
O efeito do alumínio no crescimento e na actividade de (Na ... · requisitos necessários à...
62
Universidade de Aveiro 2007 Departamento de Biologia O efeito do alumínio no crescimento e na actividade de (Na + /K + )ATPase da Escherichia coli
Transcript of O efeito do alumínio no crescimento e na actividade de (Na ... · requisitos necessários à...

Universidade de Aveiro 2007
Departamento de Biologia
O efeito do alumínio no crescimento e na actividade de (Na+/K+)ATPase da Escherichia coli

Universidade de Aveiro 2007
Departamento de Biologia
Patrícia Moraes
O efeito do alumínio no crescimento e na actividade de (Na+/K+)ATPase da Escherichia coli
Dissertação apresentada à Universidade de Aveiro para cumprimento dos requisitos necessários à obtenção do grau de Mestre em Biologia Molecular e Celular, realizada sob a orientação científica da Doutora Virgília Sofia Almeida de Azevedo e Silva, Professora Auxiliar Convidada do Departamento de Biologia da Universidade de Aveiro e da Doutora Maria Paula Polónia Gonçalves, Professora Associada do Departamento de Biologia da Universidade de Aveiro.

o júri
presidente Prof. Dr. António Carlos Matias Correia Professor Associado com Agregação da Universidade de Aveiro
Prof. Dra. Maria Paula Polónia Gonçalves Professora Associada da Universidade de Aveiro (Co-orientadora)
Prof. Dr. Rui Pedro Ramos Cardoso Professor Auxiliar da Universidade de Aveiro
Prof. Dra. Virgília Sofia Almeida de Azevedo e Silva Professora Auxiliar Convidada da Universidade de Aveiro (Orientadora)

agradecimentos
À Prof. Dra. Virgília Sofia Almeida de Azevedo e Silva minha orientadora,
obrigada pelo constante apoio, sempre indicando a direcção a ser tomada
nos momentos de maior dificuldade e pelas valiosas sugestões dadas.
Agradeço, principalmente, pela sua dedicação depositada no meu trabalho
de dissertação.
À minha co-orientadora, Prof. Dra. Maria Paula Polónia Gonçalves, pela
colaboração, paciência e por seus conhecimentos repassados, durante
todo o desenvolvimento desse trabalho.
Aos colegas do laboratório de Microbiologia e Bioquímica, obrigada pela
ajuda constante.
Aos amigos distantes, que sempre mantive, obrigada pela força e pelo
estímulo que me deram, desde o início.
Aos amigos que fiz em Aveiro, que ficarão para sempre em minha
memória e principalmente em meu coração.
À minha eterna irmã e amiga Cintia, obrigada pelo incentivo e
principalmente por ter acreditado em mim!
Aos meus Pais devo um agradecimento especial com todo o amor, carinho
e compreensão, sei que mesmo não estanto presentes sempre deram seu
“tudo” por isso, esse sonho realizado também é um sonho vosso.
- Vocês foram o estímulo do início, uma ajuda constante, concreta e
infatigável.
Acalento a esperança de que tudo isto sirva para lhes mostrar não ter sido
perdida a educação, que se esforçaram por me proporcionar.
- Obrigada por vocês existirem!
Espero que Deus me dê a alegria de tê-los em minha vida por muito tempo
ainda.
A Deus, agradeço por Seus ensinamentos, por ter me dado a oportunidade
de estar aqui, por ajudar a superar minhas próprias e humanas limitações.
Obrigada por estar ao meu lado sempre, sei que se não fosse seu apoio
jamais poderia ter forças para caminhar até o final.

palavras-chave
Toxicidade do alumínio, Escherichia coli (estirpe JM 109), (Na+/K+)ATPase.
resumo
A toxicidade do alumínio está bem documentada em situações de exposição massiva ou prolongada. O alumínio é ubíquo no nosso ambiente e em condições acídicas a sua biodisponibilidade aumenta. O mecanismo de acção tóxica do alumínio continua por esclarecer. In vivo e in vitro o efeito inibitório do alumínio na (Na+/K+)ATPase dos mamíferos mantém as suas características principais, e parece envolver a redução do grau de oligomerização da enzima. A (Na+/K+)ATPase é uma proteína integrante da membrana plasmática que hidrolisa ATP e medeia o transporte activo de Na+ e K+ através da membrana celular de todos os eucariontes. Esta enzima também está presente na membrana interna da Escherichia coli (E. coli), uma bactéria Gram negativa, que se encontra no intestino grosso dos animais de sangue quente. Visto que a (Na+/K+)ATPase de diferentes espécies exibe sensibilidades distintas a alumínio, é imperativo descobrir qual a base molecular da susceptibilidade desta enzima a alumínio. O principal objectivo deste trabalho consistiu em determinar se o alumínio inibe ou não a (Na+/K+)ATPase da E. coli, pois a composição lipídica da sua membrana interna é diferente da da membrana plasmática das células de mamífero. Por exemplo, não há nem esteróis nem esteróides na membrana interna de E. coli, sendo a fluidez de membrana assegurada pela presença de ácidos gordos mais insaturados e de cadeia mais curta. A toxicidade de alumínio para a E. coli (estirpe JM 109) foi estudada em meio líquido de crescimento Boillon – Miller (pH 7,0) durante a fase estacionária a 37ºC. A adição de AlCl3, na gama de concentrações milimolares, reduziu drasticamente a capacidade de sobrevivência das células de E. coli (reflectida pela redução de UFC*mL-1 ao longo do tempo). A actividade de (Na+/K+)ATPase de esferoplastos preparados a partir de células de E. coli previamente expostas a AlCl3 9,5*10-3 M permaneceu idêntica à medida em condições controlo (esferoplastos preparados a partir de células de E. coli não expostas a AlCl3). Além do mais, não se observou inibição da (Na+/K+)ATPase induzida por alumínio quando se adicionou, directamente ao meio de reacção, AlCl3 (concentração final até 1,0*10-3 M). Em conclusão, a (Na+/K+)ATPase de E. coli não parece ser sensível a alumínio, apesar do alumínio ser tóxico para E. coli.

keywords
Aluminium toxicity, Escherichia coli (strain JM 109), (Na+/K+)ATPase.
abstract
Aluminium toxicity has been recognized in many situations where exposure is heavy or prolonged. Aluminium is ubiquitous in our environment and its bioavailability is increased under acidic conditions. The mechanism of toxic action of aluminium continues to remain poorly understood. The in vivo and in vitro inhibitory effect of aluminium on mammalian (Na+/K+)ATPase share the same main characteristics, which seems to involve the reduction of the degree of oligomerization of the enzyme. The (Na+/K+)ATPase is an integral membrane protein that hydrolyses ATP and mediates the active transport of Na+ and K+ across the plasma membrane of all eukaryotic cells. This enzyme is also present in the inner membrane of Escherichia coli (E. coli), a Gram negative bacterium that is commonly found in the lower intestine of warm-blooded animals. Given that differences on the sensitivity of the (Na+/K+)ATPase to aluminium exposure among species have been shown, it is imperative to figure out the molecular basis of the enzyme susceptibility to aluminium. The aim of this work is to investigate whether aluminium also inhibits the (Na+/K+)ATPase of E. coli, since the lipidic composition of its inner membrane differs from the plasma membrane of mammalian cell. For instance, there are no sterols or steroids in the inner membrane of E. coli, where membrane fluidity tends to be maintained by fatty acids with shorter chains and more unsaturated fatty acids. In Boillon – Miller growth liquid medium (pH 7.0) the toxicity of aluminium to E. coli (strain JM 109) has been studied during the stationary phase at 37ºC. Addition of AlCl3, in the millimolar range, drastically reduced the survival ability of E. coli (as reflected in the reduction of CFU*mL-1 over time). The (Na+/K+)ATPase activity of spheroplast prepared from E. coli cells previously exposed to 9.5*10-3 M AlCl3 remained identical to the measured in control conditions (spheroplast prepared from E. coli cells not exposed to AlCl3). Moreover, no aluminium-induced inhibition of the (Na+/K+)ATPase was observed when AlCl3 (final concentration up to 1.0*10-3 M) was added directly to the reaction medium. In conclusion, the (Na+/K+)ATPase of E. coli does not seem to be sensitive to aluminium, besides aluminium being toxic to E. coli.

Índice
Página
Abreviaturas 1
Lista de Figuras 2
1. Introdução 3
1.1. O alumínio 5
1.2. A Escherichia coli 5
1.3. Efeito do alumínio na Escherichia coli 8
1.4. A bomba de (Na+/K+)ATPase 9
1.5. Efeito do alumínio na actividade da (Na+/K+)ATPase 11
1.6. Objectivo do trabalho 11
2. Material e Métodos 15
2.1. Material biológico e meios de cultura 17
2.2. Preparação da cultura de Escherichia coli 17
2.3. Contagem das UFC 18
2.4. Preparação de esferoplastos 19
2.5. Quantificação da proteína 21
2.6. Medição da actividade da adenosinatrifosfatase 22
2.7. Análise de dados 22
3. Resultados e Discussão 25
3.1. Efeito do AlCl3 na capacidade de sobrevivência de E. coli 27
3.2. Efeito do AlCl3 na actividade da (Na+/K+)ATPase de E. coli 39
4. Conclusões 45
5. Bibliografia 49

1
Abreviaturas
ADN Ácido desoxirribonucleico
ATP Adenosina 5’- trifosfato
ATPase Adenosinatrifosfatase
D.O. Densidade óptica
E. coli Escherichia coli
EC50 Concentração de AlCl3 que produz 50% do efeito máximo
EDTA Ácido [etilenodiamino]tetracético
E.P. Erro padrão
HEPES Ácido N-[2-hidroxietil]piperazina-N-[2- etanossulfónico]
JM 109 Estirpe de Escherichia coli (espécie)
LB Meio líquido Boillon – Miller pH 7,0
M Taxa de morte
N Densidade das células viáveis
N0 Densidade (inicial) máxima observada
(Na+/K+)ATPase Adenosina 5’-trifosfatase dependente de Na+ e K+
Pi Fosfato inorgânico
R2 Quadrado do coeficiente de relação momentânea
rpm Rotações por minuto
SDS Dodecilsulfato de sódio
UFC Unidades formadoras de colónias

2
Lista de Figuras
Figura 1.1. Escherichia coli observada ao microscópio electrónico de varrimento.
Figura 1.2. Intermediários do transporte mediado pela (Na+/K+)ATPase segundo o modelo
de Alberts-Post.
Figura 1.3. Topologia da subunidade α1 da (Na+/K+)ATPase de carneiro.
Figura 2.1. Fotografias com os passos para preparação da cultura de E. coli.
Figura 2.2. Esquema do método de preparação de esferoplastos de E. coli.
Figura 2.3. Observação ao microscópio óptico de cultura de E. coli.
Figura 2.4. Observação ao microscópio óptico da preparação de esferoplastos de E. coli.
Figura 2.5. Equipamento usado para medir a actividade da (Na+/K+)ATPase.
Figura 2.6. Representação esquemática do modelo farmacodinâmico do efeito do AlCl3 na
cultura de E. coli em sistema fechado.
Figura 3.1. Curva de crescimento de E. coli em meio LB, monitorizada pelo método
turbidimétrico e por contagem de UFC.
Figura 3.2. Crescimento e declínio da população de E. coli em condição de cultura
controlo.
Figura 3.3. Efeito da exposição a diferentes concentrações de AlCl3 no número de células
viáveis de E. coli em cultura.
Figura 3.4. Morte de células de E. coli em cultura em função do tempo de exposição a
AlCl3.
Figura 3.5. Variação da taxa de morte em função da concentração de AlCl3.
Figura 3.6. Variação da área sob a curva de morte-tempo em função da concentração de
AlCl3.
Figura 3.7. Variação da área sob a curva de morte-tempo em função do tempo de
exposição a AlCl3.
Figura 3.8. Simulação do efeito do AlCl3 na dinâmica populacional de E. coli.
Figura 3.9. Registo da actividade ATPásica de esferoplastos de E. coli.
Figura 3.10. Efeito da concentração de proteína na actividade da (Na+/K+)ATPase.
Figura 3.11. Efeito in vivo do alumínio na actividade da (Na+/K+)ATPase e nas actividades
ATPásicas total e basal.
Figura 3.12. Efeito in vitro do alumínio na actividade da (Na+/K+)ATPase e nas
actividades ATPásicas total e basal.

