
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIAbdtd.inpa.gov.br/bitstream/tede/1714/5/Dissertação_...
48
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA PROGRAMAS DE PÓS-GRADUAÇÃO DO INPA Programa de Pós-Graduação em Genética, Conservação e Biologia Evolutiva Isolamento e Caracterização de Locos Microssatélites para o vetor da malária Anopheles nuneztovari sensu lato. TATIANA BACRY CARDOZA Dissertação apresentada ao Programa de Pós- Graduação do INPA como parte dos requisitos para obtenção do título de Mestre em Genética, Conservação e Biologia Evolutiva. MANAUS-AM Março, 2011
Transcript of INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIAbdtd.inpa.gov.br/bitstream/tede/1714/5/Dissertação_...

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA
PROGRAMAS DE PÓS-GRADUAÇÃO DO INPA
Programa de Pós-Graduação em Genética, Conservação e Biologia
Evolutiva
Isolamento e Caracterização de Locos Microssatélites para o vetor da malária Anopheles
nuneztovari sensu lato.
TATIANA BACRY CARDOZA
Dissertação apresentada ao Programa de Pós-
Graduação do INPA como parte dos requisitos para
obtenção do título de Mestre em Genética,
Conservação e Biologia Evolutiva.
MANAUS-AM
Março, 2011

ii
TATIANA BACRY CARDOZA
Isolamento e Caracterização de Locos Microssatélites para o vetor da malária Anopheles
nuneztovari sensu lato.
ORIENTADOR(A): Profa. Dra. Vera Margarete Scarpassa
Dissertação apresentada ao Programa de Pós-
Graduação do INPA como parte dos requisitos para
obtenção do título de Mestre em Genética,
Conservação e Biologia Evolutiva.
MANAUS-AM
Março, 2011

iii
Ficha Catalográfica
Cardoza, Tatiana Bacry
Isolamento e caracterização de locos microssatélites para o vetor da malária Anopheles nuneztovari sensu lato. ̸ Tatiana Bacry Cardoza. – Manaus : [s.n.], 2011.
Dissertação (Mestrado em Genética, Conservação e Biologia Evolutiva)—INPA, Manaus, 2011. Orientadora: Dra. Vera Margarete Scarpassa Área de concentração: 1. Anopheles 2. DNA microssatélites 3. Malária
Sinopse: Anopheles nuneztovari é o principal vetor da malária na Colômbia e Venezuela. Na Amazônia brasileira, seu papel na transmissão da malária ainda não está completamente elucidado. Foram isolados e caracterizados 18 locos microssatélites específicos para A. nuneztovari por meio de uma biblioteca genômica enriquecida com regiões microssatélites. Palavras-chave: Malária, microssatélites, Genética evolutiva, Espécies cripticas.

iv
Dedicatória
A Deus;
A minha amada família.

v
Agradecimentos
A Deus, por tornar meus sonhos possíveis e por sempre me surpreender.
Ao Instituto Nacional de Pesquisas da Amazônia (INPA), pela estrutura laboratorial.
Ao programa de pós graduação em Genática, Conservação e Biologia Evolutiva e aos
seus professores por todo apoio e aprendizado.
A Fundação de Amparo a Pesquisa do Estado do Amazonas (FAPEAM) pela bolsa
concedida durante o mestrado.
A FAPEAM-PIPT, CNPq-CT-Amazônia, PROCAD-Amazônia-INPA ̸ UNICAMP
pelo apoio financeiro a este estudo.
A Dra Vera Margarete Scarpassa pela orientação neste trabalho e por todo o
aprendizado durante esses anos.
A Dra Anete Pereira de Sousa, por ter me recebido em seu laboratório, no qual pude
realizar a etapa de genotipagem em gel de poliacrilamida.
A Dra Adna Sousa, por ter me ensinado a técnica de genotipagem em poliacrilamida,
pela amizade, pelo carinho e prestatividade sempre.
A técnica do CBMEG Patricia Zambon, pela disposição e auxílio.
Aos colegas de trabalho Simone Batista, Jéssica Pereira, Ronildo Baiatone, Alex
Sandro e Ledayane e aos ex colegas Joel Cordeiro e Bárbara Alessandra por proporcionarem
um ambiente de trabalho divertido e prazeroso e, por estarem sempre dispostos a ajudar
quando preciso.
A Jéssica e Simone, pelo auxílio durante a criação das amostras em insetário e
trabalho de bancada.
Aos meus pais, Maria do Socorro Bacry e Jorge Alexander Sosa, por incentivarem a
minha educação. Se o conhecimento é parte do que somos, então essa parte devo a vocês.
Amor eterno.
As minhas irmãs, Alessandra e Tayana, e ao meu lindo sobrinho João Davi, agradeço
simplesmente por sempre estarem aqui.
Ao meu querido Mário Said por sempre acreditar na minha capacidade. Agradeço toda
a admiração.
A todos que contribuíram direta ou indiretamente nesta etapa da minha vida.

vi
Resumo
Anopheles nuneztovari é considerada um importante vetor da malária humana no norte e oeste da Colômbia e oeste da Venezuela. Na Amazônia brasileira, embora esta espécie tenha sido encontrada infectada com o parasito da malária, o seu envolvimento na transmissão desta doença é bastante controverso. Estudos genéticos recentes indicam a existência de cinco “linhagens” dentro do táxon A. nuneztovari, que podem diferir quanto à capacidade de transmitir o agente etiológico que causa a malária. Neste sentido, estudos genéticos adicionais com marcadores microssatélites podem auxiliar a esclarecer as relações genéticas entre estas linhagens e proporcionar uma melhor compreensão da dinâmica de transmissão da malária envolvendo esta espécie na Amazônia brasileira. Para isso foram desenvolvidos marcadores microssatélites a partir de uma biblioteca genômica enriquecida com seqüências de CT8 e GT8. A biblioteca genômica foi desenvolvida utilizando amostras procedentes das periferias das cidades de Manaus (Amazonas) e de Nova Mazagão (Amapá). Um total de 192 clones positivos foram sequenciados, dos quais foi possível desenhar 58 pares de primers flanqueadores das regiões de microssatélites. Observou-se microssatélites com diferentes motivos de repetições. Dentre eles, 74,36% foram dinucleotídeos, 15,38% trinucleotídeos, 6,41% tetranucleotídeos e 3,85% pentanucleotídeos, sendo 79,49% microssatélites simples perfeitos, 6,41% simples imperfeitos e 14,01% compostos perfeitos e imperfeitos. Foram sintetizados 41 pares de primers, dos quais 28 amplificaram com sucesso e 18 foram caracterizados em uma amostra de Manaus, com um número amostral que variou de 34 a 39 indivíduos. Todos os locos microssatélites foram polimórficos. O número de alelos por loco variou de 2 a 13. A heterozigosidade observada (HO) variou entre 0,079 e 0,921, enquanto a heterozigosidade esperada (HE) variou entre 0,078 e 0,872. O conteúdo de polimorfismo informativo (PIC) variou entre 0,088 e 0,854. Após a correção de Bonferroni seis locos mostraram-se em desequilíbrio de Hardy-Weinberg. Alem disso, sete locos polimórficos foram testados para amplificação heteróloga em cinco espécies de Anopheles: Anopheles rangeli, Anopheles dunhami, Anopheles konderi, Anopheles oswaldoi sensu lato e Anopheles darlingi. Os sete locos amplificaram com sucesso para todas as espécies, exceto em A. darlingi, que não houve amplificação para nenhum loco. Esses resultados comprovam a eficiência dos marcadores microssatélites para determinar o polimorfismo e futuramente caracterizar, no nível molecular, as linhagens de A. nuneztovari.

vii
Abstract
Anopheles nuneztovari is considered an important vector of human malaria in northern and western Colombia and western Venezuela. In the Brazilian Amazon, although this species has been found infected with the malaria parasite, their involvement in transmission is quite controversial. Recent genetic studies indicate that there are five lineages within the taxon A. nuneztovari, which may differ in their capacity to transmit the etiologic agent that causes malaria. In this sense, additional genetic studies using microsatellite markers may help to clarify the genetic relationships among these strains and provide a better understanding of the dynamics of malaria transmission involving this species in the Brazilian Amazon. Microsatellite markers were developed from a genomic library enriched with sequences CT8 and GT8. The genomic library was developed with samples obtained from the outskirts of the cities of Manaus (Amazonas) and Nova Mazagão (Amapá). A total of 192 positive clones were sequenced and a total of 58 pairs of primers were drawn. Among them, 74.36% were dinucleotide, 15.38% trinucleotide, 6.41% tetranucleotide and 3.85% pentanucleotide, and 79.49% are simple perfect microsatellites, imperfect and mere 6.41% and 14.01% compound perfect and imperfect. From these, 41 primers pairs were synthesized, and 23 were successfully amplified. All 18 microsatellite loci were characterized were polymorphic. The number of alleles per locus ranged from 2 to 13. The observed heterozygosity (Ho) ranged from 0.079 to 0.921, while the expected heterozygosity (He) ranged from 0.078 to 0.872. The polymorphism information content (PIC) ranged from 0.088 to 0.854. We tested seven polymorphic markers for heterologous amplification in five species of Anopheles: Anopheles rangeli, Anopheles dunhami, Anopheles konderi, Anopheles oswaldoi sensu lato and Anopheles darlingi. The seven loci were successfully amplified for all species, except in An. darlingi, that did not amplify any locus. These results demonstrate the efficiency of microsatellite markers to determine polymorphism for future molecular characterization of A. nuneztovari lineages.

viii
SUMÁRIO 1. INTRODUÇÃO .................................................................................................................. 10
1.1 Considerações gerais sobre a malária ............................................................................. 10
1.2 – Considerações sobre o Anopheles nuneztovari ............................................................ 12
1.2.1 – Relação com a Malária ............................................................................................ 14
1.3 Estudos genéticos e morfológicos com Anopheles nuneztovari ..................................... 14
1.4 Marcadores Moleculares Microssatélites ....................................................................... 17
2. OBJETIVOS ....................................................................................................................... 20
2.1 Objetivo Geral ................................................................................................................. 20
2.2 Objetivo Específicos ....................................................................................................... 20
3. MATERIAL E MÉTODOS ............................................................................................... 21
3.1 Obtenção das amostras de Anopheles nuneztovari ......................................................... 21
3.2 Extração e quantificação de DNA.................................................................................. 23
3.3 Construção do banco enriquecido com microssatélites .................................................. 24
3.3.1 Digestão do DNA genômico com enzimas de restrição ............................................ 24
3.3.2 Ligação dos fragmentos a adaptadores ....................................................................... 24
3.3.3 Pré-amplificação, via PCR ........................................................................................... 25
3.3.4 Purificação ................................................................................................................... 25
3.3.5 Seleção de fragmentos contendo as regiões de Microssatélites .................................. 25
3.3.6 Clonagem em vetor ...................................................................................................... 25
3.3.7 Desenho dos primers .................................................................................................. 26
3.4 Validação dos primers ................................................................................................... 27
3.5 Genotipagem ................................................................................................................... 28
3.6 Análise Estatística ........................................................................................................... 31
4. RESULTADOS ................................................................................................................... 32
5. DISCUSSÃO ....................................................................................................................... 36
6. CONCLUSÕES ................................................................................................................... 38
7. REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................. 39

ix
Lista de Figuras
Figura 1: Esquema da dinâmica da transmissão da malária....................................... 10
Figura 2: Distribuição global dos vetores principais e auxiliares da malária............. 11
Figura 3: Mapa da distribuição geográfica do Anopheles nuneztovari...................... 13
Figura 4: Ciclo biológico do Anopheles nuneztovari................................................. 13
Figura 5: Localização do local de coleta no mapa da cidade de Manaus................... 21
Figura 6: Material utilizado para coleta de Anopheles nuneztovari........................... 22
Figura 7: Identificação de Anopheles nuneztovari.................................................... 23
Figura 8: Quantificação do DNA extraído em gel de agarose 1%............................. 24
Figura 9: Programa Primer Select. Possíveis primers e suas características para um
tipo de microssatélite..................................................................................
27
Figura 10: Revelação do gel de poliacrilamida em nitrato de prata............................. 30
Figura 11: Produto de PCR submetido a eletroforese em gel de poliacrilamida......... 30
Figura 12: Genotipagem manual.................................................................................. 31
Figura 13: Frequência dos motivos de repetição dos microssatélites.......................... 32

