
JULIA QUARTI CARDOSO - ppgn.ufrj.br€¦ · universidade federal do rio de janeiro centro de...
143
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO CENTRO DE CIÊNCIAS DA SAÚDE INSTITUTO DE NUTRIÇÃO JOSUÉ DE CASTRO PROGRAMA DE PÓS-GRADUAÇÃO EM NUTRIÇÃO EFEITO DA PIPERINA SOBRE MECANISMOS DE RESISTÊNCIA A QUIMIOTERÁPICOS EM UM MODELO EXPERIMENTAL DE LEUCEMIA MIELOIDE CRÔNICA JULIA QUARTI CARDOSO RIO DE JANEIRO 2019
Transcript of JULIA QUARTI CARDOSO - ppgn.ufrj.br€¦ · universidade federal do rio de janeiro centro de...

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
CENTRO DE CIÊNCIAS DA SAÚDE
INSTITUTO DE NUTRIÇÃO JOSUÉ DE CASTRO
PROGRAMA DE PÓS-GRADUAÇÃO EM NUTRIÇÃO
EFEITO DA PIPERINA SOBRE MECANISMOS DE RESISTÊNCIA A QUIMIOTERÁPICOS
EM UM MODELO EXPERIMENTAL DE LEUCEMIA MIELOIDE CRÔNICA
JULIA QUARTI CARDOSO
RIO DE JANEIRO
2019

Julia Quarti Cardoso
EFEITO DA PIPERINA SOBRE MECANISMOS DE RESISTÊNCIA A QUIMIOTERÁPICOS
EM UM MODELO EXPERIMENTAL DE LEUCEMIA MIELOIDE CRÔNICA
Orientadoras: Eliane Fialho de Oliveira
Vivian Mary Barral Dodd Rumjanek
RIO DE JANEIRO
2019
Tese de Doutorado apresentada ao Programa de
Pós-Graduação em Nutrição do Instituto de
Nutrição Josué de Castro da Universidade Federal
do Rio de Janeiro, como parte dos requisitos
necessários à obtenção do Título de Doutor em
Ciências Nutricionais.

FICHA CATALOGRÁFICA
Cardoso, Julia Quarti.
Efeito da piperina sobre mecanismos de resistência a quimioterápicos em um
modelo experimental de leucemia mieloide crônica. / Julia Quarti Cardoso. - Rio de
Janeiro: UFRJ / Centro de Ciências da Saúde, Instituto de Nutrição Josué de Castro,
Programa de Pós-Graduação em Nutrição, 2019.
142 f.: il.; 31 cm.
Orientadoras: Eliane Fialho de Oliveira e Vivian Mary Barral Dodd Rumjanek.
Tese (doutorado) – Universidade Federal do Rio de Janeiro / Instituto de
Nutrição Josué de Castro, Programa de Pós-Graduação em Nutrição, 2019.
Referências: f. 102-129.
1. Leucemia Mielogênica Crônica BCR-ABL Positiva. 2. Resistência a
Múltiplos Medicamentos. 3. Células - efeitos de drogas. 4. Técnicas In Vitro. 5.
Piper nigrum. 6. Nutrição - tese. I. Oliveira, Eliane Fialho de. II. Rumjanek, Vivian
Mary Barral Dodd. III. Universidade Federal do Rio de Janeiro, Centro de Ciências
da Saúde, Instituto de Nutrição Josué de Castro, Programa de Pós-Graduação em
Nutrição. IV. Título.


Dedico esta tese ao meu tio, Marcus Quarti,
pela força e pelo otimismo com que vence
diariamente a leucemia.

AGRADECIMENTOS
Ao CNPq, FAPERJ e Fundação do Câncer, pelos auxílios financeiros recebidos, essenciais para a
realização deste trabalho.
Às minhas orientadoras, Lili e Vivian. Não tenho palavras para agradecer tudo que fizeram por
mim, por toda confiança e por tudo que aprendi durante todos esses anos. Agradeço pela
oportunidade de poder acompanhar de perto o trabalho que vocês fazem com tanto amor, carinho,
cuidado e dedicação, pois não há maneira mais fácil de aprender do que observando o exemplo de
vocês. Vocês são minhas inspirações! Obrigada pela amizade, por cada conselho e ensinamento.
Sem dúvidas vocês se complementam como orientadoras e a minha formação foi ainda mais
preciosa por conta disso. Obrigada por tudo que representam para mim!
Às professoras que tanto colaboraram em muitos dos experimentos desta tese, Ana Rossini
(UERJ), Luciana Chiarini (UFRJ), Mariana Monteiro (UFRJ), Anna Votto (FURG), Karina
Machado (FURG), Sandra Mascarenhas (UFPB), Cecília Carpinella (Córdoba, Argentina).
Agradeço por toda paciência e acolhimento (à mim ou às minhas alunas de IC). A ajuda de vocês
foi essencial para a realização deste trabalho e para o meu aprendizado como pesquisadora.
À minha “piperete”, Larissa, por aparecer justamente no momento em que eu mais precisava.
Que sorte a minha ter te encontrado nesse finalzinho tão tumultuado. Desde o início você foi uma
aluna maravilhosa, esperta, ágil e interessada, por isso conseguiu me ajudar tanto e em tão pouco
tempo (nem acredito que você está há menos de 1 ano comigo - e olha que ainda quebrou o dedo
no meio do caminho...rs). E, além de ser uma ótima aluna, ainda tem um sorriso que ilumina
meus dias e um dos corações mais doces que eu conheço. Agradeço muito por toda sua ajuda e
empenho! Você tem um lugarzinho especial no meu coração!
Aos meus ex “piperetes” que me ajudaram ao longo desses 4 anos, Erika, Juliana, Marina,
Giovanna, André, Mariana, Gabriela (emprestada...rs) e Emanoela (quase aluna..rs). Agradeço
pela amizade, conversas e risadas. Obrigada pelo empenho em me ajudar da melhor maneira
possível com os experimentos. Eu não poderia ter tido melhores alunos! Muito obrigada!
À Profª. Danielly Ferraz, por não medir esforços para me ajudar, no que fosse preciso. Agradeço
imensamente pela sua amizade, por todos os ensinamentos desde a minha IC e pelos conselhos.
Obrigada sempre por me motivar a seguir em frente e por compartilhar sua experiência comigo,
me dando forças quando eu mais precisei durante todo esse período da vida acadêmica. Que
honra ter você como revisora desta tese, muito obrigada por todas as suas contribuições ao longo
do trabalho!

À Profª. Tatiana El-Bacha, por ser sempre tão solícita. Obrigada por cada contribuição desde que
eu estava no mestrado. Agradeço também por ter aceitado o convite para participar da banca
avaliadora, mesmo estando fora do país.
À Profª. Fátima Sardinha, por ter contribuído para minha formação como professora e
pesquisadora. Agradeço, principalmente, por ter ajudado a desenvolver minha capacidade de me
expressar de forma mais clara e objetiva. Obrigada também por ter aceitado o convite para
participar da banca avaliadora.
À Profª. Luciana Rangel, pela disponibilidade em me ajudar, sempre. Obrigada pela paciência em
responder minhas dúvidas e por ter aceitado o convite para participar da banca avaliadora.
À pesquisadora Ana Cecilia, por ter aceitado o convite para participar da banca avaliadora.
Aos Profs. Marcia Soares e Antônio Ferreira, por terem aceitado participar como suplentes da
banca.
À tão querida Dra. Otília, por ser uma verdadeira inspiração para mim. Obrigada pelas conversas
e conselhos tão sábios sobre a vida e sobre o meu trabalho.
Aos técnicos de laboratório Giselle e Sebastião, pelo convívio e eficiência no preparo do material
para os meus experimentos. Muito obrigada pela compreensão nos momentos de pressa e correria
e por se empenharem em me ajudar, sempre.
Ao Paulinho e à Jacque, por serem sempre tão solícitos e me ajudarem no que fosse preciso.
Ao meu verdadeiro anjo da guarda, Kim. Que sorte (ou merecimento... rs) de ter você na minha
vida! Obrigada por ser minha best e por iluminar meu caminho, tornando tudo mais leve.
Obrigada pelos melhores conselhos e palavras de consolo, por poder contar com você nos
momentos mais importantes e de maior dificuldade, por sempre me lembrar que devo ser grata
por tudo que tenho, por ser como uma irmã pra mim e por me fazer sentir “felicidade extrema” só
de lembrar que tenho você ao meu lado! Obrigada por tudo e mais um pouco! Amo você, amiga!
À minha amiga, Iris Leite, por toda ajuda e empenho com os experimentos do HPLC, mesmo
tendo pouco tempo disponível para fazer as análises. Obrigada pela sua amizade, por ouvir meus
desabafos e por alegrar meus dias com tantas risadas. Você mora no meu coração!
À Daianne Torres, por toda ajuda com as imunos e por ter se envolvido tanto com o meu
trabalho, me ajudando a entender os resultados dos experimentos. Obrigada pela ótima
companhia, pelas conversas e por ser sempre tão divertida!

Às minhas queridas amigas da “velha guarda” do LABAF’s, Fabi, Chris Angel, Renatinha e Lú.
Não tem um dia que eu não sinta falta da parceria e da companhia de vocês no laboratório. Mas
agradeço por todos os ensinamentos que, mesmo depois de anos, continuam me ajudando e por
todo apoio e incentivo que vocês me dão, mesmo de longe. Obrigada por termos formado o
melhor grupo de pesquisa de todos! Amo vocês!
Às Profs. Ana Luisa, Maria Lúcia e Denise pelo carinho, pela força e por contribuírem tanto para
o meu aprendizado, como professora e como pesquisadora.
A todos do LABAF's, em especial, ao Christian e Luan. Muito obrigada por compartilharem seus
conhecimentos e experiência, sempre dando ótimas sugestões para o meu trabalho.
A todos do Laboratorio Química Fina y Productos Naturales por me acolherem tão bem durante o
período que estive em Córdoba. Obrigada por serem tão gentis e também por todos os
ensinamentos no laboratório.
A todos do Laboratório de Cultura Celular (Rio Grande, FURG), em especial, Michele, Estela,
Mari e Fran. Agradeço imensamente pela parceria nos experimentos e por acolherem tão bem
minhas queridas alunas. À Mari e Fran por todo carinho e sintonia que tivemos desde que nos
conhecemos. Vocês são muito especiais para mim. Muito obrigada por tudo, meninas!
A todos do Laboratório de Toxicologia e Biologia Molecular (UERJ), em especial, Orlando e
Pedro. Obrigada pela ajuda e conversas durante os experimentos.
A todos do LTPV, em especial, Giulia e Samir. Obrigada pelas conversas, pela troca de
experiências e por toda ajuda e empréstimo de material e de reagentes (doação, na verdade, né?
rs...). Agradeço muitíssimo!
A todos do LIT, pelo agradável convívio. Agradeço, especialmente, a Maria, Eduardo e Raphael
Valente por muitas vezes ouvirem meus desabafos e por sempre tentarem me dar ideias para os
experimentos. Obrigada por tudo!
Às minhas “amigas de faculdade” que vou levar para a vida, Didini, Dani, Malu e Ana. Obrigada
pela amizade, compreensão, apoio e por torcerem tanto por mim! Como eu sinto falta do nosso
convívio diário! Vocês sempre vão ser as minhas “nutris” preferidas!
Às minhas queridas e amadas “amigas de escola” (Thais, Camila, Gabi, Mari e Mayara) por
serem tão especiais; por todo amor, amizade e conselhos; pelos nossos encontros que renovam as
minhas energias. Tenho certeza de que a nossa amizade e nossos “pijamões” serão eternos!
Obrigada por todo apoio, sempre! Amo vocês, sinzi?!

A todos do Centro Espírita Amigos no Caminho (CEAC), em especial aos meus pais (Denise e
Luis Paulo), Mauro, Marluci, Sérgio e Celina por todas as preces e boas vibrações emanadas nos
momentos em que eu mais precisei. Certamente me ajudaram a ficar mais serena. Muito obrigada
por tudo!
À minha segunda família, Mônica, Daniel, Gi, Gê, Florinha e Heitor. Obrigada pela compreensão
nos momentos em que estive ausente, pelo apoio e pela torcida, sempre. Obrigada pelo carinho,
pelas comidas deliciosas, pelas risadas. Muito obrigada por me acolherem como parte da família.
Vocês conquistaram um grande espaço no meu coração! Amo vocês!
À minha família que tanto se orgulha de mim a cada conquista. Obrigada pelo apoio! Amo vocês!
Aos meus bichinhos, Mila Burns, Elis Flor de Lis e Tom Zé por serem meus companheiros e por
acalmarem meu coração nos momentos de estresse com a tese. Vocês são minhas paixões! Amo
muito vocês!
Ao Roldan, por ser meu amor de vida. Obrigada por alegrar meus dias; por me fazer tão feliz, me
conquistando cada dia mais; por muitas vezes se preocupar mais comigo do que com você
mesmo. Obrigada por sentir tanto orgulho de mim que me faz sentir especial. Seu amor, incentivo
e apoio me permitiram chegar até aqui e tenho certeza que, juntos, iremos ainda mais longe. Te
amo muito!
À irmã, Bela, por ser mais do que uma melhor amiga, sempre me ajudando e apoiando em todos
os momentos. Só o “amor de irmã” explica tantas coisas que fazemos uma pela outra. Obrigada,
especialmente, pela formatação das referências em todos os meus trabalhos! Como sempre, você
me salva! Agradeço por você ser a melhor irmã do mundo! Te amo S2!
Aos meus tão preciosos pais, Denise e Luis Paulo, por serem meus alicerces nessa jornada, por
me amarem incondicionalmente, sofrerem as minhas angustias e comemorarem as minhas
vitórias. Obrigada por me guiarem pelo caminho do bem. Obrigada por não medirem esforços e
se dedicarem incansavelmente para que eu pudesse ir em busca dos meus sonhos e,
principalmente, por acreditarem que eu seria capaz de alcançá-los, isso me dá forças para
continuar a minha trajetória. Essa conquista, sem dúvida, também é de vocês! Sem o amor de
vocês, eu nada seria. Amo vocês mais do que tudo!
A Deus, por me dar forças para vencer os desafios, superar os obstáculos e ir em busca da minha
felicidade. Obrigada por colocar em meu caminho pessoas maravilhosas que me ajudam nessa
caminhada. Muito obrigada pela vida!

“Crê em ti mesmo, age e verá os resultados.
Quando te esforças, a vida também
se esforça para te ajudar.”
Chico Xavier

RESUMO
A resistência a múltiplas drogas (MDR) é o principal obstáculo ao tratamento da leucemia mieloide
crônica (LMC) em crise blástica. Diferentes mecanismos podem ser induzidos durante o desenvolvimento
do fenótipo MDR, dentre eles: superexpressão de bombas de efluxo de drogas, como a glicoproteína-P (P-
gp); alteração do ciclo celular; inibição da morte celular e aumento do metabolismo de xenobióticos. Uma
estratégia promissora é a identificação de compostos cuja ação seja seletiva sobre células MDR, fenômeno
conhecido como sensibilidade colateral (CS). A piperina, fitoquímico presente na pimenta-do-reino,
apresenta efeito anticâncer e anti-MDR, no entanto, ainda não foi explorada sua ação na promoção de CS.
O objetivo desse estudo foi investigar os efeitos da piperina sobre diferentes mecanismos de resistência a
quimioterápicos em células de LMC em crise blástica. Os ensaios de redução de MTT demonstraram que
a piperina promoveu maior efeito citotóxico sobre as linhagens de leucemia com fenótipo MDR, Lucena-1
e FEPS, do que sobre a linhagem sensível a drogas, K562, caracterizando a ocorrência da CS. Em
contrapartida, o efeito citotóxico da piperina sobre macrófagos peritoneais de camundongos foi menor do
que nas células MDR. Ensaios de coloração com panótico rápido e por citometria de fluxo indicaram que
a piperina provocou importantes alterações morfológicas nas linhagens de leucemia, como indução da
formação de vacúolos no citoplasma, que, por sua vez, pode ter gerado aumento da granulosidade celular,
em maior grau na FEPS. Experimentos de citometria de fluxo indicaram que embora a piperina não tenha
alterado a distribuição do ciclo celular, induziu apenas as células MDR ao processo de apoptose, sendo
que esse efeito foi mais pronunciado sobre a linhagem FEPS. Tais dados foram confirmados pela
marcação de caspase-3 clivada por imunofluorescência. Apesar de as células MDR superexpressarem a
enzima glicolítica gliceraldeído-3-fosfato desidrogenase (GAPDH), por RT-PCR em tempo real e Western
blotting, indicando maior resistência à morte celular independente de caspases (MCIC), a piperina não foi
capaz de induzir MCIC via inibição de GAPDH nessas linhagens. A enzima poli (ADP-ribose)
polimerase-1 (PARP-1) possui diversas funções celulares relacionadas tanto à sobrevivência, como
indução de morte celular. As análises por Western blotting indicaram que a linhagem FEPS apresentou os
menores níveis dessa proteína, bem como a menor atividade (inferida pelos níveis do polímero poli (ADP-
ribose) – PAR), além disso, foi a única que apresentou níveis detectáveis do fragmento de 24 kDa,
sugerindo o desenvolvimento de um mecanismo capaz de impedir a superativação de PARP-1, o que
poderia proteger essas células contra morte por necrose. A piperina foi capaz de reduzir os níveis de
PARP-1 e do fragmento de 24 kDa, indicando uma possível relação com a indução de CS. A fim de
investigar se os níveis de PARP-1 teriam correlação com o fenótipo MDR, foi realizada análise da
expressão de P-gp, por citometria de fluxo. Apesar de a piperina ter aumentado a expressão de P-gp em
ambas as linhagens MDR, somente na FEPS haveria uma possível correlação entre a diminuição da
expressão de PARP-1 e o aumento da expressão de P-gp após incubação com piperina. Também
investigamos a possibilidade da piperina estar sofrendo metabolização nas células MDR, gerando
metabólitos com potencial ainda mais citotóxico. Primeiramente, analisamos o nível de genes relacionados
ao metabolismo de fase I de xenobióticos por RT-PCR em tempo real. As linhagens não apresentaram
níveis detectáveis de CYP3A4, CYP1B1 e PXR. No entanto, a linhagem FEPS apresentou a maior
expressão de CYP3A5, seguida da Lucena-1, sendo que a piperina foi capaz de induzir ainda mais a
expressão desse gene nessas células MDR e, ainda, demonstrou interagir com a proteína CYP3A5, por
docking molecular. Em seguida, avaliamos a metabolização de piperina, por CLAE-DAD. No entanto, não
foram observadas diferenças entre as linhagens de leucemia, sugerindo que, provavelmente, CYP3A5 não
estaria metabolizando esse fitoquímico. Sendo assim, piperina parece promover CS por diferentes vias de
sinalização, como modulando PARP-1 (na FEPS), P-gp e CYP3A5. A identificação de mecanismos
relacionados à indução de CS pode auxiliar no delineamento de estratégias contra células com fenótipo
MDR.
Palavras-chave: Leucemia Mieloide Crônica, Resistência a Múltiplas Drogas, Piperina, Sensibilidade
Colateral, PARP-1, Glicoproteína-P, CYP3A5.

ABSTRACT
Multiple drug resistance (MDR) is the main obstacle to the treatment of chronic myeloid leukemia (CML)
in blast crisis. Different mechanisms may be induced during the development of the MDR phenotype,
among them: overexpression of drug efflux pumps, such as P-glycoprotein (P-gp); cell cycle alteration;
cell death inhibition and increased xenobiotics metabolism. One promising strategy is the identification of
compounds whose action is selective on MDR cells, the phenomenon known as collateral sensitivity (CS).
Piperine, phytochemical present in black pepper, has anti-cancer and anti-MDR effects, however, its effect
on CS induction was not investigated yet. The aim of this study was to investigate the effects of piperine
on different mechanisms of drug resistance in CML cells in blast crisis. The MTT reduction assay showed
that piperine promoted a greater cytotoxic effect on the leukemia cells with the MDR phenotype, Lucena-
1 and FEPS than on the drug-sensitive cells, K562, characterizing the occurrence of CS. In contrast, the
cytotoxic effect of piperine on mouse peritoneal macrophages was lower than in MDR cells. Rapid
panoptic staining and flow cytometry indicated that piperine caused significant morphological changes in
leucemia cells, such as induction of cytoplasmic vacuoles formation, which, in turn, may increase cellular
granularity, to a greater degree in FEPS. Flow cytometric experiments indicated that although piperine did
not undergo a change in the cell cycle, piperine induced only MDR cells to apoptosis and the effect was
more pronounced on FEPS cells. These data were confirmed by immunofluorescence of cleaved caspase-
3. MDR cells overexpress a glycolytic enzyme glycerol dehydrogenase-3-phosphate dehydrogenase
(GAPDH), by real-time RT-PCR and Western blotting, suggesting a caspase-independent cell death
(CICD) resistance. However piperine was not able to induce CICD via inhibition of GAPDH in these cell
lines. Poly (ADP-ribose) polymerase-1 (PARP-1) has several survival and cell death-related functions.
Western blotting analyzes indicated that FEPS cells presented a small amount of this protein, as well as
the lower activity (inferred by the levels of the polymer Poly (ADP-ribose) - PAR). In addition, it was
detected only in FEPS cells the PARP-1 24 kDa fragment, suggesting the development of a mechanism
capable of preventing the overactivation of PARP-1, which could protect these cells from death by
necrosis. Piperine reduced the levels of PARP-1 and its 24 kDa fragment, establishing a relationship with
the induction of CS. In order to investigate whether PARP-1 levels is correlated with the MDR phenotype,
analysis of the P-gp expression was performed by flow cytometry. Although piperine increased P-gp
expression in both MDR cells, only in FEPS would have a possible correlation between the decrease in
PARP-1 expression and the increase in P-gp expression after incubation with piperine. We also
investigated the possibility that piperine is undergoing metabolism in MDR cells, generating metabolites
with even more cytotoxic potential. First, we analyzed the level of genes related to phase I metabolism of
xenobiotics by real time RT-PCR. The cell lines showed no detectable levels of CYP3A4, CYP1B1 and
PXR. However, FEPS cells presented the highest expression of CYP3A5, followed by Lucena-1, and
piperine was able to further induce expression of this gene in these MDR cells and also demonstrated to
interact with the CYP3A5 protein by molecular docking. Next, we evaluated the metabolism of piperine
by HPLC-DAD. However, no differences were observed among leukemia cells, suggesting that CYP3A5
would probably not be metabolizing this phytochemical. Thus, piperine appears to promote CS by
different signaling pathways, such as modulating PARP-1 (in FEPS), P-gp and CYP3A5. The
identification of mechanisms related to CS induction may help in the design of strategies against cells with
MDR phenotype.
Key-words: Chronic Myeloid Leukemia, Multiple Drug Resistance, Piperine, Collateral Sensitivity,
PARP-1, P-glycoprotein, CYP3A5.

LISTA DE ABREVIATURAS
ABC - do inglês, ATP-Binding Cassette
ABL - do inglês, Abelson Leukemia
AIF - do inglês, Apoptosis Inducing Factor
APAF-1 - do inglês, Apoptosis Protease Activating Fator-1
BCR - do inglês, Breakpoint Cluster Region
BCRP - do inglês, Breast Cancer Resistance-Protein
Caspase - do inglês, Cysteine-dependent Aspartate-specific Proteases
CLAE-DAD - Cromatografia Líquida com Detecção por Arranjo de Diodos
CS - do inglês, Collateral Sensitivity
CYP450 - do inglês, Cytochrome P450
DMSO - Dimetilsulfóxido
EDTA - do inglês, Ethylenediamine Tetraacetic Acid
FITC - do inglês, Fluorescein Isothiocyanate
FSC - do inglês, Forward Scatter
G1 - do inglês, Gap phase 1
G2 - do inglês, Gap phase 2
GAPDH - do inglês, Glyceraldehyde 3-phosphate dehydrogenase
HEM - do inglês, Protoporphyrin IX containing Fe
HIT - Hitonavir
Hsp70 - do inglês, Heat Shock Protein 70
IAP - do inglês, Inhibitors of Apoptosis Proteins
IC50 - do inglês, Half Maximal Inhibitory Concentration
LC-MS/MSn
- do inglês, Liquid Chromatography-electrospray Ionization/multi-stage Mass
Spectrometry
LMC - Leucemia Mieloide Crônica
MCIC - Morte Celular Independente de Caspase
MDR - do inglês, Multidrug Resistance
MDR1 - Gene de Resistência a Múltiplas Drogas 1
MIF - Média da Intensidade de Fluorescência

MRP-1 - do inglês, Multidrug Resistance-Related Protein 1
MTT - Brometo de 3-(4,5-dimetiltiazol-2-il)-2,5-difeniltetrazol
NAD+ - Nicotinamida Adenina Dinucleotídeo
PAR - Polímeros poli (ADP-ribose)
PARP-1 - Poli (ADP-ribose) polimerase-1
PBMC - do inglês, Peripheral Blood Mononuclear Cells
PBS - do inglês, Phosphate-Buffered Saline
PDB - do inglês, Protein Data Bank
P-gp - do inglês, P-glycoprotein
Ph1 - do inglês Philadelphia Chromosome
PI - do inglês, Propidium Iodide
PIP - Piperina
PMSF - do inglês, Phenylmethylsulfonyl Fluoride
PVDF - do inglês, Polyvinylidene Difluoride
PXR - do inglês, Pregnane X Receptor
ROS - do inglês, Reactive Oxygen Species
rTNF - do inglês, Tumor Necrosis Factor Receptor
RT-PCR - do inglês, Reverse Transcription Polymerase Chain Reaction
S - Fase de Síntese de DNA
SDS - do inglês, Sodium Dodecyl Sulfate
SFB - Soro Fetal Bovino
SSC - do inglês, Side Scatter
TBS - do inglês, Tris-buffered Saline
TCA - do inglês, Trichloroacetic Acid
TEMED - do inglês, Tetramethylethylenediamine
UA - unidade arbitrária
UHPLC/QTOF-MS - do inglês, Ultra-high Performance Liquid Chromatography-quadrupole
Time-of-flight Mass Spectrometry

LISTA DE FIGURAS
Figura 1. Distribuição proporcional dos dez tipos de câncer mais incidentes estimados para
2018 por sexo, exceto pele não melanoma....................................................................................20
Figura 2. Translocação recíproca entre os cromossomos 9 e 22, formando o cromossomo
Philadelphia (Ph1)........................................................................................................................21
Figura 3. Modelo hipotético bidimensional da glicoproteína-P humana, baseada na análise da
seqüência de aminoácidos e seus domínios funcionais.................................................................23
Figura 4. Alimentos funcionais e seus compostos bioativos........................................................34
Figura 5. Estruturas químicas dos isômeros da piperina..............................................................36
Figura 6. Proposta das vias metabólicas da piperina em ratos.....................................................37
Figura 7. Estruturas químicas dos metabólitos da piperina em humanos....................................38
Figura 8. Efeito da piperina sobre a viabilidade das células de leucemia K562, Lucena-1 e FEPS
pelo método de redução de MTT..................................................................................................59
Figura 9. Efeito da piperina sobre a viabilidade das células de leucemia K562, Lucena-1 e FEPS
pelo método de exclusão de azul de tripan...................................................................................61
Figura 10. Efeito da piperina sobre a viabilidade de macrófagos peritoneais..............................62
Figura 11. Efeito da piperina sobre a morfologia das células de leucemia K562, Lucena-1 e
FEPS..............................................................................................................................................63
Figura 12. Efeito da piperina sobre o tamanho e granulosidade das células de leucemia K562,
Lucena-1 e FEPS...........................................................................................................................64
Figura 13. Efeito da piperina sobre a distribuição do ciclo celular das células de leucemia K562,
Lucena-1 e FEPS...........................................................................................................................67
Figura 14. Efeito da piperina sobre a morte das células de leucemia K562, Lucena-1 e FEPS..70
Figura 15. Efeito da piperina sobre a clivagem de caspase-3 na linhagem K562........................72
Figura 16. Efeito da piperina sobre a clivagem de caspase-3 na linhagem Lucena-1..................73
Figura 17. Efeito da piperina sobre a clivagem de caspase-3 na linhagem FEPS........................74
Figura 18. Efeito da piperina sobre a expressão gênica de GAPDH nas células de leucemia
K562, Lucena-1 e FEPS................................................................................................................75

Figura 19. Efeito da piperina sobre a expressão de GAPDH nas células de leucemia K562,
Lucena-1 e FEPS...........................................................................................................................76
Figura 20. Efeito da piperina sobre a expressão de PARP-1 e PAR nas células de leucemia
K562, Lucena-1 e FEPS................................................................................................................77
Figura 21. Efeito da piperina sobre a expressão de P-gp nas linhagens de leucemia..................79
Figura 22. Expressão gênica de CYP3A5 nas células de leucemia K562, Lucena-1 e
FEPS..............................................................................................................................................81
Figura 23. Efeito da piperina sobre a expressão gênica de CYP3A5 nas células de leucemia
K562, Lucena-1 e FEPS................................................................................................................82
Figura 24. Interação da piperina com CYP3A5...........................................................................83
Figura 25. Curva de degradação de piperina no meio de cultura celular.....................................84
Figura 26. Metabolização da piperina nas células de leucemia K562, Lucena-1 e FEPS...........85

LISTA DE TABELAS
Tabela 1. Efeito citotóxico da piperina (IC50) sobre as linhagens celulares K562, Lucena-1 e
FEPS pelo método de redução de MTT.........................................................................................60
Tabela 2. Percentual de células em alterações no seu tamanho e/ ou granulosidade em relação ao
controle...........................................................................................................................................65
Tabela 3. Percentual das células K562, Lucena-1 e FEPS nas fases Sub-G1, G1, S ou G2/M após
tratamento com piperina.................................................................................................................68
Tabela 4. Percentual das células K562, Lucena-1 e FEPS em necrose, apoptose tardia e
apoptose..........................................................................................................................................71

SUMÁRIO
1. INTRODUÇÃO........................................................................................................................19
1.1. Câncer......................................................................................................................................19
1.1.1. Leucemias.................................................................................................................20
1.2. Mecanismos relacionados à Resistência a Múltiplas Drogas..................................................21
1.2.1. Transportadores ABC...............................................................................................22
1.2.2. Ciclo celular..............................................................................................................23
1.2.3. Morte celular.............................................................................................................24
1.2.4. Poli (ADP-ribose) polimerase-1 PARP-1)................................................................26
1.2.4.1 Reparo do DNA..........................................................................................26
1.2.4.2 Apoptose.....................................................................................................28
1.2.4.3 Partanatos....................................................................................................28
1.2.4.4 P-gp.............................................................................................................29
1.2.5. Metabolismo de xenobióticos e substâncias endógenas...........................................29
1.3. Sensibilidade Colateral............................................................................................................31
1.4. Fitoquímicos............................................................................................................................32
1.4.1 Piperina......................................................................................................................35
1.4.1.1 Biodisponibilidade e metabolismo..............................................................35
1.4.1.2 Efeito antitumoral.......................................................................................39
1.5. Modelo experimental de LMC................................................................................................42
2. JUSTIFICATIVA....................................................................................................................45
3. OBJETIVOS.............................................................................................................................46
3.1. Objetivo geral..............................................................................................................46
3.2. Objetivos específicos...................................................................................................46
4. MÉTODOS................................................................................................................................47
4.1. Culturas de células.......................................................................................................47
4.2. Ensaio de citotoxicidade - Redução de MTT (3-(4,5 dimethyl thiazole-2yl)-2,5
diphenyl tetrazolium bromide) com as células de leucemia...........................................................47
4.3. Ensaio de citotoxicidade - Teste de exclusão com Azul de Tripan.............................48
4.4. Ensaio de citotoxicidade - Redução de MTT com macrófagos peritoneais................48
4.5. Coloração por Panótico Rápido...................................................................................49

4.6. Análise do ciclo celular.............................................................................................50
4.7. Ensaio de apoptose.......................................................................................................50
4.8. Imunofluorescência......................................................................................................51
4.9. Investigação da expressão de GAPDH, PARP-1 e PAR.............................................52
4.9.1. Preparação do extrato protéico das células...................................................52
4.9.2. Eletroforese em gel de poliacrilamida em condições desnaturantes.............52
4.9.3. Western blotting............................................................................................53
4.10. Análise da expressão de P-gp....................................................................................53
4.11. Análise da influência da piperina sobre a expressão gênica de GAPDH e de
marcadores do metabolismo de xenobióticos.................................................................................54
4.12. Docking molecular.....................................................................................................55
4.13. Determinação da piperina intracelular e extracelular por Cromatografia Líquida com
Detecção por Arranjo de Diodos (CLAE-DAD)............................................................................56
4.14. Análise estatística......................................................................................................57
5. RESULTADOS.........................................................................................................................58
5.1. Efeito da piperina sobre a viabilidade das células de leucemia...................................58
5.2. Efeito da piperina sobre a viabilidade de células não-tumorais...................................61
5.3. Efeito da piperina sobre a morfologia das células de leucemia...................................62
5.4. Efeito da piperina sobre o ciclo celular e a morte celular nas células de leucemia.....66
5.5. Efeito da piperina sobre a ativação de caspase-3 nas células de leucemia..................71
5.6. Efeito da piperina sobre os níveis de GAPDH nas células de leucemia......................74
5.7. Efeito da piperina sobre os níveis de PARP-1 e PAR nas células de leucemia...........76
5.8. Efeito da piperina sobre a expressão de P-gp nas linhagens MDR, Lucena-1 e
FEPS...............................................................................................................................................78
5.9. Efeito da piperina sobre enzimas metabolizadoras de xenobióticos nas células de
leucemia..........................................................................................................................................80
5.10. Análise da metabolização de piperina nas células de leucemia.................................84
6. DISCUSSÃO.............................................................................................................................87
7. CONCLUSÕES......................................................................................................................100
8. REFERÊNCIAS.....................................................................................................................102
9. ANEXOS.................................................................................................................................130

19
1. INTRODUÇÃO
1.1. Câncer
O câncer é uma doença conhecida, essencialmente, pela rápida proliferação e caráter
invasivo das células, em decorrência do acúmulo progressivo de mutações no seu genoma. Essas
alterações na expressão ou função de genes essenciais para a manutenção da homeostasia celular
provoca uma perda da capacidade da célula de reparar danos no DNA e de responder aos sinais
de proliferação e morte, adquirindo autonomia de crescimento e tornando-se independentes de
controles externos (National Institutes of Health, 2019).
Além disso, pesquisas sobre o desenvolvimento do câncer têm destacado a complexidade
dessa doença. Atualmente sabe-se que as células cancerígenas recrutam/ liberam um conjunto de
células e moléculas, constituindo o microambiente tumoral. Com isso, as células de câncer
podem influenciar as células normais que envolvem e suprem as demandas do tumor, induzindo
alterações na vascularização e oxigenação, garantindo vantagens de sobrevivência e crescimento
em relação às células normais (SIEMANN e HORSMAN, 2015).
De acordo com a Organização Mundial da Saúde (OMS), o câncer é a segunda principal
causa de mortalidade no mundo, sendo responsável por uma estimativa de 9,6 milhões de mortes
em 2018. Aproximadamente 70% das mortes por essa doença ocorrem em países em
desenvolvimento, dentre eles, o Brasil (OMS, 2018).
No Brasil, foi divulgada pelo Instituto Nacional de Câncer José Alencar Gomes da Silva
(INCA) a estimativa para os anos de 2018 e 2019. Foi estimada a ocorrência de cerca de 420 mil
casos novos de câncer por ano, excetuando-se o câncer de pele não melanoma (aproximadamente
170 mil casos novos). Segundo essa mesma previsão, as leucemias estarão entre os 10 tipos de
cânceres mais incidentes entre a população, com aproximadamente 5.940 casos novos em
homens e 4.860 em mulheres (Figura 1) (INCA, 2018).

20
Figura 1. Distribuição proporcional dos dez tipos de câncer mais incidentes estimados para
2018 por sexo, exceto pele não melanoma (INCA, 2018).
1.1.1. Leucemias
As leucemias representam um grupo de neoplasias malignas derivadas das células
hematopoiéticas na medula óssea. De acordo com a célula de origem, as leucemias podem ser
classificadas em mieloide ou linfoide e de acordo com a velocidade de sua evolução, são
classificadas em aguda ou crônica (INCA, 2018). Portanto, os tipos mais comuns de leucemia
são: leucemia linfoide aguda, leucemia mieloide aguda, leucemia linfoide crônica e leucemia
mieloide crônica (LMC), sendo esta última o foco desta tese.
A LMC é uma doença mieloproliferativa caracterizada pelo excesso de granulócitos
(neutrófilos, basófilos, eosinófilos). Esse tipo de câncer pode se apresentar em três fases distintas:
fase crônica ou estável, fase acelerada (de metamorfose ou de transformação) e fase aguda (crise
blástica) (ANKATHIL e cols., 2018). Os pacientes que evoluem para crise blástica apresentam o
pior prognóstico com mediana de sobrevida de menos de 1 ano (HEHLMANN e cols., 2016).
A LMC é caracterizada por uma anormalidade citogenética adquirida, o cromossomo
Philadelphia (Ph1) ou t(9;22) (q34;q11). O Ph1 resulta de uma translocação recíproca entre os
cromossomos 9 e 22, por meio da transferência do proto-oncogene ABL (do inglês,
Abelson Leukemia) do cromossomo 9 para uma região do cromossomo 22 denominada BCR (do
inglês, breakpoint cluster region) (Figura 2). Com isso, é formado o gene híbrido BCR-ABL, que
codifica a proteína de fusão bcr-abl e, esta, por sua vez, apresenta atividade constitutiva de
tirosina quinase aumentada, estando associada à inibição de apoptose, à indução da proliferação,
a alterações no citoesqueleto celular e a modificações dos processos de sinalização em células

21
portadoras do gene híbrido. Portanto, a expressão do gene BCR-ABL em células hematopoiéticas
é responsável por liderar a manifestação clínica da LMC (ASNAFI e cols., 2019).
Cromossomo 9
normal
Cromossomo 22
normal
Cromossomo 9
alterado
Cromossomo 22
alterado
(cromossomo
Philadelphia)
Figura 1. Translocação recíproca entre os cromossomos 9 e 22, formando o cromossomo
Philadelphia (Ph1) (adaptado de National Cancer Institute, 2015).
Com relação às opções de tratamento dos pacientes com LMC, o transplante alogênico de
medula óssea apresenta potencial curativo, sendo capaz de erradicar o cromossomo Ph1.
Contudo, este procedimento ainda apresenta riscos de morbidade e mortalidade. Por esse motivo,
deve ser indicado apenas em casos selecionados, sendo uma opção apenas para pacientes com
quadro de saúde relativamente estável e que possuem doador compatível (JABBOUR e
KANTARJIAN, 2018).
Outras estratégias no tratamento da LMC incluem a utilização de inibidores seletivos da
proteína bcr-abl (como mesilato de imatinibe, desatinibe e nilotinibe) e quimioterapia (como
vincristina e daunorrubicina) (MASCHAN e cols., 2016; SINGH e COUMAR, 2018; CHIBA e
cols., 2018). No entanto, a resistência das células tumorais aos quimioterápicos é um dos
principais obstáculos ao sucesso do tratamento da LMC em crise blástica (ZHOU e cols., 2018).
1.2. Mecanismos relacionados à Resistência a Múltiplas Drogas
A resistência a múltiplas drogas (MDR - do inglês, Multidrug Resistance) é um caso de
resistência adquirida, observada em células tumorais, in vivo e in vitro, que consiste na ausência
de resposta aos quimioterápicos, tanto àquele que foi utilizado inicialmente, quanto a outros

22
empregados como alternativa. A MDR ocorre quando as células tumorais apresentam resistência
a classes de fármacos com mecanismos de ação diferentes, que não possuem estrutura molecular
relacionada, nem farmacológica (GOTTESMAN e PASTAN, 1993).
Existem vários mecanismos envolvidos com a resistência, que incluem a superexpressão
de bombas de efluxo de drogas (como transportadores ABC), alteração do ciclo celular, inibição
da morte celular, aumento do reparo do DNA e do metabolismo de xenobióticos (CREE e
CHARLTON, 2017).
1.2.1. Transportadores ABC
A superexpressão de bombas de efluxo de droga, como os transportadores ABC (ATP-
Binding Cassette), tais como P-gp (P-glycoprotein), MRP-1 (Multidrug Resistance-associated
Protein) e BCRP (Breast Cancer Resistance-protein) causa resistência a diversos agentes
quimioterápicos durante o tratamento contra o câncer. Dentre estes, a P-gp foi a primeira a ser
caracterizada por sua capacidade de conferir um fenótipo de resistência a quimioterápicos em
células cancerígenas (JULIANO e LING, 1976). A superexpressão de P-gp tem sido relatada
como um fator independente de pior prognóstico no desfecho clínico de pacientes com diferentes
tipos de cânceres como, orteossarcoma, câncer de mama e leucemia mieloide (JARAMILLO e
cols., 2018). No entanto, sabe-se que esse transportador também é constitutivamente expresso em
vários órgãos envolvidos na absorção e eliminação de fármacos, como intestino, fígado, rim e
cérebro, contribuindo para o sistema de defesa do nosso organismo contra substâncias tóxicas e
xenobióticos (PAN e cols., 2016).
A P-gp (Figura 3) é uma proteína de membrana, codificada em humanos pelo gene de
resistência a múltiplas drogas 1 (MDR1). Possui peso molecular de 170 kD e é composta por 12
hélices transmembranares organizadas em dois domínios citoplasmáticos intercalados ligantes de
ATP (GOTTESMAN e LING, 2006).

