
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA ...§ão versão... · RESUMO Os ciclídeos...
82
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA, CONSERVAÇÃO E BIOLOGIA EVOLUTIVA CÓDIGO DE BARRAS DE DNA COMO FERRAMENTA PARA O ESTUDO DA BIODIVERSIDADE DE PEIXES DA FAMÍLIA CICHLIDAE NA BACIA AMAZÔNICA ANA PAULA COSTA DE CARVALHO Manaus, Amazonas Junho 2014
Transcript of INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA ...§ão versão... · RESUMO Os ciclídeos...

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA,
CONSERVAÇÃO E BIOLOGIA EVOLUTIVA
CÓDIGO DE BARRAS DE DNA COMO FERRAMENTA PARA O
ESTUDO DA BIODIVERSIDADE DE PEIXES DA FAMÍLIA
CICHLIDAE NA BACIA AMAZÔNICA
ANA PAULA COSTA DE CARVALHO
Manaus, Amazonas
Junho 2014

i
Ana Paula Costa de Carvalho
CÓDIGO DE BARRAS DE DNA COMO FERRAMENTA PARA O
ESTUDO DA BIODIVERSIDADE DE PEIXES DA FAMÍLIA
CICHLIDAE NA BACIA AMAZÔNICA
Orientador: Ph.D. Tomas Hrbek
Dissertação apresentada ao Programa
de Pós-Graduação em Genética,
Conservação e Biologia Evolutiva do
Instituto Nacional de Pesquisas da
Amazônia como parte dos requisitos
para obtenção de título de Mestre em
Genética, Conservação e Biologia
Evolutiva.
Manaus, Amazonas
Junho 2014

ii
C331c Carvalho, Ana Paula Costa de
Código de barras de DNA como ferramenta para o estudo da
biodiversidade de peixes da família Cichlidae na bacia Amazônica /
Ana Paula Costa de Carvalho. --- Manaus: [s.n.], 2014.
xii, 86 f. : il. color.
Dissertação (Mestrado) --- INPA, Manaus, 2014.
Orientador: Tomas Hrbek.
Área de concentração: Genética, Conservação e Biologia
Evolutiva.
1. DNA Barcoding. 2. Conservação. 3. Ictiofauna. 4.
Neotropical. I.Título
CDD 597.58
Sinopse:
Estudou-se a metodologia do código de barras de DNA (gene
COI) para identificar e delimitar a ictiofauna diversificada de
ciclídeos em água doce da região Neotropical. O código de barras
de DNA conseguiu discriminar a maioria das espécies analisadas e
sinalizar espécies crípticas e complexos de espécies.
Palavras-chave: Citocromo oxidase I, espécies crípticas,
identificação.

iii
AGRADECIMENTOS
Primeiramente a Deus pela minha existência, pela dádiva de viver e por abençoar sempre o
meu caminho.
Aos meus Familiares, principalmente aos meus Avós (Raimundo Ferreira da Costa e
Maria Mendes da Costa) e aos meus Pais (Domingos Marques de Carvalho e Euzemar
Costa de Carvalho) que moram no meu coração, pelo afeto e carinho, pelos conselhos que
me ajudaram a vencer na estrada da vida e pelo incentivo na minha determinação, devo tudo
isso à vocês.
Ao Francisco Martins de Castro, meu esposo, pelo companheirismo, amor, carinho,
dedicação, apoio e compreensão nas fases difíceis da minha vida e pela vitória conquistada.
Ao Professor Dr. Tomas Hrbek pela orientação, confiança e ajuda durante a execução deste
trabalho.
A Profª. Dra. Izeni Pires Farias pelo apoio e ensinamentos durante o desenvolvimento da
minha dissertação.
Agradeço o Programa de Pós-graduação em Genética, Conservação e Biologia
Evolutiva/INPA pela oportunidade de mestrado. Os Professores do programa que se
dedicaram em repassar conhecimentos essenciais ao meu aprendizado e para a minha
formação profissional durante as disciplinas. E também a secretária Alessandra, sempre
disposta a atender nossas solicitações.
A Universidade de Federal do Amazonas (UFAM) pelo espaço utilizado do Laboratório de
Genética Animal (LEGAL) para execução das análises do meu mestrado.
A CAPES, pela concessão da bolsa de estudos e pelo financiamento deste projeto de pesquisa
juntamente com a CNPq e FAPEAM.
Aos colegas (José, Valéria, Deyla, Natasha, Sarah, Gabi, Mário, Adriano, Júlia, Olavo,
Concy, Elciomar, Fabrício, Fabinho e Emanuel) do Laboratório de Evolução e Genética

iv
Animal (LEGAL)/UFAM que voluntariamente me ajudaram a realizar este trabalho. Obrigada
pela colaboração, incentivo e atenção!
Aos pesquisadores e amigos do INPA e do LEGAL pela receptividade e colaboração em
realizar as coletas e identificar as amostras.
Enfim, agradeço a todos, que de alguma maneira, contribuíram para a realização deste
trabalho.

v
"Há homens que lutam um dia e são bons.
Há outros que lutam um ano e são melhores.
Há os que lutam muitos anos e são muito
bons. Porém, há os que lutam toda a vida.
Esses são os imprescindíveis."
Bertolt Brecht.

vi
RESUMO
Os ciclídeos neotropicais constituem um grupo variado, no qual compreendem cerca de 60
gêneros e pelo menos 1300 espécies são reconhecidas. São distribuídas na América Central e
do Sul, Texas, Índias Ocidentais, África, Madagascar, Síria, Israel, Irã, Sri Lanka, litoral do
sul da Índia. A diversidade encontrada na família Cichlidae abrange os mais diferentes
aspectos morfológicos e ecológicos, que facilitam na adaptação dos peixes desta família.
Além disso, evidências com base nos dados moleculares mostraram que a maior parte da
diversificação da ictiofauna Neotropical ocorreu recentemente. Estas características tornam
difícil a compreensão da taxonomia e identificação dessa fauna, tornando um grande desafio
para as ferramentas moleculares. Neste contexto, o presente estudo teve como objetivo testar a
eficácia da metodologia do código de barras de DNA (gene COI) para identificar e delimitar a
ictiofauna diversificada de ciclídeos em água doce da região Neotropical. Para esta finalidade,
foram analisadas 59 espécies de peixes na bacia Amazônica, sendo amostrado por até cinco
espécimes, identificadas a priori morfologicamente, e obtidas 147 sequências de código de
barras de DNA. Foram combinados os dados morfológicos com os moleculares, onde apenas
nove espécies apresentaram concordância em ambos os métodos de identificação. Das 45
espécies analisadas, 40 (88,8%) foram corretamente identificadas pelas sequências de códigos
de barras. Os principais valores de divergência genética intraespecífica e interespecífica pelo
K2P nos agrupamentos moleculares foram 0% e 55%. E verificou-se sete espécies crípticas
(15,5%), sete complexos de espécies (15,5%) e aproximadamente cinco espécies (11,2%)
mostraram ausência de monofilia recíproca. Este estudo é o primeiro analisar um grande
número de espécies de peixes de água doce da região Neotropical, incluindo um grande
número de espécies estreitamente relacionadas. Os resultados confirmaram a eficácia do
código de barras para sinalizar espécies crípticas e complexos de espécies, que passaram
recentemente por um processo de irradiação, além de discriminar a maioria das espécies
analisadas. Os resultados também revelaram divergências genéticas sugestivas de isolamento
reprodutivo e especiação críptica em sete espécies. Enfim, os resultados obtidos contribuirão
significamente para o projeto Internacional do Código de Barras da Vida, e para a campanha
FISH-BOL fornecendo as sequências de código de barras para uso na identificação dessas
espécies por especialistas e não-especialistas, e permitindo-lhes estar disponível para uso em
outras aplicações.

vii
ABSTRACT
The Neotropical cichlids are a diverse group, which comprises about 60 genera and at least
1300 species are recognized. They are distributed in Central and South America, Texas, West
Indies, Africa, Madagascar, Syria, Israel, Iran, Sri Lanka, southern coast of India. The
diversity found in the Cichlid family covers the most different morphological and ecological
aspects that facilitate the adaptation of this fish family. In addition, evidence based on
molecular data showed that most of the diversification of the Neotropical fish fauna occurred
recently. These features make it difficult to understand the taxonomy and identification of this
fauna, becoming a major challenge for molecular tools. In this context, this study aimed to
test the effectiveness of DNA barcode methodology (COI gene) to identify and delineate the
diverse ictiofauna of freshwater cichlids in the Neotropics. For this purpose, 59 species of
Amazon basin fish were analyzed, being sampled by up to five specimens, morphologically
identified beforehand, and 147 DNA barcode sequences obtained. Morphological data were
combined with the molecular where only nine species showed agreement in both methods of
identification. Of the 45 species analyzed, 40 (88.8%) were correctly identified by the barcode
sequencing. The main values of intraspecific and interspecific genetic divergence by K2P in
molecular clusters were 0% and 55%. Seven cryptic species (15.5%) as well as seven
complexes of species (15.5%) were verified and approximately five species (11.2%) showed
absence of mutual monophyly. This study is the first to analyze a large number of freshwater
fish species in South America, including a large number of closely related species. The results
confirmed the effectiveness of barcode to signal cryptic species and species complexes, which
have recently undergone a process of irradiation, in addition to discriminate most species
analyzed. The results also revealed genetic differences suggesting reproductive isolation and
cryptic speciation in seven species. Finally, the results will contribute significantly for the
Life Barcode International Project, and for the FISH-BOL campaign providing sequences of
barcodes on the identification of these species by specialists and non-specialists, as well as
allowing them to be available for use in other applications.

viii
SUMÁRIO
Pág.
RESUMO……………………………………………………………………….. vi
ABSTRACT..…………………………………………………………………… vii
1. INTRODUÇÃO GERAL……………………………………………………. 13
1.1 Biologia e Ecologia dos Ciclídeos................................................................... 13
1.2 História Biogeográfica dos Ciclídeos.............................................................. 18
1.3 Código de Barras de DNA............................................................................... 19
CAPÍTULO I........................................................................................................ 23
Artigo I................................................................................................................... 24
2. CONSIDERAÇÕES GERAIS........................................................................ 47
3. REFERÊNCIAS BIBLIOGRÁFICAS........................................................... 47

ix
LISTA DE TABELAS
Pág.
Tabela 1. Nomes das espécies, baseadas nas identificações morfológicas, que
foram analisadas no presente estudo. 59
Tabela 2. Nome e referências das seqüências dos primers usados na
amplificação e seqüenciamento. 67
Tabela 3. Número de espécies que foram identificadas pelo banco de dados
online BOLD (Barcode of life) e NCBI (National Center Biotechnology
Information).
71
Tabela 4. Valores médios das divergências intraespecífica e interespecífica do
gene COI (divergência mínima, média, máxima e desvio padrão) das 147
sequências obtidas pelo método de Kimura-2-parâmetros.
71
Tabela 5. Porcentagem média da distância intraespecífica (K2P) entre os
genêros de Ciclídeos amazônicos representados por mais que uma espécie. 72
Tabela 6. Porcentagem média da distância intraespecífica (K2P) para as espécies
de ciclídeos classificados morfologicamente e representados por mais de um
espécime.
72
Tabela 7. Porcentagem média da distância intraespecífica (K2P) para as espécies
de ciclídeos classificados molecularmente e representados por mais de um
espécime.
74
Tabela 8. Número de linhagens biológicas encontradas para cada espécie de
ciclídeos na Bacia Amazônica. 75
Tabela 9. Distância média da distância interespecífica (K2P) para as espécies de
ciclídeos delimitados molecularmente pelo modelo Generalized Mixed Yule
Coalescent (GMYC).
77
Tabela 10. Distância média da distância interespecífica (K2P) para as espécies
de ciclídeos classificados morfologicamente.
82

x
LISTA DE FIGURAS
Pág.
Figura 1. Representação da morfologia externa de um Ciclídeo. 14
Figura 2. Placas dentígeras faringianas em Cichlasoma minckleyi e Crenicichla chicha. 14
Figura 3. Mapa do genoma mitocondrial de Arapaima gigas representando os principais
genes. O citocromo c oxidase é mostrada em vermelho. 18
Figura 4. Esquema do barcoding gap. Em vermelho mostra a distribuição da variação
intraespecífica e em amarelo a variação interespecífica. 20
Capítulo I.
Figura 1. Localidades onde foram realizadas as amostragens das espécies de ciclídeos
realizados na bacia hidrográfica da Amazônia. Os números correspondem aos locais
de amostragem: 1 – Rio Araguaia; 2 – Rio Xingu; 3 – Rio Tapajós; 4 – Rio Purus; 5 –
Rio Jatapu; 6 – Rio Negro.
28
Figura 2. Árvore com as espécies delimitadas pelo modelo GMYC, construídas a
partir de 147 sequências de código de barras de DNA. 68

13
1. INTRODUÇÃO GERAL
1.1 Biologia e Ecologia dos Ciclídeos
Os primeiros estudos com a família Cichlidae foram realizados por Heckel em 1840, e
posteriormente por Jardine em 1843. Em 1904, Pellegrin conseguiu revisar todos os gêneros e
espécies da família conhecidas por ele com base em diagnósticos morfológicos. Entre 1905 a
1906 Regan fez uma série de revisões taxonômicas nas espécies e gêneros da família, o que
melhorou significativamente as classificações dos táxons Neotropicais (Kullander, 2003).
Porém, a primeira revisão filogenética para o grupo dos ciclídeos neotropicais foi feita apenas
em 1976 por Cichocki (Kullander, 2003). Mais recentemente, revisões taxonômicas com base
em dados morfológicos e moleculares foram feitos por Kullander (1998), Farias et al. (1999),
Farias et al. (2000), Sparks e Smith (2004), Kocher (2004), López-Fernandez et al. (2005),
Smith et al. (2008), López-Fernández et al. (2010), Hulsey et al. (2010), Hulsey et al. (2011) e
McMahan et al. (2013) proporcionando hipóteses filogenéticas com base na morfologia e
dados moleculares respectivamente.
Os peixes ciclídeos são conhecidos pela sua rápida especiação simpátrica e radiação
adaptativa - processos biológicos - nos quais permitiram a colonização dos lagos africano
(Malawi, Victoria e Tanganyika) e das regiões neotropicais (Genner et al., 2007; López-
Fernández et al. 2013; Hilary e López-Fernández, 2013). Somente nesses lagos, nos últimos
milhões de anos, surgiram mais de 1.000 espécies de peixes de água doce (Allender et al.,
2003).
Dessa forma, os ciclídeos formam um dos grupos mais diversos de peixes de água
doce do mundo, com aproximadamente 1900 espécies descritas (Kullander, 1998, 2003). A
diversidade encontrada neste grupo abrange os mais diferentes aspectos que variam desde as
características morfológicas e ecológicas, que facilitam na adaptação dos peixes desta família
(Kullander, 2003). Os ciclídeos neotropicais também constituem um grupo variado, no qual
compreendem cerca de 60 gêneros e estima-se que mais de 1300 espécies são reconhecidas
(López-Fernandez et al., 2010, Kullander, 2003). Apresentam 7-24 espinhos na nadadeira
dorsal e 2-12 na nadadeira anal, uma única narina em cada lado da cabeça e corpo
comprimido lateralmente. A linha lateral é geralmente dividida em uma porção anterior-
superior terminando abaixo da extremidade da base da nadadeira dorsal, e uma porção
posterior-inferior correndo ao longo do meio do pedúnculo caudal (Figura 1) (Kullander,

14
2003; Froese e Pauly, 2012).
As maxilas são geralmente móveis e apresentam protrusão (Froese e Pauly, 2012;
Sampaio e Guolart, 2011). Há uma variação considerável na forma da placa dentária
associada à dieta especializada. Também possuem placas dentígeras faringianas (Varela et al.,
2012) (Figura 2) utilizadas na mastigação que sustentam a monofilia do grupo (Kullander,
2003). Esse complexo maxilar da faringe que se localiza atrás da cavidade bucal é
funcionalmente dissociado das maxilas, e apresenta grande diversificação de formas e funções
entre as espécies desta família (Meyer 1993 apud Sampaio e Goulart, 2011).
De um modo geral, os ciclídeos possuem cuidado parental (incubação oral ou depósito
de ovos em superfícies) e o seu comprimento do corpo varia de 25-30 mm para as espécies
dos gêneros Apistogramma e Taeniacara a aproximadamente um metro em Cichla temensis
(Kullander, 2003).
A família também é caracterizada por diversos padrões de cores entre as espécies do
mesmo gênero, que são explicados pela atuação da seleção divergente ou disruptiva nas suas
colorações, nos quais permitiram formar uma ampla variação no grupo de ciclídeos (Allender
Figura 1. Representação da morfologia externa de um Ciclídeo. Fonte: Fishbase.
Figura 2. Placas dentígeras faringianas em Cichlasoma minckleyi (A e B) e Crenicichla chicha (C). Fonte: Figura
adaptada de Trapani, 2003 in: Sampaio e Goulart 2012; Varela et al., 2012.

15
et al., 2003). Essas colorações auxiliam na escolha do parceiro, onde os machos, geralmente
têm cores mais evidentes do que as fêmeas (Dalton et al., 2010).
Os ciclídeos apresentam diferenças de diversificação entre os clados de espécies
“fenotipicamente crípticas” e radiações que muitas vezes contêm espécies “geneticamente
crípticas” (Hulsey, 2009; Perazzo, 2010; Martins-Santos et al., 2014). Esses complexos de
espécies muitas das vezes, dificultam na delimitação das espécies de um grupo, pois em
alguns grupos distintos aparentam pouca ou nenhuma diferença fenotípica, enquanto que em
outros casos, apresentam grandes diferenças fenotípicas que facilitam na delimitação das
diferentes linhagens (Hulsey, 2009).
Até a década de 90, estimava-se que cerca de 160 táxons não tinham sido descritos na
América do Sul (Kullander, 1998, 2003). Porém, recentemente diversas espécies de ciclídeos
foram descritas morfologicamente, na bacia amazônica, entre elas: Apistogramma angayuara
e Apistogramma salpinction no rio Trombetas (Kullander e Ferreira, 2005); Geophagus sveni
e G. neambi na drenagem do rio Tocantins (Lucinda et al., 2010); Dicrossus foirni e
Dicrossus warzeli nos rios Tapajós e rio Negro (Römer et al., 2010); Krobia xinguensis no rio
Xingu (Kullander, 2012) e Crenicichla chicha no rio Tapajós (Varella et al., 2012). Em outras
localidades do Brasil, também já foram descritas novas espécies de ciclídeos: Laetacara
araguaiae na cidade de Rio Verde no estado de Goiás (Ottoni e Costa, 2009); Australoheros
perdi no Vale do Rio Doce no sudeste do Brasil (Ottoni et al., 2011); Australoheros tavaresi,
Australoheros mattosi e Australoheros montanus nas bacias dos rios São Francisco, Paraná e
Paraíba do Sul, sudeste do Brasil (Ottoni, 2012); Laetacara flamannellus no estado do Amapá
(Ottoni et al., 2012).
Quanto à distribuição da família, podem ser encontrados na América Central e do Sul,
Texas, Índias Ocidentais, África, Madagascar, Síria, Israel, Irã, Sri Lanka, e no litoral do sul
da Índia (Kullander, 2003; Froese e Pauly, 2012). Na América do Sul, habitam locais lênticos
dentro de rios e córregos, e algumas espécies dos gêneros Crenicichla, Teleocichla e
Retroculus são considerados reofílicos (Kullander, 2003).
Na América do Sul, o sistema fluvial amazônico passou por grandes transformações
geológicas, destacando-se a mudança no sentido da drenagem dos rios de leste-oeste para
oeste-leste (Lundberg et al., 1998; Albert et al., 2006). Ocorreu a formação do Lago Pebas
(15-10 Ma) que ocupou a Amazônia Ocidental, no qual foi separada da Amazônia Central e
Leste pelo Arco Purus, promovendo possivelmente a diversificação alopátrica. Com o
surgimento da cordilheira andina venezuelana e depois o surgimento do arco Uapes (12 Ma)
ocorreu o rompimento do Arco Purus, possibilitando a reconexão entre a Amazônia Ocidental