10
1. INTRODUÇÃO
1.1 Considerações gerais sobre a malária
A malária, também conhecida como maleita ou impaludismo, é uma doença cuja
dinâmica de transmissão envolve um parasito, um vetor e o homem (figura 1). O parasito
causador da malária é um protozoário unicelular do gênero Plasmodium. Quatro espécies de
Plasmodium são causadoras da malária em seres humanos: Plasmodium falciparum,
Plasmodium vivax, Plasmodium malariae e Plasmodium ovale. Destas, apenas a última não
ocorre nas Américas (Forattini, 2002). O homem contrai a doença por meio da picada de
mosquitos fêmeas do gênero Anopheles, quando estas estão infectadas por uma das quatro
espécies de Plasmodium. Essa enfermidade caracteriza-se por acessos de febre de 24, 48 ou
72 horas, provocando danos no fígado e anemia.
Figura 1: Esquema da dinâmica de transmissão da malária. Fonte: Imagens google
As espécies de Anopheles envolvidas na transmissão da malária humana variam de
acordo com a região geográfica (figura 2). Por exemplo, em países da África, os principais
vetores são Anopheles gambiae e Anopheles funestus (ambos pertencentes a complexos de
espécies), na Ásia, os impotantes vetores são Anopheles maculatus, Anopheles minimus,
Anopheles dirus e Anopheles culicifacies, e na Austrália e Nova Guiné, Anopheles
punctulatus (Touré et al., 2004). Considerando as Américas, Anopheles darlingi, Anopheles

11
albimanus, Anopheles nuneztovari, Anopheles pseudopunctipennis e Anopheles aquasalis são
os mais importantes vetores de malária. Contudo, outras espécies podem ter importante papel
na transmissão da malária em nível regional (Touré et al., 2004).
Figura 2: Distribuição global dos vetores principais e auxiliares da malária. Fonte:
http://www.cdc.gov/malaria/about/biology/mosquitoes/map.html
A malária permanece ainda uma das principais causas de morbidade e mortalidade
nas regiões tropicais e subtropicais do mundo e, em particular, nos países pobres. A África,
Ásia e as Américas são os continentes que apresentam a maior incidência desta doença.
Estima-se que entre 350 e 500 milhões de pessoas são infectadas anualmente em todo o
mundo. Destas, cerca de um a três milhões morrem, principalmente na África, onde as
maiores vítimas são crianças com idade menor que cinco anos (Roll Back Malaria, 2005). Nas
Américas, a malária é transmitida em 22 países, onde é estimado que aproximadamente 137
milhões de pessoas estejam sob o risco de contrair esta doença (Roll Back Malaria, 2010). O
Brasil é o maior responsável pelo número de casos, com 50,7% do total de registros,
correspondente a cerca de 300 mil casos por ano. Destes, aproximadamente 99,5% dos casos
são registrados na Amazônia Legal, constituída pelos Estados do Acre, Rondônia, Amazonas,

12
Roraima, Amapá, Pará, Maranhão, Mato Grosso e Tocantins (OPAS, 2006; SIVEP-Malária,
2009; Ministério da Saúde, 2010).
No Brasil, depois de um incremento importante no número de casos nos anos 1998 e
1999 (635.646 casos em 1999), houve uma redução muito expressiva durante os dois anos
subsequentes (348.259 casos em 2002) quando foi implementado o Plano de Intensificação
das Ações de Controle da Malária. Porém, a partir de 2003 houve novamente um crescimento
no número de casos atingindo um pico de 603.532 casos em 2005. No ano seguinte, registrou-
se novamente uma redução importante no número de casos e, em 2007 foi registrado uma
incidência de aproximadamente 458 mil casos, passando para 315 mil casos em 2008 e 306
mil casos em 2009 (SVS, 2010).
Na Amazônia brasileira, só em 2009, foram resgistrados mais de 105 mil casos de
malária e em 2010 mais de 132 mil casos, sendo os Estados mais afetados o Amazonas e o
Pará, este último ultrapassando 50 mil casos (SVS, 2010). Este elevado números de casos se
deve a fatores ambientais que favorescem o desenvolvimento dos mosquitos e à manutenção
do ciclo de transmissão da doença. Os padrões de ocupação do espaço, o uso da terra e as
variações climáticas são, por fim, fatores decisivos no comportamento desta enfermidade.
No entanto, apesar da redução geral no número de casos, a incidência da malária ainda
permanece elevada na periferia de muitas cidades da região amazônica, decorrente destes
fatores ambientais e também de fatores sociais, como: migrações das populações humanas,
assentamentos desordenados, instalações de moradias precárias, entre outros fatores,
favorescendo o contato entre os vetores e as populações humanas (Tadei et al., 1998;
Susam/Devis, 2004; Scarpassa e Conn, 2007).
1.2 – Considerações sobre o Anopheles nuneztovari
Anopheles nuneztovari Gabaldón, 1940 apresenta ampla distribuição geográfica no
norte da América do Sul, incluindo Bolívia, Brasil, Colômbia, Equador, Guiana, Guiana
Francesa, Suriname e Venezuela, e parte do Panamá, na América Central (Faran, 1980;
Consoli e Lourenço de Oliveira, 1994). No Brasil, é encontrada nos Estados da região
amazônica ( Scarpassa 1996) (figura 3).

13
Figura 3: Mapa da distribuição geográfica do A. nuneztovari (ícones em vermelho). Fonte: Google Maps
Esta espécie está classificada na Ordem Diptera, Família Culicidae, Subfamília
Anophelinae; Gênero Anopheles e Subgênero Nyssorhynchus. Este subgênero constitui um
grupo de anofelinos que representa o maior número de vetores principais ou auxiliares da
malária humana na região Neotropical (Forattini, 2002).
O ciclo biológico ovo-adulto desta espécie é relativamente rápido, em condições de
temperatura de 26 a 28º C, com uma média de 12,02 ± 0,004 dias (Scarpassa e Tadei, 1990).
Apresentam metamorfose completa, ou seja, são holometábulos, sendo seu ciclo de vida
dividido em quatro estágios: ovo, larva, pupa e adulto (figura 4).
Figura 4: Ciclo biológico do A. nuneztovari. Estágios de Ovo, larva, pupa e adulto. Fonte: Imagens Google

14
1.2.1 – Relação com a Malária
A. nuneztovari é considerado um importante vetor da malária humana no norte e oeste
da Colômbia e oeste da Venezuela, apresentando graus elevados de antropofilia (Gabaldón,
1981). Rey e Renjifo (1950) foram os primeiros autores a descrever infecção por Plasmodium
sp. para A. nuneztovari, com a presença de oocistos no estômago, em áreas de Cucutá (norte
da Colômbia). Posteriormente, muitos outros estudos entomológicos, de dinâmica de
transmissão e taxas de infecção foram realizados com populações desta espécie em diferentes
localidades da Colômbia e Venezuela, e os resultados destes estudos confirmaram o
envolvimento desta espécie na transmissão da malária humana naquelas localidades
(Gabaldón, 1981; Fajardo e Alzate, 1987; Rubio-Palis e Curtis, 1992; Gautret et al., 1995;
Scarpassa et al., 1999; Scarpassa et al., 2000). Na Amazônia brasileira, no entanto, A.
nuneztovari apresenta um comportamento predominantemente zoofílico e seu envolvimento
na transmissão da malária não está ainda esclarecido, embora vários estudos descreveram esta
espécie infectada com Plasmodium vivax, P. falciparum e P. malarie. A. nuneztovari foi
encontrada infectada com o parasito da malária nos Estados do Amazonas, Amapá, Pará e
Roraima, por meio da técnica de ELISA e IRMA (Arruda et al., 1986; Tadei et al., 1998;
Tadei e Thatcher, 2000; Póvoa et al., 2001; Silva-Vasconcelos et al., 2002). Mais
recentemente, Galardo et al., (2007) relataram infecção para A. nuneztovari com Plasmodium
vivax, variante VK210, P. vivax variante VK247 e P. malariae em comunidades rurais do Rio
Matapí (Estado do Amapá). Estes autores, com base nos resultados de infecção, densidade e
outros parâmetros entomológicos, consideraram A. nuneztovari um importante vetor,
conjuntamente com A. darlingi e A. marajoara naquelas localidades.
1.3 Estudos genéticos e morfológicos com Anopheles nuneztovari
Considerando o envolvimento de A. nuneztovari na transmissão da malária, vários
estudos comportamentais, genéticos e morfológicos foram realizados com populações
procedentes da Colômbia, Venezuela e Brasil (Amazônia brasileira) (Elliott, 1968; 1972;
Kitzmiller et al., 1973; Conn et al., 1993; Scarpassa et al., 1996; 1999; 2000; Hribar, 1994;
Linley et al., 1996; Fritz et al., 1994; Onyabe & Conn, 1999; Conn et al., 1998; ; Mirabello e
Conn, 2008) com o objetivo de compreender o status taxonômico e historia evolutiva e,
consequentemente, a dinâmica de transmissão da malária causada por este vetor.

15
Elliott (1968; 1972) sugeriu duas formas alopátricas para A. nuneztovari, com base nos
padrões comportamentais. Populações do oeste da Venezuela e norte da Colômbia apresentam
antropofilia e endofagia acentuadas, com um pico de atividade hematofágica em torno da
meia-noite e foram consideradas importantes vetores de malária. Por outro lado, populações
de outras localidades, incluindo o Brasil, apresentam zoofilia e exofilia marcantes, picam nas
primeiras horas da noite e não foram consideradas vetores de malária. Subsequentemente,
populações consideradas vetoras e não vetoras foram analisadas para o polimorfismo de
inversões paracêntricas dos cromossomos politênicos (Kitzmiller et al., 1973). As populações
de Tibú (norte da Colômbia) e de Barinas (oeste da Venezuela) apresentaram uma inversão
homozigota (fixa) no cromossomo X (braço R) e ausência deste rearranjo em populações de
Manaus e imediações (Brasil), sendo está ultima considerada, pelos autores, como arranjo
padrão. Posteriormente, outras análises citogenéticas propuseram a existência de três
citótipos. O citótipo A, considerado padrão, encontra-se distribuído por toda a Amazônia
brasileira; o citótipo B, além da presença da inversão no cromossomo X, apresentou uma
inversão polimórfica (2La) e ocorre na Venezuela, ao sudeste da Cordilheira dos Andes; o
citótipo C, que além da inversão no cromossomo X, contém um nítido cromocentro e um
complexo de inversões, constituído por duas inversões menores (2Lc e 2Ld), incluídas em
uma inversão maior denominada de 2Lb. Este último é encontrado no oeste da Colômbia e no
oeste da Venezuela, ao oeste dos Andes (Conn et al., 1993).
Fritz et al. (1994) compararam as seqüências do ITS2 do DNA ribossomal de nove
amostras de A. nuneztovari procedentes do Brasil, Bolívia, Suriname, Venezuela e Colômbia
e observaram que os espécimes da Colômbia e Venezuela tem sequências idênticas, mas
diferentes daquelas da Amazônia brasileira. Estes dados sugeriram que as populações de A.
nuneztovari da Colômbia/Venezuela e Brasil podem ser dintinguidas por diferenças mínimas
na sequência de DNA desta regiões. Onyabe e Conn (1999) ao estudarem o mesmo marcador
em amostras do Brasil, Colômbia e Venezuela, também observaram diferenças entre as
populações destas regiões. Os autores sugeriram ser consistente com pelo menos duas
espécies crípticas, uma na Venezuela/Colômbia e outra no Brasil, suportanto os resultados de
Fritz et al. (1994).
Scarpassa et al. (1999) ao analisarem seis populações de A. nuneztovari procedentes
da Colômbia e da Amazônia brasileira, por meio de isoenzimas, encontraram pouca
estruturação genética e diferenciação geográfica nas populações da Amazônia brasileira,
sugerindo que as mesmas podem ser recentes. Por outro lado, quando estas populações foram
comparadas com as da Colômbia, os resultados indicaram elevada diferenciação genética,