23
Meio extracelular
Meio intracelular
Quimioterápicos
Sítio ativo
(hidrólise de ATP)NH2
NBD1 NBD2 COOH
1 2 3 4 86 75 9 10 11 12
A B A B
S S
Figura 3. Modelo hipotético bidimensional da glicoproteína-P humana, baseada na análise
da sequência de aminoácidos e seus domínios funcionais (adaptado de DI PIETRO e cols.,
1999).
Os substratos da P-gp são substâncias hidrofóbicas,como os quimioterápicos vincristina e
daunorrubicina. Por sua característica hidrofóbica, esses substratos atravessam a membrana
plasmática por difusão passiva. Entretanto, devido à atividade transportadora da P-gp, ocorre o
efluxo dessas substâncias, o que impede seu acúmulo dentro da célula. Como os quimioterápicos
devem atingir concentrações suficientes nas células tumorais para exercerem seus efeitos,
alterações no efluxo da droga podem ser responsáveis pela aquisição de quimiorresistência contra
uma variedade de drogas anticâncer (JOYCE e cols., 2015).
1.2.2. Ciclo celular
O ciclo celular é um processo por meio do qual uma célula somática duplica seu material
genético e o reparte igualmente às suas células-filhas. A etapa inicial desse processo de divisão
celular é conhecida como intérfase. A intérfase é dividida em três fases: Gap phase 1 (G1);
síntese de DNA (S); Gap phase 2 (G2), durante a qual a célula se prepara para a mitose. Na
mitose ocorre a separação dos cromossomos e, consequentemente, a divisão celular propriamente
dita, finalidade maior do ciclo celular (VERMEULEN e cols., 2003; SALAZAR-ROA e cols.,
2017).
A progressão do ciclo celular de uma fase para outra é controlada pela ativação e
inativação sequencial de muitas proteínas que monitoram o estado da célula. Caso ocorram
mutações nos genes que codificam tais proteínas, a progressão do ciclo celular será

24
comprometida, podendo gerar resistência aos agentes neoplásicos. Com isso, proteínas
regulatórias do ciclo celular são importantes alvos na terapia do câncer (ABOU-ZAHR
e BORTHAKUR, 2017). A diminuição da proliferação da célula tumoral por meio do bloqueio
do ciclo celular em alguma fase específica permite o reparo do DNA ou a indução de proteínas
pró-apoptóticas (PANI e NUDLER, 2017).
1.2.3. Morte celular
A morte celular programada é extremamente importante para todos os organismos
multicelulares, uma vez que apresenta diversas funções, incluindo eliminação de forma eficaz de
células danificadas, sendo a apoptose o tipo mais conhecido.
A apoptose caracteriza-se por alterações morfológicas tais como condensação da
cromatina (pyknosis), degradação do DNA e blebbling de membrana que culminam na liberação
de pequenas partículas envoltas por membrana contendo componentes celulares conhecidos como
corpos apoptóticos que são rapidamente identificados pelas células vizinhas ou fagócitos e,
geralmente, eliminados sem indução de inflamação ou tecido cicatricial (SCHWEICHEL e cols.,
1973; XU e cols., 2019). A cascata de sinalização para apoptose leva à ativação de proteases
como as caspases (do inglês, Cysteine-dependent Aspartate-specific Proteases), responsáveis pela
execução da morte celular (XU e cols., 2019).
As caspases são sintetizadas como zimogênios inativos e, por isso, necessitam de ativação
para exercerem suas funções. A ativação completa é alcançada por meio da clivagem de um pró-
domínio por outra caspase. As caspases envolvidas na apoptose são classificadas em caspases
iniciadoras e caspases efetoras. As caspases iniciadoras (caspases-2, -8, -9 e -10) estão envolvidas
na iniciação da cascata proteolítica, enquanto as caspases efetoras (caspases-3, -6 e -7) são
responsáveis pela clivagem de diversos substratos citoplasmáticos e nucleares relacionados a
sobrevivência celular (CLARK, 2016). Entre os diversos substratos das caspases é possível citar a
enzima poli (ADP-ribose) polimerase-1 (PARP-1), cuja clivagem é um importante marcador da
morte celular por apoptose.
A apoptose pode ser iniciada por meio de duas vias: extrínsica (ou via do receptor de
morte) ou intrínsica (ou mitocondrial). Apoptose por via extrínseca é induzida por sinais
extracelulares que resultam na interação de ligantes específicos com receptores de morte
pertencentes à família dos receptores do fator de necrose tumoral (rTNF - do inglês, Tumor

25
Necrosis Factor Receptor). Essa interação é capaz de promover a ativação da cascata de
sinalização de caspases: pro-caspase-8 sofre auto-clivagem, gerando caspase-8 ativa que inicia a
fase de execução da morte celular; caspase-8, então,cliva a pro-caspase-3, gerando caspase-3
ativa (XU e cols., 2019).
A via intrínseca é ativada em resposta a diversas situações de estresse, incluindo danos no
DNA. Esses estímulos causam aumento da permeabilidadeda membrana mitocondrial, o que
libera algumas proteínas do espaço intermembranal para o citosol que, por sua vez, vão contribuir
para ativação das caspases. As proteínas da família Bcl-2 são reguladoras essenciais dessa via,
sendo classificadas em anti-apoptóticas (Bcl-2 e Bcl-xL) e pró-apoptóticas (Bax e Bak). As
proteínas pró-apoptóticas se oligomerizam e formam poros na membrana mitocondrial, por meio
do qual ocorre liberação de citocromo c. No citosol, o citocromo c se liga a APAF-1 (do inglês,
Apoptosis Protease Activating Fator-1), induzindo a formação de um complexo, conhecido como
apoptossoma, que requer caspase-9. No apoptossoma, a caspase-9 é clivada e ativada, levando a
ativação de outras caspases, como a caspase-3 (XU e cols., 2019).
No entanto, mesmo que a apoptose seja um mecanismo extremamente bem regulado, é
bastante comum que as células de câncer desenvolvam mecanismos de evasão a esse tipo de
morte, ocasionando falha do tratamento quimioterápico (HANAHAN e WEINBERG, 2011). Por
exemplo, muitas células com fenótipo MDR expressam níveis aumentados de IAP (do inglês,
Inhibitors of Apoptosis Proteins) (SHI e cols., 2007), inclusive em células de leucemia (SILVA e
cols., 2013). Por isso, a reativação das vias apoptóticas nas células MDR consiste em um
importante mecanismo de defesa capaz de promover a eliminação dessas células de uma maneira
eficiente (PAN e cols., 2016).
Outra abordagem contra as células MDR é a identificação de compostos capazes de
induzir morte não-apoptótica. Já foi descrito que a P-gp seria capaz de inibir a apoptose, mas não
teria a capacidade de modular a Morte Celular Independente de Caspase (MCIC) (ZU e cols.,
2014). Sendo assim, caso um composto tenha ação sobre a via de sinalização independente de
caspases poderia ser seletivamente citotóxico para as células com fenótipo MDR que
superexpressam P-gp.
Apesar de ainda não ter sido descrito um marcador específico para a MCIC, algumas
modificações na morfologia celular já foram observadas: membrana plasmática irregular,

26
encolhimento de núcleo e ausência de corpos apoptóticos, fragmentação do DNA e blebbing
celular (LAVALLARD e cols., 2009).
De maneira semelhante a apoptose, a MCIC também causa aumento da permeabilidade da
membrana mitocondrial, no entanto, de maneira contrária a apoptose, esse fenômeno ocorre
mesmo na ausência da ativação de caspases (GIAMPAZOLIAS e cols., 2018). Apesar de ainda
não ter sido totalmente elucidado quais seriam as consequências das alterações na membrana da
mitocôndria (TAIT e cols., 2014), uma das hipóteses sugere a ocorrência de prejuízos na
respiração celular, causando deficiência na produção energética (LARTIGUE e cols., 2009).
Já foi descrito que a disfunção da cadeia respiratória poderia ser compensada pela
melhoria da eficiência da glicólise. Algumas linhagens celulares, inclusive de LMC que
superexpressam uma enzima da via glicolítica, GAPDH (do inglês, Glyceraldehyde 3-phosphate
dehydrogenase), apresentam proteção contra estresses provocados pela MCIC, uma vez que
possuem maior eficiência glicolítica, o que permite que continuem se proliferando
(LAVALLARD e cols., 2009). Por isso, a superexpressão de GAPDH parece estar envolvida com
a maior resistência das células a MCIC.
1.2.4. Poli (ADP-ribose) polimerase-1 (PARP-1)
A poli (ADP-ribose) polimerase-1 (PARP-1) é a mais abundante da superfamília PARP
que é constituída por 18 proteínas (SODHI e cols., 2010). Essa enzima apresenta diversas funções
relacionadas tanto à promoção do reparo do DNA e sobrevivência celular, bem como indução de
morte celular. Além disso, parece haver uma correlação entre a inibição da PARP-1 e o
desenvolvimento do fenótipo MDR.
Esses mecanismos serão abordados com mais detalhes nos subtópicos a seguir.
1.2.4.1. Reparo do DNA
As células de câncer são capazes de desenvolver diversos mecanismos para detectar e
reparar danos no DNA, permitindo a formação de proteínas íntegras e garantindo o bom
funcionamento e proliferação celulares. Por isso, a elevada capacidade de reparo do DNA em
células tumorais pode limitar a eficácia de abordagens terapêuticas cujos mecanismos de ação
envolvem a indução de danos no DNA, gerando resistência celular durante a quimioterapia e
radioterapia (RAJAWAT e cols., 2017).

27
A PARP-1 é uma proteína nuclear de 116 kDa que promove o reparo de quebras no DNA
por excisão de base, mantendo a integridade do genoma celular. Essa enzima se liga a estruturas
danificadas do DNA e recruta outras enzimas de reparo, servindo como um fator de
sobrevivência para a célula. PARP-1 catalisa a transferência intracelular de unidades de ADP-
ribose para as proteínas nucleares, a partir da nicotinamida adenina dinucleotídeo (NAD+),
levando à formação de polímeros poli (ADP-ribose) (PAR) (RAY-CHAUDHURI e
NUSSENZWEIG, 2017).
Nesse contexto, os inibidores de PARP-1 têm sido utilizados em combinação com agentes
genotóxicos no tratamento de diferentes tipos de câncer, como: mama, ovário, próstata, pâncreas
(KAMEL e cols., 2018) e leucemias (WU e cols., 2019), os quais apresentam deficiência no
reparo do DNA. A inibição de PARP-1 é uma estratégia capaz de melhorar a atividade de drogas
antitumorais por meio da inibição da necrose e ativação de apoptose, resultando na diminuição da
dose utilizada do fármaco e dos efeitos colaterais provocados pelo tratamento (WU e cols., 2019).
Além de serem utilizados como co-adjuvantes no tratamento do câncer, os inibidores de PARP-1
também podem ser empregados como agentes isolados capazes de eliminar as células de câncer
com defeito no reparo do DNA (RAJAWAT e cols., 2017). Sendo assim, esses compostos
representam um grande avanço na terapia-alvo do câncer por serem capazes de atingir
seletivamente as células tumorais através da sua deficiência em uma via de reparo especifica.
No entanto, já foi demonstrado que a utilização de inibidores de PARP-1 em células que
superexpressam P-gp constitui um desafio, uma vez que essa bomba de efluxo extrui alguns
desses compostos para o meio extracelular, reduzindo sua ação (MONTONI e cols., 2013).
O inibidor de PARP-1 Olaparibe já foi descrito como substrato da P-gp em um modelo
experimental in vivo de câncer de mama em camundongos, sendo capaz de induzir a expressão
desse transportador (ROTTENBERG e cols., 2008). Outro estudo realizado com células de
câncer de ovário com fenótipo MDR também observou resistência à ação do Olaparibe. No
entanto, foi possível reverter esse mecanismo por meio da utilização de inibidores de P-gp, como
elacridar, zosuquidar e valspodar. Por isso, os inibidores de PARP-1 que não são substratos de P-
gp, como o Veliparibe, parecem ser mais promissores no tratamento de células MDR (LAWLOR
e cols., 2014).

28
1.2.4.2. Apoptose
O nível de dano ao DNA parece ser crucial na escolha entre a sobrevivência celular ou
indução de um mecanismo de morte. Quando o dano no DNA é demasiadamente extenso e
irreparável, as caspases-3 e 7 sofrem ativação e estas, por sua vez, podem clivar e inativar a
PARP-1. A clivagem de PARP-1 por essas caspases gera os fragmentos de 89 kDa e 24 kDa,
importantes marcadores bioquímicos da ocorrência de apoptose (SOLDANI e SCOVASSI, 2002;
CHAITANYA e cols., 2010; BOOZARI e cols., 2015).
A inibição da PARP-1 torna-se necessária para impedir o reparo do DNA, e,
consequentemente, a sobrevivência celular. Além disso, a clivagem da PARP-1 impede a
depleção de energia (NAD+ e ATP) na célula (BERGER, 1985), e, consequentemente, a morte
celular por necrose. Para a célula manter a PARP-1 ativa ocorre elevado consumo de NAD+ e
ATP. Como várias vias metabólicas dependem de NAD+, é fundamental repor os seus níveis, no
entanto, para que o NAD+ seja reciclado, ocorre aumento do gasto de ATP. Com isso, a
inativação da PARP-1 promove redução da demanda energética celular, o que possibilita que a
célula tenha ATP suficiente para ocorrência de apoptose (D'AMOURS e cols., 2001).
1.2.4.3. Partanatos
Partanatos é uma forma de morte celular programada, estudada há relativamente menos
tempo do que os outros tipos, mediada pela super-ativação da PARP-1, cuja ativação de caspases
não é obrigatória. Diversos trabalhos recentes têm sugerido a indução de partanatos como uma
nova estratégia capaz de eliminar células cancerígenas (AKHIANI e cols., 2014; MA e cols.,
2016; ZHENG e cols., 2016; JIANG e cols., 2018).
Caracteriza-se pela rápida depleção de energia (NAD+ e ATP) e pela síntese e acúmulo do
polímero PAR, levando à externalização de fosfatidilserina e dissipação do potencial de
membrana mitocondrial. Além disso, PAR também atua como um sinal de morte capaz de
provocar a liberação de AIF (do inglês, Apoptosis Inducing Factor) da mitocôndria. AIF, então,
sofre translocação para o núcleo, causando condensação nuclear, fragmentação do DNA em larga
escala e morte celular (YU e cols., 2002; ANDRABI e cols., 2006; DAVID e cols., 2009).

29
1.2.4.4. P-gp
Como abordado previamente, a utilização de inibidores de PARP-1 em células de câncer,
que apresentam deficiência de mecanismos de reparo do DNA, pode auxiliar durante o
tratamento quimioterápico. Em contrapartida, já foi descrito que células de camundongos knock
down para PARP-1 seriam capazes de desenvolver o fenótipo MDR, uma vez que poderiam
apresentar aumento concomitante da expressão e atividade da P-gp, embora ainda não se saiba ao
certo qual seria a razão para essa correlação (WURZER e cols., 2000; WESIERSKA-GADEK,
2005). No entanto, PARP-1 parece ser um promissor alvo terapêutico contra o câncer, seja por
meio da sua inibição ou ativação, dependendo do modelo experimental em questão.
1.2.5. Metabolismo de xenobióticos e substâncias endógenas
Durante o processo de transformação de uma célula normal em cancerígena, ocorrem
alterações no metabolismo celular, podendo levar ao aparecimento do fenótipo MDR (CREE,
2017). A ativação de enzimas responsáveis pela biotransformação de quimioterápicos
(xenobióticos) em metabólitos inativos, que podem ser facilmente excretados, é uma das
estratégias da célula tumoral capaz de gerar a resistência ao tratamento (LOLODI e cols., 2017).
Em geral, o metabolismo ocorre em duas fases, conhecidas como fase I e fase II. As
reações químicas mais comuns na fase I são oxidação, redução e hidrólise (GUENGERICH,
2007; GUENGERICH, 2008). Já na fase II, geralmente ocorrem reações de conjugação entre os
fármacos ou metabólitos formados na fase I com um substrato endógeno hidrofílico, como ácido
glicurônico, sulfato, acetatos endógenos, glutationa e aminoácidos (JANCOVA, 2010). A
principal função dessas reações é a transformação dos compostos em metabólitos, a fim de
facilitar sua excreção e eliminação do organismo.
Recentemente, a importância dos transportadores ABC para o metabolismo de drogas
também passou a ser reconhecida, sendo classificados como metabolismo de fase III. Essas
proteínas regulam a entrada e a saída de compostos exógenos e, consequentemente, seus efeitos
farmacológicos. Com isso, a inibição dessas bombas de efluxo poderia restabelecer as
concentrações dos quimioterápicos no interior da célula de câncer, revertendo o fenótipo de
resistência (DÖRING e PETZINGER, 2014).
O fígado é o principal órgão responsável pelo metabolismo de compostos, no entanto,
outros órgãos e tecidos também desempenham este papel, como intestino, rins, pulmões, plasma,

30
hemácias, placenta, pele e cérebro (FAN e LANNOY, 2014). Dentre as enzimas fundamentais
para esse processo de biotransformação, destacam-se as proteínas do complexo CYP450 (do
inglês, Cytochrome P450) envolvidas no metabolismo de fase I (AKHDAR e cols., 2012). O
CYP450 é considerado o principal responsável pela ativação e eliminação de fármacos. Apesar
de ser uma superfamília, com 57 isoformas distribuídas em 18 famílias e 44 subfamílias, apenas
cerca de 12 CYP's, pertencentes às famílias 1, 2 e 3, são responsáveis pelo metabolismo da
maioria de compostos (ZANGER e SCHWAB, 2013).
Alguns substratos podem induzir ou inibir essas enzimas. Quando ocorre inibição
enzimática pode haver a diminuição do metabolismo do composto, levando ao aumento de sua
concentração plasmática e, consequentemente, prolongando seu efeito, além de poder provocar
toxicidade farmacológica. E, caso ocorra indução enzimática, poderá haver um aumento do
metabolismo, levando à redução da concentração plasmática do composto, reduzindo seu efeito
terapêutico (ZANGER e SCHWAB, 2013).
Diversos xenobióticos induzem a expressão de enzimas por meio da via de sinalização do
PXR (do inglês, Pregnane X Receptor), um receptor nuclear que regula transcricionalmente
enzimas envolvidas no metabolismo de fármacos e proteínas transportadoras de drogas (MOORE
e cols., 2000). Uma vez que o PXR controla a expressão de genes relacionados ao metabolismo
de compostos anticâncer, pesquisas recentes têm investigado seu potencial papel na aquisição da
MDR (CHEN, 2010). Gupta e cols. (2008) demonstraram que a ativação de PXR induz a
expressão de diferentes genes como CYP2B6 e CYP3A4, o que contribui para a resistência de
células de câncer de ovário a ação de quimioterápicos. Além disso, PXR regula a expressão dos
genes CYP3A4 e MDR1 em várias linhagens celulares, aumentando o metabolismo e eliminação
de agentes anticâncer (AKHDAR e cols., 2012).
Em outros casos, porém, esse processo de metabolização também pode gerar metabólitos
com atividade biológica ainda mais potente do que sua molécula de origem, como ocorre com os
pró-fármacos (ZHANG e cols, 2017). A descoberta de pró-fármacos é uma estratégia promissora
contra o câncer, sendo capaz de melhorar a eficiência e a seletividade dos compostos citotóxicos.
Um clássico exemplo é a ciclofosfamida, um agente alquilante utilizado no tratamento do câncer
que precisa ser metabolizado pelas CYP's no fígado a fim de que possa ter atividade terapêutica
(RODRIGUEZ-ANTONA e INGELMAN-SUNDBERG, 2006).

31
Existem evidências de que a indução de alguns membros da CYP450 pode ser
responsável pela metabolização de compostos, tornando-os mais efetivos contra células tumorais
(SWANSON e cols., 2010). Já foi demonstrado que a expressão de CYP3A4 em tumor cerebral
de ratos confere sensibilidade dessas células ao quimioterápico nemorrubicina (LU e cols., 2009).
No entanto, a inibição da expressão de CYP1B1 em uma variedade de cânceres humanos poderia
impedir a progressão tumoral, mostrando ter benefício terapêutico (CASTRO e cols., 2008). Por
isso, a ativação e inibição diferencial dos membros da família do CYP450 pode gerar
citotoxicidade seletiva em células de câncer.
Sendo assim, pesquisas sobre o papel do metabolismo de compostos anticâncer na
aquisição de resistência à quimioterapia têm contribuído parao desenvolvimento de novas
abordagens, beneficiando-se da especificidade do metabolismo das células tumorais.
Além da importante função das CYP’s sobre o metabolismo de fase I de xenobióticos,
essas enzimas também apresentam papel relevante sobre o metabolismo de substratos endógenos,
como esteroides e hormônios. Esse sistema pode funcionar como um ciclo fútil de NADPH
oxidase, oxidando NADPH na ausência de substrato, produzindo superóxido e outros ROS (do
inglês, Reactive Oxygen Species) (HANUKOGLU e cols., 2006; JIANG e cols., 2015).
Posteriormente, essas espécies reativas de oxigênio, particularmente os superóxidos, são
protonados e formam o radical perhidroxila, contribuindo para a peroxidação lipídica e
desestabilização das membranas celulares (SINHA e cols., 2013).
Com isso, a modulação das CYP’s poderia provocar o aumento/ diminuição da
metabolização de diversos compostos, influenciando nos seus efeitos biológicos, ou então seria
capaz de contribuir para a geração de ROS nas células, prejudicando seu funcionamento.
1.3. Sensibilidade Colateral
Uma estratégia para superar a MDR é a identificação de substâncias que, em
concentrações inócuas a células denominadas sensíveis a quimioterapia, sejam capazes de reduzir
a viabilidade de células MDR. Esse fenótipo de sensibilidade apresentado pelas células MDR é
chamado sensibilidade colateral (CS - do inglês, Collateral Sensitivity) (HALL e cols., 2009). A
identificação de agentes promotores de CS em células MDR é um grande desafio para os
pesquisadores por ser um fenômeno considerado raro, uma vez que existem poucas substâncias
conhecidas. Dentre estas, destacam-se as que possuem maiores efeitos promissores: verapamil,

32
tiopronina, NSC73306, Dp44mT, Desmosdumotina B e 2-Deoxy-D-glicose(PLUCHINO e cols.,
2012). No entanto, os mecanismos pelos quais esses compostos induzem CS ainda não foram
completamente elucidados.
Atualmente, existem apenas algumas hipóteses sobre os possíveis mecanismos de CS
sobre células que apresentam expressão de transportadores ABC. Alguns estudos indicam que a
membrana das células MDR é menos estável, uma vez que células que superexpressam P-gp
apresentam alterações nas propriedades biofísicas da membrana plasmática, resultando numa
estrutura menos rígida. Portanto, os agentes promotores de CS causariam, preferencialmente,
perturbações na membrana dessas células resistentes, como aumento da fluidez e da
permeabilidade, comprometendo a integridade celular e, consequentemente, a viabilidade celular
(CALLAGHAN e RIORDAN, 1995).
Outra hipótese relacionada com a CS é baseada em observações de que diversos agentes
promotores de CS são substratos da P-gp, e, por isso, seriam capazes de promover o estímulo da
atividade ATPásica dessa bomba de efluxo. Esses compostos estariam sendo constantemente
internalizados e extruídos para o meio intra e extracelular, em um processo conhecido como
"ciclo fútil", levando, portanto, a um aumento na taxa de depleção de ATP intracelular
(LABERGE e cols., 2009; GOTTESMAN e cols., 2009). Isso, possivelmente, promoveria um
aumento na taxa de fosforilação oxidativa e um aumento de ROS, podendo levar a célula
resistente a apoptose (PLUCHINO e cols., 2012).
1.4. Fitoquímicos
Os ensaios clínicos com os clássicos agentes reversores, moduladores da MDR ou, ainda,
quimiossensibilizadores, como o bloqueador de canais de cálcio, verapamil (CORNWELL e
cols., 1987), foram limitados devido a sua toxicidade (KIM e cols., 2014). Por isso, nos últimos
anos, buscou-se descobrir compostos efetivos e com baixa citotoxicidade em células normais,
capazes de modular o fenótipo MDR.
Nesse contexto, os fitoquímicos são de vital importância para a saúde humana, uma vez
que são capazes de atingir diferentes alvos moleculares em células de câncer e, ainda, apresentam
baixo custo, baixa toxicidade em células normais e fácil disponibilidade (NYAMAI e cols.,
2016).

33
Os compostos bioativos ou fitoquímicos são metabólitos secundários presentes nas
plantas, sendo considerados substâncias não-nutricionais, porém de vital importância para a
saúde, uma vez que podem atuar no metabolismo e na fisiologia humana, reduzindo o risco de
desenvolvimento de doenças crônicas não transmissíveis, como doenças cardiovasculares,
diabetes mellitus e câncer (KIBE e cols., 2017). São classificados de acordo com suas estruturas
químicas (Figura 4) e podem ser divididos em cinco grandes grupos: carotenoides, compostos
fenólicos, alcaloides, compostos nitrogenados e compostos organossulfurados (SAMY e
GOPALAKRISHNAKONE, 2010).
34
Figura 4. Alimentos funcionais e seus compostos bioativos (adaptado de SURH, 2003).
Os compostos bioativos são encontrados nos alimentos funcionais. De acordo com a
Agência Nacional de Vigilância Sanitária em 1999, o alimento ou ingrediente que alegar
propriedades funcionais ou de saúde deve, além de exercer funções nutricionais básicas, quando
se tratar de nutriente, produzir efeitos metabólicos e fisiológicos, e ou efeitos benéficos à saúde,
devendo ser seguro para consumo sem supervisão médica (ANVISA, 1999).

35
O órgão que regulamenta os alimentos funcionais nos Estados Unidos é a Academia de
Nutrição e Dietética. Sua definição inclui os alimentos integrais, fortificados, enriquecidos ou
restauradores, que apresentam, potencialmente, efeitos benéficos para a saúde, quando
consumidos como parte de uma dieta variada. Para produzir o efeito desejado, os compostos
bioativos devem estar presentes nos alimentos funcionais, em quantidades suficientes e
adequadas. No entanto, as evidências científicas ainda são escassas para assegurar a quantidade
desses alimentos que deve ser ingerida diariamente para prevenir determinadas doenças, por isso,
poucos alimentos apresentam uma quantidade recomendada diária (HASLER e cols., 2009;
CROWE e cols., 2013).
Dentre os milhares de compostos bioativos presentes em alimentos, muitos são
identificados em ervas e especiarias, as quais têm sido alvos frequentes de pesquisas científicas
relacionando com suas propriedades benéficas para a saúde (BOWER e cols., 2016; YASHIN e
cols., 2017).
1.4.1. Piperina
A piperina (Figura 4) é um alcaloide que pode ser isolado, principalmente, da espécie
Piper nigrum, cujos frutos são conhecidos tradicionalmente como pimenta-do-reino (NISHA e
cols., 2009). O teor de piperina nesses frutos varia em torno de 5-9%, sendo considerada o
principal alcaloide e composto bioativo presente nos grãos de pimenta-preta, o qual confere o
sabor pungente (MAHDY e cols., 2012; MEGHWAL e GOSWAMI, 2013).
1.4.1.1. Biodisponibilidade e metabolismo
A piperina pode apresentar-se, de forma minoritária, nas três formas isoméricas:
isopiperina, isochavicina e chavicina (Figura 5). A exposição da piperina à luz leva ao aumento
da sua isomerização e, consequentemente, perda da sua atividade biológica, uma vez que os
benefícios associados à saúde são atribuídos, em sua maioria, à piperina (KOZUKUE e cols.,
2007).

36
Figura 5. Estruturas químicas dos isômeros da piperina (BUDAVARI, 1996).
De acordo com alguns estudos conduzidos em ratos, foi observada meia-vida de cerca de
2 horas para a piperina, no entanto, o tempo necessário para atingir a concentração máxima desse
fitoquímico no sangue variou entre 1 e 6 horas devido as diferentes dosagens e vias de
administração (ZHONG FENG e cols., 2010; SAHU e cols., 2014; SHAO e cols., 2015; LI e
cols., 2016). Além disso, piperina apresentou biodisponibilidade em torno de 25 % em ratos (LI e
cols., 2016).
Estudos sobre a farmacocinética da piperina em humanos são escassos, no entanto, uma
pesquisa com voluntários saudáveis observou que após a ingestão de duas doses de 24 mg de
piperina durante um dia, sua concentração sérica foi em torno de 6 µM (VOLAK e cols., 2013).
Considerando que na pimenta-do-reino teria em torno de 9% de piperina (MAHDY e cols., 2012;
MEGHWAL e GOSWAMI, 2013), os voluntários desse estudo ingeriram o equivalente a 267 mg
de pimenta-preta duas vezes ao dia.
Um estudo realizado nos Estados Unidos da América estimou o consumo médio de
pimenta-do-reino, baseando-se em populações que utilizam regularmente esse condimento em
sua dieta. O consumo diário desses indivíduos variou entre 17 mg e 31 mg (KINDELL, 1984). Na
população indiana este consumo é estimado em torno de 330 mg, quase dez vezes maior do que o
observado para os norte-americanos (PRADEEP e cols., 1993). Essas estimativas indicam que
seria inviável o consumo de 267 mg de pimenta-do-reino duas vezes ao dia a fim de atingir 6 µM
de piperina no sangue, mesmo considerando como padrão a população que mais ingere esse
condimento, os indianos.

37
Um recente trabalho publicado por Gao e cols. (2017) identificou, em ratos, os
metabólitos da piperina em amostras de sangue, bile e fezes após sua administração via oral.
Foram detectados e identificados 12 metabólitos, sendo que alguns destes foram reportados pela
primeira vez na literatura científica. Além disso, também foram descritas as principais vias
metabólicas da piperina nesse modelo experimental: redução, hidroxilação, glucuronidação e a
sulfatação (Figura 6). Esses dados sugerem que a piperina estaria sendo metabolizada por
enzimas de fase I e II, apesar desse estudo não ter identificado quais enzimas específicas
participariam desse processo.
Oxidação
Oxidação
Oxidação
Glucuronidação
Redução
Demetilação
Redução
Piperina
Figura 6. Proposta das vias metabólicas da piperina em ratos (Adaptado de GAO e cols.,
2017).

38
Recentemente, Zabela e cols. (2018) realizaram um estudo in vitro a fim de investigar o
metabolismo da piperina e de seus análogos utilizando microssomos hepáticos humanos. Dentre
as enzimas de fase I que foram testadas (CYP3A4, CYP2D6, CYP2C9, CYP1A2, CYP2C8 e
CYP2A6) somente a enzima hepática CYP1A2 foi responsável pela metabolização desse
composto.
Com relação ao metabolismo da piperina em humanos, CLAUS e GERHARD (1984)
detectaram e caracterizaram três metabólitos da piperina na urina de atletas (Figura 7).
Figura 7. Estruturas químicas dos metabólitos da piperina em humanos (Adaptado de
CLAUS e GERHARD, 1984).
É provável que os metabólitos encontrados em humanos, por Claus e Gerhard (1984)
sejam os mesmos que foram identificados pelas siglas M06, M07 e M08 (Figura 6) no estudo de
Gao e cols. (2017) com ratos. Esses dados sugerem que a metabolização da piperina gere alguns
metabólitos semelhantes em humanos e ratos.
A identificação de metabólitos ainda é um desafio devido à complexidade da matriz
biológica e à baixa concentração de metabólitos encontrados em amostras biológicas (GAO e

39
cols., 2017). A relação entre o metabolismo da piperina e os seus efeitos biológicos em humanos
ainda é de extrema relevância, uma vez que ainda não está claro se a sua biotransformação em
metabólitos é necessária para que possa apresentar efeitos para a saúde ou se sua própria
molécula original já seria responsável pelas suas propriedades benéficas. Por isso, novos estudos
devem ser conduzidos a fim de elucidar essas questões a respeito da metabolização desse
fitoquímico.
Enquanto os estudos sobre o metabolismo da piperina são raros, existem diversas
pesquisas, inclusive ensaios clínicos, que buscam compreender a capacidade desse composto em
aumentar a biodisponibilidade e eficácia de outras substâncias, incluindo, antibióticos, anti-
inflamatórios e compostos bioativos de outros alimentos. A piperina foi a primeira molécula a ser
validada cientificamente, em 1979, como um bioenhancer. Essa substância, mesmo em baixas
concentrações, é capaz de aumentar a biodisponibilidade de diferentes medicamentos em 30-
200%, sendo, portanto considerada um biopotencializador (ATAL e BEDI, 2010). Com relação
aos mecanismos de ação relacionados com esse efeito, alguns autores já relataram que a piperina
seria capaz de inibir a bomba de efluxo P-gp e enzimas metabolizadoras de xenobióticos, como
CYP3A4 e CYP2B1, e, assim, promoveria o aumento da absorção e do tempo de ação de
inúmeros compostos (REEN e WIEBEL, 1997; BHARDWAJ e cols., 2002; SINGH e cols.,
2013; SHAMSI e cols., 2017; LEE e cols., 2018).
1.4.1.2. Efeito antitumoral
Acredita-se que os estudos científicos sobre o efeito da piperina na saúde tenham sido
iniciados há quase 200 anos atrás (CARPENTER, 1828). Atualmente, existem inúmeras
comprovações dos benefícios desse composto para a saúde, incluindo: efeito anti-hipertensivo,
antiplaquetário, analgésico, anti-inflamatório, antidiarreico, antiespasmódico, antipirético,
hepato-protetor, bactericida, fungicidade, larvicida e inseticida (SRIVASTAVA e SINGH, 2017).
Piperina também apresenta importante ação quimiopreventiva e quimioterapêutica em
estudos in vitro e in vivo. Uma recente revisão publicada por Manayi e cols. (2018) concluiu,
ainda, que essas evidências sobre o efeito anticâncer da piperina poderiam subsidiar futuros
ensaio clínicos.
Foi encontrado um total de 144 trabalhos científicos na base de dados Pubmed (acesso em
27 de fevereiro de 2019) sobre o efeito da piperina contra o câncer. Entre os mecanismos de ação

40
sugeridos, destacam-se: a modulação de enzimas metabolizadoras de xenobióticos, atividade
antioxidante e pró-oxidante, efeito antiproliferativo, regulação do ciclo celular, indução de
apoptose e autofagia, potencial anti-metastático e reversão da MDR (RATHER e cols., 2018).
Um dos mecanismos responsáveis pelo efeito quimiopreventivo da piperina está
relacionado à sua capacidade de modulação de enzimas metabolizadoras de xenobióticos,
promovendo inibição de enzimas de fase I (CYP450), responsáveis pela ativação de pró-
carcinógenos, ou então indução de enzimas de fase II (glutationa peroxidase) que possuem
atividade antioxidante (SELVENDIRAN e cols., 2005).
Outros estudos têm evidenciado que a piperina contribui para a redução do
desenvolvimento do câncer devido à prevenção de danos às células e aos tecidos pela inibição de
reações de oxidação causadas por ROS (MANOHARAN e cols., 2009; VELLAICHAMY e cols.,
2009; LIU e cols., 2010).
Piperina apresentou efeitos contra os danos ao DNA causados pelo carcinógeno
benzo(a)pireno em células do pulmão, reduzindo a formação de radicais livres e a proliferação de
células cancerígenas em camundongos (SELVENDIRAN e cols., 2004a; SELVENDIRAN e
cols., 2004b).
Enquanto o efeito antioxidante da piperina protege as células normais do nosso
organismo, prevenindo o desenvolvimento do câncer, esse composto também é capaz de atuar
como um potente pró-oxidante em células tumorais, contribuindo para o tratamento contra essa
doença. Normalmente altas concentrações de piperina levam ao efeito pró-oxidativo. O estudo de
Gunasekaran e cols. (2017) sugeriu que a piperina induziu células de câncer hepático a apoptose
via geração de ROS mitocondrial. A piperina também foi capaz de gerar ROS em carcinoma oral
de células escamosas, levando a perda de potencial da membrana mitocondrial, ativação de
caspases e bloqueio do ciclo celular (SIDDIQUI e cols., 2017). Além disso, esse composto
induziu apoptose via geração de ROS em células de adenocarcinoma retal (YAFFE e cols., 2013).
Estudos in vitro sugerem que a piperina seria capaz de inibir a proliferação por meio do
bloqueio do ciclo celular na fase G1 em células de câncer de próstata (OUYANG e cols., 2013;
BA e MALHOTRA, 2018), cólon (YAFFE e cols., 2015) e melanoma (FOFARIA e cols., 2014)
ou então na fase G2/M em carcinoma oral de células escamosas (SIDDIQUI e cols., 2017) e em
osteossarcoma (ZHANG e cols., 2015).

41
Além disso, a piperina se mostrou capaz de induzir apoptose em vários modelos
experimentais de câncer, incluindo de cólon (YAFFE e cols., 2015), melanoma (FOFARIA e
cols., 2014), pulmão (LIN e cols., 2014), mama (GREENSHIELDS e cols., 2015), carcinoma oral
de células escamosas (SIDDIQUI e cols., 2017), carcinoma hepatocelular (GUNASEKARAN e
cols., 2017), próstata (BA e MALHOTRA, 2018) e células de câncer de ovário (SI e cols., 2018).
A piperina também parece modular a autofagia, processo de conservação celular em que
materiais citoplasmáticos são degradados e reciclados, a fim de atender as demandas energéticas
da célula e de manter a homeostasia celular (CHIANG e cols., 1989). No entanto, sabe-se que
esse mecanismo também pode levar as células de câncer à morte celular nos casos de ativação
excessiva dessa via (LI e cols., 2017). A piperina inibiu o crescimento e a proliferação de células
de câncer de próstata por meio da indução da autofagia (OUYANG e cols., 2013). Além disso, a
piperina mostrou ser capaz de inibir mTOR, um regulador negativo da autofagia, em células de
câncer de cólon (MOREAU e KAUR, 2017).
Estudos recentes indicam que a piperina teria, ainda, um papel de inibição da transição
epitélio-mesenquimal, processo biológico que permite que uma célula epitelial sofra
modificações bioquímicas, moleculares e morfológicas, resultando na aquisição de um fenótipo
de célula mesenquimal, com maior capacidade de migração, invasão e resistência a apoptose
(CHAFFER e cols., 2016; KIM e cols., 2017; SHIBUE e WEINBERG, 2017). Além disso, a
piperina também inibe a angiogênese por meio da modulação do Fator de Crescimento Vascular
Endotelial, que promove o crescimento de novos vasos sanguíneos (TALIB e cols., 2017).
Já foi demonstrado também que a piperina pode atuar de maneira sinérgica com a
quimioterapia, radioterapia ou com outros compostos bioativos de alimentos. A piperina
sensibiliza células de câncer de mama, que superexpressam o proto-oncogene HER2, para ação
da droga anticâncer paclitaxel (DO e cols., 2013); potencializa o efeito citotóxico da radioterapia
em camundongos com melanoma e em células de câncer de cólon (TAK e cols., 2012);
potencializa o efeito anticâncer do tamoxifeno contra células de câncer de mama (KHAMIS e
cols., 2018); apresenta efeito sinérgico com paclitaxel em células de adenocarcinoma de ovário
(PAL e cols., 2016); o tratamento com a combinação de curcumina com piperina em ratos
provoca inibição do carcinoma hepatocelular induzido pelo carcinógeno dietilnitrosamina
(PATIAL e cols., 2015). Esses dados sugerem um possível uso desse composto como adjuvante
na terapia do câncer.

