
Liliana Andrea Villegas Sierra Avaliação dos … Os valores de dispersão ... Escala: o corpo da...
126
Liliana Andrea Villegas Sierra Avaliação dos Parâmetros de Transporte de um Biocolóide através de uma Areia Saturada Dissertação de Mestrado Dissertação apresentada ao Programa de Engenharia da PUC-Rio como requisito parcial para obtenção do título de Mestre em Engenharia Civil. Orientador: Prof. Tácio Mauro Pereira de Campos Co-orientador: Prof a . Denise Maria Mano Pessôa Rio de janeiro Setembro de 2010
Transcript of Liliana Andrea Villegas Sierra Avaliação dos … Os valores de dispersão ... Escala: o corpo da...

Liliana Andrea Villegas Sierra
Avaliação dos Parâmetros de Transporte de um Biocolóide
através de uma Areia Saturada
Dissertação de Mestrado
Dissertação apresentada ao Programa de Engenharia da PUC-Rio como requisito parcial para obtenção do título de Mestre em Engenharia Civil.
Orientador: Prof. Tácio Mauro Pereira de Campos Co-orientador: Profa. Denise Maria Mano Pessôa
Rio de janeiro
Setembro de 2010
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Liliana Andrea Villegas Sierra
Avaliação dos Parâmetros de Transporte de um Biocolóide
através de uma Areia Saturada
Dissertação apresentada como requisito parcial para obtenção do título de Mestre pelo Programa de Pós-Graduação em Engenharia Civil da PUC-Rio. Aprovada pela Comissão Examinadora abaixo assinada.
Prof. Tácio Mauro Pereira de Campos Orientador
Departamento de Engenharia Civil - PUC-Rio
Profa. Denise Maria Mano Pessôa Co-orientador
Departamento de Engenharia Civil - PUC-Rio
Prof. Eurípedes de Amaral Vargas Jr. Departamento de Engenharia Civil - PUC-Rio
Profa. Maria Claudia Barbosa Universidade Federal do Rio de Janeiro
Prof. José Tavares Araruna Júnior Departamento de Engenharia Civil - PUC-Rio
Prof. José Eugenio Leal Coordenador(a) Setorial do Centro Técnico Científico - PUC-Rio
Rio de Janeiro, 27 de setembro de 2010
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Todos os direitos reservados. É proibida a reprodução total ou parcial do trabalho sem autorização da universidade, da autora e do orientador.
Liliana Andrea Villegas Sierra
Graduou-se em Engenharia Civil pela Universidad Mayor de San Andrés, Bolívia em 2006. Ingressou no curso de mestrado em Engenharia Civil em 2008 na área de Geotecnia. Principais áreas de interesse e linhas de pesquisa: Geotecnia Ambiental, Geotecnia Experimental, Engenharia Sanitária e Hidráulica.
Ficha Catalográfica
Sierra, Liliana Andrea Villegas
Avaliação dos parâmetros de transporte de um biocolóide através de uma areia saturada / Liliana Andrea Villegas Sierra ; orientador: Tácio Mauro Pereira de Campos ; co-orientador: Denise Maria Mano Pessôa. – 2010.
126 f. : il. (color.) ; 30 cm
Dissertação (mestrado)-Pontifícia Universidade Católica do Rio de Janeiro, Departamento de Engenharia Civil, 2010.
Inclui bibliografia
1. Engenharia civil – Teses. 2. Transporte de contaminantes. 3. Escherichia coli. 4. Areia de quartzo. 5. Filtração mecânica. I. Campos, Tácio Mauro Pereira de. II. Pessôa, Denise Maria Mano. III. Pontifícia Universidade Católica do Rio de Janeiro. Departamento de Engenharia Civil. IV. Título.
CDD: 624
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Para meus pais, Félix e Lupe
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Agradecimentos
À CAPES pelo apoio financeiro e à PUC-Rio pela oportunidade de cursar o
mestrado.
Ao professor Tácio Mauro Pereira de Campos, pela orientação e confiança, para
a realização deste trabalho.
À professora Denise Mano Pessôa, obrigada pela ajuda constante, a paciência,
e dedicação que me brindou ao longo de todo este período.
Aos Professores do Departamento de Engenharia Civil da PUC-Rio, pelo ensino
transmitido.
A minha família que com amor incondicional, sempre me apóia e incentiva em
cada etapa da minha vida.
A todos os meus amigos pela amizade sincera, que com alegria, entusiasmo e
companheirismo me ajudaram a superar todos os obstáculos e fizeram do
mestrado uma das melhores etapas da minha vida.
Em especial ao Gerardo, pela compreensão e carinho, principalmente naqueles
momentos em que todo parecia dar errado, obrigada por me ajudar a tornar este
trabalho possível.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Resumo
Sierra, Liliana Andrea Villegas; Campos, Tácio Mauro Pereira de; Pessôa, Denise Maria Mano. Avaliação dos Parâmetros de Transporte de um Biocolóide através de uma Areia Saturada. Rio de janeiro, 2010. 126p. Dissertação de Mestrado - Departamento de Engenharia Civil, Pontifícia Universidade Católica do Rio de Janeiro.
Neste trabalho são avaliados os parâmetros de transporte da bactéria
Escherichia coli ATCC11229, microrganismo indicador de contaminação fecal,
através de colunas de areia de quartzo saturadas. Com este propósito foi
desenvolvido o equipamento para a execução dos ensaios Advecção-Dispersão-
Sorção (ADS), com injeção contínua de uma suspensão bacteriana em água
destilada em uma concentração inicial de 108 bactérias/mL, por até doze horas
em temperatura ambiente e sob condições que permitiram desprezar os efeitos
do crescimento e decaimento dos microrganismos. Para a determinação dos
parâmetros de transporte foram executados ensaios microbiológicos, ensaios de
batelada e oito ensaios ADS para diferentes valores de gradiente hidráulico. Os
ensaios de batelada mostraram uma baixa adsorção bactéria-solo nas condições
avaliadas. As curvas de chegada obtidas nos ensaios ADS, mostraram elevados
valores para o fator de retardamento entre três e nove, variando
proporcionalmente com o incremento de gradiente hidráulico. Os valores de
dispersão hidrodinâmica variaram entre 1,44x10 2 cm2/min e 5,47x102 cm2/min.
Os resultados dos ensaios sugerem que os processos físico-químicos têm pouca
influência no transporte deste microrganismo em areia de quartzo, enquanto que
o processo de filtração mecânica, forças hidrodinâmicas, a forma e distribuição
de tamanho dos grãos parecem explicar mais adequadamente o comportamento
das curvas de chegada.
Palavras-chave
Transporte de contaminantes; Escherichia coli; Areia de quartzo; Filtração
mecânica.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Abstract
Sierra, Liliana Andrea Villegas; Campos, Tácio Mauro Pereira de (Advisor); Pessôa, Denise Maria Mano (Advisor). Evaluation of the Transport Parameters of a Biocolloid through a Saturated Sand. Rio de janeiro, 2010. 126p. MSc. Dissertation – Departamento de Engenharia Civil, Pontifícia Universidade Católica do Rio de Janeiro.
This study deals with the assessment of the transport parameters of the
Escherichia coli ATCC11229 bacteria, a fecal contamination indicator, through
saturated quartz sand columns. With this purpose, it was developed an
equipment to carrying out Advection-Dispersion-Sorption (ADS) experiments,
with continuous input of bacterial distilled water suspension at an influent
concentration of 108 cell/mL for about twelve hours, at environmental temperature
and under conditions that allow disregard the effect of the growth and decay of
the microorganisms. Microbiological experiments, batch tests, and eight ADS
experiments at several hydraulic gradient values, were executed for determine
the transport parameters. The batch tests indicated a low bacterium-soil
adsorption under evaluated conditions. The breakthrough curves, obtained by
ADS experiments, showed high values for the retardation factor, ranged from
three to nine with increasing the hydraulic gradient. The hydrodynamic dispersion
values ranged between 1,44x10-2cm2/min and 5,47x10-2 cm2/min. The results of
the experiments suggested that the physical-chemical processes have a little
influence on the transport of this microorganism in quartz sand, whereas the
straining process, hydrodynamic forces, the shape and grain distribution, would
can explain more appropriately the behavior of the breakthrough curves.
Keywords
Transport of contaminants; Escherichia coli; Quartz sand; Straining.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Sumário
1 INTRODUÇÃO 14
1.1. Objetivo 16
1.2. Estrutura da Dissertação 17
2 REVISÃO BIBLIOGRÁFICA 18
2.1. Generalidades 18
2.2. Áreas de proteção 21
2.3. Sobrevivência de bactérias e vírus na água subterrânea 22
2.4. Escherichia coli 24
2.5. Transporte de Biocolóides 27
2.6. Processos de Transporte 29
2.6.1. Advecção 29
2.6.2. Dispersão Hidrodinâmica 30
2.6.3. Adsorção 32
2.6.3.1. Sorção de equilíbrio 34
2.6.3.2. Sorção Dinâmica 35
2.6.4. Filtração mecânica (straining) 43
2.6.5. Exclusão 46
2.6.6. Bloqueamento e Ripening 47
2.7. Ensaios experimentais 48
3 MATERIAIS E MÉTODOS 50
3.1. Considerações Gerais 50
3.2. Meio poroso 50
3.2.1. Caracterização geotécnica do meio poroso 51
3.3. Bactéria 55
3.3.1. Manutenção da cepa de Escherichia Coli 55
3.3.2. Determinação do número de bactérias 56
3.3.2.1. Contagem celular em Câmara de Neubauer 56
3.3.2.2. Leitura da densidade óptica por espectrofotometria 57
3.3.2.3. Contagem de microrganismos pela Técnica dos Tubos Múltiplos 58
3.3.2.4. Contagem de Número de Unidades Formadoras de Colônias em
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Placas de Petri 59
3.4. O equipamento para o ensaio ADS 60
3.4.1. Características do frasco de Mariotte 62
3.4.2. Características da coluna 63
3.5. Ensaios Microbiológicos 65
3.5.1. Correlação entre Número de Células e Absorbância 65
3.5.2. Curva de crescimento 67
3.5.3. Ensaio de Sobrevivência 68
3.6. Ensaio de Adsorção em Batelada 69
3.7. Ensaio Advecção-Dispersão-Sorção (ADS) 70
3.7.1. Saturação da areia 72
3.7.2. Injeção contínua da suspensão de E. coli 73
3.7.3. Obtenção das curvas de chegada 76
3.8. Ensaios de controle 77
4 APRESENTAÇÃO E DISCUSSÃO DOS RESULTADOS 79
4.1. Ensaios Microbiológicos 79
4.1.1. Curvas de correlação entre absorbância e número de células 79
4.1.2. Curva de Crescimento Escherichia coli em meio TSB 81
4.1.3. Avaliação da sobrevivência de E. coli em água 82
4.2. Ensaio de Adsorção em Batelada 83
4.3. Ensaio ADS 85
4.3.1. Curvas de chegada 86
4.3.2. Fator de Retardamento e Dispersão Hidrodinâmica 89
4.3.3. Ensaios de controle 94
4.4. Discussão 95
4.4.1. Retardamento 95
4.4.2. Filtração Mecânica como processo dominante no transporte de E. coli
nos ensaios ADS 97
4.4.3. Influencia das propriedades do meio poroso 100
5 CONCLUSÕES E SUGESTÕES 103
5.1. Conclusões 103
5.2. Sugestões 104
REFERÊNCIAS BIBLIOGRÁFICAS 105
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

APÊNDICE A 112
A.1. Determinação do número de células durante o ensaio de adsorção em
batelada 112
A.2. Cálculo do coeficiente de distribuição KD 115
APÊNDICE B 117
B.1. Dados para a construção da curva de chegada 117
B.2. Determinação gráfica dos parâmetros “b” e “R” 122
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Lista de Figuras
Figura 2.1. Fontes de contaminação biológica das águas subterrâneas
(Adaptado de Corapcioglu e Haridas, 1984) 18
Figura 2.2. Micrografia eletrônica da E. coli. Escala: o corpo da célula é de
aproximadamente 1 μm de comprimento (Berg, 2004) 25
Figura 2.3. Mecanismos básicos de transporte em filtração de água (Adaptado
de Yao et al., 1971) 37
Figura 2.4. Teoria de energia DVLO. As forças combinadas de atração e
repulsão governam a agregação de partículas com relação à
distância entre elas (Gwyn, 2003) 42
Figura 2.5. Formas em que a filtração limita a migração das partículas,
a) Filtração completa, b) Filtração incompleta ou straining, e
c) Filtração físico química (McDowell-Boyer et al., 1986) 43
Figura 2.6. a) Cinética inicial de deposição, b) Efeito do bloqueamento e
c) Efeito do ripening (Kretzschmar et al., 1999) 48
Figura 3.1. Areia de quartzo. Lupa ZEISS Stereo Discovery V8 x10 de
aumento 51
Figura 3.2. Distribuição de tamanhos dos grãos de quartzo 52
Figura 3.3. Aparato para a determinação da condutividade hidráulica saturada 53
Figura 3.4. a) Câmara de Neubauer utilizada, b) Esquema da CNB com
a lâminula sobre a área de contagem 56
Figura 3.5. Área de Contagem da CNB e esquema da metodologia
de contagem. 57
Figura 3.6. Esquema da Técnica de Contagem em Placa 60
Figura 3.7. Esquema de funcionamento do equipamento para os ensaios ADS
a) Δh=0 , b) Δh>0 61
Figura 3.8. Corte transversal e detalhe da tampa do Mariotte 62
Figura 3.9. Corte transversal e detalhe da base do Mariotte 63
Figura 3.10. Esquema dos componentes da coluna 63
Figura 3.11. Corte transversal da coluna 64
Figura 3.12. Esquema da metodologia seguida para a obtenção das curvas de
correlação entre absorbância e a) Suspensão bacteriana em
meio TSB; b) Suspensão bacteriana em água destilada 67
Figura 3.13. Crescimento das colônias de E. coli no ensaio de adsorção
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

para diferentes concentrações iniciais 70
Figura 3.14. Foto do equipamento utilizado no ensaio ADS 71
Figura 3.15. Vista superior e lateral da coluna após do processo de
conformação da coluna de areia 73
Figura 3.16. Crescimento do inóculo de E. coli em 500 mL de meio de
cultura TSB 74
Figura 3.17. Linha continua - Forma da curva de chegada sem retardamento
nem decaimento; Linha tracejada - a) Forma da curva de chegada
com retardamento b) Forma da curva de chegada com
decaimento 76
Figura 4.1. Curvas de correlação entre Número de Células e Absorbância
(a) para meio TSB; (b) para água destilada 79
Figura 4.2. Curva de Correlação para células suspendidas em meio de
cultura TSB 80
Figura 4.3. Curva de Correlação para células suspendidas em água destilada 80
Figura 4.4. Curvas de crescimento da Escherichia coli ATCC 11229 82
Figura 4.5. Isoterma de adsorção da E. coli em areia de quartzo 84
Figura 4.6. Curvas de chegada para cada ensaio ADS 88
Figura 4.7. Curvas de chegada para diferentes gradientes hidráulicos 90
Figura 4.8. Variação do fator de Retardamento com relação ao gradiente
hidráulico 91
Figura 4.9. Variação da Dispersão Hidrodinâmica com relação à velocidade
de poros 91
Figura 4.10. Variabilidade da condutividade hidráulica no ensaio ADS 93
Figura 4.11. Distribuição do tamanho dos grãos ao longo da coluna de areia 95
Figura 4.12. Tipos de locais dentro do meio poroso, susceptíveis à retenção
de bactérias devido ao processo de Filtração mecânica 99
Figura 4.13. Fotos ampliadas 600 vezes, da deposição de E.coli O157: H7
em areia de 150 μm (Bradford et al., 2006) 100
Figura 4.14. Imagens ESEM (Environmental Scanning Electron Microscope)
de a) leito de esferas de vidro (ampliadas 200 vezes, b) quartzo
com forma irregular (ampliada 500 vezes) (Tufenkji et al., 2004) 102
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Lista de Tabelas
Tabela 2.1. Doenças causadas pelo consumo de água contaminada por
excretos humanos ou animais (Bactérias, Protozoários e Vírus)
(Cajazeiras, 2007) 20
Tabela 2.2. Fatores físico-químicos que afetam a sobrevivência de bactérias
entéricas e vírus no solo (Abu-Ashour et al., 1993) 23
Tabela 2.3. Componentes utilizados em experimentos realizados para o
estudo do transporte de microrganismos 49
Tabela 3.1. Determinação da condutividade hidráulica saturada 54
Tabela 3.2. Características físicas da areia de quartzo limpa 54
Tabela 4.1. Cálculo da concentração de células viáveis suspensas em água
destilada pela Técnica dos Tubos Múltiplos 83
Tabela 4.2. Resultados obtidos através da técnica de contagem em placa 83
Tabela 4.3. Condições hidráulicas durante a saturação da areia 85
Tabela 4.4. Condições hidráulicas do sistema experimental durante a injeção
da suspensão de E. coli 86
Tabela 4.5. Parâmetros de transporte: Retardamento e Dispersão
hidrodinâmica 90
Tabela 4.6. Concentração de E. coli retida ao longo da coluna de areia
(Ensaio ADS 1) 94
Tabela 4.7. Numero de volume de poros necessários para preencher com
bactérias os espaços de poro destinados à filtração mecânica 98
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

1 INTRODUÇÃO
Um crescente esforço de pesquisa tem sido destinado ao estudo da
migração e retenção de microrganismos em ambientes granulares naturais ou
modelados. O entendimento desses processos é de grande interesse em várias
aplicações ambientais tais como a proteção de fontes naturais de água potável,
tratamentos de filtração natural de água de rios, como a filtração em margem e a
biorremediação de áreas contaminadas.
Para grande parte da população mundial, e especialmente em países em
desenvolvimento e regiões áridas, a principal fonte de água potável é a água
subterrânea, captada de poços perfurados ou escavados. No Brasil, segundo o
Instituto Brasileiro de Geografia e Estatística (IBGE, 2010), para o ano 2002,
aproximadamente 58,8% da população brasileira na área rural era abastecida,
para fins domésticos, com água de poço ou nascentes.
Considerando a relação direta entre água usada para beber e surtos de
doenças e epidemias, a contaminação da água subterrânea (em particular a
contaminação de origem microbiológica) constitui-se em uma potencial ameaça
para a saúde publica. Esta é uma das principais razões da necessidade de se
estudar o comportamento do transporte de microrganismos patogênicos no
subsolo.
Nem todos os países possuem legislação estabelecendo perímetros de
proteção em torno de captações de água subterrâneas, porém, a preocupação
em preservar os recursos hídricos é crescente e, vários países já os adotam
como medida preventiva para manter a qualidade destas águas (Coelho e
Duarte, 2003).
Esses perímetros de proteção, todavia estão frequentemente baseados em
distâncias arbitrárias, que podem subestimar ou sobreestimar a proteção dos
recursos de água. Pang et al. (2003), em um estudo relacionado com a
determinação dessas distâncias mínimas, concluíram que estimativas feitas
através da determinação em laboratório da taxa de decaimento dos
microrganismos livres, sem considerar o material do aquífero e outros processos,
podem resultar em distâncias muito maiores do que necessário.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

15
Para definir mais adequadamente os perímetros de proteção é necessária
informação referente à atenuação do transporte de patogênicos ou de
indicadores microbiológicos em sistemas de água subterrânea. Esta informação
resulta mais apropriada se efetuada considerando as características próprias do
local como o clima, o material do aquífero e a composição da água subterrânea.
Entretanto, devido à escassa informação referente a investigações
envolvendo, especificamente, esse problema no Brasil, através deste estudo,
tenta-se iniciar um melhor entendimento dos processos que influenciam o
transporte de microrganismos no solo sob as condições ambientais locais.
Com este propósito foi implementado no Laboratório de Geotecnia e Meio
Ambiente da PUC–Rio, um equipamento para a realização de ensaios de
transporte de contaminantes considerando as particularidades que exige o
trabalho com microrganismos, principalmente na minimização do potencial de
contaminação externa durante a execução dos ensaios.
A revisão bibliográfica com relação ao problema exposto mostrou que,
para fins de pesquisa, sobre transporte, microrganismos como bactérias e vírus
são considerados, por suas dimensões, como partículas suspensas ou coloidais,
mais especificamente como biocolóides. Nesses estudos são incluídos
processos conhecidos de advecção, dispersão e processos de filtração
baseados na teoria clássica da filtração do colóide, como também na teoria
DLVO (Derjaguin and Landau, and Verwey and Overbeek).
Experimentalmente, o transporte de microrganismos em meios porosos
saturados tem sido tradicionalmente investigado através da aplicação do método
de colunas empacotadas (packed column), termo utilizado em cromatografia,
onde a concentração dos microrganismos suspensos no efluente é monitorada
em função do tempo.
Estudos realizados sob condições controladas em laboratório estão
dirigidos a avaliar a influência sobre os processos de transporte das
propriedades do biocolóide, do meio poroso e da solução. Materiais como areia
de quartzo seja limpa ou revestida ou leitos de esferas de vidro vêm sendo
implementados em tais estudos como meios granulares. Como microrganismos,
são utilizadas bactérias como as do gênero Pseudomonas (Barton et al., 1995;
Martin et al., 1996; Camesano et al., 1999), Escherichia coli (Pang et al., 2003;
Redman et al., 2004; Foppen et al., 2005; Foppen et al., 2007; Jiang et al.,
2007), Cryptosporidium oocysts (Tufenkji et al., 2004) e vírus como fagos MS2
e PRD1 (Shijven et al., 2002). Em muitos casos também são usadas
microesferas de látex como um modelo representativo das bactérias.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

16
Modelos matemáticos do transporte de bactérias em meio saturado,
geralmente envolvem a forma simplificada da equação Advecção-Dispersão-
Sorção, que pode ser derivada a partir dos princípios básicos de balanço de
massa. Poucos esforços na modelagem levam em conta a influência de
numerosos fatores físicos, químicos e biológicos, que em função das atuais
pesquisas, sabe-se que afetam o transporte dos microrganismos na
subsuperfície.
Contudo, a capacidade de prever com precisão o destino de bactérias no
ambiente de subsuperfície, está atualmente limitada pela compreensão
incompleta sobre os mecanismos de deposição (Bradford et al., 2006) e o
entendimento sobre o comportamento dos microrganismos quando fluem através
de meios naturais, ainda constitui uma meta para futuras investigações.
Como ponto de partida para o início do estudo experimental do transporte
de microrganismos em meios porosos foi escolhida água destilada como
solução, areia de quartzo comercial como meio granular, e a bactéria
Escherichia coli ATCC11229 como biocolóide, por se tratar de um importante
indicador de contaminação de origem fecal, de rápido crescimento, fácil
manipulação e fácil detecção nos procedimentos laboratoriais.
1.1. Objetivo
Esta pesquisa tem como objetivo geral estudar o transporte do
microrganismo Escherichia coli ATCC11229 através de areia de quartzo
saturada, sem a interferência de fatores biológicos e sob condições
desfavoráveis à adesão.
Após definir as condições do ensaio, para alcançar esse objetivo foi
necessário o desenvolvimento dos seguintes objetivos específicos:
Elaborar procedimentos microbiológicos necessários para a
quantificação de E. coli, nos diferentes ensaios do trabalho.
Projetar e montar um equipamento apropriado para o estudo de
transporte de microrganismos nas condições experimentais
previstas.
Determinar parâmetros de transporte, retardamento e dispersão
hidrodinâmica de E. coli através de ensaios Advecção-Dispersão-
Sorção com diferentes gradientes hidráulicos.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

17
Determinar a sobrevivência de E. coli em água.
Determinar o máximo potencial de adsorção de E. coli em areia
através do ensaio de batelada, sob as mesmas condições dos
ensaios ADS.
1.2. Estrutura da Dissertação
Esta dissertação foi desenvolvida em 5 capítulos que são descritos a
seguir.
O Capítulo 1 trata da introdução da dissertação, na qual se registra a
importância de estudar o transporte dos microrganismos patogênicos em meios
granulares, e uma breve descrição do que tem sido feito nesta área de estudo
com ênfase na parte experimental. Também são apresentados os principais
objetivos e a estrutura geral da dissertação.
No Capítulo 2 é apresentada uma revisão sobre a problemática abordada e
os fundamentos teóricos referidos aos processos de transporte e retenção de
biocolóides no solo, e finalmente as bases teóricas necessárias para a
compreensão dos procedimentos aplicados em ensaios de transporte.
No Capítulo 3 faz-se uma descrição das características relevantes do
biocolóide e do meio poroso utilizados neste trabalho, assim como também, uma
descrição em detalhe do projeto para a construção do equipamento destinado à
execução dos ensaios de transporte. Os procedimentos seguidos na realização
dos ensaios microbiológicos, de batelada e de transporte também são expostos
neste capítulo.
No Capítulo 4 apresentam-se os resultados obtidos para cada ensaio
realizado, e a seguir a discussão destes para, finalmente, no Capítulo 5
apresentar as conclusões do trabalho e as propostas para trabalhos futuros
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

2 REVISÃO BIBLIOGRÁFICA
2.1. Generalidades
No mundo inteiro, os aquíferos - formações geológicas capazes de
armazenar as águas - estão sob perigo cada vez maior de contaminação em
decorrência da urbanização, do desenvolvimento industrial, das atividades
agrícolas e das empresas de mineração.
Uma grande variedade de microrganismos patogênicos pode ser
encontrada em excrementos de seres humanos e de outros animais de sangue
quente. Por isso, a contaminação de águas por material de origem fecal é uma
das formas mais importantes de introdução de microrganismos patogênicos nas
águas (Cajazeiras, 2007). A introdução destes microrganismos na água
subterrânea pode ser atribuída às seguintes causas: ruptura acidental das redes
de esgoto, vazamentos de tanques sépticos residenciais e latrinas, água
infiltrada através de aterros sanitários, recarga artificial de aquíferos por água de
esgoto tratada, e a aplicação de esgoto no solo (Corapcioglu e Haridas, 1984).
Poços mal construídos são particularmente propensos a apresentar esse tipo de
contaminação.
Figura 2.1. Fontes de contaminação biológica das águas subterrâneas
(Adaptado de Corapcioglu e Haridas, 1984)
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

19
A recarga artificial de fontes de água subterrânea por infiltração de águas
residuais e uma proposta amplamente usada para disposição de águas residuais
domesticas e municipais, por exemplo, na aplicação do sistema alternativo on-
site para o tratamento de águas residuais em áreas costeiras denominado
Marshland Upwelling System (Richardson, 2002).
Outra fonte de bactérias em solos ou em aquíferos de água subterrânea é
a aplicação controlada ou injeção de cepas de bactéria para biorremediação in
situ (bioaumentação) de locais contaminados ou como organismos de
biocontrole contra certas doenças das plantas (Kretzschmar et al., 1999).
Conhecimento sobre transporte de bactérias no meio poroso é assim um
requerimento chave para a disposição segura de águas residuais e para o
desenvolvimento de estratégias de biorremediação efetiva de solos
contaminados e aquíferos utilizando cepas de bactéria.
Existem perfis naturais de solo que podem degradar alguns poluentes até
atingirem a água subterrânea. Mas também, estudos confirmaram que
patogênicos penetram facilmente, rapidamente e profundamente durante
eventos de chuvas fortes (Chu et al., 2001; citado por Knappett, 2006).
Foppen et al. (2008) estudaram os mecanismos de transporte vertical de águas
residuais em um aquífero superficial aluvial pobremente consolidado e os
resultados indicaram a presença de E. coli em uma concentração de
106 UFC/100 mL, em profundidades de 50 m abaixo da superfície do solo.
Por tanto, a preocupação com a contaminação da água subterrânea
concentra-se principalmente nos aquíferos não confinados, especialmente nas
áreas em que a zona vadosa é pouco espessa e o lençol freático é raso.
Entretanto, há riscos significativos de poluição em pontos em que o aquífero é
semiconfinado, se os aquitardes confinantes forem relativamente pouco
espessos e permeáveis (Foster et al., 2006).
De um modo geral os microrganismos patogênicos podem ser agrupados
como: Vírus, Bactérias, Protozoários e Ovos de Helmintos. As bactérias e vírus
mais relevantes em termos de patogenicidade que poderiam possivelmente ser
transportados pelas águas subterrâneas são: Salmonella sp., Shigella sp.,
Yersinia enterocolitica, Y. pseudotuberculosis, Leptospira sp., Francisella
tularensis, Dyspepsia coli, E. coli enterotóxica, Pseudomonades, Vibrio sp.,
Legionella sp. E os vírus infecciosos hepatitis vírus, pólio vírus, Coxsackie
viruses, adenovirus, rotavirus, e Norwalklike vírus (Pekdeger e Matthess, 1983).
Organizações internacionais como a Organização Mundial de Saúde
(OMS) e outras, tem repetidamente expressado preocupação sobre a
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

20
disponibilidade e qualidade dos recursos de água, e apontado que doenças
relacionadas com a água constituem ainda sérios problemas de saúde causados
pela poluição microbiológica, saneamento inadequado e falta de água limpa. A
Tabela 2.1 mostra as principais doenças de veiculação hídrica, tendo como
agente causal a ingestão de água contaminada.
Tabela 2.1. Doenças causadas pelo consumo de água contaminada por
excretos humanos ou animais (Bactérias, Protozoários e Vírus)
(Cajazeiras, 2007)
Doença Agente Causal Sintomas
Bac
téri
a
Febre tifóide Disenteria bacilar Cólera Diarréia Leptospirose Salmonelose
Salmonella typhi Bacilo disentérico Víbrio cholerae Escherichia coli Leptospira interrogans Salmonela
Febre elevada, diarréia, ulceração do intestino delgado Forte Diarréia Diarréia extremamente forte, desidratação, alta taxa de mortalidade Icterícia, febre Febre, náusea, diarréia.
Pro
tozo
ári
os
Disenteria amebiana Giardíase
Entamoeba histolytica Giárdia lamblia
Diarréia prolongada, com sangramento, abscessos no fígado e intestino fino Diarréia leve a forte, náusea, indigestão, flatulência
Vír
us
Hepatite infecciosa Gastroenterite Paralisia infantil
Vírus da hepatite A Enterovírus, parvovírus, rotavirus Poliomielites vírus
Icterícia, febre Diarréia leve a forte Paralisia
É evidente então, a suprema importância de prevenir a ingestão de água
subterrânea contaminada. Nesse sentido, para avaliar a proteção das zonas
subterrâneas contra microrganismos patogênicos (principalmente bactérias e
vírus), os mecanismos de controle que devem ser estudados são principalmente,
a persistência das bactérias e de vírus sob condições biológicas e químicas da
água subterrânea e os processos físicos, físico-químicos, e microbiológicos que
controlam o transporte de microrganismos na água subterrânea (Pekdeger e
Matthess, 1983).
O interesse na determinação dos fatores que controlam o transporte
através de materiais geológicos e a derivação de uma teoria adequada para
descrever o transporte tem sido reforçado nas últimas duas décadas, devido à
preocupação referida à polução dos recursos de água subterrânea. As questões
incluem a contaminação das águas subterrâneas por patogênicos, o potencial do
transporte bacteriano em conjunção com a biodegradação de contaminantes
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