3
1. Introdução

4

5
1.1. O alumínio
O alumínio, de símbolo químico Al, é um elemento metálico trivalente, leve, sólido
cristalino, de cor branca-prateada e sem odor característico; é o metal mais comum e o terceiro
elemento mais abundante da crosta terrestre. Este elemento químico é constituinte de solos
rochas, e minerais, como os feldspatos e a bauxite, entre outros. O alumínio é uma das espécies
mais reactivas que se conhece. Quando exposto ao oxigénio, forma-se rapidamente à sua
superfície uma película contínua e fina de óxido de alumínio, que limita a posterior corrosão do
alumínio metálico (Klaassen et al., 2003). O metal alumínio, simultaneamente maleável e dúctil,
para além da referida resistência à corrosão, possui outras propriedades, tais como alta
condutividade eléctrica e térmica, baixa densidade e boa reflectividade. Por conseguinte, são
numerosas as suas aplicações industriais (Bokovay, 1988).
A exposição humana ao alumínio ocorre por várias vias, das quais se salientam a oral,
inalatória e dérmica (Caballero et al., 2003). A principal fonte adicional de alumínio na dieta são
geralmente os aditivos alimentares. A utilização de alumínio na embalagem alimentar e no
fabrico de utensílios de cozinha também contribui para elevar os teores em alumínio nos
alimentos ingeridos. Por outro lado, compostos de alumínio são usados no tratamento da água da
rede pública, na preparação de medicamentos, vacinas e antitranspirantes (Miller, 1984; Driscoll,
1988; Domingo, 1993). A absorção do alumínio no tracto gastrointestinal é reduzida. A sua
absorção é favorecida em condições ácidas e na presença de ligandos complexantes que
aumentam a sua biodisponibilidade. Porém, quando os principais mecanismos que protegem
contra a acumulação de alumínio no organismo (barreira gastrointestinal e excreção renal) estão
comprometidos pode ocorrer intoxicação. Por exemplo, são bem conhecidas as consequências
clínicas da intoxicação por alumínio nos pacientes submetidos a diálise, que incluem anemia
microcítica, osteomalácia, osteodistrofia, encefalopatia e demência.
1.2. A Escherichia coli
A Escherichia coli (E. coli) é, talvez, o organismo mais conhecido em termos genéticos e
fisiológicos (Fig. 1.1). Esta bactéria é uma bactéria simbionte que se encontra no cólon e no
intestino grosso da maioria dos animais de sangue quente, incluindo o Homem. Por ouro lado, é
também a causa frequente de várias doenças infecciosas (intestinais e extra-intestinais). A
barreira entre comensalismo e a virulência é o resultado de um complexo equilíbrio entre o
estado do hospedeiro e a presença e expressão de factores de virulência nas bactérias (Berg,

6
1996). Outros parâmetros ambientais, bem como a quantidade do inóculo, podem influenciar o
curso de uma infecção (Ochman et al., 1984).
Figura 1.1. Escherichia coli observada ao microscópio electrónico de varrimento. A Escherichia
coli é uma bactéria bacilar Gram-negativa que pertence à família Enterobacteriaceae (http://pt.wikipedia.org/wiki/Escherichia_coli).
A E. coli é uma bactéria bacilar Gram-negativa, pertencente à família Enterobacteriaceae,
com aproximadamente 2 µm de comprimento e 0,8 µm de largura, que se divide por fissão
binária. O envelope de bactérias Gram-negativas é uma estrutura complexa (Glauert & Thornley,
1969). A membrana externa da E. coli, com cerca de 10-15 nm de espessura, é composta por um
folheto externo de lipopolissacarídeos, um folheto interno de fosfolípidos e proteínas, das quais
as porinas são as mais abundantes. A parede celular, com cerca de 1,3 nm de espessura, é
fundamentalmente constituída por peptidoglicano (lipoproteína de Braun), que se encontra ligado
covalentemente à membrana externa, conferindo forma à célula da E. coli. A membrana interna,
com cerca de 8 nm de espessura, é composta essencialmente por fosfolípidos e proteínas. A
fosfatidiletanolamina, o fosfatidilglicerol, a cardiolopina e a fosfatidilserina, na proporção de
75:18:5:2, são os fosfolípidos maioritários. A ausência de esteróis e esteróides é uma
característica que distingue esta membrana das membranas celulares dos eucariontes (Razin,

7
1975). A fluidez de membrana é, neste caso, assegurada pela presença maioritária de ácidos
gordos insaturados de cadeia curta. Finalmente, o espaço entre a membrana externa e interna,
periplasma, com cerca de 10 nm de espessura, contém ~80.000 proteínas essenciais para o
metabolismo e quimiotaxia.
As células da E. coli podem também apresentar forma esférica, quando perdem
parcialmente a parede celular. A bacteriólise ocorre por acção de autolisinas (autólise) ou por
acção de lisozimas (heterólise). A optimização das condições de preparação de esferoplastos de
E. coli por acção conjunta de lisozima e EDTA, um quelante de catiões divalentes, foi proposta
por Birdsell & Cota-Robles (1967). Estes investigadores observaram que a adição de EDTA a
células de E. coli tratadas com lisozima promove a conversão rápida das células em estruturas
esféricas sensíveis a choque osmótico. O uso de esferoplastos é vantajoso para muitos estudos,
pois a ruptura da membrana externa expõe grandes áreas da membrana interna ao ambiente
(Hofschneider & Wandstructur, 1960).
O crescimento de uma população bacteriana é condicionado por muitos factores, incluindo
o tipo de ambiente, número de indivíduos na população, interacções dinâmicas com outras
populações bacterianas, presença de outros microrganismos predadores, factores químicos e
físicos tais como disponibilidade de nutrientes essenciais, temperatura, pH, osmolaridade,
pressão hidrostática, concentração de oxigénio, luz, radiação ionizante ou ultravioleta, presença
de metabolitos tóxicos resultantes do metabolismo das células da população em crescimento ou
presença de agentes antimicrobianos tais como bacteriocinas e antibióticos. As células de E. coli
não requerem factores específicos de crescimento, podendo sintetizar todos os seus componentes
macromoleculares a partir de glicose. Quando em condições óptimas, estas células possuem um
ciclo de vida bastante curto, aproximadamente de 30 minutos. Em cultura pura, uma população
de E. coli pode atingir uma densidade final de 2-5*109 células por mL em cerca de 12 a 18 horas,
quando em meio de cultura rico e sob condições aeróbicas. A representação gráfica do aumento
do número de indivíduos em função do tempo de cultura designa-se por curva de crescimento.
Entre outros factores, o esgotamento de nutrientes e a densidade populacional são considerados
determinantes para a dinâmica populacional das culturas bacterianas (Martin, 1932; Moyer,
1936; Dagley & Hinshelwood, 1938; Elliker & Frazier, 1938; Delpy et al., 1956; Ecker &
Lockhart, 1961). Em condições experimentais, quando se inocula uma população bacteriana num
sistema fechado, o crescimento dessa população passa por quatro fases características:
1. Fase lag – fase de adaptação metabólica ao novo ambiente; o metabolismo celular está
direccionado para sintetizar as enzimas requeridas para o crescimento nas novas condições

8
ambientais encontradas pelas células. O número de indivíduos não aumenta nesta fase, podendo
até mesmo decrescer.
2. Fase exponencial – fase na qual o número de células da população duplica a cada
geração. Esta taxa de crescimento não pode ser mantida indefinidamente num sistema fechado.
Após um determinado período de crescimento exponencial, as condições ambientais tornam-se
desfavoráveis pela escassez de nutrientes essenciais, acumulação de metabolitos tóxicos e
limitação de espaço.
3. Fase estacionária – fase em que a taxa de crescimento diminui significativamente devido
às condições limitantes do meio. As células continuam metabolicamente activas e em divisão,
mas parte das células torna-se inviável e a taxa de divisão celular é muito próxima da taxa de
morte celular, o que mantém constante o número de células viáveis na população.
4. Fase de declínio – fase em que as células perdem a capacidade de se dividir, a taxa de
morte celular torna-se maior que a taxa de divisão e o número de células viáveis decresce
exponencialmente até à completa extinção da população.
1.3. Efeito do alumínio na Escherichia coli
O interesse pelo impacto da presença de alumínio nos ecossistemas cresceu a partir dos
anos setenta do século passado, em virtude da acidificação dos solos promover a sua
biodisponibilidade (Cronan et al., 1986; Wood, 1988). Por conseguinte, grande parte das
pesquisas sobre efeitos biológicos do alumínio tem sido dedicada ao estudo dos mecanismos de
tolerância a alumínio em plantas. Por outro lado, também nos anos setenta do século passado
Alfrey e colaboradores (1972; 1976) estabeleceram ser o alumínio o principal agente etiológico
responsável pela Demência de Diálise, incentivando o estudo da relação entre exposição a
alumínio e desenvolvimento de desordens neurológicas.
Os estudos relativos aos efeitos do alumínio em microrganismos são escassos (Piña &
Cervantes, 1996). A toxicidade do alumínio foi estudada em E. coli (Guida et al., 1991; Bojic et
al., 2002). No primeiro estudo (Guida et al., 1991), observou-se inibição do crescimento na
presença de concentrações de nitrato de alumínio na ordem dos milimolar. A análise por
espectrometria de absorção atómica mostrou ocorrer ligação de alumínio a um local intracelular,
bem como a sua ligação à superfície de célula com cooperatividade evidente. Este estudo
demonstrou ainda ser a toxicidade do alumínio mais elevada em meio ácido e na ausência de
ferro. A acidificação favorece a formação de Al(H2O)63+, sugerida como sendo a espécie tóxica.
Por outro lado, o alumínio utiliza o sistema de transporte do ferro para aceder ao espaço

9
intracelular. A importância do pH para acção tóxica do alumínio em E. coli foi posteriormente
confirmada por Bojic e colaboradores (2002). Estes investigadores estudaram os efeitos tóxicos
do alumínio em E. coli em função do pH, concentração de alumínio e tempo de exposição, em
meio de crescimento (peptona 10,0 g/L; glicose 5,0 g/L; NaCl 3,0 g/L; NH4Cl 1,0 g/L; e CaCl2
0,01 g/L) e em meio fisiológico (NaCl 0,85%). Nas condições experimentais usadas, o alumínio
é tóxico para a E. coli na gama de concentrações entre 0,1 e 10,0 mg/L. A toxicidade é mais
acentuada a pH ácidos e no meio fisiológico do que no meio de crescimento bacteriano. Este
facto deve-se provavelmente à elevação da biodisponibilidade de Al3+ no meio fisiológico, bem
como às restrições ao nível de nutrição, crescimento e multiplicação que este meio representa.
1.4. A bomba de (Na+/K+)ATPase
A bomba de sódio foi descoberta por Jens Christian Skou em 1957. A sua descoberta foi
publicada na revista Biochimica et Biophysica Acta (vol. 23, pp. 394-401) num artigo intitulado
"The influence of some cations on an adenosine triphosphatase from peripheral nerves". Em
1997, pela descoberta da Bomba de Sódio, recebeu o Prémio Nobel da Química (juntamente com
Paul D. Boyer e John E. Walker).
Skou descobriu uma enzima que hidrolisa ATP somente se estiverem presente Na+ e K+
para além do Mg2+. Assim, esta enzima foi denominada (Na+/K+)ATPase. Skou propôs que a
(Na+/K+)ATPase é parte integrante da bomba de Na+ e K+ e que o ATP fornece a energia
necessária ao transporte desses catiões. Na presença de Na+, Mg2+ e ATP ocorre fosforilação da
cadeia lateral de um aspartato específico. Este intermediário fosforilado (E-P) é então hidrolisado
se o K+ estiver presente. A fosforilação não requer K+, enquanto que a desfosforilação não requer
Na+ ou Mg2+. A fosforilação dependente de Na+ e a desfosforilação dependente de K+ não são as
únicas reacções críticas. A bomba também se interconverte entre, pelo menos, duas
conformações diferentes, indicadas por E1 e E2. Assim, pelo menos quatro estados
conformacionais (E1, E1-P, E2-P e E2) participam no transporte de Na+ e K+ e concomitante
hidrólise de ATP. Três iões Na+ e dois iões K+ são transportados por ATP hidrolisado. Desse
modo, a bomba gera uma corrente eléctrica através da membrana citoplasmática. Ou seja, a
(Na+/K+)ATPase é electrogénica. Na conformação E1 os locais de ligação dos iões apresentam-se
expostos ao interior da célula, enquanto que na conformação E2, se encontram expostos ao
espaço extracelular. E1 tem uma alta afinidade para Na+, enquanto E2 tem uma alta afinidade
para K+. O ciclo da bomba de sódio e potássio encontra-se representado na Figura 1.2.

10
Figura 1.2. Intermediários do transporte mediado pela (Na+/K+)ATPase segundo o modelo de Alberts-Post (Lingrel & Kuntzweiler, 1994).
A bomba de sódio e potássio é constituída por dois tipos de subunidades obrigatórias, α e
β. A maior parte da cadeia α, incluindo o centro de ATPase, está localizada no lado citosólico da
membrana (Fig. 1.3). A pequena porção de α do lado extracelular contém o centro de ligação
para a ubaína e outros esteróides. A ubaína é um inibidor específico da (Na+/K+)ATPase que, por
sua vez, é o único receptor fisiológico conhecido para a ubaína (Jorgensen et al., 2003).
Figura 1.3. Topologia da subunidade α1 da (Na+/K+)ATPase de carneiro (Lingrel & Kuntzweiler, 1994).

