
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA INPA …§ão... · DANILO GABRIEL DA ROCHA CASTANHO...
57
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA DE ÁGUA DOCE E PESCA INTERIOR – BADPI CONTRIBUIÇÃO DE ESPÉCIES ORNAMENTAIS PARA A ESTRUTURA FUNCIONAL DE ASSEMBLEIAS DE PEIXES DE IGARAPÉS DO MÉDIO RIO NEGRO, AMAZÔNIA BRASILEIRA DANILO GABRIEL DA ROCHA CASTANHO MANAUS – AMAZONAS JUNHO/2017
Transcript of INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA INPA …§ão... · DANILO GABRIEL DA ROCHA CASTANHO...

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA DE ÁGUA DOCE E
PESCA INTERIOR – BADPI
CONTRIBUIÇÃO DE ESPÉCIES ORNAMENTAIS PARA A ESTRUTURA
FUNCIONAL DE ASSEMBLEIAS DE PEIXES DE IGARAPÉS DO MÉDIO RIO
NEGRO, AMAZÔNIA BRASILEIRA
DANILO GABRIEL DA ROCHA CASTANHO
MANAUS – AMAZONAS
JUNHO/2017

i
DANILO GABRIEL DA ROCHA CASTANHO
CONTRIBUIÇÃO DE ESPÉCIES ORNAMENTAIS PARA A ESTRUTURA
FUNCIONAL DE ASSEMBLEIAS DE PEIXES DE IGARAPÉS DO MÉDIO RIO
NEGRO, AMAZÔNIA BRASILEIRA
ORIENTADORA: Dra. Cláudia Pereira de Deus
COORIENTADORES: Dr. Jansen A. S. Zuanon
Dr. Rafael Pereira Leitão
Dissertação de Mestrado apresentada ao
Instituto Nacional de Pesquisas da
Amazônia – INPA, como parte dos
requisitos para obtenção do título de
Mestre em Ciências Biológicas, área de
concentração em Biologia de Água Doce
e Pesca Interior.
MANAUS – AMAZONAS
JUNHO/2017

ii
B Castanho, Danilo Gabriel da Rocha. Contribuição de espécies ornamentais para a estrutura funcional de assembleias de peixes de igarapés do médio rio negro, Amazônia Brasileira / Danilo Gabriel da Rocha Castanho. - Manaus, 2017, 55f.: il, color.
Dissertação (mestrado) – INPA, Manaus, 2017 Orientadora: Cláudia Pereira de Deus Coorientador: Jansen Alfredo Sampaio Zuanon; Rafael Pereira Leitão Área de concentração: Biologia de Água Doce e Pesca Interior
1.Diversidade Funcional. 2. Ictiofauna. 3. Pesca Ornamental. 4. Igarapés. 5. Amazônia. I. Título
CDD.
Sinopse
Foi analisada a estrutura funcional das assembleias de peixes em 13 igarapés
utilizados pela pesca ornamental na porção média da bacia do rio Negro. A partir de
cenários hipotéticos de subtração de abundâncias, foi avaliada a contribuição das
espécies ornamentais na estrutura funcional das assembleias amostradas.
Palavras-chave: Diversidade Funcional; Ictiofauna; Pesca Ornamental; Igarapés;
Amazônia.
Keywords: Functional Diversity; Ichthyofauna; Ornament fishing; streams, Amazon.

iii
Agradecimentos
Agradeço aos notáveis pesquisadores, professores, orientadores e
amigos Dr. Cláudia Pereira de Deus, Dr. Jansen Zuanon e Dr. Rafael Leitão por
todos os conselhos, ensinamentos e apoio concedido, pela paciência e tempo
desprendido e pela oportunidade de realizar essa etapa de minha formação em
ótima companhia.
Agradeço, também, ao Dr. Fabrício Teresa por aceitar entrar nessa “empreitada”,
por toda ajuda prestada e pelo tempo cedido.
Agradeço ao INPA - Instituto Nacional de Pesquisas da Amazônia e ao
Programa de Pós-Graduação em Biologia de Água Doce e Pesca Interior, pelo
auxílio técnico e administrativo, pela infraestrutura proporcionada. Agradeço
ainda ao CNPQ pela bolsa concedida.
Gostaria de agradecer às amigas Paulinha, Akemi, Thatyla, Cris, Camila
e Natália pelos cafés com muita prosa, os almoços compartilhados e pelas
“enroladas” no laboratório. À Dra. Sidinéia pelas conversas sempre
divertidíssimas, pelos conselhos e reflexões que ultrapassam a vida profissional
(no mínimo). À companheiríssima Marina por me apresentar o sistema médico
manauara e pelas inúmeras risadas regadas com suco de cevada. Aos
companheiros de Catalão, Fabiano, Oliveira e Frank pelas coletas sempre muito
produtivas.
Agradeço aos amigos Tiago e Sergio pelo tempo e muita paciência
quando o assunto é o R e, pelas sugestões sempre bem-vindas.
Agradeço aos amigos da melhor turma de Pós-Graduação que já houve:
Adria, Marcele, Susana, Paola, Carla, Felipe, Danniel, Valesca, Raize, Maiby,
Camila, Andressa, Cris, Eduardo, Elio e Enzio pela companhia, experiências
trocadas, ajudas de última hora e principalmente pelos churrascos, festas,
viagens, risadas e muita união.
Agradeço aos amigos de república Nayana, Kaue, Gringo #1 (Scott),
Gringo #2 (Cameron), Tai e Fer pelos almoços em família, pelos podrões e
principalmente por dividir o tempo comigo no Botafogo, o melhor bar da região
norte.

iv
Agradeço aos companheiros de coleta Gonô e Eudes por me
apresentarem a real face da maior floresta tropical do mundo, pelas pescarias
sempre muito divertidas e por fazerem das coletas a melhor experiência que eu
poderia ter.
Por fim, agradeço imensamente minha família por todo apoio, confiança e
incentivo dado a mim.

v
Dedico este trabalho aos meus pais e
toda minha família, pela confiança cedida,
pelo apoio incondicional, pelo incentivo pleno
e pela união sempre presente em nossa
família.

vi
“Cada um de nós compõe a sua história
cada ser em si carrega o dom de ser capaz e ser
feliz”
Almir Sater & Renato Teixeira

vii
Resumo
O estudo da biodiversidade através de sua faceta funcional traz vantagens
relacionadas à compreensão dos mecanismos estruturadores, às relações entre
as espécies e seus respectivos papeis ecossistêmicos, e à predição de
alterações nos serviços biológicos frente às alterações ambientais. Uma das
principais fontes de pressões sobre as populações naturais de peixes na
Amazônia é a pesca, que tem sido realizada mais intensivamente ao longo das
últimas décadas. Visto que a pesca ornamental realizada na porção média da
bacia do rio Negro é uma atividade extrativista com foco direcionado a certas
espécies, é possível que a pressão exercida pela atividade gere perturbações na
estrutura funcional das comunidades e suprima funções ecológicas específicas.
Com esse cenário, o presente estudo buscou estimar os efeitos da pesca
ornamental na estrutura funcional de assembleias de peixes de igarapés, por
meio da elaboração de cenários hipotéticos de subtração de abundâncias de
espécies ornamentais e não ornamentais. Por meio da avaliação de três índices
funcionais complementares (Redundância, Divergência e Dispersão funcional)
constatamos que as assembleias estudadas possuem alta diversidade funcional
relacionada ao uso do ambiente e de recursos alimentares. As espécies
exploradas pela pesca ornamental contribuem significativamente para a
redundância funcional das assembleias, e a retirada simulada do Tetra cardinal
Paracheirodon axelrodi causa maiores perdas para a assembleia do que a
retirada de espécies não ornamentais. Tais resultados indicam que a atividade
deve ser executada de forma ordenada e com manejo adequado dos estoques,
a fim de não comprometer a estrutura funcional das assembleias locais de
peixes. Uma vez que a atividade não gera alterações significativas nas condições
físicas e químicas do ambiente, e que a inviabilidade econômica diminui as
chances de extinção local, a pesca ornamental na região de Barcelos tem
condições propícias a se manter viável sem alterar a estabilidade do
ecossistema.
.
Palavras-chave: Diversidade Funcional; Ictiofauna; Pesca Ornamental; Igarapés;
Amazônia.

viii
Abstract
The study of biodiversity through its functional facet brings advantages related to
the understanding of the structuring mechanisms, the relationships between the
species and their respective ecosystemic roles, and the prediction of changes in
biological services in the face of environmental changes. One of the main sources
of pressures on natural fish populations in the Amazon is fishing, which has been
more intensively carried out over the last decades. Seeing that the ornamental
fishery carried out in the middle portion of the Negro River basin is an extractive
activity focused on certain species, it is possible that the pressure exerted by the
activity generates disturbances in the functional structure of the communities and
suppresses specific ecological functions. With this scenario, the present study
sought to estimate the effects of ornamental fishing on the functional structure of
streams fish assemblages, through the elaboration of hypothetical scenarios of
subtraction of abundances of ornamental and non - ornamental species. Through
the evaluation of three complementary functional indexes (Redundancy,
Divergence and Functional Dispersion) we found that the assemblies studied
have high functional diversity related to the use of the environment and food
resources. The species exploited by ornamental fisheries contribute significantly
to the functional redundancy of the assemblages, and the simulated withdrawal
of the Tetra cardinal Paracheirodon axelrodi causes greater losses to the
assembly than the removal of non-ornamental species. These results indicate
that the activity should be performed in an orderly manner and with adequate
management, in order not to spoil the functional structure of local fish
assemblages. As the activity does not generate significant changes in the
physical and chemical conditions of the environment, and that economic
unfeasibility decreases the chances of local extinction, ornamental fishery in the
Barcelos region is prone to remain viable without altering the stability of the
ecosystem.
Keywords: Functional Diversity; Icthyofauna; Ornament fishing; streams,
Amazon.

ix
Sumário
1. Introdução geral ......................................................................................... 1
2. Objetivos .................................................................................................... 5
2.1 Objetivo Geral ........................................................................................... 5
2.2 Objetivos Específicos ................................................................................ 5
3. Referências bibliográficas..........................................................................6
4. Contribuição Funcional de Espécies Ornamentais Em Assembleias de
Peixes do Médio Rio Negro, Amazônia brasileira........................................ 10
5. Introdução ................................................................................................ 11
6. Material e Métodos................................................................................... 12
6.1 Área de estudo ........................................................................................ 12
6.2 Delineamento experimental .................................................................... 13
6.4 Material ictiológico ................................................................................... 14
6.5 Estrutura funcional .................................................................................. 14
6.6 Análises estatísticas ................................................................................ 16
7. Resultados ............................................................................................... 18
7.1 Estrutura taxonômica............................................................................... 18
7.2 Estrutura funcional................................................................................... 19
7.3 Efeito da retirada de espécies ornamentais na estrutura funcional das
assembleias (Análise A).................................................................................... 21
7.4 Comparação do efeito gerado pela subtração das abundâncias de espécies
ornamentais e não ornamentais na estrutura funcional das assembleias (Análise
B)....................................................................................................................... 23
8. Discussão................................................................................................. 27
9. Considerações finais............................................................................... 30
10. Referências Bibliográficas...................................................................... 30
11. Material suplementar............................................................................... 35

x
Lista de Figuras
Figura 1. Área de estudo e locais de coleta (círculos pretos) na porção média da
bacia do rio Negro, região de Barcelos-AM (estrela
cinza).................................................................................................................13
Figura 2. Volume ocupado pelas espécies categorizadas como ornamentais no
espaço multidimensional. Polígono branco: volume ocupado pelo total de
espécies amostradas; polígono rosa: volume ocupado por espécies ornamentais;
círculos brancos: espécies não ornamentais; círculos pretos: espécies
ornamentais.......................................................................................................20
Figura 3. Resultados obtidos no índice de redundância funcional, nos cenários
1 e 2, análise A, em condições originais e com 100% das abundâncias das
espécies ornamentais subtraídas.......................................................................22
Figura 4. Resultados obtidos no índice de divergência funcional, no cenário 2,
análise A, em diferentes porcentagens de subtração de abundâncias de P.
axelrodi. ............................................................................................................ 23
Figura 5. Resultados obtidos no cenário 1 de seleção de espécies nos índices
de Redundância, Divergência e Dispersão funcional, nas diferentes
porcentagens de subtração de abundâncias de espécies ornamentais (preto) e
não ornamentais (cinza). .................................................................................. 24
Figura 6. Resultados obtidos no cenário 2 de seleção de espécies nos índices
de Redundância, Divergência e Dispersão funcional, nas diferentes
porcentagens de subtração de abundâncias de P. axelrodi (preto) e espécies não
ornamentais (cinza). ......................................................................................... 26
Figura S1. Fotografia de um rapiché, aparelho de coleta de peixes ornamentais
típico da região do médio rio Negro. Fonte: Danilo G. R. Castanho.. ............... 35
Figura S2. Vista lateral de um cacuri, tipo de armadilha para peixes ornamentais
utilizada na região do médio rio Negro. Fonte: Danilo G. R. Castanho. ........... 35
Figura S3. Esquema das medidas ecomorfológicas obtidas com auxílio de
paquímetro digital e análise fotográfica, através do programa computacional
Image J. Siglas: Cp comprimento padrão, Amc altura máxima do corpo, Aco
Altura da cabeça, passando pelo centro do olho, Do diâmetro do olho, Doc
distância do centro do olho até a parte inferior da cabeça, Lc largura do corpo,
Lb largura da boca, Ab altura máxima da boca, Po distância entre a pré-maxila
e a base da cabeça, Aip altura da inserção da nadadeira peitoral, Acp altura do
corpo passando pela inserção da nadadeira peitoral, Cnp comprimento máximo
da nadadeira peitoral, Anp área da nadadeira peitoral, Hnc altura da nadadeira
caudal, Anc área da nadadeira caudal, Apc altura do pedúnculo caudal, Prt

