
MORCEGOS NEOTROPICAIS - sbe.com.br Neotropicais.pdf · Pertencem à ordem Chiroptera, ... na...
85
MORCEGOS NEOTROPICAIS BIOLOGIA, ECOLOGIA E TÉCNICAS DE COLETA ROBERTO LEONAN MORIM NOVAES
Transcript of MORCEGOS NEOTROPICAIS - sbe.com.br Neotropicais.pdf · Pertencem à ordem Chiroptera, ... na...

MORCEGOS NEOTROPICAIS BIOLOGIA, ECOLOGIA E TÉCNICAS DE COLETA
ROBERTO LEONAN MORIM NOVAES

Apostila básica
ROBERTO LEONAN MORIM NOVAES
Rio de Janeiro
2008
MORCEGOS NEOTROPICAIS BIOLOGIA, ECOLOGIA E TÉCNICAS DE COLETA
Esta é uma obra independente e foi desenvolvida para fins educacionais, não sendo autorizada a sua
reprodução e comercialização. A distribuição digital é feita gratuitamente pelo autor e Projeto Pró-Morcegos.

_______________________________________________________
QUEM SOMOS?
O Projeto Pró-Morcegos é uma organização de ação ambiental que surgiu em
2004 no Rio de Janeiro com a finalidade de promover pesquisas sobre a biologia,
conservação e ecologia dos morcegos do Brasil. Além do desenvolvimento de pesquisas
científicas o Projeto oferece cursos, palestras e aulas em universidades, empresas,
órgãos públicos e organizações privadas. Este projeto é independente e sem fins
lucrativos, que visa apenas conscientizar e informar a população sobre a importância
de se preservar esses animais para se manter um ecossistema equilibrado.
COORDENAÇÃO
Luis Fernando Menezes Jr.
Ana Carolina Duarte
PESQUISA E AÇÃO AMBIENTAL
Roberto Leonan Morim Novaes
COLABORAÇÃO
Ana Cristina Façanha
Carla Clarissa Nobre de Oliveira
Carolinne Isabella Dias Gomes
Flávio Augusto Pereira Mello
Mariana Vieira Pinto Aguiar
PROJETO PRÓ-MORCEGOS Tel.: (21) 3624 0628
E-mail: [email protected] www.promorcegos.org
PROJETO PRÓ-MORCEGOS

Índice
I Aspectos gerais sobre a Ordem Chiroptera...................................................................1 Introdução.................................................................................................................2 Classificação..............................................................................................................2 Nota biológica............................................................................................................3 Distribuição...............................................................................................................4 Morfologia..................................................................................................................5 Ecolocalizador............................................................................................................7 Taxonomia.................................................................................................................8 Evolução....................................................................................................................9 Referências bibliográficas.......................................................................................11 II Introdução aos morcegos neotropicais........................................................................15 Referências bibliográficas.......................................................................................17 III Hábitos alimentares e importância ecológica...........................................................18 Introdução...............................................................................................................19 Frugivoria e dispersão de sementes.......................................................................19 Nectarivoria e polinização......................................................................................21 Insetivoria e controle de pragas.............................................................................22 Piscivoria e controle de populações........................................................................23 Carnivoria e controle de populações......................................................................24 Hematofagia............................................................................................................25 Referências bibliográficas.......................................................................................26 IV Raiva em quirópteros..................................................................................................33 Introdução...............................................................................................................34 Patogenia.................................................................................................................34 Sintomatologia em mamíferos................................................................................35 Sintomatologia em morcegos hematófagos............................................................35 Sintomatologia em morcegos não hematófagos.....................................................36 Referências bibliográficas.......................................................................................37 V Abrigos diurnos............................................................................................................39 Introdução...............................................................................................................40 Abrigos naturais......................................................................................................40 Abrigos artificiais....................................................................................................42 Referências bibliográficas.......................................................................................45 VI Reprodução..................................................................................................................49 Introdução...............................................................................................................50 Monoestria...............................................................................................................51 Poliestria.................................................................................................................51 Gestação..................................................................................................................52 Referências bibliográficas.......................................................................................53 VII Conservação...............................................................................................................56 Introdução...............................................................................................................57 Conservação dos morcegos do Brasil......................................................................58 Refêrencias bibliográficas.......................................................................................60 VIII Técnicas de coleta....................................................................................................62 Introdução...............................................................................................................63 Métodos de captura.................................................................................................63 Triagem...................................................................................................................69 Referências bibliográficas.......................................................................................74

I
ASPECTOS GERAIS SOBRE A ORDEM CHIROPTERA

Reino: Metazoa
Filo: Chordata
Classe: Mammalia
Ordem: Chiroptera
Subordem: Megachiroptera
Subordem: Microchiroptera
INTRODUÇÃO
Os morcegos são animais noturnos e os únicos mamíferos capazes de
realizar um vôo mecânico verdadeiro, sendo isso possível graças às grandes
adaptações que estes animais sofreram nos membros anteriores (HILL &
SMITH, 1988). Pertencem à ordem Chiroptera, que provém do grego “Cheir”
(mão) e “Pteron” (asa), que indica que suas asas são, na verdade, uma mão
altamente modificada (PERACCHI et al., 2006). Chiroptera é a segunda maior
ordem entre os mamíferos, possuindo 1113 espécies (SIMMONS, 2005), sendo
ultrapassada apenas pela ordem dos roedores (Rodentia).
CLASSIFICAÇÃO
Embora alguns autores considerem que a Ordem Chiroptera possua uma
origem monofilética (SIMMONS, 1994; HUTCHEON & KIRSCH, 2006), os
quirópteros, como também são chamados, são divididos em duas subordens
distintas, Megachiroptera e Microchiroptera. A primeira é encontrada
exclusivamente no Velho Mundo, e compreende uma única família
(Pteropodidae) com 42 gêneros e 185 espécies, enquanto a segunda está
amplamente distribuída por todo o globo, envolvendo 17 famílias, 157 gêneros
e 928 espécies (SIMMONS, 2005).
Classificação taxonômica dos Chiroptera
2

NOTA BIOLÓGICA
Megachiroptera
Dentre os Megachiroptera estão os maiores morcegos conhecidos, sendo o
maior de todos o Pteropus vampyrus (raposa-voadora). Este morcego, nativo de
algumas ilhas da Ásia e Oceania, pode apresentar uma envergadura de 1,7
metros e ter até 1,5 quilos (KUNZ & JONES, 2000). Apesar do grande tamanho,
estes morcegos são frugívoros, podendo apresentar também hábitos
nectarívoros em menor freqüência. Além disso, não possuem ecolocalizador e
guiam-se unicamente pela visão, que na maior parte das espécies é bem
desenvolvida (NOWAK, 1994). A maioria das espécies de Megachiroptera tem
olhos grandes e focinho alongado próprio para o consumo de frutos e néctar
(PERACCHI et al., 2006).
Microchiroptera
Nesta subordem se encontra o menor de todos os morcegos,
Craseonycteris thonglongyai. Conhecido como morcego-abelha, habita as
florestas do sudeste da Ásia, sendo encontrado principalmente na Tailândia.
Pesa 2g e tem uma envergadura de asas de cerca de 10cm (HILL & SMITH,
1981). Os Microchiroptera apresentam um tamanho relativamente pequeno se
comparados aos grandes morcegos frugívoros do Velho Mundo. Neste grupo se
encontra a maior diversidade de morcegos e a maior variedade de hábitos
alimentares (PERACCHI et al., 2006). Estão amplamente distribuídos por todo
globo, habitando todos os continentes. Os Microchiroptera apresentam um
sistema de ecolocalização que auxilia na locomoção e na procura e captura de
alimentos (ARITA & FENTON, 1997; GRIFFIN, 2001). Mesmo possuindo um
complexo ecolocalizador os microquirópteros não descartam o uso da visão. A
maior parte das espécies possui um campo visual bem desenvolvido e bastante
usual.
3

DISTRIBUIÇÃO
Morcegos estão amplamente distribuídos por todo o globo e ocupam quase
todos os ambientes terrestres, sendo encontrados em todos os continentes,
exceto em algumas ilhas do Pacífico e regiões muito frias como as calotas
polares ou regiões de altitude extrema (HILL & SMITH, 1988). Os
Megachiroptera têm ocorrência restrita e são encontrados na Ásia, África,
Oceania e diversas ilhas dos oceanos Índico e Pacífico. Por outro lado os
Microchiroptera são distribuídos por todos os continentes e diversas faixas
insulares (DRUMMOND, 2004).
Aqui no Brasil encontramos apenas os morcegos da subordem
Microchiroptera com 168 espécies distribuídas em nove famílias (REIS et al.,
2007; MIRANDA et al., 2007). São elas: Emballonuridae, Vespertilionidae,
Molossidae, Mormoopidae, Noctilionidae, Furipteridae, Thyropteridae,
Natalidae e Phyllostomidae. Segundo JONES (2002), a maior diversidade e
abundância de quirópteros está localizado nos trópicos, sendo ainda maior nos
neotrópicos, e por isso essas regiões recebem o nome de Hotspots.
Esquematização de gradientes de cores aplicado ao grau de riqueza de morcegos. Branco: Nenhuma espécie por 500km² / Vermelho: 120 espécies por 500km².
(original de JONES, 2002)
4

MORFOLOGIA
Os quirópteros apresentam uma das mais ricas variedades de formas
entre os mamíferos, apresentando fisionomias e morfologia bem diferenciadas.
A maior parte dos morcegos tem tamanho corporal pequeno, apresenta o corpo
coberto por pêlos, sendo estes muito finos, podendo ser mais ou menos densos
ou alongados, dependendo da espécie. A coloração pode variar bastante, e,
embora predomine um padrão oscilante entre o marrom-escuro e pardo
acinzentado, há morcegos amarelos, completamente brancos, vermelhos,
pretos, alaranjados e até mesmo multicoloridos (GRAHAM, 1994)
As asas dos morcegos são, na verdade, uma adaptação especial dos
membros anteriores e são formados pelos braços, mãos e pelo prolongamento
da pele (chamada de patágio). Os dedos das mãos são constituídos
basicamente pelos mesmos ossos de uma mão humana, porém sofreram um
notável alongamento, mais precisamente no segundo, terceiro, quarto e quinto
dedos. Os ossos do antebraço (ulna e rádio) também sofreram um alongamento
e juntamente com o braço (úmero) são responsáveis por pouco menos da
metade da extensão das asas no seu comprimento. O conjunto ósseo das asas é
leve e a maior parte dos músculos se concentra no antebraço do animal. Desde
os ombros até os membros posteriores há um prolongamento da pele, o
patágio, composto por uma fina camada de tecido conectivo envolvido
superficialmente por uma também fina pele elástica e bastante irrigada. É
esse conjunto que dá forma à asa do morcego. Apenas o polegar fica fora do
prolongamento do patágio. Algumas espécies possuem uma membrana
prolongada entre os membros posteriores chamada de uropatágio.
Os membros posteriores dos morcegos sofreram diversas adaptações, e
com isso não se tornaram capazes de sustentar o próprio corpo como em outros
mamíferos (NEUWEILER, 2000). Em decorrência da adaptação ao vôo, as pernas
dos morcegos não possuem uma estrutura óssea que favoreça a locomoção
íntegra. A curvatura do joelho possui uma rotação inversa a dos demais
mamíferos, de tal forma que a rótula se encontra voltada para trás em relação
ao corpo. Ao mesmo tempo, tornou-se vantajosa a postura de repouso com a
cabeça voltada para baixo, possibilitando ao morcego abandonar o ponto de
5

pouso e se deixar cair para iniciar o vôo, sem gasto de energia com impulsos
para cima (NEUWEILER, 2000).
Uma questão interessante nesse comportamento de repouso é que os
morcegos desenvolveram um mecanismo de contra-peso por meio de um
dispositivo de “trava” automático ativado pelo próprio peso do animal, que faz
com que os tendões dos dedos mantenham estes involuntariamente
tensionados, sem contrações musculares, permitindo ao morcego suportar o
peso do corpo de cabeça para baixo com um gasto bastante reduzido de
energia, o que é bastante vantajoso e oportuno (NEUWEILER, 2000).
A anatomia interna dos morcegos é basicamente igual a dos outros
mamíferos placentados, embora possa haver algumas diferenciações quanto ao
sistema digestório, especialmente estômago e intestino, que são os mais
adaptados a dietas específicas.
Em resposta ao ecolocalizador, alguns Microchiroptera desenvolveram
uma pequena estrutura no rosto denominada de folha nasal. Acredita-se que a
folha nasal atue como um receptor para as ondas emitidas pelo seu biosonar.
Outra estrutura, chamada Trago (tragus), encontrada em todos os
microquirópteros, atua também como um receptor do ecolocalizador. O trago
fica localizado na entrada do pavilhão auricular dos morcegos e constitui-se de
uma protuberância cartilaginosa bem maleável.
Morfologia externa (http://www.morcegolivre.vet.br)
6

ECOLOCALIZADOR
Além da visão, os Microchiroptera utilizam outra ferramenta para
auxiliar no vôo e na procura de alimentos, o ecolocalizador. Existem duas
teorias que abordam o surgimento do ecolocalizador. A primeira defende que
os morcegos teriam desenvolvido a eco-orientação como uma resposta à
captura de insetos na escuridão, quando tais mamíferos começaram a
desenvolver atividade de vôo, necessitando de um mecanismo que auxiliasse
na percepção e perseguição dos insetos predados (NEUWEILER, 2000;
SPEAKMAN, 2001). A segunda teoria, mais difundida no meio científico, defende
que o sistema de ecolocalização teria surgido não como uma adaptação ao
comportamento alimentar, mas como uma adaptação à escuridão do ambiente
cavernícola adotado num segundo estágio da evolução (FENTON, 1992). A
grosso modo, o ecolocalizador constitui-se de ondas de alta freqüência
produzidas por estalidos feitos na laringe dos morcegos. Estas ondas são
emitidas freqüentemente enquanto o morcego voa e, ao se chocar com algum
corpo (objeto, planta ou animal), retornam e são receptadas pelo tragus e pela
folha nasal (nos morcegos que possuem). As ondulações das ondas são
codificadas e transportadas ao cérebro, onde o animal obtém informações sobre
a textura, distância, forma e tamanho do corpo no qual as ondas se chocaram.
A partir destas informações o morcego conseguirá encontrar possíveis presas
para alimentação e o possibilitará desviar de objetos a sua frente enquanto
voa .
Devido ao seu hábito alimentar os morcegos insetívoros são os que
possuem o ecolocalizador mais sensível, já que para capturar insetos em pleno
vôo o morcego necessita de uma grande capacidade para localizá-los
(SCHNITZLER & KALKO, 2001).
Representação do ecolocalizador dos morcegos: envio das ondas, choque com um corpo, retorno e recepção.
(http://www.morcegolivre.vet.br)
7
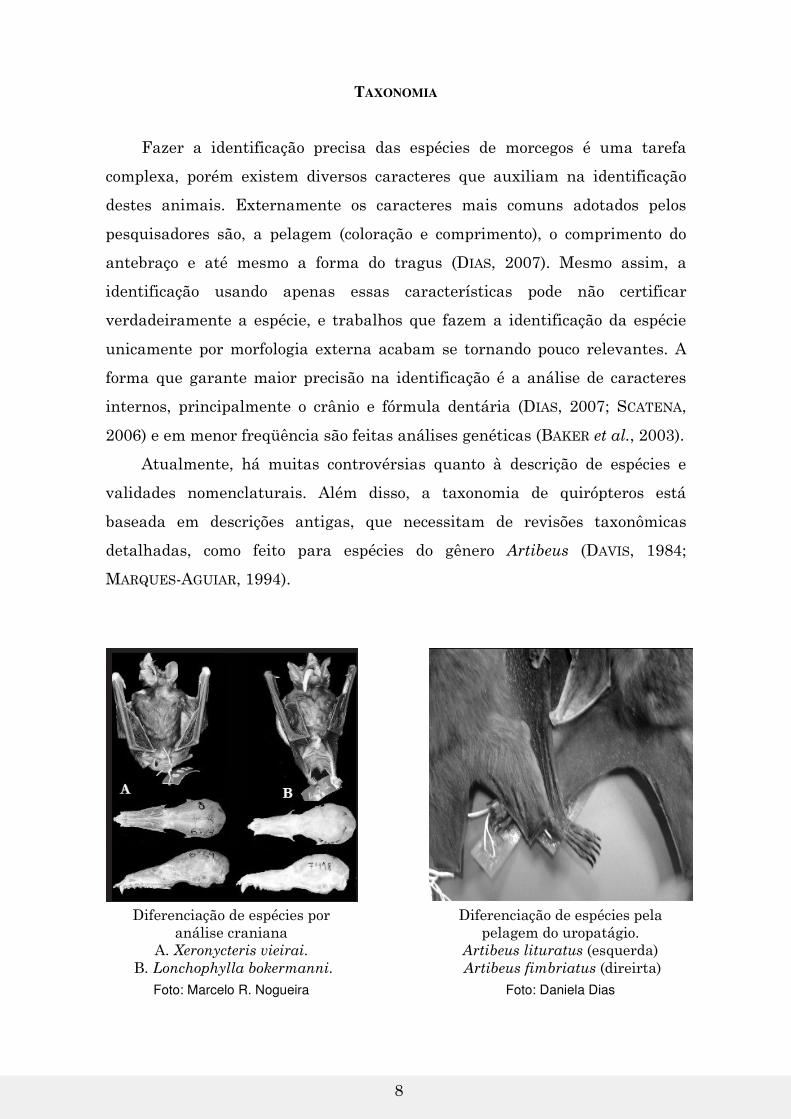
TAXONOMIA
Fazer a identificação precisa das espécies de morcegos é uma tarefa
complexa, porém existem diversos caracteres que auxiliam na identificação
destes animais. Externamente os caracteres mais comuns adotados pelos
pesquisadores são, a pelagem (coloração e comprimento), o comprimento do
antebraço e até mesmo a forma do tragus (DIAS, 2007). Mesmo assim, a
identificação usando apenas essas características pode não certificar
verdadeiramente a espécie, e trabalhos que fazem a identificação da espécie
unicamente por morfologia externa acabam se tornando pouco relevantes. A
forma que garante maior precisão na identificação é a análise de caracteres
internos, principalmente o crânio e fórmula dentária (DIAS, 2007; SCATENA,
2006) e em menor freqüência são feitas análises genéticas (BAKER et al., 2003).
Atualmente, há muitas controvérsias quanto à descrição de espécies e
validades nomenclaturais. Além disso, a taxonomia de quirópteros está
baseada em descrições antigas, que necessitam de revisões taxonômicas
detalhadas, como feito para espécies do gênero Artibeus (DAVIS, 1984;
MARQUES-AGUIAR, 1994).
Diferenciação de espécies por análise craniana
A. Xeronycteris vieirai. B. Lonchophylla bokermanni.
Foto: Marcelo R. Nogueira
Diferenciação de espécies pela pelagem do uropatágio.
Artibeus lituratus (esquerda) Artibeus fimbriatus (direirta)
Foto: Daniela Dias
8