42
A piperina também é capaz de modular o fenótipo MDR em alguns modelos
experimentais, como de câncer de mama, pulmão, cólon e linfoma. Esse composto inibiu a
expressão gênica e atividade das bombas de efluxo P-gp e BCRP (em células de câncer de mama)
e da bomba de efluxo MRP-1 (em células de câncer de pulmão) (LI e cols., 2011). Um trabalho
recente também demonstrou a ação desse fitoquímico sobre células de câncer de cólon e linfoma
com fenótipo MDR, por meio da potencialização do efeito citotóxico dos quimioterápicos
doxorrubicina e mitoxantrona, respectivamente. Além de ter inibido a atividade da P-gp nesses
dois tipos de cânceres (LI e cols., 2018). Outra pesquisa recentemente publicada por Morsy e
cols. (2018) constatou, por meio de estudos in silico e in vitro, que a piperina foi capaz de inibir a
P-gp em células de câncer de ovário, sugerindo que esse fitoquímico seria um promissor
adjuvante no tratamento com doxorrubicina.
O efeito da piperina em leucemias humanas foi pouco investigado. Apenas um trabalho
foi encontrado na literatura científica (base de dados Pubmed), ressaltando, mais uma vez, a
importância da presente pesquisa para a área em questão. No estudo citado, foi visto que
concentrações de 20 ou 40 μM de piperina levaram as células K562 à diferenciação em
macrófagos/ monócitos, sendo que a indução desse mecanismo pode ser considerada uma
estratégia eficaz para o tratamento da leucemia (SONG e cols., 2008).
1.5. Modelo experimental de LMC
Nesta tese foram utilizadas três linhagens celulares de LMC: K562, linhagem parental
sensível à quimioterapia e as linhagens MDR, Lucena-1 e FEPS, que foram derivadas da K562.
As células K562 pertencem a uma linhagem originada de uma LMC em crise blástica
eritroleucemia humana e caracteriza-se por expressar o oncogene quimérico BCR-ABL no
cromossomo Philadelphia (KOEFFLER e GOLDE, 1980).
As linhagens celulares Lucena-1 e FEPS foram desenvolvidas no Laboratório de
Imunologia Tumoral da Universidade Federal do Rio de Janeiro (RUMJANEK e cols., 2001;
DAFLON-YUNES e cols., 2013) e foram selecionadas a partir da linhagem K562 devido à
exposição a concentrações crescentes dos quimioterápicos vincristina e daunorrubicina,
respectivamente.
Essas linhagens diferem entre si tanto sob o aspecto funcional (DAFLON-YUNES e cols.,
2013), quanto o seu transcriptoma (MOREIRA e cols., 2014) e o seu proteoma (VIDAL, 2017).

43
Podem ser destacadas algumas diferenças entre essas células: Lucena-1 e FEPS possuem um
proteoma menor (VIDAL, 2017), o que poderia indicar que se encontram mais diferenciadas do
que a K562; também possuem um ciclo mais lento, isto é, se dividem menos (DAFLON-YUNES
e cols., 2013), sugerindo que apresentam maior resistência à ação de quimioterápicos que atuam
durante a etapa de proliferação celular; as linhagens de leucemia apresentam grandes diferenças
na expressão de genes relacionadas à morte celular (MOREIRA e cols., 2014), apontando para a
possibilidade de ocorrência de mecanismos de resistência à morte nas células MDR.
As linhagens Lucena-1 e FEPS também são mais resistentes ao estresse oxidativo que a
linhagem sensível a drogas, K562, provavelmente devido a maior capacidade antioxidante das
células com fenótipo MDR. Diversas enzimas antioxidantes estão diferentemente expressas nas
linhagens Lucena-1 e FEPS, em comparação com a K562. Um exemplo disso é o sistema da
tiorredoxina/peroxirredoxina que se encontra aumentado nas células com fenótipo MDR, Lucena-
1 e FEPS (MOREIRA e cols., 2014; VIDAL, 2017), e é considerado crucial para a defesa contra
o estresse oxidativo em células de câncer (MONTEIRO e cols., 2017).
Lucena-1 também apresenta alta atividade da catalase, enzima envolvida com a proteção
de ROS, especificamente com a conversão do peróxido de hidrogênio em água e oxigênio,
provavelmente conferindo a essas células proteção contra a radiação UVA, contra H202 (peróxido
de hidrogênio) (TRINDADE e cols., 1999) e contra agentes quimioterápicos (RUMJANEK e
col., 2001). Um recente trabalho realizado por Soares e cols. (2019) também identificou na FEPS
maior resistência a UVA e H202, em relação a K562, sendo que essa linhagem MDR é ainda mais
resistente à ação de quimioterápicos do que a Lucena-1 (DAFLON-YUNES e cols., 2013). Além
disso, a atividade da catalase também está elevada na linhagem FEPS (VIDAL, 2017). Dessa
forma, diversos compostos anticâncer que agem por meio da indução de estresse oxidativo
tornam-se ineficazes.
No entanto, Lucena-1 e FEPS se mostraram sensíveis a outro tipo de radiação (UVB)
(TRINDADE e cols., 1999; SOARES e cols., 2019). A indução de ROS pela UVB é causada pelo
aumento de moléculas de oxigênio reativas como oxigênio singlete e peróxido de hidrogênio,
além de radicais livres hidroxila (SCHARFFETTER-KOCHANEK e cols., 2000; RAVANAT e
cols., 2001). Além disso, esse tipo de radiação também causa dano direto no DNA da célula,
principalmente devido à interação com bases heterocícilicas, formando dímeros de pirimidina

44
(RAVANAT e cols., 2001). Sendo assim, a radiação UVB parece atuar por diferentes
mecanismos de ação, sendo capaz de afetar até mesmo as células MDR.
Além disso, a linhagem Lucena-1 apresenta cinco vezes mais cópias do gene MDR1 (que
codifica a P-gp) que sua linhagem parental K562 (RUMJANEK e col., 2001), sendo que a
linhagem FEPS possui expressão e atividade da P-gp ainda maior do que a Lucena-1 (DAFLON-
YUNES e cols., 2013). Com isso, em ambas linhagens MDR, a expressão de P-gp parece ser o
principal mecanismo de resistência (DAFLON-YUNES e cols., 2013; RUMJANEK e cols.,
2013). Contudo, sabe-se que o fenótipo MDR tem origem multifatorial, podendo gerar alterações
no ciclo celular, inibição da morte celular, aumento do reparo do DNA e do metabolismo de
xenobióticos (CREE e CHARLTON, 2017).
A fim de investigar a importância da expressão de P-gp para o desenvolvimento do
fenótipo MDR, foi analisado o perfil proteômico das linhagens Lucena-1 e FEPS silenciadas para
MDR1 (que codifica a P-gp) (VIDAL, 2017). Foi demonstrado que o silenciamento da P-gp
provocou diversas alterações em proteínas de outras vias de sinalização e metabólicas, sugerindo
uma interconexão entre os diferentes mecanismos de resistência (VIDAL, 2017).
Portanto, os mecanismos de resistência a quimioterápicos desenvolvidos durante a
aquisição do fenótipo MDR parecem ser extremamente complexos e vão muito além da
superexpressão de transportadores de efluxo. Por isso, são necessárias múltiplas abordagens
experimentais a fim de identificar as vias de sinalização alteradas.

45
2. JUSTIFICATIVA
O fenômeno da MDR é considerado o principal obstáculo no tratamento de vários tipos de
câncer, dentre eles a LMC em crise blástica. Os pacientes que evoluem para crise blástica
apresentam o pior prognóstico com mediana de sobrevida de menos de 1 ano. Uma das
estratégias mais promissoras contra a MDR é a identificação de substâncias capazes de atuar
seletivamente sobre células MDR. Esse fenótipo de sensibilidade apresentado pelas células MDR
é chamado CS, um fenômeno considerado raro, uma vez que poucas substâncias apresentam essa
ação.
Além disso, a identificação de compostos capazes de modular a resistência por meio de
mecanismos como regulação do ciclo celular, indução de morte celular e ativação de enzimas
metabolizadoras de xenobióticos (promovendo ativação de compostos anticâncer), pode ser uma
estratégia contra o câncer.
Entre os compostos capazes de reverter a resistência de células a quimioterápicos está a
piperina. Apesar de já terem sido demonstradas evidências científicas promissoras sobre sua ação
anticâncer e anti-MDR, ainda não foi investigado seu efeito na promoção de CS.
Dessa forma, torna-se relevante investigar os efeitos da piperina na indução de CS em
células de LMC em crise blástica com fenótipo MDR.

46
3. OBJETIVOS
2.1. Objetivo geral
Investigar os efeitos da piperina sobre diferentes mecanismos de resistência a
quimioterápicos em células de LMC em crise blástica.
2.2. Objetivos específicos
Avaliar o efeito da piperina sobre a viabilidade e morfologia das células de LMC humana
sensível a drogas, K562, e sobre linhagens de LMC com fenótipo MDR, Lucena-1 e
FEPS;
Verificar, nas linhagens de leucemia K562, Lucena-1 e FEPS, a ação da piperina sobre a:
distribuição do ciclo celular;
indução de morte celular;
modulação de GAPDH, PARP-1, PAR e P-gp;
expressão gênica do receptor PXR e de enzimas de fase I (CYP3A4, CYP3A5 e
CYP1B1);
Investigar a interação da piperina com a proteína CYP3A5;
Analisar nas linhagens de leucemia K562, Lucena-1 e FEPS a metabolização da piperina;

47
4. MÉTODOS
4.1. Culturas de células
As linhagens K562 e Lucena-1 foram mantidas em uma concentração de 2 × 104 céls/mL
e foram cultivadas em um volume final de 5 mL de meio RPMI-1640, suplementado com 10% de
soro fetal bovino (SFB), 100 unidades/mL de penicilina G e 100 μg/mL de sulfato de
estreptomicina, 5 μM de β-mercaptoetanol e 2 g/L de bicarbonato de sódio. As células da
linhagem FEPS foram mantidas como descrito para K562 e Lucena-1, porém em um volume final
de 10 mL de meio de cultivo. Na cultura de Lucena-1 foi adicionada vincristina a uma
concentração final de 60 nM e na cultura de FEPS foi adicionada daunorrubicina a uma
concentração de 466 nM, a fim de manter suas características de MDR. As células foram
mantidas a 37 °C em uma atmosfera umidificada contendo 5% de CO2 e foram subcultivadas a
cada três ou quatro dias (DAFLON-YUNES e cols., 2013).
4.2. Ensaio de citotoxicidade - Redução de MTT (3-(4,5 dimethyl thiazole-2yl)-2,5 diphenyl
tetrazolium bromide) com as células de leucemia
As células de leucemia foram plaqueadas em placas de 96 poços com fundo chato.
Posteriormente, as células foram tratadas com diferentes concentrações de piperina dissolvida em
0,25% de dimetilsulfóxido (DMSO) ou com os respectivos volumes de DMSO 0,25% (v/v;
controle).
Após o tratamento por 48, 72 e 96 horas foi realizado o ensaio de redução de MTT. O
MTT é um composto que é reduzido pelas desidrogenases mitocondriais, formando os cristais de
formazan, que possuem uma coloração roxa e podem ser quantificados por densidade ótica. Para
isso, foram adicionados 20 µL de MTT, diluído em Phosphate-Buffered Saline (PBS), a uma
concentração final de 0,5 mg/mL em cada poço. As placas foram mantidas na estufa a 37 °C e
com 5% de CO2 por 3 horas. Posteriormente, as placas foram centrifugadas a 200 × g, durante 7
minutos, o sobrenadante foi descartado e os cristais de formazan formados foram dissolvidos em
200 µL de DMSO. A intensidade de coloração foi medida em um leitor de microplacas
(Biochrom Anthos Zenyth 200 ST), a um comprimento de onda de 492 nm (adaptado de
MOSMANN, 1983). Para a análise da viabilidade celular, foi realizado o cálculo do percentual de
células viáveis em relação aos respectivos controles dos experimentos. Os valores de IC50 (do
inglês, Half Maximal Inhibitory Concentration) foram calculados a partir das curvas dose-

48
resposta, sendo definido como a concentração da droga necessária para reduzir o número de
células viáveis a 50% em relação ao controle. O programa GraphPad Software 5.0 foi utilizado
para os cálculos de IC50.
4.3. Ensaio de citotoxicidade - Teste de exclusão com azul de tripan
O número de células viáveis foi avaliado após 72 horas de tratamento das células de
leucemia com 50 e 100 μM de piperina dissolvida em 0,25% de DMSO ou com o respectivo
volume de DMSO 0,25% (v/v; controle). Foram transferidos 10 μL da suspensão de células para
eppendorfs, em seguida, foi adicionado 10 μL de solução de azul de tripan a 0,4% (v/v). Após
homogeneização, 10 μL do total do volume foram utilizados para a contagem das células em
câmara de Neubauer. Para a análise da viabilidade celular, foi realizado o cálculo do percentual
de células viáveis em relação aos respectivos controles dos experimentos. Esse método de
contagem de células é baseado no princípio de que as células vivas possuem membrana intacta e
conseguem excluir o azul de tripan, já as células mortas, em necrose ou apoptose tardia,
incorporam esse corante e ficam marcadas, permitindo contagem diferenciada entre células vivas
e mortas (BARILE, 1994).
4.4.Ensaio de citotoxicidade - Redução de MTT com macrófagos peritoneais
Animais: Duas fêmeas de camundongos albinos suíços (6-8 semanas) foram obtidas do
Biotério Thomas George do Instituto de Pesquisa em Fármacos e Medicamentos da Universidade
Federal da Paraíba. Os animais foram mantidos em condições controladas de temperatura (21 ± 1
ºC), ciclo de luminosidade (12 horas claro/ 12 horas escuro) e livre acesso à ração e à água. Após
a manipulação, a eutanásia foi realizada por meio de deslocamento cervical. Todos os
procedimentos adotados neste estudo foram aprovados pelo Comitê de Ética em Pesquisa do
Centro de Ciências da Saúde – CEP/ CCS/ UFPB (número de protocolo: 111/2016 - Anexo D).
Obtenção de macrófagos peritoneais: A inflamação peritoneal foi induzida pela injeção
intraperitoneal de 4 mL de tioglicolato a 4% (Sigma Chemical Co., St. Louis, USA). Após quatro
dias, os animais foram eutanasiados por deslocamento cervical e a cavidade peritoneal foi lavada
com 8 mL de PBS e 3% de SFB. A suspensão celular obtida da lavagem peritoneal foi
centrifugada (390 × g; 5 minutos; 4 ºC) e, em seguida, o sobrenadante foi descartado e as células
foram ressuspendidas em 1 mL de meio de cultura (RPMI + 10% SFB). As células foram

49
contadas com o auxílio de uma câmara de Neubauer utilizando azul de Tripan para identificação
somente das células viáveis.
Cultura de macrófagos: As células peritoneais foram plaqueadas em placas de 96 poços na
concentração de 4 × 105 células/ poço e incubadas por 24 horas na estufa a 37 °C e com 5% de
CO2. Em seguida, as células não aderentes foram removidas e as células remanescentes
(macrófagos) foram ainda incubadas por 24 horas na presença de diferentes concentrações de
piperina dissolvida em 0,25% de DMSO ou com o respectivo volume de DMSO 0,25% (v/v;
controle). Após 24 e 48 horas de cultura, a viabilidade celular foi analisada pelo método de
redução de MTT.
Ensaio de redução de MTT: O sobrenadante foi descartado e foi adicionado 90 µL de
meio de cultura e 10 µL de MTT, diluído em PBS, a uma concentração final de 0,5 mg/mL em
cada poço. As placas foram mantidas na estufa a 37 °C e com 5% de CO2 por 4 horas.
Posteriormente, as placas foram centrifugadas a 200 × g, durante 7 minutos, o sobrenadante foi
descartado e os cristais de formazan formados foram dissolvidos em 200 µL de DMSO. A
intensidade de coloração foi medida em um leitor de microplacas (Leitor Universal de
Microplacas EL 800, Bio-TEK), a um comprimento de onda de 570 nm (adaptado de
MOSMANN, 1983). Para a análise da viabilidade celular, foi realizado o cálculo do percentual de
células viáveis em relação aos respectivos controles dos experimentos.
4.5. Coloração por Panótico Rápido
A fim de identificar possíveis alterações na morfologia das células de leucemia após 72
horas de tratamento com 100 µM de piperina dissolvida em 0,25% de DMSO ou com os
respectivos volumes de DMSO 0,25% (v/v; controle) por 72 horas, foram realizados
citoesfregaços por 3 minutos a 500 rpm, utilizando a centrífuga Cytospin (Thermo Fisher
Scientific, USA) e, posteriormente, foi feita coloração com Panótico Rápido utilizando kit
hematológico comercial (Laborclin® produtos para laboratórios Ltda., Pinhais, PR, Brasil).
Seguindo as recomendações do fabricante, cada lâmina foi submetida à ação de um fixador
(solução de triarilmetano a 0,1%) por 1 minuto. Em seguida, as lâminas foram sequencialmente
imersas em duas soluções corantes (solução de xantenos a 0,1% e solução de tiazinas a 0,1%) por
1 minuto cada. Após a última imersão as lâminas foram lavadas em água destilada e secas ao ar

50
previamente à avaliação em um microscópio óptico de luz transmitida (OLYMPUS IX81). As
imagens foram analisadas usando o programa Olympus cellSens Standard.
4.6. Análise do ciclo celular
A distribuição do ciclo celular foi analisada por citometria de fluxo, como descrito
previamente por Pozo-Guisado e cols. (2002). As células de leucemia foram plaqueadas em
placas de 6 poços, com volume final de 3 mL/ poço na concentração final de 2 × 104 céls/mL. Em
seguida, foi feito o tratamento com 50 e 100 µM de piperina dissolvida em 0,25% de DMSO ou
com os respectivos volumes de DMSO 0,25% (v/v; controle). Após incubação por 72 horas, 1 ×
105 células foram coletadas, centrifugadas a 220 × g por 7 minutos à 4 °C e lavadas 2 vezes com
PBS, e então ressuspendidas em PBS contendo Triton X-100 0,1% (v/v), 50 µg/mL de PI (do
inglês, Propidium Iodide) e 1 mg/mL de RNase A. Em seguida, o conteúdo de DNA foi medido
em citômetro de fluxo (FACScalibur) equipado com o programa CellQuest (Becton Dickinson,
CA). A leitura do PI foi realizada pelo detector FL-2 área. A porcentagem de células nas fases
Sub-G1, G1, S e G2/M do ciclo celular foi quantificada usando o programa Summit v4.3®
(DAKO).
4.7. Ensaio de apoptose
A caracterização da morte celular foi investigada pela dupla marcação com PI e Anexina-
V-FITC (do inglês, Fluorescein isothiocyanate). A marcação por PI reflete um rompimento da
membrana plasmática, caracterizando necrose. Já a Anexina-V-FITC se liga ao fosfolipídio de
membrana, fosfatildiserina, que sofre externalização durante a apoptose. De forma similar,
quando ocorre a marcação dupla, considera-se apoptose tardia. O termo técnico é necrose
secundária, pois a marcação por PI ocorre após a permanência da célula apoptótica no meio de
cultura, de tal forma, que é secundária à marcação por Anexina-V-FITC. As células viáveis são
representadas pela ausência de qualquer marcação.
As células das linhagens de leucemia foram plaqueadas em placas de 6 poços, com
volume final de 3 mL/ poço na concentração final de 2 × 104 céls/mL. Em seguida, foi feito o
tratamento com 50 e 100 µM de piperina dissolvida em 0,25% de DMSO ou com os respectivos
volumes de DMSO 0,25% (v/v; controle). Etoposídeo (25 µM) foi utilizado como controle
positivo de indução de apoptose (SEN e cols., 2004; MOLAVI e cols., 2008). Após incubação

51
por 72 horas, as células foram centrifugadas a 220 × g por 7 minutos à 4 °C e ressuspendidas no
tampão de ligação do fabricante do kit de detecção de apoptose da eBioscience. Posteriormente,
foram transferidas para uma placa de 96 poços com fundo em U (200 µL/ poço) e foram
novamente centrifugadas a 220 × g por 7 minutos à 4 °C. Em seguida, foram incubadas com
tampão de ligação (96 µL) contendo Anexina-V-FITC (3 µL) por 20 minutos em temperatura
ambiente e protegidas da luz. Após o período de tempo referido, foram acrescentados 200 μL do
tampão de ligação e, no momento da leitura foram adicionados 4 μL de PI. A análise foi
realizada, em seguida, em citômetro de fluxo (FACScalibur) equipado com o programa CellQuest
(Becton Dickinson, CA). A leitura do PI e da Anexina-V-FITC foi realizada pelo detector FL-3 e
FL-1, respectivamente. A porcentagem de células marcadas com PI e/ou Anexina-V-FITC foi
quantificada usando o programa Summit v4.3® (DAKO).
4.8. Imunofluorescência
Os experimentos foram realizados de acordo com as instruções do fabricante. As células
de leucemia (2 × 104 céls/mL) foram tratadas com 100 µM de piperina por 72 horas.
Posteriormente, foram realizados citoesfregaços por 3 minutos a 500 rpm, utilizando a centrífuga
Cytospin (Thermo Fisher Scientific, USA) seguido de fixação com 4% de paraformaldeído por 15
minutos em temperatura ambiente e lavagem com PBS previamente à marcação por
imunofluorescência. Em seguida, as células foram permeabilizadas com 0,5% Triton X-100 em
PBS por 10 minutos. As células foram então mantidas com 1% albumina em PBS por 30 minutos
em temperatura ambiente e, posteriormente, incubadas com anticorpo contra capase-3 clivada
(1:400; Cell Signaling) diluído em 1% albumina overnight à 37 °C. Após sucessivas lavagens
com PBS, as células foram incubadas por 2 horas à temperatura ambiente com anticorpo
secundário anti-coelho conjugado com Alexa Fluor® 555 (Thermo Fisher Scientific) diluído em
PBS (1:1000) com DAPI (1 mg/ mL, Sigma Chemical Co., St. Louis, USA), para marcação
nuclear. As células foram então lavadas com PBS e montadas com para-fenilenodiamina. Cada
lâmina foi observada e fotografada em pelo menos três diferentes campos em um microscópio de
fluorescência (Spinning Disk) e as imagens foram então analisadas utilizando o programa Zen
Pro 2012 (blue edition).

52
4.9. Investigação da expressão de GAPDH, PARP-1 e PAR
4.9.1. Preparação do extrato proteico das células
Após o tratamento com 100 µM de piperina dissolvida em 0,25% de DMSO ou com os
respectivos volumes de DMSO 0,25% (v/v; controle) por 72 horas, as células foram centrifugadas
a 220 × g por 15 minutos a 4 °C, o sobrenadante foi removido, as células foram lavadas com PBS
e lisadas com nitrogênio líquido. Antes que as células descongelassem foi acrescentado um
tampão, contendo Tris-HCl 5 mM pH 7,4, EDTA (do inglês, Ethylenediamine Tetraacetic Acid)
10 mM, coquetel de inibidores de proteases (aprotinina A, leupeptina, bestatina, pepstatina A e E-
64) em concentração final de 1 mM, diluído 100 vezes, PMSF (do inglês, Phenylmethylsulfonyl
Fluoride) 1 mM, ortovanadato de sódio 1 mM, fluoreto de sódio 5 mM, óxido de fenilarsina 1
mM, ácido ocadáico 1 mM. Em seguida, este extrato obtido foi congelado e descongelado,
sonicado por 10 minutos e centrifugado por 5 minutos à 10.000 rpm. O sobrenadante resultante
da centrifugação foi considerado como o extrato proteico das células. A dosagem de proteína foi
realizada seguindo o protocolo descrito por Lowry, utilizando albumina bovina como padrão
(LOWRY e cols., 1951).
4.9.2. Eletroforese em gel de poliacrilamida em condições desnaturantes
Os extratos protéicos celulares (100 µg) foram submetidos à eletroforese em gel de
poliacrilamida à 10% na presença de SDS (do inglês, Sodium Dodecyl Sulfate) (LAEMMLI,
1970). O gel de corrida em condições desnaturantes continha Tris-HCl 375 mM, SDS 0,1 g%,
acrilamida 9,7 g%, bisacrilamida 0,3 g%, persulfato de amônio 0,033 g%, TEMED (do inglês,
Tetramethylethylenediamine) 0,055% (v/v), pH 8,8. O gel de empacotamento continha Tris-HCl
83 mM, SDS 0,07 g%, acrilamida 3,2 g%, bisacrilamida 0,09 g%, persulfato de amônio 0,073
g%, TEMED 0,123 % (v/v), pH 6,8. O tampão de corrida continha Tris-HCl 24,7 mM, glicina
190 mM, SDS 0,1 g%, pH 8,3. As proteínas foram ressuspendidas em tampão de amostra
contendo Tris-HCl 62,5 mM, glicerol 10% (v/v), azul de bromofenol 0,001 g%, SDS 2 g%, β-
mercaptoetanol 10%. Os padrões de massa molecular utilizados foram: miosina (204 kDa), β-
galactosidase (116 kDa), fosforilase B (97,4 kDa), albumina bovina (66 kDa), ovoalbumina (45
kDa) e anidrase carbônica (29 kDa). Após as corridas, utilizando corrente constante de 15 mA, os
géis foram submetidos a eletrotransferência.

53
4.9.3. Western blotting
As proteínas foram eletrotransferidas do gel para uma membrana de PVDF (do inglês,
Polyvinylidene Difluoride). A eletrotransferência foi realizada em tampão Tris-glicina 25 mM e
metanol 20% (v/v), pH 8,3 a 210 mA por 90 minutos à 4 ºC (TOWBIN e cols., 1979) utilizando-
se o sistema de transferência da Bio-Rad. A eficiência da transferência foi monitorada corando-se
a membrana com vermelho de Ponceau 0,5% (p/v) em TCA (do inglês, Trichloroacetic Acid)
10% (p/v) por 10 minutos. Posteriormente, a membrana foi descorada por lavagem com Tris-HCl
0,01 M pH 7,6 contendo 0,15 M de NaCl (TBS - do inglês, Tris-buffered Saline). A membrana
totalmente descorada do Ponceau, foi então bloqueada com Tris-HCl 0,01 M pH 7,6 contendo
0,15 M NaCl, 5% albumina e 0,1% de Tween 20 (TBS-Tween-Albumina) por 18 horas a 4 ºC.
Após 18 horas, o tampão de bloqueio foi desprezado e a membrana incubada com anticorpo
primário contra a proteína de interesse numa diluição de 1:1.000 por 2 horas à temperatura
ambiente. Em seguida, a membrana foi lavada 5 vezes com TBS-Tween e incubada por 1 hora à
temperatura ambiente com anti-IgG coelho conjugada com peroxidase diluída 1:5.000 na solução
de bloqueio. A membrana foi então lavada 5 vezes por 5 minutos com TBS-Tween e 2 vezes com
TBS. A imunodetecção das bandas foi visualizada por quimioluminescência, utilizando o kit ECL
Immobilon Western (Millipore Corporation, Bilerica, MA, USA), de acordo com as instruções
do fabricante. As membranas foram então digitalizadas em escaner para blot (C-Digit®, Li-Cor
Biosciences, Lincoln, Reino Unido). Os níveis de Heat Shock Protein 70 (Hsp70) foram
utilizados como controle interno para verificação do carregamento de proteínas no gel. Os
seguintes anticorpos primários foram utilizados: anti-PARP (#9542) (Cell Signaling Technology,
CA, EUA), anti-PAR (ab14459) (Abcam, Cambridge, MA, USA), anti-Hsp70 (sc33575) e anti-
GAPDH (0411) (Santa Cruz Biotechnology, Santa Cruz, CA, EUA). A quantificação das bandas
foi determinada por análise densitométrica, utilizando o programa Image J versão 1.43p (NIH,
EUA) e os resultados foram expressos em unidades arbitrárias (UA) conforme a sua comparação
ao controle Hsp70.
4.10. Análise da expressão de P-gp
As linhagens MDR, Lucena-1 e FEPS foram plaqueadas em placas de 6 poços, com
volume final de 5 mL/poço na concentração final de 2 × 104 céls/mL. Em seguida, foi feito o
tratamento com 50 e 100 µM de piperina dissolvida em 0,25% de DMSO ou com os respectivos

54
volumes de DMSO 0,25% (v/v; controle). Após incubação por 72 horas as células (1 × 105)
foram lavadas com PBS com 5% de SFB, e marcadas com anticorpo anti-P-gp (BD
Pharmingen™ Mouse Anti-Human P-glycoprotein - FITC) na diluição 1:5, no volume final de 10
µL, e mantidas a temperatura de 4 °C por 30 minutos. Posteriormente, as células foram
centrifugadas a 220 × g por 7 minutos, o sobrenadante foi descartado e as células foram
ressuspendidas em 300 µL de PBS com 5% de SFB para avaliação em citômetro de fluxo
(FACScalibur) equipado com o programa CellQuest (Becton Dickinson, CA). A leitura da
fluorescência foi realizada pelo detector FL-1. Os dados foram analisados utilizando o programa
Summit v4.3®(DAKO).
4.11. Análise da influência da piperina sobre a expressão gênica de GAPDH e de
marcadores do metabolismo de xenobióticos.
Após o tratamento com 100 µM de piperina dissolvida em 0,25% de DMSO ou com os
respectivos volumes de DMSO 0,25% (v/v; controle) por 72 horas foram realizadas análises por
RT-PCR (Reverse transcription polymerase chain reaction) em tempo real da expressão dos
genes que codificam GAPDH, PXR, e as enzimas de fase I, como citocromos P450 (CYP3A4,
CYP3A5, CYP1B1). Para isso, foram realizadas extrações dos mRNAs das amostras com reagente
Trizol®, de acordo com as instruções do fabricante. A concentração de RNA total foi
determinada em espectrofotômetro Nanovue (GE Healthcare, WI, EUA). Após o tratamento com
DNase, o cDNA foi sintetizado usando SuperScript III Reverse Transcriptase com 4 μg de RNA
purificado. A reação de transcrição foi realizada utilizando o kit Platinum® SYBR® Green qPCR
SuperMix em um termociclador Rotor-Gene® Q, aplicando 2 µL de cDNA concentrado em uma
mistura de reação de 10 μL de volume final contendo 0,30 μM de cada primer e 5 μL de SYBR
Green qPCR. As especificidades das reações foram avaliadas por meio do registro contínuo da
intensidade de fluorescência durante o melting na faixa de 65 °C a 95 °C. As sequências dos
oligonucleotídeos iniciadores foram obtidas da literatura científica (CHI e cols., 2009; AYED-
BOUSSEMA e cols., 2011) ou determinadas pelos pesquisadores envolvidos, sendo utilizados os
seguintes pares de oligonucleotídeos: GAPDH (senso, 5’-
CAACAGCCTCAAGATCATCAGCAA-3’ e anti-senso, 5’-
AGTGATGGCATGGACTGTGGTCAT-3’); PXR (senso, 5’-CATGAGGGGGGTAGCAAAGC-
3’ e anti-senso, 5’-TGCAGGGGATCTCCCTCTTC-3’); CYP3A4 (senso, 5’-

55
GCCTGGTGCTCCTCTATCTA-3’ e anti-senso, 5’-GGCTGTTGACCATCATAAAAG-3’);
CYP3A5 (senso, 5’-TGACCCAAAGTACTGGACAG-3’ e anti-senso, 5’-
TGAAGAAGTCCTTGCGTGTC-3’); CYP1B1 (senso, 5′-GGCCACTATCACTGACATCT-3′ e
anti-senso, 5′-ACAGTGTCCTTGGGAATGTG-3′); Hsp70 (5′-
TGTAACCCCATCATCAGCGG-3′ e 5′-TCCCAACAGTCCACCTCAAAG-3′). Os níveis de
expressão foram normalizados em relação àquele do gene Hsp70. Os controles negativos foram
realizados na ausência de cDNA. A expressão relativa foi analisada pelo método 2-Ct
(LIVAK e
SCHMITTGEN, 2001).
4.12. Docking molecular
A estrutura tridimensional do alvo enzimático CYP3A5 (receptor) foi obtida da base de
dados Protein Data Bank (PDB ID: 5VEU) (HSU e cols., 2018), enquanto a estrutura da piperina
(ligante) foi obtida da base de dados ZINC (ZINC ID: 1529772). O software AutoDock Vina
(TROTT e OLSON, 2010) foi utilizado para conduzir os estudos e o módulo AutoDock Tools foi
empregado para preparar e analisar as simulações computacionais (HUEY e MORRIS, 2008).
Durante o preparo do arquivo PDB do receptor foram removidos heteroátomos, ligantes e
moléculas de água. Além disso, foram adicionados átomos de hidrogênio polar ao receptor da
proteína e foram calculadas as cargas atômicas parciais. Para o preparo da molécula do ligante,
foram adicionados átomos de hidrogênio polares e não polares e foram calculadas as cargas de
Gasteiger para o ligante inteiro.
Para a realização de experimentos de docking molecular utilizando o Autodock Vina é
necessário a determinação de uma caixa de simulação (do inglês, grid box). Essa caixa delimita
onde o algoritmo de busca da ferramenta explorará as melhores posições e conformações do
ligante no sítio de ligação da proteína. Como para esta proteína-alvo não tínhamos a informação
exata do sítio de ligação, nas simulações realizadas neste trabalho, a caixa de simulação foi
posicionada envolvendo completamente a cadeia A da enzima CYP3A5. Assim, configuramos a
caixa com dimensão nos eixos X-, Y- e Z- de 40 Å; e as coordenadas do centro da caixa foram
definidas como x = 10,21 Å; y = 40,735 Å; z = 21,462 Å; considerando o parâmetro
exhaustiveness como 128. Esse parâmetro da ferramenta AutoDock Vina está relacionado com o
número de tentativas que o algoritmo de docking fará para buscar o melhor encaixe e energia de
interação estimada entre proteína-ligante. A proteína e o ligante foram mantidos rígidos durante o

56
processo de docking. Durante o processo de busca do algoritmo de docking, a energia livre de
ligação do ligante-receptor é estimada em Kcal/ mol para selecionar as melhores posições.
Quanto mais a energia livre de ligação for negativa, possivelmente maior a estabilidade do
complexo ligante-receptor. A análise visual dos resultados do docking foi realizada com o auxílio
do programa VMD. Por ocasião da análise visual dos resultados, comparou-se a posição ocupada
pela piperina com a dos ligantes cristalográficos Protoporphyrin IX containing Fe (HEM) e
Hitonavir (HIT) para inferir um possível sítio de alta afinidade.
4.13. Determinação da piperina intracelular e extracelular por Cromatografia Líquida com
Detecção por Arranjo de Diodos (CLAE-DAD)
Extração
As células de leucemia foram plaqueadas em placas de 6 poços com volume final de 5
mL/poço na concentração final de 2 × 104 céls/mL e, em seguida, foram tratadas com 100 µM de
piperina dissolvida em 0,25% de DMSO ou com os respectivos volumes de DMSO 0,25% (v/v;
controle) por 48 horas. Posteriormente, as células foram coletadas e centrifugadas a 220 × g por 7
minutos a 4 °C e 1 mL do sobrenadante foi coletado e armazenado a -20 °C para posterior análise
da piperina extracelular, enquanto o restante do sobrenadante foi descartado. A piperina presente
no interior das células do precipitado foi extraída com 1 mL de metanol, seguido de agitação em
vórtex durante 1 minuto, centrifugação a 4.000 rpm por 5 minutos e eliminação do solvente em
um concentrador à vácuo SpeedVac (Eppendorf, Hamburgo, Alemanha) a uma temperatura
constante de 30 °C. O resíduo foi então ressuspendido em 500 µL de fase móvel e filtrado em
filtro com diâmetro de 0,45 µm (adaptado de BAJAD e cols., 2002).
Condições cromatográficas
O sistema de cromatografia líquida (Shimadzu®, Japão) incluiu duas bombas paralelas
LC-20AD, injetor automático (SIL-20AHT), detector de arranjo de diodos (DAD) SPD-M20A e
sistema CBM-20A como controlador.
A separação cromatográfica da piperina e seus metabólitos foi realizada utilizando-se
coluna de fase reversa C18 (5 μm, 250 mm × 4,6 mm, Phenomenex®
), de acordo com a
metodologia adaptada de Bajad e cols. (2002). A fase móvel consistiu numa mistura de
acetonitrila com KH2PO4 (25 mM, pH 4,5, ajustado com ácido ortofosfórico) (45:55 v/v). As

57
análises foram realizadas em modo isocrático, com fluxo de 1 mL/minuto e volume de injeção de
20 µL. O eluente foi monitorado a 340 nm.
A identificação da piperina foi realizada por comparação com o tempo de retenção e
espectro de absorção do respectivo padrão. Foram calculados para cada linhagem celular de
leucemia os percentuais relativos à área dos picos detectados em relação à área total do
cromatograma. Posteriormente, esses percentuais foram comparados entre as linhagens a fim de
verificar se houve diferença em relação à transformação de piperina em outros compostos/
metabólitos. Os dados de integração foram adquiridos pelo software Lab Solutions (Shimadzu
Corporation®, version 5.82 SP1, 2008-2015).
4.14. Análise estatística
Os resultados dos experimentos acima descritos foram submetidos à análise estatística
descritiva e foram calculadas medidas de tendência central (média) e dispersão (erro padrão). As
diferenças entre as médias foram avaliadas por meio do emprego do Teste t de Student, não
pareado (ou pareado) ou análise de variância (ANOVA) seguida de pós-teste de Tukey ou
Dunnett, usando o programa GraphPad Prism 5. Resultados com valores de p < 0,05 foram
considerados estatisticamente significativos.

58
5. RESULTADOS
5.1. Efeito da piperina sobre a viabilidade das células de leucemia
Primeiramente, foi analisado o efeito da piperina sobre a linhagem celular sensível a
drogas, K562, e sobre as linhagens celulares com fenótipo MDR, Lucena-1 e FEPS, por meio do
ensaio de redução do MTT (Figura 8). Após o tratamento com diferentes concentrações de
piperina, que variaram de 2,5 a 300 µM, por 48 (Figura 8A), 72 (Figura 8B) e 96 horas (Figura
8C), os ensaios foram realizados.

59
48 horas
CTRL 2,5 5 10 25 50 75 100 150 300
0
20
40
60
80
100
120
140
Piperina (M)
K562
Lucena-1
FEPS
**
*
*
* *
*
*
**
*% C
élu
las v
iáveis
72 horas
CTRL 2,5 5 10 25 50 75 100 150 300
0
20
40
60
80
100
120
140
Piperina (M)
K562
Lucena-1
FEPS
*
*
*
**
*
** *
*
*
*
*
*
*
% C
élu
las v
iáveis
96 horas
CTRL 2,5 5 10 25 50 75 100 150 300
0
20
40
60
80
100
120
140
Piperina (M)
K562
Lucena-1
FEPS
**
*
*
*
*
*
*
**
*
***
% C
élu
las v
iáveis
A
B
C
Figura 8. Efeito da piperina sobre a viabilidade das células de leucemia K562, Lucena-1 e
FEPS pelo método de redução de MTT. As células foram incubadas com 0,5 mg/mL de MTT
por 3 horas a 37 °C e 5% de CO2. Após a incubação, os cristais de formazan foram dissolvidos
em DMSO. Viabilidade das células tratadas por 48 (A), 72 (B) e 96 horas (C) com concentrações
até 300 µM de piperina. O % de células viáveis foi calculado em relação ao controle com DMSO.
Os resultados são expressos como média ± erro padrão (n=3). * p < 0,05 (ANOVA com pós-teste
de Dunnett), comparado às células controle com DMSO. CTRL, controle.

60
De acordo com os resultados apresentados na Figura 8, foi observado que a piperina
reduziu a viabilidade das linhagens K562, Lucena-1 e FEPS de forma dose-dependente. Além
disso, a piperina demonstrou ser mais citotóxica nas linhagens MDR, Lucena-1 e FEPS, do que
na linhagem parental, K562, em todos os tempos de tratamento. A piperina em concentrações
iguais ou abaixo de 75 µM não afetou a K562 em nenhum dos tempos estudados, ao passo que a
concentração de 75 µM em 48 horas já foi capaz de diminuir a viabilidade da FEPS em 50%.
Além disso, as diferenças mais marcantes entre as linhagens foram percebidas após incubação
por 96 horas com piperina. Após esse período de tratamento, o efeito sobre a viabilidade das
células K562, Lucena-1 e FEPS foi significante (p < 0,05) a partir de 100 µM, 50 µM e 10 µM
de piperina, respectivamente. Estes resultados sugerem que as células MDR Lucena-1 e FEPS
apresentam maior sensibilidade a piperina que as células K562, caracterizando o fenômeno de
CS.
Também foi constatado, pelos valores de IC50, que a piperina foi mais tóxica na linhagem
celular FEPS, em relação a Lucena-1, de forma estatisticamente significativa (p < 0,05), após os
tratamentos por 72 e 96 horas. Os valores de IC50 da piperina para as células Lucena-1 nos
tempos de 72 e 96 horas foram praticamente o dobro dos encontrados para as células FEPS nos
mesmos períodos de tempo analisados (Tabela 1).
IC50 da piperina (µM)
Linhagem celular 48 horas 72 horas 96 horas
K562 106,8 ± 26,2 A,a 99,0 ± 2,3 A,a 104,4 ± 9,6 A,a
Lucena-1 102,5 ± 9,5 A,a 87,9 ± 3,9 A,ab 75,4 ± 2,9 B,b
FEPS 73,0 ± 5,8 A,a 41,5 ± 1,6 B,b 34,7 ± 4,1 C,b
Tabela 1. Efeito citotóxico da piperina (IC50) sobre as linhagens celulares K562, Lucena-1 e FEPS pelométodo de redução de MTT. Os valores de IC50foram calculados a partir das curvas dose-resposta.
Letras maiúsculas diferentes denotam diferença significativa entre as linhagens celulares no mesmotempo de incubação. p < 0,05 (ANOVA com pós-teste de Tukey).Letras minúsculas diferentes denotam diferença significativa entre os tempos de incubação na mesma
linhagem celular. p < 0,05 (ANOVA com pós-teste de Tukey).
A fim de confirmar estes resultados, foram realizados testes de citotoxicidade por ensaio
de exclusão do azul de tripan, que se baseiam na incorporação do corante azul de tripan pelas
células em apoptose tardia ou necrose, possibilitando a diferenciação entre células vivas e mortas.