11
A (Na+/K+)ATPase é uma proteína ubíqua na membrana plasmática das células animais e
que se encontra também na membrana interna da E. coli, sendo sensível a ubaína e requerendo
Mg2+ e ATP. No entanto a sua actividade é relativamente baixa quando comparada com a enzima
existente na maioria dos tecidos dos vertebrados (Hafkenscheid & Bonting, 1968).
1.5. Efeito do alumínio na actividade da (Na+/K+)ATPase
A capacidade do alumínio para modificar a actividade da (Na+/K+)ATPase de mamíferos
foi observada por vários investigadores (Lai et al., 1980; King et al., 1983; Caspers et al., 1990,
1994; Rao, 1990; 1992; Lal et al., 1993; Zatta et al., 1995; Sarin et al., 1997; Silva & Gonçalves,
2003; Silva et al., 2005, 2007). A observação mais frequentemente descrita na literatura é a
inibição de actividade de adenosinatrifosfatase na presença de alumínio. Porém, a activação
desta actividade enzimática também foi observada por outros investigadores (Zatta et al., 1995).
1.6. Objectivo do trabalho
O efeito inibitório do alumínio na (Na+/K+)ATPase tem sido observado por vários
investigadores desde os anos oitenta do século passado. Até à data foram propostos quatro
mecanismos explicativos da acção inibitória do alumínio na actividade hidrolítica de ATP da
bomba de Na+ e K+. O primeiro assenta na possibilidade de formação de complexos estáveis de
ATP-Al3+ e consequente redução da concentração do substrato da enzima, o ATP-Mg2+ (Nelson,
1996). Lal e colaboradores (1993) sugeriram que alterações na estrutura da membrana induzidas
por alumínio estariam na origem da redução da actividade hidrolítica da (Na+/K+)ATPase.
Caspers e colaboradores (1990; 1994) verificaram que o alumínio, em concentrações
micromolares, aumenta a capacidade de ligação de alta afinidade de ubaína, sem alterar a
afinidade dos locais de ligação. Por fim, Silva e colaboradores (2003; 2005) observaram que o
alumínio impede a activação adicional da actividade de (Na+/K+)ATPase produzida pela ligação,
de baixa afinidade, de ATP e reduz a sensibilidade da actividade enzimática a concentrações
submicromolares de ubaína. Os mecanismos enunciados parecem revelar aspectos
complementares da acção do alumínio, que podem contribuir para o seu efeito inibitório na
actividade hidrolítica de ATP da bomba de Na+ e K+. A (Na+/K+)ATPase possui uma estrutura
quaternária complexa e as suas isozimas exibem heterogeneidade estrutural e um padrão de
expressão que é regulado de modo a permitir que a actividade da enzima seja ajustada de acordo
com as necessidades da célula. Existem várias isoformas da subunidade catalítica (α), cujas
propriedades cinéticas, sensibilidade a ubaína e afinidades para Na+, K+, e ATP são diferentes

12
(Sweadner, 1979; 1985; Berrebi-Bertrand et al., 1990; Blanco et al., 1990; Jewell & Lingrel,
1991). Relativamente à subunidade reguladora β, importante para a biogénese do complexo
enzimático, também são conhecidas isoformas (Serluca et al., 2001), que concedem às isozimas
propriedades cinéticas e sensibilidade a esteróides cardiotónicos distintas. Nos últimos anos, tem
sido evidenciada a formação na membrana plasmática de estruturas oligoméricas constituídas por
protómeros αβ, que conferem à (Na+/K+)ATPase a capacidade de activação adicional por altas
concentrações de ATP (Maunsbach et al., 1991; Boldyrev, 2001; Taniguchi et al., 2001; Kaplan,
2002). Em suma, a actividade de (Na+/K+)ATPase das membranas biológicas depende, entre
outros factores, da estrutura oligomérica da enzima, do empacotamento da bicamada lipídica e da
microviscosidade do ambiente lipídico (Kimelberg & Papahadjopoulos, 1972; Silvius &
McEhhaney, 1982; Blanco & Mercer, 1998; Boldyrev, 2001).
O efeito do alumínio na actividade de (Na+/K+)ATPase tem sido estudado essencialmente
em preparações enriquecidas em membranas plasmáticas obtidas a partir de tecidos de
mamíferos. Recentemente, a análise comparativa do efeito do alumínio na (Na+/K+)ATPase
presente em membranas sinápticas de córtex cerebral do rato, do lobo óptico do choco e do órgão
eléctrico da tremelga revelou sensibilidades distintas da actividade enzimática a concentrações
submilimolares de AlCl3 (Silva et al., 2007). Por conseguinte, a heterogeneidade estrutural e
funcional da bomba de Na+ e K+ e das membranas onde se encontra inserida parecem
condicionar a acção inibitória do alumínio na actividade de (Na+/K+)ATPase.
Para esclarecer o mecanismo de acção do alumínio na (Na+/K+)ATPase é crucial a
existência de modelos membranares que exibam sensibilidades distintas a alumínio. O objectivo
principal deste trabalho consistiu em avaliar o efeito do alumínio (AlCl3) na actividade de
(Na+/K+)ATPase de E. coli. Tal como referido anteriormente, a composição lipídica da
membrana interna de E. coli é distinta da presente em eucariontes (Razin, 1975) e a actividade
ATPásica sensível a ubaína é relativamente baixa quando comparada com a das isozimas
existentes na maioria dos tecidos dos vertebrados (Hafkenscheid & Bonting, 1968). Por outro
lado, estudos anteriores (Guida et al., 1991; Bojic et al., 2002) evidenciaram a acumulação de
alumínio em células de E. coli e a inibição do crescimento populacional em sistema fechado na
presença de alumínio. O efeito do alumínio (AlCl3) na actividade de (Na+/K+)ATPase de E. coli
foi avaliado em esferoplastos submetidos a ciclo de congelamento e descongelamento e
preparados a partir de culturas puras de E. coli em fase estacionária. Dois regimes de exposição a
AlCl3 foram comparados: exposição in vitro e in vivo. No primeiro caso, esferoplastos obtidos a
partir de culturas E. coli controlo (ausência de alumínio adicionado ao meio de cultura) foram
incubados na presença de concentrações crescentes de AlCl3 e actividade de (Na+/K+)ATPase

13
comparada com a de esferoplastos controlo (ausência de alumínio adicionado ao meio de
reacção). Para efectuar a avaliação da exposição in vivo a AlCl3 na actividade de
(Na+/K+)ATPase de E. coli, mediu-se, em meio de reacção sem alumínio adicionado, a
actividade enzimática de esferoplastos preparados a partir de culturas E. coli expostas, durante a
fase estacionária, a concentrações AlCl3 superiores à concentração tóxica mínima. Para o efeito,
caracterizou-se inicialmente a capacidade de sobrevivência da E. coli em meios de cultura
suplementados com diferentes concentrações de AlCl3.

14

15
2. Material e Métodos

16

17
2.1. Material biológico e meios de cultura
Meio de cultura é uma substância líquida ou sólida, simples ou complexa, que permite a
nutrição, o crescimento e a multiplicação dos microrganismos. Neste trabalho foi usado meio de
cultura líquido LB Boillon – Miller pH 7,0 (Merck) constituído por peptona de caseína 10,0 g/L;
extracto de levedura 5,0 g/L e cloreto de sódio 10,0 g/L. O meio de cultura LB foi adquirido
pronto a usar, bastando somente dissolver 25,0 g/L de água destilada (meio líquido). Para
preparar o meio sólido (usado nas placas) foi necessário adicionar agar (20,0 g/L) (Formedium).
Todo o material e meios usados foram esterilizados a 126ºC durante 20 minutos. A esterilização
foi efectuada em autoclave da marca Prestige – modelo Medical 2100 classic.
Foi usada a espécie de bactéria Escherichia coli, estirpe JM 109, gentilmente oferecida
pelo Prof. Dr. Manuel Santos do Departamento de Biologia da Universidade de Aveiro.
2.2. Preparação da cultura de Escherichia coli
Realizou-se uma sementeira por exaustão de E. coli estirpe JM 109, que foi incubada a
37ºC durante 24 horas numa estufa Heraeus – Fuction Line.
Os pré-inóculos de E. coli foram preparados retirando colónias isoladas na sementeira por
exaustão e repicando para tubos contendo 5 mL de meio LB. De seguida, os tubos foram
incubados durante 24 horas, a 37ºC, com agitação de 200 rotações por minuto (rpm) numa estufa
com agitação rotativa da marca Infors Unitron.
As culturas foram preparadas em erlenmeyers contendo 50 mL de meio de cultura LB
estéril. Para isso, foi adicionada uma quantidade de pré-inóculo suficiente para que a densidade
óptica (D.O.) inicial fosse de 0,1. Essas culturas foram mantidas na estufa a 37ºC, com agitação
de 200 rpm, até ao final da experiência.
Para realizar a leitura da D.O. foi necessário diluir as amostras 10x em meio LB estéril. As
referidas leituras foram efectuadas num espectrofotómetro “Beckman modelo DU 530”, no
comprimento de onda de 600 nm.
Com o objectivo de estudar o efeito do alumínio no crescimento bacteriano, adicionou-se
alumínio às culturas de E. coli. As culturas foram preparadas como descrito anteriormente. Após
17 horas de incubação, diferentes quantidades de alumínio foram adicionadas às culturas
(concentrações finais entre 1,0*10-4 e 3,0*10-1 M). Foi preparada uma solução stock de AlCl3 em
meio LB estéril. De seguida essa solução foi esterilizada por filtração. A solução foi passada por
um filtro com 0,2 µm de diâmetro de malha, com o auxílio de uma seringa, permitindo a
retenção dos microrganismos presentes no meio.

18
As culturas que se destinavam ao isolamento de esferoplastos foram deixadas a crescer
durante 19 horas na estufa a 37ºC, com agitação de 200 rpm.
Figura 2.1. Fotografias com os passos para preparação da cultura de E coli. A: tubos com 5 mL de meio LB, líquido (pré-inóculos). B: culturas incubadas a 37ºC numa estufa com agitação de 200 rpm da marca Infors Unitron. C e D: leitura da D.O.. As referidas leituras foram efectuadas a 600,0 nm num espectrofotómetro “Beckman modelo DU 530”.
2.3. Contagem das UFC
A contagem das unidades formadoras de colónias (UFC) permite estimar o número de
células viáveis, isto é, o número de células capazes de se reproduzirem numa amostra. A
metodologia envolve a colheita de alíquotas de uma cultura microbiana em diferentes tempos de
crescimento. A técnica utilizada foi a das diluições decimais, isto é, foram realizadas diluições
sucessivas de 1:10 em meio de cultura LB. Alíquotas de 50 µL, foram semeadas por
espalhamento à superfície em placas de petri (ø 8 cm) contendo meio LB agarizado. As placas
foram incubadas invertidas a 37ºC. Após 24 horas foram contadas as colónias visíveis para
determinar o número de UFC presentes em cada tempo de amostragem. Foram consideradas
apenas as placas que continham entre 30 e 300 colónias.

19
2.4. Preparação de esferoplastos
Culturas de E. coli controlo e na presença de diferentes concentrações de alumínio foram
preparadas como descrito anteriormente. Os esferoplastos foram isolados a partir de células de E.
coli, segundo adaptação do método proposto por Birdsell & Cota-Robles (1967) e Sullivan et al.
(2005) (Fig. 2.2).
Figura 2.2. Esquema do método de preparação de esferoplastos de E. coli.
A cultura de E. coli (Fig. 2.1) foi centrifugada a 12.000xg durante 5 min a 4ºC. O
sedimento obtido foi ressuspenso em meio LB estéril. Esta suspensão foi centrifugada a
12.000xg durante 5 min a 4ºC. O sedimento obtido foi ressuspenso num volume total de 40 mL
de meio contendo sacarose 0,5 M e Tris 0,2 M (pH 8,0). Após uma pré-incubação de 5 min a
25ºC com agitação, adicionou-se lisozima (10 µg/mL) e 10 mM de EDTA (pH 8,0) e incubou-se

20
durante mais 10 min. De seguida procedeu-se a uma centrifugação a 12.000xg durante 5 min a
4ºC. O sedimento foi ressuspenso em 1 mL de meio de reacção constituído por NaCl 110 mM,
KCl 15 mM, MgCl2 10 mM, EGTA 100 µM, HEPES-Na 2 mM a pH 7,4.
Figura 2.3. Observação ao microscópio óptico de cultura de E. coli (ampliação de 1.000x). As setas indicam células de E. coli.
Todas as centrifugações foram feitas numa centrífuga refrigerada “Beckman, modelo
Avant J-251” no rotor JA 25.15.
Os esferoplastos obtidos foram observados e fotografados ao microscópio óptico com a
ampliação de 1.000x (Fig. 2.4), divididos em alíquotas e congelados a -80ºC.

21
Figura 2.4. Observação ao microscópio óptico da preparação de esferoplastos de E. coli
(ampliação de 1.000x). As setas indicam esferoplastos de E. coli.
2.5. Quantificação da proteína
A quantificação da proteína total foi efectuada segundo o método colorimétrico do biureto,
descrito por Layne (1957).
As amostras (50 µL) foram solubilizadas com dodesilsulfato de sódio (SDS) a 10%. A
reacção colorimétrica (formação de complexos violeta cupro-proteicos em meio básico) foi
iniciada com adição de 2 mL de reagente de biureto (sulfato de cobre hidratado a 0,15% tartarato
de sódio e potássio a 0,6%, hidróxido de sódio 3% e iodeto de potássio a 0,1%). A reacção
decorreu durante 15 min a 30ºC. Simultaneamente, nas mesmas condições das amostras, foram
preparados padrões de albumina sérica bovina (0,4%) de concentrações conhecidas (10 e 20
mg/mL). As absorvâncias dos padrões e das amostras foram determinadas a 540 nm num
espectrofotómetro UV/VIS “Perkin-Elmer, modelo Lambda 14P”. Os valores das absorvâncias
permitiram traçar a recta padrão. As concentrações de proteína foram obtidas por interpolação
dos valores na recta padrão.