16
cujos valores estavam dentro dos limites observados para membros de complexos de
espécies.
Com relação ao DNA mitocondrial, Conn et al. (1998) analisaram 12 populações de A.
nuneztovari, procedentes de 5 países, e os resulados revelaram a presença de três linhagens
distintas, uma na Venezuela/Colômbia e duas dentro da bacia amazônica. A análise de
variância molecular e estatística F demonstraram significativa subdivisão e corroboraram
fortemente com a existência de múltiplas linhagens. Scarpassa et al. (2000), ao examinar o
DNA mitocondrial por meio de endonucleases de restrição, de três populações procedentes da
Amazônia brasileira e uma da Colômbia, encontraram elevado valor de divergência entre as
amostras dessas localidades, sugerindo que estas populações geográficas podem
eventualmente constituir espécies separadas.
Posso et al. (2003), analisaram três populações de A. nuneztovari da Colômbia, por
meio da técnica de RAPD, e encontraram uma elevada similaridade genética entre estas
populações que pode estar sendo causada por um grande fluxo gênico de indivíduos entre as
mesmas. Subsequentemente, Sierra et al. (2004) analisaram a região ITS2 de amostras da
Colômbia, em ambos os lados dos Andes, que correspondem aos citótipos B e C e,
compararam seus resultados com os obtidos por Fritz et al. (1994). A combinação destes
dados mostrou pouca divergência genética e ambos os citótipos foram considerados como co-
específicos.
Mirabello e Conn (2008) utilizaram o gene white do DNA nuclear para analisar a
estrutura populacional do A. nuneztovari de 13 localidades da Colômbia, Venezuela,
Suriname, Brasil e Bolívia. Os resultados indicaram cinco linhagens distintas, com elevada
diferenciação genética entre elas. Os autores hipotetizaram que esta diversificação é o
resultado de uma interação da invasão do mar durante o Mioceno/Plioceno e das mudanças
climáticas durante o Pleistoceno. As localidades em que estas linhagens são simpátricas, os
autores interpretaram, como zonas de contato secundário.
Diferenças morfológicas na genitália masculina são amplamente utilizadas para
identificar espécies do subgênero Nyssorhynchus. Hribar (1994) propôs estudar se estas
diferenças podiam ser usadas para distinguir os três citótipos de A. nuneztovari. Com base nas
análises de quatrocentas genitálias masculinas procedentes do Brasil e Suriname (citótipo A),
Venezuela (citótipo B) e Venezuela e Colômbia (citótipo C) foi possível diferenciar o citótipo
B dos citótipos A e C, por meio de um conjunto de características da genitália, sendo que as
diferenças mais óbvias foram no comprimento e no grau de esclerotização dos espículos do
mesósoma. Mais recentemente, Bergo et al. (2007), por meio da análise preliminar da

17
morfologia do ápice do mesósoma, propuseram que amostras analisadas do Estado do Amapá
pode ser espécie distinta, sendo provavelmente o A. goeldii, que foi previamente considerada
um sinônimo do A. nuneztovari (Lane, 1953 apud Bergo et al. 2007). Subsequentemente,
Calado et al. (2008) utilizaram além da morfologia da genitália masculina, seqüências do
ITS2 do DNA ribossomal, gene COI do DNA mitocondrial e gene white do DNA nuclear de
amostras dos Estados do Amapá, Pará e Amazonas (Brasil), além da comparação com
seqüências de estudos anteriores, para testar a hipótese de que A. goeldii é uma espécie válida.
Com base nos resultados das análises morfológicas e moleculares, os autores consideraram A.
goeldii como espécie válida e distinta de A. nuneztovari.
Recentemente, Scarpassa & Conn (2011) sequenciaram um fragmento do gene COI
em amostras de 17 localidades abrangendo toda a distribuição geográfica. Os resultados
evidenciaram monofilia para a espécie, com quatro subclados distribuídos em dois clados
maiores. O subclade basal (II-C) encontrado nas amostras da Colômbia/Venezuela/Bolivia é
provavelmente A. nuneztovari s. s., enquanto os outros três [clado (I) e subclados (II-A, II-B)]
provavelmente representam linhagens distintas ou espécies, sendo que uma delas pode
representar A. goeldii. A diversificação e a expansão destes clados provavelmente ocorreram
no Pleistoceno.
1.4 Marcadores Moleculares Microssatélites
O DNA satélite foi descoberto na década de 1960 quando amostras de DNA genômico
foram submetidas a centrifugação com cloreto de césio. Nesta análise, observou-se bandas
distintas, sendo que uma delas continha genes e a outra, localizada numa posição superior, o
DNA satélite, que apresentava repetições contendo de dois a milhares de pares de bases (pb),
localizadas principalmente no centrômero (Tautz, 1989). Estas repetições foram subdivididas
em dois grupos: os minissatélites, com repetições constituídas de 9 a 100 pb e os
microssatélites, de 1 a 6 pb (Weber, 1990).
Os microssatélites, também denominados de SSR (Simple Sequence Repeats) ou STR
(Short Tandem Repeats), são regiões repetitivas não codificadoras de DNA, compostas de um
a seis nucleotídeos repetidos em tandem, distribuídas no genoma de eucariotos e procariotos
(Li et al., 2004). Os microssatélites são classificados de acordo com o tipo de repetição em:
perfeitos (seqüência sem nenhuma interrupção), imperfeitos (seqüência interrompidas por
bases que não pertencem ao motivo de repetição do microssatélite), interrompidos (quando
ocorre uma pequena seqüência dentro do microssatélite distinta da sequência do mesmo) e

18
compostos (duas ou mais repetições com motivos diferentes dispostas de forma adjacente)
(Oliveira et al., 2006).
As sequências de DNA que flanqueiam os microssatélites são geralmente conservadas
entre os indivíduos de uma mesma espécie, permitindo seleção de primers específicos que
amplificam, via PCR, fragmentos contendo o DNA repetitivo. Os produtos da amplificação
podem ser visualizados em gel de agarose ou poliacrilamida, ou por meio da utilização de
primers fluorescentes e então analisados no analisador de DNA (Borém e Caixeta, 2006).
Estes marcadores estão substituindo rapidamente outros marcadores em vários tipos de
estudos genéticos, principalmente devido a sua reprodutibilidade e simplicidade técnica, à
pequena quantidade de DNA requerida, ao baixo custo, ao grande poder de resolução e aos
altos níveis de polimorfismo. Além dessas características, os microssatélites são co-
dominantes e parecem ter distribuição frequente e aleatória, permitindo uma cobertura ampla
do genoma (Borém e Caixeta, 2006).
A origem, o polimorfismo e a evolução dos microssatélites podem ser explicados por
meio de dois modelos: o“crossing-over” desigual e o “DNA slippage” (Eisen, 1999). O
primeiro resulta da recombinação entre cromossomos homólogos, os quais não são alinhados
corretamente, resultando em deleções ou expansões (Eisen, 1999; Mello et al., 2002). O
segundo processo podeser decorrente de escorregões da DNA polimerase que ocorre durante a
replicação do DNA, quando esta avança ou recua em uma região repetitiva, diminuindo ou
aumentando o número de repetições, sendo este o mecanismo mais aceito para explicar o
polimorfismo em microssatélites. Muitos microssatélites tem uma alta taxa de mutação, o que
gera altos níveis de diversidade alélica, muito úteis para estudos de genética de
populações(Schlötterer, 2000).
Os microssatélites são utilizados para inferir testes de paternidade, grau de parentesco,
estrutura genética de populações, fluxo gênico, tamanho efetivo de população (Oliveira et al.,
2006), além de mapeamento genético, diagnóstico de doenças, investigação forense, estudos
ecológicos e biologia da conservação (Borém e Caixeta, 2006), para avaliar o impacto do
sistema reprodutivo, estrutura social e dispersão em populações em extinção (Diniz et al.,
2007).
Devido a estas características, estes marcadores vem sendo desenvolvidos e
amplamente utilizados em genética de populações de mosquitos vetores para avaliar a
estrutura genética, os níveis de fluxo gênico entre as populações e ajudar no esclarecimento
do status taxonômico de espécies de mosquitos (Mirabello et al., 2008; Silva et al., 2008;
Midega et al., 2010). O trabalho pioneiro de desenvolvimentos deste marcador para espécies

19
de Anopheles foi realizado por Zheng et al (1993), a fim de construir um mapa genético para
A. gambie, no qual isolaram 24 marcadores microssatélites. Muitos outros trabalhos com este
marcador foram feitos para espécies de Anopheles, com por exemplo: A. gambiae (Lehman et
al., 1996, 1997), A. albimanus (Molina-Cruz et al., 2004), A. arabiensis (Temu & Yan,
2005), A. darlingi (Conn et al., 2006; Scarpassa & Conn, 2007; Mirabello et al., 2008), A.
funestus (Michel et al., 2006), A. moucheti (Antonio-Nkondjio et al., 2008); e, também em
outras espécies de mosquito como em Aedes aegypti (Chambers et al., 2007).
1.4.1 Isolamento e caracterização de Marcadores Microssatélites em espécies de Anopheles.
Devido as características dos microssatélites, os mesmos vem sendo isolados e
caracterizados para diversas espécies de Anopheles e, amplamente utilizados em genética de
populações de mosquitos vetores para avaliar a estrutura genética, níveis de fluxo gênico entre
populações e ajudar no esclarecimento do status taxonômico de espécies de mosquitos. Para o
vetor da malária A. funetus foram primeiramente caracterizados 22 locos microssatelites
(Sinkins et al., 2000) e, em 2002, Couhet et al. isolaram e caracterizaram mais 18 locos que
juntamente com os primeiros auxiliariam na investigação da distribuição da variabilidade,
dentro e entre populações deste vetor.
Para o vetor da malária na Índia, Paquistão, Iran e Afeganistão, A. stephensi, devido a
variação morfológica e cromossomal descrita para populações rurais e urbanas 16 locos
microssatélites foram desenvolvidos para auxiliar na definição da estrutura populacional e
para compreender a importância epidemiológica deste mosquito (Verardi et al., 2002). Outro
importante vetor da Índia, A. culicifacies, faz parte de um complexo de espécies, no qual
existem cinco formas isomórficas, designadas de A, B, C, D e E, no qual o tipo A é
amplamente distribuído e responsável pela malária causada pelo Plasmodium falciparum e P.
vivax. Para examinar o fluxo gênico e entender a estrutura populacional deste vetor foram
isolados e caracterizados 31 locos microssatélites (Sunil et al., 2004).
Para o A. marajoara e A. darlingi, importantes vetores na Amazônia (Brasil), foram
isolados e caracterizados 11 locos microssatélites para a primeira espécie (Li et al., 2005) e 32
locos microssatélites para a segunda, sendo 8 desenvolvidos em 2001 por Conn e
colaboradores e 24 em 2010 por Lima e colaboradores.

