero Salmonella, a presença de tais
elementos é frequentemente estudada e associada a genes de resistência a
antimicrobianos, sendo geralmente referida como ilhas de resistência
antimicrobiana. Desta forma são importantes indicativos do desenvolvimento e
distribuição desse mecanismo entre os diversos sorotipos de Salmonella (16).
2.2 Resistência de Salmonella a agentes antimicrobianos e biocidas
Os antimicrobianos são metabólitos de origem microbiana com baixo
peso molecular, que têm a capacidade de, em pequenas doses, inibir o
crescimento ou destruir microrganismos causadores de doenças. Os efeitos
bacteriostático e bactericida são determinados por mecanismos de ação, sendo

7
eles exercidos essencialmente por inibição da síntese da parede celular, inibição
da duplicação cromossômica ou transcrição, alterações na permeabilidade da
membrana citoplasmática, interferência na replicação dos cromossomos e
interferência na síntese proteica (6) (Figura 3).
FIGURA 3: Representação esquemática dos principais modos de ação de antibióticos sobre uma célula bacteriana. Fonte: http://revistaescola.abril.com.br67.
A atividade de agentes antimicrobianos é influenciada pelo sítio de
ação, taxa de absorção e poder de penetração na célula e pelo metabolismo do
agente específico. Por isso, a atividade de determinado agente antibacteriano
pode ser totalmente afetada pela interação entre patógeno e droga, bem como
entre o hospedeiro e o patógeno (2).
No tratamento e controle de infecções por Salmonella spp., os
principais agentes antimicrobianos utilizados são as fluoroquinolonas e terceira
geração de cefalosporinas. O aparecimento de resistência a tais compostos leva a
grande preocupação, visto que para o gênero há relatos de resistência à maioria
dos antimicrobianos disponíveis (17), limitando as possibilidades de fármacos
eficazes frente a Salmonella spp., gerando portanto, um problema de alta
relevância em saúde pública (18).

8
Diferentemente de antimicrobianos, que atuam sobre um processo
fisiológico ou estrutura da bactéria, os agentes biocidas possuem mais de um sítio
de ação normalmente. No entanto, essa potencialidade pode ser reduzida em
caso de concentrações subinibitórias. Se um antimicrobiano é limitado a um sítio
de ação apenas, e um processo adaptativo da célula bloqueia o acesso a este, há
o aparecimento de resistência para com determinado agente (19). Os diferentes
mecanismos de ação de agentes biocidas são variáveis de acordo com a
estrutura química deste e da composição bacteriana, podendo-se elencar como
principais mecanismos: Interação com componentes celulares externos; Interação
com a membrana citoplasmática e interação com constituintes citoplasmáticos. Os
biocidas podem atuar em um nível de interação ou em vários com a célula
bacteriana, a fim de produzir atividade antimicrobiana (20).
A resistência bacteriana aos biocidas já fora descrita em 1950 e 1960,
e nos últimos anos vem aumentando. Resposta bacteriana a biocidas é
determinada essencialmente pela natureza de agentes químicos e o tipo de
organismo envolvido. Outros fatores como temperatura, contato, pH ambiental e
presença de matéria orgânica, podem exercer considerável efeito sobre as
propriedades do agente empregado. Os mecanismos de resistência ou tolerância
aos biocidas são relacionados a características estruturais, intrínsecas ou
adquiridas pelas bactérias (21).
A resistência aos compostos de amônio quaternário, um biocida
amplamente utilizado na desinfecção de ambientes, é relatada para diversos
sorotipos de Salmonella enterica (17). Entretanto alguns biocidas que são
amplamente utilizados como Cloreto de benzalcônico ou Iodofor ainda são
eficazes na desinfecção de instrumentos e instalações (22).
Os relatos de resistência a agentes antimicrobianos e biocidas em
Salmonella spp., entre outras bactérias, estão relacionados à existência de genes
específicos que podem determinar diferentes habilidades a célula bacteriana. Tais
genes são denominados genes de resistência e fazem parte dos chamados genes
virulência. O conhecimento sobre sua localização, composição e caracterização
são alvos de muitas pesquisas na última década (6, 7, 10, 11, 15).

9
2.3 Genes de virulência em Salmonella enterica
Genes são segmentos de DNA que codificam produtos funcionais. Os
genes estão organizados em arranjo linear dentro do cromossomo de um
organismo, conhecido como locus genético. Cada gene possui uma função
específica que pode ou não ser expressa, a depender de inúmeros fatores
intrínsecos ao próprio DNA. As bactérias, que são microrganismos de menor
complexidade genética, possuem algo em torno de 500 genes compondo seu
DNA (3) (Figura 4).
FIGURA 4: Representação esquemática ilustrando a
quantidade de genes encontrados em Salmonella Typhi. A cada gene, é atribuída uma cor de acordo com a função exercida na célula bacteriana. Fonte: LANGRIDGE24 et al., 2009.
O DNA de vários sorotipos de Salmonella enterica já foi sequenciado e
caracterizado por métodos bioquímicos e moleculares, portanto a maioria dos
genes é descrita e também se conhece em grande parte as funções que cada
gene determina para a bactéria (23). Alguns genes possuem funções necessárias
à sobrevivência da bactéria, pois codificam proteínas funcionais para a
respiração, locomoção, aquisição de nutrientes entre outras. Outros genes podem

10
estar presentes na bactéria para determinar funções que definem vantagens
evolutivas e competitivas para o microrganismo, como invasão da célula
hospedeira, resistência a determinados antimicrobianos e enterotoxinas (Quadro
1) (24).
QUADRO 1: Genes essenciais/fisiológicos e genes de virulência/adaptativos que podem
ser encontrados em Salmonella enterica. Fonte: Adaptado de
http://www.salmonella.org68.
GENES ESSENCIAIS/ FISIOLÓGICOS GENES DE VIRULÊNCIA/ADAPTATIVOS
dnaB- síntese de DNA InvD- invasão da célula
In1-A- fermentação bacteriana mviM- virulência no hospedeiro
nit- metabolismo de Nitrogênio pho-P- invasão de macrófagos
sinR- inserção da bactéria sapA- resistência a antimicrobianos
fimI- fímbria spvB- plasmídeo de virulência
flgA- flagelo stx- enterotoxina
Os genes de virulência podem estar presentes em diversas regiões do
cromossomo e/ou elementos genéticos móveis da bactéria, sendo responsáveis
pela codificação de produtos e propriedades que podem determinar a
patogenicidade e consequentemente a virulência de Salmonella spp. (25). As
regiões e elementos mais importantes na compreensão dos genes de virulência
serão abordados a seguir.
2.3.1 Ilhas de patogenicidade
Ilhas de patogenicidade são ilhas genômicas, constituídas por amplas
regiões cromossômicas (10.000 a 200.000 pares de bases) de alta instabilidade e
com características distintas do restante do genoma bacteriano (Figura 5). Estas
são diferentes do restante do cromossomo e estão comumente associadas a
genes que codificam RNA transportador. Em suas extremidades possuem regiões
denominadas Hot spots, que são elementos envolvidos na mobilidade genética
(26).

