
MICHAEL RUBEM MIRANDA TIAGO - bdtd.inpa.gov.brbdtd.inpa.gov.br/bitstream/tede/2541/5/Michael Rubem...
55
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA, CONSERVAÇÃO E BIOLOGIA EVOLUTIVA – PPG GCBEv Microbiota fúngica cultivável associada às colmeias e castas de Melipona interrupta e Melipona seminigra (Apidae, Meliponini) MICHAEL RUBEM MIRANDA TIAGO Manaus, Amazonas Agosto/2017
Transcript of MICHAEL RUBEM MIRANDA TIAGO - bdtd.inpa.gov.brbdtd.inpa.gov.br/bitstream/tede/2541/5/Michael Rubem...

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA, CONSERVAÇÃO E
BIOLOGIA EVOLUTIVA – PPG GCBEv
Microbiota fúngica cultivável associada às colmeias e castas de Melipona
interrupta e Melipona seminigra (Apidae, Meliponini)
MICHAEL RUBEM MIRANDA TIAGO
Manaus, Amazonas
Agosto/2017

Michael Rubem Miranda Tiago
Microbiota fúngica cultivável associada às colmeias e castas de Melipona
interrupta e Melipona seminigra (Apidae, Meliponini)
Orientadora: Dra. Gislene Almeida Carvalho-Zilse.
Co-Orientador: Dr. João Vicente Braga de Souza.
Financiamento do Projeto: PPI – INPA/MCTI; PRJ – 016/2012.
Bolsa: CAPES
Manaus, Amazonas
Agosto/2017
Dissertação apresentada ao Programa de Pós-Graduação
em Genética, Conservação e Biologia Evolutiva do
Instituto Nacional de Pesquisas da Amazônia como parte
dos requisitos para a obtenção do título de Mestre em
Genética, Conservação e Biologia Evolutiva.

iii
Microbiota fúngica cultivável associada às colmeias e castas de Melipona
interrupta e Melipona seminigra (Apidae, Meliponini)
Dissertação apresentada ao Programa de Pós-
Graduação em Genética, Conservação e Biologia
Evolutiva do Instituto Nacional de Pesquisas da
Amazônia, como requisito para a obtenção do título
de Mestre em Genética, Conservação e Biologia
Evolutiva.
APROVADA EM: 18 / 08 / 2017
BANCA EXAMINADORA
Dr. Carlos Gustavo Nunes da Silva
Universidade Federal do Amazonas
Dra. Míriam Silva Rafael
Instituto Nacional de Pesquisas da Amazônia
Dr. Wanderli Pedro Tadei
Instituto Nacional de Pesquisas da Amazônia

iv
T551 Tiago , Michael Rubem Miranda Tiago
Microbiota fúngica cultivável associada às colmeias e castas de
Melipona interrupta e Melipona seminigra (Apidae, Meliponini)/
Michael Rubem Miranda Tiago . --- Manaus: [s.n.], 2017.
53 f.: il.
Dissertação (Mestrado) --- INPA, Manaus, 2017.
Orientador: Gislene Almeida Carvalho-Zilse
Coorientador: João Vicente Braga de Souza
Área de concentração: Genética, Conservação e Biologia
Evolutiva
1. Abelhas . 2.Fungos. 3. Diversidade . I. Título.
CDD 638.1

v
DEDICATÓRIA
Ao Deus Eterno invisível, mas real, no qual deposito toda a minha confiança e
crença, que me formou desde o ventre de minha mãe e que de acordo com o beneplácito
de Sua vontade permitiu que eu atingisse esta meta terrena. O Deus a quem glorifico é o
Deus das Escrituras Sagradas que derramou sua Graça e misericórdia sobre a minha
vida, por isso, guardarei as suas Palavras no meu coração e farei delas uma lâmpada
para os meus pés e uma luz para o meu caminho.
Aos meus Pais, que com um Amor incondicional se dedicaram e se sacrificaram
por minha felicidade e vida profissional, por terem sido meus apoiadores desde minha
infância me ofertando uma sólida educação e uma forte disciplina. Ao meu Irmão que
muitas vezes foi o meu grande companheiro durante o colegial e hoje faz parte desta
história.
A minha Esposa e Filha, por hoje serem a minha tão sonhada família e por
terem me apoiado e me animado nos momentos difíceis demonstrando um Amor
Verdadeiro que fortalece os nossos laços e a nossa aliança. Que a nossa união seja para
toda a vida!

vi
AGRADECIMENTOS
Às instituições INPA e CAPES pelo apoio estrutural e financeiro.
À minha orientadora Dra. Gislene Almeida Carvalho Zilse, por sua clareza
científica e por todos os ensinamentos que contribuíram significativamente para a
conclusão desde Mestrado.
Ao meu co-orientador Dr. João Vicente Braga de Souza, que foi imensamente
generoso em disponibilizar a infraestrutura necessária para o desenvolvimento do
trabalho além de suas contribuições científicas.
À Dra. Ana Cláudia Alves Cortez, por ser a minha grande professora na área de
Micologia desde quando ingressei no INPA ainda como aluno de Iniciação Científica.
Para verificação da diversidade, agradeço a todos os colegas do laboratório de
Micologia que me ajudaram de alguma forma, como o Diego Fernando que me ajudou
com os testes moleculares, a “Dona Lili” e “Seu Ray” que já me ajudam há muitos anos.
Aos meus caros colegas do Grupo de Pesquisas em Abelhas (GPA) que ao longo
desta jornada me apoiaram e ajudaram da forma como lhes foi possível. Ao técnico
Diego Albuquerque, que me acompanhou em todas as coletas no Meliponário e sempre
se demonstrou uma pessoa acessível e disposta para ajudar. Ao companheiro André
Mascarenhas que compartilhou de seus recursos para o trabalho, à colega Diana Brito
pelas boas discussões e incentivo. A todos os demais que compõem o GPA e que
sempre me trataram com respeito e simpatia.
Por que assim diz o Senhor Deus, o Santo de Israel:
Em vos converter e em sossegar, está a vossa salvação;
na tranquilidade e na confiança a vossa força...
Parafraseado de Isaías 30: 15.

vii
RESUMO
Às colmeias de abelhas Amazônicas da tribo Meliponini, Melipona interrupta e M.
seminigra, que possuem grande importância no ecossistema como polinizadoras de
diversas espécies de plantas e na produção de mel, estão associadas uma grande
variedade de entomofauna e microbiota. A fim de conhecer a diversidade da microbiota
associada às colmeias e castas de M. interrupta e M. seminigra este estudo realizou a
identificação morfológica e molecular de fungos presentes no ambiente interno das
colmeias e nas células de cria com larva ou pupa (de rainha ou de operária) para os
períodos chuvoso e seco em Manaus-AM. Aplicou-se a técnica de sedimentação de
esporos a fim de amostrar fungos do ambiente interno da colmeia. Fungos internos às
células de cria em fase larval foram isolados após incubação dos discos de cria sendo
que a massa fúngica crescida no interior destas células foi coletada com alça
microbiológica, e fungos do interior das células com pupa de operária ou de rainha
foram obtidos pela lavagem individual destas células para isolamento da microbiota
interna. Os fungos isolados foram identificados pela análise das estruturas morfológicas
e, em seguida, pela amplificação e sequenciamento das regiões ITS1-5.8S e ITS2 do
DNA ribossomal. Por meio destas análises foram identificados 13 taxa a partir de 1.542
isolados fúngicos: os filamentosos foram Aspergillus fumigatus, A. nomius, A. niger, A.
ochraceus, Aspergillus sp. (T1), Penicillium citrinum, Cladosporium cladosporioides e
Monascus ruber; as leveduras foram Cystobasidium minutum, Candida apicola,
Candida versatilis, Candida etchellsii e Candida famata. Em geral, o táxon mais
abundante foi Aspergillus spp.. As leveduras foram isoladas apenas nas células de cria.
A maior diversidade de fungos em M. seminigra foi observada para a estação chuvosa
enquanto em M. interrupta não houve variação significativa entre as estações.
Ocorrência exclusiva de fungos foi registrada tanto por espécies quanto por ambiente e
por casta: Candida etchellsii ocorreu apenas em células de operárias de M. interrupta;
enquanto em M. seminigra Aspergillus ochraceus foi restrito ao ambiente interno,
Cystobasidium minutum às células de operárias e Aspergillus nomius às células de
rainhas. No entanto, tais achados requerem maior investigação sobre a possível relação
desta microbiota com estes ambientes e com as castas.
Palavras-chave: Diversidade, fungos, abelhas Amazônicas.

viii
ABSTRACT
The amazonian bee colonies of the Meliponini tribe, Melipona interrupta and M.
seminigra, which have great importance in the ecosystem as pollinators of diverse
species of plants and in the production of honey, are associated a great variety of
entomofauna and microbiota. In order to know the diversity of the microbiota associated
to the hives and castes of M. interrupta and M. seminigra, this study carried out the
morphological and molecular identification of fungi present in the internal environment
of the hives and in the cells of larva or pupa (queen or of workers) for the rainy and dry
seasons in Manaus-AM. The spore sedimentation technique was applied to collect
sample fungi from the internal environment of the hive. The fungi of the internal
environment of the breeding cells at the larval stages were isolated from the fungal mass
grown inside the craet cells and collected with a microbiological loop, the fungi on the
pupa cells (queens and workers) were isolated by washing the pupal cells. The isolated
fungi were identified by the analysis of the morphological structures and then by the
amplification and sequencing of the ITS1-5.8S and ITS2 regions of the ribosomal DNA.
By means of these analyzes 13 taxa were identified from 1,542 fungal isolates: the
filaments were Aspergillus fumigatus, A. nomius, A. niger, A. ochraceus, Aspergillus sp.
(T1), Penicillium citrinum, Cladosporium cladosporioides and Monascus ruber; the
yeasts were Cystobasidium minutum, Candida apicola, Candida versatilis, Candida
etchellsii and Candida famata. In general, the most abundant taxon was Aspergillus
spp.. Yeasts were isolated only in the rearing cells. The greatest diversity of fungi in M.
seminigra was observed for the rainy season while in M. interrupta there was no
significant variation between seasons. Exclusive occurrence of fungi was recorded both
by species and by environment and caste: Candida etchellsii occurred only in worker
cells of M. interrupta; while in M. seminigra Aspergillus ochraceus was restricted to the
internal environment, Cystobasidium minutum to worker cells and Aspergillus nomius to
queen cells. However, such findings require further research on the possible relationship
of this microbiota to these environments and to the castes.
Key-words: Diversity, fungus, Amazon bee.

ix
SUMÁRIO
1. INTRODUÇÃO .......................................................................................................... 14
2. OBJETIVOS ............................................................................................................... 19
2.1 Geral ...................................................................................................................... 19
2.2 Específicos ............................................................................................................ 19
3. MATERIAL E MÉTODOS ........................................................................................ 20
3.1 Coletas .................................................................................................................. 20
3.1.1 Fungos cultiváveis anemófilos do ambiente interno das colmeias ................ 21
3.1.2 Fungos cultiváveis dos lavados do interior das células de cria ...................... 23
3.1.3 Fungos cultiváveis das células de cria na fase de ovo ou larva...................... 24
3.2 Conservação dos fungos isolados ......................................................................... 25
3.3 Identificação .......................................................................................................... 25
3.3.1 Macro e micromorfologia............................................................................... 25
3.3.2 Molecular - regiões ITSs do rDNA ................................................................ 26
3.4 Análise estatística ................................................................................................. 27
3.4.1. Diversidade de fungos obtidos por espécies, casta e período sazonal .......... 27
4. RESULTADOS .......................................................................................................... 29
4.1 Isolamento dos fungos das colmeias de Melipona interrupta e M. seminigra ..... 29
4.2 Identificação morfológica dos fungos isolados .................................................... 32
4.3 Identificação molecular dos fungos isolados ........................................................ 37
4.3.1 Amplificação das regiões Its-1 e Its-2 do rDNA ............................................ 37
4.3.2 Sequenciamento das regiões Its-1 e Its-2 do rDNA ....................................... 37
4.4 Índices de diversidade fúngica .............................................................................. 39
5. DISCUSSÃO .............................................................................................................. 45
6. CONCLUSÃO ............................................................................................................ 49
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................... 50