16
e a Amazônia Central e Leste, permitindo uma colonização recíproca de faunas endêmicas
nessas bacias. Além desses eventos geomorfológicos, as incursões marinhas também
influenciaram na diversificação e colonização de vertebrados na Amazônia (Lovejoy et al.,
2006; Lundberg, 1998; Farias e Hrbek, 2008).
A bacia amazônica é composta por três estruturas geológicas diferentes: (i) a
Cordilheira dos Andes, à Oeste; os (ii) Escudos Cristalinos, da Guiana ao Norte e do Brasil ao
Sul; e (iii) a planície sedimentar (Ruffino, 2004). Os rios são classificados de acordo com suas
condições físico-químicas, no qual se diferencia nas cores de águas brancas, claras e pretas
(Sioli, 1950). Os rios de águas brancas possuem suas nascentes na região andina e os
tributários são os afluentes da margem direita, como os rios Juruá, Purus e Madeira e na
margem esquerda, o rio Içá e Japurá. Os de águas claras, são transparentes e esverdeadas
devido as baixas quantidades de sedimentos e de sólidos dissolvidos, entre eles estão os rios
Tapajós, Xingu, Trombetas, Branco, Nhamudá, Tocantins, Araguaia e Paru se originam nas
encostas dos escudos das Guianas e do Brasil. A bacia sedimentar Amazônica possui cerca de
2 milhões de km2 e possui sedimentos dos Escudos Cristalinos e dos Andes que se
depositaram no vale Amazônico, durante o Terciário e Quaternário. Ela drena águas escuras
do rio Negro, Jutaí, Tefé e Coari (Junk et al., 2011; Junk et al., 2012; Ruffino, 2004).
Apesar dos diferentes processos geológicos e climáticos terem influenciados na
distribuição e diversificação dos ciclídeos amazônicos, estimativas a cerca da biodiversidade
do grupo ainda são incertas e muitas espécies necessitam ser identificadas e descritas (Farias
et al., 2001). Até o momento, a maioria das espécies foi identificada por meio de análises
morfológicas utilizando chaves de identificações (www.barcoding.si.edu), porém, caracteres
morfológicos podem não ser eficientes para diagnosticar as espécies de ciclídeos (Colatreli et
al., 2012). Pois, existem evidências de compartilhamento de haplótipos entre os diferentes
grupos fenotípicos e espécies (Farias e Hrbek, 2008; Colatreli et al., 2012; Willis et al., 2007;
Willis et al., 2012).
Os ciclídeos além de apresentarem rápido tempo de geração e idade reprodutiva,
possuem alta diversidade e adaptação, ampla distribuição, ocorrência de espécies crípticas,
podendo levar a subestimação de suas espécies. Também sofrem com a influência das
variações sazonais e fatores geológicos, no aspecto biológico, ecológico e genético do grupo,
o que pode afetar as suas populações (Crampton, 2008). Portanto, torna-se importante
entender a organização, distribuição e diferenciação genética das espécies deste grupo de
peixes numa determinada bacia hidrográfica, como a bacia Amazônica, de modo, que
posteriormente possam ser definidas melhores estratégias conservacionistas (Frankham et al.,

17
2008).
1.2 História Biogeográfica dos Ciclídeos
Os fósseis mais antigos dessa família datam do Eoceno na América do Sul e do
Oligoceno da África (Murray, 2001). Assim, acredita-se que surgiram ainda no Cretáceo e sua
a filogenia molecular e morfológica demonstra que os ciclídeos são monofiléticos (Stiassny,
1991; Farias et al., 1999; Chakrabarty 2004; Sparks e Smith, 2004, 2005).
Existem duas abordagens a cerca da distribuição disjunta dos ciclídeos nos
continentes, uma está relacionada com a dispersão que ocorreu pela transposição de uma
barreira geográfica, que colaborou na exploração de novas áreas e no processo de especiação.
Esta hipótese pode ser mais provável, devido à evidência do registro fóssil, estimativas de
divergência do relógio molecular e a tolerância ao sal de alguns ciclídeos (Murray, 2001;
Vences et al 2001; Briggs 2003).
De acordo com Murray (2001), com a fragmentação da Gondwanna (entre 135 a 65
milhões de anos), dois grupos teriam se formado distintamente: os ciclídeos africanos (Velho
Mundo) e os ciclídeos sulamericanos. A linhagem africana migrou para a América do Sul e
depois se espalhou facilmente para a América Central e Norte (Murray, 2001). Segundo esse
mesmo autor, existem muitos exemplos de espécies de ciclídeos que toleram a salinidade o
qual não seria uma barreira à dispersão desses ciclídeos no passado, e, portanto, a distribuição
desse grupo não é estritamente Gondwana.
E a outra abordagem se atribui ao evento vicariante que separou as populações por
uma fragmentação (separação do supercontinente da Gondwana), determinando assim, a
distribuição da fauna de ciclídeos (Farias et al., 2000; Chakrabarty, 2004; Sparks e Smith,
2005). Segundo essa hipótese, os estudos realizados com filogenia dos ciclídeos mostram
coerências com a hipótese da fragmentação da Gondwana. Como por exemplo, Farias et al.
(1999), mostraram que os ciclídeos de Madagascar e Indianos formavam linhagens basais,
além de serem grupo irmãos com os clados africanos e neotropicais, cuja relação coincide
com a separação do continente Madagáscar e Indiano. Os ciclídeos neotropicais e africanos
divergiram posteriormente após a abertura do Oceano Atlântico Sul. Já Spark e Smith (2004),
verificaram que o ancestral comum sofreu divisão, formando dois clados: o Etroplinae no
continente Madagáscar-Índia, e o clado Ptychochrominae formado Madagáscar e África +
América do Sul. O grupo formado por Madagáscar e Índia (Etroplinae) foi separado no

18
Cretáceo, dessa forma, baseado nessas relações e nas evidências geológicas de peixes
ciclídeos, eles argumentaram que a atual distribuição de Cichlidae é congruente com
vicariância Gondwana (Spark e Smith, 2004).
1.3 Código de Barras de DNA
Atualmente existem limitações em estudos taxonômicos que são baseados apenas em
análises morfológicas, sendo assim, a inclusão de dados moleculares torna-se oportuno e
necessário para discriminar e identificar os táxons (Moritz e Cicero, 2004), como por
exemplo, os ciclídeos (Ribeiro, 2007).
Em 2003, pesquisadores da Universidade de Guelph, em Ontário, Canadá, propuseram
um "código de barras de DNA", que poderia auxiliar no processo de identificação e
delimitação das espécies (Savolainen et al., 2005). Esse código de barras se baseia num
pequeno fragmento padronizado, com apenas 648 pares de bases, do genoma mitocondrial
(DNAmt), o citocromo c oxidase subunidade I (COI) (figura 3) (Hebert et al., 2003a; Hebert
et al., 2003b; Stoeckle, 2003).
A escolha pelo DNAmt para distinguir as espécies visou pelas suas maiores diferenças
nas sequências dos ácidos nucléicos entre as espécies e pela abundância de cópias em relação
ao DNA nuclear, tornando-o bastante informativo e fácil de ser recuperado a partir de
amostras pequenas ou parcialmente degradas (Stoeckle e Hebert, 2008). Ele é compartilhado
entre táxons variados, possui elevadas substituições neutras na terceira posição do nucleotídeo
Figura 3. Mapa do genoma mitocondrial de Arapaima gigas representando os principais genes. O citocromo c
oxidase é mostrada em vermelho (Hrbek e Farias, 2008).

19
e não contêm íntrons, os quais podem dificultar a amplificação usando a reação em cadeia da
polimerase (PCR) (Stoeckle, 2003; Hebert et al., 2003a).
O código de barras de DNA é caracterizado pela presença de primers universais de
uma região específica que flanqueiam regiões das sequências conservadas; codifica parte da
enzima terminal da cadeia respiratória mitocondrial e gera sinais filogenéticos (Hebert et al.,
2003b; Hebert e Gregory, 2005). Entre os fatores que determinam a efetividade deste sistema
taxonômico estão: baixa divergência intraespecífica, estruturação populacional e o tamanho
efetivo populacional (Meyer e Paulay, 2005; Ortiz, 2010).
A variação intraespecífica consiste nas diferenças genéticas comuns entre indivíduos
da mesma espécie. A divergência interespecífica são as diferenças que se acumulam entre as
espécies (Hartl, 2008). A sobreposição entre as distâncias genéticas entre espécies
estreitamente relacionadas (divergência interespecífica) e maior variação intraespecífica
originam espécies geneticamente polifiléticas e parafiléticas, dificultando a delimitação dos
organismos pelo marcador COI (Meyer e Paulay, 2005; Tofolli et al., 2008). Assim, as
variações intraespecíficas devem ser sempre menores que as divergências interespecíficas
para que as espécies sejam reconhecidas através deste método (Figura 4) (Hebert 2003b;
Stoeckle, 2003; Ortiz, 2010).
Dessa forma, a variação intraespecífica é inversamente proporcional ao tamanho do
barcoding gap. E a estrutura populacional também pode afetar o barcoding gap na medida em
que ele irá afetar a variação intraespecífica (Ortiz, 2010). Por isso, é importante a amostragem
para melhor desempenho do código de barras, uma vez que o número amostral e dos pontos
de coletas forem insuficientes, pode-se interferir nos resultados e acabar subestimando as
variações intraespecíficas e superestimar as divergências interespecíficas e facilitar na
visualização do barcoding gap (Meyer e Paulay, 2005; Ortiz, 2010).
Assim, o barcoding gap é uma diferença clara entre as distâncias genéticas
interespecíficas e intraespecíficas que facilita a delimitação dos táxons. O critério consiste em
que a distância genética interespecífica deve ser 10 vezes maior do que a distância
intraespecífica (Hebert et al., 2004b; Wiemers e Fiedler, 2007). Em casos de alta variação
intraespecífica o barcoding gap torna-se ausente (Ortiz, 2010). Para Barret e Hebert (2005) é
fundamental determinar esses intervalos de variação intra e interespecífica entre os diferentes
grupos taxonômicos.
Outro critério importante para a existência do barcoding gap é o tamanho efetivo
populacional (Ne). Pois, em casos que existam Ne grande necessita-se de mais tempo para a
separação total e diferenciação entre as populações, além de aumentar a probabilidade de se

20
encontrar alta variação intraespecífica, e consequentemente, a não visualização do barcoding
gap (Ortiz, 2010; Toffoli et al., 2008).
Desde o início da descoberta e utilização do código de barras de DNA em diversas
pesquisas, essa metodologia vem sofrendo algumas críticas. Os críticos argumentam algumas
falhas nesse novo método molecular, entre elas, o uso de distâncias genéticas para a
construção de árvores e utilizar árvores a partir de um único gene para inferir filogenias
(DeSalle et al. 2005); divergência genética interespecífica para delimitar as espécies (Moritz e
Cicero 2004; Wiemers e Fiedler 2007); além do baixo desempenho numa amostragem
insuficiente (Meyer e Paulay, 2005). Outros autores afirmam que um pequeno segmento do
DNA mitocondrial como uma única fonte de dados, não pode representar o genoma completo
para estudo da biodiversidade e substituir a taxonomia clássica (Dunn, 2003; Lipscomb et al.,
2003; Mallet e Willmott, 2003; Will e Rubinoff, 2004; Ebach e Holdrege, 2005; Will et al.,
2005).
Porém, os defensores afirmam que o código de barras de DNA está cada vez mais
refinado e a caminho de tornar-se uma ferramenta essencial para identificação e delimitação
das espécies (Hajibabaei et al. 2006; Kress e Erickson 2008; Janzen et al., 2009). A
abordagem permite delimitar grupos geneticamente distintos, principalmente espécies
crípticas (Hebert et al. 2004a), de forma rápida e precisa (Hebert e Gregory, 2005; Gómez et
al., 2007), que são espécies indistinguíveis morfologicamente que vivem em localidades
A
A
B
Distância Genética
Figura 4. Esquema do barcoding gap. Em vermelho mostra a distribuição da variação intraespecífica e
em amarelo a variação interespecífica. (A) Para efetividade do DNA Barcoding, com distribuições discretas e sem
sobreposição. (B) Uma versão alternativa do mundo com significativa sobreposição e sem gap (Fonte: Meyer e
Paulay, 2005).

21
diferentes, porém não se intercruzam (Futuyma, 2002). E também os complexos de espécies
que são morfologicamente semelhantes e vivem numa mesma área, mas não se intercruzam.
Para Rach et al. (2008), a utilização dos códigos de barras de DNA baseado em
presença ou ausência de substituições de nucleotídeos distintos, mostrou-se ser uma
ferramenta eficaz para a identificação, discriminação e delimitação dos grupos em qualquer
nível taxonômico.
Essa nova ferramenta já foi utilizada em diversos trabalhos com grupos de
invertebrados e vertebrados, tais como: insetos (Hebert et al., 2004a; Hajibabaei et al., 2006;
Rach et al., 2008; Janzen et al., 2009; Silva-Brandão et al., 2009; Dincã et al., 2011; Park et
al., 2011; Webb et al., 2012); anelídeos (Oceguera-Figueroa et al., 2010); aves (Hebert et al.,
2004b); morcegos (Clare et al., 2011) e principalmente para o grupo dos peixes (Ward et al.,
2005, 2009; Frederico et al. 2012; Chakrabarty, 2006). Pereira et al. (2011a) observaram que a
espécie de Piabina argentea é formada por mais de uma unidade biológica. Colatreli et al.
(2012), verificaram a existência de cinco espécies hipotéticas para o ciclídeo do gênero
Astronotus. Pereira et al. (2011b) utilizaram o código de barras de DNA em peixes de água
doce da bacia Paraíba do Sul no estado de São Paulo e conseguiram discriminar todas as 58
espécies analisadas, entre elas a espécie Geophagus proximus. León-Romero et al. (2012)
utilizaram o código de barras de DNA no grupo de espécies Herichthys bartoni para testar a
monofilia e eficência desta ferramenta para discriminar as espécies. Nwani et al. (2011)
aplicaram o código de barras em 38 gêneros de famílias diferentes em peixes de água doce na
Nigéria e verificaram que o COI conseguiu separar a maioria das espécies de ciclídeos, com
exceção para as espécies Oreochromis niloticus e Sarothrodon boulengeri que houve
compartilhamento de haplótipos.
As ferramentas moleculares vêm contribuindo cada vez mais para a identificação das
espécies crípticas e a utilização das sequências das diversas pesquisas conservacionistas com
o código de barras de DNA, são atualmente, inseridas no banco de dados de código de barras
da vida (BOLD, www.barcodinglife.org), juntamente com uma imagem e informação de
garantia para cada espécime voucher (Hajibabaei et al., 2006). O Banco de Dados do Código
de Barras da Vida (em BOLD) consiste numa plataforma de informática que auxilia na
aquisição, armazenamento, análise e publicação de códigos de barras de DNA (Ratnasingham
e Hebert, 2007).
A análise dos dados, do projeto código de barras da vida é realizada pelo CBOL
(Consórcio para o Código de Barras da Vida), que é uma colaboração internacional de 170
organizações com membros de 50 países. O objetivo é estabelecer uma biblioteca pública de

22
sequências de DNA ligadas aos espécimes nomeados (www.barcoding.si.edu).
Existe uma necessidade de estabelecer e fazer cumprir as normas dos dados e para
isso, a CBOL iniciou diálogo com os grandes repositórios genômicos, como por exemplo,
National Center for Biotechnology Information – NCBI. Esse sistema estabelece orientações
formais que devem ser cumpridas para os registros obter a designação de código de barras de
DNA.
Este trabalho constitui-se então num aporte importante para consolidação da base de
dados no projeto universal Barcode da Vida, não só para a delimitação e identificação de
espécies no nível mundial, mas, também para o conhecimento do estado local da nossa
subestimada biodiversidade de ciclídeos no Amazonas, aspecto que limita a execução de
políticas protecionistas para as espécies, especialmente os peixes, fonte importante de renda e
alimento.
2. OBJETIVO
2.1. Objetivo geral:
Analisar a diversidade dos peixes da família Cichlidae oriundos de oito rios da bacia
Amazônica por meio do gene mitocondrial CO1.
2.2. Objetivos específicos:
- Gerar códigos de barras de DNA para as espécies de peixes da família Cichlidae
amostradas em seis rios amazônicos (Rio Negro, Purus, Jatapu, Tapajós, Xingu e
Araguaia/Tocantins);
- Verificar a hipótese da ferramenta de código de barras de DNA para delimitar e
identificar espécies de ciclídeos;
- Identificar presença de espécies crípticas e complexos de espécies nos ciclídeos;
- Observar casos de não monofilia nas espécies de ciclídeos amostrados.

23
CAPÍTULO I1
1 Manuscrito formatado de acordo com as normas da revista Mitochondrial DNA. Manuscrito a ser
submetido.

24
Código de barras de DNA para identificar espécies de ciclídeos da bacia Amazônica, 1
Brasil. 2
ANA PAULA C. CARVALHO 1, JOSÉ G. MARTÍNEZ 1, IZENI P. FARIAS 1 & TOMAS 3
HRBEK 1 4
1 Laboratório de Evolução e Genética Animal, Departamento de Biologia, Universidade 5
Federal do Amazonas, Av. Rodrigo Octávio Jordão Ramos, 3000, 69077-000, Manaus, AM, 6
Brasil. 2 7
Resumo 8
Nesse estudo foram analisadas 45 espécies de peixes de água doce da família Cichlidae em 9
seis localidades da bacia Amazônica, Brasil, por meio do código de barras de DNA. As 10
delimitações moleculares apresentaram distâncias genéticas médias interespecíficas que 11
variaram de 0%-55% e distância média intraespecífica com variação de 0%-1%. Dentre os 12
táxons analisados molecularmente foi possível identificar sete espécies crípticas (15,5%) e 13
sete complexos de espécies (15,5%) e aproximadamente cinco espécies (11,2%) mostraram 14
ausência de monofilia recíproca. Os dados demonstram que a taxonomia dos ciclídeos 15
Amazônicos ainda não está totalmente resolvida, sendo necessária a inclusão da taxonomia 16
integrativa para a resolução dos complexos de espécies aqui encontrados. 17
18
Palavras-chave: taxonomia, peixes de água doce, mtDNA, Cichlidae. 19
20
Correspondência: A. P. C de Carvalho, Laboratório de Evolução e Genética Animal, Departamento de Biologia,
Universidade Federal do Amazonas, Av. Rodrigo Octávio Jordão Ramos, 3000, 69077-000, Manaus, AM,
Brasil. Tel: +55 92 981057917. E-mail: [email protected]

25
Introdução 21
A família Cichlidae é composta por aproximadamente 1900 espécies (Kullander, 22
2003), com variadas características morfológicas e ecológicas que permitem a fácil adaptação 23
nos mais diferentes hábitats (Kullander 2003; Genner et al. 2007). Os grupos desta família 24
podem ser encontrados desde a América Central e Sul até o litoral sul da Índia (Kullander 25
2003; Froese e Pauly 2012). 26
Cerca de 60 gêneros e pelo menos 600 espécies são encontradas na região Neotropical 27
(López-Fernandez et al. 2010). Destas, 257 espécies são encontradas na bacia Amazônica 28
(Thompson 2010), com ampla distribuição contemplando toda a calha principal do rio 29
Amazonas e em seus afluentes (Kullander 2003). 30
As espécies dessa família sofrem pressão de pesca significativa em decorrência de sua 31
importância econômica (Chao 2001; Fernandez Acosta et al. 2008), que coloca em risco as 32
suas populações. Porém poucos estudos têm sido realizados para descrever diversas espécies 33
na Amazônia (Lewinsohn e Prado 2005). 34
Dessa forma, a alta diversidade morfológica e genética, favorece a presença espécies 35
crípticas e complexo de espécies, tornando a taxonomia instável e proporcionando diversas 36
mudanças na nomenclatura e na indefinição entre as espécies (Hulsey 2009; Amado et al. 37
2011; Anseeuw et al. 2012; Colatreli et al. 2012, Mejía et al. 2015). Além dessa grande 38
riqueza, o compartilhamento de haplótipos entre os diferentes grupos fenotípicos das espécies 39
(Willis et al. 2007; Farias e Hrbek 2008; Colatreli et al. 2012; Willis et al. 2012a) e a pouca 40
amostragem em diversas localidades dos diferentes rios amazônicos, acabam subestimando o 41
número evidente de espécies existentes (Kullander 1998). Isto faz com que se torne difícil à 42
aplicação de práticas conservacionistas das suas populações (Crampton 2008). Vários estudos 43
filogeográficos têm sido realizados para distinguir espécies crípticas e elucidar os casos de 44
polimorfismos intraespecíficos em ciclídeos (Willis et al. 2012a; 2012b; Musilová et al. 45

26
2008). Desta maneira, trabalhos adicionais são necessários com o fim de determinar não só 46
novas metodologias para o reconhecimento das espécies, como também a sua aplicabilidade e 47
eficiência na delimitação de espécies de ciclídeos como base para a sua conservação e 48
manejo. 49
Considerando as limitações da taxonomia tradicional para a delimitação de espécies, 50
torna-se necessário a inclusão de dados do código de barras de DNA para detectar as unidades 51
geneticamente distintas e facilitar na descoberta de novas espécies (Ward et al. 2009; Mejía et 52
al. 2015), especialmente em ambientes cada vez mais vulneráveis à pressão antrópica na bacia 53
Amazônica. 54
O código de barras tem surgido com sucesso na aplicação em várias espécies de peixes 55
ao redor do mundo (Ward et al. 2005; Ward et al. 2009), permitindo criar grandes bases de 56
dados como FISH-BOL (Campanha do Código de Barras da Vida para Peixes) 57
(http://www.fishbol.org), envolvendo dados de referência para sequências de DNA para 58
espécies do grupo (Becker et al. 2011; Léon-Romero et al. 2012, Bhattacharya et al. 2015). 59
Aliado a isso, esse novo método de identificação (COI) tem facilitado no controle (Steinke et 60
al. 2009) e regulamentação de práticas conservacionistas de peixes da bacia amazônica 61
(Ardura et al. 2010; Aquilino et al. 2011). 62
Uma região padronizada e curta com apenas 648 pares de base do gene mitocondrial, 63
citocromo oxidase I (COI), denominado código de barras de DNA, é utilizada atualmente para 64
delimitações rápidas e precisas de espécies de animais (Hebert et al. 2003a; Hebert et al. 65
2004; Hebert e Gregory 2005; Stoeckle e Hebert 2008). 66
Essa metodologia se baseia em dois principais pressupostos que mostram a eficácia do 67
código de barras de DNA: i) monofilia recíproca das espécies (Meyer e Paulay 2005) e ii) as 68
diferenças genéticas intraespecíficas que são sempre menores do que as diferenças genéticas 69
entre as espécies (divergência intraespecífica 10x maior do que a divergência interespecífica) 70