xi
comprimento da protrusão bucal, Fo comprimento do focinho, Massa peso total.
Fonte: Leitão et al., 2016. ................................................................................. 36
Figura S4. Qualidade do espaço funcional baseado no protocolo proposto por Maire et al. (2015)..............................................................................................38 Lista de Tabelas
Tabela S1. Lista de atributos funcionais e seus respectivos significados ecológicos. Códigos para medidas morfológicas são apresentados na figura S3.......................................................................................................................36 Tabela S2. Atributos funcionais que mais influenciam cada eixo da PCoA.
Valores obtidos através da correlação entre os atributos funcionais e os eixos da
PCoA..................................................................................................................39
Tabela S3. Lista das espécies amostradas e respectivas abundâncias.
*Espécies categorizadas como ornamentais.....................................................40
Tabela S4. Variáveis ambientais amostradas....................................................44
Tabela S5. Resultados encontrados nas condições originais de cada igarapé. Ne. Número total de espécies; Nt. Número total de exemplares capturados; NeO. Número de espécies ornamentais; No. Número de exemplares ornamentais; FR. Redundância Funcional; FDiv. Divergência funcional; FDis. Dispersão funcional............................................................................................................ 45

1
Introdução geral
Dentre as temáticas da ecologia, a compreensão dos mecanismos estruturantes
das comunidades e dos ecossistemas sempre esteve em foco. Com a
degradação ambiental em ritmo acelerado (Vitousek, Mooney, Lubchneco, &
Melillo, 1997) e elevada taxa atual de extinção de espécies (Barnosky et al.,
2011), essa é uma questão de grande importância para o manejo e conservação
da biodiversidade. A maior parte dos estudos com foco na conservação biológica
das espécies são baseados em aspectos taxonômicos da biodiversidade,
gerando informações limitadas sobre o funcionamento das comunidades, ao se
basearem principalmente na análise de alterações na quantidade de indivíduos
por espécie e sua distribuição espaço-temporal (Magurran, 2004; Cianciaruso,
Silva, & Batalha, 2009; Teresa & Casatti, 2012).
Há algum tempo essas formas tradicionais de mensuração da biodiversidade
vêm sendo questionadas com o pressuposto de que não refletem
adequadamente a estrutura e funcionamento das comunidades (Villéger, Mason,
& Mouillot, 2008). Desta forma, métricas que consideram relações filogenéticas
e características funcionais vêm ganhando espaço no meio científico (Webb,
2000; Diaz & Cabido, 2001; Ricotta, Avena, & Chiarucci, 2005; Petchey &
Gaston, 2006; Cianciaruso et al., 2009).
Uma alternativa para se quantificar a diversidade de uma comunidade é a
utilização de medidas de diversidade filogenética. Estas possuem como base a
história evolutiva das espécies e levam em consideração as relações de
parentesco entre elas (Magurran, 2004). Uma comunidade que apresenta níveis
elevados de diversidade filogenética tende a possuir espécies distantes entre si
em termos evolutivos. Neste caso, essa comunidade poderia ser considerada
mais diversa do que uma outra composta por igual número de espécies, mas
filogeneticamente mais próximas entre si (Cianciaruso et al., 2009).
Outra forma de caracterizar uma comunidade é através de sua estrutura e
diversidade funcional. Essa abordagem faz uso de índices obtidos por
características do indivíduo que expressem algum tipo de funcionalidade
(Villéger et al., 2008; Cianciaruso et al., 2009; Mouillot, Villéger, Scherer-

2
Lorenzen, & Mason, 2011; Mouillot, Graham, Villéger, Mason, & Bellwood, 2013;
Leitão et al., 2016). Uma definição amplamente aceita para diversidade funcional
é o valor e a variação dos atributos ecológicos das espécies que influenciam o
funcionamento da comunidade (Tilman, 2001; Petchey & Gaston, 2002; 2006;
Cianciaruso et al., 2009).
As medidas de diversidade e estrutura funcional das comunidades podem ser
feitas a partir do espaço multidimensional, no qual as espécies estão contidas e
posicionadas de acordo com o conjunto de suas características funcionais
(Villéger et al., 2008). Nele são calculadas as distâncias entre espécies, a
distribuição de suas abundâncias, e o volume ocupado por diferentes conjuntos
de espécies (i.e. comunidades locais).
Os índices funcionais estão ganhando visibilidade crescente no meio científico,
uma vez que com esse tipo de medida é possível fazer inferências sobre o nicho
ocupado pelas espécies e as características ambientais que favorecem sua
existência local. De acordo com Hutchinson (1958), é possível mensurar o nicho
de uma espécie a partir da definição de um espaço multidimensional. Nesta
perspectiva, as características ecológicas do organismo que permitem sua
sobrevivência e reprodução são representadas pela posição no espaço
multidimensional (Hutchinson & MacArthur, 1959; Maguire, 1973; Whittaker,
Levin, & Root, 1973).
Com o uso desse tipo de métrica é possível analisar a similaridade de funções
exercidas por espécies diferentes (redundância funcional), inferindo sobre a
estabilidade potencial de uma comunidade ou ecossistema. Um exemplo é o
estudo realizado por Laliberté et al. (2010), onde inferiu que a resiliência é
dependente da redundância funcional da comunidade. A redundância funcional
também é associada à garantia de continuidade no desempenho de certas
funções ecológicas, mesmo com a extinção de espécies (Ricotta et al., 2016).
Com os índices funcionais pode-se avaliar, ainda, o nível de especialização das
funções exercidas e o quão originais elas são (Villéger, Miranda, Hernández, &
Mouillot, 2010; Leitão et al., 2016). É possível, também, analisar eventuais
diferenças nas respostas de espécies similares a determinadas alterações
ambientais, o que pode ser fundamental para a manutenção da estabilidade da
comunidade frente a distúrbios (Laliberté et al., 2010). A abordagem funcional

3
também permite avaliar como a comunidade explora os recursos ambientais, a
partição e a ocupação de nichos (divergência e dispersão funcional), e quais
características predominam na comunidade (identidade funcional) (Moreno,
Arita, & Solis, 2006; Ricklefs, 2012; Villéger et al., 2008; Laliberté et al., 2010;
Laliberté & Legendre, 2010; Mouillot et al., 2013).
A diferenciação na forma como as espécies utilizam o ambiente e os recursos
disponíveis, é um aspecto importante na viabilidade da enorme diversidade
encontrada na bacia amazônica, principalmente em regiões pobres em
nutrientes, como em ambientes aquáticos da bacia do rio Negro. Nesta bacia, a
coexistência de mais de 1.000 espécies de peixes (Chao, Prang, & Petry, 2001)
em uma região de baixa produtividade primária, elevada acidez e baixas
concentrações de oxigênio dissolvido é explicada, em partes, pela grande
diversidade de habitats encontrados e pela variedade de itens alimentares,
autóctones e alóctones, explorados de diferentes formas (Goulding, Carvalho, &
Ferreira, 1988).
A maior diversidade de peixes de água doce do mundo está localizada na Bacia
Amazônica, superando a marca de 3.000 espécies (Santos, Ferreira, & Zuanon
2006; Reis et al., 2016), e expectativas de que ainda existam cerca de 2.000
espécies por serem descritas (Queiroz et al., 2013). A enorme biodiversidade da
bacia é associada a diversas características ecológicas da região, como por
exemplo a grande diversidade de ambientes aquáticos, tipos de água, e a
estabilidade climática da região (Goulding et al., 1988; Reis et al., 2016). Em
regiões de várzea, o grande aporte de sedimentos carreados das regiões
andinas atuam como fertilizantes, aumentando as taxas de produção primária,
enriquecendo a base da cadeia trófica (Irion, 1978; Junk, 1984; Downing, Plante,
& Lalonde, 1990; Assis, Wittman, Piedade, & Haugaasen, 2014). A grande
quantidade de matéria orgânica alóctone e sua rápida ciclagem, favorecida pelo
clima tropical e pela diversa fauna decompositora, fazem com que os nutrientes
retornem rapidamente ao sistema. Aliado a isto, o regime de inundação sazonal,
que regula a dinâmica dos sistemas de grandes rios e planícies de inundação
desta área, também favorece a alta disponibilidade de nutrientes e de hábitats
temporários, que podem ser explorados por um grande número de espécies
(Junk, Bayley, & Sparks, 1989; Junk & Wantzen, 2004).

4
O grande potencial biológico da região vem sendo submetido a intensas
pressões antrópicas, das mais variadas fontes. Tais distúrbios ambientais têm
sido ocasionados por grandes obras de infraestrutura (Junk et al., 2014) e pelas
diversas formas de uso da terra (i.e. atividades agropecuárias, madeireiras e
urbanização), que modificam a paisagem; pela poluição, que descaracteriza as
condições físico-químicas de ambientes terrestres e aquáticos (Couceiro,
Fosberg, Hamada, & Ferreira, 2006); e pela explotação não sustentável dos
recursos naturais (Isaac & Bathem, 1995).
Dentre os diversos tipos de atividades extrativistas na Amazônia, a pesca é um
dos principais. Movimentando de 100 a 200 milhões de dólares por ano e
empregando cerca de 250.000 pessoas, estima-se que a produção pesqueira
nas águas interiores da região alcance a marca de 217.000 toneladas por ano
(Santos et al., 2006). Outra atividade extrativista importante no cenário
econômico regional é a pesca ornamental, sendo considerada a terceira maior
atividade extrativista do estado do Amazonas (Souza & Mendonça, 2009). A
atividade é realizada pela população ribeirinha e tem por finalidade o comércio
de peixes vivos para fins de aquariofilia.
No estado do Amazonas, foram comercializados mais de 20 milhões de
exemplares, somente no ano de 2003, destes, 90% são representados por
apenas onze espécies. A porção média da bacia do rio Negro é o principal polo
extrator, representando mais de 80% do peixe ornamental comercializado no
estado (Chao et al., 2001; Instituto Brasileiro do Meio Ambiente e dos Recursos
Renováveis [IBAMA], 2008; Anjos, Amorim, Siqueira, & Anjos, 2009). A intensa
explotação direcionada a um número reduzido de espécies, e agrupada em uma
área específica, potencialmente, pode ocasionar alterações na estruturação das
assembleias de peixes e extinguir serviços ecossistêmicos executados pelas
espécies explotadas.
Com base no exposto acima, o presente trabalho busca avaliar a estrutura
funcional das assembleias de peixes em áreas com histórico de pesca
ornamental e estimar o potencial efeito da retirada de espécies ornamentais no
funcionamento dessas assembleias. A questão chave a ser respondida é: A
retirada de espécies ocasionada pela explotação direcionada de espécies de
interesse ornamental poderia levar à perda de funções dentro das assembleias?

5
Ou existe um nível de redundância funcional capaz de compensar os efeitos
dessa sobre-explotação?
Objetivos
Objetivo geral
Avaliar a contribuição de espécies ornamentais e o possível impacto de sua
retirada para a estrutura funcional de assembleias de peixes em igarapés
sujeitos à explotação pela pesca ornamental na região do médio rio Negro,
Amazonas, Brasil.
Objetivos específicos
Determinar a composição e abundância das assembleias de peixes em
igarapés sujeitos à explotação de espécies pela pesca ornamental;
Analisar a estrutura funcional das assembleias de peixes nesses
igarapés;
Estimar o efeito potencial da extirpação local de espécies de peixes de
interesse ornamental na estrutura funcional das assembleias.
A presente dissertação é apresentada a seguir em formato de artigo científico, a
fim de otimizar o processo de publicação do estudo, intitulado “Contribuição de
espécies ornamentais para a estrutura funcional de assembleias de peixes
de igarapés do médio rio Negro, Amazônia brasileira”. Tal artigo será
submetido a revista Ecology of Freshwater Fish.