22
2.6. Medição da actividade da adenosinatrifosfatase
A actividade da (Na+/K+)ATPase nos esferoplastos isolados a partir de culturas de E. coli
controlo e culturas contendo diferentes concentrações de alumínio foi determinada por
quantificação dos protões produzidos, segundo o método potenciométrico descrito por Madeira
et al. (1974). As reacções enzimáticas decorreram em 2 mL de meio de reacção contendo NaCl
110 mM, KCl 15 mM, MgCl2 10 mM, EGTA 100 µM, HEPES-Na 2 mM a pH 7,4, a 35ºC com
agitação magnética constante. As variações da concentração de H+ foram detectadas por um
eléctrodo “Ingold U 402 – M6 – S7 – 100”, conectado a um potenciómetro “Crison micro pH
2002”. A reacção decorreu na presença de 1 mg de proteína por mL e foi iniciada pela adição de
1 mM de Mg-ATP, e calibradas com 37,5 nmoles de HCl. A actividade que resultou dos ensaios
foi designada como actividade de ATPase total. A ATPase basal foi medida na presença de
ubaína (20 mM). A actividade de (Na+/K+)ATPase corresponde à diferença entre a ATPase total
e a basal e foi expressa em nmoles de H+/ min/ mg de proteína.
Figura 2.5. Equipamento usado para medir a actividade da (Na+/K+)ATPase, quantificando a variação da concentração de H+
.
Para determinação do efeito in vitro do alumínio na actividade da (Na+/K+)ATPase,
diferentes concentrações de alumínio (3*10-4, 5*10-4, 10-3 M) foram adicionadas ao meio de
reacção e a reacção iniciada pela adição de 1 mM de Mg-ATP, após 15 min de pré-incubação.
Para determinação do efeito in vivo do alumínio na actividade da (Na+/K+)ATPase os
esferoplastos foram isolados a partir de culturas de E. coli expostas a alumínio (6,0*10-4 e
9,5*10-3 M) durante 2 horas, tal como descrito anteriormente.
2.7. Análise dos dados
As curvas de crescimento das culturas de E. coli foram analisadas com a ajuda do
programa “MicroFit© version 1.0”. As curvas de “dose-resposta” dos diferentes parâmetros de
sobrevivência/morte para AlCl3 foram analisadas com a ajuda do programa “Microcal™ Origin

23
6.0”, pelo método de regressão não linear, introduzindo as respectivas funções de ajuste. As
curvas de “tempo-morte” em resposta a exposição a AlCl3 foram analisadas segundo o modelo
farmacodinâmico proposto por Yano et al. (1998) e modificado por nós (Fig. 2.6).
Figura 2.6. Representação esquemática do modelo farmacodinâmico do efeito do AlCl3 na cultura de E. coli em sistema fechado. A população total de E. coli, x, é considerada como sendo constituída por duas sub-populações, x1 e x2. O efeito da exposição a diferentes concentrações de AlCl3 (c) na contagem de células viáveis (UFC/mL) das duas sub-populações ao longo do tempo
é representado pelas funções dt
dx1 e dt
dx2 , onde: β’ e β’’ são as constantes de velocidade de,
respectivamente, crescimento e morte naturais das sub-populações; Ni e N0 e são a densidade de células viáveis inicial e antes da adição de AlCl3;
'maxV e ''
maxV são as velocidades máximas de
morte induzida; '50EC e ''
50EC são as concentrações de AlCl3 que causam 50% de efeito; e K1 e
K2 são as constantes de transferência entre as duas sub-populações.
Os resultados deste trabalho foram expressos como a média ± o erro padrão (E.P.) para o
número de ensaios indicados na legenda das figuras. O significado estatístico das diferenças foi
determinado pelo teste t de Student; valores de p<0,05 foram considerados representativos de
diferenças estatisticamente significativas.

24

25
3. Resultados e Discussão

26

27
3.1. Efeito do AlCl3 na capacidade de sobrevivência de E. coli
Antes de se iniciar o estudo do efeito do AlCl3 na capacidade de sobrevivência de E. coli,
procedeu-se à selecção do método de monitorização da dinâmica populacional de E. coli, nas
condições de cultura usadas.
Tal como se pode observar na figura 3.1, a turbidimetria possibilita verificar o crescimento
bacteriano avaliado pelo método da contagem das unidades formadoras de colónias (UFC).
0,1
0,3
0,5
0,7
0,9
1,1
1,3
1,5
1,7
1,9
2,1
2,3
2,5
2,7
2,9
0 10 20 30 40 50
TEMPO (horas)
CR
ES
CIM
EN
TO
(D
.O.)
7,9
8,1
8,2
8,4
8,5
8,7
8,8
9,0
9,1
9,3
9,4
9,6
9,7
CR
ES
CIM
EN
TO
(L
OG
UF
C*m
L-1)
Figura 3.1. Curva de crescimento de E. coli em meio LB , monitorizada pelo método turbidimétrico (○) e por contagem de UFC (●). As culturas foram preparadas como descrito em “Materiais e Métodos”. O meio de cultura (50 mL) foi inoculado com 4,0*10-5 células viáveis (D.O.600 nm = 0,1) e as culturas foram mantidas na estufa a 37ºC, com agitação de 200 rpm, até ao final da experiência. A tabela inclusa apresenta os parâmetros do crescimento bacteriano estimados segundo o modelo de Baranyi (“MicroFit© version 1.0”). Ni e N0, densidade inicial e densidade máxima de células viáveis na população bacteriana; Mumax = taxa específica de crescimento máximo; t-lag, duração da fase de adaptação; t-d, tempo de duplicação.
Em meio LB e à temperatura de 37ºC com agitação de 200 rpm, a população inicial de E.
coli (8,0*10-7 células viáveis por mL) aumentou rapidamente durante as primeiras horas de
incubação (fase exponencial). Durante este período, a densidade óptica da cultura bacteriana

28
aumentou proporcionalmente ao número de células viáveis na cultura. A avaliação da densidade
populacional pelo método turbidimétrico e pelo método da contagem de UFC conduziu à
obtenção de resultados concordantes. Nas condições de cultura usadas, o crescimento bacteriano
foi caracterizado por uma fase de adaptação curta (t-lag = 0,85±2,54 h), após a qual a taxa
específica de crescimento atingiu o seu valor máximo de 1,45±1,36 h (tempo de duplicação de
0,48±0,65 h) durante a fase de crescimento exponencial. A densidade populacional atingiu o seu
valor máximo, 7,24*10-9±1,26 UFC*mL-1, após ~10 h de incubação, mantendo-se o número
global de células constante durante pelo menos 24 h (fase estacionária). Após este período,
observou-se uma fase de declínio, que se prolongou por vários dias (Fig. 3.2). Apesar de Li et al.
(1993) terem demonstrado ser possível a quantificação do decréscimo do número de células
viáveis de E. coli por turbidimetria, nas condições experimentais usadas, o método de contagem
de UFC revelou-se mais sensível na monitorização da fase de declínio (resultados não
apresentados). Por conseguinte, a turbidimetria foi usada para confirmar o crescimento
bacteriano até à fase estacionária, enquanto que a dinâmica populacional durante as fases
estacionária e de declínio foi avaliada pelo método de contagem de UFC.
Com o objectivo de caracterizar a dinâmica populacional das culturas de E. coli em
condições controlo, procedeu-se à colheita de alíquotas (50 µL) de cultura microbiana em
diferentes tempos durante 30 dias. As alíquotas, após realização de diluições decimais
sucessivas, foram semeadas por espalhamento à superfície em placas de petri contendo meio LB
agarizado. As contagens de UFC*mL-1, após incubação das placas invertidas a 37ºC durante 24
horas, para cada tempo de amostragem encontram-se representadas na figura 3.2(A). O método
utilizado permite, com rigor, quantificar densidades de células viáveis na cultura superiores a
6*102 células*mL-1, ou seja, um decréscimo de densidade populacional até 99,99999% do valor
máximo atingido no início da fase estacionária.
Na figura 3.2 é visível que a fase de declínio das culturas de E. coli em condições controlo
teve início no 3º dia de incubação, durante a qual o número de células viáveis diminuiu
exponencialmente. No final dos 30 dias de incubação, a densidade de células viáveis na cultura
era de 3,17*10-6±8,27*10-5 UFC*mL-1 (Fig.3.2(A)), o que corresponde a uma percentagem de
sobrevivência de 0,07% (Fig.3.2(B)) ou de morte de 99,93% (Fig.3.2(D)). Em virtude de se ter
verificado que a densidade de células viáveis atingida no início da fase estacionária apresentava
oscilações de cultura para cultura, na figura 3.2.(C) encontra-se representada a fracção da
densidade de células viáveis relativamente à densidade máxima observada em cada cultura para
cada tempo de amostragem. A variação do valor deste parâmetro ao longo do tempo ilustra
claramente o decréscimo exponencial da densidade populacional que conduz à redução da

29
população de células viáveis de E. coli em três ordens de grandeza durante 29 dias em condições
controlo.
1,E+06
1,E+07
1,E+08
1,E+09
1,E+10
1,E+11
1,E+12
0 10 20 30
TEMPO (dias)
UF
C*m
L-1
A B
C D
0
20
40
60
80
100
0 10 20 30
% D
E M
OR
TE
0
20
40
60
80
100
120
0 10 20 30
TEMPO (dias)
% D
E S
OB
RE
VIV
ÊN
CIA
-3,2
-2,8
-2,4
-2,0
-1,6
-1,2
-0,8
-0,4
0,0
0 10 20 30
LO
G (
N/N
0)
N0 1,51*108 ± 3,72 UFC*mL-1 Nmax 7,24*109 ± 1,26 UFC*mL-1 Mumax 1,45 ± 1,36 h t-lag 0,85 ± 2,45 h t-d 0,48 ± 0,65 h RSS 36,13 RMS 5,25 UFC*mL-1
Figura 3.2. Crescimento e declínio da população de E. coli em condição de cultura controlo. Curva de crescimento e morte (A), percentagem de sobrevivência (B) fracção da densidade de células viáveis, N, relativamente à densidade máxima observada, N0, (C), percentagem de morte (D). As condições experimentais são as mesmas das descritas na legenda da figura 3.1. Os valores representam a média ± E.P. de pelo menos dois ensaios independentes realizados em duplicado.
O perfil da curva de crescimento e declínio da cultura de E coli observada em condições
controlo é consentâneo com o descrito em muitos estudos de crescimento bacteriano em
condições análogas (Delpy et al., 1956; Amanatidou et al., 1999; Oscar, 1999; Koseki & Isobe,
2005) e característico das culturas bacterianas que crescem com nutrientes limitados (Shehata &
Marr, 1971). Num sistema fechado, o crescimento exponencial não pode ocorrer