21
orgânicos na subsuperfície, o potencial de descarga de microrganismos gerados
geneticamente no meio geológico e a participação dos microrganismos como
facilitadores do transporte dos radionuclídeos na água subterrânea (Hornberger
et al., 1992).
2.2. Áreas de proteção
Reconhecido o problema da contaminação da água subterrânea, segundo
Foster et al. (2006) a abordagem mais lógica para este problema é, considerá-lo
como a interação entre a vulnerabilidade do aquífero à contaminação e a carga
contaminante que é, será ou pode ser aplicada no meio como resultado da
atividade humana.
Um fator importante que influencia o perigo representado por uma
atividade antrópica é sua proximidade de uma fonte de abastecimento
subterrânea (poço ou nascente). Sendo a avaliação dos perigos de
contaminação do aquífero dependente em primeiro lugar da definição da zona de
captura (perímetros de proteção do manancial) e, em segundo lugar, da
mobilidade, persistência e dispersão dos contaminantes dentro do regime de
fluxo do aquífero (Foster et al., 2006).
Uma área de proteção interna baseada na distância equivalente a um
tempo de trânsito horizontal médio na zona saturada tem sido amplamente
adotada para a proteção contra atividades que potencialmente infiltram vírus,
bactérias e parasitas patogênicos, como é o caso, por exemplo, das águas
residuais e dejetos espalhados nas lavouras. No entanto, o tempo de trânsito
real adotado em diferentes países em diferentes épocas apresentou variações
significativas (de 10 a 400 dias) (Foster et al., 2006).
O conceito de proteção das fontes de abastecimento de água subterrânea
foi estabelecido já há muitas décadas, fazendo parte dos códigos jurídicos dos
países europeus, no trabalho realizado por Coelho e Duarte, (2003) são
apresentados os perímetros de proteção estabelecidos para alguns esses
países. No entanto, o dimensionamento de zonas de proteção de águas
subterrâneas está baseado em observações empíricas da eliminação de
microrganismos na água subterrânea.
Por exemplo, no caso da Nova Zelândia, segundo Pang et al. (2003), é
estabelecida de forma arbitrária, como proteção, uma distância que varia de 30 a
50 metros para a água subterrânea. Na Áustria, na Alemanha, e na Holanda, as
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

22
zonas de proteção de águas subterrâneas são estabelecidas através do cálculo
do tempo de fluxo até o poço, de 50 a 60 dias, período considerado que cobre o
tempo de sobrevivência de bactérias patogênicas e vírus. Na Suíça, onde os
processos físicos e físico-químicos são mais enfatizados, as áreas de proteção
são calculadas com base de 10 dias de tempo de fluxo ou pelo menos uma
trajetória de fluxo de 100 m de comprimento através do meio poroso (Pekdeger e
Matthess, 1983).
Na América Latina e no Caribe, de acordo com a Pan American Health
Organization (PAHO, 1998) a poluição das águas subterrâneas ameaça
diretamente a saúde de grandes segmentos da população. Como uma
estimativa, 50% das comunidades depende exclusivamente de águas
subterrâneas como fonte de água. E, no entanto, apesar da importância deste
recurso como fonte de água potável, a proteção de aquíferos não recebeu a
devida atenção na Região.
Conforme Toscano et al. 2008, no Brasil, sendo as águas subterrâneas
bens dos estados, não são objeto de leis federais que regulamentem seu uso.
No entanto, podem ser encontrados alguns dispositivos legais referentes, tais
como a Resolução 396 de 03 de abril de 2008 do CONAMA (Conselho Nacional
de Meio Ambiente) que prevê a implementação de Áreas de Proteção de
Aquíferos e Perímetros de Proteção de Poços de Abastecimento e de Áreas de
Restrição e Controle do Uso da Água Subterrânea. Apenas onze estados
brasileiros dispõem de legislação que criam áreas de proteção dos aquíferos e
áreas de proteção sanitária, não sendo sugeridos, em nenhuma legislação,
métodos de delimitação de áreas de proteção.
Finalmente, ao definir o nível de controle necessário para proteger a
qualidade da água subterrânea, vale mais a pena em termos de custo e será
menos prejudicial ao desenvolvimento econômico, utilizar a capacidade de
atenuação natural dos estratos de cobertura do aqüífero, em vez de aplicar ao
terreno controles universais sobre o uso do solo e sobre a emissão de efluentes
(Foster et al., 2006).
2.3. Sobrevivência de bactérias e vírus na água subterrânea
As oportunidades de contaminação da água subterrânea incrementam-se
devido à capacidade de sobrevivência dos microrganismos no meio
subsuperficial por longos períodos de tempo. Segundo Pekdeger e Matthess
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

23
(1983) dois grupos de microrganismos devem ser diferenciados quando é
considerada a sobrevivência de bactérias e vírus na água subterrânea; os
microrganismos autóctones e os microrganismos alóctones, estes últimos
podendo ser patogênicos ou produtores de enterotoxinas.
As bactérias aloctônicas ingressam dentro da água subterrânea devido à
contaminação e são eliminadas geralmente nesse ambiente. Durante a fase
inicial (fase Lag), sob condições estéreis e oligotróficas, podem sobreviver por
vários meses sem um decrescimento substancial ou ainda incrementar o número
de células, enquanto que, em águas contendo bactérias predadoras, esta fase
pode-se limitar a poucos dias (Matthess et al., 1988)
As enterobactérias, em geral, persistem no solo durante dois ou três
meses. Entretanto, já foi detectado tempo de sobrevivência maior que cinco anos
(Corapcioglu e Haridas, 1984). Estudos de laboratório reportados por Gerba et
al. (1975), citado por Abu-Ashour et al. (1993), indicam um tempo de
sobrevivência para a E. coli maior a 4,5 meses em água subterrânea mantida na
escuridão.
É assim que a sobrevivência de bactérias no solo, normalmente varia a
partir de umas poucas semanas até poucos meses. São muitos os fatores que
afetam a sobrevivência dos microrganismos nos ambientes subsuperficiais. A
Tabela 2.2 resume os efeitos individuais destes fatores sobre a sobrevivência de
bactérias e vírus em um ambiente subsuperficial.
Tabela 2.2. Fatores físico-químicos que afetam a sobrevivência de
bactérias entéricas e vírus no solo (Abu-Ashour et al., 1993)
Fator Impacto na sobrevivência
pH
Menor tempo de sobrevivência em solos ácidos (pH 3-5) do que em solos alcalinos
Conteúdo de água no solo
Maior tempo de sobrevivência em solos úmidos e durante períodos de chuvas fortes
Conteúdo de matéria orgânica
Aumento da sobrevivência e possibilidade de crescimento quando existe quantidade suficiente de matéria orgânica
Textura, tamanho e distribuição das partículas do solo
Principalmente solos finos, minerais de argila e sustâncias húmicas incrementam a retenção de água no solo que aumenta o tempo de sobrevivência
Temperatura Baixas temperaturas incrementam o tempo de sobrevivência
Disponibilidade de nutrientes Aumento do tempo de sobrevivência
Propriedades de adsorção Os microrganismos parecem melhorar sua persistência quando se encontram em estado de sorção
Luz solar Tempos de sobrevivência são curtos na presença de luz solar direta
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

24
Informação sobre as características de sobrevivência dos microrganismos
é geralmente referida como decaimento, eliminação ou inativação, que é a
destruição irreversível do contaminante através de processos físicos, químicos
ou biológicos. A eliminação de bactérias e vírus pode ser aproximadamente
descrita por uma função exponencial:
)tt(eCC o
o)t(
(2.1)
onde t ≥ t0 e t0 ≤ 7 dias; C0 e C é a concentração inicial e a concentração no
tempo t, e λ é a constante de eliminação calculada através da equação (2.2).
2/1
2ln
(2.2)
onde τ1/2 é a vida media do microrganismo, que varia geralmente entre 1 e 20
dias (Pekdeger e Matthess, 1983).
A equação (2.1) expressa que depois de uma eliminação muito rápida no
início, bactérias e vírus podem ainda existir em pequenas quantidades na água
subterrânea, mesmo depois de longos períodos (i.e. 6 meses). A constante de
eliminação depende de parâmetros físicos, químicos, e biológicos e é específico
para diferentes espécies microbianas, que corresponde a coeficientes de
decaimento, variando desde 0,035 até 0,69 d-1, baseado na equação (2.1)
(Richardson, 2002). Ainda não é possível prever com precisão necessária as
constantes de eliminação com base nos fatores de controle, que devem ser
determinadas para cada espécie e ambiente específico.
Normalmente o crescimento está incluído no coeficiente de decaimento,
resultando em um valor neto de decaimento (Yates e Yates, 1991). Enquanto
ocorre decaimento bacteriano, algum nível de crescimento acontece
simultaneamente. Durante condições de baixa disponibilidade de nutrientes, a
limitada energia disponível é aplicada à manutenção da célula para o
crescimento (Metcalf e Eddy, 1991). Em tal caso, o crescimento pode ser
considerado desprezível.
2.4. Escherichia coli
Para predizer a presença de patogênicos na água, normalmente é usado
um grupo específico de microrganismos, geralmente conhecidos como
organismos indicadores fecais. Os mais importantes indicadores microbianos de
contaminação fecal (poluição) usados são: Escherichia Coli e bactérias
coliformes termotolerantes. Ambos são amplamente utilizados, porque sua
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

25
detecção é simples, rápida, e confiável. A E. coli é o indicador preferido de
contaminação fecal, pois é o único membro do grupo coliformes termotolerantes
que é invariavelmente encontrada em fezes de animais de sangue quente e que
ultrapassa outros coliformes termotolerantes em ambos os excrementos
humanos e animais (Foppen e Shijven, 2006).
O gênero Escherichia é típico da família Enterobacteriaceae, que tem
como hábitat o intestino de humanos e animais. A Escherichia coli é um bacilo
Gram-negativo, anaeróbico, reto, com forma de bastão de aproximadamente
2,5 μm de comprimento e 0,8 μm de diâmetro e está presente em pares ou
isoladamente. Possui órgãos externos como os chamados pili, filamentos finos e
retos, que lhe permitem captar o substrato específico e um filamento mais fino,
helicoidal e longo denominado flagelo (um o mais) que lhe permite nadar (Berg,
2004). A Figura 2.2 apresenta a imagem de uma célula de E. coli, captada
através de microscopia eletrônica.
Figura 2.2. Micrografia eletrônica da E. coli. Escala: o corpo da célula é de
aproximadamente 1 μm de comprimento (Berg, 2004)
A maioria dos membros da espécie E. coli são considerados organismos
inofensivos, enquanto que algumas estirpes são causadoras de doenças. Três
síndromes clínicas gerais podem resultar de infecção com espécies (patótipos):
doenças entéricas/ diarréicas, infecções do trato urinário, e sepsis / meningite.
Estirpes patogênicas têm sido categorizadas dentro de seis grupos com base em
características sorológicas e de virulência (Kaper et al., 2004, citado por Foppen
e Schijven, 2006).
Geralmente, quando bactérias E. coli são introduzidas dentro de ambientes
aquáticos, morrem gradualmente e este processo é acompanhado por mudanças
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

26
em suas características. Daubner (1975), citado por Foppen e Schijven (2006),
realizou experimentos de sobrevivência utilizando estirpes frescas de E. coli em
água estéril demineralizada e água altamente mineralizada. Foi observado que
cinco a vinte horas depois que a E. coli é transferida na água estéril, ocorre
encolhimento da célula e redução do conteúdo citoplasmático (na água
destilada) ou dano à integridade da célula em meio mineralizado. A questão
obvia é descobrir qual o efeito dessas mudanças morfológicas e fisiológicas na
adesão bacteriana.
Vários estudos sobre a influência da hidrofobicidade sobre o processo de
adesão, concluíram que E. coli é pobremente adesiva e que é mais um
organismo hidrofílico do que hidrofóbico, e que a hidrofobicidade não determina
a adesão. Em vez disso, o modulador primário de aderência parece ser o
potencial zeta (Foppen e Shijven, 2006).
A carga superficial da E. coli não é medida diretamente, mas é
frequentemente determinada via o que é chamado de mobilidade electroforética,
que é uma media do potencial zeta (definido como a queda de potencial através
da parte móvel da dupla camada responsável pelo fenômeno eletrocinético
(Stumm e Morgan, 1996). Foppen e Shijven (2006) determinaram o potencial
zeta da E. coli ATCC 25922 em varias soluções e os resultados de sua pesquisa
indicaram que para soluções com eletrólitos monovalentes (Na+) e divalentes
(Ca++), com valores de força iônica variando desde 10-5 até 400 mM, na faixa de
pH de 7,19 – 8,81, o potencial zeta variou desde -20 até -170 mV.
No entanto, o potencial zeta é tipicamente aplicado a partículas esféricas
duras, e no caso de células biológicas, elas formam uma camada permeável,
macia de íons em torno da célula, que controla a distribuição espacial do
potencial elétrico da superfície. Além disso, o potencial zeta reflete as
propriedades eletrocineticas resultantes da célula e não é sensível a variações a
pequena escala. Portanto, misturar esta distribuição heterogênea de carga em
um valor de potencial zeta, pode resultar em previsões erradas das interações
da superfície da bactéria e o coletor (Foppen e Shijven, 2006).
Walker et al. (2005) demonstrou que valores de potencial zeta para E. coli
D21g coletadas no meio da fase de crescimento exponencial e na fase
estacionária em soluções para várias concentrações de KCl foram similares,
enquanto que a taxa de deposição bacteriana em colunas de areia de quartzo
limpo (com carga negativa) e força iônica moderada (0,003 – 0,03 M KCl), foi 10
vezes mais alta na fase estacionária do que no meio da fase exponencial. Isto
pode ser atribuído à distribuição não uniforme de carga (negativa) nas células da
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

27
fase estacionária e consequente menor repulsão e maior adesão, do que as
células da fase exponencial que estão cobertas por proteínas relativamente
simples.
Resumindo, a bactéria E. coli, frequentemente encontrada na água
subterrânea indicando contaminação fecal, é fácil de quantificar, é hifrofílica e
possui forte carga negativa. Estas propriedades fazem que esta bactéria seja
amplamente utilizada como indicador de contaminação fecal, especialmente em
países em desenvolvimento que não dispõem de recursos suficientes de
laboratório. Além do mais, algumas estirpes de E. coli são enteropatogênicas
(Foppen e Shijven, 2006).
2.5. Transporte de Biocolóides
Colóides são comumente definidos como partículas e outras entidades
com dimensões entre aproximadamente 1nm e 1µm (Kretzschmar et al., 1999)
(até 10 µm segundo Bradford et al., 2002). Esses limites de moléculas
dissolvidas por um lado e partículas maiores suspensas por outro são graduais,
e em certa medida arbitrários.
Devido a seu pequeno tamanho, os colóides têm área especifica muito
grande (>10m2/g) tornando-se eficientes coletores de contaminantes ambientais.
Partículas coloidais permanecem estáveis em suspensão por longos períodos de
tempo até que formem agregados maiores ou que se depositem sobre
superfícies de grãos maiores (Kretzschmar et al., 1999).
Muitos contaminantes ambientais são encontrados como colóides
(organismos patogênicos) ou estão associados a colóides (metais pesados,
radionuclídeos, pesticidas, hidrocarbonetos aromáticos policíclicos, etc.).
Portanto, o transporte de vírus e bactérias, chamados de “biocolóides”
(Kretzschmar et al., 1999) é abordado como um problema de transporte de
colóides.
Porém, pelo fato de se tratar de organismos vivos, o estudo do transporte
de microrganismos é mais complexo do que no caso de colóides abióticos. Não
só eles estão sujeitos aos mesmos fenômenos físico-químicos dos colóides, mas
há também um número de processos biológicos que afetam seu transporte (Ginn
et al., 2002).
O transporte de bactérias através de um meio poroso saturado, foco
principal do presente trabalho, é governado através de numerosas relações entre
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

28
solo, a bactéria e os mecanismos e as propriedades ambientais. Uma descrição
aproximada do transporte bacteriano como um todo, requer a compreensão do
impacto de cada fator na progressão da bactéria através do espaço de poro da
matriz de solo.
A descrição do transporte biocolóides pode ser formulada como um
problema geral de transporte de solutos. Para derivar uma equação que
descreva o transporte de microrganismos no meio poroso, um enunciado
matemático da conservação da massa deve ser formulado, ou seja, a quantidade
do material deixando o sistema deve ser igual à quantidade de material entrando
no sistema, mais ou menos (±) qualquer mudança no armazenamento.
bacteriana
massa de perda
ou Ganho
VEno entrando
bacteriana
Massa
VEdo saindo
bacteriana
Massa
VEno bacteriana
massa de v ariação
de neta Taxa
(2.3)
A equação (2.3) representa o esquema do movimento do contaminante
através de um volume elementar (VE) e mudanças na quantidade do
contaminante. Assume-se que o meio poroso é homogêneo, isotrópico,
incompressível, e totalmente saturado (Yates e Yates, 1991).
Corapcioglu e Haridas (1984, 1986) foram os primeiros em desenvolver um
modelo matemático específico para o transporte de vírus e bactérias através do
meio granular poroso. Este modelo inclui os processos de advecção, dispersão
hidrodinâmica e um número de mecanismos associados à retenção de células
bacterianas no meio poroso, como também os efeitos de crescimento e
decaimento tanto dos microrganismos livres como adsorvidos.
A equação de balanço de massa macroscópico da equação (2.3) para
solutos adsorvidos (microrganismos) fluindo através de um meio poroso, na
forma unidimensional pode ser expressa como:
F2
2
hbulk R
x
Cv
x
CθD
t
θC
t
Sρ
(2.4)
onde C é a massa (ou número) de microrganismos na fase aquosa para uma
distância x e tempo t; S denota a massa (ou número) de microrganismos
adsorvidos por unidade de massa da parte sólida do meio poroso; Dh é o
coeficiente de dispersão hidrodinâmica; v é a velocidade intersticial do fluxo; ρbulk
densidade total seca do meio poroso; θ representa o volume ocupado pela
suspensão por unidade de volume total e RF a taxa de consumo (ou decaimento)
dos microrganismos.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

29
Neste trabalho, o estudo está focado no transporte da bactéria, eliminando
as complexidades derivadas dos processos biológicos, tais como crescimento,
morte e predação, restringindo a bactéria a seu estado estacionário (sem
crescimento) durante períodos de tempo para os quais o decaimento não é
observado e o sistema é desprovido de predadores.
Assim, desprezando os efeitos dos processos biológicos, para um meio
poroso homogêneo e saturado (θ=n), a equação (2.4) na forma unidimensional
pode ser simplificada como se apresenta a seguir:
x
Cv
x
CD
t
C
t
S
n 2
2
hbulk
(2.5)
Enquanto os microrganismos são transportados através do meio poroso,
eles são removidos do fluido dos poros por meio de filtração físico-química
(adsorção nas superfícies do grão) representado pelo termo tnSbulk / .
2.6. Processos de Transporte
Como foi explicado previamente, no item 2.5, os microrganismos como
vírus e bactérias são considerados, por suas dimensões e comportamento, como
colóides, mais especificamente biocolóides. Devido à complexidade, que implica
o estudo do efeito combinado dos fenômenos físico-químicos e biológicos sobre
o transporte de organismos vivos nos ambientes subsuperficiais, nesta seção
são estudados os processos de transporte, sem considerar os processos
biológicos em um meio poroso saturado. Dessa forma os principais processos
abordados são; a advecção e a dispersão, processos básicos que controlam o
transporte de solutos e de partículas coloidais suspensas; a filtração mecânica
(straining), que descreve a remoção de colóides baseado no tamanho físico; a
adsorção, resultante de várias forças de origem eletrostática entre o colóide e o
grão, quando a distância entre eles é pequena (da ordem dos nanômetros) e
finalmente, processos como a exclusão, fenômeno observado principalmente no
caso de vírus (Knapett et al., 2008) e o bloqueamento e ripenning, fenômenos
referidos às variações da taxa de deposição com relação ao tempo.
2.6.1. Advecção
Advecção é o processo pelo qual um contaminante é transportado junto
com a massa de fluxo da água subterrânea (Freeze e Cherry, 1979), sendo a
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

30
taxa média de migração de um soluto igual à velocidade linear média da água
(Fitts, 2002). Assim, a massa de fluxo de um soluto devido unicamente a
advecção é simplesmente:
CqF xax (2.6)
onde Fax é o fluxo de massa de soluto por advecção (massa/tempo/área normal à
direção x), qx é a descarga específica (volume/tempo/área) e C a concentração
do soluto (massa/volume).
Nesse sentido, para quantificar as forças de advecção presentes em um
sistema dado, é preciso determinar a velocidade da água sendo transportada
através do meio poroso. As velocidades da água subterrânea em aquíferos
porosos são geralmente menor do que um metro/dia e até poucos metros/dia.
Velocidades acima 10 m/d são restritas a sedimentos muito grossos e elevados
gradientes hidráulicos (Pekdeger e Matthess, 1983).
Em uma configuração a escala de laboratório, a descarga específica qx é
conhecida, e com base na porosidade do sistema n, a velocidade intersticial v
pode ser calculada através da seguinte relação:
n
qv x (2.7)
Na maioria dos ambientes subsuperficiais, advecção é considerada como o
principal mecanismo de transporte bacteriano (Peterson e Ward, 1987; citado por
Richardson, 2002). Corapcioglu e Haridas (1986) no estudo do transporte de
microrganismos incluíram dentro do termo de advecção, a velocidade de
sedimentação e a velocidade devido à quimiotaxia.
2.6.2. Dispersão Hidrodinâmica
A distribuição real da velocidade das águas subterrâneas varia
significativamente tanto no espaço quanto no tempo, em decorrência da não
uniformidade da velocidade de fluxo e consequentemente a massa de fluido
tende a se espalhar ou dispersar. Este processo é conhecido como dispersão
mecânica (Fitts, 2002).
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

31
Estas variações na velocidade do fluido são causadas pela resistência ao
longo das paredes dos canais de poros, mudanças da seção transversal dos
canais do poro e da tortuosidade (Frezze e Cherry, 1979).
No transporte real de um soluto na água subterrânea existem dois
processos básicos: a dispersão mecânica e a difusão molecular, ambas as quais
contribuem para uma dispersão transversal (Bear, 1972; Fitts, 2002).
A difusão molecular é a mistura que ocorre como consequência do
movimento aleatório das moléculas dentro do fluido. As moléculas que colidem
ou se impulsionam dentro do liquido tendem, ao longo do tempo, a espalhar os
solutos de modo que as concentrações de soluto se tornam mais uniformemente
distribuídas no espaço. Como outros colóides, bactérias e vírus estão sujeitos ao
movimento Browniano, através do qual a trajetória da partícula individual parece
bastante errática, enquanto o fluxo médio de partícula é proporcional ao
gradiente de concentração (Corapcioglu e Haridas, 1986).
Assim, a difusão move a massa de soluto de regiões com altas
concentrações para regiões com baixas concentrações segundo a primeira lei de
Fick (Fitts, 2002), que para um meio poroso saturado é governada pela equação
seguinte:
x
CnDF mdx
(2.8)
onde Fdx é o fluxo de massa de soluto por difusão (massa/tempo/área) e Dm é a
constante denominada coeficiente de difusão molecular. O coeficiente de difusão
para partículas suspensas (bactérias e vírus) pode ser estimado através da
equação de Stokes-Eistein (Corapcioglu e Haridas, 1986).
O fluxo de um soluto em uma direção qualquer é proporcional ao gradiente
de concentração nessa direção. O sinal negativo na equação (2.8) se deve a que
a massa de soluto se movimenta na direção que as concentrações decrescem,
na direção oposta ao gradiente de concentração.
Os efeitos da dispersão mecânica adicionada à difusão molecular
normalmente são agrupados no que é chamado de dispersão hidrodinâmica. O
coeficiente de dispersão hidrodinâmica Dh é definido pela seguinte equação:
mLh DvD (2.9)
onde αL é o coeficiente de dispersividade, v a velocidade da água subterrânea,
Dm a difusão molecular e τ a tortuosidade.
O coeficiente de dispersividade αL é uma função da falta de
homogeneidade do aquífero, assim este coeficiente cresce com a escala dos
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

32
experimentos. O meio poroso usado em experimentos de laboratório tem
coeficientes de dispersividade na ordem de 0,1 cm-1 m, em experimentos de
campo na ordem de 0,1 a 100 m, e em rocha cárstica e fissurada 10 – 1000 m
(Pekdeger e Matthess, 1983).
Tanto o coeficiente de dispersão hidrodinâmica como a dispersividade, não
são verdadeiras propriedades físicas do meio poroso, se trata de parâmetros de
ajuste que permitem ao modelo matemático, simular a dispersão de solutos de
um modo mais ou menos similar ao que ocorre dentro de um sistema real (Fitts,
2002).
A significância relativa da difusão em relação à dispersão mecânica pode
ser estimada utilizando um parâmetro adimensional conhecido como número de
Peclet (Pe) para a difusão molecular:
m
eD
vdP (2.10)
onde d é o comprimento médio característico neste caso o diâmetro médio do
grão do meio poroso. Para valores muito baixos de Pe (<0,4) unicamente a
difusão governa o processo de transporte. Para valores entre 0,4 e 5
(aproximadamente) a influência da difusão molecular e a dispersão mecânica
são da mesma ordem de grandeza. Para valores superiores a 5 a propagação do
soluto é dominada pela dispersão mecânica (Bear, 1972).
Pekdeger e Matthess (1983) consideraram que a mobilidade ativa da
bactéria pode ser representada como uma força dispersiva e incluída como uma
componente da dispersão hidrodinâmica, que decresce ao diminuir as
temperaturas (i.e. para E. coli 0,1 m/d a 20°C).
2.6.3. Adsorção
A adsorção, também denominada como filtração físico química (McDowell-
Boyer et al., 1986), é um processo reversível ou irreversível em que a célula
bacteriana é sujeita nas superfícies do solo por várias forças físico-químicas
(carga superficial, forças eletrostáticas, e forças de London-vander Waals)
(Yates e Yates, 1991).
Consequentemente, a adsorção da bactéria na superfície dos grãos é
função das propriedades físicas e químicas do meio e da água dos poros,
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

33
características da bactéria, e condições hidráulicas do sistema (Corapcioglu e
Haridas, 1984).
Propriedades do meio tal como tamanho do grão, carga superficial, e
estrutura dos poros (i.e. presença de macroporos) podem ter um efeito
significativo no grau de adsorção da bactéria. Em geral, solos de textura fina tais
como argilas, são muito efetivos na retenção de bactérias, em decorrência do
seu pequeno tamanho, grande área superficial específica e carga superficial
(Corapcioglu e Haridas, 1984). Contrariamente, meios com grãos de tamanho
maiores tais como areias, têm baixo potencial de adsorção bacteriana (Peterson
e Ward, 1989 citado em Richardson, 2002). O meio poroso, opostamente
carregado à bactéria que reside, tende a estimular elevados níveis de adsorção
de bactérias. Scholl et al. (1990) encontraram maior adesão de bactéria indígena
(com carga superficial negativa) às superfícies de meio positivamente carregado
(i.e. calcita, quartzo e moscovita cobertos de hidróxidos de ferro e moscovita) do
que no caso de meio negativamente carregado (i.e. quartzo limpo e moscovita).
A estrutura do poro da matriz do solo pode também influenciar o grau de
adesão bacteriana. Sistemas naturais de solo são raramente, ou nunca,
homogêneos em todas as direções, apresentando um grande número de
heterogeneidades e macroporos. A recompactação de solos diminui a presença
de macroporos dentro da estrutura do solo obrigando à bactéria a se infiltrar nos
espaços de poros menores, assim aumentando o potencial de adsorção
bacteriana (Richardson, 2002).
Um elevado conteúdo de sal na água subterrânea poderia incrementar a
adsorção devido à compressão da dupla camada elétrica, permitindo às células
bacterianas se aproximar dos locais carregados no meio. O incremento na
disponibilidade de íons na solução promove a formação de pontes entre o meio
carregado e as superfícies das células bacterianas (Fontes et al., 1991).
Também pode se incrementar a adsorção reduzindo o pH por baixo de 8.0 ou
adicionando cátions, especialmente de espécies divalentes (Corapcioglu e
Haridas, 1986).
A forma e tamanho de uma dada célula bacteriana podem influenciar a
adsorção nas proximidades dos grãos do meio. Em geral, diâmetros maiores
experimentam maiores interações (colisões) com o meio, aumentando assim a
possibilidade de adesão e elevadas velocidades de poros tendem a reduzir as
potenciais oportunidades de contato das bactérias suspensas nas superfícies
dos grãos (Richardson, 2002).
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

34
A filtração físico-química tem sido modelada tanto como um processo
irreversível quanto reversível em função da análise do termo tnSbulk / da
equação (2.5). No caso do processo reversível, podem ser aplicados tanto
mecanismos de equilíbrio quanto mecanismos cinéticos, descritos a seguir.
2.6.3.1. Sorção de equilíbrio
A adsorção de células bacterianas na superfície dos grãos pode ser
representada através de isotermas de equilíbrio (isoterma de Freundlich ou de
Langmuir), que descrevem a relação entre a concentração de bactérias
adsorvidas S e a concentração de bactérias suspensas C (Domenico e Schwartz,
1990). Assumindo que a adsorção de partículas é um processo que atinge o
equilíbrio rapidamente, pode ser descrito através da forma linear da isoterma de
Freundlich CKS D , expressão que derivada com relação ao tempo, pode ser
facilmente incorporada na equação de transporte equação (2.5) através da
seguinte expressão:
t
CK
t
SD
(2.11)
onde KD é o coeficiente de distribuição ou partição (L3/M) e pode ser obtido
empiricamente em ensaios em batelada no laboratório. Uma vez substituída a
equação (2.11) na equação (2.5), pode-se expressar o fator de retardamento R
como segue:
nK1R bulk
D
(2.12)
Desta forma a equação de transporte é simplificada na forma seguinte:
x
Cv
x
CD
t
CR
2
2
h
(2.13)
A representação linear da adsorção bacteriana implica que está presente
uma única capa de adsorção, onde só as células bacterianas podem ocupar um
local de adsorção dado, a tempo determinado.
A adsorção à superfície do grão impede a migração da bactéria, forçando
assim que ela seja transportada mais lentamente do que a água em torno dos
poros. O termo “retardamento” é geralmente usado para descrever o impacto da
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