EVOLUÇÃO
A história evolutiva dos morcegos ainda é bastante discutida, existindo
teorias variadas sobre a origem e os percursos evolutivos pelos quais
transitaram os primeiros morfo-morcegos. Os estudos em torno dos registros
fósseis indicam que os morcegos já possuíam um grau de diferenciação dos
outros mamíferos há cerca de 60 milhões de anos, no período Eoceno, quando
os mamíferos se encontravam em processo de transição evolutiva em
diferentes estágios. Há muitos registros fósseis de morcegos (SIMMONS &
GEISLER, 1998; SIMMONS et al., 2008), entretanto, mesmo os mais antigos
fósseis encontrados, datando de estágios diversos do Eoceno, já apresentam
uma estrutura morfológica similar dos morcegos atuais, com presença de uma
estrutura óssea, inclusive dos membros superiores para a formação das asas,
semelhante dos morcegos conhecidos nos dias de hoje (NEUWEILER, 2000;
NOWAK, 1997). Estudos de exemplares fósseis bem conservados mostram que
no período Eoceno os morcegos já apresentavam membrana alar
(HABERSETZER & STORCH, 1987), e detalhes da articulação dos ombros com o
alargamento da escápula para a inserção de músculos de vôo evidenciam que
naquele período os morcegos já realizavam a atividade de vôo batido
(HABERSETZER & STORCH, 1987; NORBERG, 1989). A teoria mais aceita no
meio científico defende o surgimento dos primeiros morcegos a partir do
memento em que começaram a surgir alterações comportamentais no nicho de
forrageamento e hábitat de um pequeno mamífero insetívoro de hábitos
noturnos, terrestre e com tendências arborícolas (HILL & SMITH, 1988; NOWAK,
1997). De acordo com esta teoria, o pequeno mamífero teria gradativamente
transitado para o nicho arborícola. Dessa alteração, modificações adaptativas
foram surgindo e assim, para compensar a habilidade de captura de insetos à
noite, o pequeno animal passaria a ganhar um comportamento saltador. Este
comportamento, pouco a pouco, num longo estágio, transitaria para o
comportamento planador, surgindo daí a tendência ao vôo, com modificações e
adaptações da membrana alar e estrutura óssea, para permitir a captura de
insetos que transitavam próximo aos ramos das árvores onde tais mamíferos
se encontravam (SPEAKMAN, 2001).
9

Além disso, estudos mais aprofundados sobre a anatomia dos morcegos
fizeram surgir uma outra teoria sobre a evolução da ordem Chiroptera,
segundo a qual os morcegos tiveram duas origens evolutivas distintas (origem
polifilética), sugerindo que um grupo teria evoluído a partir dos primatas e o
outro da ordem Insectivora (SPEAKMAN, 2001; PETTIGREW, 1986; NEUWEILER,
2000). No entanto, seja qual for a origem evolutiva, os morcegos são grandes
exemplos do sucesso evolutivo de um grupo adaptado aos diversos tipos de
nichos e ambientes, e sua ampla abrangência geográfica evidencia esse
aspecto. A ordem dos quirópteros está atualmente distribuída por todos os
continentes, em praticamente todos os ecossistemas terrestres e regiões geo-
climáticas. Trata-se de um dos grupos mais representativos dos mamíferos,
não só pelo número de espécies catalogadas, mas também pela abundância de
indivíduos existentes, existindo espécies que formam as maiores colônias de
mamíferos do mundo (NOWAK, 1994).
Representação evolutivas dos morcegos: origem monofilética a partir de um pequeno mamífero arborícola.
(http://www.tparents.org)
Onychonycteris finneyi (SIMMONS et al., 2008)
Paleochiropteryx tupaiodon (SIMMONS & GEISLER, 1998)
10

REFERÊNCIAS BIBLIOGRÁFICAS
ARITA, H.T. & FENTON, M.B. 1997. Fligth and echolocation in the ecology and
evolution of bats. Trends in Ecology and Evolution 12(2): 53-58.
BAKER, R.J., HOOFER, S.R., PORTER, C.A. & VAN DEN BUSSCHE, R.A. 2003.
Diversification among New World Leaf-Nosed Bats: An evolution
hypothesis and classification Inferred from Digenomic Congruence of
DNA sequence. Occasional Papers, Museum of Texas Tech
University 230: i+1-32p.
DAVIS, W.B. 1994. Review of the large-fruit eating bats of the Artibeus
‘lituratus’ complex (Chiroptera, Phyllostomidae) in middle America.
Occasional Papers of the Museum of Texas Tech University 93:
1-16.
DIAS, D. 2007. Quirópteros da Reserva Biológica do Tinguá, Nova
Iguaçu, Rio de Janeiro, Brasil (Mammalia, Chiroptera). Tese de
Doutorado. Universidade Federal Rural do Rio de Janeiro. 108p.
DRUMMOND, S.M. 2004. Morcegos, verdades e mitos. Uma análise acerca
do conhecimento sobre os morcegos na sociedade: ciência,
folclore e cultura. Monografia de graduação. Universidade Estadual
do Sudoeste da Bahia. 109p.
FENTON, M.B. 1992. Bats. New York: Facts on File. 2007p.
GRAHAM, G.L. 1994. Bats of the world. A golden guide from St. Martin‘s
Press. 160p.
GRIFFIN, D.R. 2001. Return to the magic well: echolocation behavior of bats
and responses of insect prey. BioScience 51(7): 555-556.
11

HABERSETZER, J. & STORCH, G. 1987. Klassifikation und funktionelle
Flügelmorphologie paläogener Fledermäuse (Mammalia, Chiroptera).
Courier Forschungsinstitut Senckenberg 91: 117-150.
HILL, J.E. & SMITH, J.D. 1988. Bats: a community perspective. Cambridge
University Press. 167p.
HILL, J.E. & SMITH, S.E. 1981. Craseonycteris thonglongyai. Mammalian
Species 160: 1-4.
HUTCHEON, J.M. & KIRSCH, J.A.W. 2006. A moveable face: deconstructing the
Microchiroptera and a new classification of extant bats. Acta
Chiropterologica 8(1): 1–10.
JONES, K.E. 2002. Chiroptera (Bats). Encyclopedia of Life Sciences.
Macmillan Publishers Ltd., Nature Publishing Group. 5pp.
KUNZ, T.H. & JONES, D.P. 2000. Pteropus vampyrus. Mammalian Species
642: 1-6.
MARQUES-AGUIAR, S.A. 1994. A systematic review of the large species of
Artibeus Leach, 1821 (Mammalia, Chiroptera), with some phylogenetic
inferences. Boletin do Museu Paraense Emílio Goeldi, sér.
Zoologia 10: 3-83.
MIRANDA, J.M.D., AZEVEDO-BARROS, M.F.M. & PASSOS, F.C. 2007. First record
of Histiotus laephotis Thomas (Chiroptera, Vespertilionidae) from
Brazil. Revista Brasileira de Zoologia 24(4): 1188-1191.
NEUWEILER, G. 2000. The biology of bats. New York: Oxford University
Press. 310p.
12

NORBERG, U.M. 1989. Vertebrate flight. Springer-Verlag, Berlin Heidelberg.
123p.
NOWAK, R.M. 1994. Walker’s bats of the world. Introduction por: KUNZ,
T.H. & PIERSON, E.D. (Eds). Jonhs Hopkins University Press. 287p.
NOWAK, R.M. 1997. Walker’s Mammals of the World. Jonhs Hopkins
University Press. 287p.
PERACCHI, A.L., LIMA, I.P., REIS, N.L., NOGUEIRA, M.L. & ORTÊNCIO-FILHO, H.
2006. Ordem Chiroptera. In: REIS, N.R., PERACCHI, A.L., PEDRO, W.A. &
LIMA, I.P (Eds). Mamíferos do Brasil. Editora da Universidade
Estadual de Londrina. p.153-230.
PETTIGREW, J.D. 1986. Are flying foxes really primates? Bat Magazines 3(2):
1-2.
REIS, N.R., PERACCHI, A.L., PEDRO, W.A. & LIMA, I.P. 2007. Morcegos do
Brasil. Editora da Universidade Estadual de Londrina. 253p.
SCATENA, M.P. 2006. Análise molecular em morcegos do gênero Artibeus
Leach, 1821 (Chiroptera, Phyllostomidae) a partir de espécimes
depositados em coleção científica. Dissertação de Mestrado.
Universidade Estadual Paulista. 60p.
SCHNITZLER, H.U. & KALKO, E.K.V. 2001. Echolocation by insect-eating bats.
BioScience 51(7): 557-569.
SIMMONS, N.B. 1994. The case for Chiropteran monophyly. American
Museum Novitates 3103: 54p.
13

SIMMONS, N.B. 2005. Order Chiroptera. In: WILSON, D.E., REEDER, D.M. (Eds).
Mammals Species of the World: a taxonomic and geographic
reference. 3.ed. v.1. Jonhs Hopkins University Press. p.312-529.
SIMMONS, N.B. & GEISLER, J.H. 1998. Phylogenetic relationships of
Icaronycteris, Archaeonycteris, Hassianycteris, and Palaeochiropterix to
extant bat lineages, with comments on the evolution of echolocation and
strategies in Microchiroptera. Bulletin of American Museum of
Natural History 235: 1-182.
SIMMONS, N.B., SEYMOUR, K.L., HABERSETZER, J. & GUNNELL, G.F. 2008.
Primitive Early Eocene bat from Wyoming and the evolution of flight
and echolocation. Nature 451: 818-822.
SPEAKMAN, J.R. 2001. The evolution of flight and acholocation in bats: another
leap in the dark. Mammal Review 31: 111-130.
14

II
INTRODUÇÃO AOS
MORCEGOS NEOTROPICAIS

SOBRE AS FAMÍLIAS NEOTROPICAIS
Nos neotrópicos existe apenas a subordem Microchiroptera, contendo
dez famílias: Emballonuridea, Furipteridae, Molossidae, Momoopidae,
Natalidae, Noctilionidae, Phyllostomidae, Thyropteridae, Vespertilionidae
(SIMMONS, 2005). Destas famílias, seis são endêmicas do Novo Mundo:
Furipteridae, Mormoopidae, Natalidae, Noctilionidae, Phyllostomidae,
Thyropteridae, dentre as quais, estão presente todos os hábitos alimentares da
Ordem Chiroptera.
A fitofagia (ingestão de partes vegetais), por exemplo, é praticada por
grande parte das espécies de Phyllostomidae, sendo esta família a principal
responsável pela dispersão de sementes e polinização de flores, especialmente
em zonas áridas (PERACCHI et al., 2006).
A insetivoria é praticada por espécies de todas as famílias encontradas
em regiões neotropicais. A maior parte das espécies são insetívoras e formam
as maiores colônias de morcegos do planeta, com milhares de indivíduos
(GRAHAM, 1994). A insetivoria é de inestimável importância para o controle de
pragas e populações de insetos nos neotrópicos (LONG, 1996).
A piscivoria, consumo de peixes e artrópodes marinhos, é praticada por
duas famílias, Noctilionidae e Vespertilionidae, e são importantes no controle
de populações de peixes e artrópodes (BORDIGNON, 2006).
A carnivoria e a hematofagia são praticadas por espécies da família
Phyllostomidae. Morcegos carnívoros podem consumir anfíbios, répteis, aves,
pequenos roedores e até mesmo outros morcegos (PERACCHI et al., 2006). Os
morcegos hematófagos pertencem à subfamília Desmondontinae, e são os
principais responsáveis pela disseminação de raiva em herbívoros domésticos
(MAYEN, 2003).
A região neotropical é considerada a maior zona biodiversa do mundo, e
isso também se aplica aos quirópteros. Das dezoito famílias de morcegos
existentes no planeta, nove estão presentes nos neotrópicos, com cerca de 290
espécies (HUTSON et al., 2001).
16

REFERÊNCIAS BIBLIOGRÁFICAS
BORDIGNON, M.O. 2006. Diet of the fishing bat Noctilio leporinus (Linnaeus)
(Mammalia, Chiroptera) in a mangrove area of southern Brazil.
Revista Brasileira de Zoologia 23(1): 256-260.
GRAHAM, G.L. 1994. Bats of the world. A golden guide from St. Martin‘s
Press. 160p.
HUTSON, A.M., MICKLEBURGH, S.P. & RACEY, P. 2001. Microchiropteran
bats: Global status survey and conservation action plan.
IUCN/SSC Chiroptera Specialist Group. IUCN. x+258p.
LONG, R.F. 1996. Bats for insect biocontrol in agriculture. IPM-Practitioner
18(9): 1-6.
MAYEN, F. 2003. Haematophagous bats in Brazil, their role in rabies
transmission, impact on public health, livestock industry and
alternatives to an indiscriminate reduction of bat population. Journal
of Veterinary Medicine B. 50: 469-472.
PERACCHI, A.L., LIMA, I.P., REIS, N.L., NOGUEIRA, M.L. & ORTÊNCIO-FILHO, H.
2006. Ordem Chiroptera. In: REIS, N.R., PERACCHI, A.L., PEDRO, W.A. &
LIMA, I.P (Eds). Mamíferos do Brasil. Editora da Universidade
Estadual de Londrina. p.153-230.
SIMMONS, N.B. 2005. Order Chiroptera. In: WILSON, D.E., REEDER, D.M. (Eds).
Mammals Species of the World: a taxonomic and geographic
reference. 3.ed. v.1. Jonhs Hopkins University Press. p.312-529.
17