11
FIGURA 5: Representação esquemática do cromossomo de Salmonella
enterica. Ilhas de patogenicidade (SPI) são mostradas fora do cromossomo enquanto genes de virulência e reguladores transcricionais são mostrados na parte interna. Fonte: MARCUS1 et al. (2000).
Nas ilhas de patogenicidade estão presentes vários genes que podem
contribuir para um determinado fenótipo de virulência, o qual é manifestado em
um período específico durante o curso da infecção. A aquisição de uma ilha de
patogenicidade por um organismo não garante a sua transformação em um
patógeno de fato, a virulência é determinada não só pelo microrganismo, mas
também pelo hospedeiro suscetível (1).
Existem 17 ilhas de patogenicidade reconhecidas no gênero
Salmonella, a SPI-1 é a mais bem caracterizada, principalmente por estar
presente em todas as espécies de Salmonella e conter um gene muito utilizado na
identificação do gênero: invA (1). Na SPI-1 estão localizados genes necessários
para invasão, característica importante para virulência (27).
A capacidade de Salmonella em sobreviver no interior de fagócitos e de
replicar dentro de vesículas de células eucarióticas é um processo complexo,
requerendo o envolvimento de muitos genes, incluindo aqueles que auxiliam na
sobrevivência a formas reativas de oxigênio, baixo pH e defensinas. A maioria
destes genes está localizada na SPI-2, essencial para habilidade de proliferar em
tecido extra-intestinal e causar infecções sistêmicas (28).
Na SPI-3 estão presentes os genes mgtB e mgtC, necessários para

12
sobrevivência no interior de macrófagos. O gene mgtB é responsável pelo
transporte de magnésio (Mg2+) quando este se encontra em baixas
concentrações. Este sistema de captação de Mg2+ é importante para adaptação a
limitações nutricionais no interior do fagossomo. A função do mgtC ainda não está
claro. Mutantes para estes genes são deficientes na proliferação intracelular e na
infecção sistêmica (29).
SPI-4 codifica o Sistema de Secreção do Tipo 1, que transloca
proteínas necessárias para colonização intestinal. É especulado que esta ilha está
envolvida na secreção de citotoxinas, responsáveis por induzir a apoptose de
macrófagos infectados. Um lócus dentro da SPI – 4 também tem sido relacionado
com a sobrevivência no interior de macrófagos. No entanto, sua função principal
ainda não foi totalmente determinada (1).
SPI-5 codifica proteínas efetoras que em conjunto a outras proteínas
são necessárias a enteropatogenicidade no hospedeiro suscetível. Estudos
indicam a sua presença em vários sorotipos de S. enterica enterica incluindo os
sorotipos Dublin e Typhimurium (28).
As ilhas de patogenicidade seis a 17 ainda carecem de estudos, sendo
algumas funções muito importantes já identificadas. Em isolados de S. Typhi a
produção de cápsula polissacarídica Vi está relacionada com a ilha de
patogenicidade SPI-7, tal antígeno protege o patógeno dos mecanismos de
imunidade inata e mediada por anticorpos do hospedeiro (30). A SPI-11 e SPI-13
possuem um papel importante que é a permanência no interior de macrófagos
(31).
2.3.2 Plasmídeos
Os plasmídeos são elementos citoplasmáticos que podem se auto-
replicar de forma independente da célula bacteriana. A maioria dos plasmídeos é
circular e composta de DNA de fita dupla, possuindo tamanho variável. A
importância dos plasmídeos incide na sua capacidade de possuir genes de
virulência e codificar fatores de virulência e resistência a antibióticos (2). Os
plasmídeos, como parte de um genoma flutuante de organismos bacterianos,

13
aparentam ser uma miniatura de pan-genoma bacteriano, que é composto por um
genoma intrínseco (essencial) e um genoma dispensável (32).
De maneira evidente, os plasmídeos detém uma grande importância
para a bactéria podendo desempenhar inúmeras funções. Entretanto, sabe-se
que nem todos os sorotipos de Salmonella spp. possuem estas estruturas (33). A
especificidade de hospedeiro de Salmonella indica que os diferentes sorotipos
exploram diferentes ecologias. Portanto, espera-se que os plasmídeos
responsáveis por virulência, lidem com diferentes tipos de seleções naturais de
acordo com os ambientes intra e extracelulares relacionados a cada sorotipo (32).
Para efeito de classificação, os plasmídeos, podem ser agrupados de
acordo com sua funcionalidade, como plasmídeos de virulência sorotipo-
específico, ou plasmídeos de alto peso molecular e os de baixo peso molecular.
Ainda, a troca destas estruturas pode ser observada em diversos sorotipos, sendo
importante essa classificação para também identificar a sua origem (33).
A presença de pequenos e grandes plasmídeos em sorotipos de
Salmonella é foco de um grande número de estudos, determinando também os
diferentes genes codificados e as respectivas regiões responsáveis por
resistência a antimicrobianos, invasão na célula hospedeira e outras funções que
podem ser observadas (34).
No sorotipo Typhimurium existe um pequeno plasmídeo (93kb) de
resistência bem reconhecido denominado pSLT, que desde a década de 70 já é
foco de estudos (35). No sorotipo Montevideo, plasmídeos pequeno e grande
foram identificados carregando genes de resistência (estreptomicina, sulfonamida,
tetraciclina): pS5403-1 (53kb) e pS5403-2 (299kb) (36).
Os plasmídeos podem possuir fatores R (resistência) e os fatores F
(fimbriais). O fator R se relaciona à habilidade de intermediar a transferência de
resistência a antimicrobianos entre bactérias que são resistentes para outras que
não são. Normalmente carreiam um grupo de genes responsáveis pela resistência
cuja função é inativar a ação de determinada droga. Os plasmídeos R, como
exemplo R1 e R100, são responsáveis por controlar o processo de replicação e
conjugação, denominados fatores de transferência de resistência (37) (Figura 6a).