27
(Hebert et al 2003b; Stoeckle 2003). 71
Neste sentido, o presente estudo teve como objetivo, gerar códigos de barras de DNA 72
para as espécies e verificar a hipótese que ferramenta molecular (COI) pode discriminar e 73
delimitar as espécies de ciclídeos na bacia Amazônica. 74
75
Material e métodos 76
Amostragem 77
Foram obtidas 147 sequências do código de barras de DNA (COI) de 16 gêneros, 45 78
espécies da família Cichlidae (Tabela 1) de seis rios da bacia amazônica (Figura 1), com 79
amostragem de até cinco espécimes por espécies. Os espécimes foram coletados por meio da 80
licença do IBAMA n° 11325-1. 81
Para cada exemplar amostrado, coletou-se uma pequena amostra de tecido do 82
pedúnculo caudal ou da nadadeira peitoral esquerda. Os tecidos foram armazenados em etanol 83
a 95% até o processamento em laboratório e os vouchers foram preservados em formaldeído a 84
10% e conservados em etanol a 70%. 85
Todos os tecidos foram depositados na Coleção de Tecidos de Genética 86
Animal/CTGA–ICB/UFAM (CGEN) do Laboratório de Evolução e Genética Animal, 87
Universidade Federal do Amazonas (LEGAL/UFAM) e os vouchers foram depositados na 88
coleção de peixes do Instituto Nacional de Pesquisas da Amazônia (INPA). 89

28
90
Figura 1. Localidades das amostragens das espécies de ciclídeos da bacia Amazônica. Os 91
números correspondem aos locais de amostragem: 1 – rio Araguaia; 2 – rio Xingu; 3 – rio 92
Tapajós; 4 – rio Purus; 5 – rio Jatapu; 6 – rio Negro. 93
94
Extração, PCR e Sequenciamento 95
O DNA foi isolado por meio do método fenol-clorofórmio (Sambrook e Russel, 2001). 96
Uma região parcial do gene mitocondrial citocromo oxidase subunidade 1 (COI) foi 97
amplificado via reação em cadeia da polimerase (PCR) com um volume final de 15 μL 98
contendo: 6,8 μL de H2O ultrapura; 1,5 μL de MgCl2 (25 mM); 1,5 μL de dNTPs (10 mM); 99
1,5 μL de tampão 10x (100mM Tris-HCl, 500mM KCl); 1,2 μL de cada primer (2 μM) 100
(Tabela 2); 0,3 μL de Taq DNA Polymerase (1 U/μL) e 1 μL de DNA (concentração variando 101
de 30 a 60 ng) em termociclador Veriti® Thermal Cycler (Applied Biosystems). 102
As condições da PCR consistiram em 35 ciclos de desnaturação a 93°C por 10 103

29
segundos, anelamento a 50°C por 35 segundos e extensão a 68°C por 1 minuto e 30 segundos. 104
A amplificação foi verificada em gel de agarose 1%, acrescido de gel red (Biotium) e 105
marcador (GeneRuler 1kb DNA Ladder). 106
Os produtos de PCR foram purificados com o kit ExoSap de acordo com as 107
recomendações do fabricante e submetido as reações de sequenciamento foi utilizado o Kit 108
Big Dye Terminator Cycle Sequencing Standart Version 3.1 (Applied Biosystems, Inc), para 109
um volume final de 10 µL por amostra, composta por: 4,4 µL de água ultrapura; 1,3 µL de 110
tampão de sequenciamento 5X; 2,0 µL primers forward (M13F -21) (2 μM) e/ou reverse 111
(M13R -27) (2 μM) (Tabela 2); 0,3 µL de BigDye Terminator v 3.1 (Applied Biosystems, 112
Foster City, CA, USA) e 2 µL de DNA purificado. 113
Os produtos resultantes dessa reação foram precipitados com etanol (100% e 70%) e 114
EDTA (125mM), em seguida ressuspendidas com 10 µL Hi-Di formamida e submetidas ao 115
sequenciador automático ABI 3130xl (Applied Biosystems Inc.). 116
117
Análises dos dados 118
As sequências foram editadas, alinhadas e traduzidas no programa Geneious v. 5.4 119
(Drummond et al. 2011). Posteriormente foram submetidas ao programa BLAST (Altschul et 120
al. 1997) para confirmação e comparação da região sequenciada com os dados disponíveis no 121
National Center for Biotechnology Information (Genbank) (http://www.ncbi.nlm.nih.gov) e 122
do Barcoding of Life Database (BOLD) (Ratnasingham e Hebert 2007). 123
A delimitação das espécies foi realizada pelo modelo Generalized Mixed Yule 124
Coalescent (GMYC) (Fujisawa e Barraclough 2013), no programa estatístico R versão 3.0.2 125
(R Development Core Team 2013). As espécies delimitadas pelo GMYC e as espécies 126
identificadas morfologicamente foram obtidos valores de distâncias médias intraespecíficas e 127
interespecíficas e distância par-a-par pelo método de Kimura-2-parâmetros no programa 128

30
MEGA 5.05 (Tamura et al. 2011). 129
130
Resultados 131
Foram obtidas sequências do código de barras de DNA (COI) de 16 gêneros e 45 132
espécies da família Cichlidae. Sendo que Crenicichla foi o gênero que apresentou maior 133
número de espécies (11) e os gêneros Acarichthys, Acaronia, Biotodoma, Caquetaia, Heros, 134
Laetacara, Mesonauta e Symphysodon apresentaram menores números de espécies (1) 135
analisadas (Tabela 5). 136
Foram encontradas sequências de COI no Genbank e BOLD para apenas 13 espécies, 137
sendo elas: Apistogramma agassizi, Geophagus argyrostictus, Geophagus altifrons, Heros 138
efasciatus, Symphysodon aequafasciatus, Acaronia nassa, Cichla piquiti, Acarichthys 139
heckelii, Retroculus lapidifer, Retroculus xinguensis, Satanoperca lilith, Teleocichla sp., 140
Crenicichla percna (Tabela 3). 141
Cerca de 40 espécies apresentaram monofilia, com exceção de duas espécies do gênero 142
Teleocichla (T. preta e T. cf. gephyrogramma) do rio Xingu e três espécies de Crenicichla (C. 143
jegui, C. lugubris e C. aff. strigata) do rio Araguaia, que apresentaram parafilia (Figura 2). 144
As divergências genéticas interespecíficas variaram de 0% a 17%, com média de 7% 145
entre os gêneros representados por mais de uma espécie (Tabela 5). As divergências genéticas 146
médias intraespecíficas para as espécies identificadas morfologicamente variaram de 0% a 147
8%, com média de 2%. De acordo com as identificações morfológicas, foram identificadas 45 148
espécies, sendo que 24 espécies (53,3%) apresentaram distâncias médias intraespecíficas ≤ 149
2% e 7 espécies (15,5%) tanto da mesma localidade quanto de locais diferentes foram 150
encontrados valores de divergências genéticas médias intraespecíficas >2% e separados em 151
clados pela árvore GMYC (Tabela 6 e Figura 2). 152
Para os indivíduos amostrados dentro das mesmas localidades, tem-se o exemplo da 153

31
espécie Teleocichla cinderela, amostradas nos pedrais de São Miguel e São Bento, 154
localizados no rio Araguaia, apresentaram distância média intraespecífica de 6% e pelas 155
análises GMYC apresentaram-se em diferentes clados. A espécie Geophagus argyrostictus da 156
localidade Xingu, também apresentaram distância média intraespecífica >2% e foram 157
separados em dois clados pelas análises moleculares. 158
E as espécies Apistogramma agassizi (3%), Acaronia nassa (3%), Crenicichla regani 159
(3%), Geophagus altifrons (7%) e Geophagus sp. (3%), amostrados de diferentes localidades 160
(Purus, Jatapu, Tapajós, Xingu) e com tipos de águas diferentes (branca, preta e clara), 161
também apresentaram valores de divergências genéticas médias intraespecíficas maiores que 162
2% e foram delimitados em clados de acordo com a respectiva localidade pelo modelo 163
GMYC (Figura 2). 164
Observou-se que algumas espécies identificadas pela taxonomia morfológica como 165
espécies diferentes, apresentaram valores de distâncias par-a-par menores que 2%. Como por 166
exemplo, as espécies de Cichla temensis do rio Jatapu e Cichla melaniae do rio Xingu; 167
Crenicichla lugubris, C. jegui, C. strigata do rio Araguaia; Teleocichla cf. gephyrogramma e 168
T. n sp. preta do rio Xingu, identificadas morfologicamente como duas ou mais espécies e 169
delimitadas pelo GMYC também como espécies diferentes (Figura 2), porém a distância 170
interespecífica foi de 0-2% para as duas delimitações (Tabela 9 e 10). Em outras situações o 171
modelo GMYC delimitou algumas espécies com maior número de linhagens (Tabela 8 e 9) do 172
que identificadas morfologicamente (Tabela 10), porém os valores de divergências 173
interespecíficos foram < 2%. 174
Comparando as duas ferramentas (taxonomia tradicional e molecular) neste estudo 175
para delimitar as espécies de ciclídeos, apenas nove espécies apresentaram congruência pelas 176
técnicas utilizadas na delimitação, havendo, portanto, concordância entre elas. 177
As divergências médias intra (0%) e interespecíficas (32%) foram encontradas para as 178

32
espécies identificadas molecularmente (Tabela 4), apresentando um gap cerca de 30 vezes 179
maior entre espécies do que dentro das espécies. 180
Foram identificadas de 1 a 4 linhagens biológicas entre as 45 espécies analisadas neste 181
estudo, sendo as espécies Apistogramma agassizi, Aequidens pallidus e Crenicichla regani 182
com maiores números de linhagens (4) (Tabela 8). 183
184
Discussão 185
A ictiofauna Amazônica é bastante diversa e por isso é importante conhecer essa 186
riqueza, para que seja realizado o monitoramento dos peixes comercializados que sofrem 187
riscos de extinção. Para isso é necessário práticas conservacionistas que visem uma gestão nos 188
recursos pesqueiros da região. E a técnica do código de barras de DNA pode ser utilizada para 189
identificar com precisão as espécies ameaçadas comercialmente e na descrição das espécies 190
subestimadas (Benzaquem et al. 2015; Kress et al. 2015). 191
Os ciclídeos apresentam ampla distribuição geográfica e uma grande diversidade de 192
espécies que se adaptam rapidamente em diferentes habitats, onde muitas delas ainda não 193
foram descritas (Reis et al. 2003). Além disso, o compartilhamento de haplótipos entre os 194
diferentes grupos fenotípicos das espécies (Willis et al. 2007; Farias e Hrbek 2008; Colatreli 195
et al. 2012; Willis et al. 2012a) e os complexos de espécies podem levar a subestimação dessa 196
diversidade (Albert et al. 2006; Kullander et al. 2003; Farias e Hrbek 2008). A utilização 197
somente da morfometria tradicional e contagens merísticas não é possível separar algumas 198
espécies de ciclídeos, devido aos altos níveis de sobreposição entre caracteres taxonômicos 199
(McMahan et al. 2011; Soria-Barreto et al. 2011; Mejía et al. 2015). 200
Nas análises moleculares foi possível identificar 14 espécies a mais em relação às 201
identificações morfológicas. O que corrobora com o estudo de Winterbottom et al. (2014) que 202

33
identificaram 42 espécies a mais através do código de barras de DNA em peixes do gênero 203
Trimma. 204
Os valores de distâncias para as espécies identificadas morfologicamente que 205
apresentaram valores intraespecíficos > 2% também foram delimitados em diferentes clados, 206
indicando possíveis novas espécies, tais como Apistogramma agassizi, Aequidens pallidus, 207
Acaronia nassa, Crenicichla regani, Geophaugs altifrons, Geophagus sp., Teleocichla 208
cinderela (Tabela 6 e Figura 2). 209
Para essas mesmas espécies juntamente com Mesonauta festivus, e com exceção da 210
espécie Teleocichla cinderela, provenientes de localidades diferentes (Purus, Jatapu, Tapajós 211
e Xingu) e com tipos de água diferentes (branca, preta e clara) foram delimitados em 212
diferentes clados pelo modelo GMYC (Figura 2), formando mais de uma linhagem biológica. 213
Segundo Hulsey (2009) os complexos de espécies muitas das vezes, dificultam na delimitação 214
das espécies de um grupo, pois em alguns grupos distintos aparentam pouca ou nenhuma 215
diferença fenotípica. Pereira et al. (2011a; 2011b) e Pazian et al. (2012) identificaram 216
complexos de espécies de Piabina argentea (Characidae) em águas doce neotropicais. Mejía 217
et al. (2015) utilizaram o código de barras de DNA no complexo de espécies de ciclídeos 218
Herichthys bartoni e verificaram a existência de três grupos genéticos composto por seis 219
espécies. 220
Neste estudo as espécies Apistogramma agassizi, Aequidens pallidus, Geophagus 221
argyrostictus, Cichla melaniae, Crenicichla n sp. preta e Teleocichla cinderela da mesma 222
localidade (Jatapu, Xingu e Araguaia) foram delimitados em mais de um clado (Figura 2). A 223
presença de espécies crípticas pode subestimar o número evidente de espécies existentes em 224
uma mesma localidade (Colatreli et al. 2012, Kadarusman et al. 2012). Winterbottom et al. 225
(2014) utilizaram a ferramenta molecular do código de barras de DNA e descobriram uma 226
grande diversidade oculta de peixes do Canadá. Enquanto que na região Amazônica, 227

34
Benzaquem et al. (2015) verificaram a diversidade críptica em Nannostomus (Lebiasinidae). 228
Na Amazônia, a alta diversidade críptica dentro de áreas alagadas que não apresentam 229
barreiras, proporcionam a divergência da população e o processo de especiação (Crampton 230
2008; Piggoti et al. 2011), propiciando grandes desafios para o reconhecimento das espécies 231
pelas ferramentas moleculares. 232
A bacia Amazônica é um mosaico com diferentes tipos de água conectados pelo rio 233
Amazonas. As condições hidrográficas (variáveis físicas e químicas) podem agir como 234
barreiras para o fluxo gênico para a dispersão de peixes atuando como forças seletivas que 235
propiciam a especiação alopátrica (Lovejoy e Araújo 2000; Toffoli et al. 2008; Ducan e 236
Fernandes 2010). 237
Foram observadas as distâncias médias interespecíficas, nas análises moleculares < 238
2% entre espécies diferentes, sinalizando compartilhamento de haplótipos. Os resultados 239
apontaram que duas espécies dos gêneros Teleocichla (Teleocichla preta e Teleocichla cf. 240
gephyrogramma) e três espécies de Crenicichla (Crenicichla jegui, Crenicichla lugubris e 241
Crenicichla lugubris) apresentaram parafilia. Para Meyer e Paulay (2005) as espécies 242
parafiléticas são aspectos limitantes para a ferramenta molecular discriminar as espécies. 243
Resultados semelhantes de parafilia foram encontrados por Musilová et al. (2008) para os 244
gêneros de ciclídeos Aequidens e Cichlasoma e por Toffoli et al. (2008) nas espécies de 245
arraias de água doce. 246
Quando o processo de coalescência não é suficiente para separar as espécies, ocorrem 247
às sobreposições da variação intraespecífica com a interespecífica originando espécies 248
parafiléticas, causadas pela retenção do polimorfismo ancestral ou introgressão após o 249
processo de hibridação (Hickerson et al. 2010; Avise 2004; Moritz e Cícero 2004). Esses 250
fatores podem explicar o compartilhamento de haplótipos entre as espécies de Teleocichla e 251
Crenicichla, observados neste trabalho, como também pode ter havido erro no processo de 252

35
identificação baseados em caracteres morfológicos para estas espécies. 253
Hubert et al. (2012) observaram compartilhamento de haplótipos em 8% das 190 254
espécies de peixes canadenses, suspeitando-se de que as espécies não poderiam ser separadas 255
devido à radiação recente e hibridação, ou até mesmo alguns pares de espécies com 256
sobreposição de haplótipos seriam talvez uma única espécie. 257
Para Ward et al. (2009) e Pereira et al. (2013) a radiação recente na ictiofauna 258
neotropical para diversas espécies de peixes e a hibridação podem interferir no uso correto do 259
código de barras de DNA. A delimitação das espécies de alguns gêneros da família Cichlidae 260
também tem sido problemática, devido ao compartilhamento de haplótipos de DNA 261
mitocondrial entre as espécies (Farias e Hrbek 2008; Willis et al. 2012a; Willis et al. 2012b). 262
A metodologia do código de barras de DNA é simples, cujas seqüências são obtidas a 263
partir de vários indivíduos. Os indivíduos semelhantes e relacionados são agrupados e existe 264
uma variação genética dentro e entre as espécies, no qual as distâncias genéticas entre as 265
espécies são maiores do que dentro das espécies (Damasphatra e Mallet 2006). O modelo de 266
distância Kimura-2-Parâmetros (K2P) é utilizado no nível de espécie e é o melhor modelo 267
utilizado, quando as distâncias intraespecíficas são baixas (Miyaki et al. 2001). 268
Hebert et al. 2004, propôs um limiar de 2% para delimitar as espécies através do 269
código de barras de DNA, este limite baseia-se na distribuição de valores de distância 270
genética (K2P) intra e inter-específicos. De acordo com Hebert et al. (2003), Johns e Avise 271
(1998) as divergências intraespecíficas raramente são maiores que 2%, e as divergências 272
interespecíficas para a maioria dos vertebrados são superiores a 2%. Assim, para as espécies 273
com distâncias menores que 2% podem consideradas a mesma espécie, e acima de 2% 274
consideradas como espécies diferentes. Mesmo assim, o limiar proposto por Hebert et al. 275
2004 deve ser cuidadosamente analisado para cada grupo (Ward et al. 2009). 276
O valor de 2% foi o mais utilizado para delimitar as espécies de peixes por alguns 277

36
pesquisadores (Pereira et al. 2013; Hubert et al. 2008; Ward et al. 2009; Carvalho et al. 2011), 278
portanto, para algumas espécies de ciclídeos esse limiar não foi tão eficiente, como por 279
exemplo, para as espécies de Cichla temensis do rio Jatapu e Cichla melaniae do rio Xingu 280
identificadas morfologicamente como duas espécies e delimitadas pelo GMYC também como 281
espécies diferentes, porém a distância interespecífica foi de 2%. Em outras situações o modelo 282
GMYC delimitou alguns gêneros com maior número de espécies do que identificadas 283
morfologicamente, porém os valores de divergências interespecíficos foram < 2%. Isso 284
significa que para as análises que abrange uma grande diversidade de espécies, a utilização de 285
um limiar padrão para todas elas não seja muito adequado. 286
A maioria das espécies (88,8%), foi discriminada pela ferramenta do código de barras 287
de DNA, apresentando divergência média (K2P) intraespecífica de 0% e interespecífica de 288
32%, estabelecendo um gap cerca de 30 vezes entre a variação intra e interespecífica. 289
Resultados semelhantes foram encontrados por Hubert et al. (2012) que verificaram em 290
espécies de peixes de água doce da fauna canadense uma divergência média de 27 vezes 291
maior entre as espécies do que dentro de cada espécie, com estimativas de distância (K2P) 292
média de 0,3% entre as espécies e 8,3% entre os gêneros. Ward et al. (2005) encontraram 293
0,39% e 9,93%, respectivamente para espécies de peixes australianos. Ward et al. (2009) 294
estudaram espécies de peixes marinhos e de água doce, encontraram distância média 295
intraespecífica para as 294 espécies no valor de 0,35%, enquanto que a média interespecífica 296
foi 8,11%. 297
Como a monofilia recíproca é um critério para a efetividade do código de barras de 298
DNA (Meyer e Paulay, 2005), as espécies que formaram clados reciprocamente monofiléticos 299
puderam ser identificadas utilizando esta ferramenta, no qual se pôde verificar a sua eficiência 300
em quase 90% das espécies utilizadas neste estudo. Peixes de água doce e salgada têm sido 301
facilmente identificados usando o código de barras, com 60-98% das espécies exibindo clados 302

37
monofiléticos (Hubert et al. 2008; Steinke et al. 2009; Mccusker et al. 2012; Young et al. 303
2013; Paz et al. 2014) 304
De acordo com Hebert et al. (2002) o código de barras de DNA atribui corretamente 305
até mesmo os táxons poucos amostrados aos seus respectivos filo ou ordem, e também 306
demonstra as atribuições a nível de espécie. Atualmente, com a aplicação dessa ferramenta em 307
diversos trabalhos, construiu-se uma biblioteca para o armazenamento dos resultados dos 308
códigos de barras para facilitar no conhecimento da biodiversidade (Savolainen et al. 2005; 309
Hebert et al. 2010), esse banco de dados, denominado Sistema de Dados do Código de Barras 310
da Vida (BOLD) auxilia na aquisição, armazenamento, análise e publicação de códigos de 311
barras de DNA (Ratnasingham e Hebert 2007). 312
Do total (59) de espécies identificadas pela ferramenta molecular, apenas 28,8% 313
apresentaram sequências de código de barras de DNA depositadas no banco de dados do 314
BOLD e Genbank (Tabela 3), o que significa que existem poucas sequências do código de 315
barras de DNA de ciclídeos depositadas nesses bancos de dados, podendo está relacionado 316
com a falta de estudos moleculares com as espécies de peixes da Amazônia. 317
Comparando os resultados morfológicos e moleculares, observou-se que apenas nove 318
espécies foram identificadas corretamente por ambas as metodologias. Baldwin et al. (2011) 319
combinaram dados moleculares de COI e morfológicos para analisar espécimes de Starksia no 320
Atlântico ocidental e verificaram que existe incongruência entre as duas ferramentas de 321
identificação, porém as linhagens genéticas são reconhecidos como espécies se forem 322
suportado pela morfologia. Ping et al. (2015) utilizaram as ferramentas morfológicas e 323
moleculares (COI) e observaram duas linhagens separadas de Nemipterus japonicus, isolados 324
geograficamente no sul da China. 325
326
327