35
adsorção sobre o transporte global (Yates e Yates, 1991). Ainda fatores
adicionais (i.e. filtração mecânica e sedimentação) podem também ser incluídos.
Experimentos de campo têm registrado fatores de retardamento entre 1 e 2
para bactérias (E. coli e Serratia marcescens) (Pekdeger e Matthess, 1983).
Muitos outros estudos revelaram fatores de retardamento menores que 1 (Fontes
et al., 1991), sugerindo velocidades bacterianas mais elevadas que a dos
traçadores introduzidos simultaneamente. Este fenômeno pode ser explicado
através da teoria de exclusão, que será visto no item 2.6.5.
Os vírus em geral, especialmente os poliovírus, têm valores elevados de
KD (até 500), dependendo das propriedades da água e as propriedades dos vírus
respectivos. Assim, a passagem subterrânea pode proporcionar uma proteção
muito efetiva contra a contaminação de vírus. Porém os vírus podem ser
novamente desorvidos, quando as concentrações dos cátions decrescem (i.e.,
devido a uma chuva intensa) sendo assim ativados para maior deslocamento
(Pekdeger e Matthess, 1983).
2.6.3.2. Sorção Dinâmica
Em um sistema composto de microrganismos em suspensão e grãos de
solo, um estado de sorção de equilíbrio não é atingido instantaneamente.
Inicialmente, um mecanismo de sorção cinética em duas etapas controla a
remoção de microrganismos da fase aquosa. No primeiro passo, descrito como
transporte da massa, os microrganismos são transferidos a partir da massa de
fluido para a superfície dos grãos de solo. Em um segundo passo, os
microrganismos são aderidos sobre a superfície como resultado de interações
físico-químicas (Tufenkji, 2004; Kretzschmar et al., 1999). O subsequente
desprendimento pode ser também controlado por processos cinéticos. Sob estas
condições, a taxa de modificação da concentração dos microrganismos retidos
pode ser descrita do seguinte modo:
Skn
Ckt
S
ndet
bulkatt
bulk
(2.14)
onde katt e kdet representam as taxas constante de adsorção e desorção
respectivamente, (T-1). A equação anterior representa um modelo cinético de no-
equilibrio (Jiang et al., 2007).
A aproximação mais frequentemente utilizada em estudos de laboratório e
campo para avaliar o transporte de microrganismos, é a teoria clássica de
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

36
filtração CFT (Colloid Filtration Theory). Na realidade, este modelo de filtração
denominado “clean-bed” ou leito limpo, trata-se de um caso especial do processo
cinético de adsorção anteriormente explicado, mediante o qual a adesão dos
microrganismos sobre as superfícies dos grãos é considerada irreversível,
desprezando o processo de desprendimento (Tufenkji, 2007). Desta forma a
equação (2.14) se reduz como segue:
Ckt
S
natt
bulk
(2.15)
De acordo com este modelo de transporte, assume-se que a cinética de
deposição de colóides (bactérias), acontece de forma irreversível em duas
etapas:
Primeira etapa; as partículas coloidais são transportadas na direção das
superfícies dos grãos mediante difusão Browniana, intercepção, e
sedimentação resultando em colisões do colóide contra o coletor.
Segunda etapa; se produz a adesão de partículas coloidais nas superfícies
dos grãos em decorrência da ação de forças inter-partículas que incluem,
forças de atração de vander Waals e forças repulsivas da dupla camada
elétrica, esta última fortemente afetada pela composição química da
solução e a carga das superfícies (Kretzschmar et al., 1999).
A cinética da etapa de transporte, que depende primariamente de fatores
físicos foi inicialmente descrita por Yao et al. (1971). Considerando inicialmente
uma única partícula do meio filtrante, esférica, chamada de “coletor”, fixada na
suspensão sem ser afetada pelas partículas vizinhas como é mostrado na Figura
2.3, onde também se mostra a trajetória de uma partícula suspensa (biocolóide),
que pode ser retida sobre o coletor (grãos de areia) devido a fenômenos físicos
de difusão, sedimentação e intercepção.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

37
Figura 2.3. Mecanismos básicos de transporte em filtração de água
(Adaptado de Yao et al., 1971)
Para a obtenção da taxa de adesão katt, inicialmente deverá ser
determinada a eficiência do coletor η, definida como a relação entre a taxa na
qual as partículas colidem com o coletor, e a taxa na qual as partículas fluem na
direção da área projetada do coletor como mostra a equação (2.16), onde dg é o
diâmetro médio do grão (coletor), v a velocidade intersticial e Co a concentração
das partículas na suspensão (Yao et al., 1971).
4
dvC
2g
o
colidem partículas ás qual na taxa (2.16)
De acordo com Yao et al. (1971), a eficiência global do meio filtrante para
um filtro granular composto por esferas uniformes, obtida a partir de um balanço
de massa, é relacionada com a eficiência do coletor η, e através da seguinte
relação:
Ld2
)n1(3
C
Cln
go
(2.17)
onde L é a profundidade do meio granular filtrante, n é a porosidade, dg o
diâmetro médio do grão (coletor), C e Co as concentrações do influente e do
efluente da coluna respectivamente. A equação (2.17) pode ser usada para
estimar a distância da viagem de colóides em um meio poroso saturado tal como
os aquíferos de água subterrânea.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

38
O objetivo da teoria de filtração (CFT) é determinar η sob condições
físicas e químicas conhecidas. Porém, devido à presença de forças repulsivas
decorrentes da dupla camada elétrica, nem todas as colisões resultam em
adesão de partículas sobre o coletor e, consequentemente se faz necessário
combinar um fator empírico na predição de η. Neste sentido, a eficiência do
coletor teórica denominada como ηo, determinada com base a fatores físicos
como será indicado mais adiante, é multiplicada pelo fator de eficiência de
colisão α, como indica a equação seguinte:
o (2.18)
A eficiência de colisão α para uma dada suspensão coloidal e meio filtrante
pode ser determinado através de ensaios de coluna e varia entre 10-3 e 1 (Ryan
e Elimelech, 1996).
Yao et al. (1971) descreveram quantitativamente os processos incluídos na
teoria da filtração (CFT), considerando que estes atuam na direção principal de
fluxo (a direção da força gravitacional), sendo os colóides transportados a partir
da massa de água para a superfície externa dos grãos. Estes processos são
descritos a seguir:
Difusão: torna-se mais importante como um mecanismo de transporte para
organismos patogênicos pequenos tais como vírus comparados com outros
maiores, como se pode observar na equação (2.19), que descreve o
movimento Browniano. Esta equação é conhecida como a equação de
Einstein (Yao et al., 1971).
d3
kTDg
(2.19)
onde Dg é a difusividade do grão (coletor) na água, k é a constante de
Boltzman (1,38048x10-23 J/K), T é a temperatura absoluta (K), μ é a
viscosidade dinâmica da água (1,002 x 10-3 N·s/m2), e d é o diâmetro do
biocolóide. A efetividade da difusão se incrementa ao decrescer o tamanho
da partícula.
Intercepção: inversamente à difusão que tem um grande efeito sobre
organismos patogênicos menores, a intercepção baseia-se na tendência
de objetos a permanecerem em movimento, uma propriedade pertencente
a colóides maiores (Richardson, 2002). A probabilidade de colisões que
ocorrem devido à intercepção pode ser descrita pela relação abaixo:
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

39
go
d
d (2.20)
A equação (2.20) mostra que quanto maior a relação entre o diâmetro da
partícula ou biocolóide d com relação ao diâmetro do coletor dg tem-se
maior probabilidade de colisão (Ryan e Elimelech, 1996). Assume-se que
tanto a partícula quanto o coletor são perfeitamente esféricos. Também é
assumida que para uma distância infinita entre o coletor e a partícula, a
concentração C é igual à Co, e C igual a zero para uma distância (d+dg)/2
medida desde o centro do coletor esférico (Yao et al., 1971).
Sedimentação: como o processo de intercepção, a sedimentação também
é proporcional ao tamanho de patogênico enquanto a difusão é
inversamente proporcional. Isso significa que a difusão tenderá a ser o
processo de transporte dominante para patogênicos de menor porte e a
sedimentação e a intercepção dominará o transporte dos maiores.
A sedimentação é o processo de movimento descendente devido à
diferença de densidade e é descrito através da Lei de Stoke’s:
18
gd)(v
2f
s
(2.21)
onde vs é a velocidade de sedimentação, ρ e ρf as densidades da partícula
e do fluido respectivamente, d o diâmetro médio do biocolóide e μ a
viscosidade dinâmica do fluido.
A Lei de Stoke’s assume que a partícula é esférica, macia e rígida, sendo
essas presunções não inteiramente válidas para microrganismos.
Este mecanismo é muito importante na acumulação de uma suspensão de
mineral inorgânico (densidade aproximadamente de 2,5 g/cm3), mas não
para microrganismos, que apresentam tamanho de partícula menor do que
5 μm e densidade aproximadamente de 1 g/cm3 (Pekdeger e Matthess,
1983).
Os três processos citados acima (difusão, interceptação e sedimentação),
que contribuem à eficiência de colisão têm sido integradas dentro de uma
equação por Yao et al. (1971) e pode ser aplicada para a filtração inicial “clean
bed” ou leito limpo, onde a deposição no interior dos poros não altera
significativamente o padrão de fluxo e as características do meio (Yao et
al.,1971). Esta equação incluindo a correção de Happel pode ser escrita como
segue (Hornberger et al., 1992):
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

40
v18
gd
d
dA
2
3
dvd
kTA90
2f
2
g
s
3
2
g
31sGIDo
)(, /
(2.22)
Na equação acima, ηD, ηI, ηG corresponde aos processos de difusão,
intercepção e sedimentação respectivamente e As é o fator de correção de
Happel que depende da porosidade de acordo com as equações seguintes:
65
5
sp2p3p32
p12A
)( (2.23)
31n1p /)( (2.24)
É também importante salientar que a equação (2.22) foi deduzida para um
coletor único e perfeitamente esférico. Além disso, assume-se que o diâmetro do
coletor não muda o que sim, de fato, ocorre devido ao acúmulo de colóides na
superfície dos grãos e a formação de biofilme (ou bio-fouling) (Knappett, 2006).
Para a determinação da taxa de adesão irreversível katt baseada na CFT,
considerada como um caso especial do processo geral de adsorção cinética, o
sistema é assumido como estacionário, inicialmente livre de microrganismos e a
influência da dispersão hidrodinâmica desprezível (i.e. o termo de dispersão e
pequeno comparado com o termo de advecção, que pode ser estimado através
do valor do número de Peclet (Pe) como foi explicado no item 2.6.2. A solução
para a equação (2.5) sob estas condições, para uma injeção contínua do
biocolóide com uma concentração Co (x=0) em um período de tempo t, e
considerando a relação dada na equação (2.15), está dada pela equação
seguinte:
x
v
kexpCC att
o (2.25)
onde a taxa de adesão katt dos microrganismos está relacionada com η e α,
através da equação (2.17) como segue:
g
attd2
v)n1(3k
(2.26)
É importante apontar que tanto η quanto a taxa de adesão katt podem ser
determinadas diretamente a partir da curva de chegada, seja com o experimento
de fonte contínua ou fonte pulso, considerando o valor de C/Co para o qual a
curva de chegada torne-se estável, através das seguintes equações:
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

41
oC
Cln
L)n1(
gd
3
2n (2.27)
o
attC
Cln
L
vk (2.28)
Como foi exposto anteriormente o valor da eficiência de colisão α depende
principalmente das condições químicas do sistema. Uma vez que a colisão
acontece, à adesão dependerá do efeito da força resultante da interação entre
partículas (força de atração ou repulsão).
Esta força ou potencial de energia total depende da distância entre
partículas e pode ser calculada como a soma das forças interpartículas tais
como, forças de atração de van der Waals e forças de repulsão eletrostática
(Verwey e Overbeek, 1948; citado por Kretzschmar et al., 1999).
A teoria de energia DLVO (Derjaguin and Landau, and Verwey and
Overbeek) estuda, de forma quantitativa, a estabilidade coloidal e foi
desenvolvida para explicar a coagulação rápida de colóides, observada em
presença de uma concentração eletrolítica alta e mais lenta, no caso de uma
baixa concentração, com uma transição de uma para a outra, conhecida como a
concentração crítica de coagulação (Gwyn, 2003). Desta forma a teoria DLVO
explica as alterações de energia que acontecem entre microrganismos e
superfícies sólidas e envolve uma estimativa da energia potencial de atração e
de repulsão, versus a distância entre partículas Essas quantidades são
mostradas na Figura 2.4 e descritas embaixo. Porém, a teoria DLVO assume a
superfície do colóide como lisa a nível molecular, sólida e inerte. Assim, as
inadequações desta teoria são sustentadas através de medições diretas das
forças de interações entre a bactéria e as superfícies sólidas através de
microscopia atômica (Ginn et al., 2002).
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

42
Figura 2.4. Teoria de energia DVLO. As forças combinadas de atração e
repulsão governam a agregação de partículas com relação à distância
entre elas (Gwyn, 2003)
As partículas sólidas de um aquífero estão quase sempre carregadas
negativamente. Uma exceção são os hidróxidos de ferro e de manganês e
substâncias orgânicas com baixos valores de pH. Bactérias e vírus, geralmente
carregados negativamente, são fortemente adsorvidos por adsorventes
aniônicos e fracamente por adsorventes catiônicos. Partículas carregadas
negativamente permanecem estáveis em suspensão em filtros de areia,
conforme as forças eletrostáticas de repulsão sejam mais fortes que as forças de
van Der Waals. Os cátions dissolvidos na água reduzem as forças repulsivas das
superfícies dos grãos. Cátions monovalentes são adsorvidos pela substância
sólida diminuindo sua carga negativa. Sob essas condições, as forças da massa
são mais efetivas e a acumulação de partículas acontece. Cátions bivalentes
podem também causar deficiência de carga positiva de modo que as forças
eletrostáticas podem ser mais eficientes para adsorção de bactérias e vírus
(Pekdeger e Matthess, 1983).
As bactérias podem se aderir ativamente e irreversivelmente sobre as
superfícies dos materiais sólidos dos aquíferos. As bactérias aderidas ficam
protegidas contra outras influencias e encontram altas concentrações de
nutrientes, decrescendo a taxa de eliminação. A adesão é mais intensiva na fase
de crescimento exponencial e pode ser de importância imediatamente depois da
intrusão da água poluída (Pekdeger e Matthess, 1983).
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

43
2.6.4. Filtração mecânica (straining)
A filtração é um mecanismo que pode limitar a migração de partículas.
McDowell-Boyer et al. (1986) consideram três formas de filtração, em função da
relação entre o tamanho da partícula e o tamanho do grão do meio poroso:
1. No caso de partículas ou agregados de partículas de tamanho comparável
ou maior do que o diâmetro médio do grão, a penetração dentro do meio
não acontecerá, mas sim a formação de uma membrana superficial sobre o
meio, conduzindo a uma diminuição na condutividade hidráulica, como
ilustra a Figura 2.5 (a). Esta forma de filtração é denominada em Bradford et
al. (2002) como “complete straining”.
2. O mecanismo de retenção de partículas dentro do meio poroso denominado
como “incomplete straining” (Bradford et al., 2002) ou simplesmente
“straining” (Corapcioglu e Haridas, 1986), acontece quando as partículas são
pequenas o suficiente para entrar no meio poroso sendo capturadas em
espaços de poros menores, como mostrado na Figura 2.5 (b). A retenção
ocorrerá em canais de poro e/ou uniões entre grãos que sejam menores que
uma dimensão crítica (Bradford e Bettahar, 2006).
3. Finalmente, a filtração físico química ou adsorção, processo que foi já
descrito no parágrafo 2.6.3 se refere à colisão e posterior retenção de
colóides em decorrência de forças físico químicas. Este mecanismo tem
maior importância quando o tamanho do colóide é pequeno em relação ao
tamanho de poro do solo (Bradford et al., 2002).
(a) (b) (c)
Figura 2.5. Formas em que a filtração limita a migração das partículas, a)
Filtração completa, b) Filtração incompleta ou straining, e c) Filtração físico
química (McDowell-Boyer et al., 1986)
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

44
Esta seção está focada no estudo do mecanismo de filtração incompleta
ou straining que, daqui por diante, será referido como filtração mecânica (termo
utilizado em Matthess et al., 1988 e Corapcioglu e Haridas, 1984).
Herzig et al. (1970) citado em Foppen et al. (2005) estimaram o potencial de
filtração mecânica utilizando relações geométricas entre o diâmetro do colóide e
o tamanho médio do grão (d/dg) assumindo os colóides e os grãos esféricos, a
través da equação seguinte:
1
2
d
d1
2
d
dZ)n1(
2
1
gg
(2.29)
onde σ(-) representa o volume de partículas que poderiam ser retidas, por
unidade de volume do total do meio poroso, n a porosidade inicial, Z é o número
de coordenação ou número de contatos entre os grãos do meio poroso. Em
função aos valores de n=0,4 e Z=7 (considerando um arranjo ortorrômbico de
esferas) foi determinado que quando a relação d/dg supera o valor de 0,05 a
filtração mecânica tem uma grande influência no processo de retenção de
partículas (citado por Corapcioglu e Haridas, 1984; Foppen et al., 2005; Bradford
et al., 2006). No entanto, observações experimentais recentes indicaram que a
filtração mecânica se dá para relações muito mais baixas, i.e. 0,002, valor citado
por Tukenkji et al. (2004) e Tufenkji (2007).
Considerando a passagem contínua de uma suspensão de bactérias
através de uma coluna de meio poroso, Foppen et al. (2005) estabeleceram que
o tempo t necessário para preencher a porção de volume de poros destinado à
deposição de bactérias σ, depende da concentração inicial Co a través da
seguinte relação:
o
bact
o Cnt
t
(2.30)
onde to é o tempo requerido para a passagem de um volume de poro (PV), (t/to)
o número de volume de poros que permitirá o preenchimento dos espaços
destinados à filtração mecânica e ρbact a densidade bacteriana (g/mL).
No entanto, em geral o solo não é composto de grãos esféricos e a
distribuição de tamanho não é uniforme. Nesse sentido Foppen et al. (2005)
propõem a utilização da função de densidade de poro f(r) dada na equação
(2.32), sendo r o radio do poro (m) e d o diâmetro do biocolóide. Esta função
permite uma estimativa mais aproximada de σ na equação (2.31). Como
resultado deste estudo Foppen et al. (2005) concluíram que a equação (2.29)
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

45
ainda parece proporcionar uma base valiosa para avaliar a importância da
filtração mecânica e para calcular o volume disponível para este processo.
drrfd
2
0 (2.31)
A distribuição de tamanho de poro pode ser determinada no laboratório
através da curva de retenção. Contudo, foi proposta uma solução baseada na
função de retenção solo-água de Van Genuchten (1980) (citado em Foppen et
al., 2005) para a determinação da função de densidade de poro através da
expressão seguinte:
)()()(
1mn1n
vv
vvv Ar1Arnmrf
(2.32)
e
vn
w
v
g
2A
cos (2.33)
onde mv, nv e αv são chamados parâmetros de van Genuchten e dependem das
propriedades do solo, δ é a tensão superficial da água 0,0728 N/m a 20°C (valor
extraído de Myers, 1999), β o ângulo de contato na interface que para o caso de
areia saturada é zero, ρw densidade da água (1,0 g/cm3) e g a aceleração da
gravidade (9,81 m/s2).
Foppen et al. (2005) e Foppen et al. (2007) incluem a filtração mecânica
na equação geral de transporte ADS, assumindo que as bactérias são retidas na
parte sólida do meio poroso em duas frações, uma delas está disponível para a
deposição devido à filtração mecânica (S2) e outra fração para a retenção
reversível (S1). Desta forma, a equação para a retenção de bactérias pode ser
expressa como:
t
S
t
S
t
S 21
(2.34)
1att
bulk
1 SkCkn
t
Sdet
(2.35)
Cn
kt
S
bulk
str2
(2.36)
Foppen et al. (2005) observaram experimentalmente que o coeficiente de
filtração mecânica Kstr é função da velocidade de fluxo v, além da relação d/dg
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

46
antes indicada por Bradford et al. (2003), através da seguinte equação de ajuste
onde a e b são parâmetros de ajuste:
vd
dak
b
g
str
(2.37)
Considerando o processo de filtração mecânica irreversível, sob
condições do meio poroso livre de microrganismos (clean bed), o valor da
dispersão hidrodinâmica desprezível a taxa de deposição devido à filtração
mecânica denominada como kstr, pode ser considerada igual a katt estimada
através da teoria clássica de filtração CFT do colóide (citado por Tufenkji, 2007).
Porém, estudos experimentais indicam que esta aproximação resulta inválida no
caso que o sistema apresenta condições desfavoráveis à adesão (Martin et al.,
1996; Kretzschmar et al., 1999; Tufenkji e Elimelech, 2004; Tufenkji, 2007).
É importante salientar que as equações apresentadas não consideram o
potencial de crescimento de um biofilme ou a formação de aglutinações de
bactérias. Em geral, as bactérias não estão presentes na solução como
partículas individuais, mas podem ser aderidas aos sólidos suspensos.
(Mc Dowell–Boyer et al., 1986). A formação de flocos de bactérias aumenta o
impacto da filtração mecânica reduzindo a relação do diâmetro do meio poroso e
o diâmetro da bactéria (Richardson, 2002).
2.6.5. Exclusão
Denomina-se exclusão ao processo físico que se refere ao incremento da
velocidade de transporte de um colóide ou biocolóide em relação à velocidade
média da água, envolvendo um incremento da taxa de transporte devido ao
tamanho ou carga eletrostática do material transportado (Ginn et al., 2002).
Com relação aos processos que originam a exclusão em Ginn et al. (2002)
são identificados o efeito aniônico e o efeito do tamanho. A exclusão aniônica
acontece em escala molecular, devido a cargas eletrostáticas repulsivas entre o
meio poroso e o soluto, sendo direcionado a se afastar das paredes de poro.
Este efeito pode se ampliar como exclusão de tamanho (size exclusion) no caso
dos microrganismos considerando a distribuição parabólica das velocidades nos
poros. Os microrganismos e colóides maiores, em virtude ao seu tamanho,
experimentam, preferencialmente, as maiores velocidades na proximidade da
linha de fluxo central (Murphy, 2000), atingindo uma velocidade média muito
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

47
mais elevada que um traçador dissolvido. E pode ser drasticamente mais
pronunciado na observação de uma escala maior em meio natural (Murphy,
2000).
O efeito do tamanho se produz quando a partícula coloidal tem a mesma
escala de uma fração significativa de poros, sendo as partículas redirecionadas
para outros canais de poro maiores, onde os colóides experimentaram
velocidades maiores. Este processo é denominado exclusão de poro (poro
exclusion) ou mais especificamente exclusão de tamanho de poro (poro size
exclusion).
O processo de exclusão no caso de microrganismos é frequentemente
observado em estudos de campo (Hornberger et al., 1992; Pang et al., 2003) e
principalmente no caso de transporte de vírus (Woessner et al., 2001). De
acordo com Sirivithyapakornanand e Keller (2003) os colóides parecem ter uma
maior probabilidade de viajar nas proximidades da linha de corrente central no
espaço de poro, mas os colóides maiores (1-3μm) se espalham em um número
maior de linhas de corrente, desta forma não sendo transportados com a
velocidade máxima.
2.6.6. Bloqueamento e Ripening
A dependência dos coeficientes de adesão com relação ao tempo é
resultado de diferencias no comportamento de adesão de colóides sobre o meio
poroso “limpo” e sobre o meio que já contém colóides aderidos (Bradford et
Bettahar, 2006). Uma diminuição da adesão ocorre quando os locais favoráveis
à adesão são completamente ocupados (bloqueio ou blocking). Por outro lado,
um aumento ou melhoria da adesão pode acontecer quando as partículas
aderidas servem como lugares favoráveis para uma adesão subsequente
(Ripening).
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

48
a) b) c)
Figura 2.6. a) Cinética inicial de deposição, b) Efeito do bloqueamento e c)
Efeito do ripening (Kretzschmar et al., 1999)
Estes dois processos foram estudados por Camesano et al. (1999)
incluindo um coeficiente na equação (2.35) como segue:
1detattatt
bulk
1 SkCkn
t
S
(2.38)
onde (ψatt) é a função de adesão adimensional que considera o processo de
bloqueamento (blocking). Quando (ψatt) é igual a 1, é assumida a adesão em leito
limpo “clean bed” e a concentração dos colóides retidos no meio poroso
decresce exponencialmente com a profundidade. O valor de (ψatt) torna-se
função de S1 para levar em conta o processo de bloqueamento com (ψatt)
diminuindo enquanto S1 aumenta.
2.7. Ensaios experimentais
Experimentalmente, o transporte de microrganismos em meios porosos
saturados tem sido tradicionalmente investigado através da aplicação do método
de colunas empacotadas (packed column), termo utilizado em cromatografia,
onde a concentração dos microrganismos suspensos no efluente é monitorada
em função do tempo. Com o objetivo de entender os mecanismos de transporte
e retenção de organismos patogênicos, a literatura mostra que foram realizados
vários estudos em laboratório e campo indicados na Tabela 2.3.
O ensaio Advecção-Dispersão-Sorção (ADS) ou também denominado
ensaio de coluna, utilizado no campo da geotecnia para o estudo de transporte
de contaminantes, é aplicado no presente estudo ao estudo do transporte de
microrganismos como se verá em detalhe nos capítulos seguintes.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

49
Tabela 2.3. Componentes utilizados em experimentos realizados para o
estudo do transporte de microrganismos
Autor Ano Coloide Meio poroso Solução
HUYSMAN F. e
VERSTRAETE W.1992
Escherichia Coli Strain Ec1 e
Streptococcus faecalis Sf3Solo arenoso e solo argiloso Água desmineralizada
MCCAULOU D. R., et al . 1994Cepas de bactérias isoladas a partir
de um lençol freático S5 e S139
Quartzo, quartzo revestido
com hematita, quartzo
revestido com polimero
Água Artificial
BARTON J. W. 1995 Pseudomonas putida PRS2000 Água Artificial
RYAN Joseph N. et al. 1996partículas de látex (i.e., bacteria e
virus)Leito de esferas de vidro
MARTIN M.J. , et al. 1996Pseudomonas Fluorescens Strain
P17Quartzo medio Água deionizada
HENDRY L. et al. 1999Klebsiella oxytoca Burkholdeira
cepaciaAreia de Sílica Água Artificial
CAMESANO T.A., et al. 1999Pseudomonas putida e carboxilated
latexSolo do sul de Arizona Água Artificial
LI Q. e LOGAN B.E. 1999 Alcaligenes paradoxus bacteria Solo Ringold Água Artificial
BRADFORD S.A., et al. 2002 Partículas de látexLeito de vidro, varias areias de
OttawaÁgua deionizada
LIPING Pang 2003 Escherichia coli e F-RNA phages Areia de Pedra Pomes Água nativa
TUFENKJI N. e
ELIMELECH M.2004
Partículas de látex (e.g., bacteria e
virus)Leitos de vidro Água deionizada
TUFENKJI et al . 2004 Cryptosporidium parvum oocysts Areia de Quartzo Água deionizada
REDMAN J.A. 2004 E. coli K12 strain, D21 Areia de quartzo ultrapura Água deionizada
FOPPEN J.W.A. et al. 2005 E. coli Areia de QuartzoÁgua de torneira
estéril
FOPPEN et al. 2005 E. coli strain ATCC25922 Areia de Quartzo Água estéril
JIANG G. et al. 2005 Bacillus Subtilis Areia de Sílica Água estéril
BRADFORD S.A. et al. 2006 carboxyl latex Areia de Quartzo Água deionizada
LEVY J. 2007 E. coli Sedimentos glaciais Água de aquifero
KUZNAR Z.A. e
ELIMELECH M.2007
partículas de látex e
Cryptosporidium parvumLeitos de vidro Água deionizada
FOPPEN et al. 2007 E. coli ATCC25922 Areia de Quartzo ultra pura Água desmineralizada
JIANG G. et al. 2007 Escherichia coli strain D Areia de Sílica Água deionizada
KIM H.N. et al. 2009Escherichia coli strains (D21g e XL1-
Blue)Areia de Quartzo ultra pura Água artificial
LUTTERODT et al. 2009E. coli strains (UCFL-71, UCFL-94,
UCFL-131, UCFL- 167, UCFL-263 e
UCFL-348)
Areia de Quartzo ultra de
99.1% de pureza
Água desmineralizada
e Água artificial
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

3 MATERIAIS E MÉTODOS
3.1. Considerações Gerais
Como foi apresentado na Tabela 2.3, são diversos os trabalhos que se
referem ao estudo sobre a interação de microrganismos com o solo, em escala
de laboratório, sob condições controladas, utilizando diversos materiais como
meio poroso e como biocolóides; microrganismos ou modelos representativos
deles.
O estudo do transporte no presente trabalho é realizado através de
ensaios Adveção-Dispersão-Sorção (ADS). Com esta finalidade, inicialmente
foram escolhidos os materiais a serem utilizados nos ensaios de transporte, e
desenvolvido no Laboratório de Geotecnia e Meio Ambiente da PUC-Rio, um
equipamento destinado à realização dos ensaios ADS. Nesta seção serão
apresentadas as propriedades dos materiais escolhidos, os métodos e as
técnicas microbiológicas utilizadas e a descrição do equipamento.
Previamente à realização dos ensaios ADS foi necessária a execução de
ensaios microbiológicos, assim como de ensaios de batelada. A metodologia
utilizada nestes ensaios preliminares e nos ensaios ADS propriamente ditos
também será apresentada nesta seção.
3.2. Meio poroso
O meio poroso utilizado foi areia de quartzo industrial, limpa e seca,
utilizada por Pessôa (2006). Esta areia é resultado da mistura homogênea de
vários tipos de areias com distribuição uniforme de grãos com a finalidade de
obter uma areia bem graduada. As imagens da Figura 3.1, feitas em microscópio
estereoscópico (lupa) mostram a morfologia e tamanho dos grãos. O aspecto da
areia foi descrito por Pessôa (2006) como uma areia formada por grãos de
quartzo hialinos (incolor, transparente) e leitosos (branco translúcido,
embaçado), em geral angulosa.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