61
Para isso, as linhagens K562, Lucena-1 e FEPS foram tratadas com 50 ou 100 µM de piperina
por 72 horas (Figura 9). Após esse período foi realizada a contagem celular com o corante.
K562 Lucena-1 FEPS0
20
40
60
80
100CTRL
PIP (50 M)
PIP (100 M)*
*
*
* *
*% C
élu
las v
iáv
eis
Figura 9. Efeito da piperina sobre a viabilidade das células de leucemia K562, Lucena-1 e
FEPS pelo método de exclusão de azul de tripan. As células foram incubadas com piperina nas
concentrações indicadas por 72 horas. Em seguida, foram marcadas com azul de tripan (0,4%)
(v/v). Os resultados são expressos como média e erro padrão (n=3). * p < 0,05 (ANOVA com
pós-teste de Dunnett), comparado às células controle com DMSO. CTRL, controle. PIP, piperina.
Foi observado que a piperina foi mais citotóxica para a linhagem FEPS do que para as
linhagens K562 e Lucena-1, ratificando os dados anteriores. No entanto, foi observado que as
diferenças de efeito desse composto entre as linhagens celulares foram maiores pelo ensaio de
redução de MTT. Por exemplo, a concentração de 50 µM, que não apresentava diferença na K562
após 72 horas de incubação em ensaios de MTT, pelo presente método, mostrou reduzir o número
total de células viáveis, se aproximando mais do percentual de células viáveis das demais
linhagens celulares após o tratamento com piperina. Como o MTT reflete o metabolismo da
mitocôndria, por meio da avaliação da atividade das desidrogenases mitocondriais, sugere-se que
a piperina tenha provocado menos alterações sobre o metabolismo celular da K562.
5.2. Efeito da piperina sobre a viabilidade de células não-tumorais
Uma vez que foi constatado o efeito citotóxico da piperina em células de leucemia
mieloide crônica, que contém precursores da série macrofágica, foi investigada a citotoxicidade
desse composto sobre macrófagos peritoneais de camundongos. Após incubação com piperina
por 24 ou 48 horas, foi realizado o ensaio de redução de MTT (Figura 10).
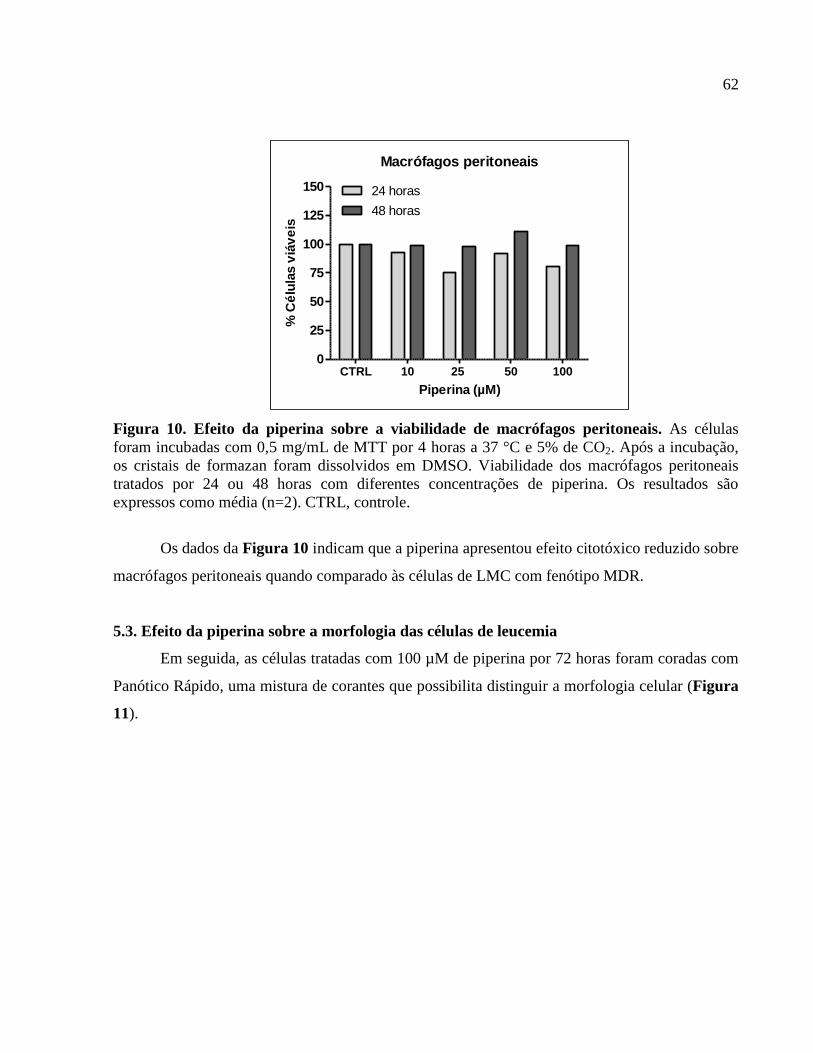
62
Macrófagos peritoneais
CTRL 10 25 50 1000
25
50
75
100
125
150 24 horas
48 horas
Piperina (µM)
% C
élu
las v
iáv
eis
Figura 10. Efeito da piperina sobre a viabilidade de macrófagos peritoneais. As células
foram incubadas com 0,5 mg/mL de MTT por 4 horas a 37 °C e 5% de CO2. Após a incubação,
os cristais de formazan foram dissolvidos em DMSO. Viabilidade dos macrófagos peritoneais
tratados por 24 ou 48 horas com diferentes concentrações de piperina. Os resultados são
expressos como média (n=2). CTRL, controle.
Os dados da Figura 10 indicam que a piperina apresentou efeito citotóxico reduzido sobre
macrófagos peritoneais quando comparado às células de LMC com fenótipo MDR.
5.3. Efeito da piperina sobre a morfologia das células de leucemia
Em seguida, as células tratadas com 100 µM de piperina por 72 horas foram coradas com
Panótico Rápido, uma mistura de corantes que possibilita distinguir a morfologia celular (Figura
11).

63
K5
62
Luce
na-
1FE
PS
CTRL PIP (100 µM)
10 µm 10 µm
10 µm
10 µm
10 µm
10 µm
Figura 11. Efeito da piperina sobre a morfologia das células de leucemia K562, Lucena-1 e
FEPS. As células foram incubadas com 100 µM de piperina por 72 horas. Em seguida, foram
coradas com Panótico Rápido e observadas em microscópio de luz transmitida (aumento de 63×).
As imagens são representativas de 3 experimentos independentes. CTRL, controle. PIP, piperina.

64
As imagens por microscopia de campo claro indicam que a piperina provocou alterações
morfológicas importantes nas células K562, Lucena-1 e, de forma ainda mais marcante, sobre a
FEPS. Na FEPS, foi observado aumento da formação e do volume de vacúolos no citoplasma em
comparação com as demais linhagens celulares (Figura 11).
Em seguida, por meio de citometria de fluxo, analisamos os parâmetros de tamanho (FSC-
H) e granulosidade (SSC-H) a fim de confirmar as modificações morfológicas induzidas pela
piperina nas diferentes linhagens celulares (Figura 12).
Controle
K562
Piperina (50 µM)
Piperina (100 µM)
Lucena-1
Controle
Piperina (50 µM)
Piperina (100 µM)
FEPS
Controle
Piperina (50 µM)
Piperina (100 µM)
A
B
D
E
G
H
C F I
Figura 12. Efeito da piperina sobre o tamanho e granulosidade das células de leucemia
K562, Lucena-1 e FEPS. As células foram incubadas com piperina nas concentrações indicadas
por 72 horas. Em seguida, foram analisadas em citômetro de fluxo. Linhagem K562 (A) controle
(B) incubada com 50 μM de piperina (C) ou incubada com 100 μM de piperina. Linhagem
Lucena-1 (D) controle (E) incubada com 50 μM de piperina (F) ou incubada com 100 μM de
piperina. Linhagem FEPS (G) controle (H) incubada com 50 μM de piperina (I) ou incubada com
100 μM de piperina. As imagens são representativas de 3 experimentos independentes. FSC
(Forward Scatter) - tamanho relativo da célula; SSC (Side Scatter) - granulosidade ou
complexidade celular.

65
Observa-se na Figura 12 que as células leucêmicas foram afetadas pelo tratamento com
piperina, de acordo com os parâmetros de tamanho e granulosidade. No entanto, a concentração
de 50 µM de piperina já foi suficiente para provocar alterações morfológicas na linhagem FEPS,
enquanto que nas linhagens K562 e Lucena-1, isso só foi mais marcante na concentração de 100
µM de piperina.
Além disso, foi realizada a quantificação do percentual de células com alterações em sua
morfologia, quando comparadas aos seus respectivos controles (Tabela 2).
K562
ControlePiperina(50 µM)
Piperina(100 µM)
0,7 ± 0,1 2,2 ± 0,6 1,7 ± 0,5
2,1 ± 0,2 4,7 ± 0,7 7,8 ± 1,7*
2,4 ± 0,2 7,4 ± 1,2 21,1 ± 4,0*
4,0 ± 0,4 11,8 ± 1,5 26,3 ± 4,5*
1,2 ± 0,2 2,2 ± 0,5 5,2 ± 1,3*
Lucena-1
ControlePiperina(50 µM)
Piperina(100 µM)
0,9 0,2 1,0 0,4 0,9 0,4
2,3 0,4 3,2 0,9 7,1 2,1
3,2 0,3 7,0 0,7 19,6 2,5*
4,6 0,4 8,9 0,5 26,1 2,5*
1,4 0,2 1,8 0,4 5,0 1,4*
FEPS
ControlePiperina(50 µM)
Piperina(100 µM)
0,7 ± 0,2 0,6 ± 0,2 0,5 ± 0,2
1,6 ± 0,3 3,7 ± 1,4 6,9 ± 2,6
1,9 ± 0,2 21,0 ± 3,2* 35,7 ± 3,8*
2,8 ± 0,3 24,2 ± 3,8* 42,1 ± 4,6*
0,9 ± 0,2 3,2 ± 1,2 6,4 ± 2,4*
Tabela 2. Percentual de células com alterações no seu tamanho e/ ou granulosidade em relação ao controle.
Os resultados são expressos como média ± erro padrão (n=3). * p < 0,05 (ANOVA com pós-teste de Dunnett), comparado àscélulas controle com DMSO. FSC (Forward Scatter) - tamanho relativo da célula; SSC (Side Scatter) - granulosidade oucomplexidade celular.
Região
R4 (FSC)
R4 + R2 (FSC)
R1 (SSC)
R1 + R2 (SSC)
R2 (FSC + SSC)
Foi observada uma tendência de aumento do percentual de células com o tamanho
alterado (R4 + R2 → FSC). Por exemplo, a concentração de 50 µM de piperina provocou
aumento do tamanho das células K562 e FEPS em pouco mais de duas vezes, e a concentração de
100 µM de piperina aumentou o tamanho das células K562 e Lucena-1 em torno de três vezes e
FEPS em quatro vezes. Apesar disso, somente a linhagem K562 tratada com 100 µM de piperina
apresentou diferença estatisticamente significativa (p < 0,05) em relação ao seu controle.
O principal parâmetro alterado nas três linhagens celulares foi a granulosidade (R1 ou R1
+ R2 → SSC). A linhagem FEPS foi a que apresentou o maior percentual de células com essa
característica, tanto após o tratamento com 50 µM (aumento de 8,6 em relação ao controle)
quanto com 100 µM de piperina (aumento de 15 vezes em relação ao controle). Além disso, o
percentual de células com alterações no tamanho e na granulosidade (R2 → FSC + SSC),
concomitantemente, foi semelhante entre as linhagens estudadas.

66
Sendo assim, esses dados estão de acordo com o observado na citologia, uma vez que
ambos sugerem que o efeito da piperina é mais expressivo na FEPS. Além disso, o acúmulo de
vacúolos citoplasmáticos poderia estar refletindo no aumento da granulosidade principalmente na
FEPS tratada com piperina.
5.4. Efeito da piperina sobre o ciclo celular e a morte celular nas células de leucemia
Em seguida, foram realizados experimentos com o intuito de verificar o efeito da piperina
sobre a distribuição do ciclo celular das linhagens K562, Lucena-1 e FEPS por citometria de
fluxo (Figura 13).

67
Controle
K562
Piperina (50 µM)
Piperina (100 µM)
Lucena-1
Controle
Piperina (50 µM)
Piperina (100 µM)
FEPS
Controle
Piperina (50 µM)
Piperina (100 µM)
A
B
D
E
G
H
C F I
Figura 13. Efeito da piperina sobre a distribuição do ciclo celular das células de leucemia
K562, Lucena-1 e FEPS. As células foram incubadas com piperina nas concentrações indicadas
por 72 horas. Após o tratamento, 1 105 células foram ressuspendidas em PBS contendo Triton
X-100 0,1% (v/v), 50 μg/mL de iodeto de propídio e 1 mg/mL de RNase A. O conteúdo de DNA
foi medido usando citômetro de fluxo. Linhagem K562 (A) controle (B) incubada com 50 μM de
piperina (C) ou incubada com 100 μM de piperina. Linhagem Lucena-1 (D) controle (E)
incubada com 50 μM de piperina (F) ou incubada com 100 μM de piperina. Linhagem FEPS (G)
controle (H) incubada com 50 μM de piperina (I) ou incubada com 100 μM de piperina. As
imagens são representativas de 3 experimentos independentes.
As condições testadas nesse estudo, de concentração e de tempo de exposição ao
tratamento, não foram suficientes para gerar acúmulo de células em qualquer fase do ciclo celular
nas linhagens estudadas (Figura 13). Além disso, confirmamos esses resultados por meio da

68
análise quantitativa das células nas fases G1, S e G2/M (Tabela 3). Esses dados sugerem que a
piperina não possui ação citostática, uma vez que não demonstrou ter efeito antiproliferativo.
K562
ControlePiperina(50 µM)
Piperina(100 µM)
3,2 ± 0,6 3,9 ± 0,3 6,0 ± 0,8*
43,2 ± 3,3 55,2 ± 5,3 47,7 ± 8,1
29,4 ± 1,2 26,6 ± 2,9 31,0 ± 2,1
21,6 ± 1,6 18,2 ± 3,0 14,9 ± 1,5
Lucena-1
ControlePiperina(50 µM)
Piperina(100 µM)
2,8 0,4 4,0 0,8 7,9 0,6*
43,7 1,4 44,4 3,5 42,8 1,7
22,9 1,1 24,9 2,0 26,5 1,2
28,8 0,8 29,9 2,3 20,6 1,2*
FEPS
ControlePiperina(50 µM)
Piperina(100 µM)
2,5 ± 0,6 3,5 ± 0,4 7,0 ± 0,5*
46,7 ± 2,8 50,9 ± 2,8 47,6 ± 5,0
22,3 ± 0,6 24,4 ± 1,2 19,9 ± 1,1
24,7 ± 1,0 19,2 ± 2,2 20,0 ± 2,0
Tabela 3. Percentual das células K562, Lucena-1 e FEPS nas fases Sub-G1, G1, S ou G2/M após tratamento com piperina.
Os resultados são expressos como média ± erro padrão (n=3). * p < 0,05 (ANOVA com pós-teste de Dunnett), comparado àscélulas controle com DMSO.
Fase
Sub-G1
G1
S
G2/M
Além da investigação da atuação da piperina sobre o bloqueio do ciclo celular, foram
analisadas as células com o DNA fragmentado, o que seria indicativo de morte celular (Figura
13). Para isso, foram quantificadas as células na fase Sub-G1. Apenas o tratamento com 100 µM
de piperina promoveu acúmulo de células nessa fase nas linhagens K562, Lucena-1 e FEPS
(Tabela 3). Além disso, pôde ser observada uma maior tendência em acumular células nessa fase
Sub-G1 nas linhagens MDR Lucena-1 e FEPS (2,8 vezes em relação ao controle) do que na
linhagem sensível a drogas, K562 (1,9 vezes em relação ao controle) após o tratamento com 100
µM de piperina.
Em seguida, foi investigado o mecanismo de morte celular induzido pela piperina nas
linhagens de leucemia. Primeiramente avaliamos a apoptose, forma de morte celular mais
estudada. Algumas características desse tipo de morte envolvem mudanças morfológicas do
núcleo celular (que se apresenta fragmentado), um encolhimento do volume celular e mudanças
na membrana plasmática com externalização do fosfolipídio de membrana fosfatidilserina. Em
modelos in vitro, a célula apoptótica não pode ser rapidamente destruída por macrófagos, por
isso, passa a perder o controle da permeabilidade da membrana, podendo vir a morrer por
necrose, fenômeno conhecido por apoptose tardia. Além disso, avaliamos também a possibilidade
de ocorrência de necrose, que pode ser caracterizada por inchaço, disfunção mitocondrial, ruptura
da membrana celular, derramamento do conteúdo citoplasmático para o espaço extracelular e lise
celular.

69
Nesse experimento foram utilizados os marcadores fluorescentes Anexina-V FITC e PI. A
Anexina-V é uma lectina que se liga à fosfatidilserina (indicando apoptose) e o PI tem a
capacidade de se ligar ao DNA, desde que a célula apresente sua membrana permeável
(indicando necrose) (Figura 14).

70
Controle negativo
K562
Controle positivo
Piperina (50 µM)
A
B
C
Lucena-1
Controle negativo
Controle positivo
Piperina (50 µM)
E
F
G
I
J
K
FEPS
Controle negativo
Controle positivo
Piperina (50 µM)
Piperina (100 µM)D Piperina (100 µM)H L Piperina (100 µM)
Figura 14. Efeito da piperina sobre a morte das células de leucemia K562, Lucena-1 e
FEPS. As células foram incubadas com piperina nas concentrações indicadas por 72 horas. Em
seguida, foram marcadas com Anexina V-FITC e iodeto de propídio e o resultado foi analisado
em citômetro de fluxo. Linhagem K562 (A) controle negativo (B) controle positivo (25 µM de
etoposídio) (C) incubada com 50 µM de piperina (D) ou incubada com 100 µM de piperina.
Linhagem Lucena-1 (E) controle negativo (F) controle positivo (25 µM de etoposídio) (G)
incubada com 50 µM de piperina (H) ou incubada com 100 µM de piperina. Linhagem FEPS (I)
controle negativo (J) controle positivo (25 µM de etoposídio) (K) incubada com 50 µM de
piperina (L) ou incubada com 100 µM de piperina. As imagens são representativas de 3
experimentos independentes.

71
Após o tratamento das células K562 com piperina por 72 horas, não houve aumento
significativo das células em apoptose, apoptose tardia ou necrose, sugerindo que essas células
apresentam algum grau de resistência à piperina (Tabela 4).
Com relação ao efeito da piperina nas células Lucena-1, é possível observar dupla
marcação com Anexina-V e PI após o tratamento com 100 μM de piperina por 72 horas (Tabela
4). Esse resultado sugere que, nestas condições, possivelmente ocorreu morte celular via
apoptose.
Na linhagem FEPS, incubada por 72 horas com 100 μM de piperina, foi observado
aumento da marcação com Anexina-V e da dupla marcação com Anexina-V e PI, sugerindo que
esse composto também foi capaz de induzir apoptose nessa linhagem celular (Tabela 4). No
entanto, que esse perfil de marcação foi mais intenso do que o observado para o controle positivo
(Figura 14J).
K562
Controlenegativo
Controle positivo
Piperina(50 µM)
Piperina(100 µM)
1,2 ± 0,4 14,0 ± 5,5 2,2 ± 0,9 5,4 ± 3,0
3,8 ± 1,3 32,5 ± 2,4* 2,4 ± 1,3 8,2 ± 1,2
94,3 ± 1,8 37,7 ± 2,2* 94,6 ± 1,6 85,0 ± 2,0*
0,8 ± 0,1 15,8 ± 5,0* 0,8 ± 0,4 1,4 ± 0,6
Lucena-1
Controlenegativo
Controle positivo
Piperina(50 µM)
Piperina(100 µM)
2,3 ± 0,4 30,0 ± 7,5* 2,0 ± 0,9 2,2 ± 1,2
2,8 ± 0,5 15,0 ± 3,4* 4,8 ± 0,4 11,8 ± 1,1*
93,5 ± 0,1 50,7 ± 4,9* 91,6 ± 1,0 83,2 ± 1,9
1,4 ± 0,0 4,3 ± 2,3 1,6 ± 0,3 2,7 ± 0,6
FEPS
Controlenegativo
Controle positivo
Piperina(50 µM)
Piperina(100 µM)
0,8 ± 0,2 1,1 ± 0,3 0,9 ± 0,3 1,1 ± 0,6
2,9 ± 0,4 3,8 ± 0,8 5,6 ± 1,2 16,0 ± 2,8*
95,8 ± 0,3 94,0 ± 0,8 92,4 ± 1,0 79,9 ± 3,1*
0,6 ± 0,1 1,2 ± 0,3 1,0 ± 0,2 3,1 ± 0,8*
Tabela 4. Percentual das células K562, Lucena-1 e FEPS em necrose, apoptose tardia e apoptose.
Os resultados são expressos como média ± erro padrão (n=3). * p < 0,05 (ANOVA com pós-teste de Dunnett), comparado às células controle com DMSO.
Região
R1 (Necrose)
R2 (Apoptose tardia)
R3 (Células viáveis)
R4 (Apoptose)
Apesar de ambas as linhagens MDR terem sido alvos da piperina, quando são comparados
os percentuais de células em apoptose ou apoptose tardia, observa-se que esse fitoquímico
apresenta maior ação sobre a linhagem FEPS, em relação à Lucena-1. O fato de a FEPS ter mais
células em apoptose ou apoptose tardia é, provavelmente, um reflexo do que foi observado nos
outros resultados que sugerem que a FEPS começa a sofrer os efeitos da piperina antes das outras
linhagens. Esses dados reforçam a propriedade da piperina de ser seletivamente mais citotóxica
para a linhagem FEPS, a mais resistente.
5.5. Efeito da piperina sobre a ativação de caspase-3 nas células de leucemia
Posteriormente, foi investigado, por meio de Imunofluorescência, se a indução de
apoptose pela piperina estaria relacionada à clivagem de caspase-3 (Figuras 15, 16 e 17).

72
20 µm 20 µm 20 µm
20 µm 20 µm 20 µm
CTR
LP
IP (
10
0 µ
M)
K562
DAPI Capase-3 clivada MergeA
Figura 15. Efeito da piperina sobre a clivagem de caspase-3 na linhagem K562. As células
foram incubadas com 100 µM de piperina por 72 horas. Em seguida, foram marcadas com
anticorpo contra caspase-3 clivada e DAPI e analisadas em microscópio de fluorescência. As
imagens são representativas de 3 experimentos independentes. CTRL, controle. PIP, piperina.

73
20 µm 20 µm 20 µm
20 µm 20 µm 20 µm
CTR
LP
IP (
10
0 µ
M)
Lucena-1
DAPI Capase-3 clivada MergeB
Figura 16. Efeito da piperina sobre a clivagem de caspase-3 na linhagem Lucena-1. As
células foram incubadas com 100 µM de piperina por 72 horas. Em seguida, foram marcadas com
anticorpo contra caspase-3 clivada e DAPI e analisadas em microscópio de fluorescência. As
imagens são representativas de 3 experimentos independentes. CTRL, controle. PIP, piperina.

74
20 µm 20 µm 20 µm
20 µm 20 µm 20 µm
CTR
LP
IP (
10
0 µ
M)
FEPS
DAPI Capase-3 clivada MergeC
Figura 17. Efeito da piperina sobre a clivagem de caspase-3 na linhagem FEPS. As células
foram incubadas com 100 µM de piperina por 72 horas. Em seguida, foram marcadas com
anticorpo contra caspase-3 clivada e DAPI e analisadas em microscópio de fluorescência. As
imagens são representativas de 3 experimentos independentes. CTRL, controle. PIP, piperina.
Como esperado, a linhagem K562 tratada com 100 µM de piperina por 72 horas não
apresentou marcação para caspase-3 clivada, uma vez que esse composto não foi capaz de induzir
apoptose nessas células (Figura 15). Somente as linhagens Lucena-1 e, de forma mais acentuada,
FEPS, apresentaram células com núcleo apoptótico e ativação de caspase-3, indicando, mais uma
vez, a seletividade da piperina sobre as células MDR (Figuras 16 e 17).
5.6. Efeito da piperina sobre os níveis de GAPDH nas células de leucemia
A detecção de cerca de 20% de células em apoptose (por Anexina-V e PI, pela presença
de núcleos com corpos apoptóticos e pela clivagem de caspase-3) apenas justifica parcialmente a
diminuição da viabilidade celular nas células MDR tratadas com piperina (por MTT ou azul de

75
tripan). Por isso, sugere-se o envolvimento de outros mecanismos de morte celular,
concomitantemente. Um exemplo disso seria a indução da MCIC.
Já foi demonstrado que células MDR podem superexpressar GAPDH, se tornando mais
resistentes a MCIC (LAVALLARD e cols., 2009). Com isso, a fim de testar se a piperina poderia
modular esse tipo de morte celular, seletivamente nas células MDR, foi investigada a expressão
gênica de GAPDH nas células de leucemia após 72 horas de tratamento com 100 µM de piperina
(Figura 18).
K562 Lucena-1 FEPS0
50
100
150
200
250
300 CTRL
PIP (100 M)
*Exp
ressão
de G
AP
DH
(un
idad
es a
rbit
rári
as)
Figura 18. Efeito da piperina sobre a expressão gênica de GAPDH nas células de leucemia
K562, Lucena-1 e FEPS. As células foram incubadas com 100 µM de piperina por 72 horas. A
expressão gênica de GAPDH foi avaliada por RT-PCR em tempo real utilizando SYBR Green. Os
resultados são expressos como média ± erro padrão (n=3). * p < 0,05 (Teste t de Student,
pareado) comparado às células controles. CTRL, controle. PIP, piperina.
No presente trabalho também foi possível observar uma relação entre o grau de resistência
das células e o aumento da expressão de GAPDH, sendo que a linhagem FEPS apresentou maior
expressão desse gene. No entanto, a piperina não foi capaz de modular a expressão de GAPDH
nas linhagens MDR, mas reduziu significativamente a expressão desse gene na K562 (Figura
18). Por isso, o mecanismo de CS induzido pela piperina não parece envolver a MCIC mediada
por GAPDH.
Posteriormente, esses resultados foram confirmados por meio da avaliação da expressão
proteica por Western blotting (Figura 19).

76
GAPDH(37 kDa)
Hsp70 (70 kDa)
CTRL PIP CTRL PIP CTRL PIP
K562 Lucena-1 FEPS
GAPDH/ Hsp70 0,57 0,18 0,70 0,73 1,37 1,50
Figura 19. Efeito da piperina sobre a expressão de GAPDH nas células de leucemia K562,
Lucena-1 e FEPS. As células foram incubadas com 100 µM de piperina por 72 horas. Os
extratos protéicos das células tratadas (100 μg) foram submetidos à eletroforese em gel de
poliacrilamida à 10% na presença de SDS. Em seguida, Western blotting foi realizado utilizando
anticorpos contra GAPDH e Hsp70 (controle de carregamento). Os imunoblottings são
representativos de 2 diferentes experimentos. A análise densitométrica de cada banda foi
realizada utilizando o software Image J e os resultados expressos como unidade arbitrária (UA)
em relação ao controle de carregamento Hsp70. CTRL, controle. PIP, piperina.
Os efeitos da piperina sobre a modulação proteica e gênica do GAPDH foram
semelhantes, ou seja, afetou mais a K562 do que as linhagens com fenótipo MDR, sugerindo,
mais uma vez, que esses dados não parecem ter relação com o fenômeno de CS.
5.7. Efeito da piperina sobre os níveis de PARP-1 e PAR nas células de leucemia
Dados do nosso grupo de pesquisa demonstraram que apenas a linhagem MDR FEPS
apresenta teores detectáveis de PARP clivada no fragmento de 24 kDa, capaz de inibir a PARP-1
(116 kDa). Essa peculiaridade da FEPS poderia contribuir para sua própria sobrevivência, uma
vez que a inibição da PARP-1 poderia salvar as células da depleção de energia (VIDAL e cols.,
2018). Esses dados indicam que a PARP-1 poderia ser um alvo importante contra a MDR. Por
isso, foi investigado, nas linhagens de leucemia, se a piperina poderia modular a PARP-1, pelo
método de Western blotting (Figura 20A).

77
PAR(40 kDa)
PARP-1 (116 kDa)
PARP-1 clivada (24 kDa)
Hsp70 (70 kDa)
CTRL PIP CTRL PIP CTRL PIP
K562 Lucena-1 FEPS
CTRL PIP CTRL PIP CTRL PIP
K562 Lucena-1 FEPSA B
Hsp70 (70 kDa)
PARP-1/ Hsp70
PARP-1/ Hsp70 0,93 0,70 0,70 0,81 1,02 0,82
0,88 0,84 0,96 0,87 0,54 0,47
PAR(50 kDa)
PAR/ Hsp70
PAR/ Hsp70
PARP-1/ Hsp70
PARP-1/ Hsp70
PARP-1 clivada (89 kDa)
PARP-1 clivada (40 kDa)
1,07 0,70 1,01 0,88 0,69 0,37
0,95 0,00 0,31 0,41 0,03 0,01
0,92 0,47 0,35 0,18 0,24 0,18
0,81 0,26
Figura 20. Efeito da piperina sobre a expressão de PARP-1 e PAR nas células de leucemia
K562, Lucena-1 e FEPS. As células foram incubadas com 100 µM de piperina por 72 horas. Os
extratos proteicos das células tratadas (100 μg) foram submetidos à eletroforese em gel de
poliacrilamida à 10% na presença de SDS. Em seguida, Western blotting foi realizado utilizando
anticorpos contra PARP-1 (A), PAR (B) e Hsp70 (controle de carregamento) (A) (B). A análise
densitométrica de cada banda foi realizada utilizando o software Image J e os resultados
expressos como unidade arbitrária (UA) em relação ao controle de carregamento Hsp70. CTRL,
controle. PIP, piperina.
Foram observados teores semelhantes de PARP-1 (116 kDa) entre K562 (1,07 UA) e
Lucena-1 (1,01 UA), mas FEPS apresentou a menor expressão dessa enzima (0,69 UA). Além
disso, apenas as linhagens K562 e Lucena-1 apresentaram níveis detectáveis de PARP-1 clivada
no fragmento de 89 kDa (0,95 e 0,31 UA, respectivamente), sendo a marcação na K562 a mais
intensa. Foi identificada a presença de um fragmento de PARP-1 pouco conhecido (40 kDa) de
maneira também mais marcante na linhagem K562 (0,92 UI na K562 X 0,35 UI na Lucena-1 X
0,24 UI na FEPS). Por último, apenas a linhagem FEPS apresentou expressão do fragmento de 24
kDa (0,81 UI) (Figura 20A).
Com relação ao efeito da piperina, foi observada redução mais relevante dos teores de
PARP-1 (116 kDa) na linhagem FEPS. Além disso, esse composto ainda reduziu a quantidade de
PARP-1 no fragmento de 24 kDa nessa mesma linhagem celular. Já na linhagem K562, a piperina
teve efeito na redução do conteúdo de PARP-1 (89 kDa) e PARP-1 (40 kDa) (Figura 20A).
Sendo assim, as alterações provocadas pela piperina foram mais perceptíveis na linhagem

78
sensível a drogas (K562) e na linhagem mais resistente desse modelo experimental (FEPS),
apesar de ter apresentado efeitos e/ ou intensidades distintas entre si (Figura 20A).
Em seguida, foi realizado um experimento, por Western blotting, a fim de verificar se
além da modulação da PARP-1, piperina poderia estar alterando a atividade dessa enzima. Para
isso, quantificamos a expressão do polímero PAR, importante marcador da super-ativação da
PARP-1 (Figura 20B).
Não foram observadas alterações importantes nos teores de PAR (50 kDa) entre as
linhagens ou após o tratamento com 100 µM de piperina. No entanto, a FEPS apresentou menor
conteúdo de PAR (40 kDa) em comparação com as demais linhagens, sendo que a piperina
provocou pouca ou nenhuma alteração na expressão desse polímero nas linhagens celulares
estudadas (Figura 20B).
5.8. Efeito da piperina sobre a expressão de P-gp nas linhagens MDR, Lucena-1 e FEPS
Foi demonstrado por Wurzer e cols. (2000) e Wesierska-Gadek (2005) uma possível
correlação entre a PARP-1 e o fenótipo MDR, uma vez que células deficientes em PARP-1
apresentaram maior expressão de P-gp e foram mais resistentes à ação de quimioterápicos do que
células controle. Por isso, investigamos se a piperina também seria capaz de modular a expressão
de P-gp. Para isso, foram realizados experimentos em citômetro de fluxo com as linhagens
Lucena-1 e FEPS após o tratamento por 72 horas com 50 e 100 µM de piperina (Figura 21).

79
Lucena-1
CTRL PIP (50 M) PIP (100 M)0
5
10
15
20
*
Exp
ressão
de
P-g
p
(MIF
)
FEPS
CTRL PIP (50 M) PIP (100 M)0
10
20
30
40
50
*
Exp
ressão
de
P-g
p
(MIF
)
A
B
Figura 21. Efeito da piperina sobre a expressão de P-gp nas linhagens de leucemia. As
células foram incubadas com piperina por 72 horas e, em seguida, com o anticorpo, anti-P-
gp/FITC, por 30 minutos. Após a lavagem, a intensidade de fluorescência foi medida por
citometria de fluxo. Linhagens Lucena-1 (A) e FEPS (B) incubadas com 50 e 100 µM de
piperina. Os resultados são expressos como média e erro padrão (n=3). * p < 0,05 (ANOVA com
pós-teste de Dunnett), comparado às células controle com DMSO. MIF, média de intensidade de
fluorescência. CTRL, controle. PIP, piperina.
Como era esperado, a linhagem FEPS apresentou a maior expressão dessa bomba de
efluxo (cerca de 3,9 vezes em relação a Lucena-1). Além disso, nossos experimentos indicaram
que 100 µM de piperina provocou aumento significativo da expressão da P-gp em ambas as
linhagens MDR (cerca de 2 vezes em relação ao controle da Lucena-1 e 1,4 em relação ao
controle da FEPS).
Como não foram observadas alterações importantes nos teores de PARP-1 na linhagem
Lucena-1 após o tratamento com piperina, o aumento da expressão de P-gp nessa linhagem não
parece ter relação com a modulação dessa enzima. No entanto, os resultados da FEPS indicam

80
uma possível relação entre a diminuição da expressão de PARP-1 e o aumento da expressão de P-
gp após incubação com piperina.
5.9. Efeito da piperina sobre enzimas metabolizadoras de xenobióticos nas células de
leucemia
Um dos mecanismos relacionados ao fenótipo MDR é o aumento da capacidade da célula
de câncer de metabolizar compostos, por meio da indução de enzimas fundamentais para o
processo de biotransformação, gerando moléculas mais facilmente excretadas (LOLODI e cols.,
2017). Em outros casos, porém, esse processo de metabolização também pode gerar metabólitos
com atividade biológica ainda mais potente do que sua molécula de origem (ZHANG e cols.,
2017). A fim de testar se o efeito citotóxico seletivo da piperina sobre as células MDR estaria
relacionado a uma dessas hipóteses, foram realizados experimentos para avaliar expressão gênica
dos seguintes marcadores do metabolismo de fase I: enzimas do complexo citocromo P450
(CYP3A4, CYP3A5, CYP1B1) e PXR.
Experimentos por RT-PCR em tempo real demonstraram que as linhagens de leucemia
não apresentaram expressão detectável dos genes CYP3A4, CYP1B1 e PXR. Com relação ao gene
CYP3A5, a linhagem FEPS apresentou a maior expressão, dentre as demais linhagens estudadas
(cerca de 240 vezes, em relação a K562, e cerca de 5 vezes, em relação a Lucena-1) (Figura 22).
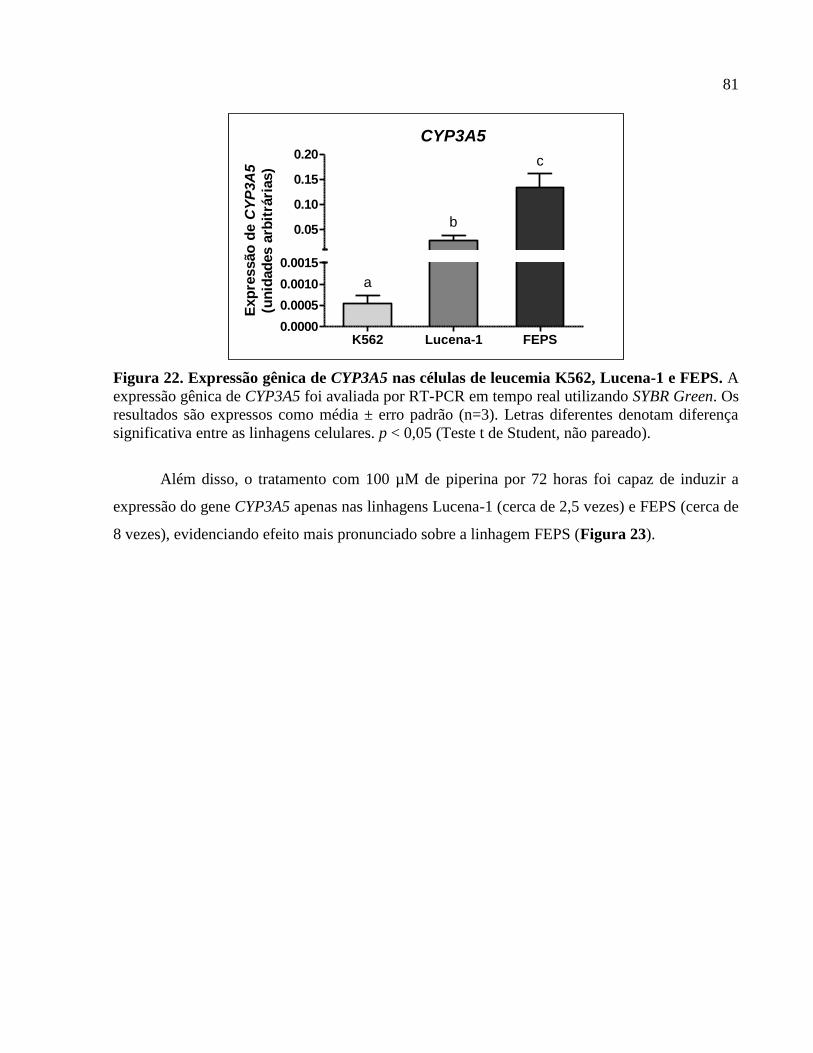
81
K562 Lucena-1 FEPS0.0000
0.0005
0.0010
0.0015
0.05
0.10
0.15
0.20
a
b
c
CYP3A5
Exp
ressão
de C
YP
3A
5
(un
idad
es a
rbit
rári
as)
Figura 22. Expressão gênica de CYP3A5 nas células de leucemia K562, Lucena-1 e FEPS. A
expressão gênica de CYP3A5 foi avaliada por RT-PCR em tempo real utilizando SYBR Green. Os
resultados são expressos como média ± erro padrão (n=3). Letras diferentes denotam diferença
significativa entre as linhagens celulares. p < 0,05 (Teste t de Student, não pareado).
Além disso, o tratamento com 100 µM de piperina por 72 horas foi capaz de induzir a
expressão do gene CYP3A5 apenas nas linhagens Lucena-1 (cerca de 2,5 vezes) e FEPS (cerca de
8 vezes), evidenciando efeito mais pronunciado sobre a linhagem FEPS (Figura 23).

82
K562
CTRL PIP (100 M)0.0000
0.0002
0.0004
0.0006
0.0008
0.0010
Exp
ressão
de C
YP
3A
5
(un
idad
es a
rbit
rári
as)
Lucena-1
CTRL PIP (100 M)0.00
0.02
0.04
0.06
0.08
0.10
*E
xp
ressão
de C
YP
3A
5
(un
idad
es a
rbit
rári
as)
FEPS
CTRL PIP (100 M)0.0
0.5
1.0
1.5
2.0
*
Exp
ressão
de C
YP
3A
5
(un
idad
es a
rbit
rári
as)
A
B
B
Figura 23. Efeito da piperina sobre a expressão gênica de CYP3A5 nas células de leucemia
K562, Lucena-1 e FEPS. As células foram incubadas com 100 µM de piperina por 72 horas. Em
seguida, a expressão gênica de CYP3A5 foi avaliada por RT-PCR em tempo real utilizando SYBR
Green. Os resultados são expressos como média ± erro padrão (n=3). * p < 0,05 (Teste t de
Student, pareado) comparado às células controles. CTRL, controle. PIP, piperina.
Sendo assim, dentre os genes avaliados, CYP3A5 foi o único identificado nas células de
leucemia. Além disso, CYP3A5 se apresentou mais expresso nas linhagens com fenótipo MDR,
comparado à K562. A linhagem mais resistente, FEPS, obteve os maiores níveis, seguida da
linhagem Lucena-1 e, por último, a célula sensível a drogas, K562, e a piperina ainda induziu a
expressão desse gene, em maior grau na FEPS, sugerindo possível participação na promoção de
CS.

