O papel do contacto célula a célula na manutenção da...
54
UNIVERSIDADE DE LISBOA FACULDADE DE CIÊNCIAS DEPARTAMENTO DE BIOLOGIA ANIMAL O papel do contacto célula a célula na manutenção da diversidade bacteriana Ana Sofia Nunes Mestrado em Biologia Evolutiva e do Desenvolvimento 2012
Transcript of O papel do contacto célula a célula na manutenção da...

UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
O papel do contacto célula a célula na manutenção da diversidade bacteriana
Ana Sofia Nunes
Mestrado em Biologia Evolutiva e do Desenvolvimento
2012


UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
O papel do contacto célula a célula na manutenção da diversidade bacteriana
Ana Sofia Nunes
Dissertação orientada por: Professor Doutor Francisco Dionísio
Mestrado em Biologia Evolutiva e do Desenvolvimento
2012


AGRADECIMENTOS
Gostava de agradecer a todos aqueles que me permitiram chegar aqui, de uma outra forma.
Em primeiro lugar, quero agradecer ao Professor Francisco Dionísio por ter
aceite ser meu orientador. Por todo o apoio que me deu, pela motivação e entusiasmo mesmo quando as bactérias não cooperaram. Pelas ideias sugeridas ao longo do trabalho, pela paciência quando apareciam resultados estranhos. Agradeço a confiança que depositou em mim, todo o incentivo e disponibilidade constante.
Em segundo lugar, queria agradecer a todo o grupo de Ecologia Evolutiva de Microrganismos, em especial à Iolanda por toda a paciência que teve comigo, por me ter ensinado o abc das bactérias e por toda a ajuda durante este ano. Ao Luís Carvalho pela preciosa ajuda na Estatística e por todas as sugestões.
Queria também agradecer a todos os meus colegas de mestrado, pelo bom ambiente e espírito de camaradagem. Queria em especial agradecer à Raquel e às Joanas, pelos desabafos constantes e por todo o apoio. Por terem feito com que o ano curricular tivesse passado muito mais depressa!
Agradeço à minha família e amigos, em especial aos meus pais por me terem sempre apoiado nas minhas decisões, por me terem dado sempre todas as oportunidades e por saber que posso contar com eles para o que der e vier. À minha irmã Carla, por todas as boleias matinais e pelas gargalhadas logo pela manhã. Aos meus (futuros) sogros por me tratarem como uma filha. À Dora, por, como eu, amar os não humanos incondicionalmente.
A uma Vida muito especial na minha vida, pelo privilégio da sua companhia durante quase 15 anos da minha existência e que fez de mim uma pessoa muito mais Feliz. Que não está mais por cá mas que estará sempre comigo. Que me ensinou o que é o amor incondicional.
Por fim agradeço ao Bruno. Por Tudo.


i
RESUMO
Vários mecanismos ecológicos podem explicar a coexistência de diferentes
estirpes bacterianas. Com este trabalho, propomos que uma forma de interacção bacteriana mediada pelo contacto poderá ser um desses mecanismos. Propomos assim um novo mecanismo – Vantagem de crescimento Dependente de Contacto (VDC) – que permite a coexistência entre diferentes estirpes.
Os nossos resultados sugerem que o contacto com uma população em alta densidade celular permite às bactérias uma taxa de crescimento superior em comparação com o seu crescimento numa população em baixa densidade de bactérias. Observámos que este mecanismo está presente em três espécies distintas, evolutivamente tão distantes como Escherichia coli e Staphylococcus aureus, sugerindo que este mecanismo está amplamente distribuído no mundo bacteriano, permitindo a colonização de novos habitats ao interagirem com as estirpes já residentes.
Ao utilizarmos estirpes resistentes a antibióticos também verificámos que o contacto com uma população com uma alta densidade bacteriana lhes confere vantagem, permitindo a sua manutenção na população em baixas frequências. Este mecanismo junta-se assim a outros (p. ex. epistasia, mutações compensatórias) tornando a eliminação da resistência a antibióticos bastante mais complexa que o esperado.
PALAVRAS CHAVE: Interacções bacterianas, Contacto célula a célula, Resistência a antibióticos, Selecção dependente de frequência

ii

iii
ABSTRACT
Several ecological mechanisms have been suggested to explain the
coexistence of different bacterial strains. We propose a new mechanism that may enable the invasion of previously established populations by another population and its maintenance.
We propose a new mechanism involving cell-to-cell contact – contact-dependent growth advantage (CDA) that allows the coexistence of different strains. Here, we use antibiotic resistant clones and its susceptible ancestral and we show that the contact between resistant clone and the wild type clone provides an advantage in the resistant clone’s growth when rare.
This phenomenon was observed in three different species, both within and between species. CDA was in fact observed in two evolutionary distant bacterial species – Escherichia coli and Staphylococcus aureus –, suggesting that this interaction may probably be widely spread in the bacteria world enabling the colonization of a new habitat.
We must underline that this mechanism makes the elimination of antibiotic resistance more difficult, maintaining antibiotic resistant strains in low frequencies within the populations.
KEYWORDS: Antibiotic Resistance, Bacterial Interactions, Cell-to-cell contact, Frequency dependent selection.

iv

v
ÍNDICE RESUMO i ABSTRACT iii I. INTRODUÇÃO 1 I.1. Bactérias, luta pela sobrevivência e manutenção da diversidade bacteriana 1 I.2. Interacções bacterianas 2
I.2.1. Interacções mediadas pelo contacto célula a célula 2 I.3. Bactérias como organismos modelo em evolução experimental 3 I.4. Objectivos 4
II. MATERIAIS E MÉTODOS 7 II.1. Estirpes bacterianas utilizadas e condições de crescimento 7 II.2. Selecção de mutantes espontâneos resistentes a antibióticos 7 II.3. Ensaios com e sem contacto entre bactérias resistentes e bactérias ancestrais sensíveis 8
II.3.1. Meio líquido 8 II.3.1.1. Tubos 8 II.3.1.2. Seringas 8 II.3.1.3. Placas multipoços para cultura de células 9
II.3.2. Meio sólido 10 II.3.2.1. Meio sólido directamente em LA 10 II.3.2.2. Meio sólido em membrana porosa 10
II.4. Curvas de crescimento em Meio Mínimo 11 II.5. Ensaios em alta e baixa densidade 11 II.6. Evolução experimental 11 II.7. Estimativa do número de gerações e do fitness relativo 11 II.8. Análise estatística 12
III. RESULTADOS 13 III.1. Interacção com outras bactérias aumenta a taxa de crescimento 13
III.1.1. Meio líquido – tubos 13 III.1.2. Meio sólido 14
III.2. O contacto célula a célula é necessário para o mecanismo VDC 15 III.3. VDC actua quando o contacto se estabelece com células em estado de latência 16 III.4. O fenómeno VDC é independente da mutação que confere resistência ao ácido nalidíxico 18 III.5. Mecanismo VDC como consequência do crescimento em populações bacterianas com uma alta densidade populacional 19 III.6. VDC entre espécies diferentes 22 III.7. O caso de Enterococcus faecalis: inexistência do fenómeno VDC 22 III.8. O papel dos genes rpoS e lrp no fenómeno VDC 23 III.9. Testando o mecanismo fisiológico por detrás do fenómeno VDC: o efeito do dióxido de carbono 25 III.10. Evolução experimental 26
III.10.1. Controlo da evolução experimental e mutante ancestral 26 III.10.2. Réplicas sujeitas à pressão selectiva 27 III.10.3. Evolução - ensaios em alta e baixa densidade 28
IV. DISCUSSÃO 31 IV.1. VDC como um novo mecanismo que permite a manutenção da diversidade bacteriana 31 IV.2 Fenómeno VDC amplamente distribuído no mundo bacteriano 31

vi
IV.3. VDC e selecção dependente de frequência 33 IV.4. VDC e fases do crescimento bacteriano 33 IV.5. O papel dos genes rpoS e lrp no fenómeno VDC 34 IV.6. Fenómeno VDC: uma perspectiva evolutiva 36 IV.7. Perspectivas futuras 38
V. REFERÊNCIAS 39

1
I. INTRODUÇÃO
I.1. Bactérias, luta pela sobrevivência e manutenção da diversidade bacteriana
(W)hen the physiologist J. B. S. Haldane once commented that even the Archbishop of Canterbury is 65% water, he omitted to mention that the head of the Church of England also consists of more bacterial cells than eukaryotic cells and that the combined number of genes in these bacteria exceeds the total number of eukaryotic genes by a factor of at least 100.
(Macpherson e Harris, 2004)
Na natureza, as populações bacterianas vivem geralmente em comunidades muito complexas, nas quais várias estirpes e várias espécies bacterianas co-habitam, existindo consequentemente uma forte competição tanto por espaço como por recursos (Hibbing et al., 2010).
Um exemplo disso é o corpo humano, sendo colonizado por microrganismos logo após o nascimento, microrganismos estes que colonizam as membranas mucosas e o epitélio da pele (Macpherson e Harris, 2004). O intestino humano é colonizado por várias espécies bacterianas, atingindo cerca de 500 espécies (Berg, 1996). A colonização por algumas destas espécies continua ao longo da vida, sendo que algumas estirpes substituem as então residentes.
No caso específico de Escherichia coli, assiste-se a uma sucessão de eventos de colonização do tracto gastrointestinal humano (Berg, 1996). As células bacterianas de E. coli permanecem no intestino de mamíferos durante toda a sua vida mas, enquanto algumas estirpes podem permanecer no intestino durante meses ou anos, outras estirpes são transientes, permanecendo apenas durante poucos dias ou poucas semanas (Sears et al., 1950). Caugant et al. (1981) analisaram uma população de E. coli num hospedeiro humano durante um período de 11 meses, tendo conseguido distinguir pelo menos 12 tipos electroforéticos num único dia e um total de 53 tipos no período de 11 meses (Caugant et al., 1981). Assim, apesar da estabilidade das comunidades bacterianas ao nível da espécie (Ley et al., 2006), cada estirpe bacteriana está exposta à competição intra e interespecífica. Esta competição leva à actuação da Seleção Natural que providencia a força motora para a inovação e diversificação entre os organismos competidores (Schluter, 1996 citado por Hibbing et al., 2010).
Já foi demonstrado (Levin, 1972; Helling et al.,1987; Alves, 2010) que a selecção dependente de frequência estabilizante é muito importante na manutenção da variabilidade em populações bacterianas, sendo que poderá também desempenhar um papel fundamental na manutenção da diversidade genética observada na maior parte das espécies bacterianas (Levin, 1988).
Especificamente em E. coli, Levin (1972) e Helling et al., (1987) verificaram a ocorrência de selecção dependente de frequência, mostrando que diferentes genótipos conseguem coexistir mesmo em ambientes simples. Alves (2010), mostrou uma elevada ocorrência de selecção dependente de frequência estabilizante em estirpes de E. coli resistentes a antibióticos. Estes resultados sugerem assim que a selecção dependente de frequência poderá ser crucial para a manutenção de estirpes resistentes a antibióticos

2
que, apresentando custos de fitness na ausência de drogas e na ausência de outras mutações (p. ex. mutações compensatórias (Bjorkman et al., 2000), epistasia (Trindade et al., 2009), mutações sem custo (Ramadhan & Hegedus, 2005)), seriam eliminadas da população.
Para além das populações mencionadas anteriormente, a ocorrência de selecção dependente de frequência já foi demostrada em Drosophila (Ayala, 1971), e em vírus (Elena et al., 1997; Yuste et al., 2002).
Dois mecanismos ecológicos têm sido sugeridos para explicar a vantagem em raridade de determinadas estirpes de E. coli: cross-feeding (Turner et al., 1996) e trade-off demográfico (Levin, 1972, Turner et al., 1996). De acordo com a hipótese do cross-feeding, a vantagem do genótipo em minoria poderá advir da excreção de um metabolito resultante do metabolismo ou de morte celular do genótipo em maioria, sendo que este genótipo é eficiente a utilizar este metabolito (Turner, et al., 1996). No que concerne ao trade-off demográfico, cada genótipo é especializado num diferente nicho. Como exemplo, um dos genótipos possui um maior fitness quando existe abundância de glucose no meio, enquanto que o outro genótipo possui um fitness mais elevado quando a glucose é escassa (Turner, et al., 1996).
I.2. Interacções bacterianas The history of evolution and biodiversity is fundamentally a history of the evolution of species
interactions. Species in pure isolation simply do not make sense. (Thompson, 1999)
Há inúmeras formas de interacções bacterianas, desde interacções
mutualistas, antagonistas e ainda comportamentos sociais muito complexos (p. ex. Myxobacteria).
Não pretendendo enveredar por uma descrição exaustiva das formas de interacção descritas em bactérias, importa aqui sublinhar que as interacções bacterianas podem ser divididas em interacções “à distância” (como interacções através de quorum sensing – comunicação cooperativa, guias e manipulação química (Keller e Surette, 2006) –, produção de colicinas e de antibióticos) e interacções mediadas pelo contacto directo célula a célula. Uma vez que este trabalho trata de um mecanismo de interacção mediado pelo contacto celular, colocaremos o enfoque neste.
I.2.1. Interacções mediadas pelo contacto célula a célula Interacções antagonistas entre bactérias foram, já há muito, identificadas
(Gratia, 1925). No entanto, apenas recentemente foram descobertas interacções antagonistas envolvendo o contacto directo célula a célula. Aoki et al. (2005) descreveram um tipo de interacção intercelular no qual o crescimento bacteriano é regulado através desse contacto directo célula a célula. Os autores verificaram que determinadas estirpes de E. coli, incluindo estirpes uropatogénicas, apresentavam um sistema de inibição do crescimento bacteriano através do contacto directo entre células, observando que a inibição dependia do estado de crescimento do inibidor, ocorrendo na fase logarítmica não ocorrendo, contudo, na fase estacionária. No entanto, o

3
crescimento das células alvo era inibido independentemente da sua fase de crescimento (Aoki et al., 2005). Lemonnier et al. (2008) descreveram um tipo diferente de interacção antagonista, no qual apenas durante a fase estacionária, um dos genótipos – devido a uma mutação ou a um efeito pleiotrópico de uma mutação –, conseguia inibir o genótipo mais frequente, interferindo com o seu crescimento máximo. Estes autores verificaram que a inibição exigia também o contacto entre as bactérias.
Em bactérias também foram descritas interacções não antagonistas mediadas pelo contacto. Dubey e Ben-Yehuda (2011) verificaram a existência de nanotubos que ligam células bacterianas vizinhas, funcionando como uma rede de troca de moléculas. Através de microscopia electrónica, verificaram a presença destas extensões tubulares que ligam as células em Bacillus subtilis e entre B. subtilis e Staphylococcus aureus e ainda entre B. subtilis e E. coli, sendo esta última uma bactéria evolutivamente distante de B. subtilis. Foi assim possível constatar que estes nanotubos permitem a troca de moléculas intracelulares dentro da mesma espécie e entre espécies distintas. Complementarmente, Wei et al. (2011), verificaram a existência de transferência de lipoproteínas entre células de Myxococcus xanthus dependente do contacto célula a célula. Sabe-se ainda que esta espécie possui um programa de desenvolvimento complexo durante a formação do corpo de frutificação, programa este também dependente da comunicação directa célula a célula (Kaiser, 2004). Um outro tipo de troca molecular que envolve interacções físicas entre células vizinhas é a conjugação bacteriana (Lederberg e Tatum, 1946), responsável pela transferência de plasmídeos conjugativos.
Outros contributos são também fundamentais para compreendermos a importância de interacções mediadas pelo contacto célula a célula em bactérias. Kolenbrander et al. (2002) destacam que nas superfícies de esmalte de dentes, o contacto físico entre as bactérias colonizadoras e entre as bactérias e o seu hospedeiro é essencial para a colonização inicial e a subsequente formação de biofilmes. Os biofilmes são comunidades microbianas que conferem protecção ao crescimento bacteriano, permitindo a sobrevivência num ambiente hostil. As estruturas que formam biofilmes contêm canais em que os nutrientes circulam e em que as células das diferentes regiões do biofilme exibem padrões de expressão genética diferentes. A complexidade da estrutura e metabolismo destes conduziu mesmo à analogia entre biofilmes e tecidos de organismos superiores (Costerton et al., 1999).
I.3. Bactérias como organismos modelo em evolução experimental
Why else would we study evolution? It is true that evolutionary biology has some practical relevance – for example, to animal breeding or to the origins of antibiotic resistance – but the real reason why we study it is that we are interested in origins. We want to know where we
came from. (Maynard Smith e Szathmary, 1995)
A evolução experimental permitiu que muitas das controvérsias que visassem colocar em causa a Biologia Evolutiva enquanto ciência fossem