51
Figura 3.1. Areia de quartzo. Lupa ZEISS Stereo Discovery V8 x10 de
aumento
3.2.1. Caracterização geotécnica do meio poroso
A caracterização geotécnica do meio poroso foi feita através da
determinação das seguintes propriedades: massa específica, distribuição do
tamanho dos grãos, relação de vazios e permeabilidade. O resumo dos
resultados obtidos está disposto na Tabela 3.2, ao final desta seção.
As análises foram realizadas no Laboratório de Geotecnia e Meio
Ambiente da PUC-Rio, utilizando protocolos padrão indicados para cada ensaio.
A massa específica dos grãos (G) ou densidade relativa dos grãos foi
determinada de acordo com o procedimento indicado na norma brasileira
ABNT/NBR 6508/84. O valor médio de 2,64, assim determinado, considera-se
coerente com a densidade teórica do quartzo (2,65).
Devido à dependência do tamanho dos grãos sobre os diferentes
processos de transporte, foi feita a medida da distribuição do tamanho dos grãos
através do ensaio de análise granulométrica, de acordo com o procedimento
especificado pela Norma Brasileira ABNT/NBR 7181/84, utilizando uma mesa
vibratória horizontal.
A Figura 3.2 ilustra a curva granulométrica da areia de quartzo usada nos
experimentos deste estudo. De acordo com a curva granulométrica, o solo
apresenta 12% de areia fina (0,06 até 0,2 mm), 22% de areia média (0,2 até
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA
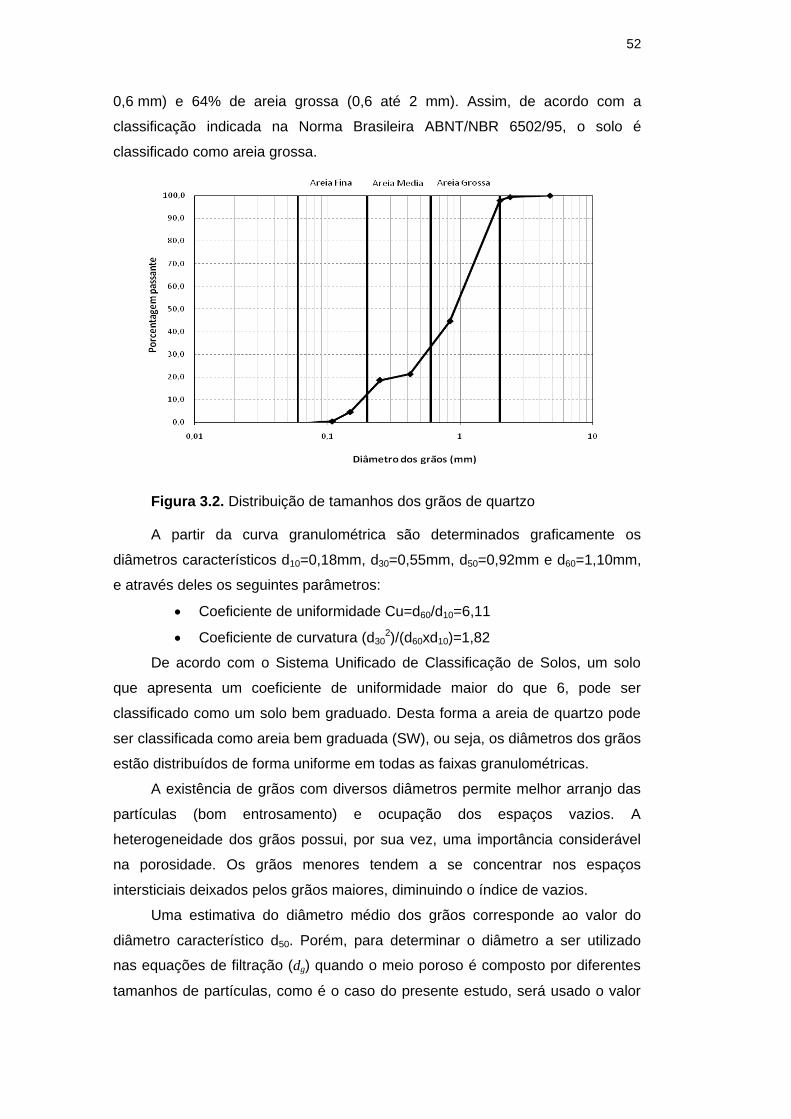
52
0,6 mm) e 64% de areia grossa (0,6 até 2 mm). Assim, de acordo com a
classificação indicada na Norma Brasileira ABNT/NBR 6502/95, o solo é
classificado como areia grossa.
Figura 3.2. Distribuição de tamanhos dos grãos de quartzo
A partir da curva granulométrica são determinados graficamente os
diâmetros característicos d10=0,18mm, d30=0,55mm, d50=0,92mm e d60=1,10mm,
e através deles os seguintes parâmetros:
Coeficiente de uniformidade Cu=d60/d10=6,11
Coeficiente de curvatura (d302)/(d60xd10)=1,82
De acordo com o Sistema Unificado de Classificação de Solos, um solo
que apresenta um coeficiente de uniformidade maior do que 6, pode ser
classificado como um solo bem graduado. Desta forma a areia de quartzo pode
ser classificada como areia bem graduada (SW), ou seja, os diâmetros dos grãos
estão distribuídos de forma uniforme em todas as faixas granulométricas.
A existência de grãos com diversos diâmetros permite melhor arranjo das
partículas (bom entrosamento) e ocupação dos espaços vazios. A
heterogeneidade dos grãos possui, por sua vez, uma importância considerável
na porosidade. Os grãos menores tendem a se concentrar nos espaços
intersticiais deixados pelos grãos maiores, diminuindo o índice de vazios.
Uma estimativa do diâmetro médio dos grãos corresponde ao valor do
diâmetro característico d50. Porém, para determinar o diâmetro a ser utilizado
nas equações de filtração (dg) quando o meio poroso é composto por diferentes
tamanhos de partículas, como é o caso do presente estudo, será usado o valor
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

53
do diâmetro efetivo d10, que de acordo com Martin et al. (1996), descreve de
modo mais exato o transporte de bactérias através de um meio heterogêneo.
O índice de vazios máximo e mínimo da areia de quartzo foi determinado
através dos métodos indicados pelas normas brasileiras ABNT/NBR 12004/90 e
ABNT/NBR 12051/91 respectivamente, sendo obtidos os resultados de 0,31 para
o emin e 0,66 para o emax. Nos ensaios ADS, o índice de vazios se manteve em
torno de 0,4, valor que permitiu a determinação da porosidade do meio poroso
através da equação (3.1), obtendo o valor de porosidade (n) igual a 0,29.
e1
en
(3.1)
O coeficiente de permeabilidade ou condutividade hidráulica saturada Ks foi
determinado usando o método da carga constante e parede rígida, de acordo
com a norma ABNT/NBR 13292/95. De forma sucinta, o procedimento envolveu
o uso da Garrafa de Mariotte, a partir da qual o fluxo de saída foi conectado à
válvula de ingresso no fundo do permeâmetro de parede rígida, gerando fluxo
ascendente e constante através da coluna de areia, conforme ilustra a
Figura 3.3. O volume de saída do efluente foi medido através da determinação
do peso do efluente em intervalos de tempo constante.
Figura 3.3. Aparato para a determinação da condutividade hidráulica
saturada
O valor da condutividade hidráulica Ks foi determinado a partir da Lei de
Darcy:
H
ALQK s
(3.2)
Mariotte
Permeâmetro de
Parede rígida
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

54
onde Q é a vazão de saída medida por peso de água através do tempo, L é o
comprimento, A é a seção transversal da amostra, e Δh a diferença de carga
entre o ingresso e saída da água do permeâmetro. O ensaio foi realizado três
vezes, sendo o valor adotado a média dos resultados dos três ensaios, como é
apresentado na Tabela 3.1.
Tabela 3.1. Determinação da condutividade hidráulica saturada
Ensaio i [-] Ks [cm/s]
1 0,25 2,91E-02
2 0,22 3,05E-02
3 0,20 2,75E-02
Média 2,91E-02
A condutividade hidráulica pode ser calculada também antecipadamente
utilizando a equação de Hazem Ks = d102, que para a areia de quartzo utilizada
corresponde ao valor de 3,24E-02 cm/s. Esta equação determina que para
areias soltas, a condutividade hidráulica é igual ao quadrado do diâmetro do
grão, para o qual só 10% da amostra tem um diâmetro menor. É interessante
notar que Pekdeger e Matthess (1988), citado em Foppen et al., (2005)
relacionam a filtração mecânica com a inversa do d10. Assim, o percentual de
10% da areia mais fina impacta tanto a condutividade como na filtração
mecânica. Esta relação tem lógica desde que estes grãos mais finos irão a
preencher os espaços entre aqueles maiores e assim influenciar o tamanho
médio do poro da coluna compacta inteira.
Tabela 3.2. Características físicas da areia de quartzo limpa
Propriedade Valor
Massa específica dos Grãos, G (-)
Relação de vazios máxima, emax (-)
Relação de vazios mínima, emin (-)
Diâmetro médio, d50 (mm)
Diâmetro efetivo, d10 (mm)
Condutividade hidráulica saturada, Ks (cm/s)
2,64
0,66
0,31
0,92
0,18
2,91E-2
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

55
3.3. Bactéria
Neste estudo irão ser avaliados, os principais processos conhecidos que
determinam a movimentação e distribuição da E. coli em aquíferos sob
condições de saturação, através da determinação de parâmetros de transporte
como a adsorção e a filtração, com o objetivo de esclarecer a importância
relativa dos vários fatores que influenciam o transporte.
O biocolóide usado como objeto de estudo foi a bactéria Escherichia coli
ATCC (American Type Culture Colletion) 11229, gentilmente cedida pelo
Professor Doutor Marco Antônio Lemos Miguel do Instituto de Microbiologia
Paulo de Góes da Universidade Federal do Rio de Janeiro. A determinação das
propriedades das células, como densidade específica, densidade seca e
determinação do tamanho das células foram realizadas, mas não fazem parte
dessa dissertação.
As dimensões da bactéria foram obtidas do estudo realizado por Matthess
et al. (1991b) citado em Foppen et al., (2005) e Foppen e Shijven, (2006), que
estudaram o transporte de E. coli ATCC 11229, determinando um comprimento
médio de 2-4 μm, diâmetro médio de 1 µm e a densidade da bactéria variando
entre de 1-1,05 g/cm3.
3.3.1. Manutenção da cepa de Escherichia Coli
A cepa de E. coli foi mantida e transferida por repique mensal, para tubos
de ensaio de 20 mL, contendo 5 mL de meio sólido Trypticase Soy Agar (TSA)
inclinado e também para tubos contendo 10 mL de meio líquido Trypticase Soy
Broth (TSB). As células dos repiques, conservados em geladeira, foram ativadas
para serem usadas nos ensaios, através de preparo do pré-inóculo, pela
transferência de alíquotas dos repiques para tubos de ensaio contendo 10 mL de
meio TSB. Após a inoculação, os tubos (pré-inóculo) eram incubados a
temperatura ambiente por 48 horas após as quais eram imediatamente usados
nos ensaios.
A partir destes tubos contendo o pré-inóculo, foram preparados inóculos,
número de bactérias necessárias para a realização dos ensaios, multiplicando
numericamente a população bacteriana. Para tal inoculou-se uma alíquota do
pré-inóculo, de volume igual ao 1% (v/v) do volume de meio TSB, necessário em
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

56
função do ensaio que será realizado. Esse conjunto é incubado até que as
bactérias atinjam o final da fase exponencial da curva de crescimento.
3.3.2. Determinação do número de bactérias
A estimativa numérica de células bacterianas foi feita através da contagem
de células por microscopia em Câmara de Neubauer, pela densidade óptica da
suspensão celular e por contagem de células viáveis em meio de cultura através
da técnica dos Tubos Múltiplos e por Contagem em Placas, dependendo do
ensaio a ser realizado. Essas técnicas serão descritas a seguir.
3.3.2.1. Contagem celular em Câmara de Neubauer
A câmara de Neubauer (CNB) consiste numa lâmina de microscopia com
espessura bem maior que uma lâmina normal, onde existe marcação de área
que permite, através de uso de uma lamínula de cobertura, determinar o volume
da amostra contida em cada espaço determinado por quadrantes. Por
conseguinte é possível, após contagem do número de bactérias nesses
quadrantes, se determinar o número de bactérias em cada mL da amostra
referentes a cada ensaio ou condição. Neste estudo foi utilizada uma câmara de
Neubauer modelo 1/400 mm2 x 0,1 mm (Figura 3.4).
a) b)
Figura 3.4. a) Câmara de Neubauer utilizada, b) Esquema da CNB com a
lâminula sobre a área de contagem
A seguir, a Figura 3.5 mostra que a área de contagem de 1 mm2 está
dividida em 25 quadrados cada um deles com área de 0,04 mm2 (1/25 mm2). A
contagem é realizada em pelo menos 5 desses quadrados, escolhidos
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

57
aleatoriamente. Por sua vez, estes últimos quadrados estão subdivididos em 16
quadrados menores de área igual a 0,0025 mm2 (1/400 mm2).
Figura 3.5. Área de Contagem da CNB e esquema da metodologia de
contagem.
O número total de bactérias é calculado multiplicando por 25 a média do
número de bactérias contadas nos 5 quadrados escolhidos, obtendo assim o
número de bactérias em 1 mm2. Considerando a profundidade da câmara de 0,1
mm o valor pode ser calculado para 1 mm3.
3.3.2.2. Leitura da densidade óptica por espectrofotometria
A espectrofotometria é uma técnica utilizada para determinar a quantidade
de uma substância ou elementos em solução ou suspensão em uma amostra.
Ela é baseada na absorção de radiação dos comprimentos de onda entre as
regiões que variam desde a faixa do ultravioleta, passando pela luz visível, até a
zona do infravermelho.
Desse modo, a absorbância A é uma medida indireta da concentração
bacteriana numa suspensão. A relação linear que existe entre absorbância e
concentração é conhecida como a relação de Beer-Lambert, que pode ser
expressa como:
A abc
(3.3)
onde a é uma constante que depende do tamanho e índice de refração da
bactéria, e do comprimento de onda da luz incidente (Deshpande e Shonnard,
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

58
1999); b é distância percorrida pelo feixe luminoso através da amostra e
finalmente c a concentração bacteriana.
Assim, a absorbância da luz a cada comprimento de onda é diretamente
proporcional à concentração da solução contida na cubeta. Esta linearidade
deixa de ocorrer em concentrações muito baixas e muito elevadas da
substância, podendo-se, neste último caso, diluir previamente a amostra a medir.
Uma curva padrão deve ser estabelecida para que seja possível correlacionar o
valor da absorbância com a quantidade numérica de bactérias, determinada por
contagem em Câmara de Neubauer ou por contagem de unidades formadoras
de colônias em meio de cultura.
Neste estudo a absorbância foi determinada utilizando o espectrofotômetro
Spectronic Genesys (Spectronic Instruments Inc.) e como recipientes, cubetas
retangulares de quartzo, de 1 cm de comprimento.
3.3.2.3. Contagem de microrganismos pela Técnica dos Tubos Múltiplos
A técnica dos Tubos Múltiplos também é conhecida como Técnica do
Número Mais Provável (NMP) e permite estimar o número de células viáveis
numa suspensão celular através do uso de indução matemática. O princípio
deste método consiste na inoculação de várias réplicas de diluições em série de
uma mesma amostra, em tubos de ensaio com meio de cultura líquido e na
detecção e contagem do número de réplicas nas quais for detectado
crescimento. Assume-se que cada tubo de ensaio onde se detecta crescimento
terá recebido pelo menos uma célula microbiana viável e cultivável. Interpreta-se
que os tubos nos quais não ocorre crescimento microbiano não devem ter
recebido nenhuma célula viável. Os tubos com crescimento distinguem-se
facilmente daqueles onde não ocorreu crescimento, pela presença de turbidez
e/ou mudanças de coloração apresentada pelo meio de cultura específico para
cada espécie ou grupo a ser detectado. A determinação do número mais
provável de células é feita através da aplicação de tabelas publicadas,
preparadas através de cálculos estatísticos, levando-se em consideração uma
combinação do número de réplicas, onde se detecta crescimento e as
respectivas diluições.
Para a determinação do NMP, as amostras foram diluídas sucessivas
vezes e inoculado 1 mL de cada diluição, em triplicata, em tubos com 10 mL de
meio líquido (TSB ou Caldo Fluorocult), de tal forma que se obtivesse uma última
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

59
diluição de 3 tubos que apresentasse pelo menos um resultado negativo. O
arranjo de número de tubos positivos das 3 diluições foi transposto para Tabelas
estatísticas que informam o NMP para as diferentes combinações de tubos
positivos e que também incluem os limites de confiança dos números mais
prováveis dos microrganismos pesquisados em função da Tabela em questão.
3.3.2.4. Contagem de Número de Unidades Formadoras de Colônias em Placas de Petri
A contagem em placas permite a visualização de colônias de
microrganismos, que são consideradas, cada uma como tendo sido originária de
uma única célula. Como é possível que algumas colônias se originem de células
não separadas, considera-se então não o número de células, mas sim o número
de Unidades formadoras de colônias.
Neste trabalho foi utilizada a técnica de contagem em placa por
derramamento em profundidade ou “pour plate”. Neste, a suspensão da amostra
contendo microrganismos em água sofria diluições sucessivas e uma alíquota de
0,1 mL dessas diluições era aplicada, em triplicata, no fundo de placas de Petri
de plástico, previamente esterilizadas como mostra o esquema da Figura 3.6.
Sobre esta alíquota era aplicado uma quantidade de meio TSA (tripticase soy
agar) fundido, a temperatura de aproximadamente 45º- 50ºC e o conjunto
cuidadosamente homogeneizado, através de movimentos circulares. Após a
solidificação do meio, as placas tampadas eram invertidas e incubadas a
temperatura ambiente (25°C) por 24 a 48 horas, para proceder à contagem do
número de colônias.
Na técnica de contagem em placa, as placas adequadas para contagem
devem ter entre 30 a 300 colônias. Assim, após a contagem, o resultado médio
do número de colônias das placas referentes a cada diluição era registrado e
multiplicado corrigido pelo fator da diluição.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

60
Figura 3.6. Esquema da Técnica de Contagem em Placa
3.4. O equipamento para o ensaio ADS
O estudo de laboratório implementado para determinar os parâmetros de
transporte da bactéria E. coli ATCC11229 em areia saturada, denominado
ensaio ADS (Advecção-Dispersão-Sorção), adotou um sistema baseado na
passagem contínua de uma suspensão bacteriana através de uma coluna de
solo, com fluxo vertical constante da base para o topo da coluna, reduzindo os
efeitos gravitacionais sobre o transporte bacteriano e facilitando a saída de
gases na coluna durante sua saturação.
Com este objetivo, no laboratório de Geotecnia da PUC-Rio e como parte
do presente estudo, foi desenvolvido o equipamento necessário para a
realização do ensaio ADS.
Para que fosse possível realizar os ensaios sob condições assépticas
foram utilizados materiais resistentes ao ataque químico e a temperaturas
maiores do que 121°C, visando à esterilização do equipamento por
autoclavação. Sendo assim foram escolhidos o teflon, resistente a uma grande
variedade de produtos químicos usados durante os ensaios, podendo ser usado
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

61
entre -200°C e +200°C em serviços intermitentes, o polipropileno, que tem boa
resistência química e resistência ao calor de até 120°C e finalmente o silicone.
O equipamento, semelhante ao utilizado para o ensaio de permeabilidade
de solos granulares à carga constante, consiste basicamente de uma coluna de
parede rígida e um reservatório (Frasco de Mariotte), ligados entre si por
mangueiras de silicone. O reservatório está destinado ao armazenamento de
água destilada estéril na etapa de saturação da coluna ou da suspensão
bacteriana durante os ensaios.
O Frasco de Mariotte é um dispositivo usado em laboratório para manter
constante a carga hidráulica do líquido a ser inserido na coluna, sem precisar de
qualquer sistema suplementar ou de bombeamento (Thorel et al. 2002). Assim,
este dispositivo foi utilizado para gerar o fluxo ascendente através da coluna de
areia.
Agitador
magnético
Mariotte
Permeâmetro de
parede rígida
a) b)
Figura 3.7. Esquema de funcionamento do equipamento para os ensaios
ADS a) Δh=0 , b) Δh>0
Devido ao funcionamento hidráulico do equipamento, mostrado na
Figura 3.7 é preciso determinar a posição da coluna com relação ao Mariotte, de
forma que a solução contida no frasco de Mariotte flua até a coluna, da base
para o topo, por efeito da diferença de carga hidráulica Δh. Para isto, a parte
inferior do tubo do frasco de Mariotte deve ficar por cima da válvula de saída da
coluna. Esse sistema permite que a pressão, no final do tubo dentro do frasco,
seja o valor da pressão atmosférica, enquanto que a pressão acima do nível de
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

62
água no interior do frasco de Mariotte se torna negativa. A Figura 3.7 (a) mostra
a configuração do equipamento para a condição estacionária do sistema (Δh=0)
e na . Figura 3.9 (b) no caso que o gradiente hidráulico é maior que um (Δh>0).
A seguir são descritas as características do Frasco de Mariotte e da
coluna.
3.4.1. Características do frasco de Mariotte
Os Mariottes consistiram de cilindros de teflon com 15,8 cm de diâmetro
interno e comprimento de 29 cm. Foram construídos para conter
aproximadamente 5 litros de material fluido (água ou suspensão bacteriana). A
tampa de cada Mariotte, também de teflon, é acoplada à abertura superior do
Mariotte de forma a não permitir nenhuma passagem de ar. Para garantir o
fechamento hermético, foi disposto um lenço de silicone entre a tampa e o
cilindro. Essa tampa é atravessada por um tubo de polipropileno de 5/8” de
diâmetro sujeita à tampa através de um O-ring como mostra a Figura 3.8. É
importante apontar que foi colocada uma peça adicional no extremo exterior do
tubo em forma de U, com o objetivo de minimizar a entrada de ar contaminado,
durante a execução dos ensaios.
Figura 3.8. Corte transversal e detalhe da tampa do Mariotte
Assume-se que o valor da concentração da suspensão bacteriana se
mantém constante no ingresso da coluna para o ensaio inteiro, nesse sentido
para manter a suspensão bacteriana homogeneizada foi acoplada como base do
Mariotte, na parte inferior do cilindro, uma placa agitadora com um bastão
magnético (Agitador magnético Fisatom mod. 753), conseguindo-se a agitação
contínua da suspensão e, portanto sem variações da concentração inicial. Entre
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

63
o cilindro e a base também foi colocado um lenço de silicone para a vedação do
reservatório (ver Figura 3.9).
Figura 3.9. Corte transversal e detalhe da base do Mariotte
A saída do fluxo do Mariotte está situada na parte lateral inferior do cilindro
através de uma válvula de aço inoxidável conectada a uma mangueira de
silicone (ϕint= ¼ ̎ ), que conduz o fluxo até a base da coluna.
3.4.2. Características da coluna
O diâmetro das colunas atendeu a relação mínima de d/d10 de 50,
recomendada para estudos de filtração de água. A finalidade do uso dessa
relação é minimizar os efeitos sobre a porosidade e a distribuição de fluxo em
colunas empacotadas (Knappett et al., 2008).
Figura 3.10. Esquema dos componentes da coluna
Como se observa na Figura (3.10) as colunas eram constituídas por
cilindros de teflon com 6,35 cm de diâmetro interno e 20 cm de comprimento.
Discos de alumínio perfurado foram colocados no topo e base das colunas para
uma distribuição uniforme do fluxo e sustentação da coluna de areia. Para evitar
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

64
a perda de grãos de areia durante o funcionamento da coluna, foi utilizada uma
tela de aço inoxidável com abertura de 0,109 mm (equivalente a malha #150)
também no topo e base da coluna.
A base e a tampa encaixam perfeitamente nos extremos da coluna através
de O-ring’s. A válvula de ingresso do fluxo está situada no centro da tampa
inferior, uma vez que a solução ingressa à base da coluna é acumulada em um
espaço de dimensões relativamente pequenas antes de iniciar sua passagem
através da coluna de areia. Isto permite que a solução seja uniformemente
distribuída, além da placa de alumínio perfurada colocada também com este
mesmo objetivo. A mesma configuração foi projetada para a tampa superior, com
a única diferença que a válvula de saída está situada na parte lateral da tampa.
A Figura (3.11) mostra a seção transversal da coluna completamente montada.
Figura 3.11. Corte transversal da coluna
Durante a montagem da coluna os comprimentos das mangueiras do
influente e efluente foram mantidos constantes para todos os ensaios, para
reduzir a variabilidade entre eles devido às perdas nas tubulações.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

65
3.5.Ensaios Microbiológicos
Os ensaios microbiológicos foram realizados sob condições de assepsia,
utilizando para isto uma capela de fluxo laminar durante a manipulação das
culturas bacterianas. Os materiais usados durante os ensaios, tais como meios
de cultura, vidrarias, água, etc., foram previamente esterilizados em autoclave a
121°C, equivalente a uma pressão de 1 atm, por 15 a 20 minutos.
A esterilização da areia foi realizada com a finalidade de destruir a
população de microrganismos com a mínima alteração das propriedades físicas
e químicas do solo, sendo a esterilização com calor úmido (em autoclave) um
dos métodos mais utilizados.
Com este objetivo foram colocados aproximadamente 1250 gramas de
areia seca numa bandeja de vidro coberta por papel alumínio. As dimensões da
bandeja permitiram que a espessura de solo já distribuído não fosse maior do
que 2 cm como recomendado pelo procedimento padronizado no Methods of Soil
Analysis (SSSA, 1994), permitindo a penetração de vapor durante a
autoclavação. O solo foi assim, autoclavado a 0,10 MPa = 1 atm = 1 kgf/cm2 a
121ºC durante 1 hora. Como controle e para verificar a eficiência do processo de
esterilização, alíquotas do solo já autoclavado foram retiradas com espátula
estéril, colocadas em três tubos de ensaio contendo 10 mL de meio TSB cada
um deles, e incubadas por 2 dias (48 horas) a 25ºC não sendo observado
crescimento bacteriano nestes meios.
3.5.1. Correlação entre Número de Células e Absorbância
A determinação do número de células pela densidade óptica da suspensão
bacteriana é um método simples, de fácil manipulação e de resposta rápida.
Entretanto os valores de absorbância não resultam diretamente no número de
células existentes nas culturas. Assim, para se conhecer o número de células
presentes em cada uma das amostras avaliadas nas diferentes etapas de cada
ensaio, a partir de determinações espectrofotométricas, é necessário estabelecer
uma correlação gráfica entre o valor da densidade óptica por absorbância e o
número de células de uma suspensão determinado por uma metodologia que
possibilite a contagem de células, como por exemplo, a contagem em Câmara
de Neubauer.
Foram estabelecidas curvas de correlação tanto para células suspensas
em meio TSB, como para aquelas ressuspensas em água destilada, após
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

66
centrifugação. As análises da água usada para a saturação das colunas foram
feitas para serem usadas como referência nas determinações do número de
células nos efluentes dos ensaios de batelada e ADS sem a introdução do
inoculo de E. coli.
Para a obtenção de massa celular para as determinações a serem
efetuadas, alíquotas de 1,5 mL de pré-inóculo de E. coli foram inoculadas em
erlenmeyers de 250 mL contendo 150 mL de meio TSB estéril. Esse conjunto foi
incubado em banho-maria a 25°C, com agitação constante de 100 rpm por
24 horas. Experimentos prévios, realizados no laboratório de Geotecnia e Meio
Ambiente, apontaram que a massa celular da cultura de E.coli ATCC 11229 se
estabilizava a partir de 24 horas de incubação nas condições descritas neste
item.
O meio TSB apresenta uma cor amarelada e absorve melhor a luz em
comprimentos de onda diferentes daqueles absorvidos com água destilada.
Como, para alguns ensaios a absorbância foi determinada diretamente dos tubos
de TSB com crescimento bacteriano e, em outros, as células estavam suspensas
em água, foi necessário realizar a correlação entre a absorbância e o número de
células nas duas condições.
No caso do estabelecimento da correlação entre o número de células e a
absorbância medida diretamente em meio TSB, uma alíquota de 10 mL retirada
do crescimento de 24 horas foi diluída sucessivas vezes em meio estéril, e a
absorbância relativa a cada diluição lida em espectrofotômetro a 600 nm (Ling et
al., 2002). As mesmas diluições foram lidas em Câmara de Neubauer para a
determinação da concentração celular, como se mostra na Figura 3.12 a.
Para a construção da curva de correlação entre a concentração de células
e a absorbância, em amostras suspensas em água destilada, alíquotas de 10 mL
do crescimento de 24 horas foram retiradas, centrifugadas a 1225 xg equivalente
3500 rpm na centrífuga Bio Eng Modelo BE-4000 por 25 minutos e ressuspensas
no mesmo volume de água destilada, como é mostrado no esquema da
Figura 3.12 b. Essa suspensão foi então diluída sucessivas vezes para a
contagem de células em Câmara de Neubauer e para a leitura da absorbância a
410 nm (Foppen, et al., 2005).
Com os dados obtidos foram traçadas curvas, onde se procurou
estabelecer a faixa de correlação linear entre os dois parâmetros, para que as
leituras dos ensaios fossem realizadas dentro desses limites de confiança, onde
a Lei de Beer- lambert é válida.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

67
a) b)
Figura 3.12. Esquema da metodologia seguida para a obtenção das
curvas de correlação entre absorbância e a) Suspensão bacteriana em
meio TSB; b) Suspensão bacteriana em água destilada
3.5.2. Curva de crescimento
A curva de crescimento da E. coli ATCC 11229 foi feita com o objetivo de
conhecer a cinética do crescimento da referida bactéria sob as condições
determinadas nessa dissertação. O ensaio consistiu na determinação da
concentração de células para diferentes períodos crescentes de incubação.
Para a obtenção da curva de crescimento, alíquotas de 1,5 mL do pré-
inóculo de 24 horas, foram introduzidas em erlenmeyers de 250 mL contendo
150 mL de TSB estéril. Esse conjunto foi incubado em banho-maria a 25°C, com
agitação constante de 100 rpm pelo período de duração do ensaio. Para cada
ensaio foram feitas triplicatas desses conjuntos.
Imediatamente após o procedimento de inoculação foram, então, retiradas,
de forma asséptica, em fluxo laminar, alíquotas de 5 mL de cada frasco, para
determinar o número de bactérias correspondente ao tempo inicial (To) e,
novamente após diferentes e crescentes períodos de incubação, para a
determinação do crescimento bacteriano ao longo do tempo. Esse crescimento
foi avaliado através da determinação da densidade óptica, lendo-se a
absorbância a 600 nm diretamente no material retirado dos erlenmeyers. Caso a
absorbância lida ultrapassasse a faixa do momento linear, a alíquota era diluída
até que caísse numa faixa de leitura confiável.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

68
O ensaio foi realizado em triplicata, sendo preparados simultaneamente
três inóculos e incubados paralelamente por um tempo superior a 24 horas. A
concentração final de células para cada ponto da curva de crescimento é
resultante do valor médio de absorbância dos três valores obtidos.
3.5.3. Ensaio de Sobrevivência
O ensaio de sobrevivência da bactéria E. coli ATCC11229 em água
destilada, foi realizado com o objetivo de estabelecer o tempo que, as células
que seriam introduzidas na coluna de areia, permaneceriam na fase estacionária
e, desta forma garantir que o número de células não experimentaria variações
devido ao crescimento ou decaimento, durante a execução do ensaio ADS.
Consistiu na determinação do número de células viáveis coletadas do final
da fase logarítmica de crescimento, ressuspensas em água destilada, através da
contagem de microrganismos pela técnica de tubos múltiplos, para diferentes
períodos de tempo.
Previamente foi preparado o inóculo, adicionando 1 mL do pré-inóculo em
um erlenmeyer de 250 mL contendo 125 mL de meio TSB estéril, este conjunto
foi incubado em banho maria a 25º C e 100 rpm durante 24 horas. Após
24 horas de incubação foi retirada uma alíquota de 5 mL deste crescimento para
realizar determinação da biomassa através da leitura de absorbância no
espectrofotômetro. Imediatamente o inóculo é transferido para tubos falcon
estéreis, para ser centrifugado a 1225 xg equivalente 3500 rpm por 25 minutos.
Após este tempo o sobrenadante foi descartado, e o precipitado ressuspenso em
120 mL de água destilada em um erlenmeyer de 250 mL. A suspensão assim
preparada foi homogeneizada, para posteriormente iniciar a preparação das
diluições sucessivas, requeridas para efetuar a contagem através da técnica dos
tubos múltiplos, correspondentes ao tempo To. Neste instante também foi
retirada uma alíquota de 5 mL para controle das quantidades de células totais
presentes na ressuspensão.
Uma vez realizadas as diluições sucessivas, para cada diluição
considerada, foram inoculados 3 tubos contendo 10 mL de caldo Fluorocult para
a identificação de E. coli, através da transferência de 1 mL para cada um dos
três tubos contendo o meio específico. Finalmente os tubos são incubados em
banho-maria à temperatura de 36°C. As leituras dos tubos que apresentaram
crescimento são realizadas a 24 e 48 horas. Esta leitura corresponde ao tempo
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