83
Por isso, além da análise da expressão gênica, foi investigada a interação da piperina com
a proteína CYP3A5 por docking molecular (in silico) (Figura 24).
A B
Figura 24. Interação da piperina com CYP3A5. Foram realizados experimentos para avaliar a
interação da piperina com CYP3A5 por docking molecular. (A) Ligante Protoporphyrin IX
containing Fe (HEM): vermelho; Ligante Hitonavir (HIT): laranja; Piperina: verde. Proteína
CYP3A5: cinza (B) Aproximação do local de interação Piperina-CYP3A5- e comparação entre a
posição final da Piperina após o docking molecular com os dois ligantes cristalográficos HEM e
HIT.
Nos experimentos de docking proteína-ligante foi estimada a energia livre de ligação, para
a interação piperina-CYP3A5, em -8,6 Kcal/ mol. Além disso, foi observada sobreposição da
posição ocupada pela piperina (verde) e pelos ligantes cristalográficos depositados no PDB
(HEM e HIT), indicando que a piperina ocupa sítio semelhante a esses ligantes cristalográficos
naturais, o que, provavelmente, estaria relacionado ao seu efeito biológico. A sobreposição da
piperina nos sítios dos ligantes cristalográficos pode ser reforçada pelo fato de que a caixa de
interação para simulação do docking envolveu toda a cadeia A da CYP3A5, aumentando a
probabilidade da piperina ancorar em qualquer região dessa proteína. No entanto, mesmo diante
das grandes dimensões da caixa de interação, a molécula localizou-se, preferencialmente, em um
sítio de possível alta afinidade. Esses resultados sugerem que a piperina foi capaz de interagir
com a CYP3A5, provavelmente em um sítio de importância biológica ou de grande afinidade.

84
5.10. Análise da metabolização de piperina nas células de leucemia
Dados sobre o metabolismo da piperina são escassos e ainda não está claro se esse
composto poderia ser metabolizado, gerando compostos com ação citotóxica ainda mais
pronunciada, ou se a sua própria molécula original já apresentaria efeitos potenciais contra o
câncer. Buscando compreender o efeito seletivamente citotóxico da piperina sobre células MDR,
foram quantificados, por CLAE-DAD, a piperina e seus possíveis metabólitos no interior das
células (piperina intracelular) de leucemia e no sobrenadante (piperina extracelular) das culturas
celulares.
Primeiramente, foi testado se a piperina se manteria estável no meio de cultura, sem sofrer
degradação, nas mesmas condições de cultivo das linhagens celulares (37 °C em uma atmosfera
umidificada contendo 5% de CO2) (Figura 25).
Meio de cultura + Piperina
0 8 12 24 48 72 0
20
40
60
80
100
120
Tempo (horas)
Pip
eri
na (
M)
Figura 25. Curva de degradação de piperina no meio de cultura celular. O meio de cultura
(RPMI + 10% SFB) foi incubado com 100 µM de piperina pelos períodos de tempo indicados e
mantido a 37 °C em uma atmosfera umidificada contendo 5% de CO2. Em seguida, foi realizada
análise em CLAE-DAD utilizando-se coluna de fase reversa C18 e fase móvel constituída de
acetonitrila com KH2PO4 (45:55 v/v). Os resultados são expressos como média (n=2).
Os dados indicaram que apenas o tempo de incubação de 72 horas provocou degradação
da piperina de maneira relevante (cerca de 50% em relação ao tempo 0). Assim, a análise dos
metabólitos de piperina produzidos pelas células de leucemia poderia ser realizada com até 48
horas de incubação com esse composto, de forma a impedir que a degradação de piperina no
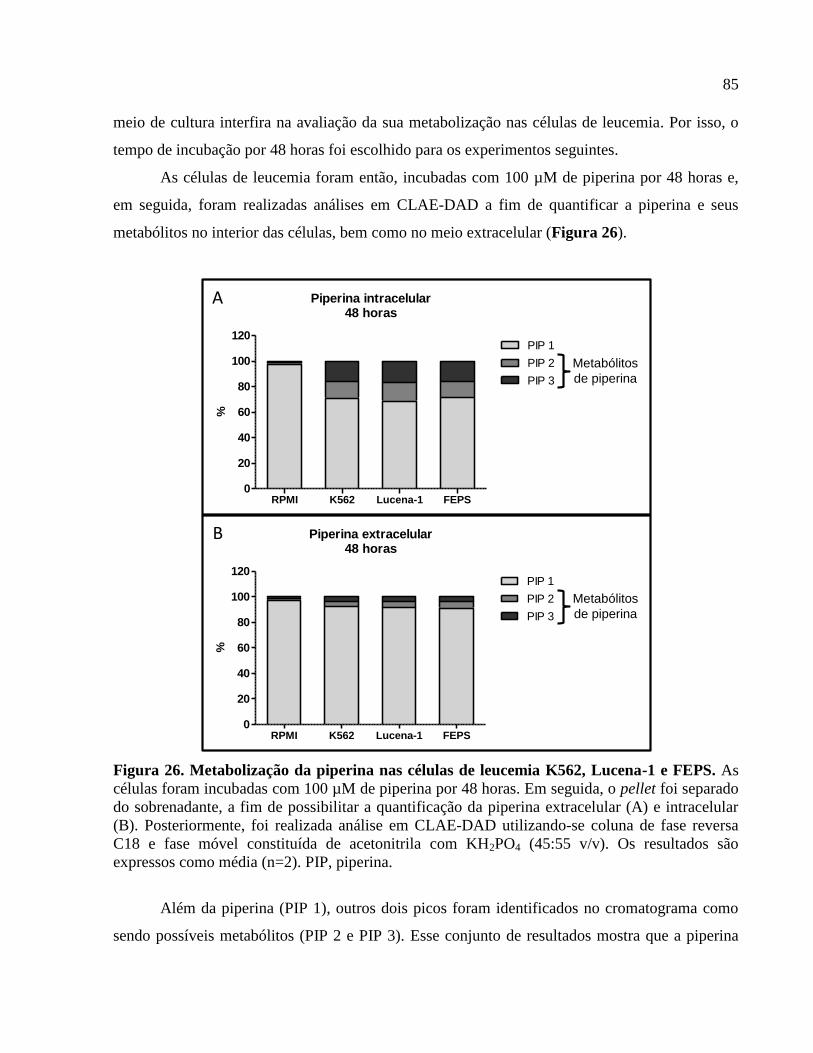
85
meio de cultura interfira na avaliação da sua metabolização nas células de leucemia. Por isso, o
tempo de incubação por 48 horas foi escolhido para os experimentos seguintes.
As células de leucemia foram então, incubadas com 100 µM de piperina por 48 horas e,
em seguida, foram realizadas análises em CLAE-DAD a fim de quantificar a piperina e seus
metabólitos no interior das células, bem como no meio extracelular (Figura 26).
Piperina extracelular48 horas
RPMI K562 Lucena-1 FEPS0
20
40
60
80
100
120PIP 1
PIP 2
PIP 3
%
Piperina intracelular48 horas
RPMI K562 Lucena-1 FEPS0
20
40
60
80
100
120PIP 1
PIP 2
PIP 3
%
Metabólitos
de piperina
Metabólitos
de piperina
B
A
Figura 26. Metabolização da piperina nas células de leucemia K562, Lucena-1 e FEPS. As
células foram incubadas com 100 µM de piperina por 48 horas. Em seguida, o pellet foi separado
do sobrenadante, a fim de possibilitar a quantificação da piperina extracelular (A) e intracelular
(B). Posteriormente, foi realizada análise em CLAE-DAD utilizando-se coluna de fase reversa
C18 e fase móvel constituída de acetonitrila com KH2PO4 (45:55 v/v). Os resultados são
expressos como média (n=2). PIP, piperina.
Além da piperina (PIP 1), outros dois picos foram identificados no cromatograma como
sendo possíveis metabólitos (PIP 2 e PIP 3). Esse conjunto de resultados mostra que a piperina

86
foi internalizada pelas linhagens de leucemia e, mesmo nas células MDR, que poderiam extruir
esse composto, foi possível sua identificação no meio intracelular (Figura 26A). Além disso, a
piperina demonstrou sofrer metabolização de maneira semelhante entre as linhagens celulares, de
forma mais intensa no meio intracelular (cerca de 30% de metabólitos) (Figura 26A) do que no
meio extracelular (cerca de 10% de metabólitos) (Figura 26B). Esses dados sugerem que as
células de leucemia metabolizaram piperina, mas essa metabolização parece não estar relacionada
com a indução de CS, uma vez que não foram observadas diferenças na proporção de metabólitos
de piperina entre as células leucêmicas.

87
6. DISCUSSÃO
Nesse trabalho foi investigado o efeito da piperina sobre células de leucemia mieloide
crônica sensíveis e resistentes a drogas. Os nossos resultados indicam que esse fitoquímico
apresentou efeito citotóxico mais pronunciado sobre as células MDR, Lucena-1 e FEPS. Até onde
se sabe, essa propriedade da piperina em induzir CS ainda não tinha sido descrita na literatura
científica para nenhum outro modelo experimental de câncer, o que demonstra o caráter inédito
deste estudo.
Diversos estudos têm descrito que, quando uma molécula é capaz de induzir CS em
determinada linhagem MDR, quanto mais P-gp essas células expressarem, mais citotóxico será o
composto (HALL e cols., 2009; KELLY e cols., 2010; NOBILI e cols., 2011). A citotoxicidade
da piperina foi mais acentuada na linhagem celular Lucena-1 em relação a K562, sendo que a
Lucena-1 apresenta cinco vezes mais cópias do gene MDR1 (que codifica a P-gp) do que sua
linhagem parental, K562 (RUMJANEK e cols., 2001). Além disso, a piperina foi ainda mais
citotóxica na linhagem FEPS, cuja expressão de P-gp é duas vezes maior que a apresentada pela
linhagem Lucena-1 (RUMJANEK e cols., 2001).
Estes resultados demonstram possível correlação entre o aumento da expressão de P-gp e
a indução do efeito citotóxico da piperina, sugerindo que o mecanismo da piperina em induzir CS
seria dependente da presença de P-gp na membrana dessas células tumorais. Dados experimentais
prévios obtidos pela autora durante o mestrado corroboram essa hipótese sobre o envolvimento
da P-gp na indução de CS, uma vez que as células Lucena-1 e FEPS silenciadas para o gene
MDR1 foram menos sensíveis a ação da piperina do que as células controle (QUARTI, 2015).
Apesar do efeito mais pronunciado sobre células resistentes, a piperina não apresentou
efeito citotóxico sobre macrófagos peritoneais de camundongos, um tipo de célula mieloide
normal, já diferenciada, e que pouco prolifera. De acordo com outros estudos, Lucena-1 e FEPS
também parecem ser mais diferenciadas (CARRETT-DIAS e cols., 2016) e possuir ciclo celular
mais lento (DAFLON-YUNES e cols., 2013) em relação à K562. Por isso, ainda que as células
normais e MDR se assemelhem quanto à velocidade de seu ciclo celular, a piperina induziu
efeitos aparentemente contrários nesses dois modelos, sugerindo que a diferenciação celular e a
baixa proliferação não estejam relacionadas à maior sensibilidade das células MDR à ação da
piperina.

88
Esses dados estão de acordo com outros trabalhos que destacaram o efeito tóxico reduzido
da piperina sobre células humanas normais como osteoblastos, fibroblastos e células mamárias
quando comparado com células de câncer, sugerindo que esse composto poderia constituir
promissor agente terapêutico contra o câncer (ZHANG e cols., 2015; YAFFE e cols., 2015;
GREENSHIELDS e cols., 2015). Chuchawankul e cols. (2012) também avaliaram o efeito da
piperina sobre células do sistema imune humano (Peripheral blood mononuclear cells - PBMC),
principal modelo experimental utilizado para investigar a seletividade de compostos sobre células
de leucemia. Foi constatado que a piperina foi pouco tóxica para essas células, inclusive
apresentando IC50 de aproximadamente 350 µM (8,5 vezes maior do que o encontrado para a
linhagem FEPS).
A fim de comprovar os efeitos relacionados à indução de CS, foram realizados
experimentos para avaliar os efeitos da piperina sobre a morfologia celular, ciclo celular e morte
celular.
Ao observar a morfologia celular após o tratamento com a piperina, foram verificadas
diferenças entre as três linhagens de leucemia, mas sem evidências de que as células K562
estivessem se diferenciando em macrófagos, diferentemente do que foi encontrado por Song e
cols. (2008). Esses autores demonstraram aumento da diferenciação da K562 em macrófagos/
monócitos após o tratamento com 20 ou 40 μM de piperina e, com isso, inibição da sua
proliferação. No entanto, os aspectos morfológicos delineados no presente estudo reforçam os
dados de MTT que indicam que a linhagem K562 apresenta certa resistência ao efeito citotóxico
da piperina. Além disso, também não foi possível identificar perfil mais diferenciado nas células
MDR, como foi visto anteriormente por Carrett-Dias e cols. (2016).
A avaliação de alterações morfológicas também revelou o aumento da formação e do
volume de vacúolos no citoplasma das células, especialmente na FEPS tratada com piperina.
Posteriormente esses dados ainda foram confirmados utilizando-se o parâmetro de granulosidade
(SSC) no citômetro de fluxo.
Os vacúolos citoplasmáticos observados nas células de leucemia, especialmente na FEPS
tratada com piperina, poderiam tratar-se de corpúsculos lipídicos: organelas formadas por um
centro rico em lipídio neutro (principalmente triacilglicerol e ésteres de esterol) e colesterol,
cercado por uma membrana monocamada de fosfolipídios, com composição peculiar e exibindo
diversas proteínas associadas (ZWEYTICK e cols., 2000; MURPHY, 2001; TAUCHI-SATO e

89
cols., 2002; MARTIN e PARTON, 2005). Essas organelas são dinâmicas e estão envolvidas no
metabolismo de lipídios e sinalização celular (BOZZA e VIOLA, 2010) e podem aumentar em
número e tamanho em diferentes tipos de células cancerígenas (HAGER e cols., 2006; STRAUB
e cols., 2010; SUNDELIN e cols., 2012; GUILLAUMOND e cols., 2015), dentre elas, em
leucemias (VERBRUGGE e cols., 2016) e em foam cells (macrófagos ricos em lipídeos)
(BECHOR e cols., 2017).
Já foi demonstrado que a formação de corpúsculos lipídios seria importante para a
sobrevivência das células expostas a um agente estressor, como um quimioterápico (KOIZUME e
MIYAGI, 2016; WELTE e GOULD, 2017). Cotte e cols. (2018) sugeriram, inclusive, possível
correlação entre a elevada formação de corpúsculos lipídicos e o aumento da resistência a
quimioterápicos.
Entretanto, ao mesmo tempo em que essas organelas teriam a função de proteger as
células de câncer submetidas ao estresse, um excessivo turnover dos corpúsculos lipídicos e um
desbalanço entre a biogênese e o colapso dessas estruturas poderia liberar centenas de espécies
lipídicas com diferentes características, causando danos celulares (PETAN e cols., 2018). Sendo
assim, a piperina pode ter gerado desequilíbrio entre a produção e destruição dessas organelas,
aumentando o número e o volume de corpúsculos lipídicos na linhagem mais sensível a sua ação,
FEPS.
Considerou-se também que o efeito mais pronunciado da piperina sobre as linhagens
Lucena-1 e FEPS pudesse refletir uma parada ou alongamento do ciclo celular. No entanto, a
piperina não foi capaz de bloquear o ciclo celular em nenhuma das linhagens leucêmicas
estudadas. Ademais, a piperina induziu, preferencialmente, as células de leucemia à morte
celular, ao invés de ter provocado a inibição da proliferação celular, sendo a linhagem FEPS a
mais afetada, corroborando com os experimentos descritos anteriormente e com a ocorrência do
fenômeno de CS.
Não foi possível encontrar relatos na literatura científica sobre a indução de apoptose pela
piperina em células MDR. No entanto, a piperina promoveu apoptose em inúmeros modelos
experimentais de câncer, incluindo de cólon (YAFFE e cols., 2015), melanoma (FOFARIA e
cols., 2014), pulmão (LIN e cols., 2014), mama (GREENSHIELDS e cols., 2015), carcinoma oral
de células escamosas (SIDDIQUI e cols., 2017), carcinoma hepatocelular (GUNASEKARAN e
cols., 2017), próstata (BA e MALHOTRA, 2018) e células de câncer de ovário (SI e cols., 2018).

90
Esses dados fortalecem os resultados apresentados neste trabalho e sugerem que a piperina é um
composto com propriedades anticâncer importantes.
Os experimentos para identificação das proteínas relacionados à morte celular indicaram
que a piperina induziu apoptose por um mecanismo dependente de caspase-3 nas linhagens MDR
Lucena-1 e FEPS, uma vez que houve ativação dessa proteína. Outros autores também
constataram esse mesmo efeito da piperina em células de melanoma (FOFARIA e cols., 2014),
carcinoma oral de células escamosas (SIDDIQUI e cols., 2017) e em células de câncer de ovário
(SI e cols., 2018).
A apoptose é o tipo de morte celular mais estudada, no entanto, existem também outras
formas de morte que ocorrem de maneira independente da ativação de caspases (LAVALLARD e
cols., 2009). Morfologicamente, em alguns aspectos, podem se assemelhar ao que foi observado
no presente estudo, isto é, membrana plasmática irregular, encolhimento de núcleo e reduzida
fragmentação nuclear. MCIC não envolve necessariamente ativação de caspases, mas a
superexpressão de GAPDH em células de câncer parece conferir resistência a esse tipo de morte
devido a maior eficiência do metabolismo glicolítico (LAVALLARD e cols., 2009).
Neste trabalho foi demonstrado que as células MDR expressam mais GAPDH do que a
K562, de forma mais evidente na FEPS do que na Lucena-1 (VIDAL e cols., 2018).
Originalmente considerou-se que a piperina seria capaz de diminuir a expressão de GAPDH nas
células resistentes a quimioterápicos, tornando-as mais sensíveis a MCIC. No entanto, a piperina
induziu efeito contrário ao esperado, uma vez que modulou tanto a expressão gênica como a
proteica de GAPDH na K562, enquanto que nas linhagens Lucena-1 e FEPS, células mais
sensíveis à morte celular, não foi observado efeito. Por isso, esta relação precisará ser explorada
futuramente.
A detecção de células em apoptose e marcadas para caspase-3 clivada apenas justifica
parcialmente a diminuição da viabilidade celular nas células MDR tratadas com piperina. No
entanto, a piperina não parece induzir MCIC. Por isso, sugere-se o envolvimento de outro tipo de
morte celular. Um exemplo disso seria a modulação da autofagia.
A autofagia pode ser definida como um processo de conservação celular em que porções
citoplasmáticas são englobadas por uma membrana com dupla camada lipídica, resultando em um
compartimento delimitado, denominado autofagossomo. Posteriormente, a porção externa do
autofagossomo se funde com endossomos e lisossomos originando o autofagolisossomo onde o

91
material interno e a membrana serão completamente digeridos por hidrolases lisossomais (LEVY
e cols., 2017).
Tanto a indução da autofagia como sua inibição poderiam ser estratégias contra o câncer,
uma vez que esse processo pode contribuir para a proteção das células de câncer, promovendo a
resistência a quimioterápicos, e também pode levar a morte de células MDR nos casos de
ativação excessiva da via autofágica (LI e cols., 2017). Por isso, a piperina poderia estar
modulando a autofagia, afetando de forma mais pronunciada a FEPS.
Outra diferença marcante entre a célula MDR, FEPS, e a célula sensível a
quimioterápicos, K562, é em relação a expressão da PARP-1. Além da imprescindível atuação
dessa enzima sobre o reparo do DNA celular, PARP-1 também apresenta outras inúmeras
funções na célula de câncer, como por exemplo, envolvimento na morte celular. Por isso, foram
avaliados, por Western blotting, os teores de PARP-1 e PAR, importante marcador da super-
ativação dessa enzima, nas células de leucemia na presença ou não de piperina.
Nossos experimentos indicaram que a FEPS apresentou a menor expressão de PARP-1
(116 kDa) e, ainda, os teores mais baixos do polímero PAR. Esses dados sugerem que essa
linhagem poderia ter desenvolvido mecanismo capaz de atenuar a super-ativação de PARP-1,
diferentemente da K562 e Lucena-1.
Apenas a linhagem MDR FEPS apresentou quantidades detectáveis de PARP clivada no
fragmento de 24 kDa. Já foi demonstrado que PARP-1 clivada no fragmento de 24 kDa tem papel
importante no processo de resistência das células de câncer a morte celular por necrose. Este
fragmento se liga ao DNA, competindo com a enzima de reparo PARP-1 e, consequentemente,
atenua a sua superativação e reduz os teores de PAR (YUNG e cols., 2001). Portanto, a PARP-1
clivada no fragmento de 24 kDa seria capaz de atenuar a superativação de PARP-1,
possivelmente bloqueando a morte celular por necrose (CHAITANYA e cols., 2010).
As células com DNA severamente danificado amplificam a atividade da PARP-1,
resultando em alto consumo de NAD+, levando a depleção de ATP e induzindo a morte celular
por necrose (ou partanatos). Por isso, a inativação da PARP-1 salvaria as células da depleção de
energia (NAD+ e ATP) (HERCEG e WANG, 1999; D'AMOURS e cols., 2001). A atenuação da
atividade da PARP-1 na FEPS, observada no presente estudo, seria um importante mecanismo de
proteção contra necrose, contribuindo para o fenótipo de resistência dessa linhagem, uma vez que

92
possui elevada demanda energética devido à expressão e atividade das bombas de efluxo (P-gp e
MRP-1) (VIDAL e cols., 2018).
Nossos experimentos também identificaram um fragmento de PARP-1 de 40 kDa nas
linhagens de leucemia. No entanto, esse fragmento pareceu estar mais acentuado na K562, sendo
que a piperina reduziu sua expressão nas linhagens K562 e Lucena-1. Estudo realizado com
células de neuroblastoma, após tratamento com uma toxina denominada maitotoxina, demonstrou
que a PARP-1 seria substrato da calpaína, uma protease dependente de cálcio, gerando o
fragmento de 40 kDa (MCGINNIS e cols., 1999). No entanto, ainda não se sabe ao certo o papel
desse fragmento sobre vias específicas de morte celular, apesar de existirem indícios de que
estaria relacionado com a morte celular por necrose, pelo menos em doenças neurológicas
(CHAITANYA e cols., 2010).
Além disso, foi demonstrado que a piperina reduziu o fragmento de 89 kDa da PARP-1 na
K562, possivelmente diminuindo sua suscetibilidade à apoptose. No entanto, esse composto
reduziu a expressão de PARP-1 em maior grau na FEPS, sugerindo relação com a indução de
apoptose. Caspase-3 é considerada importante para a clivagem de PARP-1 nas células de câncer,
originando os fragmentos de 24 kDa e de 89 kDa (SOLDANI e SCOVASSI, 2002;
CHAITANYA e cols., 2010; BOOZARI e cols., 2015). No entanto, apesar de a piperina ter
reduzido os teores de PARP-1 na FEPS, não foi observado aumento concomitante dos fragmentos
de 89 kDa (sem alterações) e 24 kDa (houve diminuição, inclusive). Como já foi mencionado
anteriormente, o fragmento de 24 kDa parece ser um fator protetor para a FEPS e, de alguma
maneira, piperina prejudica esse mecanismo de sobrevivência, considerando que essa é a célula
mais sensível a sua ação. Além da caspase-3 e calpaína, citadas anteriormente, existem outras
proteases como granzima e metaloproteinases de matriz que também podem ser responsáveis pela
formação de fragmentos de PARP-1 (CHAITANYA e cols., 2010). Por isso, a piperina poderia
estar modulando algumas dessas proteases, aumentando a clivagem de PARP-1 em fragmentos
que não estariam sendo reconhecidos pelo anticorpo utilizado no presente trabalho.
O tipo de morte celular programada que é mediada pela super-ativação da PARP-1 é
denominado “Partanatos”. Partanatos está associado à síntese e ao acúmulo do polímero PAR que
atua como um "sinal de morte" capaz de provocar a liberação de AIF da mitocôndria e este, por
sua vez, sofre translocação para o núcleo, causando condensação nuclear, fragmentação do DNA
e morte celular (YU e cols., 2002; ANDRABI e cols., 2006; DAVID e cols., 2009). No entanto, a

93
piperina não foi capaz de induzir esse tipo de morte celular no nosso modelo experimental, uma
vez que não aumentou os teores de PARP-1 (116 kDa) ou PAR.
Além disso, a piperina também não parece atuar de maneira semelhante a um “inibidor de
PARP-1”. Um dos mecanismos de resistência das células de câncer a ação de quimioterápicos é a
sua capacidade de promover aumento do reparo do DNA, mediado principalmente pela PARP-1.
Por isso, destaca-se a importância dos inibidores de PARP-1 a fim de impedir o reparo do DNA
dessas células, o que poderia levá-las a morte celular por apoptose. Apesar de a piperina ter
provocado diminuição da expressão da PARP-1 na FEPS, não foi capaz de alterar a atividade
dessa enzima, como foi visto pelos teores de PAR que não sofreram alteração.
Uma hipótese que também se buscou investigar na presente tese foi a possível correlação
entre PARP-1 e P-gp. A deficiência de PARP-1 pode ter provocado aumento da expressão de P-
gp em fibroblastos de camundongos (WURZER e cols., 2000) e em células de mieloma humano
que superexpressam P-gp (WESIERSKA-GADEK, 2005). Nesses trabalhos foi constatada uma
redução drástica na expressão da proteína supressora de tumores p53, nas células deficientes de
PARP-1. Por isso, esses autores sugeriram que a expressão de P-gp poderia ser regulada
negativamente pela p53 e, assim, a ausência dessa proteina supressora de tumores poderia
contribuir para o aumento dos teores dessa bomba de efluxo (WURZER e cols., 2000;
WESIERSKA-GADEK, 2005).
No presente estudo também foi observado que a célula com maior expressão de P-gp
(FEPS) apresentou a menor expressão e atividade da PARP-1, sugerindo possível envolvimento
dessa enzima no desenvolvimento do fenótipo MDR no modelo experimental empregado. Apesar
de a piperina ter provocado aumento da expressão de P-gp nas duas linhagens MDR, esse
composto só foi capaz de modular a expressão de PARP-1 na FEPS, sendo que conforme a
piperina reduziu a expressão de PARP-1 na FEPS, parece ter aumentado, concomitantemente, a
expressão de P-gp nessa linhagem. No entanto, como as células K562, Lucena-1 e FEPS não
apresentam níveis detectáveis da proteína p53 (DAFLON-YUNES e cols, 2013), não foi possível
estabelecer o mecanismo responsável pelo aumento de P-gp e redução do conteúdo de PARP-1
na FEPS. Além disso, também não foi identificado de que forma a modulação da P-gp e da
PARP-1 poderia ter correlação com o aumento da sensibilidade das células MDR para a ação da
piperina.

94
A superexpressão de P-gp em células MDR pode provocar alterações, de forma indireta,
na propriedade físico-química da membrana plasmática, conferindo maior instabilidade
(SHAROM, 2014). Por isso, algumas substâncias promotoras de CS seriam capazes de aumentar
a permeabilidade e fluidez da membrana de células MDR que superexpressam P-gp, exercendo
efeito citotóxico seletivo e revertendo o fenótipo MDR (ZINZI e cols., 2014). Há um relato de
que a piperina seria capaz de aumentar a permeabilidade das células intestinais, e, com isso,
promover o aumento da biodisponibilidade de diversas substâncias (KHAJURIA e cols., 2002).
Assim, é possível supor que nas linhagens de leucemia MDR, empregadas no presente estudo, a
piperina também promove aumento da permeabilidade, o que justificaria o efeito da piperina em
induzir CS nessas células.
Buscou-se testar a hipótese de que diferenças no metabolismo da piperina, entre a
linhagem sensível a drogas e as linhagens MDR, justificariam seu efeito de indução de CS. Caso
a piperina estivesse sendo mais metabolizada na K562, indicaria que os metabólitos teriam menor
atividade do que sua molécula original. Em contrapartida, se a piperina fosse convertida em
metabólitos em maior grau nas células Lucena-1 e FEPS, indicaria que esse composto seria um
pró-fármaco, uma vez que sua metabolização melhoraria seus efeitos biológicos.
Primeiramente, foram realizados experimentos a fim de avaliar algumas enzimas
responsáveis pelo metabolismo de fármacos. Alguns marcadores importantes do metabolismo de
xenobióticos como as enzimas de fase I, CYP3A4 e CYP1B1, não foram identificados nos
experimentos por RT-PCR em tempo real em nenhuma das linhagens de leucemia utilizadas no
presente estudo.
Apesar disso, diferentes trabalhos, desenvolvidos por um mesmo grupo de pesquisa no
Japão, detectaram teores basais de mRNA de CYP3A4 (NAGAI e cols., 2002; NAGAI e cols.,
2004; NAGAI e cols., 2007) e CYP1B1 (NAGAI e cols., 2002; NAGAI e cols., 2007) na
linhagem K562. No entanto, vale ressaltar que os pares de primers utilizados nos estudos citados
não eram específicos para os genes de interesse, o que poderia ter gerado resultados falso-
positivos. De outro modo, os resultados de Wang e cols. (2004) estão de acordo com os que
foram apresentados nessa tese em relação à não detecção de expressão gênica de CYP3A4 na
linhagem K562.
Experimentos realizados pela técnica microarray conduzidos por Moreira e cols. (2014),
utilizando as mesmas linhagens do presente trabalho, identificaram diferenças de expressão de

95
CYP3A5 entre as células de leucemia (6 vezes maior na FEPS, em relação a K562), sugerindo
possível relação com a MDR.
Em geral, a exposição das células a xenobióticos pode desencadear aumento da expressão
gênica de enzimas do complexo CYP450 como um mecanismo de defesa contra essas
substâncias, de modo a permitir a sua eliminação de maneira mais eficiente (LOLODI e cols.,
2017). Em contrapartida, algumas substâncias denominadas de pró-fármacos podem se beneficiar
desse mecanismo de defesa, uma vez que nesses casos, o processo de metabolização seria capaz
de gerar metabólitos com atividade biológica ainda mais potente do que sua molécula original,
afetando seletivamente as células com essa característica (ZHANG e cols, 2017). A fim de
investigar se CYP3A5 estaria relacionada com o efeito mais pronunciado da piperina na FEPS,
foram executados experimentos para avaliar a expressão desse gene.
Foram validados os resultados de Moreira e cols. (2014), no entanto, foi constatada
diferença expressivamente maior entre K562 e FEPS (cerca de 240 vezes em relação a K562),
provavelmente por conta da maior precisão do método utilizado no presente estudo (RT-PCR em
tempo real). Além disso, também foi observada maior expressão do gene CYP3A5 na FEPS, em
relação à Lucena-1 (cerca de 5 vezes).
O aumento da expressão gênica basal de CYP3A5 nas linhagens Lucena-1 e FEPS pode
ser consequência do processo de seleção das células resistentes. A linhagem Lucena-1 foi
selecionada a partir da exposição à concentrações crescentes de vincristina, que é
preferencialmente metabolizada pela CYP3A5 (DENNISON e cols., 2007; SABA e cols., 2015).
Com relação à FEPS, que foi selecionada por daunorrubicina, existem evidências de que esse
quimioterápico também seria metabolizado pela CYP3A5 (HUANG e cols., 2017).
Wang e cols. (2004) investigaram o envolvimento de CYP3A5 na aquisição de resistência
a drogas nas linhagens de leucemia K562 e U937. Foi demonstrado que a expressão gênica de
CYP3A5, em células de leucemia, poderia causar a metabolização in situ de diversas drogas
anticâncer, levando à MDR. Dados desse mesmo grupo de pesquisa revelaram, ainda, possível
correlação entre CYP3A5 e o fenótipo MDR, uma vez que a transfecção das células de leucemia
HL-60 com o gene CYP3A5 foi capaz de induzir diretamente a resistência a antraciclinas e
alcaloides (WANG e cols., 2005).
Atualmente, a terapia mais efetiva para LMC é a utilização de inibidores seletivos da bcr-
abl, como mesilato de imatinibe. Meta análise envolvendo diversos estudos clínicos indicou a

96
importância de CYP3A5 na metabolização do mesilato de imatinibe em seu principal metabólito
ativo (CARGNIN e cols., 2018), indicando que compostos que modifiquem ou induzam CYP3A5
possam vir a ter grande importância clínica.
Os experimentos realizados na presente tese indicaram que a piperina induziu a expressão
gênica de CYP3A5. No entanto, ainda não foi possível identificar o mecanismo de indução dessa
enzima. Foi avaliada a expressão de PXR, principal regulador transcricional da família CYP3A,
mas nenhuma linhagem de leucemia apresentou expressão basal desse receptor. Por isso, sugere-
se que a indução de CYP3A5 tenha ocorrido de maneira independente de PXR, embora já tenha
sido sugerido o envolvimento desse receptor na indução de CYP3A4 após o tratamento com
piperina (WANG e cols., 2013).
Tem sido observado efeito dicotômico da piperina na modulação de CYP, uma vez que
esse composto poderia inibir ou estimular a atividade dessas enzimas dependendo da dose
utilizada, frequência e/ ou duração do tratamento. Alguns estudos indicam que a piperina teria a
capacidade de inibir múltiplas enzimas do complexo CYP450 (BHARDWAJ e cols., 2002;
SHAMSI e cols., 2017; LEE e cols., 2018), sugerindo possível aumento da biodisponibilidade de
diversos medicamentos (DUDHATRA e cols., 2012). No entanto, outros autores observaram que
a piperina também seria capaz de induzir a expressão de enzimas metabolizadoras de
xenobióticos. No estudo de Singh e Rao (1993) foram observados teores elevados de CYPB5 no
fígado de camundongos após a ingestão de pimenta-do-reino. Uma pesquisa publicada por Wang
e cols. (2013) demonstrou que esse composto induziu a expressão gênica de CYP3A4 em
hepatócitos humanos e em células intestinais.
Como a piperina provocou indução do gene CYP3A5 e, ainda, demonstrou interagir com
essa enzima, por docking molecular, foi possível supor que esse composto estaria sofrendo
biotransformação por CYP3A5, sendo convertido em metabólitos mais citotóxicos, de forma
mais pronunciada nas linhagens MDR. No entanto, não foi possível observar diferenças na
metabolização de piperina, pelo método CLAE-DAD, entre as linhagens MDR e a linhagem
sensível a drogas, K562. Recentemente, Zabela e cols. (2018) investigaram o metabolismo da
piperina e seus análogos utilizando microssomos hepáticos humanos. Foi constatado que a
piperina foi o composto mais estável metabolicamente, embora tenha sido metabolizada
exclusivamente pela enzima hepática CYP1A2. Contudo, nesse trabalho não foi investigado o
papel da CYP3A5 no metabolismo da piperina. Dados do presente trabalho indicam que a

97
piperina não estaria sendo preferencialmente metabolizada por CYP3A5, pois, na teoria, a célula
com a maior expressão desse gene (FEPS), deveria apresentar maiores proporções de
metabólitos, o que não foi possível comprovar na presente tese.
No experimento para avaliação dos metabólitos de piperina utilizamos a técnica de
CLAE-DAD e detectamos dois picos como possíveis metabólitos, mas não identificamos essas
substâncias devido à ausência de padrões comerciais que possibilitasse a comparação dos perfis
cromatográficos. Adicionalmente, também não foi possível determinar se esses compostos são os
mesmos que já foram descritos na literatura científica por Gao e cols. (2017), já que esses autores
identificaram os metabólitos por UHPLC/QTOF-MS (do inglês, Ultra-high Performance Liquid
Chromatography-quadrupole Time-of-flight Mass Spectrometry), com base em seu espectro de
massas e sua fórmula molecular.
Além disso, é possível que o método utilizado na presente tese não tenha sido sensível o
suficiente para detecção de todos os metabólitos produzidos a partir da piperina. Por isso, não é
possível excluir a possibilidade de terem sido gerados outros metabólitos, além dos que foram
descritos. Técnicas cromatográficas mais sofisticadas e sensíveis, como a LC-MS/MSn (do inglês,
Liquid Chromatography-electrospray Ionization/multi-stage Mass Spectrometry), seriam mais
indicadas para essa pesquisa, uma vez que permitiriam a identificação de metabólitos presentes
em baixas concentrações, com base na sua fórmula molecular e perfil de fragmentação (PITT,
2009).
No entanto, a identificação de percentuais semelhantes de piperina (em torno de 70%) nas
três linhagens celulares foi fundamental para sustentar a hipótese de que a metabolização desse
composto seria semelhante nas células de leucemia testadas no presente estudo. Essa constatação
foi a mais importante dessa análise, uma vez que se pretendia identificar se esse poderia ser o
mecanismo responsável pelo efeito diferenciado da piperina nas células de leucemia com
fenótipo MDR. No entanto, essa correlação não foi confirmada.
Além das CYP’s serem essenciais para a metabolização de xenobióticos, também têm
papel importante na biossíntese de esteroides que dependem da transferência de elétrons do
NADPH. No entanto, esse sistema pode funcionar como um ciclo fútil de NADPH oxidase,
oxidando NADPH na ausência de substrato, produzindo superóxido e outros ROS
(HANUKOGLU e cols., 2006; JIANG e cols., 2015). Por isso, existe possibilidade da piperina

98
estar gerando ROS via CYP3A5, de forma mais intensa na FEPS, uma vez que essa linhagem
apresenta maior expressão desse gene.
Outros trabalhos identificaram possível correlação entre a indução da expressão de CYP e
o aumento da formação de ROS. A superexpressão de CYP3A5 mostrou desempenhar papel
importante impedindo a progressão do carcinoma hepatocelular, provocando aumento da geração
de ROS e reduzindo metástases. A indução de ROS via CYP3A5 inibiu seletivamente a
fosforilação de AKT por meio do bloqueio da atividade da quinase mTORC2 (JIANG e cols.,
2015). Já havia sido demonstrado que o antifúngico propiconazole, utilizado na agricultura, seria
capaz de levar à formação de tumores hepáticos em camundongos. No entanto, ainda não se sabia
se este composto atuaria induzindo ROS via CYP450 (NESNOW e cols., 2011). Nesnow e cols.
(2011) buscaram avaliar o mecanismo de ação desse antifúngico e revelaram que os teores
aumentados de ROS, após o tratamento com propiconazol, são devido ao aumento da expressão
de enzimas do complexo CYP450, conhecidas por desacoplarem, produzindo elevação de ROS.
Recente revisão publicada por Rather e Bhagat (2018) destacou diversos estudos que
apontaram que piperina seria capaz de alterar o balanço redox celular, influenciando a fisiologia
da célula por diferentes mecanismos de ação. Dependendo do modelo biológico, piperina
aumenta a sobrevivência celular ou induz a morte celular, atuando como antioxidante (em
menores concentrações) ou como pró-oxidante (em maiores concentrações). Por isso, a piperina é
caracterizada como potente agente anticâncer, seja na prevenção ou no tratamento dessa doença.
Embora exista a possibilidade de relação entre a ação da piperina e a modulação de
CYP3A5 e ROS, estudos do nosso grupo de pesquisa indicam que as linhagens Lucena-1 e FEPS
são mais resistentes ao estresse oxidativo que a linhagem sensível a drogas, K562, provavelmente
devido a maior capacidade antioxidante das células com fenótipo MDR (TRINDADE e cols.,
1999; MOREIRA e cols., 2014; VIDAL, 2017; SOARES e cols., 2019). Por isso, essa hipótese
deve ser investigada mais detalhadamente no futuro.
Experimentos prévios realizados pela autora durante o mestrado sugerem que o
mecanismo da piperina em induzir CS seria dependente da presença de P-gp na membrana dessas
células tumorais, uma vez que as células Lucena-1 e FEPS silenciadas para o gene MDR1 foram
menos sensíveis a ação da piperina do que as células controle (QUARTI, 2015). No entanto, a
piperina poderia promover CS por exercer efeitos diretos sobre a P-gp, como foi demonstrado na

99
presente tese por meio do aumento da expressão deste transportador nas células MDR, ou então
essa modulação na P-gp poderia estar provocar outras alterações na célula.
Além da superexpressão de P-gp, as células MDR apresentam outras importantes
diferenças em relação à K562, como as que foram demonstradas na presente pesquisa: expressão
aumentada de GAPDH, diminuição da expressão de PARP-1 e PAR (apenas na FEPS) e aumento
do gene CYP3A5. A piperina também parece atuar por diferentes vias, induzindo CS, como
promovendo maior degradação da PARP-1 (apenas na FEPS), aumentando a expressão de P-gp e
induzindo CYP3A5. No entanto, ainda são necessários novos estudos a fim de estabelecer os
mecanismos causais dessas alterações.