6
Referências bibliográficas
Anjos, H.D.B. dos, Amorim, R.M.S. de, Siqueira, J.A., & Anjos, C. R. dos. (2009),
Exportação de peixes ornamentais do estado do Amazonas, bacia amazônica,
Brasil. Instituto de Pesca, São Paulo, Brasil, 35 (2): 259-274.
Assis, R. L., Wittmann, F., Piedade, M. T. F., & Haugaasen, T. (2014), Effects of
hydroperiod and substrate properties on tree alpha diversity and composition in
Amazonian. Plant Ecology 215 (10). doi:10.1007/s11258-014-0415-y.
Barnosky, A. D., Matzke, N., Tomiya, S., Wogan, G. O. U., Swartz, B., Quental,
T. B., Marshall, C., McGuire, J. L., Lindsey, E. L., Maguire, K. C., Mersey, B.,
Ferrer, E. (2011), Has the Earth’s sixth mass extinction already arrived? Nature
471: 51–57.
Chao, N. L.; Prang, G.; Petry, P. (2001), The Fishery diversity and conservation
of ornamental fishes in the Rio Negro Basin, Brasil: a review of Project Piaba
(1989-1999). In: Chao N. L., Prang, P., Sonnenschein, L. & Tlusty, M. T. (Eds.)
Conservation and Management of Ornamental Fish Resources of the Rio Negro
Basin, Amazônia, Brazil Project Piaba (pp. 161-204). EDUA Manaus.
Cianciaruso, M.V., Silva, I.A., & Batalha, M.A. (2009), Phylogenetic and
functional diversities: new approaches to community Ecology. Biota Neotropica
9(3): 93-103. http://www.biotaneotropica.org.br.
Couceiro, S. R. M., Forsberg, B. R., Hamada, N., Ferreira, R. L. M. (2006), Effects
of an oil spill and discharge of domestic sewage on the insect fauna of cururu
stream, Manaus, AM, Brazil. Brazilian Journal of Biology, 66(1A):35-44.
Diáz, S., & Cabido, M. (2001), Vive la différence: plant functional diversity matters
to ecosystem processes. Trends in Ecology & Evolution, 16: 646-655.
Downing, J. A, Plante, C., Lalonde, S. (1990), Fish production correlates with
primary productivity, not the morphoedaphic Index. Canadian Journal of Fisheries
and Aquatic Sciences. 47: 1929-1936.
Goulding, M., Carvalho, M. L., & Ferreira, E. J. G. (1988), Rio Negro, rich life in
poor water: Amazonian diversity and foodchain ecology as seen through fish
communities. The Hague, NED: SPB Academic Publishing.
Hutchinson, G. E. (1958), Concluding remarks. Cold Spring Harbor Symposia on
Quantitative Biology. 22: 415-427.
Hutchinson, G. E., & MacArthur, R. (1959), A theoretical ecological model of size
distributions among species of animals. The American Naturalist. 93: 117- 126.
Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis.
(2008), Diagnóstico geral das práticas de controle ligadas a exploração, captura,

7
comercialização, exportação e uso de peixes para fins ornamentais e de
aquariofilia. Brasília, DF: Diretoria de Uso Sustentável da Biodiversidade e
Florestas.
Irion, G. (1978), Soil infertility in the Amazonian rain forest. Naturwissenschaften
65:515–519.
Isaac, V. J., & Barthem, R. B. (1995), Os recursos pesqueiros da Amazônia
brasileira. Boletim do Museu Paraense Emílio Goeldi, série Antroplogia 11(2).
Junk, W. (1984), Ecology of the varzea of Amazonian White Water Rivers. In:
Sioli, H. (Eds.). The Amazon: Limnology and landscape ecology of a mighty
tropical river and its basin. (pp. 215-244) Dordrecht, W. Junk Publishers.
Junk W.J., Bayley P.B., Sparks R.E. (1989), The flood pulse concept in river-
floodplain systems. Canadian Journal of Fisheries and Aquatic Sciences
106:110–127.
Junk, W. J., & Wantzen K. M. (2004) The flood pulse concept: new aspects,
approaches and applications – na update. In: Welcomme, R. L., & Petr T., [Eds]
Proceedings of the second international symposium on the management of large
rivers of fisheries (pp. 117 – 149) vol. 2 Food and Agriculture Organization &
Mekong River Comission FAO Regional Office for Asia and the Pacific, Bangkok.
Rap Publication.
Junk, W. J., Piedade, M. T. F., Lourival, R., Wittmann, F., Kandus, P., Lacerda,
L. D., Bozelli, R. L., Esteves, F. A., Nunes da Cunha, C., Maltchik, L., Schongart,
J., Schaeffer-Novelli, Y., Agostinho, A. A. (2014), Brazilian wetlands: their
definition, delineation, and classification for research, sustainable management,
and protection. Aquatic Conservation Marine and Freshwater Ecossystems. 24:
5-22.
Laliberté, E., & Legendre, P. (2010), A distance-based framework for meansuring
functional diversity from multiple traits. Ecology, 91(1): 299–305.
Laliberté, E., Wells, J. A., DeClerck, F., Metcalfe, D. J., Catterall, C. P., Queiroz,
C., Aubin, I., Bonser, S. P., Ding, Y., Fraterrigo, J. M., McNamara, S., Morgan, J.
W., Merlos, D. S., Vesk, P. A., Mayfield, M. M. (2010), Land-use intensification
reduces functional redundancy and response diversity in plant communities.
Ecology Latters, 13: 76-86.
Leitão, R. P., Zuanon, J., Villéger, S., Williams, S. E., Baraloto, C., Fortunel, C.,
Mendonça, F. P., Mouillot, D. (2016), Rare species contribute disproportionately
to the functional struture of species assemblages. Proceedings Royal Society B
283: 20160084. http://dx.doi.org/10.1098/rspb.2016.0084.

8
Maguire jr., B. (1973), Niche response structure and the anajytical potentials ofits
relationship to the habitat. The American Naturalist l07 (954): 213-246.
Magurran, A.E. (2004), Measuring biological diversity. Blackwell, Oxford.
Moreno, C. E., Arita, H. T., & Solis, L.. (2006), Morphological assembly
mechanisms in Neotropical bat assemblages and ensembles within a landscape.
O ecologia, 149: 133-140.
Mouillot, D., Villéger, S., Scherer-Lorenzen, M., & Mason, N. W. H. (2011),
Functional structure of biological communities predicts ecosystem
multifunctionality. PLoS ONE 6:e17476.
Mouillot, D., Graham, N. A. J., Villéger, S., Mason, N. W. H., & Bellwood, D. R.
(2013), A functional approach reveals community responses to disturbances.
Trends in Ecology and Evolution 28:167–177.
Petchey, O.L., & Gaston, K.J. (2002), Functional diversity (FD), species richness
and community composition. Ecology Letters, 5: 402-411.
Petchey, O.L., & Gaston, K.J. (2006), Functional diversity: back to basics and
looking forward. Ecology Letters, 9(6):741-758.
Queiroz, L.J., Torrente-Vilara, G., Ohara, W.M., Pires, T.H.S., Zuanon, J., Doria,
C.R.C. (2013). Peixes do rio Madeira. Santo Antônio Energia, São Paulo.
Reis, R. E., Albert, J. S., Di Dario, F., Mincarone, M. M., Petry, P., Rocha, L. A.
(2016), Fish biodiversity and conservation in South America. Journal of Fish
Biology. 89 (1): 12-47.
Ricklefs, R. E. (2012), Species richness and morphological diversity of passerine
birds. Proceedings of the National Academy of Sciences, 109(36): 14482-14487.
Ricotta, C., Avena, G., & Chiarucci, A. (2005), Quantifying the effects of nutrient
addition on the taxonomic distinctness of serpentine vegetation. Plant Ecology
179(1):21-29.
Ricotta, C., Bello, F. de, Moretti, M., Caccianiga, M., Cerabolini, B. E. L., Pavoine,
S. (2016), Measuring the functional redundancy of biological communities: a
quantitative guide. Methods in Ecology and Evolution. 7: 1386-1395.
Santos, G. M., Ferreira, E. J. G., Zuanon, J. A. S. (2006), Peixes comerciais de
Manaus. Manaus: Ibama/AM, ProVárzea.
Souza, R. L. de, & Mendonça, M. R. de. (2009), Caracterização da pesca e dos
pescadores de peixes ornamentais da região de Tefé/AM. UAKARI. 5(2): 7-17.

9
Teresa, F. B. (2012), Diversidade funcional de comunidades de peixes de
riachos. (Tese de Doutorado não publicada) Programa de Pós-Graduação em
Biologia Animal – UNESP. São José do Rio Preto.
Teresa, F. B., & Casatti, L. (2012), Influence of forest cover and mesohabitats
types on functional and taxonomic diversity of fish communities in Neotropical
lowland streams. Ecology of Freshwater Fish. 21(3): 1-10. Doi: 10.1111/j.1600-
0633.2012.00562.x
Tilman, D. 2001. Functional diversity. In: Levin, S. A. (Eds.) Encyclopedia of
Biodiversity (pp. 109-120). Academic Press, San Diego.
Villéger, S., Mason, N. W. H., & Mouillot, D. (2008), New multidimensional
functional diversity indices for a multifaceted framework in functional ecology.
Ecology 89:2290–2301.
Vitouse, P. M., Mooney, H. A., Lubchneco, J., & Melillo, J.M. (1997), Human
domination of Earth’s ecosystems. Science. 277: 494-499
Webb, C.O. (2000), Exploring the phylogenetic structure of ecological
communities: an example for rain forest trees. The American Naturalist
156(1):145-155.
Whittaker, R. H., Levin, S. A., & Root, R. B. (1973), Niche, Habitat, and Ecotope.
The American Naturalist. 107(955): 321-338.

10
Contribuição de espécies ornamentais para a estrutura
funcional de assembleias de peixes de igarapés do médio rio
Negro, Amazônia brasileira
DANILO G. R. CASTANHO¹*, CLÁUDIA P. DE DEUS¹, JANSEN ZUANON¹, RAFAEL P. LEITÃO²,
FABRÍCIO B. TERESA³
¹Instituto Nacional de Pesquisas da Amazônia, Manaus, Brasil
²Universidade Federal de Minas Gerais, Belo Horizonte, Brasil
³Universidade Estadual de Goiás, Anápolis, Brasil
*Correspondência: E-mail: [email protected]
Resumo
Uma atividade extrativista só é considerada sustentável quando não interfere na
estabilidade e resiliência das comunidades biológicas e do ambiente explorado.
A pesca ornamental é uma das principais atividades econômicas do médio rio
Negro desde a década de 50, retirando milhões de exemplares anualmente. Tal
atividade é direcionada a um pequeno número de espécies e tem o Tetra cardinal
(Paracheirodon axelrodi) como principal representante. Visto os potenciais
efeitos da pressão exercida em um número reduzido de espécies e a falta de
informações sobre os impactos gerados, o presente estudo caracterizou a
estrutura funcional das assembleias de peixes de igarapés explotados pela
atividade e analisou, por meio de cenários hipotéticos, os possíveis efeitos da
depleção das populações de peixes ornamentais sobre a estrutura funcional
destas assembleias. Por meio da avaliação de três índices funcionais
(redundância funcional, divergência funcional e dispersão funcional)
constatamos que (1) a exclusão completa de todas as espécies ornamentais,
simultaneamente, pode diminuir em até 12% a redundância funcional das
assembleias estudadas; (2) a divergência funcional tende a aumentar com a
retirada de espécies ornamentais; e (3) a diversidade de respostas frente à
alterações ambientais, analisada através da dispersão funcional, não sofreria
alterações significativas com a exclusão das espécies ornamentais. Uma
comparação entre os efeitos potenciais da retirada do Tetra Cardinal versus
espécies não ornamentais indica que a retirada desta espécie ornamental possui
efeito específico, reduzindo em maior intensidade a redundância funcional das
assembleias. Com esses resultados é possível inferir que a pesca ornamental
na região do médio rio Negro necessita de manejo adequado, e poderia continuar
a ser realizada sem comprometer a estabilidade e resiliência das assembleias
de peixes e dos ambientes explotados.
Palavras chave: Diversidade Funcional; Ictiofauna; Pesca Ornamental; Igarapés; Amazônia