69
inicial T0. Posteriormente o procedimento foi repetido para diferentes períodos de
tempo.
3.6. Ensaio de Adsorção em Batelada
Este ensaio foi realizado com o objetivo de analisar a participação do
processo de adsorção físico-química na retenção de bactérias E. coli na areia de
quartzo, sob condições de ensaio escolhidas. Os ensaios foram executados de
acordo com o procedimento indicado por Jiang et al., (2007) e modificado para
as concentrações de 106, 107, 108, e 109 cel/mL, conforme descrito a seguir.
A areia foi homogeneizada com auxílio de uma espátula, para que fossem
acrescentados 2 gramas deste material em tubos plásticos cônicos de 50 mL
com tampa de rosca (tubos Falcon), procedendo em seguida à esterilização do
conjunto em autoclave a 121ºC por 1 hora.
A massa de células de E.coli foi obtida a partir do crescimento do inóculo,
durante 24 horas em banho-maria a 25ºC e agitação constante de 100 rpm,
preparado utilizando 1 mL de pré-inóculo em 100 mL de meio TSB. Ao final da
incubação, uma alíquota de aproximadamente 5 mL era retirada assepticamente,
para leitura da absorbância a 600 nm e posterior correlação entre Absorbância e
Número de células, para a determinação dos volumes e das diluições da cultura
bacteriana a serem utilizados para a obtenção do número de células necessário
para cada concentração celular testada. Esse volume de suspensão bacteriana
era distribuído em tubos falcon estéreis, centrifugação, por 25 minutos a 1225 xg
equivalente 3500 rpm e ressuspenso em água destilada estéril.
Em seguida, 20 mL da ressuspensão bacteriana, preparada na
concentração desejada, foram adicionados aos tubos falcon com os 2 g de areia
estéril. Esse conjunto foi homogeneizado vigorosamente em um vortex durante
10 segundos e incubado a temperatura ambiente em agitação constante de
100 rpm durante uma hora. Após este período, os tubos foram centrifugados por
30 segundos a 120 xg equivalentes a 1100 rpm na centrífuga Bio Eng Modelo
BE-4000. Após a centrifugação, foram retirados 10 mL do sobrenadante e
colocados em outro tubo falcon de 50 mL vazio e estéril. A concentração de
células neste sobrenadante foi determinada através de diluição serial para a
contagem em placa com a técnica de derramamento em profundidade, conforme
descrito no item 3.3.2.4. Este ensaio foi realizado em duplicata para cada
concentração. A Figura 3.13 mostra a aparência das placas de Petri após o
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

70
crescimento das colônias (UFC) de E. coli em meio sólido TSA para duas
diluições diferentes.
Figura 3.13. Crescimento das colônias de E. coli no ensaio de adsorção
para diferentes concentrações iniciais
O coeficiente de partição KD (mL/g) ou coeficiente de distribuição é um
valor relativo que representa a razão entre o número de bactérias aderidas à
superfície da fase sólida Cs (UFC/g) e o número de bactérias que permanece em
suspensão Cw. (UFC m/L), calculado segundo a seguinte equação:
w
sD
C
CK (3.4)
A concentração de células na fase sólida, a concentração de células em
suspensão, e a porcentagem de adsorção Pa, são dadas pelas seguintes
equações:
W
NNC st
s
(3.5)
V
NC s
w (3.6)
100
N
NNP
t
sta
(3.7)
onde Nt é o número total de bactérias adicionadas ao solo (UFC), Ns o número
total de bactérias no sobrenadante (UFC), W a massa do solo utilizada (g) e V o
volume de fluido na mistura (mL).
3.7. Ensaio Advecção-Dispersão-Sorção (ADS)
O objetivo dos ensaios ADS foi avaliar os processos de transporte da
bactéria E.coli em areia saturada, através da obtenção da curva de chegada
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

71
(Breakthrough curve) para diferentes velocidades de fluxo. Os ensaios foram
executados no equipamento projetado neste estudo (Figura 3.14),
exclusivamente com esta finalidade e cujas características foram descritas no
item 3.4.
Figura 3.14. Foto do equipamento utilizado no ensaio ADS
O procedimento do ensaio é semelhante ao ensaio de permeabilidade de
solos granulares à carga constante, porém com percolação contínua da
suspensão bacteriana em água destilada, numa concentração inicial conhecida.
Na medida em que a solução permeia através do solo (areia de quartzo), o
efluente é coletado e determinado o valor da concentração celular. O grande
diferencial deste ensaio com outro de transporte de contaminantes, reside nos
cuidados específicos que exige a manipulação da bactéria, devendo se cumprir
um protocolo de esterilização antes de cada ensaio e prever qualquer tipo de
contaminação externa durante o ensaio. A areia e a água destilada e
componentes da coluna tais como, mangueiras, discos perfurados, malha de
aço, tubo de polipropileno foram esterilizados com antecedência a cada ensaio,
assim como toda a vidraria e materiais a serem utilizados.
Basicamente cada ensaio foi dividido em duas etapas: saturação da areia
e injeção contínua da suspensão bacteriana, cada uma destas descritas a
seguir.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

72
3.7.1. Saturação da areia
O procedimento inclui a montagem do equipamento e conformação da
coluna de areia.
Inicialmente o interior do frasco de Mariotte foi esterilizado utilizando uma
solução de acetona, preparado assim para verter imediatamente após este
tratamento, 3,5 litros de água destilada já estéril, reservada para a saturação da
areia. Em seguida a tampa era colocada, e ajustados os parafusos para seu
fechamento hermético, introduzindo finalmente o tubo de polipropileno.
Posteriormente a base da coluna também foi esterilizada com acetona,
mantendo fechada a válvula da base da coluna e colocados o disco perfurado, a
malha de aço, e o cilindro de teflon. Em seguida a mangueira de silicone era
conectada na válvula de saída do Mariotte. Abrindo-se esta válvula deixava-se
fluir a água destilada até que toda a mangueira estivesse preenchida e sem a
presença de bolhas de ar, para finalmente ligar a mangueira à base da coluna. A
partir daqui inicia-se o processo de conformação da coluna de areia.
A areia já esterilizada e previamente seca na estufa a 105°C e passada
pela malha #150, foi vertida dentro da coluna em 4 camadas de
aproximadamente 300 g (5 cm) com ajuda de um Becker e um funil de vidro.
Antes de colocar a primeira camada de areia deixou-se fluir um volume de água
equivalente ao volume de poros necessários para preencher os poros da areia
desta primeira camada, considerando o volume adicional que é mantido na base
da coluna. Para as camadas subsequentes, antes de colocar a camada de areia
correspondente, água estéril foi adicionada aos poucos, com o auxílio de uma
proveta graduada de 100 mL. Entre camada e camada foram aplicadas 25
batidas laterais na coluna. Esta prática minimizou a formação de regiões
heterogêneas no empacotamento da coluna.
Concluída a conformação da coluna de areia, o topo da coluna era
nivelado utilizando uma barra metálica apoiada nas bordas do cilindro, para a
imediata colocação da malha de aço, disco metálico perfurado e a tampa
superior da coluna. As fotografias da Figura 3.15 mostram a coluna antes de
colocar a tampa superior. Finalmente eram ajustados os parafusos para o
fechamento hermético da coluna, e a partir de aqui, iniciava-se a saturação da
areia, através da passagem contínua de aproximadamente 4 volume de poros de
água destilada, com a aplicação de uma carga hidráulica constante, determinada
pela posição do tubo de polipropileno no interior do Mariotte.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

73
Figura 3.15. Vista superior e lateral da coluna após do processo de
conformação da coluna de areia
Concluída a saturação eram medidos o pH e a condutividade elétrica da
água. E antes de dar inicio ao ensaio ADS propriamente, a válvula superior e
inferior da coluna eram fechadas até efetuar a conexão da mangueira que
permitirá a injeção da suspensão bacteriana.
3.7.2. Injeção contínua da suspensão de E. coli
Paralelamente à etapa de saturação foi preparada a suspensão de E. coli a
ser injetada na coluna, com o objetivo de obter uma concentração da suspensão
em torno de 1 x 108 cel/mL. Esta foi considerada a concentração inicial de
células injetadas para cada ensaio. Para isto, primeiro foi preparado o inóculo
bacteriano de E. coli mediante a inoculação de 5 mL do pré-inóculo em 500 mL
de meio TSB, sendo o conjunto incubado em agitação constante de 100 rpm e
temperatura aproximada de 25°C. A quantidade de inóculo preparado foi
determinada com base no número de células crescidas até 24 horas de
incubação e em função da quantidade de células necessárias para obter a
concentração inicial desejada (1 x 108 cel/mL). A foto da Figura 3.16 mostra o
inóculo sendo incubado no banho Maria e a turvação do meio, relativa ao
crescimento no final do tempo de incubação. A determinação da concentração
de bactérias no inóculo foi feita através da medida de densidade óptica e
posterior correlação na curva de calibração.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

74
Figura 3.16. Crescimento do inóculo de E. coli em 500 mL de meio de
cultura TSB
Após o crescimento, o inóculo foi distribuído uniformemente em tubos
falcon estéreis de 50 mL de capacidade, para serem centrifugados, sob
condições assépticas, a 1225 xg equivalente 3500 rpm por 25 minutos. Através
desse processo as células são precipitadas no fundo do tubo e desse modo a
suspensão separada em material precipitado e o sobrenadante. Após a
centrifugação o sobrenadante era descartado assepticamente em algum
recipiente dentro da capela de fluxo laminar. Em seguida as células precipitadas
eram ressuspensas com auxílio de um agitador mecânico (vortex), adicionando-
se 1 ou 2 mL de água destilada estéril. Após a total ressuspensão do
precipitado, era acrescentada água destilada para completar um volume de 500
mL de suspensão bacteriana.
Todos os 500 mL da suspensão bacteriana eram colocados no interior do
Frasco de Mariotte, previamente desinfetado com uma solução de acetona e
com o bastão magnético no interior. Imediatamente depois era adicionada água
estéril até completar os 4 litros de suspensão de E. coli. O reservatório então era
fechado e a altura do tubo regulada em função do gradiente a ser imposto, de
acordo com cada ensaio. A partir deste momento era ligado o agitador
magnético para a homogeneização contínua dos 4 litros de suspensão
bacteriana a ser injetada na coluna de areia.
Antes do início da injeção da suspensão bacteriana, porém, e
encontrando-se a coluna já saturada, todas as válvulas eram fechadas para
proceder-se à troca da mangueira utilizada para a saturação da coluna, por outra
que conduzia a suspensão de bactérias do Mariotte até a base da coluna. Da
mesma forma que durante a etapa de saturação a mangueira era colocada de
forma a evitar a formação de bolhas de ar. A partir de aqui o equipamento estava
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

75
preparado para iniciar a passagem contínua da suspensão de E. coli através da
coluna de areia. A injeção da suspensão bacteriana foi efetuada de forma
contínua, passando entre 12 e 14 volume de poros, variando de acordo com
cada ensaio. A duração de cada ensaio foi determinada pela forma da curva de
chegada (Breakthrough curve) quando a concentração normalizada (C/Co)
tornava-se constante.
O fluxo foi medido frequentemente durante todo o ensaio
(aproximadamente a cada 10 minutos), com o objetivo de controlar o volume de
saída do efluente da coluna e de forma simultânea observar o comportamento da
condutividade hidráulica durante a injeção da suspensão bacteriana. Para tal,
utilizou-se uma proveta graduada e um cronômetro, sendo calculado o valor da
vazão (Q) através da relação Volume/ Tempo. Posteriormente, com o valor do
gradiente hidráulico constante ao longo da execução do ensaio e através da Lei
de Darcy, foram calculados os valores de velocidade de Darcy (vd) e
condutividade hidráulica (KS).
Durante todo o ensaio o agitador magnético se manteve em contínuo
funcionamento e na mesma velocidade de agitação, isto com a finalidade de
causar o mínimo de interferências no fluxo de saída.
A coleta de amostras foi realizada em provetas de vidro graduadas, de
25 mL, previamente esterilizadas. O volume coletado foi de 15 mL a intervalos
de 15 ou 30 minutos variando em função do gradiente imposto. Para a
determinação da concentração do efluente, foi medida a absorbância no
espectrofotômetro, durante cada intervalo de amostragem. Uma vez lida a
absorbância as amostras foram mantidas na geladeira para a um posterior
controle através da contagem na câmara de Neubauer seguindo o procedimento
detalhado no item 3.3.2.1.
É importante salientar que foi realizado um controle da concentração do
influente da coluna, extraindo para isso, com uma seringa, uma alíquota da
mangueira no setor de saída do Mariotte e lida a absorbância para posterior
correlação na curva de calibração. Esse controle foi feito em decorrência que é
assumido que o valor da concentração inicial é mantido constante durante a
totalidade do ensaio.
Para a determinação da densidade seca total do solo (ρBulk) uma vez
desmontada a coluna, a areia foi esterilizada e depois seca na estufa a 105°C,
para a obtenção do peso seco. Sendo conhecido o volume ocupado pela areia, o
valor da densidade seca total foi calculado dividindo o peso da areia seca entre o
volume da coluna.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

76
3.7.3. Obtenção das curvas de chegada
Como foi explicado na revisão bibliográfica, o transporte de biocolóides
através de um meio poroso saturado é representado matematicamente através
da equação (2.5). A solução analítica desta equação, proposta por Ogata e
Banks (1961), citada em Azevedo et al. (2002), é apresentada a seguir:
tRD2
vtRLerfc
D
vLexp
tRD2
vtRLerfc
2
1
C
C
hhho
(3.8)
onde L é o comprimento da coluna de solo, erfc é a função de erro
complementar, R o fator de retardamento, Dh a dispersão hidrodinâmica, t o
tempo e v a velocidade intersticial.
A representação gráfica da equação de transporte ADS equação (2.5) é
denominada curva de chegada ou curva de transporte (Breakthrough curve), a
forma da curva e mostrada na Figura 3.17.
a) b)
Figura 3.17. Linha continua - Forma da curva de chegada sem
retardamento nem decaimento; Linha tracejada - a) Forma da curva de
chegada com retardamento b) Forma da curva de chegada com
decaimento
O valor de retardamento pode ser obtido da curva de transporte,
assumido como igual ao valor de volume de poro PV, correspondente à
concentração relativa (C/Co) igual a 0,5 na curva de chegada. Esta suposição foi
definida a partir da solução analítica apresentada na equação (3.8), na qual se
negligencia o segundo termo do lado direito da equação e substituindo alguns
termos da mesma, tem-se, segundo explicação matemática mostrada por
Azevedo et al. (2002):
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

77
2
1
o RT4
PTRerfc
2
1
C
C (3.9)
onde T é igual a v·t/L e P é igual a v·L/Dh. Tomando T=R na equação (3.9), o
argumento da função de erro complementar toma-se nulo e a função é, então,
igualada à unidade. Assim tem-se a concentração relativa igual a 0,5. Na
determinação do coeficiente de dispersão hidrodinâmica, para o ponto de
concentração relativa de 0,5 é traçada uma tangente a este ponto. Com isto é
possível estabelecer o valor de b, como dado por Azevedo et al. (2002):
h
x
RT
o
RD4
Lvb
dT
CCd
)/( (3.10)
A equação acima mostra que a representação matemática do lado
esquerdo da equação nada mais é do que uma derivada, obtida a partir da
equação (3.9). Observa-se, portanto, que o valor de b é a tangente tomada na
curva de chegada na concentração relativa de 0,5. Com o valor b é possível, a
partir da igualdade mostrada na equação (3.10), obter o coeficiente de dispersão
hidrodinâmica, dado por:
22
xh
bR4
LvD
(3.11)
3.8. Ensaios de controle
No final dos ensaios ADS observou-se a necessidade de realizar alguns
controles com uma forma de verificar os procedimentos realizados e auxiliar na
posterior análise dos resultados.
Foi realizado o controle da concentração de ingresso da suspensão
durante os últimos ensaios ADS, aproximadamente na metade e no final do
ensaio, através da retirada de alíquotas de aproximadamente 5 mL, com a
utilização de uma seringa, para a posterior determinação da concentração inicial
mediante a medida da densidade óptica e posterior correlação entre absorbância
e número de células.
Um segundo controle foi realizado, imediatamente após a finalização do
ensaio ADS, que consistiu na determinação da concentração de bactérias retidas
na areia e a distribuição granulométrica dos grãos de areia, ao longo da coluna.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

78
Para isto, foram determinados três pontos de amostragem, na base, na metade e
no topo da coluna (este controle foi realizado para o ensaio ADS 1).
Para a retirada das amostras de forma asséptica, a coluna foi levada para
a capela de fluxo laminar, onde foi cuidadosamente desmontada. Uma vez
retirada a tampa superior, foram coletadas as amostras de areia do topo da
coluna, em torno de 1 grama para a determinação da concentração de bactérias
e aproximadamente 100 gramas para a análise granulométrica. A seguir foi
colocada uma tampa de encaixe perfeito no interior da coluna, ocupando o
espaço deixado pela areia que foi retirada, e assim, a coluna foi virada para a
obtenção das amostras correspondentes à base, e por último à metade da
coluna, da mesma maneira como foi feito no topo da coluna. Imediatamente
depois foram realizadas as diluições sucessivas requeridas para a aplicação da
técnica dos Tubos múltiplos e para determinar as concentrações da bactéria na
areia. Posteriormente, foi obtida a curva granulométrica mediante peneiramento
para cada ponto de amostragem através do procedimento padronizado pela
Norma Brasileira ABNT/NBR 7181/84.
Finalmente, uma vez concluídos todos os ensaios programados, foi
realizado um último ensaio, que permitiu determinar o diâmetro médio dos poros
da coluna de areia. Dessa forma, foram montadas quatro colunas de areia com
densidade seca média de 1,74 g/cm3, em cilindros de acrílico de dimensões de
aproximadamente 6 cm de diâmetro interno e 9 cm de comprimento. Os cilindros
foram colocados dentro de uma bandeja, cada um deles disposto sobre uma
pedra porosa e papel filtro situado na base do cilindro de areia.
No inicio do ensaio, a bandeja foi preenchida com água de forma que a
pedra porosa fosse totalmente coberta para a água começar a ascender nas
colunas, por capilaridade. Depois de observada a estabilização foi medida a
altura de capilaridade hc. Considerando a média dessa altura foi calculado o
diâmetro médio dm de poro conforme a equação seguinte:
ghd
c
m
(3.12)
onde, δ é a tensão superficial da água 0,0728 N/m a 20°C (valor extraído de
Myers, 1999), e g a aceleração da gravidade (9,81 m/s2).
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA
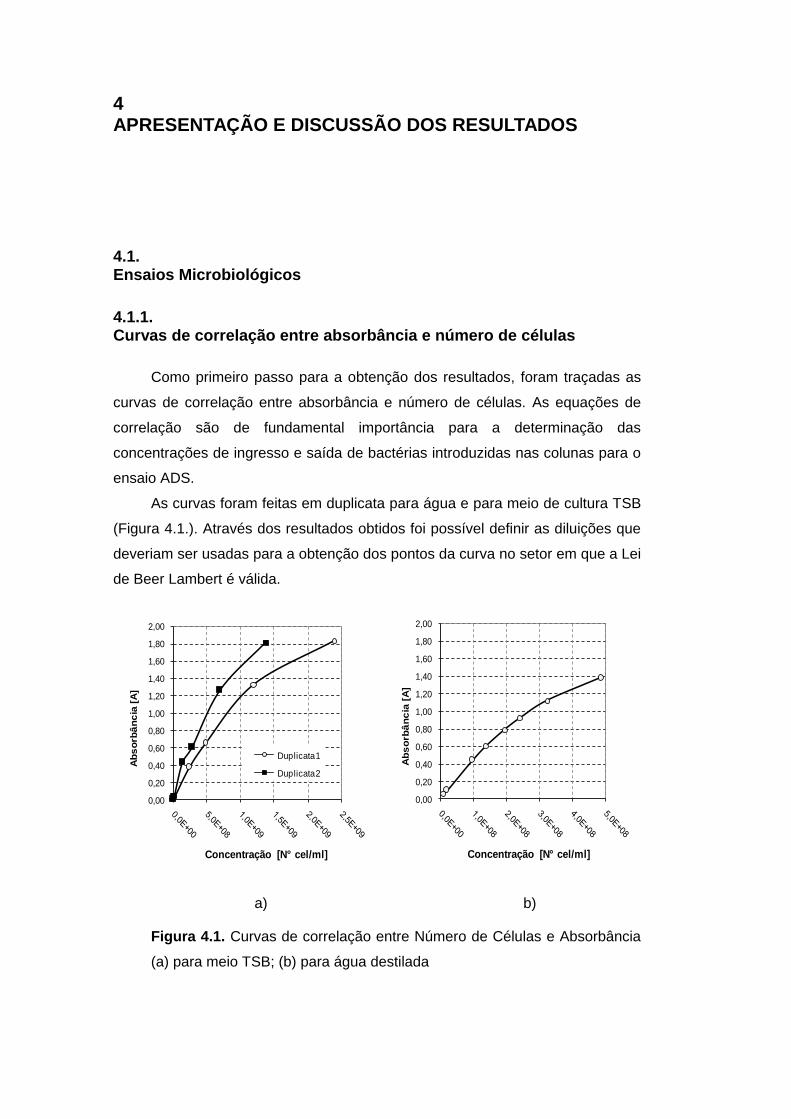
4 APRESENTAÇÃO E DISCUSSÃO DOS RESULTADOS
4.1. Ensaios Microbiológicos
4.1.1. Curvas de correlação entre absorbância e número de células
Como primeiro passo para a obtenção dos resultados, foram traçadas as
curvas de correlação entre absorbância e número de células. As equações de
correlação são de fundamental importância para a determinação das
concentrações de ingresso e saída de bactérias introduzidas nas colunas para o
ensaio ADS.
As curvas foram feitas em duplicata para água e para meio de cultura TSB
(Figura 4.1.). Através dos resultados obtidos foi possível definir as diluições que
deveriam ser usadas para a obtenção dos pontos da curva no setor em que a Lei
de Beer Lambert é válida.
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
1,60
1,80
2,00
Ab
so
rbâ
nc
ia [
A]
Concentração [N° cel/ml]
Duplicata 1
Duplicata 2
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
1,60
1,80
2,00
Ab
so
rbâ
nc
ia [
A]
Concentração [N° cel/ml]
a) b)
Figura 4.1. Curvas de correlação entre Número de Células e Absorbância
(a) para meio TSB; (b) para água destilada
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

80
A seguir, as Figuras 4.2 e 4.3 mostram as curvas de ajuste linear para as
correlações Absorbância versus Número de células, respectivamente para; a
leitura de absorbância das diferentes diluições da suspensão celular em meio
TSB a 600 nm e para a leitura da absorbância do mesmo material em água a
410 nm.
y = 3E-09x + 0,019R² = 0,996
0
0,2
0,4
0,6
0,8
1
1,2
0,00E+00 5,00E+07 1,00E+08 1,50E+08 2,00E+08 2,50E+08 3,00E+08 3,50E+08 4,00E+08
Ab
so
rbâ
nc
ia [
A]
Concentração [N° cel/ml]
Meio de Cultura TSB Lineal (Meio de Cultura TSB)
Figura 4.2. Curva de Correlação para células suspendidas em meio de
cultura TSB
y = 4E-09x + 0,017R² = 0,998
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,0E+00 2,0E+07 4,0E+07 6,0E+07 8,0E+07 1,0E+08 1,2E+08 1,4E+08 1,6E+08 1,8E+08
Ab
so
rbâ
nc
ia [
A]
Concentração [N° cel/ml]
Ensaio 1 Ensaio 2 Ensaio 3 Ensaio 4 Ensaio 5
Figura 4.3. Curva de Correlação para células suspendidas em água
destilada
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

81
Foram realizados de 2 a 5 ensaios para cada condição e após o ajuste da
curva, obtidos os coeficientes de correlação, R=0,996 para a leitura da
suspensão bacteriana em meio TSB e de R= 0,998 para a leitura da suspensão
bacteriana em água.
No caso da suspensão de células bacterianas em água destilada, foram
realizados 5 ensaios, para uma ampla gama de diluições dentro do ramo inicial
da curva apresentada no gráfico da Figura 4.1.b. Foi calculada a média dos
resultados obtidos e ajustados os pontos a uma curva linear neste caso a faixa
de validez da equação de ajuste, está definido entre 0 e 0,6.
Obtidas assim as equações de regressão e todas as condições
permanecendo constantes é suficiente ler a absorbância da suspensão e
calcular, usando essas equações, a concentração de células.
Para Meio de Cultura TSB 91076942
01930
ml
celNC
,
,aAbsorbânci
Para Água Destilada 91012654
01720
ml
celNC
,
,aAbsorbânci
4.1.2. Curva de Crescimento Escherichia coli em meio TSB
As curvas de crescimento obtidas para os ensaios realizados são
apresentadas no gráfico da Figura 4.4. São observados, para ambas as curvas,
limites de tempo coincidentes entre as fases de crescimento lag, exponencial e
estacionária.
Observa-se, no gráfico apresentado, o fim da fase logarítmica em
aproximadamente 24 horas após o início da incubação, seguida da fase
estacionária. O fim da fase logarítmica foi escolhido para ser usado como tempo
de incubação porque nesta fase o número de células que se dividem equivale a
quantidade de células que morrem, mantendo assim, o número de células
estável durante o tempo no qual os ensaios irão transcorrer. O número de
bactérias determinado após este período mostrou-se também constante, tanto
para estes, como para os ensaios seguintes, na ordem de 109 cel/mL, sendo
possível, após centrifugar as células obtidas do crescimento do inóculo,
ressuspendê-las no mesmo volume ou em volume maior, obtendo concentrações
iniciais para introduzir na coluna com células nas mesmas condições numéricas
para todos os ensaios ADS (entre 1,0 x 108 até 1,79 x 108 cel/mL). Este tempo
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

82
de incubação foi o mesmo utilizado por Foppen et al. (2005) para a obtenção de
aproximadamente 109 cel/mL de Escherichia coli ATCC25922.
0,000
0,500
1,000
1,500
2,000
2,500
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75
Ab
so
rvâ
nc
ia (A
)
Tempo (h)
Ensaio 1 Ensaio 2
Figura 4.4. Curvas de crescimento da Escherichia coli ATCC 11229
4.1.3. Avaliação da sobrevivência de E. coli em água
Esses ensaios foram realizados com o objetivo de avaliar a constância
numérica da população bacteriana presente na suspensão que iria ser
introduzida na coluna, durante os ensaios ADS.
Os Resultados apresentados na Tabela 4.1 indicam que o número de
células viáveis se manteve constante até o último período de tempo analisado
(35 horas), superior ao tempo de duração dos ensaios que tiveram até, no
máximo, aproximadamente 12 horas de duração. Este resultado garantiu que o
número de células da bactéria E. coli ATCC11229 suspensas em água destilada,
não sofreria variações no tempo, sendo desprezado o efeito do crescimento e de
morte dos microrganismos durante a execução dos ensaios de batelada e
principalmente nos ensaios ADS.
Fase lag
Fase log
Fase Estacionaria
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

83
Tabela 4.1. Cálculo da concentração de células viáveis suspensas em
água destilada pela Técnica dos Tubos Múltiplos
Tempo NMP/mL
* 9,59E+08
2 2,40E+08
11 2,40E+08
25 2,40E+08
35 2,40E+08
Tempo de incubação 24 horas
* Concentração inicial determinada através de leitura de densidade óptica (Número de
células Totais.
4.2. Ensaio de Adsorção em Batelada
Os ensaios de adsorção foram executados sob condições de temperatura
entre 22 e 24°C; pH neutro; e baixa força iônica, entre 10 e 20 µS/cm.
Os resultados finais, dos ensaios de adsorção da E. coli ATCC11229 em
areia de quartzo limpa, obtidos para cada concentração (109, 108, 107 e 106)
encontram-se resumidos na Tabela 4.2. Os dados determinados no laboratório
se encontram dispostos no Apêndice A.
Tabela 4.2. Resultados obtidos através da técnica de contagem em placa
Co Nt Ns Pa Cs Cw
UFC/mL UFC UFC % cel/g cel/mL
3,08E+09 6,15E+10 1,35E+10 78,0 2,40E+10 6,77E+08
3,91E+08 3,91E+08 3,91E+08 31,9 1,25E+09 2,66E+08
2,34E+07 4,68E+08 3,37E+08 25,2 6,56E+07 1,68E+07
2,13E+06 4,26E+07 3,81E+07 10,6 2,26E+06 1,90E+06
Co :concentração Inicial da suspensão bacteriana; Nt :número total de células na suspensão
bacteriana; Ns: número total de células no sobrenadante; Pa :percentagem de adsorção;
Cs:concentração de células sorvidas no solo; Cw:concentração de células na solução.
Os dados obtidos para as concentrações de células da bactéria no solo e
no sobrenadante, Cs e Cw respectivamente, foram plotados em um gráfico de
escala logarítmica dupla (Figura 4.5) e, ajustados à isoterma de Freundlich
(Cs=K Cw m) para a determinação do potencial máximo de sorção da bactéria na
areia de quartzo limpa.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

84
y = 0,001x1,465
R² = 0,972
1,00E+03
1,00E+06
1,00E+09
1,00E+12
1,00E+05 1,00E+07 1,00E+09
Cs
[c
el/g
]
Cw [cel/mL]
Figura 4.5. Isoterma de adsorção da E. coli em areia de quartzo
A partir da equação de ajuste são obtidos os parâmetros KD=0,001 mL/g e
m=1,465, com um coeficiente de determinação r2=0,972. O valor do expoente
reflete a forma da curva, e neste caso, mostra que a relação entre Cs e Cw não foi
completamente linear. O valor de KD reflete o alcance da sorção, assim
KD= 0,001 mL/g significa que de cada célula que ficou adsorvida na areia, 1000
se mantiveram no sobrenadante, mostrando uma baixa adesão da bactéria a
este solo.
Jiang et al. (2007), estudaram a adsorção das células da bactéria E. coli
(strain D) suspensas em água deionizada em areia de sílica fina e grossa
separadamente, e obtiveram valores de KD de 10 e 0,01 mL/g respectivamente,
apontando uma diferença de até 3 ordens de grandeza entre ambos os valores.
No caso deste estudo foi obtido um valor de KD de uma ordem de grandeza
menor que o valor obtido por Jiang et al. (2007) para areia grossa. Esta diferença
pode ser atribuída à pequena massa de 2 gramas utilizada nos ensaios de
batelada, além de se tratar com uma areia de quartzo bem graduada.
Observa-se que a percentagem de adsorção se incrementa na medida em
que se aumenta a concentração inicial da suspensão bacteriana (Tabela 4.2)
mostrando a dependência da adsorção com relação à concentração da
suspensão, ainda utilizando areia de quartzo da mesma distribuição
granulométrica para todos os casos.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