x
LISTA DE TABELAS
Tabela 1. Origem das amostras de fungos cultiváveis isolados em colmeias de Melipona
interrupta (MI) e M. seminigra (MS) mantidas no Meliponário GPA-INPA, Manaus-
AM nos anos de 2015 e 2016. ........................................................................................ 21
Tabela 2. Número (n) e frequência relativa (%) dos fungos isolados dos discos de cria,
células de cria e do ambiente interno das colmeias de Melipona interrupta e M.
seminigra. ....................................................................................................................... 29
Tabela 3. Fungos isolados de colmeias de Melipona interrupta e M. seminigra no
Meliponario GPA-INPA, Manaus-AM, identificados por análise molecular das regiões
ITS1-5.8S e ITS2 do rDNA. ........................................................................................... 38
Tabela 4. Número e frequência relativa dos fungos identificados e não identificados
isolados do ambiente interno (AI) das colmeias, das células de cria com larva (L) e das
células de cria com pupas de rainha (R) ou operária (O) de Melipona interrupta e M.
seminigra no Meliponário GPA-INPA, Manaus-AM. ................................................... 39
Tabela 5. Número e frequência relativa dos fungos isolados nas colmeias (C1 a C5) de
Melipona interrupta nas estações chuvosa e seca no Meliponário GPA-INPA, Manaus-
AM. ................................................................................................................................. 40
Tabela 6. Número e frequência relativa dos fungos isolados nas colmeias (C1 a C5) de
Melipona seminigra nas estações chuvosa e seca no Meliponário GPA-INPA, Manaus-
AM. ................................................................................................................................. 41

xi
LISTA DE FIGURAS
Figura 1. Esquema da região ITS1-5.8S-ITS2 do rDNA fúngico (BRASILEIRO et al.
2004). .............................................................................................................................. 15
Figura 2. Colmeias de Melipona interrupta e M. seminigra do Meliponário do INPA,
Manaus, AM. A - Melipona interrupta e B - M. seminigra. .......................................... 20
Figura 3. Coleta de fungos do ambiente interno das colmeias de Melipona interrupta
(A e B) e M. seminigra (C e D). ..................................................................................... 22
Figura 4. Colônias de fungos, em meio BDA, isolados do ambiente interno das colmeias
de Melipona interrupta e M. seminigra mantidas no Meliponário GPA-INPA, Manaus-
AM. ................................................................................................................................. 23
Figura 5. Vista geral de um disco de cria de Melipona seminigra com destaque para
massa fúngica presente na parede interna da célula de cria contendo ovo. .................... 25
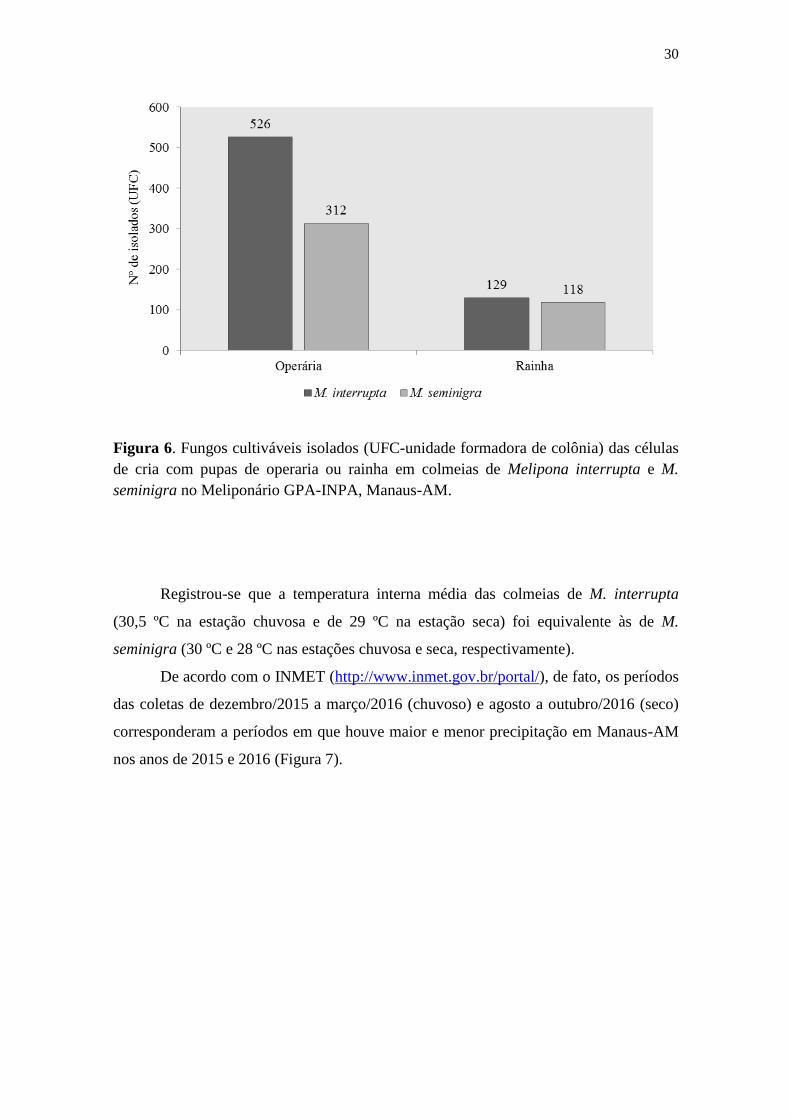
Figura 6. Fungos cultiváveis isolados (UFC-unidade formadora de colônia) das células
de cria com pupas de operaria ou rainha em colmeias de Melipona interrupta e M.
seminigra no Meliponário GPA-INPA, Manaus-AM. ................................................... 30
Figura 7. Precipitação pluvial em Manaus – AM durante os meses de coleta de amostras
fúngicas em colmeias de Melipona interrupta e M. seminigra. Estação chuvosa:
dezembro/2015 e março/2016; estação seca: agosto, setembro e outubro/2016. ........... 31
Figura 8. Abundância de fungos isolados em colmeias de Melipona interrupta e M.
seminigra nas estações seca e chuvosa no Meliponário GPA-INPA, Manaus-AM. ...... 31
Figura 9. Aspergillus niger isolados das colmeias de Melipona interrupta e M.
seminigra do Meliponário GPA-INPA/Manaus-AM. .................................................... 32
Figura 10. Aspergillus ochraceus isolado das colmeias de Melipona interrupta e M.
seminigra do Meliponário GPA-INPA/Manaus-AM. .................................................... 33
Figura 11. Aspergillus sp. (T1) isolado das colmeias de Melipona interrupta e M.
seminigra do Meliponário GPA-INPA, Manaus-AM. ................................................... 33
Figura 12. Aspergillus nomius (T2) isolado de M. interrupta e M. seminigra do
Meliponário GPA-INPA, Manaus-AM. ......................................................................... 34
Figura 13. Aspergillus fumigatus (T3) isolado de Melipona interrupta e M. seminigra do
Meliponário GPA-INPA, Manaus-AM. ......................................................................... 34
Figura 14. Cladosporium cladosporioides isolado das colmeias de Melipona interrupta
e M. seminigra do Meliponário GPA-INPA, Manaus-AM. ........................................... 35

xii
Figura 15. Penicillium citrinum isolado de colmeias de Melipona interrupta e M.
seminigra do Meliponário GPA-INPA, Manaus-AM. ................................................... 35
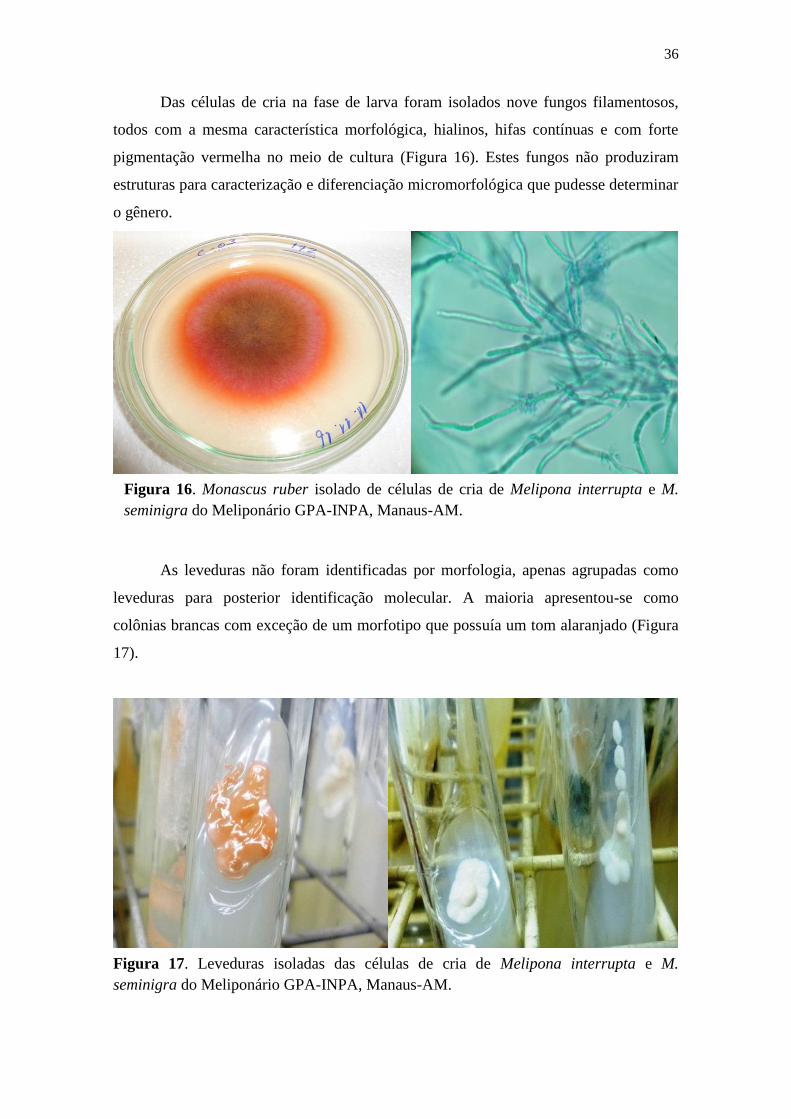
Figura 16. Monascus ruber isolado de células de cria de Melipona interrupta e M.
seminigra do Meliponário GPA-INPA, Manaus-AM. ................................................... 36
Figura 17. Leveduras isoladas das células de cria de Melipona interrupta e M. seminigra
do Meliponário GPA-INPA, Manaus-AM. .................................................................... 36
Figura 18. Perfil de amplificação da região Its-1 e Its-2 de fungos isolados de colmeias
de Melipona interrupta (amostras 1 a 3) e M. seminigra (amostras 4 a 5) do Meliponário
GPA-INPA, Manaus-AM. C+ = controle positivo. C- = controle negativo sem DNA. . 37
Figura 19. Frequência (%) dos fungos isolados de Melipona interrupta e M. seminigra
nas estações seca e chuvosa no Meliponário GPA-INPA, Manaus-AM. ....................... 44

xiii
LISTA DE ABREVIAÇÕES E SIGLAS
INPA – Instituto Nacional de Pesquisas da Amazônia
COSAS – Coordenação de Sociedade, Ambiente e Saúde
GPA – Grupo de Pesquisas em Abelhas
CAPES - Coordenação de Aperfeiçoamento de Pessoal de Nível Superior
ITS – Internal Transcribed Spacer
rDNA – DNA ribossomal
BDA – Batata, Dextrose, Agar
UFC – Unidade Formadora de Colônia

14
1. INTRODUÇÃO
Fungos são seres vivos eucariontes que vivem em simbiose ou em vida
parasitária com outros organismos. São por isso, importantes para a sobrevivência de
outras espécies. Em caso de mutualismo também são importantes decompositores
primários assumindo um papel de destaque nos ecossistemas (RIBEIRO et al. 2012).
Para sua disseminação muitos fungos dependem do ar. Os fungos encontrados
neste ambiente atmosférico são chamados de anemófilos. Os fungos anemófilos são
responsáveis por várias doenças alérgicas como asma e rinite, por essa razão sua
ocorrência é sempre importante de ser notificada, pois são causadores de doenças a
seres humanos e/ou a animais como as espécies de abelhas utilizadas na produção de
mel (LACAZ et al. 2002; CASTAGNINO et al. 2006a; O’CONNOR et al. 2014).
Estudos anteriores com fungos anemófilos demonstraram a diversidade destes
microrganismos na cidade de Manaus. Os estudos de FONSECA e CONCEIÇÃO
(1977) e de FURTADO e FERRARONI (1998) investigaram a frequência e a
diversidade destes fungos em hospitais, sendo registrado que o gênero Cladosporium foi
o mais frequente atingindo o ápice nos meses de Setembro, Outubro e Novembro.
A identificação dos fungos baseia-se convencionalmente nas suas características
macro e micromorfológicas, porém, em muitos casos, há insuficiência de informações
para distinguir duas ou mais espécies que apresentam alto grau de semelhança, o que
torna o processo de identificação convencional limitado para se determinar com clareza
uma espécie ou um gênero fúngico. Com a intenção de suprir essa deficiência, as
ferramentas moleculares passaram a ser utilizadas na identificação destes
microrganismos e, atualmente, se constituem ferramenta importante para a
caracterização taxonômica dos fungos (SANDERS et al. 1995; PORTER e GOLDING
2012).
O ácido desoxirribonucleico ribossomal (rDNA) é uma molécula que possui
sequências codificantes e sequências não codificantes. Atualmente, sabe-se que as
sequências codificantes são conservadas em cada espécie de fungo e não apresentam
variações, no entanto, as sequências não codificantes denominadas de regiões ITSs
(Internal Transcribed Spacer) variam de uma espécie para outra conferindo a cada
espécie uma sequencia ITS especifica (MENEZES et al. 2015). Desta forma, as regiões
ITS1 e ITS2 do rDNA de fungos têm sido utilizadas como ferramentas taxonômicas na
identificação de fungos por meio da identidade das sequencias ITSs. Essas regiões ITSs
estão localizadas entre os genes 18S e 28S do rDNA contendo uma pequena região