30
indefinidamente, devido ao consumo de nutrientes essenciais e à produção de metabolitos. Com
a acumulação de metabolitos e de produtos tóxicos e a carência de nutrientes no meio de cultura,
a manutenção das funções vitais das células é comprometida, verificando-se, então, a fase de
declínio da cultura, caracterizada pela redução do número de células viáveis.
Seguidamente, procedeu-se ao estudo do impacto do AlCl3 na dinâmica populacional de E.
coli nas condições de cultura anteriormente descritas. Para o efeito, após 17 horas de incubação,
diferentes quantidades de AlCl3 foram adicionadas às culturas e a densidade de células viáveis
avaliada ao longo do tempo. A fracção da densidade de células viáveis relativamente à densidade
máxima observada para cada tempo de amostragem encontram-se representadas na figura 3.3. A
densidade máxima de células viáveis foi sempre avaliada em cada cultura e corresponde ao valor
médio de UFC*mL-1 das amostragens realizadas em duplicado uma e duas horas antes da adição
de AlCl3.
Tal como se pode observar na figura 3.3, a dependência da fracção da densidade de células
viáveis relativamente à densidade máxima observada em função do tempo durante a fase de
declínio da cultura na ausência de alumínio parece ter a forma duma curva sigmoidal
decrescente. Na realidade, o ajuste à função exponencial MteNN =0 revela que o
desaparecimento de células viáveis foi lento (M = 0,038±0,002 h-1; R2 = 0,94173).
O AlCl3, quando presente no meio de cultura em concentrações inferiores a ~9,0*10-3 M,
não parece modificar a forma em “S invertido” da dependência de 0NN em função do tempo
de cultura, enquanto que em concentrações superiores causou uma alteração marcada na forma
da curva que descreve o desaparecimento de células viáveis na fase de declínio. Nomeadamente,
observou-se uma transição entre a forma “S invertido” para a forma “L”, que pode ser
interpretada como um aumento da taxa de morte de células viáveis induzido por alumínio. De
facto, na gama de concentrações de AlCl3 entre 9,0*10-3 M e 9,0*10-2 M, os valores de M são
cerca de 10 a 100 vezes superiores aos registados nas culturas de E. coli em condições controlo.
Por outro lado, para concentrações de AlCl3 inferiores a 9,0*10-3 M observou-se,
tendencialmente, uma redução dos valores de M, acompanhada por diminuição dos valores de R2
(quadrado do coeficiente de correlação momentânea).
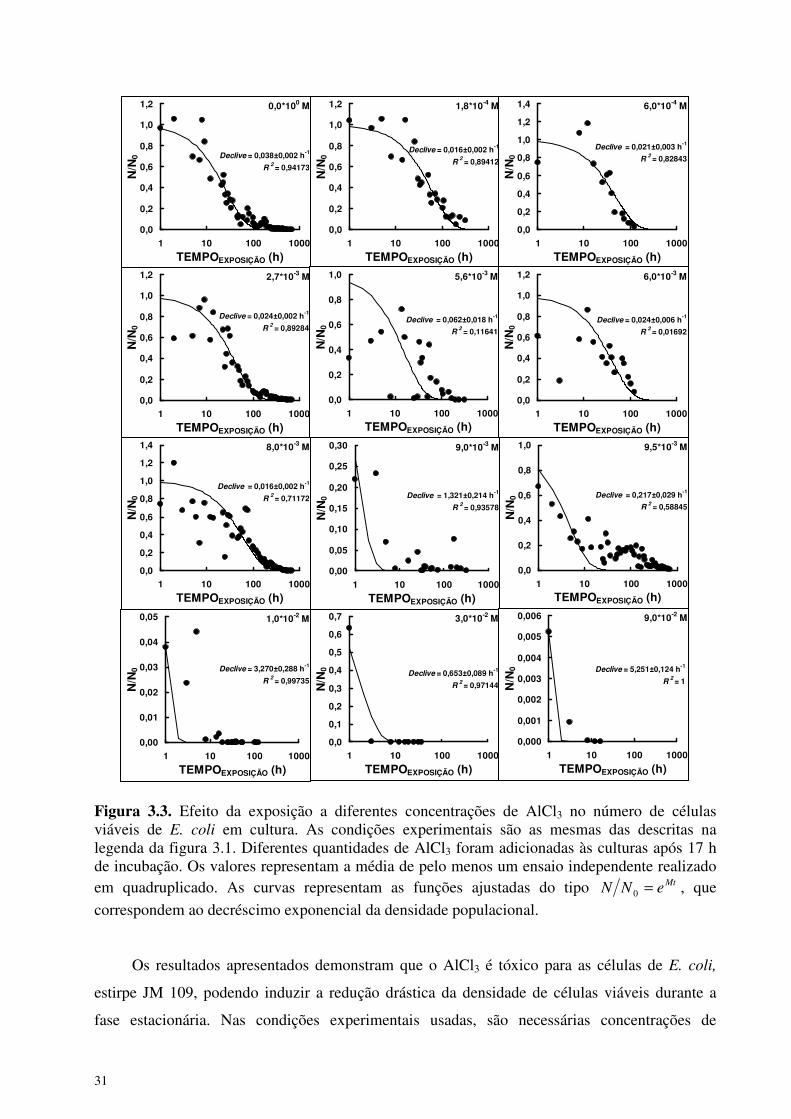
31
3,0*10-2 M
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0
Declive = 0,653±0,089 h-1
R2 = 0,97144
0,0*100 M
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0 Declive = 0,038±0,002 h-1
R2 = 0,94173
1,8*10-4 M
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0
Declive = 0,016±0,002 h-1
R2 = 0,89412
6,0*10-4 M
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0
Declive = 0,021±0,003 h-1
R2 = 0,82843
2,7*10-3 M
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0
Declive = 0,024±0,002 h-1
R2 = 0,89284
5,6*10-3 M
0,0
0,2
0,4
0,6
0,8
1,0
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0Declive = 0,062±0,018 h-1
R2 = 0,11641
6,0*10-3 M
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0
Declive = 0,024±0,006 h-1
R2 = 0,01692
8,0*10-3 M
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0
Declive = 0,016±0,002 h-1
R2 = 0,71172
9,0*10-3 M
0,00
0,05
0,10
0,15
0,20
0,25
0,30
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0 Declive = 1,321±0,214 h-1
R2 = 0,93578
9,5*10-3 M
0,0
0,2
0,4
0,6
0,8
1,0
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0 Declive = 0,217±0,029 h-1
R2 = 0,58845
1,0*10-2 M
0,00
0,01
0,02
0,03
0,04
0,05
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0 Declive = 3,270±0,288 h-1
R2 = 0,99735
9,0*10-2 M
0,000
0,001
0,002
0,003
0,004
0,005
0,006
1 10 100 1000
TEMPOEXPOSIÇÃO (h)
N/N
0 Declive = 5,251±0,124 h-1
R2 = 1
Figura 3.3. Efeito da exposição a diferentes concentrações de AlCl3 no número de células viáveis de E. coli em cultura. As condições experimentais são as mesmas das descritas na legenda da figura 3.1. Diferentes quantidades de AlCl3 foram adicionadas às culturas após 17 h de incubação. Os valores representam a média de pelo menos um ensaio independente realizado em quadruplicado. As curvas representam as funções ajustadas do tipo Mt
eNN =0 , que
correspondem ao decréscimo exponencial da densidade populacional. Os resultados apresentados demonstram que o AlCl3 é tóxico para as células de E. coli,
estirpe JM 109, podendo induzir a redução drástica da densidade de células viáveis durante a
fase estacionária. Nas condições experimentais usadas, são necessárias concentrações de

32
alumínio superiores a 9,0*10-3 M no meio de cultura para induzir a morte massiva de células.
Concentrações de AlCl3 inferiores a 1,8*10-4 M não causaram qualquer alteração significativa na
densidade bacteriana durante as fases estacionária e de declínio das culturas, bem como
concentrações de AlCl3 superiores a 1,0*10-1 M causaram o desaparecimento total das células
viáveis (resultados não apresentados). Esta observação está de acordo com os resultados obtidos
por Guida et al. (1991), Bojic et al. (2002) e Harrison et al. (2005). Estes investigadores, usando
diferentes estirpes de E. coli, verificaram que a toxicidade do alumínio é influenciada pela
duração da exposição, pela composição do meio de cultura e pelo pH. Assim, a concentração
mínima bactericida (CMB) de alumínio para a E. coli (estirpe JM 109) após 2 e 24 h de
incubação em meio Luria-Bertani enriquecido com vitamina B1 é de, respectivamente,
>3,04*10-1 M e 1,90*10-2 M (Harrison et al., 2005). Na gama de concentrações de 7,5*10-7 -
7,5*10-5 M, o AlCl3 é tóxico para E. coli (estirpe KF-4) em NaCl a 0,85%, sendo a sua
toxicidade acentuada pela acidificação do meio de cultura (Bojic et al., 2002). Os mesmos
investigadores observaram que em meio de cultura constituído por peptona 10,0 g/L; glicose 5,0
g/L, e NaCl 3,0 g/L, NH4Cl 1,0 g/L e CaCl2 0,010 g/L, a toxicidade do AlCl3, avaliada pelo
decréscimo de UFC*mL-1, apenas é detectável na presença de AlCl3 7,5*10-5 M (a concentração
mais elevada das testadas) e em meio ácido (pH=5,2). Por fim, Guida et al. (1991) observaram
que concentrações milimolares de AlNO3 (9,0*10-4 M e 2,5*10-3 M) reduzem significativamente
a taxa específica de crescimento máximo de E. coli (estirpe KF-4) em meio de cultura rico em
nutrientes, não se observando qualquer aumento na densidade bacteriana após inoculação na
presença de 2,5*10-3 M a pH 5,4.
A análise mais detalhada dos resultados apresentados na figura 3.3 revela que o AlCl3, na
gama de concentrações de 1,8*10-4 M - 9,0*10-3 M, provoca uma dispersão acentuada dos
valores de 0NN durante as primeiras horas de exposição. Este efeito é mais facilmente
visualizado na figura 3.4, onde se apresenta a variação da percentagem de morte de células
viáveis em função do tempo de exposição a diferentes concentrações de AlCl3.
Após adição de AlCl3, em concentrações finais superiores a 1,8*10-4 M, às culturas
bacterianas, registou-se sempre um decréscimo na densidade de células viáveis. Porém, a
diminuição inicial da população de células viáveis de E. coli foi sempre acompanhada de uma
fase subsequente de crescimento bacteriano, quando as concentrações de AlCl3 não
ultrapassaram o valor de 1,0*10-2 M. A alteração induzida na dinâmica populacional é mais
evidente quando

33
6,0*10-4
-60
-30
0
30
60
90
120
0 1 2 3
120
90
60
30
0
-30
1,8*10-4
-60
-30
0
30
60
90
120
0 1 2 3
0,0*100
-60
-30
0
30
60
90
120
0 1 2 3
120
90
60
30
0
-30-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
6,0*10-3
-60
-30
0
30
60
90
120
0 1 2 3
120
90
60
30
0
-30
998988
5,6*10-3
-60
-30
0
30
60
90
120
0 1 2 3
120
90
60
30
0
-30
998988
2,7*10-3
-60
-30
0
30
60
90
120
0 1 2 3
120
90
60
30
0
-30-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
9,5*10-3
-60
-30
0
30
60
90
120
0 1 2 3998988
9,0*10-3
-60
-30
0
30
60
90
120
0 1 2 3998988
8,0*10-3
-60
-30
0
30
60
90
120
0 1 2 3998988
120
90
60
30
0
-30-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
-8
-6
-4
-2
0
2
0 1 2 3
LO
G (
N/N
0)
8 98 998
9,0*10-2
-60
-30
0
30
60
90
120
0 1 2 3998988
3,0*10-2
-60
-30
0
30
60
90
120
0 1 2 3998988
1,0*10-2
-60
-30
0
30
60
90
120
0 1 2 3998988
120
90
60
30
0
-30
TEMPOEXPOSIÇÃO (h)
%
DE
MO
RT
E
Figura 3.4. Morte de células de E. coli em cultura em função do tempo de exposição a AlCl3. As condições experimentais são as mesmas das descritas na legenda da figura 3.3. Os valores representam a média de pelo menos um ensaio independente realizado em quadruplicado. Os valores apresentados nos gráficos inclusos são os que se encontram na figura 3.2.
as culturas foram expostas a concentrações de AlCl3 compreendidas entre 6,0*10-4 M e 8,0*10-3
M, onde se observou que durante as primeiras 12 h após o início da exposição, a densidade
34
populacional atingiu valores iguais aos registados antes da adição de AlCl3 às culturas. Na
ausência de alumínio, este período corresponde à fase estacionária, durante a qual a densidade
populacional se manteve sensivelmente igual à registada no final da fase exponencial.
Com o objectivo de caracterizar o efeito tóxico do alumínio nas condições experimentais
utilizadas, procedeu-se à análise das curvas de “dose-efeito”, usando dois parâmetros distintos,
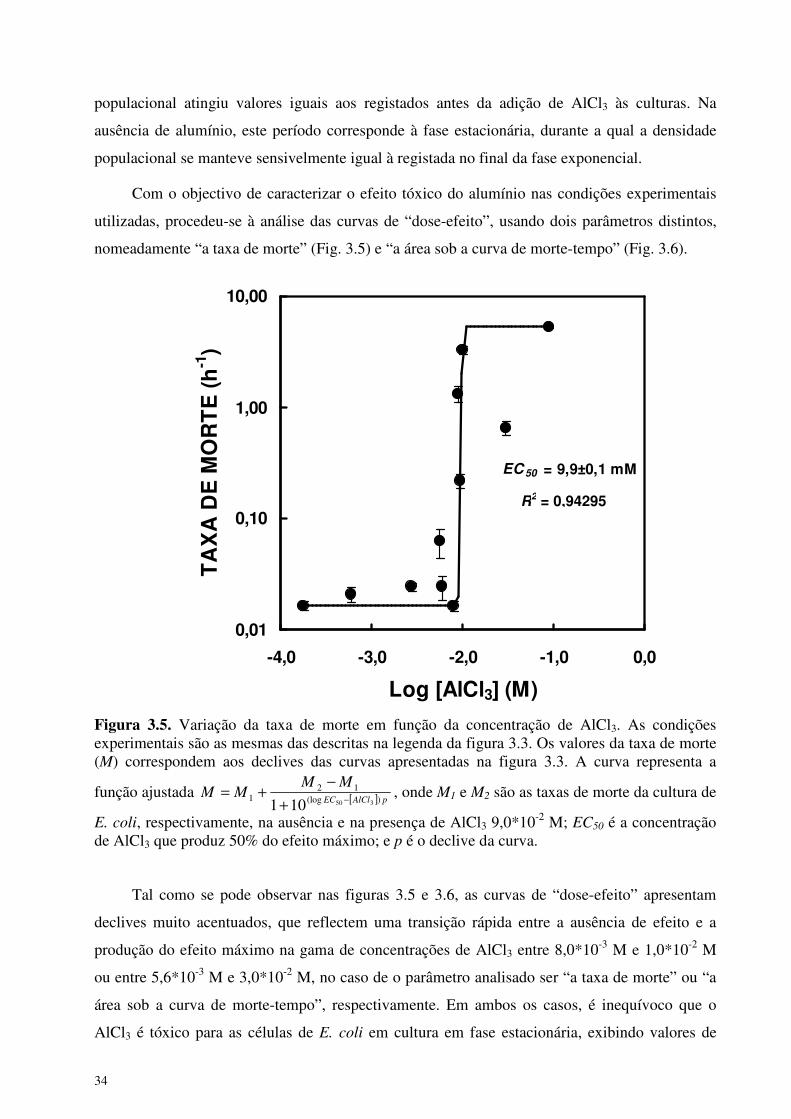
nomeadamente “a taxa de morte” (Fig. 3.5) e “a área sob a curva de morte-tempo” (Fig. 3.6).
0,01
0,10
1,00
10,00
-4,0 -3,0 -2,0 -1,0 0,0
Log [AlCl3] (M)
TA
XA
DE
MO
RT
E (h
-1)
EC 50 = 9,9±0,1 mM
Figura 3.5. Variação da taxa de morte em função da concentração de AlCl3. As condições experimentais são as mesmas das descritas na legenda da figura 3.3. Os valores da taxa de morte (M) correspondem aos declives das curvas apresentadas na figura 3.3. A curva representa a
função ajustada [ ] pAlClEC
MMMM
)(log12
1350101 −+
−+= , onde M1 e M2 são as taxas de morte da cultura de
E. coli, respectivamente, na ausência e na presença de AlCl3 9,0*10-2 M; EC50 é a concentração de AlCl3 que produz 50% do efeito máximo; e p é o declive da curva.
Tal como se pode observar nas figuras 3.5 e 3.6, as curvas de “dose-efeito” apresentam
declives muito acentuados, que reflectem uma transição rápida entre a ausência de efeito e a
produção do efeito máximo na gama de concentrações de AlCl3 entre 8,0*10-3 M e 1,0*10-2 M
ou entre 5,6*10-3 M e 3,0*10-2 M, no caso de o parâmetro analisado ser “a taxa de morte” ou “a
área sob a curva de morte-tempo”, respectivamente. Em ambos os casos, é inequívoco que o
AlCl3 é tóxico para as células de E. coli em cultura em fase estacionária, exibindo valores de
R2 = 0,94295