85
O valor do coeficiente de distribuição KD próximo a zero obtido a partir do
ajuste, indica que a adesão das bactérias na areia de quartzo é nula, o que é de
se esperar devido à carga elétrica negativa da E. coli e da areia de quartzo,
criando uma força eletrostática repulsiva. A utilização de água destilada como
solução, contribui a criar condições desfavoráveis para a adesão em decorrência
de sua baixa força iônica. Isto sugere que a presença de qualquer oxido na areia
de quartzo tem efeito desprezível na deposição de bactérias, que poderia ter
sido originado pelo processo de peneiramento mecânico (Brown et al., 2002).
Em função destes resultados também se desprezam os efeitos de
bloqueamento (blocking), já que este processo está presente em um sistema
onde tanto o colóide quanto o meio poroso estão opostamente carregados
(Deshpande e Shonnard,1999).
4.3. Ensaio ADS
Foram conduzidos 8 ensaios ADS, um de cada vez, devido ás exigências
do procedimento do ensaio. Todos os ensaios foram executados sob condições
de fluxo em regime permanente e na temperatura ambiente entre 22 e 24°C, pH
e força iônica similares às condições de execução dos ensaios de batelada. Na
Tabela 4.3 são apresentadas as condições hidráulicas do sistema durante a
etapa de saturação.
Tabela 4.3. Condições hidráulicas durante a saturação da areia
ENSAIO i Q K vd pV pH EC ml/min cm/d cm/d μS/cm
1 0,09 4,70 2,89E-02 213,48 4,0 7,3 9,8
2 0,09 3,26 2,01E-02 148,24 3,0 6,5 18,1
3 0,09 3,62 2,23E-02 164,47 4,6 _ _
4 0,09 4,70 2,89E-02 213,48 4,2 6,7 9,0
5 0,09 5,46 3,36E-02 248,14 4,0 6,8 8,4
6 0,10 5,02 2,76E-02 228,05 4,4 7,0 11,4
7 0,09 4,29 2,64E-02 194,88 5,8 7,1 12,5
8 0,09 3,09 1,90E-02 140,50 4,9 6,9 11,4 i: gradiente hidráulico; Q: vazão; Ks: condutividade hidráulica saturada; vd :velocidade de Darcy; PV: volume de
poro; EC: condutividade elétrica.
A saturação da areia foi realizada aplicando o mesmo gradiente hidráulico
de 0,09, considerando-se esse valor baixo suficiente para uma boa saturação da
areia e alto o suficiente para permitir a passagem de 3 a 4 volume de poros em
um tempo apropriado para proceder à preparação da seguinte etapa do ensaio.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

86
Na Tabela 4.3 também são apresentados os valores de pH e condutividade
elétrica da água destilada medida no final da etapa de saturação, e considerados
os valores iniciais na etapa de injeção da suspensão bacteriana.
No ensaio ADS 1 o gradiente hidráulico imposto foi o mínimo possível
devido às baixas velocidades que apresenta um aquífero natural (<1m/d). Assim,
nos ensaios ADS posteriores o gradiente foi sendo incrementado, sendo
realizados os ensaios ADS dentro de uma faixa de gradiente hidráulico de 0,03
até 0,36.
A Tabela 4.4 apresenta os valores de gradiente definidos para cada ensaio
e os valores médios de vazão de saída do efluente (Q), da velocidade de Darcy
(vd) e da condutividade hidráulica saturada (Ks), embora os valores de
condutividade hidráulica não tenham se mantido constantes, particularmente
para gradientes maiores que 0,24. Também são apresentados os valores de
densidade seca (ρbulk), determinada no final dos ensaios e com base a este valor
calculados os valores de porosidade (n) e velocidade intersticial (v). A variação
da condutividade hidráulica é analisada no item 4.3.2.
Tabela 4.4. Condições hidráulicas do sistema experimental durante a
injeção da suspensão de E. coli
ENSAIO Co i *vd *KS *Q ρbulk n v
cel/mL cm/d cm/s mL/min g/cm3 cm/d
1 1,07E+08 0,03 70,17 2,66E-02 1,54 1,89 0,28 247,36
2 1,65E+08 0,06 205,57 3,93E-02 4,52 1,86 0,29 698,46
3 1,55E+08 0,06 179,05 3,43E-02 3,94 1,86 0,30 604,93
4 1,79E+08 0,06 174,01 3,33E-02 3,83 1,86 0,29 592,55
5 1,63E+08 0,16 374,39 2,79E-02 8,23 1,87 0,29 1280,91
6 1,40E+08 0,24 544,06 2,67E-02 11,97 1,87 0,29 1861,41
7 1,31E+08 0,24 411,13 2,02E-02 9,04 1,89 0,28 1442,92
8 1,31E+08 0,36 681,32 2,19E-02 14,98 1,90 0,28 2446,46 * Valor médio para cada ensaio
4.3.1. Curvas de chegada
As curvas de chegada para E. coli construídas para 8 ensaios ADS com
injeção continua são discutidos nesta seção. As Tabelas de amostragem,
controle de saída de volume de poros e determinação do número de
microrganismos são apresentados no Apêndice C1.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

87
Na Figura 4.6, são plotadas as curvas de chegada de células de E. coli,
para cada ensaio, junto com os pontos de controle medidos com a técnica de
contagem em câmara de Neubauer. Observa-se a proximidade dos pontos
determinados em CNB com os pontos definidos através das leituras de
absorbância, o que é de se esperar em virtude dos valores de absorbância
medidos estiver entre 0,002 até 0,707, ou seja, dentro da faixa de validade da
equação de correlação.
A curva de transporte caracteriza-se por apresentar forma de uma
sigmóide, esta configuração é observada apenas nas curvas correspondentes a
baixos gradientes hidráulicos isto é para 0,03, 0,06 até 0,16. Para gradientes
maiores, a curva e mais parecida com uma função quadrática.
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
0 2 4 6 8 10 12 14 16
C/C
o
volume de poro
Ensaio 1 (i=0,03) Contagem em CNB
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
0 2 4 6 8 10 12 14 16
C/C
o
volume de poro
Ensaio 2 (i=0,06) Contagem em CNB
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
0 2 4 6 8 10 12 14 16
C/C
o
volume de poro
Ensaio 3 (i=0,06) Contagem em CNB
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
0 2 4 6 8 10 12 14 16
C/C
o
volume de poro
Ensaio 4 (i=0,06) Contagem em CNB
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

88
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
0 2 4 6 8 10 12 14 16
C/C
o
volume de poro
Ensaio 5 (i=0,16) Contagem em CNB
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
0 2 4 6 8 10 12 14 16
C/C
o
volume de poro
Ensaio 6 (i=0,24) Contagem na CNB
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
0 2 4 6 8 10 12 14 16
C/C
o
volume de poro
Ensaio 7 (i=0,24) Contagem em CNB
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
0 2 4 6 8 10 12 14 16
C/C
o
volume de poro
Ensaio 8 (i=0,36) Contagem em CNB
Figura 4.6. Curvas de chegada para cada ensaio ADS
Pode-se observar uma estabilização da concentração do efluente antes da
curva superar o valor de C/Co igual à unidade, para as curvas que apresentam o
formato de uma sigmóide. A formação deste patamar sugere a capacidade de
atenuação de E. coli pela areia de quartzo. Porém observa-se também nos
ensaios 1 e 3, uma tendência ao desprendimento das células que puderam ficar
retidas na medida que o fluxo continua.
No entanto, a determinação de C/Co no patamar da curva permite calcular
a eficiência real do coletor único (η) através da equação (2.27), valor
adimensional que representa as oportunidades que o biocolóide tem para colidir
com os grãos de areia. O valor teórico da eficiência do coletor calculada através
da equação (2.22) comparado com o valor real ou experimental determina a
eficiência da adesão (α).
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

89
Neste caso, o patamar claramente definido só foi atingido na curva que
corresponde ao ensaio 4, sendo o valor de C/Co igual a 0,9 aproximadamente.
Nesse caso o valor da eficiência de colisão (α) é de 0,05, o que é de se esperar
devido a que o ensaio foi realizado sob condições desfavoráveis (presença da
barreira de energia repulsiva) e que ainda mais reforça o resultado obtido no
ensaio de adsorção exposto no item 4.2.
A curva correspondente ao menor gradiente (i=0,03) mostra um patamar
temporário para o valor de C/Co igual a 0,5. A formação deste patamar parece
estar relacionada com a baixa velocidade do ensaio, contrário da análise
apresentada por Foppen et al. (2005), que sugeriram que o patamar não é
afetado pelas variações da taxa de fluxo e sim pela concentração inicial, sendo
que neste estudo a concentração inicial foi mantida constante. Mas também, de
acordo com Foppen et al. (2005), a formação do patamar na curva de chegada
pode ser atribuída à presença de poros pequenos demais para a passagem da
bactéria através de eles, o que pode ter influenciado nos resultados do presente
estudo.
Esse processo conhecido como filtração mecânica, pressupõe que o
volume destinado à retenção de partículas (bactérias) é fixo para um
determinado meio poroso devido a sua dependência com a geometria do grão e
da bactéria. A determinação da quantidade de bactérias que o meio poroso é
capaz de reter pode ser expressa pelo valor do volume de bactérias retidas por
unidade de volume de meio poroso (σ) e os cálculos, a análise e a possível
explicação para o comportamento das curvas de chegada, com base ao
processo associado, são discutidas no item 4.4.2.
4.3.2. Fator de Retardamento e Dispersão Hidrodinâmica
Os indicadores de transporte: retardamento (R) e dispersão hidrodinâmica
(Dh) foram determinados para cada ensaio conforme o método indicado em
Azevedo et al. (2002).
Esse método permite obter, de forma gráfica e direta, o valor do
retardamento a partir das curvas de chegada, como também calcular o valor da
dispersão hidrodinâmica através da determinação gráfica do parâmetro b. Os
gráficos mostrando a aplicação do método descrito em Azevedo et al. (2002),
são apresentados no Apêndice B2.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA
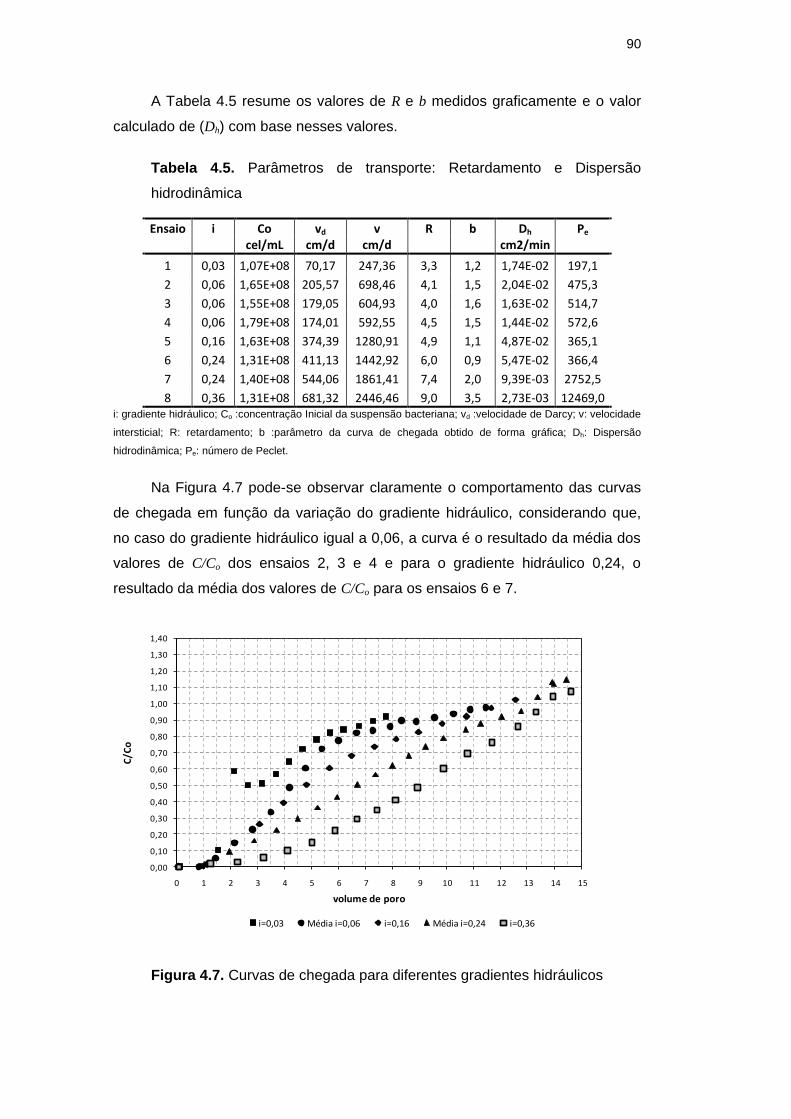
90
A Tabela 4.5 resume os valores de R e b medidos graficamente e o valor
calculado de (Dh) com base nesses valores.
Tabela 4.5. Parâmetros de transporte: Retardamento e Dispersão
hidrodinâmica
Ensaio i Co vd v R b Dh Pe cel/mL cm/d cm/d cm2/min
1 0,03 1,07E+08 70,17 247,36 3,3 1,2 1,74E-02 197,1
2 0,06 1,65E+08 205,57 698,46 4,1 1,5 2,04E-02 475,3
3 0,06 1,55E+08 179,05 604,93 4,0 1,6 1,63E-02 514,7
4 0,06 1,79E+08 174,01 592,55 4,5 1,5 1,44E-02 572,6
5 0,16 1,63E+08 374,39 1280,91 4,9 1,1 4,87E-02 365,1
6 0,24 1,31E+08 411,13 1442,92 6,0 0,9 5,47E-02 366,4
7 0,24 1,40E+08 544,06 1861,41 7,4 2,0 9,39E-03 2752,5
8 0,36 1,31E+08 681,32 2446,46 9,0 3,5 2,73E-03 12469,0 i: gradiente hidráulico; Co :concentração Inicial da suspensão bacteriana; vd :velocidade de Darcy; v: velocidade
intersticial; R: retardamento; b :parâmetro da curva de chegada obtido de forma gráfica; Dh: Dispersão
hidrodinâmica; Pe: número de Peclet.
Na Figura 4.7 pode-se observar claramente o comportamento das curvas
de chegada em função da variação do gradiente hidráulico, considerando que,
no caso do gradiente hidráulico igual a 0,06, a curva é o resultado da média dos
valores de C/Co dos ensaios 2, 3 e 4 e para o gradiente hidráulico 0,24, o
resultado da média dos valores de C/Co para os ensaios 6 e 7.
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
1,40
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
C/C
o
volume de poro
i=0,03 Média i=0,06 i=0,16 Média i=0,24 i=0,36
Figura 4.7. Curvas de chegada para diferentes gradientes hidráulicos
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

91
Observa-se que todas as curvas atingem a metade da concentração inicial,
(C/Co=0,5) para valores de volume de poro maiores do que a unidade, ou seja,
todas as curvas de chegada apresentam retardamento. Este incremento do fator
de retardamento com relação ao gradiente imposto apresenta um
comportamento linear contrário ao comportamento esperado para um problema
de transporte de contaminantes, como mostra o gráfico da Figura 4.8, onde na
medida em que a velocidade aumenta o retardamento diminui. E ainda mais,
considerando o resultado do ensaio de adsorção, poderia se esperar que as
curvas não apresentem retardamento nenhum.
R = 16,47 i+ 2,893R² = 0,932
0,00
2,00
4,00
6,00
8,00
10,00
0 0,05 0,1 0,15 0,2 0,25 0,3 0,35 0,4
R
Gradiente i
Figura 4.8. Variação do fator de Retardamento com relação ao gradiente
hidráulico
y = 0,053x - 0,001R² = 0,887
0,00
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2
D [c
m2/m
in]
Velocidade [cm/min]
Figura 4.9. Variação da Dispersão Hidrodinâmica com relação à
velocidade de poros
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

92
No gráfico da Figura 4.9, mostra-se a relação entre a dispersão e a
velocidade. Os valores de dispersão hidrodinâmica foram calculados em função
de R e b apresentados na Tabela 4.5, assim como também em função da
velocidade de poros, que corresponde à média dos valores medidos para cada
ensaio. Através da equação de correlação pode-se obter o valor de
dispersividade αL=0,053 cm.
Teoricamente existe uma relação linear dada pela equação (2.9) entre
ambos os parâmetros dispersão hidrodinâmica e velocidade de poros, sendo a
inclinação da reta o fator da dispersividade longitudinal (αL) e a interseção com o
eixo vertical a difusão molecular (Dm). No caso do presente estudo, os valores
que apresentam esta tendência teórica são os primeiros pontos do gráfico, ou
seja, para os ensaios com gradiente menor que 0,24 e como será visto mais
adiante, para aqueles ensaios em que não houve diminuição notável da
condutividade hidráulica. Assim, a redução da dispersividade hidrodinâmica a
partir da velocidade de 14,43 m/d (1 cm/min) pode estar relacionada com a
diminuição de canais de poro disponíveis para a propagação do fluxo, que se
manifesta através da redução da condutividade hidráulica.
Nos gráficos da Figura 4.10 apresentam-se as variações de condutividade
hidráulica com relação ao número de volume de poros. Os valores de
condutividade hidráulica foram obtidos durante a execução do ensaio ADS
através da medida da vazão do efluente. Note-se que esses valores assim
determinados no início dos ensaios encontram-se muito próximos ao valor de
condutividade hidráulica determinado durante a caracterização da areia de
quartzo, valor este representado pela linha tracejada nos gráficos da Figura 4.10.
Cada um dos ensaios ADS foi realizado aplicando um gradiente hidráulico
constante, porém os ensaios executados com valores de gradiente superiores a
0,24 (ensaios 7 e 8) mostraram uma notável diminuição da condutividade
hidráulica (de 0,03 até 0,01 cm/s), como pode ser observado na Figura 4.10 (c) e
em consequência também da velocidade de percolação. Esta observação,
permite afirmar que durante os ensaios 7 e 8 houve uma diminuição da
porosidade, mudando as condições iniciais desses ensaios, e
consequentemente conduzindo a um erro no cálculo da dispersão hidrodinâmica
correspondente a esses ensaios, conforme valores plotados no gráfico da
Figura 4.9.
Nos casos de gradientes mais baixos, a permeabilidade também
apresentou variações, mas em menor proporção e com uma tendência distinta,
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

93
às vezes aumentando outras diminuindo, que pode se atribuir a variações da
velocidade de agitação da suspensão dentro do frasco de Mariotte.
0,005
0,015
0,025
0,035
0,045
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Ks
cm
/s
volume de poro
Ensaio 1 (i=0,03)
0,005
0,015
0,025
0,035
0,045
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Ks
cm
/s
volume de poro
Ensaio 5 (i=0,16)
0,005
0,015
0,025
0,035
0,045
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Ks
cm/s
volume de poro
Ensaio 6 (i=0,24) Ensaio 7 (i=0,24) Ensaio 8 (i=0,36)
Figura 4.10. Variabilidade da condutividade hidráulica no ensaio ADS
(c)
(a)
(b)
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

94
De acordo com McDowell-Boyer et al. (1986), a mais importante
manifestação dos mecanismos de filtração é a redução da condutividade
hidráulica como o entupimento dos poros, o que explica a diminuição da
condutividade hidráulica observada.
4.3.3. Ensaios de controle
Na Tabela 4.6 são apresentados os resultados obtidos através da técnica
dos Tubos Múltiplos correspondentes ao ensaio ADS 1, para a determinação da
concentração de bactérias retidas ao longo da coluna de areia após a passagem
da suspensão bacteriana de forma contínua e constante. Pode-se observar que
a maioria da massa de E. coli foi depositada próximo ao local de ingresso da
suspensão na coluna.
Tabela 4.6. Concentração de E. coli retida ao longo da coluna de areia
(Ensaio ADS 1)
Seção da
coluna NMP/100mL NMP/10mL
Peso do solo úmido (g)
umidade %ω
Peso solo
seco Ws (g)
Concentração de células por grama de
solo (cel/g)
Base 2,10E+10 2,10E+09 1,600 6,8 1,50 1,40E+09
Metade 9,30E+08 9,30E+07 1,139 6,9 1,07 8,73E+07
Topo 4,00E+07 4,00E+06 1,631 21,0 1,35 2,97E+06
As determinações da concentração de bactérias ao longo da coluna de
areia indicaram que, à medida que esses microrganismos percolam através da
coluna, sua concentração diminui, com presença de uma concentração celular
mais elevada nas primeiras camadas e menor concentração no final da coluna.
Este comportamento é de se esperar considerando a equação da eficiência do
filtro formulada conforme a teoria da filtração equação (2.25).
Na Figura 4.11 são apresentadas as curvas granulométricas obtidas
através da análise por peneiramento, efetuada para amostras correspondentes a
três seções ao longo da coluna de areia comparada com a curva granulométrica
determinada para a caracterização do meio poroso (linha tracejada). Os
resultados mostram que na houve diferença significativa entre as curvas
granulométricas correspondentes a porções de areia retiradas da base, metade
e topo da coluna, o que permite afirmar que a distribuição dos grãos foi mantida
constante ao longo da coluna de areia durante a execução dos ensaios ADS.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

95
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
90,0
100,0
0,1 1 10
% q
ue
pas
a
abertura (mm)
Coluna Base Mitade Topo
Figura 4.11. Distribuição do tamanho dos grãos ao longo da coluna de
areia
4.4. Discussão
4.4.1. Retardamento
Valores do fator de retardamento para Escherichia coli, determinados
através de experimentos de laboratório, utilizando colunas de areia com injeção
contínua são reportados pela literatura como próximos à unidade, a partir de
0,56 até 2,06 (Pang et al., 2003; Powelson e Mills., 2001 citado em Foppen e
Schijven, 2006).
Como foi visto para este estudo, os valores de retardamento obtidos a
partir da curva de transporte, variam proporcionalmente com a velocidade, a
partir de valores de 3 até 9, sendo muito superiores aos valores encontrados na
literatura.
Normalmente a existência de fatores de retardamento muito elevados é
atribuída à participação de processos de adsorção físico-química durante o
transporte, ou seja, a interações entre os grãos do solo e o contaminante
(bactéria), devido a forças físicas (forças de van der Waals e forças
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

96
eletrostáticas) ou, em alguns casos ligações químicas, que originam a fixação
das células da bactéria aos grãos do solo.
Contudo, no presente estudo os ensaios foram executados sob condições
desfavoráveis à adsorção físico-química, devido à utilização de água destilada,
que apresenta baixa condutividade elétrica e pH próximo ao neutro, além da
carga eletrostática negativa do quartzo e da E. coli, esta última por sua natureza
hidrofílica. Este fato foi confirmado através dos resultados obtidos nos ensaios
de batelada obtendo um coeficiente de partição próximo a zero (KD=0,001mL/g)
permitindo afirmar que, sob ás condições dos ensaios de transporte as bactérias
são muito pouco retidas no meio poroso.
Portanto, o fator de retardamento calculado com base ao coeficiente de
partição, através da equação (2.12) resulta ser aproximadamente igual à
unidade, contrariamente às observações realizadas com base nas curvas de
chegada nos ensaios ADS. Nesse sentido, a participação do processo de
adsorção físico-química no transporte de E. coli suspensa em água destilada, em
areia de quartzo nas condições avaliadas, é considerada desprezível.
Por outro lado, a teoria da filtração desenvolvida para estudar a deposição
de partículas em uma primeira etapa, na qual o leito de areia encontra-se limpo,
está baseada na proporcionalidade entre o incremento da taxa de deposição de
primeira ordem com relação ao incremento da velocidade dos poros de acordo
com a equação (2.28) (Tufenkji, 2004; Kretzschmar et al., 1999). Resultados
obtidos por Pang et al. (2003) para E. coli, utilizando água subterrânea e solos
nativos, verificaram esta relação entre a taxa de deposição e a velocidade de
fluxo, com valores de retardamento próximos à unidade, concluindo que a
filtração é o processo dominante na remoção ou atenuação de bactérias.
Contudo, a teoria da filtração não prediz adequadamente o comportamento da
deposição, em presença de interações repulsivas da dupla camada eletrostática
(Kretzschmar et al., 1999), condições impostas no presente trabalho. Além disso,
não foi encontrada na literatura nenhuma relação entre a taxa de deposição e o
retardamento.
Outros estudos (Li et al., 2004, Bradford et al., 2006) avaliaram a retenção
da bactéria E. coli para diferentes velocidades de fluxo, sob condições não
favoráveis. Os resultados destas pesquisas mostraram tendência de produzir
elevadas concentrações no efluente para maiores velocidades de fluxo,
contrariamente a teoria da filtração anteriormente descrita. Observaram também
que a forma da curva de chegada é afetada pela ocorrência da filtração
mecânica, principalmente no caso de areias finas.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

97
Consequentemente, uma explicação para a obtenção de fatores de
retardamento muito elevados, como os obtidos no presente trabalho, pode ser
atribuída à influência da filtração mecânica, como é descrito a seguir.
4.4.2. Filtração Mecânica como processo dominante no transporte de E. coli nos ensaios ADS
De acordo com Foppen et al. (2005), a capacidade do meio poroso para
capturar as bactérias pode ser determinada através do cálculo do volume de
poros disponíveis para a retenção de bactérias por unidade de volume de meio
poroso (σ). De acordo com a literatura, a significância do processo de filtração
mecânica está associada à relação d/dg, considerando os diâmetros das
partículas e dos grãos completamente esféricos. Devido ao fato da E. coli ser
uma bactéria em forma de bastão, para este estudo foi utilizado como diâmetro
da bactéria (d), o valor médio da dimensão maior, isto é 3 µm. Utilizar a
dimensão menor de (1µm) subestimaria a influência do processo de filtração
mecânica. No caso do diâmetro de grão foram utilizados para os cálculos o
diâmetro característico d50= 920 µm e o diâmetro efetivo d10=180 µm. Com estas
definições, a relação d/dg foi igual a 0,003 e 0,02 para d50 e d10, respectivamente,
que resulta ser maior do que 0,002, para valores inferiores a este. De acordo
com Tufenkji (2004), o processo de filtração mecânica não tem impacto sobre o
transporte. Devido ao valor muito baixo para d/dg comparado com valores
referenciais antigamente definidos como 0,05 (Pekdeger e Matthess, 1983)
esperava-se que filtração mecânica tivesse pouca significância. Porém o
comportamento das curvas de chegada mostra o contrário.
Para o melhor entendimento do processo de filtração mecânica, no
presente estudo, foi calculado o valor de (σ) utilizando a equação (2.29),
proposta por Herzig (1970) e também através da integração da função de
densidade de poro, proposta por Wise et al. (1994), ambas as estratégias citadas
em Foppen et al. (2005) e descritas no item 2.6.4. Os parâmetros de van
Genuchten utilizados para o cálculo dos coeficientes da função de poro foram
determinados pelo método de redes neuronais Schaap et al. (1998).
Considerando a velocidade de poros constante para cada ensaio, como
também a concentração inicial, haverá um tempo para o qual todos os sítios
disponíveis estejam preenchidos e, consequentemente, também haverá um
número definido de volume de poros, que pode ser calculado através da relação
dada pela equação (2.30). A partir do estudo realizado por Foppen et al. (2005)
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

98
foi deduzido o valor do número de células por grama de biomassa igual a
6,36E+11, que permite transformar as unidades de concentração inicial de
(cel/mL) para (g/mL) e assim calcular o número de volume de poros que são
necessários passar através da coluna de areia para preencher os canais de poro
que teoricamente estão disponíveis para a retenção de células. Os resultados
são apresentados na Tabela 4.7.
Tabela 4.7. Número de volume de poros necessários para preencher com
bactérias os espaços de poro destinados à filtração mecânica
σ [-] % σ Co (cel/mL) Co (g/mL) pV Smax
Equação Herzig et al., 1970
dg=d50 6,71E-06 0,001 1,46E+08 2,30E-04 0,10 3,58E-06
dg=d10 3,98E-04 0,040 1,46E+08 2,30E-04 6,16 2,12E-04
Função de densidade de poro
2,47E-03 0,247 1,46E+08 2,30E-04 38,15 1,32E-03
Co:valor médio de todos os ensaios. Para a determinação da função de densidade de poro foram
considerados foram considerados os parâmetros de van Genuchten αv=3,01 (1/m) e nv=3,9966
com base nas propriedades da areia: ρbulk=1,89 e n=0,3.
Note-se que o valor do volume de bactéria que pode ser retido no meio
poroso (σ) de acordo com a equação da função de densidade de poros é
superior ao valor calculado através das aproximações baseadas na relação de
aspecto (d/dg), que pode ser interpretado pelo fato de que esta última
aproximação considera os grãos do solo como esferas do mesmo diâmetro
enquanto que a função de densidade de poro considera a distribuição do
tamanho dos poros.
Assim, os resultados obtidos indicam que, para uma areia bem graduada
como a utilizada neste estudo considerar o valor de diâmetro efetivo está mais
próximo do comportamento real do que considerar o diâmetro característico d50.
Por outro lado, com base ao ensaio de capilaridade, foi determinado um
diâmetro médio de poro igual a 352 μm, que resulta maior do que 124 μm, este
último calculado a partir da função de densidade de poro. Isto significa que o
valor de (σ) real será menor que 0,25% valor apresentado na Tabela 4.7. O valor
mais próximo à realidade pode ser então calculado mediante a determinação da
curva característica da areia no laboratório.
Contudo, as curvas de chegada obtidas neste trabalho mostraram que
houve um tempo para o qual a concentração do efluente superou o valor da
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

99
concentração inicial, o que sugere que ainda existam espaços disponíveis à
deposição devido ao processo de filtração mecânica, a deposição resulta ser
reversível devido ao elevado gradiente hidráulico aplicado nos ensaios. Só foi
possível observar a formação do patamar para os ensaios realizados com
gradientes menores que 0,06. Com maior definição para os ensaios 2 e 4 (v=5,9
m/d) e, no caso do ensaio 1 (v =2,47 m/d), foi observado um patamar temporário
para um valor de C/Co de aproximadamente 0,5.
Este comportamento pode ser explicado mediante a análise da Figura
4.12, onde pode se observar a forma como o solo pode reter bactérias, tanto nas
contrições dos poros como nos canais de poro que apresentam uma
extremidade com diâmetro igual a zero, esses últimos denominados por Foppen
et al. (2005) como DEP (Dead End Pore).
a) Tipo A. Constrição de poro b) Tipo B. DEP (Dead End Pore)
Figura 4.12. Tipos de locais dentro do meio poroso, susceptíveis à
retenção de bactérias devido ao processo de Filtração mecânica
Com base a essas considerações, para ensaios realizados com elevadas
velocidades (>6 m/d), a areia é capaz de reter bactérias mais rapidamente, seja
nas regiões tipo A ou tipo B. Porém, forças hidrodinâmicas, consequência da
elevada velocidade do ensaio, podem atuar com maior intensidade sobre as
bactérias retidas na região tipo A, forçando-as a atravessar a estreiteza do canal
de poro. Esta suposição tem como base a afirmação feita por Bradford et
al.(2006), que assumiram que as células de E. coli podem se agregar quando
um grande número delas monodispersas são depositadas na constrição dos
poros ou nos locais de filtração mecânica. Quando a E. coli depositada atinge
uma concentração crítica no local de filtração mecânica, a E. coli O157:H7
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