14
FIGURA 6: A: Representação esquemática de célula bacteriana, plasmídeos e seus respectivos métodos de resistência a antimicrobianos. be – Bomba de efluxo; ed- Enzimas degradam o antimicrobiano; el- Enzimas se ligam ao antimicrobiano e inibem sua ação. Fonte: Adaptada de scienceblogs.com.br69. B: Representação esquemática de vários genes e funções em um plasmídeo, cada cor representa um gene diferente. Fonte: SWITT36 et al. (2013).
Genes como dhfrA1, aadA1, qacED1 e sul1, relacionados à resistência
a antimicrobianos, são frequentemente encontrados em plasmídeos como pSLT e
pSD107 (Figura 6b). A identificação e sequenciamento de plasmídeos são
utilizados não apenas para avaliar possíveis mecanismos de virulência, mas
também com intuito de determinar a sua semelhança como uma ferramenta de
epidemiologia, onde se pode determinar a origem e grau de parentesco entre
variantes de Salmonella (34).
A maioria dos plasmídeos apenas consegue adquirir os genes de
resistência por meio de Transposons, seja por um plasmídeo diferente, do
cromossomo ou plasmídeos presentes em outras bactérias num mesmo
hospedeiro (38).
be
el
ed

15
2.3.3 Transposons
Os transposons são chamados de “genes saltadores”, pois podem
mover-se de um lugar para outro no genoma do organismo. Podem também
tornar-se integrados ao DNA plasmidial. Transposons simples chamados de
sequências de inserção têm somente aqueles genes necessários a incorporação
em novos locais (Figura 7). Os transposons complexos têm genes adicionais,
como aqueles que codificam resistência a antibióticos, que podem garantir a
sobrevivência durante a terapia antimicrobiana (6).
FIGURA 7: Representação esquemática de um transposon codificando um gene bacteriano e após a ação de uma enzima específica, o mesmo é realocado em outra posição na mesma célula bacteriana. Fonte: Adaptada de www.chrisdellavedova.com70.
A inserção de um transposon em um gene essencial à sobrevivência
da bactéria resulta na morte da célula. A replicação dos transposons ocorre
somente durante o processo replicativo do cromossomo bacteriano ou do
plasmídeo no qual estão inseridos, portanto não podem se replicar de forma
OUTRO GENE
GENE PARA ENZIMA
TRANSPOSON
FIM DA SEQUÊNCIA
ENZIMA TRANSPOSON
CORTE E INSERÇÃO DE TRANSPOSON EM OUTRO LOCAL
TRANSPOSON
GENE ALTERADO

16
independente. Plasmídeos e transposons codificam funções adicionais que
podem trazer vantagens à sobrevivência de célula. Genes que codificam funções
como produção de toxinas e resistência a antibióticos podem ser transportados
por esses elementos genéticos (2).
Os transposons podem representar um ou mais genes que determinam
um fator de virulência ou resistência como é caso do gene tet(A) localizado no
transposon Tn1721, que determina resistência a tetraciclina. Sua ocorrência foi
relatada em Salmonella Cholerasuis e Typhimurium isolados de animais na
Alemanha. Entretanto um determinante maior de virulência relacionado aos
transposons, é a propriedade de elevar a expressão fenotípica de outros genes
como strA e strB, que definem resistência a estreptomicina às bactérias que os
possuem (38).
Em alguns transposons (Tn21, Tn1403, Tn1404, Tn1696, Tn1412 e
Tn2000) são encontrados integrons que determinam funções de virulência. Esses
transposons podem estar localizados no cromossomo ou em plasmídeos (39).
2.3.4 Integrons
Integrons são definidos como unidades genéticas que possuem um
sistema de recombinação sítio-específica por meio da reorganização dos ORF’s
(Open Reading Frames) presentes em cassetes gênicos, convertendo-os em
genes funcionais. Excisão e inserção de genes cassete são facilitadas por uma
integrase presente nos integrons. Em adição, os integrons fornecem promotores
para a expressão de genes transportados pelo gene cassete (40).
A habilidade para adquirir e expressar novos genes permite aos
integrons contribuir para a variabilidade genética tanto no DNA cromossomal
como no não cromossomal. Além disso, eles parecem desempenhar papel
importante na transmissão de resistência a antibióticos entre bactérias Gram
negativas por meio de conjugação (2).
A grande dispersão dos integrons é provavelmente uma consequência
da sua associação a outros elementos móveis, que também aumenta as

17
possibilidades genéticas. A sua diversidade e abundância está relacionada ao tipo
de pressão seletiva que pode abranger o ambiente e o hospedeiro (41).
Geralmente os integrons possuem o gene intI responsável por codificar
integrase além de um sítio de recombinação denominado attI em que são
inseridos os cassetes gênicos. Vários cassetes podem ser capturados em
conjunto pelo mesmo integron, e sua inserção ou excisão é determinada por meio
da recombinação realizada pela integrase entre attI e/ou qualquer outro sítio-
específico (42) (Figura 8).
FIGURA 8: Representação esquemática da conformação de integron e genes cassete. Fonte: Adaptada de BOUCHER15 et al., 2007.
Os integrons são divididos em classes de acordo com a sequência de
aminoácidos presentes na integrase (IntI). Existem cinco classes de integrons
identificadas, onde cada um é responsável pelo carregamento de cassetes
codificadores de resistência a antibióticos, onde no gênero Salmonella são
associados somente integrons de classe 1 e 2 (39,43).
Apesar de diversos estudos apontarem os integrons de classe 1 como
os principais carreadores de genes de resistência à antimicrobianos (44), é
evidenciado por alguns pesquisadores que na maioria dos isolados de Salmonella
spp. multirresistentes, existe a presença de integrons de classe 1 e 2 (15; 45,46).
GENE CASSETE
CIRCULAR LIVRE
INTEGRAÇÃO/
EXCISÃO

18
Na avaliação de 187 cepas de Salmonella enterica sorotipo
Schwarzengrund coletadas de aves e suínos, foi observada a presença de
integrons de classe 1 na maioria das amostras (84,49%). Foram também
encontrados genes cassetes clássicos como aadA2 (resistência a estreptomicina);
dfrA1 (resistência ao trimetoprim) blaPSE-1 (resistência a ampicilina), além de
seis isolados positivos para SGI-1 (47).
2.3.5 Genes cassete
Os genes cassete são a menor forma de DNA móvel, encontrados
junto a um integron e inseridos normalmente como sequências lineares
constituindo moléculas de DNA maiores como plasmídeos e cromossomos
bacterianos, frequentemente associados ao transporte de genes de resistência a
antibióticos (4) (Figura8).
Diversos sorotipos de Salmonella possuem muitos cassetes gênicos
conhecidos que transportam genes como aadA1, aadA2, aadA4 e aadA5
(conferem resistência a estreptomicina e espectinomicina); blaOXA-30 e bla PSE-
1 (resistência a ampicilina); dfrA1, dfrA7, dfrA12 e dfrA17 (resistência ao
trimetoprim) . Estão frequentemente associados à transposons e plasmídeos
conjugativos, contribuindo para a dispersão dessa característica via transferência
horizontal (48).
Em estudo comparativo entre amostras obtidas de suínos foram
encontrados oito genes cassete diferentes (aadA1, aadA2, AA dA3, aadA4,
aadA5, B, C, D, E1 e E2) associados a integrons, entre os sorotipos Cholerasuis,
Typhimurium, Enteretidis, Heron, Bredeney, Treforest, Albany e Newport. A
resistência à estreptomicina e ampicilina foi a mais observada na maioria dos
sorotipos (49). Uma ocorrência relevante está na grande quantidade de isolados
do sorotipo Cholerasuis com resistência a fluoroquinolona, fato este que enfatiza
a importância dos genes cassete e suas possíveis resistências, visto que
atualmente tal antimicrobiano é o de eleição no tratamento de salmonelose em
humanos (49,50).