11
Introdução
Em um cenário de extinção de espécies comparado aos grandes eventos de
extinção em massa (Barnosky et al., 2011), uma preocupação difundida são as
consequências geradas na integridade e funcionamento dos ecossistemas.
Quando a perda de espécies é acompanhada pela perda de funções e serviços
ecossistêmicos, os prejuízos deixam de ser somente taxonômicos e afetam a
estabilidade das comunidades ou dos ecossistemas. O desempenho de funções
similares por espécies diferentes, garante a continuidade na execução de
funções ecossistêmicas mesmo com eventos de extinção, tal propriedade é
usualmente conhecida como redundância funcional (Ricotta et al., 2016).
Os ambientes de água doce concentram grande parte da biodiversidade global
e são facilmente afetados por mudanças ambientais (Taylor et al., 2006). Dentre
os veículos de extinção em ambientes aquáticos, a descaracterização do habitat
(Casatti, Ferreira, & Carvalho, 2009) e a sobre-explotação dos recursos estão
entre os principais. Realizada, na maioria das vezes, de maneira desordenada,
a pesca é um importante agente causador de declínio nos estoques
populacionais de organismos aquáticos, atingindo de macro invertebrados
(camarões) à peixes (Isaac & Barthem, 1995).
A pesca ornamental na Amazônia é uma atividade extrativista que retira de forma
seletiva milhões de exemplares de peixes anualmente (Anjos, Amorim, Siqueira,
& Anjos, 2009). A categorização de uma dada espécie como “ornamental”
envolve uma perspectiva antropocêntrica, pois o que torna uma espécie atrativa
ou não pode envolver aspectos da forma do corpo, padrão de colorido,
comportamento, ou uma combinação desses fatores. Do ponto de vista
ecológico, as espécies-alvo da pesca ornamental desempenham funções que
contribuem para a manutenção do ecossistema local. Entretanto, são escassas
as informações sobre as funções desempenhadas por essas espécies em seus
ambientes naturais, o que dificulta a avaliação das possíveis consequências
geradas pela sua retirada seletiva das comunidades biológicas. Um estudo
realizado por Gasparini, Floeter, Ferreira & Sazima (2005) evidenciou uma
redução na densidade de indivíduos jovens de peixes-anjo (Pomacanthidae),
potencialmente, decorrente da pesca ornamental. A espécie, em seu estágio

12
jovem, é associada a relação mutualística de “limpeza” à outras espécies de
peixes recifais e tem como instrumento de reconhecimento a coloração
conspícua, característica também associada ao interesse ornamental. A perda
dessa função ecossistêmica pode afetar a saúde da comunidade e até influenciar
a diversidade local (Floeter, Ferreira, & Gasparini, 2007).
A análise da estrutura funcional de assembleias biológicas permite relacionar
diretamente as espécies com determinadas funções ecológicas. Além disso,
essa abordagem permite a avaliação de respostas potenciais das espécies às
múltiplas pressões antrópicas sobre os ecossistemas. Por exemplo, estudos vêm
demostrando que o desmatamento da zona ripária favorece espécies detritívoras
e onívoras com maior tolerância a hipóxia (Teresa & Casatti, 2012). A predição
de potenciais respostas às alterações ambientais é de grande valia no
direcionamento de ações de manejo e na criação de áreas de proteção.
O potencial crescimento da pesca ornamental direcionada a um número reduzido
de espécies, e concentrada em uma área específica, gera preocupações em
relação as eventuais consequências no funcionamento do ecossistema em nível
local. O presente trabalho buscou analisar a estrutura funcional das assembleias
de peixes em áreas com histórico de pesca ornamental e avaliar o efeito da
explotação direcional ocasionada pela atividade.
Material e Métodos
Área de estudo
O presente estudo foi realizado em igarapés de terceira a quinta ordem,
historicamente explorados pela pesca ornamental. Apresentam água preta ou
clara, com largura variando de 5,7 a 19,2 m; profundidade média de 0,4 a 1,6 m;
abertura média de dossel variando de 14,6 a 100%; e velocidade da corrente de
0,06 a 0,32 m.s-1. A temperatura da água variou de 25 a 30.6°C; condutividade
de 0,017 a 0,114 S.s-1 e pH variando entre 4,02 e 5,27 (dados pessoais). Os
igarapés estudados pertencem às bacias dos rios Negro, Itu, Aracá e Cuiuni, e

13
estão localizados na região do município de Barcelos, estado do Amazonas
(figura 1).
A bacia do Rio Negro cobre uma área aproximada de 700.000 km² (Latrubesse,
2008), sendo 118.000 km² constituídos de áreas alagáveis (Melack & Hess,
2010). Essas incluem as planícies de inundação ao longo do rio principal e as
regiões de cabeceira localizadas na Amazônia colombiana, venezuelana e
brasileira. O rio Negro é o segundo maior afluente da bacia do rio Solimões-
Amazonas, ficando atrás, apenas, do rio Madeira. Sua ictiofauna ultrapassa a
marca de 1.000 espécies (Chao, Prang, & Petry, 2001) e abriga mais de 80%
dos peixes ornamentais comercializados no estado do Amazonas (Anjos et al.,
2009).
Figura 1. Área de estudo e locais de coleta (círculos pretos) na porção média da bacia do rio
Negro, região de Barcelos-AM (estrela cinza) (QGIS Development Team, 2016).
Delineamento experimental
Foram selecionados 13 igarapés, sendo que cinco deságuam diretamente no rio
Negro, quatro no rio Aracá, três no rio Itu e um no rio Cuiuni. Em cada igarapé
foi delimitado um trecho de 100 metros de extensão, em locais historicamente
utilizados pela pesca ornamental (definidos por meio de informações obtidas
diretamente com pescadores ornamentais de Barcelos). As amostragens foram

14
realizadas uma única vez, no período de vazante-seca, em abril de 2016. O
período de coleta foi selecionado por seu maior rendimento na pesca
ornamental.
Material ictiológico
As coletas de peixes foram conduzidas com base na metodologia estabelecida
pelo Projeto Igarapés (Mendonça et al., 2005), com modificações. Foram
estabelecidos trechos não bloqueados de 100 metros, onde os peixes foram
coletados com auxílio de rapiché (apetrecho semelhante a um puçá, largamente
utilizado pelos pescadores ornamentais da região do médio rio Negro. Figura
S1), rede de cerco, malhadeiras e cacuri (armadilha do tipo “covo”, com
orientação vertical, utilizada pelos pescadores ornamentais da região do médio
rio Negro. Figura S2).
Em cada igarapé amostrado foram utilizadas cinco malhadeiras de cinco metros
de comprimento, com malhas de 30, 50, 70, 90 e 120 mm entre nós opostos, por
um período de duas horas. No caso dos cacuris, foram utilizadas três unidades
por um período de duas horas. O esforço de coleta utilizando rapiché foi
padronizado como um coletor por um período de uma hora. As amostragens com
rede de cerco se deram por meio de um arrasto por trecho, utilizando uma rede
de 11 m de comprimento e malha de 2 mm entre nós adjacentes.
Todos os exemplares capturados foram eutanasiados com uma dose letal de
anestésico (Eugenol), fixados em formalina a 10%, e transferidos para solução
alcóolica (etanol 70%), para posterior identificação, pesagem, contagem e
realização das medidas ecomorfológicas.
Estrutura funcional das assembleias de peixes
Para mensurar a diversidade e a estrutura funcional das assembleias,
primeiramente foi realizada uma análise ecomorfológica de todas as espécies
coletadas nos igarapés. Para tal, foram tomadas 21 medidas ecomorfológicas de
cada exemplar, obtidas diretamente dos exemplares, com uso de paquímetro,
balança e estereomicroscópio (para classificação de dentes e rastros
branquiais), e, também, por meio de medidas ponto a ponto obtidas em imagens
fotográficas dos mesmos exemplares (Figura S3). Sempre que possível, as
medidas foram tomadas em dez exemplares adultos por espécie; para aquelas

15
que não atingiram a abundância de dez indivíduos, foram mensurados todos os
exemplares adultos capturados (variando de 1 a 9 exemplares). As medidas
foram combinadas em 16 atributos funcionais que refletem aspectos da
locomoção, forrageamento e uso do habitat (Villéger, Miranda, Hernândez, &
Mouillot, 2010; Leitão et al., 2016) (Tabela S1). Tais aspectos representam o
potencial nicho ecológico ocupado pelas espécies, e estão intimamente ligados
às respostas das espécies frente a alterações no ambiente (Ribeiro, Teresa, &
Casatti, 2016; Teresa & Casatti, 2017)
A possível existência de colinearidade entre os atributos funcionais foi testada
através de correlações de Pearson; e como não houve valores do coeficiente de
correlação acima de 0,65, todos foram utilizados nas análises. A partir dos
valores dos atributos foi calculada uma matriz de distância de Gower e, em
seguida, realizada uma Análise de Coordenadas Principais (PCoA). As
coordenadas de cada espécie nos primeiros quatro eixos da PCoA foram
utilizadas para construir o espaço funcional multidimensional. A seleção dos
quatro primeiros eixos se baseou na qualidade do espaço funcional, estimada
pelo protocolo proposto por Maire et al. (2015). Com isso, foi selecionada a
quantidade mínima de eixos que representasse fidedignamente as distâncias
funcionais iniciais entre pares de espécies (Figura S4).
Para representar a estrutura funcional das assembleias, utilizamos os índices FR
(Functional Redundancy – Redundância funcional), FDiv (Functional Divergence
- Divergência funcional) e FDis (Functional Dispersion - Dispersão funcional). O
índice FR reflete a coocorrência de espécies funcionalmente idênticas ou
similares. Este é obtido através da diferença entre o índice de diversidade de
Simpson, baseado na faceta taxonômica da diversidade, e o índice de
diversidade quadrática de Rao, que utiliza a diversidade dos atributos funcionais
(de Bello, Lep, Lavorel, & Moretti, 2007). Dessa forma, uma assembleia com
espécies totalmente diferentes, em termos funcionais, possui valores de FR
tendendo a zero. O contrário se espera em uma comunidade com espécies
totalmente redundantes em termos funcionais, onde FR tende ao valor de um.
Dessa forma, valores mais próximos a um indicam uma assembleia mais
redundante e valores mais próximos a zero indicam uma assembleia com baixa
redundância.

16
O índice FDiv retrata o desvio das espécies na distância média da comunidade
em relação ao centroide do espaço multidimensional (Villéger, Mason, & Mouillot,
2008). Levando em consideração as abundâncias, pode-se analisar se as
espécies mais próximas à média (ou mais comuns em termos de padrões
ecomorfológicos), são também as mais abundantes, o que faz os valores se
aproximarem de zero. Quando as espécies mais distantes à média apresentam
maior abundância, a tendência dos resultados é se aproximar de 1. Neste caso,
infere-se que comunidades com altos valores de FDiv apresentam maior
diferenciação na utilização dos recursos, uma vez que as características
mensuradas são relacionadas à utilização do ambiente.
E por fim, o índice FDis representa o quão distantes as espécies estão da média
dos atributos funcionais da comunidade, deste modo, valores elevados indicam
elevada dispersão de atributos funcionais na comunidade e valores baixos
indicam baixa dispersão, padrão agregado de atributos funcionais na
comunidade. FDis é calculado através da média das distâncias de cada espécie
em relação ao centroide do espaço multidimensional. Diferente do índice FDiv,
nesse caso, o centroide é estimado através das posições das espécies e suas
abundâncias. Desse modo, espécies com maiores abundâncias possuem maior
influência no deslocamento do centroide. FDis é pouco influenciado pela riqueza
de espécies, sendo que a abundância das espécies tem maior peso para este
cálculo (Villéger et al., 2008; Laliberté & Legendre, 2010; Colzoni, 2013; Meira
Junior, 2015). Desse modo, pode ser associado à diversidade de respostas de
uma comunidade frente a alterações ambientais (Laliberté et al., 2010).
Análises estatísticas
Para a decisão de quais espécies seriam consideradas como “ornamentais”, e,
portanto, avaliadas diferenciadamente no estudo, foi confeccionada uma lista
com as espécies presentes na Instrução Normativa IN 001/2012 do Ministério da
Pesca e Aquicultura e Ministério do Meio Ambiente; a lista apresentada no
estudo de Anjos et al. (2009), e espécies citadas pelos pescadores ornamentais
da região de Barcelos (observações pessoais dos autores).