100
7. CONCLUSÕES
As linhagens de LMC utilizadas no presente estudo diferiram quanto à resposta à piperina.
Ao invés das células multirresistentes exibirem resistência à ação da piperina, apresentaram
sensibilidade maior a esse composto, caracterizando, pela primeira vez, a propriedade desse
fitoquímico em induzir CS. Em contrapartida, a piperina apresentou efeito citotóxico reduzido
sobre macrófagos peritoneais de camundongos.
O efeito mais pronunciado da piperina sobre as linhagens MDR ficou evidenciado por
meio das análises de morfologia celular, apoptose e clivagem de caspase-3, mas não pelo ciclo
celular:
- a piperina promoveu indução da formação de vacúolos no citoplasma das células de
leucemia, que, por sua vez, pode ter gerado aumento da granulosidade celular, em maior
grau na FEPS.
- a piperina não alterou a distribuição do ciclo celular, mas induziu apenas as células
MDR ao processo de apoptose e aumento de clivagem de caspase-3, sendo que esse efeito
foi mais pronunciado sobre a linhagem FEPS.
Além disso, a piperina também modulou PARP-1, P-gp e CYP3A5, mas não afetou a
expressão de GAPDH e não sofreu metabolização em maior grau nas células MDR:
- as células MDR superexpressaram a enzima glicolítica GAPDH, indicando maior
resistência à MCIC, no entanto, a piperina não foi capaz de induzir MCIC via inibição de
GAPDH nessas linhagens.
- a linhagem FEPS apresentou os menores teores de PARP-1, bem como a menor
atividade (inferida pelo conteúdo do polímero PAR), além disso, foi a única que
apresentou quantidades detectáveis do fragmento de 24 kDa, sugerindo o
desenvolvimento de mecanismo capaz de impedir a superativação de PARP-1, o que
poderia proteger essas células contra morte por necrose. No entanto, a piperina foi capaz
de reduzir os teores de PARP-1 e do fragmento de 24 kDa, indicando possível relação
com a indução de CS.
- a piperina aumentou a expressão de P-gp em ambas as linhagens MDR, no entanto,
somente na FEPS haveria possível relação entre a diminuição da expressão de PARP-1 e o
aumento da expressão de P-gp após incubação com esse fitoquímico.

101
- as linhagens de leucemia não apresentaram quantidades detectáveis de CYP3A4,
CYP1B1 e PXR. No entanto, a linhagem FEPS apresentou a maior expressão de CYP3A5,
seguida da Lucena-1, sendo que a piperina foi capaz de induzir ainda mais a expressão
desse gene nessas células MDR. Além disso, a piperina ainda demonstrou interagir com a
proteína CYP3A5, indicando mais uma possível relação com a indução de CS.
- não foram observadas diferenças de metabolização de piperina entre as linhagens de
leucemia, sugerindo que, provavelmente, CYP3A5 não estaria metabolizando esse
fitoquímico.
Esse conjunto de resultados sugere que a piperina atuou por diferentes vias de sinalização,
sendo que algumas delas, como a modulação de PARP-1 e de CYP3A5, ainda não tinham sido
descritas como possíveis alvos para a indução de CS na literatura científica.
A identificação de agentes promotores de CS, bem como dos seus mecanismos de ação,
pode levar a descoberta de compostos que sejam efetivos em prevenir a MDR por meio da
administração adjuvante desde o início da quimioterapia ou tornar o tratamento quimioterápico
novamente eficiente por meio da morte seletiva de células com fenótipo MDR.

102
8. REFERÊNCIAS
ABOU-ZAHR, A.; BORTHAKUR, G. Emerging cell cycle inhibitors for acute myeloid
leukemia. Expert Opinion on Emerging Drugs, v. 22, p. 137-148, 2017.
AGÊNCIA NACIONAL DE VIGILÂNCIA SANITÁRIA (ANVISA). Disponível em:
http://portal.anvisa.gov.br/ Acessado em: 02 de março de 2019 (1999).
AKHDAR, H.; LEGENDRE, C.; ANINAT, C.; MOREL, F. Anticancer Drug Metabolism:
Chemotherapy Resistance and New Therapeutic Approaches. Rijeka: INTECH Open Access
Publisher, p. 138-170, 2012.
AKHIANI, A. A.; WERLENIUS, O.; AURELIUS, J.; MOVITZ, C.; MARTNER,
A.; HELLSTRAND, K.; THORÉN, F. B. Role of the ERK pathway for oxidant-induced
parthanatos in human lymphocytes. PLoS ONE, v. 9, e89646, 2014.
ANDRABI, S. A.; KIM, N. S.; YU, S. W.; WANG, H.; KOH, D. W; SASAKI, M. Poly(ADP-
ribose) (PAR) polymer is a death signal. Proceedings of the NationalAcademy of Sciences, v.
103, p. 18308-18313, 2006.
ANKATHIL, R.; AZLAN, H.; DZARR, A. A.; BABA, A. A. Pharmacogenetics and the
treatment of chronic myeloid leukemia: how relevant clinically? An update. Pharmacogenomics,
v. 19, p. 475-493, 2018.
ASNAFI, A. A.; DERIS, Z. Z.; SHAHRABI, S.; ZIBARA, K.; VOSUGHI, T. Chronic myeloid
leukemia with complex karyotypes: Prognosis and therapeutic approaches. Journal of Cellular
Physiology, v. 234, p. 5798-5806, 2018.
ATAL, N.; BEDI, K. L. Bioenhancers: revolutionary concept to market. Journal of Ayurveda &
Integrative Medicine, v. 1, p. 96-99, 2010.
https://www.ncbi.nlm.nih.gov/pubmed/?term=Akhiani%20AA%5BAuthor%5D&cauthor=true&cauthor_uid=24586933

103
AYED-BOUSSEMA, I.; PASCUSSI, J. M.; MAUREL, P.; BACHA, H.; HASSEN,
W. Zearalenone activates pregnane X receptor, constitutive androstane receptor and aryl
hydrocarbon receptor and corresponding phase I target genes mRNA in primary cultures of
human hepatocytes. Environmental Toxicology and Pharmacology, v. 31, p. 79-87, 2011.
BA, Y.; MALHOTRA, A. Potential of piperine in modulation of voltage-gated K+ current and its
influences on cell cycle arrest and apoptosis in human prostate cancer cells. European Review
for Medical and Pharmacological Science, v. 22, p. 8999-9011, 2018.
BAJAD, S.; JOHRI, R. K.; SINGH, K.; SINGH, J.; BEDI, K. L. Simple high-performance liquid
chromatography method for the simultaneous determination of ketoconazole and piperine in rat
plasma and hepatocyte culture. Journal of Chromatography A, v. 949, p. 43-47, 2002.
BARILE, F. A. In vitro cytotoxicology: mechanisms and methods (1ª Ed.). Florida: CRC Press,
p. 222, 1994.
BECHOR, S.; NACHMIAS, D.; ELIA, N.; HAIM, Y.; VATARESCU, M.; LEIKIN-FRENKEL,
A.; GERICKE, M.; TARNOVSCKI, T.; LAS, G.; RUDICH, A. Adipose tissue conditioned
media support macrophage lipid-droplet biogenesis by interfering with autophagic flux.
Biochimica et Biophysica Acta - Molecular and Cell Biology of Lipids, v. 1862, p. 1001-1012,
2017.
BERGER, N. A. Poly(ADP-ribose) in the cellular response to DNA damage. Radiation
Research, v. 101, p. 4-15, 1985.
BHARDWAJ, R. K.; GLAESER, H.; BECQUEMONT, L.; KLOTZ, U.; GUPTA, S. K.;
FROMM, M. F. Piperine, a major constituent of black pepper, inhibits human P-glycoprotein and
CYP3A4. The Journal of Pharmacology and Experimental Therapeutics, v. 302, p. 645-650,
2002.

104
BOOZARI, M.; MOHAMMADI, A.; ASILI, J.; EMAMI, S.A.; TAYARANI-NAJARAN, Z.
Growth inhibition and apoptosis induction by Scutellaria pinnatifida A. Ham. on HL-60 and
K562 leukemic cell lines. Environmental Toxicology and Pharmacology, v. 39, p. 307-312,
2015.
BOWER, A.; MARQUEZ, S.; DE MEJIA, E. G. The Health Benefits of Selected Culinary Herbs
and Spices Found in the Traditional Mediterranean Diet. Critical Reviews in Food Science and
Nutrition, v. 56, p. 2728-2746, 2016.
BOZZA, P. T.; VIOLA, J. P. Lipid droplets in inflammation and cancer. Prostaglandins,
Leukotrienes & Essential Fatty Acids, v. 82, p. 243-250, 2010.
BUDAVARI, S. The merck index: an encyclopedia of chemicals, drugs, and biological (12ª Ed.).
Nova Jersey: Editora Merck, 1996.
CALLAGHAN, R.; RIORDAN, J. R. Collateral sensitivity of multidrug resistant cells to narcotic
analgesics is due to effects on the plasma membrane. Biochimica et Biophysica Acta-
Molecular and Cell Biology of Lipids, v. 24, p. 155-162, 1995.
CARGNIN, S.; RAVEGNINI, G.; SOVERINI, S.; ANGELINI, S.; TERRAZZINO, S. Impact of
SLC22A1 and CYP3A5 genotypes on imatinib response in chronic myeloid leukemia: A
systematic review and meta-analysis. Pharmacological Research, v. 131, p. 244-254, 2018.
CARRETT-DIAS, M.; ALMEIDA, L. K.; PEREIRA, J. L.; ALMEIDA, D. V.; FILGUEIRA, D.
M.; MARINS, L. F.; VOTTO, A. P.; TRINDADE, G. S. Cell differentiation and the multiple
drug resistance phenotype in human erythroleukemic cells. Leukemia Research, v. 42, p. 13-20,
2016.
CASTRO, D. J.; BAIRD, W. M.; PEREIRA, C. B.; GIOVANINI, J.; LOHR, C. V.; FISCHER,
K. A.; YU, Z.; GONZALEZ, F. J.; KRUEGER, S. K.; WILLIAMS, D. E. Fetal mouse Cyp1b1

105
and transplacental carcinogenesis from maternal exposure to dibenzo(a,l)pyrene. Cancer
Prevention Research, v. 1, p. 128-134, 2008.
CHAFFER, C. L.; SAN JUAN, B. P.; LIM, E.; WEINBERG, R. A. EMT, cell plasticity and
metastasis. Cancer and Metastasis Reviews, v. 35, p. 645-654, 2016.
CHAITANYA, G. V.; STEVEN, A. J.; BABU, P. P. PARP-1 cleavage fragments: signatures of
cell-death proteases in neurodegeneration. Cell Communication and Signaling, v. 8, p. 31,
2010.
CHEN, T. Overcoming drug resistance by regulating nuclear receptors. Advanced Drug
Delivery Reviews, v. 62, p. 1257-1264, 2010.
CHI, A. C.; APPLETON, K.; HENRIOD, J. B.; KRAYER, J. W.; MARLOW, N. M.;
BANDYOPADHYAY, D.; SIGMON, R. C.; KURTZ, D. T. Differential Induction of CYP1A1
and CYP1B1 by Benzo[a]pyrene in Oral Squamous Cell Carcinoma Cell Lines and by Tobacco
Smoking in Oral Mucosa. Oral Oncology, v. 45, p. 980-985, 2009.
CHIANG, H. L.; TERLECKY, S. R.; PLANT, C. P.; DICE, J. F. A role for a 70-kilodalton heat
shock protein in lysosomal degradation of intracellular proteins. Science, v. 246, p. 382-385,
1989.
CHIBA, A.; TOYA, T.; MIZUNO, H.; TOKUSHIGE, J.; NAKAMURA, F.; NAKAZAKI, K.;
KUROKAWA, M. Chronic myelogenous leukemia presenting with central nervous system
infiltration, successfully treated with central nervous system-directed chemotherapy followed by
allogeneic stem cell transplantation. International Journal of Hematology, v. 108, p. 640-646,
2018.
CHUCHAWANKUL, S.; KHORANA, N.; POOVORAWAN, Y. Piperine inhibits cytokine
production by human peripheral blood mononuclear cells. Genetics and Molecular Research, v.
11, p. 617-627, 2012.

106
CLARK, A. C. Caspase Allostery and Conformational Selection. Chemical Reviews, v. 116, p.
6666-6706, 2016.
CLAUS, H.; GERHARD, S. Piperine - an example of individually different (polymorph)
metabolism of an omnipresent nutrition component. Liebigs Annalen der Chemie, p. 1319-
1331, 1984.
CORNWELL, M. M.; PASTAN, I.; GOTTESMAN, M. M. Certain calcium channel blockers
bind specifically to multidrug resistant human KB carcinoma membrane vesicles and inhibit drug
binding to P-glycoprotein. The Journal of Biological Chemistry, v. 262, p. 2166-2170, 1987.
COTTE, A. K.; AIRES, V.; FREDON, M.; LIMAGNE, E.; DERANGÈRE, V.; THIBAUDIN,
M.; HUMBLIN, E.; SCAGLIARINI, A.; DE BARROS, J. P. P.; HILLON, P.
Lysophosphatidylcholine acyltransferase 2-mediated lipid droplet production supports colorectal
cancer chemoresistance. Nature Communications, v. 9, p. 322, 2018.
CREE, I. A.; CHARLTON, P. Molecular chess? Hallmarks of anti-cancer drug resistance. BMC
Cancer, v. 17, p. 10, 2017.
CROWE, K. M.; FRANCIS, C.; Academy of Nutrition and Dietetics. Position of the academy of
nutrition and dietetics: functional foods. Journal of the Academy of Nutrition and Dietetics, v.
113, p. 1096-1103, 2013.
DAFLON-YUNES, N.; PINTO-SILVA, F. E.; VIDAL, R. S.; NOVIS, B. F.; BERGUETTI, T.;
LOPES, R. R.; POLYCARPO, C.; RUMJANEK, V. M. Characterization of a multidrug-resistant
chronic myeloid leukemia cell line presenting multiple resistance mechanisms. Molecular and
Cellular Biochemistry, v. 383, p. 123-135, 2013.
D'AMOURS, D.; SALLMANN, F. R.; DIXIT, V. M; POIRIER, G. G. Gain-of-function of poly
(ADP-ribose) polymerase-1 upon cleavage by apoptotic proteases: implications for apoptosis.
Journal of Cell Science, v. 114, p. 3771-3778, 2001.

107
DAVID, K. K; ANDRABI, S. A.; DAWSON, T. M.; DAWSON, V. L. Parthanatos, a messenger
of death. Frontiers in Bioscience, v. 14, p. 1116-1128, 2009.
DENNISON, J. B.; JONES, D. R.; RENBARGER, J. L.; HALL, S. D. Effect of CYP3A5
expression on vincristine metabolism with human liver microsomes. Journal of Pharmacology
and Experimental Therapeutics, v. 321, p. 553-563, 2007.
DI PIETRO, A.; DAYAN, G.; CONSEIL, G.; STEINFELS, E.; KRELL, T.; TROMPIER, D.;
BAUBICHON-CORTAY, H.; JAULT, J. P-glycoprotein-mediated resistance to chemotherapy in
cancer cells: using recombinant cytosolic domains to establish structure function relationships.
Brazilian Journal of Medical and Biological Research, v. 32, p. 925-939, 1999.
DO, M. T.; KIM, H. G.; CHOI, J. H.; KHANAL, T.; PARK, B. H.; TRAN, T. P.; JEONG, T. C.;
JEONG, H. G. Antitumor efficacy of piperine in the treatment of human HER2-overexpressing
breast cancer cells. Food Chemistry, v. 141, p. 2591-2599, 2013.
DÖRING, B.; PETZINGER, E. Phase 0 and phase III transport in various organs: combined
concept of phases in xenobiotic transport and metabolism. Drug Metabolism Reviews, v. 46, p.
261-282, 2014.
DUDHATRA, G. B.; MODY, S. K.; AWALE, M. M.; PATEL, H. B.; MODI, C. M.; KUMAR,
A.; KAMANI, D. R.; CHAUHAN, B. N. A comprehensive review on pharmacotherapeutics of
herbal bioenhancers. The Scientific World Journal, v. 2012, p. 1-33, 2012.
FAN, J.; LANNOY, I. A. M. Pharmacokinetics. Biochemical Pharmacology, v. 87, p. 93-120,
2014.
FOFARIA, N. M.; KIM, S. H.; SRIVASTAVA, S. K. Piperine causes G1 phase cell cycle arrest
and apoptosis in melanoma cells through checkpoint kinase-1 activation. PLoS One, v. 9,
e94298, 2014.

108
GAO, T.; XUE, H.; LU, L.; ZHANG, T,.; HAN, H. Characterization of piperine metabolites in
rats by ultra-high-performance liquid chromatography with electrospray ionization quadruple
time-of-flight tandem mass spectrometry. Rapid Communications in Mass Spectrometry, v.
31, p. 901-910, 2017.
GIAMPAZOLIAS, E.; TAIT, S. W. G. Caspase-independent cell death: An anti-cancer double
whammy. Cell Cycle, v. 17, p. 269-270, 2018.
GOTTESMAN, M. M.; AMBUDKAR, S. V.; XIA, D. Structure of a multidrug transporter.
Nature Biotechnology, v. 27, p. 546-547, 2009.
GOTTESMAN, M. M.; LING, V. The molecular basis of multidrug resistance in cancer: the
early years of P-glycoprotein research. FEBS letters, v. 580, p. 998-1009, 2006.
GOTTESMAN, M. M.; PASTAN, I. Biochemistry of multidrug resistance mediated by the
multidrug transporter. Annual Review of Biochemistry, v. 62, p. 385-427, 1993.
GREENSHIELDS A. L.; DOUCETTE, C. D.; SUTTON, K. M.; MADERA, L.; ANNAN, H.;
YAFFE, P. B.; KNICKLE, A. F.; DONG, Z.; HOSKIN, D. W. Piperine inhibits the growth and
motility of triple-negative breast cancer cells. Cancer Letters,v. 357, p. 129-140, 2015.
GUENGERICH, F. P. Cytochrome P450 and chemical toxicology. Chemical Research in
Toxicology, v. 21, p. 70-83, 2008.
GUENGERICH, F. P. Mechanisms of cytochrome P450 substrate oxidation: Mini Review.
Journal of Biochemical and Molecular Toxicology, v. 21, p. 163-168, 2007.
GUILLAUMOND, F.; BIDAUT, G.; OUAISSI, M.; SERVAIS, S.; GOUIRAND, V.;
OLIVARES, O.; LAC, S.; BORGE, L.; ROQUES, J.; GAYET, O. Cholesterol uptake disruption,
in association with chemotherapy, is a promising combined metabolic therapy for pancreatic

109
adenocarcinoma. Proceedings of the National Academy of Sciences, v. 112, p. 2473-2478,
2015.
GUNASEKARAN, V.; ELANGOVAN, K.; NIRANJALI DEVARAJ, S. Targeting
hepatocellular carcinoma with piperine by radical-mediated mitochondrial pathway of apoptosis:
An in vitro and in vivo study. Food and Chemical Toxicology, v. 105, p. 106-118, 2017.
GUPTA, D.; VENKATESH, M.; WANG, H.; KIM, S.; SINZ, M.; GOLDBERG, G. L.;
WHITNEY, K.; LONGLEY, C.; MANI, S. Expanding the roles for pregnane X receptor in
cancer: proliferation and drug resistance in ovarian cancer. Clinical Cancer Research, v. 14, p.
5332-5340, 2008.
HAGER, M. H.; SOLOMON, K. R.; FREEMAN, M. R. The role of cholesterol in prostate
cancer. Current Opinion Clinical Nutrition & Metabolic Care, v. 9, p. 379-385, 2006.
HALL, M. D.; HANDLEY, M. D.; GOTTSMAN, M. M. Is resistance useless? Multidrug
resistance and collateral sensitivity. Trends in Pharmacological Sciences, v. 30, p. 546–556,
2009.
HANAHAN, D.; WEINBERG, R. A. Hallmarks of cancer: the next generation. Cell, v. 144, p.
646-674, 2011.
HANUKOGLU, I. Antioxidant protective mechanisms against reactive oxygen species (ROS)
generated by mitochondrial P450 systems in steroidogenic cells. Drug Metabolism Reviews, v.
38, p.171-196, 2006.
HASLER, C. M.; BROWN, A. C; American Dietetic Association. Position of American Dietetic
Association: functional foods. Journal of the American Dietetic Association, v. 104, p. 814-
826, 2009.

110
HEHLMANN, R.; SAUBELE, S.; VOSKANYAN, A.; SILVER, R. T. Management of CML-
blast crisis. Best Practice & Research Clinical Haematology, v. 29, p. 295-307, 2016.
HERCEG, Z.; WANG, Z. Q. Failure of poly(ADP-ribose) polymerase cleavage by caspases leads
to induction of necrosis and enhanced apoptosis. Molecular and Cellular Biology, v. 19, p.
5124-5133, 1999.
HSU, M. H; SAVAS, U.; JOHNSON, E. F. The X-Ray Crystal Structure of the Human Mono-
Oxygenase Cytochrome P450 3A5-Ritonavir Complex Reveals Active Site Differences between
P450s 3A4 and 3A5. Molecular Pharmacology. v. 93, p. 14-24, 2018.
HUANG, Z.; WANG, J.; QIAN, J.; LI, Y.; XU, Z.; CHEN, M.; TONG, H. Effects of cytochrome
P450 family 3 subfamily A member 5 gene polymorphisms on daunorubicin metabolism and
adverse reactions in patients with acute leukemia. Molecular Medicine Reports, v. 15, p. 3493-
3498, 2017.
HUEY, R., MORRIS, G. M. Using AutoDock 4 with AutoDocktools: a tutorial. The Scripps
Research Institute, p. 54-56, 2008.
INSTITUTO NACIONAL DO CÂNCER (INCA). Estimativa 2018 - Incidência de Câncer no
Brasil. Ministério da Saúde, 2018. Disponível em:
https://www.inca.gov.br/publicacoes/livros/estimativa-2018-incidencia-de-cancer-no-brasil.
Acessado em: 08 de março de 2019 (2018).
JABBOUR, E.; KANTARJIAN, H. Chronic myeloid leukemia: 2018 update on diagnosis,
therapy and monitoring. Americal Journal of Hematology, v. 93, p. 442-459, 2018.
JANCOVA, P.; ANZENBACHER, P.; ANZENBACHEROVA, E. Phase II drug metabolizing
enzymes. Biomedical Papers of the Medical Faculty of the University Palacky, v. 154, p. 103-
116, 2010.

111
JARAMILLO, A. C.; SAIG, F. A.; CLOOS, J.; JANSEN, G.; PETERS, G. J. How to overcome
ATP-binding cassette drug efflux transporter-mediated drug resistance? Cancer Drug Resist, v.
1, p. 6-29, 2018.
JIANG, F.; CHEN, L.; YANG, Y. C.; WANG, X. M.; WANG, R. Y.; LI, L.; WEN, W.;
CHANG, Y. X.; CHEN, C. Y.; TANG, J.; LIU, G. M.; HUANG, W. T.; XU, L.; WANG, H. Y.
CYP3A5 Functions as a Tumor Suppressor in Hepatocellular Carcinoma by Regulating
mTORC2/Akt Signaling. Cancer Research, v. 75, p. 1470-1481, 2015.
JIANG, H. Y.; YANG, Y.; ZHANG, Y. Y.; XIE, Z.; ZHAO, X. Y.; SUN, Y.; KONG, W. J. The
dual role of poly(ADP-ribose) polymerase-1 in modulating parthanatos and autophagy under
oxidative stress in rat cochlear marginal cells of the stria vascularis. Redox Biology, v. 14, p.
361-370, 2018.
JOYCE, H.; MCCANN, A.; CLYNES, M.; LARKIN, A. Influence of multidrug resistance and
drug transport proteins on chemotherapy drug metabolism. Expert Opinion on Drug
Metabolism & Toxicology, v. 11, p. 795-809, 2015.
JULIANO, R. L.; LING, V. A surface glycoprotein modulating drug permeability in Chinese
hamster ovary cell mutants. Biochimica et Biophysica Acta, v. 455, p. 152-162, 1976.
KAMEL, D.; GRAY, C.; WALIA, J. S.; KUMAR, V. PARP Inhibitor Drugs in the Treatment of
Breast, Ovarian, Prostate and Pancreatic Cancers: An Update of Clinical Trials. Current Drug
Targets, v. 19, p. 21-37, 2018.
KELLY, R. J.; DRAPER, D.; CHEN, C. C.; ROBEY, R. W.; FIGG, W. D.; PIEKARZ, R. L.;
CHEN, X.; GARDNER, E. R.; BALIS, F. M.; VENKATESAN, A. M.; STEINBERG, S. M.;
FOJO, T.; BATES, S. E. A pharmacodynamic study of docetaxel in combination with the P-
glycoprotein antagonist tariquidar (XR9576) in patients with lung, ovarian, and cervical cancer.
Clinical Cancer Research, v. 17, p. 569-580, 2010.

112
KHAJURIA, A.; THUSU, N.; ZUTSHI, U. Piperine modulates permeability characteristics of
intestine by inducing alterations in membrane dynamics: influence on brush border membrane
fluidity, ultrastructure and enzyme kinetics. Phytomedicine: International Journal of
Phytotherapy and Phytopharmacology, v. 9, p. 224-231, 2002.
KHAMIS, A. A. A.; ALI, E. M. M.; EL-MONEIM, M. A. A.; ABD-ALHASEEB, M. M.; EL-
MAGD, M. A.; SALIM, E. I. Hesperidin, piperine and bee venom synergistically potentiate the
anticancer effect of tamoxifen against breast cancer cells. Biomedice & Pharmacotherapy, v.
105, p. 1335-1343, 2018.
KIBE, M. N.; KONYOLE, S. O.; KATHURE, D.; NGUKA, G.; OLOO, M. O.; WANGARI, P.
M. The role of phytochemicals in prevention and control of chronic diseases. International
Journal of Current Research, v. 9, p. 62540-62543, 2017.
KIM, D. H.; XING, T.; YANG, Z.; DUDEK, R.; LU, Q.; CHEN, Y. H. Epithelial mesenchymal
transition in embryonic development, tissue repair and cancer: a comprehensive overview.
Journal of Clinical Medicine, v. 7, pii: E1, 2017.
KIM, S. S.; SEONG, S.; KIM, Y. S. Synergistic effect of ginsenoside Rg3 with verapamil on the
modulation of multidrug resistance in human acute myeloid leukemia cells. Oncology Letters, v.
7, p. 1265-1269, 2014.
KINDELL, S. S. Studies on selected genotoxic and toxic properties of piperine. Tese - Drexel
University, Philadelphia, 1984.
KOEFFLER, H. P.; GOLDE, D. W. Human myeloid leukemia cell lines: a review. Blood, v. 56,
p. 344-350, 1980.
KOIZUME, S.; MIYAGI, Y. Lipid Droplets: A Key Cellular Organelle Associated with Cancer
Cell Survival under Normoxia and Hypoxia. International Journal of Molecular Science, v. 17,
p. 1430, 2016.

113
KOZUKUE, N.; PARK, M. S.; CHOI, S. H.; LEE, S. U.; OHNISHI-KAMEYAMA, M.; LEVIN,
C. E.; FRIEDMAN, M. Kinetics of light-induced cis-trans isomerization of four piperines and
their levels in ground black peppers as determined by HPLC and LC/MS. Journal of
Agricultural and Food Chemistry, v. 55, p. 7131-7139, 2007.
LABERGE, R. M.; AMBADIPUDI, R.; GEORGES, E. P-glycoprotein (ABCB1) modulates
collateral sensitivity of a multidrug resistant cell line to verapamil. Archives of Biochemistry
and Biophysics, v. 491, p. 53-60, 2009.
LAEMMLI, U. K. Cleavage of structural proteins during the assembly of the head of
bacteriophages T4. Nature, v. 227, p. 680-685, 1970.
LARTIGUE, L.; KUSHNAREVA, Y.; SEONG, Y.; LIN, H.; FAUSTIN, B.; NEWMEYER, D.
D. Caspase-independent mitochondrial cell death results from loss of respiration, not cytotoxic
protein release. Molecular Biolology of the Cell, v. 20, p. 4871-4884, 2009.
LAVALLARD, V. J.; PRADELLI, L. A.; PAUL, A.; BÉNÉTEAU, M.; JACQUEL,
A.; AUBERGER, P.; RICCI, J. E. Modulation of caspase-independent cell death leads to
resensitization of imatinib mesylate-resistant cells, Cancer Research, v. 69, p. 3013-3020, 2009.
LAWLOR, D.; MARTIN, P.; BUSSCHOTS, S.; THERY, J.; O'LEARY, J. J.; HENNESSY, B.
T.; STORDAL, B. PARP Inhibitors as P-glyoprotein Substrates. Journal of Pharmaceutical
Sciences, v. 103, p. 1913-1920, 2014.
LEE, S. H.; KIM, H. Y.; BACK, S. Y.; HAN, H. K. Piperine-mediated drug interactions and
formulation strategy for piperine: recent advances and future perspectives. Expert Opinion on
Drug Metabolism & Toxicology, v. 14, p. 43-57, 2018.
LEVY, J. M. M.; TOWERS, C. G.; THORBURN, A. Targeting autophagy in cancer. Nature
Reviews Cancer, v. 17, p. 528-542, 2017.

114
LI, C.; WANG, Q.; REN, T.; ZHANG, Y.; LAM, C. W. K.; CHOW, M. S. S.; ZUO, Z. Non-
linear pharmacokinetics of piperine and its herb-drug interactions with docetaxel in Sprague-
Dawley rats. Journal of Pharmaceutical and Biomedical Analysis, v. 128, p. 286-293, 2016.
LI, H.; KRSTIN, S.; WANG, S.; WINK, M. Capsaicin and Piperine Can Overcome Multidrug
Resistance in Cancer Cells to Doxorubicin. Molecules, v. 23, pii: E557, 2018.
LI, S.; LEI, Y.; JIA, Y.; LI, N.; WINK, M.; MA, Y. Piperine, a piperidine alkaloid from Piper
nigrum re-sensitizes P-gp, MRP1 and BCRP dependent multidrug resistant cancer cells.
Phytomedicine, v. 19, p. 83-87, 2011.
LI, Y.; LEI, Y.; YAO, N.; WANG, C.; HU, N.; YE, W.; ZANG, D.; CHEN, Z. Autophagy and
multidrug resistance in cancer. Chinese Journal of Cancer, v. 36, p. 52-62, 2017.
LIN, Y.; XU, J.; LIAO, H.; LI, L.; PAN, L. Piperine induces apoptosis of lung cancer A549 cells
via p53-dependent mitochondrial signaling pathway. Tumor Biology, v. 35, p. 3305-3310, 2014.
LIU, Y.; YADEV, V. R.; AGGARWAL, B. B.; NAIR, M. G. Inhibitory effects of black pepper
(Piper nigrum) extracts and compounds on human tumor cell proliferation, cyclooxygenase
enzymes, lipid peroxidation and nuclear transcription factor-kappa-B. Natural Product
Communications, v. 5, p. 1253-1257, 2010.
LIVAK, K.J.; SCHMITTGEN, T.D. Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods, v. 25, p. 402-408, 2001.
LOLODI, O.; WANG, Y. M.; WRIGHT, W. C.; CHEN, T. Differential Regulation of CYP3A4
and CYP3A5 and Its Implication in Drug Discovery. Current Drug Metabolism, v. 18, p. 1095-
1105, 2017.
LOWRY, O. H.; ROSEBRUOGH, N. J.; FARR, A. L.; RANDALL, R. J. Protein measurement
with the Folin phenol reagent. The Journal of Biological Chemistry, v. 193, p. 265-275, 1951.

115
LU, H.; CHEN, C. S.; WAXMAN, D. J. Potentiation of methoxymorpholinyl doxorubicin
antitumor activity by P450 3A4 gene transfer. Cancer Gene Therapy, v. 16, p. 393-404, 2009.
MA, D.; LU, B.; FENG, C.; WANG, C.; WANG, Y.; LUO, T.; FENG, J.; JIA, H.; CHI, G.;
LUO, Y.; GE, P. Deoxypodophyllotoxin triggers parthanatos in glioma cells via induction of
excessive ROS. Cancer Letters, v. 371, p. 194-204, 2016.
MAHDY, K.; SHAKER, O.; WAFAY, H.; NASSAR, Y.; HASSAN, H.; HUSSEIN, A. Effect of
some medicinal plant extracts on the oxidative stress status in Alzheimer's disease induced in
rats. European Review for Medical and Pharmacological Sciences, v. 3, p. 31-42, 2012.
MANAYI, A.; NABAVI, S. M.; SETZER, W. N.; JAFARI, S. Piperine as a potential anti-
cancer agent: a Review on Preclinical studies. Current Medicinal Chemistry, v. 25, p. 4918-
4928, 2018.
MANOHARAN, S.; BALAKRISHNAN, S.; MENON, V. P.; ALIAS, L. M.; REENA, A. R.
Chemopreventive efficacy of curcumin and piperine during 7,12-dimethylbenz[a]anthracene-
induced hamster buccal pouch carcinogenesis. Singapore Medical Journal, v. 50, p. 139-146,
2009.
MARTIN, S.; PARTON, R. G. Caveolin, cholesterol, and lipid bodies. Seminars in Cell
Developmental Biology, v. 16, p. 163-174, 2005.
MASCHAN, A.; NOVICHKOVA, G.; MIAKOVA, N.; PERSIANTSEVA, M. Very long
survival in complete cytogenetic remission in an adolescent with lymphoid blast crisis of chronic
myeloid leukemia after treatment with intensive ALL-directed chemotherapy combined with
continuous imatinib. Pediatric Blood Cancer, v. 63, p. 2243-2245, 2016.
MCGINNIS, K. M.; GNEGY, M. E.; PARK, Y. H.; MUKERJEE, N.; WANG, K. K. Procaspase-
3 and poly(ADP)ribose polymerase (PARP) are calpain substrates. Biochemical Biophysical
Research Communications, v. 263, p. 94-99, 1999.

116
MEGHWAL, M.; GOSWAMI, K. T. Piper nigrum and Piperine: An Update. Phytotherapy
Research, v. 27, p. 1121-1130, 2013.
MOLAVI, O.; MA, Z.; HAMDY, S.; LAI, R.; LAVASANIFAR, A.; SAMUEL, J. Synergistic
antitumor effects of CpG oligodeoxynucleotide and STAT3 inhibitory agent JSI-124 in a mouse
melanoma tumor model. Immunology & Cell Biology, v. 86, p. 506-514, 2008.
MONTEIRO, H. P.; OGATA, F. T.; STERN, A. Thioredoxin promotes survival signaling events
under nitrosative/oxidative stress associated with cancer development. Biomedical Journal, v.
40, p. 189-199, 2017.
MONTONI, A.; ROBU, M.; POULIOT, E.; SHAH, G. M. Resistance to PARP-inhibitors in
cancer therapy. Frontiers Pharmacology, v. 4, p. 18, 2013.
MOORE, L. B.; PARKS, D. J.; JONES, S. A.; BLEDSOE, R. K.; CONSLER, T. G.; STIMMEL,
J. B.; GOODWIN, B.; LIDDLE, C.; BLANCHARD, S. G.; WILLSON, T. M.; COLLINS, J. L.;
KLIEWER, S. A. Orphan nuclear receptors constitutive androstane receptor and pregnane X
receptor share xenobiotic and steroid ligands. The Journal of Biological Chemistry, v. 275, p.
15122-15127, 2000.
MOREAU, R.; KAUR, H. Curcumin and piperine inhibit mTORC1 signaling in intestinal
epithelial cells. TheFASEB Journal, v. 31, p. 135.8-135.8, 2017.
MOREIRA, M. A.; BAGNI, C.; DE PINHO, M. B.; MAC-CORMICK, T. M.; DOS SANTOS,
M. M.; PINTO-SILVA, F. E.; DAFLON-YUNES, N.; RUMJANEK, V. M. Changes in gene
expression profile in two multidrug resistant cell lines derived from a same drug sensitive cell
line. Leukemia Research, v. 38, p. 983-987, 2014.
MORSY, M. A.; EL-SHEIKH, A. A. K.; IBRAHIM, A. R. N.; KHEDR, M. A.; AL-TAHER, A.
Y. In silico comparisons between natural inhibitors of ABCB1/P-glycoprotein to overcome

117
doxorubicin-resistance in the NCI/ADR-RES cell line. European Journal of Pharmaceutical
Science, v. 112, p. 87-94, 2018.
MOSMANN, T. Rapid colorimetric assay for cellular growth and survival: application to
proliferation and cytotoxicity assays. Journal of Immunological Methods, v. 65, p. 55-63,
1983.
MURPHY, D. The biogenesis and functions of lipid bodies in animals, plants and
microorganisms. Progress in Lipid Research, v. 40, p. 325-438, 2001.
NAGAI, F.; HIYOSHI, Y.; SUGIMACHI, K.; TAMURA, H.O. Cytochrome P450 (CYP)
expression in human myeloblastic and lymphoid cell lines. Biological and Pharmaceutical
Bulletin, v. 25, p. 383-385, 2002.
NAGAI, F.; KATO, E.; TAMURA, H. O. Oxidative stress induces GSTP1 and CYP3A4
expression in the human erythroleukemia cell line, K562. Biological and Pharmaceutical
Bulletin, v. 27, p. 492-495, 2004.
NAGAI, F.; SHIMIZU, M.; SAKAMOTO, T.; KOBAYASHI, T.; TAMURA, H. Changes in the
expression of cytochrome P450 genes in hemin-induced differentiated K562 cells. Biological
and Pharmaceutical Bulletin, v. 30, p. 1954-1957, 2007.
NATIONAL CANCER INSTITUTE (NIH). Disponível em: https://www.cancer.gov/about-
cancer/understanding/what-is-cancer. Acessado em: 02 de fevereiro de 2019 (2015).
NESNOW, S.; GRINDSTAFF, R. D.; LAMBERT, G.; PADGETT, W. T.; BRUNO, M.; GE, Y.;
CHEN, P. J.; WOOD, C. E.; MURPHY, L. Propiconazole increases reactive oxygen species
levels in mouse hepatic cells in culture and in mouse liver by a cytochrome P450 enzyme
mediated process. Chemico-Biological Interactions, v. 194, p. 79-89, 2011.

118
NISHA, P.; SINGHAL, R. S.; PANDIT, A. B. The degradation kinetics of flavor in black pepper
(Piper nigrum L.). Journal of Food Engineering, v. 92, p. 44-49, 2009.
NOBILI, S.; LANDINI, I.; MAZZEI, T.; MINI, E. Overcoming tumor multidrug resistance using
drugs able to evade P-glycoprotein or to exploit its expression. Medicinal Research Reviews, v.
32, p. 1220-1262, 2012.
NYAMAI, D. W.; ARIKA, W.; OGOLA, P. E.; NJAGI, E. N. M.; NGUGI, M. P. Medicinally
Important Phytochemicals: An Untapped Research Avenue. Research and Reviews: Journal of
Pharmacognosy and Phytochemistry, v. 4, p. 35-49, 2016.
ORGANIZAÇÃO MUNDIAL DA SAÚDE (OMS). Disponível em: https://www.who.int/news-
room/fact-sheets/detail/cancer. Acessado em: 02 de março de 2019 (2018).
OUYANG, D. Y.; ZENG, L. H.; PAN, H.; XU, L. H.; WANG, Y.; LIU, K. P.; HE, X.H. Piperine
inhibits the proliferation of human prostate cancer cells via induction of cell cycle arrest and
autophagy. Food and Chemical Toxicology, v. 60, p. 424-430, 2013.
PAL, M. K.; JAISWAR, S. P.; SRIVASTAV, A. K.; GOYAL, S.; DWIVEDI, A.; VERMA, A.;
SINGH, J.; PATHAK, A. K.; SANKHWAR, P. L.; RAY, R. S. Synergistic effect of piperine and
paclitaxel on cell fate via cyt-c, Bax/Bcl-2-caspase-3 pathway in ovarian adenocarcinomas
SKOV-3 cells. European Journal of Pharmacology, v. 791, p. 751-762, 2016.
PAN, S. T.; LI, Z. L.; HE, Z. X.; QIU, J. X.; ZHOU, S. F. Molecular mechanisms for tumour
resistance to chemotherapy. Clinical and Experimental Pharmacology Physiology, v. 43, p.
723-737, 2016.
PANI, B.; NUDLER, E. Mechanistic insights into transcription coupled DNA repair. DNA
Repair (Amst), v. 56, p. 42-50, 2017.