19
Os genes cassete possuem uma variabilidade notável, em muitos
estudos onde foi realizada a sua caracterização foi possível a observação da
presença de centenas de sequências diferentes. Até pouco tempo não havia
muito interesse nestes genes, mas várias descobertas têm evidenciado a sua
importância em Salmonella e em outras bactérias. A constituição destas
estruturas singulares intrínsecas a várias bactérias é determinante de diversos
fatores de persistência no ambiente e resistência aos antimicrobianos (41).
2.4 Métodos de detecção de genes de virulência
O estudo de métodos para detecção da presença de fatores de
virulência em bactérias patogênicas é foco de muitas pesquisas e trabalhos no
intuito de facilitar a sua identificação e reduzir os custos finais nesse processo.
Métodos como sequenciamento de DNA, hibridização de DNA e Reação em
cadeia da polimerase são bastante utilizados na detecção de genes de resistência
a antimicrobianos em numerosos gêneros de bactérias. A detecção de genes
relacionados a transposons, integrons e plasmídeos é realizada principalmente
por técnicas de biologia molecular (38). Ainda, a caracterização de ilhas de
patogenicidade é utilizada para descrever sorotipos e linhagens específicos de
Salmonella spp. (51).
2.4.1 Reação em cadeia da polimerase
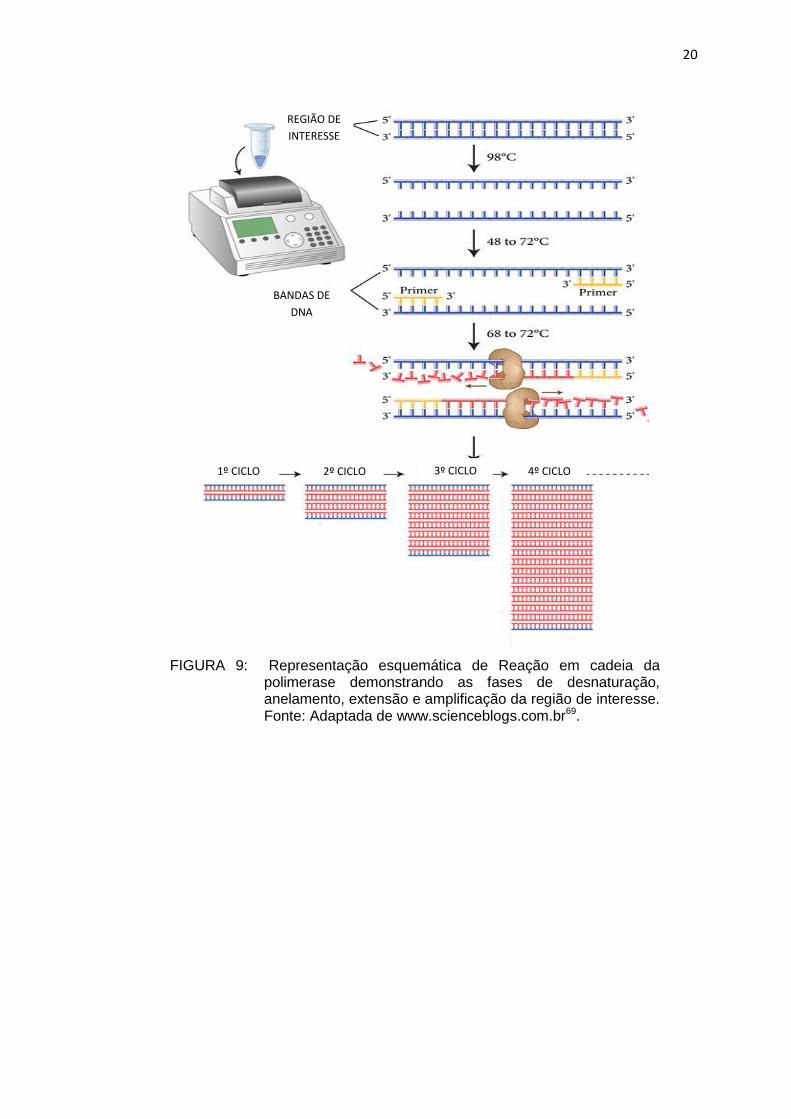
A reação em cadeia da polimerase (PCR) é uma técnica em que
pequenas amostras de DNA podem ser rapidamente amplificadas em
quantidades suficientes para que a análise seja feita. Iniciando com um fragmento
de DNA do tamanho de um gene, a PCR pode ser utilizada para produzir
literalmente bilhões de cópias em poucas horas (6) (Figura 9, 10). Esta técnica,
em associação ou não à técnica de Southern blotting, é a mais utilizada na
identificação de genes de virulência em Salmonella, entre outros gêneros de
bactérias (50, 52).
20
FIGURA 9: Representação esquemática de Reação em cadeia da polimerase demonstrando as fases de desnaturação, anelamento, extensão e amplificação da região de interesse. Fonte: Adaptada de www.scienceblogs.com.br69.
BANDAS DE
DNA
REGIÃO DE
INTERESSE
1º CICLO 2º CICLO 3º CICLO 4º CICLO

21
FIGURA 10: Gel de agarose corado demonstrando
perfil plasmidial de vários sorotipos de
Salmonella enterica. Fonte: del CERRO
53 et al., 2003.
A utilização da PCR para amplificação do gene invA é um método
muito utilizado na detecção de bactérias do gênero Salmonella, isso porque
muitos estudos apontam a sua presença em todos os isolados de Salmonella spp.
testados (53). Esta é uma ferramenta muito utilizada em pesquisas na qual se
pode também detectar genes de resistência em ilhas de patogenicidade (25, 52),
plasmídeos (49), transponsons (38), integrons de classe 1 e classe 2 e
respectivos genes cassete (45).
Outras modalidades de PCR são focos de muitos estudos no intuito de
aumentar a sensibilidade, elaborar métodos quantitativos e reduzir os custos
dessa técnica (54, 55,56, 57, 58, 59, 60, 61).
A PCR multiplex pode combinar numerosos primers para amplificar
diversos genes alvo, para deduzir informação desejável e diminuir o tempo para
obter os resultados (5). A sua utilização pode ser feita para determinar a presença
de fato de Salmonella e outras bactérias como Shigella e Escherichia
simultaneamente com primers específicos para cada gênero e gene alvo (54).
Sua aplicação também pode ser diretamente voltada à determinação de genes de
virulência como fliC, purE e sucA nas amostras identificadas de Salmonella (5) ou