17
Para analisar a contribuição das espécies ornamentais na estrutura funcional das
assembleias, foram criados dois cenários:
Cenário 1. Neste cenário, todas as espécies categorizadas como “ornamentais”
presentes em cada um dos igarapés foram selecionadas e tiveram suas
abundâncias subtraídas em intervalos de 5%, até o limite de 100%, simulando
um aumento da pesca ornamental. A fim de testar o efeito da subtração das
abundâncias foram realizadas duas análises: A) Foram comparados através do
teste de Wilcoxon para amostras pareadas os resultados obtidos nos índices
funcionais com as assembleias em condições originais e com 100% das
abundâncias das espécies ornamentais de cada igarapé subtraídas. B) Para
verificar se a subtração das espécies ornamentais ocasiona um efeito específico,
foram selecionadas de forma aleatória através da função Sample do programa
R (R Core Team, 2016) um número de espécies “não-ornamentais” (espécies
que não se enquadraram na categoria “ornamental”) igual ao número de
espécies ornamentais de cada igarapé. As espécies selecionadas tiveram suas
abundâncias subtraídas no intervalo de 5% até o limite de 100%. Os resultados
obtidos nos índices funcionais no gradiente de subtração das abundâncias de
espécies “ornamentais” e “não ornamentais” foram comparados através do teste
de Wilcoxon para amostras pareadas.
Todas as análises foram realizadas individualmente em cada igarapé, dessa
forma os 13 igarapés foram utilizados como réplicas.
Cenário 2. Como a principal espécie explorada pela pesca ornamental é o Tetra
cardinal, Paracheirodon axelrodi (Schultz, 1956), foram avaliados os possíveis
impactos gerados pela redução da abundância dessa espécie nos igarapés.
Para isso, foram selecionados os igarapés onde a espécie ocorreu (n= 9) e sua
abundância foi subtraída em intervalos de 5%, até o limite de 100%,
individualmente em cada igarapé. A fim de testar o efeito da subtração das
abundâncias foram realizadas duas análises: A) Foram comparados através do
teste de Wilcoxon para amostras pareadas os resultados obtidos nos índices
funcionais com as assembleias em condições originais e com 100% da
abundância do Tetra cardinal de cada igarapé subtraída. B) Para verificar se a
subtração dessa espécie ocasiona um efeito específico, foi selecionada de forma
aleatória através da função Sample do programa R (R Core Team, 2016) uma

18
espécie “não-ornamental” (espécie que não se enquadrou na categoria
“ornamental”) presente em cada igarapé. As espécies selecionadas tiveram suas
abundâncias subtraídas no intervalo de 5% até o limite de 100%. Os resultados
obtidos nos índices funcionais no gradiente de subtração das abundâncias do
Tetra cardinal e de espécies “não ornamentais” foram comparados através do
teste de Wilcoxon para amostras pareadas.
Todas as análises foram realizadas individualmente em cada igarapé, dessa
forma os 13 igarapés foram utilizados como réplicas.
Hemigrammus vorderwinkleri apresentou abundância 224% maior que a
segunda espécie mais abundante (Hemigrammus analis). Em um único evento
de coleta foi capturado cerca de 50% de sua abundância total, o que aparenta
ser efeito da captura de um cardume, influenciando a amostragem e, não
representando de fato a proporcionalidade real entre as espécies. Assim, a fim
de evitar tendências errôneas nos cálculos dos índices, essa espécie foi excluída
das análises.
Resultados
Estrutura taxonômica e proporção de espécies ornamentais nas
assembleias
Foram capturados 4.286 peixes, pertencentes a sete ordens, 24 famílias e 110
espécies. A ordem mais abundante foi Characiformes (89,92 % dos exemplares
coletados), seguida por Perciformes (5,62 %) e, Siluriformes (2,70 %). A família
Characidae foi a mais representativa, com 34 espécies e 3.147 exemplares,
representando 73,42% do total de peixes amostrados.
Das 110 espécies coletadas, 22 foram classificadas como ornamentais, sendo
sete da família Cichlidae, sete Lebiasinidae, cinco Characidae, duas
Gasteropelecidae e uma Potamotrygonidae, totalizando 1.148 exemplares
(Tabela S3). Em todos os igarapés amostrados foi encontrada pelo menos uma
espécie ornamental.

19
Dentre as espécies ornamentais, o Tetra cardinal (Paracheirodon axelrodi)
apresentou a maior abundância (396 exemplares) e a maior frequência de
ocorrência (9 dos 13 igarapés). Carnegiella marthae foi a segunda espécie
ornamental mais importante, com 297 exemplares e ocorrência em quatro
igarapés. Hemigrammus analis foi a espécie mais representativa fora da
categoria ornamental, com 492 indivíduos coletados, seguida de H.bellottii, com
275 indivíduos.
Estrutura funcional das assembleias
Considerando o total de espécies amostradas nos 13 igarapés (110), 13,11% do
volume do espaço funcional foi preenchido pelas 22 espécies enquadradas na
categoria ornamental (figura 2).

20
Figura 2. Volume ocupado pelas espécies categorizadas como ornamentais no espaço
multidimensional considerando o total de espécies amostradas. Polígono branco: volume
ocupado pelo total de espécies amostradas; polígono rosa: volume ocupado por espécies
ornamentais; círculos brancos: espécies não ornamentais; círculos pretos: espécies
ornamentais.
Os valores do índice de Divergência Funcional (FDiv) foram altos em todos os
igarapés, variando de 0,53 a 0,91. Foi observado no índice de Dispersão
Funcional (FDis) valores variando de 0,05 a 0,14. A Redundância Funcional (FR)
total das assembleias variou de 0,15 no igarapé Zalala, o qual apresentou 11
espécies, a 0,50 no igarapé Mamulé, com 17 espécies amostradas. O igarapé

21
Bui-Bui apresentou o maior número de espécies (34) e redundância funcional de
0,47. A maior concentração de ornamentais foi encontrada no igarapé Daracuá
(10) com redundância funcional de 0,44 (tabela S5).
Efeito da retirada de espécies ornamentais na estrutura funcional
das assembleias (Análise A)
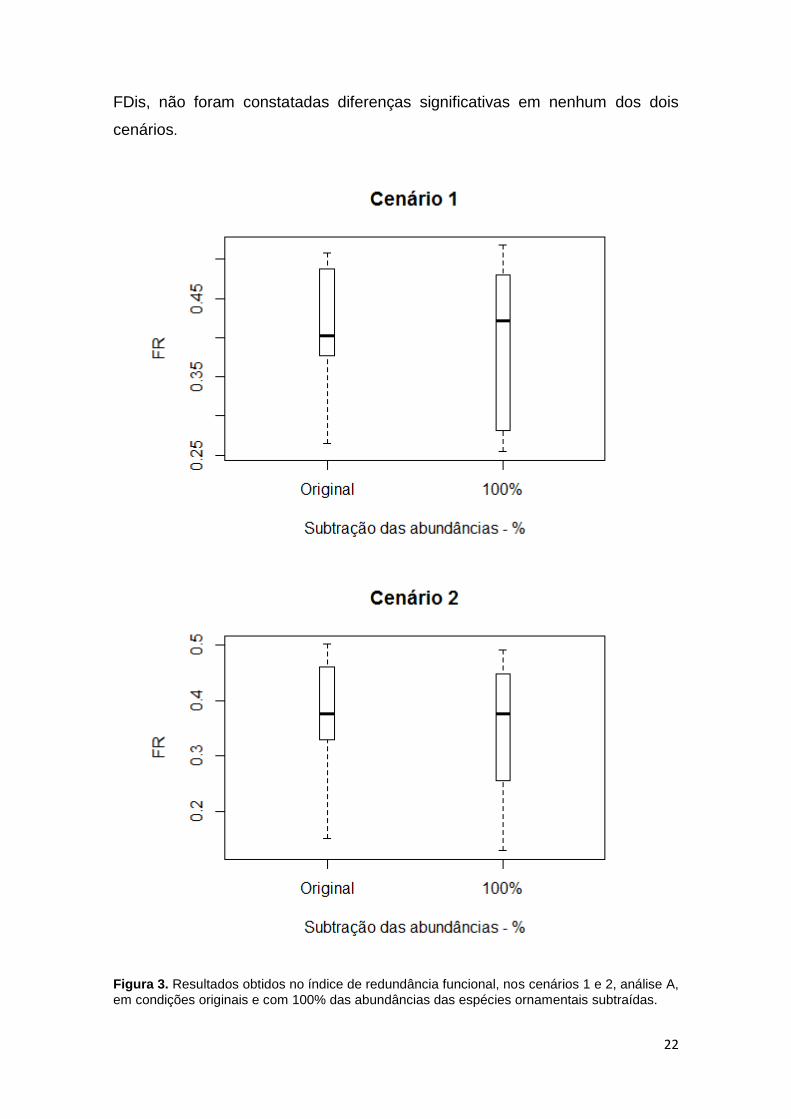
O teste de comparação de medianas mostrou diminuição significativa nos
valores de redundância funcional (FR) das assembleias nos cenários 1 (P= 0,03)
e 2 (P= 0,003), para análise A (figura 3). A queda média nos valores do índice
foi de 3,19% no cenário 1, para os 13 igarapés. A partir da subtração de 35%
das abundâncias do cardinal, foi possível constatar diminuição significativa nos
valores de FR no cenário 2 (P= 0,042). No cenário 1, não houve diferenças
significativas nos resultados de FDiv, com exclusão completa das espécies
ornamentais. Já no cenário 2, foi constatado aumento significativo nos valores
de FDiv a partir da retirada inicial de 5% das abundâncias do cardinal, P. axelrodi
(P= 0,036) até 95% (P=0,027). Com a exclusão completa da espécie o teste de
comparação de medianas mostrou valor de P= 0,054 (figura 4). Para o índice
22
FDis, não foram constatadas diferenças significativas em nenhum dos dois
cenários.
Figura 3. Resultados obtidos no índice de redundância funcional, nos cenários 1 e 2, análise A,
em condições originais e com 100% das abundâncias das espécies ornamentais subtraídas.

23
Figura 4. Resultados obtidos no índice de divergência funcional, no cenário 2, análise A, em diferentes porcentagens de subtração de abundâncias de P. axelrodi.
Comparação do efeito gerado pela subtração das abundâncias de
espécies ornamentais e não ornamentais na estrutura funcional
das assembleias (Análise B)
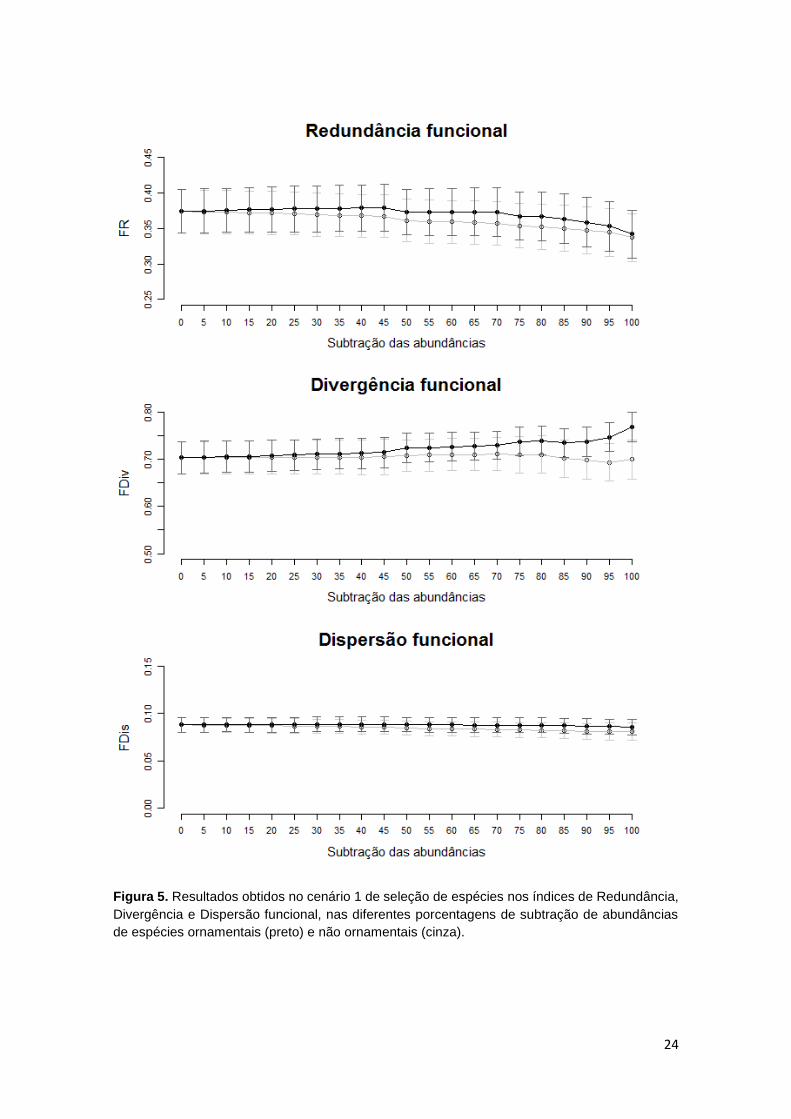
No cenário 1, análise B, onde todas as espécies ornamentais de cada igarapé
tiveram suas abundâncias subtraídas no intervalo de 5%, a perda de
redundância funcional foi significativamente menor do que quando comparada
com a subtração das abundâncias de espécies não ornamentais (P< 0,001.
Figura 5). A dispersão e a divergência funcional tiveram um aumento
significativamente maior com a retirada de espécies ornamentais do que com a
retirada de espécies não ornamentais (P< 0,001 em ambos os índices. Figura 5).
24
Figura 5. Resultados obtidos no cenário 1 de seleção de espécies nos índices de Redundância,
Divergência e Dispersão funcional, nas diferentes porcentagens de subtração de abundâncias
de espécies ornamentais (preto) e não ornamentais (cinza).