38
Conclusão 328
Esse estudo foi o primeiro a examinar um grande número de gêneros (15) 329
representados por diversas espécies de ciclídeos em seis rios da bacia Amazônica. 330
Pôde-se verificar a eficiência da técnica do código de barras de DNA em discriminar 331
aproximadamente 90% das espécies analisadas, revelando uma diversidade oculta relacionada 332
à presença de complexo de espécies e espécies crípticas. A descoberta das diferentes espécies 333
crípticas é possível conhecer as principais espécies de ciclídeos que podem estar sendo 334
comercializados sem ao menos serem descritas, além de auxiliar novas pesquisas 335
morfológicas, verificando as diferenças sutis que as espécies apresentam. 336
A obtenção dos códigos de barras de DNA para as espécies da família Cichlidae 337
proporcionará contribuições importante sobre a ictiofauna Amazônica, para os bancos de 338
dados do BOLD e na Campanha FISH-BOL. 339
340
Agradecimentos 341
Esta pesquisa faz parte do projeto da rede SISBIOTA/Rede BIOPHAM (Sistema 342
Nacional de Pesquisa em Biodiversidade/Rede de pesquisa para ampliação do conhecimento 343
sobre a biodiversidade de vertebrados da Amazônia brasileira com aplicações sobre seu uso e 344
conservação) com apoio financeiro da Fundação de Amparo à Pesquisa do Estado do 345
Amazonas (FAPEAM) e do Conselho Nacional de Desenvolvimento Científico e Tecnológico 346
(CNPq), sob a coordenação de I. P. Farias. A permissão para realizar o trabalho de campo e 347
para a coleta de amostras de tecido foi concedida pelo IBAMA (Licença nº 11325-1). 348
Agradecemos os especialistas Ph. D. Lúcia Helena Rapp Py-Daniel e D. Jansen Alfredo 349
Sampaio Zuanon, do Instituto Nacional de Pesquisas da Amazônia e os demais colaboradores 350
que auxiliaram no processo de coleta e identificações morfológicas. Aos colegas de 351
Laboratório de Genética Animal da Universidade Federal do Amazonas (UFAM) que 352

39
ajudaram nas análises laboratoriais e nas correções do manuscrito. Este artigo constitui uma 353
parte da Dissertação de Mestrado do Programa de Genética, Conservação e Biologia 354
Evolutiva do INPA / UFAM. 355
356
Referências 357
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. 1997. 358
Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. 359
Nucleic Acids Research, 25: 3389-402. 360
Amado MV, Farias IP, Hrbek T. 2011. A Molecular Perspective on Systematics, Taxonomy 361
and Classification Amazonian Discus Fishes of the Genus Symphysodon. International Journal 362
of Evolutionary Biology.16 p. 363
Anseeuw D, Nevado B, Busselen P, Snoeks J, Verheyen E. 2012. Extensive Introgression 364
among Ancestral mtDNA Lineages: Phylogenetic Relationships of the Utaka within the Lake 365
Malawi Cichlid Flock. International Journal of Evolutionary Biology. 366
Aquilino SVL, Tango JM, Fontanilla IC, Pagulayan RC, Basiao ZU, Ong PS, Quilang JP. 367
2011. DNA barcoding of the ichthyofauna of Taal Lake, Philippines. Molecular Ecology 368
Resources, 11: 612–619. 369
Ardura A, Planes S, Garcia-Vazquez E. 2011. Beyond Biodiversity: Fish Metagenomes, 370
Public Library of Science ONE, 6 e22592. 371
Avise JC. 2004. Molecular markers, natural history, and evolution. 2ª ed. 684 p. 372
Baldwin CC, Castillo CI, Weigt LA, Victor BC. 2011. Seven new species within western 373
Atlantic Starksia atlantica, S. lepicoelia, and S. sluiteri (Teleostei, Labrisomidae), with 374
comments on congruence of DNA barcodes and species. ZooKeys 79: 21–72. 375
Becker S, Hanner R, Steinke D. 2011. Five years of FISH-BOL: Brief status report. 376
Mitochondrial DNA, 22: 3–9. 377

40
Benzaquem DC, Oliveira C, Batista JS, Zuanon J, Porto JIR. 2015. DNA Barcoding in 378
Pencilfishes (Lebiasinidae: Nannostomus) Reveals Cryptic Diversity across the Brazilian 379
Amazon. PLoS ONE 10(2): e0112217. 380
Bhattacharya M, Sharma, AR, Patra BC, Sharma G, Seo EM, Nam JS, Chakraborty C, Lee 381
SS. 2015. DNA barcoding to fishes: current status and future directions. Mitochondrial DNA, 382
Early Online: 1–9. 383
BOLD Systems. Available at: (www.barcodinglife.org). Accessed on 12 March 2014. 384
Carvalho DC, Oliveira DAA, Pompeuc OS, Leal CG, Oliveira C, Hanne R. 2011. Deep 385
barcode divergence in Brazilian freshwater fishes: the case of the São Francisco River basin, 386
22: 80-86. 387
Colatreli OP, Meliciano NV, Toffoli D, Farias IP, Hrbek T. 2012. Deep Phylogenetic 388
Divergence and Lack of Taxonomic Concordance in Species of Astronotus (Cichlidae). 389
International Journal of Evolutionary Biology: 1-8. 390
Crampton WGR. 2008. Ecology and life history of an Amazon floodplain cichlid: the discus 391
fish Symphysodon (Perciformes: Cichlidae). Neotropical Ichthyology, 6: 599-612. 392
Chao, NL. 2001. The fishery, diversity, and conservation of ornamental fishes in the Rio 393
Negro basin, Brazil - A review of Project Piaba (1989-99). In: Chao NL, Petry P, Prang G, 394
Sonneschein L, Tlusty MF, eds. Conservation and Management of Ornamental Fish 395
Resources of the Rio Negro Basin, Amazonia, Brazil - Project Piaba. University of Amazonas 396
Press, Manaus, Brazil. 303 p. 397
Drummond AJ, Ashton B, Buxton S, Cheung M, Cooper A, Duran C, Field M, Heled J, 398
Kearse M, Markowitz S, Moir R, Stones-Havas S, Sturrock S, Thierer T, Wilson A. 2011. 399
Geneious v5.4 (www.geneious.com). 400
Duncan WP, Fernandes MN. 2010. Physicochemical characterization of the white, black, and 401
clearwater rivers of the Amazon Basin and its implications on the distribution of freshwater 402

41
stingrays (Chondrichthyes, Potamotrygonidae). Pan-American Journal of Aquatic Sciences, 5: 403
454-464. 404
Farias IP, Hrbek T. 2008. Patterns of diversification in the discus fishes (Symphysodon spp. 405
Cichlidae) of the Amazon basin. Molecular Phylogenetics and Evolution, 49: 32-43. 406
Fernandez-Acosta A, Pereira-Bonilla DJ, Senior-Mojica A, Telles-Murcia C. 2008. Pesca y 407
acuicultura Colombia, Informe Técnico Regional Cuencas del Orinoco y Amazonas. Bogota, 408
Colombia, 78 p. 409
Fish barcode of life. Available at: http://www.fishbol.org/. Accessed on 25 May 2014. 410
Froese R, D Pauly. 2012. FishBase. World Wide Web electronic publication. 411
www.fishbase.org, version (08/2012). 412
Fujisawa T, Barraclough TG. 2013. Delimiting Species Using Single-locus Data and the 413
Generalized Mixed Yule Coalescent (GMYC) Approach: A Revised Method and Evaluation 414
on Simulated Datasets. Systematic Biology Advance. 49 p. 415
Genner MJ, Seehausen O, Lunt DH, Joyce DA, Shaw PW, Carvalho GR, Turner GF. 2007. 416
Age of Cichlids: New Dates for Ancient Lake Fish Radiations. Molecular Biology and 417
Evolution, 24:1269–1282. 418
Hebert, PDN, Cywinska A, Ball SL, deWaard JR. 2002. Biological identifications through 419
DNA barcodes. Proceedings Royal Society of London: 96-99. 420
Hebert PDN, Ratnasingham S, deWaard JR. 2003a. Barcoding animal life: cytochrome c 421
oxidase subunit 1 divergences among closely related species. Proceedings Royal Society of 422
London B, 270: S96–S99. 423
Hebert, PDN, Cywinska A, Ball SL, deWaard JR. 2003b. Biological identifications through 424
DNA barcodes. Proc. R. Soc. Lond. B, 270, 313–321 425
Hebert PDN, Penton EH, Burns JM, Janzen DH, Hallawachs W. 2004. Ten species in one: 426
DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes 427

42
fulgerator. Proceedings of the National Academy of Sciences, 101: 14812–14817. 428
Hebert PDN, Gregory R. 2005. The Promise of DNA Barcoding for Taxonomy. Systematic 429
Biology, 54: 852–859. 430
Hebert PDN, deWaard JR, Landry JF. 2010. DNA barcodes for 1/1000 of the animal 431
kingdom. Biology Letters, 6: 359-362. 432
Hickerson MJ, Carstens BC, Cavender-Bares J, Crandall KA, Graham CH, Johnson JB, 433
Rissler L, Victoriano PF, Yoder AD. 2010. Phylogeography’s past, present, and future: 10 434
years after Avise, 2000. Molecular Phylogenetics and Evolution, 54: 291–301. 435
Hubert N, Hanner R, Holm E, Mandrak NE, Taylor E, Burridge M,Watkinson D, Dumont P, 436
Curry A, Bentzen P, Zhang J, April J, Bernatchez L. 2008. Identifying Canadian Freshwater 437
Fishes through DNA Barcodes, 3: 1-8. 438
Hubert N, Hanner R, Holm E, Mandrak NE, Taylor E, Burridge M, Watkinson D, Dumont P, 439
Curry A, Bentzen P, Zhang J, April J, Bernatchez L. 2012. Identifying Canadian Freshwater 440
Fishes through DNA Barcodes. Public Library of Science ONE, 3: e2490. 441
Hulsey CD. 2009. Cichlid genomics and phenotypic diversity in a comparative context. 442
Integrative and Comparative Biology, 49: 618-29. 443
Johns GC, Avise JC. 1998. A comparative summary of genetic distances in the vertebrates 444
from the mitochondrial cytochrome b gene. Molecular Biology and Evolution, 15:1481–1490. 445
Kadarusman, Hubert N, Hadiaty RK, Sudarto, Paradis E, Pouyaud L. 2012. Cryptic Diversity 446
in Indo-Australian Rainbow fishes Revealed by DNA Barcoding: Implications for 447
Conservation in a Biodiversity Hotspot Candidate, 7: 1-8. 448
Kress WJ, García-Robledo C, Uriarte M, Erickson DL. 2015. DNA barcodes for ecology, 449
evolution, and conservation, Trends in Ecology & Evolution, 30: 25-35. 450
Kullander SO. 2003. Family Cichlidae. In: Reis RE, Kullander SO, Ferraris Jr CJ. Check list 451
of the freshwater fishes of South and Central America. Porto Alegre: EDIPUCRS, 2003, p. 452

43
605-647. 453
León-Romero Y, Mejía O, Soto-Galera E. 2012. DNA barcoding reveals taxonomic conflicts 454
in the Herichthys bartoni species group (Pisces: Cichlidae). Molecular Ecology Resources, 455
12: 1021–1026. 456
Lewinsohn TM, Prado PI. 2005. How Many Species Are There in Brazil? Conservation 457
Biology, 19, 619–624. 458
López-Fernández H, Winemiller KO, Honeycutt RL. 2010. Multilocus phylogeny and rapid 459
radiations in Neotropical cichlid fishes (Perciformes: Cichlidae: Cichlinae). Molecular 460
Phylogenetics and Evolution, 55: 1070–1086. 461
Lovejoy NR, De Araújo ML. 2000. Molecular systematics, biogeography and population 462
structure of neotropical freshwater needlefishes of the genus Potamorrhaphis. Mol Ecol, 9: 463
259-68. 464
Mccusker MR, Denti D, Guelpen LV, Kenchington E, Bentzen P. 2012. Barcoding Atlantic 465
Canada’s commonly encountered marine fishes. Molecular Ecology Resources, 12p. 466
Mcmahan CD, Murray CM, Geheber AD, Boeckman CD, Piller KR. 2011. Paraneetroplus 467
synspilus is a Junior Synonym of Paraneetroplus melanurus (Teleostei: Cichlidae), Zootaxa 468
2833: 1–14. 469
Mejía O, Pérez-Miranda F, León-Romero Y, Soto-Galera, Luna E. 2015. Morphometric 470
variation of the Herichthys bartoni (Bean, 1892) species group (Teleostei: Cichlidae): How 471
many species comprise H. labridens (Pellegrin, 1903)? Neotropical Ichthyology, 131: 61-76. 472
Miyaki CY, Russo CAM, Pereira SL. 2001. Reconstrução filogenética: Introdução e o método 473
da máxima parcimômia. In: Matioli SR. Biologia Molecular e Evolução. Ribeirão Preto: 474
Holos, 202 p. 475
Meyer CP, Paulay G. 2005. DNA Barcoding: Error Rates Based on Comprehensive Sampling. 476
PloS Biology, 3: 2229-2238. 477

44
Moritz C, Cicero C. 2004. DNA Barcoding: Promise and Pitfalls. PLoS Biology, 2: 1529-478
1531. 479
Musilová Z, Rícan O, Janko K, Novák J. 2008. Molecular phylogeny and biogeography of the 480
Neotropical cichlid fish tribe Cichlasomatini (Teleostei: Cichlidae: Cichlasomatinae). 481
Molecular Phylogenetics and Evolution, 46: 659–672. 482
Paz FPC, Batista JS, Porto JIR. 2014. DNA Barcodes of rosy tetras and allied species 483
(Characiformes: Characidae: Hyphessorycon) from the Brazilian Amazon basin, PLOS ONE, 484
9: 1-8. 485
Pazian MF, Pereira LHG, Shimabukuru-Dias CK, Oliveira C, Foresti F. 2012. Cytogenetic 486
and molecular markers reveal the complexity of the genus Piabina Reinhardt, 1867 487
(Characiformes: Characidae), Neotropical Ichthyology, 10: 329-340. 488
Pereira LHG, Hanner R, Foresti F, Piggoti CO, Chao NL, Beheregaray LB. 2013. Can DNA 489
barcoding accurately discriminate megadiverse Neotropical freshwater fish fauna? BMC 490
Genetics, 14: 20. 491
Pereira LHG, Pazian MF, Hanner R, Foresti F, Oliveira C. 2011a. DNA barcoding reveals 492
hidden diversity in the Neotropical freshwater fish Piabina argentea (Characiformes: 493
Characidae) from the Upper Paraná Basin of Brazil. Mitochondrial DNA, 22: 87–96. 494
Pereira LHG, Maia GMG, Haneer R, Foresti F, Oliveira C. 2011b. DNA barcodes 495
discriminate freshwater fishes from the Paraíba do Sul River Basin, São Paulo, Brazil. 496
Mitochondrial DNA, 22: 71–79. 497
Piggoti MP, Chao NL, Beheregaray LB. 2011. Three fishes in one: cryptic species in an 498
Amazonian floodplain forest specialist. Biological Journal of the Linnean Society, 102: 391–499
403. 500
Ping N, Zhongli S, Hebert PDN, Russell B. 2015. The Taxonomic status of japanese threadfin 501
bream Nemipterus japonicus (Bloch, 1791) (Perciformes: Nemipteridae) with a redescription 502
of this species from the South China sea based on morphology and DNA barcodes. J. Ocean 503

45
Univ. China, 14 (1): 178-184. 504
R Development Core Team. 2013. R: A Language and Environment for Statistical 505
Computing. Vienna: R Foundation for Statistical Computing. Available at: http://www.R-506
project.org. Accessed on 20 April 2014. 507
Ratnasingham S, Hebert PDN. 2007. Barcoding Bold: The Barcode of Life Data System 508
(www.barcodinglife.org). Molecular Ecology Notes, 1-10. 509
Reis RE, Kullander SO, Ferraris Jr. CJ. 2003. Check list of the freshwater fishes of South and 510
Central America. Porto Alegre: EDIPUCRS, p. 605-647. 511
Sambrook J, Russell DW. 2001. Molecular Cloning. Cold Spring Harbor Laboratory Press, 512
Cold Spring Harbor, NY, USA. 3rd edition. 513
Savolainen V, Cowan RS, Vogler AP, Roderick GK, Lane R. 2005. Towards writing the 514
encyclopaedia of life: an introduction to DNA barcoding. Philosophical Transactions of the 515
Royal Society B, 360: 1805-1811. 516
Soria-Barreto M, Rodiles-Hernández R, González-Díaz AA. 2011. Revista Mexicana de 517
Biodiversidad 82: 569-579. 518
Steinke D, Zemlak TS, Heber PDN. 2009. Barcoding Nemo: DNA-Based Identifications for 519
the Ornamental Fish Trade. Public Library of Science ONE 4:1-5. 520
Stoeckle MY, Hebert PDN. 2008. Barcode of Life. Scientific American, 2008: 82–88. 521
Toffoli D, Hrbek T, Araújo MLG, Almeida MP, Charvet-Almeida P, Farias IP. 2008. A test 522
of the utility of DNA barcoding in the radiation of the freshwater stingray genus 523
Potamotrygon (Potamotrygonidae, Myliobatiformes). Genetics and Molecular Biology, 31: 524
324-336. 525
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA 5: Molecular 526
Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and 527
Maximum Parsimony Methods. Molecular Biology and Evolution. 528

46
Thompson C. 2010. Amazon Alive! A decade of Discovery 1999-2009. WWF Living 529
Amazon Initiative. 57 p. 530
Ward RD, Hanner , Heber PDN. 2009. The campaign to DNA barcode all fishes, FISH-BOL. 531
Journal of Fish Biology, 74: 329–356. 532
Ward R D, Zemlak TS, Innes BH, Last PR, Heber PDN. 2005. DNA barcoding Austrália fish 533
species. Philosophical Transactions of the Royal Society B, 360: 1845-1857. 534
Winterbottom R, Hanner RH, Burridge M, Zur M. 2014. A cornucopia of cryptic species - a 535
DNA barcode analysis of the gobiid fish genus Trimma (Percomorpha, Gobiiformes), 536
ZooKeys 381: 79–111. 537
Willis SC, Macrander J, Farias IP, Ortí G. 2012a. Simultaneous delimitation of species and 538
quantification of interspecific hybridization in Amazonian peacock cichlids (genus Cichla) 539
using multi-locus data. BMC Evolutionary Biology, 12: 96. 540
Willis SC, López-Fernandez H, Montana CG, Farias IP, Ortí G. 2012b. Species-level 541
phylogeny of ‘Satan’s perches’ based on discordant gene trees (Teleostei: Cichlidae: 542
Satanoperca Günther 1862). Molecular Phylogenetics and Evolution, 63: 798–808. 543
Willis SC, Nuñes MS, Montaña CG, Farias IP, Lovejoy NR. 2007. Systematics, 544
biogeography, and evolution of the Neotropical peacock basses Cichla (Perciformes: 545
Cichlidae). Molecular Phylogenetics and Evolution, 44: 291–307. 546
Young M, Mckelvey KS, Pilgrim KL, Schwartz MK. 2013. DNA barcoding at riverscape 547
scales: ass essing biodiversity among fishes of the genus Cottus (Teleostei) in northern Rocky 548
Mountain streams. Molecular Ecology Resources, 1-13. 549
550
551
552
553

47
CONSIDERAÇÕES FINAIS 554
555
Os fatores ecológicos e históricos que ocorreram na bacia Amazônica proporcionaram 556
a especiação, onde atualmente ainda não é possível saber a verdadeira diversidade de peixes 557
existentes nos rios amazônicos por causa espécies crípticos. 558
As espécies crípticas de ciclídeos foram descobertas nesta pesquisa pelo código de 559
barras de DNA, no qual estavam sendo negligenciadas, porque não era possível separá-las 560
baseadas apenas em caracteres morfológicos. 561
Algumas espécies da família Cichlidae não são monofiléticas e apresentam taxonomia 562
mal resolvida. E o compartilhamento de haplótipos para algumas espécies está relacionado 563
aos processos de irradiação recente que ocasionam a retenção do polimorfismo ancestral ou 564
introgressão após o processo de hibridação. Essas características dificultam na eficácia do 565
código de barras de DNA na identificação e delimitação das espécies de maneira precisa. 566
As sequências obtidas, a partir do código de barras de DNA de várias espécies de 567
ciclídeos, irão contribuir positivamente no banco de dados do BOLD e FISH-BOL, auxiliando 568
na identificação dessas espécies por pesquisadores e não-especialistas em todo o mundo. 569
570
Referências Bibliográficas 571
572
A Era de Aquários - Peixes Ornamentais (www.aquahobby.com). Acesso: 29/10/2012. 573
574
Albert, J. S.; Lovejoy, N. R.; Crampton, W. G. R. 2006. Miocene tectonism and the separation 575
of cis- and trans-Andean river basins: Evidence from Neotropical fishes. Journal of South 576
American Earth Sciences, 21: 14–27. 577
578
Amado, M. V.; Farias, I. P.; Hrbek, T. 2011. A Molecular Perspective on Systematics, 579
Taxonomy and Classification Amazonian Discus Fishes of the Genus Symphysodon. 580