4
cabalmente postas de parte, permitindo a formulação de hipóteses e previsões possíveis de testar. A evolução experimental dedica-se ao estudo da evolução das populações em tempo real, incluindo elementos fundamentais tais como a manutenção de populações controlo, populações replicadas, observação de múltiplas gerações e a perspectiva de análise genética detalhada. A evolução experimental é assim a biologia evolutiva no seu senso mais empírico (Garland e Rose, 2009).
Existem vários organismos modelo utilizados em evolução experimental, desde organismos assexuados e sexuados, procariotas ou eucariotas, vertebrados ou invertebrados. No presente trabalho, colocaremos o enfoque em bactérias e na sua utilização em evolução experimental.
Foram já realizados inúmeros estudos de evolução experimental utilizando bactérias (ver Behe, 2010). Assim, algumas características essenciais destes organismos consistem nos seus tempos de geração curtos e na sua facilidade de propagação, permitindo estudos de longo termo com populações de grandes dimensões e com várias réplicas. As bactérias podem ainda ser armazenadas indefinidamente e mais tarde utilizadas, permitindo a comparação entre o ancestral e os indivíduos que foram sujeitos a evolução, tornando possível a comparação de características genotípicas e fenotípicas, incluindo o fitness relativo baseado em competições directas. Variáveis ambientais, assim como factores genéticos, podem ainda ser manipulados de modo a testar os seus efeitos nas dinâmicas evolutivas (Schneider e Lenski, 2004).
É de salientar que a evolução experimental não tem como objectivo mimetizar, reproduzir ou prever a evolução na natureza. A sua principal utilidade reside na sua capacidade em testar a teoria evolutiva e hipóteses, o que outras abordagens alternativas não possibilitam (Bennett e Lenski, 1999). I.4. Objectivos
Neste trabalho pretendeu-se estudar um novo mecanismo de interacção mediado pelo contacto célula a célula – Vantagem de crescimento Dependente de Contacto (VDC) – que possibilita a manutenção da diversidade bacteriana. Pretendemos assim verificar se este mecanismo permite que bactérias em baixa densidade que crescem em contacto com uma outra população em alta densidade apresentem uma vantagem relativamente a bactérias que crescem isoladas em baixa densidade. Tencionamos assim determinar se o crescimento com outras estirpes é vantajoso para populações que se encontram em baixa densidade, facilitando a invasão de uma população já residente e a sua manutenção em baixas frequências, viabilizando consequentemente a coexistência entre estirpes.
Tencionámos ainda verificar se este mecanismo actua em bactérias Gram-negativas e Gram-positivas, sendo ainda um objectivo determinar se o mecanismo descrito actua exclusivamente de forma intra-específica ou também entre espécies distintas.
Por último, pretendeu-se, através de evolução experimental, determinar se existe variabilidade genética que responda a pressões selectivas no que concerne ao mecanismo por nós descrito. Procurámos assim determinar se um mutante sujeito a evolução, apresentou um aumento de fitness

5
comparativamente com o mutante ancestral, quando em contacto com a estirpe ancestral sensível.
Com este trabalho pretendeu-se assim determinar a importância de uma forma de interacção que envolve o contacto célula a célula, na manutenção da diversidade bacteriana, visando-se verificar se esta permite a manutenção e evolução de estirpes resistentes a antibióticos.

6

7
II. MATERIAIS E MÉTODOS
II.1. Estirpes bacterianas utilizadas e condições de crescimento
Foram utilizadas quatro estirpes bacterianas distintas, pertencentes a quatro espécies: E. coli K12 MG1655, Salmonella enterica serovar Typhimurium LT2, Enterococcus faecalis LN68 e Staphylococcus aureus ATCC 25923. Foram também utilizados mutantes espontâneos resistentes a ácido nalidíxico, a rifampicina e a estreptomicina derivados da estirpe de E. coli K12, clones espontâneos resistentes a ácido nalidíxico derivados de S. aureus ATCC 25923 e Salmonella enterica serovar Typhimurium LT2 e clones espontâneos resistentes a rifampicina derivados de E. faecalis LN68 (uma vez que esta é intrinsecamente resistente a ácido nalidíxico).
Adicionalmente, e nos ensaios referidos, foram utilizadas as estirpes E. coli JW0334 lacY::kan, JW5437 rpoS::kan e JW0872 lrp::kan (designadas ao longo do trabalho por lacY-, rpoS- e lrp-, respectivamente) assim como mutantes espontâneos resistentes a ácido nalidíxico destas estirpes. Estas estirpes integram a Keio collection (Baba et al., 2006), que resulta de uma construção sistemática de um conjunto de mutantes knockout, mutantes estes que apresentam uma delecção definida precisamente num único gene. A Keio collection engloba mutantes para todos os genes não essenciais da estirpe E. coli K12 BW25113 (estirpe com um pedigree bem definido que não foi sujeito a mutagénese e que está estreitamente relacionada com a estirpe MG1655) (Baba et al., 2006).
Relativamente às condições de crescimento, as estirpes foram incubadas em meio Luria Bertani líquido (LB) ou em LB suplementado com agar (LA) (meio sólido) a uma temperatura de 37ºC com uma agitação constante (170 rpm).
Quando necessário, os meios foram suplementados com as seguintes concentrações de antibióticos: 40 µg/ml para o ácido nalidíxico, 100 µg/ml para a rifampicina e 100 µg/ml para a estreptomicina.
Todas as diluições foram realizadas numa solução tampão de MgSO4 10-2 M e os clones foram armazenados em glicerol a 15% (V/V) a -20 °C de modo a poderem ser reutilizados em experiências futuras. II.2. Selecção de mutantes espontâneos resistentes a antibióticos
Os mutantes espontâneos resistentes a ácido nalidíxico, rifampicina e estreptomicina derivados da estirpe E. coli K12 tinham já sido isolados, pelo que foram seleccionados apenas mutantes espontâneos de E. faecalis LN68 resistentes a rifampicina e de S. aureus, Salmonella e E. coli lacY-, rpoS- e lrp-, resistentes a ácido nalidíxico.
Para a selecção de mutantes espontâneos resistentes a antibióticos, foram inoculados vários tubos contendo 10 ml de LB com cada uma das 6 estirpes ancestrais anteriormente referidas, que cresceram durante 24 horas a 37°C e com agitação constante (170 rpm). Foram plaqueados 100µl de cada cultura

8
em LA suplementado com o respectivo antibiótico, com posterior incubação a 37°C, overnight.
Após a obtenção de estirpes mutantes, procedeu-se ao isolamento de colónias (2 isolamentos) de modo a assegurar que todos os mutantes são monoclonais. Após o seu isolamento, cada mutante foi incubado em LB suplementado com o respectivo antibiótico a 37°C e com agitação constante. II.3. Ensaios com e sem contacto entre bactérias resistentes e bactérias ancestrais sensíveis
Foram realizados ensaios em que se permitiu o contacto entre a estirpe
resistente e a estirpe ancestral sensível e ensaios onde o contacto foi impedido. Nestes ensaios foram utilizadas culturas bacterianas que se encontravam em fase estacionária. Em cada ensaio o rácio inicial de bactérias resistentes e de bactérias sensíveis foi de 1:105 (aproximadamente
102 para 107 células). Estes ensaios foram realizados em meio líquido (LB) usando três métodos distintos e em meio sólido (LA) usando dois métodos diferentes (ver explicação em baixo).
II.3.1. Meio líquido Em meio líquido foram realizados ensaios em tubos de 50 ml, em seringas
e em placas com multipoços.
II.3.1.1. Tubos No caso dos ensaios em tubos, procedeu-se às diluições necessárias e
estes foram inoculados com uma mistura das duas estirpes (possibilitando o contacto e competição entre estirpes) e com as duas estirpes separadas (não existindo contacto nem competição).
Nestes ensaios as estirpes foram incubadas durante 3 horas, a 37ºC e com agitação de 170 rpm.
II.3.1.2. Seringas No caso de ensaios com seringas, utilizaram-se pares de seringas tendo
sido colocados filtros no meio, não permitindo o contacto entre bactérias sensíveis e resistentes, e pares de seringas ligadas directamente sem filtros, havendo contacto entre estirpes.
Este sistema permitiu testar o efeito do sobrenadante, já que o meio nos dois compartimentos das seringas foi misturado a cada 30 minutos durante o período de crescimento, permitindo a passagem do meio (LB) das estirpes resistentes para as estirpes ancestrais e vice-versa, existindo portanto, uma partilha do meio. Esta mistura foi realizada pressionando os êmbolos de cada lado várias vezes, sendo o êmbolo do lado da estirpe em alta densidade o último a ser pressionado.

9
Em cada seringa colocou-se 2 ml de ar estéril e 5 ml de LB, sendo depois cada seringa inoculada com a respectiva diluição de cada estirpe. Nestes ensaios as estirpes foram incubadas durante 3 horas, a 37ºC e com agitação de 170 rpm.
Este set up experimental está ilustrado nas figuras 1A e 1B.
Figura 1A e 1B – Seringas sem filtros e com filtros, respectivamente. Nas seringas sem filtros, as duas estirpes estão misturadas, existindo contacto. Quando as seringas estão separadas por dois filtros, a estirpe resistente é colocada numa das seringas, sendo a estirpe sensível colocada na seringa do lado oposto. Os filtros não permitem a passagem das bactérias de uma seringa para a outra, permitindo contudo a passagem do meio e, consequentemente, dos metabolitos produzidos pelas células.
II.3.1.3. Placas multipoços para cultura de células Este procedimento foi baseado no método descrito por Aoki et al. (2005). Estes ensaios foram realizados em placas com seis poços, tendo-se
realizado experiências com e sem contacto entre a estirpe resistente e a estirpe sensível.
Para inibir o contacto entre as estirpes mencionadas, utilizaram-se membranas de Polyetileno tereftalato com 23mm de tamanho e com um poro de 0,4 µm, de forma a serem colocadas as duas estirpes separadas no mesmo poço. Colocou-se 3,2 ml de LB inoculado com 102 células da estirpe resistente na parte inferior dos poços e colocou-se 2,5 ml do mesmo meio com 107 células da estirpe sensível na parte superior do poço.
As competições com contacto foram realizadas utilizando o mesmo rácio das estirpes resistente/sensível, tendo sido utilizado o mesmo volume de LB (5,7 ml) mas sem a presença da membrana.
Nos dois casos, houve crescimento durante 3 horas, a 37ºC e com agitação de 130 rpm.
Este set up experimental está ilustrado na figura 2.
Figura 2 – Set up experimental de placas multipoços para cultura de células. Como é possível observar através da figura, as placas contêm 6 poços nos quais são colocadas membranas (sem contacto) ou não (com contacto).
1A
1B

10
II.3.2. Meio sólido
II.3.2.1. Meio sólido directamente em LA
Inocularam-se placas de Petri contendo LA com uma mistura da estirpe sensível com a estirpe resistente (existindo contacto entre bactérias) e inocularam-se placas de Petri divididas ao meio, colocando-se de um lado a estirpe sensível e do outro a estirpe resistente (não existindo contacto). Foram colocadas 102 células da estirpe resistente e 107 células da estirpe sensível.
As estirpes foram incubadas durante 6 horas, sendo a cultura resultante raspada e ressuspendida em 1 ml de MgSO4 (10-2 M).
Nestes ensaios o período de incubação foi de 6 horas em vez das 3 horas dos ensaios anteriores uma vez que, em 3 horas apenas, o número de células não atingia valores contáveis. Assim, realizámos uma experiência semelhante, mas colocando as estirpes em membrana porosa, sendo essas membranas colocadas numa caixa de Petri sobre o LA.
II.3.2.2. Meio sólido em membrana porosa
Nestes ensaios colocaram-se duas membranas (0,45 µm) em cada placa de Petri contendo LA, sendo que uma das membranas foi inoculada com uma mistura da estirpe sensível com a estirpe resistente e a outra inoculada apenas com a estirpe resistente. Uma vez mais, foram colocadas 102 células da estirpe resistente e 107 células da estirpe sensível.
Ao fim de 3 horas de incubação a 37ºC, os filtros foram removidos das placas de Petri e foram colocados em tubos contendo 2 ml de MgSO4 10-2 M. Para ressuspender as células bacterianas, os tubos foram vigorosamente agitados durante 2 minutos.
Este set up experimental está ilustrado na figura 3.
Figura 3 – Set up experimental do meio sólido em membrana porosa. Como é possível observar através da figura, numa placa contendo LA colocam-se duas membranas em cada lado da placa. Estas são inoculadas com 102 células isoladas ou com 102 células resistentes a ácido nalidíxico misturadas com 107 células ancestrais sensíveis.