20
2. OBJETIVOS
2.1 Objetivo Geral
Isolar e caracterizar marcadores moleculares microssatélites para A. nuneztovari da
Amazônia brasileira.
2.2 Objetivo Específicos
• Construir duas bibliotecas genômicas enriquecidas com regiões de DNA
microsatélites de A. nuneztovari;
• Gerar a sequência nucleotídica dos clones recombinantes das bibliotecas genômicas;
• Desenhar primers a partir das seqüências que possuírem as regiões microssatélites;
• Validar os primers microssatélites para A. nuneztovari e verificar os locos
polimórficos que possam ser utilizados como marcadores em estudos de genética de
populações;
• Verificar o alcance da Amplificação heteróloga em cinco espécies do gênero
Anopheles.

21
3. MATERIAL E MÉTODOS
3.1 Obtenção das amostras de Anopheles nuneztovari
Para o desenvolvimento das bibliotecas enriquecidas com regiões microssatélites, as
amostras de A. nuneztovari foram procedentes dos Estados do Amazonas, município de
Manaus (03º 4ʹ S , 59 º 51ʹ W) (figura 5A), e Amapá, município de Nova Mazagão (00° 07’ S,
51° 17’ W) (figura 5B).
Figura 5 : Localização dos locais de coleta. A: Sitio Santa Vitória, Bairro Puraquequara (Manaus, Amazonas); B:
Em vermelho, Nova Mazagão (Amapá).
Os mosquitos foram coletados no período noturno, entre 18:00 e 21:00 horas, com
auxílio de um capturador manual e imediatamente transferidos para copos de papelão
parafinados. Em seguida, o material foi transportado em isopor fechado para o Instituto
A
B

22
Nacional de Pesquisas da Amazônia (INPA) e, então, os mosquitos (fêmeas) separados
individualmente em copos de oviposição, previamente montados, para obtenção das posturas
(figura 6).
Figura 6: Material utilizado para coleta de A. nuneztovari. A: capturadores manuais; B: copos de papelão
parafinados contendo espécimes coletados; C: copos de plásticos para oviposição, contendo uma fêmea em cada.
Após a oviposição, as fêmeas que ovipuseram e seus ovos foram identificados sob
Microscópio estereoscópico e Microscópio Óptico, com aumento de 32X e 400X,
respectivamente, seguindo-se as chaves de Gorham et al. (1967) e Cova-Garcia (1961) (figura
7). Algumas das características utilizadas na identificação foram: presença de flutuadores
abertos e sem coroa (para os ovos) e padrão de coloração da asa onde a pré- humeral, de cor
escura, está na mesma proporção que a humeral, de cor clara; e a presença de um anel de cor
escura no quinto segmento do terceiro par de pernas (fêmea adulta). Posteriormente, os ovos
foram transferidos para copos de eclosão e, em seguida, as larvas recém eclodidas foram
mantidas em bandejas para completar o desenvolvimento até o estágio de pupa, conforme o
método de Scarpassa e Tadei (1990). Após o estágio pupal, foram transferidas para copos
fechados com filó, no qual continha água destilada, para a emergência dos adultos. Os
espécimes para análises foram então acondicionados em microtubos eppendorfs, que foram
etiquetados e congelados em freezer -80° C, onde permaneceram até o momento das
extrações do DNA genômico.

23
Figura 7: Identificação de A. nuneztovari. Parte superior: Identificação de ovos (A-D) sob microssópio óptico A:
aumento de 20X; B: aumento de 40X; C: aumento de 100X; D: aumento de 400X. Parte inferior: Identificação
do adulto fêmea (E-G) sob microscópio esteroscópico. E: adulto fêmea; F: Asa do adulto fêmea; G: perna adulto
fêmea.
3.2 Extração e quantificação de DNA
Para a construção das bibliotecas, o DNA genômico foi extraído de um pool de 7 a 8
larvas, procedentes dos Estados do Amazonas e Amapá, utilizando o protocolo de extração
com fenol e clorofórmio, conforme método de Sambrook e Russell (2001). No final da
extração, o DNA foi ressuspendido em tampão TE 1X.
O DNA foi quantificado em eletroforese de gel de agarose a 1%, por comparação com
o marcador de peso molecular conhecido O corante fluorescente utilizado foi o GelRed
(Biotium), estando o mesmo junto com o azul de bromofenol, sendo ambos aplicados
conjuntamente com a amostra de DNA. Em seguida o gel foi fotodocumentado (figura 8).

24
Figura 8: Quantificação do DNA extraído em gel de agarose 1%. MM: Marcador Molecular; 25-47: amostras de
DNA.
3.3 Construção do banco enriquecido com regiões microssatélites de A. nuneztovari.
Para o desenvolvimento das duas bibliotecas enriquecidas com marcadores
microssatélites foi necessário realizar uma série de procedimentos, os quais serão
suscintamente descritos a seguir. A metodologia empregada foi confome as descrições de
Billotti et al. (1999).
3.3.1 Digestão do DNA genômico com enzimas de restrição
Inicialmente, o DNA genômico, previamente quantificado, foi digerido com a enzima
de restrição Rsa I (procedente de Rhodopseudomonas sphaeroides), para a obtenção de
fragmentos de tamanhos adequados. Este procedimento ocorreu a temperatura de 37°C
overnight. Posteriormente, a digestão foi checada em gel de agarose e fotodocumentada.
3.3.2 Ligação dos fragmentos a adaptadores
A digestão do DNA genômico gerou fragmentos de diversos tamanhos, mas com as
mesmas sequências terminais complementares, as quais foram ligadas aos adaptadores Rsa21
e Rsa25, contendo as seguintes sequências 5’CTCTTGCTTACGCGTGGACTA3’ e 5’
TAGTCCACGCGTAAGCAAGAGCACA 3’, respectivamente. Esta ligação possibilitou que
todos os fragmentos digeridos pudessem ter uma terminação comum e conhecida. A reação

25
preparada continha água milliQ, tampão, adaptadores, DNA ligase e o DNA digerido. Esta
reação foi incubada em termociclador a temperatura de 20°C, durante 2 horas.
3.3.3 Pré-amplificação, via PCR
Nesta etapa foi realizada uma reação de PCR para amplificar os fragmentos e garantir
que a ligação do DNA aos adaptadores tenha ocorrido. A reação foi preparada com água
milliQ, tampão, cloreto de magnésio, dNTP, Rsa21, Taq DNA polimerase e o DNA ligado
aos adaptadores.
3.3.4 Purificação
A purificação foi realizada com o emprego do kit “Quiaquick PCR purification”
(Qiagen), conforme as recomendações do fabricante, com o objetivo de preparar a seleção
dos fragmentos de interesse.
3.3.5 Seleção de fragmentos contendo as regiões de Microssatélites
Esta etapa foi realizada através de um processo denominado de hibridização, o qual
envolveu a ligação de sondas marcadas com biotina à esferas magnéticas recobertas por
estreptavidina. As sequências de microssatélites foram ligadas às sondas e à esferas
magnéticas e a seleção dos fregmentos se deu com o uso de um ímã. Após esta etapa de
hibridização ocorreram várias lavagens do DNA para remoção de fregmentos não específicos
e, em seguida, os fragmentos selecionados foram amplificados através da técnica de PCR.
3.3.6 Clonagem em vetor
Para esta etapa foi utilizado como vetor o plasmídeo pGEM-T (Promega, Madison,
WI), aos quais os fragmentos amplificados via PCR (inserto) foram ligados com o auxílio da
DNA ligase. A reação foi incubada a temperatura de 4°C, overnight, gerando moléculas
“híbridas” de DNA contendo o plasmídeo ligado ao inserto (DNA a ser clonado).
Em seguida estas moléculas “híbridas” (vetor+inserto) foram incorporadas por
bactérias hospedeiras de Echerichia coli XL1 Blue através de um choque térmico que tornou
estas bactérias temporariamente permeáveis para estas moléculas híbridas . Esta técnica

26
proporcionou a amplificação do inserto. Uma vez transformadas se deu a seleção das colônias
recombinantes, ou seja, aquelas que incorporaram o plasmídeo ligado ao inserto. Para isso
uma solução foi plaqueada num meio de cultura contendo ampicilina, IPTG (isopropyl-beta-
D-thiogalactopyranoside) e X-Gal (5-bromo-4-chloro-3-indolyl-b-D-galactopyranoside) e,
transportada para a estufa a 37ºC, para o crescimento das colônias por aproximadamente 20
horas. Uma vez comprovado crescimento das colônias, estas foram transferidas para a
geladeira para posterior seleção das colônias de coloração branca (bactérias que carregavam o
plasmideo recombinante). Em seguida, foram repicadas e novamente colocadas em meio de
cultura contendo 2YT-HMFM e ampicilina para crescimento e manutenção dos clones. Após
esta etapa foi feita a inoculação e extração do DNA plasmidial (Sambrook e Russel, 2001)
com objetivo de isolar o DNA plasmidial das colônias recombinantes para reação de
sequenciamento dos clones e obtenção das sequências.
3.3.7 Preparo das Reações de Sequenciamento
Nesta etapa, o inserto foi amplificado por meio de uma reação contendo: água estéril,
tampão save money, primer, DNA do plasmídeo e Big Dye. Em seguida as amostras foram
purificadas, a fim de eliminar possíveis interferentes ou excessos de reagentes, com o uso de
isopropanol. Posteriormente, as amostras de ambas as bibliotecas (Amazonas e Amapá) foram
sequenciadas utilizando-se os primers T7 e SP6, juntamente com o kit Big Dye e
eletroinjetadas no Analisador de DNA, ABI 3130 (Applied Biosystems) para obtenção das
sequências.
3.3.8 Desenho dos primers,
No desenho dos primers foram utilizados os programas MICROSAT (CIRAD) e o
pacote de programas LASER GENE versão 5.03 (DNAstar Inc.), nos quais as sequências
geradas pelo sequenciador automático foram editadas. Inicialmente os adaptadores foram
removidos e, então, separadas as sequências que apresentaram tamanhos acima de 150 pares
de bases (pb). Foi realizada uma busca dos microssatélites em cada sequência. Nestas, os
microssatélites foram localizados, assim como os possíveis primers nas regiões flanqueadoras
(figura 9). Adotou-se como critérios para desenho dos primers as seguintes características:
distância mínima de 10 pb dos microssatélites tanto para o primer forward como o primer
reverso; diferença de temperatura de anelamento menor que 3°C para ambos os primers;