22
ainda, a diferenciação de alguns sorotipos (Typhimurium, Choleraesuis, Infantis,
Hadar, Enteritidis, Dublin e Gallinarum) sabidamente de importância em saúde
pública e animal (62).
2.4.2 Hibridização de ácidos nucleicos
A hibridização de ácidos nucleicos é um procedimento que considera
que se duas espécies são similares ou relacionadas, uma porção maior das
sequências dos seus ácidos nucleicos também será similar. O método mede a
habilidade das fitas de DNA de um organismo de ligar-se por pareamento de
bases complementares com fitas de DNA de outro organismo. Experimentalmente
utiliza-se uma sonda marcada, para que seja visualizado o alvo de pesquisa (6).
Algumas técnicas bastante eficiente utilizadas na detecção de genes
de virulência, entre outras aplicações, são baseadas na hibridização de ácidos
nucleicos.
I. Southern blotting
A utilização de Southern blotting irá identificar se determinadas
sequências de DNA estão presentes ou não em uma amostra de DNA analisada.
O método consiste em seis etapas inicialmente: extração de DNA; digestão de
DNA; preparo da sonda; transferência; hibridização e detecção do DNA. Para
separar os fragmentos de acordo com seu tamanho, utiliza-se a eletroforese em
gel. A detecção do DNA será com base num filtro de nitrocelulose com DNA e sua
posterior exposição a uma sonda marcada radioativamente específica para um
gene (6). Ainda, pode-se amplificar o DNA com a PCR para que na amostra se
realize o Southern blotting (49) (Figura 11).

23
FIGURA 11: Representação esquemática da técnica de Southern blotting demonstrando as etapas necessárias para a sua realização. Extração de DNA; digestão de DNA; preparo da sonda; transferência; hibridização e detecção do DNA Fonte: www.icb.ufmg.br/big/genmed/southblot.html72.
Em estudo realizado na última década, foi observada uma alteração no
padrão de plasmídeos de isolados de Salmonella Cholerasuis de humanos em
relação aos isolados de suínos com a utilização da técnica de Southern blotting.
Quando foram comparados entre si, as marcações observadas nos sorotipos
isolados de humanos apresentavam plasmídeos sem qualquer ligação com os
observados nos isolados de suínos (Figura 12). Os resultados sugerem que cada
plasmídeo evolui de forma independente, ainda que pertençam ao mesmo
sorotipo (50).
FIGURA 12: Resultado de Southern blotting de plasmídeos de
virulência de Salmonella Cholerasuis isoladas de humanos e suínos. Fonte: TZENG50 et al. (2012).
DNA SEPARADO EM GEL DE AGAROSE TRANSFERÊNCIA DE FRAGMENTOS
DO GEL PARA MEMBRANA
MEMBRANA COM BANDAS DE DNA TRANSFERIDAS
SONDA INCUBADA COM MEBRANA
BANDAS COM DNA LIGADO SÃO EXPOSTAS NO FILME
SONDA RADIOMARCADA

24
II. Microarray ou Chip de DNA
Um chip de DNA pode ser fabricado para conter centenas de milhares
de sequências de DNA de fitas simples sintéticas. Cada sequência de DNA é
exclusiva para um gene diferente. Um DNA desconhecido é separado em fitas
simples, cortado enzimaticamente e marcado com um corante fluorescente, o
DNA desconhecido é inserido no chip e hibridiza com o DNA no chip. O DNA
marcado vai se ligar somente com o DNA complementar no CHIP. O DNA ligado
vai ser detectado pela fluorescência e analisada por um computador (6).
A utilização da técnica de Microarray primeiramente foi testada para
diferenciar apenas a espécie e em seguida os sorotipos de Salmonella.
Inicialmente essa técnica era muito dispendiosa tanto em tempo quanto em
custos para os laboratórios. No entanto, constantes esforços determinaram uma
viabilidade da técnica e conseguiram introduzir vários genes de importância nos
chips, como os genes de virulência de Salmonella stm, sty, e stv. (62) (Figura 13).
A Microarray oferece uma alternativa viável à PCR para caracterização
genética de bactérias. Vem sendo utilizada com relativo sucesso para detectar
genes de resistência, caracterizar a virulência de isolados, Fingerprinting
genômico e detecção de polimorfismos genéticos (49).
FIGURA 14: Representação esquemática de Microarray de genes de virulência e resistência de Salmonella spp. e E. coli. Cada marcação determina um gene padrão. Fonte: CHEN51 et al. (2005)

25
2.4.3 Outros métodos de detecção de genes de virulência em Salmonella enterica
I. Fingerprinting
Nos últimos anos a identificação e sequenciamento completo de bases
do DNA se tornou possível com métodos utilizando dezenas de testes
bioquímicos modernos, mas isso é impraticável para a identificação laboratorial
porque demanda uma grande quantidade de tempo (6).
A técnica de Fingerprinting é capaz de determinar uma ampla
sequência de genes, podendo ser utilizada tanto na detecção de genes de
virulência em diversas regiões do cromossomo, como também na caracterização
de plasmídeos presentes nas bactérias e diferenciação de sorotipos, incluindo
Salmonella spp. (62).
A utilização de Fingerprinting é uma ferramenta valiosa para determinar
os genes presentes e sua relação genética com diversas bactérias isoladas. É um
dos principais métodos moleculares utilizados em agências regulatórias e
laboratórios federais de saúde pública na vigilância sanitária de rotina e para
traçar as fontes de surtos de doenças (63).
II. Tipificação de sequência de multilocus
Um método usado para realizar estudos genéticos em populações
bacterianas é a Tipificação de sequência de multilocus (Multilocus sequence
typing – MLST). Essa técnica é baseada na determinação de sequências de DNA,
e, tem sido aplicada para muitos patógenos importantes fornecendo informações
sobre a evolução e a diversificação de espécies, sendo útil para entender a
evolução de clones e a patogenicidade da Salmonella. No entanto, possui
desvantagens como a acessibilidade limitada e o custo elevado (64).
A MLST pode ser utilizada como uma ferramenta de epidemiologia
molecular de alta relevância podendo detectar muitos genes de resistência (fimA,
tgm, hisC, icdA, atpB) de maneira simultânea (Figura 14). Todos os dados obtidos

26
são determinados de maneira digital e diminuem a taxa de erro, inclusive entre
laboratórios diferentes (5).
FIGURA 14: Tipificação de sequência multilocus (MLST), cada quadrado branco condiz com a presença de um gene alvo determinado na abscissa.
Fonte: SINGH5 et al. 2012.
III. Dendograma
A elaboração de dendogramas é um método realizado a partir de
software específico, onde um conjunto de dados obtidos em testes como
eletroforese, PCR, identificação de plasmídeos e suscetibilidade a
antimicrobianos, é analisado em conjunto como ferramenta para determinar a
similaridade genética bem como outras semelhanças (Figura16). Essa análise
ajuda no entendimento da presença ou não de estruturas como plasmídeos e
genes de virulência, correlacionando à localização geográfica e fontes de
contaminação (64).