11
II.4. Curvas de crescimento em Meio Mínimo
Foram realizados ensaios com o objectivo de obter curvas de crescimento do mutante Nal48 (mutante derivado da estirpe E. coli K12 MG1655) quando este foi incubado com e sem 107 células da estirpe lacY- em meio mínimo (meio Mínimo M9 com agar 1,5% suplementado com MgSO4 (3 mM)) complementado com as seguintes fontes de carbono selectivas: glucose (4 g/l), substrato utilizado por ambas as estirpes e lactose (4 g/l), substrato utilizável apenas pelo mutante Nal48.
As estirpes foram inoculadas em tubos contendo 5 ml de meio, tendo sido incubadas durante 8 horas a 37ºC e com agitação de 170 rpm. A cada 30 minutos procedeu-se ao plaqueamento em LA com ácido nalidíxico. II.5. Ensaios em alta e baixa densidade
Estes ensaios foram realizados em meio líquido, inoculando-se 10 ml de LB
em tubos de 50 ml com 102 células (baixa densidade) e com 107 células (alta densidade).
As estirpes foram incubadas durante 3 horas, a 37ºC e com agitação de 170 rpm.
Foram também realizados ensaios em meio sólido em membrana porosa, sendo o protocolo semelhante ao descrito anteriormente. II.6. Evolução experimental
O mutante Nal48 foi sujeito a evolução experimental durante 960 gerações. Para tal, em cada ciclo de evolução, procedeu-se a dois dias de competição
entre o mutante e a estirpe ancestral sensível, com um rácio inicial de 1:105 (pressão selectiva) e dois dias de selecção de mutantes (em que se adicionou ácido nalidíxico ao meio). Os mutantes seleccionados em cada réplica foram sucessivamente sujeitos a este ciclo de evolução. Para além disto, efectuou-se um controlo da evolução do Nal48, no qual cada ciclo de evolução foi igual ao anterior, exceptuando a introdução da estirpe ancestral sensível. Cada condição foi replicada 5 vezes.
Ao fim de 16 ciclos de evolução (cerca de 960 gerações), foi isolado um clone de cada réplica da evolução assim como dos controlos e foram realizados testes experimentais. Para tal, procedeu-se à competição entre os mutantes evoluídos e o wt. As mesmas experiências foram realizadas com as réplicas controlo e com o ancestral. As réplicas que sugeriram acção da selecção foram sujeitas adicionalmente a ensaios com e sem contacto com 107 células do wt e a ensaios com o mutante em alta e em baixa densidade. O mesmo foi realizado para os mutantes controlo e ancestral. II.7. Estimativa do número de gerações e do fitness relativo
Sendo N1(0) e N2(0) o número inicial de unidades formadoras de colónias (ufc’s) das estirpes 1 e 2 respectivamente (quando t = 0) e sendo N1(T) e

12
N2(T) o número final de ufc’s das estirpes 1 e 2 (quando t = T), então o número de gerações decorridas desde t=0 até t=T pelas estirpes 1 e 2 serão dadas por:
O fitness relativo foi determinado de acordo com Lenski et al. (1991), sendo o fitness da estirpe 1 relativamente à estirpe 2 dado pelo rácio: II.8. Análise estatística
Todos os dados obtidos neste trabalho foram resultado de ensaios realizados, pelo menos em triplicado. Após a obtenção destes dados, procedeu-se às seguintes análises estatísticas:
• testes t de Student bilaterais para amostras independentes, para determinar: (1) se o número de gerações completado por uma estirpe resistente a um antibiótico é diferente quando esta estirpe cresce na presença ou na ausência de 107 células da estirpe ancestral; (2) se o número de gerações completado por uma estirpe que cresce em baixa densidade, é diferente do número de gerações completado quando esta cresce em alta densidade.
• ANOVA (Análise de Variância) de dois factores para determinar: (1) se o número de gerações completado por um mutante é significativamente dependente do contacto e da estirpe utilizada (lacY-, rpoS- e lrp-; mutante evoluído, ancestral e controlo); (2) se o número de gerações completado por um mutante é dependente do contacto e da adição de dióxido de carbono. Posteriormente, para as análises que revelaram diferenças significativas, foi realizada uma ANOVA de um factor e um teste de Tuckey post hoc para verificar onde se encontravam essas diferenças.
Antes da realização das ANOVAs acima mencionadas, foram testados os pressupostos desta análise (distribuição normal – testes de Shapiro-Wilk e Kolmogorov-Smirnov – e homogeneidade das variâncias – teste de Levene). No incumprimento de algum dos pressupostos, procedeu-se à transformação dos dados.
Foi utilizado o programa estatístico SPSS 20.0 (Chicago, IL) para a realização de todas as ANOVAs e o Excel mac 2011 para a realização de todos os testes t de Student.
Ao longo do capítulo referente aos resultados, é apresentado o valor médio do número de gerações completado pelas estirpes em 3 ou 6 horas de incubação ± 2xErro Padrão.
ng1 = Log2[N1(T)/N1(0)] ng2 = Log2[N2(T)/N2(0)]
ng1/ng2
13
III. RESULTADOS
III.1. Interacção com outras bactérias aumenta a taxa de crescimento
No que diz respeito às estirpes bacterianas E. coli K12, Salmonella enterica serovar Typhimurium LT2 e S. aureus ATCC 25923, utilizando a estirpe ancestral sensível e um mutante espontâneo resistente a ácido nalidíxico (designado Nal48, S57 e SA7, respectivamente), foi possível verificar que todos os mutantes completaram um maior número de gerações quando cresceram em competição com o seu ancestral, comparativamente ao seu crescimento isolado, sem competição. Este fenómeno verificou-se quando aproximadamente 102 células da estirpe resistente cresceram com aproximadamente 107 células da estirpe sensível.
Este fenómeno foi observado tanto em meio líquido, como em meio sólido.
III.1.1. Meio líquido – tubos
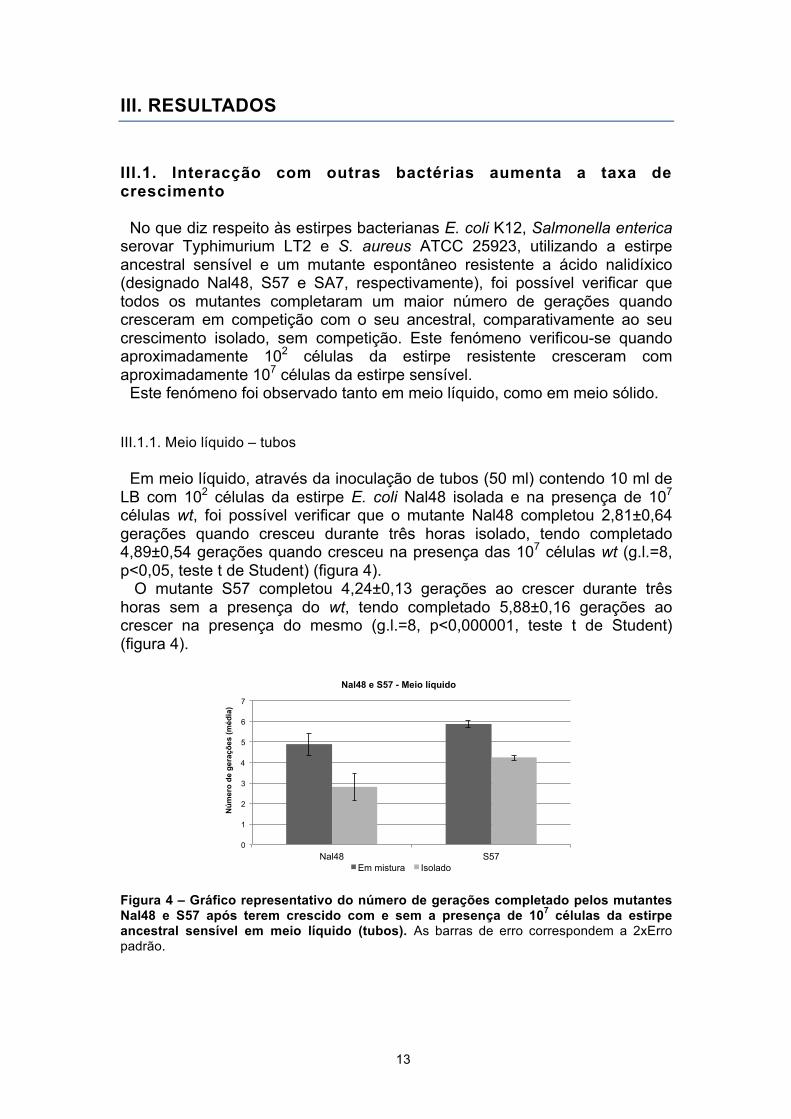
Em meio líquido, através da inoculação de tubos (50 ml) contendo 10 ml de LB com 102 células da estirpe E. coli Nal48 isolada e na presença de 107 células wt, foi possível verificar que o mutante Nal48 completou 2,81±0,64 gerações quando cresceu durante três horas isolado, tendo completado 4,89±0,54 gerações quando cresceu na presença das 107 células wt (g.l.=8, p<0,05, teste t de Student) (figura 4).
O mutante S57 completou 4,24±0,13 gerações ao crescer durante três horas sem a presença do wt, tendo completado 5,88±0,16 gerações ao crescer na presença do mesmo (g.l.=8, p<0,000001, teste t de Student) (figura 4).
Figura 4 – Gráfico representativo do número de gerações completado pelos mutantes Nal48 e S57 após terem crescido com e sem a presença de 107 células da estirpe ancestral sensível em meio líquido (tubos). As barras de erro correspondem a 2xErro padrão.
0
1
2
3
4
5
6
7
Nal48 S57
Núm
ero
de g
eraç
ões
(méd
ia)
Nal48 e S57 - Meio líquido
Em mistura Isolado
14
III.1.2. Meio sólido
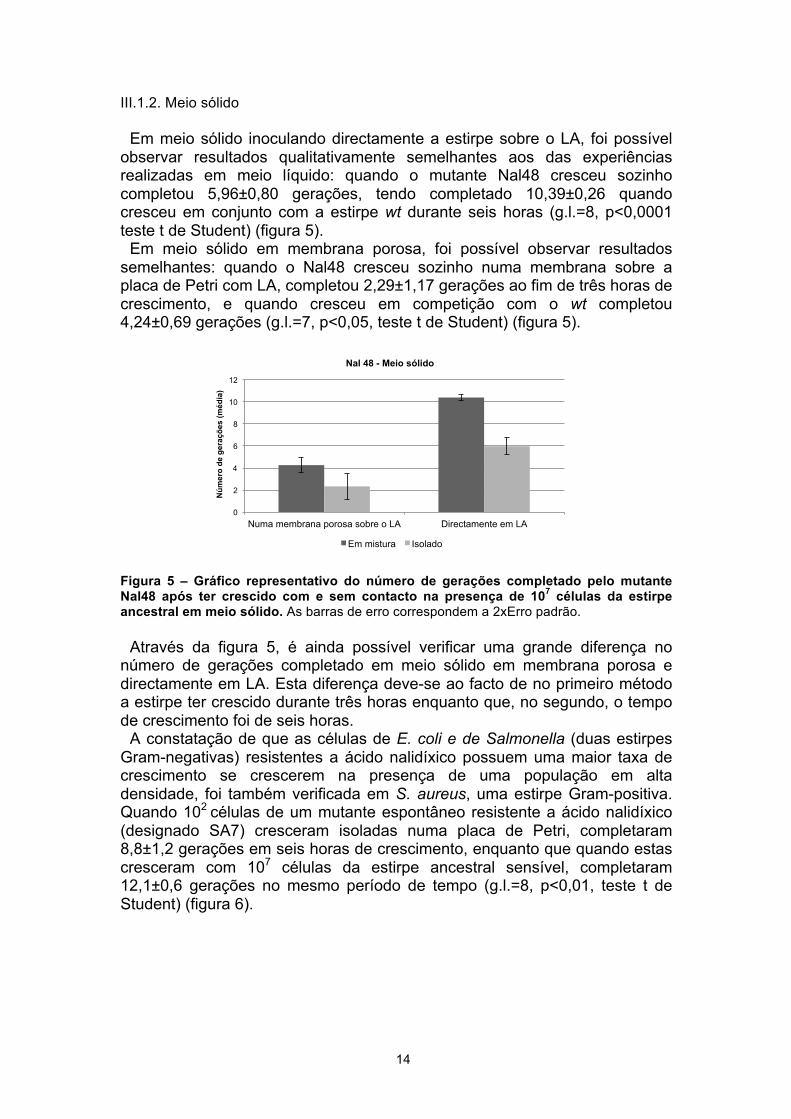
Em meio sólido inoculando directamente a estirpe sobre o LA, foi possível observar resultados qualitativamente semelhantes aos das experiências realizadas em meio líquido: quando o mutante Nal48 cresceu sozinho completou 5,96±0,80 gerações, tendo completado 10,39±0,26 quando cresceu em conjunto com a estirpe wt durante seis horas (g.l.=8, p<0,0001 teste t de Student) (figura 5).
Em meio sólido em membrana porosa, foi possível observar resultados semelhantes: quando o Nal48 cresceu sozinho numa membrana sobre a placa de Petri com LA, completou 2,29±1,17 gerações ao fim de três horas de crescimento, e quando cresceu em competição com o wt completou 4,24±0,69 gerações (g.l.=7, p<0,05, teste t de Student) (figura 5).
Figura 5 – Gráfico representativo do número de gerações completado pelo mutante Nal48 após ter crescido com e sem contacto na presença de 107 células da estirpe ancestral em meio sólido. As barras de erro correspondem a 2xErro padrão.
Através da figura 5, é ainda possível verificar uma grande diferença no número de gerações completado em meio sólido em membrana porosa e directamente em LA. Esta diferença deve-se ao facto de no primeiro método a estirpe ter crescido durante três horas enquanto que, no segundo, o tempo de crescimento foi de seis horas.
A constatação de que as células de E. coli e de Salmonella (duas estirpes Gram-negativas) resistentes a ácido nalidíxico possuem uma maior taxa de crescimento se crescerem na presença de uma população em alta densidade, foi também verificada em S. aureus, uma estirpe Gram-positiva. Quando 102 células de um mutante espontâneo resistente a ácido nalidíxico (designado SA7) cresceram isoladas numa placa de Petri, completaram 8,8±1,2 gerações em seis horas de crescimento, enquanto que quando estas cresceram com 107 células da estirpe ancestral sensível, completaram 12,1±0,6 gerações no mesmo período de tempo (g.l.=8, p<0,01, teste t de Student) (figura 6).
0
2
4
6
8
10
12
Numa membrana porosa sobre o LA Directamente em LA
Núm
ero
de g
eraç
ões
(méd
ia)
Nal 48 - Meio sólido
Em mistura Isolado

15
Figura 6 – Gráfico representativo do número de gerações completado pelo mutante SA7 após ter crescido com e sem contacto com 107 células da estirpe ancestral em meio sólido. As barras de erro correspondem a 2xErro padrão.
Assim, foi possível verificar que três estirpes distintas apresentam um maior número de gerações ao crescer em contacto com uma população constituída por uma alta densidade de bactérias. Surge então uma nova questão: esta vantagem deve-se à secreção de algum metabolito pela estirpe que se encontra em alta densidade? III.2. O contacto célula a célula é necessário para o mecanismo VDC
Para testar se a vantagem descrita anteriormente é resultado da secreção de um metabolito pela estirpe em alta densidade (estirpe ancestral sensível), foi testado o efeito do sobrenadante destas células. Para tal, realizaram-se ensaios nos quais foi utilizado o sobrenadante da estirpe wt. Este sobrenadante resultou do crescimento de 107 células em LB durante três horas, tendo o meio sido filtrado de forma a assegurar que todas as bactérias tinham sido eliminadas.
Foi possível verificar que a estirpe de E. coli resistente a ácido nalidíxico completou um número de gerações semelhante em sobrenadante (2,58±0,69) e em LB (2,81±0,64) (g.l.=8, p=0,813, teste t de Student). Estes resultados sugerem que a vantagem da estirpe que cresce com o wt não resulta da secreção de metabolitos por parte deste último.
Assim, colocou-se a hipótese de que o contacto célula a célula será necessário para que exista a vantagem atrás mencionada.
Para testar esta hipótese, foram realizados ensaios utilizando pares de seringas, permitindo e inibindo o contacto entre a estirpe resistente e a estirpe susceptível. Quando não existe contacto entre as estirpes, o meio é partilhado por elas, testando-se também o efeito do sobrenadante.
Foi possível verificar que em E. coli, quando o mutante Nal48 cresceu sem contacto com o wt, completou 3,25±0,68 gerações em três horas de
0
2
4
6
8
10
12
14
Núm
ero
de g
eraç
ões
(méd
ia)
SA7 - meio sólido directamente em LA
Em mistura Isolado