35
EC50 na ordem de concentrações milimolares, nomeadamente entre 5,3±0,5 mM e 9,9±0,1 mM,
em função do parâmetro usado para estimar o decréscimo, induzido por alumínio, da densidade
de células viáveis na cultura.
0
20
40
60
80
100
-4,0 -3,5 -3,0 -2,5 -2,0 -1,5 -1,0
LOG [AlCl3] (M)
ÁR
EA
SO
B A
CU
RV
A D
E T
EM
PO
-M
OR
TE
(%
do
máx
imo
)
EC50 = 5,3±0,5 mM
R2 =0,97477
TEMPOEXPOSIÇ ÃO (h) EC50 (mM) R2
1 5,5 ± 5,6 0,465073 5,9 ± 5,2 0,519948 5,3 ± 0,5 0,97477
12 7,9 ± 1,9 0,9210616 5,8 ± 0,5 0,9488736 6,3 ± 1,3 0,76957100 7,3 ± 2,9 0,55163
Figura 3.6. Variação da área sob a curva de morte-tempo em função da concentração de AlCl3. As condições experimentais são as mesmas das descritas na legenda da figura 3.3. Os valores da área sob a curva de morte-tempo (A) correspondem à percentagem das áreas sob a curvas de morte-tempo apresentadas na figura 3.3 para as diferentes concentrações de AlCl3, considerando a área sob a curva de morte-tempo na ausência de AlCl3 igual a 100% (A1) e a da respectiva curva na presença de AlCl3 9,0*10-2 M (A2) igual a 0%. A curva representa a função ajustada
[ ] pAlClEC
AAAA
)(log12
1350101 −+
−+= para 8 h de exposição. Na tabela inclusa são apresentados os valores
de EC50 calculados para os tempos de exposição indicados.
As áreas limitadas pelas funções integráveis das curvas de morte-tempo e pela recta
definida pelo limite do método de detecção de células viáveis usado correspondem aos
respectivos integrais definidos para cada tempo de exposição. Por conseguinte, quanto maior é o
incremento da área sob a curva morte-tempo ao longo do tempo mais células viáveis existem na
população bacteriana, enquanto que a sua morte é reflectida pela inalteração do valor deste
parâmetro.

36
A título ilustrativo da variação da área sob a curva de morte-tempo durante as primeiras
100 horas de exposição a AlCl3, apresenta-se na figura 3.7 a dependência deste parâmetro em
função do tempo de exposição a AlCl3 6,0*10-4 M, 6,0*10-3 M e 9,0*10-2 M, bem como na
condição controlo (ausência de AlCl3 adicionado). As concentrações de AlCl3 escolhidas podem
ser consideradas como exemplares dos efeitos do alumínio na gama de concentrações do valor de
EC50 determinado (6,0*10-3 M) e aproximadamente dez vezes inferior (6,0*10-4 M) e superior
(9,0*10-2 M).
0
5
10
15
20
25
30
35
40
0 20 40 60 80 100
TEMPO (h)
ÁR
EA
SO
B A
CU
RV
A D
E M
OR
TE
-T
EM
PO
(u
.a.)
0,00E+00
6,00E-04
6,00E-03
9,00E-02
0,0*100 M
6,0*10-4 M
6,0*10-3 M
9,0*10-2 M
0
3
6
0 3 6 9
Figura 3.7. Variação da área sob a curva de morte-tempo em função do tempo de exposição a AlCl3. As condições experimentais são as mesmas das descritas na legenda da figura 3.3. Os valores da área sob a curva de morte-tempo foram calculados a partir da curvas de morte-tempo apresentadas na figura 3.3, na ausência (●) e na presença de AlCl3 6,0*10-4 M (▲), 6,0*10-3 M (■) e 9,0*10-2 M (♦). O gráfico incluso amplia as primeiras 10 h de exposição a AlCl3.
Em condições controlo, é notória a proporcionalidade directa entre o valor da área sob a
curva de morte-tempo e o tempo de exposição até às 16 h, isto é, desde as 17 h e as 33 h após
inoculação dos meios de cultura com E. coli. A partir desse momento, o declive da curva é
gradualmente menor, reflectindo o início da fase de declínio da cultura bacteriana, caracterizada
pelo decréscimo exponencial da densidade de células viáveis. Na presença de AlCl3 9,0*10-2 M o

37
valor da área sob a curva de morte-tempo é muito baixo e constante ao longo do tempo, o que
traduz a morte rápida de células de E. coli induzida por alumínio, tal como já descrito
anteriormente (Figs. 3.3 e 3.4). Quando as culturas de E. coli foram expostas a AlCl3 6,0*10-4 M
e 6,0*10-3 M, a área da curva de morte-tempo manteve-se inferior à característica da condição
controlo durante 8 e 36 h, respectivamente. Porém, para tempos de exposição mais longos o seu
valor foi superior ao registado na ausência de AlCl3 adicionado ao meio de cultura. Por outro
lado, as formas das dependências da área sob a curva de morte-tempo em função do tempo de
exposição a AlCl3 6,0*10-4 M e 6,0*10-3 M são diferentes entre si e diferentes das obtidas na
ausência e na presença de AlCl3 9,0*10-2 M. Ambas exibem um “plateau” inicial, isto é, um
período durante o qual ocorreu diminuição da densidade de células viáveis na cultura bacteriana,
seguido duma fase de aumento do número de células viáveis, mais rápido na presença de AlCl3
6,0*10-4 M do que de 6,0*10-3 M.
Os resultados apresentados nas figuras 3.3 a 3.7 ilustram o efeito do AlCl3 na dinâmica
populacional de E. coli (estirpe JM 109) nas condições experimentais usadas. A adição de AlCl3,
na gama de concentrações milimolares, reduz a densidade de células viáveis em cultura. As
culturas de E. coli em fase estacionária, quando injuriadas por AlCl3, exibem a capacidade de
recuperar a densidade populacional máxima quando injuriadas por concentrações de AlCl3,
inferiores a ~8,0*10-3M. Os resultados obtidos sugerem que tanto a velocidade de morte induzida
como a magnitude da redução inicial da densidade de células viáveis são directamente
proporcionais à concentração de AlCl3 adicionado. São vários os modelos que se podem propor
para explicar o efeito do AlCl3 na dinâmica populacional de E. coli. Neste trabalho, testou-se o
modelo apresentado em “Materiais e Métodos”, que consiste numa adaptação do modelo
farmacodinâmico proposto por Yano et al. (1998) para explicar o mecanismo de resistência
microbiana a antibióticos.
A ocorrência de subpopulações discretas em populações geneticamente homogéneas de E.
coli está documentada na literatura (Balaban et al., 2004; Kussell et al., 2005; Halász et al.,
2007). Estas subpopulações, designadas de persistentes, parecem ser caracterizadas por taxas
específicas de crescimento máximo mais reduzidas e menor sensibilidade a agentes stressantes.
Em contraste com as células de E. coli resistentes, as células produzidas pelas subpopulações
persistentes, são sensíveis aos agentes stressantes. O aparecimento de persistentes em cultura
parece ser favorecido durante a fase estacionária (persistentes do tipo I), mas ocorre também
durante as fases de crescimento bacteriano anteriores (persistentes do tipo II ou espontâneos)
com a frequência de 1 em cada 106 células de E. coli. A existência de subpopulações persistentes
numa cultura bacteriana pode explicar a existência de ciclos aleatórios de “declínio-crescimento-

38
declínio-crescimento” tal como observado nas figuras 3.2 a 3.4. Por conseguinte, considerou-se,
para efeitos de modelização do efeito do AlCl3 na dinâmica populacional de E. coli, a existência
de uma subpopulação de persistentes com menor sensibilidade a AlCl3 que, em resposta à sua
adição ao meio de cultura, continuariam a assegurar a subsistência da população de E. coli com
as características iniciais. Os resultados da simulação efectuada são apresentados na figura 3.8.
-8
-7
-6
-5
-4
-3
-2
-1
0
1
0 25 50 75 100 125
TEMPOEXPOSIÇÃO (h)
LO
G (N
/N0)
0.00E+00
0.00E+00
6.00E-04
6.00E-04
6.00E-03
6.00E-03
1.00E-02
1.00E-02
3.00E-02
3.00E-02
9.00E-02
9.00E-02
Figura 3.8. Simulação do efeito do AlCl3 na dinâmica populacional de E. coli . A simulação foi efectuada de acordo com o modelo apresentado em “Materiais e Métodos”. Os valores dos parâmetros de simulação foram os seguintes: β1’-β1’’ e β2’-β2’’, respectivamente, -1 e 2 (durante a fase estacionária) e -9 (durante a fase de declínio); Ni 8,00*10-7; N0 3,18*10-9; '
maxV e ''maxV
5,25; '50EC e ''
50EC , respectivamente, 9,0*10-3 e 1,8*10-4. As linhas e os símbolos representam,
respectivamente, os valores gerados pela simulação e os dados experimentais para as concentrações molares de AlCl3 indicadas.

39
Tal como se pode observar na figura 3.8, os valores atribuídos às constantes parecem
adequados para simular o efeito do AlCl3 na dinâmica populacional de E. coli na ausência de
alumínio, bem como nas gamas de concentrações submicromolares e superiores a AlCl3 9,0*10-3
M. No entanto, para a gama intermédia de concentrações é ainda necessário proceder à afinação
dos parâmetros de simulação para reproduzir o comportamento observado experimentalmente.
3.2 Efeito do AlCl3 na actividade de (Na+/K+)ATPase de E. coli
A segunda parte deste trabalho foi dedicada ao estudo do efeito do AlCl3 na actividade de
(Na+/K+)ATPase das células de E. coli. Para facilitar o acesso do ATP ao centro catalítico da
enzima, procedeu-se à preparação de esferoplastos a partir de culturas de E. coli em fase
estacionária, que foram congelados e descongelados antes dos ensaios, tal como descrito em
“Materiais e Métodos”. A (Na+/K+)ATPase utiliza Mg-ATP como fonte energética para o
transporte activo de Na+e K+ e é inibida especificamente por ubaína. Por cada molécula de ATP
hidrolisada é libertado um protão. Por conseguinte, a actividade de hidrolítica de ATP dos
esferoplastos foi medida por quantificação dos H+ produzidos em resposta a adição de Mg-ATP
na ausência e na presença de ubaína (Fig. 3.9).
A B
A
Figura 3.9. Registo da actividade ATPásica de esferoplastos de E. coli. Os esferoplastos (0,1 mgproteína/mL) foram previamente incubados na ausência (A) e na presença de ubaína 20 mM (B) em NaCl 110 mM, KCl 15 mM, MgCl2 10 mM, EGTA 100 µM e HEPES-Na 2 mM) a pH 7,4, a 35ºC e com agitação magnética constante. As reacções foram iniciadas pela adição de Mg-ATP 1 mM (concentração final) e finalizadas por adições sucessivas de HCl para calibração do sinal. As