27
FIGURA 15: Dendograma de Salmonella Javiana. Presença de genes semelhantes (linhas vermelhas) ou distintos a partir do peso molecular em plasmídeos. Presença de resistência a antimicrobianos testados (quadrado vermelho), baixa suscetibilidade a antimicrobianos testados (quadrado verde) e suscetibilidade plena a antimicrobianos testados (azul claro). Presença de genes de virulência, amarelo presente, azul ausente. Fonte: Adaptada de MEZAL64 et al. 2013.
IV. Sequenciamento genômico
O sequenciamento de todo o genoma bacteriano é possível de ser
realizado, apesar de representar uma técnica laboriosa e cara. A respeito das
bactérias do gênero Salmonella, existem dados publicados de sequenciamento
dos sorotipos Typhimurium, Cholerasuis, Enteritidis, Dublin, Infantis, Hadar e
Gallinarum e podem ser obtidos no chamado GenBank que é administrado pelo
Centro Nacional para Informação de Biotecnologia (NCBI). A partir dos dados
presentes no GenBank pode-se comparar um ou mais fragmentos de DNA
obtidos de Salmonella sem haver necessidade de sequenciar todo o seu genoma
(60).

28
3. CONSIDERAÇÕES FINAIS
Para entender os fatores que envolvidos na virulência de Salmonella, é
importante admitir a presença dos genes de virulência que auxiliam na
proliferação, disseminação e a patogenicidade nos hospedeiros suscetíveis ao
microrganismo patogênico. Na verdade, esses genes são compostos por várias
sequências que determinam capacidades para a sobrevivência das bactérias em
meio a tantos desafios evolutivos e ambientais.
A presença de dezenas de linhagens de Salmonella multirresistentes a
antimicrobianos comuns é uma realidade preocupante para a saúde pública e
animal. Sabe-se que o uso intensivo de antimicrobianos de maneira
indiscriminada e profilática é um fator de grande relevância na seleção natural de
bactérias mais resistentes. Conhecer os mecanismos, propriedades e estruturas
bacterianas que têm importância na virulência de Salmonella, compõem pontos
necessários no entendimento do comportamento da bactéria em seus
hospedeiros.
A alta demanda por estudos e métodos de detecção de genes de
virulência abrange a pesquisa não só em microbiologia, mas especialmente nas
áreas de bioquímica e biologia celular. Ainda, esses estudos representam passos
largos essenciais para os avanços no entendimento de patógenos e doenças que
acometem animais e seres humanos.

29
REFERÊNCIAS BIBLIOGRÁFICAS
1. Marcus SL, Brumell JH, Pfeifer CG, Finlay BB. Salmonella Pathogenicity
islands: big virulence in small packages. MicrobesInfect. 2000; 2:145−156.
2. Quinn PJ, Markey BK, Carter, ME, Donnely WJ, Leonard FC. Microbiologia
Veterinária e Doenças Infecciosas. 1ª ed. Porto Alegre: Artmed; 2005, 512p.
3. Lewin B. Genes VIII. Pearson Education Inc: Philadelphia; 2004, 1056p.
4. Michael CA, Gilling MR, Holmes AJ, Hughes l, Andrews NR, Holley MP;
Strokes HW. Mobile Gene Cassettes: A Fundamental Resource for Bacterial
Evolution, The Amer nat. 2004; 164: 1-12.
5. Singh P, Foley S, Nayak R, Kwon YM. Multilocus sequence typing of
Salmonella strains by high-throughput sequencing of selectively amplified
target gene, J Microbiol Methods. 2012; 88: 127–133
6. Tortora GJ, Funcke, BR, Case C.L. Microbiologia. 10.ed. Porto
Alegre:Artmed, 2012, 894p.
7. BAUMAN, R. W. Microbial Genetics. In: Microbiology. 2.ed. San Francisco:
Pearson, 2009. cap. 7, p. 197-328.
8. Ochman H, Lawrence JG., Groisman EA. Lateral gene transfer and the
nature of bacterial innovation, Nat. 2000; 405: 299–304.
9. Thomas CM, Nielsen KM. Mechanisms of, and barriers to, horizontal gene
transfer between bacteria. Nat. Rev. Microbiol, 2005; 3: 711–721.
10. Van Elsas J D; Bailey MJ. The ecology of transfer of mobile genetic
elements, FEMS Microb Eco. 2002; 42: 187–197.
11. Yang B, Zheng J, Brown EW, Zhao S, Meng J. Characterization of
antimicrobial resistance-associated integrons and mismatch repair gene
mutations in Salmonella serotypes, International J. of Antimic Ag. 2009; 33:
120-124.

30
12. Hirsh DD. Samonella.In: Microbiologia Veterinária. 2 ed. Rio de Janeiro:
Granabara Koogan SA, 2003. cap. 9, p. 56-63.
13. Fierer J, Guiney DG. Diverse virulence traits underlying different clinical
outcomes of Salmonella infection. The Jour of Clin Investig. 2001; 107: 21-
32.
14. Hsua M, Tang CY, Lina H, Chena YH, Chena YL, Sua YH, Chenb DS, Linc
JH, Chang CC. Comparative study of class 1 integron, ampicillin,
chloramphenicol, streptomycin, sulfamethoxazole, tetracycline (ACSSuT) and
fluoroquinolone resistance in various Salmonella serovars from humans and
animals, Comp. Immunol. Microbiol. Infect. Dis. 2013; 36: 9– 16.
15. Boucher Y, Labbate M, Koenig JE, Stokes HW. Integrons: mobilizable
platforms that promote genetic diversity in bacteria. Tren in Microb. 2007;
v.15 (7): 301-309.
16. Miriagou V, Caratolli A, Fanning S. Antimicrobial resistance islands:
resistance gene clusters in Salmonella chromosome and plasmids, Microb
and Infect. 2006; 8: 923-1930.
17. Kich JD, Coldebella A, Morés N, Nogueira MG, Cardoso M, Fratamico PM,
Call JE, Fedorka-cray P, Luchansky JB. Prevalence, distribution, and
molecular characterization of Salmonella recovered from swine finishing
herds and a slaughter facility in Santa Catarina, Brazil, Int J Food Microbiol.
2011; 151:307-313.
18. Pérez-Moreno MA, Picó-Plana E, Toro M, Grande-Armas J, Quiles-Fortuny
V, Ponse MJ, Gomes C, Sáenz Y, Torres C, Ruiz J. BLactamases,
transferable quinolone resistance determinants, and class 1 integron-
mediated antimicrobial resistance in human clinical Salmonella enterica
isolates of non-Typhimurium serotypes. Int Jour Med Microb. 2013; 303: 25–
31.
19. Poole,K. Mechanisms of bacterial biocide and antibiotic resistance, J Appl
Microbio 2002; 92: 55–64.