III
HÁBITOS ALIMENTARES
E IMPORTÂNCIA ECOLÓGICA

INTRODUÇÃO
Os morcegos podem apresentar hábitos alimentares muito variados, e
essa diversidade de dietas não é encontrada em nenhum outro grupo de
mamíferos. São utilizados como alimento insetos e outros artrópodes, frutos,
sementes, folhas, flores, pólen, néctar, pequenos vertebrados e sangue
(PERACCHI et al., 2006), por isso os quirópteros são considerados animais de
extrema importância para a manutenção do meio ambiente.
A maior parte das espécies de morcego existentes nos neotrópicos é
insetívora, com representantes de todas as famílias encontradas aqui. No
entanto, os morcegos mais abundantes possuem hábitos frugívoros, sendo
esses pertencentes a uma única família, Phyllostomidae (REIS et al., 2007).
FRUGIVORIA E DISPERSÃO DE SEMENTES
Os morcegos frugívoros encontrados aqui no Brasil e em todo o Novo
Mundo pertencem a uma única família, Phyllostomidae. Estes morcegos
podem consumir uma grande variedade de frutos, infrutescências, folhas,
sementes e outras partes vegetais (ZORTÉA, 2007). GARDNER (1977) cita 95
espécies vegetais sendo consumidas por uma única espécie frugívora (Artibeus
jamaicensis). Entre os principais frutos consumidos estão os das famílias
Piperaceae, Solanaceae, Cecropiaceae e Moraceae (MELLO, 2002; PASSOS &
SILVA, 2003; PASSOS et al., 2003; PASSOS & GRACIOLLI, 2004; MELLO, 2006).
Alguns estudos indicam que morcegos têm preferência por frutos macios e
carnosos (PASSOS et al., 2003; NOVAES et al. no prelo), o que pode estar ligado
ao valor nutricional deles (MORRISON, 1980) e ao paladar (HERRERA, 1982).
Por possuir grande mobilidade (BERNARD & FENTON, 2003), e não
destruírem as sementes ingeridas (FLEMING & SOSA, 1994), morcegos
frugívoros são considerados importantes na dispersores de sementes (MELLO
et al., 2008), o que é crucial na dinâmica e regeneração de florestas
(WHITTAKER & JONES, 1994).
Morcegos costumam utilizar trilhas e caminhos abertos no interior das
matas como rota de locomoção, e com isso as sementes que atravessam o tubo
19

digestivo desses morcegos são eliminadas junto com as fezes durante o vôo,
gerando novas plantas (BARRESE, 2005). Os frutos maiores, que possuem
sementes impossíveis de serem ingeridas, acabam sendo carregadas pelos
morcegos para serem comidas em locais longe da planta mãe, e após o
consumo da polpa os morcegos abandonam a semente (NOVAES et al. no prelo).
Por possuir um comportamento voador, morcegos são capazes de ultrapassar
barreiras geográficas difíceis ou impossíveis para outros mamíferos e por isso
conseguem, em pouco tempo, disseminar uma grande quantidade de sementes
em uma vasta área.
Alguns vegetais co-evoluíram com os morcegos (quiropterocoria),
desenvolvendo estruturas especiais e infrutescências atrativas aos morcegos,
facilitando a sua dispersão (BARRESE, 2005). Um exemplo de planta
quiropterocórica são as Piperaceae (especialmente o gênero Piper), que
segundo MELLO (2002), são os principais recursos alimentares de Carollia
perspicillata. Outros exemplos de quiropterocoria e interações específicas são
as plantas Solanaceae (especialmente Solanum) e os morcegos Sturnira lilium
(MELLO, 2006), Moraceae (especialmete Ficus) e Cecropiaceae com espécies dos
gêneros Artibeus, Platyrrhinus e Chiroderma (NOGUEIRA & PERACCHI, 2002;
2003; ZORTÉA, 2007).
Artibeus lituratus Foto: Roberto Novaes
Carollia perspicillata Foto: Roberto Novaes
Ficus (Moraceae) Foto: Marco Mello
Piper (Piperaceae) Foto: Roberto Novaes
Sturnira lilium Foto: Marco Mello
Solanum (Solanaceae) Foto: Marco Mello
Espécies de morcegos freqüentes no Brasil e as principais plantas utilizadas como ali-mento por elas.
20

NECTARIVORIA E POLINIZAÇÃO
Morcegos nectarívoros (ou polinívoros) pertencem à família
Phyllostomidae e são os principais dispersores de pólen, sendo responsáveis
por grande parte da reprodução de diversas espécies de flores (SAZIMA et al.,
1999; SILVA & PERACCHI, 1999). Essas espécies têm especial destaque na
polinização de plantas raras e endêmicas, como Dyssochroma viridiflorum,
solanácea epífita e endêmica da Mata Atlântica (SAZIMA et al., 2003), e
Pticairnia albiflos, bromeliácea rara e endêmica de afloramentos rochosos no
Rio de Janeiro (WENDT et al., 2001). A dispersão de pólen é quase de total
exclusividade dos Chiroptera já que aves, como os beija-flores, têm sua
atividade alimentar restrita à parte do dia, enquanto morcegos têm longa
atividade desde o crepúsculo até o amenhacer. Acredita-se que algumas flores
noturnas co-evoluíram com os morcegos, desenvolvendo estruturas atrativas
especiais e adaptadas à morfologia destes morcegos, adaptação que é chamada
de quiropterofilia (TSCHAPKA et al., 1999; STROO, 2000; FENSTER et al., 2004).
As flores que fazem parte da síndrome da quiropterofilia normalmente
apresentam cores mais claras, odor atrativo, e nectários desenvolvidos que
facilitem o acesso dos morcegos ao seu interior. Esse mutualismo é mais
comum do que parece, já que mesmo morcegos de dieta predominantemente
frugívora ou até mesmo onívora acabam por utilizar as flores como fontes de
nutrição (SILVA & PERACCHI, 1999). Parte das flores nortunas, principalmente
de zonas áridas, dependem quase que exclusivamente dos morcegos para sua
reprodução (SORIANO et al., 2000), sendo os morcegos nectarívoros os
principais agentes polinizadores de cactáceas presentes nestes ambientes
(PETIT, 1995; NASSAR et al., 1997).
Glossophaga soricina Foto: Roberto Novaes
Bombacaceae Foto: Roberto Novaes
Espécie de morcego com ampla distribuição no Brasil, sendo muito comum em áreas florestais e ambientes urbanos; e uma espécie de bombacácea, planta considerada como quiropterofílica
21

INSETIVORIA E CONTROLE DE PRAGAS
A maior parte das espécies possui hábito alimentar insetívoro
(ALTRINGHAM, 1996). Todas as famílias presentes na região neotropical
apresentam espécies com dieta parcial ou exclusivamente composta por
insetos, de inúmeras Ordens: Diptera, Lepidoptera, Orthoptera, Coleoptera,
Hemiptera, Hymenoptera, Isoptera, Neuroptera, Odonata, entre outras
(DEBELICA et al., 2006; DECHMANN et al., 2006).
Apesar do termo “insetívoro”, os morcegos com essa dieta podem
consumir outros artrópodes, como aracnídeos, diplópodos e quilópodos
(ALTRINGHAM, 1996). Morcegos insetívoros, podem consumir o dobro do seu
peso em insetos em uma única noite e por isso são os principais controladores
naturais de artrópodes (ROMANO et al., 1999), sendo extremamente
importantes para a agricultura e o meio ambiente. LONG (1996) cita que a
introdução de morcegos em zonas rurais reduziu drasticamente as pragas
agrícolas, possibilitando a diminuição no uso de agrotóxicos.
ROMANO et al. (1999) estimaram o consumo de cerca de 385kg de
insetos/noite por uma colônia com aproximadamente 64.000 morcegos
molossídeos da espécie Tadarida brasiliensis, e segundo ALTRINGHAM (1996),
essa mesma colônia é responsável por consumir 13.000 toneladas de insetos
durante o verão.
Analisando os hábitos alimentares das espécies insetívoras, é possível
observar a incidência de diversas ordens de insetos que podem ser prejudiciais
à sociedade, tanto como pragas urbanas, quanto agrícolas. A grande
quantidade de insetos consumidos por noite indica que morcegos são de
inestimável importância para o controle das populações desses insetos e das
pragas causadas por eles.
Molossus molossus Foto: Marco Mello
Eptesicus brasiliensis Foto: Roberto Novaes
Espécies consideradas “comuns” na maior parte do território brasileiro. São freqüententemente capturadas em áreas florestais e urbanas e são consideradas como grandes contro-dores de insetos e pragas.
22

PISCIVORIA E CONTROLE DE POPULAÇÕES
Na região neotropical existem duas espécies piscívoras (ou ictiófagas
como citam alguns autores): Noctilio leporinus (Noctilionidae) e Myotis vivesi
(Vespertilionidae). Estas espécies se alimentam de alevinos de água doce ou
salgada, ártropodes aguáticos e terrestres (ALTENBACH, 1989; BORDIGNON,
2006). Dentre os muitos peixes consumidos por estes morcegos, estão os das
famílias Clupeidae, Atherinopsidae, Mugelidae, Englaulidae, Centropomidae e
Carangidae, e dentre os artrópodes utilizados na alimentação estão
principalmente os das Classes Insecta, Crustacea e Arachnida (BORDIGNON &
FRANÇA, 2002; BORDIGNON, 2006).
Devido à plasticidade de seus hábitos alimentares e ao metabolismo
acelerado, o que os leva consumo de grandes quantidades alevidos por noite
(BORDGNON, 2006), morcegos piscívoros são de grande importância ambiental,
já que são grandes controladores de população, seja de peixes ou de
artrópodes.
No Brasil, há registro apenas para Noctilio leporinus, que costuma ser
visto em ambientes costeiros, principalmente baías, prais de baixo
hidrodinamismo, lagoas e lagos.
Noctilio leporinus Foto: Marco Mello
Noctilio leporinus tem ampla distribuição em território brasileiro, porém poucos estudos sobre sua biologia e ecologia foram feitos.
23

CARNIVORIA E CONTROLE DE POPULAÇÕES
Dentre todas as famílias, apenas Phyllostomidae apresenta espécies com
uma dieta carnívora. O comportamento alimentar de tais morcegos é baseado
no consumo de pequenos vertebrados como pequenos roedores, anfíbios
(Anuros), pequenas aves, e até mesmo outros morcegos (PERACCHI et al., 1982;
FENTON, 1992; ESBÉRARD & BERGALLO, 2004; BORDIGNON, 2005; OPREA et al.,
2006). No entanto, boa parte das espécies carnívoras também podem consumir
artrópodes regularmente (PERACCHI & ALBUQUERQUE, 1976) e eventualmente
frutos (UIEDA et al., 2007) e por isso, muitas vezes são consideradas onívoras.
Segundo FENTON et al. (1992), morcegos pertencentes à guilda dos
animalívoros são indicadores de ambientes preservados, e por isso raros em
ambientes urbanos e áreas fragmentadas.
Sendo assim, os morcegos que compõem este grupo são responsáveis,
juntamente com outros animais, pelo controle das populações de pequenos
vertebrados e artrópodes de áreas florestais.
Chrotopterus auritus Foto: Marco Mello
Tonatia bidens Foto: Marco Mello
Vampyrum spectrum Foto: Maricélio Guimarães
Os morcegos carnívoros brasileiros pertencem à subfamília Phyllostominae. Chroropterus auritus e Tonatia bidens, possuem grande distribuição, ocorrendo em grande parte do país. Já Vampyrum spectrum possui distribuição restrita, com ocorrência predominante no ecossistema amazônico.
24
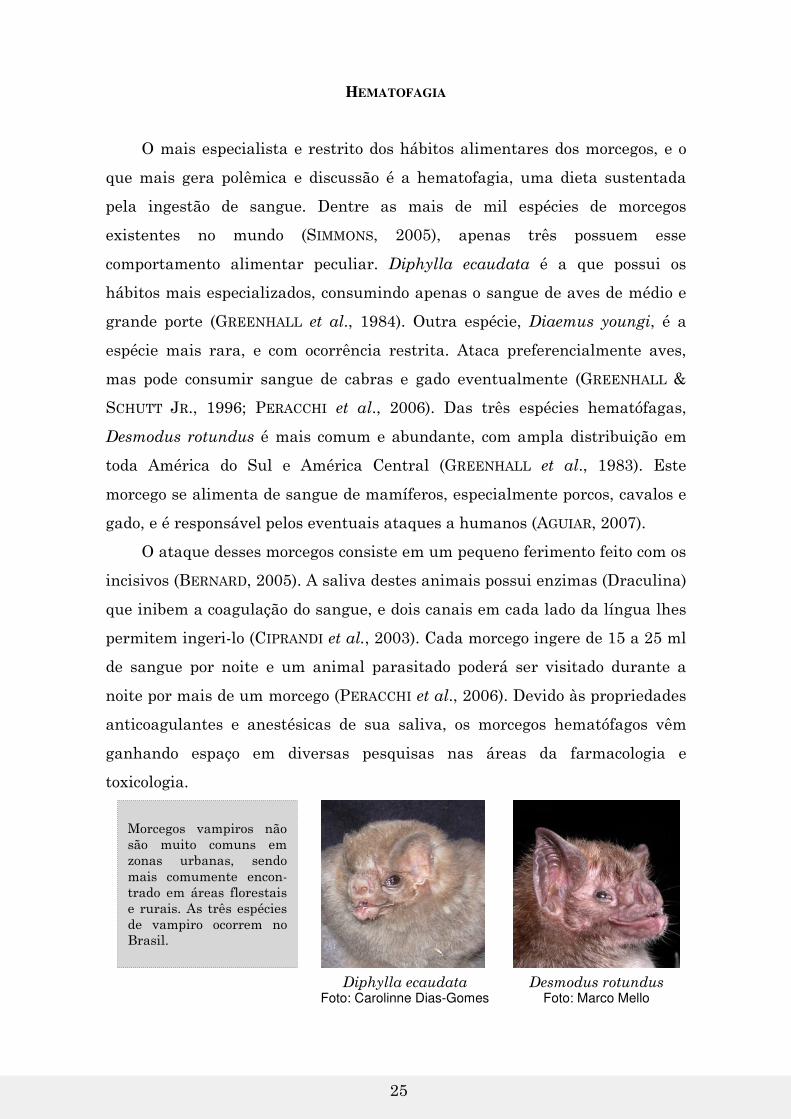
HEMATOFAGIA
O mais especialista e restrito dos hábitos alimentares dos morcegos, e o
que mais gera polêmica e discussão é a hematofagia, uma dieta sustentada
pela ingestão de sangue. Dentre as mais de mil espécies de morcegos
existentes no mundo (SIMMONS, 2005), apenas três possuem esse
comportamento alimentar peculiar. Diphylla ecaudata é a que possui os
hábitos mais especializados, consumindo apenas o sangue de aves de médio e
grande porte (GREENHALL et al., 1984). Outra espécie, Diaemus youngi, é a
espécie mais rara, e com ocorrência restrita. Ataca preferencialmente aves,
mas pode consumir sangue de cabras e gado eventualmente (GREENHALL &
SCHUTT JR., 1996; PERACCHI et al., 2006). Das três espécies hematófagas,
Desmodus rotundus é mais comum e abundante, com ampla distribuição em
toda América do Sul e América Central (GREENHALL et al., 1983). Este
morcego se alimenta de sangue de mamíferos, especialmente porcos, cavalos e
gado, e é responsável pelos eventuais ataques a humanos (AGUIAR, 2007).
O ataque desses morcegos consiste em um pequeno ferimento feito com os
incisivos (BERNARD, 2005). A saliva destes animais possui enzimas (Draculina)
que inibem a coagulação do sangue, e dois canais em cada lado da língua lhes
permitem ingeri-lo (CIPRANDI et al., 2003). Cada morcego ingere de 15 a 25 ml
de sangue por noite e um animal parasitado poderá ser visitado durante a
noite por mais de um morcego (PERACCHI et al., 2006). Devido às propriedades
anticoagulantes e anestésicas de sua saliva, os morcegos hematófagos vêm
ganhando espaço em diversas pesquisas nas áreas da farmacologia e
toxicologia.
Desmodus rotundus Foto: Marco Mello
Diphylla ecaudata Foto: Carolinne Dias-Gomes
Morcegos vampiros não são muito comuns em zonas urbanas, sendo mais comumente encon-trado em áreas florestais e rurais. As três espécies de vampiro ocorrem no Brasil.
25