40
setas indicam a adição de 20 µL de Mg-ATP 100 mM e duas adições consecutivas de 25 µL de HCl 3 mM. A velocidade do registador foi fixada a 10 mm/min.
A adição de Mg-ATP à suspensão de esferoplastos em meio iónico de composição
adequada para a (Na+/K+)ATPase causa a acidificação do meio de reacção (Fig. 3.9 (A)) que foi
claramente menor na presença de ubaína (Fig. 3.9 (B)). A actividade de (Na+/K+)ATPase foi
calculada como a diferença entre a actividade de ATPase na ausência e na presença de 20 mM de
ubaína e expressa em nmoles de H+/ min/ mgproteína. Para o efeito, procedeu-se inicialmente à
determinação da concentração de Mg-ATP saturante e da concentração de ubaína que inibia
maximamente a actividade hidrolítica de Mg-ATP (resultados não apresentados), bem como à
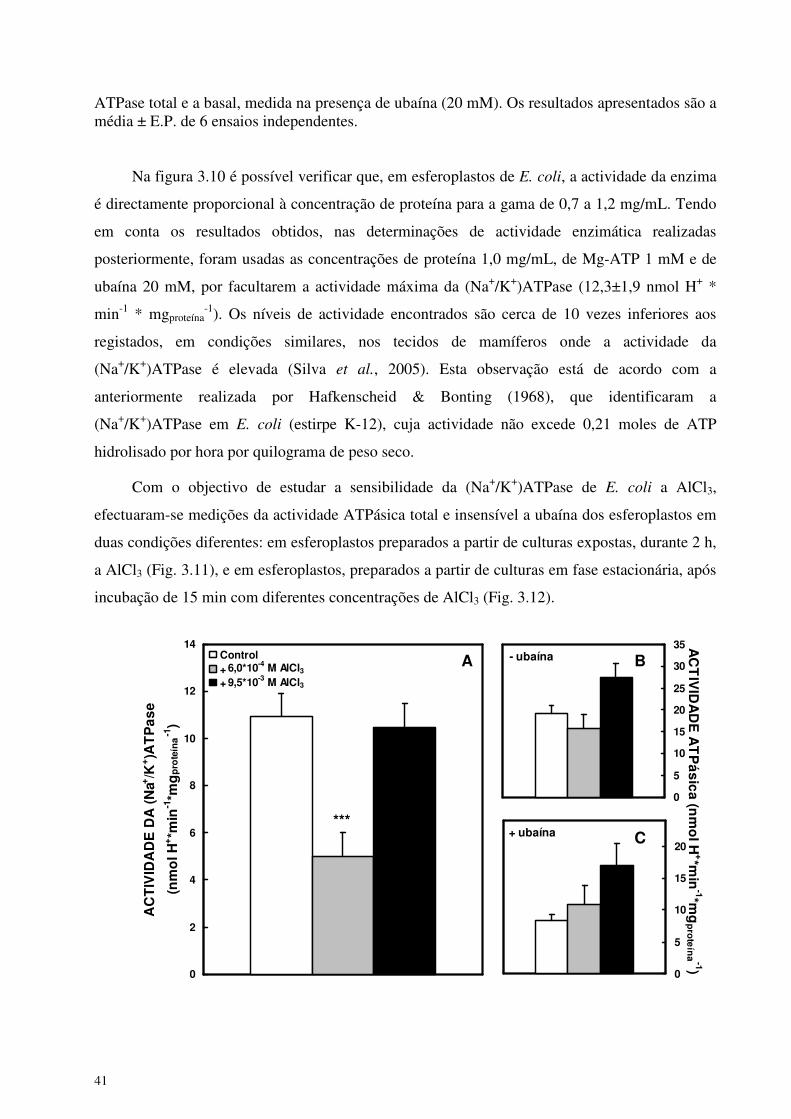
determinação da concentração óptima de proteína no ensaio (Fig. 3.10). A quantidade de H+ foi
sempre avaliada durante o período em que a sua produção era directamente proporcional ao
tempo de reacção.
0
5
10
15
20
25
30
35
0 1 2 3 4 5
PROTEÍNA (mg)
AC
TIV
IDA
DE
DA
(Na+
/K+)A
TP
ase
(nm
ol H
+*m
in-1
)
[Proteína] = 0,7 mg*ml-1
- ubaína
+ ubaína
0 5 10 15 20
PRODUÇÃO DE H+ (nmol*min-1)
[Proteína] = 1,0 mg*ml-1
+ ubaína
- ubaína
0 5 10 15 20 25 30 35 40 45
PRODUÇÃO DE H+ (nmol*min-1)
[Proteína] = 1,2 mg*ml-1
- ubaína
+ ubaína
0 5 10 15 20 25 30 35 40 45 50 55
PRODUÇÃO DE H+ (nmol*min-1)
Figura 3.10. Efeito da concentração de proteína na actividade da (Na+/K+)ATPase. A actividade da (Na+/K+)ATPase foi medida em esferoplastos isolados a partir de cultura de E coli. As reacções decorreram a 35ºC em 2 ml de meio (NaCl 110 mM, KCl 15 mM, MgCl2 10 mM, EGTA 100 µM e HEPES-Na 2mM a pH 7,4). As reacções foram iniciadas pela adição de 1 mM de Mg-ATP. A actividade da (Na+/K+)ATPase foi determinada considerando a diferença entre a
41
ATPase total e a basal, medida na presença de ubaína (20 mM). Os resultados apresentados são a média ± E.P. de 6 ensaios independentes.
Na figura 3.10 é possível verificar que, em esferoplastos de E. coli, a actividade da enzima
é directamente proporcional à concentração de proteína para a gama de 0,7 a 1,2 mg/mL. Tendo
em conta os resultados obtidos, nas determinações de actividade enzimática realizadas
posteriormente, foram usadas as concentrações de proteína 1,0 mg/mL, de Mg-ATP 1 mM e de
ubaína 20 mM, por facultarem a actividade máxima da (Na+/K+)ATPase (12,3±1,9 nmol H+ *
min-1 * mgproteína-1). Os níveis de actividade encontrados são cerca de 10 vezes inferiores aos
registados, em condições similares, nos tecidos de mamíferos onde a actividade da
(Na+/K+)ATPase é elevada (Silva et al., 2005). Esta observação está de acordo com a
anteriormente realizada por Hafkenscheid & Bonting (1968), que identificaram a
(Na+/K+)ATPase em E. coli (estirpe K-12), cuja actividade não excede 0,21 moles de ATP
hidrolisado por hora por quilograma de peso seco.
Com o objectivo de estudar a sensibilidade da (Na+/K+)ATPase de E. coli a AlCl3,
efectuaram-se medições da actividade ATPásica total e insensível a ubaína dos esferoplastos em
duas condições diferentes: em esferoplastos preparados a partir de culturas expostas, durante 2 h,
a AlCl3 (Fig. 3.11), e em esferoplastos, preparados a partir de culturas em fase estacionária, após
incubação de 15 min com diferentes concentrações de AlCl3 (Fig. 3.12).
- ubaína
0
0,2
0,4
0,6
0,8
1
1,2
0
5
10
15
20
25
30
35
+ ubaína
0
0,2
0,4
0,6
0,8
1
1,2
0
5
10
15
20
***
0
2
4
6
8
10
12
14
AC
TIV
IDA
DE
DA
(N
a+/K
+)A
TP
as
e
(nm
ol H
+*m
in-1
*mg
pro
teín
a-1
)
Control+ +
6,0*10-4 M AlCl39,5*10-3 M AlCl3
AC
TIV
IDA
DE
AT
Pá
sic
a (n
mo
l H+*m
in-1*m
gp
rote
ína
-1)
A B
C
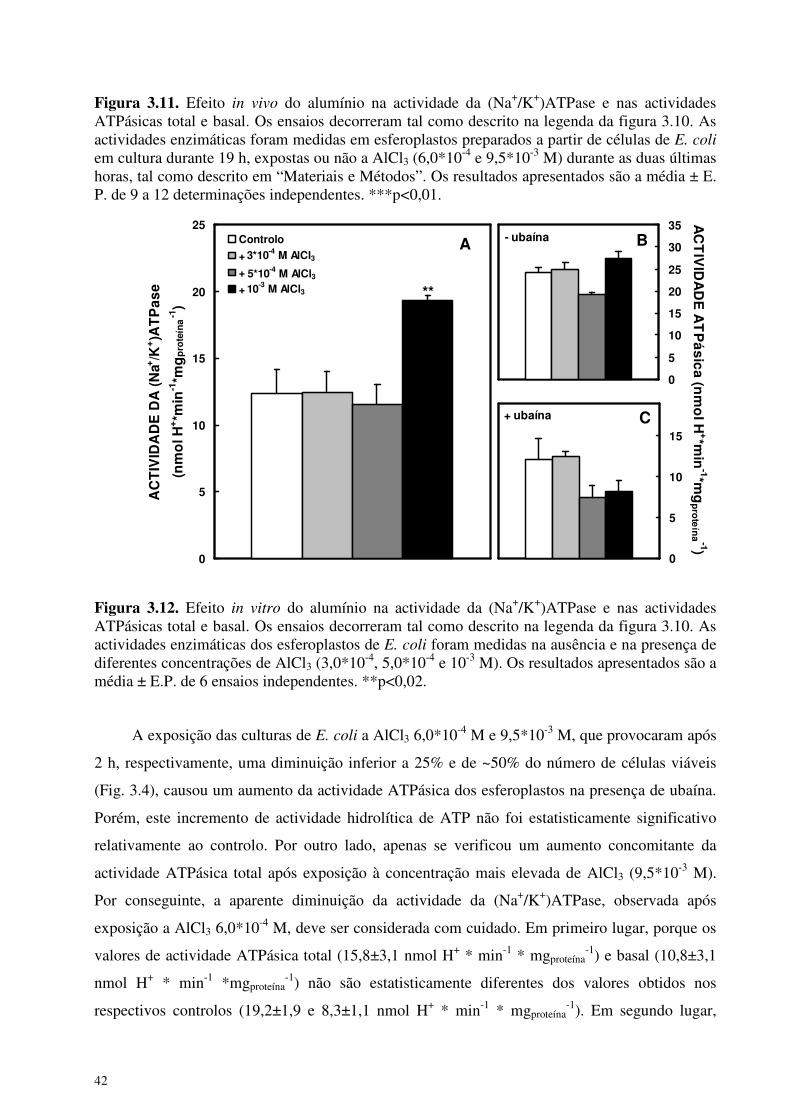
42
Figura 3.11. Efeito in vivo do alumínio na actividade da (Na+/K+)ATPase e nas actividades ATPásicas total e basal. Os ensaios decorreram tal como descrito na legenda da figura 3.10. As actividades enzimáticas foram medidas em esferoplastos preparados a partir de células de E. coli em cultura durante 19 h, expostas ou não a AlCl3 (6,0*10-4 e 9,5*10-3 M) durante as duas últimas horas, tal como descrito em “Materiais e Métodos”. Os resultados apresentados são a média ± E. P. de 9 a 12 determinações independentes. ***p<0,01.
- ubaína
0
0,2
0,4
0,6
0,8
1
1,2
0
5
10
15
20
25
30
35
+ ubaína
0
0,2
0,4
0,6
0,8
1
1,2
0
5
10
15
**
0
5
10
15
20
25
AC
TIV
IDA
DE
DA
(Na+
/K+)A
TP
ase
(nm
ol H
+*m
in-1
*mg
pro
teín
a-1
)
Controlo
+
+
+
3*10-4 M AlCl3
5*10-4 M AlCl310-3 M AlCl3
AC
TIV
IDA
DE
AT
Pá
sic
a (n
mo
l H+*m
in-1*m
gp
rote
ína
-1)
A BA
CA
Figura 3.12. Efeito in vitro do alumínio na actividade da (Na+/K+)ATPase e nas actividades ATPásicas total e basal. Os ensaios decorreram tal como descrito na legenda da figura 3.10. As actividades enzimáticas dos esferoplastos de E. coli foram medidas na ausência e na presença de diferentes concentrações de AlCl3 (3,0*10-4, 5,0*10-4 e 10-3 M). Os resultados apresentados são a média ± E.P. de 6 ensaios independentes. **p<0,02.
A exposição das culturas de E. coli a AlCl3 6,0*10-4 M e 9,5*10-3 M, que provocaram após
2 h, respectivamente, uma diminuição inferior a 25% e de ~50% do número de células viáveis
(Fig. 3.4), causou um aumento da actividade ATPásica dos esferoplastos na presença de ubaína.
Porém, este incremento de actividade hidrolítica de ATP não foi estatisticamente significativo
relativamente ao controlo. Por outro lado, apenas se verificou um aumento concomitante da
actividade ATPásica total após exposição à concentração mais elevada de AlCl3 (9,5*10-3 M).
Por conseguinte, a aparente diminuição da actividade da (Na+/K+)ATPase, observada após
exposição a AlCl3 6,0*10-4 M, deve ser considerada com cuidado. Em primeiro lugar, porque os
valores de actividade ATPásica total (15,8±3,1 nmol H+ * min-1 * mgproteína-1) e basal (10,8±3,1
nmol H+ * min-1 *mgproteína-1) não são estatisticamente diferentes dos valores obtidos nos
respectivos controlos (19,2±1,9 e 8,3±1,1 nmol H+ * min-1 * mgproteína-1). Em segundo lugar,

43
porque o aumento da actividade hidrolítica de ATP na presença de ubaína pode ser devido não só
à activação de ATPases insensíveis a ubaína, como também à diminuição da sensibilidade da
(Na+/K+)ATPase a este inibidor específico (Caspers et al., 1990; 1994).
O efeito da exposição in vivo a AlCl3 na actividade hidrolítica de ATP de E.coli foi
comparado com o efeito produzido pela presença de AlCl3 no meio de reacção. Tal como se pode
observar na figura 3.12, e à semelhança do verificado nos ensaios de exposição in vivo (Fig.
3.11), não se registaram diferenças, estatisticamente significativas relativamente ao controlo, da
actividade ATPásica na ausência e na presença de ubaína. Contrariamente ao observado
anteriormente, a actividade hidrolítica de ATP insensível a ubaína parece ser tendencialmente
menor na presença de concentrações de AlCl3 superiores a 3*10-4 M. Pelas mesmas razões
anteriormente invocadas, a aparente estimulação da (Na+/K+)ATPase de E. coli por AlCl3 10-3 M
deve ser interpretada com reservas. Os resultados obtidos parecem sugerir que a actividade da
(Na+/K+)ATPase de E. coli não é significativamente alterada por concentrações submilimolares
de AlCl3.

44

45
4. Conclusões

46

47
A toxicidade de alumínio para a E. coli (estirpe JM 109) foi estudada em meio líquido de
crescimento LB (Boillon – Miller, pH 7,0) a 37ºC com agitação de 200 rpm.
• O AlCl3, na gama de concentrações milimolares, reduziu drasticamente a capacidade de
sobrevivência das células de E. coli durante a fase estacionária.
o A adição de AlCl3, após 17 horas de incubação, diminui o número de UFC presentes,
de uma maneira dependente do tempo de amostragem.
o A análise das curvas de “dose-efeito” que o AlCl3 é tóxico para as células de E. coli
em cultura em fase estacionária, exibindo valores de EC50 na ordem de concentrações
milimolares, nomeadamente entre 5,3*10-3 M e 9,9*10-3 M, em função do parâmetro
usado para estimar o decréscimo, induzido por alumínio, da densidade de células
viáveis na cultura. A aplicação dum modelo farmacodinâmico de simulação das curvas
de morte-tempo na presença de diferentes concentrações de AlCl3 revelou ainda a
possibilidade de existência de duas subpopulações nas culturas de E. coli, cujas
sensibilidades a alumínio foram estimadas em 9,0*10-3 M e 1,8*10-4 M,
respectivamente.
• Apesar do alumínio ser tóxico para E. coli, a (Na+/K+)ATPase de E. coli não parece ser
sensível a alumínio.
o A actividade de (Na+/K+)ATPase de esferoplastos preparados a partir de células de E.
coli previamente expostas a AlCl3 9,5*10-3 M permaneceu idêntica à medida em
condições controlo (esferoplastos preparados a partir de células de E. coli não expostas
a AlCl3).
o Não se observou inibição da (Na+/K+)ATPase induzida por alumínio quando se
adicionou, directamente ao meio de reacção, AlCl3 (concentração final até 1,0*10-3
M).