15
codificadora a 5.8S (WHITE et al. 1990; SANDERS et al. 1995; BRASILEIRO et al.,
2004; PORTER e GOLDING, 2012) (Figura 1).
Apesar de seu enorme potencial como ferramenta molecular para a identificação
de fungos, estudos sobre a diversidade de fungos associados às colmeias de abelhas são
raros. Ainda, os pouquíssimos estudos são voltados à caracterização da microbiota
fúngica restrita ao mel, ao pólen, as células de cria ou ao corpo da abelha (MENEZES et
al. 2015; FERRAZ et al. 2008; STUART et al. 2004) e se baseiam em analises
morfológicas. Os primeiros autores que usaram a região ITS para fins taxonomicos de
fungos em abelhas sem ferrão foram MENEZES et al. (2010). Estes autores registraram
a presença do fungo Monascus no interior das celulas de cria o qual é comido pelas
larvas de abelhas Scaptotrigona depilis antes de iniciarem o consumo do alimento larval
propriamente dito. Estes autores demonstraram que em condições in vitro, a taxa de
sobrevivência de larvas é maior na presença do fungo Monascus comparada a larvas
cultivadas na ausência do fungo.
No Brasil há registros da presença de leveduras associadas com abelhas sem
ferrão, as leveduras mais abundantes já descritas são Starmerella meliponinorum,
Candida apicola, Zygosaccharomyces machadoi, Starmerella neotropicalis assim como
outras espécies do clado Starmerella (ROSA e LACHANCE 2005; DANIEL et al.
2013; ROSA et al. 2013). Há ainda os estudos realizados com Apis mellifera que
identificam a microbiota intestinal das abelhas abrindo novas perspectivas para a
compreensão das relações dos microrganimos com as abelhas (ENGEL et al. 2012;
ENGEL e MORAN 2013).
A interação entre fungos e insetos vem sendo estudada com objetivo de
beneficiar as sociedades humanas. Novas ferramentas biotecnológicas estão ganhando
espaço para a resolução de problemas, como a utilização de fungos no controle
Figura 1. Esquema da região ITS1-5.8S-ITS2 do rDNA fúngico (BRASILEIRO et al.
2004).

16
biológico de insetos praga e vetores de doenças, com isso, levando a produção de
substâncias bioativas de interesse comercial (SILVA et al. 2012; FONSECA 2016).
Fungos associados a abelhas podem ser uma nova fonte de substâncias de interesse
industrial, o conhecimento e exploração desta possibilidade tem grande importância na
atualidade, pois os compostos produzidos a partir de microrganismos causam menor
dado à natureza e possuem um menor custo (BUENO 2016; FONSECA 2016).
Apesar de relatos constantes na literatura da existência de fungos em colmeias
de abelhas sem ferrão (OLIVEIRA et al. 2013; SOUZA 2014) até o momento não há
registros bibliográficos sobre a identificação da diversidade fúngica associada a
colmeias de abelhas sem ferrão amazônicas.
As abelhas são insetos da Ordem Hymenoptera juntamente com as formigas e as
vespas (DANFORTH 2007). Apresentam como característica peculiar o habito de se
alimentar de pólen e néctar sendo de enorme importância para os ecossistemas
(CARVALHO-ZILSE e NUNES-SILVA 2012). As abelhas sem ferrão, objeto deste
estudo, pertencem à subfamília Apinae e tribo Meliponini. Das 418 espécies
encontradas no mundo, 244 espécies válidas estão presentes no Brasil (PEDRO 2014)
distribuídas em 29 gêneros. Das 116 espécies existentes no Amazonas, merecem
destaque a Melipona interrupta e M. seminigra por serem as mais comumente
cultivadas para a Meliponicultura (CARVALHO-ZILSE e NUNES-SILVA 2012).
Melipona (Michmelia) seminigra Friese, 1903 e Melipona (Melikerria)
interrupta Latreille, 1811, são espécies que tem fundamental importância na polinização
e na produção de mel por meio da atividade de Meliponicultura. M. seminigra é
polinizadora de espécies vegetais como tucumã (Vetix cymosa - Verbenacea), Mouriria
ulei (Melastomaceae), Myrcia amazonica e outros (MARQUES-SOUZA et al. 2002).
FERREIRA e ABSY (2015) identificaram a origem do pólen carregado por M.
interrupta e M. seminigra, em Manaus/AM, sendo estes originados de cinco espécies
vegetais. Os polens eram oriundos de Mimosa sensitiva (Fabaceae/Mimosoideae);
Mimosa pudica (Fabaceae/Mimosoideae); Miconia e Bellucia (Melastomataceae);
Myrciaria cauliflora (Myrtaceae).
Durante as atividades de coleta de matéria prima para as necessidades da
colmeia, propágulos fúngicos podem ser transportados aderidos ao corpo da abelha.
STUART et al. (2004) identificaram 11 gêneros de fungos aderidos ao corpo de
operárias do gênero Trigona uma espécie de abelha Meliponini. As abelhas foram
coletadas durante o processo de forrageamento levantando a hipótese de que elas podem

17
ser disseminadoras de fungos, incluindo os patogênicos, para culturas vegetais
(STUART et al. 2004).
As informações disponíveis a respeito de fungos associados a abelhas são
escassas, sendo os relatos ocasionais voltados para Apis mellifera de onde foi
identificado o fungo Ascosphaera apis como sendo o causador da morte destas abelhas
durante o desenvolvimento larval (CASTAGNINO et al. 2006). Outros registros
indicam fungos associados ao pólen sendo transportado para a colmeia pelas abelhas
durante o processo de forrageamento, o que acarreta a possivel contaminação da
colmeia por estes propágulos fúngicos (GONZÁLEZ et al. 2005; SHUTLER et al.
2014).
Com relação à diversidade da microbiota fúngica associada especificamente às
colmeias de abelhas Melipona não há registros que suportem afirmações definitivas. O
estudo de FERRAZ et al. (2008) isolou e identificou fungos presentes no corpo de
abelhas mortas da espécie M. subnitida Ducke coletadas na região peridomiciliar da
colmeia, sendo identificados os fungos Aspergillus sp.; Aspergillus niger; A. terreus;
Penicillium sp.; Curvularia sp.; Monilia sp.; Nigrospora sp.; Cladosporium sp. e
Tricoderma sp.. No entanto, o fato de não haver outros estudos semelhantes para
comparação torna-se difícil afirmar que estes gêneros fúngicos estejam associados
exclusivamente a este grupo de abelhas.
Nenhum dos estudos anteriormente citados relata a identificação da microbiota
fúngica interna às colmeias de abelha sem ferrão, apesar de potencialmente estas
abelhas serem carreadoras destes microrganismos.
Assim, uma questão intrigante que se levanta é sobre o potencial de transporte
dessa microbiota aderida ao corpo das abelhas ou ao pólen para dentro do ambiente da
colmeia. E ainda, se haveria alguma associação desta microbiota ao microambiente das
colmeias, das espécies e até mesmo das células de cria, que apesar de terem o mesmo
tamanho produzem machos ou fêmeas, estas últimas que podem se desenvolver nas
castas rainhas ou operarias de acordo com fatores genéticos e ambientais envolvidos
(KERR 1950).
Há muito se sabe que nos insetos da ordem Hymenoptera a determinação sexual
segue o padrão de haplodiploidia no qual os machos são haploides gerados por
partenogênese arrenótoca, ou seja, os ovos não fecundados dão origem a machos,
enquanto que os ovos fecundados dão origem às fêmeas (KERR et al. 2001). Portanto, a
genética é o fator determinante dos sexos.

18
No entanto, entre as fêmeas, ao final do último estágio larval (L3 – larva pré-
defecante) após consumo de todo o alimento disponível na célula de cria individual se
diferenciam as castas: rainha ou operaria (Bonetti, 2008). De acordo com a hipótese de
KERR (1950) a diferenciação de abelhas Melipona em castas rainha ou operária se dá
por um sistema genético-alimentar que depende de uma dupla heterozigosidade dos
genes xa e xb. Operarias são homozigotas para um ou ambos os genes. Rainhas são
produzidas a partir de larvas duplo-heterozigotas que tenham sido suficientemente
alimentadas. Caso não tenham recebido alimento suficiente durante a vida larval se
desenvolvem em operarias. Assim, tem-se a indicação de que fatores ambientais podem
influenciar a diferenciação de castas. Neste sentido, o alimento larval – constituído de
mel, pólen, geleia real – é fundamental para o processo de produção de rainhas e,
consequentemente, para a reprodução da colmeia (MENEZES et al. 2007). A
possibilidade de associação fúngica ao alimento disponível às larvas abre a perspectiva
investigativa acerca da participação de fungos associados à produção de rainhas.
BRITO et al. (2013) afirmam que a produção de rainhas em Melipona é limitada
pelas condições ambientais. Eles observaram uma forte e negativa correlação entre a
proporção de rainhas e a precipitação acumulada para M. interrupta o que não ocorreu
para M. seminigra. Os autores sugerem que tais resultados provavelmente envolvem um
efeito indireto da restrição de disponibilidade de alimentos na natureza e,
consequentemente, na colmeia.
Considerando todas estas informações, a investigação da microbiota fúngica
associada torna-se de relevante importância para conhecimento taxonômico dos fungos
associados às colmeias. Não há registro de estudos da microbiota fúngica em abelhas
sem ferrão amazônicas e sua potencial relação com as espécies, às colmeias ou as
castas.
Com base nesta perspectiva de abordagem, o objetivo deste estudo foi identificar
morfológica e molecularmente a microbiota fúngica potencialmente associada às
colmeias e às castas de duas espécies de abelhas sem ferrão Melipona interrupta e
Melipona seminigra, por serem as duas principais espécies de abelhas utilizadas em
Meliponicultura no Amazonas. O conhecimento dos fungos associados às castas tem a
intenção de analisar se algum gênero fúngico específico possui relação direta com
alguma das castas produzidas. A incidência destes fungos nas duas estações sazonais
características da região Amazônica, seca e chuvosa foi verificada, pois já se sabe que
nestes a segregação das castas também varia.

19
Desta forma testou-se as seguintes hipóteses alternativas:
H0: Existe relação da microbiota fúngica cultivável em colmeias e castas de abelhas
Melipona interrupta e Melipona seminigra.
H1: Não existe relação da microbiota fúngica cultivável em colmeias de abelhas
Melipona interrupta e Melipona seminigra.
H1: Não existe relação da microbiota fúngica cultivável em castas de abelhas Melipona
interrupta e Melipona seminigra.
H1: Não existe relação da diversidade da microbiota fúngica cultivável em colmeias de
Melipona interrupta e Melipona seminigra e a sazonalidade.
2. OBJETIVOS
2.1 Geral
Identificar a diversidade dos fungos isolados das colmeias de Melipona
interrupta e M. seminigra pela macro e micromorfologia e identificação molecular pela
região ITS1-5.8S-ITS2 do rDNA.
2.2 Específicos
Investigar e comparar a diversidade dos fungos cultiváveis isolados do ambiente
interno de colmeias de Melipona interrupta e M. seminigra;
Verificar a microbiota fúngica presente no microambiente das células de cria
durante a fase larval;
Examinar a diversidade de fungos cultiváveis isolados do microambiente de
células de cria em estágio de pupa considerando sua potencial correlação com as castas
rainha e operaria;
Verificar se há influência sazonal sobre a diversidade dos fungos isolados das
espécies e dos ambientes (colmeia, célula de cria com pupa).