31
20. Maillard JY. Bacterial target sites for biocide action. J Appl Microbio. 2002;
92: 16–27.
21. Morente EO, Fernández-Fuentes MA, Burgos MJG, Abriouel H, Pulido RP,
Gálvez A. Biocide tolerance in bactéria, Int J Food Microbiol. 2012; 162 :13–
25
22. Borowsky LM, Bessa MC, Cardoso MI, Avancini CAM. Sensibilidade e
resistência de amostras de Salmonella Typhimurium isoladas de suínos
abatidos no Rio Grande do Sul/Brasil frente aos desinfetantes químicos
quaternário de amônio e iodofor. Ciência Rural. 2006;36: 1474-1479.
23. MARQUES NDB, 2011. Caracterização molecular e fenotípica de
Salmonella Typhi ISOLADA DE CASOS DE FEBRE TIFÓIDE NO ESTADO
DO PARÁ, NO PERÍODO DE 1970 A 2009. Universidade Federal do Pará
Dissertação apresentada ao Programa de Pós-Graduação em Biologia de
Agentes Infecciosos e Parasitários 89 p
24. Langridge GC, Phan MD, Turner DJ, Perkins TT, Parts L, Haase J, Charles I,
Maskell DJ, Peters SE, Dougan G, Wain J, Parkhill J, Turner
AK.Simultaneous assay of every Salmonella Typhi gene using one million
transposon mutants, Genome Research. 2009;19:2308–2316.
25. Oliveira AP, Sola MC, Feistel JC, Moreira NM, Oliveira JJ. Salmonella
enterica: genes de virulência e ilhas de patogenicidade. Enciclopédia
Biosfera, Centro Científico Conhecer. 2013; 9 (16):1947-1972.
26. Vieira MAM. Ilhas de patogenicidade. O mundo da saúde, 2009; 33: 406-
414.
27. Hensel, M. Evolution of pathogenicity islands of Salmonella enterica. I. J Med
Microbiol. 2004; 294:95-102.
28. Schmidt H.; Hensel M. Pathogenicity islands in bacterial pathogenesis. Clinl
Microb Rev. 2004; 19(1): 14-56.

32
29. Groisman EA, Ochman H. The path to Salmonella, Features. 2000; 66: 21-
27.
30. Ferreira E O, Campos LC. Salmonella. In: Trabulsi LR, Althernum F.
Microbiologia. 5.ed. São Paulo:Ed.Atheneu, 2008. cap. 43, p. 329-338.
31. Shah DH, Lee, M, Park J, Lee J, Kwon J, Chae J. Identification of Salmonella
gallinarum virulence genes in a chicken infection model using PCR based
signature-tagged mutagenesis, Microbiology. 2005; 151: 3957-3968.
32. Chu C, Feng Y, Chien AC, Hu S, Chu CH, Chiu CH. Evolution of genes on
the Salmonella Virulence plasmid phylogeny revealed from sequencing of the
virulence plasmids of S. enterica serotype Dublin and comparative analysis,
Genomics. 2008; 92: 339–343.
33. Rychlik I, Gregorova D, Hradecka H. Distribution and function of plasmids in
Salmonella enterica, J. Vet. Microbiol. 2006; 112: 1-10.
34. Bleicher A, Schofl G, Rodicio MR, SALUZ, H.P. The plasmidome of
Salmonella enterica serovar Derby isolated from pork meat, Plasmid. 2013;
69: 2012-210.
35. García-Quintanilla M, Casadesús J. Virulence plasmid interchange between
strains ATCC 14028, LT2, and SL1344 of Salmonella enterica serovar
Typhimurium, Plasmid. 2011; 65: 169–175.
36. Switt AM, Ranieri M, Bakker HC, Peters J, Degoricija L, Cummings CA,
Govoni G, Bolchacova E, Furtado MR, Wiedmann M. Cornell University,
2013. Disponível em: www3.appliedbiosystems.com. Acesso em: 19 set.
2013.
37. Pezzella C, Ricci A, Digiannatale E, Luzzi, I, Carattoli A. Tetracycline and
streptomycin resistance genes, transposons, and plasmids in Salmonella
enterica isolates from animals in Italy, Antimicrob Agents Chemother
(Bethesda). 2004; 48: 903-908.

33
38. Lázaro NS, Tibana A, Rodrigues DP, Reis, EMF, Quintaes BR, Hofer.
Antimicrobial resistance and R-plasmid inSalmonella spp. from swine and
abattoir, Pesq. Vet. Bras. 2004; 24(2): 57-60
39. Fluit AC, Schmitz FJ. Resistance integrons and super-integrons.Review. Clin
Microbiol Infect. 2004;10: 272–288.
40. Cambray G, Guerout AM, Mazel D. Integrons. Annual Review of Genetics,
Palo Alto. 2010; 44: 141–166.
41. Moura A, Pereira C, Henriques I, Correia A.Novel gene cassettes and
integrons in antibiotic-resistant bacteria isolated from urban wastewaters,
Research in Microbiology. 2012;163: 92-100.
42. Partridge SR,Tsafnat, G, Coiera E; Iredell J. Gene cassettes and cassette
arrays in mobile resistance integrons. FEMS Microbiology Review, 2009; 33:
757–784.
43. Mazel, D. Integrons: agents of bacterial evolution. Nat. Rev. Microbiol. 2006;
4: 608–620.
44. Huang SC, Chiu CH, Chiou CS, Yang YJ. Multidrug-resistant Salmonella
enterica serovar Panama carrying class 1 integrons is invasive in Taiwanese
children, J Formos Med Assoc. 2013; 112: 269-275
45. Macedo-Vin˜as M, Cordeiro NF, Bado I, Herrera-Leon S, Vola M, Robino L,
Gonzalez-Sanz R, Mateos S, Schelotto F, Algorta G, Ayala JA, Echeita A,
Vignoli R. Surveillance of antibiotic resistance evolution and detection of
class 1 and 2 integrons in human isolates of multi-resistant Salmonella
Typhimurium obtained in Uruguay between 1976 and 2000 Int Jour Inf Dis.
2009; 13: 342—348
46. Mokracka J, Koczura R, Kaznowski A. Multiresistant Enterobacteriaceae
with class 1 and class 2 integrons in a municipal wastewater treatment plant
Water Research. 2012; 46:3353-3363.