48
International Journal of Evolutionary Biology. 581
582
Barrett, R. D. H.; Hebert, P. D. N. 2005. Identifying spiders through DNA barcodes. Can. J. 583
Zool. 83: 481–491. 584
585
BOLD Systems (www.barcodinglife.org). Acesso em: 28/08/2012. 586
587
Briggs, J. C. 2003. Fishes and Birds: Gondwana Life Rafts Reconsidered. Systematic 588
Biology, 52 (4): 548-553. 589
590
Chakrabarty, P. 2004. Cichlid biogeography: comment and review. Fish and Fisheries, 5: 97–591
119. 592
593
Chakrabarty, P. 2006. Systematics and historical biogeography of Greater Antillean 594
Cichlidae. Molecular Phylogenetics and Evolution, 39: 619–627. 595
596
Clare, E. L.; Lim, B. K.; Fenton, M. B.; Hebert, P. D. N. 2011. Neotropical Bats: Estimating 597
Species Diversity with DNA Barcodes. Plos One, 6(7):1-14. 598
Colatreli, O. P.; Meliciano, N. V.; Toffoli, D.; Farias, I. P.; Hrbek, T. 2012. Deep 599
Phylogenetic Divergence and Lack of Taxonomic Concordance in Species of Astronotus 600
(Cichlidae). International Journal of Evolutionary Biology: 1-8. 601
DeSalle, R.; Egan, M. G.; Siddall, M. 2005. The unholy trinity: taxonomy, species 602
delimitation and DNA barcoding. Phil. Trans. R. Soc. B, 360: 1905-1916. 603
604
Dincã, V.; Zakharov, E. V.; Hebert, P. D. N.; Vila, R. 2011. Complete DNA barcode 605
reference library for a country’s butterfly fauna reveals high performance for temperate 606
Europe. Proceedings of the Royal Society B, 278: 347–355. 607
Dunn, C. P. 2003. Keeping taxonomy based in morphology. Trends in Ecology and Evolution, 608
18(6): 270-271. 609
610
Drummond, A. J.; Ashton, B.; Buxton, S.; Cheung, M.; Cooper, A.; Duran, C.; Field, M.; 611
Heled, J.; Kearse, M.; Markowitz, S.; Moir, R.; Stones-Havas, S.; Sturrock, S.; Thierer, T.; 612
Wilson, A. 2011. Geneious v5.4 (www.geneious.com). 613

49
Ebach, M. C.; Holdrege, C. 2005. DNA barcoding is no substitute for taxonomy. Nature, 434: 614
697. 615
Farias, I. P.; Hrbek, T. 2008. Patterns of diversification in the discus fishes (Symphysodon 616
spp. Cichlidae) of the Amazon basin. Molecular Phylogenetics and Evolution, 49: 32-43. 617
618
Farias, I. P.; Ortí, G.; Meyer, A. 2000. Total Evidence: Molecules, Morphology, and the 619
Phylogenetics of Cichlid Fishes. Journal of Experimental Zoology (Mol Dev Evol), 288:76–620
92. 621
622
Farias, I. P.; Orti, G.; Sampaio, I.; Schneider, H.; Meyer, A. 2001. The cytochrome b gene as 623
a phylogenetic marker: the limits of resolution for analyzing relationships among cichlid 624
fishes. J. Mol. Evol., 53: 89-103. 625
626
Farias, I. P.; Orti, G. O.; Sampaio, I.; Schneider, H.; Meyer, A. 1999. Mitochondrial DNA 627
Phylogeny of the Family Cichlidae: Monophyly and Fast Molecular Evolution of the 628
Neotropical Assemblage. J Mol Evol (1999) 48:703–711. 629
630
Felsenstein, J. 1985. Confidence limits on phylogenies: an approach using the bootstrap. 631
Evolution, 39: 783–791. 632
633
Frederico, R. G.; Farias, I. P.; Araújo, M. L. G.; Charvet-Almeida, P.; Alves-Gomes, J. A. 634
2012. Phylogeography and conservation genetics of the Amazonian freshwater stingray 635
Paratrygon aiereba Müller & Henle, 1841 (Chondrichthyes: Potamotrygonidae). Neotropical 636
Ichthyology, 10(1): 71-80. 637
638
Froese, R. and D. Pauly. 2012. FishBase. World Wide Web electronic publication. 639
www.fishbase.org, version (08/2012). 640
Gómez, A.; Wright, P. J.; Lunt, D. H.; Cancino, J. M.; Carvalho, G. R.; Hughes, R. N.. 2007. 641
Mating trials validate the use of DNA barcoding to reveal cryptic speciation of a marine 642
bryozoan táxon. Proc. R. Soc. B, 274: 199-207. 643
644
Hajibabaei, M.; Janzen, D. H.; Burns, J. M.; Hallwachs, W.; Hebert, P. D. N. 2006. DNA 645

50
barcodes distinguish species of tropical Lepidoptera. Proceedings of the National Academy of 646
Sciences, 103 (4): 968-971. 647
648
Hartl, D. L. 2008. Princípios de genética de população. Tradução de Iulo Feliciano Afonso. 649
FUNPEC Editora, Ribeirão Preto, SP. 217p. 650
651
Hebert, P. D. N.; Cywinska, A.; Ball, S. L and deWaard, J. R. 2003a. Biological 652
identifications through DNA barcodes. Proceedings Royal Society of London: 96-99. 653
654
Hebert, P. D. N.; Gregory, R. 2005. The Promise of DNA Barcoding for Taxonomy. 655
Systematic Biology, 54(5): 852–859. 656
657
Hebert, P. D. N.; Penton, E. H.; Burns, J. M.; Janzen, D. H.; Hallawachs, W. 2004a. Ten 658
species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly 659
Astraptes fulgerator. Proceedings of the National Academy of Sciences, 101(41): 14812–660
14817. 661
662
Hebert, P. D. N.; Ratnasingham, S.; deWaard, J.R. 2003b. Barcoding animal life: cytochrome 663
c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. B (Suppl.) 664
270, S96–S99. 665
666
Hebert, P. D. N.; Stoeckle, M. Y.; Zemlak, T. S.; Francis, C. M. 2004b. Identification of Birds 667
through DNA Barcodes. PloSBiology, October, 2: 1658-1663. 668
669
Hebert, P. D. N.; deWaard, J. R..; Landry, J. F. 2010. DNA barcodes for 1/1000 of the animal 670
kingdom. Biol. Lett. 2010 6, 359-362. 671
672
Hberk, T. Farias, I. 2008. The complete mitochondrial genome of the pirarucu (Arapaima 673
gigas, Arapaimidae, Osteoglossiformes). Genetics and Molecular Biology, 31, 1 (suppl), 293-674
302. 675
676
Janzen, D. H.; Hallwachs, W.; Blandin, P.; Burns, J. M.; Cadiou, J.; Chacon, I.; Dapkey, T.; 677

51
Deans, A. R.; Epstein, M. E.; Espinoza, B.; Franclemont, J. G.; Haber, W. A.; Hajibabaei, M.; 678
Hall, J. P. W.; Hebert, P. D. N.; Gauld, I. D.; Harvey, D. J.; Hausmann, A.; Kitching, I. J.; 679
Lafontaine, D.; Landry, J. F.; Lemaire, C.; Miller, J. Y.; Miller, J. S.; Miller, L.; Miller, S. E.; 680
Montero, J.; Munroe, E.; Green, S. R.; Ratnasingham, S.; Rawlins, J. E.; Robbins, R. K.; 681
Rodriguez, J. R.; Rougerie, R.; Sharkey, M. J.; M . Smith, M. A.; Solis, M. A.; Sullivan, J. B.; 682
Thiaucourt, P., Wahl, D. B.; Weller, S. J.; Whitfield, J. B.; Willmott, K. R.; Wood, D. M.; 683
Woodley, N. E.; Wilson, J. J. 2009. Integration of DNA barcoding into an ongoing inventory 684
of complex tropical biodiversity. Molecular Ecology Resources, 9: 1–26. 685
Johns G. C; Avise, J. C. 1998. A comparative summary of genetic distances in the vertebrates 686
from the mitochondrial cytochrome b gene. Mol Biol Evol, 15:1481–1490. 687
688
Kress, W. J.; Erickson, D. L. 2008. DNA barcodes: Genes, genomics, and bioinformatics. 689
PNAS, 105(8): 2761–2762. 690
691
Kullander, S. O. 1998 apud Kullander, S. O. Family Cichlidae. In: Reis, R. E.; Kullander, S. 692
O.; Ferraris Jr., C.J. Check list of the freshwater fishes of South and Central America. Porto 693
Alegre: EDIPUCRS, 2003, p. 605-647. 694
Kullander, S. O. 2009. Crenicichla mandelburgeri, a new species of cichlid fish (Teleostei: 695
Cichlidae) from the Paraná river drainage in Paraguay. Zootaxa 2006: 41–50. 696
697
Kullander, S. O.; Ferreira, E. J. G. 2005. Two new species of Apistogramma Regan 698
(Teleostei: Cichlidae) from the rio Trombetas, Pará State, Brazil. Neotropical Ichthyology, 699
3(3):361-371. 700
701
Kullander, S. O. 2012. Krobia xinguensis, a new species of cichlid fish from the Xingu River 702
drainage in Brazil (Teleostei: Cichlidae). Zootaxa 3197: 43–54 (2012). 703
704
Kullander, S. O. Family Cichlidae. In: Reis, R. E.; Kullander, S. O.; Ferraris Jr., C.J. Check 705
list of the freshwater fishes of South and Central America. Porto Alegre: EDIPUCRS, 2003, p. 706
605-647. 707
León-Romero, Y.; Mejía, O.; Soto-Galera, E. 2012. DNA barcoding reveals taxonomic 708

52
conflicts in the Herichthys bartoni species group (Pisces: Cichlidae). Molecular Ecology 709
Resources, 12: 1021–1026. 710
711
Lipscomb, D.; Platnick, N.; Wheeler, Q. 2003. The intellectual content of taxonomy: a 712
comment on DNA taxonomy. TRENDS in Ecology and Evolution,18(2): 65-66. 713
714
López-Fernández, H.; Winemiller, K. O.; Honeycutt, R. L. 2010. Multilocus phylogeny and 715
rapid radiations in Neotropical cichlid fishes (Perciformes: Cichlidae: Cichlinae). 716
Molecular Phylogenetics and Evolution, 55: 1070–1086. 717
718
Lovejoy, N. R.; Albert, J. S.; Crampton, W. G. R. 2006. Miocene marine incursions and 719
marine/freshwater transitions: evidence from Neotropical fishes. Journal of South American 720
Earth Sciences, 21: 5-13. 721
722
Lucinda, P. H. F.; Lucena, C. A. S.; Assis, N. C. 2010. Two new species of cichlid fish genus 723
Geophagus Heckel from the Rio Tocantins drainage (Perciformes: Cichlidae). Zootaxa, 2429: 724
29–42 . 725
726
Lundberg, J. G.; Marshall, L. G.; Guerrero, J.; Horton, B.; Malabarba, M. C. S. L.; Wesseling, 727
F. 1998. The stage for neotropical fish diversification: A history of tropical South American 728
Rivers. In: Malabarba, L.R.; Reis, R.E.; Vari, R.P.; Lucena, Z.M.; Lucena, C.A.S. Phylogeny 729
and classification of Neotropical fishes. EDIPUCRS, Porto Alegre, Rio Grande do Sul, Brasil, 730
603p. 731
732
Mallet, J.; K. Willmott. 2003. Taxonomy: Renaissance or Tower of Babel? Trends Ecol. Evol. 733
18:57–59. 734
735
Meyer, C. P.; Paulay, G. 2005. DNA Barcoding: Error Rates Based on Comprehensive 736
Sampling. PloS Biology, 3(12): 2229-2238. 737
738
Mora, C.; Tittensor, D. P.; Adl, S.; Simpson, A. G. B.; Worm, B. 2011. How Many Species 739
Are There on Earth and in the Ocean? PLoS Biology, 9(8): 1-8. 740
741
Moritz, C.; Cicero, C. 2004. DNA Barcoding: Promise and Pitfalls. PLoS Biology, 2 (10): 742

53
1529-1531. 743
744
Murray, A. M. 2001. The fossil record and biogeography of the Cichlidae (Actinopterygii: 745
Labroidei). Biological Journal of the Linnean Society, 74: 517–532. 746
747
Nwani, C. D.; Becker, S.; Braid, H. E.; Ude, E. F. Okogwu, O. I.; Hanner, R. 2011. DNA 748
barcoding discriminates freshwater fishes from southeastern Nigeria and provides river 749
system-level phylogeographic resolution within some species. Mitochondrial DNA, 22(S1): 750
43– 51. 751
752
Oceguera-Figueroa, A.; León-Régagnon, V.; Siddall, M. E. 2010. DNA barcoding reveals 753
Mexican diversity within the freshwater leech genus Helobdella (Annelida: Glossiphoniidae). 754
Mitochondrial DNA, 21(S1): 24–29. 755
756
Ortiz, M. F. 2010. Validação do DNA Barcode como identificador de espécies: um estudo de 757
ampla amostragem com o gênero Pseudoplatystoma (Siluriformes, Pimelodidae) na 758
Amazônia. Dissertação de Mestrado, Instituto Nacional de Pesquisas da Amazônia/Manaus : 759
[s.n.], 2010. 73f. 760
Ottoni, F. P.; Bragança, P. H. N.; Amorim, P. A.; Gama, C. S. 2012. A new species of 761
Laetacara from the northern Brazil coastal floodplains (Teleostei: Cichlidae). Vertebrate 762
Zoology, 62 (2): 181 – 188. 763
764
Ottoni, F. P. 2012. Three new species of Australoheros from southeastern Brazil, with 765
taxonomic notes on Chromys oblonga, Heros autochton and H. jenynsii (Teleostei: Labroidei: 766
Cichlidae). Vertebrate Zoology, 62 (1): 83 – 96. 767
768
Ottoni, F. P.; Lezama, A. Q.; Triques, M. L.; Fragoso-Moura, E. N.; Lucas, C. C. T.; Barbosa, 769
F. A. R. 2011. Australoheros perdi, new species (Teleostei: Labroidei: Cichlidae) from the 770
lacustrine region of the Doce River Valley, southeastern Brazil, with biological information. 771
Vertebrate Zoology, 61 (1): 137 – 145. 772
773
Ottoni, F. P.; Costa, W. J. E. M. 2009. Description of a new species of Laetacara 774
KULLANDER, 1986 from central Brazil and re-description of Laetacara dorsigera 775

54
(HECKEL, 1840) (Labroidei: Cichlidae: Cichlasomatinae), Vertebrate Zoology, 59 (1): 41-776
48. 777
778
Park, D. S.; Foottit, R.; Maw, E.; Hebert, P. D. N. 2011. Barcoding Bugs: DNA-Based 779
Identification of the True Bugs (Insecta: Hemiptera: Heteroptera). PloS ONE, v. 6, i. 4: 1-9. 780
781
Pereira, L. H. G.; Pazian, M. F.; Hanner, R.; Foresti, F.; Oliveira, C. 2011a. DNA barcoding 782
reveals hidden diversity in the Neotropical freshwater fish Piabina argentea (Characiformes: 783
Characidae) from the Upper Paraná Basin of Brazil. Mitochondrial DNA, 22(S1): 87–96. 784
785
Pereira, L. H. G.; Maia, G. M. G.; Hanner, R.; Foresti, F.; Oliveira, C. 2011b. DNA barcodes 786
discriminate freshwater fishes from the Paraíba do Sul River Basin, São Paulo, Brazil. 787
Mitochondrial DNA, 22(S1): 71–79. 788
789
Piggoti, M. P.; Chao, N. L.; Beheregaray, L. B. 2011. Three fishes in one: cryptic species in 790
an Amazonian floodplain forest specialist. Biological Journal of the Linnean Society, 102: 791
391–403. 792
793
Rach, J; DeSalle, R., Sarkar, I. N., Schierwater, B e Hadrys, H. 2008. Character-based DNA 794
barcoding allows discrimination of genera, species and populations in Odonata. Proc. R. Soc. 795
B, 275, 237–24. 796
797
Ratnasingham, S.; Hebert, P. D. N. 2007. Barcoding Bold: The Barcode of Life Data System 798
(www.barcodinglife.org). Molecular Ecology Notes, 1-10. 799
800
Römer, U.; Hahn, I.J.; Vergara, P. M. 2010. Description of Dicrossus foirni sp. n. and 801
Dicrossus warzeli sp. n. (Teleostei: Perciformes: Cichlidae), two new cichlidae species from 802
the Rio Negro and the Rio Tapajós, Amazon drainage, Brazil. Vertebrate Zoology, 60 (2): 803
123-138. 804
805
Ruffino, M. L. (Coord.) 2004. A pesca e os recursos pesqueiros na Amazônia brasileira. 806
Ibama/ProVárzea, Manaus, AM. 272p. 807
808
Savolainen, V.; Cowan, R. S.; Vogler, A. P.; Roderick, G. K.; Lane, R. 2005. Towards 809

55
writing the encyclopaedia of life: an introduction to DNA barcoding. Phil. Trans. R. Soc. B, 810
360: 1805-1811. 811
812
Scitable by nature education (www.nature.com/scitable). Acesso em: 27/11/2012. 813
814
Silva-Brandão, K. L.; Lyra, M. L.; Freitas, A. V. L. 2009. Barcoding Lepidoptera: Current 815
Situation and Perspectives on the Usefulness of a Contentious Technique. Neotropical 816
Entomology, 38(4): 441-451. 817
Sparks, J. S.; Smith, Wm. L. 2004. Phylogeny and biogeography of cichlid fishes (Teleostei: 818
Perciformes: Cichlidae). Cladistics, 20: 501–517. 819
820
Sparks, J. S.; Smith, Wm. L. 2005. Freshwater Fishes, Dispersal Ability, and Nonevidence: 821
“Gondwana Life Rafts” to the Rescue. Syst. Biol. 54(1): 158–165. 822
823
Stearns, S. C.; Hoekstra, R. F. 2003. Evolução: uma introdução. Traduzido por Blum, M. São 824
Paulo: Atheneu Editora: 214-231. 825
826
Stiassny, M. L. J. 1991. Phylogenetic intrarelationships of the family Cichlidae: Na overview. 827
In: Keenleyside, M.H.A. Cichlid fishes: Behaviour ecology and evolution. London, Chapman 828
and Hall, p.378. 829
Stoeckle, M. Y; Hebert, P. D. N. 2008. Barcode of Life. Scientific American, 2008: 82–88. 830
Stoeckle, M. 2003. Taxonomy, DNA and the Bar Code of Life. BioScience, 53(9):796-797. 831
Tamura K.; Peterson D.; Peterson N.; Stecher G.; Nei M.; Kumar, S. 2011. MEGA 5: 832
Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary 833
Distance, and Maximum Parsimony Methods. Molecular Biology and Evolution. 834
The Cichlidroom companion (www.cichlidae.com). Acesso: 29/10/2012. 835
836
Varella, H. R.; Kullander, S. O.; Lima, F. C. T. 2012. Crenicichla chicha, a new species of 837
pike cichlid (Teleostei: Cichlidae) from the rio Papagaio, upper rio Tapajós basin, Mato 838
Grosso, Brazil. Neotropical Ichthyology, 10(2): 233-244. 839
840

56
Ward, R. D.; Hanner, R.; Hebert, P. D. N. 2009. The campaign to DNA barcode all fishes, 841
FISH-BOL. Journal of Fish Biology, 74: 329–356. 842
843
Ward, R. D.; Zemlak, T. S.; Innes, B. H.; Last, P. R.; Hebert, P. D. N. 2005. DNA barcoding 844
Austrália fish species. Phil Trans. R. Soc. B, 360: 1845-1857. 845
846
Webb, J. M.; Jacobus, L. M.; Funk, D. H.; Zhou, X.; Kondratieff, B.; Geraci, C. J.; DeWalt, 847
R. E.; Baird, D. J.; Richard, B.; Phillips, I.; Hebert, P. D. N. 2012. A DNA Barcode Library 848
for North American Ephemeroptera: Progress and Prospects. PLoS ONE, 7 (5): 1-6. 849
850
Website for the Consortium for the Barcode of Life (CBOL) (www.barcoding.si.edu). 851
Acesso: 20/09/2012. 852
853
Wiemers, M.; Fiedler, K. 2007. Does the DNA barcoding gap exist? – a case study in blue 854
butterflies (Lepidoptera: Lycaenidae). Frontiers in Zoology, 4(8): 1-16. 855
856
Will, K. W.; D. Rubinoff. 2004. Myth of the molecule: DNA barcodes for species cannot 857
replace morphology for identification and classification. Cladistics, 20:47–55. 858
859
Will, K. W.; Mishler, B. D.; Wheeler, Q. D. 2005. The Perils of DNA Barcoding and the 860
Need for Integrative Taxonomy. Syst. Biol. 54(5):844–851. 861
862
Willis S. C., Macrander J., Farias I. P. & Ortí G. 2012. Simultaneous delimitation of species 863
and quantification of interspecific hybridization in Amazonian peacock cichlids (genus 864
Cichla) using multi-locus data. BMC Evolutionary Biology, 12: 96. 865
866
Willis S. C., Nuñes M. S., Montaña C. G., Farias I. P. & Lovejoy N. R. 2007. Systematics, 867
biogeography, and evolution of the Neotropical peacock basses Cichla (Perciformes: 868
Cichlidae). Molecular Phylogenetics and Evolution, 44: 291–307. 869
870
Junk, W. J.; Piedade, M. T. F.; Schöngart, J.; Wittmann, F. A classification of major natural 871
habitatis of Amazonian white-water river floodplains (várzeas). Wetlands Ecology and 872
Management (2012) 20:461–475. 873
874

57
Junk, W. J.; Piedade, M. T. F.; Schöngart, J.; Wittmann, F, Cohn-Haft, M.; Adeney, J. M.; 875
Wittmann, F. A Classification of Major Naturally-Occurring Amazonian Lowland Wetlands. 876
Wetlands (2011) 31:623–640. 877
878
879
880