119
PATIAL, V. S. M.; SHARMA, S.; PRATAP, K.; SINGH, D.; PADWAD, Y. S. Synergistic
effect of curcumin and piperine in suppression of DENA-induced hepatocellular carcinoma in
rats. Environmental Toxicology and Pharmacology, v. 40, p. 445-452, 2015.
PETAN, T.; JARC, E.; JUSOVIĆ, M. Lipid Droplets in Cancer: Guardians of Fat in a Stressful
World. Molecules, v. 23, pii: E1941, 2018.
PITT, J. J. Principles and applications of liquid chromatography-mass spectrometry in clinical
biochemistry. Clinical Biochemistry Reviews, v. 30, p. 19-34, 2009.
PLUCHINO, K. M.; HALL, M. D.; GOLDSBOROUGH, A. S.; CALLAGHAN, R.;
GOTTESMAN, M. M. Collateral sensitivity as a strategy against cancer multidrug resistance.
Drug Resistance Updates, v. 15, p. 98-105, 2012.
POZO-GUISADO, E.; ALVAREZ-BARRIENTOS, A.; MULERO-NAVARRO, S.;
SANTIAGO-JOSEFAT, B.; FERNADEZ-SALGUERO, P. M. The antiproliferative activity of
resveratrol results in apoptosis in MCF-7 but not MDA-MB231 in human breast cancer cells:
cell-specific alteration of the cell cycle. Biochemical Pharmacology, v. 64, p. 1357-1366, 2002.
PRADEEP, K. U.; GEERVANI, P.; EGGUM, B. O. Common Indian spices: Nutrient
composition, consumption and contribution to dietary value. Plant Foods for Human Nutrition,
v. 44, p. 137-148, 1993.
QUARTI, J. Efeito da piperina na indução de sensibilidade colateral e na reversão da resistência à
vincristina em células de leucemia resistentes a quimioterápicos. Dissertação (Mestrado em
Nutrição) – Instituto de Nutrição Josué de Castro, Universidade Federal do Rio de Janeiro, Rio de
Janeiro, 2015.
RAJAWAT, J.; SHUKLA, N.; MISHRA, D. P. Therapeutic Targeting of Poly(ADP-Ribose)
Polymerase-1 (PARP1) in Cancer: Current Developments, Therapeutic Strategies, and Future
Opportunities. Medicinal Research Reviews, v. 37, p. 1461-1491, 2017.

120
RATHER, R. A.; BHAGAT, M. Cancer Chemoprevention and Piperine: Molecular Mechanisms
and Therapeutic Opportunities.Frontiers in Cell and Developmental Biology, v. 6, p. 10, 2018.
RAVANAT, J. L.; DOUKI, T.; CADET, J. Direct and indirect effects of UV radiation on DNA
and its components. Photochemistry Photobiology, v. 63, p. 88-102, 2001.
RAY-CHAUDHURI, A.; NUSSENZWEIG, A. The multifaceted roles of PARP1 in DNA repair
and chromatin remodelling. Nature Review Molecular Cell Biology, v. 18, p.610-621, 2017.
REEN, R. K.; WIEBEL, F. J.; SINGH, J. Piperine inhibits aflatoxin B-1-induced cytotoxicity and
genotoxicity in V79 Chinese hamster cells genetically engineered to express rat cytochrome
P4502B1. Journal of Ethnopharmacology, v. 58, p. 165-173, 1997.
RODRIGUEZ-ANTONA, C.; INGELMAN-SUNDBERG, M. Cytochrome P450
pharmacogenetics and cancer. Oncogene, v. 25, p. 1679-1691, 2006.
ROTTENBERG, S.; JASPERS, J. E.; KERSBERGEN, A.; VAN DER BURG, E.; NYGREN, A.
O.; ZANDER, S. A.; DERKSEN, P. W.; DE BRUIN, M.; ZEVENHOVEN, J.; LAU, A.;
BOULTER, R.; CRANSTON, A.; O'CONNOR, M. J.; MARTIN, N. M.; BORST, P.; JONKERS,
J. High sensitivity of BRCA1-deficient mammary tumors to the PARP inhibitor AZD2281 alone
and in combination with platinum drugs. Proceedings of the National Academy of Sciences, v.
105, p. 17079-17084, 2008.
RUMJANEK, V. M.; TRINDADE, G. S.; WAGNER-SOUZA, K.; DE-OLIVEIRA, M. C.;
MARQUES-SANTOS, L. F.; MAIA, R. C.; CAPELLA, M. A. Multidrug resistance in tumour
cells: characterization of the multidrug resistant cell line K562-Lucena 1. Anais da Academia
Brasileira de Ciências, v. 73, p. 57-69, 2001.
RUMJANEK, V. M.; VIDAL, R. S.; MAIA, R. C. Multidrug resistance in chronic myeloid
leukaemia: how much can we learn from MDR-CML cell lines? Bioscience Reports, v. 33, p. 6.
pii: e00081, 2013.

121
SABA, N.; BHUYAN, R.; NANDY, S. K.; SEAL A. Differential Interactions of Cytochrome
P450 3A5 and 3A4 with Chemotherapeutic Agent-Vincristine: A Comparative Molecular
Dynamics Study. Anti-cancer Agents in Medicinal Chemistry, v. 15, p. 475-483, 2015.
SAHU, P. K.; SHARMA, A.; RAYEES, S.; KOUR, G.; SINGH, A.; KHULLAR,M.;
MAGOTRA, A.; PASWAN, S. K.; GUPTA, M.; AHMAD, I.; ROY, S.; TIKOO, M. K.;
SHARMA, S. C.; SINGH, S.; SINGH, G. Pharmacokinetic study of Piperine in wistar rats after
oral and intravenous administration. International Journal of Drug Delivery, v. 6, p. 82-87,
2014.
SALAZAR-ROA, M.; MALUMBRES, M. Fueling the Cell Division Cycle. Trends in Cell
Biology, v. 27, p. 69-81, 2017.
SAMY, R. P.; GOPALAKRISHNAKONE, P. Therapeutic potential of plants as anti-microbials
for drug discovery. Evidence-Based Complementary and Alternative Medicine, v. 3, p. 283-
294, 2010.
SCHARFFETTER-KOCHANEK, K.; BRENNEISEN, P.; WENK, J.; HERMANN, G.; KUHR,
M. A.; KUHR, L.; MEEWES, C.; WLASCHEK, M. Photoaging of the skin from phenotype to
mechanisms. Experimental Gerontology, v. 35, p. 307-316, 2000.
SCHWEICHEL, J. U.; MERKER, H. J. The morphology of various types of cell death in prenatal
tissues. Teratology, v. 7, p. 253-66, 1973.
SELVENDIRAN, K.; BANU, S. M.; SAKTHISEKARAN, D. Protective effect of piperine on
benzo(a)pyrene-induced lung carcinogenesis in swiss albino mice. Clinica Chimica Acta, v.
350, p. 73-78, 2004b.
SELVENDIRAN, K.; SENTHILNATHAN, P.; MAGESH, V.; SAKTHISEKARAN, D.
Modularory effect of piperine on mitochondrial antioxidant system in benzo(a)pyrene- induced
experimental drug carcinogenesis. Phytomedicine, v. 14, p. 85-89, 2004a.

122
SELVENDIRAN, K.; THIRUNAVUKKARASU, C.; SINGH, J. P.; PADMAVATHI, R.;
SAKTHISEKARAN, D. Chemopreventive effect of piperine on mitochondrial TCA cycle and
phase-I and glutathione-metabolizing enzymes in benzo(a)pyrene induced lung carcinogenesis in
Swiss albino mice. Molecular and Cellular Biochemistry, v. 271, p. 101-106, 2005.
SEN, N.; DAS, B. B.; GANGULY, A.; MUKHERJEE, T.; TRIPATHI, G.;
BANDYOPADHYAY, S.; RAKSHIT, S.; SEN, T.; MAJUMDER, H. K. Camptothecin induced
mitochondrial dysfunction leading to programmed cell death in unicellular hemoflagellate
Leishmania donovani. Cell Death & Differentiation, v. 11, p. 924-936, 2004.
SHAMSI, S.; TRAN, H.; TAN, R. S.; TAN, Z. J.; LIM, L. Y. Curcumin, Piperine, and Capsaicin:
A Comparative Study of Spice-Mediated Inhibition of Human Cytochrome P450 Isozyme
Activities. Drug Metabolism & Disposition, v. 45, p. 49-55, 2017.
SHAO, B.; CUI, C.; JI, H.; TANG, J.; WANG, Z.; LIU, H.; QIN, M.; LI, X.; WU, L. Enhanced
oral bioavailability of piperine by self-emulsifying drug delivery systems: in vitro, in vivo and in
situ intestinal permeability studies. Drug Delivery, v. 22, p. 740-747, 2015.
SHAROM, F. J. Complex Interplay between the P-Glycoprotein Multidrug Efflux Pump and the
Membrane: Its Role in Modulating Protein Function. Frontiers in Oncology, v. 4, p. 41, 2014.
SHI, Z.; LIANG, Y. J.; CHEN, Z. S.; WANG, X. H.; DING, Y.; CHEN, L. M.; FU, L. W.
Overexpression of Survivin and XIAP in MDR cancer cells unrelated to P-glycoprotein.
Oncology Reports, v. 17, p. 969-976, 2007.
SHIBUE, T., WEINBERG, R. A. EMT, CSCs, and drug resistance: the mechanistic link and
clinical implications. Nature Reviews Clinical Oncololy, v. 14, p. 611-629, 2017.
SI, L.; YANG, R.; LIN, R.; YANG, S. Piperine functions as a tumor suppressor for human
ovarian tumor growth via activation of JNK/p38 MAPK-mediated intrinsic apoptotic pathway,
Bioscience Reports, v. 38, pii: BSR20180503, 2018.

123
SIDDIQUI, S.; AHAMAD, M. S.; JAFRI, A.; AFZAL, M.; ARSHAD, M. Piperine triggers
apoptosis of human oral squamous carcinoma through cell cycle arrest and mitochondrial
oxidative stress. Nutrition and Cancer, v. 69, p. 791-799, 2017.
SIEMANN, D. W.; HORSMAN, M. R. Modulation of the tumor vasculature and oxygenation to
improve therapy. Pharmacology & Therapeutics, v. 153, p. 107-124, 2015.
SILVA, K. L.; DE SOUZA, P. S.; NESTAL DE MORAES, G.; MOELLMANN-COELHO, A.;
VASCONCELOS FDA, C.; MAIA, R. C. XIAP and P-glycoprotein co-expression is related to
imatinib resistance in chronic myeloid leukemia cells. Leukemia Research, v. 37, p. 1350-1358,
2013.
SINGH, A.; RAO, A. R. Evaluation of the modulatory influence of black pepper (Piper nigrum,
L.) on the hepatic detoxication system. Cancer Letters, v. 72, p. 5-9, 1993.
SINGH, D. V.; GODBOLE, M. M.; MISRA, K. A plausible explanation for enhanced
bioavailability of P-gp substrates in presence of piperine: simulation for next generation of P-gp
inhibitors. Journal of Molecular Modeling, v. 19, p. 227-238, 2013.
SINGH, V.K.; COUMAR, M.S. Chronic myeloid leukemia: Existing therapeutic options and
strategies to overcome drug resistance. Mini-Reviews in Medical Chemistry, v. 19, p. 333-345,
2018.
SINHA, K.; DAS, J.; PAL, P. B.; SIL, P. C. Oxidative stress: the mitochondria-dependent and
mitochondria-independent pathways of apoptosis. Archives of Toxicology, v. 87, p. 1157-1180,
2013.
SOARES, F. S.; LETTNIN, A. P.; WAGNER, E. F.; MATTOZO, F. H.; CARRETT-DIAS, M.;
RUMJANEK, V. M. B. D.; FILGUEIRA, D. M. V. B.; DE SOUZA VOTTO, A. P. Multidrug
resistance phenotype: Relation between phenotype induction and its characteristics in
erythroleukemia cells. Cell Biology International, v. 43, p. 214-219, 2019.

124
SODHI, R. K.; SINGH, N.; JAGGI, A. S. Poly (ADP-ribose) polymerase-1 (PARP-1) and its
therapeutic implications. Vascular Pharmacology, v. 53, p. 77-87, 2010.
SOLDANI, C.; SCOVASSI, A. I. Poly (ADP-ribose) polymerase-1 cleavage during apoptosis: an
update. Apoptosis, v. 7, p. 321-328, 2002.
SONG, Q. F.; QU, Y. C.; ZHENG, H. B.; ZHANG, G. H.; LIN, H. G.; YANG, J. L.
Differentiation of erythroleukemia K562 cells induced by piperine. Chinese Journal of Cancer,
v. 27, p. 571-574, 2008.
SRIVASTAVA, A. K.; SINGH, V. K. Biological action of Piper nigrum - the king of spices.
European Journal of Biological Research, v. 7, p. 223-233, 2017.
STRAUB, B. K.; HERPEL, E.; SINGER, S.; ZIMBELMANN, R.; BREUHAHN, K.; MACHER-
GOEPPINGER, S.; WARTH, A.; LEHMANN-KOCH, J.; LONGERICH, T.; HEID, H.; Lipid
droplet-associated PAT-proteins show frequent and differential expression in neoplastic
steatogenesis. Modern Pathology, v. 23, p. 480-492, 2010.
SUNDELIN, J. P.; STÅHLMAN, M.; LUNDQVIST, A.; LEVIN, M.; PARINI, P.;
JOHANSSON, M. E.; BORÉN, J. Increased expression of the very low-density lipoprotein
receptor mediates lipid accumulation in clear-cell renal cell carcinoma. PLoS ONE, v. 7, e48694,
2012.
SURH, Y. L. Cancer chemoprevention with dietary phytochemicals. Nature, v. 3, p. 768-780,
2003.
SWANSON, H. I.; NJAR, V. C.; YU, Z.; CASTRO, D. J.; GONZALEZ, F. J.; WILLIAMS, D.
E.; HUANG, Y.; KONG, A. N.; DOLOFF, J. C.; MA, J.; WAXMAN, D. J.; SCOTT, E. E.
Targeting drug-metabolizing enzymes for effective chemoprevention and chemotherapy. Drug
Metabolism and Disposition, v. 38, p. 539-544, 2010.

125
TAIT, S. W.; ICHIM, G.; GREEN, D. R. Die another way--non-apoptotic mechanisms of cell
death. Journal of Cell Science, v. 127, p. 2135-2144, 2014.
TAK, J. K.; LEE, J. H.; PARK, J. W. Resveratrol and piperine enhance radiosensitivity of tumor
cells. BMB Reports, v. 45, p. 242-246, 2012.
TALIB, W. H. Regressions of breast carcinoma syngraft following treatment with piperine in
combination with thymoquinone. Scientia Pharmaceutica, v. 85, E27, 2017.
TAUCHI-SATO, K.; OZEKI, S.; HOUJOU, T.; TAGUCHI, R.; FUJIMOTO, T. The surface of
lipid droplets is a phospholipid monolayer with a unique Fatty Acid composition. The Journal of
Biological Chemistry, v. 277, p. 44507-44512, 2002.
TOWBIN, H.; STAEHELIN, T.; GORDON, J. Electrophoretic transfer of proteins from
polyacrilamide gels to nitrocellulose sheets: procedure and some applications. Proceedings of
the National Academy of Sciences, v. 76, p. 4350-4354, 1979.
TRINDADE, G. S.; CAPELLA, M. A.; CAPELLA, L.; AFFONSO-MITIDIERI, O. R.;
RUMJANEK, V. M. Differences in sensitivity to UVC, UVB and UVA radiation of a multidrug-
resistant cell line overexpressing P-glycoprotein. Photochemistry Photobiology, v. 69, p. 694-
699, 1999.
TROTT, O., OLSON, A. J. AutoDock Vina: improving the speed and accuracy of docking with a
new scoring function, efficient optimization and multithreading. Journal of Computacional
Chemistry, v. 31, p. 445-461, 2010.
VELLAICHAMY, L.; BALAKRISHNAN, S.; PANJAMURTHY, K.; MANOHARAN, S.;
ALIAS, L. M. Chemopreventive potential of piperine in 7,12-dimethylbenz[a]anthracene-induced
skin carcinogenesis in Swiss albino mice. Environmental Toxicology and Pharmacology, v.
28, p. 11-18, 2009.

126
VERBRUGGE, S. E.; AL, M.; ASSARAF, Y. G.; KAMMERER, S.; CHANDRUPATLA, D.
M.; HONEYWELL, R.; MUSTERS, R. P.; GIOVANNETTI, E.; O'TOOLE, T.; SCHEFFER, G.
L.; KRIGE, D.; DE GRUIJL, T. D.; NIESSEN, H. W.; LEMS, W. F.; KRAMER, P.
A.; SCHEPER, R. J.; CLOOS, J.; OSSENKOPPELE, G. J.; PETERS, G. J.; JANSEN, G.
Multifactorial resistance to aminopeptidase inhibitor prodrug CHR2863 in
myeloid leukemia cells: down-regulation of carboxylesterase 1, drug sequestration in lipid
droplets and pro-survival activation ERK/Akt/mTOR. Oncotarget, v. 7, p. 5240-5257, 2016.
VERMEULEN, K.; VAN BOCKSTAELE, D. R.; BERNEMAN, Z. N. The cell cycle: a review
of regulation, deregulation and therapeutic targets in cancer. Cell Proliferation, v. 36, p. 131-
149, 2003.
VIDAL, R. S. Alvos Terapêuticos no Fenótipo de Resistência a Múltiplas Drogas. Tese
(Doutorado em Química Biológica) – Instituto de Bioquímica Médica, Universidade Federal do
Rio de Janeiro, Rio de Janeiro, 2017.
VIDAL, R. S.; QUARTI, J.; RODRIGUES, M. F.; RUMJANEK, F. D.; RUMJANEK, V. M.
Metabolic Reprogramming During Multidrug Resistance in Leukemias. Frontiers in Oncology,
v. 8, p. 90, 2018.
VOLAK, L. P.; HANLEY, M. J.; MASSE, G.; HAZARIKA, S.; HARMATZ, J. S.; BADMAEV,
V.; MAJEED, M.; GREENBLATT, D. J.; COURT, M. H. Effect of a herbal extract containing
curcumin and piperine on midazolam, flurbiprofen and paracetamol (acetaminophen)
pharmacokinetics in healthy volunteers. British Journal of Clinical Pharmacology, v. 75, p.
450-462, 2013.
WANG, T.; CHEN, F. Y; HAN, J. Y; ZHONG, J. H.; TENG, Y.; OUYANG, R. R. The
relationship between cytochrome P450, subfamily IIIA, polypeptide 5 gene and drug resistance in
leukemia cell lines. Zhonghua Nei Ke Za Zhi, v. 43, p. 527-529, 2004.
https://www.ncbi.nlm.nih.gov/pubmed/?term=Assaraf%20YG%5BAuthor%5D&cauthor=true&cauthor_uid=26496029
https://www.ncbi.nlm.nih.gov/pubmed/?term=Kammerer%20S%5BAuthor%5D&cauthor=true&cauthor_uid=26496029
https://www.ncbi.nlm.nih.gov/pubmed/?term=Musters%20RP%5BAuthor%5D&cauthor=true&cauthor_uid=26496029
https://www.ncbi.nlm.nih.gov/pubmed/?term=Niessen%20HW%5BAuthor%5D&cauthor=true&cauthor_uid=26496029

127
WANG, T.; CHEN, F. Y.; GU, C. H.; ZHONG, H.; TENG, Y.; OUYANG, R. R. Transfection of
HL 60 cells with CYP3A5 gene induces drug-resistant phenotype. Zhonghua Nei Ke Za Zhi, v.
27, p. 461-464, 2005.
WANG, Y. M.; LIN, W.; CHAI, S. C.; WU. J.; ONG, S. S.; SCHUETZ, E. G.; CHEN, T.
Piperine activates human pregnane X receptor to induce the expression of cytochrome P450 3A4
and multidrug resistance protein 1. Toxicology and Applied Pharmacology, v. 272, p. 96-107,
2013.
WELTE, M. A.; GOULD, A. P. Lipid droplet functions beyond energy storage. Biochimica et
Biophysica Acta, v. 1862, p. 1260-1272, 2017.
WESIERSKA-GADEK, J. Major Contribution of the Multidrug Transporter P-Glycoprotein to
Reduced Susceptibility of Poly (ADP-Ribose) Polymerase-1 Knock-Out Cells to Doxorubicin
Action. Journal of Cellular Biochemistry, v. 95, p. 1012-1028, 2005.
WU, J.; XIAO, S.; YUAN, M.; LI, Q.; XIAO, G.; WU, W.; OUYANG, Y.; HUANG, L.; YAO,
C. PARP inhibitor re-sensitizes Adriamycin resistant leukemia cells through DNA damage and
apoptosis. Molecular Medicine Reports, v. 19, p. 75-84, 2019.
WURZER, G.; HERCEG, Z.; WESIERSKA-GADEK, J. Increased Resistance to Anticancer
Therapy of Mouse Cells Lacking the Poly(ADP-ribose) Polymerase Attributable to Up-
Regulation of the Multidrug Resistance Gene Product P Glycoprotein. Cancer Research, v. 60,
p. 4238-4244, 2000.
XU, X.; LAI, Y.; HUA, Z. C. Apoptosis and apoptotic body: disease message and therapeutic
target potentials. Bioscience Reports, v. 39, BRS20180992, 2019.
YAFFE, P. B.; DOUCETTE, C. D.; WALSH, M.; HOSKIN, D. W. Piperine impairs cell cycle
progression and causes reactive oxygen species-dependent apoptosis in rectal cancer cells.
Experimental and Molecular Pathology, v. 94, p. 109-114, 2013.

128
YAFFE, P. B.; POWER COOMBS, M. R.; DOUCETTE, C. D.; WALSH, M.; HOSKIN, D. W.
Piperine, an alkaloid from black pepper, inhibits growth of human colon cancer cells via G1
arrest and apoptosis triggered by endoplasmic reticulum stress. Molecular Carcinogenesis, v.
54, p. 1070-1085, 2015.
YASHIN, A.; YASHIN, Y.; XIA, X.; NEMZER, B. Antioxidant Activity of Spices and Their
Impact on Human Health: A Review. Antioxidants (Basel), v. 6, pii: E70, 2016.
YU, S. W.; WANG, H.; POITRAS, M. F.; COOMBS, C.; BOWERS, W. J.; FEDEROFF, H. J.
Mediation of poly (ADP-ribose) polymerase-1-dependent cell death by apoptosis-inducing factor.
Science, v. 297, p. 259-263, 2002.
YUNG, T. M. C., SATOH, M. S. Functional Competition between Poly (ADP-ribose)
Polymerase and Its 24-kDa Apoptotic Fragment in DNA Repair and Transcription. The Journal
of Biological Chemistry, v. 276, p. 11279-11286, 2001.
ZABELA, V.; HETTICH, T.; SCHLOTTERBECK, G.; WIMMER, L.; MIHOVILOVIC, M. D.;
GUILLET, F.; BOUAITA, B.; SHEVCHENKO, B.; HAMBURGER, M.; OUFIR, M. GABAA
receptor activity modulating piperine analogs: In vitro metabolic stability, metabolite
identification, CYP450 reaction phenotyping, and protein binding. Journal of Chromatography
B - Analytical Technologies in the Biomedical and Life Science, v. 1072, p. 379-389, 2018.
ZANGER, U. M.; SCHWAB, M. Cytochrome P450 enzymes in drug metabolism: Regulation of
gene expression, enzyme activities, and impact of genetic variation. Pharmacology &
Therapeutics, v. 138, p. 103-141, 2013.
ZHANG, J.; ZHU, X.; LI, H.; LI, B.; SUN, L.; XIE, T.; ZHU, T.; ZHOU, H.; YE, Z. Piperine
inhibits proliferation of human osteosarcoma cells via G2/M phase arrest and metastasis by
suppressing MMP-2/-9 expression. International Immunopharmacology, v. 24, p. 50-58, 2015.

129
ZHANG, X.; LI, X.; YOU, Q. Prodrug strategy for cancer cell-specific targeting: A recent
overview. European Journal of Medicinal Chemistry, v. 139, p. 542-563, 2017.
ZHENG, L.; WANG, C.; LUO, T.; LU, B.; MA, H.; ZHOU, Z.; ZHU, D.; CHI, G.; GE, P.; LUO,
Y. JNK Activation Contributes to Oxidative Stress-Induced Parthanatos in Glioma Cells via
Increase of Intracellular ROS Production. Molecular Neurobiology, v. 54, p. 3492-3505, 2017.
ZHONG FENG, G. J.; LI JING, Z. L.; LIAO JINGLI, Y. F. Pharmacokinetics of piperine from
Piperlongum in rats. Pharmacology and Clinics of Chinese Materia Medica, 2010.
ZHOU, T.; MEDEIROS, L. J; HU, S. Chronic Myeloid Leukemia: Beyond BCR-ABL1. Current
Hematologic Malignancy Reports, v. 13, p. 435-445, 2018.
ZINZI, L.; CAPPARELLI, E.; CANTORE, M.; CONTINO, M.; LEOPOLDO, M.; COLABUFO,
N. A. Small and Innovative Molecules as New Strategy to Revert MDR. Frontiers in Oncology,
v. 4, p. 2, 2014.
ZU, Y.; YANG, Z.; TANG, S.; HAN, Y.; MA, J. Effects of P-glycoprotein and its inhibitors
on apoptosis in K562 cells. Molecules, v. 19, p. 13061-13075, 2014.
ZWEYTICK, D.; ATHENSTAEDT, K.; DAUM, G. Intracellular lipid particles of eukaryotic
cells. Biochimica et Biophysica Acta, v. 1469, p. 101-120, 2000.

130
9. ANEXOS
Anexo A
0,0 2,5 5,0 7,5 10,0 min0
50
100
150
200
250
300
350
400mAU
RPMI + PIP λ= 340 nm
Cromatograma do meio de cultura incubado com piperina por 48 horas.

131
Anexo B
0,0 2,5 5,0 7,5 10,0 min
0
10
20
30
40
50mAU
0,0 2,5 5,0 7,5 10,0 min0
10
20
30
40
50mAU
0,0 2,5 5,0 7,5 10,0 min
0
10
20
30
40
50mAU
FEPS + PIP
K562 + PIP
Lucena-1 + PIP
A
B
C
λ= 340 nm
λ= 340 nm
λ= 340 nm
Cromatogramas da piperina no meio intracelular das células K562, Lucena-1 e FEPS por 48
horas de incubação.

132
Anexo C
0,0 2,5 5,0 7,5 10,0 min0
50
100
150
200
250
300
350
400mAU
0,0 2,5 5,0 7,5 10,0 min0
50
100
150
200
250
300
350
400 mAU
λ= 340 nm
λ= 340 nm
0,0 2,5 5,0 7,5 10,0 min0
50
100
150
200
250
300
350
400mAU
λ= 340 nmFEPS + PIP
K562 + PIP
Lucena-1 + PIP
A
B
C
Cromatogramas da piperina no meio extracelular das células K562, Lucena-1 e FEPS por 48
horas de incubação.

133
Anexo D

April 2018 | Volume 8 | Article 901
HypotHesis and tHeorypublished: 04 April 2018
doi: 10.3389/fonc.2018.00090
Frontiers in Oncology | www.frontiersin.org
Edited by: Michael Breitenbach,
University of Salzburg, Austria
Reviewed by: Johannes A. Mayr,
Paracelsus Medical University Salzburg, Austria
Hamid Morjani, Université de Reims Champagne
Ardenne, France Carlo Vito Bruschi,
CVBiotechnologies, E.U., Austria
*Correspondence:Vivian M. Rumjanek
Specialty section: This article was submitted to
Molecular and Cellular Oncology, a section of the journal
Frontiers in Oncology
Received: 10 January 2018Accepted: 15 March 2018
Published: 04 April 2018
Citation: Vidal RS, Quarti J, Rumjanek FD and
Rumjanek VM (2018) Metabolic Reprogramming During Multidrug
Resistance in Leukemias. Front. Oncol. 8:90.
doi: 10.3389/fonc.2018.00090
Metabolic reprogramming during Multidrug resistance in LeukemiasRaphael Silveira Vidal1, Julia Quarti1,2, Franklin D. Rumjanek1 and Vivian M. Rumjanek1*
1Instituto de Bioquímica Médica Leopoldo de Meis, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil, 2Instituto de Nutrição Josué de Castro, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil
Cancer outcome has improved since introduction of target therapy. However, treatment success is still impaired by the same drug resistance mechanism of classical chemo-therapy, known as multidrug resistance (MDR) phenotype. This phenotype promotes resistance to drugs with different structures and mechanism of action. Recent reports have shown that resistance acquisition is coupled to metabolic reprogramming. High-gene expression, increase of active transport, and conservation of redox status are one of the few examples that increase energy and substrate demands. It is not clear if the role of this metabolic shift in the MDR phenotype is related to its maintenance or to its induction. Apart from the nature of this relation, the metabolism may represent a new target to avoid or to block the mechanism that has been impairing treatment success. In this mini-review, we discuss the relation between metabolism and MDR resistance focusing on the multiple non-metabolic functions that enzymes of the glycolytic pathway are known to display, with emphasis with the diverse activities of glyceraldehyde-3- phosphate dehydrogenase.
Keywords: multidrug resistance, glycolysis, glyceraldehyde-3-phosphate dehydrogenase, leukemia, reactive oxygen species
BaCKGroUnd
Multidrug resistance (MDR) in cancer is the major factor impairing the success of conventional chemotherapy (1). Originally, many hopes were placed on the possibility that by inhibiting the activity of ABC transporters a reversal of the resistance would be attained. This was based on the knowledge that the transporters were shown to be capable of mediating the efflux of many chemo-therapeutic drugs. Among the efflux transporters of the ABC superfamily, ABCB1 (P-gp), ABCC1 (MRP1), and ABCG2 (BCRP or MXR) have been described as major players in the development of MDR (2–4), with particular emphasis being given to ABCB1. However, it soon became apparent that inhibition of these transporters was not an effective approach since normal cells may also express these transporters and, therefore, inhibitors could sometimes generate an unacceptable toxicity. Furthermore, in experimental situations, where the down regulation of ABCB1 was achieved, a number of other effects were still observed. Now, it is quite clear that MDR is a multifactorial phenomenon that is involved in the regulation of survival and apoptosis, as well as a number of other cellular pathways. ABCB1 transporter expression, in cells with the MDR phenotype, is but one factor linked to pharmacological evasion of chemotherapeutic drugs (1, 5).
A case in point is the observation that hypoxia participates in the regulation of drug resistance. For instance, ABCB1 gene expression and synthesis of functional proteins are induced by hypoxic environments (6). Furthermore, ABC transporters are expressed not only in MDR cancer cells, but

2
Vidal et al. MDR Cells and Metabolic Reprogramming
Frontiers in Oncology | www.frontiersin.org April 2018 | Volume 8 | Article 90
also in a number of stem and progenitor cells. Additionally, it has been reported that hypoxia promotes an undifferentiated cell state in various stem and precursor cell populations, as well as in cancer stem cells (7–9). In this respect, it has also been suggested that NOTCH signaling is involved. However, it must be recalled that when cells are under hypoxic conditions, there is a metabolic shift from oxidative phosphorylation to glycolysis (10). This situ-ation contrasts with cells under normoxia, in which glucose is first anaerobically catabolized to pyruvate which is then further catabolized along the Krebs cycle where NADH and FADH2 are reoxidized by the respiratory chain associated to the electron transport system. Incidentally, glycolysis is a hallmark in many types of tumor cells (11). This phenotype is in fact the basis of the so-called Warburg effect, also known as aerobic glycolysis. The Warburg effect describes a situation in which the glycolytic pathway is fully activated even in the presence of adequate oxygen supply (12). Although Warburg originally proposed that cancer was due to an impairment of mitochondrial function, it is accepted today that these organelles retain full oxidative capacities.
It must be mentioned, however, that apart from red blood cells, aerobic glycolysis is prevalent in highly proliferative cells, whether tumoral or not. Stem cells are a case in point (13). The common belief that cells undergoing glycolysis selected an inefficient form of energy production is misguided. Barring the comparative stoi-chiometry of ATP formation between glycolysis and OXPHOS, aerobic glycolysis is in fact an efficient form of ATP production due to the kinetic properties of the enzymes participating in the pathway which afford very fast fluxes compatible with the ATP demand of the rapidly growing cells. Beyond its role in bioener-getics, glycolysis constitutes a branch of the pentose phosphate pathway (PPP), since glucose-6-phosphate is also the substrate for glucose-6-phosphate dehydrogenase, the first enzyme of that pathway. Thus, glycolysis also contributes to the production of precursors for the biosynthesis of nucleotides (via generation of ribulose-5-phosphate). In addition, the PPP pathway promotes the formation of NADPH, an essential coenzyme for reductive biosynthetic processes such as that of fatty acids. NADPH also has an important role in maintaining the redox equilibrium. Similarly, glycolysis can be considered as an anaplerotic pathway by way of its participation in amino acids synthesis (via 3-phosphoglycerate or via pyruvate). Thus, from an energetic stand point glycolysis more than compensates the relatively small amounts of ATP produced when compared with oxidative phosphorylation.
However, it must be emphasized that tumors are in fact con-stituted by a mosaic of different cellular subpopulations. As such, from the biochemical point of view tumors can also be envis-aged as being functionally heterogeneous. Accordingly, within the context of types of metabolism, tumors can be perceived as composed of subsets of resistant quiescent/slow-cycling cells that occasionally rely more on mitochondrial respiration and less on glycolysis. Likewise the same tumor could also harbor cells that are exclusively glycolytic (14, 15). Interestingly, the possibility of a switch that regulates mitochondrial function in the case of metas-tasis has been proposed. The results of Porporato et al. showed that overburdening the electron transport system may be an essential step in enhancing migration of cells in vitro and in vivo (16). The authors concluded that in order to achieve metastasis,
mitochondria must be active, although not necessarily functional. By extension such findings suggest that in tumor cells there may be switches that constantly activate/inactivate mitochondrial function depending on changes dictated by the microenviron-ment that affect, for example, the availability of metabolites. The intermittent switching between anaerobic and oxidative metabo-lism seems to be a feature of metastasis. According to this scheme, accumulating data show that there is a tradeoff involving growth versus migration, i.e., cells which are proliferating prevalently exhibit a glycolytic type of metabolism, whereas migrating cells which proliferate less, rely more on mitochondria (16). Within this framework, it is known that the switch between the two main types of energy metabolism may be regulated by ATP demand. For example, cells expressing ABC transporters on their surfaces require a considerable amount of ATP in order to sustain the drug efflux activity. Additionally, it has been suggested that the transporter activity might suffer the impact of alterations in pH gradient due to the glycolytic phenotype (17). Normal cells maintain a gradient between acidic vesicular compartments and an alkaline cytoplasm a situation not observed in tumor cells that have an acidic cytoplasm. Nevertheless, it has been observed that MCF-7adr, that has an MDR phenotype, presents a similar pH gradient to that of normal cells (18, 19). This gradient contributes to the sequestration of drugs in acidic organelles and subsequent extrusion from the cell (19).
The mitochondrial electron transport chain generates reactive oxygen species (ROS) (20). In some instances, such as partial disintegration of complex I (21–23), oxidative stress may result from excessive production of ROS which is buffered by redox homeostasis. In turn, homeostasis is achieved by the participation of a number of enzymes such as catalase, superoxide dismutase, gluthatione peroxidase, and cofactors such as reduced glutha-tione and NADPH. Control of redox equilibrium is important since an imbalance between the amount of ROS produced and of antioxidant systems may lead to DNA damage, particularly mitochondrial DNA, and other cell lesions.
Notwithstanding, the effect of ROS as agents of oxidative stress affecting the expression of ABCB1 is controversial. Both, downregulation and upregulation of the transporter have been reported (24, 25). Antioxidants (26), as well as products resulting from glycolysis can act as scavengers of free radicals. In this way, the energy metabolism could also play an indirect role as modula-tors of ABCB1 expression. For instance, it is known that under conditions leading to glycolysis inhibition, ABCB1 expression was observed to be downregulated. Conversely, when exogenous pyruvate was added to the tumor cells there was increased drug resistance and transporter expression (25).
Therefore, there is consensus that under normal conditions, glycolysis, and mitochondrial oxidative phosphorilation operate in concert in so far as energy production in the form of ATP is concerned.
Originally, glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was thought to function exclusively as part of the gly-colytic process in the cytoplasm, where its role is well established. However, under conditions of oxidative stress GAPDH may also redirect glyceraldehyde-3-P to glucose 6 phosphate (G6P) as a result from the reversal of part of the glycolytic reactions.