REFERÊNCIAS BIBLIOGRÁFICAS
AGUIAR, L.M.S. 2007. Subfamília Stenodermatinae. In: REIS, N.R., PERACCHI,
A.L. PEDRO, W.A. & LIMA, I.P. (Eds). Morcegos do Brasil. Editora da
Universidade Estadual de Londrina. P. 39-43.
ALTENBACH, J.S. 1989. Prey capture by the fishing bats Noctilio leporinus and
Myotis vivesi. Journal of Mammalogy 70: 421-424.
ALTRINGHAM, J.D. 1996. Bats: Biology and Behaviour. Oxford University
Press. New York. 262p.
BERNARD, E. & FENTON, M.B. 2003. Bat mobility and Roosts in a fragmented
landscape in Central Amazonia, Brazil. Biotropica 35(2): 262-277.
BARRESE, C. 2005. Fenologia de plantas do gênero Piper (Linnaeus,
1737) (Piperales, Piperaceae): Implicações em quiropterocoria.
Monografia de graduação. Centro Universitário da Fundação de Ensino
Octávio Bastos. 40p.
BERNARD, E. 2005. Morcegos Vampiros: Sangue, raiva e preconceito. Ciência
Hoje 36(214): 44-49.
BORDIGNON, M.O. 2005. Predação de morcegos por Chrotopterus auritus
(Peters) (Mammalia, Chiroptera) no pantanal de Mato Grosso do Sul,
Brasil. Revista Brasileira de Zoologia 22(4): 1207-1208.
BORDIGNON, M.O. 2006. Padrão de atividade e comportamento de
forrageamento do morcego-pescador Noctilio leporinus (Linnaeus)
(Chiroptera, Noctilionidae) na Baía de Guaratuba, Paraná, Brasil.
Revista Brasileira de Zoologia 23(1): 50-57.
26

BORDIGNON, M.O. & FRANÇA, A.O. 2002. Fish consumption by Noctilio
leporinus (Linnaeus, 1758) in Guaratuba Bay, southern Brazil.
Chiroptera Neotropical 8(1-2): 148-150.
CIPRANDI, A., HORN, F. & TERMIGNONI, C. 2003. Saliva de animais
hematófagos: fonte de novos anticoagulantes. Revista Brasileira de
Hematologia e Hemoterapia 25(4): 250-262.
DEBELICA, A., MATTHEWS, A.K. & AMMERMAN, L.K. 2006. Dietary study of big
free-tailed bats (Nyctinomops macrotis) in Big Bend National Park,
Texas. The Southwestern Naturalist 51(3): 414-418.
DECHMANN, D.K.N., SAFI, K. & VONHOF, M.J. 2006. Matching morphology and
diet in the disc-winged bat, Thyroptera tricolor (Chiroptera). Journal of
Mammalogy 87(5): 1013-1019.
ESBÉRARD, C.E.L. & BERGALLO, H.G. 2004. Aspectos sobre a biologia de
Tonatia bidens (Spix) no estado do Rio de Janeiro, sudeste do Brasil
(Mammalia, Chiroptera, Phyllostomidae). Revista Brasileira de
Zoologia 21(2): 253-259.
FENSTER, C.B., ARMBRUSTER, W.S., WILSON, P., DUDASH, M.R. & THOMSON, J.D.
2004. Pollination syndromes and floral specialization. Annual Review
Ecology, Evolution and Systematic 35: 375–403.
FENTON, M.B. 1992. Bats. Facts on File. New York. 2007p.
FENTON, M.B., ACHARYAS, L., ÁUDET, D., HICKEY, M.B.C., MERRIMAN, C.,
OBRIST, M.K. & SYME, D.M. 1992. Phyllostomid bats (Chiroptera:
Phyllostomidae) as indicators of habitat disruption in the Neotropics.
Biotropica 24(3): 440-446.
27

FLEMING, T.H. & SOSA, V.J. 1994. Effects of nectarivorous and frugivorous
mammals on reproductive success of plants. Journal of Mammalogy
75: 845-851.
GARDNER, A.L. 1977. Feeding habits. In: BAKER, R.J; JONES-JR, J.K. & CARTER,
D.C. (Eds). Biology of Bats of the New World, Family Phyllostomidae.
Part II. Special Publication of the Museum, Texas Tech University
V.13. P. 239-350.
GREENHALL, A.M. & SCHUTT JR., W.A. 1996. Diaemus youngi. Mammalian
Species 533: 1-7.
GREENHALL, A.M., JOERMANN, G. & SCHMIDT, U. 1983. Desmodus rotundus.
Mammalian Species 202: 1-6.
GREENHALL, A.M., SCHMIDT, U. & JOERMANN, G. 1984. Diphylla ecautada.
Mammalian Species 227: 1-3.
HERRERA, C.M. 1982. Seasonal variations in the quality of fruits and diffuse
coevolution between plants and avian disperses. Ecology 63: 773-785.
LONG, R.F. 1996. Bats for insect biocontrol in agriculture. IPM-Practitioner
18(9): 1-6.
MELLO, M.A.R. 2002. Interações entre o morcego Carollia perspicillata
(Linnaeus, 1758) (Chiroptera: Phyllostomidae) e plantas do
gênero Piper (Linnaeus, 1737) (Piperales: Piperaceae) em uma
área de Mata Atlântica. Dissertação de Mestrado. Universidade do
Estado do Rio de Janeiro. 61p.
MELLO, M.A.R. 2006. Interações entre o morcego Sturnira lilium
(Chiroptera: Phyllostomidae) e plantas da família Solanaceae.
Tese de Doutorado. Universidade Estadual de Campinas. 144p.
28

MELLO, M.A.R., KALKO, E.K.V. & SILVA, W.R. 2008. Movements of the bat
Sturnira lilium and its role as a seed disperser of Solanaceae in the
Brazilian Atlantic Forest. Journal of Tropical Ecology 24: 225-228.
MORRISON, D.W. 1980. Efficiency of food utilization by fruit bats. Oecologia
45: 270-273.
NASSAR, J.M., RAMÍREZ, N. & LINARES, O. 1997. Comparative pollination
biology of Venezuela columnar cacti and the role of nectar-feeding bats in
their sexual reprodution. American Journal of Botany 84: 918-927.
NOGUEIRA, M.R. & PERACCHI, A,.L. 2002. The feeding specialization in
Chiroderma doriae (Phyllostomidae, Stenodermatinae) with comments on
its conservation implications. Chiroptera Neotropical 8(1-2): 143-148.
NOGUEIRA, M.R. & PERACCHI, A.L. 2003. Fig-seed predation by 2 species of
Chiroderma: Discovety of a new feeding strategy in bats. Journal of
Mammalogy 84(1): 225-233.
NOVAES, R.L.M., OLIVEIRA, C.C.N., MENEZES JR., L.F., DUARTE, A.C. no prelo.
Dieta de Artibeus lituratus (Olfers, 1818) (Mammalia, Chiroptera) em
área urbana: frugivoria e novo registro de folivoria. Revista Brasileira
de Zoociências.
OPREA, M., VIEIRA, T.B., PIMENTA, V.T., MENDES, P., BRITO. D., DITCHFIELD,
A.D., KNEGT, L.V. & ESBÉRARD, C.E.L. 2006. Bat predation by
Phyllostomus hastatus. Chiroptera Neotropical 12(1): 255-258.
PASSOS, F.C. & GRACIOLLI, G. 2004. Observações da dieta de Artibeus lituratus
(Olfers) (Chiroptera: Phyllostomidae) em duas áreas do sul do Brasil.
Revista Brasileira de Zoologia 21(3): 487-489.
29

PASSOS, F.C. & SILVA, W.R. 2003. Frugivoria e ecologia alimentar em
morcegos. In: Anais do IV Encontro Brasileiro para o Estudo de
Quirópteros - IV EBEQ.
PASSOS, F.C., SILVA, W.R., PEDRO, W.A. & BONIN, M.R. 2003. Frugivoria em
morcegos (Mammalia, Chiroptera) no Parque Estadual Intervales,
sudeste do Brasil. Revista Brasileira de Zoologia 20(3): 511-517.
PERACCHI, A.L. & ALBUQUERQUE, S.T. 1976. Sobre os hábitos alimentares de
Chrotopterus auritus australis Thomas, 1905 (Mammalia, Chiroptera,
Phyllostomidae). Revista Brasileira de Biologia 36(1): 179-184.
PERACCHI, A.L., ALBUQUERQUE, S.T. & RAIMUNDO, S.D.L. 1982. Contribuição
ao conhecimento dos hábitos alimentares de Trachops cirrhosus (Spix,
1823) (Mammalia: Chiroptera: Phyllostomidae). Arquivos da
Universidade Federal Rural do Rio de Janeiro 5(1): 1-5.
PERACCHI, A.L., LIMA, I.P., REIS, N.L., NOGUEIRA, M.L. & ORTÊNCIO-FILHO, H.
2006. Ordem Chiroptera. In: REIS, N.R., PERACCHI, A.L., PEDRO, W.A. &
LIMA, I.P (Eds). Mamíferos do Brasil. Universidade Estadual de
Londrina. p.153-230.
PETIT, S. 1995. The pollinators of two species of Columnar Cacti on Curação,
Netherlands Antilles. Biotropica 27: 538-541.
REIS, N.R., PERACCHI, A.L., PEDRO, W.A., LIMA, I.P. (Eds). 2007. Morcegos do
Brasil. Editora da Universidade Estadual de Londrina. 253p.
ROMANO, M.C., MAIDAGAN, J.I. & PIRE, E.F. 1999. Behaviour and demography
in urban colony of Tadarida brasiliensis (Chiroptera, Molossidae) in
Rosario, Argentine. Revista de Biologia Tropical 47(4): 1121-1127.
30

SAZIMA, M., BUZATO, S. & SAZIMA, I. 1999. Bat-pollinated Flower Assemblages
and Bat Visitors at Two Atlantic Forest Sites in Brazil. Annals of
Botany 83: 705-712.
SAZIMA, M., BUZATO, S. & SAZIMA, I. 2003. Dyssochroma viridiflorum
(Solanaceae): a reproductively bat-dependent epiphyte from the Atlantic
Rainforest in Brazil. Annals of Botany 92: 725-730
SILVA, S.S.P. & PERACCHI, A.L. 1999. Visits of bats to flowers of Lafoensia
glyptocarpa Koehne (Lythraceae). Revista Brasileira Biologia 59(1):
19-22.
SIMMONS, N.B. 2005. Order Chiroptera. In: WILSON, D.E., REEDER, D.M. (Eds).
Mammals Species of the World: a taxonomic and geographic
reference. 3.ed. v.1. Jonhs Hopkins University Press. p.312-529.
SORIANO, P.J. RUIZ, A. & NASSAR, J.M. 2000. Notas sobre la distribuición e
importancia ecologica de los murciélagos Leptonycteris curasoe y
Glossophaga longirostris en zonas áridas andinas. Ecotropicos 13(2):
31-35.
STROO, A. 2000. Pollen morphological evolution in bat pollinated plants. Plant
Systematic and Evolution 222: 225-242.
TSCHAPKA, M. & VON HELVERSEN, O. 1999. Pollinators of syntopic Marcgravia
species in Costa Rican Lowland Rain Forest: bats and opossums. Plant.
Biology 1: 382-388.
UIEDA, W., SATO, T.M., CARVALHO, M.C. & BONATO, V. 2007. Fruit as unusual
food items of the carnivorous bat Chrotopterus auritus (Mammalia,
Phyllostomidae) from southeastern Brazil. Revista Brasileira de
Zoologia 24(3): 844-847.
31

WENDT, T., CANELA, M.B.F., FARIA, A.P.G. & RIOS, R.I. 2001. Reproductive
biology and natural hybridization between two endemic species of
Pitcairnia (Bromeliaceae). American Journal of Botany 88(10): 1760-
1767.
WHITTAKER, R.J. & JONES, S.H. 1994. The role of frugivorous bats and birds in
the rebuilding of a tropical forest ecosystem, Krakatau, Indonesia.
Journal of Biogeography 21:245-258.
ZORTÉA, M. 2007. Subfamília Stenodermatinae. In: REIS, N.R., PERACCHI, A.L.
PEDRO, W.A. & LIMA, I.P. (Eds). Morcegos do Brasil. Editora da
Universidade Estadual de Londrina. p.107-128.
32

IV
RAIVA EM QUIRÓPTEROS

INTRODUÇÃO
A enfermidade mais comumente associada aos morcegos é a Raiva,
doença virótica aguda e letal (BREDT et al., 1998). Diversas espécies de pelo
menos três famílias (Phyllostomidae, Molossidae e Vespertilionidae) já
tiveram a Raiva diagnosticada (ALMEIDA et al., 1994; CUNHA et al., 2006).
A Raiva quando transmitida por morcegos hematófagos, principalmente
Desmodus rotundus, pode trazer grandes prejuízos à percuária da América
Latina devido à contaminação de herbívoros domésticos (GREENHALL et al.,
1983; ACHA & MÁLAGA-ALBA, 1988). O vírus rábico parece ter encontrado nos
morcegos hematófagos o melhor e o mais eficiente veículo de propagação, uma
vez que estes agridem diariamente outros animais usados como alimentação e
até mesmo outros morcegos nas interações sociais (BREDT et al., 1998).
MAYEN (2003) alerta sobre o impacto do vírus rábico sobre a saúde
pública, entretando, a vacinação de animais domésticos reduz o risco de
epidemia, e mesmo os morcegos sendo potenciais transmissores da Raiva
(CUNHA et al., 2006), acredita-se que eles não oferecem grandes riscos à
sociedade de um modo geral.
PATOGENIA
O vírus da Raiva ataca o sistema nervoso central de mamíferos e, para
que haja a infecção, ele deve entrar em contato com a corrente sangüínea do
animal, seja por meio de uma ferida, corte, mordedura, arranhão ou contato
com líquidos corpóreos contaminados (BERNARD, 2005). Um dos locais de maior
concentração do vírus são nas glândulas salivares do animal, o que aumenta a
possibilidade de transmissão por contato da saliva contaminada (BREDT et al.,
1998; BERNARD, 2005). É bom enfatizar que não só morcegos são vetores da
Raiva, pelo contrário, a maior partes das infecções são constatadas em
carnívoros silvestres e domésticos, embora qualquer animal de sangue quente
seja susceptível à Raiva, em menor ou maior grau (BREDT et al., 1998).
34

SINTOMATOLOGIA DE RAIVA EM MAMÍFEROS
Um mito comum é de que morcegos são imunes ao vírus, porém isso não é
verdade (BERNARD, 2005). Segundo BREDT et al. (1998), em quirópteros a
Raiva apresenta três fases: prodrômica, excitativa e paralítica.
A fase prodrômica possui curta duração, de 2 a 3 dias, caracterizada pelos
sinais iniciais da doença. O animal apresenta pequenas alterações
comportamentais como hiperexcitabilidade à luz, ruídos, deslocamentos de ar
e outros fatores externos.
Na fase excitativa os sintomas são os mais associados à doença. Durante
esta fase, encontram-se exacerbados os sinais de hiperexcitabilidade
observados durante a fase prodrômica e o animal pode se tornar agressivo. A
fase excitativa pode durar de 3 a 7 dias, quando a transmissão do vírus é mais
freqüente.
SINTOMATOLOGIA EM MORCEGOS HEMATÓFAGOS
E COMPORTAMENTO SOCIAL
A sintomatologia da raiva em morcegos hematófagos, especificamente em
D. rotundus, é relativamente bem conhecida. O comportamento e os sintomas
mais freqüentes são: atividade alimentar diurna, hiperexcitabilidade,
agressividade, tremores, falta de coordenação dos movimentos, contrações
musculares e paralisia. No começo da enfermidade, os indivíduos doentes
afastam-se da colônia, deixam de realizar asseio corporal, seus pêlos tomam-se
desalinhados e sujos. Tremor generalizado pode ser observado em vários deles.
Feridas frescas são freqüentes e provocadas por agressões de seus
companheiros sadios a cada tentativa de reintegração ao grupo, de onde são
expulsos violentamente. O morcego enfermo perde a capacidade de voar e pode
cair ao chão. A incapacidade de vôo é o primeiro sintoma motriz observado nos
morcegos raivosos, porém isto não os impede de caminhar pelo chão ou pelas
paredes (BREDT et al., 1998).
Num estágio mais avançado da doença os morcegos infectados começam a
ter mais dificuldades de caminhar e de sustentar seu corpo sobre os pés e
35

polegares das asas. Sinais de desidratação são percebidos e há um aumento
gradativo dos sintomas paralíticos, com maior intensidade nas asas do que nas
extremidades posteriores. A morte do indivíduo pode ocorrer cerca de 48 horas
após o aparecimentos dos sintomas.
SINTOMATOLOGIA EM MORCEGOS NÃO HEMATÓFAGOS
Nestes morcegos a raiva manifesta-se principalmente sob a forma
paralítica sem a visualização da fase excitável. Há alguns relatos de casos de
morcegos insetívoros americanos voando e perseguindo outros morcegos, casos
atribuídos a indivíduos raivosos em fase agressiva. Em outras regiões, não há
relatos semelhantes. Relatos de paralisia das asas e retenção urinária foram
descritos para a espécie Tadarida brasiliensis, no Estado do Texas, EUA. A
paralisia progressiva das asas dificulta o vôo e em uma fase mais adiantada
da doença os morcegos deixam de voar. Nessas situações os morcegos podem
deixar de sair para se alimentar, ou quando fora do abrigo, enfrentam
dificuldades para retomar. Morcegos encontrados em locais não habituais, no
chão, sobre móveis, pendurados em cortinas, paredes, janelas e muros, têm
grandes chances de estarem contaminados pelo vírus da Raiva, assim como
aqueles que estejam voando durante o dia (BRASS, 1994; BREDT et al., 1998).
No Brasil, 48,1% das espécies infectadas com vírus rábico são insetívoras, 18,5% são frugívoras, nectarívoras e hematófagas somam 11,1% cada, carnívoras 7,4% e onívoras 3,7%. Mesmo assim, devido ao seu hábito alimentar, os morcegos vampiros ainda são os principais transmissores do vírus rábico para outros animais.
(BREDT et al., 1998)
Segundo UIEDA et al. (1996), cerca de 27 espécies de morcegos já foram diagnosticadas com Raiva no Brasil, no entanto, em 80% dos casos a transmissão para humanos é feita por cães e gatos (CUNHA et al., 2005).
36