25
O teste de comparação de medianas mostrou para o cenário 2, análise B,
(subtração das abundâncias de P. axelrodi) uma perda de redundância funcional
maior do que para a remoção de espécies não ornamentais (P< 0,001. Figura 6).
FDiv e FDis apresentaram comportamentos semelhantes ao cenário 1, onde a
subtração das abundâncias de espécies ornamentais gerou um aumento maior
que a subtração das abundâncias de espécies não ornamentais (figura 6).

26
Figura 6. Resultados obtidos no cenário 2 de seleção de espécies nos índices de Redundância,
Divergência e Dispersão funcional, nas diferentes porcentagens de subtração de abundâncias
de P. axelrodi (preto) e espécies não ornamentais (cinza).

27
Discussão
O presente estudo revelou que as espécies categorizadas como ornamentais
exploram o ambiente e utilizam os recursos de forma similar à maioria das
espécies amostradas, proporcionando maior redundância à estrutura funcional
das assembleias de peixes em igarapés explorados pela pesca ornamental, na
porção média da bacia do rio Negro. Segundo a hipótese da garantia (Insurance
hypothesis, Yachi & Loreau, 1999), assembleias com maior redundância
funcional são menos vulneráveis às alterações ambientais. Neste sentido, a fim
de garantir maior resiliência aos ambientes e comunidades biológicas explotadas
pela pesca ornamental, é fundamental que haja um ordenamento da atividade e
a manutenção dos estoques dessas espécies (Naeem & Li, 1997; Yachi &
Loreau, 1999, Laliberté et al., 2010).
Embora a perda de redundância funcional seja, em média, de apenas 3,192%
com a retirada completa das espécies ornamentais nos igarapés, em alguns
ambientes esse valor ultrapassa os 12%, como é o caso do igarapé Daracuá. Ao
compararmos a condição original e com todas as espécies ornamentais
excluídas, existe uma queda significativa de redundância.
Numerosos estudos apontam a redução de redundância funcional como um dos
mecanismos causadores de aumento da vulnerabilidade de comunidades
biológicas e ecossistemas (Joner, 2008; Jonsson, 2011; Laliberté et al., 2010).
Uma assembleia em que diversas espécies desempenham a mesma função é,
teoricamente, mais resistente às mudanças estruturais do que assembleias em
que as funções são desempenhadas por uma ou poucas espécies (Fonseca &
Ganade, 2001). A garantia de que uma determinada função ecológica estará
presente em uma assembleia, mesmo com a extinção de uma espécie, é refletida
na estabilidade do ecossistema, a qual é relacionada à capacidade do
ecossistema de evitar qualquer alteração (resistência) e a habilidade de retornar
ao estado prévio após sofrer alguma alteração (resiliência) (Harrison, 1979;
Carpenter, Walker, Anderies, & Abel, 2001).
Através da comparação com a retirada de espécies não ornamentais, foi
constatado que a exclusão do Tetra cardinal (P. axelrodi) possui efeito específico
na redundância funcional das assembleias. Por apresentar grande abundância,

28
explorar os recursos comumente encontrados nos igarapés do médio rio Negro
e utilizar o ambiente de forma semelhante à maioria das espécies da família
Characidae (Walker, 2004), o cardinal contribui significativamente com a garantia
de desempenho de serviços ecológicos, favorecendo a resiliência do
ecossistema.
O presente estudo foi baseado em um conjunto de características morfológicas
relacionadas à utilização do ambiente e exploração de recursos, não tendo
incluído características comportamentais, relações filogenéticas ou padrões de
coloração. Apesar do cardinal apresentar uma morfologia “generalizada” nas
assembleias amostradas, a espécie apresenta um padrão único de coloração.
Entretanto, a função eventualmente exercida por essa propriedade, presente em
ambos os sexos, ainda é desconhecida. Via de regra, toda característica fixada
em uma população possui valor adaptativo positivo ou neutro.
Estudos realizados com ciclídeos africanos mostram que o padrão de coloração
é um importante mecanismo de reconhecimento intraespecífico, funcionando
como isolamento reprodutivo entre espécies próximas vivendo em sintopia
(Maan & Sefc, 2013). A coloração pode ser relacionada, também, à pressão de
predação, como em Poecilia reticulata, onde a coloração conspícua atua de
forma negativa, aumentando as taxas de predação (Magurran, 2005), e ao
reconhecimento de espécies que desempenham relações mutualísticas de
“limpeza” à outras espécies de peixes em recifes de coral (Gasparini et al., 2005).
O comportamento social e o forrageamento são outros aspectos relacionados
com os padrões de coloração, onde certas características demostram
dominância e favorecimento na utilização de recursos alimentares (Ochi &
Awata, 2009).
Os valores originalmente obtidos na dispersão funcional (FDis), em ambos os
cenários, são reflexos da concentração de espécies próximas ao centroide. Com
a elevada abundância concentrada nas espécies com morfologia “generalizada”
(morfotipo semelhante a maioria das espécies amostradas), o centroide é
posicionado a curtas distâncias da maioria das espécies, aproximando de zero
os valores do índice. Isso explica por que em ambos os cenários de seleção de
espécie não houve variações significativas nos valores de FDis com a subtração
das abundâncias de espécies ornamentais. Com atributos ecológicos similares

29
a maioria das espécies, as ornamentais se concentraram, em sua maior parte,
no centro do espaço multidimensional, colaborando em pequenas proporções
com a diversidade de respostas a possíveis modificações ambientais. O
posicionamento das espécies e a distribuição das abundâncias fizeram com que
os valores de FDis aumentassem com a retirada das espécies ornamentais, isso
ocorre devido ao distanciamento do centroide em relação a grupos de espécies
não ornamentais próximas às ornamentais.
Quando a retirada de espécies ornamentais é comparada com a retirada de
espécies não ornamentais (análise B), há um incremento significativo na
divergência funcional das assembleias, nos dois cenários de seleção de
espécies. Como a FDiv é obtida através do desvio das espécies na distância
média da comunidade em relação ao centroide do espaço funcional, e a
abundância é considerada, a retirada de espécies ornamentais abundantes e
próximas a distância média da comunidade, aumenta os valores desses índices.
Este aumento está ligado, provavelmente, à retirada de grandes quantidades de
P. axelrodi e Carnegiella marthae, ambas espécies ornamentais com grande
peso no cálculo do índice, devido sua elevada abundância e proximidade à
distância média da assembleia. Sob esse cenário, espécies mais divergentes e
menos abundantes, como Potamotrygon wallacei e Trichomycterus hasemani,
ganham relevância no cálculo do índice.
Mesmo com baixas taxas de produtividade primária e baixa concentração de
nutrientes em suas águas, a região do médio rio Negro apresenta elevada
riqueza de espécies de peixes (Chao et al., 2001; Goulding, Carvalho, & Ferreira,
1988) e sustenta uma intensa pesca ornamental. A considerável diferenciação
na utilização do ambiente e na exploração de recursos, evidenciada pelos altos
valores de divergência funcional, é um aspecto importante que permite a
coexistência de uma grande diversidade de espécies. A exemplo disso, estudos
realizados no lago Victoria, no leste africano, sugerem que a grande riqueza de
espécies de ciclídeos do gênero Hoplachromis (aproximadamente 500 espécies)
é viabilizada, em partes, pelas diferentes formas e períodos de uso do habitat e
exploração de recursos alimentares (Meyer, Verheyen, Salzburger, & Smoeks,
2003).

30
Considerações finais
Na região do médio rio Negro, a pesca ornamental explora espécies com baixo
valor monetário individual; com isso, para se tornar rentável, os pescadores
necessitam capturar grandes quantidades de indivíduos dessas espécies,
diferentemente da pesca ornamental realizada no rio Xingu, por exemplo, onde
a comercialização de poucos peixes de elevado valor individual é rentável.
A captura de grandes quantidades de indivíduos induz a uma queda nos
estoques populacionais, inviabilizando as expedições de pesca do médio rio
Negro. Devido a inviabilidade econômica, os pescadores são forçados a
substituir o ponto de pesca, com isso, as chances de extinção local das espécies
ornamentais são diminuídas.
A pesca ornamental, realizada de maneira ordenada e com manejo adequado
dos estoques, tem condições propícias para se manter viável e sem alterar a
estabilidade do ecossistema, uma vez que as assembleias apresentam
considerável redundância, na maioria dos casos, não suprime funções
ecológicas, e a atividade não gera alterações significativas nas condições físicas
e químicas do ambiente.
Referências bibliográficas
Anjos, H.D.B. dos, Amorim, R.M.S. de, Siqueira, J.A., & Anjos, C. R. dos. (2009),
Exportação de peixes ornamentais do estado do Amazonas, bacia amazônica,
Brasil. Instituto de Pesca, São Paulo, Brasil, 35 (2): 259-274.
Barnosky, A. D., Matzke, N., Tomiya, S., Wogan, G. O. U., Swartz, B., Quental,
T. B., Marshall, C., McGuire, J. L., Lindsey, E. L., Maguire, K. C., Mersey, B.,
Ferrer, E. (2011), Has the Earth’s sixth mass extinction already arrived? Nature
471: 51–57.
de Bello, F., Lep_s, J., Lavorel, S., & Moretti, M. (2007), Importance of species
abundance for assessment of trait composition: an example based on pollinator
communities. Community Ecology. 8: 163–170.
Carpenter, S. Walker, B. Anderies, J.M., & Abel, N. (2001), From metaphor to
measurement: resilience of what to what? Ecosystems 4: 765-781.

31
Casatti, L., Ferreira, C. P. de, & Carvalho, F. R. (2009), Grass-dominated stream
sites exhibit low fish species diversity and dominance by guppies: an assessment
of tropical pasture river basins. Hydrobiologia. 632: 273-283.
Chao, N. L.; Prang, G.; Petry, P. (2001), The Fishery diversity and conservation
of ornamental fishes in the Rio Negro Basin, Brasil: a review of Project Piaba
(1989-1999). In: Chao N. L., Prang, P., Sonnenschein, L. & Tlusty, M. T. (Eds.)
Conservation and Management of Ornamental Fish Resources of the Rio Negro
Basin, Amazônia, Brazil Project Piaba (pp. 161-204). EDUA Manaus.
Colzoni, E. (2013), Análises funcionais, métodos estatísticos para se mensurar
a diversidade funcional. Disponível em:
http://diversidadefuncionaldeinvertebrados.blogspot.com.br. Acesso em: 07,
novembro de 2016.
Figuerola, J., Muñoz, E., Gutiérrez, R, & Ferrer, D. (1999), Blood parasites,
leucocytes and plumage brightness in the Cirl Bunting, Emberiza cirlus.
Functional Ecology. 13: 594-601.
Floeter, S. R., Ferreira, C. E. L., & Gasparini, J. (2007), Os efeitos da pesca e da
proteção através de UC’s marinhas: Três estudos de caso e implicações para os
grupos funcionais de peixes recifais no Brasil. Áreas Aquáticas Protegidas Como
Instrumento de Gestão Pesqueira. Ministério do Meio Ambiente, Brasília, pp
183–194.
Fonseca, C. R., & Ganade, G. (2001), Functional redundancy, random
extinctions and the stability of ecosystems. Journal of Ecology. 89: 118-125.
Gasparini, J. L., Floeter, S. R., Ferreira, C. E. L., & Sazima, I. (2005), Marine
ornamental trade in Brazil. Biodiversity and Conservation. 14: 2883–2899.
Goulding, M., Carvalho, M. L., & Ferreira, E. J. G. (1988), Rio Negro, rich life in
poor water: Amazonian diversity and foodchain ecology as seen through fish
communities. The Hague, NED: SPB Academic Publishing.
Harrison, G.W. (1979), Stability under environmental stress: resistance,
resilience, persistence, and variability. The American Naturalist . 113: 659-669.
Isaac, V. J., & Barthem, R. B. (1995), Os recursos pesqueiros da Amazônia
brasileira. Boletim do Museu Paraense Emílio Goeldi, série Antroplogia 11(2).
Joner, F. (2008), Redundância funcional em comunidades campestres.
(Dissertação de Mestrado não publicada) Programa de Pós-graduação em
Ecologia, UFRGS.