20
3. MATERIAL E MÉTODOS
3.1 Coletas
As amostras fúngicas foram obtidas a partir de cinco colmeias de Melipona
interrupta e cinco de M. seminigra (Figura 2) sorteadas do plantel mantido no
Meliponário do Grupo de Pesquisas em Abelhas (GPA) do Instituto Nacional de
Pesquisas da Amazônia (INPA), Manaus, AM, Brasil.
Figura 2. Colmeias de Melipona interrupta e M. seminigra do Meliponário do INPA,
Manaus, AM. A - Melipona interrupta e B - M. seminigra.
As coletas foram realizadas em dois períodos distintos: dezembro/2015 a
março/2016 correspondente ao período chuvoso, e em agosto a outubro/2016
correspondente ao período seco (Tabela 1).
A B

21
Tabela 1. Origem das amostras de fungos cultiváveis isolados em colmeias de
Melipona interrupta (MI) e M. seminigra (MS) mantidas no Meliponário GPA-INPA,
Manaus-AM nos anos de 2015 e 2016.
Local Amostras Método de coleta Período MI MS Ambiente
interno da
colmeia
Fungos dispersos
no ar
Microplacas com
furos na tampa
contendo Ágar
Sabouraud
Chuvoso
(02 e
03/2016)
Seco
(08 e
10/2016)
10 placas em
duplicata
10 placas em
duplicata
Interior
das
células de
cria
Fungos do
alimento das
células contendo
ovo ou larva
Fragmentos da
massa fúngica sobre
o alimento coletados
com alça
microbiológica
3 discos de
cria
3 discos de
cria
Fungos obtidos
por lavados das
células contendo
pupa defecante
5 discos de cria de
cada espécie.
200 µL de
lavado/célula.
Mix de 3 células de
operaria e 3 de
rainhas por disco
Chuvoso
(12/2015)
Seco
(09/2016)
10 placas em
duplicata
10 placas em
duplicata
Total de Amostras 23 23
A temperatura interna das colmeias foi registrada com auxílio de termômetro
digital para ambos os períodos chuvoso e seco.
3.1.1 Fungos cultiváveis anemófilos do ambiente interno das colmeias
Para coleta dos fungos anemófilos no ambiente interno das colmeias foram
expostas placas de petri descartáveis de diâmetro 60x15 mm contendo Ágar Sabouraud
20 % (Dicf®
) (Dextrose 20 g; Peptona 10 g; Ágar 20 g para 1.000 mL de água destilada)
com antibiótico (Cloranfenicol 250 g/L) (MINAMI 2003) sobre os discos de cria
(Figura 3). Tal procedimento de coleta foi realizado em fevereiro e março/2016 (período
chuvoso) e entre agosto e outubro/2016 (período seco).

22
Figura 3. Coleta de fungos do ambiente interno das colmeias de Melipona interrupta
(A e B) e M. seminigra (C e D).
As placas descartáveis foram furadas na parte superior (tampa) para possibilitar
a entrada das células de reprodução fúngicas, possivelmente presente no ambiente
interno das colmeias, sem retirada da tampa das placas durante o tempo de exposição (1
hora). Observou-se que as abelhas tendem a eliminar qualquer material estranho dentro
da colmeia; por isso a tampa das placas foi mantida para se evitar a degradação do meio
de cultura pelas abelhas.
Após uma hora de exposição das placas no ambiente interno das colmeias, estas
foram retiradas e transferidas para estufa à 28 ºC. Após cinco dias foi possível visualizar
ao olhar o crescimento de colônias fúngicas que foram isoladas em tubos de ensaio
contendo BDA (LACAZ et al. 2002) (Figura 4).
A
C
B
D

23
3.1.2 Fungos cultiváveis do interior das células de cria com pupa
Um disco de cria por colmeia de ambas as espécies (cinco colmeias por espécie)
destinados ao isolamento de fungos na fase de pupa, os discos de cria foram coletados
em Dezembro/2015 (período chuvoso) e em Setembro/2016 (período seco). Os discos
contendo abelhas na fase de pupa foram coletados e acondicionados em potes plásticos
estéreis e submetidos à exposição de luz ultravioleta em cabine de segurança durante 15
minutos para descontaminação externa do cerume. Com as abelhas no estágio de pupa,
os discos de cria hermeticamente fechados nos potes plásticos foram transportados para
o Laboratório de Micologia do INPA da Coordenação de Sociedade, Ambiente e Saúde
(COSAS-INPA), onde foi realizado o isolamento e a identificação por microscopia das
colônias fúngicas.
Como a diferenciação de castas ocorre no último instar larval e depende da
alimentação total da larva (KERR et al. 1996), os indivíduos foram retirados dos discos
de cria na fase de pupa, fase imediatamente posterior ao último estagio larval e onde já é
possível identificar morfologicamente a casta para se investigar possível relação da
microbiota presente nas células de cria com as castas rainha e operária.
Assim, para o isolamento de fungos das células de cria com os indivíduos já na
fase de pupa, foram selecionadas três células, por disco de cria coletado, contendo
rainhas e três células contendo operárias para ambas as espécies. As pupas de rainhas e
Figura 4. Colônias de fungos, em meio BDA, isolados do ambiente interno das
colmeias de Melipona interrupta e M. seminigra mantidas no Meliponário GPA-INPA,
Manaus-AM.

24
operárias foram aleatoriamente selecionadas e transferidas para placas de Elisa, sendo
armazenadas em estufa com temperatura controlada à 30 ºC onde continuaram o seu
desenvolvimento para serem devolvidas às colmeias de origem. Inicialmente, cada
célula de cria selecionada, já sem a pupa, foi preenchida com 200 µL de água destilada
esterilizada, sendo levemente agitada com a ponteira da pipeta constituindo-se assim os
lavados do interior das células de cria. Estes lavados das três células de uma mesma
casta foram homogeneizados em uma única amostra sendo o volume final dos lavados
de aproximadamente 600 µL, o mesmo foi feito com os lavados da outra casta. Assim,
cada amostra correspondeu ao lavado das células de três indivíduos de uma mesma
casta de uma mesma colmeia. Ao todo foram retiradas duas amostras (2 pools de 3
células de cria) por disco de cria ou por colmeia, uma de rainha e outra de operária. Os
lavados foram realizados uma vez em cada período sazonal.
Do volume final dos lavados homogeneizados, 100 µL foram plaqueados por
esgotamento em Ágar Sabouraud 20 % (Dicf®) com antibiótico (Cloranfenicol 250 g/L)
e incubados a 27 ºC para a obtenção das unidades formadoras de colônias (UFC). Após
o crescimento das colônias foi realizada a contagem das UFC e o isolamento em tubos
de ensaio contendo BDA (HIMEDIA®) (Infusão de batata 200 g; Dextrose 20 g; Agar
15 g) (LACAZ et al. 2002). Todo o processo de isolamento foi realizado em câmara de
fluxo laminar.
3.1.3 Fungos cultiváveis das células de cria com ovo ou larva
Para o isolamento de fungo que cresce dentro das células de cria contendo
indivíduos na fase larval foram coletados discos de cria nesta fase. Os discos foram
submetidos à descontaminação por exposição à luz ultravioleta durante 15 minutos,
sendo desoperculados e incubados à temperatura ambiente, durante aproximadamente
20 dias, para observação diária e monitoramento da possível presença de fungos. Com o
auxílio de um estereomicroscópio Stemi DV4 (Zeiss, Alemanha) foi possível observar
estruturas fúngicas, cujas hifas foram visualizadas aderidas à parede das células de cria
(Figura 5). Com auxílio de alça microbiológica fragmentos dessa massa fúngica foram
iscados e inoculados em placas de Petri contendo Ágar Sabouraud 20 % (Dicf®) com
antibiótico (Cloranfenicol 250 g/L). As colônias que cresceram foram mantidas nestas
placas de Petri e posteriormente preservadas em água destilada.

25
3.2 Conservação dos fungos isolados Todos os fungos isolados (do ambiente interno da colmeia e das células de cria)
foram preservados em água destilada pelo método de Castellani (RODRIGUES et al.
1992) e em óleo mineral (TERAMOTO et al. 2011).
3.3 Identificação dos fungos isolados
3.3.1 Pela Macro e micromorfologia
Para identificação dos isolados, as colônias fúngicas foram observadas
preliminarmente entre lâmina e lamínula de microscopia contendo água destilada ou
azul de lactofenol (Técnica de fragmentação). Posteriormente, as colônias foram
submetidas ao microcultivo sobre lâmina (Método de Riddell) que consiste em transferir
um fragmento da colônia fúngica para uma placa de petri contendo um bloco de meio
sólido sobre uma lâmina de microscopia onde foi inoculado o fungo e, em seguida, o
bloco foi coberto com uma lamínula e incubado durante sete dias (CORABI-ADELL et
al. 2003). Após este período as estruturas fúngicas foram coradas com azul de
lactofenol e submetidas à análise microscópica em microscópio AxiosKop 40 (Zeiss,
Germany). Para identificação dos fungos foram utilizadas as chaves de identificação de
(BANERTT e HUNTER 1972) e (SEIFERT et al. 2011).
Massa fúngica
Figura 5. Vista geral de um disco de cria de Melipona seminigra com destaque para massa
fúngica presente na parede interna da célula de cria contendo ovo.

26
3.3.2 Por analise molecular - sequência da região ITS1-5.8S-ITS2 do rDNA
Após as análises morfológicas, os gêneros identificados foram submetidos à
confirmação por meio do sequenciamento da região ITS (Internal Transcribed Spacer)
do rDNA (ITS1 - 5.8S - ITS2). Foram selecionadas quatro colônias dos gêneros que
possuíam maior número de isolados, para os gêneros que não atingiram o número de
quatro isolados foram utilizadas todas as amostras isoladas.
O procedimento de extração de DNA fúngico foi realizado de acordo com a
descrição de (FERRER et al. 2011) com modificações. Os fungos selecionados para as
análises moleculares foram cultivados em placas de Petri contendo BDA durante 10
dias a temperatura ambiente. Após esse período as amostras foram transferidas para
microtubos (2 mL) contendo 500 µL de tampão de lise e 5 µL de β-mercaptoetanol. Os
microtubos foram então submetidos a congelamento -20º C overnight, e em seguida
aquecidos a 85º C por 15 minutos e agitados vigorosamente em vórtex por 30 segundos,
sendo incubados a 56º C overnight. Em seguida foram adicionados 500 µL
fenol:clorofórmio:álcool isoamílico, seguindo para agitação no vórtex para a suspensão
homogeneizar. A suspensão foi centrifugada à 143.500 rpm durante 15 minutos e o
sobrenadante retirado e transferido para outro microtubo (1,5 mL). Foram adicionados
500 µL de álcool isopropílico (isopropanol) ao sobrenadante, a mistura foi
homogeneizada e incubada a -20º C overnight, para haver a precipitação do DNA. Com
o DNA precipitado foi realizada outra centrifugação a 13.500 rpm por 15 minutos e
duas lavagens com álcool 70 %, sendo suspendido novamente em 100 µL do tampão de
eluição.
Para amplificação gênica da região ITS1-5.8S-ITS2 do rDNA, foi realizada a
Reação em Cadeia da Polimerase – PCR usando-se os pares iniciadores Its-1 (5´-
TCCGTAGGTGAACCTGCGG-3') e Its-4 (5´-TCCTCCGCTTATTGATATGC-3') (WHITE
et al. 1990) nas seguintes condições: o volume final de cada reação foi de 30 µL, foram
adicionados tampão 1X, MgCl2 5mM, dNTPs 200 µM, primers 1,0µM, Taq DNA
polimerase 1,5 U/µL e DNA fúngico 20ng, o volume da reação foi completado com
água ultrapura. As amostras foram levadas ao termociclador para desnaturação inicial a
uma temperatura de 94 ºC durante 5 minutos. Os parâmetros de PCR foram: 40 ciclos
de desnaturação do DNA a 94 ºC por 30 segundos, anelamento dos primers a 53 ºC por
30 segundos e extensão a 72 ºC durante 1 minutos; terminando com uma extensão final
a 72 ºC durante 10 minutos, baixando a temperatura para 20 ºC para concluir a PCR. A
confirmação do rendimento e qualidade da amplificação, foi verificada por eletroforese

27
em gel de agarose a 1.0 %, em tampão TBE 1X, acrescido de 10 µL/100 mL de
GelRed®, o resultado da amplificação foi visualizado em fotodocumentador sob luz
ultravioleta.
Os produtos da PCR foram purificados usando-se uma solução de
polietilenoglicol-PEG de acordo com o descrito por (DUNN e BLATTNER 1987). Em
seguida os fragmentos amplificados foram submetidos a reação de sequenciamento pelo
método de terminação de cadeia usando-se o kit Big Dye®
Terminator Cycle Sequencing
Standart Version 3.1 (Applied Biosystems). A leitura das sequências foi feita no
sequenciador ABI 3130 Genetic Analyzer (Applied Biosystems). Os fragmentos
sequenciados foram analisados e editados no programa Bioedit. A qualidade das
sequências obtidas foi verificada com o programa Geneious R10. As sequencias obtidas
foram comparadas com a base de dados existentes no GenBank e a identidade dos
transcritos foi confirmada por meio da comparação dessas sequências com as
sequências depositadas no GenBank do NCBI, por meio da ferramenta Blastn
(http://blast.ncbi.nlm.nih.gov/Blast.cgi).
3.4 Análise estatística
3.4.1. Diversidade de fungos obtidos por espécies, casta e período sazonal
Índice de diversidade de Shannon-Weaver
Neste estudo o conceito de táxon foi utilizado para determinar um agrupamento
de indivíduos morfologicamente ou filogeneticamente semelhantes. A frequência e a
diversidade de fungos foram calculadas para cada uma das espécies, castas e período
sazonal (estação chuvosa e estação seca).
Para caracterizar a diversidade da comunidade de fungos foi empregado o índice
de diversidade de Shannon-Weaver (H') (KREBS 1989). Este índice determina a
heterogeneidade de espécies, quanto maior o valor, mais heterogênea é a comunidade,
significando que é maior a sua diversidade. O índice de Shannon-Weaver foi calculado
segundo a fórmula:
H' = - (Pi) * (log2 Pi) onde, Pin = ni
/N
Onde: ni= número de exemplares da i-ésima espécie; N é o número total de exemplares
da amostra.