REFERÊNCIAS BIBLIOGRÁFICAS
ACHA, P.N. & MÁLAGA-ALBA, M. 1988. Economic losses due to Desmodus
rotundus. In: GREENHALL, A.M. & SCHIMIDT, U. (Eds). Natural history
of vampire bats. CRC Press. pp. 208-213.
ALMEIDA, M.F., AGUIAR, E.A.C., MARTORELLI, L.F.A. & SILVA, M.M.S. 1994.
Diagnóstico laboratorial de raiva em quirópteros realizado em área
metropolitana na região sudeste do Brasil. Revista de Saúde Pública
28(5): 341-344.
BERNARD, E. 2005. Morcegos Vampiros: sangue, raiva e preconceito. Ciência
Hoje 36(214): 44-49.
BRASS, D.A. 1994. Rabies in bats: natural history and public health
implications. Livia Press. 335p.
BREDT, A., ARAÚJO, F.A.A., CAETANO-JR, J., RODRIGUES, M.G.R., YOSHIZAWA,
M., SILVA, M.M.S., HARMANI, N.M.S., MASSUNAGA, P.N.T., BÜRER, S.P.,
PORTO, V.A.R. & UIEDA, W. 1998. Morcegos em áreas urbanas e
rurais: Manual de manejo e controle. 2ªEd. Fundação Nacional de
Saúde. 117p.
CUNHA, E.M.S., LARA, M.C.C.S.H., NASSAR, A.F.C., SODRÉ, M.M. & AMARAL,
L.F.V. 2005. Isolamento do vírus da raiva em Artibeus fimbriatus no
Estado de São Paulo. Revista de Saúde Pública 39 (4): 683-684.
CUNHA, E.M.S., SILVA, L.H.Q., LARA, M.C.C.S.H., NASSAR, A.F.C., ALBAS, A.,
SODRÉ, M.M. & PEDRO, W.A. 2006. Bat rabies in the north-nortwestern
regions of the state of São Paulo, Brazil: 1997-2002. Revista de Saúde
Pública 40(6): 1082-1086.
37

GREENHALL, A.M., JOERMANN, G. & SCHMIDT, U. 1983. Desmodus rotundus.
Mammalian Species 202: 1-6.
MAYEN, F. 2003. Haematophagous bats in Brazil, their role in rabies
transmission, impact on public health, livestock industry and
alternatives to an indiscriminate reduction of bat population. Journal of
Veterinary Medicine B 50: 469-472.
UIEDA, W., HAYASHI, M.M., GOMES, L.H. & SILVA, M.M.S. 1996. Espécies de
quirópteros diagnosticados com Raiva no Brasil. Boletim do Instituto
Pasteur 1: 17-35.
38

V
ABRIGOS DIURNOS

INTRODUÇÃO
Morcegos passam a metade de suas vidas dentro dos abrigos, por isso não
é surpreendente que as condições e eventos relacionados aos abrigos tenham
um papel importante na ecologia e evolução deles (KUNZ,1982). Os abrigos
provém locais para acasalamento, criação de filhotes, interações sociais e
digestação alimentar, além de protegerem os morcegos das adversidades
climáticas e de predadores (KUNZ, 1982; KUNZ & MCCRACKEN, 1996). Assim, as
condições de um abrigo têm influência direta sobre as taxas de natalidade e
mortalidade, tornando-o especialmente importante para a sobrevivência de
uma espécie (KUNZ, 1982).
Normalmente, fatores como estabilidade da temperatura ambiente,
umidade e luminosidade, determinam a ocupação (BREDT et al., 1998).
A ecologia dos abrigos de morcegos pode ser vista como uma interação
complexa de adaptações fisiológicas, comportamentais, morfológicas e
respostas demográficas (KUNZ, 1982), que podem envolver a fidelidade a um
mesmo abrigo (ESBÉRARD et al., 2003), ou comportamento nômade, o que pode
variar de acordo com a espécie ou com as condições ambientais do local.
São muitos os abrigos diurnos utilizados por morcegos, desde folhas, ocos
de árvores e cavernas (BROOKE, 1987; SIMMONS & VOSS, 1998; TRAJANO &
GIMENEZ, 1998), até forros de casas e frestas de edifícios (PERACCHI et al.,
2006).
ABRIGOS NATURAIS
Ambientes florestais oferecem ilimitados recursos de hábitat, entre os
quais abrigos com diferentes características, que são escolhidas de acordo com
a espécie de morcego ou mesmo o hábito alimentar. Morcegos de todos os
hábitos alimentares utilizam refúgios naturais (FENTON et al., 2000; PERACCHI
et al., 2006) e praticamente todas as famílias que ocorrem nos neotrópicos
possuem espécies que já foram encontradas se abringando em locais naturais
(SIMMONS & VOSS, 1998), não só em ambientes florestais, como em perímetros
urbanos.
40

Ocos de árvore
Freqüentemente morcegos são encontrados se abrigando em ocos de
árvores de inúmeras espécies (KUNZ, 1983), e em praticamente todas as
guildas alimentares há espécies que já foram encontradas se abrigando neste
tipo de refúgio (SIMMONS & VOSS, 1998). FENTON et al. (2000) indicaram a
presença do morcego filostomídeo frugívoro Sturnira lilium em ocos de árvore
em Belize. Morcegos insetívoros das família Emballonuridae, Molossidae,
Phyllostomidae e Vespertilionidae também já foram capturados em cavidades
em árvores e troncos ocos (SIMMONS & VOSS, 1998; BIANCONI & PEDRO, 2007;
FABIAN & GREGORIN, 2007). O morcego piscívoro Noctilio leporinus também já
foi reportado se abrigando dentro do tronco de árvores ocas (HOOD & JONES
JR., 1984). Morcegos carnívoros e onívoros são comumente encontrados dentro
de árvores (NOGUEIRA et al., 2007), já morcegos hematófagos não costumam
utilizar estes abrigos (GREENHALL et al., 1983; AGUIAR, 2007), exceto Diaemus
youngi.
Folhas
Muitos morcegos, de todos os hábitos alimentares, se abrigam sob as
folhas de árvore (KUNZ, 1983), no entanto esse tipo de refúgio ainda é mais
comum entre morcegos de dieta frugívora (BROOKE, 1987; NOVAES et al. no
prelo). Comumente morcegos frugívoros são vistos se abrigando em folhas de
árvores em zonas urbanas (NOVAES et al. no prelo) e ambientes florestais
(SIMMONS & VOSS, 1998). Algumas espécies frugívoras desenvolveram
habilidades especiais para construções de tendas (KUNZ et al., 1994), e esse
tipo de abrigo consiste no dobramentos de folhas de plantas, formando uma
espécie de cabana, onde os morcegos se abrigam sob elas (KUNZ, 1983; KUNZ &
MCCRACKEN, 1994; KUNZ et al., 1994).
O uso de folhas como abrigo já foi reportado também para morcegos
insetívoros, principalmente das famílias Emballonuridae e Phyllostomidae
(SIMMONS & VOSS, 1998), além de outras espécies carnívoras, onívoras
(NOGUEIRA et al., 2007) e nectarívoras (PERACCHI et al., 2006).
41

Cavernas
De todos os abrigos naturais usados pelos quirópteros, as cavernas são as
mais comuns e associadas aos morcegos (EZZELL, 1992), e espécies de
absolutamente todas as famílias e hábitos alimentares já foram capturados em
ambientes cavernícolas (KUNZ, 1983; PERACCHI et al., 2006).
Dentre as várias guildas alimentares, os insetívoros e hematófagos
mantém presença destacada nesses ambientes (PERACCHI et al., 2006),
seguidos por carnívoros e onívoros (NOGUEIRA et al., 2007), nectarívoros
(COELHO, 2005) e frugívoros (ZORTÉA, 2007).
BREDT et al. (1999) encontraram 22 espécies de morcegos habitando
cavernas do Brasil central. TRAJANO & GIMENEZ (1998) listaram 13 espécies
em uma única caverna no sudeste do Brasil. ESBÉRARD et al. (2005) citam
presença confirmada de 29 espécies distribuídas em sete famílias em cavernas
de um único fragmento de Cerrado.
Além da grande diversidade, algumas cavernas abrigam as maiores
colônias de morcegos do mundo. Nos neotrópicos essas colônias são compostas
por morcegos insetívoros das famílias Molossidae e Vespertilionidae e podem
abrigar mais de 60.000 morcegos (GRAHAM, 1994).
ABRIGOS ARTIFICIAIS
Morcegos são bem adaptados aos ambientes urbanos e em algumas
cidades a diversidade é consideravelmente alta (REIS et al., 2002) e por isso
esses animais são vistos comumente se abringando em edificações humanas
(PERACCHI et al., 2006).
Boa parte dos morcegos encontrados nas cidades são insetívoros, e talvez
isto aconteça porque estes morcegos costumam forragear perto de postes de
iluminação para se alimentar dos insetos atraídos pelas luzes (LEE &
MCCRACKEN, 2002). E apesar de haver muitos morcegos frugívoros em cidades
(REIS et al., 2002), esses costumam ocupar abrigos naturais na maioria dos
casos, diferente de morcegos insetívoros, que representam cerca de 75% das
ocupações de abrigos artificiais por morcegos (ESBÉRARD, 2003).
42

Minas, porões e pontes.
Por manter características físicas semelhantes aos ambientes
cavernícolas, minas e porões também podem ser potenciais abrigos para
quirópteros (JOHNSON et al., 2006). Na maioria dos casos, os morcegos que
ocupam esses ambientes são insetívoros, mas morcegos de outros hábitos
alimentares também podem ser encontrados (BREDT et al., 1998).
Segundo PERACCHI et al. (2006), morcegos de todos os hábitos alimentares
já foram observados se abrigando e minas e porões e NOGUEIRA & POL (1998),
constataram a coabitação de duas espécies insetívoras, Rinchonycteris naso
(Emballonuridae) e Noctilio albiventris (Noctilionidae) se abrigando sob uma
ponte no sudeste do Brasil. Nos EUA, grandes colônias de morcegos
insetívoros da família Vespertilionidae se abrigam sob pontes durante
períodos sazonais (GRAHAM, 1994).
Forros, frestas e outros abrigos.
Muitas casas possuem morcegos usando componentes das edificações
como abrigo, e a maior parte desde morcegos possui dieta insetívora, embora
quirópteros com outras dietas também sejam encontrados nesses locais.
YEE (2000) cita a coabitação de seis espécies de morcegos com diferentes
hábitos alimentares - os insetívoros Saccopteryx bilineata (Emballonuridae) e
Myotis nigricans (Vespertilionidae), o frugívoro Carollia perspicillata
(Phyllostomidae), o nectarívoro Glossophaga soricina (Phyllostomidae) e os
hematófagos Desmodus rotundus e Diphylla ecaudata (Phyllostomidae) - todos
dividindo o mesmo abrigo em uma construção humana. ESBÉRARD (2003)
reporta também a utilização do forro e frestas de uma casa pelo morcego
insetívoro Molossus rufus (Molossidae) e pelo onívoro Phyllostomus hastatus
(Phyllostomidae).
E segundo REGIDOR et al. (2003), uma grande colônia do molossídeo
Tadarida brasilensis abriga as frestas de uma represa na Argentina,
indicando a grande adaptabilidade dos morcegos em ocupar diversos tipos de
abrigos.
43

Chaminé
Frestas nas telhas
Forro
Vão no telhado
Morcegos são extremamente versáteis quanto à habitação e uso de abrigos. Boa parte das casas com forro possuem colônias de morcegos se abrigando em seu interior e segundo ESBÉRARD (2003) 75% dos morcegos em que habitam construções humanas são insetívoros e pertencentes às famílias Molossidae e Vespertilionidae. Edificações humanas constituem bons locais para moradia de morcegos, e normalmente os morcegos abrigados nos forros utilizam chaminés, frestas nas telhas e vãos no telhado para entrada e saída. É bom ressaltar que morcegos são animais silvestres e por isso protegidos pela Consituição Federal, e por mais que possam ser inconvenientes e trazer prejuízos para proprietários de casas com morcegos no forro, sua retirada, apanha ou destruição é crime previsto por lei e implicará em multa ou reclusão.
44

REFERÊNCIAS BIBLIOGRÁFICAS
AGUIAR, L.M.S. 2007. Subfamília Stenodermatinae. In: REIS, N.R., PERACCHI,
A.L. PEDRO, W.A. & LIMA, I.P. (Eds). Morcegos do Brasil. Editora da
Universidade Estadual de Londrina. p.39-43
BIANCONI, G.V. & PEDRO, W.A. 2007. Família Vespertilionidae. In: REIS, N.R.,
PERACCHI, A.L. PEDRO, W.A. & LIMA, I.P. (Eds). Morcegos do Brasil.
Editora da Universidade Estadual de Londrina. p.167-195.
BREDT, A., ARAÚJO, F.A.A., CAETANO-JR, J., RODRIGUES, M.G.R., YOSHIZAWA,
M., SILVA, M.M.S., HARMANI, N.M.S., MASSUNAGA, P.N.T., BÜRER, S.P.,
PORTO, V.A.R. & UIEDA, W. 1998. Morcegos em áreas urbanas e
rurais: Manual de manejo e controle. 2ªEd. Fundação Nacional de
Saúde. 117p.
BREDT, A., UIEDA, W. & MAGALHÃES, E.D. 1999. Morcegos cavernícolas da
região do Distrito Federal. Centro-oeste do Brasil (Mammalia,
Chiroptera). Revista Brasileira de Zoologia 16(3): 731-770.
BROOKE, A.P. 1987. Tent construction and social organization in Vampyressa
nymphaea (Chiroptera: Phyllostomidae) in Costa Rica. Journal of
Tropical Ecology 3(2): 171-175.
COELHO, D.C. 2005. Ecologia e conservação da quiropterofauna no
corredor Cerrado-Pantanal. Tese de Doutorado. Universidade de
Brasília. 116p.
ESBÉRARD, C.E.L. 2003. Armadilha para retirada de morcegos abrigados em
telhados. Chiroptera Neotropical 9(1-2): 164-166.
45