34
47. Wang Y, Chang YC, Chuang HL, Chiu CC, Yeh KS, Chang C C, Hsuan SL,
Chen TH. Antibiotic resistance, integrons and Salmonella genomic island 1
among Salmonella Schwarzengrund in broiler chicken and pig, Afr J
Microbiol Res. 2010; 4: 677-681.
48. Van TTH, Nguyen HNK, Smooker PM, Coloe PJ. The antibiotic resistance
characteristics of non-typhoidal Salmonella enterica isolated from food-
producing animals, retail meat and humans in South East Asia. International
Int J Food Microbiol. 2012; 154: 98-106.
49. Hsu YM, Tanga CY, Lin H, Chena YH, Chen YL, Su YH,. Chen DS, Linc JH,
Chang CC. Comparative study of class 1 integron, ampicillin,
chloramphenicol, streptomycin, sulfamethoxazole, tetracycline (ACSSuT) and
fluoroquinolone resistance in various Salmonella serovars from humans and
animals, Comp. Immunol.Microbiol. Infec Dis. 2013; 36:9– 16.
50. Tzeng JI, Chu CH, Chen SW, Yeh CM, Chiu CH, Chiou CS, Lin JH, Chu C.
Reduction of Salmonella enterica serovar Choleraesuis carrying large
virulence plasmids after the foot and mouth disease outbreak in swine in
southern Taiwan, and their independent evolution in human and pig, J
Microbiol Immunol Infect. 2012; 45: 418-425
51. Chen S , Zhaob S, McDermottb PF, SchroederaCM, Whiteb DG, Menga J. A
DNA microarray for identification of virulence and antimicrobial resistance
genes in Salmonella serovars and Escherichia coli, Mol. Cell.Probes. 2005;
19: 195–201
52. Mezal EH, Sabol A, Khan MA, Ali N, Stefanova R, Khan AA. Isolation and
molecular characterization of Salmonella enterica serovar Enteritidis from
poultry house and clinical samples during 2010. Food Microbiol. 2014; 38:
67-74
53. del Cerro A, Soto SM, Mendoza MC. Virulence and antimicrobial-resistance
gene profiles determined by PCR-based procedures for Salmonella isolated
from samples of animal origin, Food Microbiol. 20 (2003) 431–438

35
54. Amini K, Salehi TZ, Nikbakht G, Ranjbar R, Amini J, Ashrafganjooei SB.
Molecular detection of invA and spv virulence genes in Salmonella enteritidis
isolated from human and animals in Iran, Afr J Microbiol Res. 2010; 4(21):
2202-2210.
55. Jamshidi A, Bassami MR, Afshari-Nic S. Identification of Salmonella serovars
Typhimurium by a multiplex PCR-Based assay from Poultry carcasses in
Mashhad-Iran. Int. J. Vet. Res. 2008; 3(1): 43-49.
56. Shao Y, Zhu S, Jin C, Chen F. Development of multiplex loop-mediated
isothermal amplification-RFLP (mLAMP-RFLP) to detect Salmonella spp..
and Shigella spp.. in milk, Int J Food Microbiol. 2011;148: 75–79
57. Cheung PY, Kam KM. Salmonella in food surveillance: PCR, immunoassays,
and other rapid detection and quantification methods. Food Res Intern. 2012;
45: 802–808
58. Zahraei T, Mahzoonae MR, Ashrafi A. Ampllification of invA gene of
Salmonella by polymerase chain reaction (PCR) as a specific method for
detection of Salmonella. J. Fac. Vet. Med. Univ. Tehran. 2006; 61(2): 195-
199.
59. Zhang Y.; LeJeune JT. Transduction of bla(CMY-2), tet(A), and tet(B) from
Salmonella enterica subspecies enterica serovar Heidelberg to S.
Typhimurium. Vet Microb. 2008; 129: 418-425..
60. Akiba M, MKusumoto, Iwata T. Rapid identification of Salmonella enterica
serovars, Typhimurium, Choleraesuis, Infantis, Hadar, Enteritidis, Dublin and
Gallinarum, by multiplex PCR, J Microbiol Methods. 2011; 85: 9–15.
61. Peterson G, Gerdes B, Berges J, Nagaraja TG, Frye JG, Boyle DS,
Narayanan S. Development of microarray and multiplex polymerase chain
reaction assays for identification of serovars and virulence genes in
Salmonella enterica of human or animal origin, Vet Diagn Invest. 2010;
22:559–569.

36
62. Sajid SU, Schwarz S. Plasmid fingerprinting and virulence gene detection
among indigenous strains of Salmonella enterica serovar enteritidis. Ayub
Med Coll Abbottabad. 2009;21(2): 83-86.
63. Akiyama T, Khan AA, Cheng CM, Stefanova R. Molecular characterization of
Salmonella enterica serovar Saintpaul isolated from imported seafood,
pepper, environmental and clinical samples, Food Microbiol . 2011; 28:1124-
1128.
64. Shahada F, Amamoto A, Chuma T, Shirai A, Okamoto K. Antimicrobial
susceptibility phenotypes, resistance eterminants and DNA fingerprints of
Salmonella enterica serotype Typhimurium isolated from bovine in Southern
Japan, Int J Antimicrob Agents. 2007; 30: 150–156
65. Lan R, Reeves PR, Octavia S. Population structure, origins and evolution of
major Salmonella enterica clones. Infect. Genet. Evol. 2009; 9(5): 996-1005.
66. Ezat H. Mezal, Rossina Stefanova d, Ashraf A. Khan . Isolation and
molecular characterization of Salmonella enterica serovar Javiana from food,
environmental and clinical samples, Int J Food Microbiol. 2013; 164:113–118.
67. Student Consult: Bacteria, 2011. Disponível em:www.studentconsult.com.
Acesso em: 20 set. 2013.
68. Revista Escola: Humano x bactérias: como ambos se defendem? Disponível
em: http://revistaescola.abril.com.br. Acesso em: 20 set. 2013.
69. Salmonella Organization: List of Salmonella genes. Disponível em
http://www.salmonella.org. Acesso em: 20 set. 2013.
70. Science blog: Blog de Ciência. Disponível em: www.scienceblogs.com.br.
Acesso em: 20 set. 2013.
71. Lavedova CD, página pessoal, 2008. Disponível em
www.chrisdellavedova.com. Acesso em: 20 set. 2013.

37
72. Instituto de Ciências Biológicas, UFMG: Hibridização de ácidos nucleicos.
Disponível em www.icb.ufmg.br/big/genmed/southblot.html. Acesso em: 20
set. 2013.


