3
Vidal et al. MDR Cells and Metabolic Reprogramming
Frontiers in Oncology | www.frontiersin.org April 2018 | Volume 8 | Article 90
The formed G6P then becomes the initial substrate of the PPP leading to the increase of NADPH production (27). GAPDH is particularly sensitive to H2O2-induced oxidation and it has been suggested that cytosolic GAPDH might function as a sensor for redox signals and an information hub to transduce these signals (28). Oxidative stress may also promote GAPDH aggregation leading to mitochondrial dysfunction and necrotic cell death via the permeability transition pore (29). Interestingly, under some circumstances GAPDH was shown to bind to band 3 protein, an anion transporter located on the inner side of the red blood cell membrane. In this membrane bound state, GAPDH along with other glycolytic enzymes contributed toward ATP channeling thus allowing its direct consumption by ion pumps without release into the cytoplasm (30). Such a role for GAPDH would be in keeping with an accessory function within the context of drug efflux. GAPDH, however, plays a number of other roles in different cell compartments, where several pools of GAPDH sense cellular stresses and activate cognate pathways to maintain homeostasis or activate cell death (31). It has also been reported that following exposure to stressors GAPDH translocate into the nucleus (32) where it may suffer ADP-ribosylation by NO (33). Apart from GAPDH, other enzymes of the glycolytic pathway are known to display multiple non-metabolic functions. In fact, of the 10 enzymes that constitute glycolysis, at least 7 have been shown to display extra-glycolytic activities that may bear on the MDR phenotype (34). For example, hexokinase II (HKII) binds to the mitochondrial voltage-gated anion channels and thus relieves the negative feedback effect of G6P (35). HKII overexpression is regulated by HIF-1α and also by c-Myc oncogene. HKII is also under the control of many miRNAs. When phosphohexosei-somerase, or phosphoglucoisomerase (PGI) is secreted by cells it acquires the status of a cytokine and is renamed as the autocrine motility factor (AMF). As such, AMF stimulates cell motility and so PGI is thought to be one of the factors driving metastasis (36). Besides, PGI is involved in many other activities such as apoptosis and EMT. Other glycolytic enzymes such as phosphofructokinase 1, aldolase and triose phosphate isomerase, phosphoglycerate mutase, and pyruvate kinase have all been reported to take part in several cellular functions that have a direct relation to tumorigen-esis. Hence, it is entirely plausible that individually or collectively, the glycolytic enzymes and in particular GAPDH may constitute integral parts of the MDR phenotype by acting in a non-canonical fashion (37).
Alternatively, the metabolic rewiring of tumor cells may also result from gene rearrangement. Using yeast as a model system, genomic instability and the reprogramming of central metabolism have been approached (38). Regarding gene rear-rangement, factors that normally contribute toward the integrity of the replicative process may be compromised. Drug resistance has been observed in yeast in which genome translocants were investigated (39). A parallel between yeast and tumor cells could thus be established even though the detailed mechanisms are still not understood. Indeed in MDR cells the over expression of mini chromosome maintenance 7 (MCM7) was detected (40).
Oxidative stress induces severe damage to proteins, lipids, and DNA. Normally, the generation of oxyradicals is prevented by the mitochondrial antioxidant system; however, the degree of
damage will also depend on the repair capacity of the cell. DNA damage leads to the activation of Poly(ADP-ribose) polymerases or PARPs that are nuclear enzymes responsible for catalyzing the attachment of ADP-ribose units from nicotinamide adenine dinucleotide (NAD+) to acceptor proteins involved in the recogni-tion and repair of DNA strands breaks. After repair is completed the Poly(ADP-ribose) (PAR) chains are degraded (41). During caspase mediated apoptosis PARP is cleaved into fragments that in turn inactivate the enzyme inhibiting repair to proceed (42, 43). It has been suggested that PARP inhibitors (PARPi) could be used in combination with chemotherapeutic drugs, but PARPi tends to be extruded from the cell by the ABCB1 transporter in MDR tumors (44).
Cell survival and energy metabolism in MDR tumor cells will be discussed in the next section.
Mdr, CHroniC MyeLoid LeUKeMia, and enerGy MetaBoLisM
Multidrug resistant cells may be very heterogeneous in their characteristics and survival pathways. In the present review, we will highlight, including our own data, some aspects related to the energy metabolism of two MDR chronic leukemia cell lines derived from the CML cell line, K562. The two cell lines were selected after exposure to different chemotherapeutic drugs. One cell line was exposed to vincristine, originating Lucena-1 (45), and the other exposed to daunorubicin, originating FEPS (46). Despite arising from the same parental cell, they were quite distinct. Comparative microarray analysis identified 130 differentially expressed genes between K562 versus Lucena-1, 1,932 between K562 versus FEPS, and 1,211 between Lucena-1 versus FEPS. ABCB1 was overexpressed in both MDR cell lines, but highly overexpressed in FEPS, which is the most resistant line (47). Similarly, comparative proteomics of the parental cell line and the MDR counterparts indicated that K562 presented 560 unique proteins, Lucena-1 had 38 and FEPS 63 unique proteins. Lucena-1 and FEPS shared 929 proteins. From the latter, 112 were common only to Lucena-1 and FEPS (48). Results from another survey investigating the proteomic profiles of K562 and Lucena-1, identified 36 differentially expressed proteins between these two cell lines (40). From those, the leucine-rich PPR motif-containing protein and MCM7, as well as the expression of ABCB1 could be used as markers to identify patients that would respond or fail to therapy with the tirosine kinase inhibitor, Imatinib (40). The importance of ABCB1 in the resistance to Imatinib in the clini-cal setting has been demonstrated (49, 50), but it is not the only transporter involved. Tumor cells expressing ABCG2 are capable of Imatinib extrusion with great affinity (51, 52). In a different CML model, comparing the proteomic analysis of another MDR cell line derived from K562 selected with doxorubicin, Qinghong et al. highlighted 44 differentially expressed proteins. Some of the differentially expressed proteins were common to those observed in FEPS (53).
The possibility that overexpression of ABCB1 in these cell lines might reflect stem cell characteristics was analyzed by look-ing at the NOTCH pathway. Notch was overexpressed in FEPS,

FiGUre 1 | High-resolution respirometry of K562, Lucena-1, and FEPS cells. Respiratory parameters of intact K562, Lucena-1, and FEPS cells. Routine respiration—basal respiration of intact cells; leak respiration—rate of oxygen consumption after the addition of oligomycin, that is, uncoupled respiration; coupled respiration (routine–leak); ETS—maximum respiratory capacity (induced by the addition of FCCP); ROX—rate of oxygen consumption in the presence of rotenone and antimycin A, that is, respiration not associated with electron transport system. All the parameters were corrected by ROX values. The bars marked with asterisks denote the values that are significantly different with reference to K562 cells.
4
Vidal et al. MDR Cells and Metabolic Reprogramming
Frontiers in Oncology | www.frontiersin.org April 2018 | Volume 8 | Article 90
the most resistant cell line displaying the highest expression of ABCB1, and slow replication time (47). Another pathway present in both, normal stem cells and CML stem cells is the canonical Wnt pathway. This signaling pathway was shown to be more strongly activated to positively regulate ABCB1 in Lucena-1 cells when compared with the non-MDR K562. However, FEPS was not studied on this occasion (40). Conversely, both MDR cell lines had an increased expression of carbonic anhydrase and hemoglobin (47). This was confirmed by proteomics (48) sug-gesting that these cells are more differentiated compared with the parental erythroleukemic cell line K562 (54). Furthermore, they have the potential to maintain the intracellular pH (17). It has been reported that Imatinib induces erythrocytic differentiation in K562 cells and this is independent of blockade of apoptosis being also observed in resistant clones (55).
One characteristic of the two MDR cell lines described above (Lucena-1 and FEPS) is their antioxidant capacity. The elevated catalase activity observed in Lucena-1 provides these cells with protection against cytotoxic chemicals as well as UV radiation (56). Catalase activity is also elevated in FEPS and these cells also present increased Glucose 6 phosphate dehydrogenase (G6PD) activity (Vidal RS, Faria G, Maia RC, and Rumjanek VM, unpub-lished data). Despite the well-known role in redox homeostasis, G6PD is also involved in cell growth and signaling and this might be an equally important role in resistant cells (57). Many redox changes are now perceived to allow, in a localized compartment, a rapid and physiological signaling event that may regulate the activity of certain proteins (26, 58). Important players in regulating these intracellular effects are members of the thioredoxin family, including thioredoxin that has increased levels in Lucena-1 and FEPS (48), glutaredoxins, and peroxiredoxins. The third most upregulated gene in Lucena-1 when compared with K562 is SESN3 that catalyzes peroxiredoxins leading to ROS detoxifica-tion (47). Furthermore, several members of peroxiredoxins, a family of antioxidant enzymes, are increased in FEPS (PRDX1, PRDX2, PRDX3, and PRDX6) or Lucena-1 (PDRX1) compared with K562 (47, 48). Therefore, ROS generated by the MDR tumor cell lines Lucena-1 and FEPS, are rapidly reduced. In this way, many anticancer drugs that act via generation of oxidative stress become ineffective.
When the MDR cell lines and their parental counterpart, K562, were tested for oxygen consumption measured by high-resolution respirometry, the most resistant cell line, FEPS, repro-ducibly displayed comparatively lower values in all parameters measured (Figure 1). This result suggested that in FEPS, energy was probably being obtained via glycolysis. Gene expression (47) and protein expression (48) indicate an increase in pyruvate kinase levels in FEPS, whereas in Lucena-1 only increased protein expression was observed (48). Differences in the glycolytic path-way, with higher expression of hexoquinase 2, GAPDH and LDH, were observed using SKOV3 TR, an ovarian cell line transfected with the ABCB1 gene (59).
Proteomic analysis of another MDR line obtained by doxoru-bicin selection using K562 as the parental cell, described upregu-lation of fructose-biphosphate aldolase A, fructose-biphosphate aldolase C, transaldolase, and alpha-enolase suggesting that the cells need more energy to survive chemical stress (53). FEPS cells,
selected with daunorubicin also presented upregulated fructose-biphosphate aldolase A (48).
GapdH, Mdr, and CeLL deatH
Among the differences observed in the present study was the variation found in the levels of GAPDH between K562 and its MDR counterparts. Using a proteomic approach (48), it was possible to verify that in relation to K562, the levels of GAPDH were 2.4 times higher in FEPS and 1.2 times in Lucena-1. These results were validated by western blot (Figure 2). Overexpression of GAPDH in Imatinib-resistant cells has been observed by other authors (60, 61). However, these results differ from those found by Cerezo et al. where no difference in GAPDH was observed in daunorubicin resistant cell lines overexpressing ABCB1 (62). Presumably the inconsistency reflects the well-known intrinsic differences encountered in tumor cells of different origin.
Interestingly, a recent report describes a strategy to circumvent the ABCB1 transporter activity by transferring constructs that specifically inhibited GAPDH into target tumor cells by using liposomes. Such treatment was effective in vitro and in vivo (63).
Therefore, the possibility exists that GAPDH plays a role in the resistance observed in some MDR tumors. Various non-glycolytic roles have been assigned to GAPDH (64). Post-translational modifications of GAPDH may dictate subcellular localization and different functions (31, 65). It has been proposed that Sirtuin1 expression retains GAPDH in the cytosol (66). However, no dif-ferences in gene or protein expression of Sirtuin 1, were observed in Lucena-1 or FEPS when compared with their parental cell line K562. As mentioned before, GAPDH translocates to the nucleus where it binds to DNA and participates in a number of DNA-dependent processes (67). However, the lack of differential expression of Sirtuin 1 in Lucena-1 and FEPS does not invali-date the hypothesis of GAPDH mediated action in the nucleus. GAPDH trafficking may occur by a number of mechanisms.

FiGUre 2 | Expression of proteins related to cell death-resistance on K562, Lucena-1, and FEPS cells. Equal amounts of total cellular proteins (100 µg) were loaded in each lane for the detection of glyceraldehyde-3-phosphate dehydrogenase (GAPDH), pro-caspase-7, cleaved PARP, and Hsp70 (loading control) by western blot. Densitometric analysis of each lane was calculated using ImageJ Software. The data are expressed as arbitrary units and represent the mean of two independent experiments.
5
Vidal et al. MDR Cells and Metabolic Reprogramming
Frontiers in Oncology | www.frontiersin.org April 2018 | Volume 8 | Article 90
For example, it is known that GAPDH can bind to microtubules and may thus get access to several intracellular organelles, includ-ing the nucleus (68).
It has been suggested that GAPDH is an inhibitor of caspase independent cell death (CID) (61, 69, 70). Death evasion to a number of chemotherapeutic drugs is the characteristic of cells displaying the MDR phenotype. Defects in the apoptotic process involving caspase activation to induce cell death have been observed in a number of multidrug resistant cell lines (62, 71).
Apoptosis is a result of caspase activation as a consequence of mitochondrial permeabilization and cytochrome c release.
However, cells might also be killed following mitochondrial permeabilization even when caspase activation is inhibited, in the process known as CID. It has been suggested that, in such cases, cell death might occur as a result of a collapse of mitochondrial function, or the release of other proteins that could mediate the death process such as apoptosis-inducing factor (AIF), Smac/Diablo, HtrA2/Omi, Endonuclease G (72).
Mitochondrial permeabilization leading to cytochrome c release, and subsequent caspase activation, involves Bcl-2 family members having pro and anti-apoptotic properties. No difference in Bcl-2 levels could be detected when Lucena-1 and FEPS were compared with their parental non-MDR cell line K562 (46). On the other hand, the inhibitor of apoptosis survivin is increased in these cells, similarly to what has been described in K562/ADR (73) and other MDR cell lines (74, 75). Survivin suppresses cell death via caspase inhibition. Therefore, in a situation where caspases are inhibited, cell death could be a result of CID.
In a study, using CML cells, Lavallard et al. described that the tyrosine kinase inhibitor, Imatinib, was able to induce cell death in Bcr-Abl-positive cells by both caspase-dependent and independent manner (61). To induce CID, Imatinib was added to CML cells treated with caspase-inhibitors. In such situa-tion cells transfected with GAPDH were protected from CID, however, these same transfected cells were not protected from Imatinib-induced apoptosis when no caspase-inhibitors were used. Furthermore, Imatinib-resistant K562 cells spontaneously overexpressed GAPDH compared with parental K562 and were protected from CID (61). This finding is in agreement with the results obtained with both Lucena-1 and FEPS MDR cell lines that also showed resistance to Imatinib and displayed higher levels of GAPDH compared with the parental K562 (Figure 2).
In experiments where caspase activity was inhibited but cyto-chrome c release occurred, it has been suggested that GAPDH acted by increasing the glycolytic metabolism and generating ATP as well as translocating to the nucleus where it is involved in the expression of Atg12 (70). Using GAPDH mutants, that either supported ATP production, but did not translocate to the nucleus or presented nuclear function albeit unable to produce ATP, these authors verified the requirement for the dual role played by GAPDH (70). The induction of Atg12 expression following mitochondrial permeabilization leads to autophagy with the removal of damaged mitochondria and subsequent cell survival (69).
Another death process involves the release of AIF from the mitochondrial intermembrane followed by its translocation to the nucleus. This step is a result of overactivation of the nuclear enzyme poly (ADP-ribose) synthetase 1 (PARP-1) (76). In the mitochondrial intermembrane space, AIF co-localizes with Hsp60. Due to its oxyredutase activity, AIF might act as a scanveger in the mitochondrial electron transport. However, AIF nuclear activity is independent of oxyredutase activity as mutations in the oxyredutase domain do not inhibit death-induction when it translocates to the nucleus (77). Once present in the nucleus AIF induces chromatin condensation and large-scale DNA fragmen-tation. The use of AIF mutants lacking the DNA-binding property abrogated cell death in spite of the preservation of AIF’s nuclear translocation (78).

6
Vidal et al. MDR Cells and Metabolic Reprogramming
Frontiers in Oncology | www.frontiersin.org April 2018 | Volume 8 | Article 90
In response to oxidative/nitrosative stress GAPDH binds to Siah and translocates to the nucleus where it activates PARP-1 (79). In the nucleus PARP-1 functions as a sensor to regulate celular DNA repair. PARP-1 overactivation, leading to AIF translocation to the nucleus, is a result of an attempt to restore damaged DNA. Usually, after repair is completed, the PAR chains are degraded. However, in cells with severely damaged DNA, nuclear PARP-1 is extensively activated and promotes the synthesis of an excess of PAR polymer. This reaches toxic levels and in the cytosol constitutes a death signal inducing AIF nuclear translocation (76, 80). The activation of PARP-1 results in the depletion of the cellular NAD+ and ATP pools. PARP-1 may be cleaved in vitro and in vivo by caspase-3 and caspase-7 originating two fragments an 89-kD catalytic fragment and a 24-kD DNA binding domain, capable of arresting the process (81). Similar fragments of 89 and 24 kD were detected in the MDR cell line FEPS, whereas only the 89 was observed in Lucena-1 cells (Figure 2). The relationship between PARP-1 activity and drug resistance is unclear but ABCB1 activity occurs at the expense of ATP. It has been described that cells from animals knockout for PARP-1 showed and increased ABCB1 expression and activity (82).
intraCeLLULar CaLCiUM and Mdr
The homeostatic control of cytosolic calcium concentration is of fundamental importance and changes in intracellular microenvi-ronmental calcium levels can impact on cell survival, diverse cell functions, and cell death.
Proteins of the S100 family are calcium sensing proteins important in maintaining the homeostasis of the cell (83). Reports show that the calcium binding protein S100A6, also known as calcyclin or CACYBP, interacts with GAPDH (84). S100A6 is present in tumor cells as well as in normal fibroblasts, smooth, and heart muscle cells (83). This protein, as well as its gene expression, is increased twofold in FEPS compared with K562 (48). However, it is not clear how the interaction of S100A6 and GAPDH might play a role inhibiting CID.
Sorcin is another calcium–protein associated with MDR (85–87). Using K562 cells overexpressing sorcin, it was observed that ABCB1 was upregulated. The opposite was also true. In MDR-K562 cells selected by exposure to doxorubicin it was found that sorcin was upregulated, suggesting that ABCB1 and sorcin may regulate the expression of each other (85). When sorcin was analyzed using the MDR cell line Lucena-1 selected by vincristine, an increased gene and protein expression was also observed compared with the parental K562 (47, 48). This result is in agreement with the proteomics of another doxorubicin-induced K562 where the protein sorcin was also increased (53).
Despite the fact that the relationship between sorcin and ABCB1 has been known for a number of years (88), the mean-ing of such relation is still unknown. Differences in intracel-lular calcium levels have been reported in cells presenting the MDR phenotype related to ABCB1 overexpression (89). Some of these differences could be attributed to increased sorcin levels but in most cases, a causal relationship was not established.
Cells transfected with cDNA for ABCB1 did not mobilize calcium when the SERCA inhibitor thapsigargin was used (90). The explanation proposed by Gutheil et al. was that thapsigargin was being extruded. However, no calcium mobilization could be induced in Lucena-1 cells using thapsigargin in a condition in which the inhibitor could not be extruded from the cell, suggest-ing that drug efflux could not fully explain the lack of mobiliza-tion (91). Considering that the main intracellular calcium store is the endoplasmic reticulum (ER) and that thapsigargin elicits ER-stress inhibiting the ER calcium pump, this is another death-inducing mechanism that is altered in MDR cells. Similarly, other workers using two types of resistant cells, one selected with vincristine and one with a stable transfection with a gene encod-ing ABCB1, also described the reduced sensitivity of MDR cells to thapsigargin (92, 93). Moreover, using immunofluorescence, they observed differences in the localization of the ER proteins ryanodine receptor (RyR), inositol 1,4,5-trisphosphate receptor and calnexin, between the MDR cells and their parental (92). Despite the fact that no statistical difference was observed in the amount of GAPDH comparing the two types of resistant cells with the parental one, treatment with thapsigargin decreased the protein content of GAPDH as well as ABCB1, suggesting a relationship between the two (93).
ConCLUsion
The MDR phenotype is a very complex phenomenon. There is now a growing awareness that drug resistance transcends the ABCB1 transporters and involves other elements associated to metabolic reprogramming. Accordingly, the enzymes of the glycolytic path-way have been shown to exert several regulatory roles that bear on drug resistance. Therefore, pharmacological interference studies on the MDR phenotype may have a better chance to succeed if they are expanded by include as potential targets GAPDH and other enzymes of the glycolytic pathway. In this review, many ancillary roles of such enzymes have been commented and under-lined those that affect, for instance, the supply of ATP for drug extrusion purposes. Furthermore, attention was called to the fact that cells displaying the MDR phenotype also displayed enhanced expression/activity of enzymes involved in the maintenance of redox homeostasis of tumor cells. Thus, chemotherapy aiming at the combined abrogation of drug resistance and careful modula-tion of the redox equilibrium may open up alternative avenues for the control of tumor growth and metastasis.
aUtHor ContriBUtions
VR wrote and reviewed the article. RV and JQ contributed to the writing and data gathering. JQ contributed to the editing and submission of this manuscript. FR contributed to the writing and reviewed the article.
FUndinG
This work received financial support from the following Brazilian agencies: Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação Carlos Chagas Filho

7
Vidal et al. MDR Cells and Metabolic Reprogramming
Frontiers in Oncology | www.frontiersin.org April 2018 | Volume 8 | Article 90
de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES-PROCAD). RV and JQ were recipients of PhD
fellowships from CNPq. The authors would like to thank Mariana Figueiredo Rodrigues for help in performing the high-resolution respirometry experiments.
reFerenCes
1. Holohan C, Van Schaeybroeck S, Longley DB, Johnston PG. Cancer drug resis-tance: an evolving paradigm. Nat Rev Cancer (2013) 13:714–26. doi:10.1038/nrc3599
2. Borst P, Evers R, Kool M, Wijnholds J. A family of drug transporters: the mul-tidrug resistance-associated proteins. J Natl Cancer Inst (2000) 92:1295–302. doi:10.1093/jnci/92.16.1295
3. Litman T, Druley TE, Stein WD, Bates SE. From MDR to MXR: new understanding of multidrug resistance systems, their properties and clinical significance. Cell Mol Life Sci (2001) 58:931–59. doi:10.1007/PL00000912
4. Gottesman MM, Fojo T, Bates SE. Multidrug resistance in cancer: role of ATP-dependent transporters. Nat Rev Cancer (2002) 2:48–58. doi:10.1038/nrc706
5. Tsuruo T, Naito M, Tomida A, Fujita N, Mashima T, Sakamoto H, et al. Molecular targeting therapy of cancer: drug resistance, apoptosis and survival signal. Cancer Sci (2003) 94:15–21. doi:10.1111/j.1349-7006.2003.tb01345.x
6. Comerford KM, Wallace TJ, Karhausen J, Louis NA, Montalto MC, Colgan SP. Hypoxia-inducible factor-1-dependent regulation of the multidrug resistance (MDR1) gene. Cancer Res (2002) 62:3387–94.
7. Schöning JP, Monteiro M, Gu W. Drug resistance and cancer stem cells: the shared but distinct roles of hypoxia-inducible factors HIF1α and HIF2α. Clin Exp Pharmacol Physiol (2017) 44:153–61. doi:10.1111/1440-1681.12693
8. Liu ZH, Dai XM, Du B. Hes1: a key role in stemness, metastasis and multidrug resistance. Cancer Biol Ther (2015) 16:353–9. doi:10.1080/15384047.2015.1016662
9. Gustafsson MV, Zheng X, Pereira T, Gradin K, Jin S, Lundkvist J, et al. Hypoxia requires notch signaling to maintain the undifferentiated cell state. Dev Cell (2005) 9:617–28. doi:10.1016/j.devcel.2005.09.010
10. Milane L, Duan Z, Amiji M. Role of hypoxia and glycolysis in the development of multi-drug resistance in human tumor cells and the establishment of an orthotopic multi-drug resistant tumor model in nude mice using hypoxic pre-conditioning. Cancer Cell Int (2011) 11:3. doi:10.1186/1475-2867-11-3
11. Ganapathy-Kanniappan S, Geschwind JF. Tumor glycolysis as a target for cancer therapy: progress and prospects. Mol Cancer (2013) 12:152. doi:10.1186/1476-4598-12-152
12. Warburg O. On respiratory impairment in cancer cells. Science (1956) 124: 269–70.
13. Folmes CD, Terzic A. Energy metabolism in the acquisition and main-tenance of stemness. Semin Cell Dev Biol (2016) 52:68–75. doi:10.1016/j.semcdb.2016.02.010
14. Viale A, Corti D, Draetta GF. Tumors and mitochondrial respiration: a neglected connection. Cancer Res (2015) 75:3685–6. doi:10.1158/0008-5472.CAN-15-0491
15. Moreno-Sánchez R, Marín-Hernández A, Saavedra E, Pardo JP, Ralph SJ, Rodríguez-Enríquez S. Who controls the ATP supply in cancer cells? Biochemistry lessons to understand cancer energy metabolism. Int J Biochem Cell Biol (2014) 50:10–23. doi:10.1016/j.biocel.2014.01.025
16. Porporato PE, Payen VL, Pérez-Escuredo J, De Saedeleer CJ, Danhier P, Copetti T, et al. A mitochondrial switch promotes tumor metastasis. Cell Rep (2014) 8:754–66. doi:10.1016/j.celrep.2014.06.043
17. Omran Z, Scaife P, Stewart S, Rauch C. Physical and biological characteristics of multi drug resistance (MDR): an integral approach considering pH and drug resistance in cancer. Semin Cancer Biol (2017) 43:42–8. doi:10.1016/j.semcancer.2017.01.002
18. Schindler M, Grabski S, Hoff E, Simon SM. Defective pH regulation of acidic compartments in human breast cancer cells (MCF-7) is normalized in adriamycin-resistant cells (MCF-7adr). Biochemistry (1996) 35:2811–7. doi:10.1021/bi952234e
19. Altan N, Chen Y, Schindler M, Simon SM. Defective acidification in human breast tumor cells and implications for chemotherapy. J Exp Med (1998) 187:1583–98. doi:10.1084/jem.187.10.1583
20. Filomeni G, De Zio D, Cecconi F. Oxidative stress and autophagy: the clash between damage and metabolic needs. Cell Death Differ (2015) 22:377–88. doi:10.1038/cdd.2014.150
21. Ricci JE, Muñoz-Pinedo C, Fitzgerald P, Bailly-Maitre B, Perkins GA, Yadava N, et al. Disruption of mitochondrial function during apoptosis is mediated by caspase cleavage of the p75 subunit of complex I of the electron transport chain. Cell (2004) 117:773–86. doi:10.1016/j.cell.2004.05.008
22. Martinvalet D, Dykxhoorn DM, Ferrini R, Lieberman J. Granzyme A cleaves a mitochondrial complex I protein to initiate caspase-independent cell death. Cell (2008) 133:681–92. doi:10.1016/j.cell.2008.03.032
23. Jacquemin G, Margiotta D, Kasahara A, Bassoy EY, Walch M, Thiery J, et al. Granzyme B-induced mitochondrial ROS are required for apoptosis. Cell Death Differ (2015) 22:862–74. doi:10.1038/cdd.2014.180
24. Cen J, Zhang L, Liu F, Zhang F, Ji BS. Long-term alteration of reactive oxygen species led to multidrug resistance in MCF-7 cells. Oxid Med Cell Longev (2016) 2016:7053451. doi:10.1155/2016/7053451
25. Wartenberg M, Richter M, Datchev A, Günther S, Milosevic N, Bekhite MM, et al. Glycolytic pyruvate regulates P-Glycoprotein expression in multicellular tumor spheroids via modulation of the intracellular redox state. J Cell Biochem (2010) 109:434–46. doi:10.1002/jcb.22422
26. Cort A, Ozben T, Saso L, De Luca C, Korkina L. Redox control of multidrug resistance and its possible modulation by antioxidants. Oxid Med Cell Longev (2016) 2016:4251912. doi:10.1155/2016/4251912
27. Ralser M, Wamelink MM, Kowald A, Gerisch B, Heeren G, Struys EA, et al. Dynamic rerouting of the carbohydrate flux is key to counteracting oxidative stress. J Biol (2007) 6:10. doi:10.1186/jbiol61
28. Hildebrandt T, Knuesting J, Berndt C, Morgan B, Scheibe R. Cytosolic thiol switches regulating basic cellular functions: GAPDH as an information hub? Biol Chem (2015) 396:523–37. doi:10.1515/hsz-2014-0295
29. Nakajima H, Itakura M, Kubo T, Kaneshige A, Harada N, Izawa T, et al. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) aggregation causes mitochondrial dysfunction during oxidative stress-induced cell death. J Biol Chem (2017) 292:4727–42. doi:10.1074/jbc.M116.759084
30. Campanella ME, Chu H, Low PS. Assembly and regulation of a glycolytic enzyme complex on the human erythrocyte membrane. Proc Natl Acad Sci U S A (2005) 102:2402–7. doi:10.1073/pnas.0409741102
31. Tristan C, Shahani N, Sedlak TW, Sawa A. The diverse functions of GAPDH: views from different subcellular compartments. Cell Signal (2011) 23:317–23. doi:10.1016/j.cellsig.2010.08.003
32. Sawa A, Khan AA, Hester LD, Snyder SH. Glyceraldehyde-3-phosphate dehydrogenase: nuclear translocation participates in neuronal and nonneu-ronal cell death. Proc Natl Acad Sci U S A (1997) 94:11669–74. doi:10.1073/pnas.94.21.11669
33. Brüne B, Lapetina EG. Nitric oxide-induced covalent modification of glyco-lytic enzyme glyceraldehyde-3-phosphate dehydrogenase. Methods Enzymol (1996) 269:400–7. doi:10.1016/S0076-6879(96)69041-7
34. Lincet H, Icard P. How do glycolytic enzymes favour cancer cell prolifera-tion by nonmetabolic functions? Oncogene (2015) 34:3751–9. doi:10.1038/onc.2014.320
35. Bustamante E, Pedersen PL. High aerobic glycolysis of rat hepatoma cells in culture: role of mitochondrial hexokinase. Proc Natl Acad Sci U S A (1977) 74:3735–9. doi:10.1073/pnas.74.9.3735
36. Watanabe H, Takehana K, Date M, Shinozaki T, Raz A. Tumor cell autocrine motility factor is the neuroleukin/phosphohexose isomerase polypeptide. Cancer Res (1996) 56:2960–3.
37. Jung DW, Kim WH, Williams DR. Chemical genetics and its application to moonlighting in glycolytic enzymes. Biochem Soc Trans (2014) 42:1756–61. doi:10.1042/BST20140201
38. Tosato V, Grüning NM, Breitenbach M, Arnak R, Ralser M, Bruschi CV. Warburg effect and translocation-induced genomic instability: two yeast models for cancer cells. Front Oncol (2013) 2:212. doi:10.3389/fonc. 2012.00212

8
Vidal et al. MDR Cells and Metabolic Reprogramming
Frontiers in Oncology | www.frontiersin.org April 2018 | Volume 8 | Article 90
39. Nikitin DV, Bruschi CV, Sims J, Breitenbach M, Rinnerthaler M, Tosato V. Chromosome translocation may lead to PRK1-dependent anticancer drug resistance in yeast via endocytic actin network deregulation. Eur J Cell Biol (2014) 93:145–56. doi:10.1016/j.ejcb.2014.03.003
40. Corrêa S, Pizzatti L, Du Rocher B, Mencalha A, Pinto D, Abdelhay E. A com-parative proteomic study identified LRPPRC and MCM7 as putative actors in imatinib mesylate cross-resistance in Lucena cell line. Proteome Sci (2012) 10:23. doi:10.1186/1477-5956-10-23
41. De Vos M, Schreiber V, Dantzer F. The diverse roles and clinical relevance of PARPs in DNA damage repair: current state of the art. Biochem Pharmacol (2012) 84:137–46. doi:10.1016/j.bcp.2012.03.018
42. Nicholson DW, Ali A, Thornberry NA, Vaillancourt JP, Ding CK, Gallant M, et al. Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature (1995) 376:37–43. doi:10.1038/376037a0
43. Boulares AH, Yakovlev AG, Ivanova V, Stoica BA, Wang G, Iyer S, et al. Role of poly(ADP-ribose) polymerase (PARP) cleavage in apoptosis. J Biol Chem (1999) 274:22932–40. doi:10.1074/jbc.274.33.22932
44. Montoni A, Robu M, Pouliot E, Shah GM. Resistance to PARP-inhibitors in cancer therapy. Front Pharmacol (2013) 4:18. doi:10.3389/fphar.2013.00018
45. Rumjanek VM, Trindade GS, Wagner-Souza K, de-Oliveira MC, Marques-Santos LF, Maia RC, et al. Multidrug resistance in tumour cells: characteriza-tion of the multidrug resistant cell line K562-Lucena 1. An Acad Bras Cienc (2001) 73:57–69. doi:10.1590/S0001-37652001000100007
46. Daflon-Yunes N, Pinto-Silva FE, Vidal RS, Novis BF, Berguetti T, Lopes RR, et al. Characterization of a multidrug-resistant chronic myeloid leukemia cell line presenting multiple resistance mechanisms. Mol Cell Biochem (2013) 383:123–35. doi:10.1007/s11010-013-1761-0
47. Moreira MA, Bagni C, de Pinho MB, Mac-Cormick TM, dos Santos Mota M, Pinto-Silva FE, et al. Changes in gene expression profile in two multidrug resistant cell lines derived from a same drug sensitive cell line. Leuk Res (2014) 38:983–7. doi:10.1016/j.leukres.2014.06.001
48. Vidal SR. Alvos Terapêuticos no Fenótipo de Resistência a Múltiplas Drogas [Doctoral/PhD’s thesis]. Rio de Janeiro (RJ): Universidade Federal do Rio de Janeiro (2017).
49. da Cunha Vasconcelos F, Mauricio Scheiner MA, Moellman-Coelho A, Mencalha AL, Renault IZ, Rumjanek VM, et al. Low ABCB1 and high OCT1 levels play a favorable role in the molecular response to imatinib in CML patients in the community clinical practice. Leuk Res (2016) 51:3–10. doi:10.1016/j.leukres.2016.10.005
50. Maia RC, Vasconcelos FC, Souza PS, Rumjanek VM. Towards comprehension of the ABCB1/P-glycoprotein role in chronic myeloid leukemia. Molecules (2018) 23:E119. doi:10.3390/molecules23010119
51. Burger H, van Tol H, Boersma AW, Brok M, Wiemer EA, Stoter G, et al. Imatinib mesylate (STI571) is a substrate for the breast cancer resistance protein (BCRP)/ABCG2 drug pump. Blood (2004) 104:2940–2. doi:10.1182/blood-2004-04-1398
52. Ozvegy-Laczka C, Hegedus T, Várady G, Ujhelly O, Schuetz JD, Váradi A, et al. High-affinity interaction of tyrosine kinase inhibitors with the ABCG2 multidrug transporter. Mol Pharmacol (2004) 65:1485–95. doi:10.1124/mol.65.6.1485
53. Qinghong S, Shen G, Lina S, Yueming Z, Xiaoou L, Jianlin W, et al. Comparative proteomics analysis of differential proteins in respond to doxo-rubicin resistance in myelogenous leukemia cell lines. Proteome Sci (2015) 13:1. doi:10.1186/s12953-014-0057-y
54. Rutherford T, Clegg JB, Higgs DR, Jones RW, Thompson J, Weatherall DJ. Embryonic erythroid differentiation in the human leukemic cell line K562. Proc Natl Acad Sci U S A (1981) 78:348–52. doi:10.1073/pnas.78.1.348
55. Jacquel A, Colosetti P, Grosso S, Belhacene N, Puissant A, Marchetti S, et al. Apoptosis and erythroid differentiation triggered by Bcr-Abl inhibitors in CML cell lines are fully distinguishable processes that exhibit different sensitivity to caspase inhibition. Oncogene (2007) 26:2445–58. doi:10.1038/sj.onc.1210034
56. Trindade GS, Capella MA, Capella LS, Affonso-Mitidieri OR, Rumjanek VM. Differences in sensitivity to UVC, UVB and UVA radiation of a multidrug- resistant cell line overexpressing P-glycoprotein. Photochem Photobiol (1999) 69:694–9. doi:10.1111/j.1751-1097.1999.tb03348.x
57. Ho HY, Cheng ML, Chiu DT. Glucose-6-phosphate dehydrogenase—beyond the realm of red cell biology. Free Radic Res (2014) 48:1028–48. doi:10.3109/10715762.2014.913788
58. Hanschmann EM, Godoy JR, Berndt C, Hudemann C, Lillig CH. Thioredoxins, glutaredoxins, and peroxiredoxins—molecular mechanisms and health sig-nificance: from cofactors to antioxidants to redox signaling. Antioxid Redox Signal (2013) 19:1539–605. doi:10.1089/ars.2012.4599
59. Milane L, Ganesh S, Shah S, Duan ZF, Amiji M. Multi-modal strategies for overcoming tumor drug resistance: hypoxia, the Warburg effect, stem cells, and multifunctional nanotechnology. J Control Release (2011) 155:237–47. doi:10.1016/j.jconrel.2011.03.032
60. Ohmine K, Nagai T, Tarumoto T, Miyoshi T, Muroi K, Mano H, et al. Analysis of gene expression profiles in an imatinib-resistant cell line, KCL22/SR. Stem Cells (2003) 21:315–21. doi:10.1634/stemcells.21-3-315
61. Lavallard VJ, Pradelli LA, Paul A, Bénéteau M, Jacquel A, Auberger P, et al. Modulation of caspase-independent cell death leads to resensitization of imatinib mesylate-resistant cells. Cancer Res (2009) 69:3013–20. doi:10.1158/ 0008-5472.CAN-08-2731
62. Cerezo D, Cánovas M, García-Peñarrubia P, Martín-Orozco E. Collateral sensitivity to cold stress and differential Bcl-2 family expression in new daunomycin-resistant lymphoblastoid cell lines. Exp Cell Res (2015) 331: 11–20. doi:10.1016/j.yexcr.2014.11.017
63. Guan J, Sun J, Sun F, Lou B, Zhang D, Mashayekhi V, et al. Hypoxia-induced tumor cell resistance is overcome by synergistic GAPDH-siRNA and che-motherapy co-delivered by long-circulating and cationic-interior liposomes. Nanoscale (2017) 9:9190–201. doi:10.1039/c7nr02663c
64. Nicholls C, Li H, Liu JP. GAPDH: a common enzyme with uncommon functions. Clin Exp Pharmacol Physiol (2012) 39:674–9. doi:10.1111/j.1440- 1681.2011.05599.x
65. Sirover MA. Subcellular dynamics of multifunctional protein regulation: mechanisms of GAPDH intracellular translocation. J Cell Biochem (2012) 113:2193–200. doi:10.1002/jcb.24113
66. Joo HY, Woo SR, Shen YN, Yun MY, Shin HJ, Park ER, et al. SIRT1 interacts with and protects glyceraldehyde-3-phosphate dehydrogenase (GAPDH) from nuclear translocation: implications for cell survival after irradiation. Biochem Biophys Res Communn (2012) 424:681–6. doi:10.1016/j.bbrc.2012.07.006
67. Kosova AA, Khodyreva SN, Lavrik OI. Role of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) in DNA repair. Biochemistry (Mosc) (2017) 82:643–54. doi:10.1134/S0006297917060013
68. Tisdale EJ, Talati NK, Artalejo CR, Shisheva A. GAPDH binds Akt to facilitate cargo transport in the early secretory pathway. Exp Cell Res (2016) 349:310–9. doi:10.1016/j.yexcr.2016.10.025
69. Rathmell JC, Kornbluth S. Filling a GAP(DH) in caspase-independent cell death. Cell (2007) 129:861–3. doi:10.1016/j.cell.2007.05.030
70. Colell A, Ricci JE, Tait S, Milasta S, Maurer U, Bouchier-Hayes L, et al. GAPDH and autophagy preserve survival after apoptotic cytochrome c release in the absence of caspase activation. Cell (2007) 129:983–97. doi:10.1016/ j.cell.2007.03.045
71. Cerezo D, Lencina M, Ruiz-Alcaraz AJ, Ferragut JA, Saceda M, Sanchez M, et al. Acquisition of MDR phenotype by leukemic cells is associated with increased caspase-3 activity and a collateral sensitivity to cold stress. J Cell Biochem (2012) 113:1416–25. doi:10.1002/jcb.24016
72. Tait SW, Green DR. Caspase-independent cell death: leaving the set without the final cut. Oncogene (2008) 27:6452–61. doi:10.1038/onc.2008.311
73. Wang YJ, Zhao HD, Zhu CF, Li J, Xie HJ, Chen YX. Tuberostemonine reverses multidrug resistance in chronic myelogenous leukemia cells K562/ADR. J Cancer (2017) 8:1103–12. doi:10.7150/jca.17688
74. Wang L, Zhang GM, Feng ZH. Down-regulation of survivin expression reversed multidrug resistance in adriamycin-resistant HL-60/ADR cell line. Acta Pharmacol Sin (2003) 24:1235–40.
75. Liu F, Xie ZH, Cai GP, Jiang YY. The effect of survivin on multidrug resistance mediated by P-glycoprotein in MCF-7 and its adriamycin resistant cells. Biol Pharm Bull (2007) 30:2279–83. doi:10.1248/bpb.30.2279
76. Fatokun AA, Dawson VL, Dawson TM. Parthanatos: mitochondrial- linked mechanisms and therapeutic opportunities. Br J Pharmacol (2014) 171:2000–16. doi:10.1111/bph.12416
77. Daugas E, Susin SA, Zamzami N, Ferri KF, Irinopoulou T, Larochette N, et al. Mitochondrio-nuclear translocation of AIF in apoptosis and necrosis. FASEB J (2000) 14:729–39. doi:10.1096/fasebj.14.5.729
78. Ye H, Cande C, Stephanou NC, Jiang S, Gurbuxani S, Larochette N, et al. DNA binding is required for the apoptogenic action of apoptosis inducing factor. Nat Struct Biol (2002) 9:680–4. doi:10.1038/nsb836

9
Vidal et al. MDR Cells and Metabolic Reprogramming
Frontiers in Oncology | www.frontiersin.org April 2018 | Volume 8 | Article 90
79. Nakajima H, Kubo T, Ihara H, Hikida T, Danjo T, Nakatsuji M, et al. Nuclear-translocated glyceraldehyde-3-phosphate dehydrogenase promotes poly(ADP-ribose) polymerase-1 activation during oxidative/nitrosative stress in stroke. J Biol Chem (2015) 290:14493–503. doi:10.1074/jbc.M114.635607
80. Sousa FG, Matuo R, Soares DG, Escargueil AE, Henriques JAP, Larsen AK, et al. PARPs and the DNA damage response. Carcinogenesis (2012) 33:1433–40. doi:10.1093/carcin/bgs132
81. Chaitanya GV, Steven AJ, Babu PP. PARP-1 cleavage fragments: signatures of cell-death proteases in neurodegeneration. Cell Commun Signal (2010) 8:31. doi:10.1186/1478-811X-8-31
82. Wurzer G, Herceg Z, Wesierska-Gadek J. Increased resistance to anticancer therapy of mouse cells lacking the poly(ADP-ribose) polymerase attributable to up-regulation of the multidrug resistance gene product P-glycoprotein. Cancer Res (2000) 60:4238–44.
83. Donato R. Functional roles of S100 proteins, calcium-binding proteins of the EF-hand type. Biochim Biophys Acta (1999) 1450:191–231. doi:10.1016/S0167-4889(99)00058-0
84. Filipek A, Wojda U, Leśniak W. Interaction of calcyclin and its cyanogen bromide fragments with annexin II and glyceraldehyde 3-phosphate dehydrogenase. Int J Biochem Cell Biol (1995) 27:1123–31. doi:10.1016/1357- 2725(95)00096-8
85. Qi J, Liu N, Zhou Y, Tan Y, Cheng Y, Yang C, et al. Overexpression of sorcin in multidrug resistant human leukemia cells and its role in regulating cell apoptosis. Biochem Biophys Res Commun (2006) 349:303–9. doi:10.1016/j.bbrc.2006.08.042
86. Zhou Y, Xu Y, Tan Y, Qi J, Xiao Y, Yang C, et al. Sorcin, an important gene associated with multidrug-resistance in human leukemia cells. Leuk Res (2006) 30:469–76. doi:10.1016/j.leukres.2005.08.024
87. Colotti G, Poser E, Fiorillo A, Genovese I, Chiarini V, Ilari A. Sorcin, a calcium binding protein involved in the multidrug resistance mechanisms in cancer cells. Molecules (2014) 19:13976–89. doi:10.3390/molecules190913976
88. Sugawara I, Mizumoto K, Ohkochi E, Hamada H, Tsuruo T, Mori S. Immunocytochemical identification and localization of the Mr 22,000 calcium- binding protein (sorcin) in an adriamycin-resistant myelogenous leukemia
cell line. Jpn J Cancer Res (1989) 80:469–74. doi:10.1111/j.1349-7006.1989.tb02338.x
89. Sulová Z, Seres M, Barancík M, Gibalová L, Uhrík B, Poleková L, et al. Does any relationship exist between P-glycoprotein-mediated multidrug resistance and intracellular calcium homeostasis. Gen Physiol Biophys (2009) 28 Spec No Focus:F89–95.
90. Gutheil JC, Hart SR, Belani CP, Melera PW, Hussain A. Alterations in Ca2+ transport ATPase and P-glycoprotein expression can mediate resistance to thapsigargin. J Biol Chem (1994) 269:7976–81.
91. Wagner-Souza K, Echevarria-Lima J, Rodrigues LA, Reis M, Rumjanek VM. Resistance to thapsigargin-induced intracellular calcium mobilization in a multidrug resistant tumour cell line. Mol Cell Biochem (2003) 252:109–16. doi:10.1023/A:1025586225941
92. Sereš M, Poláková E, Krizanová O, Hudecová S, Klymenko SV, Breier A, et al. Overexpression of P-glycoprotein in L1210/VCR cells is associated with changes in several endoplasmic reticulum proteins that may be partially responsible for the lack of thapsigargin sensitivity. Gen Physiol Biophys (2008) 27:211–21.
93. Sereš M, Ditte P, Breier A, Sulová Z. Effect of thapsigargin on P-glycoprotein-negative and P-glycoprotein-positive L1210 mouse leukaemia cells. Gen Physiol Biophys (2010) 29:396–401. doi:10.4149/gpb_2010_04_396
Conflict of Interest Statement: The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer CVB and the handling Editor declared their past shared affiliation.
Copyright © 2018 Vidal, Quarti, Rumjanek and Rumjanek. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.


