ESBÉRARD, C.E.L., MOTTA, A.G., OLIVEIRA, D.M., AREAS, A.F., RODRIGUES,
R.T.V. & BERGALLO, H.G. 2003. Observação de fidelidade ao abrigo em
Molossus rufus no Estado do Rio de Janeiro, sudeste do Brasil.
Chiroptera Neotropical 9(1-2): 175-178.
ESBÉRARD, C.E.L., MOTTA, J.A. & PERIGO, C. 2005. Morcegos cavernícolas da
Área de Proteção Ambiental (APA) Nascentes do Rio Vermelho, Goiás.
Revista Brasileira de Zoociências 7(2): 311-325.
EZZELL, C. 1992. Caves creatures. Science News 141(6): 88-90.
FENTON, M.B., VONHOF, M.J., BOUCHARD, S., Gill, S.A., JOHNSTON, D.S., REID,
F.A., RISKIN, D.K., STANDING, K.L., TAYLOR, J.R. & WAGNER, R. 2000.
Roosts used by Sturnira lilium (Chiroptera: Phyllostomidae) in Belize.
Biotropica 32(4a): 729-733.
GRAHAM, G.L. 1994. Bats of the world. A golden guide from St. Martin‘s
Press. 160p.
GREENHALL, A.M., JOERMANN, G. & SCHMIDT, U. 1983. Desmodus rotundus.
Mammalian Species 202: 1-6.
HOOD, C.S. & JONES JR., J.K. 1984. Noctilio leporinus. Mammalian Species
216: 1-7.
JOHNSON, J.B., WOOD, P.B. & EDWARDS, J.W. 2006. Are external mine
entrance characteristics related to bat use? Wildlife Society Bulletin
34(5): 1368-1375.
KUNZ, T.H. 1982. Roosting ecology of bats. In: KUNZ, T.H. (Eds). Ecology of
bats. Plenum Press. p.1-55.
46

KUNZ, T.H. & MCCRACKEN, G.F. 1994. Tents and harems: alteration of leaves
by foliage roosting bats. Living World 1993-1994: 32-37.
KUNZ, T.H. & MCCRACKEN, G.F. 1996. Tents and harems: Apparent defence of
foliage roost by tent-making bats. Journal of Tropical Ecology 12(1):
121-137.
KUNZ, T.H., MARTY, S.F., BROOKE, A.P. & MCCRACKEN, G.F. 1994.
Convergence in tent architecture and tent-making behavior among
neotropical and paleotropical bats. Journal of Mammalian Evolution
2(1): 57-78.
LEE, Y.F. & MCCRACKEN, G.F. 2002. Foraging activity and food resourse use
od Brazilian free-tailed bats, Tadarida brasiliensis (Molossidae).
Ecoscience 9(3): 306-313.
NOGUEIRA, M.R. & POL, A. 1998. Observações sobre os hábitos de
Rhynchonycteris naso (Wied-Neuwied, 1820) e Noctilio albiventris
Desmarest, 1818 (Mammalia, Chiroptera). Revista Brasileira de
Biologia 58(3): 473-480.
NOGUEIRA, M.R., PERACCHI, A.L. & MORATELLI, R. 2007. Subfamília
Phyllostominae. In: REIS, N.R., PERACCHI, A.L. PEDRO, W.A. & LIMA, I.P.
(Eds). Morcegos do Brasil. Editora da Universidade Estadual de
Londrina. p.67-97.
NOVAES, R.L.M., OLIVEIRA, C.C.N., MENEZES JR., L.F., DUARTE, A.C. no prelo.
Dieta de Artibeus lituratus (Olfers, 1818) (Mammalia, Chiroptera) em
área urbana: frugivoria e novo registro de folivoria. Revista Brasileira
de Zoociências.
47

PERACCHI, A.L., LIMA, I.P., REIS, N.L., NOGUEIRA, M.L. & ORTÊNCIO-FILHO, H.
2006. Ordem Chiroptera. In: REIS, N.R., PERACCHI, A.L., PEDRO, W.A. &
LIMA, I.P. (Eds). Mamíferos do Brasil. Editora da Universidade
Estadual de Londrina. p.153-230.
REGIDOR, H., MOSA, S. & NUÑEZ, A. 2003. Confinamiento de una colonia de
Tadarida brasiliensis, una alternativa de manejo compatible con la
conservación. Chiroptera Neotropical 9(1-2): 157-162.
REIS, N.R., LIMA, I.P., PERACCHI, A.L. 2002. Morcegos (Chiroptera) da área
urbana de Londrina, Paraná - Brasil. Revista Brasileira de Zoologia
19(3): 739-746.
SIMMONS, N.B. & VOSS, R.S. 1998. The mammals of Paracou, French Guiana:
A neotropical lowland rainforest fauna part 1. Bats. Bulletin of the
American Museum of Natural History 237: 219p.
TRAJANO, E. & GIMENEZ, E.A. 1998. Bat community a cave from Eastern
Brazil, including a new record of Lionycteris (Phyllostomidae,
Glossophaginae). Study of Neotropical Fauna & Environment 33:
69-75.
YEE, D.A. 2000. Peropteryx macrotis. Mammalian Species 643: 1-4.
ZORTÉA, M. 2007. Subfamília Stenodermatinae. In: REIS, N.R., PERACCHI, A.L.
PEDRO, W.A. & LIMA, I.P. (Eds). Morcegos do Brasil. Editora da
Universidade Estadual de Londrina. p.107-128.
48

VI
REPRODUÇÃO

INTRODUÇÃO
Os morcegos, assim como outros mamíferos placentados, apresentam
desenvolvimento do embrião no corpo da fêmea, o que ofere condições
adequadas ao seu desenvolvimento como temperatura, fluxo constante de
nutrientes e proteção imunitária (NEUWEILER, 2000). Algumas espécies
possuem adaptações de armazenamento de esperma para induzir a época de
fecundação e o nascimento do filhote em períodos sazonais favoráveis
(BALMORI, 1999). Após o nascimento, o cuidado parental e a lactação
demandam, juntamente com a gravidez, um alto custo energético pela fêmea,
necessitando um incremento de energia durante o período de gestação e
lactação (VOIGT, 2003). Por esta razão, os ciclos reprodutivos nos quirópteros
parecem estar associados à disponibilidade de alimentos e às condições do
ambiente no qual estão inseridos (ZORTÉA, 2003; OLIVEIRA, 2007).
O aparelho reprodutor dos morcegos é bastante similar a dos outros
mamíferos, no entanto possui algumas peculiaridades surgidas de adaptações
fisiológicas e etológicas (BALMORI, 1999). Dentro do grupo dos quirópteros há
tantas diferenças reprodutivas de acordo com as espécies e famílias que pode-
se dizer que esta Ordem retém grande parte das variações reprodutivas da
Classe Mammalia (FRAZER, 1969; BALMORI, 1999). A maturação das gônadas
acontece graças aos hormônios gonadotrópicos secretados pelo glôbulo interior
da hipófise, que por sua vez são controladas pelos centros neurosecretores do
cérebro (hipotálamo). Os ciclos reprodutivos e as características sexuais
secundárias estão regulados da mesma forma, influenciando os demais
estímulos externos através do córtex cerebral (KOLB, 1971; BALMORI, 1999).
O orgão reprodutor dos machos compreende um par de testículos, vários
pares de glândulas, um sistema condutor de esperma e um orgão copulador
(pênis). Na maior parte das espécies os testículos só descem ao escroto no
período reprodutivo para facilitar a espermatogênese, permanecendo interno
em uma cavidade abdominal durante a maior parte do repouso sexual. O orgão
reprodutor feminino é formado por ovários, ovidutos, útero e vagina. Possuem
dois ovários como os outros mamíferos e o útero é bastante variável de acordo
com a espécie (BALMORI, 1999).
50

Muitos morcegos possuem também glândulas odoríferas que são usadas
em atividade de acasalamento (PERACCHI et al., 2006). Além disso, em muitas
espécies há formações sociais de grupos reprodutivos e de haréns (KUNZ et al.,
1983), caracterizados pela hierarquia de dominância (WILKINSON, 1990).
A maior parte do conhecimento sobre a reprodução dos quirópteros
neotropicais está concentrado na América Central, em estudos de FLEMING et
al. (1972), GANNON & WILLIG (1992), TSCHAPKA (2005) e CHAVERRI & KUNZ
(2006). No Brasil, estudos no Cerrado e Caatinga foram conduzidos por WILLIG
(1985), ZORTÉA (2003), e na Mata Atlântica por MELLO & FERNANDEZ (2000),
GOMES & UIEDA (2004), COSTA et al. (2007). O período reprodutivo dos
quirópteros neotropicais é variável de acordo com o ambiente e a espécie, com
morcegos se reproduzindo uma ou mais vezes por ano.
MONOESTRIA
A monoestria é caracterizada por um único pico de nascimento de filhores
por ano para uma determinada espécie (RACEY & ENTWISTLE, 2000).
Normalmente esse padrão reprodutivo está ligado às condições ambientais ou
fisiológicas, acontecendo em um períodos determinados e por isso é chamada
de monoestria sazonal.
A monoestria sazonal já foi registrada, entre outras espécies, para o
morcego fitófago Anoura geoffroyi (Phyllostomidae), estando diretamente
interligada à variação climática e disponibilidade de recursos (ZORTÉA, 2003).
WILLIG (1985) também encontrou monoestria sazonal para o morcego píscivoro
Noctilio leporinus.
POLIESTRIA
De maneira geral, a poliestria pode ser subdividida em três tipos
diferentes. Poliestria bimodal e estro pós-parto, que é caracterizada pela
estação reprodutiva apresentando dois picos de nascimento em um mesmo ano
e a fêmea se apresentando fértil e receptiva logo após o parto; poliestria
bimodal, que apresenta estação reprodutiva com dois picos de nascimento por
51

ano e poliestria multimodal, apresentando um comportamento contínuo, com
reprodução durante todo o ano (FLEMING et al., 1972; WILSON, 1973;
NEUWEILER, 2000; RACEY & ENTWISTLE 2000). A maior parte dos morcegos
fitófagos apresenta poliestria bimodal (WILLIG, 1985; TSCHAPKA, 2005;
CHAVERRI & KUNZ, 2006). Já morcegos hematófagos apresentam poliestria
assincrônica, em um padrão de tempo indefinido (TADDEI et al., 1991; ALENCAR
et al., 1994).
GESTAÇÃO
Normalmente as fêmeas têm um único filhote por parição, mas fêmeas de
alguns vespertilionídeos, especialmente o gênero Lasiurus, podem parir de
dois a cinco filhotes em uma única parição (PERACCHI et al., 2006).
Morcegos fitófagos costumam ter uma gestação de três a cinco meses,
variando de acordo com a espécie. Já morcegos insetívoros possuem gestações
mais curtas, de dois e três meses. Os morcegos hematófagos são os que
possuem a gestação mais longa, em torno de sete meses (SCHMIDT, 1988),
talvez relacionado ao seu metabolismo. A grande maioria das espécies dá a luz
a um filhote por gestação, entretanto há alguns registros de gêmeos, apesar de
raros (PERACCHI et al., 2006).
Fêmea grávida de Glossophaga soricina Foto: Roberto L.M. Novaes
Morcegos dão a luz à filhotes pesando de 15% a 25% do peso da mãe, considerados grandes para recém nascidos. O filhote nasce sem pêlos e sem capacidade de vôo e de se defenderer sozinho, e por isso permanece agarrado ao corpo da mãe. Com cerca de seis semanas o filhote já adqüire capacidade de vôo. Morcegos alcançam a maturidade sexual entre um e dois anos de vida, dependendo da espécie.
52

REFERÊNCIAS BIBLIOGRÁFICAS
ALENCAR, A.O., SILVA, G.A.P., ARRUDA, M.M., SOARES, A.J. & GUERRA, D.Q.
1994. Aspectos biológicos e ecológicos de Desmodus rotundus (Chiroptera)
no nordeste do Brasil. Pesquisa Veterinária Brasileira 14(4): 95-103.
BALMORI, A. 1999. La reproducción en los quiropteros. Galemys, Revisiones
en Mastozoologia 11(2): 17-34.
CHAVERRI, G. & KUNZ, T. H. 2006. Roosting ecology of the tent-roosting bat
Artibeus watsoni (Chiroptera: Phyllostomidae) in southwestern Costa
Rica. Biotropica 38(1): 77-84.
COSTA, L.M., ALMEIDA, J.C. & ESBÉRARD, C.E.L. 2007. Dados de reprodução de
Platyrrhinus lineatus em estudo de longo prazo no Estado do Rio de
Janeiro (Mammalia, Chiroptera, Phyllostomidae). Inheringia, Série
Zoologia 97(2): 152-156.
FLEMING, T.H., HOOPER, E.T. & WILSON, D.E. 1972. Three Central American
bat communities: structure, reproductive cycles and movement patterns.
Ecology 53(4): 555-569.
FRAZER, J.F.D. 1969. Los ciclos sexuales de los vertebrados. Labor. 141p.
GANNON, M.R. & WILLIG, M.R. 1992. Bat reproduction in the Luquillo
experimental forest of Puerto Rico. The Southern Naturalist 37(4):
414-419.
GOMES, M.N. & UIEDA, W. 2004. Abrigos diurnos, composição de colônias,
dismorfismo sexual e reprodução do morcego hematófago Desmodus
rotundus (E. Geoffroy) (Chiroptera, Phyllostomidae) no Estado de São
Paulo, Brasil. Revista Brasileira de Zoologia 21 (3): 629-638.
53

KOLB, E. (Eds). 1971. Fisiología veterinaria. Acribia. 122p.
KUNZ, T.H., AUGUST, P.Y. & BURNETT, C.D. 1983. Harem social organization
in cave roosting Artibeus jamaicensis (Chiroptera: Phyllostomidae).
Biotropica 15(2): 133-138.
MELLO, M.A.R. & FERNANDEZ, F.A.S. 2000. Reproductive ecology of the bat
Carollia perspicillata (Chiroptera, Phyllostomidae) in a fragment of the
Brazilian Atlantic Coastal forest. Säugetierkunde 65: 340-349.
NEUWEILER, G. 2000. The biology of bats. New York: Oxford University
Press. 310p.
OLIVEIRA, C.C.N. 2007. Aspectos reprodutivos de Artibeus planirostris
Spix, 1823 no Campus do Pici, Fortaleza, Ceará. Monografia de
graduação. Universidade Federal do Ceará. 39p.
PERACCHI, A.L., LIMA, I.P., REIS, N.L., NOGUEIRA, M.L. & ORTÊNCIO-FILHO, H.
2006. Ordem Chiroptera. In: Reis, N.R., Peracchi, A.L., Pedro, W.A. &
Lima, I.P (Eds). Mamíferos do Brasil. Editora da Universidade
Estadual de Londrina. p.153-230.
RACEY, P.A. & ENTWISTLE, A.C. 2000. Life-history and reproductive strategies
of bats.. In: CRICHTON, E.C. & KRUTZCHS, P.H. (Eds). Reproductive
biology of bats. Academic Press. p.607-626.
SCHMIDT, C. 1988. Reproduction. In: GREENHALL, A.M. & SCHMIDT, U. (Eds).
Natural history of vampire bats. CRC Press. 246p.
TSCHAPKA, M. 2005. Reproduction of the bat Glossophaga commissarisi
(Phyllostomidae: Glossophaginae) in the Costa Rican Rain Forest during
frugivorous and nectarivorous periods. Biotropica 37(3): 409-415.
54

TADDEI, V.A., GONÇALVES, C.A., PEDRO, W.A., TADEI, W.J., KOTAIT, I. &
ARIETA, C. 1991. Distribuição do morcego vampiro Desmodus
rotundus no Estado de São Paulo e a raiva dos animais
domésticos. Impressos Especiais da CATI. 107p.
VOIGT. C.C. 2003. Reproductive energetics of the nectar-feeding bat
Glossophaga soricina (Phyllostomidae). Journal of Composition and
Physiology B 173: 79-85
WILKINSON, G.S. 1990. Food sharing in vampire bats. Scientific American
262(2): 64-70.
WILLIG, M.R. 1985. Reproductive patterns of bats from caatingas and cerrado
biomes in Northeast Brazil. Journal of Mammalogy 66(4): 668-681.
WILSON, D.E. 1973. Bat faunas: a trophic comparison. Systematic Zoology
22(1): 4-29
ZORTÉA, M. 2003. Reproductive patterns and feeding habits of three
nectarivorous bats (Phyllostomidae: Glossophaginae) from the brazilian
cerrado. Brazilian Journal of Biology 63(1): 159-168.
55