28
Índice de Equitabilidade
O índice de equitabilidade (E) foi calculado para representar a uniformidade do
número de indivíduos por táxon (MAGURRAN 2004). Este índice tende para 0 quando
um táxon é dominante na comunidade, e tende para 1 quando todos os táxons
apresentam a mesma abundância na comunidade. O índice foi calculado pela seguinte
fórmula:
E = H'/ 1n S
Onde: H' é o índice de Shannon- Weaver baseado no número de indivíduos e S é o
número de táxons presentes na amostra.
Índice de Sorensen (Similaridade)
A similaridade foi calculada pelo Índice de Sorensen (S') avaliando as diferenças
entre os fungos isolados de M. interrupta e M. seminigra e entre as estações sazonais
(BEGON et al. 2006) e entre as castas. Os valores variáveis foram 0 (nenhuma
semelhança) e 1 (similaridade absoluta). A fórmula para o cálculo foi:
S' = ______
Onde: “a” é o número total de táxons coletadas na condição A, “b” é o número
total de táxons coletadas na condição B e “c” é o número de táxons iguais em ambas às
condições.
Índice de Riqueza
É o número de táxons identificados por espécies (M. interrupta X M. seminigra),
por estação (chuvosa X seca), por ambiente (ambiente interno da colmeia x ambiente da
célula de cria), por casta (célula de rainha X de operaria).
2c
a + b

29
4. RESULTADOS
Em ambas as espécies foi possível o isolamento e identificação de diversidade
fúngica presente em todos os ambientes (ambiente interno, célula de larva, célula de
pupa rainha ou operária) tanto na estação chuvosa quanto na estação seca.
4.1 Isolamento dos fungos das colmeias de Melipona interrupta e M. seminigra
Observou-se maior número total de isolados fúngicos em colmeias de M.
interrupta que em M. seminigra. Quanto aos ambientes, a maior quantidade de fungos
isolados foi registrada em células de cria contendo pupas para ambas as espécies
(Tabela 2).
Tabela 2. Número (n) e frequência relativa (%) dos fungos isolados dos discos de cria,
células de cria e do ambiente interno das colmeias de Melipona interrupta e M.
seminigra.
Quanto aos ambientes, a maior quantidade de fungos isolados foi registrada em
células de operárias para ambas as espécies, sendo que nas células de operarias de M.
interrupta isolou-se quase o dobro de isolados que de M. seminigra (Figura 6).
Espécie de
Abelha
Ambiente
interno
(UFC) %
Disco de cria
(Pupa)
(UFC) %
Células de cria
(Larva)
(UFC) %
Total
(UFC) %
M. interrupta
Colmeia 1 27 6,1 162 14,9 1 11,1 190 12,3
Colmeia 2 24 5,4 3 0,3 - - 27 1,8
Colmeia 3 42 9,5 12 1,1 - - 54 3,5
Colmeia 4 87 19,6 466 42,8 - - 553 35,9
Colmeia 5 12 2,7 14 1,3 - - 26 1,7
Total 192 43,2 657 60,3 1 11,1 850 55,1
M. seminigra
Colmeia 1 69 15,5 364 33,4 5 55,6 438 28,4
Colmeia 2 55 12,4 29 2,7 - - 84 5,4
Colmeia 3 61 13,7 19 1,7 2 22,2 82 5,3
Colmeia 4 44 10,0 16 1,5 - - 60 3,9
Colmeia 5 23 5,2 4 0,4 1 11,1 28 1,82
Total 252 56,8 432 39,7 8 88,9 692 44,9
Total Geral 444 100 1.089 100 9 100 1.542 100
30
Figura 6. Fungos cultiváveis isolados (UFC-unidade formadora de colônia) das células
de cria com pupas de operaria ou rainha em colmeias de Melipona interrupta e M.
seminigra no Meliponário GPA-INPA, Manaus-AM.
Registrou-se que a temperatura interna média das colmeias de M. interrupta
(30,5 ºC na estação chuvosa e de 29 ºC na estação seca) foi equivalente às de M.
seminigra (30 ºC e 28 ºC nas estações chuvosa e seca, respectivamente).
De acordo com o INMET (http://www.inmet.gov.br/portal/), de fato, os períodos
das coletas de dezembro/2015 a março/2016 (chuvoso) e agosto a outubro/2016 (seco)
corresponderam a períodos em que houve maior e menor precipitação em Manaus-AM
nos anos de 2015 e 2016 (Figura 7).

31
Figura 7. Precipitação pluvial em Manaus – AM durante os meses de coleta de
amostras fúngicas em colmeias de Melipona interrupta e M. seminigra. Estação
chuvosa: dezembro/2015 e março/2016; estação seca: agosto, setembro e outubro/2016.
Quanto ao período climático foi verificado que, para ambas as espécies, o maior
número de isolados ocorreu no período chuvoso (Figura 8).
Figura 8. Abundância de fungos isolados em colmeias de Melipona interrupta e M.
seminigra nas estações seca e chuvosa no Meliponário GPA-INPA, Manaus-AM.
36,80%
63,20%
Melipona interrupta
Seca Chuvosa
21,10%
78,90%
Melipona seminigra
Seca Chuvosa

32
4.2 Identificação morfológica dos fungos isolados
A análise morfológica permitiu separar os 1.542 fungos isolados (UFC) de
acordo com as características morfológicas. Inicialmente foram separados em fungos
filamentosos (904 isolados) e leveduriformes (638 isolados).
Dentre os fungos filamentosos, foi possível identificar como base na macro e
micromorfologia três gêneros, não sendo possível identificar todos os fungos
filamentosos com base nas características morfológicas. Os gêneros identificados pela
macro e micromorfologia foram Aspergillus spp.; Penicillium sp. e Cladosporium sp..
Foi possível ainda diferenciar duas espécies do gênero Aspergillus sendo
inferidas como A. niger e A. ochraceus (Figura 9 e 10). A. niger apresentou uma colônia
negra com a cabeça conidial globosa de conídios negros; enquanto A. ochraceus
apresentou uma colônia amarela com cabeça conidial globosa de conídios amarelos. Os
demais morfotipos do gênero Aspergillus foram agrupados como Aspergillus sp. (T1),
Aspergillus sp. (T2) e Aspergillus sp. (T3) (Figura 11, 12 e 13).
Figura 9. Aspergillus niger isolados das colmeias de Melipona interrupta e M.
seminigra do Meliponário GPA-INPA/Manaus-AM.

33
Figura 10. Aspergillus ochraceus isolado das colmeias de Melipona interrupta e M.
seminigra do Meliponário GPA-INPA/Manaus-AM.
Figura 11. Aspergillus sp. (T1) isolado das colmeias de Melipona interrupta e M.
seminigra do Meliponário GPA-INPA, Manaus-AM.

34
Figura 12. Aspergillus nomius (T2) isolado de M. interrupta e M. seminigra do
Meliponário GPA-INPA, Manaus-AM.
Figura 13. Aspergillus fumigatus (T3) isolado de Melipona interrupta e M. seminigra
do Meliponário GPA-INPA, Manaus-AM.

35
O gênero identificado como Cladosporium apresentou cor característica única
comum para todas as colônias isoladas, sendo colônia verde escuro, com hifas e
conídios (Figura 14).
O gênero Penicillium apresentou colônia verde clara, conidióforo em forma de
pincel. A única variação entre os isolados foi que algumas amostras produziram
pigmento vermelho no meio de cultura, no entanto, não foram diferenciadas em
morfotipos (Figura 15).
Figura 14. Cladosporium cladosporioides isolado das colmeias de Melipona
interrupta e M. seminigra do Meliponário GPA-INPA, Manaus-AM.
Figura 15. Penicillium citrinum isolado de colmeias de Melipona interrupta e M.
seminigra do Meliponário GPA-INPA, Manaus-AM.
36
Das células de cria na fase de larva foram isolados nove fungos filamentosos,
todos com a mesma característica morfológica, hialinos, hifas contínuas e com forte
pigmentação vermelha no meio de cultura (Figura 16). Estes fungos não produziram
estruturas para caracterização e diferenciação micromorfológica que pudesse determinar
o gênero.
As leveduras não foram identificadas por morfologia, apenas agrupadas como
leveduras para posterior identificação molecular. A maioria apresentou-se como
colônias brancas com exceção de um morfotipo que possuía um tom alaranjado (Figura
17).
Figura 17. Leveduras isoladas das células de cria de Melipona interrupta e M.
seminigra do Meliponário GPA-INPA, Manaus-AM.
Figura 16. Monascus ruber isolado de células de cria de Melipona interrupta e M.
seminigra do Meliponário GPA-INPA, Manaus-AM.

37
Os fungos que não apresentaram estruturas de reprodução foram agrupados
como não identificados, para serem investigados pela análise molecular.
4.3 Identificação molecular dos fungos isolados
4.3.1 Amplificação da região ITS1-5.8S-ITS2 do rDNA
O perfil eletroforético do produto amplificado da região ITS1-5.8S-ITS2 do
rDNA confirmou a amplificação positiva da região alvo pela presença de uma banda
com aproximadamente 600 pares de bases, conforme esperado (Figura 18).
4.3.2 Sequenciamento da região ITS1-5.8S-ITS2 do rDNA
O sequenciamento das amostras teve como prioridade os fungos
morfologicamente identificados em nível de gênero para confirmação deste e pretensa
discriminação dos mesmos em nível de espécies, incluindo as leveduras. Algumas
amostras dos fungos não identificados pela morfologia (n=28) também foram
submetidas à identificação molecular.
Com base nas sequencias obtidas para a região ITS1-5.8S-ITS2 do rDNA,
foram identificadas cinco espécies de fungos filamentosos Aspergillus fumigatus,
Aspergillus nomius, Cladosporium cladosporioides, Penicillium citrinum, Monascus
ruber e cinco leveduras, Candida apicola, Candida etchellsii, Candida versatilis,
Cystobasidium minutum e Derbaryomyces hansenii que é o teleomorfo de Candida
famata, com base na identificação in silico comparativamente as sequencias existentes
no GenBank (Tabela 3).
600 pb
Figura 18. Perfil de amplificação da região Its-1 e Its-2 de fungos isolados de colmeias
de Melipona interrupta (amostras 1 a 3) e M. seminigra (amostras 4 a 5) do
Meliponário GPA-INPA, Manaus-AM. C+ = controle positivo. C- = controle negativo
sem DNA.

38
Tabela 3. Fungos isolados de colmeias de Melipona interrupta e M. seminigra no
Meliponario GPA-INPA, Manaus-AM, identificados por análise molecular da região
ITS1-5.8S-ITS2 do rDNA comparados in silico a sequencias do GenBank.
Isolado Morfologia Espécie Identificada/ Acesso
GenBank
Identidade
(%)
61 – R (L) 3 Filamentoso Aspergillus fumigatus (acesso KY926853.1) 99
172 (An) 5 Filamentoso Aspergillus nomius (acesso KJ634091.1) 100
61 – O (L) 3 Filamentoso Cladosporium cladosporioides (acesso KX960612.1) 100
53 (An) 3 Filamentoso Penicillium citrinum (acesso LT558892.1) 99
C – 01 Filamentoso Monascus ruber (acesso KY635850.1) 100
61 – R (L) 2 Levedura Candida apícola (acesso EU926479.1) 99
31E – O (L) 3 Levedura Candida etchellsii (acesso KY102074.1) 99
172 – R (L) 1 Levedura Candida versatilis (acesso KY102502.1) 99
172 – O (L) 1 Levedura Cystobasidium minutum (acesso KT329195.1) 100
31E – R (L) 1 Levedura Debaryomyces hansenii
(Candida famata) (acesso KY103209.1)
99
Todos os gêneros identificados pelas estruturas morfológicas foram submetidos
à análise molecular, porém não foi possível corroborar os resultados para A. niger e A.
ochraceus. Os dados de sequenciamento agruparam as sequencias entre os fungos
leveduriformes, porém a análise da macro e micromorfologia destes isolados claramente
comprovam que eles não se encaixam nesta identidade. Assim manteve-se o resultado
da análise morfológica para posterior reanalise molecular.
As amostras agrupadas pela morfologia como Aspergillus sp. (T2) foram
identificadas como Aspergillus nomius, Aspergillus sp. (T3) como Aspergillus
fumigatus e o grupo Aspergillus sp. (T1) sem discriminação da espécie.
O fungo isolado das células de cria na fase de larva não pôde ser identificado
pelas estruturas morfológicas, foi, portanto, identificado pela análise molecular como
Monascus ruber (Tabela 3).
Para o gênero Penicillium as sequencias indicaram apenas uma espécie
Penicillium citrinum, assim como para o gênero Cladosporium tendo apenas a espécie
Cladosporium cladosporioides identificada (Tabela 3).
As leveduras brancas foram identificadas como Candida apicola, Candida
etchellsii, Candida versatilis e Candida famata. As leveduras alaranjadas foram
identificadas como Cystobasidium minutum (Tabela 3).
O grupo dos fungos não identificados é um grupo diversificado refere-se aos
fungos que não puderam ser identificados pela macro e micromorfologia. Algumas