100
agregada pode ser liberada dentro a solução aquosa como resultado de forças
hidrodinâmicas de cisalhamento.
Este fenômeno pode explicar também as elevadas concentrações no
efluente determinadas para velocidades maiores a 6m/d (C/Co>1), evidenciando
que o processo de filtração mecânica para velocidades de fluxo muito elevadas é
menos eficiente como processo de atenuação de patogênicos.
O comportamento observado confirma a afirmação, realizada por Bradford
et al. (2002), que um modelo de remoção de partículas que considera tanto a
adsorção cinética (irreversível) junto com a filtração mecânica é mais real,
particularmente para sistemas com partículas e tamanhos de grão
intermediários.
No entanto, o processo de filtração mecânica tem uma clara influência
sobre a atenuação de bactérias, vários estudos (Bradford et al., 2006; Li et al.,
2004) demonstraram que quanto mais baixa a velocidade de fluxo, maior a
capacidade de atenuação do meio poroso, o que é observado através da
estabilização da curva para um determinado valor de C/Co menor que a unidade.
Este tipo de deposição devido à filtração mecânica foi mostrado por Bradford
(2006) através de fotos ampliadas 600 vezes (Figura 4.13). É por esta razão que
o processo de filtração mecânica é modelado também como um processo
irreversível (Pang et al., 2003).
Figura 4.13. Fotos ampliadas 600 vezes, da deposição de E.coli O157: H7
em areia de 150 μm (Bradford et al., 2006)
4.4.3. Influencia das propriedades do meio poroso
Foi mostrado que a filtração mecânica pode explicar o comportamento das
curvas de chegada, mas por outro lado, nesta seção é discutida a influência da
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

101
forma do grão, além do tamanho como sobre os processos de transporte e
retenção.
No estudo realizado por Bradford et al. (2002), utilizando colóides
carboxílicos, carregados negativamente e areia quartzosa de diferentes
tamanhos, os autores concluíram que o pico do efluente decresce com o
incremento do tamanho dos colóides e diminuição do tamanho dos grãos,
variáveis que foram mantidas constantes no presente estudo. Também
concluíram, que a concentração de colóides no efluente é altamente dependente
da distribuição do tamanho dos grãos, porém utilizaram areias com coeficientes
de uniformidade entre 1 e 3, menores que o valor de 6,11 correspondente à areia
utilizada neste trabalho.
Embora os experimentos realizados por Bradford et al. (2002) não tenham
sido dirigidos à determinação do fator de retardamento, não foi observado
retardo nas curvas de chegada, o que poderia estar relacionado com a utilização
de meios porosos com distribuição uniforme dos grãos.
Foppen e Shijven (2006) apresentam resultados obtidos por Mathess et al.
(1995a), que demonstram que o coeficiente de uniformidade não afeta o valor do
coeficiente de filtração (ou coeficiente de adesão katt) que está diretamente
relacionado com o valor de C/Co (patamar da curva de chegada). Entretanto, a
influência deste parâmetro na forma da curva de chegada e consequentemente
no retardamento, não foi analisada.
Portanto, pode-se afirmar que embora o coeficiente de uniformidade não
afete a eficiência de filtração, pode, sim, influenciar a forma da curva chegada.
Quanto à análise da forma dos grãos, experimentos realizados por Tufenkji
et al. (2004) comparam o transporte de Criptosporidium parvum oocysts e
microesferas de látex, através de areia de quartzo e de esferas de vidro de
tamanho comparável com o tamanho da areia. Estes estudos revelaram que a
forma do grão contribui consideravelmente no potencial de filtração mecânica do
meio poroso, como pode ser observado na Figura 4.14. Deste modo, Tufenkji et
al. (2004) demonstraram que comparado com a sílica esférica, a sílica britada
resulta numa maior atenuação de microesferas e Cryptosporidium devido ao
efeito de filtração mecânica.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

102
Figura 4.14. Imagens ESEM (Environmental Scanning Electron
Microscope) de a) leito de esferas de vidro (ampliadas 200 vezes, b)
quartzo com forma irregular (ampliada 500 vezes) (Tufenkji et al., 2004)
Com base neste estudo, pode se afirmar que a forma predominantemente
angulosa da areia de quartzo comercial utilizada neste trabalho, pode ter
contribuído ainda mais na retenção de bactérias devido ao processo de filtração
mecânica.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

5 CONCLUSÕES E SUGESTÕES
5.1. Conclusões
Este estudo avaliou os parâmetros de transporte do microrganismo
patogênico Escherichia coli ATCC11229 suspenso em água destilada, através
de colunas saturadas de areia de quartzo.
Os ensaios de sobrevivência permitiram demonstrar que a massa de
células injetadas na coluna de areia foi mantida constante o tempo de execução
dos ensaios ADS, ou seja, o efeito de fatores biológicos não interferiu no
transporte deste microrganismo na areia de quartzo.
O ensaio de batelada, através da obtenção do coeficiente de partição KD
igual a 0,001 mL/g, demonstrou que sob condições desfavoráveis à adesão (i.e.
pH próximo ao neutro, baixa condutividade elétrica e coletor e microrganismo
com igual carga elétrica) o processo de filtração físico-química pode ser
considerado desprezível no transporte da E. coli em areia de quartzo.
Sem a interferência do processo de filtração físico-química, os valores para
o fator de retardamento R, entre 3 e 9, obtidos diretamente da curva de chegada
não representa a capacidade de adsorção da bactéria na areia durante o
transporte, sugerindo a intervenção do processo de filtração mecânica.
A diminuição da condutividade hidráulica observada para os ensaios
executados com velocidade de poros maior que 14,4 m/d, evidencia o
entupimento dos poros da areia, alterando as propriedades hidráulicas do meio
poroso, sendo possível estabelecer uma relação linear entre a dispersão
hidrodinâmica e a velocidade de poro, só para valores de dispersão
hidrodinâmica compreendidos entre 1,44 x10-2 cm2/min e 5,47 x10- 2 cm2/min.
Não foi observada a remoção de bactérias devido ao processo de filtração
mecânica nos ensaios realizados com velocidades de poro maiores que 6m/d.
Contrariamente foi observada uma concentração no efluente da coluna maior do
que a concentração inicial no final destes ensaios, que sugere que as bactérias
inicialmente retidas foram arrastadas em decorrência da ação de forças
hidrodinâmicas cisalhantes.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

104
O processo dominante neste estudo foi a filtração mecânica, sendo
observado que este processo, além de depender da concentração inicial,
depende também da variação da velocidade, da distribuição do tamanho e da
forma dos grãos.
Uma descrição precisa do transporte de bactérias requer a compreensão
dos efeitos combinados de sorção, filtração física, velocidade intersticial e
dispersão.
5.2. Sugestões
O estudo e o comportamento de microrganismos em solos é ainda um
desafio. A seguir são apresentadas algumas sugestões e recomendações para
pesquisas futuras, com o objetivo de complementar e comprovar os dados
apresentados.
Determinar a distribuição real do tamanho de poros através da obtenção
da curva de retenção solo-água em laboratório e estudar o comportamento da E.
coli ATCC11229 utilizando areia de quartzo com distribuição granulométrica
uniforme, para confirmar o efeito do coeficiente de uniformidade no processo de
filtração mecânica.
Avaliar o sistema experimental proposto utilizando como solução água com
elevada força iônica que permita avaliar a influência do processo de adsorção
físico-química e, consequentemente determinar a taxa de desorção.
Realizar ensaios de transporte de microrganismos aplicando velocidades
de fluxo menores a 6m/d, representado melhor as velocidades dos aquíferos
naturais.
Definir a velocidade crítica que causa o desprendimento das bactérias que
são retidas pelo processo de filtração mecânica.
Pesquisas recentes sugerem que o monitoramento da concentração da
fase fluida é insuficiente para a identificação dos mecanismos fundamentais
controlando o transporte dos microrganismos, devendo ser complementados
pela distribuição espacial dos microrganismos retidos (Tufenkji, 2007).
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

REFERÊNCIAS BIBLIOGRÁFICAS
ABU-ASHOUR J.; JOY D.M.; LEE H.; WHITELEY H.R.; ZELIN S. Transport of Micororganisms through soil. Water, Air and Soil Pollution, Vol.75, 1994, pp. 141-158.
AZEVEDO, I.D.; AZEVEDO, R.F.; CARVALHO, C.A. A Numerical Study of Heavy Metal Transport in the Dump Area of the Municipal District of Viçosa. MG, Brasil: Environmental Geotechnics (4th ICEG). 2002, pp. 27-32.
BARTON J.W.; FORD R.M. Determination of effective Transport Coefficients for Bacterial migration in Sand Columns. American Society for Microbiology, Applied and Environmental Microbiology, Vol. 61, No. 9, 1995, pp. 3329-3335.
BEAR J. Dynamics of Fluids in Porous Media. Ed. American Elsevier, New York, 1972, 784 pp.
BERG H.C. E. coli in Motion. Biological and medical physics/ Biomedical Engineering. Springer-verlag, New York, Inc., 2004, 129 pp.
BRADFORD S.A.; BETTAHAR M. Concentration Dependent Transport of Colloids in Saturated Porous Media. Elsevier, Journal of Contaminant Hydrology 82, 2006, pp. 99-117.
BRADFORD S.A.; BETTAHAR M.; SIMUNEK J.; VAN GENUCHTEN M.T. Straining and Attachment of Colloids in Physically Heterogeneous Porous Media. Soil Science Society of America, Vadose Zone Journal, vol. 3, 2004, pp. 384-394.
BRADFORD S.A.; SIMUNEK J.; BETTAHAR M.; VAN GENUCHTEN M.T.; YATES S.R. Modeling Colloid Attachment, Straining, and Exclusion in Saturated Porous Media. American Chemical Society, Environmental Science & Technology, vol. 37 No. 10, 2003, pp. 2242-2250.
BRADFORD S.A.; YATES S.R.; BETTAHAR M.; SIMUNEK J. Physical Factors Affecting the Transport and Fate of Colloids in Saturated Porous Media. Water Resources Research, vol. 38, No. 12, 1327, 2002, pp. 63(1)-63(12).
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

CAJAZEIRAS C.C.A. Qualidade e Uso das Águas Subterrâneas e a Relação com Doenças de Veiculação Hídrica, Região de Crajubar/CE. Universidade Federal do Ceará, Departamento de Geologia, Fortaleza, Brasil, Tese de Doutorado, 2007, 131 pp.
CAMESANO T.A.; UNICE K.M.; LOGAN B.E. Blocking and Ripening of Colloids in Porous Media and their Implications for Bacterial Transport. Elsevier Science B.V. Colloids and Surfaces A: Physicochemical and Engineering Aspects 160, 1999, pp. 291-308.
CORAPCIOGLU M.Y.; HARIDAS A. Transport and Fate of Microorganisms in Porous Media: A Theoretical Investigation. Elsevier Science Publisher, Journal of Hydrology, 72, 1984, pp. 149-169
CORAPCIOGLU M.Y.; HARIDAS A. Microbial Transport in Soils and Groundwater: A Numerical Model. Advances Water Resources, vol. 8, 1986, pp. 188-200.
COELHO V.M.T. Potencial de Contaminação de Aquífero Freático por Esgoto Domestico – Quantificação do Decaimento Bacteriológico. Universidade de São Paulo, Instituto de Geociências, SP, Brasil: 2007, 129 pp.
COELHO V.M.T.; Duarte U. Perímetros de Proteção para Fontes Naturais de Aguas Minerais. Rev. Águas Subterrâneas, No. 17, 2003, pp. 77-91.
DESHPANDE P.A.; SHONNARD D.R. An Improved Spectrophotometric Method to Study Transport, Attachment, and Breakthrough of Bacteria through Porous Media. American Society for Microbiology, Applied Environmental Microbiology, 2000, pp. 763-768.
DESHPANDE P.A.; SHONNARD D.R. Modeling the Effects of Systematic Variation in Ionic Strength on the Attachment Kinetics of Pseudomonas Fluorescens UPER-1 in Saturated Sand Columns. American Geophysical Union, Water Resources Research, Vol.35, No.5, 1999, pp. 1619-1627.
DOMENICO P.A.; SCHWARTZ F.W. Physical and Chemical Hydrogeology. John Wiley & Sons, Inc., Canada: 1990, 824 pp.
FITTS C.R. Groundwater Science. Academic Press, Elsevier Science Ltd., California: 2002, 467 pp.
FONTES D.E.; MILLS A.L.; HORNBERGER G.M.; HERMAN J.S. Physical and Chemical Factors Influencing Transport of Microorganisms through Porous Media. American Society for Microbiology, Applied and Environmental Microbiology, Vol.57, No.9, 1991, pp. 2473-2481.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

FOPPEN J.W.A.; MPOROKOSO A.; SCHIJVEN J.F. Determining Straining of Escherichia Coli from Breakthrough Curves. Elsevier, Journal of Contaminant Hydrology 76, 2005, pp. 191-210.
FOPPEN J.W.A.; SCHIJVEN J.F. Transport of E. Coli in Columns of Geochemically Heterogeneous Sediment. Elsevier, Water Research 39, 2005, pp. 3082-3088.
FOPPEN J.W.A.; SCHIJVEN J.F. Evaluation of Data from the Literature on the Transport and Survival of Escherichia Coli and Thermotolerant Coliforms in Aquifers under Saturated Conditions. Elsevier, Water Research 40, 2006, pp. 401-426.
FOPPEN J.W.A.; VAN HERWERDEN M.; SCHIJVEN J.F. Measuring and Modeling Straining of Escherichia Coli in Saturated Porous Media. Elsevier, Journal of Contaminant Hydrology 93, 2007 pp. 236-254.
FOPPEN J.W.A.; VAN HERWERDEN M.; SCHIJVEN J.F. Transport of Escherichia Coli in Saturated Porous Media: Dual Mode Deposition and Intra-Population Heterogeneity. Elsevier, Water Research 41, 2007, pp. 1473-1753.
FOPPEN J.W.A.; VAN HERWERDEN M.; KEBTIE M.; NOMAN A.; SCHIJVEN J.F.; STUYFZAND P.J.; UHLENBROOK S. Transport of Escherichia Coli and Solutes during Waste Water Infiltration in an Urban Alluvial Aquifer. Elsevier, Journal of Contaminant Hydrology 95, 2008 pp. 1–16.
FOSTER S.; HIRATA R.; GOMES D.; D'ELIA.; PARIS M. Proteção da Qualidade da Água Subterrânea: um guia para empresas de abastecimento de água, órgãos municipais e agências ambientais. Banco internacional de Reconstrução e Desenvolvimentot/Banco Mundial. Edição Brasileira: Servmar – Serviços Técnicos Ambientais Ltda, São Paulo, SP, Brasil: 2006, 104 pp.
FREEZE R.A; CHERRY J.A. Groundwater. Prentice-Hall Inc., Englewood Cliffs, New Jersey, USA: 1979, 604 pp.
GINN T.R.; WOOD B.D.; NELSON K.D.; SCHEIBE T.D.; MURPHY E.M.; CLEMENT T.P. Processes in Microbial Transport in the Natural Subsurface. Advances in Water Resources 25, 2002, pp. 1017-1042.
GWYN D. Influences of Flocculation on Bed Properties for Fine-Grained Cohesive Sediment. Oxford University, Department of Engineering Science, PhD Thesis, 2003, 217 pp.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

HORNBERGER G.M.; MILLS A.L.; HERMAN J.S. Bacterial Transport in Porous Media: Evaluation of a Model Using Laboratory Observations. Water Resources Research, Vol.28, No.3, 1992, pp. 915-938.
IBGE Instituto Brasileiro de Geografia e Estatística. Indicadores de Desenvolvimento sustentável, Dimensão Ambiental – Saneamento. 2004.
Disponível em: http://www.ibge.gov.br/home/geociencias/recursosnaturais/ids/defaulttab.shtm Acesso em: 23 de julho 2010.
JIANG G.; NOONAN M.J.; BUCHAN G.D.; SMITH N. Transport of Escherichia Coli through Variably Saturated Sand Columns and Modeling Approaches. Elsevier, Journal of Contaminant Hydrology 93, 2007, pp. 2-20.
KIM H.N.; WALKER S.L. Escherichia Coli Transport in Porous Media: Influence of Cell Strain, Solution Chemistry, and Temperature. Elsevier, Colloids and Surfaces B: Biointerfaces 71, 2009, pp. 160–167.
KNAPPETT P.S.K.; EMELKO M.B.; ZHUANG J.; McKAY L.D. Transport and Retention of a Bacteriophage and Microspheres in Saturated, Angular Porous Media: Effects of Ionic Strength and Grain Size. Elsevier, Water Research 42, 2008, pp. 4368-4378.
KNAPPETT P.S.K. Evaluating the Effects of Grain Size and Divalent Cation Concentration on the Attenuation of Viruses and Microspheres through Crushed Silica Sand. University of Waterloo, Department of Civil Engineering, Master Thesis, Ontario, Canada: 2006, 192 p.
KRETZSCHMAR R.; BORKOVEC M.; GROLIMUND D.; ELIMELECH M. Mobile Subsurface Colloids and Their Role in Contaminant Transport. Academic Press., Advances in Agronomy, vol. 66, 1999, pp. 121-193.
LI X.; SCHEIBE T.D.; JOHNSON W.P. Apparent Decreases in Colloid Deposition Rate Coefficients with Distance of Transport under Unfavorable Deposition Conditions: A General Phenomenon. Environmental Science & Technology, vol. 38, No. 21, 2004, pp. 5616-5625.
LING T.Y; ACHBERGER E.C.; DRAPCHO C.M; BENGTSON R.L. Quantifying Adsorption of an Indicator Bacteria in a Soil-Water System. American Society of Agricultural Engineers, vol. 45, No. 3, 2002, pp. 669-674.
MACIAS J.E. “Modelagem Numérica do Transporte de Vírus em aquíferos Fraturados-Porosos”. Pontifícia Universidade católica do Rio de janeiro, Departamento de Engenharia Civil, Tese de Doutorado, RJ, Brasil: 2008, 257 pp.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

MARTIN M.J.; LOGAN B.E.; JOHNSON W.P.; JEWETT D.G.; ARNOLD R.G. Scaling Bacterial Filtration Rates in Different Sized Porous Media. Journal of Environmental Engineering, 1996, pp. 407-415.
MATTHESS G.; PEKDEGER A.; SCHROETER J. Persistence and Transport of Bacteria and Viruses in Groundwater – A Conceptual Evaluation. Elsevier Science Publishers B.V., Amsterdam – Printed in The Netherlands, Journal of Contaminant Hidrogeology 2, 1988, pp. 171-188.
METCALF & Eddy, Inc. Wastewater Engineering: Treatment, Disposal, and Reuse. 3rd Edition, Irwin McGraw-Hill. Boston: 1991, 1334 pp.
McCAULOU D.R.; BALES R.C.; McCARTHY J.F. Use of Short-Pulse Experiments to Study Bacteria Transport through Porous Media. Elsevier Science Publishers B.V., Water Resources Research, Vol.22, No.13, 1986, pp. 1901-1921.
McDOWELL-BOYER L.M.; HUNT J.R.; SITAR N. Particle Transport through Porous Media. American Geophysical Union, Journal of Contaminant Hydrology 15, 1994, pp. 1-14.
MYERS D. Surfaces, Interfaces, and Colloids: principles and Applications. Second Edition. Jhon Wiley & Sons, Inc., 1999, 519 pp.
PAHO Pan American Health Organization. Health in the Americas. Scientific Publication No. 569, Vol. 1, Washington D.C., USA: 1998, 359 pp.
PANG L.; CLOSE M.; GOLTZ M.; SINTON L.; DAVIES H.; HALL C.; STANTON G. Estimation of Septic Tank Setback Distances Based on Transport of E. Coli and F-RNA Phages. Elsevier, Environment International 29, 2003, pp. 907–921.
PEKDEGER A.; MATTHESS G. Factors of Bacteria and Virus Transport in Groundwater. Springer-Verlag, New York Inc., Environmental Geology, vol. 5, No. 2, 1983, pp. 49-53.
PESSÔA T.O.A. Avaliação da Influência da Mineralogia, Índice de Vazios e Teor de Umidade em Propriedades Térmicas de Solos. Pontifícia Universidade Católica do Rio de Janeiro, Departamento de Engenharia Civil, Dissertação de mestrado, 2006, 163 p.
REDMAN J.A.; WALKER S.L.; ELIMELECH M. Bacterial Adhesion and Transport in Porous Media: Role of the Secondary Energy Minimum. American Chemical Society, Environmental Science & Technology, Vol.38, No.6, 2004, pp. 1777-1785.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

RICHARDSON S.D. Wastewater Plume Dynamics and Bacteria Transport within the Marshland Upwelling System. Louisiana State University, Department of Civil and Environmental Engineering, Louisiana, US, Thesis, Master of Science, 2002, 219 pp.
RYAN J.N.; ELIMELECH M. Colloid Mobilization and Transport in Groundwater. Elsevier, Colloid and Surfaces A: Physicochemical and Engineering Aspects 107, 1996, pp. 1-56.
SCHAAP, M. G.; LEIJ F. J.; VAN GENUCHTEN M. Th. Neural Network Analysis for Hierarchical Prediction of Soil Hydraulic Properties”. Soil Sci. Soc. Am. J. Vol.4, No. 52, 1998, pp. 847- 855.
SCHIJVEN J.F.; HASSANIZADEH S.M.; DE BRUIN H.A.M. Column Experiments to Study Nonlinear Removal of Bacteriophages by Passage trough Saturated Dune Sand. Elsevier, Journal of Contaminant Hydrology 58, 2002, pp. 243-259.
SCHINNER T.; LETZNER A.; LIEDTKE S.; CASTRO F.D.; EYDELNANT I.A.; TUFENKJI N. Transport of Selected Bacterial Pathogens in Agricultural Soil and Quartz Sand. Elsevier, Water Research, 2009, 11 pp.
SCHOLL M.A.; MILLS A.L.; HERMAN J.S.; HORNBERGER G.M. The Influence of Mineralogy and Solution Chemistry on the Attachment of Bacteria to Representative Aquifer Materials. Elsevier Science Publication, Journal of Contaminant Hydrology 6, 1990, pp. 321-336.
SIRIVITHAYAPAKORN S.; KELLER A. Transport of colloids in saturated porous media: A pore-scale observation of the size exclusion effect and colloid acceleration. American Geophysical Union, Water Resources Research, Vol.39, No.4, 2003, pp.1109.
SSSA Soil Science Society of America Inc. Methods of Soil Analysis, Part 2: Microbial and Biochemical Properties”. 1994, 1121 pp.
STUMM W.; MORGAN J.J. AQUATIC CHEMISTRY: Chemical Equilibria an Rates In Natural Waters. 3rd ed. Wiley-Interscience Series of Texts and Monographs, John Wiley and Sons, Inc., New York: 1996, 1022 pp.
THOREL L.; FAVRAUD C.; GARNIER J. Mariotte Bottle in a Centriguge: A Device for Constant Water Table Level. International Journal of Physical Modelling in Geotechnics 1, 2002, pp.23-26.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

TOSCANO G.; DOS SANTOS K.; ALMEIDA C.; DA SILVA T. Uma Analise Sintética sobre a Legislação das Águas Subterrâneas no Brasil. IX Simpósio de Recursos Hídricos do Nordeste, 2008, 20 pp.
TUFENKJI N.; ELIMELECH M. Deviation from the Classical Colloid Filtration Theory in the Presence of Repulsive DLVO Interactions. American Chemical Society, Langmuir 20, 2004, pp. 10818-10828.
TUFENKJI N.; MILLER G.F.; RYAN J.N.; HARVEY R.W.; ELIMELECH M. Transport of Cryptosporidium Oocysts in Porous Media: Role of Straining and Physicochemical Filtration. American Chemical Society, Environmental Science & Technology, Vol.38, No.22, 2004, pp. 5932-5938.
TUFENKJI N. Modeling Microbial Transport in porous Media: Traditional Approaches and recent Developments. Elsevier, Advances in Water Resources 30, 2007, pp. 1455-1469.
WALKER S.L.; HILL J.E.; REDMAN J.A.; ELIMELECH M. Influence of Growth Phase on Adhesion Kinetics of Escherichia coli D21g. American Society for Microbiology, Applied and Environmental Microbiology, 2005, pp. 3093–3099.
WALL K.; PANG L.; SINTON L.; CLOSE M. Transport and Attenuation of Microbial Tracers and Effluent Microorganisms in Saturated Pumice Sand Aquifer Material. Springer Science, Water Air Soil Pollut 188, 2008, pp. 213-224.
WOESSNER W.W.; BALL P.N.; DeBORDE D.C.; TROY T.L. Viral Transport in a Sand and Gravel Aquifer Under field Pumping Conditions. Groundwater, Vol.39, No.6, 2001, pp. 886-894.
YAO K.M.; HABIBIAN M.T.; O’MELIA CH.R. Water and Waste Water Filtration: Concepts and Applications. Department of Environmental Sciences and Engineering, University of North Carolina, vol. 5, No. 11, 1971, pp. 1105-1112.
YATES M.V.; YATES S.R. Modeling microbial transport in the subsurface: A mathematical discussion. Washington, DC: American Society for Microbiology. 1991, pp. 48-76.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA
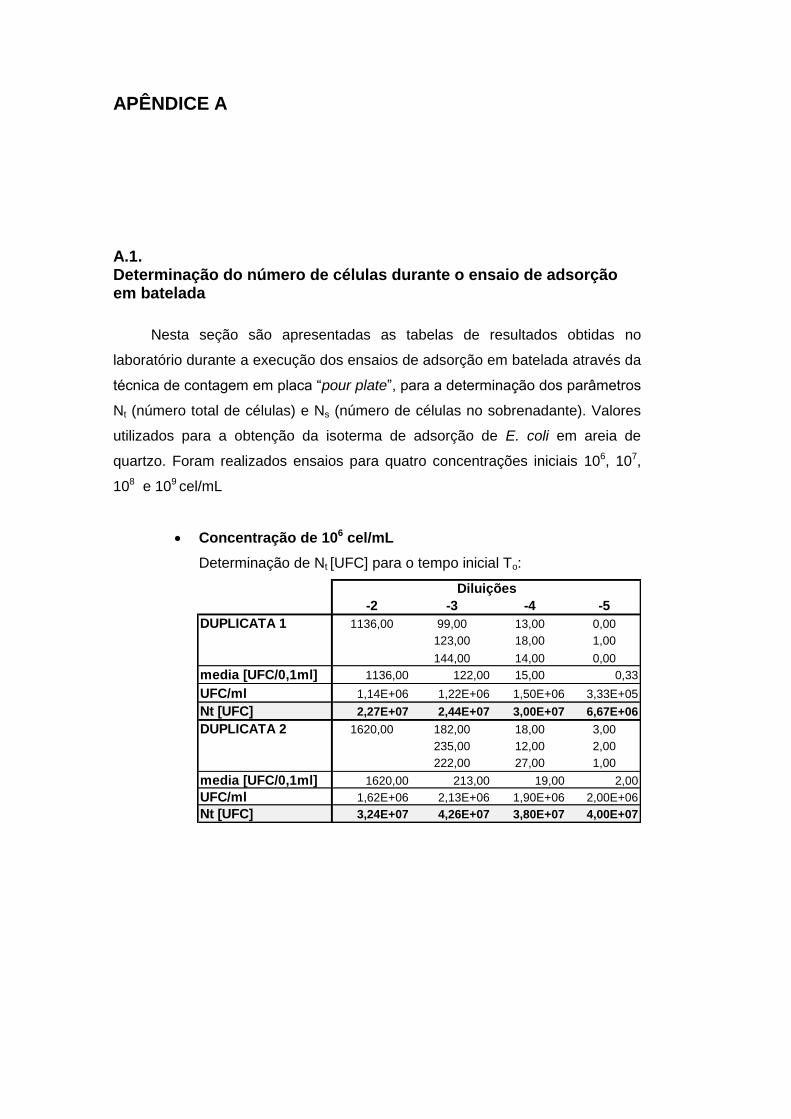
APÊNDICE A
A.1. Determinação do número de células durante o ensaio de adsorção em batelada
Nesta seção são apresentadas as tabelas de resultados obtidas no
laboratório durante a execução dos ensaios de adsorção em batelada através da
técnica de contagem em placa “pour plate”, para a determinação dos parâmetros
Nt (número total de células) e Ns (número de células no sobrenadante). Valores
utilizados para a obtenção da isoterma de adsorção de E. coli em areia de
quartzo. Foram realizados ensaios para quatro concentrações iniciais 106, 107,
108 e 109 cel/mL
Concentração de 106 cel/mL
Determinação de Nt [UFC] para o tempo inicial To:
-2 -3 -4 -5
DUPLICATA 1 1136,00 99,00 13,00 0,00
123,00 18,00 1,00
144,00 14,00 0,00
media [UFC/0,1ml] 1136,00 122,00 15,00 0,33
UFC/ml 1,14E+06 1,22E+06 1,50E+06 3,33E+05
Nt [UFC] 2,27E+07 2,44E+07 3,00E+07 6,67E+06
DUPLICATA 2 1620,00 182,00 18,00 3,00
235,00 12,00 2,00
222,00 27,00 1,00
media [UFC/0,1ml] 1620,00 213,00 19,00 2,00
UFC/ml 1,62E+06 2,13E+06 1,90E+06 2,00E+06
Nt [UFC] 3,24E+07 4,26E+07 3,80E+07 4,00E+07
Diluições
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Determinação de NS [UFC] para o tempo final T1:
-2 -3 -4 -5
DUPLICATA 1 1564,00 182,00 35,00 1,00
176,00 20,00 3,00
195,00 16,00 4,00
media [UFC/0,1ml] 1564,00 184,33 23,67 2,67
UFC/ml 1,56E+06 1,84E+06 2,37E+06 2,67E+06
Ns [UFC] 3,13E+07 3,69E+07 4,73E+07 5,33E+07
DUPLICATA 2 1124,00 195,00 20,00 1,00
190,00 32,00 4,00
186,00 25,00 3,00
media [UFC/0,1ml] 1124,00 190,33 25,67 2,67
UFC/ml 1,12E+06 1,90E+06 2,57E+06 2,67E+06
Ns [UFC] 2,25E+07 3,81E+07 5,13E+07 5,33E+07
Diluições
Concentração de 107 cel/mL
Determinação de Nt [UFC] para o tempo inicial To:
-3 -4 -5 -6
DUPLICATA 1 1528,00 288,00 13,00 4,00
1944,00 249,00 9,00 2,00
276,00 21,00 6,00
media [UFC/0,1ml] 1736,00 271,00 14,33 4,00
UFC/ml 1,74E+07 2,71E+07 1,43E+07 4,00E+07
Nt [UFC] 3,47E+08 5,42E+08 2,87E+08 8,00E+08
DUPLICATA 2 836,00 172,00 6,00 1,00
468,00 204,00 2,00 0,00
800,00 215,00 6,00 1,00
media [UFC/0,1ml] 701,33 197,00 4,67 0,67
UFC/ml 7,01E+06 1,97E+07 4,67E+06 6,67E+06
Nt [UFC] 1,40E+08 3,94E+08 9,33E+07 1,33E+08
Diluições
Determinação de NS [UFC] para o tempo final T1:
-3 -4 -5 -6
DUPLICATA 1 1836,00 16,00 18,00 5,00
1240,00 34,00 29,00 2,00
45,00 40,00 5,00
media [UFC/0,1ml] 1538,00 31,67 29,00 4,00
UFC/ml 1,54E+07 3,17E+06 2,90E+07 4,00E+07
Ns [UFC] 3,08E+08 6,33E+07 5,80E+08 8,00E+08
DUPLICATA 2 456,00 167,00 9,00 3,00
399,00 130,00 12,00 1,00
472,00 252,00 12,00 3,00
media [UFC/0,1ml] 442,33 183,00 11,00 2,33
UFC/ml 4,42E+06 1,83E+07 1,10E+07 2,33E+07
Ns [UFC] 8,85E+07 3,66E+08 2,20E+08 4,67E+08
Diluições
Não foram considerados os valores que correspondem à diluição 10-4
para a
Duplicata 1.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Concentração de 108 cel/mL
Determinação de Nt [UFC] para o tempo inicial To:
-4 -5 -6 -7
DUPLICATA 1 1560,00 332,00 68,00 1,00
1744,00 347,00 57,00 12,00
339,00 42,00 6,00
media [UFC/0,1ml] 1652,00 339,33 55,67 6,33
UFC/ml 1,65E+08 3,39E+08 5,57E+08 6,33E+08
Nt [UFC] 3,30E+09 6,79E+09 1,11E+10 1,27E+10
DUPLICATA 2 2264,00 325,00 32,00 8,00
2116,00 302,00 37,00 3,00
345,00 34,00 6,00
media [UFC/0,1ml] 2190,00 324,00 34,33 5,67
UFC/ml 2,19E+08 3,24E+08 3,43E+08 5,67E+08
Nt [UFC] 4,38E+09 6,48E+09 6,87E+09 1,13E+10
Diluições
Determinação de NS [UFC] para o tempo final T1:
-4 -5 -6 -7
DUPLICATA 1 1716,00 177,00 39,00 8,00
1240,00 199,00 42,00 3,00
203,00 42,00 7,00
media [UFC/0,1ml] 1478,00 193,00 41,00 6,00
UFC/ml 1,48E+08 1,93E+08 4,10E+08 6,00E+08
Ns [UFC] 2,96E+09 3,86E+09 8,20E+09 1,20E+10
DUPLICATA 2 792,00 96,00 37,00 10,00
772,00 116,00 35,00 8,00
147,00 30,00 14,00
media [UFC/0,1ml] 782,00 119,67 34,00 10,67
UFC/ml 7,82E+07 1,20E+08 3,40E+08 1,07E+09
Ns [UFC] 1,56E+09 2,39E+09 6,80E+09 2,13E+10
Diluições
Concentração de 109 cel/mL
Determinação de Nt [UFC] para o tempo inicial To:
-5 -6 -7 -8
DUPLICATA 1 1508,00 317,00 32,00 5,00
267,00 31,00 3,00
339,00 23,00 1,00
media [UFC/0,1ml] 1508,00 307,67 28,67 3,00
UFC/ml 1,51E+09 3,08E+09 2,87E+09 3,00E+09
Nt [UFC] 3,02E+10 6,15E+10 5,73E+10 6,00E+10
DUPLICATA 2 1708,00 464,00 1188,00 748,00
media [UFC/0,1ml] 1708,00 464,00 1188,00 748,00
UFC/ml 1,71E+09 4,64E+09 1,19E+11 7,48E+11
Nt [UFC] 3,42E+10 9,28E+10 2,38E+12 1,50E+13
Diluições
Não foram considerados os valores que correspondem às diluições 10-7
e 10-8
para
a Duplicata 2.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Determinação de NS [UFC] para o tempo final T1:
-5 -6 -7 -8
DUPLICATA 1 1868,00 452,00 189,00 65,00
394,00 207,00 50,00
416,00 216,00 57,00
media [UFC/0,1ml] 1868,00 420,67 204,00 57,33
UFC/ml 1,87E+09 4,21E+09 2,04E+10 5,73E+10
Ns [UFC] 3,74E+10 8,41E+10 4,08E+11 1,15E+12
DUPLICATA 2 724,00 59,00 21,00 1,00
89,00 26,00 2,00
55,00 21,00 3,00
media [UFC/0,1ml] 724,00 67,67 22,67 2,00
UFC/ml 7,24E+08 6,77E+08 2,27E+09 2,00E+09
Ns [UFC] 1,45E+10 1,35E+10 4,53E+10 4,00E+10
Diluições
Não foram considerados os valores que correspondem às diluições 10-7
e 10-8
para
a Duplicata 1.
A.2. Cálculo do coeficiente de distribuição KD
Concentração de 106 cel/mL
Dil Co Nt Ns Pa Cs Cw KD
UFC/ml UFC UFC % cel/g cel/ml ml/g
DUPLICATA 1 -2 1,14E+06 2,27E+07 3,13E+07 -37,7 -4,27E+06 1,56E+06 -2,7
-3 1,22E+06 2,44E+07 3,69E+07 -51,1 -6,21E+06 1,84E+06 -3,4
-4 1,50E+06 3,00E+07 4,73E+07 -57,8 -8,64E+06 2,37E+06 -3,6
-5 3,33E+05 6,67E+06 5,33E+07 -700,0 -2,33E+07 2,67E+06 -8,7
DUPLICATA 2 -2 1,62E+06 3,24E+07 2,25E+07 30,6 4,96E+06 1,12E+06 4,4
-3 2,13E+06 4,26E+07 3,81E+07 10,6 2,27E+06 1,90E+06 1,2
-4 1,90E+06 3,80E+07 5,13E+07 -35,1 -6,67E+06 2,57E+06 -2,6
-5 2,00E+06 4,00E+07 5,33E+07 -33,3 -6,67E+06 2,67E+06 -2,5
O valor de KD determinado para a concentração 106 cel/mL corresponde ao valor da tabela
calculado para a diluição 10-3
da Duplicata 2 igual a 1,2 mL/g.
Concentração de 107 cel/mL
Dil Co Nt Ns Pa Cs Cw KD
UFC/ml UFC UFC % cel/g cel/ml ml/g
DUPLICATA 1 -3 1,74E+07 3,47E+08 3,08E+08 11,4 1,97E+07 1,54E+07 1,3
-4 2,71E+07 5,42E+08 6,33E+07 88,3 2,38E+08 3,17E+06 75,3
-5 1,43E+07 2,87E+08 5,80E+08 -102,3 -1,46E+08 2,90E+07 -5,0
-6 4,00E+07 8,00E+08 8,00E+08 0,0 0,00E+00 4,00E+07 0,0
DUPLICATA 2 -3 7,01E+06 1,40E+08 8,85E+07 36,9 2,59E+07 4,42E+06 5,9
-4 1,97E+07 3,94E+08 3,66E+08 7,1 1,40E+07 1,83E+07 0,8
-5 4,67E+06 9,33E+07 2,20E+08 -135,7 -6,33E+07 1,10E+07 -5,8
-6 6,67E+06 1,33E+08 4,67E+08 -250,0 -1,67E+08 2,33E+07 -7,1
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Dil Co Nt Ns Pa Cs Cw KD
UFC/ml UFC UFC % cel/g cel/ml ml/g
DUPLICATA 1 -4 2,71E+07 5,42E+08
-3 3,08E+08
media 5,42E+08 3,08E+08 43,2 1,17E+08 1,54E+07 7,6
DUPLICATA 2 -4 1,97E+07 3,94E+08 3,66E+08
media 3,94E+08 3,66E+08 7,1 1,40E+07 1,83E+07 0,8
2,34E+07 4,68E+08 25,2 6,56E+07 1,68E+07 4,2
O valor de KD determinado para a concentração 107 cel/mL corresponde ao valor médio,
dos valores calculados para as diluições 10-4
e 10-3
da Duplicata 1 e a diluição 10-4
da
Duplicata 2. Assim o valor de KD resulta ser igual a 4,2 mL/g.
Concentração de 108 cel/mL
O valor de KD determinado para a concentração 108 cel/mL corresponde ao valor médio,
dos valores calculados para a Duplicata 1 e a Duplicata 2, utilizando a média das diluições
10-5
e 10-6
. Assim o valor de KD resulta ser igual a 4,7 mL/g.
Concentração de 109 cel/mL
Dil Co Nt Ns Pa Cs Cw KD
UFC/ml UFC UFC % cel/g cel/ml ml/g
DUPLICATA 1 -5 1,51E+09 3,02E+10 3,74E+10 -23,9 -3,61E+09 1,87E+09 -1,9
-6 3,08E+09 6,15E+10 8,41E+10 -36,7 -1,13E+10 4,21E+09 -2,7
-7 2,87E+09 5,73E+10
-8 3,00E+09 6,00E+10
DUPLICATA 2 -5 1,71E+09 3,42E+10 1,45E+10 57,6 9,84E+09 7,24E+08 13,6
-6 4,64E+09 9,28E+10 1,35E+10 85,4 3,96E+10 6,77E+08 58,6
-7 4,53E+10
-8 4,00E+10
O valor de KD determinado para a concentração 109 cel/mL corresponde ao valor calculado
considerando a diluição 10-6
(para Nt) da Duplicata 1 e 10-6
da Duplicata 2 (para Ns). Assim
o valor de KD resulta ser igual a 35,5 mL/g.
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