27
ausência de formação dos diversos tipos de dímeros entre os primers; porcentagem de guanina
e citosina na sequência acima de 40%; e comprimento do fragmento entre 150 e 300 pb. Estas
informações foram fornecidas pelo programa Primer Select (LASER GENE), o qual lista
inúmeras possibilidades de primers flanqueadores para um dado microssatélite (figura 9).
Os primers desenhados receberam nomeação de acordo com a localidade a qual
pertenciam, sendo estas AN e ANU, a primeira para amostras do Amapá e a segunda para
amostras do Amazonas.
Figura 9: Programa Primer Select. Possíveis primers (verde e vermelho) e suas caracteristicas para um tipo de
microssatélite.
3.4 Validação dos primers
Após o desenho dos primers as sequências nucleotídicas de ambas as bibliotecas
foram encaminhadas para síntese. Os primers sintetizados foram ressuspendidos conforme o
protocolo do fabricante e os testes de amplificação realizados a fim de otimizar as condições
ideais de amplificação, tomando como referência as temperaturas de anelamento fornecidas
pelo programa PRIMER SELECT do pacote LASER GENE (DNAstar Inc.).
Em seguida estes primers foram validados por meio das análises da amostra de
Anopheles nuneztovari obtida no bairro do Puraquequara, Manaus (AM). Inicialmente, as
reações de PCR foram preparadas como seguem: tampão (10mM Tris-Cl, 25mM KCl, ph
8,4), 1,4mM de cloreto de magnésio, 0,5 µM dos primers, 0,4mM de cada dNTPs, 2-5ng de
DNA genômico, 0,01U/µl de Taq DNA polimerase (Invitrogen) e água estéril, totalizando o

28
volume de 25μl. As reações de PCR foram levadas ao termociclador utilizando programas
específicos adaptados de Scarpassa e Conn (2007) e com temperaturas de anelamento
previamente padronizadas para cada loco (Tabela 1). Os produtos de PCR foram submetidos
a eletroforese em géis de agarose e fotodocumentados, a fim de verificar a amplificação para
genotipagem.
Tabela 1: Ciclagem da reação de PCR.
Passo Temperatura Tempo
1 95°C 5 minutos
2 95°C 20 segundos
3 TA 30 segundos 30 ciclos
4 72°C 20 segundos
6 72°C 10 minutos
TA: Temperatura de anelamento específica para cada loco.
Alguns dos primers validados foram escolhidos para teste de amplificação heteróloga
em outras cinco espécies de Anopheles: A. rangeli, A. dunhami, A. konderi, A. oswaldoi senso
lato e A. darlingi.
3.5 Genotipagem dos locos microssatélites em Gel de Poliacrilamida
Esta análise foi realizada em gel de poliacrilamida de acordo com metodologia
descrita em Creste et al. (2001), e revelado com nitrato de prata, conforme metodologia
adaptada de Beidler et al. (1982) (Beidler et al., 1982 apud Creste et al., 2001).
O gel de poliacrilamida foi polimerizado entre duas placas de vidro com tamanhos
diferentes, que foram limpas antes do preparo do gel, com álcool comum e acetona. A placa
de maior tamanho no qual um termômetro encontra-se aderido foi tratada com solução “repel”
(dimethyldichlorosilane in octamethyl cyclo-octasilane) evitando assim que o gel ficasse
aderido. O vidro menor foi tratado com solução “bind” (ɣ-methacryloxy-propyl-
trimethoxysilane), para a aderência do gel .
O gel foi preparado com tampão TBE 1X, poliacrilamida a 6%, persulfato de amônio e
TEMED. A solução do gel foi vertida lentamente sobre a placa de vidro, de forma contínua
para que não se formassem bolhas. Após a polimerização do gel, as placas foram colocadas na

29
cuba, na qual foi acrescido o tampão de corrida. A fonte de eletroforese foi ligada, e a placa
submetida a uma pré-corrida a 150W, até atingir temperatura entre 49°C e 51°C.
Enquanto a placa estava sendo aquecida, as amostras (produtos de PCR) juntamente
com formamida foram desnaturadas em termociclador a temperatura de 95°C, durante 5
minutos. Ao término deste tempo as amostras foram submetidas a um choque térmico (com
gelo). Em seguida, foram aplicadas no gel e a corrida eletroforética realizada a 120W por
aproximadamente duas horas.
Após a eletroforese, as placas de vidro foram separadas cuidadosamente e o gel
revelado com nitrato de prata. Este procedimento passou por dez passos que podem ser
observados no quadro abaixo e na figura 10.
1. O gel foi colocado em solução fixadora (1000 mL de solução, sendo 10% de etanol e 1% de
ácido acético), por no mínimo 10 minutos.
2. Lavado em água destilada.
3. Colocado em ácido nítrico a 1,5% por 3 minutos.
4. Lavado em água destilada.
5. O gel foi mergulhado em nitrato de prata a 0,2% por 20 minutos, com agitação constante
em shaker.
6. Lavado em água destilada.
7. Mergulhado na solução contendo carbonato de sódio e formaldeído a 37%, até a revelação
das bandas.
8. Mergulhado o gel na solução STOP (1000mL de ácido acético a 5%), por 5 minutos
9. Lavado em água destilada.
10. O gel foi colocado para secar.

30
Figura 10: Revelação do gel de poliacrilamida em nitrato de prata. Etapa realizada no CBMEG em Campinas
(SP). A: Passos de 1 a 4; B: Passos 5 e 6; C: Passo 7; D: Passos 8 e 9; E: Passo 10.
Após verificado que o gel estava seco, o mesmo foi escaneado com o auxílio de um
scanner estando o gel ainda aderido na placa de vidro (figura 11) e, em seguida foi realizado a
genotipagem manual das amostras para os locos analisados.
Figura 11: Produto de PCR submetido a eletroforese em gel de poliacrilamida. MM: Marcador Molecular.
3.6 Genotipagem Manual
Uma vez secos, as placas de vidro com os géis aderidos foram colocadas sob luz para
visualização dos padrões de bandas. O Marcador Molecular de 10 pares de bases (Invitrogen)

31
foi identificado, com seus diversos tamanhos e em seguida as amostras foram numeradas e os
alelos comparados com o marcador molecular para que fosse definido o seu tamanho (figura
12).
Após a genotipagem, foi construído um banco de dados com o tamanho dos alelos
genotipados para todos os indivíduos analisados, para as duas bibliotecas desenvolvidas, para
posterior análise estatística.
Figura 12: Genotipagem manual. Amostras enumeradas de 1 a 44; em azul estão os tamanhos do Marcador
Molecular (MM); em vermelho os diferentes alelos detectados para este loco (total de 9 alelos).
3.7 Análise Estatística
Uma vez genotipados, os locos das duas bibliotecas foram analisados estatisticamente
sobre alguns parâmetros, tais como: número de alelos por loco, conteúdo de polimorfismo
informativo (PIC), poder de discriminação (D), equilíbrio de Hardy-Weinberg, desequilíbrio
de ligação, heterozigosidade esperada e observada e presença de alelos nulos.
Os valores de PIC foram distribuídos em três classes: pouco informativos (menor que
0,25), razoavelmente informativos (de 0,25 a 0,5) e altamente informativos (acima de 0,5), de
acordo com Botstein et al., (1980). Sendo os locos com valores mais próximos de 1 são os
mais desejáveis.
Os valores de D estabelecem as melhores combinações dos marcadores microssatélites
que permitem a discriminação dos acessos selecionados. Nestas análises os seguintes
programas estatísticos foram empregados: Arlequin 3.1 (Excoffier et al., 2006), Fstat v. 2.9.3
(Goudet, 2002), MSTOOLS (Park, 2001), TFPGA (Miller, 1997) e MICRO-CHECKER
(Oosterhout et al., 2004).

32
4. Resultados
Foram sequenciados um total de 192 clones, para a biblioteca de Manaus e de Nova
Mazagão. Destes foi possível desenhar um total de 58 pares de primers flanquadores das
regiões microssatélites. Observou-se microssatélites com diferentes motivos de repetições.
Dentre eles, 74,36% foram dinucleotídeos, 15,38% trinucleotídeos, 6,41% tetranucleotídeos e
3,85% pentanucleotídeos, sendo 79,49% microssatélites simples perfeitos, 6,41% simples
imperfeitos e 14,01% compostos perfeitos e imperfeitos (Figura 13).
Figura 13: Frequência dos motivos de repetição dos microsstélites observados nas bibliotecas de A. nuneztovari.
Foram sintetizados 41 pares de primers, considerando os microssatélites
dinucleotídeos e trinucleotídeos com número de repetição igual ou superior a cinco. Estes,
foram em seguida ressuspendidos e testados na amostra de Manaus. Ao todo foi possível
amplificar 28 pares de primers, dos quais 23 foram genotipados em poliacrilamida.
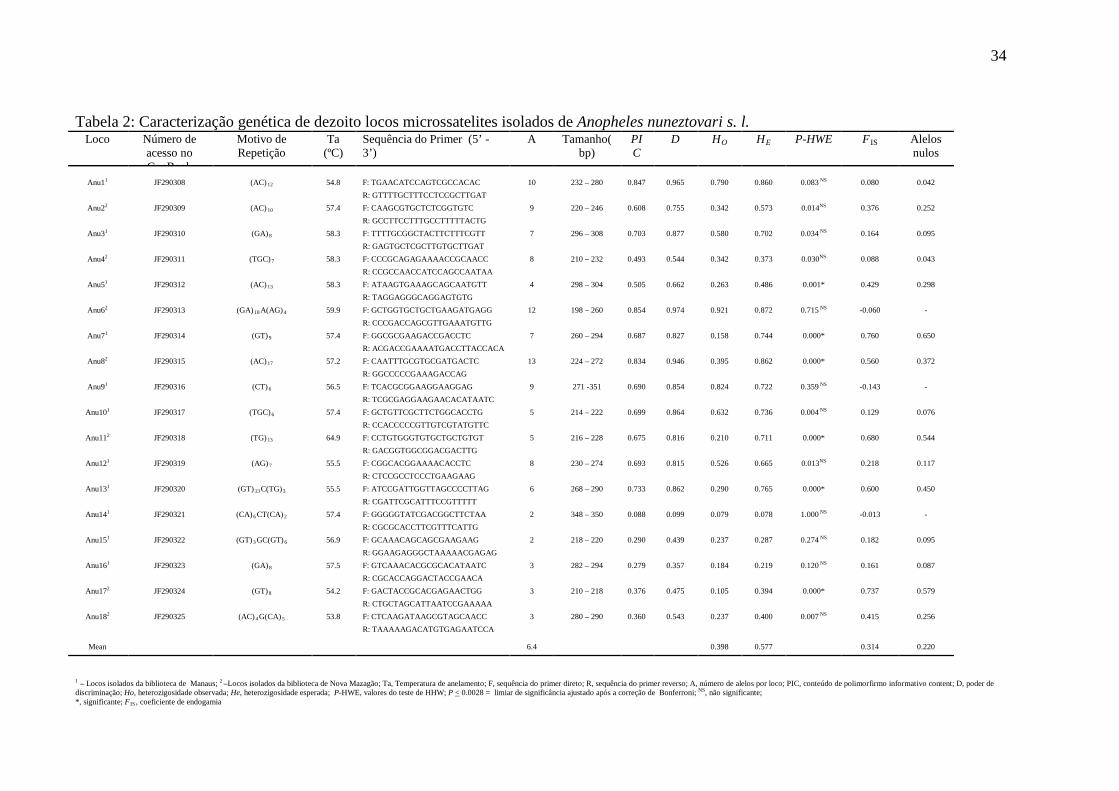
Foram obtidos 18 locos microssatélites polimórficos, avaliados com 34 a 39
indivíduos, sendo 11 locos da biblioteca de Manaus e sete da biblioteca de Nova Mazagão. A
temperatura de anelamento variou de 53,8 a 64.9 ºC. O número de alelos por loco variou de 2
(loco Anu 14 e Anu 15) a 13 (loco Anu8), e média de 6,4. A heterozigosidade observada (Ho)
variou entre 0,079 a 0,921 (Anu14 – Anu6, respectivamente) com uma média de 0,398,
enquanto a heterozigosidade esperada (He) variou entre 0,078 a 0,872 (Anu14 – Anu6,
respectivamente) com a média de 0,577. O conteúdo de polimorfismo informativo (PIC)
variou entre 0,088 (Anu14) a 0,854 (Anu6). Neste estudo, doze locos mostraram-se altamente