59
ANEXO A 881
882
Tabela 1. Nomes das 148 espécies, baseadas nas identificações morfológicas, que foram analisadas no presente estudo. 883
Espécie Localidade ID Número
CTGA
ID Número
do Voucher
Aequidens michaeli Arara, banco direito do Iriri, Rio Iriri, Xingu CTGA_1754 30954
Aequidens michaeli Cachoeira do Jericoá, Xingu CTGA_11191
Aequidens pallidus Pedral, borda da ressaca, Jatapu CTGA_100496 37196
Aequidens pallidus Igarapé do Santo Amaro, margem direita do rio Jatapu CTGA_100513 37166
Aequidens pallidus Igarapé do Santo Amaro, margem direita do rio Jatapu CTGA_100514 37166
Aequidens pallidus Igarapé 2, na margem esquerda do rio Jatapu CTGA_100240 37105
Aequidens pallidus Igarapé extra, próx. igarapé Central, Trilha central (Aiuanã), Rio Negro CTGA_105155 42363
Aequidens pallidus Igarapé extra, próx. igarapé Central, Trilha central (Aiuanã), Rio Negro CTGA_105156 42363
Aequidens cf. eape Igarapé Barreirinha, margem esquerda do rio Tapajós CTGA_104548 41457
Apistogramma sp. Rio Iriri, com 4 horas baixo boca do Rio Novo, Xingu CTGA_2031 31132
Apistogramma aff. Staecki Igarape 2, km 4 da trilha da esquerda, Lago da Velha, margem direita do rio
Jatapu
Apistogramma agassizi Igarapé dos Mutuns, Ipixuna, rio Purus CTGA_103206 42054
Apistogramma agassizi Igarapé dos Mutuns, Ipixuna, rio Purus CTGA_103207 42054
Apistogramma agassizi Igarapé dos Mutuns, Ipixuna, rio Purus CTGA_103208 42054

60
Apistogramma agassizi Igarapé dos Mutuns, Ipixuna, rio Purus CTGA_103209 42054
Apistogramma agassizi Igarapé dos Mutuns, Ipixuna, rio Purus CTGA_103210 42054
Apistogramma agassizi Igarapé Barreirinha, margem esquerda do rio Tapajós CTGA_104507 41464
Apistogramma agassizi Igarape 4, 300m da trilha da esquerda, Lago da Velha, margem direita do rio
Jatapu, bacia do baixo rio Uatumã
CTGA_100264 37404
Apistogramma agassizi Igarape 4, 300m da trilha da esquerda, Lago da Velha, margem direita do rio
Jatapu, bacia do baixo rio Uatumã
CTGA_100265 37404
Apistogramma agassizi Igarape do Monte Negro (Ig. 3 margem direita), afluente do rio Jatapu. CTGA_100539 37293
Apistogramma agassizi Igarape 2 acima do Tabocal, margem esquerda do rio Jatapu CTGA_100608 39064
Apistogramma agassizi Margem direita a montante da Vila de Sao Jose do Jabote CTGA_100581 39049
Apistogramma agassizi Igarape 4, 300m da trilha da esquerda, Lago da Velha, margem direita do rio
Jatapu, bacia do baixo rio Uatumã
CTGA_100263 37404
Apistogramma agassizi Igarape 4, 300m da trilha da esquerda, Lago da Velha, margem direita do rio
Jatapu, bacia do baixo rio Uatumã
CTGA_100266 37404
Apistogramma agassizi Igarape 4, 300m da trilha da esquerda, Lago da Velha, margem direita do rio
Jatapu, bacia do baixo rio Uatumã
CTGA_100256 37404
Apisotgramma pertensis Igarapé 2, na margem esquerda do rio Jatapu CTGA_100550 37116
Apisotgramma pertensis Igarape do Monte Negro (Ig. 3 margem direita), afluente do rio Jatapu CTGA_100538 37301
Apisotgramma pertensis Igarapé 2, na margem esquerda do rio Jatapu CTGA_100551 37116
Apisotgramma pertensis Igarapé 2, na margem esquerda do rio Jatapu CTGA_100552 37116
Apisotgramma pertensis Igarapé do Monte Negro (Ig. 3 margem direita), afluente do rio Jatapu CTGA_100527 37293
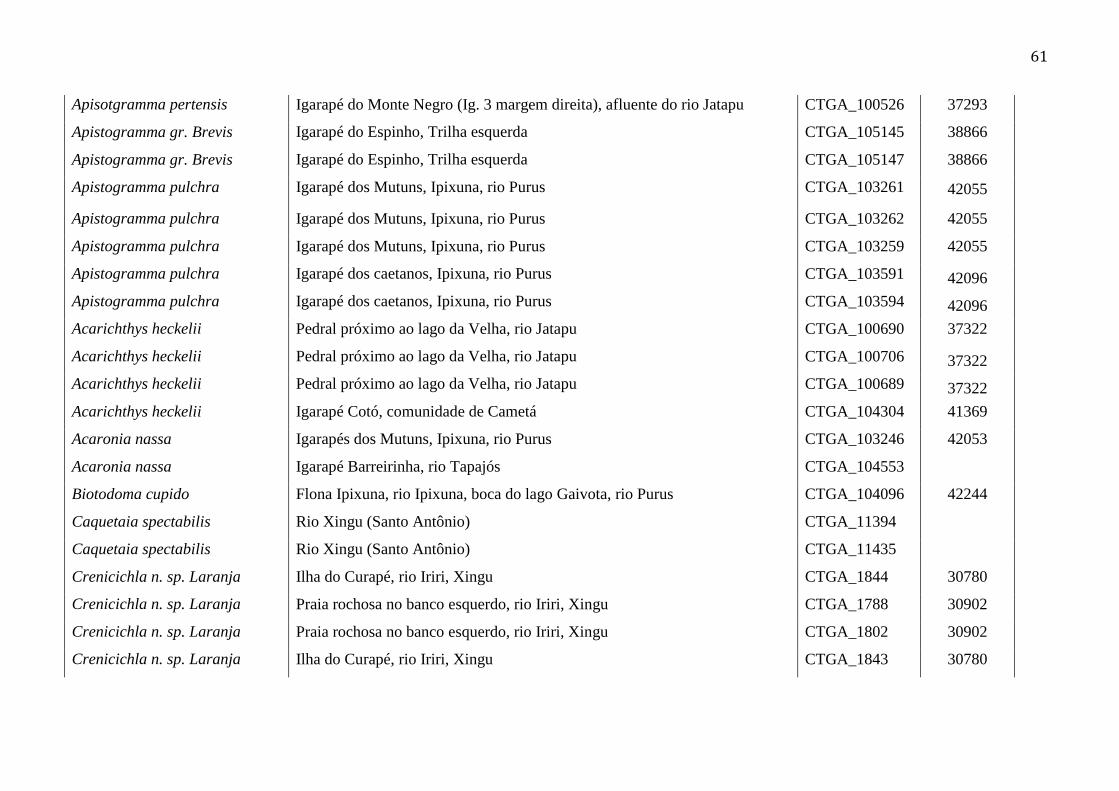
61
Apisotgramma pertensis Igarapé do Monte Negro (Ig. 3 margem direita), afluente do rio Jatapu CTGA_100526 37293
Apistogramma gr. Brevis Igarapé do Espinho, Trilha esquerda CTGA_105145 38866
Apistogramma gr. Brevis Igarapé do Espinho, Trilha esquerda CTGA_105147 38866
Apistogramma pulchra Igarapé dos Mutuns, Ipixuna, rio Purus CTGA_103261 42055
Apistogramma pulchra Igarapé dos Mutuns, Ipixuna, rio Purus CTGA_103262 42055
Apistogramma pulchra Igarapé dos Mutuns, Ipixuna, rio Purus CTGA_103259 42055
Apistogramma pulchra Igarapé dos caetanos, Ipixuna, rio Purus CTGA_103591 42096
Apistogramma pulchra Igarapé dos caetanos, Ipixuna, rio Purus CTGA_103594 42096
Acarichthys heckelii Pedral próximo ao lago da Velha, rio Jatapu CTGA_100690 37322
Acarichthys heckelii Pedral próximo ao lago da Velha, rio Jatapu CTGA_100706 37322
Acarichthys heckelii Pedral próximo ao lago da Velha, rio Jatapu CTGA_100689 37322
Acarichthys heckelii Igarapé Cotó, comunidade de Cametá CTGA_104304 41369
Acaronia nassa Igarapés dos Mutuns, Ipixuna, rio Purus CTGA_103246 42053
Acaronia nassa Igarapé Barreirinha, rio Tapajós CTGA_104553
Biotodoma cupido Flona Ipixuna, rio Ipixuna, boca do lago Gaivota, rio Purus CTGA_104096 42244
Caquetaia spectabilis Rio Xingu (Santo Antônio) CTGA_11394
Caquetaia spectabilis Rio Xingu (Santo Antônio) CTGA_11435
Crenicichla n. sp. Laranja Ilha do Curapé, rio Iriri, Xingu CTGA_1844 30780
Crenicichla n. sp. Laranja Praia rochosa no banco esquerdo, rio Iriri, Xingu CTGA_1788 30902
Crenicichla n. sp. Laranja Praia rochosa no banco esquerdo, rio Iriri, Xingu CTGA_1802 30902
Crenicichla n. sp. Laranja Ilha do Curapé, rio Iriri, Xingu CTGA_1843 30780

62
Crenicichla percna Barinha, cachoeiras no meio do canal, rio Iriri, Xingu CTGA_1889 30929
Crenicichla aff. Inpa Boca do rio Novo, Xingu CTGA_1958 31034
Crenicichla sp.1 Vila de Sta. Izabel, rio Araguaia, Pará CTGA_2852
Crencichla jegui Vila de Sta. Izabel, rio Araguaia, Pará CTGA_2853
Crenicichla lugubris Vila de Sta. Izabel, rio Araguaia, Pará CTGA_2862
Crenicichla lugubris Vila de Sta. Izabel, rio Araguaia, Pará CTGA_2863
Crenicichla astroblepa Pedral São Miguel, rio Araguaia CTGA_2970
Crenicichla astroblepa Pedral São Miguel, rio Araguaia CTGA_3069
Crenicichla astroblepa Pedral Rebojo, rio Araguaia CTGA_3417
Crenicichla astroblepa Araguatins, Tocantins CTGA_3093
Crenicichla aff strigata Pedral Rebojo, rio Araguaia CTGA_3409
Crenicichla n. sp. Preta Rio Xingu (área 03) CTGA_11096
Crenicichla n. sp. Preta Rio Xingu (área 03) CTGA_11097
Crenicichla n. sp. Preta Cachoeira do Jericoá, rio Xingu CTGA_11189
Crenicichla n. sp. Preta Cachoeira do Jericoá, rio Xingu CTGA_11197
Crenicichla n. sp. Preta Cachoeira do Espelho, rio Xingu CTGA_2100 31003
Crenicichla n. sp. Xingu Rio Xingu (área 04) CTGA_11136
Crenicichla sp. Xingu Rio Xingu (área 04) CTGA_11137
Crenicichla sp. Xingu Rio Xingu (área 04) CTGA_11138
Crenicichla sp. Xingu Rio Xingu (área 04) CTGA_11139
Crenicichla sp. Xingu Rio Xingu (Praia do caju) CTGA_11233

63
Crenicichla sp. Xingu Rio Xingu (Corredeira a jusante da Cachoeira de Belo Monte) CTGA_11425
Crenicichla regani Igarapé 4, 300m da trilha da esquerda, Lago da Velha, margem direita do rio
Jatapu
CTGA_100258 37401
Crenicichla regani Rio Jatapu. Braço do Igarapé 2 CTGA_100350
Crenicichla regani Flona Ipixuna, rio Ipixuna, comunidade Mutum, tonco CTGA_104101 41923
Crenicichla regani Flona Ipixuna, rio Ipixuna, comunidade Mutum, tonco CTGA_104100 41923
Crenicichla regani Igarapé no acesso para as trilhas da comunidade de Cametá CTGA_104256
Crenicichla regani Igarapé Jutuarana CTGA_104806 41321
Crenicichla regani Rio Xingu (Santo Antônio) CTGA_11391
Cichla melaniae Rio Xingu (Praia do caju) CTGA_11230
Cichla melaniae Rio Xingu (Corredeira a jusante da Cachoeira de Belo Monte) CTGA_11430
Cichla melaniae Rio Xingu (Praia do caju) CTGA_11231
Cichla melaniae Rio Xingu (Praia do caju) CTGA_11232
Cichla melaniae Rio Xingu (Praia do caju) CTGA_11228
Cichla melaniae Rio Iriri, Xingu CTGA_11058
Cichla melaniae Rio Xingu (Praia do caju) CTGA_11229
Cichla mirianae pitanga Rio Xingu (jusante de Belo Monte) CTGA_11433
Cichla mirianae pitanga Rio Xingu (jusante de Belo Monte) CTGA_11434
Cichla temensis Igarapé 3, na margem esquerda do rio Jatapu CTGA_100336 37155
Cichla temensis Igarapé 3, na margem esquerda do rio Jatapu CTGA_100334 37155
Cichla temensis Igarapé 3, na margem esquerda do rio Jatapu CTGA_100335 37155

64
Cichla piquiti Pedral Remanso dos Botos CTGA_3060
Geophagus altifrons Pedral Cachoeira do Zé dos gatos CTGA_2744
Geophagus altifrons Flona Ipixuna, rio Ipixuna, boca do lago Gaivota, Purus CTGA_104036 42247
Geophagus argyrostictus Rio Xingu (Praia do caju) CTGA_11350
Geophagus argyrostictus Rio Xingu (Praia do caju) CTGA_11240
Geophagus argyrostictus Rio Xingu (Praia do caju) CTGA_11351
Geophagus argyrostictus Rio Bacajá, Xingu CTGA_11312
Geophagus argyrostictus Rio Iriri, Xingu CTGA_11059
Geophagus argyrostictus Rio Iriri, Xingu CTGA_11060
Geophagus argyrostictus Rio Iriri, Xingu CTGA_11061
Geophagus sp. Boca do Irirí, Rio Iriri na confluência com Rio Xingu CTGA_1651 30852
Geophagus sp. Praia em frente a comunidade de Cametá, Tapajós CTGA_104694 41518
Geophagus sp. Praia em frente a comunidade de Cametá, Tapajós CTGA_104696 41518
Geophagus sp. Praia em frente à com. Itapuama, Tapajós CTGA_104996 41526
Geophagus proximus Em frente à comunidade Cametá, Tapajós CTGA_104573 41272
Geophagus proximus Em frente à comunidade Cametá, Tapajós CTGA_104574 41272
Geophagus proximus Em frente à comunidade Cametá, Tapajós CTGA_104576 41272
Heros efasciatus Rio Xingu (Santo Antônio) CTGA_11397
Heros efasciatus Rio Xingu (Santo Antônio) CTGA_11398
Heros efasciatus Rio Xingu (Santo Antônio) CTGA_11399
Heros efasciatus Rio Xingu (Santo Antônio) CTGA_11400

65
Heros efasciatus Pedral próximo ao lago da Velha CTGA_100685 37323
Heros efasciatus Pedral próximo ao lago da Velha CTGA_100692 37323
Heros efasciatus Pedral próximo ao lago da Velha CTGA_100704 37323
Heros efasciatus Pedral próximo ao lago da Velha CTGA_100705 37323
Heros efasciatus Pedral próximo ao lago da Velha CTGA_100716 37323
Laetacara araguaiae Praia em frente à comunidade Cametá, margem esquerda do rio Tapajós CTGA_104891 41146
Mesonauta festivus Igarapé Barreirinha, Tapajós CTGA_104529
Mesonauta festivus Igarapé Anduru, margem esquerda do rio Tapajós CTGA_104595 41254
Mesonauta festivus Pedral próximo ao lago da Velha CTGA_100732 37324
Mesonauta festivus Ressaca do Jatapu CTGA_100680 37241
Mesonauta festivus Ressaca do Jatapu CTGA_100681 37241
Retroculus lapidifer Araguatins,Tocantins CTGA_3178
Retroculus lapidifer Araguatins,Tocantins CTGA_3179
Retroculus lapidifer Araguatins,Tocantins CTGA_3180
Retroculus xinguensis Barinha, cachoeiras no meio do canal, Rio Iriri, Xingu CTGA_1903 30948
Retroculus xinguensis Rio Xingu (Praia do caju) CTGA_11235
Satanoperca jurupari Araguatins,Tocantins CTGA_3174
Satnoperca sp. Boca do Irirí, Rio Iriri na confluência com Rio Xingu CTGA_1700 30887
Satnoperca sp. Boca do Irirí, Rio Iriri na confluência com Rio Xingu CTGA_1703 30887
Satanoperca lilith Ressaca do Jatapu CTGA_100670 37267
Satanoperca lilith Ressaca do Jatapu CTGA_100418 37211

66
884
885 886 887
888
889
890
891
892
Satanoperca lilith Pedral, borda da ressaca Jatapu CTGA_100485 37437
Symphysodon aequafasciatus Rio Xingu (Santo Antônio) CTGA_11437
Symphysodon aequafasciatus Rio Xingu (Santo Antônio) CTGA_11439
Symphysodon aequafasciatus Rio Xingu (Santo Antônio) CTGA_11440
Teleocichla sp. Pedral Remanso dos Botos CTGA_3000
Teleocichla n. sp. Preta Cachoeira do Espelho, Rio Xingu CTGA_2064 31019
Teleocichla n. sp. Preta Cachoeira do Espelho, Rio Xingu CTGA_2102 31019
Teleocichla n. sp. Preta Cachoeira do Espelho, Rio Xingu CTGA_2103 31019
Teleocichla n. sp. Preta Cachoeira do Espelho, Rio Xingu CTGA_2104 31019
Teleocichla cf. gephyrogramma Rio Xingu (Cachoeira do Jericoá) CTGA_11190
Teleocichla cinderela Pedral São Miguel, Araguaia CTGA_2975
Teleocichla cinderela Pedral São Miguel, Araguaia CTGA_2976
Teleocichla cinderela Pedral Rebojo, Araguaia CTGA_3094

67
Tabela 2. Nome e referências das sequencias dos primers usados na amplificação e sequenciamento no presente estudo. 893
894
Gene Primer Primer sequence 5’ - 3’ Referências
CO1
VF2 5’gtaaaacgacggccagtCAACCAACCACAAAGACATTGGCAC3’
Ivanova et al., 2007 FishF2 5’gtaaaacgacggccagtCGACTAATCATAAAGATATCGGCAC3’
FishR2 5’caggaaacagctatgacTTCAGGGTGACCGAAGAATCAGAA3’
VR1d 5’caggaaacagctatgacCTCAGGGTGTCCGAARAAYCARAA3’
CO1
M13F (-21) TGTAAAACGACGGCCAGT Messing, 1983
M13R (-27) CAGGAAACAGCTATGAC
895 896 897 898 899 900 901 902 903 904 905 906 907 908 909 910 911 912

68
913 914 915 916 917 918 919 920 921 922 923 924 925 926 927 928 929 930 931 932 933 934 935 936 937 938 939 940 941 942 943 944 945 946 947 948 949 950 951 952 953 954 955 956 957 958 959

69
960 961 962 963 964 965 966 967 968 969 970 971 972 973 974 975 976 977 978 979 980 981 982 983 984 985 986 987 988 989 990 991 992 993 994 995 996 997 998 999 1000 1001 1002 1003 1004 1005 1006 1007 1008 1009

70
1010 1011 1012 1013 1014 1015 1016 1017 1018 1019 1020 1021 1022 1023 1024 1025 1026 1027 1028 1029 1030 1031 1032 1033 Figura 2. Árvore com as espécies delimitadas pelo modelo GMYC, construídas a partir de 1034
147 sequências de código de barras de DNA. 1035
1036
1037 1038 1039 1040 1041 1042 1043 1044 1045 1046 1047 1048 1049 1050 1051 1052 1053 1054 1055 1056 1057 1058

71
Tabela 3. Número de espécies que foram identificadas pelo banco de dados online 1059
BOLD (Barcode of life) e NCBI (National Center Biotechnology Information). 1060
1061
1062
1063
1064
1065
1066
1067
1068
1069
1070
1071
1072
1073
1074
Tabela 4. Valores médios das divergências intraespecífica e interespecífica do gene COI 1075
(divergência mínima, média e máxima) das 147 sequências obtidas pelo método de Kimura-2-1076
parâmetros. 1077
Dados Moleculares N Div. mínima Div. média Div. máx.
Intraespecífica 147 0,0 0,0 0,01
Interespecífica 147 0,0 0,32 0,55
1078
1079
1080
1081
1082
1083
Família Espécies Número de
espécimes BOLD NCBI
Cichlidae Apistogramma agassizi 14 7
Cichlidae Geophagus argyrostictus 7 7
Cichlidae Geophagus altifrons 2 1
Cichlidae Heros efasciatus 9 9
Cichlidae Symphysodon aequafasciatus 3 3
Cichlidae Acaronia nassa 2 2
Cichlidae Cichla piquiti 1 1
Cichlidae Acarichthys heckelii 4 4
Cichlidae Retroculus lapidifer 3 3
Cichlidae Retroculus xinguensis 2 2
Cichlidae Satanoperca lilith 3 3
Cichlidae Teleocichla sp. 1 1
Cichlidae Crenicichla percna 1 1
TOTAL 13 52 38 8