16
crescimento, enquanto que com contacto, o mutante completou 6,78±0,34 gerações (g.l.=8, p<0,001, teste t de Student). De notar que os filtros não mostraram ter qualquer efeito sobre o número de gerações do wt. Assim, o wt completou 4,21±0,62 gerações com filtros (ou seja, sem contacto com o Nal48) e 5,08±0,46 gerações sem filtros (com contacto com o Nal48) (g.l.=8, p>0,05, teste t de Student).
A mesma tendência foi observada em Salmonella: quando o mutante S57 cresceu sozinho completou 3,94±0,21 gerações, enquanto que, quando cresceu com 107 células da estirpe ancestral sensível completou 5,32±0,36 gerações (g.l.=8, p<0,001, teste t de Student).
Resultados semelhantes foram observados em S. aureus quando se procedeu a esta metodologia. Desta forma, quando o mutante SA7 cresceu sozinho completou 2,57±0,73 gerações, enquanto que, quando cresceu com contacto com o wt completou 5,52±0,78 gerações (g.l.=8, p<0,001, teste t de Student).
Adicionalmente, para confirmar a necessidade do contacto célula a célula, foram realizadas experiências utilizando placas com 6 poços em que, novamente, se permitiu o contacto entre a estirpe resistente e a estirpe sensível e em que não se permitiu o contacto entre elas através da utilização de membranas (com poros de 0,4 µm). Com estas membranas, e tal como sucede no caso das seringas, o meio é partilhado pelas duas estirpes sem que haja contacto entre estas.
Observámos que o número de gerações completado pelo mutante Nal48 foi 2,17±0,97 quando este cresceu sozinho. No entanto, quando cresceu com 107 células da estirpe sensível completou 5,60±0,39 gerações (g.l.=8, p<0,003, teste t de Student). O número de gerações completado pela estirpe sensível não foi significativamente diferente nas duas situações (g.l.=8, p=0,78, teste t de Student).
As mesmas experiências foram realizadas com S. aureus, sendo que, sem contacto, a estirpe resistente completou 2,63±0,99 gerações, enquanto que, com contacto, completou 5,07±0,58 gerações (g.l.=8, p<0,01, teste t de Student). Uma vez mais, a presença da membrana não afectou o número de gerações alcançado pela estirpe ancestral sensível (g.l.=8, p=0,28, teste t de Student).
Novamente, estes resultados sugerem que esta vantagem é mediada pelo contacto célula a célula, não sendo consequência da produção de um metabolito secretado pela estirpe ancestral susceptível em alta densidade.
III.3. VDC actua quando o contacto se estabelece com células em estado de latência
De forma a averiguar se o fenómeno VDC necessita que as células que se apresentam em alta densidade estejam reprodutivamente activas, realizaram-se curvas de crescimento do mutante Nal48 em que 102 células desta estirpe cresceram isoladas e na presença de 107 células da estirpe lacY- da Keio Collection (Baba et al., 2006). Esta estirpe, para além de lacY- é também lacZ-, o que não lhe permite utilizar lactose como substracto. Em E. coli, o transporte e metabolismo da lactose envolve o operão lac. Este operão
17
compreende três genes necessários para o uptake e metabolismo da lactose e açúcares relacionados: lacZ, lacY e lacA. Assim, o gene lacZ codifica para uma β-galactosidase, uma enzima responsável pela conversão da lactose em alolactose e subsequente intermediários metabólicos; o gene lacY codifica para a lactose permease, que facilita o uptake de lactose e moléculas similares e o gene lacA codifica para uma acetiltransferase que está envolvida no metabolismo do açúcar (Ozbudak et al., 2004).
Assim, o mutante Nal48 foi inoculado na presença e na ausência de 107 células da estirpe lacY- (também lacZ-) em meio mínimo com glucose (em que há crescimento das duas estirpes) e em meio mínimo com lactose (em que apenas cresce o mutante Nal48).
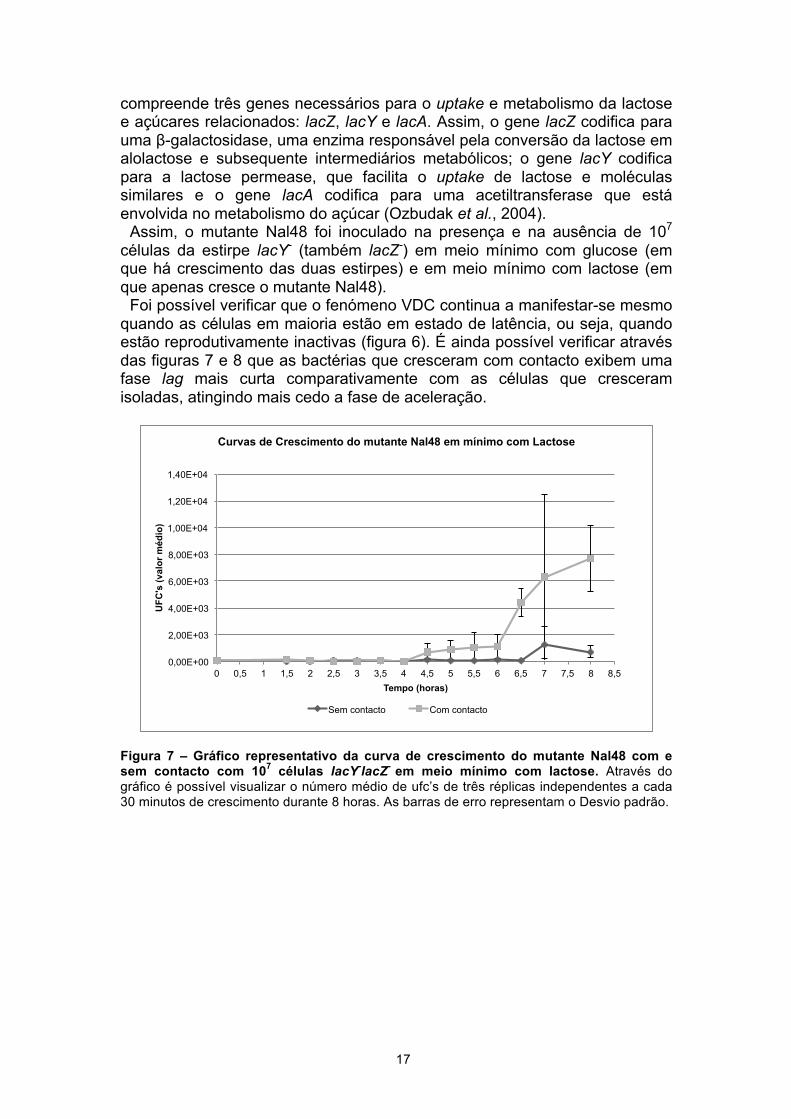
Foi possível verificar que o fenómeno VDC continua a manifestar-se mesmo quando as células em maioria estão em estado de latência, ou seja, quando estão reprodutivamente inactivas (figura 6). É ainda possível verificar através das figuras 7 e 8 que as bactérias que cresceram com contacto exibem uma fase lag mais curta comparativamente com as células que cresceram isoladas, atingindo mais cedo a fase de aceleração.
Figura 7 – Gráfico representativo da curva de crescimento do mutante Nal48 com e sem contacto com 107 células lacY-lacZ- em meio mínimo com lactose. Através do gráfico é possível visualizar o número médio de ufc’s de três réplicas independentes a cada 30 minutos de crescimento durante 8 horas. As barras de erro representam o Desvio padrão.
0,00E+00
2,00E+03
4,00E+03
6,00E+03
8,00E+03
1,00E+04
1,20E+04
1,40E+04
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5 5,5 6 6,5 7 7,5 8 8,5
UFC
's (v
alor
méd
io)
Tempo (horas)
Curvas de Crescimento do mutante Nal48 em mínimo com Lactose
Sem contacto Com contacto
18
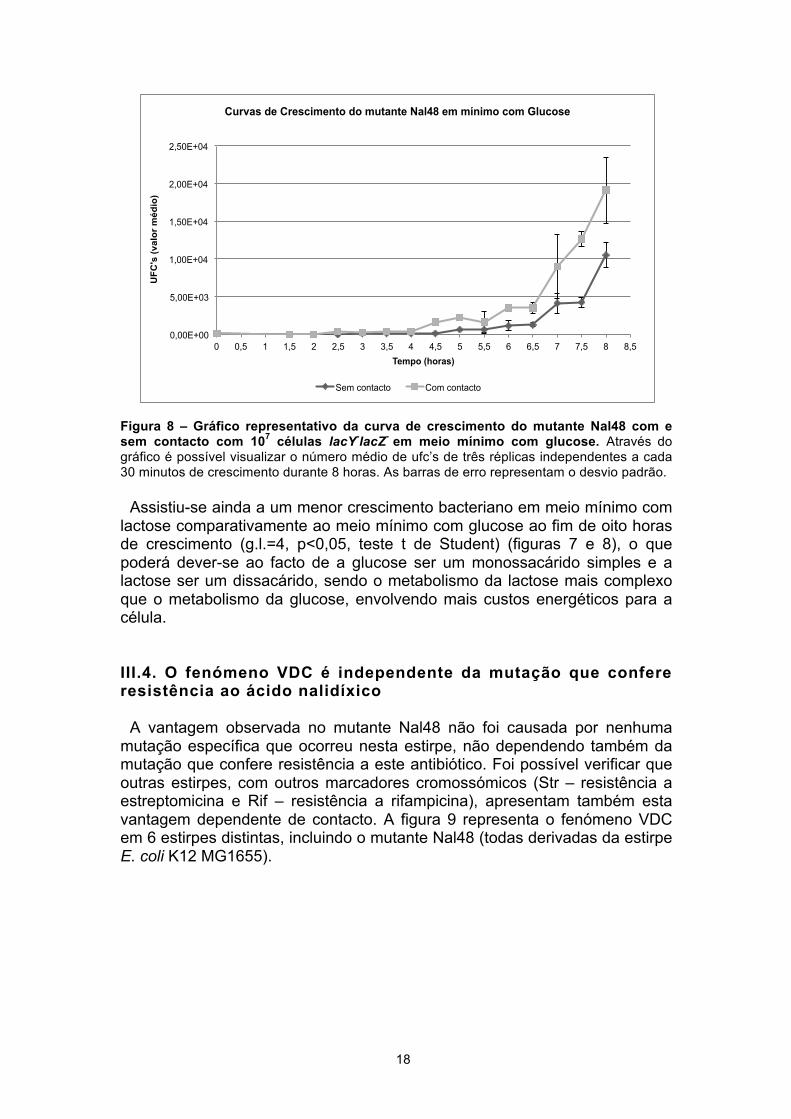
Figura 8 – Gráfico representativo da curva de crescimento do mutante Nal48 com e sem contacto com 107 células lacY-lacZ- em meio mínimo com glucose. Através do gráfico é possível visualizar o número médio de ufc’s de três réplicas independentes a cada 30 minutos de crescimento durante 8 horas. As barras de erro representam o desvio padrão.
Assistiu-se ainda a um menor crescimento bacteriano em meio mínimo com
lactose comparativamente ao meio mínimo com glucose ao fim de oito horas de crescimento (g.l.=4, p<0,05, teste t de Student) (figuras 7 e 8), o que poderá dever-se ao facto de a glucose ser um monossacárido simples e a lactose ser um dissacárido, sendo o metabolismo da lactose mais complexo que o metabolismo da glucose, envolvendo mais custos energéticos para a célula. III.4. O fenómeno VDC é independente da mutação que confere resistência ao ácido nalidíxico
A vantagem observada no mutante Nal48 não foi causada por nenhuma mutação específica que ocorreu nesta estirpe, não dependendo também da mutação que confere resistência a este antibiótico. Foi possível verificar que outras estirpes, com outros marcadores cromossómicos (Str – resistência a estreptomicina e Rif – resistência a rifampicina), apresentam também esta vantagem dependente de contacto. A figura 9 representa o fenómeno VDC em 6 estirpes distintas, incluindo o mutante Nal48 (todas derivadas da estirpe E. coli K12 MG1655).
0,00E+00
5,00E+03
1,00E+04
1,50E+04
2,00E+04
2,50E+04
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5 5,5 6 6,5 7 7,5 8 8,5
UF
C's
(va
lor
méd
io)
Tempo (horas)
Curvas de Crescimento do mutante Nal48 em mínimo com Glucose
Sem contacto Com contacto

19
Figura 9 – Gráfico representativo do fenómeno VDC em 6 mutantes distintos derivados de E. coli K12 MG1655. Através da figura é possível verificar que, para além do mutante Nal48, o fenómeno VDC está presente nos mutantes Nal45, Nal59, Str36, Rif3 e Rif4. As barras de erro representam 2*Erro Padrão.
Verifica-se então que, para além do mutante Nal48, o fenómeno VDC está também presente nas estirpes Nal45, Nal59, Str36, Rif3 e Rif4. Conclui-se assim que o mecanismo não advém de nenhuma mutação específica que ocorreu no mutante Nal48.
III.5. Mecanismo VDC como consequência do crescimento em populações bacterianas com uma alta densidade populacional
A constatação de que os mutantes completam um maior número de gerações ao crescer em contacto com o wt colocou-nos uma nova questão: será que esta vantagem depende necessariamente da presença do wt, ou depende apenas da densidade celular?
Para responder a esta questão foram realizados ensaios no sentido de confirmar se o número de gerações completado por um mutante diferia quando este crescia em baixa (102) e em alta densidade (107), não se introduzindo assim o seu ancestral. Estes ensaios foram realizados em meio líquido, em tubos de 50 ml.
Foi possível verificar que o mutante Nal48 completou 4,75±0,24 gerações quando cresceu em baixa densidade, tendo completado 6,02±0,18 gerações quando cresceu em alta densidade (g.l.=7, p<0,01, teste t de Student).
O facto deste mecanismo estar presente nos mutantes de E. coli e S. aureus – espécies evolutivamente distintas –, sugere que as estirpes ancestrais (wt) já o possuem. Para confirmar esta questão, realizaram-se ensaios semelhantes ao anterior, mas com o wt de cada estirpe.
Foi possível verificar que, ao fim de três horas de crescimento em meio líquido (tubos de 50 ml), 102 células wt de E. coli completaram 3,76±1,08 gerações, enquanto que as 107 células completaram 5,32±0,64 gerações
Nal45
Nal48
Nal59
Str36 Rif3
Rif4
Nal45
Nal48
Nal59
Str36
Rif3
Rif4
0
1
2
3
4
5
6
7
8
9
10
Núm
ero
gera
ções
(méd
ia)
Estirpes
Com contacto Sem contacto