32
Jonsson, M. (2011), Perda de Biodiversidade e Funcionamento dos
Ecossistemas. ECOLOGIA.INFO 30.
Laliberté, E., & Legendre, P. (2010), A distance-based framework for meansuring
functional diversity from multiple traits. Ecology. 91(1): 299–305.
Laliberté, E., Wells, J. A., DeClerck, F., Metcalfe, D. J., Catterall, C. P., Queiroz,
C., Aubin, I., Bonser, S. P., Ding, Y., Fraterrigo, J. M., McNamara, S., Morgan, J.
W., Merlos, D. S., Vesk, P. A., Mayfield, M. M. (2010), Land-use intensification
reduces functional redundancy and response diversity in plant communities.
Ecology Latters, 13: 76-86.
Latrubesse, E.M. (2008), Patterns of anabranching channels: the ultimate
end-member adjustment of mega rivers. Geomorphology. 101: 130–145.
Leitão, R. P. (2015) Estrutura Funcional e Conservação de Assembleias de
Peixes de Riachos na Amazônia Brasileira. (Tese de Doutorado não publicada)
Programa de Pós-Graduação em Biologia de Água Doce e Pesca Interior –
Instituto Nacional de Pesquisas da Amazônia. Manaus. 146p.
Leitão, R. P., Zuanon, J., Villéger, S., Williams, S. E., Baraloto, C., Fortunel, C.,
Mendonça, F. P., Mouillot, D. (2016), Rare species contribute disproportionately
to the functional structure of species assemblages. Proceedings Royal Society B
283: 20160084. http://dx.doi.org/10.1098/rspb.2016.0084.
Maan, M. E., & Cumming, M. E. (2012), Poison Frog Colors Are Honest Signals
of Toxicity, Particularly for Bird Predators. The American Naturalist. 179(1): E1-
E14.
Maan, M. E., & Sefc, K. M. (2013), Colour variation in cichlid fish: Developmental
mechanisms, selective pressures and evolutionary consequences. Seminars in
Cell & Developmental Biology. 24(6-7): 516-528.
Magurran, A. E. (2005), Evolutionary ecology: the Trinidadian guppy. Oxford University Press, Oxford. Maire E., Grenouillet, G., Brosse, S., & Villéger, S. (2015), How many dimensions are needed to accurately assess functional diversity? A pragmatic approach for assessing the quality of functional spaces. Global Ecology and Biogeography. 24: 728–740. doi:10.1111/geb.12299 Meira Junior, M. S. de (2015), Efeitos da diversidade funcional na produção
florestal: estudo de caso no cerrado sentido restrito. (Dissertação de Mestrado
não publicada) Programa de Pós-graduação em Ciências Florestais –
Departamento de Engenharia Florestal, UNB.
Melack, J.M., & Hess, L.L. (2010), Remote sensing of the distribution and extent
of wetlands in the Amazon basin. In Junk W.J., Piedade M.T.F., Wittmann F.,

33
Schöngart J., Parolin P. (Eds.) Amazon floodplain forests: Ecophysiology,
biodiversity and sustainable management. (pp. 43-59) Springer, Ecological
Studies.
Mendonça, F.P., Magnusson, W.E., & Zuanon, J. (2005), Relationships Between
Habitat Characteristics and Fish Assemblages in Small Streams of Central
Amazonia. Copeia, 4: 750.
Meyer, A., Verheyen, E., Salzburger, W., & Snoeks, J. (2003), Origin of the
Superflock of Cichlid Fishes from Lake Victoria, East Africa. Science. 300(5617):
325-329
Mouillot, D., Dumay, O., & Tomasini, J. A. (2006), Limiting similarity, niche filtering
and functional diversity in coastal lagoon fish communities. Estuarine, Coastal
and Shelf Science. 71: 443 - 456
Mouillot, D., Graham, N. A. J., Villéger, S., Mason, N. W. H., & Bellwood, D. R.
(2013), A functional approach reveals community responses to disturbances.
Trends in Ecology and Evolution 28:167–177.
Naeem, S., & Li, S. (1997), Biodiversity enhances ecosystem
reliability. Nature. 390: 507-509.
Ochi, H., & Awata, S. (2009), Resembling the juvenile colour of host cichlid
facilitates access of the guest cichlid to host territory. Behaviour. 146: 741–756
Pagotto, J. P. A. (2014), Padrões ecomorfológicos de assembleias de peixes em
córregos da bacia do rio Pirapó, região norte do Paraná, Brasil. (Tese de
Doutorado não publicada) Programa de Pós-Graduação em Ecologia de
Ambientes Aquáticos Continentais - Universidade Estadual de Maringá, Maringá.
85p.
R Core Team (2016), R: A language and environment for statistical computing.
R Foundation for Statistical Computing (www.R-project.org).
Ribeiro, M. D., Teresa, F. B., & Casatti, L. (2016), Use of functional traits to
assess changes in stream fish assemblages across a habitat gradiente.
Neotropical Ichthyology. 14(1): e140185
Ricotta, C., Bello, F. de, Moretti, M., Caccianiga, M., Cerabolini, B. E. L., Pavoine,
S. (2016), Measuring the functional redundancy of biological communities: a
quantitative guide. Methods in Ecology and Evolution. 7: 1386-1395.
Summers, K., & Clough, M. E. (2001), The evolution of coloration and toxicity in
the poison frog family (Dendrobatidae). PNAS. 98(11): 6227-6232.

34
Taylor, B. W., Flecker, A. S., & Hall Jr., R. O. (2006), Loss of a Harvested Fish
Species Disrupts Carbon Flow in a Diverse Tropical River. Science. 313: 833-
836. doi: 10.1126/Science. 1128223
Teresa, F. B. (2012), Diversidade funcional de comunidades de peixes de
riachos. (Tese de Doutorado não publicada) Programa de Pós-Graduação em
Biologia Animal – UNESP. São José do Rio Preto.
Teresa, F. B., & Casatti, L. (2012), Influence of forest cover and mesohabitats
types on functional and taxonomic diversity of fish communities in Neotropical
lowland streams. Ecology of Freshwater Fish. 21(3): 1-10. Doi: 10.1111/j.1600-
0633.2012.00562.x
Teresa, F. B., & Casatti, L. (2017), Trait-based metrics as bioindicators:
Responses of stream fish assemblages to a gradiente of environmental
degradation. Ecological Indicators. 75: 249-258.
Villéger, S., N. W. H. Mason, & D. Mouillot. (2008), New multidimensional
functional diversity indices for a multifaceted framework in functional ecology.
Ecology 89:2290–2301.
Villéger, S., Miranda, J. R., Hernândez, D. F., & Mouillot, D. (2010), Contrasting
changes in taxonomic vs. functional diversity of tropical fish communities after
habitat degradation. Ecological Applications, 20(6): 1512–1522.
Yachi, S., & Loreau, M. (1999), Biodiversity and ecosystem productivity in a
fluctuating environment: The insurance hypothesis. Proceedings of the National
Academy of Sciences of the United States of America. 96(4): 1463-1468.
Walker, I. (2004), The food spectrum of the cardinal – tetra (Paracheirodon
axelrodi; Characidae) in its natural habitat. Acta Amazônica. 34(1): 69-73.

35
Material suplementar
Figura S1. Fotografia de um rapiché, aparelho de coleta de peixes ornamentais típico da região do médio rio Negro. Fonte: Danilo G. R. Castanho
Figura S2. Vista lateral de um cacuri, tipo de armadilha para peixes ornamentais utilizada na região do médio rio Negro. Fonte: Danilo G. R. Castanho.

36
Figura S3. Esquema das medidas ecomorfológicas obtidas com auxílio de paquímetro digital e análise fotográfica, através do programa computacional Image J. Siglas: Cp-comprimento padrão, Amc-altura máxima do corpo, Aco-Altura da cabeça, passando pelo centro do olho, Do-diâmetro do olho, Doc-distância do centro do olho até a parte inferior da cabeça, Lc-largura do corpo, Lb-largura da boca, Ab-altura máxima da boca, Po -distância entre a pré-maxila e a base da cabeça, Aip-altura da inserção da nadadeira peitoral, Acp-altura do corpo passando pela inserção da nadadeira peitoral, Cnp-comprimento máximo da nadadeira peitoral, Anp-área da nadadeira peitoral, Hnc -altura da nadadeira caudal, Anc -área da nadadeira caudal, Apc-altura do pedúnculo caudal, Prt-comprimento da protrusão bucal, Fo-comprimento do focinho, Massa peso total. Fonte: Leitão et al., 2016.
Tabela S1. Lista de atributos funcionais e seus respectivos significados ecológicos. Códigos para medidas morfológicas são apresentados na figura S3.
Atributo funcional Cálculo/Classe Significado ecológico
Referências
Área da abertura da boca
(Lb*Ab)/(Lc*Amc) tipo/tamanho de itens alimentares capturados
Adaptado de Karpouzi e Stergiou (2003)
Formato da boca
Ab/Lb Tática alimentar Karpouzi e Stergiou (2003)
Posição da boca
Po/Aco Tática alimentar e posição na coluna d’água
Adaptado de Sibbing e Nagelkerke (2001)

37
Comprimento da protrusão bucal
Prt/Fo Tática alimentar e natureza dos itens alimentares
Leitão et al., 2016
Tamanho do olho
Do/Aco
Acuidade visual
Adaptado de Boyle e Horn (2006)
Posição do olho Doc/Aco Posição vertical ocupada na coluna d’água
Gatz (1979)
Formato do corpo
Amc/Lc Hidrodinamismo e posição ocupada na coluna d’água
Adaptado de Sibbing e Nagelkerke (2001)
Área do corpo
(ln(((Pi/4)*Lc*Amc)+1)) /(ln(Massa+1))
Manobrabilidade Adaptado de Villéger et
al., 2010
Posição da nadadeira peitoral
Aip/Acp Manobrabilidade Leitão et al.,2016
Aspecto relativo da nadadeira peitoral
(Cnp^2)/Anp Uso da nadadeira peitoral para propulsão
Leitão et al.,2016
Estrangulamento do pedúnculo caudal
Hnc/Apc Eficiência da propulsão caudal
Leitão et al.,2016
Área relativa das nadadeiras
(2*Anp)/Anc Principal tipo de propulsão entre as
nadadeideiras
Adaptado de Villéger et
al., 2010
Proporção das nadadeiras em relação ao tamanho do corpo
((2*Anp)+Anc)/ (((Pi/4)*Lc*Amc))
Eficiência da manobrabilidade e/ou aceleração
Leitão et al.,2016
Rastros branquiais Ausente, pequenos e numerosos, pequenos e esparsos, intermediários e numerosos, intermediários e esparsos, longos e numerosos e longos e esparsos
Capacidade de filtragem e proteção das brânquias
Leitão et al.,2016
Massa Log(Massa+1) Metabolismo, resistência e capacidade de natação
Villéger et. al, 2010
Formato dos dentes Ausente, viliforme Cônico, Caniniforme Incisiviforme, Espatulado largo e fino Multicúspide central, Mamiliforme, Multicúspide triangular, Multicúspide serrilhado, Reniforme
Natureza de itens alimentares capturados e tática alimentar
Adaptado de Gatz (1979), Keenleyside (1979), Sazima (1986)

38
Figura S4. Qualidade do espaço funcional baseado no protocolo proposto por Maire et al. (2015).

39
Tabela S2. Atributos funcionais que mais influenciam cada eixo da PCoA. Valores obtidos
através da correlação entre os atributos funcionais e os eixos da PCoA
Atributo funcional Eixo 1 Eixo 2 Eixo 3 Eixo 4
Formato dos Dentes
0,7924
Rastros branquiais
0,7342
Posição da nadadeira peitoral
-0,6579
Massa 0,8679
Tamanho do olho
-0,6977
Posição da boca -0,4870
Área da boca
-0,6884
Formato da boca -0,5927
Área do corpo 0,3656
Formato do corpo 0,7127
Posição do olho -0,4248
Proporção das nadadeiras em relação ao tamanho do corpo
-0,3853

40
Tabela S3. Lista das espécies amostradas e respectivas abundâncias. * Espécies
categorizadas como ornamentais
Espécies Abundância
Characiformes 3854
Acestrorhynchidae 14
Acestrorhynchus_falcirostris 3
Acestrorhynchus_microleps 10
Acestrorhynchus_minimus 1
Anastomidae 3
Leporinus_klausewitzi 3
Characidae 3147
Axelrodia_stigmatias 1
Brittanichthys_axelrodi* 6
Bryconops_giacopinii 49
Characidium_pellucidum 18
Charax_condei 4
Gnathocharax_steindachneri 15
Hemigrammus_analis 492
Hemigrammus_bellottii 275
Hemigrammus_bleheri* 50
Hemigrammus_coeruleus 78
Hemigrammus_levis 21
Hemigrammus_lunatus 10
Hemigrammus_ocellifer 3
Hemigrammus_stictus* 137
Hemigrammus_vorderwinkleri 1102
Hoplocharax_goethei 95
Hyphessobrycon_aff_melazonatus 98
Hyphessobrycon_bentosi 2
Hyphessobrycon_diancistrus 1
Hyphessobrycon_rosaceus* 10
Hyphessobrycon_sp_microschemobrycon 18
Iguanodectes_adujai 1
Iguanodectes_gesleri 3
Iguanodectes_spilurus 3
Microschemobrycon_casiquiare 3
Moenkhausia_aff_coletti 145
Moenkhausia_coletti 14
Moenkhausia_lepidura 5
Moenkhausia_mikia 2
Paracheirodon_axelrodi* 396
Parapristela_aubiygnei 79
Poptella_compressa 1