72
Tabela 5. Porcentagem média da distância intraespecífica (K2P) entre os genêros de Ciclídeos 1084
amazônicos representados por mais que uma espécie. 1085
Gênero Número de
gêneros
% média da
distância
intraespecífica
Aequidens 3 11%
Apistogramma 6 17%
Acarichthys 1 0%
Acaronia 1 3%
Biotodoma 1 N/A
Caquetaia 1 0%
Crenicichla 11 17%
Cichla 4 3%
Geophagus 4 6%
Heros 1 0%
Laetacara 1 N/A
Mesonauta 1 1%
Retroculus 2 5%
Satanoperca 3 15%
Symphysodon 1 0%
Teleocichla 4 13%
1086 1087 1088
1089
1090
Tabela 6. Porcentagem média da distância intraespecífica (K2P) para as espécies de ciclídeos 1091
classificados morfologicamente e representados por mais de um espécime. 1092
Espécie
Número de
espécimens
analisadas
% média da
distância
intraespecífica
Aequidens michaeli 2 0%
Aequidens pallidus 6 8%
Aequidens cf. eape 1 N/A
Apistogramma sp. 1 N/A
Apistogramma aff. staecki 1 N/A
Apistogramma agassizi 14 3%
Apisotgramma pertensis 6 0%
Apistogramma gr. brevis 2 1%

73
Apistogramma pulchra 5 0%
Acarichthys heckelii 4 0%
Acaronia nassa 2 3%
Biotodoma cupido 1 N/A
Caquetaia spectabilis 2 0%
Crenicichla n. sp. laranja 4 0,2%
Crenicichla percna 1 N/A
Crenicichla aff. inpa 1 N/A
Crenicichla sp.1 1 N/A
Crencichla jegui 1 N/A
Crenicichla lugubris 2 0%
Crenicichla astroblepa 4 0%
Crenicichla aff strigata 1 N/A
Crenicichla n. sp. preta 5 1%
Crenicichla sp. xingu 6 0,2%
Crenicichla regani 7 3%
Cichla melaniae 7 1%
Cichla mirianae pitanga 2 0%
Cichla temensis 3 0%
Cichla piquiti 1 N/A
Geophagus altifrons 2 7%
Geophagus argyrostictus 7 0,3%
Geophagus sp. 4 3%
Geophagus proximus 3 0,5%
Heros efasciatus 9 0%
Laetacara araguaiae 1 N/A
Mesonauta festivus 5 1%
Retroculus lapidifer 3 0,2%
Retroculus xinguensis 2 0%
Satanoperca jurupari 1 N/A
Satnoperca sp. 2 0%
Satanoperca lilith 3 0,2%
Symphysodon aequafasciatus 3 0,2%
Teleocichla sp. 1 N/A
Teleocichla n. sp. preta 4 0,1%
Teleocichla cf. gephyrogramma 1 N/A
Teleocichla cinderela 3 6%
1093 1094 1095 1096 1097 1098 1099 1100

74
Tabela 7. Porcentagem média da distância intraespecífica (K2P) para as espécies de ciclídeos 1101
classificados molecularmente e representados por mais de um espécime. 1102
1103
Espécie
Número de
espécimens
analisadas
% média da
distância
intraespecífica
Aequidens michaeli 2 0%
Aequidens pallidus_JAT3 1 N/A
Aequidens pallidus_JAT1 2 0%
Aequidens pallidus_JAT2 1 N/A
Aequidens pallidus_RN 2 0%
Aequidens cf. eape 1 N/A
Apistogramma sp. 1 N/A
Apistogramma aff staecki 1 N/A
Apistogramma agassizi_JAT1 3 0%
Apistogramma agassizi_JAT2 5 0.3%
Apistogramma agassizi_PUR 5 0.3%
Apistogramma agassizi_TAP 1 N/A
Apistogramma pertensis_JAT 6 0%
Apistogramma gr brevis_RN 2 0.5%
Apistogramma pulchra_PUR 5 0%
Acarichthys heckelli_JAT_TAP 4 0%
Acaronia nassa_PUR 1 N/A
Acaronia nassa_TAP 1 N/A
Biotodoma cupido 1 N/A
Caquetaia spectabilis 2 0%
Crenicichla n. sp. laranja 4 0.2%
Crenicichla percna 1 N/A
Crenicichla aff. Inpa 1 N/A
Crenicichla sp1 1 N/A
Crenicichla lugubris 4 0%
Crenicichla astroblepa 4 0%
Crenicichla n sp preta 5 1%
Crenicichla sp xingu_XIN 6 0%
Crenicichla regani_XIN 1 N/A
Crenicichla regani_JAT 2 0%
Crenicichla regani_PUR 2 0.3%
Crenicichla regani_TAP 2 0.3%
Cichla melaniae_XIN2 3 0%
Cichla melaniae_XIN1 4 0%
Cichla mirianae pitanga 2 0%
Cichla temensis 3 0%

75
Cichla piquiti 1 N/A
Geophagus altifrons_ARA 1 N/A
Geophagus altifrons_PUR 1 N/A
Geophagus argyrostictus_XIN2 4 0%
Geophagus argyrostictus_XIN1 3 0%
Geophagus sp_TAP 3 0.5%
Geophagus sp_XIN 1 N/A
Geophagus proximus_TAP 2 0.5%
Heros efasciatus_XIN_JAT 9 0%
Laetacara araguaiae_TAP 1 N/A
Mesonauta festivus_TAP 2 0.3%
Mesonauta festivus_JAT 3 0.4%
Retroculus lapidifer 3 0.2%
Retroculus xinguensis 2 0%
Satanoperca jurupari 1 N/A
Satanoperca sp. 2 0%
Satanoperca lilith 3 0.2%
Symphysodon aequafasciatus 3 0.2%
Teleocichla sp. 1 N/A
Teleocichla n. sp. preta 5 0.3%
Teleocichla cinderela_ARA1 1 N/A
Teleocichla cinderela_ARA2 1 N/A
Teleocichla cinderela_ARA3 1 N/A
1104 1105 1106 Tabela 8. Número de linhagens biológicas encontradas para cada espécie de ciclídeos na Bacia 1107
Amazônica. 1108
1109
Família Espécies Número de linhagens
Cichlidae Apistogramma agassizi 4
Cichlidae Apistogramma sp. 1
Cichlidae Apistogramma pertensis 1
Cichlidae Apistogramma pulchra 1
Cichlidae Apistogramma gr. brevis 1
Cichlidae Apistogramma aff. staecki 1
Cichlidae Biotodoma cupido 1
Cichlidae Geophagus argyrostictus 2
Cichlidae Geophagus altifrons 2
Cichlidae Geophagus proximus 1
Cichlidae Geophagus sp. 2
Cichlidae Caquetaia spectabilis 1
Cichlidae Heros efasciatus 1

76
Cichlidae Symphysodon aequafasciatus 1
Cichlidae Mesonauta festivus 2
Cichlidae Laetacara araguaiae 1
Cichlidae Acaronia nassa 2
Cichlidae Cichla melaniae 2
Cichlidae Cichla piquiti 1
Cichlidae Cichla temensis 1
Cichlidae Cichla mirianae pitanga 1
Cichlidae Acarichthys heckelii 1
Cichlidae Aequidens cf. epae 1
Cichlidae Aequidens pallidus 4
Cichlidae Aequidens michaeli 1
Cichlidae Retroculus lapidifer 1
Cichlidae Retroculus xinguensis 1
Cichlidae Satanoperca jurupari 1
Cichlidae Satanoperca sp. 1
Cichlidae Satanoperca lilith 1
Cichlidae Teleocichla cf. gephyrogramma 1
Cichlidae Teleocichla n. sp. preta -
Cichlidae Teleocichla sp. 1
Cichlidae Teleocichla cinderela 3
Cichlidae Crenicichla regani 4
Cichlidae Crenicichla aff. inpa 1
Cichlidae Crenicichla percna 1
Cichlidae Crenicichla sp. preta 2
Cichlidae Crenicichla sp. xingu 1
Cichlidae Crenicichla jegui 1
Cichlidae Crenicichla lugubris -
Cichlidae Crenicichla strigata -
Cichlidae Crenicichla n. sp. laranja 1
Cichlidae Crenicichla astroblepa 1
Cichlidae Crenicichla sp. 1 1
TOTAL 45 60
1110 1111 1112 1113 1114 1115 1116 1117 1118 1119

77
Tabela 9. Distância média da distância interespecífica (K2P) para as espécies de ciclídeos delimitados molecularmente pelo modelo Generalized Mixed 1120
Yule Coalescent (GMYC). 1121
1122
Aeq
_m
icha
eli_
XIN
Aeq
_p
all
idu
s_JA
T3
Aeq
_p
all
idu
s_JA
T1
Aeq
_p
all
idu
s_JA
T2
Aeq
_p
all
idu
s_R
N
Aeq
_cf
_ea
pe_
TA
P
Ap_
sp_
XIN
Ap_
aff
_st
aec
ki_JA
T
Ap_
aga
ssiz
i_JA
T1
Ap_
aga
ssiz
i_JA
T2
Ap_
aga
ssiz
i_P
UR
Ap_
aga
ssiz
i_T
AP
Ap_
per
ten
sis_
JAT
Ap_
gr_
bre
vis_
RN
Ap_
pu
lchra
_P
UR
Aca
_h
ecke
lli_
JAT
_T
AP
Ac_
na
ssa_
PU
R
Ac_
na
ssa_
TA
P
Bio
_cu
pid
o_P
UR
Caq_
spec
tab
ilis
_X
IN
Cr_
n_
sp_
lara
nja
_X
IN
Cr_
per
cna_
XIN
Cr_
aff
_in
pa_
XIN
Cr_
sp1
_A
RA
Cr_
lugu
bri
s_A
RA
Aeq_michaeli_XIN
Aeq_pallidus_JAT3 0.15
Aeq_pallidus_JAT1 0.10 0.12
Aeq_pallidus_JAT2 0.16 0.01 0.11
Aeq_pallidus_RN 0.15 0.06 0.11 0.07
Aeq_cf_eape_TAP 0.15 0.20 0.14 0.20 0.16
Ap_sp_XIN 0.34 0.32 0.35 0.32 0.30 0.37
Ap_aff_staecki_JAT 0.39 0.36 0.39 0.37 0.37 0.36 0.22
Ap_agassizi_JAT1 0.37 0.40 0.38 0.41 0.39 0.37 0.26 0.19
Ap_agassizi_JAT2 0.36 0.39 0.37 0.40 0.38 0.38 0.26 0.19 0.00
Ap_agassizi_PUR 0.34 0.35 0.36 0.37 0.33 0.33 0.25 0.21 0.06 0.06
Ap_agassizi_TAP 0.36 0.36 0.36 0.37 0.35 0.37 0.27 0.19 0.04 0.04 0.03
Ap_pertensis_JAT 0.36 0.39 0.40 0.41 0.38 0.39 0.24 0.20 0.22 0.23 0.24 0.22
Ap_gr_brevis_RN 0.37 0.34 0.35 0.35 0.35 0.37 0.29 0.20 0.24 0.24 0.21 0.21 0.25
Ap_pulchra_PUR 0.37 0.41 0.39 0.41 0.39 0.39 0.27 0.19 0.25 0.25 0.27 0.27 0.17 0.27
Aca_heckelli_JAT_TAP 0.24 0.29 0.25 0.27 0.27 0.24 0.31 0.31 0.31 0.32 0.36 0.35 0.32 0.35 0.35
Ac_nassa_PUR 0.22 0.24 0.21 0.24 0.23 0.23 0.34 0.37 0.37 0.38 0.36 0.36 0.44 0.44 0.46 0.27
Ac_nassa_TAP 0.21 0.24 0.22 0.24 0.21 0.22 0.33 0.34 0.35 0.36 0.35 0.35 0.42 0.43 0.42 0.24 0.03
Bio_cupido_PUR 0.35 0.37 0.37 0.39 0.38 0.37 0.36 0.42 0.37 0.38 0.39 0.37 0.44 0.43 0.41 0.41 0.39 0.40
Caq_spectabilis_XIN 0.26 0.26 0.28 0.26 0.24 0.26 0.35 0.36 0.36 0.37 0.35 0.37 0.38 0.38 0.41 0.19 0.21 0.18 0.42
Cr_n_sp_laranja_XIN 0.24 0.31 0.32 0.32 0.31 0.35 0.38 0.40 0.41 0.40 0.41 0.43 0.45 0.44 0.44 0.28 0.30 0.29 0.37 0.31
Cr_percna_XIN 0.26 0.32 0.32 0.32 0.31 0.33 0.37 0.34 0.41 0.41 0.42 0.42 0.42 0.40 0.42 0.31 0.31 0.31 0.34 0.34 0.08

78
Cr_aff_inpa_XIN 0.30 0.37 0.32 0.36 0.38 0.34 0.41 0.45 0.43 0.42 0.43 0.43 0.47 0.46 0.47 0.27 0.33 0.31 0.36 0.36 0.16 0.20
Cr_sp1_ARA 0.35 0.40 0.37 0.42 0.42 0.32 0.37 0.41 0.38 0.39 0.37 0.38 0.42 0.37 0.48 0.27 0.30 0.31 0.38 0.34 0.24 0.25 0.23
Cr_lugubris_ARA 0.29 0.33 0.38 0.35 0.29 0.34 0.36 0.39 0.41 0.42 0.41 0.40 0.45 0.41 0.45 0.31 0.28 0.27 0.37 0.33 0.11 0.10 0.20 0.23
Cr_astroblepa_ARA 0.38 0.40 0.39 0.41 0.41 0.33 0.37 0.37 0.35 0.36 0.34 0.35 0.41 0.36 0.47 0.30 0.29 0.28 0.35 0.32 0.26 0.26 0.26 0.08 0.22
Cr_n_sp_preta_XIN 0.29 0.36 0.34 0.36 0.34 0.37 0.37 0.38 0.38 0.38 0.43 0.41 0.42 0.43 0.42 0.30 0.26 0.26 0.36 0.29 0.08 0.07 0.16 0.27 0.12
Aeq
_m
icha
eli_
XIN
Aeq
_p
all
idu
s_JA
T3
Aeq
_p
all
idu
s_JA
T1
Aeq
_p
all
idu
s_JA
T2
Aeq
_p
all
idu
s_R
N
Aeq
_cf
_ea
pe_
TA
P
Ap_
sp_
XIN
Ap_
aff
_st
aec
ki_JA
T
Ap_
aga
ssiz
i_JA
T1
Ap_
aga
ssiz
i_JA
T2
Ap_
aga
ssiz
i_P
UR
Ap_
aga
ssiz
i_T
AP
Ap_
per
ten
sis_
JAT
Ap_
gr_
bre
vis_
RN
Ap_
pu
lchra
_P
UR
Aca
_h
ecke
lli_
JAT
_T
AP
Ac_
na
ssa_
PU
R
Ac_
na
ssa_
TA
P
Bio
_cu
pid
o_P
UR
Caq_
spec
tab
ilis
_X
IN
Cr_
n_
sp_
lara
nja
_X
IN
Cr_
per
cna_
XIN
Cr_
aff
_in
pa_
XIN
Cr_
sp1
_A
RA
Cr_
lugu
bri
s_A
RA
Cr_sp_xingu_XIN 0.26 0.34 0.32 0.34 0.32 0.35 0.36 0.38 0.37 0.37 0.41 0.41 0.42 0.44 0.43 0.29 0.25 0.24 0.35 0.31 0.07 0.08 0.16 0.27 0.11
Cr_regani_XIN 0.35 0.38 0.40 0.39 0.40 0.31 0.50 0.42 0.40 0.39 0.40 0.40 0.51 0.47 0.44 0.33 0.35 0.32 0.35 0.35 0.23 0.23 0.24 0.28 0.28
Cr_regani_JAT 0.33 0.37 0.40 0.37 0.36 0.31 0.46 0.41 0.38 0.38 0.40 0.40 0.47 0.42 0.43 0.33 0.32 0.29 0.34 0.32 0.19 0.23 0.25 0.27 0.25
Cr_regani_PUR 0.33 0.36 0.40 0.36 0.35 0.32 0.47 0.39 0.38 0.37 0.41 0.39 0.46 0.42 0.41 0.33 0.32 0.29 0.33 0.32 0.20 0.22 0.24 0.28 0.23
Cr_regani_TAP 0.34 0.38 0.40 0.38 0.39 0.31 0.50 0.41 0.40 0.39 0.41 0.40 0.50 0.47 0.44 0.31 0.34 0.31 0.36 0.35 0.22 0.23 0.23 0.27 0.27
Ci_melaniae_XIN2 0.23 0.24 0.25 0.24 0.22 0.26 0.30 0.35 0.34 0.35 0.36 0.35 0.37 0.41 0.35 0.23 0.25 0.23 0.35 0.24 0.31 0.30 0.36 0.33 0.33
Ci_melaniae_XIN1 0.25 0.25 0.25 0.25 0.24 0.27 0.34 0.34 0.38 0.37 0.40 0.39 0.39 0.41 0.37 0.24 0.24 0.24 0.37 0.23 0.31 0.29 0.36 0.34 0.33
Ci_mirianae_pitanga_XIN 0.23 0.26 0.22 0.25 0.21 0.21 0.33 0.38 0.38 0.39 0.36 0.37 0.38 0.42 0.41 0.25 0.21 0.20 0.32 0.25 0.33 0.31 0.36 0.29 0.35
Ci_temensis_JAT 0.21 0.24 0.24 0.24 0.22 0.26 0.31 0.36 0.33 0.33 0.34 0.33 0.37 0.40 0.37 0.23 0.24 0.24 0.37 0.22 0.30 0.29 0.35 0.32 0.31
Ci_piquiti_ARA 0.23 0.24 0.24 0.24 0.22 0.29 0.30 0.36 0.32 0.33 0.34 0.33 0.36 0.39 0.35 0.25 0.25 0.25 0.36 0.24 0.30 0.31 0.38 0.34 0.32
Geo_altifrons_ARA 0.23 0.29 0.29 0.30 0.31 0.32 0.31 0.45 0.39 0.39 0.36 0.39 0.33 0.38 0.35 0.29 0.28 0.28 0.37 0.31 0.28 0.28 0.29 0.38 0.35
Geo_altifrons_PUR 0.27 0.30 0.32 0.30 0.31 0.37 0.31 0.46 0.39 0.39 0.36 0.37 0.34 0.38 0.35 0.27 0.32 0.32 0.35 0.32 0.30 0.31 0.31 0.38 0.34
Geo_argyrostictus_XIN2 0.22 0.29 0.29 0.29 0.29 0.28 0.34 0.39 0.34 0.35 0.34 0.34 0.33 0.36 0.38 0.25 0.28 0.28 0.35 0.32 0.28 0.29 0.30 0.38 0.33
Geo_argyrostictus_XIN1 0.22 0.29 0.27 0.29 0.29 0.26 0.36 0.41 0.36 0.36 0.34 0.34 0.33 0.37 0.38 0.25 0.28 0.29 0.35 0.32 0.30 0.30 0.31 0.38 0.34
Geo_sp_TAP 0.22 0.29 0.29 0.30 0.30 0.32 0.31 0.46 0.40 0.41 0.38 0.40 0.35 0.39 0.37 0.28 0.30 0.30 0.36 0.30 0.26 0.26 0.28 0.38 0.34
Geo_sp_XIN 0.24 0.32 0.31 0.33 0.32 0.34 0.30 0.43 0.38 0.39 0.37 0.39 0.33 0.36 0.36 0.23 0.29 0.29 0.35 0.28 0.29 0.29 0.31 0.37 0.37
Geo_proximus_TAP 0.24 0.29 0.30 0.29 0.29 0.34 0.29 0.44 0.38 0.39 0.36 0.37 0.33 0.38 0.35 0.24 0.30 0.30 0.38 0.29 0.27 0.29 0.30 0.33 0.32
He_efasciatus_XIN_JAT 0.23 0.22 0.21 0.23 0.22 0.22 0.39 0.34 0.37 0.37 0.36 0.36 0.37 0.36 0.40 0.25 0.23 0.23 0.43 0.19 0.36 0.31 0.37 0.36 0.36
Lae_araguaiae_TAP 0.21 0.23 0.24 0.24 0.24 0.22 0.37 0.37 0.43 0.43 0.39 0.41 0.40 0.40 0.46 0.25 0.17 0.19 0.37 0.25 0.31 0.31 0.28 0.30 0.29
Mes_festivus_TAP 0.25 0.23 0.25 0.23 0.22 0.25 0.37 0.41 0.42 0.41 0.37 0.41 0.49 0.37 0.50 0.30 0.25 0.24 0.42 0.20 0.31 0.30 0.38 0.33 0.32