20
(g.l.=8, p<0,05, teste t de Student). Foi verificada a mesma tendência em meio sólido em membrana porosa: em três horas, 102 células completaram 2,85±0,23 gerações, enquanto que 107 células completaram 4,54±0,34 gerações (g.l.=8, p<0,0001, teste t de Student).
Verificaram-se resultados semelhantes em Salmonella: após três horas de crescimento em meio líquido (tubos de 50 ml), 102 células wt completaram 4,67±0,42 gerações, enquanto que as 107 células completaram 6,21±0,39 gerações (g.l.=8, p<0,001, teste t de Student). Em meio sólido em membrana porosa, nas células que cresceram em baixa densidade (102 células), não obtivemos qualquer colónia nas cinco réplicas após as três horas de crescimento e plaqueamento, o que sugere a ocorrência de morte celular.
Realizando os mesmos ensaios para S. aureus, verificaram-se resultados semelhantes: em tubos, 102 células do wt desta estirpe completou 0,13±0,75 gerações, enquanto que 107 células completaram 5,36±0,69 gerações (g.l.=8, p<0,0001, teste t de Student). Em meio sólido em membrana porosa, 102 células completaram 2,10±0,88 gerações, enquanto que 107 células completaram 3,41±0,65 gerações (g.l.=8, p<0,05, teste t de Student).
Em conjunto com os resultados anteriores, estes resultados mostram-nos que este mecanismo depende apenas do contacto com uma população constituída por uma alta densidade de células. O facto de este mecanismo se encontrar nas estirpes ancestrais de E. coli K12, Salmonella enterica serovar Typhimurium LT2 e S. aureus, sugere que o(s) gene(s) envolvido(s) neste mecanismo estão amplamente presente no mundo bacteriano.
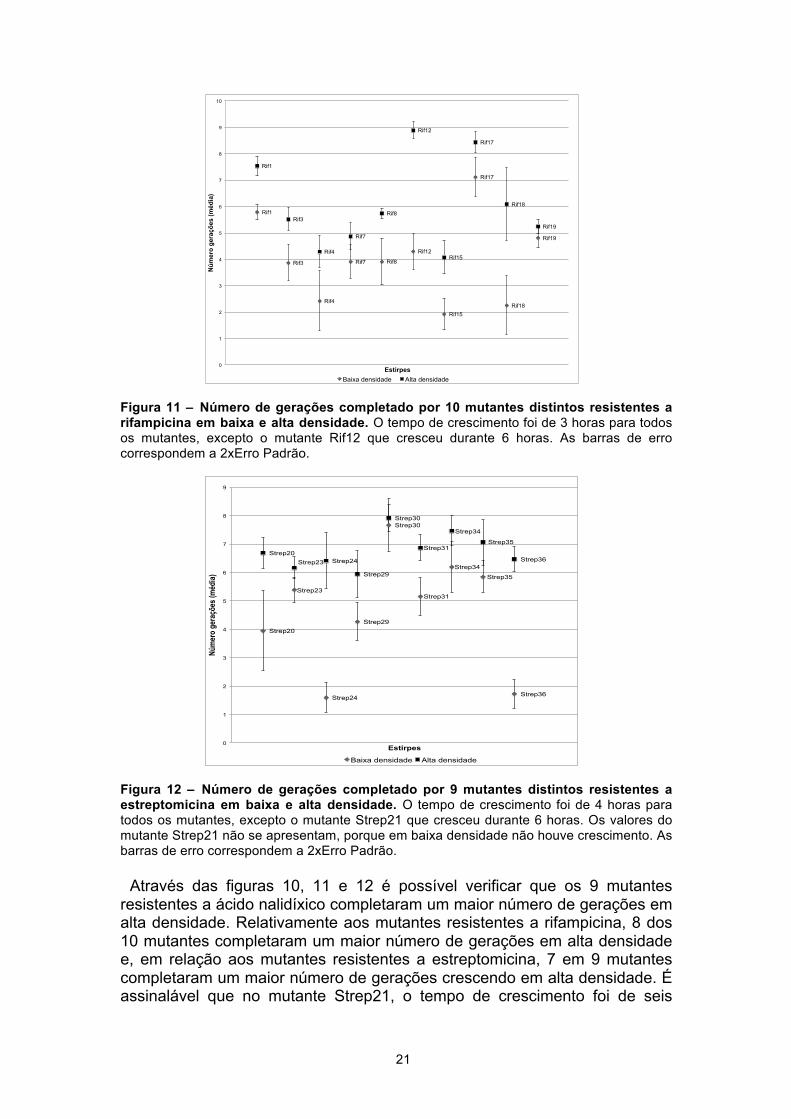
Foram ainda realizados ensaios em meio líquido (tubos) com 29 mutantes adicionais espontâneos da estirpe E. coli K12, sendo 9 resistentes a ácido nalidíxico, 10 resistentes a rifampicina e 10 resistentes a estreptomicina. Os gráficos seguintes (figuras 10, 11 e 12) representam o número de gerações completado pelos 29 mutantes ao crescer em baixa e alta densidade.
Figura 10 – Número de gerações completado por 9 mutantes distintos resistentes a ácido nalidíxico em baixa e alta densidade. O tempo de crescimento foi de 3 horas para todos os mutantes. As barras de erro correspondem a 2xErro Padrão.
Nal41 Nal43
Nal45
Nal50
Nal51
Nal54 Nal56
Nal58
Nal59
Nal41
Nal43
Nal45
Nal50 Nal51
Nal54
Nal56
Nal58
Nal59
0
1
2
3
4
5
6
7
8
Núm
ero
gera
ções
(méd
ia)
Estirpes Baixa Densidade Alta Densidade
21
Figura 11 – Número de gerações completado por 10 mutantes distintos resistentes a rifampicina em baixa e alta densidade. O tempo de crescimento foi de 3 horas para todos os mutantes, excepto o mutante Rif12 que cresceu durante 6 horas. As barras de erro correspondem a 2xErro Padrão.
Figura 12 – Número de gerações completado por 9 mutantes distintos resistentes a estreptomicina em baixa e alta densidade. O tempo de crescimento foi de 4 horas para todos os mutantes, excepto o mutante Strep21 que cresceu durante 6 horas. Os valores do mutante Strep21 não se apresentam, porque em baixa densidade não houve crescimento. As barras de erro correspondem a 2xErro Padrão.
Através das figuras 10, 11 e 12 é possível verificar que os 9 mutantes resistentes a ácido nalidíxico completaram um maior número de gerações em alta densidade. Relativamente aos mutantes resistentes a rifampicina, 8 dos 10 mutantes completaram um maior número de gerações em alta densidade e, em relação aos mutantes resistentes a estreptomicina, 7 em 9 mutantes completaram um maior número de gerações crescendo em alta densidade. É assinalável que no mutante Strep21, o tempo de crescimento foi de seis
Rif1
Rif3
Rif4
Rif7 Rif8
Rif12
Rif15
Rif17
Rif18
Rif19
Rif1
Rif3
Rif4
Rif7
Rif8
Rif12
Rif15
Rif17
Rif18
Rif19
0
1
2
3
4
5
6
7
8
9
10
Núm
ero
gera
ções
(méd
ia)
Estirpes Baixa densidade Alta densidade
Strep20
Strep23
Strep24
Strep29
Strep30
Strep31
Strep34
Strep35
Strep36
Strep20
Strep23 Strep24
Strep29
Strep30
Strep31
Strep34
Strep35
Strep36
0
1
2
3
4
5
6
7
8
9
Núm
ero
gera
ções
(méd
ia)
Estirpes
Baixa densidade Alta densidade

22
horas, apresentando contudo crescimento nulo em baixa densidade (em quatro das cinco réplicas do ensaio), possivelmente devido a custos de resistência muito elevados ou devido a morte celular. III.6. VDC entre espécies diferentes
O fenómeno VDC foi também observado entre as espécies E. coli K12 e S.aureus.
Em meio sólido, 102 células do mutante Nal48 de E. coli completaram 9,94±1,06 gerações quando cresceram durante seis horas com 107 células de S. aureus wt, completando apenas 3,94±0,71 gerações quando cresceram sem a presença de S. aureus durante o mesmo período de tempo (g.l.=7, p<0,0001, teste t de Student). Em meio líquido, em seringas, verificaram-se resultados semelhantes: 102 células do mutante Nal48 completaram 5,20±1,02 gerações quando cresceram sozinhas, tendo completado 7,47±1,26 gerações crescendo em contacto com 107 células da estirpe wt de S. aureus (g.l.=7, p<0,05, teste t de Student).
De igual modo, em meio sólido, 102 células do mutante SA7 de S. aureus completaram 12,93±0,17 gerações quando cresceram com 107 células de E. coli wt, tendo completado apenas 2,78±1,1 gerações quando cresceram isoladas (g.l.=7, p<0,01, teste t de Student). Em meio líquido, em seringas, 102 células da estirpe SA7 completaram 3,27±0,80 gerações quando cresceram sozinhas e completaram 4,89±1,01 gerações quando cresceram com 107 células wt de E. coli (g.l.=8, p<0,05, teste t de Student). III.7. O caso de Enterococcus faecalis: inexistência do fenómeno VDC
Foi também testada a presença do fenómeno VDC em E. faecalis LN68. Foram testados três mutantes espontâneos distintos derivados desta estirpe
(designados LN7, LN10 e LN47), resistentes a rifampicina. Estes ensaios foram realizados em meio líquido, em seringas, verificando-se que o número de gerações completado por estes mutantes não é diferente quando o mutante cresceu com ou sem contacto com 107 células da estirpe ancestral sensível. Assim, 102 células do mutante LN7 completaram 7,70±0,37 gerações crescendo isoladas e completaram 8,04±0,47 gerações quando cresceram na presença de 107 células do wt (g.l.=8, p=0,29, teste t de Student). O mutante LN10 completou 6,89±0,68 gerações crescendo sozinho, completando 6,71±0,65 gerações crescendo com o wt (g.l.=8, p=0,71, teste t de Student). Por fim, o mutante LN47 completou 7,98±0,63 gerações quando cresceu sozinho, completando 7,65±0,34 gerações quando cresceu com o wt (g.l.=8, p=0,40, teste t de Student).
Para confirmar estes resultados, foi utilizado o método de placas com seis poços para o mutante LN7. O número de gerações completado ao fim de três horas após o crescimento isolado, foi de 4,69±0,15, tendo completado 5,26±0,51 gerações na presença de 107 células do wt (g.l.=4, p=0,15, teste t

23
de Student). Confirmando-se os resultados com este mutante, esta metodologia não foi repetida para os restantes.
Foram também realizados ensaios com células wt desta estirpe, em que estas cresceram em baixa e em alta densidade durante três horas. Em meio líquido (tubos de 50 ml) 102 células completaram 6,55±0,36 gerações, enquanto que 107 células completaram 6,04±0,26 gerações (g.l.=8, p=0,05, teste t de Student). Em meio sólido em membrana porosa, 102 células completaram 3,67±0,49 gerações, enquanto que 107 células completaram apenas 1,23±0,10 gerações (g.l.=8, p<0,001, teste t de Student). Assim, verificou-se que em meio sólido em membrana porosa, as células apresentam uma maior taxa de crescimento em baixa densidade, fenómeno contraditório relativamente a E. coli, Salmonella e S. aureus.
Posteriormente, pretendeu-se verificar se células de E. coli apresentavam vantagem ao crescer na presença de E. faecalis LN68 em alta densidade, pretendendo-se verificar se o mecanismo VDC actuaria entre E. faecalis e uma espécie distinta. Foi possível observar que o número de gerações completadas por 102 células de um mutante espontâneo de E. coli resistente a rifampicina (Rif3) não aumentou com a presença de 107 células de E. faecalis, nem em meio líquido (tubos) (g.l.=8, p=1,0, teste t de Student) nem em meio sólido em membrana porosa (g.l.=7, p=0,67, teste t de Student). De igual forma, foi possível observar que o número de gerações completadas por 102 células do mutante Nal48 não aumentou com a presença desta estirpe em meio líquido (tubos) (g.l.=8, p=0,43, teste t de Student).
Conclui-se que o fenómeno de VDC não está presente em E. faecalis LN68. Para além disto, estes resultados sugerem que, para este fenómeno actuar entre espécies distintas é necessário que as duas espécies sejam portadoras do(s) gene(s) envolvido(s) no fenómeno VDC. III.8. O papel dos genes rpoS e lrp no fenómeno VDC
Para compreender que possíveis mecanismos moleculares estão envolvidos neste mecanismo, testou-se o efeito do contacto em três estirpes de E. coli: lacY-, rpoS- e lrp-. Estas estirpes têm perda de função dos genes lacY, rpoS e lrp, respectivamente. Foram gerados mutantes espontâneos resistentes a ácido nalidíxico para cada uma das estirpes (y3 derivado de lacY-, r19 derivado de rpoS- e l15 derivado de lrp-). Posteriormente realizaram-se experiências com e sem contacto em meio sólido, utilizando as três estirpes resistentes e o mutante ancestral sensível correspondente.
Verificou-se que 102 células do mutante y3 completaram 3,77±0,36 gerações crescendo durante seis horas isoladas, tendo completado 10,81±0,31 gerações na presença de 107 células da estirpe lacY- ancestral no mesmo período de tempo (g.l.=8, p<0,001, teste t de Student). O mutante r19 completou 1,78±0,91 gerações ao crescer isolado, completando 10,87±0,55 gerações na presença de 107 células da estirpe rpoS- ancestral (g.l.=8, p<0,00001, teste t de Student). Por último, o mutante l15 completou 2,11±0,83 gerações quando cresceu sozinho, tendo completado 9,76±0,66 gerações quando cresceu em contacto com 107 células da estirpe lrp-
ancestral (g.l.=8, p<0,00001, teste t de Student).

24
Conclui-se que o fenómeno VDC continua a manifestar-se nestas estirpes, pelo que os genes rpoS e lrp não parecem ser essenciais para este mecanismo. No entanto, após a realização de uma ANOVA de dois factores, verificou-se que as variáveis contacto (p<0,0001) e estirpe (p<0,01) têm efeitos significativos, assim como a interacção das duas (p<0,05) (tabela 1). Realizaram-se posteriores ANOVAS com um factor de modo a averiguar qual a estirpe responsável por estas diferenças. Verificou-se que o número de gerações sem contacto com 107 células varia com a estirpe (p<0,05, ANOVA de um factor), sendo este valor inferior nas estirpes lrp- e rpoS- comparativamente com a estirpe lacY- (p<0,05, teste de Tukey). Entre as estirpes lrp- e rpoS- não há diferenças significativas (p>0,05, teste de Tukey) (figura 13).
Verificou-se ainda que o número de gerações com contacto com 107 células varia também com a estirpe (p<0,05, ANOVA de um factor), sendo este valor superior nas estirpes rpoS- e lacY- comparativamente com a estirpe lrp- (p<0,05, teste de Tukey). É de salientar que, com contacto com 107 células, não há diferenças significativas entre as estirpes lacY- e rpoS- (p>0,05, teste de Tukey) (figura 13).
TABELA 1 – ANOVA de dois factores para o número de gerações com e sem contacto nas estirpes lacY-, lrp- e rpoS-.
Figura 13 – Número de gerações completado pelos mutantes lacY-, lrp- e rpoS- com e sem contacto com 107 células wt. Como é possível observar, sem contacto, os mutantes lrp- e rpoS- completaram um menor número de gerações que a estirpe lacY- e, com contacto, o mutante lrp- completou um menor número de gerações que os mutantes lacY- e rpoS-.
Fonte de Variação g.l. F pEstirpe 2 8,491 0,002
Contacto 1 851,137 0,000Estirpe*Contacto 2 5,021 0.016
Erro 23
lacY-
lrp-
rpoS-
lacY-
lrp- rpoS-
0
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
Núm
ero
gera
ções
(méd
ia)
Com contacto Sem contacto