41
Priocharax_sp 2
Rhinobrycon_negrensis 8
Crenuchidae 14
Ammocryptocharax_minutus 1
Crenuchus_spilurus 3
Elachocharax_junki 6
Elachocharax_puncher 1
Odontocharacidium_aphanes 2
Poecilocharax_weitzmani 1
Ctenoluciidae 2
Boulengerella_lateristriga 2
Curimatidae 93
Curimatopsis_evelynae 90
Curimatopsis_macroleps 3
Erythrinidae 21
Hoplerythrinus_unitaeniatus 6
Hoplias_curupira 1
Hoplias_malabaricus 14
Gasteropelecidae 329
Carnigiella_marthae* 297
Carnigiella_strigata* 32
Hemiodontidae 27
Hemiodus_gracilis 27
Lebiasinidae 196
Copella_nattereri* 33
Copella_nigrofasciata 11
Nannostomus_digramus* 96
Nannostomus_eques* 12
Nannostomus_marginatus* 3
Nannostomus_marilynae* 7
Nannostomus_trifasciatus* 1
Nannostomus_unifaciatus* 29
Pyrrulina_semifasciata 4
Serrasalmidae 8
Metynnis_hypsauchen 1
Metynnis_sp1_linha lateral preta 4
Metynnis_sp2 2
Pygopristis_denticulata 1
Chondrichthyes 3
Potamotrygonidae 3
Potamotrygon_wallacei* 3
Cyprinodontiformes 53
Poecilidae 52
Fluviphylax_obscurus 16

42
Fluviphylax_pigmeus 36
Rivulidae 1
Rivulus_ornatus 1
Gymnotiformes 18
Hypopomidae 17
Hypopygus_lepturus 12
Microsternachus_cf_bilineatus 4
Steatogenys_elegans 1
Ramphicthyidae 1
Hypopygus_hoedeman 1
Perciformes 241
Cichlidae 240
Acarichthys_heckelii 3
Acaronia_nassa 1
Acaronia_vultuosa 1
Aequidens_cf._pallidus 10
Aequidens_pallidus 3
Aequidens_sp 2
Aequidens_sp2 1
Apistogramma_gephyra* 1
Apistogramma_gibbiceps* 3
Apistogramma_pauciscranis* 11
Apistogramma_pertensis* 1
Apistogramma_uaupesi* 5
Biotecus_opercularis 12
Biotodoma_wavrini 3
Cichla_orinocensis 3
Crenicichla_notophthalmus 4
Dicrossus_filamentosus* 11
Heros_notatus* 4
Hoplarchus_psitticus 143
Laetacara_fulvipinis 2
Mesonauta_insignis 3
Satanoperca_lilith 2
Satanoperca_sp 1
Taeniacara_cf_candidi 9
Uaru_amphiacanthoides 1
Eleotridae 1
Microphilypnus_ternetzi 1
Siluriformes 116
Doradidae 70
Acanthodoras_spinosissimus 3
Ambliodoras_afinis 5
Scorpiodoras_heckelii 62
Heptapteridae 1

43
Gladioglanis_conquistador 1
Loricariidae 9
Acestridium_discus 2
Ancistrus_dolichopterus 6
Dekeyseria_scaphyryncha 1
Pimelodidae 3
Rhamdia_laukidi 3
Trichomycteridae 33
Trichomycterus_hasemani 33
Synbranchiformes 1
Synbranchidae 1
Synbrancus_sp 1
Total Geral 4286
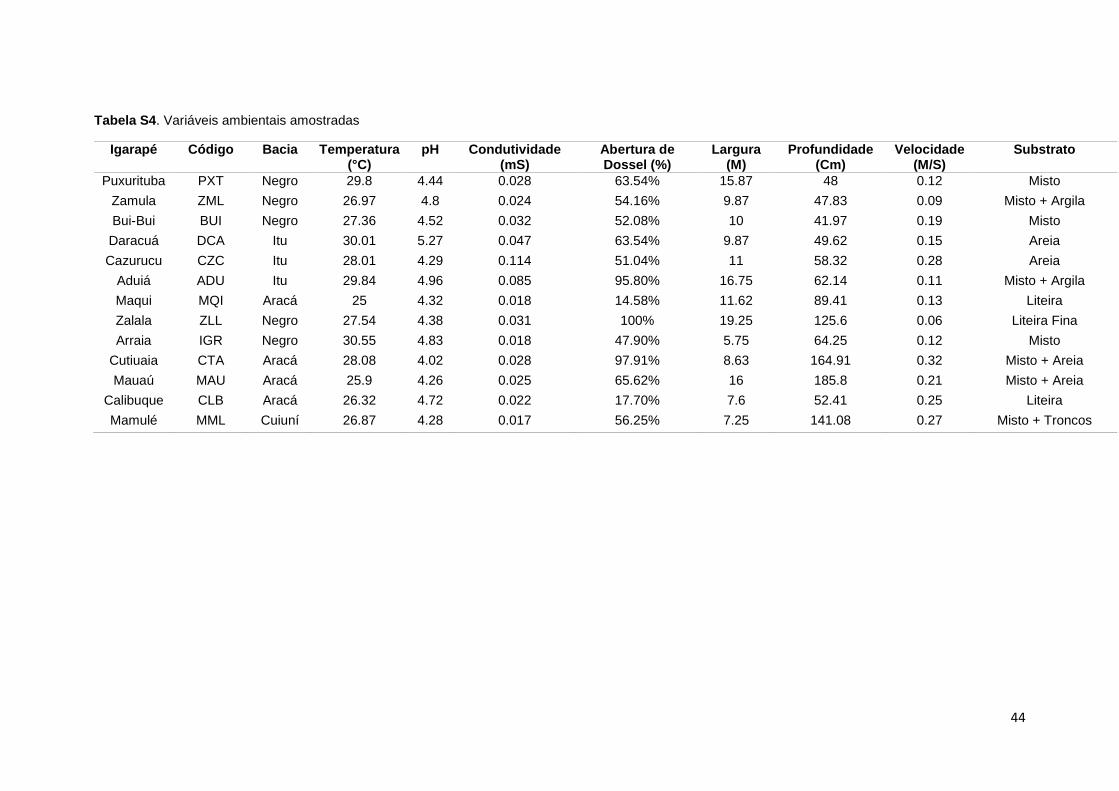
44
Tabela S4. Variáveis ambientais amostradas
Igarapé Código Bacia Temperatura (°C)
pH Condutividade (mS)
Abertura de Dossel (%)
Largura (M)
Profundidade (Cm)
Velocidade (M/S)
Substrato
Puxurituba PXT Negro 29.8 4.44 0.028 63.54% 15.87 48 0.12 Misto
Zamula ZML Negro 26.97 4.8 0.024 54.16% 9.87 47.83 0.09 Misto + Argila
Bui-Bui BUI Negro 27.36 4.52 0.032 52.08% 10 41.97 0.19 Misto
Daracuá DCA Itu 30.01 5.27 0.047 63.54% 9.87 49.62 0.15 Areia
Cazurucu CZC Itu 28.01 4.29 0.114 51.04% 11 58.32 0.28 Areia
Aduiá ADU Itu 29.84 4.96 0.085 95.80% 16.75 62.14 0.11 Misto + Argila
Maqui MQI Aracá 25 4.32 0.018 14.58% 11.62 89.41 0.13 Liteira
Zalala ZLL Negro 27.54 4.38 0.031 100% 19.25 125.6 0.06 Liteira Fina
Arraia IGR Negro 30.55 4.83 0.018 47.90% 5.75 64.25 0.12 Misto
Cutiuaia CTA Aracá 28.08 4.02 0.028 97.91% 8.63 164.91 0.32 Misto + Areia
Mauaú MAU Aracá 25.9 4.26 0.025 65.62% 16 185.8 0.21 Misto + Areia
Calibuque CLB Aracá 26.32 4.72 0.022 17.70% 7.6 52.41 0.25 Liteira
Mamulé MML Cuiuní 26.87 4.28 0.017 56.25% 7.25 141.08 0.27 Misto + Troncos
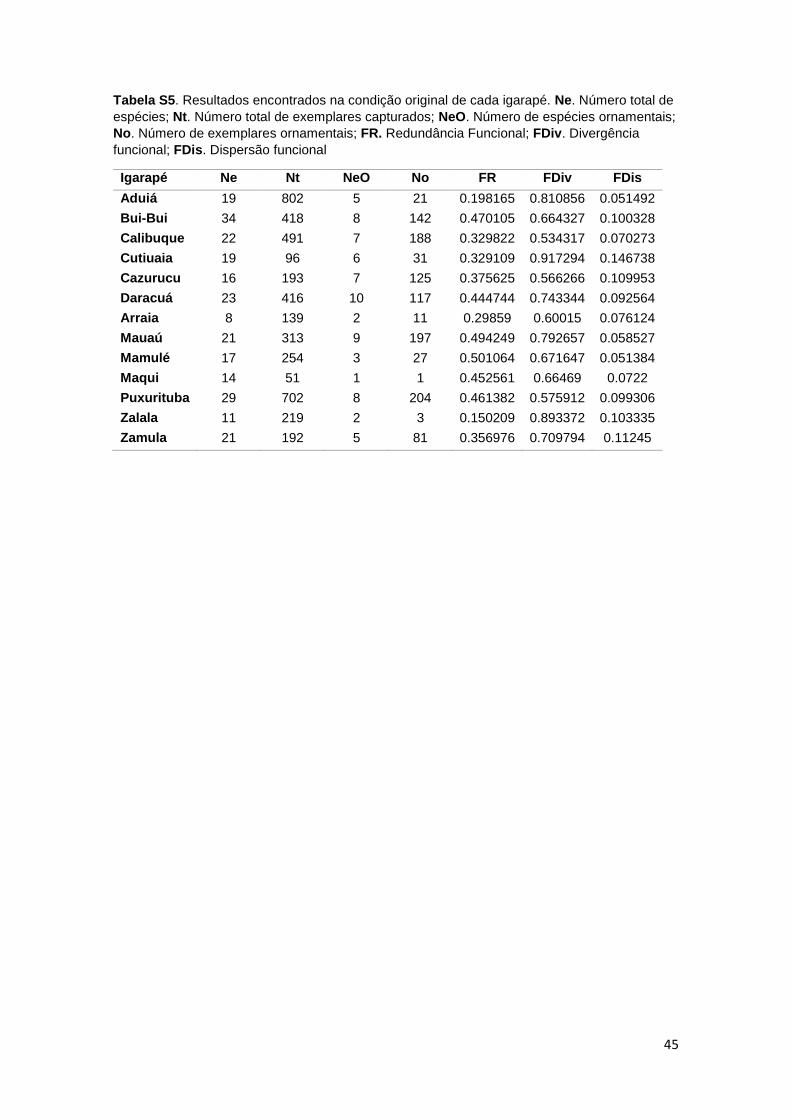
45
Tabela S5. Resultados encontrados na condição original de cada igarapé. Ne. Número total de
espécies; Nt. Número total de exemplares capturados; NeO. Número de espécies ornamentais;
No. Número de exemplares ornamentais; FR. Redundância Funcional; FDiv. Divergência
funcional; FDis. Dispersão funcional
Igarapé Ne Nt NeO No FR FDiv FDis
Aduiá 19 802 5 21 0.198165 0.810856 0.051492
Bui-Bui 34 418 8 142 0.470105 0.664327 0.100328
Calibuque 22 491 7 188 0.329822 0.534317 0.070273
Cutiuaia 19 96 6 31 0.329109 0.917294 0.146738
Cazurucu 16 193 7 125 0.375625 0.566266 0.109953
Daracuá 23 416 10 117 0.444744 0.743344 0.092564
Arraia 8 139 2 11 0.29859 0.60015 0.076124
Mauaú 21 313 9 197 0.494249 0.792657 0.058527
Mamulé 17 254 3 27 0.501064 0.671647 0.051384
Maqui 14 51 1 1 0.452561 0.66469 0.0722
Puxurituba 29 702 8 204 0.461382 0.575912 0.099306
Zalala 11 219 2 3 0.150209 0.893372 0.103335
Zamula 21 192 5 81 0.356976 0.709794 0.11245


