33
informativos (Anu1, Anu2, Anu3, Anu5, Anu6, Anu7, Anu8, Anu9, Anu10, Anu11, Anu12 e
Anu13), cinco razoavelmete informativos (Anu4, Anu15, Anu16, Anu17, Anu18) e apenas
um locos mostrou-se pouco informativo (Anu14). O poder de discriminação (D) foi estimado
para cada loco e variou de 0,099 (Anu14) a 0,974 (Anu6) (tabela 2 ).
O coeficiente de endogamia apresentou o valor mais elevado para o loco Anu7 (0,760)
(tabela 1). Seis locos (Anu5, Anu7, Anu8, Anu11, Anu13 e Anu17) mostraram desvio
significativo para EHW após a correção de Bonferroni. Consequentemente, foi utilizado o
programa MICRO-CHECKER e os resultados dos desvios é provavelmente decorrente da
presença de alelos nulos.
Dos 18 marcadores polimórficos, 7 foram testados para amplificação heteróloga em
outras cinco espécies de Anopheles: A. rangeli, A. dunhami, A. konderi, A. oswaldoi sensu
lato e A. darlingi (tabela 3). Todos os locos testados amplificaram com sucesso para todas as
espécies, exceto em A. darlingi, que não amplificou para nenhum loco testado.
34
Tabela 2: Caracterização genética de dezoito locos microssatelites isolados de Anopheles nuneztovari s. l. Loco Número de
acesso no GenBank
Motivo de Repetição
Ta (ºC)
Sequência do Primer (5’ - 3’)
A Tamanho(bp)
PIC
D HO HE P-HWE F IS Alelos nulos
Anu11 JF290308 (AC) 12 54.8 F: TGAACATCCAGTCGCCACAC 10 232 – 280 0.847 0.965 0.790 0.860 0.083 NS 0.080 0.042
R: GTTTTGCTTTCCTCCGCTTGAT Anu22 JF290309 (AC) 10 57.4 F: CAAGCGTGCTCTCGGTGTC 9 220 – 246 0.608 0.755 0.342 0.573 0.014NS 0.376 0.252
R: GCCTTCCTTTGCCTTTTTACTG Anu31 JF290310 (GA) 8 58.3 F: TTTTGCGGCTACTTCTTTCGTT 7 296 – 308 0.703 0.877 0.580 0.702 0.034 NS 0.164 0.095
R: GAGTGCTCGCTTGTGCTTGAT Anu42 JF290311 (TGC) 7 58.3 F: CCCGCAGAGAAAACCGCAACC 8 210 – 232 0.493 0.544 0.342 0.373 0.030NS 0.088 0.043
R: CCGCCAACCATCCAGCCAATAA Anu51 JF290312 (AC) 13 58.3 F: ATAAGTGAAAGCAGCAATGTT 4 298 – 304 0.505 0.662 0.263 0.486 0.001* 0.429 0.298
R: TAGGAGGGCAGGAGTGTG Anu62 JF290313 (GA) 18 A(AG) 4 59.9 F: GCTGGTGCTGCTGAAGATGAGG 12 198 – 260 0.854 0.974 0.921 0.872 0.715 NS -0.060 -
R: CCCGACCAGCGTTGAAATGTTG Anu71 JF290314 (GT) 9 57.4 F: GGCGCGAAGACCGACCTC 7 260 – 294 0.687 0.827 0.158 0.744 0.000* 0.760 0.650
R: ACGACCGAAAATGACCTTACCACA Anu82 JF290315
(AC) 17 57.2 F: CAATTTGCGTGCGATGACTC 13 224 – 272 0.834 0.946 0.395 0.862 0.000* 0.560 0.372
R: GGCCCCCGAAAGACCAG Anu91 JF290316 (CT) 6 56.5 F: TCACGCGGAAGGAAGGAG
9 271 -351 0.690 0.854 0.824 0.722 0.359 NS -0.143 -
R: TCGCGAGGAAGAACACATAATC Anu101 JF290317 (TGC) 6 57.4 F: GCTGTTCGCTTCTGGCACCTG 5 214 – 222 0.699 0.864 0.632 0.736 0.004 NS 0.129 0.076
R: CCACCCCCGTTGTCGTATGTTC Anu112 JF290318 (TG) 13 64.9 F: CCTGTGGGTGTGCTGCTGTGT 5 216 – 228 0.675 0.816 0.210 0.711 0.000* 0.680 0.544
R: GACGGTGGCGGACGACTTG Anu121 JF290319 (AG) 7 55.5 F: CGGCACGGAAAACACCTC 8 230 – 274 0.693 0.815 0.526 0.665 0.013NS 0.218 0.117
R: CTCCGCCTCCCTGAAGAAG Anu131 JF290320 (GT) 13 C(TG)3 55.5 F: ATCCGATTGGTTAGCCCCTTAG 6 268 – 290 0.733 0.862 0.290 0.765 0.000* 0.600 0.450
R: CGATTCGCATTTCCGTTTTT Anu141 JF290321 (CA) 6 CT(CA) 2 57.4
F: GGGGGTATCGACGGCTTCTAA 2 348 – 350 0.088 0.099 0.079 0.078 1.000 NS -0.013 -
R: CGCGCACCTTCGTTTCATTG Anu151 JF290322
(GT) 3 GC(GT) 6 56.9 F: GCAAACAGCAGCGAAGAAG 2 218 – 220 0.290 0.439 0.237 0.287 0.274 NS 0.182 0.095
R: GGAAGAGGGCTAAAAACGAGAG Anu161 JF290323 (GA) 8 57.5 F: GTCAAACACGCGCACATAATC 3 282 – 294 0.279 0.357 0.184 0.219 0.120 NS 0.161 0.087
R: CGCACCAGGACTACCGAACA Anu172 JF290324 (GT) 8 54.2 F: GACTACCGCACGAGAACTGG 3 210 – 218 0.376 0.475 0.105 0.394 0.000* 0.737 0.579
R: CTGCTAGCATTAATCCGAAAAA Anu182 JF290325 (AC) 4 G(CA) 5 53.8 F: CTCAAGATAAGCGTAGCAACC
3 280 – 290 0.360 0.543 0.237 0.400 0.007 NS 0.415 0.256 R: TAAAAAGACATGTGAGAATCCA
Mean 6.4 0.398 0.577 0.314 0.220
1 – Locos isolados da biblioteca de Manaus; 2 –Locos isolados da biblioteca de Nova Mazagão; Ta, Temperatura de anelamento; F, sequência do primer direto; R, sequência do primer reverso; A, número de alelos por loco; PIC, conteúdo de polimorfirmo informativo content; D, poder de discriminação; Ho, heterozigosidade observada; He, heterozigosidade esperada; P-HWE, valores do teste de HHW; P < 0.0028 = limiar de significância ajustado após a correção de Bonferroni; NS, não significante; *, significante; F IS , coeficiente de endogamia

35
Tabela 3: Amplificação heteróloga Loco
Espécies Anopheles
rangeli Anopheles dunhami
Anopheles konderi
Anopheles oswaldoi
Anopheles darlingi
ANU1 + + + + - ANU2 + + + + - ANU3 + + + + - ANU5 + + + + - ANU7 + + + + - ANU8 + + + + - ANU17 + + + + - + amplificou com sucesso; - não amplificou

36
5. DISCUSSÃO
Estudos de genética de populações em mosquitos vetores são importantes, pois
possibilitam quantificar a variabilidade, conhecer a estrutura genétia de populações, inferir o
grau de fluxo gênico entre as subpopulações e o tamanho efetivo (Ne) da população,
permitindo uma melhor compreensão da dinâmica das populações de vetores e, como
consequência, fornecendo subsídios para aprimorar as estratégias de controle de vetores
(Scarpassa et al., 1999; 2008). Resultados de vários estudos tem demonstrado que populações
geneticamente estruturadas, diferentes linhagens e membros de complexos de espécies
crípticas podem diferir na competência vetorial.
Tais estudos podem ser realizados através de diversos marcadores moleculares, como
marcadores isoenzimáticos, DNA mitocondrial e também com os marcadores microssatélites,
que podem medir a variabilidade genética através de parâmetros como: grau de
polimorfismo, número de alelos, heterozigosidade observada e esperada. Diversos estudos
demonstram a eficiência dos marcadores microssatélites frente a outros marcadores, sendo
estes últimos mais variáveis. Um exemplo disso, pode ser verificado ao observar dois estudos
com A. darlingi na Amazônia brasileira com marcadores diferentes, um isoenzimático e o
outro com microssatélites. O primeiro detectou baixa diversidade alélica (Santos et al., 1999)
enquanto o segundo uma elevada diversidade alélica (Scarpassa e Conn, 2007), demonstrando
portanto uma maior eficiência.
Estes marcadores também vem sendo preferidos para analisar complexo de espécies
pois além de auxiliar no entendimento da estrutura de populações de mosquitos podem
auxiliar também no entendimento das variantes das relações genéticas, como por exemplo a
diferença na variável epidemiológica, que faz com que mosquito prefira realizar o repasto
sanguíneo em um animal, em relação a outro (Keyghobadi et al., 2004).
O número de alelos por loco aqui caracterizados, estão numa proporção semelhante a
outras espécies de mosquitos vetores de doenças, tais como: Aedes aegypti, vetor da dengue,
cujo número de alelos para 17 locos microssatélites variou de 3 a 10 (Slotman et al., 2007);
Ae. polynesensis, vetor da filariose nas reagiões do Pacífico Sul, para 6 locos microssatélites
variou de 4 a 12 alelos (Behbahni et al., 2004); e para outros vetores da malária, A. funestus e
A. stephensi, para o primeiro variou de 5 a 15 alelos em 14 locos microssatélites (Cohuet et

37
al., 2002) e para o segundo de 2 a 13 alelos em 16 locos microssatélites (Verardi et al., 2002).
Número de alelos superior a estes podem ser observados em Culex pipens, vetor do vírus do
west do nilo, com 8 locos microssatélites, variou de 7 a 19 alelos (Keyghobadi et al., 2004) e
para A. marajoara variou de 11 a 52 alelos para 11 locos (Li et al., 2005). Conn et al. (2001)
ao isolar e caracterizar 8 locos microssatélites para A. darlingi, principal vetor da malária na
América Latina, encontraram um amplitude de 7 a 44 alelos por loco em amostras do Estado
do Pará (Brasil). Recentemente, foram caracterizados mais 24 locos microssatélites no Estado
do Amazonas, com 4 a 11 alelos por loco, para esta espécie (Lima et al., 2010).
Outras características detectadas em alguns locos microsstatelites aqui descritos,
também podem ser visualizadas em outras espécies de Anopheles, por exemplo, a deficiência
de heterozigotos em realação ao equilíbrio de Hardy-Weinberg (EHW) também foi detectado
em A. arabiensis (Temu & Yan, 2005) e A. moucheti (Antonio-Nkondjio et al., 2008).
Desvios para HWE podem ser causados por diversos fatores, tais como: endocruzamento,
efeito gargalo de garrafa, deriva genética, presença de alelos nulos ou ainda efeito de
amostragem. Esses fatores podem refletir o acúmulo de mutações nas regiões flanqueadoras
dos primers que resultam em defictis de heterozigotos. Os desvios apresentados nos oito locos
caracterizados neste trabalho, são devido ao excesso de homozigotos, indicados pelos valores
do coeficiente de endogamia (Fis) desses locos, que variaram de 0,429 (Anu8) chegando ate
0,760 (Anu10).
A estimativa de PIC fornece o conteúdo de polimorfismo de cada loco, levando em
consideração o número de alelos por loco e suas frequências relativas (Botstein et al., 1980).
Doze locos mostraram-se altamente informativos, com valores próximos a 1. Os índices de
PIC variaram de 0,088 a 0,854. Para estabelecer melhores combinações de marcadores
microssatélites que permitissem a discriminação dos acessos selecionados, foi utilizada a
metodologia de Tessier et al. (1999) para calcular o D (poder de discriminação). Os índices do
D variaram de 0,099 a 0,974. O maior índice de PIC e D foi observado no loco Anu6, o qual
apresentou 12 alelos.
Neste estudo foram desenvolvidos e caracterizados 18 locos microssatélites para A.
nuneztovari que poderão auxiliar no melhor entendimento da estrutura genética do vetor A.
nuneztovari. Estudos populacionais com esta espécie poderão auxiliar no melhor
entendimento da diferença comportamental desta espécie nas diferentes localidades de sua
distribuição geográfica, e elucidar sua participação na transmissão da malária na Amazônia
brasileira.