25
III.9. Testando o mecanismo fisiológico por detrás do fenómeno VDC: o efeito do dióxido de carbono
Repaske e Clayton (1978) demonstraram que E. coli necessita de CO2 para o crescimento. Verificaram assim que quando uma concentração saturada estava presente no momento da inoculação, o crescimento ocorria à taxa máxima, tendo ainda verificado que concentrações limitantes de CO2 restringiam a taxa de crescimento proporcionalmente à concentração de CO2 presente. Walker (citado por Repaske e Clayton, 1978) tinha já demonstrado que a adição de CO2 encurtava a fase lag em E. coli. Colocámos assim a hipótese de o fenómeno VDC dever-se a uma maior concentração de dióxido de carbono na situação em que 102 células crescem na presença de 107 células bacterianas.
Para responder a esta questão, inocularam-se tubos (50 ml) com uma mistura de 102 células do mutante y3 com 107 células da estirpe ancestral (lacY-) e tubos apenas com 102 células do mutante y3, com e sem adição de 10mM de Na2CO3, para averiguar os efeitos do carbono no crescimento bacteriano nas duas situações.
Através da tabela 2 é possível verificar que há efeito do contacto com 107 células, independentemente da adição de Na2CO3 (g.l.=1, p<0,01, ANOVA de dois factores), sendo que o Na2CO3 não tem efeito (g.l.=1, p>0,05, ANOVA de dois factores). A interacção contacto*carbono também não foi significativa (g.l.=1, p=0,702, ANOVA de dois factores).
Dado que a variável Na2CO3 se aproximou de um valor significativo, realizaram-se posteriores ANOVAs com um factor, tendo-se verificado que, com contacto com 107 células não houve diferenças no número de gerações quando se adicionou Na2CO3 ao meio (p=0,246, ANOVA de um factor). O mesmo se verificou sem contacto com 107 células (p=0,120, ANOVA de um factor). TABELA 2 – ANOVA de dois factores para o efeito das variáveis contacto e Na2CO3 no número de gerações completado pela estirpe y3.
De acordo com estes resultados, verifica-se que a adição de Na2CO3, não tem efeito no crescimento bacteriano quando as células crescem com ou sem contacto com 107 células.
Aparentemente o efeito do dióxido de carbono reflecte-se num ligeiro aumento do número de gerações nas duas condições (com e sem contacto com 107 células), sendo que este aumento não é suficiente para explicar o mecanismo VDC.
Através da figura 14 verifica-se o efeito do dióxido de carbono no aumento do número de gerações médio do mutante y3 com e sem contacto com 107 células ancestrais.
Fonte de Variação g.l. F pContacto 1 16,476 0,001Na2CO3 1 4,506 0,050
Contacto*Na2CO3 1 0,152 0,702Erro 16

26
Figura 14 – Representação do número médio de gerações com e sem contacto com
107 células ancestrais e com e sem a adição de 10mM de Na2CO3. Verifica-se que a adição de Na2CO3 provocou um aumento do número médio de gerações quando o mutante cresceu com ou sem contacto com 107 células da estirpe ancestral, pelo que o dióxido de carbono parece favorecer o crescimento das bactérias, não estando ligado ao fenómeno VDC em si. A interacção dos factores contacto e CO2 não é significativa (p=0,702). III.10. Evolução experimental
III.10.1. Controlo da evolução experimental e mutante ancestral
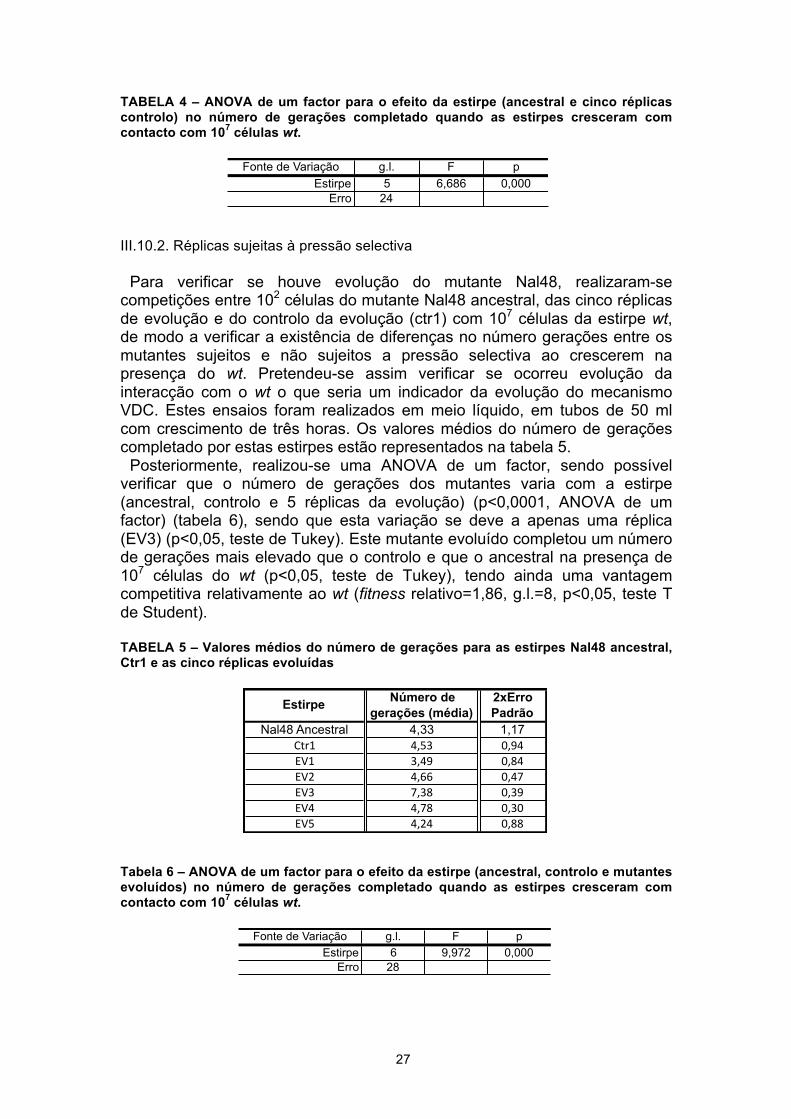
Começámos por realizar ensaios de competição em meio líquido (10 ml de LB em tubos de 50 ml), em que, tanto as cinco réplicas de controlo da evolução como o mutante ancestral (Nal48 não evoluído) cresceram durante três horas com contacto com 107 células da estirpe sensível. Os valores médios do número de gerações completado por estas estirpes estão representados na tabela 3.
Através da realização de uma ANOVA de um factor, foi possível verificar que o número de gerações completado depende da estirpe (mutante ancestral e as cinco réplicas controlo) (p<0,0001, ANOVA de um factor) (tabela 4). Estas diferenças devem-se apenas à réplica Ctr4 que completou um número de gerações maior que as restantes réplicas (p<0,05, teste de Tukey). Assim, seleccionámos apenas uma das réplicas de controlo em que não se observaram diferenças no número de gerações completado após o crescimento com 107 células wt relativamente ao mutante ancestral, tendo sido selecionado o Controlo 1 (ctr1).
TABELA 3 – Valores médios do número de gerações para as estirpes Nal48 ancestral e as cinco réplicas controlo da evolução
Sem adição de Na2CO3
Com adição de Na2CO3
Sem adição de Na2CO3
Com adição de Na2CO3
2
3
4
5
6
7
8
9
10
Núm
ero
gera
ções
(méd
ia)
Com contacto Sem contacto
Estirpe Número de gerações (média)
2xErro Padrão
Nal48 Ancestral 4,33 1,17!"#$ %&'( )&*%!"#+ %&+, )&*+!"#( '&+* )&-,!"#% -&.- )&%.!"#' (&,* )&('
27
TABELA 4 – ANOVA de um factor para o efeito da estirpe (ancestral e cinco réplicas controlo) no número de gerações completado quando as estirpes cresceram com contacto com 107 células wt.
III.10.2. Réplicas sujeitas à pressão selectiva
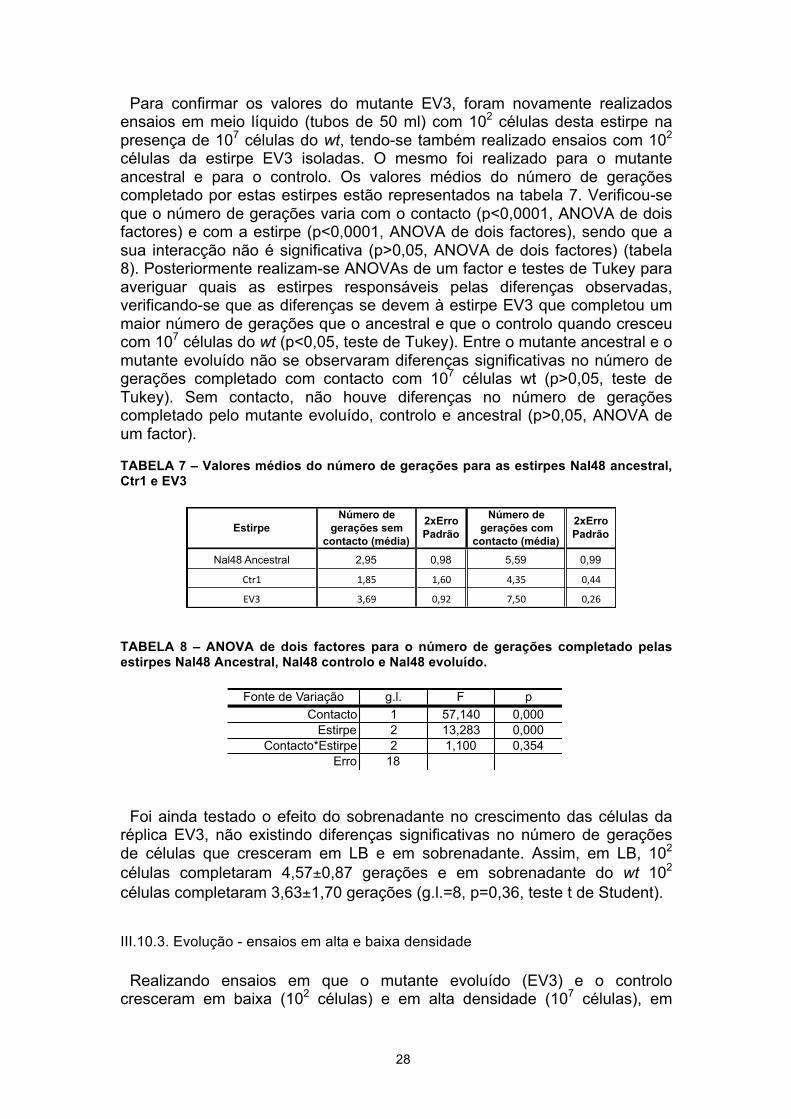
Para verificar se houve evolução do mutante Nal48, realizaram-se competições entre 102 células do mutante Nal48 ancestral, das cinco réplicas de evolução e do controlo da evolução (ctr1) com 107 células da estirpe wt, de modo a verificar a existência de diferenças no número gerações entre os mutantes sujeitos e não sujeitos a pressão selectiva ao crescerem na presença do wt. Pretendeu-se assim verificar se ocorreu evolução da interacção com o wt o que seria um indicador da evolução do mecanismo VDC. Estes ensaios foram realizados em meio líquido, em tubos de 50 ml com crescimento de três horas. Os valores médios do número de gerações completado por estas estirpes estão representados na tabela 5.
Posteriormente, realizou-se uma ANOVA de um factor, sendo possível verificar que o número de gerações dos mutantes varia com a estirpe (ancestral, controlo e 5 réplicas da evolução) (p<0,0001, ANOVA de um factor) (tabela 6), sendo que esta variação se deve a apenas uma réplica (EV3) (p<0,05, teste de Tukey). Este mutante evoluído completou um número de gerações mais elevado que o controlo e que o ancestral na presença de 107 células do wt (p<0,05, teste de Tukey), tendo ainda uma vantagem competitiva relativamente ao wt (fitness relativo=1,86, g.l.=8, p<0,05, teste T de Student).
TABELA 5 – Valores médios do número de gerações para as estirpes Nal48 ancestral, Ctr1 e as cinco réplicas evoluídas
Tabela 6 – ANOVA de um factor para o efeito da estirpe (ancestral, controlo e mutantes evoluídos) no número de gerações completado quando as estirpes cresceram com contacto com 107 células wt.
Fonte de Variação g.l. F pEstirpe 5 6,686 0,000
Erro 24
Estirpe Número de gerações (média)
2xErro Padrão
Nal48 Ancestral 4,33 1,17!"#$ %&'( )&*%+,$ (&%* )&-%+,. %&// )&%0+,( 0&(- )&(*+,% %&0- )&()+,' %&.% )&--
Fonte de Variação g.l. F pEstirpe 6 9,972 0,000
Erro 28
28
Para confirmar os valores do mutante EV3, foram novamente realizados ensaios em meio líquido (tubos de 50 ml) com 102 células desta estirpe na presença de 107 células do wt, tendo-se também realizado ensaios com 102 células da estirpe EV3 isoladas. O mesmo foi realizado para o mutante ancestral e para o controlo. Os valores médios do número de gerações completado por estas estirpes estão representados na tabela 7. Verificou-se que o número de gerações varia com o contacto (p<0,0001, ANOVA de dois factores) e com a estirpe (p<0,0001, ANOVA de dois factores), sendo que a sua interacção não é significativa (p>0,05, ANOVA de dois factores) (tabela 8). Posteriormente realizam-se ANOVAs de um factor e testes de Tukey para averiguar quais as estirpes responsáveis pelas diferenças observadas, verificando-se que as diferenças se devem à estirpe EV3 que completou um maior número de gerações que o ancestral e que o controlo quando cresceu com 107 células do wt (p<0,05, teste de Tukey). Entre o mutante ancestral e o mutante evoluído não se observaram diferenças significativas no número de gerações completado com contacto com 107 células wt (p>0,05, teste de Tukey). Sem contacto, não houve diferenças no número de gerações completado pelo mutante evoluído, controlo e ancestral (p>0,05, ANOVA de um factor). TABELA 7 – Valores médios do número de gerações para as estirpes Nal48 ancestral, Ctr1 e EV3
TABELA 8 – ANOVA de dois factores para o número de gerações completado pelas estirpes Nal48 Ancestral, Nal48 controlo e Nal48 evoluído.
Foi ainda testado o efeito do sobrenadante no crescimento das células da
réplica EV3, não existindo diferenças significativas no número de gerações de células que cresceram em LB e em sobrenadante. Assim, em LB, 102 células completaram 4,57±0,87 gerações e em sobrenadante do wt 102 células completaram 3,63±1,70 gerações (g.l.=8, p=0,36, teste t de Student).
III.10.3. Evolução - ensaios em alta e baixa densidade
Realizando ensaios em que o mutante evoluído (EV3) e o controlo cresceram em baixa (102 células) e em alta densidade (107 células), em
EstirpeNúmero de
gerações sem contacto (média)
2xErro Padrão
Número de gerações com
contacto (média)
2xErro Padrão
Nal48 Ancestral 2,95 0,98 5,59 0,99
!"#$ $%&' $%() *%+' )%**
,-+ +%(. )%./ 0%') )%/(
Fonte de Variação g.l. F pContacto 1 57,140 0,000
Estirpe 2 13,283 0,000Contacto*Estirpe 2 1,100 0,354
Erro 18

29
tubos, verificou-se que o mutante evoluído alcançou um maior número de gerações que o controlo (ctr1) em alta densidade. Em alta densidade o mutante evoluído completou 7,77±0,59 gerações e o mutante controlo completou 5,94±0,45 gerações (g.l.=8, p<0,01, teste t de Student). Em baixa densidade não se observaram diferenças significativas no número de gerações alcançado pelo mutante evoluído e pelo controlo (g.l.=8, p=0,53, teste t de Student). Assim, o controlo completou 5,09±0,70 gerações enquanto que o mutante evoluído completou 5,53±1,12 gerações (g.l.=8, p= 0,36, teste t de Student).