APÊNDICE B
B.1. Dados para a construção da curva de chegada
Ensaio 1 (i=0,03)
N°Absorbância
[A] Nº cel/ml C [cel/ml] C/Covol percolado
medido pV
C [cel/ml]
CNB C/Co
0 0,006 -2,71E+06 0,00E+00 0,00 20,0 0,1 0,00E+00 0,00
1 0,020 6,84E+05 6,84E+05 0,01 170,0 0,9 0,00E+00 0,00
2 0,062 1,09E+07 1,09E+07 0,10 275,0 1,5 1,70E+07 0,15
3 0,276 6,27E+07 6,27E+07 0,59 380,0 2,1 8,48E+07 0,74
4 0,238 5,35E+07 5,35E+07 0,50 470,0 2,6
5 0,242 5,45E+07 5,45E+07 0,51 565,0 3,1 5,83E+07 0,51
6 0,266 6,03E+07 6,03E+07 0,57 660,0 3,7
7 0,300 6,85E+07 6,85E+07 0,64 750,0 4,2 8,88E+07 0,77
8 0,336 7,73E+07 7,73E+07 0,72 840,0 4,79 0,359 8,28E+07 8,28E+07 0,78 935,0 5,2
10 0,378 8,74E+07 8,74E+07 0,82 1025,0 5,7 1,13E+08 0,98
11 0,387 8,96E+07 8,96E+07 0,84 1115,0 6,2
12 0,399 9,25E+07 9,25E+07 0,87 1210,0 6,7 7,90E+07 0,69
13 0,412 9,57E+07 9,57E+07 0,90 1305,0 7,3
14 0,422 9,81E+07 9,81E+07 0,92 1390,0 7,7 1,22E+08 1,06
15 0,587 1,38E+08 1,38E+08 1,30 1930,0 10,7 1,42E+08 1,23
16 0,607 1,43E+08 1,43E+08 1,34 1960,0 10,9
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Ensaio 2 (i=0,06)
N°Absorbância
[A] Nº cel/ml C [cel/ml] C/Covol percolado
medido pV
C [cel/ml]
CNB C/Co
0 0,005 -2,95E+06 0,00E+00 0,00 16,0 0,1 0,00E+00 0,00
1 0,010 -1,74E+06 0,00E+00 0,00 176,0 0,9
2 0,074 1,38E+07 1,38E+07 0,08 311,0 1,7 1,36E+07 0,08
3 0,142 3,02E+07 3,02E+07 0,18 437,0 2,3
4 0,213 4,75E+07 4,75E+07 0,29 567,0 3,0
5 0,307 7,02E+07 7,02E+07 0,43 702,0 3,8 6,57E+07 0,40
6 0,423 9,83E+07 9,83E+07 0,60 852,0 4,67 0,497 1,16E+08 1,16E+08 0,70 957,0 5,1
8 0,598 1,41E+08 1,41E+08 0,85 1062,0 5,7
9 0,621 1,46E+08 1,46E+08 0,89 1212,0 6,5 1,40E+08 0,85
10 0,626 1,48E+08 1,48E+08 0,89 1362,0 7,3
11 0,635 1,50E+08 1,50E+08 0,91 1457,0 7,8
12 0,651 1,54E+08 1,54E+08 0,93 1593,0 8,5
13 0,651 1,54E+08 1,54E+08 0,93 1593,0 8,5
Ensaio 3 (i=0,06)
N°Absorbância
[A] Nº cel/ml C [cel/ml] C/Covol percolado
medido pV
C [cel/ml]
CNB C/Co
0 0,015 -5,27E+05 0,00E+00 0,00 15,0 0,1 0,00E+00 0,00
1 0,022 1,17E+06 1,17E+06 0,01 130,0 0,7
2 0,031 3,35E+06 3,35E+06 0,02 231,0 1,2 2,75E+06 0,02
3 0,090 1,76E+07 1,76E+07 0,11 355,0 1,9
4 0,142 3,02E+07 3,02E+07 0,20 490,0 2,6 3,05E+07 0,25
5 0,214 4,77E+07 4,77E+07 0,31 615,0 3,3
6 0,319 7,31E+07 7,31E+07 0,47 740,0 3,9 6,15E+07 0,51
7 0,398 9,23E+07 9,23E+07 0,60 850,0 4,5
8 0,468 1,09E+08 1,09E+08 0,71 970,0 5,2 7,68E+07 0,63
9 0,494 1,16E+08 1,16E+08 0,75 1070,0 5,7
10 0,543 1,27E+08 1,27E+08 0,82 1170,0 6,2 1,01E+08 0,83
11 0,524 1,23E+08 1,23E+08 0,79 1305,0 7,0
12 0,531 1,25E+08 1,25E+08 0,80 1415,0 7,5 9,95E+07 0,82
13 0,582 1,37E+08 1,37E+08 0,88 1525,0 8,1
14 0,583 1,37E+08 1,37E+08 0,89 1650,0 8,8 9,90E+07 0,81
15 0,611 1,44E+08 1,44E+08 0,93 1775,0 9,5
16 0,635 1,50E+08 1,50E+08 0,97 1915,0 10,2 1,12E+08 0,92
17 0,668 1,58E+08 1,58E+08 1,02 2030,0 10,8
18 0,680 1,61E+08 1,61E+08 1,04 2135,0 11,4
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Ensaio 4 (i=0,06)
N°Absorbância
[A] Nº cel/ml C [cel/ml] C/Covol percolado
medido pV
C [cel/ml]
CNB C/Co
0 0,008 -2,22E+06 0,00E+00 0,00 15,0 0,1 0,00E+00 0,00
1 0,011 -1,50E+06 0,00E+00 0,00 140,0 0,8
2 0,062 1,09E+07 1,09E+07 0,06 260,0 1,4 8,45E+06 0,04
3 0,129 2,71E+07 2,71E+07 0,15 390,0 2,1
4 0,166 3,61E+07 3,61E+07 0,20 515,0 2,8 2,90E+07 0,13
5 0,224 5,01E+07 5,01E+07 0,28 631,0 3,4
6 0,307 7,02E+07 7,02E+07 0,39 741,0 4,0 8,30E+07 0,38
7 0,400 9,28E+07 9,28E+07 0,52 859,0 4,6
8 0,475 1,11E+08 1,11E+08 0,62 969,0 5,2 1,24E+08 0,57
9 0,528 1,24E+08 1,24E+08 0,69 1074,0 5,8
10 0,575 1,35E+08 1,35E+08 0,76 1184,0 6,4 1,63E+08 0,75
11 0,614 1,45E+08 1,45E+08 0,81 1299,0 7,0
12 0,645 1,52E+08 1,52E+08 0,85 1414,0 7,6 1,85E+08 0,85
13 0,668 1,58E+08 1,58E+08 0,88 1539,0 8,3
14 0,681 1,61E+08 1,61E+08 0,90 1659,0 8,9 1,93E+08 0,89
15 0,686 1,62E+08 1,62E+08 0,91 1784,0 9,6
16 0,690 1,63E+08 1,63E+08 0,91 1910,0 10,3 2,00E+08 0,92
17 0,692 1,64E+08 1,64E+08 0,92 2030,0 10,9
18 0,693 1,64E+08 1,64E+08 0,92 2135,0 11,5
Ensaio 5 (i=0,16)
N°Absorbância
[A] Nº cel/ml C [cel/ml] C/Covol percolado
medido pV
C [cel/ml]
CNB C/Co
0 0,007 -2,47E+06 0,00E+00 0,00 15,0 0,1 0,00E+00 0,00
1 0,025 1,90E+06 1,90E+06 0,01 210,0 1,1 2,50E+05 0,00
2 0,119 2,47E+07 2,47E+07 0,15 395,0 2,1 1,93E+07 0,14
3 0,193 4,26E+07 4,26E+07 0,26 570,0 3,1 2,96E+07 0,21
4 0,281 6,39E+07 6,39E+07 0,39 737,0 4,0 4,34E+07 0,31
5 0,357 8,24E+07 8,24E+07 0,51 892,0 4,8 0,00
6 0,425 9,88E+07 9,88E+07 0,61 1047,0 5,7 6,85E+07 0,50
7 0,475 1,11E+08 1,11E+08 0,68 1202,0 6,5 0,00
8 0,513 1,20E+08 1,20E+08 0,74 1357,0 7,3 9,58E+07 0,69
9 0,544 1,28E+08 1,28E+08 0,78 1507,0 8,1 0,00
10 0,572 1,34E+08 1,34E+08 0,83 1662,0 9,0 1,02E+08 0,74
11 0,608 1,43E+08 1,43E+08 0,88 1822,0 9,8 1,07E+08 0,77
12 0,638 1,50E+08 1,50E+08 0,92 1988,0 10,7 1,40E+08 1,01
13 0,673 1,59E+08 1,59E+08 0,98 2158,0 11,7 1,17E+08 0,85
14 0,707 1,67E+08 1,67E+08 1,03 2328,0 12,6 1,60E+08 1,16
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

Ensaio 6 (i=0,24)
N°Absorbância
[A] Nº cel/ml C [cel/ml] C/Covol percolado
medido pV
C [cel/ml]
CNB C/Co
0 0,007 -2,47E+06 0,00E+00 0,00 15,0 0,1 0,00E+00 0,00
1 0,015 -5,27E+05 0,00E+00 0,00 190,0 1,0 2,50E+05 0,00
2 0,089 1,74E+07 1,74E+07 0,12 365,0 2,0 1,93E+07 0,14
3 0,133 2,81E+07 2,81E+07 0,20 536,0 2,9 2,96E+07 0,21
4 0,181 3,97E+07 3,97E+07 0,28 702,0 3,8 4,34E+07 0,31
5 0,231 5,18E+07 5,18E+07 0,37 847,0 4,6 0,00
6 0,273 6,20E+07 6,20E+07 0,44 992,0 5,4 6,85E+07 0,50
7 0,317 7,27E+07 7,27E+07 0,52 1138,0 6,1 0,00
8 0,358 8,26E+07 8,26E+07 0,59 1288,0 7,0 9,58E+07 0,69
9 0,392 9,08E+07 9,08E+07 0,65 1433,0 7,7 0,00
10 0,432 1,01E+08 1,01E+08 0,72 1568,0 8,5 1,02E+08 0,74
11 0,469 1,09E+08 1,09E+08 0,78 1698,0 9,2 1,07E+08 0,77
12 0,506 1,18E+08 1,18E+08 0,85 1833,0 9,9 1,40E+08 1,01
13 0,543 1,27E+08 1,27E+08 0,91 1973,0 10,7 1,17E+08 0,85
14 0,567 1,33E+08 1,33E+08 0,95 2113,0 11,4 1,60E+08 1,16
15 0,589 1,39E+08 1,39E+08 0,99 2248,0 12,1 1,30E+08 0,94
16 0,605 1,42E+08 1,42E+08 1,02 2378,0 12,8 1,51E+08 1,09
17 0,626 1,48E+08 1,48E+08 1,05 2503,0 13,5
18 0,659 1,56E+08 1,56E+08 1,11 2608,0 14,1
19 0,686 1,62E+08 1,62E+08 1,16 2704,2 14,6
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA
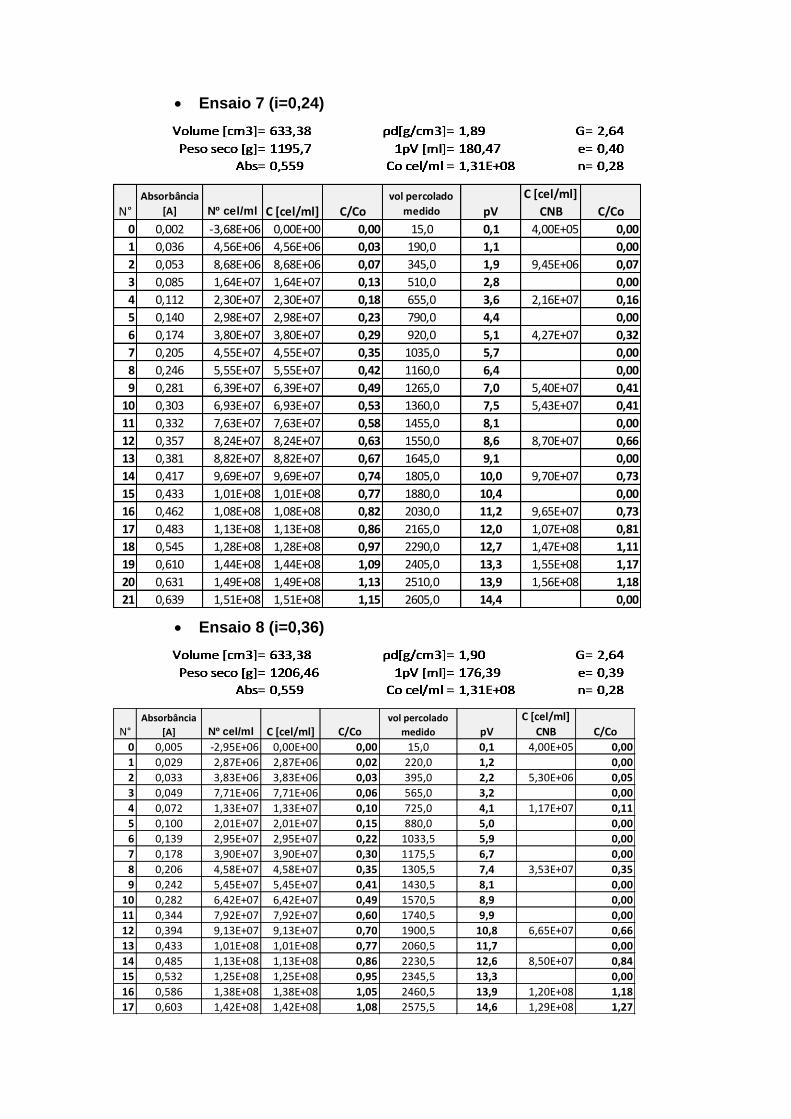
Ensaio 7 (i=0,24)
N°Absorbância
[A] Nº cel/ml C [cel/ml] C/Covol percolado
medido pV
C [cel/ml]
CNB C/Co
0 0,002 -3,68E+06 0,00E+00 0,00 15,0 0,1 4,00E+05 0,00
1 0,036 4,56E+06 4,56E+06 0,03 190,0 1,1 0,00
2 0,053 8,68E+06 8,68E+06 0,07 345,0 1,9 9,45E+06 0,07
3 0,085 1,64E+07 1,64E+07 0,13 510,0 2,8 0,00
4 0,112 2,30E+07 2,30E+07 0,18 655,0 3,6 2,16E+07 0,16
5 0,140 2,98E+07 2,98E+07 0,23 790,0 4,4 0,00
6 0,174 3,80E+07 3,80E+07 0,29 920,0 5,1 4,27E+07 0,32
7 0,205 4,55E+07 4,55E+07 0,35 1035,0 5,7 0,00
8 0,246 5,55E+07 5,55E+07 0,42 1160,0 6,4 0,00
9 0,281 6,39E+07 6,39E+07 0,49 1265,0 7,0 5,40E+07 0,41
10 0,303 6,93E+07 6,93E+07 0,53 1360,0 7,5 5,43E+07 0,41
11 0,332 7,63E+07 7,63E+07 0,58 1455,0 8,1 0,00
12 0,357 8,24E+07 8,24E+07 0,63 1550,0 8,6 8,70E+07 0,66
13 0,381 8,82E+07 8,82E+07 0,67 1645,0 9,1 0,00
14 0,417 9,69E+07 9,69E+07 0,74 1805,0 10,0 9,70E+07 0,73
15 0,433 1,01E+08 1,01E+08 0,77 1880,0 10,4 0,00
16 0,462 1,08E+08 1,08E+08 0,82 2030,0 11,2 9,65E+07 0,73
17 0,483 1,13E+08 1,13E+08 0,86 2165,0 12,0 1,07E+08 0,81
18 0,545 1,28E+08 1,28E+08 0,97 2290,0 12,7 1,47E+08 1,11
19 0,610 1,44E+08 1,44E+08 1,09 2405,0 13,3 1,55E+08 1,17
20 0,631 1,49E+08 1,49E+08 1,13 2510,0 13,9 1,56E+08 1,18
21 0,639 1,51E+08 1,51E+08 1,15 2605,0 14,4 0,00
Ensaio 8 (i=0,36)
N°Absorbância
[A] Nº cel/ml C [cel/ml] C/Covol percolado
medido pV
C [cel/ml]
CNB C/Co
0 0,005 -2,95E+06 0,00E+00 0,00 15,0 0,1 4,00E+05 0,00
1 0,029 2,87E+06 2,87E+06 0,02 220,0 1,2 0,00
2 0,033 3,83E+06 3,83E+06 0,03 395,0 2,2 5,30E+06 0,05
3 0,049 7,71E+06 7,71E+06 0,06 565,0 3,2 0,00
4 0,072 1,33E+07 1,33E+07 0,10 725,0 4,1 1,17E+07 0,11
5 0,100 2,01E+07 2,01E+07 0,15 880,0 5,0 0,00
6 0,139 2,95E+07 2,95E+07 0,22 1033,5 5,9 0,00
7 0,178 3,90E+07 3,90E+07 0,30 1175,5 6,7 0,00
8 0,206 4,58E+07 4,58E+07 0,35 1305,5 7,4 3,53E+07 0,35
9 0,242 5,45E+07 5,45E+07 0,41 1430,5 8,1 0,00
10 0,282 6,42E+07 6,42E+07 0,49 1570,5 8,9 0,00
11 0,344 7,92E+07 7,92E+07 0,60 1740,5 9,9 0,00
12 0,394 9,13E+07 9,13E+07 0,70 1900,5 10,8 6,65E+07 0,66
13 0,433 1,01E+08 1,01E+08 0,77 2060,5 11,7 0,00
14 0,485 1,13E+08 1,13E+08 0,86 2230,5 12,6 8,50E+07 0,84
15 0,532 1,25E+08 1,25E+08 0,95 2345,5 13,3 0,00
16 0,586 1,38E+08 1,38E+08 1,05 2460,5 13,9 1,20E+08 1,18
17 0,603 1,42E+08 1,42E+08 1,08 2575,5 14,6 1,29E+08 1,27
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

B.2. Determinação gráfica dos parâmetros “b” e “R”
Nesta seção apresentam-se as gráficas das curvas de chegada obtidas
durante os ensaios ADS, indicando a forma como foi determinado o fator de
retardamento R, assim como também o valor de b para posterior calculo da
dispersão hidrodinâmica Dh.
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
1,40
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
C/C
o
volume de poro
Ensaio 1 (i=0,03)
R=3,3 b=1,2 Dh=1,74E-2 [cm2/min]
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
1,40
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
C/C
o
volume de poro
Ensaio 2 (i=0,06)
R=4,1 b=1,5 Dh=2,04E-2 [cm2/min]
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
1,40
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
C/C
o
volume de poro
Ensaio 3 (i=0,06)
R=4,0 b=1,6 Dh=1,63E-2 [cm2/min]
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
1,40
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
C/C
o
volume de poro
Ensaio 4 (i=0,06)
R=4,5 b=1,5 Dh=1,44E-2 [cm2/min]
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
1,40
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
C/C
o
volume de poro
Ensaio 5 (i=0,16)
R=4,9 b=1,1 Dh=4,87E-2 [cm2/min]
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA
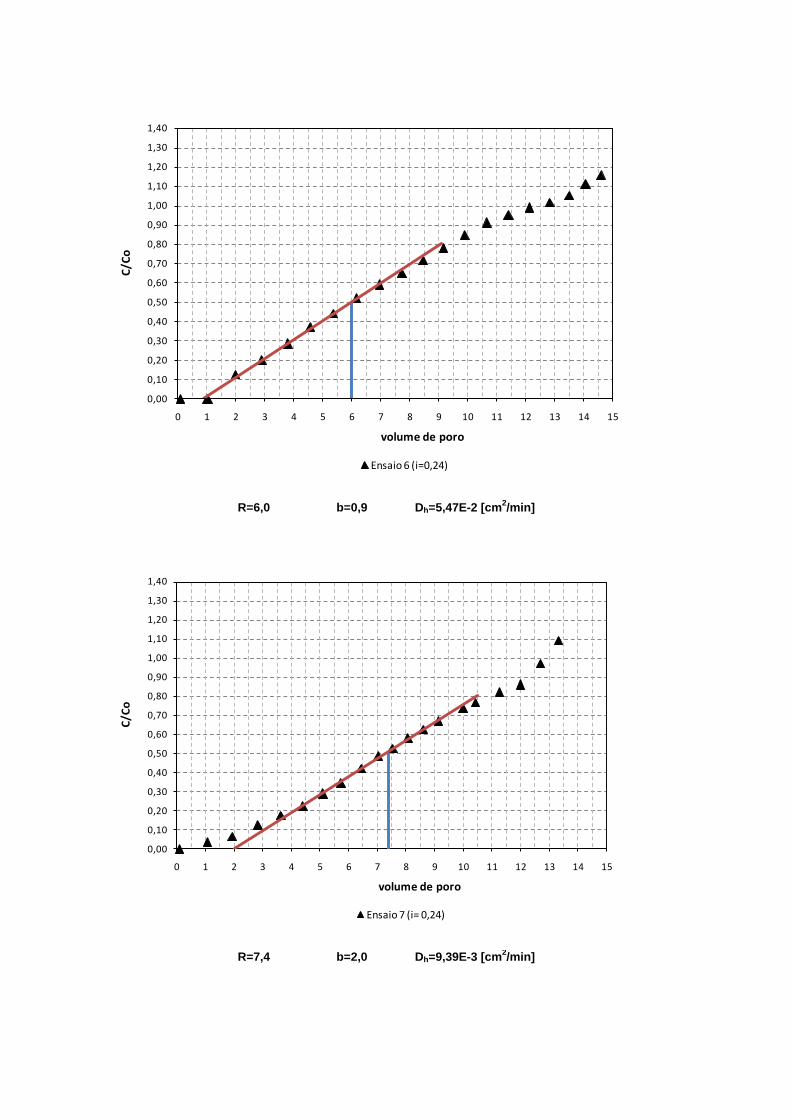
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
1,40
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
C/C
o
volume de poro
Ensaio 6 (i=0,24)
R=6,0 b=0,9 Dh=5,47E-2 [cm2/min]
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
1,40
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
C/C
o
volume de poro
Ensaio 7 (i= 0,24)
R=7,4 b=2,0 Dh=9,39E-3 [cm2/min]
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA

0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
1,40
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
C/C
o
volume de poro
Ensaio 8 (i=0,36)
R=9,0 b=3,5 Dh=7,89E-3 [cm2/min]
DBD
PUC-Rio - Certificação Digital Nº 0812404/CA


