38
6. CONCLUSÕES
Das bibliotecas genômicas de Manaus e Nova Mazagão um total de 18 locos foram
utilizados na caracterização genética para A. nuneztovari.
Todos os locos caracterizados foram polimórficos variando de 2 a 13 alelos.
A amplificação heteróloga dos locos microssatélites testados teve amplificação
positiva para todos os locos em quatro espécies de Anopheles, exceto para A. darlingi,
provavelmente pela divergência evolutiva entre eles.
Os locos microssatélites isolados poderão ser usados como excelentes marcadores para
investigar genética de populações de A. nuneztovari s.l. e fornecer informações para um
melhor entendimento da diferença comportamental dos membros deste complexo e,
consequentemente, elucidar a participação desta espécie na transmissão da malária na
Amazônia brasileira.

39
7. REFERÊNCIAS BIBLIOGRÁFICAS
Antonio-Nkondjio, C.; Ndo, C.; Kengne, P.; Mukwaya, L.; Awono-Ambene, P.; Fontenille,
D. & Simard, F. 2008. Population structure of the malaria vector Anopheles moucheti in the
equatorial forest region of Africa. Malaria Journal, 7:120.
Arruda, M.; Carvalho, M.B.; Nussenzweig, R.S.; Maracic, M.; Ferreira, A.W. & Cochrane,
A.H. 1986. Potential vectors of malaria and their different susceptibility to Plasmodium
falciparum and Plasmodium vivax in Northern Brazil identified by immunoassay. American
Journal of Tropical Medicine and Hygiene, 35(5): 873-881.
Behbahani, A.; Dutton, T.J.; Raju, A.K.; Townson, H.; Sikins, P. 2004. Polymorphic
microsatellite loci in the mosquito Aedes polynesiensis. Molecular Ecology Notes. 4:59-61.
Bergo, E.S.; Souto, R.N.P.; Galardo, A.K.R.; Nagaki, S.S.; Calado, D.C. & Sallum, M.A.M.
2007. Systematic notes on Anopheles Meigen (Diptera: Culicidae) species in the state of
Amapá, Brazil. Memórias do Instituto Oswaldo Cruz, 102(3): 373-376.
Billotte, N.; Lagoda, P.J.L.; Risterucci, A.M. & Baurens, C. 1999: Microsatellite-enriched
libraries: applied methodology for the development of SSR markers in tropical crops. Fruits.
54, 277 - 288.
Borém, A. & Caixeta, E.T. 2006. Marcadores Moleculares. Viçosa, MG, 374pp.
Botstein, D.; White, R.L.;Skolnick, M. 1980. Construction of a genetic linkage map in a man
using restriction fragment leght polumorphisms. The American Journal of Human Genetics,
32: 314-331.
Calado, D.C.; Foster, P.G.; Bergo, E.S.; dos Santos, C.L.S.; Galardo, A.K.R. & Sallum,
M.A.M. 2008. Resurrection of Anopheles goeldii from synonymy with Anopheles nuneztovari

40
(Diptera, Culicidae) and a new record for Anopheles dunhami in the Brazilian Amazon.
Memórias do Instituto Oswaldo Cruz, 103(8): 791-799
.
Chambers, E.W.; Meece, J.K.; McGowan, J.A.; Lovin, D.D.; Hemme, R.R.; Chadee, D.D.;
McAbee, K.; Brown, S.E.; Knudson, D.L. & Severson, D.W. 2007. Microsatellite isolation
and linkage group identification in the yellow fever mosquito Aedes aegypti. Journal of
Heredity, 98(3):202-210.
Couhet, A.; Simard, F.; Berthomieu, A.; Raymond, M.; Fontenille, D.; Weill, M. 2002.
Isolation and characterization of microsatellite DNA markers in the malaria vector Anopheles
funestus. Molecular Ecology Notes, 2:498-500.
Consoli, R.A.G.B. & Oliveira, R.L.D. 1994. Principais mosquitos de importância sanitária no
Brasil. Fiocruz, Rio de janeiro, Brasil, 225 pp.
Conn, J.; Puertas, Y. R. & Seawright, J.A. 1993. A new cytotype of Anopheles nuneztovari
from Western Venezuela and Colombia. American Mosquito Control Association, 9(3): 294-
301.
Conn, J.E.; Mitchell, S.E. & Cockburn, A.F. 1998. Mitochondrial DNA analysis of the
Neotropical malaria vector Anopheles nuneztovari. Genome, 41: 313-327.
Conn, J.E.; Bollback, J. P.; Onyabe, D.Y.; Robinson, T. N.; Wilkerson, R.C.; Póvoas, M.M.
2001. Isolation of polymorphic microsatellite markers from the malaria vector Anopheles
darlingi. Molecular Ecology Notes, 1: 223-225.
Conn, J.E.; Vineis, J.H.; Bollback, J.P.; Onyabe, D.Y.; Wilkerson, R.C. & Póvoa, M.M. 2006.
Population structure of the malaria vector Anopheles darlingi in a malaria-endemic region of
eastern Amazonian Brazil. American Journal of Tropical Medicine and Hygiene, 74(5): 798-
806.
Creste, S.; Tulmann Neto, A. & Figueira, A. 2001. Detection of single sequence repeat
polymorphisms in denaturing polyacrylamide sequencing gels by silver staining. Plant
Molecular Biology Reporter, 19: 299-306.

41
Cova-Garcia, P. 1961. Notas sobre los anofelinos de Venezuela y su identificación. Ed.
Grafos, Caracas.
Diniz, F.M.; Iyengar, A.; Lima, P. S. da C.; Maclean, N.; Bentzen, Paul. 2007. Application of
a double-enrichment procedure for microsatellite isolation and the use of tailed primers for
high throughput genotyping. Genetics and Molecular Biology 30 (2): 380-384.
Elliott, R. 1968. Studies on man-vector contact in some malarious areas in Colombia. Bull.
World. Health, 38: 239-253
Elliott, R. 1972. The influence of vector behavior on malaria transmission. American Journal
of Tropical Medicine and Hygiene, 21:755-63.
Eisen, J.A. 1999. Mechanistic basis for microsatellite instability. In: Goldstein, D.B.;
Schlötterer, C. (eds), Microsatellites - evolution and applications, 4: 34-48.
Excoffier L, Laval G, Schneider S (2006) An integrated software package for population
genetics data analysis, version 3.1. Computational and Molecular Population Genetics Lab.
Institute of Zoology, University of Berne, Swittzerdand. Available from
http://cmpg.unibe.ch/software/arlequin3
Faran, M.E. 1980. Mosquito studies (Diptera: Culicidae). XXXIV. A revision of the
Albimanus section of the subgenus Nyssorhynchus of Anopheles. Contributions of the
American Entomological Instute, 15:1-215.
Fajardo, P. & Alzate, A. 1987. Anopheles nuneztovari como vector de malaria en el Bajo
Calima, Buenaventura. Revista Colombia Medica., 18(1): 14-18.
Forattini, O.P. 2002. Culicidologia Médica, vol. 2: Identificação, Biologia, Epidemiologia.
Editora da Universidade de São Paulo, São Paulo, Brasil, 860 pp.

42
Fritz, G.N.; Conn, J.; Cockburn, A. & Seawright, J. 1994. Sequence analysis of the ribosomal
DNA internal transcribed spacer 2 from populations of Anopheles nuneztovari (Diptera:
Culiciade). Molecular . Biology and Evolution, 11(3): 406-416.
Galardo, A.K.R.; Arruda, M.; Couto, A.A.R.D.; Wirtz, R.; Lounibos, L.P. & Zimmerman,
R.H. 2007. Malaria vector incrimination in three rural riverine villages in the Brazilian
Amazon. American Journal of Tropical Medicine and Hygiene,76(3): 461-469.
Galbadón, A. 1981. Anopheles nuneztovari: importante vector y agente da malaria refractiva
em Venezuela. BolDir Malariol Saneam Amb, 21:28-38.
Gautret, P.; Barreto, M.; Méndez, P.; Zorrilla, G., & Carrasquila, G. 1995. High prevalence of
malaria in village of the Colombia Pacific Coast. Memórias do Instituto Oswaldo Cruz, 90(5):
559-560.
Gorham, J.R.; Stojanovich, C.J. & Scott, H.G. 1967. Clave ilustrada para los mosquitos
anofelinos de Sudamerica Oriental. Departamento f Healther, Education and Welfare Public
Health Service. 64 pp.
Goudet, J. 2002. Fstat ver. 2.9.3.2. Institute of Ecology. Biology Building, UNIL, CH-1015,
Lausanne, Switzerland.
Hribar, L.J. 1994. Geographic variation of male genitalia of Anopheles nuneztovari (Diptera:
Culicidae). Mosquito Systematics Journal, 26(3): 132-144.
Keyghobadi, N.; Matrone, M.A.; Ebel, G.D.; Kramer, L.D.; Fonseca, D.M. 2004.
Microsatellite loci from the northern house mosquito (Culex pipens), a principal vector of
west Nile virus in North America. Molecular Ecology Notes, 4:20-22.
Kitzmiller, J.B.; Kreutzer, R.D. & Tallaferro, E. 1973. Chromosomal differences in
populations of Anopheles nuneztovari. Buletin of the World Health Organization, 48: 435-
445.

43
Lehman, T.; Hawley, W.A.; Kamau, L.; Fontenille, D.; Simard, F.& Collins, F.H. 1996.
Genetic differentiation of Anopheles gambiae populations from east and west Africa:
comparison of microsatellite and allozyme loci. Heredity, 77: 192-200.
Lehman, T.; Besansky, N.J.; Hawley, W.A.; Fahey, T.G.; Kamau, L. & Collins, F.H. 1997.
Microgeographic structure of Anopheles gambiae in western Kenya based on mtDNA and
microsatellite loci. Molecular Ecology, 6: 243-253.
Li, Y.C.; Korol, A.B.; Fahima, T. & Nevo, E. 2004. Microsatellites within genes: structure,
function and evolution. Molecular Biology and Evolution, 21(6): 991-1007.
Li, C.; Wilkerson, R.C,; Fonseca, D.M. 2005. Isolation of polymorphic microsatellite markers
from malaria vector Anopheles marajoara (Diptera: Culicidae). Molecular Ecology Notes, 5:
65-67.
Lima, G.N.; Batista, J.S.; Formiga, K.M.; Cidade, F.W.; Rafael, M.S.; Tadei, W.P.; Santos,
J.M.M. 2010. New 24 polymorphic DNA microsatellite loci for the major malaria vector
Anopheles darlingi and transpeciesamplification with another anophelines. Conservation
Genetics Resources, 2:205-209.
Linley, J.R.; Lounibos, L.P.; Conn, J.; Duzak, D. & Nishimura, N. 1996. A description and
morphometric comparison of eggs from eight geographic populations of the South American
malaria vector Anopheles (Nyssorhynchus) nuneztovari (Diptera: Culicidae).
American Mosquito Control Association, 12: 275-292.
Mello, M.P. de; Bachega, T.A.S.S.; Costa-Santos, M. da; Mermejo, L.M. & Castro, M. de.
2002. Bases Moleculares da Hiperplasia Adrenal Congênita. Arquivo Brasileiro
Endocrinologia Metabólica, 46(4): 457 – 477.
Michel, A.P.; Grushko, O.; Guelbeogo, W.M.; Lobo, N.F. S.; Sagnon, N’F.; Costantini, C. &
Besansky, N. J. 2006. Divergence with gene flow in Anopheles funestus from the Sudan
Savanna of Burkina Faso, West Africa. Genetics, 173: 1389-1395.