30

31
IV. DISCUSSÃO
IV.1. VDC como um novo mecanismo que permite a manutenção da diversidade bacteriana
Com este trabalho propomos o mecanismo VDC, como um novo
mecanismo de interacção bacteriana mediado pelo contacto célula a célula que permite a manutenção de diferentes estirpes e mesmo de diferentes espécies bacterianas numa população, permitindo a sua coexistência. De notar que no presente trabalho foram utilizados, na maioria dos casos, meios nutritivos muito complexos, não se podendo aplicar directamente o Princípio da Exclusão Competitiva. O Princípio da Exclusão competitiva, primeiramente formulado por V. Volterra (1927), mais conhecido como o princípio de Gause (1934), postula que apenas uma espécie competidora sobreviverá (Rescigno e Richardson, 1965). Segundo este princípio, o número de genótipos que coexistem não pode exceder o número de recursos limitantes, sendo que as causas para a coexistência de espécies em ambientes similares não foram ainda totalmente compreendidas. Assim, o mecanismo VDC poderá facilitar a manutenção da diversidade bacteriana numa população ao permitir que células em baixa densidade apresentem uma vantagem quando em contacto com uma população em alta densidade, tendo assim a oportunidade de colonizar novos habitats e manterem-se neles em baixas frequências através da interacção com a população já residente.
Verificou-se que este mecanismo depende do contacto célula a célula, não resultando da secreção de um metabolito por outras células, sendo que os resultados aqui apresentados não são explicáveis pelo quorum sensing, dependendo contudo da densidade populacional bacteriana. Por outro lado, este mecanismo difere também dos até então descritos envolvendo contacto célula a célula (Aoki et al., 2005, Lemonnier et al., 2008), uma vez que o crescimento bacteriano não é afectado negativamente, tendo-se, contrariamente a estes autores, verificado uma vantagem dependente de contacto e de uma baixa densidade bacteriana.
Dubey e Ben-Yehuda (2011) verificaram a existência de nanotubos que ligam células bacterianas vizinhas, funcionando como uma rede de troca de moléculas. Estes autores verificaram que estes nanotubos estão presentes em meio sólido, verificando contudo a sua inexistência em meio líquido. De acordo com os nossos resultados, o fenómeno VDC actua tanto em meio sólido como em meio líquido, sendo, portanto, incerto se estes nanotubos desempenham alguma função no fenómeno aqui descrito.
IV.2 Fenómeno VDC amplamente distribuído no mundo bacteriano
Neste trabalho foram utilizadas quatro espécies bacterianas: E. coli, Salmonella enterica, E. faecalis e S. aureus, sendo as duas primeiras Gram-negativas entéricas e as duas últimas Gram-positivas, sendo E. faecalis também entérica. De notar que E. coli e E. faecalis são comensais, tendo

32
uma relação benigna com o seu hospedeiro, colonizando o tracto-gastroentestinal de humanos e outros mamíferos. S. aureus é também uma estirpe comensal, colonizando assimptomaticamente a pele e a parte anterior das narinas. No entanto, estas três espécies, apesar de comensais, são também patogénicas oportunistas para o Homem, sendo capazes de infectar uma grande variedade de locais do corpo (Willems et al., 2011). A estirpe Salmonella enterica é patogénica, causando uma doença gastrointestinal numa grande variedade de animais, sendo ainda responsável por uma doença semelhante à febre tifóide no rato. Para além disto, isolados específicos foram encontrados em casos de doença severa em pombos (Chan et al., 2003).
Verificámos que o mecanismo VDC está presente tanto em estirpes Gram-positivas como Gram-negativas, actuando de forma intra e inter-específica, sendo que actua entre espécies evolutivamente tão distantes como S. aureus e E. coli, que apresentam também habitats muito diferentes. Estes resultados sugerem que este tipo de interacção deve estar amplamente distribuído no mundo bacteriano, podendo ser de extrema importância para que as espécies consigam colonizar com sucesso novos habitats.
Este mecanismo está também presente em Salmonella. Estes resultados são extremamente importantes, já que, como referido anteriormente, se trata de uma estirpe entérica patogénica que, conseguindo tirar proveito de estirpes já residentes (como E. coli) poderá permanecer com sucesso na flora microbiana indefinidamente mesmo que em baixas frequências. Não foram realizados ensaios entre E. coli e Salmonella, mas dado que o fenómeno VDC actua entre espécies tão distantes como E. coli e S. aureus, é-nos sugerido que este fenómeno actuará também entre espécies tão estreitamente relacionadas como E. coli e Salmonella.
A eliminação de estirpes resistentes a antibióticos poderá assim ser dificultada por mecanismos de interacção bacteriana mediadas pelo contacto célula a célula, sendo que, para além do quorum sensing (Bassler, 2002) – mecanismo independente de contacto – um alvo possível para combater a resistência poderia ser dirigida a estes mecanismos. Seria crucial descobrir o gene(s) envolvido(s) no fenómeno VDC de modo a desenvolver eventuais terapêuticas neste contexto.
É de notar que o fenómeno VDC não está presente em E. faecalis. Sabe-se que esta espécie forma cadeias de células. Estas estruturas poderiam compensar assim a ausência de uma população densa, ao permitirem que as células possuam um contacto íntimo entre si. No entanto, S. aureus forma também agregados celulares embora estruturalmente diferentes (cachos de uvas) verificando-se, contudo, o fenómeno VDC, o que não explica a sua ausência em E. faecalis. Por outro lado, não explica também o facto de E. coli não conseguir adquirir vantagem ao crescer com esta espécie, já que células de E. coli conseguiriam de qualquer forma estabelecer um contacto próximo com células de E. faecalis, mesmo na presença destas cadeias.
Assim, não é possível determinar as razões por detrás da ausência do fenómeno VDC em E. faecalis, pelo que seria interessante averiguar as causas que conduzem a este resultado. Seria importante testar uma estirpe de E. faecalis diferente da utilizada neste trabalho de modo a confirmar a ausência deste mecanismo nesta espécie.

33
IV.3. VDC e selecção dependente de frequência
Foi testado o fenómeno VDC em 30 mutantes distintos de E. coli K12 resistentes a três antibióticos distintos, tendo sido anteriormente testada a acção de selecção dependente de frequência estabilizante nestes mesmos mutantes (Alves, 2010).
Alves (2010) verificou a acção deste tipo de selecção em todos os mutantes resistentes a ácido nalidíxico testados. Nos mutantes Strep36, Rif3 e Rif4 foi também detectada a acção da selecção dependente de frequência estabilizante, não tendo sido detectada nos restantes.
Verificámos que em todos os mutantes em que não se verificou o fenómeno VDC, a acção da selecção dependente de frequência não foi também verificada, sendo que o contrário não é válido (muitos dos mutantes que exibem o fenómeno VDC não estão a sofrer acção deste tipo de selecção). Estes resultados sugerem que a mutação responsável pelo fenótipo dependente de frequência é diferente do gene responsável pelo fenómeno VDC.
Estudos anteriores demonstraram (Levin, 1972; Helling et al.,1987; Alves, 2010) a importância da selecção dependente de frequência estabilizante na manutenção da variabilidade em populações bacterianas. Com este trabalho encontrámos um mecanismo que poderá explicar as causas da vantagem de certas estirpes quando se encontram em baixas frequências e que aumentará grandemente a probabilidade do sucesso da colonização de novos habitats, permitindo a invasão de populações já residentes e permanência em baixas densidades.
É de salientar que todos os mutantes resistentes a ácido nalidíxico sofrerem tanto a acção da selecção dependente de frequência, como do fenómeno VDC. A resistência a este antibiótico é adquirida através de mutações cromossomais em genes envolvidos na replicação, tal como a gyrA. Sabe-se que ocorrem interacções pleiotrópicas entre este gene e muitos outros (Drlica et al., 2009; Jeong et al., 2006). Alves (2010) concluiu assim que, se outra mutação responsável pelo fenótipo dependente de frequência ocorreu noutro local do cromossoma, o seu efeito poderia ser melhorado por mutações no gene gyrA. IV.4. VDC e fases do crescimento bacteriano
Segundo Monod (1949), o crescimento bacteriano compreende seis fases distintas do crescimento: 1- fase lag (fase de latência, em que a taxa de crescimento é nula), 2- fase de aceleração (a taxa de crescimento aumenta), 3- fase exponencial (as bactérias apresentam uma taxa de crescimento constante), 4- fase de retardamento (a taxa de crescimento decresce), 5- fase estacionária (não há crescimento bacteriano) e 6- fase de declínio (morte celular).
Após a análise das curvas de crescimento em meio Mínimo com glucose ou com lactose, verificámos que o fenómeno VDC influencia a fase lag, encurtando-a. A fase de aceleração, parece ser assim atingida mais cedo nas células que cresceram com contacto com outras células em alta densidade, atingindo mais rapidamente a fase de crescimento exponencial.

34
A fase lag é a fase menos compreendida do ciclo de crescimento bacteriano. Segundo Swinnen et al., (2004), há vários factores que influenciam a duração desta fase, como alterações nas condições ambientais, a identidade e o fenótipo das bactérias, a fase de crescimento ou a história fisiológica das células e, em caso de pequeno volume de inóculo e condições rigorosas, o volume de inóculo no momento da alteração ambiental.
Sabe-se que a maquinaria para o uptake de ferro é activada nesta fase do ciclo de crescimento bacteriano, sendo a fase lag caracterizada pelo uptake e perda específicos mas transientes de iões metálicos (Rolfe et al., 2012), sendo que as concentrações de Cálcio e o Manganésio diminuem drasticamente durante as fases exponencial e estacionária, comparativamente com a fase lag, sugerindo que a necessidade fisiológica destes metais é mais elevada na fase lag, sugerindo também que estes metais poderão desempenhar um papel crucial na transição da fase lag para a fase exponencial (Rolfe et al., 2012).
Com os nossos resultados surge uma questão fundamental: porque há um encurtamento da fase lag quando as bactérias crescem em contacto com uma alta densidade de bactérias? Os mecanismos fisiológicos do fenómeno VDC não são conhecidos, sendo o facto das bactérias ‘arrancarem’ o seu crescimento mais depressa quando estão em alta densidade ainda incompreendido. Provavelmente existirá troca de moléculas entre células que exigirá proximidade entre elas, podendo estas desencadear o crescimento bacteriano. Contudo, estas moléculas são desconhecidas, podendo-se apenas excluir o dióxido de carbono. É de referir que o efeito do contacto com uma população com uma alta densidade celular descrito neste trabalho parece resgatar as células da morte celular. Alguns resultados sugerem que sem a presença de uma população densa há um decréscimo do número inicial de células ao fim de três horas, o que, por sua vez, sugere morte celular. O mesmo não se verifica quando as células crescem com contacto com 107 células, podendo as células estar a ser efectivamente resgatadas da morte celular. IV.5. O papel dos genes rpoS e lrp no fenómeno VDC
Como verificámos anteriormente, sem contacto com 107 células, os mutantes lrp- e rpoS- completaram um menor número de gerações comparativamente ao mutante lacY-, o que poderá estar relacionado com os custos inerentes às mutações que estas estirpes contemplam relativamente à estirpe lacY-. Com contacto, o mutante lrp- completou também um menor número de gerações comparativamente ao mutante lacY-, voltando assim a reflectir os custos destas mutações.
O gene lrp codifica para o factor de transcrição leucine-responsive regulatory protein (Lrp) e foi primeiramente identificado como um locus (livR) envolvido no transporte de aminoácidos de cadeia ramificada, tendo-se posteriormente verificado que mutações neste gene afectavam a expressão de operões envolvidos na biossíntese e degradação de aminoácidos e no transporte de péptidos. Estes resultados sugeriram que o gene lrp poderá ter um papel geral no metabolismo de aminoácidos (Calvo e Matthews, 1994).

35
Este gene foi reconhecido como sendo um gene regulador global do metabolismo de E. coli, especulando-se assim o significado fisiológico e adaptativo deste gene (Calvo e Matthews, 1994).
Landgraf et al. (1996) sugeriram que há uma correlacção inversa entre a expressão de lrp e a taxa de crescimento, sendo a sua expressão mais elevada quando a taxa de crescimento é menor. No entanto, em células que entram na fase estacionária, os autores verificaram uma elevada expressão deste gene (Landgraf et al., 1996).
O gene rpoS é o gene estrutural de σs, que corresponde à segunda subunidade sigma da RNA polimerase em E. coli e está envolvida na expressão de vários genes induzidos na fase estacionária (Lange, e Hengge-Aronis, 2006). Esta subunidade da RNA polymerase é também um regulador crucial de várias respostas ao stress em E. coli. Enquanto esta subunidade está praticamente ausente quando as células estão em rápido crescimento, este é fortemente induzido durante a entrada das células na fase estacionária e em muitas outras condições de stress, sendo essencial para a expressão de múltiplas resistências ao stress. Esta subunidade tem assim um enorme impacto, não apenas na tolerância ao stress, mas também em toda a fisiologia das células quando as condições não são as óptimas (Weber et al., 2005).
Tendo estes dois genes um papel fundamental na regulação de outros genes, a sua perda de função acarreta, muito provavelmente, custos de fitness. Uma vez que no mutante lrp- estes custos se manifestaram com e sem contacto com 107 células, este gene não parece ter nenhum papel no fenómeno VDC. No entanto, verificámos que com contacto com 107 células, o mutante rpoS- completou um número de gerações idêntico ao lacY-, sugerindo que o contacto com uma alta densidade de bactérias compensa os custos da mutação no gene rpoS. Estes resultados sugerem que, quando a estirpe cresce sozinha em baixa densidade, as bactérias encontram-se em condições de stress, sendo que a perda do gene rpoS se traduz numa diminuição de fitness uma vez que as bactérias não conseguem responder a esta condição. Contudo, quando a estirpe cresce com uma população com uma alta densidade celular, deixa de estar em condições de stress, pelo que o gene rpoS poderá não ser necessário, sendo a sua função desnecessária.
Zinser e Kolter (2004) verificaram que, em E. coli, a perda de função dos genes rpoS e lrp resultam num ganho de fitness na fase estacionária – fenótipo GASP (growth advantage in stationaty phase). Estes autores verificaram que estas duas classes de mutações são seleccionadas durante a evolução na fase estacionária e sugeriram que a selecção favorece a alteração da expressão genética em vez da geração de uma nova função. Afirmaram ainda que a implicação evolutiva desta descoberta se traduz no facto de a adaptação a novos ambientes ser dominada, pelo menos inicialmente, por alterações na regulação da actividade de genes pré-existentes, em vez de gerar novas actividades de novo (Zinser e Kolter, 2004).
Sabe-se que o regulão rpoS não é activado durante a fase lag (Rolfe et al., 2012). Outra explicação para a compensação de custos da perda de função deste gene através do contacto com uma população em alta densidade celular poderá ser uma nova regulação da actividade de outros genes, que