39
amostras deste grupo foram submetidas à análise molecular, porém não houve sucesso
na amplificação do rDNA. Repetições destas análises serão realizadas posteriormente.
4.4 Índices de diversidade fúngica
Uma vez que o esforço amostral foi similar para a investigação dos isolados nas
espécies, foi possível realizar a comparação dos índices de Heterogeneidade (H' -
(Shannon-Weaver), Similaridade (S' – Sorensen) e Riqueza (R) (Tabela 4).
O índice de riqueza (R) que representa a quantidade de táxons identificados foi o
mesmo para ambas as espécies de Melíponas. No entanto, em M. interrupta, Candida
etchellsii apareceu exclusivamente em células de operaria. Em M. seminigra registrou-
se Aspergillus ochraceus apenas no ambiente interno das colmeias, Cystobasidium
minutum somente em células de operárias e Aspergillus nomius unicamente em células
de rainhas.
O índice de diversidade (H') entre as espécies de abelhas com relação à origem
dos isolados na colmeia (AI – ambiente interno, L- célula de cria com larva, R – célula
de pupa de rainha, O – célula de pupa de operária) verificou que a maior diversidade foi
obtida em M. seminigra, indicando que apesar de terem sido obtidos menos isolados a
comunidade de fungos foi mais heterogênea em M. seminigra quando comparado com
M. interrupta (Tabela 4).
O índice de Equitabilidade (E) demonstrou que houve maior predominância de
táxons em M. interrupta que em M. seminigra (Tabela 4).
O índice de similaridade de Sorensen (S') calculado entre as comunidades de
fungos identificados das colmeias de ambas as espécies de abelhas indicou uma
similaridade acima da média entre as comunidades, indicando que mais da metade dos
táxons é semelhante (Tabela 4).
Tabela 4. Número e frequência relativa dos fungos identificados e não identificados
isolados do ambiente interno (AI) das colmeias, das células de cria com larva (L) e das
células de cria com pupas de rainha (R) ou operária (O) de Melipona interrupta e M.
seminigra no Meliponário GPA-INPA, Manaus-AM.
Classe/Gêneros M. interrupta
M. seminigra
Total (%) AI L R O
AI L R O
Aspergillus sp. (T1) 7 - - 109 48 - 31 - 195 (12,6)
Aspergillus nomius 2 - - - 12 - 64 - 78 (5,1)
Aspergillus fumigatus - - 7 1 - - - 6 14 (0,9)

40
Aspergillus niger 132 - 100 3
72 - 8 43 358 (23,2)
Aspergillus ochraceus - - - -
24 - - - 24 (1,6)
Penicillium citrinum 1 - 6 46
1 - 2 - 56 (3,6)
Cladosporium cladosporioides - - 2 3
- - - 2 7 (0,5)
Monascus ruber - 1 - - - 8 - - 9 (0,6)
Cystobasidium minutum - - - - - - - 258 258 (16,7)
Candida apicola - - 11 2 - - - - 13 (0,8)
Candida etchellsii - - - 364 - - - - 364 (23,6)
Candida versatilis - - - - - - 2 - 2 (0,1)
Candida famata - - 1 - - - - - 1 (0,06)
Não identificados 50 - 2 -
95 - 13 3 163 (10,6)
Total 192 1 129 528
252 8 120 312 1.542 (100)
Colônias Isoladas 850 692
Riqueza (R) 10 10
Shannon-Weaver (H') 1,3553 1,5307
Equitabilidade (E) 0,38779 0,46214
Sorensen (S') 0,7
Quando analisados os índices da comunidade com relação a sazonalidade
verificou-se que em M. interrupta o índice de diversidade (H') e de equitabilidade (E)
foram menores na estação chuvosa em relação à seca, representando que na estação seca
houve mais heterogeneidade de fungos enquanto na chuvosa alguns táxons foram mais
dominantes na comunidade de fungos (Tabela 5).
Assim, o índice de similaridade de Sorensen para M. interrupta referente às
estações sazonais foi um índice médio representando que pelo menos metade da
comunidade dos fungos de cada estação é semelhante (Tabela 5).
Tabela 5. Número e frequência relativa dos fungos isolados nas colmeias (C1 a C5) de
Melipona interrupta nas estações chuvosa e seca no Meliponário GPA-INPA, Manaus-
AM.
Espécies Estação Chuvosa Estação Seca
C1 C2 C3 C4 C5 C1 C2 C3 C4 C5
Aspergillus sp. (T1) - 1 - 3 3 108 1 - - -
Aspergillus nomius 1 1 - - - - - - - -
Aspergillus fumigatus - - - - - - - 5 - 3
Aspergillus niger 16 4 3 115 - 6 12 24 53 2
Penicillium citrinum - 1 - - - 52 - - - -
Cladosporium cladosporioides - - - - - 1 - 4 - -

41
Monascus ruber - - - - - 1 - - - -
Candida apicola - - 3 - - - - - - 10
Candida etchellsii - - - 364 - - - - - -
Candida famata - - - 1 - - - - - -
Não identificados 2 - - 17 2 3 7 15 - 6
Total por Colmeia 19 7 6 500 5 171 20 48 53 21
Total 537 313
Riqueza (R) 7 7
Shannon-Weaver (H’) 0,7184 1,3587
Equitabilidade (E) 0,293021249 0,555875934
Sorensen (S’) 0,5714
Em M. seminigra o índice de diversidade (H') demonstrou que a maior
diversidade ocorreu na estação chuvosa. O índice de equitabilidade (E) para a estação
chuvosa indicou que ocorreram táxons com maior abundancia na comunidade quando
comparado com a estação seca onde a equitabilidade tendeu a uma abundancia mais
uniforme na comunidade de fungos (Tabela 6).
Tabela 6. Número e frequência relativa dos fungos isolados nas colmeias (C1 a C5) de
Melipona seminigra nas estações chuvosa e seca no Meliponário GPA-INPA, Manaus-
AM.
Espécies Estação Chuvosa Estação Seca
C1 C2 C3 C4 C5 C1 C2 C3 C4 C5
Aspergillus sp. (T1) 4 27 7 2 2 1 36 - - -
Aspergillus nomius 68 8 - - - - - - - -
Aspergillus fumigatus - - - - - - - - 6 -
Aspergillus niger 87 - 9 3 - 14 8 2 - -
Aspergillus ochraceus - - 24 - - - - - - -
Penicillium citrinum - 2 - - 1 - - - - -
Cladosporium cladosporioides - - 2 - - - - - - -
Monascus ruber - - - - - 5 - 2 - 1
Cystobasidium minutum 258 - - - - - - - - -
Candida versatilis - - - 2 - - - - - -
Não identificados - 3 18 10 9 1 - 18 37 15
Total por Colmeia 417 40 60 17 12 21 44 22 43 16
Total 546 146
Riqueza (R) 8 4
Shannon-Weaver (H’) 1,3727 1,1540
Equitabilidade (E) 0,493248812 0,792712745
Sorensen (S’) 0,3333

42
A similaridade de Sorensen em M. seminigra com relação às comunidades de
fungos isolados nas estações sazonais foi abaixo da média estabelecida, o que indica
pouca semelhança entre as comunidades de fungos de cada estação (Tabela 6).
O índice de diversidade (H') para as castas de M. interrupta foi de 0,8076 em
rainhas e de 0,8865 em operárias, o que indica que a diversidade de fungos foi
praticamente igual entre as castas. A equitabilidade (E) calculada foi de 0,37375 em
rainhas e de 0,34666 em operárias, indicando que para as duas castas houve um táxon
mais dominante. O índice de similaridade de Sorensen (S') foi de 0,769203, indicando
que a comunidade de fungos foi bastante similar entre as castas.
Em M. seminigra o índice de diversidade (H') para as castas foi de 1,0090 em
rainhas e de 0,5342 em operárias indicando maior diversidade de fungos nas células de
rainhas. A equitabilidade (E) foi de 0,54857 em rainhas e de 0,42652 em operárias, o
que demonstra que não ocorreram táxons dominantes na comunidade de fungos. A
similaridade de Sorensen (S') foi de 0,222, o que indicou uma baixa similaridade entre
as comunidades de fungos das castas.
As leveduras foram isoladas somente no interior das células de cria com
indivíduos na fase de pupa sendo que Candida apicola ocorreu nas duas estações
sazonais enquanto Candida famata, Candida etchellsii, Candida versatilis e
Cystobasidium minutum ocorreram somente na estação chuvosa.
Quando comparados os resultados com relação às castas observou-se que as
células de rainhas e de operárias de M. interrupta obtiveram mais táxons que as células
de operárias e rainhas de M. seminigra. Em M. interrupta a levedura Candida famata
foi isolada apenas de células de rainhas, nas células de operárias os táxons particulares
foram Aspergillus sp. (T1) e Candida etchellsii. Em M. seminigra os táxons Aspergillus
sp. (T1), A. nomius, Penicillium citrinum e Candida versatilis ocorreram apenas em
células de operárias, e nas células de rainhas os táxons exclusivos foram A. fumigatus,
Cladosporium cladosporioides e Cystobasidium minutum.
Dos 1.542 fungos isolados 850 foram isolados de M. interrupta tendo as
leveduras como o grupo mais abundante, o maior índice individual foi de Candida
etchellsii com percentual de (23,6 %). Dos táxons identificados a maior diversidade foi
de fungos filamentosos com sete táxons identificados sendo A. niger o mais frequente
com percentual de (15,2 %). Os táxons Candida apicola, Candida etchellsii e Candida
famata ocorreram somente em M. interrupta (Figura 19).

43
De M. seminigra foram isolados 692 (UFC), dos táxons identificados o maior
percentual foi de Cystobasidium minutum (16,7 %), seguido por A. niger com (8,0 %).
Assim como em M. interrupta o número de táxons entre os fungos filamentosos foi
maior, sendo oito táxons de fungos filamentosos e dois de leveduras. Os táxons que
ocorrem somente em M. seminigra foram A. ochraceus, Cystobasidium minutum e
Candida versatilis (Figura 19).

44
7,5
0,13 0,52
15,2
3,4
0,3
0,06
0,8
23,6
3,4
5,1 4,9
0,4
8,0
1,6
0,2 0,1 0,5
16,7
7,2
0
5
10
15
20
25
Fre
quên
cia
(%)
M. interrupta M. seminigra
Figura 19. Frequência (%) dos fungos isolados de Melipona interrupta e M. seminigra nas estações seca e chuvosa no Meliponário GPA-INPA,
Manaus-AM.

45
5. DISCUSSÃO
Este estudo representa o primeiro registro, por identificação molecular, da
diversidade de fungos associados à colmeias de abelhas sem ferrão amazônicas. Neste
estudo foram encontradas 13 espécies de fungos associados às colmeias de M.
interrupta e M. seminigra. Este achado acrescenta conhecimento e abre caminhos para
novas investigações sobre a microbiota fúngica cultivável associada e suas possíveis
relações com abelhas sem ferrão. O fungo filamentoso Monascus ruber e as leveduras
Candida apicola, Candida etchellsii, Candida versatilis, Candida famata e
Cystobasidium minutum estão sendo registrados pela primeira vez associados às
colmeias de M. interrupta e M. seminigra. Os demais gêneros já foram observados
nestas espécies em outros Meliponários de Manaus e Iranduba/AM (SOUZA 2014).
As metodologias de isolamento de fungos nas colmeias de M. interrupta e M.
seminigra aplicadas no presente trabalho resultaram em um número similar de isolados
para ambas as espécies. Possivelmente porque as características dos locais de coleta nas
colmeias eram similares uma vez que as espécies apresentam a mesma estrutura de
ninho (discos de cria e potes de alimento), comportamento social similar além do fato
das colmeias das duas espécies serem mantidas em um mesmo local (Meliponário) o
que possibilita a sobreposição de pasto meliponicola durante o forrageamento, além da
dispersão aérea de fungos no ambiente do Meliponário.
A técnica de sedimentação de esporos (MINAMI 2003) estabelece a exposição
das placas com o meio de cultura ao ambiente da coleta com a tampa retirada, no
entanto, no interior das colmeias as placas não puderam ser expostas abertamente
porque as abelhas invadiam e degradavam o meio de cultura, para contornar essa
desvantagem as tampas das placas foram mantidas e furadas para possibilitar a entrada
do ar, o que solucionou satisfatoriamente essa dificuldade. Assim a técnica de
sedimentação de esporos com essa adaptação é útil para se isolar fungos do interior das
colmeias de M. interrupta e M. seminigra. A identificação dos fungos pela análise das
estruturas morfológicas não foi eficiente para se determinar as espécies dos fungos
isolados, a análise morfológica possibilitou a identificação somente em nível de gênero.
Essa limitação no uso de técnicas de análise morfológica para identificação de fungos
foi observada por estudos recentes com taxonomia de fungos (CORTEZ et al. 2016;
VISAGIE et al. 2016), abrindo espaço para as técnicas moleculares que mostram-se
atualmente mais eficientes para a determinação taxonômica e filogenética dos fungos.