44
Miller MP (1997) Tools for population genetics analysis (TFPGA) 1.3. A windows program
for the analysis of allozyme and molecular population genetics data. Computer software
distributed by author.
Mirabello, L.; Vineis, J. H.; Yanoviak, S.P.; Scarpassa, V.M.; Póvoa, M.M.; Padilla, N.;
Achee, N.L. & Conn, J.E. 2008. Microsatellite data suggest significant population structure
and differentiation within the malaria vector Anopheles darlingi in Central and South
America. BMC Ecology, 8 (3):2-15
Mirabello, L. & Conn, J.E. 2008. Population analysis using the nuclear White gene detects
Piocene/Pleistocene lineage divergence within Anopheles nuneztovari in South America.
Medical and Veterinary Entomology, 22: 109-119.
Midega, J.T.; Muturi, E.J.; Balraine, F.N.; Mbogo, C.M.; Githure, J.; Beier, J.C.; Yan, G.
2010. Population structure of Anopheles gambie along the Kenyan coast. Acta Tropica, 114:
103-108.
Molina-Cruz, A.; de Mérida, A.M.P.; Mills, K.; Rodríguez, F.; Schoua, C.; Yurrita, M.M.;
Molina, E.; Palmiere, M. & Black IV, W. C. 2004. Gene flow among Anopheles albimanus
populations in Central America, South America and the Caribbean assessed by microsatellites
and mitochondrial DNA. American Journal of Tropical Medicine and Hygiene, 71(3): 350-
359.
Oliveira, E.J.; Pádua, J.G.; Zucchi, M.I.; Vencovsky, R.& Vieira, M.L.C. 2006. Origin,
evolution and genome distribution of microsatellites. Genetics and Molecular Biology, 29(2):
294-307.
Onyabe, D.Y. & Conn, J.E. 1999. Intragenomic heterogeneity of ribosomal DNA spacer
(ITS2) varies regionally in the Neotropical malaria vector Anopheles nuneztovari (Diptera:
Culicidae). Insect Molecular Biology, 8(4): 435-442.
OPAS, 2009. Malária, redução de casos. Em 2008, Brasil avança na redução do número de
casos de malária no país.

45
(http://new.paho.org/bra/index.php?option=com_content&task=view&id=401&Itemid=259)
acesso em: 20/10/2009
Park, S.D.E.2001. Trypanotolerance in West African Cattle and the population genetic effects
of selection Ph.D. thesis (in prep.), University of Dublin.
Posso, C.E.; González, R.; Cárdenas, H.; Gallego, G.; Duque, M.C. & Suarez, M.F. 2003.
Random amplified polymorphic DNA analysis of Anopheles nuneztovari (Diptera: Culicidae)
from Western and Northeastern Colombia. Memórias do Instituto Oswaldo Cruz, 98(4): 469-
476.
Póvoa, M.M.; Wirtz, R.A.; Lacerda, R.N.L.; Miles, M.A. & Warhurst, D. 2001. Malaria
vectors in the municipality of Serra do Navio, State on Amapá, Amazon Region, Brazil.
Memórias do Instituto Oswaldo Cruz, 96: 179-184.
Rey, H. & Renjifo, S. 1950. Anopheles (N.) nuneztovari infectado en la naturaleza con
Plasmodium spp. Rev. Acad. Colomb. Cienc. Exatas Fis. Quim. Nat., VII:534-538.
Rubio-Palis, Y. & Curtis, C.F. 1992. Biting and resting behavior os anophelines in Western
Venezuela and implications for control of malaria transmission. Medical and Veterinary
Entomology, 6: 325-334.
Roll Back Malaria, 2010. http://www.rbm.who.int/gmap/part3.pdf Acesso em 21/01/2011.
Sambrook, J. & Russell, D. 2001. Molecular Cloning: a laboratory manual. Gold Spring
Harbor Laboratory Press, Gold Spring Harbor, NY.
Santos, J.M.M.; Lobo, J.A.;Tadei,W.P.; Contel, E.P.B.1999. Intrapopulacional genetic
differentiation in Anopheles (N.) darlingi Root, 1926 (Diptera: Culicidae) in the Amazon
region. Genetics and Molecular Biology, 22(3): 325-331.
Scarpassa, V.M. & Tadei, W.P. 1990. Biologia de anofelinos amazônicos. XIII. Estudo do
ciclo biológico de Anopheles nuneztovari (Diptera: Culicidae). Acta Amazonica, 20 (unico):
95-118.

46
Scarpassa, V.M.; Tadei, W.P. & Suarez, M. F. 1996. Allozyme differentiation among
allopatric populations of Anopheles nuneztovari (Diptera: Culicidae). Brazilian Journal of
Genetics, 19(2): 265-269.
Scarpassa, V.M.; Tadei, W.P. & Suarez, M. F. 1999. Population structure and genetic
divergence in Anopheles nuneztovari (Diptera: Culicidae) from Brazil and Colombia.
American Journal of Tropical Medicine and Hygiene, 60(6): 1010-1018.
Scarpassa, V.M. Geurgas, S. Azeredo-Espin, A.M.L. & Tadei, W.P. 2000. Genetic divergence
in mitochondrial DNA of Anopheles nuneztovari (Diptera: Culicidae) from Brazil and
Colombia. Genetics and Molecular Biology, 23(1): 71-78.
Scarpassa, V.M. & Conn, J.E. 2007. Population genetic structure of the major malaria vector
Anopheles darlingi (Diptera: Culicidae) from the Brazilian Amazon, using microsatellite
markers. Memórias do Instituto Oswaldo Cruz, 102(3): 319-327.
Scarpassa, V.M.; Cardoza, T.B. & Cardoso Junior, R.P. 2008. Population genetics and
Phylogeography of Aedes aegypti (Diptera: Culicidae) from Brazil. American Journal of
Tropical Medicine and Hygiene, 78(6): 895-903.
Schlötterer, C. (2000). Evolutionary dynamics of microsatellite DNA. Chromosoma, 109:
365-371.
Sierra, D. M.; Velez, I. D. & Linton, Y-M. 2004. Malaria vector Anopheles (Nyssorhynchus)
nuneztovari comprises one genetic species in Colombia based on homogeneity of nuclear
ITS2 rDNA. Journal of Medical Entomology. Vol. 41(3):302-307.
Silva-Vasconcelos, A.M.Y.N.; Kató, E.N.; Mourão, R.T.L.; Souza, Lacerda, R.N.L.; Sibajev,
A.; Tsouris, P.; Póvoa, M.M.; Momem, H. & Rosa-Freitas, M.G. 2002. Biting indices, host-
seeking activity and natural infection rates of anopheline species in Boa Vista, Roraima,
Brazil from 1996 to 1998. Memórias do Instituto Oswaldo Cruz, 97: 151-161.

47
Silva, A.P.B.; Tadei, W.P.; Santos, J.M.M. 2008. Análisis de polimorfismo en poblaciones de
Anopheles darlingi de La Amazonia Brasileña. Revista Latinoamericana de Genética, Lima,
Peru. p. 228-234.
Slotman, M.A.; Kelly, N.B.; Harrington, L.C.; Kitthawee, S.; Jones, J.W.; Scott, T.W.;
Caccone, A.; Powell, J.R. 2007. Polymorphic microsatellite markers for studies of Aedes
aegypti (Diptera: Culicidae), the vector of dengue and yellow fever. Molecular Ecology
Notes, 7:168-171.
Susam/Devis – Secretaria de saúde do Amazonas e Departamento de vigilância em saúde
2004. Relatório de atividades de controle de endemias, 2002 a 2004. Manaus, Amazonas.
SIVEP –Malária, 2009. Casos de malária notificadas na FMT-AM – 2007 a 2009.
(http://www.fmt.am.gov.br/informe/2009/MURAL%20MALARIA2007A2009_Final.pdf).
Acesso em : 02/11/2009
SVS, 2010.
http://portal.saude.gov.br/portal/aplicacoes/noticias/default.cfm?pg=dspDetalheNoticia&id_ar
ea=124&CO_NOTICIA=12040. Acesso em 21/01/2011
Tadei, W.P.; Thatcher, B.D.; Santos, J.M.M.; Scarpassa, V.M.; Rodrigues, I.B. & Rafael,
M.S. 1998. Ecological observations on anopheline vectors of malaria in the Brazilian
Amazon. American Journal of Tropical Medicine and Hygiene, 59:325-35
Tadei, W.P., Thatcher, B.D. 2000. Malaria vectors in the Brazilian Amazon: Anopheles of the
subgenus Nyssorhynchus. Revista do Instituto de Medicina Tropical de São Paulo, 43(2): 86-
97.
Tautz, D. 1989. Hypervariability of simple sequence as a general source for polymorphic
DNA markers. Nucleic Acid Research, 17(16): 6463-6471.

48
Tessier C, David J, This P, Boursiquot JM, Charrier A. 1999. Optimization of the choice of
molecular markers for varietal identification in Vitis vinifera L. Theoretical and Applied
Genetics. 98: 171-177.
Temu, E. A. & Yan, G. 2005. Microsatellite and mitochondrial genetic differentiation of
Anopheles arabiensis (Diptera: Culicidae) from western Kenya, the great rift valley, and
coastal Kenya. American Journal of Tropical Medicine and Hygiene,73(4): 726-733.
Touré, Y.T.; Oduola, A.M. & Morel, C.M. 2004. The Anopheles gambiae genome: next steps
for malaria vector control. Trends in Parasitology, 20: 142-149.
Verardi, A.; Donnely, M.J.; Rowland, M.; Townson, H. 2002. Isolation and characterization
of microsatellite loci in the mosquito Anopheles stephensi Liston (Diptera: Culicidae).
Molecular Ecology Notes, 2: 488-490.
Weber, J. L. 1990. Informativeness of human (dCdA)n.(dG-dT)n polymorphisms. Genomics,
7, 524-530.
World malaria reports 2005 - http://www.rollbackmalaria.org/wmr2005/ acesso em
12/11/2009
Zheng, L.; Collins, F.H.; Kumar, V.; Kafatos, F.C.; 1993. A detailed genetic map for the
malaria vector, Anopheles gambie. Science, 261: 605-608.
FINANCIAMENTOS:
Genética de populações de vetores da malária e dengue e dos sorotipos do vírus dengue na
Amazônia Ocidental. CNPq - CT-Amazônia Edital Nº 055/2008; Coordenadora: Dra. Vera
Margarete Scarpassa
Institutos Nacionais de Ciência e Tecnologia DA BIODIVERSIDADE AMAZÔNICA CNPq -
Edital dos Institutos Nacionais de Ciência e Tecnologia: 015/2008 Coordenador: Dr. William
E. Magnusson


