48

49
5. Bibliografia

50

51
Alfrey A.C., Mishell J.M., Burks J., Contiguglia S.R., Rudolph H., Lewin E. & Holmes J.H.
(1972). Syndrome of dyspraxia and multifocal seizures associated with chronic
hemodialysis. Trans. Am. Soc. Artif. Intern. Organs. 18: 257-261.
Alfrey A.C., LeGendre G.R. & Kaehny W.D. (1976). The dialysis encephalopathy syndrome.
Possible aluminum intoxication. N. Engl. J. Med. 294: 184-188.
Amanatidou A., Smid E.J. & Gorris L.G.M. (1999). Effect of elevated oxygen and carbon
dioxide on the surface growth of vegetable-associated micro-organisms. J. Appl.
Microbiol. 86: 429-438.
Balaban N.Q., Merrin J., Chait R., Kowalik L. & Leibler S. (2004). Bacterial persistence as a
phenotypic switch. Science 305: 1622-1625.
Berrebi-Bertrand I., Lelièvre L.G., Mouas C. & Swynghedauw B. (1990). Inotropic effect of
ouabain in hypertrophied rat heart. Pflugers Arch. 417: 247-254.
Berg R.D. (1996). The indigenous gastrointestinal microflora. Trends in Microbiology. 4:
430-435.
Birdsell D.C. & Cota-Robles E.H. (1967). Production and Ultrastructure of Lysozyme and
Ethylenediaminetetraacetate-Lysozyme Spheroplasts of Escherichia coli. Department
of Life Sciences, University of California, Riverside, California. Bacteriol. 93: 427-437.
Blanco G., Berberian G. & Beaugé L. (1990). Detection of a highly ouabain sensitive isoform
of rat brainstem Na,K-ATPase. Biochim. Biophys. Acta. 1027: 1-7.
Blanco G. & Mercer R.W. (1998). Isozymes of the Na+/K+-ATPase: heterogeneity in
structure, diversity in function. Am. J. Physiol. 275: F633-650.
Bokovay G. (1988). Aluminum. (Canadian minerals yearbook). Mineral Resources Branch. 26:
8.1-8.
Bojić A., Purenović M., Kocić B., Mihailović D. & Bojić D. (2002). The comparison of
aluminium effects and its uptake by Escherichia coli in different media. Cent. Eur. J.
Public Health. 10: 66-71.
Boldyrev A.A. (2001). Na/K-ATPase as an oligomeric ensemble. Biochemistry (Mosc). 66:
821-831.
Caballero B., Trugo L.C. & Finglas P.M. (2003). Encyclopedia of Food sciences and
Nutrition. (Second edition) – Academic Press, Elsevier Science, pp. 156-165.
Caspers M.L., Kwaiser T.M. & Grammas P. (1990). Control of [3H]ouabain binding to
cerebromicrovascular (Na+ + K+)-ATPase by metal ions and proteins. Biochem.
Pharmacol. 39: 1891-1895.

52
Caspers M.L., Dow M.J., Fu M.J., Jacques P.S. & Kwaiser T.M. (1994). Aluminum-induced
alterations in [3H]ouabain binding and ATP hydrolysis catalyzed by the rat brain
synaptosomal (Na++K+)-ATPase. Mol. Chem. Neuropathol. 22: 43-55.
Cronan C.S., Walker W.J. & Bloom P.R. (1986). Predicting aqueous aluminum
concentrations in natural waters. Nature (London) 324: 140-143.
Dagley Y.S. & Hinshelwood C.N. (1938). Physicochemical aspects of bacterial growth. I.
Dependence of growth of bacterium lactis aerogenes on concentration of medium. J.
Chem. Soc. 141: 1930-1936.
De Voto E. & Yokel R.A. (1994). The Biological Speciation and Toxicokinetics of
Aluminum. Environmental Health Perspectives. 102: 940-951.
Delpy L.P., Beranger G. & Kaweh M. (1956). Bacterial growsth phases. Ann. Inst. Pasteur
(Paris) 91: 112-115.
Domingo J.L. Gomez M., Sanches D.J. & Corbella J. (1993). Effect of various dietary
constituents on gastrointestinal absoption of aluminium from drinking water and
diet. Res. Commun. Chem. Pathol. Pharmacol. 79: 377-380.
Driscoll C.T. & Schecher W.D. (1988). Aqueous chemistry of aluminium. In: Metal ions in
biological systems. Sigel H. and Sigel A. (eds). Marcel Dekker Inc., New York. 24: 63-64.
Ecker R.E. & Lockhart W.R. (1961). Specific effect of limiting nutrient on physiological
events during culture growth. J. Bacteriol. 82: 511-516.
Elliker P.R. & Frazier W.C. (1938). Influence of time and temperature of incubation on heat
resistance of Escherichia coli. J. Bact. 36: 83-98.
Glauert A.M. & Thornley M.J. (1969). The topography of the bacterial cell wall. Annu. Rev.
Microbiol. 23: 159-198.
Guida L., Ziba S., Martin N.H. & Robert K.P. (1991). Aluminium toxicity and binding to
Escherichia coli. Arch-Microbiol. 156: 507-512.
Halász Á., Kumar V. & Rubin H. (2007). E. coli persistence as random toxin activation
followed by slow deterministic recovery. Visit: http://cnls.lanl.gov/q-bio/wiki/
images/9/98/Poster-halasz.pdf
Harrison J.J., Turner R.J. & Ceri H. (2005). High-throughput metal susceptibility testing of
microbial biofilms. BMC Microbiol. 5: 53.
Hafkenscheid J.C. & Bonting S.L. (1968). Studies on (Na+-K+)-activated ATPase. XIX.
Occurrence and properties of a (Na+-K+)-activated ATPase in Escherichia coli.
Biochim. Biophys. Acta. 151: 204-211.

53
Hofschneider P.H. & Wandstructur Z. (1960). Escherichia coli B Sphaeroplasten. Proc.
European Regional Conf. Electron Microscopy Delft, vol. 2, pp. 1028-1032.
Jewell E.A. & Lingrel J.B. (1991). Comparison of the substrate dependence properties of the
rat Na+, K+-ATPase alpha 1, alpha 2, and alpha 3 isoforms expressed in HeLa cells. J.
Biol. Chem. 266: 16925-16930.
Jorgensen P.L., Hakansson K.O. & Karlish S.J.D. (2003). Structure and mechanism of Na, K-
ATPase: Functional Sites and Their Interactions. Annu. Rev. Physiol. 65: 817-49.
Kaplan J.H. (2002). Biochemistry of Na,K-ATPase. Annu. Rev. Biochem. 71: 511-535.
Kimelberg H.K. & Papahadjopoulos D. (1972). Phospholipid requirements for (Na++K+)-
ATPase activity: head-group specificity and fatty acid fluidity. Biochim. Biophys. Acta.
282: 277-292.
Klaassen C.D. & Watkins J.B. (2003). Casarett & Doull's Essentials of Toxicology (Book).
McGraw-Hill Professional. 01: 512 pages.
Koseki S. & Isobe S. (2005). Prediction of pathogen growth on iceberg lettuce under real
temperature history during distribution from farm to table. Int. J. Food Microbiol.
104: 239-248.
Kussell E., Kishony R., Balaban N.K. & Leibler S. (2005). Bacterial persistence: a model of
survival in changing environments. Genetics. 169: 1807-1814.
Lai J.C., Guest J.F., Leung T.K., Lim L. & Davison. A.N. (1980). The effects of cadmium,
manganese and aluminium on sodium-potassium-activated and magnesium-activated
adenosine triphosphatase activity and choline uptake in rat brain synaptosomes.
Biochem. Pharmacol. 29: 141-146.
Layne E. (1957). Spectrophotometric and turbidimetric methods for measuring proteins.
Meth. Enzymol. 3: 447-451.
Li R.C., Nix D.E. & Schentag J.J. (1993). New turbidimetric assay for quantitation of viable
bacterial densities. Antimicrob. Agents Chemother. 37: 371-374.
Lingrel J.B. & Kuntzweiler T. (1994). Na+/K+-ATPase. J. Biol. Chem. 269: 19659-19662.
Madeira V.M.C., Antunes-Madeira M.C. & Carvalho A.P. (1974). Activation energies of the
ATPase activity of sarcoplasmic reticulum. Biochim. Biophys. Res. Commun. 58: 897-
904.
Martin D.S. (1932). The oxygen consumption of Escherichia coli during the lag and
logarithmic phases of growth. J. Gen. Physiol. 15: 691-708.

54
Maunsbach A.B., Skriver E. & Hebert H. (1991). Two-dimensional crystals and three-
dimensional structure of Na,K-ATPase analyzed by electron microscopy. Soc. Gen.
Physiol. Ser. 46: 159-172.
Miller R.G., Kopfler F.C., Ketly K.C., Stober J.A. & Ulmer N.S. (1984). The occurrence of
aluminium in drinking water. J. Am. Water Works Assoc. 76: 84-91.
Moyer L.S. (1936). Changes in the electrokinetic potential of bacteria at various phases of
the culture cycle. J. Bact. 32: 433-464.
Nelson D.J. (1996). Aluminum complexation with nucleoside di- and triphosphates and
implication in nucleoside binding proteins. Coord. Chem. Rev. 149: 95-111.
Ochman H. & Selander R.K. (1984). Standard reference strains of Escherichia coli from
natural populations. J. Bacteriol. 157: 690-693.
Oscar T.P. (1999). Response surface models for effects of temperature and previous growth
sodium chloride on growth kinetics of Salmonella typhimurium on cooked chicken
breast. J. Food Prot. 62: 1470-1474.
Padan E., Bibi E., Ito M. & Krulwich T.A. (2005). Alkaline pH homeostasis in bacteria: New
insights. Biochim. Biophys. Acta – Biomembranes. 1717: 67-88.
Piña R.G. & Cervantes C. (1996). Microbial interactions with aluminium. Biometals 9: 311-
316.
Razin S. (1975). The mycoplasma membrane. In: Danielli J.F., Rosenberg M.D. & Cadenhead
D.A. (ed.), Progress in surface and membrane science, Academic Press Inc., New York,
vol. 9, pp. 257-312.
Serluca F.C., Sidow A., Mably J.D. & Fishman M.C. (2001). Partitioning of tissue expression
accompanies multiple duplications of the (Na+/K+)ATPase alpha subunit gene.
Genome Res. 11: 1625-1631.
Shehata E.T. & Marr G.A. (1971). Effect of Nutrient Concentration on the Growth of
Escherichia coli. J. Bacteriol. 107: 210-216.
Silva V.S. & Gonçalves P.P. (2003). The inhibitory effect of aluminium on the
(Na+/K+)ATPase activity of rat brain cortex synaptosomes. J. Inorg. Biochem. 97: 143-
150.
Silva V.S., Duarte A.I., Rego A.C., Oliveira C.R. & Gonçalves P.P. (2005). Effect of Chronic
Exposure to Aluminium on Isoform Expression and Activity of Rat (Na+/K+)ATPase.
Toxicol. Sci. 88: 485-494.
Silva V.S., Nunes M.A., Cordeiro J.M., Calejo A.I., Santos S., Neves P., Sykes A., Morgado F.,
Dunant Y. & Gonçalves P.P. (2007). Comparative effects of aluminum and ouabain on

55
synaptosomal choline uptake, acetylcholine release and (Na+/K+)ATPase. Toxicology.
236: 158-177.
Silvius J.R. & McEhhaney R.N. (1982). Molecular properties of membrane lipids and
activity of a membrane adenosine triphosphatase from Acholeplasma laidlawii B.
Rev. Infect. Dis. Suppl: S50-S58.
Skou J.C. (1957). The influence of some cations on an adenosine triphosphatase from
peripheral nerves. Biochim. Biophys. Acta 23: 394-401.
Sweadner K.J. (1979). Two molecular forms of (Na++K+)-stimulated ATPase in brain.
Separation, and difference in affinity for strophanthidin. J. Biol. Chem. 254: 6060-
6067.
Sweadner K.J. (1985). Enzymatic properties of separated isozymes of the (Na+, K+)ATPase.
Substrate affinities, kinetic cooperativity, and ion transport stoichiometry. J. Biol.
Chem. 260: 11508-11513.
Taniguchi K., Kaya S., Abe K. & Mardh S. (2001). The oligomeric nature of Na/K-transport
ATPase. J. Biochem. 129: 335-342.
Wood J.M. (1988). Transport, bioaccumulation, and toxicity of metals and metalloids in
microorganisms under environmental stress. In: Kramer J.R. & Allen H.E. (ed.), Metal
speciation: theory, analysis and application. Lewis Publishers, Chelsea, Mich. pp. 309-312.
Yano Y., Nakayama A., Ishihara K. & Saito H. (1998). Adaptive Changes in Membrane
Lipids of Barophilic Bacteria in Response to Changes in Growth Pressure. Appl.
Environ. Microbiol. 64: 479-485.
Zatta P., Zambenedetti P., Pizziuti A. & Perazzolo M. (1995). Different effects of aluminum
upon carbonic anhydrases and Na+/K+-ATPase activities in rat. Neurosci. Lett. 197:
65-68.


