46
Os iniciadores ITS-1e ITS-4 utilizados para a amplificação das regiões ITS1-
5.8S e ITS2 do rDNA obtiveram bom êxito para a amplificação das regiões de interesse,
o que possibilitou a confirmação a nível de espécie dos gêneros identificados pela
análise morfológica incluindo outros que não foram identificados pela morfologia, as
leveduras foram identificadas somente pela análise molecular. Os primers ITS-1 e ITS-2
têm sido utilizados universalmente para identificação de fungos filamentosos e
leveduras e se mostraram eficientes para esta finalidade (WHITE et al. 1990;
SANDERS et al. 1995; LUPATINI et al. 2008). Alguns trabalhos com leveduras
associadas a abelhas sem ferrão no Brasil realizados por ROSA et al. (1999, 2007,
2013); TEIXEIRA et al. (2003); PIMENTEL et al. (2005); ROSA e LACHANCE
(2005) e DANIEL et al. (2013), não utilizaram estes primers para identificação
molecular e caracterização filogenética de leveduras associadas a abelhas sem ferrão,
foram utilizados primers para amplificação nas regiões D1/D2 da subunidade maior do
rDNA.
O fungo A. nomius foi isolado do ambiente interno das colmeias de ambas as
espécies de abelhas e apenas em M. seminigra foi isolado dos discos de cria na fase de
pupa em células de rainhas. Este fungo é um importante produtor de aflatoxinas B e G
no Brasil (OLSEN et al. 2008), tendo sido isolado de abelhas como visitantes florais
tornando-se potenciais vetores deste fungo para as plantas, o que oferece risco de
contaminação as plantações polinizadas por abelhas vetores de esporos deste fungo
(MASSI et al. 2015).
O gênero Aspergillus foi o de maior frequência em ambas às colmeias, para o
qual foram identificadas cinco espécies. As demais espécies isoladas foram Penicillium
citrinum, Cladosporium cladosporioides, Monascus ruber, Cystobasidium minutum,
Candida apicola, Candida etchellsii, Candida versatilis, Candida famata. SOUZA
(2014) investigou abelhas sem ferrão em dois Meliponários (municípios de Manaus e
Iranduba) identificando 16 gêneros e 12 espécies de fungos filamentosos isolados do
corpo das abelhas. Três destas espécies também apareceram no interior das colmeias de
M. interrupta e M. seminigra aqui analisadas: em ambas Penicillium citrinum e
Cladosporium cladosporioides, e Aspergillus ochraceus apenas em M. seminigra.
Leveduras têm sido comumente encontradas associadas a diferentes ordens de
insetos, oferendo benefícios como proteção contra patógenos e a liberação de enzimas
extracelulares que atuam na metabolização de substancias de interesse para os insetos
(PROMNUAN et al. 2009). Portanto, é compreensivo o isolamento de leveduras

47
associados células de cria das abelhas como foi o caso de Candida apicola que já foi
observada associada a abelhas sem ferrão no Tocantins, incluindo a espécie M.
scutellaris (NETO 2017).
A levedura Cystobasidium minutum é um fungo que apresenta características do
gênero Rhodotorula, no entanto, Rhodotorula é um grupo que inclui muitas espécies de
relevância clinica, e filogeneticamente está subdividida em três classes:
Microbotryomycetes, Cystobasidiomycetes e Exobasidiomycetes. O táxon isolado foi
geneticamente classificado como Cystobasidium minutum da classe
Cystobasidiomycetes (YURKOV et al. 2015).
O registro de ocorrência exclusa de Candida etchellsii apenas em células de
operaria para M. interrupta e de Aspergillus ochraceus no ambiente interno, de
Cystobasidium minutum em células de operárias e Aspergillus nomius em células de
rainhas para M. seminigra requer melhor investigação sobre a possível relação desta
microbiota com estes ambientes e talvez com as castas. Até o momento ainda não foram
realizados estudos que comprovem se os táxon isolados exclusivamente das células de
rainhas e de operárias tem alguma participação na diferenciação das castas no que se
refere ao sistema genético alimentar descrito por KERR, (1950), mas tanto os táxon do
gênero Aspergillus e as leveduras identificadas possuem potencial para fermentação e
produção de metabólitos secundários (PROMNUAN et al. 2009; FOLEY 2013; VEGA-
ALVARADO et al. 2015).
As leveduras podem apresentar uma relação mais específica com as abelhas. No
Brasil existem registros de leveduras associadas a abelhas sem ferrão. Starmerella
neotropipicalis foi identificada associada ao pólen coletado por M. quinquefasciata
(DANIEL et al. 2013). Outros estudos identificaram Candida floris isolada de um
adulto de Trigona sp.; Starmerella meliponinorum foi identificada associada a M.
quadrifasciata, M. rufiventris, Tetratrigona angustula e Trigona fulviventris
(TEIXEIRA et al. 2003; ROSA et al. 2007, 2013). Muitas leveduras isoladas de abelhas
são conhecidamente associadas às flores sendo, por essa razão, cogitada a possibilidade
de existir uma relação benéfica entre os fungos e as abelhas (ROSA et al. 2013). Existe
ainda a possibilidade de que microrganismos realizem a fermentação do alimento no
intestino da abelha. O estudo de ENGEL et al. (2012) em Apis mellifera comprovou que
a relação evolutiva entre os microrganismos presentes no intestino dessas abelhas é
antiga e que o estilo de vida das abelhas sociais pode ter favorecido a diversificação de
microrganismos específicos nas abelhas.

48
Os índices de diversidade e similaridade apresentaram pequenas variações, os
menores índices de diversidade foram observados entre as castas e os mais elevados
entre as espécies de abelhas. Não há estudos que descrevam a diversidade e similaridade
de fungos associados às colmeias de abelhas sem ferrão amazônicas. Os estudos com
Meliponíneos restringem-se a fungos aderidos ao corpo das abelhas e ao mel. FERRAZ
et al.(2008) investigaram fungos aderidos ao corpo de M. subnitida, dos achados pelos
autores três gêneros em comum com os encontrados em M. interrupta e M. seminigra
podem ser citados Aspergillus sp., Penicillium sp. e Cladosporium sp., incluindo a
espécie A. niger.
A sazonalidade não influenciou a diversidade e frequência dos fungos isolados
em M. interrupta. Embora a sazonalidade tenha influência sobre a dispersão dos fungos
(CORTEZ et al. 2016), a equivalente diversidade destes organismos nas colmeias
analisadas em ambas as estações ocorreu possivelmente porque as coletas foram
realizadas em ambientes fechados onde alguns gêneros comuns no ambiente
atmosférico da região amazônica não foram isolados (FURTADO e FERRARONI
1998; FONSECA e CONCEIÇÃO 1977). O gênero Aspergillus que foi o mais frequente
é comumente encontrado em diversos ambientes (FERNÁNDEZ et al. 2014).
Apenas em M. seminigra a quantidade dos fungos isolados na estação chuvosa
foi superior em relação à estação seca. Uma provável explicação pode ser consequência
do habito mais generalista de forrageamento apresentado por M. seminigra
comparativamente a M. interrupta (FERREIRA e ABSY 2015), o que pode possibilitar
maior amplitude de contato com microrganismos. Outra possível razão para a pouca
diferença foi a influencia do fenômeno El Niño (2015-2016) que aumentou a
temperatura e diminuiu a precipitação tornando uma parte dos meses chuvosos muito
semelhantes à estação seca (CPTEC/INPE 2016a) (CPTEC/INPE 2016b). Mesmo
assim, pela experiência da equipe do GPA é notório que as colmeias de M. seminigra
são mais úmidas que as de M. interrupta (CARVALHO-ZILSE, inf. p.).
O único táxon identificado das células de cria na fase de larva foi o fungo
Monascus ruber. Este táxon foi isolado tanto de M. interrupta como em M. seminigra.
A identificação deste fungo corrobora o estudo de MENEZES et al. (2015) onde o
gênero Monascus foi isolado de células de cria na fase de larva em M. scutellaris. O fato
de apenas este fungo ter sido isolado nesta fase do desenvolvimento das abelhas pode
ser resultado da ação inibitória do fungo impossibilitando o crescimento de outros
microrganismos, e que também serve como defesa para as larvas contra patógenos.

49
Ainda, há necessidade de alimentação com este fungo para iniciar de fato a alimentação
larval provisionada pelas operarias à célula de cria.
Os fungos identificados neste estudo são fungos de importância clinica como as
Candida spp. que podem causar candidíases e os filamentosos que podem causar
manifestações alérgicas como Aspergillus spp. (OLIVEIRA e BORGES-PALUCH
2015). Também é conhecido o potencial de Monascus ruber que não é patogênico e se
destaca na produção de pigmentos para a culinária, substâncias antibióticas e enzimas;
no Oriente é utilizado para fazer vinhos vermelhos Shoa-Hsing e queijo vermelho de
soja (SÁ 2005). As leveduras são uma fonte para a produção de bioativos de interesse
industrial, como biosurfactantes, antibióticos e nos processos de metabolização
(CLEMENTE et al. 2015). Por essas razões recomenda-se o uso de EPIs nas atividades
de Meliponicultura e manuseio dos produtos das abelhas durante a colheita.
6. CONCLUSÃO
Este estudo comprovou a existência de fungos associados às colmeias, e às
castas de M. interrupta e M. seminigra. Nas células de rainhas e operárias foi observado
à presença de leveduras e fungos filamentosos, as leveduras estão restritas as células de
cria. Nas células de cria na fase de larva de M. interrupta e M. seminigra ocorre o fungo
Monascus ruber, abrindo a possibilidade de uma potencial relação de mutualismo entre
as abelhas e este fungo. Este é o primeiro registro que utiliza os iniciadores ITS-1 e ITS-
2 para identificar a diversidade fúngica em colmeias de M. interrupta e M. seminigra.
A diversidade de fungos nas duas espécies de abelhas foi similar, da mesma
forma com relação às estações sazonais, havendo uma diminuição na diversidade dos
associados às castas tanto de M. interrupta como de M. seminigra.
Com relação às hipóteses alternativas pode-se afirma que existe relação com da
microbiota fúngica cultivável em castas de abelhas M. interrupta e M. seminigra, pois
as leveduras ocorreram apenas nas células de cria com rinhas e operárias. Não existe
relação da microbiota fúngica em colmeias de abelhas M. interrupta e M. seminigra.
Existe relação da microbiota fúngica cultivável em colmeias de abelhas M. interrupta e
M. seminigra e a sazonalidade, pois em M. seminigra foi observado maior diversidade
na estação chuvosa em relação à estação seca o que não ocorreu em M. interrupta.
Este estudo contribuiu para o aumenta do conhecimento da diversidade de
fungos associados à M. interrupta, M. seminigra e ao grupo de abelhas sem ferrão

50
usadas em Meliponicultura. A identificação de táxons que podem causar manifestações
alérgicas e micoses chama a atenção para o risco de contaminação; o conhecimento de
que as colmeias podem ser uma fonte de fungos com potencial biotecnológico, incentiva
a busca de novas linhagens de fungos com o potencial de fermentação e produção de
biosurfactantes e pigmentos. Os fungos filamentosos e as leveduras que foram
identificados são fungos que apresentam potencial biotecnológico e podem ser testados
para esta finalidade.
O estudo da microbiota fúngica cultivável das abelhas deve ampliar a
investigação para uma melhor compreensão da relação desta microbiota com as abelhas
M. interrupta e M. seminigra. A identificação dos fungos presentes no corpo das
abelhas, incluindo a investigação da microbiota intestinal, pode apresentar informações
relevantes sobre a relação dos fungos com essas abelhas, relacionando tais informações
com o pólen e o mel. A origem do pólen deve ser investigada para verificar se há fungos
comumente presentes nas fontes de pólen e se estes, ao serem transportados para as
colmeias, atuam de forma benéfica paras as abelhas.
REFERÊNCIAS BIBLIOGRÁFICAS
Banertt, H. L.; Hunter, B. B. 1972. Illustrated genera of imperfect fungi. 3o ed.
Minneapolis: Burgess Publishing Company.
Begon, M.; Townsed, C. R.; Harper, J. L. 2006. Ecology from Individuals to
Ecosystems. Fourth. Australia: Blackwell publishing.
Bonetti, A. M. 2008. Genética da determinação de castas em Melipona: expressão de
genes casta-específicos. Universidade de São Paulo.
Brasileiro, B. T. R. V et al. 2004. Pcr-Fingerprinting Based on Pcr Markers. Brazilian
Journal of Microbiology, 35: 205–210.
Brito, B. B. P. et al. 2013. Parâmetros biométricos e reprodutivos de colônias em
gerações de Melipona quadrifasciata. Archivos de zootecnia, 62(238): 366.
Carvalho-Zilse, G. A.; Nunes-Silva, C. G. 2012. Threats to the Stingless Bees in the
Brazilian Amazon: How to Deal with Scarce Biological Data and an Increasing Rate of
Destruction. In: Florio, RM (Ed), Bees: Biology, Threats and Colonies. In: Nova
Science Publishers, p. 147–168.
Castagnino, G. L. B. et al. 2006. Doença Cria Giz Ascosphaera apis (Maassen ex
Claussen) Olive & Spiltoir em abelhas Apis mellifera L . na Depressão Central do Rio
Grande do Sul. Ciência Rural, 36(6): 1909–1911.