VII
CONSERVAÇÃO

INTRODUÇÃO
A região neotropical é considerada a mais biodiversa do planeta, inclusive
em quirópteros, com cerca de 290 espécies (HUTSON et al., 2001). Mesmo assim
é a que sofre mais com a fragmentação florestal, redução de ecossistemas e
destruição de hábitats por influência antrópica, feita através da extração de
madeira e outros recursos naturais, expansão agropecuária e especulação
imobiliária.
OCHOA (2000) indica que a extração de madeiras em regiões neotropicais
afeta consideravelmente a diversidade de mamíferos, e se torna ainda mais
acentuado em comunidades de morcegos. Segundo REIS et al. (2003), a
fragmentação florestal é uma das principais causas da redução da diversidade
de quirópteros.
HUTSON et al. (2001) constataram que para a região neotropical, 45
espécies de morcego são consideradas vulneráveis, 10 estão em perigo e uma
está criticamente em perigo de extinção.
STATUS FAMÍLIA VU EP CR
Emballonuridae 2 1 0 Mormoopidae 1 0 0 Phyllostomidae 24 4 0 Natalidae 1 0 0 Furipteridae 1 0 0 Thyropteridae 1 0 0 Vespertilionidae 11 3 1 Molossidae 4 2 0
TOTAL 45 10 1
Tabela feita a partir dos dados indicados por HUTSON et al. (2001). Status de conservação e nº de espécies: Legenda:
VU = Vulnerável EP = Em perigo CR = Criticamente em perigo
57

CONSERVAÇÃO DOS MORCEGOS DO BRASIL
O Brasil é o quinto maior país do mundo e ocupa quase metade da
América do Sul, sendo o primeiro país em diversidade biológica do planeta
(MITTERMEIER et al., 1997), acolhendo cerca de 14% da biota mundial
(LEWINSOHN & PRADO, 2002). Aqui se encontra também a maior diversidade
de mamíferos (REIS et al., 2006). Porém, todo esse patrimônio biológico corre
sérios riscos, já que as aceleradas taxas de extinção de espécies, muitas antes
mesmo de serem conhecidas pela ciência, acaba por reduzir a diversidade
gênica e ameaçar o equilíbrio ecológico (BERGALLO et al., 2000; RAW, 2003;
MACHADO et al., 2005).
Das 168 espécies de morcegos registradas no Brasil (MIRANDA et al.,
2007; REIS et al., 2007), cinco são consideradas vulneráveis pelo IBAMA
(2003), embora AGUIAR & TADDEI (1995) reconheçam nove espécies brasileiras
como vulneráveis e sugiram a sua inclusão na lista oficial. Além disso há
outras 26 espécies na categoria Deficientes em dados (MACHADO et al., 2005),
e esses números podem representar uma subestimativa dos números reais de
espécies ameaçadas de extinção, já que aspectos importantes da taxonomia e
biologia dos quirópteros ainda é desconhecido ou mal compreendido. Essa
escassez de informações constitui forte limitação às iniciativas de conservação
das espécies (DIAS, 2007).
A maior parte das espécies ameaçadas de extinção no Brasil, são
endêmicas e pertencentes à família Phyllostomidae, uma das famílias de
maior importância ecológica para o Brasil (PERACCHI et al., 2006; REIS et al.,
2007). De maneira geral, as principais ameaças aos morcegos brasileiros são
constituídas pelo desmatamento e fragmentação florestal (REIS et al., 2003),
na Mata Atlântica pela especulação imobiliária e plantio de monoculturas; no
Cerrado e Pantanal pela expansão agropecuária e na Amazônia pela extração
de madeira e outros recursos naturais e queimadas para implantação de
pastagens.
58

A tabela abaixo mostra a lista das espécies de morcegos ameaçados de extinção do Brasil, informando o hábito alimentar, status e a consideração por autor. Legenda: VU = vulnerável.
Considerados por:
Família / Espécie Hábito
alimentar Status
AGUIAR & TADDEI (1995)
IBAMA (2003)
Emballonuridae Saccopteryx gymnura Insetívoro VU X Phyllostomidae Chiroderma doriae Frugívoro VU X Lichonycteris obscura Nectarívoro VU X Lonchophylla bokermanni Nectarívoro VU X X Lonchophylla dekeyseri Nectarívoro VU X Platyrrhinus recifinus Frugívoro VU X X Vampyrum spectrum Carnívoro VU X Vespertilionidae Lasiurus ebenus Insetívoro VU X X Lasiurus agregius Insetívoro VU X Myotis ruber Insetívoro VU X X
Total: 10 espécies
Das 10 espécies consideradas como ameaçadas de extinção por AGUIAR & TADDEI (1995) e IBAMA (2003), quatro são endêmicas do Brasil, Lonchophylla
bokermanni, Lonchophylla dekeyseri, Platyrrhinus recifinus e Lasiurus ebenus (REIS et al., 2007) e por isso se encontram em condições ainda mais delicadas.
As espécies endêmicas de Mata Atlântica e Cerrado, como Platyrrhinus recifinus e Lonchophylla dekeyseri merecem especial atenção, já que porções pequenas destes ecossistemas se encontram protegidas, enquanto a maior parte está em declínio e sofrendo grande ação antrópica.
Platyrrhinus recifinus Foto: Roberto L.M. Novaes
59

REFERÊNCIAS BIBLIOGRÁFICAS
AGUIAR, L.M.S. & TADDEI, V.A. 1995. Workshop sobre a conservação dos
morcegos brasileiros. Chiroptera Neotropical 1(2): 24-29.
BERGALLO, H.G., ROCHA, C.F.D., ALVES, M.A.S., VAN SLUYS, M. 2000. A fauna
ameaçada de extinção do Estado do Rio de Janeiro. Editora da
Universidade do Estado do Rio de Janeiro. 168p.
DIAS, D. 2007. Quirópteros da Reserva Biológica do Tinguá, Nova
Iguaçu, Rio de Janeiro, Brasil (Mammalia, Chiroptera). Tese de
Doutorado. Universidade Federal Rural do Rio de Janeiro. 108p.
HUTSON, A.M., MICKLEBURGH, S.P. & RACEY, P. 2001. Microchiropteran
bats: Global status survey and conservation action plan.
IUCN/SSC Chiroptera Specialist Group. IUCN. X+258p.
IBAMA 2003. Lista das espécies da fauna brasileira ameaçadas de
extinção. Anexo à Instrução Normativa nº3, de 27 de maio de 2003, do
Ministério do Meio Ambiente, Brasília, Brasil.
LEWINSOHN, T.M. & PRADO, P.I. 2005. How many species are there in Brazil?
Conservation Biology 19(3): 619-624.
MACHADO, A.B.M., MARTINS, C.S. & DRUMMOND, G.M. 2005. Lista da fauna
brasileira ameaçada de extinção: incluindo as listas das espécies
quase ameaçadas e deficientes de dados. Fundação Biodiversitas.
160p.
MIRANDA, J.M.D., AZEVEDO-BARROS, M.F.M. & PASSOS, F.C. 2007. First record
of Histiotus laephotis Thomas (Chiroptera, Vespertilionidae) from Brazil.
Revista Brasileira de Zoologia 24(4): 1188-1191.
60

MITTERMEIER, R.A., WERNER, T., AYRES, J.M. & FONSECA, G.A.B. 1997. O país
da megadiversidade. Ciência Hoje 14: 20-27.
OCHOA, J. 2000. Efectos de la extracción de maderas sobre la diversidad de
mamíferos pequeños en bosques de tierras bajas de la Guayana
Venezolana. Biotropica 32(1): 146-164.
PERACCHI, A.L., LIMA, I.P., REIS, N.L., NOGUEIRA, M.L. & ORTÊNCIO-Filho, H.
2006. Ordem Chiroptera. In: REIS, N.R., PERACCHI, A.L., PEDRO, W.A. &
LIMA, I.P (Eds). Mamíferos do Brasil. Editora da Universidade
Estadual de Londrina. p.153-230.
RAW, A. 2003. A sistemática biológica no currículo universitário. Ciência
Hoje 32(190): 59-61.
REIS, N.R., BARBIERI, M.L.S., LIMA, I.P. & PERACCHI, A.L. 2003 . O que é
melhor para manter a riqueza de espécies de morcegos (Mammalia,
Chiroptera): um fragmento florestal grande ou vários fragmentos de
pequeno tamanho? Revista Brasileira de Zoologia 20(2): 225-230.
REIS, N.R., PERACCHI, A.L., PEDRO, W.A., LIMA, I.P. (Eds). 2007. Morcegos do
Brasil. Editora da Universidade Estadual de Londrina. 253p.
REIS, N.R., SHIBATTA, O.A., PERACCHI, A.L., PEDRO, W.A. & LIMA, I.P. 2006.
Sobre os mamíferos do Brasil. In: REIS, N.R., PERACCHI, A.L., PEDRO,
W.A. & LIMA, I.P. (Eds). Mamíferos do Brasil. Editora da Universidade
Estadual de Londrina. 17-25p.
61

VIII
TÉCNICAS DE COLETA

INTRODUÇÃO
Nos últimos anos o número de pesquisa envolvendo morcegos vem
crescendo significativamente, e com isso se faz necessária a adoção de métodos
de captura mais eficientes e obtenção de informações mais detalhadas.
Morcegos são capturados em vôo ou em seus abrigos e o sucesso da captura
está relacionado com o método usado de acordo com as características da
espécie que se deseja capturar (KUNZ & KURTA, 1988), além do conhecimento
sobre seus comportamentos e hábitos (DALQUEST & WALTON, 1970).
Normalmente o método usado para captura de morcegos em regiões
tropicais é feito através de redes de neblina (FINDLEY & WILSON, 1983;
HEIDEMAN & HEANY, 1989). No entando, outros métodos também são usados
como as harpas ou harp traps (KUNZ & ANTHONY, 1977). Por possuírem alto
custo e serem de difícil manuseio as harpas são pouco utilizadas, e por isso
neste capítulo serão abordados apenas os métodos de captura com redes de
neblina.
MÉTODOS DE CAPTURA
Grande parte das pesquisas feitas nas regiões neotropicais utilizam as
redes de neblina como o principal método de captura (HODGKISON et al., 2002),
e alguns estudos indicam que o sucesso desse método está ligado com à altura
em que as redes são armadas em relação ao solo (COSSON, 1995; BERNARD,
2001; KALKO & HANDLEY, 2001). Algumas espécies costumam voar acima da
copa das árvores enquanto outras forrageiam próximo ao solo (PERACCHI et al.,
2006). É partir desta análise que devem ser escolhidos o método e o local onde
as redes de neblina serão armadas.
As redes são utilizadas, na maioria das vezes, para capturar morcegos
durante o vôo, ou em menor freqüencia saindo de seus abrigos (KUNZ &
KURTA, 1988). Essas redes são de baixo peso e compactas, além de serem de
fácil transporte e montagem em campo (KUNZ & KURTA, 1988). No entanto,
HANDLEY (1967) comenta que o seu grande potencial como ferramenta para
estudos ecológicos é mal explorado.
63

Redes de neblina são feitas de malha de nylon extremamente fino (cerca
de 20mm) e coloração escura, sendo a maioria de cor preta. São dividas em
quatro compartimentos horizontais, formando espécies de bolsas onde o
morcego fica preso. As redes possuem amarras em suas extremidas laterais
que são colocadas em varas na vertical para a sua sustentação. As redes mais
usadas possuem nove metros de comprimento por dois e meio de altura,
embora redes maiores ou menores também possam ser usadas. Se utilizadas
com cuidado e guardadas de forma adequada costumam ser duráveis.
Ilustração e foto das bolsas da rede de neblima usada para captura de morcego durante o vôo. O animal fica preso dentro do compartimento, onde parmanece até a sua retirada. As vistorias de rede devem ser feitas em intervalos de tempo curtos, para evitar que o animal se enrosque demais na rede, facilitando sua retirada, se o intervalo de vistoria for muito longo o animal pode danificar a rede ou até mesmo morrer por exaustão ou estresse.
Glossophaga soricina Foto: Roberto L.M. Novaes Adaptado de KUNZ & KURTA (1988)
A rede é colocada na vertical, presa com ajuda de varas. As bolsas, como evidenciado na figura, ajudam a prender o morcego. Ilustração adaptada de JONES et al. (1996)
64

As redes podem ser armadas em diferentes locais, estando relacionados à
espécie que se deseja capturar ou com o tipo de trabalho que ser queira fazer.
A maior parte dos trabalhos de diversidade feitos em áreas florestais na região
neotropical são conduzidos com uso de redes de neblina armadas em trilhas na
mata, clareiras ou em frente à abrigos previamente identificados (SIMMONS &
VOSS, 1998; FENTON et al., 2001; ESBÉRARD et al., 2006). Mesmo assim, as
redes de neblina podem ser armadas em muitos ambientes como bordas de
mata, sobre cursos d’ água, próximos a pontos de alimentação e até mesmo na
copa das árvores (KUNZ & KURTA, 1988).
Trilhas, clareiras e bordas de mata.
Este método é especialmente eficaz para captura de morcegos fitógafos, já
que estes costumam forragear mais baixo, em oposição aos morcegos
insetívoros que costumam voar mais alto (BERNARD, 2001; PERACCHI et al.,
2006). As redes de neblina são armadas perpendicularmente às trilhas,
cruzando clareiras ou paralelamente às bordas de mata, visando otimizar a
captura de morcegos que normalmente utilizam esses espaços como rotas de
vôo.
Em ambientes florestais as redes são armadas normalmente sob árvores, o que é especialmente eficaz para a captura de morcegos frugívoros. Já a maior parte das espécies insetívoras forrageiam acima da copa das árvores, além disso possuem um ecolocalizador mais sensível, sentindo a rede com mais facilidade, dificultando assim a sua captura.
Adaptado de KUNZ & KURTA (1988) Adaptado de JONES et al. (1996)
65

Capturas em abrigos
A maior parte das coletas feitas em abrigos são destinadas à captura de
morcegos insetívoros devido à dificuldade de capturá-los durante o vôo. Pórem
morcegos de todos os hábitos alimentares são capturados em seus abrigos
(SIMMONS & VOSS, 1998). Este método consiste na colocação da rede em frente
à abertura do abrigo, natural ou artificial, na hora de saída dos morcegos. Esta
forma de captura é especialmente importante em levantamento de espécies de
determinadas áreas, pois acaba por otimizar as capturas, incrementando o
número de espécies animalívoras, mais difíceis de serem capturadas em vôo
livre.
Para a captura de morcegos em abrigos é necessário que as principais saídas do refúgio sejam cercadas por redes de neblina. As figuras ilustram o cercamento com redes de neblina de uma casa usada como abrigo por morcegos e colocação de redes ao redor de uma planta usada como refúgio. Em alguns casos a coleta de morcegos em abrigos pode ser feita manualmente ou com ajuda de puçá.
Ilustrações adaptadas de KUNZ & KURTA (1988).
66

Coleta sobre coleções de água
Muitos estudos indicam a associação de algumas espécies de morcegos à
cursos d’água (NOGUEIRA et al., 1998). Este comportamento é evidenciado
principalmente para espécies insetívoras (ADAMS et al., 2003) e em alguns
casos a água se torna um fator primordial para a ocorrência de uma
determinada espécie (HOOD & JONES JR., 1984; MEYER et al., 2005). Por isso a
coleta sobre cursos d’água ou em torno de coleções de água torna-se um
método muito importante, principalmete para estudos de diversidade de
espécies. Este método consiste em armar a rede de neblina cruzando um
determinado espelho d’água, paralelamente ou perpendicularmente, ou rodeá-
lo com redes de neblina.
Existem diversas formas de se armar redes sobre cursos de água, variando conforme as características do ambiente. As metodologias usadas podem influenciar diretamente o número de captura e até mesmo a diversidade de espécies capturadas.
Ilustrações adaptadas de KUNZ & KURTA (1988).
67

Coletas de dossel
Parte das espécies de morcegos insetívoros, principalmente das famílias
Molossidae e Emballonuridae, forrageiam acima das copas das árvore
(PERACCHI et al., 2006). Até mesmo espécies animalívoras da família
Phyllostomidae costumam forregear na copa das árvores, se alimentando de
pequenos artrópodes no alto das folhas (NOGUEIRA et al., 2007). Para a captura
destes morcegos se faz necessário a adoção de um método diferenciado, que
consiste em armar as redes de neblina no alto das árvores. Para esté método é
necessário a utilização de cabos, cordas e roldanas, além de uma montagem
específica. Existem guias especiais que ensinam o passo-a-passo para a
montagem desse tipo de método (KUNZ & KURTA, 1988; VON MATTER, 2001;
HODGKISON et al., 2002).
Apesar da montagem deste método não ser fácil, ele garante bons resultados, já que possibilita a captura de espécies que são praticamente impossíveis de serem capturadas com redes armadas em solo.
Ilustração adaptada de VON MATTER (2001).
68