36
poderá reflectir-se favoravelmente na fase lag e na presença de uma população com uma alta densidade, conseguindo-se adaptar rapidamente. IV.6. Fenómeno VDC: uma perspectiva evolutiva
Um grande problema da Biologia Evolutiva consiste na explicação da cooperação. O problema da cooperação reflecte-se no motivo para que um indivíduo exiba um comportamento que acarreta custos mas beneficia outros indivíduos ou o grupo local (Hamilton 1963). A cooperação é assim vulnerável à invasão por cheaters que não cooperam adquirindo, contudo, o benefício através daqueles que cooperam. (Diggle et al., 2007a).
Verificámos que o mecanismo VDC actua em duas situações distintas para um mesmo mutante (designaremos mutante A): quando o mutante A se encontra numa população constituída por células idênticas a si, em alta densidade; quando o mutante A se encontra numa população com células wt de outra estirpe (p. ex. B), sendo que a vantagem se verifica quando o mutante A se encontra em baixa densidade, e a outra estirpe em alta densidade.
Colocam-se assim as seguintes questões: quando o mecanismo actua entre estirpes diferentes, serão as células da estirpe em raridade cheaters que, através de uma grande proximidade, roubam algo às células da estirpe que se encontra em alta densidade? Quando o mecanismo sucede entre a mesma estirpe, estaremos perante um comportamento de cooperação que exige o contacto entre células? Neste último caso, a selecção de parentesco explicaria facilmente um comportamento de cooperação entre indivíduos, já que as células são fortemente aparentadas. Assim, os indivíduos produziriam um bem comum acessível à população através do contacto entre células. Contudo, entre estirpes diferentes, a explicação torna-se mais difícil. Possivelmente, estaremos perante uma população de cheaters (população que se encontra em baixa densidade) que tira proveito de comportamentos de cooperação da estirpe que se encontra em alta densidade, não tendo, contudo, custos do comportamento de cooperação obtendo, todavia, os seus benefícios. Isto explicaria facilmente o facto de esta vantagem entre estirpes diferentes se manifestar apenas até determinada densidade celular. Diggle et al. (2007b), no contexto da evolução social do quorum sensing, verificaram que o fitness relativo de cheaters é dependente da sua frequência, possuindo um maior fitness quando são menos comuns. Nos mutantes em que o fenómeno VDC foi testado utilizando estirpes diferentes (mutantes Nal48, S57 e SA7) verificámos também a acção da selecção dependente de frequência (dados não mostrados), sendo que os mutantes apresentam uma vantagem competitiva relativamente ao wt quando se encontram em raridade.
Embora não tenhamos verificado se a vantagem dependente de contacto dos mutantes, ao crescer com uma estirpe diferente, depende da sua densidade inicial nas três horas de ensaio experimental (testámos apenas o efeito com o mutante em baixa densidade), o facto de a selecção dependente de frequência actuar neste mutantes poderá ser indicativo de que esta vantagem só existirá enquanto estes são menos comuns. Alves (2010) verificou a acção da selecção dependente de frequência estabilizante em vários mutantes resistentes a antibióticos (competições de 24 horas cujo

37
rácio inicial da estirpe mutante e do wt foi de 1:105), sendo que estes mutantes exibiam uma vantagem competitiva relativamente ao wt quando se encontravam em raridade. No entanto, quando as competições foram realizadas com um rácio inicial de 1:1 ou 102:1 (mutante para wt), os mutantes deixaram de exibir esta vantagem, exibindo por vezes desvantagem, mostrando assim o custo de resistência.
Comportamentos de cheating que se traduzem numa vantagem mediada pelo contacto quando os cheaters se encontram apenas em baixas densidades, explicariam o facto de apenas termos verificado evolução desta vantagem numa de cinco réplicas. De facto, para a evolução experimental, recorremos a duas estirpes diferentes. Explicações como mutações com um custo muito elevado ou mutações deletérias ou ainda perda de variabilidade genética devido a bottlenecks sucessivos poderiam ser invocadas. Contudo, outra explicação deverá ser considerada: a existência de um trade-off evolutivo entre o fitness dos cheaters (população de mutantes em baixa densidade) e a densidade celular atingida. É de notar que o mutante sujeito a evolução experimental estava também sujeito à acção da selecção dependente de frequência (Alves, 2010), mantendo esta vantagem competitiva na raridade após ter sido sujeito a evolução experimental. A selecção dependente de frequência estabilizante poderá assim impedir que a vantagem mediada pelo contacto aumente de forma significativa, impondo um limite nas densidades celulares atingidas pelos cheaters. O benefício retirado pelo contacto pode assim ser limitado para estes (p. ex. moléculas, nutrientes, etc.) que, tornando-se comuns, deixam de conseguir obter estes benefícios, tornando-se a evolução deste sistema difícil.
Relativamente à réplica na qual se observou evolução, não podemos garantir que esta tenha sido dirigida ao fenómeno VDC, dado que esta poderá ter ocorrido num outro caracter relevante para o fitness, tal como uma melhor utilização dos recursos. Podemos apenas garantir que as réplicas utilizadas tiveram origem num único clone ancestral, pelo que a mutação foi a única fonte para a variação genética. Contudo, o facto de a evolução ter conduzido a um aumento do número de gerações do mutante evoluído apenas quando a estirpe cresce na presença de uma população em alta densidade (o mesmo não se verifica quando a estirpe evoluída cresce isolada), sugere que, de facto, estamos perante a evolução da interacção entre estirpes. É de salientar que a densidade bacteriana atingida encontra-se na ordem de grandeza máxima que um mutante não evoluído consegue atingir na presença de 107 células ancestrais. Neste mutante, o efeito do contacto na obtenção do benefício pode ter sido assim optimizado.
Importa referir que foi isolado apenas um mutante de cada réplica da evolução, sendo que deveriam ter sido isolados mais mutantes.
Não observámos uma diminuição do fitness em nenhuma das réplicas da evolução comparativamente com o ancestral ou com o controlo, podendo ser excluída a acumulação de mutações deletérias no curso da evolução como explicação.
Possivelmente, se a evolução experimental tivesse sido realizada utilizando apenas uma estirpe, verificaríamos uma evolução convergente das cinco réplicas num aumento de fitness dos indivíduos devido a uma optimização do contacto e dos benefícios daí resultantes. A selecção de parentesco facilmente explicaria a evolução deste sistema.

38
IV.7. Perspectivas futuras
Com este trabalho pretendemos descrever um novo mecanismo de interacção bacteriana dependente do contacto entre células, que permite que mutantes que se encontram em baixa densidade, adquiram vantagem ao crescer com uma estirpe diferente numa alta densidade celular. Este mecanismo poderá revelar-se muito importante na colonização e manutenção de diferentes estirpes num mesmo habitat, mesmo estirpes que seriam eliminadas da população devido a custos inerentes a mutações.
Aqui, utilizámos estirpes resistentes a antibióticos, mostrando que o mecanismo VDC é eficaz na sua manutenção em baixas frequências em populações com outras estirpes e mesmo com outras espécies. Seria interessante averiguar se este mecanismo permitiria a manutenção de estirpes resistentes a antibióticos em densidades mais elevadas utilizando mutantes que não estão sob a acção de selecção dependente de frequência estabilizante.
Trindade et al. (2009) mostraram que num ambiente sem antibióticos o custo da resistência múltipla é mais baixo que o esperado devido a epistasia positiva entre alelos que conferem resistência. Ensaios de competição mostraram que o custo de resistência a um dado antibiótico depende da presença de alelos de resistência para outros antibióticos. Este trabalho demonstrou que a eliminação de estirpes multirresistentes é assinalavelmente mais difícil que o previsto. Seria assim extremamente relevante averiguar a relação entre o fenómeno VDC e a epistasia, de modo a verificar se estes podem actuar em conjunto promovendo a manutenção de estirpes multirresistentes.
Ao longo destas últimas duas décadas, a nossa visão acerca das bactérias alterou-se dramaticamente. As bactérias têm sido estudadas como populações de células que actuam independentemente (Keller e Surette, 2006) contudo, temos assistido a comportamentos sociais notáveis que já foram comparados aos comportamentos exibidos por insectos, vertebrados e humanos (Diggle et al., 2007a).
Com este trabalho propomos um novo mecanismo que envolve uma forte interacção entre células. Esta interacção possibilita que estirpes resistentes a antibióticos se mantenham indefinidamente numa população em baixas frequências. Assim, contrariamente ao esperado, a sua eliminação torna-se muito mais difícil, já que estas estirpes conseguem tirar proveito das células que estão ao seu redor, tendo assim uma oportunidade para invadir e manter-se numa população já residente. Os mecanismos moleculares e fisiológicos do mecanismo aqui descrito não são ainda conhecidos, sendo essencial compreender as causas da vantagem inerentes a este mecanismo. Qual o gene(s) envolvido(s)? De que benefícios fisiológicos falamos?
Por último, qual será a relação entre o mecanismo VDC e a origem da multicelularidade?

39
V. REFERÊNCIAS
Alves, D. (2010) The role of fitness frequency-dependent in the evolution of antibiotic resistance. Dissertação de mestrado. Faculdade de Ciências da Universidade de Lisboa, Lisboa. Aoki, S. K. et al. (2005) Contact-dependent inhibition of growth in Escherichia coli. Science 309, 1245–1248. Ayala, F. J. (1971) Competition between Species: Frequency Dependence. Science 171, 820–824. Baba, T. et al. (2006) Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Molecular Systems Biology 2, 2006.0008. Bassler, B. L. (2002) Small talk: Cell-to-Cell Communication in Bacteria. Cell 109, 421–424 Behe, M. J. (2010) Experimental evolution, loss-of-function mutations and “the first rule of adaptive evolution”. The Quarterly Review of Biology 85, 419–445. Bennett, A. F. e Lenski, R. E. (1999) Experimental evolution and its role in evolutionary physiology. American Zoologist 39, 346–362. Berg, R. D. (1996) The Indigenous gastrointestinal microflora. Trends in microbiology 4, 430–435. Bjorkman J. et al. (2000) Effects of environment on compensatory mutations to ameliorate costs of antibiotic resistance. Science 287, 1479–1482. Calvo, J. M. e Matthews, R. G. (1994) The Leucine-Responsive Regulatory Protein, a Global Regulator of Metabolism in Escherichia coli. Microbiological Reviews 58, 466–490. Caugant, D. A. et al. (1981) Genetic diversity and temporal variation in the E. coli population of a human host. Genetics 98, 467–490. Chan, K. et al. (2003) Genomic Comparison of Salmonella enterica Serovars and Salmonella bongori by Use of an S. enterica Serovar Typhimurium DNA Microarray. Journal of Bacteriology 185, 553–563. Costerton, J. W. (1999) Bacterial Biofilms: A Common Cause of Persistent Infections. Science 284, 1318 – 1322. Diggle, S. P. et al. (2007a) Evolutionary theory of bacterial quorum sensing: when is a signal not a signal? Phil. Trans. R. Soc. B. 362, 1241–1249. Diggle, S. P. et al. (2007b) Cooperation and conflict in quorum-sensing

40
bacterial populations. Nature 450, 411–415. Drlica, K. et al. (2009). Quinolones: action and resistance updated. Current Topics in Medicinal Chemistry 9, 981–998. Dubey, G.P. e Ben-Yehuda, S. (2011) Intercellular nano-tubes mediate bacterial communication. Cell 144, 590–600. Elena, S. F. et al. (1997) Frequency-dependent selection in a mammalian RNA virus. Evolution 51, 984–987. Garland, T. e Rose, M. R. (2009) Darwin’s Other Mistake. In Experimental Evolution: Concepts, Methods, and Applications of Selection Experiments, Garland, T. e Rose, M. R., eds. (Berkeley: University of California Press), pp. 3–9. Gratia, A. (1925) Sur un remarquable exemple d'antagonisme entre deux souches de colibacille. C. R. Soc. Biol. 93, 1040–1041. Hamilton, W. D. (1963) The evolution of altruistic behaviour. The American Naturalist 97, 354–356. Helling, R. B., et al. (1987). Evolution of Escherichia coli during growth in a constant environment. Genetics 116, 349–358. Hibbing M. E., et al. (2010) Bacterial competition: surviving and thriving in the microbial jungle. Nature Reviews Microbiology 8, 15–25. Jeong, K. S. et al. (2006). Analysis of pleiotropic transcriptional profiles: a case study of DNA gyrase inhibition. PLoS Genetics 2, 1464–1476. Kaiser, D. (2004) Signaling in myxobacteria. Annual Review of Microbiology 58, 75–98. Keller, L. e Surette, M. G. (2006) Nature Reviews Microbiology 4, 249–258. Kolenbrander, P. E. et al. (2002) Communication among Oral Bacteria. Microbiology and Molecular Biology Reviews 66, 486–505. Landgraf, J. R. et al. (1996) Effects of Nutrition and Growth Rate on Lrp Levels in Escherichia coli. Journal of Bacteriology 178, 6930–6936. Lange, R. e Hengge-Aronis, R. (2006) The nIpD gene is located in an operon with rpoS on the Escherichia coli chromosome and encodes a novel lipoprotein with a potential function in ceil wall formation. Molecular Microbiology 13, 733–743. Lederberg, J., e Tatum, E. L. (1946) Gene recombination in Escherichia coli. Nature 158, 558.

41
Lemonnier, M. et al. (2008) The evolution of contact-dependent inhibition in non-growing populations of Escherichia coli. Proc. R. Soc. B 275, 3–10. Levin, B. R. (1972) Coexistence of Two Asexual Strains on a Single Resource. Science 175, 1272–1274. Levin, B. R. (1988). Frequency-dependent selection in bacterial populations. Philos Trans R Soc Lond B Biol Sci 319, 459–472. Lenski, R. E. et al. (1991). Long-Term Experimental Evolution in Escherichia coli. I. Adaptation Divergence During 2,000 Generations. American Naturalist 138, 1315–1341. Ley, R. E. et al. (2006) Ecological and Evolutionary Forces Shaping Microbial Diversity in the Human Intestine. Cell 124, 837–848. Macpherson, A. J. e Harris, N. L. (2004) Interactions between commensal intestinal bacteria and the immune system. Nature Reviews 4, 478–485. Maynard Smith, J. e Szathmary, E. (1995) The Major Transitions in Evolution, (Oxford: Oxford University Press). Monod, J. (1949) The growth of bacterial cultures. Annual Review of Microbiology 3, 371–394. Ozbudak E. M., et al. (2004) Multistability in the lactose utilization network of Escherichia coli. Nature 427, 737–740. Ramadhan, A. A. e Hegedus, E. (2005) Survivability of vancomycin resistant enterococci and fitness cost of vancomycin resistance acquisition. Journal of Clinical Pathology 58, 744–746. Repaske, R. e Clayton, M. A. (1978) Control of Escherichia coli Growth by CO2. Journal of Bacteriology 135, 1162–1164. Rescigno, A. e Richardson I. W. (1965) On the competitive exclusion principle. Bulletin of Mathematical Biophysics 27, 85–89. Schneider, D. e Lenski, R. E. (2004) Dynamics of insertion sequence elements during experimental evolution of bacteria. Research in Microbiology 155, 319–327. Sears, H. J., et al. (1950) Persistence of individual strains of Escherichia coli in the intestinal tract of man. Journal of Bacteriology 59, 293–301. Rolfe, M. D. et al. (2012) Lag Phase Is a Distinct Growth Phase That Prepares Bacteria for Exponential Growth and Involves Transient Metal Accumulation. Journal of Bacteriology 194, 686–701. Swinnen, I. A. M. et al. (2004) Predictive modelling of the microbial lag phase:

42
a review. International Journal of Food Microbiology 94, 137–159. Thompson, J. N. (1999) The Evolution of Species Interactions. Science 284, 2116–2118. Trindade, S. et al. (2009) Positive Epistasis Drives the Acquisition of Multidrug Resistance. Plos Genetics 5, e1000578. Turner, P. E., et al. (1996) Tests of ecological mechanisms promoting the stable coexistence of two bacterial genotoypes. Ecology 77, 2119–2129. Weber, H. et al. (2005) Genome-Wide Analysis of the General Stress Response Network in Escherichia coli: σS-Dependent Genes, Promoters, and Sigma Factor Selectivity. Journal of Bacteriology 187, 1591–1603. Wei, X. et al. (2011) Heterologous protein transfer within structured myxobacteria biofilms. Molecular Microbiology 81, 315–326. Willems, R. J. L. et al. (2011) Population biology of Gram-positive pathogens: high-risk clones for dissemination of antibiotic resistance. FEMS Microbiology Reviews 35, 872–900. Yuste, E., et al. (2002) Frequency-dependent selection in human immunodeficiency virus type 1. Journal of General Virology 83, 103–106. Zinser, E. R. e Kolter, R. (2004) Escherichia coli evolution during stationary phase. Research in Microbiology 155, 328–336.