51
Clemente, L. et al. 2015. Bioprospecção de antibióticos produzidos por fungos da
caatinga. Evidência, Joaçaba, 15(1): 37–56.
Corabi-Adell, C.; Lucon, C. M. M.; Ichikawa, A. 2003. Uma técnica simples para
observação microscópica de Trichoderma spp. (hypocreales) em cultura de lâminas.
Arq. Inst. Biol., 70: 92–95.
Cortez, A. C. A. et al. 2016. A comparison of the freshwater fungal community during
the non-rainy and rainy seasons in a small black water lake in Amazonas , Brazil.
Journal of Food, Agriculture & Environment, 14(2): 156–161.
CPTEC/INPE. 2016a. Fenômeno El Niño inicia sua fase de declínio. INFOCLIMA, 1:
1–3.
CPTEC/INPE. 2016b. Voltam, chuvas um padrão típico de anos com atuação do
fenômeno El Niño. INFICLIMA, 2: 1–3.
Danforth, B. 2007. Bees - a primer. Current Biology, 17(5): 156–161.
Daniel, H. et al. 2013. Starmerella neotropicalis f . a ., sp . nov ., a yeast species found
in bees and pollen. International Journal of Systematic and Evolutionary Microbiology,
63: 3896–3903.
Dunn, I. S.; Blattner, F. R. 1987. Charons 36 to 40: multi enzyme, high capacity,
recombination deficient replacement vectors with polylinkers and polystuffers. Nucleic
Acids Research, 15(6): 2677–2698.
Engel, P.; Martinson, V. G.; Moran, N. A. 2012. Functional diversity within the simple
gut microbiota of the honey bee. Proceedings of the National Academy of Sciences, 109
(27): 11002–11007.
Engel, P.; Moran, N. A. 2013. Functional and evolutionary insights into the simple yet
specific gut microbiota of the honey bee from metagenomic analysis. Gut microbes,
4(1): 60–65.
Fernández, M. et al. 2014. Especies de Aspergillus en ambientes hospitalarios con
pacientes pediátricos en estado crítico. Revista Iberoamericana de Micologia, 31(3):
176–181.
Ferraz, R. E. et al. 2008. Microbiota Fúngica de Melipona subnitida Ducke
(Hymenoptera : Apidae)”, Neotropical Entomology, 37(3), p. 345–346.
Ferreira, M. G.; Absy, M. L. 2015. Pollen niche and trophic interactions between
colonies of Melipona (Michmelia) seminigra merrillae and Melipona (Melikerria)
interrupta (Apidae: Meliponini) reared in floodplains in the Central Amazon.
Arthropod-Plant Interactions, 9(3): 263–279.
Ferrer, A.; Miller, A. N.; Shearer, C. A. 2011. Minutisphaera and Natipusilla: two new
genera of freshwater Dothideomycetes. Mycologia, 103(2): 411–23.

52
Foley, K. 2013. The Ecology and Evolution of Aspergillus spp . Fungal Parasites in
Honey Bees. University of Leeds.
Fonseca, M. D. P. 2016. Estudo da diversidade e atividade larvicida de Agaricomycestes
lognolíticos (Basidiomycota) no Estado do Amazonas.
Fonseca, O. J. M.; Conceição, L. A. 1977. Fungos anemófilos de Manaus. Acta
Amazonica, 7: 497–501.
Furtado, M. S. S.; Ferraroni, J. J. 1998. Airborne fungi in hospitals of the city of
Manaus, Amazonas, Brazil. Science and Culture, 34(12): 1642–1647.
González, G. et al. (2005) “Occurrence of mycotoxin producing fungi in bee pollen”,
International Journal of Food Microbiology, 105(1), p. 1–9. doi:
10.1016/j.ijfoodmicro.2005.05.001.
Kerr, W. E. 1950. Genetic determination of castes in the genus Melipona. Genetics,
35(2): 143.
Kerr, W. E. et al. 2001. Aspectos pouco mencionados da biodiversidade amazônica.
Parcerias Estratégicas, 80: 45–60.
Kerr, W. E.; Carvalho-Zilse, G. A.; Nascimento, V. A. 1996. Abelha Uruçu: Biologia,
Manejo e Conservação. Paracatu, MG: Fundação Acangaú.
Krebs, C. J. 1989. Ecological Methodology. 2ª Ed, New York.
Lacaz, C. D. S.; Port, E.; Martins, J. E. C. 2002. Tratado de Micologia Médica. 9o ed,
Savier, São Paulo.
Lupatini, M.; Mello, A. H.; Antoniolli, Z. I. 2008. Caracterização do DNA ribossômico
do isolado de Scleroderma UFSMSc1 de Eucalytus grandis W. Hill Ex-Maiden R. Bras.
Ciências. Solo, 32(1): 2677–2682.
Magurran, A. E. 2004. Measuring Biological Diversity. Oxford: Blackwell publishing
company.
Marques-Souza, A. C. et al. 2002. Características morfológicas e bioquímicas do pólen
coletado por cinco espécies de meliponíneos da Amazônia central. Acta Amazonica,
32(2): 217–229.
Massi, F. P. et al. 2015. Identification of Aspergillus nomius in bees visiting Brazil nut
flowers. Microbes and Environments, 30(3): 273–275.
Menezes, C. et al. 2007. Alimentação larval de Melipona (Hymenoptera, Apidae):
estudo individual das células de cria. Bioscience Journal, 23: 70–75.
Menezes, C. et al. 2015. A Brazilian social bee must cultivate fungus to survive.
Current Biology. Elsevier Ltd, 25(2): 1–5.
Menezes, J. P. et al. 2010. Variabilidade genética na região ITS do rDNA de isolados de

53
Trichoderma spp. (Biocontrolador) e Fusarium oxysporum f. sp. Chrysanthemi. Search,
34(1): 132–139.
Menezes P. S. R. 2014. The stingless bee fauna in Brazil (Hymenoptera: Apidae).
Sociobiology, 61(4): 348–354.
Minami, P. S. 2003. Micologia: métodos laboratoriais de diagnóstico das micoses. 1o
ed. Barueri: Manole.
Neto, D. J. C. 2017. Leveduras vetorizadas por abelhas sem ferrão em áreas de cerrado
no estado do Tocantins, Brasil. Dissertação de mestrado, p. 54.
O’Connor, D. J. et al. 2014. Atmospheric concentrations of Alternaria, Cladosporium,
Ganoderma and Didymella spores monitored in Cork (Ireland) and Worcester (England)
during the summer of 2010. Aerobiologia, 30(4): 397–411.
Oliveira, K. A. M.; Ribeiro, L. S.; Oliveira, G. V. 2013. Caracterização microbiológica,
físico-química e microscópica de Abelhas Canudo (Scaptotrigona depilis) e Jataí
(Tetragonisca angustula). Revista Brasileira de Produtos Agroindustriais, 15(66): 239–
248.
Oliveira, L. D. C.; Borges-Paluch, L. R. 2015. Alergias respiratórias: Uma revisão dos
principais fungos anemófilos e fatores desencadeantes. Rev Baiana de Saúde Pública,
39(2): 426–441.
Olsen, M. et al. 2008. Aspergillus nomius, an important aflatoxin producer in Brazil
nuts?. World Mycotoxin Journal, 1: 123–126.
Pimentel, M. R. C. et al. 2005. Candida riodocensis and Candida cellae, two new yeast
species from the Starmerella clade associated with solitary bees in the Atlantic rain
forest of Brazil. FEMS Yeast Research, 5: 875–879.
Porter, T. M.; Golding, G. B. 2012. Factors that affect large subunit ribosomal DNA
amplicon sequencing studies of fungal communities: Classification method, primer
choice, and error. PLoS ONE, 7(4): 1–12.
Promnuan, Y.; Kudo, T.; Chantawannakul, P. 2009. Actinomycetes isolated from
beehives in Thailand. World Journal of Microbiology and Biotechnology, 25(9): 1685–
1689.
Ribeiro, M. M. R. et al. 2012. Diversity of fungi associated with Atta bisphaerica
(Hymenoptera: Formicidae): the activity of Aspergillus ochraceus and Beauveria
bassiana. Psyche, p. 6.
Rodrigues, E. G.; Lírio, V. S.; Lacaz, C. D. S. 1992. Preservacao fungos interesse
médico em água destilada. Revista Inst. Med. Tropical, p. 159–165.
Rosa, C. A. et al. 1999. Candida batistae , a new yeast species associated with solitary
digger nesting bees in Brazil. Mycologia, 91(3): 426–433.

54
Rosa, C. A. et al. 2007. Candida flosculorum sp. nov. and Candida floris sp. nov., two
yeast species associated with tropical flowers. International Journal of Systematic and
Evolutionary Microbiology. 57: 2970–2974.
Rosa, C. A. et al. 2013. Yeast communities associated with stingless bees. FEMS Yeast
Research, 4: 271–275.
Rosa, C. A.; Lachance, M. C. 2005. Zygosaccharomyces machadoi sp . n ., a yeast
species isolated from a nest of the stingless bee Tetragonisca angustula. Lundiana, 6:
27–29.
Sá, R. A. 2005. Estudo da produção de metabólitos secundários por Monascus ruber
CCT 3802 em cultivo submerso. Florianópolis, SC: Universidade Federal de Santa
Catarina, p. 50–71.
Sanders, I. R. et al. 1995. Identification of ribosomal DNA polymorphisms among and
within spores of the Glomales: application to studies on the genetic diversity of
arbuscular mycorrhizal fungal communities. New Phytologist, 130(3): 419–427.
Seifert, K. A. et al. 2011. The Genera of Hyphomycetes. Netherlands: CBS-KNAW.
Shutler, D. et al. 2014. Honey bee Apis mellifera parasites in the absence of Nosema
ceranae fungi and Varroa destructor mites. PLoS ONE, 9(6): 1–8.
Silva, E. A. R. et al. 2012. Seleção de isolados de fungos entomopatogênicos para o
controle de Leptopharsa hevea (Hemiptera: Heteroptera, Tingidae). Arq. Inst. Biol.,
79(4): 549–556.
Souza, J. R. S. (2014) “Diversidade fúngica associada a abelhas sem ferrão (Melipona
spp.) em Meliponário na cidade de Manaus e Iranduba, Amazonas, Brasil.”,
Dissertação, UFAM, p. 65.
Stuart, R. M.; Lamas, C.; Pimentel, I. C. 2004. Trigona sp. como visitante floral e vetor
de esporos fúngicos para goiabeira (Psidium guajava L. – Myrtaceae). Ecology, 73(3):
959–970.
Teixeira, A. C. P. et al. 2003. Starmerella meliponinorum sp . nov ., a novel
ascomycetous yeast species associated with stingless bees. International Journal of
Systematic and Evolutionary Microbiology, 53: 339–343.
Teramoto, A.; Martins, M. C.; Cunha, M. G. 2011. Avaliação de métodos para
preservação de isolados de Corynespora cassiicola (Berk. & M. A. Curtis) C.T. Wei.
Pesquisa Agropecuária Tropical, 41(2): 296–298.
Vega-Alvarado, L. et al. 2015. High-quality draft genome sequence of Candida apicola
NRRL Y-50540. Genome Announcements, 3(3): 4–5.
Visagie, C. M. et al. 2016. A taxonomic review of Penicillium species producing
conidiophores with solitary phialides, classified in section Torulomyces. Persoonia:

55
Molecular Phylogeny and Evolution of Fungi, 36: 134–155.
White, T. J. et al. 1990. Amplification and direct sequencing of fungal ribosomal RNA
genes for phylogenetics. In: PCR Protocols: A Guide to Methods and Applications.
New York: Academic Press, p. 315–322.
Yurkov, A. M. et al. 2015. Two yeast species Cystobasidium psychroaquaticum f.a. sp.
nov. and Cystobasidium rietchieii f.a. sp. nov. isolated from natural environments, and
the transfer of Rhodotorula minuta clade members to the genus Cystobasidium. Antonie
van Leeuwenhoek, 107: 173–185.


