TRIAGEM
Após a captura do morcego, alguns procedimentos básicos são conduzidos
para obtenção de dados. Em estudos taxonômicos as espécies capturadas são
sacrificadas e levadas para o laboratório onde são fixadas em formol 10% e
conservadas em álcool 70% para posteriormente serem analisadas
morfologicamente (DIAS, 2007). Porém, em estudos ecológicos os morcegos
normalmente não são sacrificados, e as informações sobre eles são obtidas
ainda em campo.
Retirada do morcego da rede
O primeiro passo após a captura de um morcego é a sua retirada da rede.
Se feito corretamente, com cuidado e cautela, esse método não traz nenhum
prejuízo ao animal, nem danifica a rede. Para evitar a mordida, este
procedimento deve ser feito com a utilização de luvas de raspa de couro. O
morcego deve começar a ser retirado pelos mebros posteriores, e por último as
asas. Existem várias técnicas para manipulação de morcegos, porém eles só
devem ser exercidas por um profissional experiente e vacinado contra a raiva.
Retirada de Artibeus lituratus da rede. A retirada deve ser feita a partir dos membros inferiores a fim de facilitar a liberação do animal. O corte da rede dever ser feito em último caso, não sendo necessário na maioria dos casos.
Foto: Roberto L.M. Novaes
Método para imobilização segurando as asas para trás. Este método não machuca o animal e permite a sua manipulação sem permitir que ele se movimente demasiadamente e tente uma mordida defensiva.
Vampyressa pusilla
Foto: Roberto L.M. Novaes
69

Identificação e medição
Os morcegos ficam contidos dentro de sacos de pano até serem feitos os
processos de triagem, evitando assim o estresse do animal. A medição é feita
com ajuda de paquímetro e as medidas são anotadas em milímetros. Como já
citado nesta apostila, as medidas são caracteres primordiais para identificação
de uma espécie, e em campo as principais medições feitas são do antebraço,
tíbia, cabeça, comprimento total e falanges. Com as medidas anotadas é
possível se chegar até o nível específico de alguns grupos de morcego com a
utilizações de chaves de identificação, como as de VIZZOTO & TADDEI (1973),
EMMONS & FEER (1997) e GREGORIN & TADDEI, (2002).
Medição do antebraço de Artibeus lituratus. Essa medida pode ser essencial na determinação de uma espécie, porém, devido a sobreposição de medidas, em alguns casos somente as análises taxonômicas permitem se chegar ao nível mais específico.
Foto: Ana C. Façanha
Medição da cabeça de Platyrrhinus recifinus. Apesar desta medida não ser muito usual, ela pode dar indícios sobre a espécie ou idade do animal.
Foto: Roberto L.M. Novaes
70

Sexagem, estágio reprodutivo e determinação de idade
A determinação do sexo e estágio reprodutivo de morcegos é de grande
importância para o conhecimento dos períodos reprodutivos e suas relações
com as condições do ambiente em que vivem (OLIVEIRA, 2007). Segundo RACEY
(1988), as fêmeas são subdivididas em três grupos: Grávida, quando o
abdomêm está proeminente e o feto palpável; Lactante, quando as mamas
estão secretantes; e Inativas, quando não há nenhum desses caracteres
visíveis. Já os machos são classificados apenas quanto a posição dos testículos
(escrotado ou abdominal) (BALMORI, 1999). Já a idade é determinada através
do grau de ossificação das epífises (ANTHONY, 1988; COSSON et al., 1983).
Morcegos com epífises totalmente cartilaginosas são classificados como filhote
com epífises apresentando médio estágio de ossificação, porém com regiões
ainda cartilaginosas são classificados como jovem (ou subadulto) e os com
epífises totalmente ossificadas são classificados como adulto (KUNZ, 1988).
Artibeus lituratus com mamas lactantes. Normalmente essa característica é visível, no entanto poder haver a necessidade de palpação ou de leve pressão na mama.
Foto: Roberto L.M. Novaes
71

Marcação
Muitas pesquisas ecológicas utilizam marcações temporárias ou
permanentes. Este recurso é especialmente útil para estudos de movimentação
(BERNARD & FENTON, 2003) e deslocamento (COSTA et al., 2006).
Os métodos de marcação temporária mais utilizados são furos no
dactilopatágio, que de acordo com a posição indicam uma numeração e a
descoloração de pêlos, mais utilizada para estudos comportamentais (SILVA et
al., 2007).
Já as marcações permanentes mais utilizadas são as anilhas metálicas e
numeradas presas no antebraço no animal, as coleiras plásticas numeradas
(ESBÉRARD & DAEMON, 1999), e os métodos de marcação por chip ou
dispositivo guia e localização por radiotelemetria (MELLO et al., 2008). Esses
métodos costumam ser mais usuais, pois permitem avaliar padrões de
movimentação e migração, além de estimar a área e o tempo de vida de um
animal. É importante ressaltar que a marcação pode trazer prejuízos aos
animais, e a utilização de marcações permanentes deve ser feita com muito
critério por pesquisadores experientes. Os dados dos animais capturados e
marcados devem ser anotados em uma planílha de informações que deve ser
guardada e consultada após alguma recaptura.
Morcego marcado com coleira plástica numerada. Este método não costuma trazer grandes prejuízos ao animal, no entanto a sua colocação deve ser feita com cuidado e critério. Após a marcação os morcegos são liberados no próprio local.
Carollia perspicillata
Foto: Roberto L.M. Novaes
72

Planilha de informações para morcegos capturados
73

REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, R.A., PEDERSEN, S.C., THIBAULT, K.M., JADIN, J. & PETRU, B. 2003.
Calcium as a limiting resource to insectivorous bats: can water holes
provide a supplemental mineral source? Journal of Zoology 2006: 189-
194.
ANTHONY, E.L.P. 1988. Age determination in bats. In: KUNZ, T.H. (Ed).
Ecological and behavioral methods for the study of bats.
Smithsonian Instituition Press. p.47-58.
BALMORI, A. 1999. La reproducción en los quiropteros. Galemys, Revisiones
en Mastozoologia 11(2): 17-34.
BERNARD, E. 2001. Vertical stratification of bat communities in primary
forests of Central Amazon, Brazil. Journal of Tropical Ecology 17:
115-126.
BERNARD, E. & FENTON, M.B. 2003. Bat mobility and roosts in a fragmented
Landscape in Central Amazonia, Brazil. Biotropica 35(2): 262-277.
COSSON, J.M. 1995. Captures of Myonycteris torquata (Chiroptera,
Pteropodidae) in forest canopy in South Cameroon. Biotropica 27(3):
395-396.
COSSON, J.M., RODOLPHE, F. & PASCAL, M. 1983. Détermination deI'âge
individuel, croissance post-natale et ontogenèse précoce de Carollia
pespicillata (Linnaeus, 1758) (Chiroptera, Phyllostomidae). Mammalia
57: 565-578.
COSTA, L.M., PRATA, A.F.D., MORAES, D., CONDE, C.F.V., JORDÃO-NOGUEIRA,
T. & ESBÉRARD, C.E.L. 2006. Deslocamento de Artibeus fimbriatus sobre
o mar. Chiroptera Neotropical 12(2): 289-290.
74

DALQUEST, W.W. & WALTON, D.W. 1970. Diurnal retreats of bats. In:
SLAUGHTER, B.H. & WALTON, D.W. (Eds). About bats: a chiropteran
symposium. Southern Methodist University Press. p.162-187.
DIAS, D. 2007. Quirópteros da Reserva Biológica do Tinguá, Nova
Iguaçu, Rio de Janeiro, Brasil (Mammalia, Chiroptera). Tese de
Doutorado. Universidade Federal Rural do Rio de Janeiro. 108p.
EMMONS, L.H. & FEER, F. 1997. Neotropical rainforest mammals: a field
guide. 2ª Ed. The University of Chicago Press. 392p.
ESBÉRARD, C.E.L. & DAEMON, C. 1999. Um novo método para marcação de
morcegos. Chiroptera Neotropical 5(1-2): 116-117.
ESBÉRARD, C.E.L., JORDÃO-NOGUEIRA, T., LUZ, J.L., MELO, G.G.S., MANGOLIN,
R., JUCÁ, N., RAÍCES, D.S.L., ENRICE, M.C. & BERGALLO, H.G. 2006.
Morcegos da Ilha Grande, Angra dos Reis, RJ, sudeste do Brasil. Revista
Brasileira de Zoociências 8 (2): 147-153.
FENTON, M.B., BERNARD, E., BOUCHARD, S., HOLLIS, L., JOHNSTON, D.S.,
LAUSEN, C.L., RATCLIFFEE, J.M., RISKIN, D.K., TAYLOR, J.R. & ZIGOURIS, J.
2001. The bat fauna of Lamanai, Belize: roosts and trophic roles.
Journal Tropical Ecology 17: 511-524.
FINDLEY, J.S. & WILSON, D.E. 1983. Are bats rare in tropical Africa?
Biotropica 15: 299-303.
GREGORIN, R. & TADDEI, V. A. 2002. Chave artificial para a identificação de
Molossídeos brasileiros (Mammalia, Chiroptera). Mastozoología
Neotropical 9(1):13-32.
75

HANDLEY, C.O. 1967. Bats of the canopy of an Amazonian forest. Atas do
simpósio sobre biota Amazonica (Zoologia) 5: 211-215.
HEIDMAN, P.D. & HEANEY, L.R. 1989. Population biology and estimates of
abundance of fruit bats (Pteropodidae) in Philippind sub-montane rain
forest. Journal of Zoology 218: 565-586.
HODGKISON, R., AHMAD, D., BALDING, S., KINGSTON, T., ZUBAID, A. & KUNZ,
T.H. 2002. Capturing bats (Chiroptera) in tropical forest canopies. In:
Mitchell, A.W., Secoy, K. & Jackson, T. (Eds). The Global Canopy
Programme Handbook: Techniques of access and study in the
forest roof. Global Canopy Programme. p.160-167.
HOOD, C.S. & JONES JR., J.K. 1984. Noctilio leporinus. Mammalian Species
216: 1-7.
JONES, C., MCSHEA, W.J., CONROY, M.J. & KUNZ, T.H. 1996. Capturing
mammals. In: WILSON, D.E., NICHOLS, J., RUDRIN, R., COLE, R. & FOSTER,
M. (Eds). Measuring and monitoring biological diversity.
Smithsonian Institution Press. p.115-122.
KALKO, E.K.V. & HANDLEY, C.O. 2001. Neotropical bats in the canopy:
diversity, community structure, and implications for conservation. Plant
Ecology 153: 319-333.
KUNZ, T.H. 1988. Ecological and behavioral methods for the study of
bats. Smithsonian Institution Press. 533p.
KUNZ, T.H. & ANTHONY, E.L.P. 1977. On the efficiency of the Tuttle bat trap.
Journal of Mammalogy 58(3): 309-315.
76

KUNZ, T.H., & KURTA, A. 1988. Capture methods and holding devices. In:
KUNZ, T.H. (Ed). Ecological and behavioral methods for the study
of bats. Smithsonian Institution Press. p.1-29.
MELLO, M.A.R., KALKO, E.K.V. & SILVA, W.R. 2008. Movements of the bat
Sturnira lilium and its role as a seed disperser of Solanaceae in the
Brazilian Atlantic Forest. Journal of Tropical Ecology 24: 225-228.
MEYER, C.F.J., WEINBEER, M. & KALKO, E.K.V. 2005. Home-range size and
spacing patterns of Macrophyllum macrophyllum (Phyllostomidae)
foraging over water. Journal of Mammalogy 86(3): 587-598.
NOGUEIRA, M.R., PERACCHI, A.L. & MORATELLI, R. 2007. Subfamília
Phyllostominae. In: REIS, N.R., PERACCHI, A.L., PEDRO, W.A. & LIMA, I.P.
(Eds). Morcegos do Brasil. Editora da Universidade Estadual de
Londrina. p.61-97.
NOGUEIRA, M.R., POL, A. & PERACCHI, A.L. 1998. Use of water holes by
Stenodermatinae bats in western Amazonian Brazil. Bat Research
News 39(3): 95.
OLIVEIRA, C.C.N. 2007. Aspectos reprodutivos de Artibeus planirostris
Spix, 1823 no Campus do Pici, Fortaleza, Ceará, Brasil. Monografia
de graduação. Universidade Federal do Ceará. 39p.
PERACCHI, A.L., LIMA, I.P., REIS, N.L., NOGUEIRA, M.L. & ORTÊNCIO-Filho, H.
2006. Ordem Chiroptera. In: REIS, N.R., PERACCHI, A.L., PEDRO, W.A. &
LIMA, I.P (Eds). Mamíferos do Brasil. Editora da Universidade
Estadual de Londrina. p.153-230.
RACEY, P.A. 1988. Reproductive assessment in bats. In: KUNZ T.H. (Ed).
Ecological and behavioral methods for study of bats. Smithsonian
Institution Press. p.31-45.
77

SILVA, R.M., PEIXOTO, G., FRANÇA, D., LOURENÇO, E.C., COSTA, L.M. &
ESBÉRARD, C.E.L. 2007. Descoloração do pêlo como método de marcar
morcegos para observações comportamentais. Chiroptera Neotropical
13(2): 323-325.
SIMMONS, N.B. & VOSS, R.S. 1998. The mammals of Paracou, French Guiana:
A neotropical lowland rainforest fauna part 1. Bats. Bulletin of the
American Museum of Natural History 237: 219p.
Vizotto, L.D. & Taddei, V.A. 1973. Chave para determinação de quirópteros
brasileiros. Revista da Faculdade de Filosofia Ciências e Letras
São José do Rio Preto - Boletim de Ciências 1: 1- 72.
VON MATTER, S. 2001. Metodologia de montagem e manutenção de rede
suspensa ou bandeira em área de mata fechada: detalhes e dicas úteis. IX
Congresso Brasileiro de Ornitologia.
78

AGRADECIMENTOS
APOIO E PARCERIA
À Carolinne I. Dias Gomes pela leitura, sugestão e revisão dos textos. Ao Dr. Marco A.R. Mello (www.casadosmorcegos.org) por ceder algumas fotografias para ilustrar essa apostila. À toda equipe do Projeto Pró-Morcegos, em especial a Flávio Zen Mello por todo apoio à pesquisa e ação ambiental.
NOVAES, R.L.M. 2008. Morcegos neotropicais: biologia, ecologia e técnicas de coleta. Apostila básica. Projeto Pró-Morcegos. 85p.

Rua Quaxima, 45, Madureira. CEP 21310-250, Rio de Janeiro,
RJ, Brasil. Tel.: (21) 3624 0628
[email protected] www.promorcegos.org
ROBERTO LEONAN MORIM NOVAES Laboratório de Mastozoologia
Universidade Federal Rural do Rio de Janeiro. [email protected]

BIOLOGIA, ECOLOGIA E TÉCNICAS DE COLETA
Morcegos sempre foram vítimas de mitos e crendices populares, o que lhes
atribuiu má fama, criando uma corrente de preconceitos e inverdades em
torno deles. A sociedade moderna não consegue enxergar através do medo e
da aparência obscura desses animais, não percebendo o valor real que eles
têm para o equilíbrio da vida. Grande parte da dinâmica de regeneração
florestal acontece graças à dispersão de sementes feita por morcegos
frugívoros, e uma parcela significativa das pragas urbanas e agrícolas é
controlada por morcegos insetívoros. Esta apostila foi desenvolvida com o
intuito de demonstrar a importância ecológica e econômica dos morcegos.
Seu conteúdo é constituído por aspectos básicos sobre a biologia e ecologia
dos quirópteros da região neotropical, além de um capítulo sobre métodos
de coleta e pesquisa com morcegos, o que é de grande valia para novos
estudantes de Biologia. Desta forma, trabalhos de pesquisa e ação ambiental
estão sendo incentivados, revertendo a corrente de preconceito para a
corrente da consciência ambiental e do pensamento racional.
ROBERTO LEONAN MORIM NOVAES
MORCEGOS NEOTROPICAIS
Rio de Janeiro Tel.: (21) 3624 0628
[email protected] www.promorcegos.rg3.net


