79
Mes_festivus_JAT 0.23 0.23 0.23 0.23 0.21 0.25 0.35 0.40 0.39 0.39 0.36 0.39 0.46 0.38 0.47 0.29 0.22 0.22 0.39 0.19 0.30 0.30 0.38 0.31 0.31
Ret_lapidifer_ARA 0.28 0.26 0.27 0.26 0.25 0.30 0.32 0.41 0.35 0.36 0.37 0.40 0.38 0.38 0.41 0.29 0.23 0.22 0.38 0.23 0.34 0.36 0.35 0.38 0.35
Ret_xinguensis_XIN 0.28 0.31 0.27 0.31 0.27 0.32 0.35 0.40 0.36 0.37 0.37 0.40 0.41 0.41 0.46 0.29 0.24 0.23 0.42 0.23 0.35 0.38 0.35 0.41 0.40
Sat_jurupari_ARA 0.23 0.28 0.27 0.27 0.26 0.30 0.37 0.33 0.38 0.37 0.37 0.36 0.44 0.34 0.40 0.27 0.25 0.24 0.41 0.23 0.32 0.33 0.35 0.36 0.33
Sat_sp_XIN 0.26 0.27 0.27 0.27 0.28 0.31 0.32 0.34 0.37 0.36 0.36 0.35 0.42 0.30 0.38 0.27 0.28 0.26 0.40 0.23 0.32 0.31 0.35 0.34 0.35
Sat_lilith_JAT 0.26 0.28 0.30 0.29 0.30 0.24 0.33 0.39 0.38 0.39 0.36 0.40 0.42 0.36 0.41 0.26 0.24 0.22 0.40 0.25 0.32 0.32 0.34 0.32 0.31
Sym_aequafasciatus_XIN 0.25 0.21 0.21 0.23 0.23 0.26 0.37 0.40 0.40 0.41 0.36 0.38 0.40 0.41 0.46 0.29 0.22 0.25 0.42 0.22 0.31 0.33 0.33 0.36 0.37
Te_sp_ARA 0.29 0.30 0.34 0.31 0.33 0.38 0.40 0.44 0.42 0.41 0.38 0.40 0.44 0.44 0.43 0.37 0.34 0.33 0.40 0.39 0.23 0.26 0.25 0.36 0.29
Te_n_sp_preta_XIN 0.29 0.28 0.34 0.30 0.29 0.34 0.36 0.38 0.40 0.39 0.37 0.40 0.41 0.39 0.43 0.33 0.30 0.29 0.39 0.33 0.26 0.26 0.23 0.31 0.27
Te_cinderela_ARA1 0.38 0.40 0.43 0.42 0.42 0.43 0.53 0.48 0.51 0.50 0.49 0.49 0.53 0.49 0.55 0.41 0.39 0.37 0.43 0.39 0.30 0.31 0.34 0.38 0.34
Te_cinderela_ARA2 0.39 0.39 0.44 0.42 0.41 0.43 0.53 0.47 0.54 0.53 0.52 0.52 0.51 0.50 0.53 0.44 0.40 0.38 0.44 0.42 0.31 0.30 0.34 0.39 0.35
Te_cinderela_ARA3 0.34 0.36 0.38 0.38 0.38 0.38 0.48 0.44 0.49 0.49 0.46 0.46 0.48 0.46 0.49 0.39 0.35 0.33 0.40 0.38 0.28 0.27 0.29 0.35 0.30
Cr_
ast
rob
lepa_
AR
A
Cr_
n_
sp_p
reta
_X
IN
Cr_
sp_
xingu
_X
IN
Cr_
regan
i_X
IN
Cr_
regan
i_JA
T
Cr_
regan
i_P
UR
Cr_
regan
i_T
AP
Ci_
mel
an
iae_
XIN
2
Ci_
mel
an
iae_
XIN
1
Ci_
mir
iana
e_pit
an
ga_
XIN
Ci_
tem
ensi
s_JA
T
Ci_
piq
uit
i_A
RA
Geo
_alt
ifro
ns_
AR
A
Geo
_alt
ifro
ns_
PU
R
Geo
_a
rgyr
ost
ictu
s_X
IN2
Geo
_a
rgyr
ost
ictu
s_X
IN1
Geo
_sp
_T
AP
Geo
_sp
_X
IN
Geo
_p
roxi
mu
s_T
AP
He_
efasc
iatu
s_X
IN_JA
T
La
e_a
rag
uaia
e_T
AP
Mes
_fe
stiv
us_
TA
P
Mes
_fe
stiv
us_
JAT
Ret
_la
pid
ifer
_A
RA
Ret
_xi
ngu
ensi
s_X
IN
Cr_n_sp_preta_XIN 0.28
Cr_sp_xingu_XIN 0.27 0.05
Cr_regani_XIN 0.31 0.23 0.26
Cr_regani_JAT 0.30 0.22 0.25 0.04
Cr_regani_PUR 0.30 0.21 0.24 0.04 0.01
Cr_regani_TAP 0.30 0.23 0.25 0.01 0.04 0.04
Ci_melaniae_XIN2 0.33 0.33 0.32 0.33 0.33 0.35 0.32
Ci_melaniae_XIN1 0.32 0.32 0.32 0.33 0.33 0.35 0.32 0.02
Ci_mirianae_pitanga_XIN 0.27 0.35 0.34 0.35 0.33 0.36 0.34 0.06 0.07
Ci_temensis_JAT 0.32 0.32 0.31 0.35 0.33 0.35 0.33 0.02 0.03 0.07
Ci_piquiti_ARA 0.32 0.33 0.31 0.35 0.35 0.36 0.33 0.02 0.04 0.07 0.03
Geo_altifrons_ARA 0.38 0.30 0.29 0.34 0.35 0.35 0.35 0.29 0.29 0.29 0.27 0.29
Geo_altifrons_PUR 0.40 0.32 0.31 0.33 0.33 0.33 0.34 0.28 0.30 0.29 0.28 0.27 0.07
Geo_argyrostictus_XIN2 0.37 0.29 0.27 0.30 0.30 0.32 0.31 0.27 0.27 0.26 0.25 0.26 0.10 0.09

80
Geo_argyrostictus_XIN1 0.37 0.31 0.28 0.30 0.32 0.33 0.31 0.27 0.27 0.25 0.25 0.26 0.10 0.10 0.01
Geo_sp_TAP 0.39 0.28 0.27 0.32 0.33 0.33 0.33 0.29 0.29 0.28 0.28 0.29 0.02 0.06 0.09 0.09
Geo_sp_XIN 0.39 0.29 0.28 0.34 0.34 0.33 0.35 0.29 0.29 0.27 0.27 0.27 0.06 0.04 0.10 0.10 0.05
Geo_proximus_TAP 0.35 0.30 0.28 0.31 0.33 0.33 0.32 0.26 0.28 0.26 0.25 0.25 0.06 0.03 0.10 0.10 0.05 0.04
He_efasciatus_XIN_JAT 0.35 0.34 0.36 0.35 0.33 0.33 0.34 0.24 0.23 0.24 0.23 0.26 0.33 0.32 0.32 0.32 0.32 0.29 0.29
Lae_araguaiae_TAP 0.31 0.30 0.31 0.33 0.30 0.31 0.31 0.24 0.24 0.22 0.24 0.25 0.31 0.32 0.31 0.33 0.32 0.31 0.32 0.20
Mes_festivus_TAP 0.34 0.32 0.32 0.41 0.37 0.38 0.40 0.22 0.22 0.23 0.23 0.24 0.34 0.34 0.32 0.34 0.33 0.32 0.32 0.22 0.25
Mes_festivus_JAT 0.33 0.30 0.29 0.39 0.35 0.36 0.38 0.21 0.20 0.21 0.21 0.22 0.33 0.32 0.31 0.32 0.32 0.30 0.31 0.22 0.24 0.02
Ret_lapidifer_ARA 0.35 0.33 0.32 0.39 0.37 0.37 0.38 0.24 0.27 0.23 0.24 0.24 0.25 0.27 0.28 0.28 0.26 0.24 0.26 0.28 0.28 0.32 0.30
Ret_xinguensis_XIN 0.41 0.34 0.33 0.45 0.41 0.41 0.42 0.29 0.29 0.27 0.27 0.27 0.28 0.30 0.30 0.31 0.29 0.26 0.30 0.28 0.28 0.31 0.29 0.09
Sat_jurupari_ARA 0.32 0.31 0.31 0.35 0.37 0.36 0.34 0.24 0.25 0.25 0.23 0.24 0.31 0.32 0.30 0.32 0.31 0.33 0.31 0.28 0.27 0.28 0.25 0.29 0.30
Sat_sp_XIN 0.33 0.31 0.32 0.35 0.35 0.36 0.36 0.23 0.24 0.26 0.23 0.22 0.31 0.29 0.30 0.31 0.30 0.29 0.29 0.29 0.26 0.28 0.26 0.32 0.32
Sat_lilith_JAT 0.31 0.32 0.31 0.35 0.34 0.36 0.34 0.24 0.25 0.21 0.25 0.24 0.30 0.27 0.29 0.30 0.30 0.29 0.28 0.30 0.25 0.27 0.26 0.22 0.29
Sym_aequafasciatus_XIN 0.37 0.34 0.30 0.44 0.39 0.42 0.42 0.24 0.25 0.21 0.22 0.23 0.30 0.27 0.29 0.29 0.28 0.27 0.25 0.17 0.24 0.21 0.18 0.31 0.33
Te_sp_ARA 0.37 0.24 0.22 0.31 0.30 0.29 0.28 0.32 0.33 0.35 0.31 0.30 0.33 0.35 0.35 0.36 0.34 0.36 0.35 0.39 0.34 0.31 0.29 0.39 0.39
Te_n_sp_preta_XIN 0.34 0.25 0.24 0.31 0.29 0.29 0.30 0.30 0.32 0.30 0.30 0.30 0.37 0.37 0.36 0.38 0.37 0.36 0.35 0.32 0.30 0.33 0.32 0.35 0.37
Te_cinderela_ARA1 0.35 0.31 0.31 0.32 0.30 0.31 0.30 0.33 0.35 0.33 0.33 0.34 0.43 0.47 0.40 0.40 0.42 0.43 0.43 0.38 0.36 0.37 0.36 0.41 0.42
Te_cinderela_ARA2 0.36 0.31 0.31 0.31 0.29 0.29 0.29 0.34 0.35 0.33 0.34 0.35 0.43 0.46 0.42 0.42 0.43 0.44 0.43 0.40 0.35 0.39 0.38 0.40 0.44
Te_cinderela_ARA3 0.31 0.27 0.28 0.27 0.25 0.26 0.25 0.30 0.31 0.30 0.29 0.31 0.38 0.41 0.37 0.37 0.38 0.38 0.38 0.34 0.31 0.35 0.33 0.36 0.38
Sat_
juru
pa
ri_
AR
A
Sat_
sp_
XIN
Sat_
lili
th_JA
T
Sym
_aeq
ua
fasc
iatu
s_X
IN
Te_
sp_A
RA
Te_
n_
sp_
pre
ta_
XIN
Te_
cin
der
ela_
AR
A1
Te_
cin
der
ela_
AR
A2
Te_
cin
der
ela_
AR
A3
Sat_jurupari_ARA
Sat_sp_XIN 0.04
Sat_lilith_JAT 0.24 0.23
Sym_aequafasciatus_XIN 0.28 0.26 0.26
Te_sp_ARA 0.34 0.38 0.32 0.37
Te_n_sp_preta_XIN 0.32 0.33 0.33 0.34 0.15

81
Te_cinderela_ARA1 0.40 0.40 0.33 0.44 0.16 0.22
Te_cinderela_ARA2 0.43 0.43 0.34 0.43 0.16 0.24 0.05
Te_cinderela_ARA3 0.38 0.38 0.31 0.39 0.12 0.21 0.08 0.05
1123 1124 1125 1126 1127 1128 1129 1130 1131 1132 1133 1134 1135 1136 1137 1138 1139 1140 1141 1142 1143 1144 1145 1146 1147 1148 1149 1150 1151

82
1152 Tabela 10. Distância média da distância interespecífica (K2P) para as espécies de ciclídeos classificados morfologicamente. 1153
Aeq
_m
icha
eli
Aeq
_p
all
idu
s
Aeq
_cf
_ea
pe
Ap_
sp.
Ap_
aff
_st
aec
ki
Ap_
aga
ssiz
i
Ap_
per
ten
sis
Ap_
gr_
bre
vis
Ap_
pu
lchra
Aca
_h
ecke
lli
Ac_
na
ssa
Bio
_cu
pid
o
Caq_
spec
tab
ilis
Cr_
n_
sp_
lara
nja
Cr_
per
cna
Cr_
aff
_in
pa
Cr_
sp1
Cr_
jeg
ui
Cr_
lugu
bri
s
Cr_
ast
rob
lepa
Cr_
aff
_st
riga
ta
Cr_
n_
sp_p
reta
Cr_
sp_
xingu
Cr_
regan
i
Ci_
mel
an
iae
Ci_
mir
iana
e_pit
an
ga
Ci_
tem
ensi
s
Ci_
piq
uit
i
Geo
_alt
ifro
ns
Aeq_michaeli
Aeq_pallidus 0.1
Aeq_cf_eape 0.2 0.2
Ap_sp. 0.3 0.3 0.4
Ap_aff_staecki 0.4 0.4 0.4 0.2
Ap_agassizi 0.4 0.4 0.4 0.3 0.2
Ap_pertensis 0.4 0.4 0.4 0.2 0.2 0.2
Ap_gr_brevis 0.4 0.3 0.4 0.3 0.2 0.2 0.2
Ap_pulchra 0.4 0.4 0.4 0.3 0.2 0.3 0.2 0.3
Aca_heckelli 0.2 0.3 0.2 0.3 0.3 0.3 0.3 0.4 0.4
Ac_nassa 0.2 0.2 0.2 0.3 0.4 0.4 0.4 0.4 0.4 0.3
Bio_cupido 0.4 0.4 0.4 0.4 0.4 0.4 0.4 0.4 0.4 0.4 0.4
Caq_spectabilis 0.3 0.3 0.3 0.4 0.4 0.4 0.4 0.4 0.4 0.2 0.2 0.4
Cr_n_sp_laranja 0.2 0.3 0.4 0.4 0.4 0.4 0.5 0.4 0.4 0.3 0.3 0.4 0.3
Cr_percna 0.3 0.3 0.3 0.4 0.3 0.4 0.4 0.4 0.4 0.3 0.3 0.3 0.3 0.1
Cr_aff_inpa 0.3 0.4 0.3 0.4 0.4 0.4 0.5 0.5 0.5 0.3 0.3 0.4 0.4 0.2 0.2
Cr_sp1 0.3 0.4 0.3 0.4 0.4 0.4 0.4 0.4 0.5 0.3 0.3 0.4 0.3 0.2 0.2 0.2
Cr_jegui 0.3 0.3 0.3 0.4 0.4 0.4 0.5 0.4 0.5 0.3 0.3 0.4 0.3 0.1 0.1 0.2 0.2
Cr_lugubris 0.3 0.3 0.3 0.4 0.4 0.4 0.4 0.4 0.5 0.3 0.3 0.4 0.3 0.1 0.1 0.2 0.2 0.0
Cr_astroblepa 0.4 0.4 0.3 0.4 0.4 0.3 0.4 0.4 0.5 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.1 0.2 0.2
Cr_aff_strigata 0.3 0.3 0.3 0.4 0.4 0.4 0.4 0.4 0.5 0.3 0.3 0.4 0.3 0.1 0.1 0.2 0.2 0.0 0.0 0.2
Cr_n_sp_preta 0.3 0.3 0.4 0.4 0.4 0.4 0.4 0.4 0.4 0.3 0.3 0.4 0.3 0.1 0.1 0.2 0.3 0.1 0.1 0.3 0.1
Cr_sp_xingu 0.3 0.3 0.4 0.4 0.4 0.4 0.4 0.4 0.4 0.3 0.2 0.4 0.3 0.1 0.1 0.2 0.3 0.1 0.1 0.3 0.1 0.1

83
Cr_regani 0.3 0.4 0.3 0.5 0.4 0.4 0.5 0.4 0.4 0.3 0.3 0.3 0.3 0.2 0.2 0.2 0.3 0.3 0.3 0.3 0.3 0.2 0.3
Ci_melaniae 0.2 0.2 0.3 0.3 0.3 0.4 0.4 0.4 0.4 0.2 0.2 0.4 0.2 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3
Ci_mirianae_pitanga 0.2 0.2 0.2 0.3 0.4 0.4 0.4 0.4 0.4 0.2 0.2 0.3 0.2 0.3 0.3 0.4 0.3 0.4 0.3 0.3 0.3 0.3 0.3 0.3 0.1
Ci_temensis 0.2 0.2 0.3 0.3 0.4 0.3 0.4 0.4 0.4 0.2 0.2 0.4 0.2 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.02 0.1
Ci_piquiti 0.2 0.2 0.3 0.3 0.4 0.3 0.4 0.4 0.4 0.2 0.3 0.4 0.2 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.03 0.1 0.03
Geo_altifrons 0.2 0.3 0.3 0.3 0.5 0.4 0.3 0.4 0.4 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.4 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3
Geo_argysrostictus 0.2 0.3 0.3 0.3 0.4 0.3 0.3 0.4 0.4 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.4 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.1
Geo_sp 0.2 0.3 0.3 0.3 0.5 0.4 0.3 0.4 0.4 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.4 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.04
Geo_proximus 0.2 0.3 0.3 0.3 0.4 0.4 0.3 0.4 0.4 0.2 0.3 0.4 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.2 0.2 0.04
He_efasciatus 0.2 0.2 0.2 0.4 0.3 0.4 0.4 0.4 0.4 0.3 0.2 0.4 0.2 0.4 0.3 0.4 0.4 0.4 0.4 0.3 0.4 0.3 0.4 0.3 0.2 0.2 0.2 0.3 0.3
Lae_araguaiae 0.2 0.2 0.2 0.4 0.4 0.4 0.4 0.4 0.5 0.2 0.2 0.4 0.2 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.2 0.2 0.2 0.3 0.3
Mes_festivus 0.2 0.2 0.3 0.4 0.4 0.4 0.5 0.4 0.5 0.3 0.2 0.4 0.2 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.4 0.2 0.2 0.2 0.2 0.3
Ret_lapidifer 0.3 0.3 0.3 0.3 0.4 0.4 0.4 0.4 0.4 0.3 0.2 0.4 0.2 0.3 0.4 0.3 0.4 0.4 0.3 0.4 0.3 0.3 0.3 0.4 0.3 0.2 0.2 0.2 0.3
Ret_xinguensis 0.3 0.3 0.3 0.3 0.4 0.4 0.4 0.4 0.5 0.3 0.2 0.4 0.2 0.4 0.4 0.4 0.4 0.4 0.4 0.4 0.4 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.3
Sat_jurupari 0.2 0.3 0.3 0.4 0.3 0.4 0.4 0.3 0.4 0.3 0.2 0.4 0.2 0.3 0.3 0.4 0.4 0.3 0.3 0.3 0.3 0.3 0.3 0.4 0.2 0.2 0.2 0.2 0.3
Sat_sp 0.3 0.3 0.3 0.3 0.3 0.4 0.4 0.3 0.4 0.3 0.3 0.4 0.2 0.3 0.3 0.4 0.3 0.4 0.4 0.3 0.4 0.3 0.3 0.4 0.2 0.2 0.2 0.2 0.3
Sat_lilith 0.3 0.3 0.2 0.3 0.4 0.4 0.4 0.4 0.4 0.3 0.2 0.4 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.2 0.2 0.2 0.2 0.3
Sym_aequafasciatus 0.2 0.2 0.3 0.4 0.4 0.4 0.4 0.4 0.5 0.3 0.2 0.4 0.2 0.3 0.3 0.3 0.4 0.4 0.4 0.4 0.4 0.3 0.3 0.4 0.2 0.2 0.2 0.2 0.3
Te_sp 0.3 0.3 0.4 0.4 0.4 0.4 0.4 0.4 0.4 0.4 0.3 0.4 0.4 0.2 0.3 0.3 0.4 0.3 0.3 0.4 0.3 0.2 0.2 0.3 0.3 0.3 0.3 0.3 0.3
Te_n_sp_preta 0.3 0.3 0.3 0.4 0.4 0.4 0.4 0.4 0.4 0.3 0.3 0.4 0.3 0.3 0.3 0.2 0.3 0.3 0.3 0.3 0.3 0.3 0.2 0.3 0.3 0.3 0.3 0.3 0.4
Te_cf_gephyrogramma 0.3 0.3 0.3 0.4 0.4 0.4 0.4 0.4 0.4 0.3 0.3 0.4 0.3 0.3 0.3 0.2 0.3 0.3 0.3 0.3 0.3 0.3 0.2 0.3 0.3 0.3 0.3 0.3 0.4
Te_cinderela 0.4 0.4 0.4 0.5 0.5 0.5 0.5 0.5 0.5 0.4 0.4 0.4 0.4 0.3 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.4
Geo
_a
rgys
rost
ictu
s
Geo
_sp
.
Geo
_p
roxi
mu
s
He_
efasc
iatu
s
La
e_a
rag
uaia
e
Mes
_fe
stiv
us
Ret
_la
pid
ifer
Ret
_xi
ngu
ensi
s
Sat_
juru
pa
ri
Sat_
sp
Sat_
lili
th
Sym
_aeq
ua
fasc
iatu
s
Te_
sp
Te_
n_
sp_
pre
ta
Te_
cf_
gep
hyr
og
ram
ma
Te_
cin
der
ela
Geo_argysrostictus
Geo_sp. 0.1
Geo_proximus 0.1 0.05

84
1154
He_efasciatus 0.3 0.3 0.3
Lae_araguaiae 0.3 0.3 0.3 0.2
Mes_festivus 0.3 0.3 0.3 0.2 0.2
Ret_lapidifer 0.3 0.3 0.3 0.3 0.3 0.3
Ret_xinguensis 0.3 0.3 0.3 0.3 0.3 0.3 0.1
Sat_jurupari 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3
Sat_sp 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.3 0.03
Sat_lilith 0.3 0.3 0.3 0.3 0.2 0.3 0.2 0.3 0.2 0.2
Sym_aequafasciatus 0.3 0.3 0.2 0.2 0.2 0.2 0.3 0.3 0.3 0.3 0.3
Te_sp 0.4 0.3 0.3 0.4 0.3 0.3 0.4 0.4 0.3 0.4 0.3 0.4
Te_n_sp_preta 0.4 0.4 0.3 0.3 0.3 0.3 0.3 0.4 0.3 0.3 0.3 0.3 0.2
Te_cf_gephyrogramma 0.4 0.4 0.4 0.3 0.3 0.3 0.4 0.4 0.3 0.3 0.3 0.3 0.2 0.0
Te_cinderela 0.4 0.4 0.4 0.4 0.3 0.4 0.4 0.4 0.4 0.4 0.3 0.4 0.1 0.2 0.2


















